生物多样性与生态系统多功能性和多服务性的关系: 回顾与展望
井新, 贺金生
植物生态学报
2021, 45 ( 10):
1094-1111.
DOI: 10.17521/cjpe.2020.0154
近10年来, 生物多样性与生态系统多功能性(BEMF)的关系是生物多样性与生态系统功能领域新兴的热点研究方向。生态系统多功能性是指生态系统同时提供多重生态系统功能的能力, 受到群落和生态系统生态学研究者的广泛关注。该文简要回顾了生物多样性与生态系统多功能性关系研究历史, 侧重介绍了生态系统多功能性量化方法发展历程, 并总结了生物多样性与生态系统多功能性研究的主要趋势, 包括生物多样性维度、时空尺度和全球变化驱动因子等对生态系统多功能性的影响。同时, 回顾了近5年生物多样性与生态系统多功能性关系研究的新方法、新方向; 根据生态系统服务和生态系统功能的区别, 提出了生态系统多服务性(ecosystem multiserviceability, EMS)概念。最后简要介绍了生物多样性与生态系统多功能性、生物多样性与生态系统多服务性(BEMS)研究存在的不足及对未来的展望。
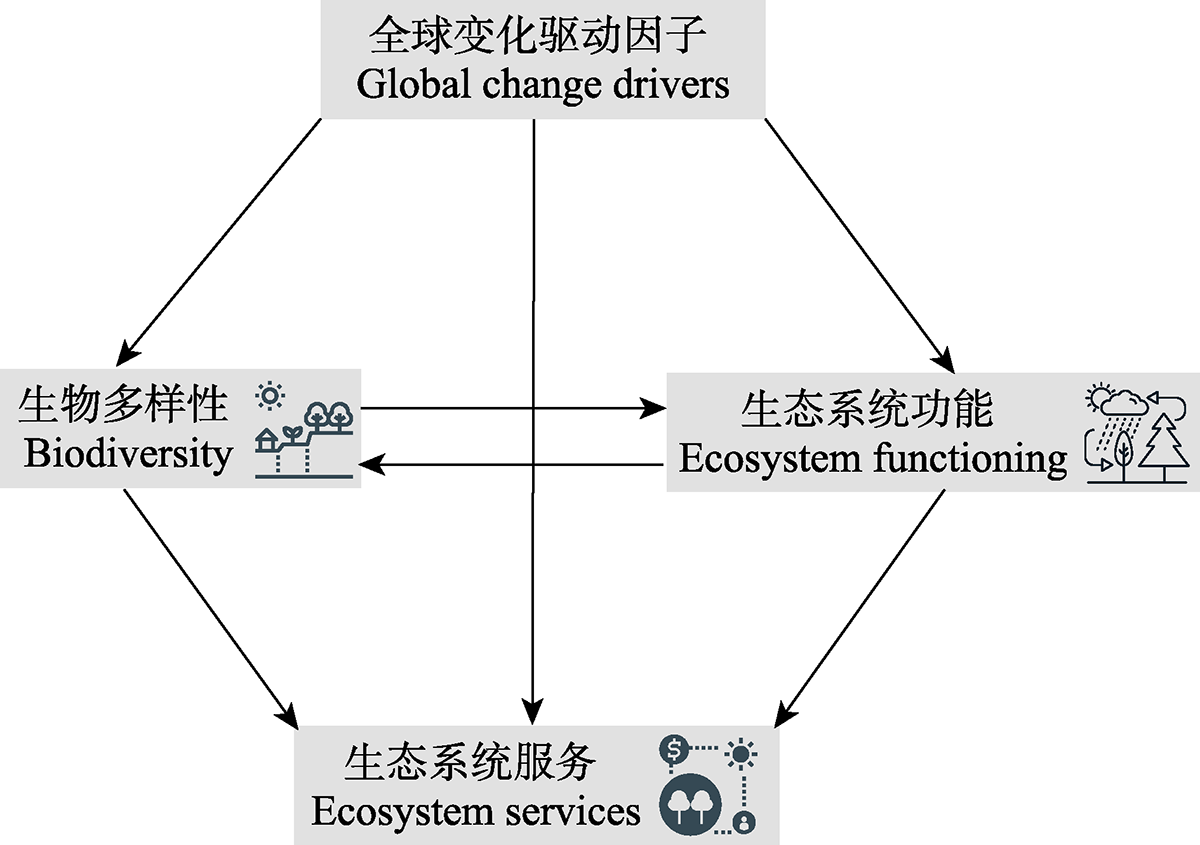
View image in article
图1
全球变化因子对生物多样性和生态系统功能和服务关系的调控作用。生物多样性和生态系统多功能性研究者常关注: 多个全球变化因子, 如气候变化、土地利用变化和氮沉降等对不同维度的生物多样性(如分类、功能和谱系多样性)、 生态系统功能和服务直接和间接影响。
正文中引用本图/表的段落
生物多样性与生态系统多功能性研究的主要特点是继承了BEF研究的传统(Hooper et al., 2012; Duffy et al., 2017; Isbell et al., 2017, 2018; Gonzalez et al., 2020), 具体可概括为关注: (1)生物多样性的维度; (2)时空依赖性; (3)驱动因子多样化等(图1)。下面将分别叙述。
除了生物多样性是生态系统多功能性的关键驱动因子, 还有其他的非生物因子和人类活动等影响BEMF关系, 例如气候、土壤、土地利用、放牧和氮沉降等。而这些因子对生态系统多功能性的影响是当前研究的难点。如图1所示, 全球变化因子对BEMF的影响主要体现在两个方面: (1)全球变化因子直接影响生物多样性和生态系统功能; (2)全球变化因子通过生物多样性间接影响生态系统功能, 或者全球变化因子通过生态系统功能间接影响生物多样性。由于缺少全球变化因子通过生态系统功能间接影响生物多样性的研究, 我们将着重介绍气候变化、土壤属性、土地利用和放牧干扰等主要的全球变化因子对生态系统多功能性的直接影响, 以及全球变化因子通过生物多样性对生态系统多功能性的间接影响。
生物多样性与生态系统多功能性研究的主要特点是继承了BEF研究的传统(Hooper et al., 2012; Duffy et al., 2017; Isbell et al., 2017, 2018; Gonzalez et al., 2020), 具体可概括为关注: (1)生物多样性的维度; (2)时空依赖性; (3)驱动因子多样化等(图1).下面将分别叙述. ...
生物多样性与生态系统多功能性研究的主要特点是继承了BEF研究的传统(Hooper et al., 2012; Duffy et al., 2017; Isbell et al., 2017, 2018; Gonzalez et al., 2020), 具体可概括为关注: (1)生物多样性的维度; (2)时空依赖性; (3)驱动因子多样化等(图1).下面将分别叙述. ...
生物多样性与生态系统多功能性研究的主要特点是继承了BEF研究的传统(Hooper et al., 2012; Duffy et al., 2017; Isbell et al., 2017, 2018; Gonzalez et al., 2020), 具体可概括为关注: (1)生物多样性的维度; (2)时空依赖性; (3)驱动因子多样化等(图1).下面将分别叙述. ... Warming and top predator loss drive ecosystem multifunctionality 2 2018 ... Key synthetic references in the field of biodiversity and ecosystem multifunctionality published in the past five years
生物多样性与生态系统多功能性研究的主要特点是继承了BEF研究的传统(Hooper et al., 2012; Duffy et al., 2017; Isbell et al., 2017, 2018; Gonzalez et al., 2020), 具体可概括为关注: (1)生物多样性的维度; (2)时空依赖性; (3)驱动因子多样化等(图1).下面将分别叙述. ...
生物多样性与生态系统多功能性研究的主要特点是继承了BEF研究的传统(Hooper et al., 2012; Duffy et al., 2017; Isbell et al., 2017, 2018; Gonzalez et al., 2020), 具体可概括为关注: (1)生物多样性的维度; (2)时空依赖性; (3)驱动因子多样化等(图1).下面将分别叙述. ...
生物多样性与生态系统多功能性研究的主要特点是继承了BEF研究的传统(Hooper et al., 2012; Duffy et al., 2017; Isbell et al., 2017, 2018; Gonzalez et al., 2020), 具体可概括为关注: (1)生物多样性的维度; (2)时空依赖性; (3)驱动因子多样化等(图1).下面将分别叙述. ...
生物多样性与生态系统多功能性研究的主要特点是继承了BEF研究的传统(Hooper et al., 2012; Duffy et al., 2017; Isbell et al., 2017, 2018; Gonzalez et al., 2020), 具体可概括为关注: (1)生物多样性的维度; (2)时空依赖性; (3)驱动因子多样化等(图1).下面将分别叙述. ...
生物多样性与生态系统多功能性研究的主要特点是继承了BEF研究的传统(Hooper et al., 2012; Duffy et al., 2017; Isbell et al., 2017, 2018; Gonzalez et al., 2020), 具体可概括为关注: (1)生物多样性的维度; (2)时空依赖性; (3)驱动因子多样化等(图1).下面将分别叙述. ... Explaining ecosystem multifunction with evolutionary models 1 2017 ... 在谱系多样性方面的研究比较少, 其中以Cadotte等(2017)利用不同的进化模型对生态系统多功能性的研究为代表, 也是国际上第一篇从谱系进化理论方面探讨BEMF关系的文章.到目前为止, 同时考虑生物多样性这三个维度的研究仅有Roger等(2016)对细菌多样性与生态系统多功能性的meta分析, Le Bagousse-Pinguet等(2019)在全球干旱区和Yan等(2020)在中国内蒙古草地的研究.其中, Yan等(2020)发现植物的功能性状多样性对生态系统多功能性的影响大于分类和谱系多样性, 同时发现干旱胁迫间接通过功能性状多样性影响生态系统多功能性.这一工作是国内学者发表的第一篇关于生物多样性三个维度与生态系统多功能性关系的文章. ... 不同利用方式下草地生态系统的多功能性与物种多样性 1 2019 ... 传统的认识认为放牧对生态系统多功能性有负面效应, 比如高强度放牧对生态系统多功能性的负面效应并不能被生物群落的正作用抵消(Sircely & Naeem, 2012; Zhang et al., 2016a).同样, Ren等(2018)也发现高强度放牧对生态系统多功能性的负效应远远大于地上和地下生物多样性的正作用.当然也有例外, 在热带半干旱生态系统的研究发现, 放牧形成的木本灌丛对保护土壤微生物功能和生态系统多功能性有至关重要的作用(Chandregowda et al., 2018).蔡艳等(2019)和Wang等(2020)的研究发现适度放牧干扰会提高生态系统多功能性和植物多样性.Wang等(2019)的研究表明牲畜的多样化能持续增强生态系统的多功能性.这些研究说明放牧强度、景观多样性以及牲畜的选择对BEMF有显著影响.尽管如此, 我们对放牧用地的生态恢复了解得很少.已有的研究表明, 停止放牧以后, 土壤质地尤其是粉粒和黏粒含量以及土壤持水能力下降, 土壤肥力和土壤有机碳含量也相应下降, 最终导致生态系统多功能性显著下降(Peco et al., 2017).而Grman等(2018)和Cruz-Alonso等(2019)的研究表明, 生态系统恢复需要时间, 长期恢复能增强生态系统多功能性, 主要取决于植物群落物种的更替、恢复策略和生态系统类型等因素. ... 不同利用方式下草地生态系统的多功能性与物种多样性 1 2019 ... 传统的认识认为放牧对生态系统多功能性有负面效应, 比如高强度放牧对生态系统多功能性的负面效应并不能被生物群落的正作用抵消(Sircely & Naeem, 2012; Zhang et al., 2016a).同样, Ren等(2018)也发现高强度放牧对生态系统多功能性的负效应远远大于地上和地下生物多样性的正作用.当然也有例外, 在热带半干旱生态系统的研究发现, 放牧形成的木本灌丛对保护土壤微生物功能和生态系统多功能性有至关重要的作用(Chandregowda et al., 2018).蔡艳等(2019)和Wang等(2020)的研究发现适度放牧干扰会提高生态系统多功能性和植物多样性.Wang等(2019)的研究表明牲畜的多样化能持续增强生态系统的多功能性.这些研究说明放牧强度、景观多样性以及牲畜的选择对BEMF有显著影响.尽管如此, 我们对放牧用地的生态恢复了解得很少.已有的研究表明, 停止放牧以后, 土壤质地尤其是粉粒和黏粒含量以及土壤持水能力下降, 土壤肥力和土壤有机碳含量也相应下降, 最终导致生态系统多功能性显著下降(Peco et al., 2017).而Grman等(2018)和Cruz-Alonso等(2019)的研究表明, 生态系统恢复需要时间, 长期恢复能增强生态系统多功能性, 主要取决于植物群落物种的更替、恢复策略和生态系统类型等因素. ... Biodiversity loss and its impact on humanity 1 2012 ... 生物多样性与生态系统功能(biodiversity and ecosystem functioning, BEF)的关系是群落生态学和生态系统生态学的研究热点, 有近30年的发展历史(Cardinale et al., 2012; Tilman et al., 2014; van der Plas, 2019).早期的BEF研究主要关注物种多样性与单个生态系统功能的关系, 比如植物物种丰富度对生态系统净初级生产力的影响(Hector et al., 1999; Loreau et al., 2001; Hooper et al., 2005).随着研究的深入, 研究者逐渐发现群落生物多样性越高, 凋落物分解越快, 抵御病虫害能力越强, 并伴随着土壤碳和养分循环等生态系统功能的提升.也就是说, 高的生物多样性能显著提升生态系统对多重生态系统功能的支持能力, 而生态系统的这一能力被定义为生态系统多功能性(ecosystem multifunctionality, EMF)(Hector & Bagchi, 2007; Gamfeldt et al., 2008). ... Woody shrubs increase soil microbial functions and multifunctionality in a tropical semi-arid grazing ecosystem 1 2018 ... 传统的认识认为放牧对生态系统多功能性有负面效应, 比如高强度放牧对生态系统多功能性的负面效应并不能被生物群落的正作用抵消(Sircely & Naeem, 2012; Zhang et al., 2016a).同样, Ren等(2018)也发现高强度放牧对生态系统多功能性的负效应远远大于地上和地下生物多样性的正作用.当然也有例外, 在热带半干旱生态系统的研究发现, 放牧形成的木本灌丛对保护土壤微生物功能和生态系统多功能性有至关重要的作用(Chandregowda et al., 2018).蔡艳等(2019)和Wang等(2020)的研究发现适度放牧干扰会提高生态系统多功能性和植物多样性.Wang等(2019)的研究表明牲畜的多样化能持续增强生态系统的多功能性.这些研究说明放牧强度、景观多样性以及牲畜的选择对BEMF有显著影响.尽管如此, 我们对放牧用地的生态恢复了解得很少.已有的研究表明, 停止放牧以后, 土壤质地尤其是粉粒和黏粒含量以及土壤持水能力下降, 土壤肥力和土壤有机碳含量也相应下降, 最终导致生态系统多功能性显著下降(Peco et al., 2017).而Grman等(2018)和Cruz-Alonso等(2019)的研究表明, 生态系统恢复需要时间, 长期恢复能增强生态系统多功能性, 主要取决于植物群落物种的更替、恢复策略和生态系统类型等因素. ... Embracing scale-dependence to achieve a deeper understanding of biodiversity and its change across communities 1 2018 ... 在空间上, 生物群落和生态系统功能都随着空间尺度的变化而变化.其中α、β和γ多样性是最常用来研究生物多样性对空间的依赖性(Chase et al., 2018).Pasari等(2013)最早研究了不同空间尺度上生物多样性对生态系统多功能性的影响, 发现α和γ多样性对生态系统多功能性有显著正效应, 而高的β多样性显著降低了生态系统多功能性的景观异质性.Yan等(2020)研究了α和β多样性对草地生态系统多功能性的影响.在森林生态系统方面的研究, van der Plas等(2016)参考生物多样性的计算方法, 利用阈值法, 研究了生物多样性对α、β和γ多功能性的影响, 发现群落物种组成的趋同, 会导致生态系统多功能性的趋同.同样, Mori等(2016)研究了不同空间尺度的生物多样性效应.与van der Plas等(2016)不同的是, Mori等(2016)利用欧式距离的算法, 研究了β多样性和多功能性的不相似性之间的关系.与Mori法相关的研究还有Barnes等(2018)和Martinez- Almoyna等(2019).Mori等(2018)进一步综述了β多样性对生态系统功能, 尤其是多功能性的影响.此外, Stürck和Verburg (2017)提出了生态系统多功能性的尺度依赖问题.Hölting等(2019b)提出了α和β多功能性的概念, 并首次提出利用Bray-Curtis指数来计算β多功能性.Thompson和Gonzalez (2016)借助集合群落的概念, 从理论上探讨了群落多样性、群落组成以及空间尺度对生态系统多功能性的影响.综合这些研究, 笔者认为对BEMF时空依赖性的研究在很大程度上得益于传统研究方法的启发和新量化方法和概念的提出. ... Rare microbial taxa as the major drivers of ecosystem multifunctionality in long-term fertilized soils 1 2020 ... 众所周知, 生物多样性由多个维度构成, 主要包括分类多样性、功能性状多样性和谱系多样性等.其中, 分类多样性是BEMF研究最多的内容, 主要是因为分类多样性数据容易获得.分类多样性研究集中在植物物种丰富度(Maestre et al., 2012a)、均匀度(Maestre et al., 2012b; Li et al., 2019)、优势度(Lohbeck et al., 2016)、种间关系(Slade & Roslin 2016; Slade et al., 2019)、功能群和群落组成(Maestre et al., 2012b; Wagg et al., 2014; Baeten et al., 2019; Li et al., 2019)等.随着高通量测序技术的发展, BEMF研究逐渐延伸到土壤微生物(Jing et al., 2015; Delgado-Baquerizo et al., 2016a; Wagg et al., 2019).此外, 多营养级的物种多样性是近年来研究的热点(Wagg et al., 2014; Jing et al., 2015; Lefcheck et al., 2015; Soliveres et al., 2016b; Martinez-Almoyna et al., 2019), 也是难点, 主要受限于物种鉴定水平和分子生物学技术的应用.在分类多样性方面的研究, 发现分类多样性是生态系统多功能性主要且关键的驱动因子, 但分类多样性效应在不同研究系统中, 有明显差异(van der Plas, 2019).比如在全球干旱区的研究, 植物物种丰富度对生态系统多功能性有较弱的正效应(Maestre et al., 2012a), 而在青藏高原的研究发现植物物种丰富度能解释42%的生态系统多功能性的空间变异(Jing et al., 2015).此外, 不同的研究对物种水平生物多样性重要性的认识也存在差异.比如, 常见种一般认为同时决定多个生态系统功能的水平(Lohbeck et al., 2016), 然而最新的研究表明稀有种对生态系统多功能性的贡献也是不可忽略的(Soliveres et al., 2016a; Chen et al., 2020).此外, 生物入侵种对生态系统多功能性的影响也值得关注(Ramus et al., 2017; Byers & Sotka, 2019; Sotka & Byers, 2019; Thomsen et al., 2019). ... Long-term recovery of multifunctionality in Mediterranean forests depends on restoration strategy and forest type 2 2019 ... 传统的认识认为放牧对生态系统多功能性有负面效应, 比如高强度放牧对生态系统多功能性的负面效应并不能被生物群落的正作用抵消(Sircely & Naeem, 2012; Zhang et al., 2016a).同样, Ren等(2018)也发现高强度放牧对生态系统多功能性的负效应远远大于地上和地下生物多样性的正作用.当然也有例外, 在热带半干旱生态系统的研究发现, 放牧形成的木本灌丛对保护土壤微生物功能和生态系统多功能性有至关重要的作用(Chandregowda et al., 2018).蔡艳等(2019)和Wang等(2020)的研究发现适度放牧干扰会提高生态系统多功能性和植物多样性.Wang等(2019)的研究表明牲畜的多样化能持续增强生态系统的多功能性.这些研究说明放牧强度、景观多样性以及牲畜的选择对BEMF有显著影响.尽管如此, 我们对放牧用地的生态恢复了解得很少.已有的研究表明, 停止放牧以后, 土壤质地尤其是粉粒和黏粒含量以及土壤持水能力下降, 土壤肥力和土壤有机碳含量也相应下降, 最终导致生态系统多功能性显著下降(Peco et al., 2017).而Grman等(2018)和Cruz-Alonso等(2019)的研究表明, 生态系统恢复需要时间, 长期恢复能增强生态系统多功能性, 主要取决于植物群落物种的更替、恢复策略和生态系统类型等因素. ...
生物多样性与生态系统多功能性研究的主要特点是继承了BEF研究的传统(Hooper et al., 2012; Duffy et al., 2017; Isbell et al., 2017, 2018; Gonzalez et al., 2020), 具体可概括为关注: (1)生物多样性的维度; (2)时空依赖性; (3)驱动因子多样化等(图1).下面将分别叙述. ...
生物多样性与生态系统多功能性研究的主要特点是继承了BEF研究的传统(Hooper et al., 2012; Duffy et al., 2017; Isbell et al., 2017, 2018; Gonzalez et al., 2020), 具体可概括为关注: (1)生物多样性的维度; (2)时空依赖性; (3)驱动因子多样化等(图1).下面将分别叙述. ...
生物多样性与生态系统多功能性研究的主要特点是继承了BEF研究的传统(Hooper et al., 2012; Duffy et al., 2017; Isbell et al., 2017, 2018; Gonzalez et al., 2020), 具体可概括为关注: (1)生物多样性的维度; (2)时空依赖性; (3)驱动因子多样化等(图1).下面将分别叙述. ...
生物多样性与生态系统多功能性研究的主要特点是继承了BEF研究的传统(Hooper et al., 2012; Duffy et al., 2017; Isbell et al., 2017, 2018; Gonzalez et al., 2020), 具体可概括为关注: (1)生物多样性的维度; (2)时空依赖性; (3)驱动因子多样化等(图1).下面将分别叙述. ...
生物多样性与生态系统多功能性研究的主要特点是继承了BEF研究的传统(Hooper et al., 2012; Duffy et al., 2017; Isbell et al., 2017, 2018; Gonzalez et al., 2020), 具体可概括为关注: (1)生物多样性的维度; (2)时空依赖性; (3)驱动因子多样化等(图1).下面将分别叙述. ...
生物多样性与生态系统多功能性研究的主要特点是继承了BEF研究的传统(Hooper et al., 2012; Duffy et al., 2017; Isbell et al., 2017, 2018; Gonzalez et al., 2020), 具体可概括为关注: (1)生物多样性的维度; (2)时空依赖性; (3)驱动因子多样化等(图1).下面将分别叙述. ...
生物多样性与生态系统多功能性研究的主要特点是继承了BEF研究的传统(Hooper et al., 2012; Duffy et al., 2017; Isbell et al., 2017, 2018; Gonzalez et al., 2020), 具体可概括为关注: (1)生物多样性的维度; (2)时空依赖性; (3)驱动因子多样化等(图1).下面将分别叙述. ...
生物多样性与生态系统多功能性研究的主要特点是继承了BEF研究的传统(Hooper et al., 2012; Duffy et al., 2017; Isbell et al., 2017, 2018; Gonzalez et al., 2020), 具体可概括为关注: (1)生物多样性的维度; (2)时空依赖性; (3)驱动因子多样化等(图1).下面将分别叙述. ...
本文的其它图/表
|

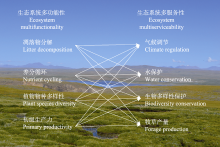
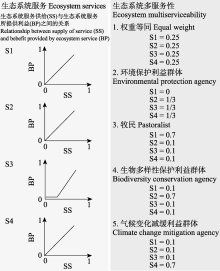
{kind=link}