花生(Arachis hypogaea )是我国重要的经济作物, 总产量居油料作物之首, 是我国重要的食用油源、食品工业理想原料和出口创汇作物。花生是需钙量大的作物, 每形成100 kg荚果, 吸钙量达2.0-2.5 kg, 缺钙会导致花生空壳、烂果(张二全等, 1994 ; 吴旭银等, 2007 )。我国东南沿海花生主要种植于旱砂地, 该区域土壤盐基饱和度较低, 酸度较大, 土壤中钙大量流失, 严重影响花生籽粒的发育和产量的形成, 甚至造成绝产。
Ca是植物生长发育过程中必需的营养元素, 参与植物从种子萌发、生长分化、形态建成到开花结果等全过程。Ca2+ 信使参与调控植物抗逆反应, 如调节酶的活性、蛋白质磷酸化和基因表达等(Monroy & Dhindsa, 1995 ; Gong et al ., 1997 ; Yang & Poovaiah, 2003 ; Hetherington & Brownlee, 2004 )。环境胁迫下植物能通过提高胞内游离Ca2+ 浓度使其与钙调素(CaM)结合, 从而启动一系列生理生化过程, 形成细胞的逆境伤害适应机制, 从而使Ca2+ 和CaM在植物对逆境胁迫的感受、传递、响应和适应过程中起中心作用。国内外对花生Ca营养的研究较多, 主要涉及Ca对花生的生长发育、产量及产量构成因素、籽仁品质的影响, 以及形态解剖特征、生理生化特性和分子生物学机理, 初步明确了Ca对花生荚果发育、抗逆性、产量构成等方面的重要作用(Adams et al ., 1992 ; 周卫和林葆, 1996 , 2001 ; Chamlong et al ., 1999 ; 刘秀梅等, 2005 ; Rahman, 2006 ; 王才斌等, 2008 ; 李岳等, 2012 )。Ca对淀粉的水解与转运、蛋白质的合成等过程具有不可或缺的调控作用, 缺Ca可使蛋白质和RNA合成能力降低。植物产量与品质的形成主要是在一系列碳、氮代谢酶催化下进行的, 有关花生代谢酶活性与产量和品质关系的研究较多(张智猛等, 2008 ; 郭峰等, 2009 ; 崔光军等, 2010 ; 张佳蕾等, 2013 ), 较高的代谢酶活性有利于产量提高和品质改善。
前人对钙肥在花生生理特性、产量及品质方面的影响已有较多的研究, 但钙肥对花生叶片碳、氮代谢酶活性的影响研究较少, 更没有针对酸性土壤增施钙肥影响产量和品质的酶学机理进行研究。因此通过研究钙肥对叶片碳、氮代谢酶活性的影响, 从酶学角度来解释增施钙肥对酸性土壤花生产量和品质的影响机理, 对花生钙营养机理研究具有重要的理论意义, 可为沿海地区酸性土花生合理施肥提供技术指导。
1 材料和方法
1.1 试验地概况
试验于2013年在威海文登市西楼社区金岭山庄的丘陵地(文登试验点)进行, 于2014年在日照东港区三庄镇三庄二村的丘陵山地(三庄实验点)进行。文登市试验点耕层土壤含有机质10.10 g·kg-1 、水解性氮77.6 mg·kg-1 、速效磷58.1 mg·kg-1 、速效钾142.0 mg·kg-1 、交换性钙0.20 g·kg-1 、交换性镁0.09 g·kg-1 , pH值4.56。三庄试验点的耕层土壤含有机质12.24 g·kg-1 、水解性氮56.4 mg·kg-1 、速效磷74.2 mg·kg-1 、速效钾98.5 mg·kg-1 、交换性钙0.64 g·kg-1 、交换性镁0.57 g·kg-1 , pH值4.73。
1.2 试验设计
选用花生品种‘花育22号’为试验材料, 文登试验点于2013年5月1日播种, 9月20日收获, 三庄试验点于2014年4月29日播种, 9月18日收获。起垄覆膜栽培, 垄距80 cm, 垄面宽50 cm, 垄上播种两行, 行距30 cm, 穴距16.6 cm, 每穴两粒, 种植密度是15万穴·hm-2 。
试验设置3个钙肥处理, 分别为每667 m2 施钙镁磷肥(P2 O5 : CaO: SiO2 : MgO = 14:28:40:5) 0、50、100 kg, 折算成CaO为0 (T0 )、14 (T1 )、28 kg (T2 )。每个处理小区面积为66.7 m2 , 3次重复。为了消除钙镁磷肥里磷、硅、镁的差异, 在T0 和T1 处理小区的肥料中分别加入1.40和0.70 kg的P2 O5 (化学纯), 4.00和2.00 kg的SiO2 (用硅酸钠折算), 0.50和0.25 kg的MgO (用硫酸镁折算), 混合均匀后施入, 使3个处理中的磷、硅、镁保持一致。每667 m2 施用三元复合肥(N:P2 O5 :K2 O = 20:15:10) 100 kg, 其他栽培管理措施按一般花生高产田要求进行。
1.3 测定项目与方法
1.3.1 荚果产量及产量构成因素
收获时每个处理选有代表性的地段取样10株, 考察单株结果数、饱果数和双仁果数, 烘干测定单株产量。各处理逐小区实测, 去掉小区的边行和两头, 测量实收面积, 刨收、摘果、去杂, 自然风干, 统计荚果产量。在风干荚果中随机选取有经济价值的荚果计算双仁果率和出仁率。
1.3.2 籽仁品质
在各处理自然风干荚果中选取均匀一致的荚果脱壳, 用多功能谷物近红外分析仪(DA7250 Perten, Hägersten, Sweden)测定各处理籽仁的蛋白质和脂肪含量, 氨基酸组分和脂肪酸组分, 以及油酸/亚油酸(O/L)值等。
1.3.3 碳、氮代谢酶活性
自开花期开始到成熟期, 每隔15天在各处理内选取长势相同、受光方向一致的主茎倒3叶, 迅速放入液氮罐中带回室内, 于-40 ℃低温冰箱中保存, 用于碳、氮代谢相关酶活性测定, 每个小区3次重复。
谷氨酰胺合成酶(GS)、谷氨酸脱氢酶(GDH)、谷氨酸合成酶(GOGAT)活性参照Lin和Kao (1996)的方法测定; 谷草转氨酶(GOT)、谷丙转氨酶(GPT)活性参照吴良欢等(1998)的方法测定; 蔗糖合成酶(SS)、蔗糖磷酸合成酶(SPS)活性参照薛应龙和上海植物生理学会(1985)的方法测定; 磷酸烯醇式丙酮酸羧化酶(PEPCase)活性参照González等(1998)的方法测定。
GS活力单位定义: 每克组织在反应体系中每30 min使540 nm下OD值变化0.01定义为一个酶活力单位。GDH、GOGAT活力单位定义: 每克组织在反应体系中每分钟消耗1 nmol·L-1 的NADH定义为一个酶活力单位。GOT、GPT活力单位定义: 每克组织在反应体系中每分钟使340 nm下OD值变化0.001定义为一个酶活力单位。SS、SPS活力单位定义: 每克组织在反应体系中每分钟催化产生1 µg蔗糖定义为一个酶活力单位。PEPCase活力单位定义: 每克组织在反应体系中每分钟消耗1 nmol·L-1 的NADH定义为一个酶活力单位。
1.4 数据处理
采用DPS 7.05软件对数据进行分析, 用最小显著极差法(LSD )对平均数进行显著性检验; 采用Origin 8软件作图。两个试验点的产量和品质数据同时进行分析, 代谢酶活性变化规律基本一致, 仅采用文登试验点的数据进行分析。
2 结果和分析
2.1 酸性土增施钙肥对花生荚果产量的影响
由表1 可知, 酸性土增施钙肥能显著增加花生的荚果产量。两个试验点增施钙肥处理的单株结果数、双仁果率、单株果重、出仁率和荚果产量均显著高于不施钙肥处理, T1 和T2 处理间除了荚果产量差异较大外, 单株结果数、双仁果率、单株果重和出仁率差异不显著。文登试验点T1 和T2 处理分别比T0 增产25.27%和18.81%, 三庄试验点T1 和T2 处理分别增产28.57%和24.49%。钙肥对酸性土花生出仁率的影响较大, 文登试验点T1 和T2 处理分别比T0 提高17.22%和16.35%, 三庄试验点T1 和T2 处理分别比T0 提高12.97%和11.60%, 两试验点T1 处理的出仁率均高于T2 处理。说明酸性土增施钙肥提高产量的原因主要是增加了单株结果数, 提高了双仁果率, 同时显著提高了籽仁饱满度。
2.2 酸性土增施钙肥对花生籽仁品质的影响
酸性土增施钙肥显著增加了花生籽仁蛋白质和脂肪含量, 提高了赖氨酸和总氨基酸含量, 增加了油酸相对含量提高了O/L值(表2 )。文登试验点T1 和T2 处理的籽仁蛋白质含量分别比T0 提高2.11%和1.85%, 脂肪含量分别提高2.46%和2.68%。三庄试验点T1 和T2 处理的籽仁蛋白质含量分别比T0 提高1.92%和1.26%, 脂肪含量分别提高3.55%和2.48%。文登试验点T1 和T2 处理的蛋白质和脂肪含量差异不显著, 而三庄试验点T1 处理的蛋白质和脂肪含量显著高于T2 处理。文登试验点T1 处理的油酸相对含量显著高于T0 和T2 处理, 亚油酸相对含量显著低于后两者, 从而使其O/L值显著增高, T0 和T2 处理差异不显著。三庄试验点T1 和T2 处理的油酸相对含量均显著高于T0 , 其亚油酸相对含量显著低于后者, T1 和T2 处理之间差异不显著。
2.3 酸性土增施钙肥对花生叶片氮代谢酶活性的影响
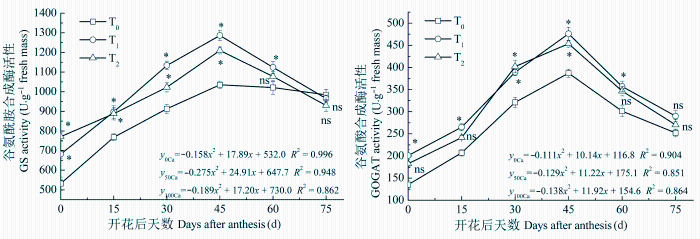
2.3.1 GS和GOGAT活性
GS和GOGAT是处于氮代谢中心的多功能酶, 参与多种氮代谢的调节。增施钙肥处理与不施钙肥处理的花生叶片GS和GOGAT活性变化趋势基本一致, 开花之后其活性先增高后降低(图1 )。酸性土增施钙肥显著提高了花生叶片的GS和GOGAT活性, 对GS活性提高幅度在生育前期较大, 到成熟期略低于不施钙肥处理。将各处理数据拟合成二次方程, 曲线拟合效果较好。方程特征系数表明, 增施钙肥处理GS活性达到峰值的时间约在花后45天, 而不施钙肥处理达到峰值的时间大约在花后56天; 增施钙肥处理GOGAT活性达到峰值的时间约在花后43天, 而不施钙肥达到峰值的时间约在花后46天。增施钙肥使GS活性达到峰值的时间提早10天左右, 使GOGAT活性峰值时间提早3天左右, 并能较长时间保持较高的活性, 有利于氮素积累。两个增施钙肥处理相比, T1 的GS活性明显高于T2 处理, 而两者的GOGAT活性差异较小。
图1 酸性土增施钙肥对花生叶片谷氨酰胺合成酶和谷氨酸合成酶活性的影响(平均值±标准误差)。T0 、T1 、T2 , 每667 m2 施CaO 0、14、28 kg。
Fig. 1 Effects of calcium fertilizer on glutamine synthetase (GS) and glutamate synthetase (GOGAT) activities in leaves of peanut in acidic soil (mean ± SE ). T0 , T1 , T2 represent the addition of CaO with 0, 14, 28 kg·667 m-2 , respectively. *, p < 0.05; ns, p > 0.05.
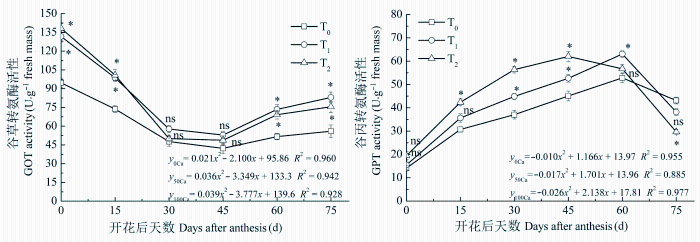
2.3.2 GOT和GPT活性
GOT和GPT是植物体内最重要的转氨酶。各处理GOT活性变化趋势表现为先降低后升高, 而GPT活性表现为先升高后降低(图2 )。增施钙肥处理的GOT和GPT活性显著高于不施钙肥处理。增施钙肥与不施钙肥处理的GOT活性在花后30天之前和花后45天之后差异较大, 而增施钙肥的GPT活性在花后60天之前较高, 花后75天时低于不施钙肥处理。GPT活性方程特征系数表明: T0 处理的GPT活性达到峰值的时间约在花后58天, T1 与T2 处理达到峰值的时间约在花后50天和41天, 增施钙肥明显早于不施钙肥处理。T1 与T2 处理的GOT活性差异较小, 而GPT活性差异较大, T2 处理花后45天之前显著高于T1 处理, 花后60天之后低于后者。
图2 酸性土增施钙肥对花生叶片谷草转氨酶和谷丙转氨酶活性的影响(平均值±标准误差)。T0 、T1 、T2 , 每667 m2 施CaO 0、14、28 kg。
Fig. 2 Effects of calcium fertilizer on glutamic-oxalacetic transaminase (GOT) and glutamate pyruvate transaminase (GPT) activities in leaves of peanut in acidic soil (mean ± SE ). T0 , T1 , T2 represent the addition of CaO with 0, 14, 28 kg·667 m-2 , respectively. *, p < 0.05; ns, p > 0.05.
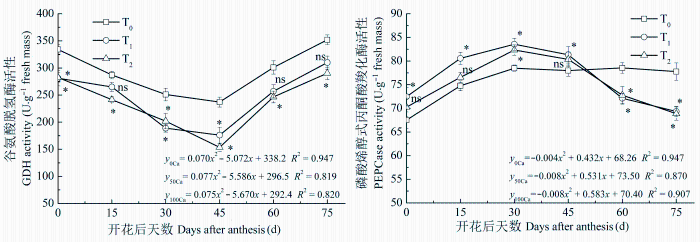
2.3.3 GDH活性
GDH对GS/GOGAT循环起辅助作用。图3 表明 酸性土花生叶片GDH活性与GS/GOGAT变化趋势相反, 开花后先降低后增高。不施钙肥处理的各时期GDH活性均显著高于施钙处理, T0 处理在花后45天时GDH活性分别比T1 和T2 处理高34.66%和54.40%。T1 与T2 处理的GDH活性差异较小。生育后期各处理的GDH活性增大的原因是花生开始衰老, GS/GOGAT同化作用变小, 植株主要依靠GDH途径进行氮素同化。
图3 酸性土增施钙肥对花生叶片谷氨酸脱氢酶和磷酸烯醇式丙酮酸羧化酶活性的影响(平均值±标准误差)。T0 、T1 、T2 , 每667 m2 施CaO 0、14、28 kg。
Fig. 3 Effects of calcium fertilizer on glutamate dehydrogenase (GDH) and phosphoenol pyruvate carboxylase (PEPCase) activities in leaves of peanut in acidic soil (mean ± SE ). T0 , T1 , T2 represent the addition of CaO with 0, 14, 28 kg·667 m-2 , respectively. *, p < 0.05; ns, p > 0.05.
2.4 酸性土增施钙肥对花生叶片碳代谢酶活性的影响
2.4.1 PEPCase活性
PEPCase是植物体内碳同化的关键酶。酸性土花生不施钙肥与增施钙肥处理的PEPCase活性变化趋势不同, 前者变化比较平稳, 从开花后活性逐渐增强, 生育后期也保持较高活性, 后者变化趋势呈抛物线型, 在花后30天达到最大后开始降低(图3 )。增施钙肥处理的PEPCase活性在花后45天之前明显高于不施钙肥处理, 之后活性降低较快, 到花后60天之后显著低于不施钙肥处理。增施钙肥的两个处理相比, T1 在花后30天之前PEPCase活性显著高于T2 , 花后45天之后差异较小。方程特征系数表明: T1 处理的PEPCase活性达到峰值的时间约在花后33天, 略早于T2 处理。
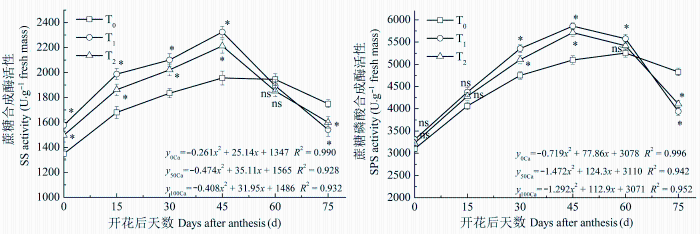
2.4.2 SS和SPS活性
SPS催化的蔗糖合成途径是叶片蔗糖合成的主要途径, 有些学者认为光合器官中SS也具有较强的催化蔗糖合成的能力。由图4 可知, 酸性土花生叶片的SS和SPS活性变化趋势基本相同, 随生育期推进表现为先增高后降低。增施钙肥处理的SS和SPS在生育前期的活性显著高于不施钙肥处理, T1 处理的SS和SPS活性要高于T2 处理。T1 和T2 处理的SS活性在花后45天之前显著高于T0 , 之后活性下降速率较快, 花后60天之后低于T0 。与SS表现基本一致, T1 和T2 处理的SPS活性在生育前期较高, 花后30天至花后45天(荚果膨大充实期)差异显著, 花后75天时其活性显著低于T0 。SS和SPS活性变化曲线拟合效果良好, 方程特征系数表明, 酸性土增施钙肥花生叶片的SS和SPS活性达到峰值的时间明显早于不施钙肥处理(SS平均提早10天, SPS平均提早12天)。
图4 酸性土增施钙肥对花生叶片蔗糖合成酶和蔗糖磷酸合成酶活性的影响(平均值±标准误差)。T0 、T1 、T2 , 每667 m2 施CaO 0、14、28 kg。
Fig. 4 Effects of calcium fertilizer on sucrose synthase (SS) and sucrose phosphate synthase (SPS) activities in leaves of peanut in acidic soil (mean ± SE ). T0 , T1 , T2 represent the addition of CaO with 0, 14, 28 kg·667 m-2 , respectively. *, p < 0.05; ns, p > 0.05.
3 讨论
近20年来, 过于重视增施增产效果明显的氮、磷、钾肥, 作为花生所需的大量元素的钙肥被忽视, 施用极少或不施, 一方面降低了土壤可交换钙的含量, 另一方面过量施肥使土壤酸化, 固态的钙得不到活化和释放, 严重影响了钙素的吸收利用。沿海酸性土壤由于钙流失严重, 花生因缺钙造成空秕率为9%-15%。施钙肥可明显增加花生荚果和籽仁产量, 主要是增加了单株结果数和出仁率、提高了果重(周录英等, 2008 ; 高芳等, 2011 )。本课题组前期研究表明酸性土花生增施钙肥后开花量高于对照, 施钙荚果产量明显提高。本研究结果表明钙肥不同用量均显著增加了酸性土花生的荚果产量, 两个试验点平均增产24.29%。增施钙肥显著提高了单株结果数和单株果重, 同时对提高双仁果率和出仁率效果显著。三庄试验点增施钙肥增产效果要好于文登试验点增施钙肥处理, 可能是试验点地力水平不同造成的, 也可能是选择的钙浓度梯度不是最合适的, 有待于进一步研究。有研究表明: 施钙肥花生的地上部营养生长比不施钙肥的差, 合理施用钙肥可以抑制花生主茎和侧枝旺长(吴文新等, 2001 ; 周录英等, 2008 )。本研究T1 处理的出仁率要高于T2 处理, 而荚果产量显著高于后者, 说明本试验条件下每667 m2 施14 kg CaO效果要好于每667 m2 施28 kg, 原因可能是高浓度钙抑制了花生的营养生长, 从而影响了产量。
施钙不仅提高了花生籽仁中脂肪和蛋白质含量, 降低了籽仁中可溶性糖含量, 而且可提高脂肪的O/L值, 增加蛋白质组分中含量不足的赖氨酸和蛋氨酸含量, 从而延长花生制品货架寿命, 改善花生蛋白质品质(周录英等, 2008 ; 高芳等, 2011 )。吴文新等(2001)的研究表明, 施钙肥比对照花生种仁粗蛋白含量提高13.78%, 粗脂肪含量提高3.08%, 而粗淀粉降低14.17%。酸性土花生增施钙肥对品质的影响与上述研究结果一致, 增施钙肥均显著提高了两个试验点花生籽仁蛋白质和脂肪含量, 对赖氨酸和总氨基酸的提高幅度也较大, 说明钙肥对酸性土花生籽仁品质改善效果良好。T1 处理显著提高了文登试验点花生的油酸相对含量, 降低了亚油酸相对含量, 从而使O/L值显著提高, T2 处理的O/L值与T0 差异较小; 而T1 和T2 处理均显著提高了三庄试验点花生的油酸含量, 降低了亚油酸含量, 两者差异不显著。
高等植物体内绝大部分NH4 + 是通过谷氨酰胺合成酶/谷氨酸合成酶(GS/GOGAT)循环同化。而谷氨酸脱氢酶(GDH)主要在植物的衰老过程及逆境如高温和水分胁迫等状况下进行NH4 + 同化功能。施用钙肥提高了花生不同生育时期叶片中硝酸还原酶(NR)、GS和GOGAT等活性, 促进植株对氮素的吸收, 增加籽仁中蛋白质含量(王媛媛等, 2014 )。鲁翠涛等(2002)研究小麦(Triticum aestivum )培养液中加入Ca2+ , 在1 mmol·L-1 浓度时, 氮素代谢关键酶NR、GS等活性随着Ca2+ 浓度的增加而增大。本研究中增施钙肥显著提高了酸性土花生叶片的GS、GOGAT和GPT活性, 酶活性达到峰值的时间要早于不施钙处理, 且峰值持续时间较长, 有利于氮素积累。王志强等(2008)通过研究不同浓度的外源Ca2+ 对小麦幼苗氮素代谢的影响, 说明小麦幼苗不同氮同化途径对Ca2+ 的响应不同, GS途径比GDH途径对小麦氮素同化量的增加作用更大。酸性土花生不施钙肥处理的GDH活性要显著高于增施钙肥处理, 说明酸性土花生缺钙时主要是靠GDH途径来进行氮素的代谢和积累。值得一提的是, T2 处理的酸性土花生叶片GPT活性要显著高于T1 处理, 这也是前者籽仁的赖氨酸和总氨基酸含量高于后者的原因。
前人研究表明, 低浓度Ca2+ 明显抑制高粱(Sorghum bicolor )、马齿苋(Portulaca oleracea )及大叶伽兰菜(Kalanchoe laciniata )叶片PEPCase活性(焦进安和施教耐, 1987 ), 5 mmol·L-1 Ca2+ 抑制花生叶片磷酸烯醇式丙酮酸羧化酶、磷酸烯醇式丙酮酸磷酸酯酶和丙酮酸激酶活性(郭振飞等, 1997 )。本研究结果有所不同, 增施钙肥提高了酸性土花生花后45天之前的叶片PEPCase活性, T1 处理的活性要高于T2 处理。SS和SPS催化的蔗糖合成途径是植物叶片蔗糖合成的主要途径。增施钙肥显著提高了酸性土花生叶片SS和SPS活性, 花后45天之前SS提高幅度较大, 花后30天至花后60天SPS提高幅度较大, T1 处理的两种酶活性要高于T2 处理。花后60天之后施钙处理的PEPCase、SS和SPS活性显著低于不施钙处理, 原因可能是缺钙花生光合产物转化运输速率低(周恩生等, 2008 ), 进入成熟期后供钙, 结实正常的花生地上部逐渐枯黄, 而缺钙的花生仍还继续开花, 出现“老来青”现象造成的。
综上所述, 酸性土增施钙肥能显著增加花生的单株结果数, 提高双仁果率, 增加荚果产量并显著提高出仁率。增施钙肥增产的同时能显著改善酸性土花生的籽仁品质, 提高蛋白质和脂肪含量, 增加赖氨酸和油酸含量, 提高O/L值。酸性土增施钙肥能显著提高花生叶片GS、GOGAT、GPT等氮代谢关键酶活性和PEPCase、SS、SPS等碳代谢关键酶活性, 延长了酶活性峰值持续期, 促进光合产物向蛋白质和脂肪转化, 同时提高了籽仁的饱满度, 这是增施钙肥提高蛋白质和脂肪含量的主要原因。本试验条件下不同钙肥施用量相比, 每667 m2 施14 kg的CaO成本较低, 其增产幅度最大, 对品质改善效果最好。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Adams JF Hartzog DL Nelson DB 1992 ). Supplemental calcium application on yield, grade, and seed quality of runner peanut.
Agronomy Journal , 85 , 86 -93 .
[本文引用: 1]
[2]
Chamlong K Sorasak M Bunlua S 1999 ). Effect of calcium rate on the decreasing of unfilled pod of peanut (Arachis hypogaea L.) grown in sandy loam soil in Yasothorn Province.
Thai Journal of Soils and Fertilizers , 21 , 184 -192 .
[本文引用: 1]
[3]
Cui GJ Liu FZ Wan YS 2010 ). Relationship between dry matter accumulation and sucrose metabolism during pod development in peanut (Arachis hypogaea L.).
Scientia Agricultura Sinica , 43 , 3965 -3973 .
[本文引用: 1]
(in Chinese with English abstract) [崔光军 , 刘风珍 , 万勇善 (2010 ). 花生荚果干物质积累与蔗糖代谢的相关性研究.
中国农业科学 , 43 , 3965 -3973 .]
[本文引用: 1]
[4]
Gao F Zhang JL Yang CT Zhang F Yang XK Lin YJ Li XD 2011 ). Effects of applying calcium on peanut physiological characteristics, its yield and kernel quality under cadmium stress.
Chinese Journal of Applied Ecology , 22 , 2907 -2912 .
[本文引用: 2]
(in Chinese with English abstract) [高芳 , 张佳蕾 , 杨传婷 , 张凤 , 杨晓康 , 林英杰 , 李向东 (2011 ). 钙对镉胁迫下花生生理特性、产量和品质的影响水.
应用生态学报 , 22 , 2907 -2912 .]
[本文引用: 2]
[5]
Gong M Chen SN Song YQ Li ZG 1997 ). Effect of calcium and calmodulin on intrinsic heat tolerance in relation to antioxidant systems in maize seedlings.
Australian Journal of Plant Physiology , 24 , 371 -379 .
[本文引用: 1]
[6]
González MC Osuna L Echevarría C Vidal J Cejudo FJ 1998 ). Expression and localization of phosphoenolpyruvate carboxylase in developing and germinating wheat grains.
Plant Physiology , 116 , 1249 -1258 .
[7]
Guo F Wan SB Wang CB Li XG Meng JJ Xu PL 2009 ). Nitrogen metabolism and relative enzyme activities of the peanut relay-cropped with wheat.
Plant Nutrition and Fertilizer Science , 15 , 416 -421 .
[本文引用: 1]
(in Chinese with English abstract) [郭峰 , 万书波 , 王才斌 , 李新国 , 孟静静 , 徐平丽 (2009 ). 麦套花生氮素代谢及相关酶活性变化研究.
植物营养与肥料学报 , 15 , 416 -421 .]
[本文引用: 1]
[8]
Guo ZF Lu SY Li MQ 1997 ). Effect of calcium on some enzyme activities of carbohydrate metabolism in peanut leaves.
Journal of Tropical and Subtropical Botany , 5 (3 ), 35 -38 .
[本文引用: 1]
(in Chinese with English abstract) [郭振飞 , 卢少云 , 李明启 (1997 ). 钙对花生叶片糖代谢有关酶活性的影响.
热带亚热带植物学报 , 5 (3 ), 35 -38 .]
[本文引用: 1]
[9]
Hetherington AM Brownlee C 2004 ). The generation of Ca2 + lcium signals in plants.
Annual Review of Plant Biology , 55 , 401 -427 .
[本文引用: 1]
[10]
Jiao JA Shi JN 1987 ). Inhibition of PEP carboxylase by calcium.
Plant Physiology Communications , (4 ), 42 -45 .
[本文引用: 1]
(in Chinese with English abstract) [焦进安 , 施教耐 (1987 ). Ca2 +对植物磷酸烯醇式丙酮酸羧化酶的抑制作用.
植物生理学通讯, (4 ), 42 -45 .]
[本文引用: 1]
[11]
Li Y Wang YF Wang ML Zhao CX Lü TZ 2012 ). Effects of calcium application rate on senescence characteristics and yield of peanut.
Journal of Qingdao Agricultural University (Natural Science) , 29 , 89 -93 .
[本文引用: 1]
(in Chinese with English abstract) [李岳 , 王月福 , 王铭伦 , 赵长星 , 吕泰宗 (2012 ). 施钙对花生衰老特性和产量的影响.
青岛农业大学学报(自然科学版) , 29 , 89 -93 .]
[本文引用: 1]
[12]
Lin CC Kao CH 1996 ). Disturbed ammonium assimilation is associated with growth inhibition of roots in rice seedlings caused by NaCl.
Plant Growth Regulation , 18 , 233 -238 .
[13]
Liu XM Zhang FD Zhang SQ He XS Wang RF Feng ZB Wang YJ 2005 ). Responses of peanut to nano-calcium carbonate.
Plant Nutrition and Fertilizer Science , 11 , 385 -389 .
[本文引用: 1]
(in Chinese with English abstract) [刘秀梅 , 张夫道 , 张树清 , 何绪生 , 王茹芳 , 冯兆滨 , 王玉军 (2005 ). 纳米碳酸钙在花生上的施用效果研究.
植物营养与肥料学报 , 11 , 385 -389 .]
[本文引用: 1]
[14]
Lu CT Li HS Wang XK 2002 ). Effect of calcium on key enzymes of nitrogen assimilation and related to protein phosphorylation in wheat.
Plant Nutrition and Fertilizer Science , 2002 , 8 (1 ), 110 -114 .
(in Chinese with English abstract) [鲁翠涛 , 李合生 , 王学奎 (2002 ). 钙对小麦氮同化关键酶活性的影响及其与蛋白质磷酸化的关系
. 植物营养与肥料学报 , 8 (1 ), 110 -114 .]
[15]
Monroy AF Dhindsa RS 1995 ). Low-temperature signal transduction: Induction of cold acclimation-specific genes of alfalfa by calcium at 25 ℃.
The Plant Cell , 7 , 321 -331 .
[本文引用: 1]
[16]
Rahman MA 2006 ). Effect of calcium and Bradyrhizobium inoculation of the growth, yield and quality of groundnut (A. hypogaea L.).
Bangladesh Journal of Scientific and Industrial Research , 41 , 181 -188 .
[本文引用: 1]
[17]
Wang CB Wu ZF Zhao PJ Zheng YP Cheng B Wan SB Sun KX 2008 ). Effects of prohexadione calcium on some physiological characteristics and pod yield of peanut.
Plant Nutrition and Fertilizer Science , 14 , 1160 -1164 .
[本文引用: 1]
(in Chinese with English abstract) [王才斌 , 吴正锋 , 赵品绩 , 郑亚萍 , 成波 , 万书波 , 孙奎香 (2008 ). 调环酸钙对花生某些生理特性和产量的影响.
植物营养与肥料学报 , 14 , 1160 -1164 .]
[本文引用: 1]
[18]
Wang YY Wang XY Ren J Zhang JL Gao B Li XD 2014 ). Effects of different Ca fertilizer applications on nitrogen metabolism in peanut.
Journal of Anhui Agricultural Sciences , 42 , 11289 -11291 .
[本文引用: 1]
(in Chinese with English abstract) [王媛媛 , 王先芸 , 任嘉 , 张佳蕾 , 高波 , 李向东 (2014 ). 钙肥不同用量对花生氮代谢的影响.
安徽农业科学 , 42 , 11289 -11291 .]
[本文引用: 1]
[19]
Wang ZQ Wang CL Lin TB 2008 ). Effects of exogenous Ca2 + on nitrogen metabolism in wheat seedlings.
Acta Ecologica Sinica , 28 , 3662 -3667 .
(in Chinese with English abstract) [王志强 , 王春丽 , 林同保 (2008 ). 外源钙离子对小麦幼苗氮素代谢的影响.
生态学报 , 28 , 3662 -3667 .]
[20]
Wu LH Jiang SH Tao QN 1998 ). Colorimetric determination of glutamate oxaloacetate transaminase (GOT) and glutamate pyruvate transaminase (GPT) activity in plants and its application.
Chinese Journal of Soil Science , 29 (3 ), 136 -138 .
(in Chinese) [吴良欢 , 蒋式洪 , 陶勤南 (1998 ). 植物转氨酶(GOT和GPT)活度比色测定方法及其应用.
土壤通报 , 29 (3 ), 136 -138 .]
[21]
Wu WX Chen JJ Zhou ES Lin HZ Lin Q You XY 2001 ). Effects of calcium and boron on the growth, yield and quality of peanut.
Subtropical Plant Science , 30 (2 ), 20 -23 .
[本文引用: 1]
(in Chinese with English abstract) [吴文新 , 陈家驹 , 周恩生 , 林汉章 , 林群 , 游雪芸 (2001 ). 钙、硼对花生生长、产量和品质的影响.
亚热带植物科学 , 30 (2 ), 20 -23 .]
[本文引用: 1]
[22]
Wu XY Wu HP Li YS Zhou YF Han JL Zhang SX 2007 ). The absorption characteristics of Ca, Mg and S in plastic mulching peanut.
Plant Nutrition and Fertilizer Science , 13 , 171 -174 .
[本文引用: 1]
(in Chinese with English abstract) [吴旭银 , 吴贺平 , 李彦生 , 周印富 , 韩金铃 , 张淑霞 (2007 ). 地膜覆盖花生对钙、镁、硫吸收特性的研究.
植物营养与肥料学报 , 13 , 171 -174 .]
[本文引用: 1]
[23]
Xue YL 1985 ). Experimental Manual of Plant Physiology . Shanghai Science and Technology Press , Shanghai .
(in Chinese) [薛应龙 , 上海植物生理学会 (1985 ). 植物生理学实验手册 . 上海科学技术出版社 , 上海 .]
[24]
Yang TB Poovaiah BW 2003 ). Calcium/calmodulin-mediated signal network in plants.
Trends in Plant Science , 8 , 505 -512 .
[本文引用: 1]
[25]
Zhang EQ Wu ZS Li YX Zhai GX Lu HS 1994 ). The effect of calcium application on peanut under soil calcium levels.
Journal of Peanut Science , (2 ), 4 -6 .
[本文引用: 1]
(in Chinese) [张二全 , 武占社 , 李英霞 , 翟桂香 , 逯怀森 (1994 ). 土壤钙素水平对花生施钙效果的影响
. 花生科技 , (2 ), 4 -6 .]
[本文引用: 1]
[26]
Zhang JL Gao F Lin YJ Wang YY Yang CT Zhang F Li YH Li XD 2013 ). Differences in seed kernel quality and related enzyme activities of different quality type peanut cultivars.
Chinese Journal of Applied Ecology , 24 , 481 -487 .
[本文引用: 1]
(in Chinese with English abstract) [张佳蕾 , 高芳 , 林英杰 , 王媛媛 , 杨传婷 , 张凤 , 李艳红 , 李向东 (2013 ). 不同品质类型花生品质性状及相关酶活性差异.
应用生态学报 , 24 , 481 -487 .]
[本文引用: 1]
[27]
Zhang ZM Wan SB Ning TY Dai LX 2008 ). Effects of nitrogen level on nitrogen metabolism and correlating enzyme activity in peanut.
Journal of Plant Ecology (Chinese Version) , 32 , 1407 -1416 .
[本文引用: 1]
(in Chinese with English abstract) [张智猛 , 万书波 , 宁堂原 , 戴良香 (2008 ). 氮素水平对花生氮素代谢及相关酶活性的影响.
植物生态学报 , 32 , 1407 -1416 .]
[本文引用: 1]
[28]
Zhou ES Chen JJ Wang F Wang HP Chen HC He Y 2008 ). Microanalysis characteristics and physiological of peanut pods and plant biochemical responses under Ca stress.
Fujian Journal of Agricultural Sciences , 23 , 318 -321 .
[本文引用: 1]
(in Chinese with English abstract) [周恩生 , 陈家驹 , 王飞 , 王煌平 , 陈惠成 , 何盈 (2008 ). 钙胁迫下花生荚果微区特征及植株生理生化反应变化.
福建农业学报 , 23 , 318 -321 .]
[本文引用: 1]
[29]
Zhou LY Li XD Wang LL Tang X Lin YJ 2008 ). Effects of different Ca applications on physiological characteristics, yield and quality in peanut.
Acta Agronomica Sinica , 34 , 879 -885 .
[本文引用: 3]
(in Chinese with English abstract) [周录英 , 李向东 , 王丽丽 , 汤笑 , 林英杰 (2008 ). 钙肥不同用量对花生生理特性及产量和品质的影响.
作物学报 , 34 , 879 -885 .]
[本文引用: 3]
[30]
Zhou W Lin B 1996 ). A study on the symptom and ultrastructure of calcium deficiency for peanut.
Scientia Agricultura Sinica , 29 (4 ), 53 -57 .
[本文引用: 1]
(in Chinese with English abstract) [周卫 , 林葆 (1996 ). 花生缺钙症状与超微结构特征的研究.
中国农业科学 , 29 (4 ), 53 -57 .]
[本文引用: 1]
[31]
Zhou W Lin B 2001 ). Study on reproductive growth and seed quality of peanut as affected by calcium supply.
Plant Nutrition and Fertilizer Science , 7 , 205 -210 .
[本文引用: 1]
(in Chinese with English abstract) [周卫 , 林葆 (2001 ). 受钙影响的花生生殖生长及种子素质研究.
植物营养与肥料学报 , 7 , 205 -210 .]
[本文引用: 1]
Supplemental calcium application on yield, grade, and seed quality of runner peanut.
1
1992
... Ca是植物生长发育过程中必需的营养元素, 参与植物从种子萌发、生长分化、形态建成到开花结果等全过程.Ca2+ 信使参与调控植物抗逆反应, 如调节酶的活性、蛋白质磷酸化和基因表达等(Monroy & Dhindsa, 1995 ; Gong et al ., 1997 ; Yang & Poovaiah, 2003 ; Hetherington & Brownlee, 2004 ).环境胁迫下植物能通过提高胞内游离Ca2+ 浓度使其与钙调素(CaM)结合, 从而启动一系列生理生化过程, 形成细胞的逆境伤害适应机制, 从而使Ca2+ 和CaM在植物对逆境胁迫的感受、传递、响应和适应过程中起中心作用.国内外对花生Ca营养的研究较多, 主要涉及Ca对花生的生长发育、产量及产量构成因素、籽仁品质的影响, 以及形态解剖特征、生理生化特性和分子生物学机理, 初步明确了Ca对花生荚果发育、抗逆性、产量构成等方面的重要作用(Adams et al ., 1992 ; 周卫和林葆, 1996 , 2001 ; Chamlong et al ., 1999 ; 刘秀梅等, 2005 ; Rahman, 2006 ; 王才斌等, 2008 ; 李岳等, 2012 ).Ca对淀粉的水解与转运、蛋白质的合成等过程具有不可或缺的调控作用, 缺Ca可使蛋白质和RNA合成能力降低.植物产量与品质的形成主要是在一系列碳、氮代谢酶催化下进行的, 有关花生代谢酶活性与产量和品质关系的研究较多(张智猛等, 2008 ; 郭峰等, 2009 ; 崔光军等, 2010 ; 张佳蕾等, 2013 ), 较高的代谢酶活性有利于产量提高和品质改善. ...
Effect of calcium rate on the decreasing of unfilled pod of peanut (Arachis hypogaea L.) grown in sandy loam soil in Yasothorn Province.
1
1999
... Ca是植物生长发育过程中必需的营养元素, 参与植物从种子萌发、生长分化、形态建成到开花结果等全过程.Ca2+ 信使参与调控植物抗逆反应, 如调节酶的活性、蛋白质磷酸化和基因表达等(Monroy & Dhindsa, 1995 ; Gong et al ., 1997 ; Yang & Poovaiah, 2003 ; Hetherington & Brownlee, 2004 ).环境胁迫下植物能通过提高胞内游离Ca2+ 浓度使其与钙调素(CaM)结合, 从而启动一系列生理生化过程, 形成细胞的逆境伤害适应机制, 从而使Ca2+ 和CaM在植物对逆境胁迫的感受、传递、响应和适应过程中起中心作用.国内外对花生Ca营养的研究较多, 主要涉及Ca对花生的生长发育、产量及产量构成因素、籽仁品质的影响, 以及形态解剖特征、生理生化特性和分子生物学机理, 初步明确了Ca对花生荚果发育、抗逆性、产量构成等方面的重要作用(Adams et al ., 1992 ; 周卫和林葆, 1996 , 2001 ; Chamlong et al ., 1999 ; 刘秀梅等, 2005 ; Rahman, 2006 ; 王才斌等, 2008 ; 李岳等, 2012 ).Ca对淀粉的水解与转运、蛋白质的合成等过程具有不可或缺的调控作用, 缺Ca可使蛋白质和RNA合成能力降低.植物产量与品质的形成主要是在一系列碳、氮代谢酶催化下进行的, 有关花生代谢酶活性与产量和品质关系的研究较多(张智猛等, 2008 ; 郭峰等, 2009 ; 崔光军等, 2010 ; 张佳蕾等, 2013 ), 较高的代谢酶活性有利于产量提高和品质改善. ...
花生荚果干物质积累与蔗糖代谢的相关性研究.
1
2010
... Ca是植物生长发育过程中必需的营养元素, 参与植物从种子萌发、生长分化、形态建成到开花结果等全过程.Ca2+ 信使参与调控植物抗逆反应, 如调节酶的活性、蛋白质磷酸化和基因表达等(Monroy & Dhindsa, 1995 ; Gong et al ., 1997 ; Yang & Poovaiah, 2003 ; Hetherington & Brownlee, 2004 ).环境胁迫下植物能通过提高胞内游离Ca2+ 浓度使其与钙调素(CaM)结合, 从而启动一系列生理生化过程, 形成细胞的逆境伤害适应机制, 从而使Ca2+ 和CaM在植物对逆境胁迫的感受、传递、响应和适应过程中起中心作用.国内外对花生Ca营养的研究较多, 主要涉及Ca对花生的生长发育、产量及产量构成因素、籽仁品质的影响, 以及形态解剖特征、生理生化特性和分子生物学机理, 初步明确了Ca对花生荚果发育、抗逆性、产量构成等方面的重要作用(Adams et al ., 1992 ; 周卫和林葆, 1996 , 2001 ; Chamlong et al ., 1999 ; 刘秀梅等, 2005 ; Rahman, 2006 ; 王才斌等, 2008 ; 李岳等, 2012 ).Ca对淀粉的水解与转运、蛋白质的合成等过程具有不可或缺的调控作用, 缺Ca可使蛋白质和RNA合成能力降低.植物产量与品质的形成主要是在一系列碳、氮代谢酶催化下进行的, 有关花生代谢酶活性与产量和品质关系的研究较多(张智猛等, 2008 ; 郭峰等, 2009 ; 崔光军等, 2010 ; 张佳蕾等, 2013 ), 较高的代谢酶活性有利于产量提高和品质改善. ...
花生荚果干物质积累与蔗糖代谢的相关性研究.
1
2010
... Ca是植物生长发育过程中必需的营养元素, 参与植物从种子萌发、生长分化、形态建成到开花结果等全过程.Ca2+ 信使参与调控植物抗逆反应, 如调节酶的活性、蛋白质磷酸化和基因表达等(Monroy & Dhindsa, 1995 ; Gong et al ., 1997 ; Yang & Poovaiah, 2003 ; Hetherington & Brownlee, 2004 ).环境胁迫下植物能通过提高胞内游离Ca2+ 浓度使其与钙调素(CaM)结合, 从而启动一系列生理生化过程, 形成细胞的逆境伤害适应机制, 从而使Ca2+ 和CaM在植物对逆境胁迫的感受、传递、响应和适应过程中起中心作用.国内外对花生Ca营养的研究较多, 主要涉及Ca对花生的生长发育、产量及产量构成因素、籽仁品质的影响, 以及形态解剖特征、生理生化特性和分子生物学机理, 初步明确了Ca对花生荚果发育、抗逆性、产量构成等方面的重要作用(Adams et al ., 1992 ; 周卫和林葆, 1996 , 2001 ; Chamlong et al ., 1999 ; 刘秀梅等, 2005 ; Rahman, 2006 ; 王才斌等, 2008 ; 李岳等, 2012 ).Ca对淀粉的水解与转运、蛋白质的合成等过程具有不可或缺的调控作用, 缺Ca可使蛋白质和RNA合成能力降低.植物产量与品质的形成主要是在一系列碳、氮代谢酶催化下进行的, 有关花生代谢酶活性与产量和品质关系的研究较多(张智猛等, 2008 ; 郭峰等, 2009 ; 崔光军等, 2010 ; 张佳蕾等, 2013 ), 较高的代谢酶活性有利于产量提高和品质改善. ...
钙对镉胁迫下花生生理特性、产量和品质的影响水.
2
2011
... 近20年来, 过于重视增施增产效果明显的氮、磷、钾肥, 作为花生所需的大量元素的钙肥被忽视, 施用极少或不施, 一方面降低了土壤可交换钙的含量, 另一方面过量施肥使土壤酸化, 固态的钙得不到活化和释放, 严重影响了钙素的吸收利用.沿海酸性土壤由于钙流失严重, 花生因缺钙造成空秕率为9%-15%.施钙肥可明显增加花生荚果和籽仁产量, 主要是增加了单株结果数和出仁率、提高了果重(周录英等, 2008 ; 高芳等, 2011 ).本课题组前期研究表明酸性土花生增施钙肥后开花量高于对照, 施钙荚果产量明显提高.本研究结果表明钙肥不同用量均显著增加了酸性土花生的荚果产量, 两个试验点平均增产24.29%.增施钙肥显著提高了单株结果数和单株果重, 同时对提高双仁果率和出仁率效果显著.三庄试验点增施钙肥增产效果要好于文登试验点增施钙肥处理, 可能是试验点地力水平不同造成的, 也可能是选择的钙浓度梯度不是最合适的, 有待于进一步研究.有研究表明: 施钙肥花生的地上部营养生长比不施钙肥的差, 合理施用钙肥可以抑制花生主茎和侧枝旺长(吴文新等, 2001 ; 周录英等, 2008 ).本研究T1 处理的出仁率要高于T2 处理, 而荚果产量显著高于后者, 说明本试验条件下每667 m2 施14 kg CaO效果要好于每667 m2 施28 kg, 原因可能是高浓度钙抑制了花生的营养生长, 从而影响了产量. ...
... 施钙不仅提高了花生籽仁中脂肪和蛋白质含量, 降低了籽仁中可溶性糖含量, 而且可提高脂肪的O/L值, 增加蛋白质组分中含量不足的赖氨酸和蛋氨酸含量, 从而延长花生制品货架寿命, 改善花生蛋白质品质(周录英等, 2008 ; 高芳等, 2011 ).吴文新等(2001)的研究表明, 施钙肥比对照花生种仁粗蛋白含量提高13.78%, 粗脂肪含量提高3.08%, 而粗淀粉降低14.17%.酸性土花生增施钙肥对品质的影响与上述研究结果一致, 增施钙肥均显著提高了两个试验点花生籽仁蛋白质和脂肪含量, 对赖氨酸和总氨基酸的提高幅度也较大, 说明钙肥对酸性土花生籽仁品质改善效果良好.T1 处理显著提高了文登试验点花生的油酸相对含量, 降低了亚油酸相对含量, 从而使O/L值显著提高, T2 处理的O/L值与T0 差异较小; 而T1 和T2 处理均显著提高了三庄试验点花生的油酸含量, 降低了亚油酸含量, 两者差异不显著. ...
钙对镉胁迫下花生生理特性、产量和品质的影响水.
2
2011
... 近20年来, 过于重视增施增产效果明显的氮、磷、钾肥, 作为花生所需的大量元素的钙肥被忽视, 施用极少或不施, 一方面降低了土壤可交换钙的含量, 另一方面过量施肥使土壤酸化, 固态的钙得不到活化和释放, 严重影响了钙素的吸收利用.沿海酸性土壤由于钙流失严重, 花生因缺钙造成空秕率为9%-15%.施钙肥可明显增加花生荚果和籽仁产量, 主要是增加了单株结果数和出仁率、提高了果重(周录英等, 2008 ; 高芳等, 2011 ).本课题组前期研究表明酸性土花生增施钙肥后开花量高于对照, 施钙荚果产量明显提高.本研究结果表明钙肥不同用量均显著增加了酸性土花生的荚果产量, 两个试验点平均增产24.29%.增施钙肥显著提高了单株结果数和单株果重, 同时对提高双仁果率和出仁率效果显著.三庄试验点增施钙肥增产效果要好于文登试验点增施钙肥处理, 可能是试验点地力水平不同造成的, 也可能是选择的钙浓度梯度不是最合适的, 有待于进一步研究.有研究表明: 施钙肥花生的地上部营养生长比不施钙肥的差, 合理施用钙肥可以抑制花生主茎和侧枝旺长(吴文新等, 2001 ; 周录英等, 2008 ).本研究T1 处理的出仁率要高于T2 处理, 而荚果产量显著高于后者, 说明本试验条件下每667 m2 施14 kg CaO效果要好于每667 m2 施28 kg, 原因可能是高浓度钙抑制了花生的营养生长, 从而影响了产量. ...
... 施钙不仅提高了花生籽仁中脂肪和蛋白质含量, 降低了籽仁中可溶性糖含量, 而且可提高脂肪的O/L值, 增加蛋白质组分中含量不足的赖氨酸和蛋氨酸含量, 从而延长花生制品货架寿命, 改善花生蛋白质品质(周录英等, 2008 ; 高芳等, 2011 ).吴文新等(2001)的研究表明, 施钙肥比对照花生种仁粗蛋白含量提高13.78%, 粗脂肪含量提高3.08%, 而粗淀粉降低14.17%.酸性土花生增施钙肥对品质的影响与上述研究结果一致, 增施钙肥均显著提高了两个试验点花生籽仁蛋白质和脂肪含量, 对赖氨酸和总氨基酸的提高幅度也较大, 说明钙肥对酸性土花生籽仁品质改善效果良好.T1 处理显著提高了文登试验点花生的油酸相对含量, 降低了亚油酸相对含量, 从而使O/L值显著提高, T2 处理的O/L值与T0 差异较小; 而T1 和T2 处理均显著提高了三庄试验点花生的油酸含量, 降低了亚油酸含量, 两者差异不显著. ...
Effect of calcium and calmodulin on intrinsic heat tolerance in relation to antioxidant systems in maize seedlings.
1
1997
... Ca是植物生长发育过程中必需的营养元素, 参与植物从种子萌发、生长分化、形态建成到开花结果等全过程.Ca2+ 信使参与调控植物抗逆反应, 如调节酶的活性、蛋白质磷酸化和基因表达等(Monroy & Dhindsa, 1995 ; Gong et al ., 1997 ; Yang & Poovaiah, 2003 ; Hetherington & Brownlee, 2004 ).环境胁迫下植物能通过提高胞内游离Ca2+ 浓度使其与钙调素(CaM)结合, 从而启动一系列生理生化过程, 形成细胞的逆境伤害适应机制, 从而使Ca2+ 和CaM在植物对逆境胁迫的感受、传递、响应和适应过程中起中心作用.国内外对花生Ca营养的研究较多, 主要涉及Ca对花生的生长发育、产量及产量构成因素、籽仁品质的影响, 以及形态解剖特征、生理生化特性和分子生物学机理, 初步明确了Ca对花生荚果发育、抗逆性、产量构成等方面的重要作用(Adams et al ., 1992 ; 周卫和林葆, 1996 , 2001 ; Chamlong et al ., 1999 ; 刘秀梅等, 2005 ; Rahman, 2006 ; 王才斌等, 2008 ; 李岳等, 2012 ).Ca对淀粉的水解与转运、蛋白质的合成等过程具有不可或缺的调控作用, 缺Ca可使蛋白质和RNA合成能力降低.植物产量与品质的形成主要是在一系列碳、氮代谢酶催化下进行的, 有关花生代谢酶活性与产量和品质关系的研究较多(张智猛等, 2008 ; 郭峰等, 2009 ; 崔光军等, 2010 ; 张佳蕾等, 2013 ), 较高的代谢酶活性有利于产量提高和品质改善. ...
Expression and localization of phosphoenolpyruvate carboxylase in developing and germinating wheat grains.
0
1998
麦套花生氮素代谢及相关酶活性变化研究.
1
2009
... Ca是植物生长发育过程中必需的营养元素, 参与植物从种子萌发、生长分化、形态建成到开花结果等全过程.Ca2+ 信使参与调控植物抗逆反应, 如调节酶的活性、蛋白质磷酸化和基因表达等(Monroy & Dhindsa, 1995 ; Gong et al ., 1997 ; Yang & Poovaiah, 2003 ; Hetherington & Brownlee, 2004 ).环境胁迫下植物能通过提高胞内游离Ca2+ 浓度使其与钙调素(CaM)结合, 从而启动一系列生理生化过程, 形成细胞的逆境伤害适应机制, 从而使Ca2+ 和CaM在植物对逆境胁迫的感受、传递、响应和适应过程中起中心作用.国内外对花生Ca营养的研究较多, 主要涉及Ca对花生的生长发育、产量及产量构成因素、籽仁品质的影响, 以及形态解剖特征、生理生化特性和分子生物学机理, 初步明确了Ca对花生荚果发育、抗逆性、产量构成等方面的重要作用(Adams et al ., 1992 ; 周卫和林葆, 1996 , 2001 ; Chamlong et al ., 1999 ; 刘秀梅等, 2005 ; Rahman, 2006 ; 王才斌等, 2008 ; 李岳等, 2012 ).Ca对淀粉的水解与转运、蛋白质的合成等过程具有不可或缺的调控作用, 缺Ca可使蛋白质和RNA合成能力降低.植物产量与品质的形成主要是在一系列碳、氮代谢酶催化下进行的, 有关花生代谢酶活性与产量和品质关系的研究较多(张智猛等, 2008 ; 郭峰等, 2009 ; 崔光军等, 2010 ; 张佳蕾等, 2013 ), 较高的代谢酶活性有利于产量提高和品质改善. ...
麦套花生氮素代谢及相关酶活性变化研究.
1
2009
... Ca是植物生长发育过程中必需的营养元素, 参与植物从种子萌发、生长分化、形态建成到开花结果等全过程.Ca2+ 信使参与调控植物抗逆反应, 如调节酶的活性、蛋白质磷酸化和基因表达等(Monroy & Dhindsa, 1995 ; Gong et al ., 1997 ; Yang & Poovaiah, 2003 ; Hetherington & Brownlee, 2004 ).环境胁迫下植物能通过提高胞内游离Ca2+ 浓度使其与钙调素(CaM)结合, 从而启动一系列生理生化过程, 形成细胞的逆境伤害适应机制, 从而使Ca2+ 和CaM在植物对逆境胁迫的感受、传递、响应和适应过程中起中心作用.国内外对花生Ca营养的研究较多, 主要涉及Ca对花生的生长发育、产量及产量构成因素、籽仁品质的影响, 以及形态解剖特征、生理生化特性和分子生物学机理, 初步明确了Ca对花生荚果发育、抗逆性、产量构成等方面的重要作用(Adams et al ., 1992 ; 周卫和林葆, 1996 , 2001 ; Chamlong et al ., 1999 ; 刘秀梅等, 2005 ; Rahman, 2006 ; 王才斌等, 2008 ; 李岳等, 2012 ).Ca对淀粉的水解与转运、蛋白质的合成等过程具有不可或缺的调控作用, 缺Ca可使蛋白质和RNA合成能力降低.植物产量与品质的形成主要是在一系列碳、氮代谢酶催化下进行的, 有关花生代谢酶活性与产量和品质关系的研究较多(张智猛等, 2008 ; 郭峰等, 2009 ; 崔光军等, 2010 ; 张佳蕾等, 2013 ), 较高的代谢酶活性有利于产量提高和品质改善. ...
钙对花生叶片糖代谢有关酶活性的影响.
1
1997
... 前人研究表明, 低浓度Ca2+ 明显抑制高粱(Sorghum bicolor )、马齿苋(Portulaca oleracea )及大叶伽兰菜(Kalanchoe laciniata )叶片PEPCase活性(焦进安和施教耐, 1987 ), 5 mmol·L-1 Ca2+ 抑制花生叶片磷酸烯醇式丙酮酸羧化酶、磷酸烯醇式丙酮酸磷酸酯酶和丙酮酸激酶活性(郭振飞等, 1997 ).本研究结果有所不同, 增施钙肥提高了酸性土花生花后45天之前的叶片PEPCase活性, T1 处理的活性要高于T2 处理.SS和SPS催化的蔗糖合成途径是植物叶片蔗糖合成的主要途径.增施钙肥显著提高了酸性土花生叶片SS和SPS活性, 花后45天之前SS提高幅度较大, 花后30天至花后60天SPS提高幅度较大, T1 处理的两种酶活性要高于T2 处理.花后60天之后施钙处理的PEPCase、SS和SPS活性显著低于不施钙处理, 原因可能是缺钙花生光合产物转化运输速率低(周恩生等, 2008 ), 进入成熟期后供钙, 结实正常的花生地上部逐渐枯黄, 而缺钙的花生仍还继续开花, 出现“老来青”现象造成的. ...
钙对花生叶片糖代谢有关酶活性的影响.
1
1997
... 前人研究表明, 低浓度Ca2+ 明显抑制高粱(Sorghum bicolor )、马齿苋(Portulaca oleracea )及大叶伽兰菜(Kalanchoe laciniata )叶片PEPCase活性(焦进安和施教耐, 1987 ), 5 mmol·L-1 Ca2+ 抑制花生叶片磷酸烯醇式丙酮酸羧化酶、磷酸烯醇式丙酮酸磷酸酯酶和丙酮酸激酶活性(郭振飞等, 1997 ).本研究结果有所不同, 增施钙肥提高了酸性土花生花后45天之前的叶片PEPCase活性, T1 处理的活性要高于T2 处理.SS和SPS催化的蔗糖合成途径是植物叶片蔗糖合成的主要途径.增施钙肥显著提高了酸性土花生叶片SS和SPS活性, 花后45天之前SS提高幅度较大, 花后30天至花后60天SPS提高幅度较大, T1 处理的两种酶活性要高于T2 处理.花后60天之后施钙处理的PEPCase、SS和SPS活性显著低于不施钙处理, 原因可能是缺钙花生光合产物转化运输速率低(周恩生等, 2008 ), 进入成熟期后供钙, 结实正常的花生地上部逐渐枯黄, 而缺钙的花生仍还继续开花, 出现“老来青”现象造成的. ...
The generation of Ca2 + lcium signals in plants.
1
2004
... Ca是植物生长发育过程中必需的营养元素, 参与植物从种子萌发、生长分化、形态建成到开花结果等全过程.Ca2+ 信使参与调控植物抗逆反应, 如调节酶的活性、蛋白质磷酸化和基因表达等(Monroy & Dhindsa, 1995 ; Gong et al ., 1997 ; Yang & Poovaiah, 2003 ; Hetherington & Brownlee, 2004 ).环境胁迫下植物能通过提高胞内游离Ca2+ 浓度使其与钙调素(CaM)结合, 从而启动一系列生理生化过程, 形成细胞的逆境伤害适应机制, 从而使Ca2+ 和CaM在植物对逆境胁迫的感受、传递、响应和适应过程中起中心作用.国内外对花生Ca营养的研究较多, 主要涉及Ca对花生的生长发育、产量及产量构成因素、籽仁品质的影响, 以及形态解剖特征、生理生化特性和分子生物学机理, 初步明确了Ca对花生荚果发育、抗逆性、产量构成等方面的重要作用(Adams et al ., 1992 ; 周卫和林葆, 1996 , 2001 ; Chamlong et al ., 1999 ; 刘秀梅等, 2005 ; Rahman, 2006 ; 王才斌等, 2008 ; 李岳等, 2012 ).Ca对淀粉的水解与转运、蛋白质的合成等过程具有不可或缺的调控作用, 缺Ca可使蛋白质和RNA合成能力降低.植物产量与品质的形成主要是在一系列碳、氮代谢酶催化下进行的, 有关花生代谢酶活性与产量和品质关系的研究较多(张智猛等, 2008 ; 郭峰等, 2009 ; 崔光军等, 2010 ; 张佳蕾等, 2013 ), 较高的代谢酶活性有利于产量提高和品质改善. ...
Ca2 +对植物磷酸烯醇式丙酮酸羧化酶的抑制作用.
1
1987
... 前人研究表明, 低浓度Ca2+ 明显抑制高粱(Sorghum bicolor )、马齿苋(Portulaca oleracea )及大叶伽兰菜(Kalanchoe laciniata )叶片PEPCase活性(焦进安和施教耐, 1987 ), 5 mmol·L-1 Ca2+ 抑制花生叶片磷酸烯醇式丙酮酸羧化酶、磷酸烯醇式丙酮酸磷酸酯酶和丙酮酸激酶活性(郭振飞等, 1997 ).本研究结果有所不同, 增施钙肥提高了酸性土花生花后45天之前的叶片PEPCase活性, T1 处理的活性要高于T2 处理.SS和SPS催化的蔗糖合成途径是植物叶片蔗糖合成的主要途径.增施钙肥显著提高了酸性土花生叶片SS和SPS活性, 花后45天之前SS提高幅度较大, 花后30天至花后60天SPS提高幅度较大, T1 处理的两种酶活性要高于T2 处理.花后60天之后施钙处理的PEPCase、SS和SPS活性显著低于不施钙处理, 原因可能是缺钙花生光合产物转化运输速率低(周恩生等, 2008 ), 进入成熟期后供钙, 结实正常的花生地上部逐渐枯黄, 而缺钙的花生仍还继续开花, 出现“老来青”现象造成的. ...
Ca2 +对植物磷酸烯醇式丙酮酸羧化酶的抑制作用.
1
1987
... 前人研究表明, 低浓度Ca2+ 明显抑制高粱(Sorghum bicolor )、马齿苋(Portulaca oleracea )及大叶伽兰菜(Kalanchoe laciniata )叶片PEPCase活性(焦进安和施教耐, 1987 ), 5 mmol·L-1 Ca2+ 抑制花生叶片磷酸烯醇式丙酮酸羧化酶、磷酸烯醇式丙酮酸磷酸酯酶和丙酮酸激酶活性(郭振飞等, 1997 ).本研究结果有所不同, 增施钙肥提高了酸性土花生花后45天之前的叶片PEPCase活性, T1 处理的活性要高于T2 处理.SS和SPS催化的蔗糖合成途径是植物叶片蔗糖合成的主要途径.增施钙肥显著提高了酸性土花生叶片SS和SPS活性, 花后45天之前SS提高幅度较大, 花后30天至花后60天SPS提高幅度较大, T1 处理的两种酶活性要高于T2 处理.花后60天之后施钙处理的PEPCase、SS和SPS活性显著低于不施钙处理, 原因可能是缺钙花生光合产物转化运输速率低(周恩生等, 2008 ), 进入成熟期后供钙, 结实正常的花生地上部逐渐枯黄, 而缺钙的花生仍还继续开花, 出现“老来青”现象造成的. ...
施钙对花生衰老特性和产量的影响.
1
2012
... Ca是植物生长发育过程中必需的营养元素, 参与植物从种子萌发、生长分化、形态建成到开花结果等全过程.Ca2+ 信使参与调控植物抗逆反应, 如调节酶的活性、蛋白质磷酸化和基因表达等(Monroy & Dhindsa, 1995 ; Gong et al ., 1997 ; Yang & Poovaiah, 2003 ; Hetherington & Brownlee, 2004 ).环境胁迫下植物能通过提高胞内游离Ca2+ 浓度使其与钙调素(CaM)结合, 从而启动一系列生理生化过程, 形成细胞的逆境伤害适应机制, 从而使Ca2+ 和CaM在植物对逆境胁迫的感受、传递、响应和适应过程中起中心作用.国内外对花生Ca营养的研究较多, 主要涉及Ca对花生的生长发育、产量及产量构成因素、籽仁品质的影响, 以及形态解剖特征、生理生化特性和分子生物学机理, 初步明确了Ca对花生荚果发育、抗逆性、产量构成等方面的重要作用(Adams et al ., 1992 ; 周卫和林葆, 1996 , 2001 ; Chamlong et al ., 1999 ; 刘秀梅等, 2005 ; Rahman, 2006 ; 王才斌等, 2008 ; 李岳等, 2012 ).Ca对淀粉的水解与转运、蛋白质的合成等过程具有不可或缺的调控作用, 缺Ca可使蛋白质和RNA合成能力降低.植物产量与品质的形成主要是在一系列碳、氮代谢酶催化下进行的, 有关花生代谢酶活性与产量和品质关系的研究较多(张智猛等, 2008 ; 郭峰等, 2009 ; 崔光军等, 2010 ; 张佳蕾等, 2013 ), 较高的代谢酶活性有利于产量提高和品质改善. ...
施钙对花生衰老特性和产量的影响.
1
2012
... Ca是植物生长发育过程中必需的营养元素, 参与植物从种子萌发、生长分化、形态建成到开花结果等全过程.Ca2+ 信使参与调控植物抗逆反应, 如调节酶的活性、蛋白质磷酸化和基因表达等(Monroy & Dhindsa, 1995 ; Gong et al ., 1997 ; Yang & Poovaiah, 2003 ; Hetherington & Brownlee, 2004 ).环境胁迫下植物能通过提高胞内游离Ca2+ 浓度使其与钙调素(CaM)结合, 从而启动一系列生理生化过程, 形成细胞的逆境伤害适应机制, 从而使Ca2+ 和CaM在植物对逆境胁迫的感受、传递、响应和适应过程中起中心作用.国内外对花生Ca营养的研究较多, 主要涉及Ca对花生的生长发育、产量及产量构成因素、籽仁品质的影响, 以及形态解剖特征、生理生化特性和分子生物学机理, 初步明确了Ca对花生荚果发育、抗逆性、产量构成等方面的重要作用(Adams et al ., 1992 ; 周卫和林葆, 1996 , 2001 ; Chamlong et al ., 1999 ; 刘秀梅等, 2005 ; Rahman, 2006 ; 王才斌等, 2008 ; 李岳等, 2012 ).Ca对淀粉的水解与转运、蛋白质的合成等过程具有不可或缺的调控作用, 缺Ca可使蛋白质和RNA合成能力降低.植物产量与品质的形成主要是在一系列碳、氮代谢酶催化下进行的, 有关花生代谢酶活性与产量和品质关系的研究较多(张智猛等, 2008 ; 郭峰等, 2009 ; 崔光军等, 2010 ; 张佳蕾等, 2013 ), 较高的代谢酶活性有利于产量提高和品质改善. ...
Disturbed ammonium assimilation is associated with growth inhibition of roots in rice seedlings caused by NaCl.
0
1996
纳米碳酸钙在花生上的施用效果研究.
1
2005
... Ca是植物生长发育过程中必需的营养元素, 参与植物从种子萌发、生长分化、形态建成到开花结果等全过程.Ca2+ 信使参与调控植物抗逆反应, 如调节酶的活性、蛋白质磷酸化和基因表达等(Monroy & Dhindsa, 1995 ; Gong et al ., 1997 ; Yang & Poovaiah, 2003 ; Hetherington & Brownlee, 2004 ).环境胁迫下植物能通过提高胞内游离Ca2+ 浓度使其与钙调素(CaM)结合, 从而启动一系列生理生化过程, 形成细胞的逆境伤害适应机制, 从而使Ca2+ 和CaM在植物对逆境胁迫的感受、传递、响应和适应过程中起中心作用.国内外对花生Ca营养的研究较多, 主要涉及Ca对花生的生长发育、产量及产量构成因素、籽仁品质的影响, 以及形态解剖特征、生理生化特性和分子生物学机理, 初步明确了Ca对花生荚果发育、抗逆性、产量构成等方面的重要作用(Adams et al ., 1992 ; 周卫和林葆, 1996 , 2001 ; Chamlong et al ., 1999 ; 刘秀梅等, 2005 ; Rahman, 2006 ; 王才斌等, 2008 ; 李岳等, 2012 ).Ca对淀粉的水解与转运、蛋白质的合成等过程具有不可或缺的调控作用, 缺Ca可使蛋白质和RNA合成能力降低.植物产量与品质的形成主要是在一系列碳、氮代谢酶催化下进行的, 有关花生代谢酶活性与产量和品质关系的研究较多(张智猛等, 2008 ; 郭峰等, 2009 ; 崔光军等, 2010 ; 张佳蕾等, 2013 ), 较高的代谢酶活性有利于产量提高和品质改善. ...
纳米碳酸钙在花生上的施用效果研究.
1
2005
... Ca是植物生长发育过程中必需的营养元素, 参与植物从种子萌发、生长分化、形态建成到开花结果等全过程.Ca2+ 信使参与调控植物抗逆反应, 如调节酶的活性、蛋白质磷酸化和基因表达等(Monroy & Dhindsa, 1995 ; Gong et al ., 1997 ; Yang & Poovaiah, 2003 ; Hetherington & Brownlee, 2004 ).环境胁迫下植物能通过提高胞内游离Ca2+ 浓度使其与钙调素(CaM)结合, 从而启动一系列生理生化过程, 形成细胞的逆境伤害适应机制, 从而使Ca2+ 和CaM在植物对逆境胁迫的感受、传递、响应和适应过程中起中心作用.国内外对花生Ca营养的研究较多, 主要涉及Ca对花生的生长发育、产量及产量构成因素、籽仁品质的影响, 以及形态解剖特征、生理生化特性和分子生物学机理, 初步明确了Ca对花生荚果发育、抗逆性、产量构成等方面的重要作用(Adams et al ., 1992 ; 周卫和林葆, 1996 , 2001 ; Chamlong et al ., 1999 ; 刘秀梅等, 2005 ; Rahman, 2006 ; 王才斌等, 2008 ; 李岳等, 2012 ).Ca对淀粉的水解与转运、蛋白质的合成等过程具有不可或缺的调控作用, 缺Ca可使蛋白质和RNA合成能力降低.植物产量与品质的形成主要是在一系列碳、氮代谢酶催化下进行的, 有关花生代谢酶活性与产量和品质关系的研究较多(张智猛等, 2008 ; 郭峰等, 2009 ; 崔光军等, 2010 ; 张佳蕾等, 2013 ), 较高的代谢酶活性有利于产量提高和品质改善. ...
钙对小麦氮同化关键酶活性的影响及其与蛋白质磷酸化的关系
0
2002
钙对小麦氮同化关键酶活性的影响及其与蛋白质磷酸化的关系
0
2002
Low-temperature signal transduction: Induction of cold acclimation-specific genes of alfalfa by calcium at 25 ℃.
1
1995
... Ca是植物生长发育过程中必需的营养元素, 参与植物从种子萌发、生长分化、形态建成到开花结果等全过程.Ca2+ 信使参与调控植物抗逆反应, 如调节酶的活性、蛋白质磷酸化和基因表达等(Monroy & Dhindsa, 1995 ; Gong et al ., 1997 ; Yang & Poovaiah, 2003 ; Hetherington & Brownlee, 2004 ).环境胁迫下植物能通过提高胞内游离Ca2+ 浓度使其与钙调素(CaM)结合, 从而启动一系列生理生化过程, 形成细胞的逆境伤害适应机制, 从而使Ca2+ 和CaM在植物对逆境胁迫的感受、传递、响应和适应过程中起中心作用.国内外对花生Ca营养的研究较多, 主要涉及Ca对花生的生长发育、产量及产量构成因素、籽仁品质的影响, 以及形态解剖特征、生理生化特性和分子生物学机理, 初步明确了Ca对花生荚果发育、抗逆性、产量构成等方面的重要作用(Adams et al ., 1992 ; 周卫和林葆, 1996 , 2001 ; Chamlong et al ., 1999 ; 刘秀梅等, 2005 ; Rahman, 2006 ; 王才斌等, 2008 ; 李岳等, 2012 ).Ca对淀粉的水解与转运、蛋白质的合成等过程具有不可或缺的调控作用, 缺Ca可使蛋白质和RNA合成能力降低.植物产量与品质的形成主要是在一系列碳、氮代谢酶催化下进行的, 有关花生代谢酶活性与产量和品质关系的研究较多(张智猛等, 2008 ; 郭峰等, 2009 ; 崔光军等, 2010 ; 张佳蕾等, 2013 ), 较高的代谢酶活性有利于产量提高和品质改善. ...
Effect of calcium and Bradyrhizobium inoculation of the growth, yield and quality of groundnut (A. hypogaea L.).
1
2006
... Ca是植物生长发育过程中必需的营养元素, 参与植物从种子萌发、生长分化、形态建成到开花结果等全过程.Ca2+ 信使参与调控植物抗逆反应, 如调节酶的活性、蛋白质磷酸化和基因表达等(Monroy & Dhindsa, 1995 ; Gong et al ., 1997 ; Yang & Poovaiah, 2003 ; Hetherington & Brownlee, 2004 ).环境胁迫下植物能通过提高胞内游离Ca2+ 浓度使其与钙调素(CaM)结合, 从而启动一系列生理生化过程, 形成细胞的逆境伤害适应机制, 从而使Ca2+ 和CaM在植物对逆境胁迫的感受、传递、响应和适应过程中起中心作用.国内外对花生Ca营养的研究较多, 主要涉及Ca对花生的生长发育、产量及产量构成因素、籽仁品质的影响, 以及形态解剖特征、生理生化特性和分子生物学机理, 初步明确了Ca对花生荚果发育、抗逆性、产量构成等方面的重要作用(Adams et al ., 1992 ; 周卫和林葆, 1996 , 2001 ; Chamlong et al ., 1999 ; 刘秀梅等, 2005 ; Rahman, 2006 ; 王才斌等, 2008 ; 李岳等, 2012 ).Ca对淀粉的水解与转运、蛋白质的合成等过程具有不可或缺的调控作用, 缺Ca可使蛋白质和RNA合成能力降低.植物产量与品质的形成主要是在一系列碳、氮代谢酶催化下进行的, 有关花生代谢酶活性与产量和品质关系的研究较多(张智猛等, 2008 ; 郭峰等, 2009 ; 崔光军等, 2010 ; 张佳蕾等, 2013 ), 较高的代谢酶活性有利于产量提高和品质改善. ...
调环酸钙对花生某些生理特性和产量的影响.
1
2008
... Ca是植物生长发育过程中必需的营养元素, 参与植物从种子萌发、生长分化、形态建成到开花结果等全过程.Ca2+ 信使参与调控植物抗逆反应, 如调节酶的活性、蛋白质磷酸化和基因表达等(Monroy & Dhindsa, 1995 ; Gong et al ., 1997 ; Yang & Poovaiah, 2003 ; Hetherington & Brownlee, 2004 ).环境胁迫下植物能通过提高胞内游离Ca2+ 浓度使其与钙调素(CaM)结合, 从而启动一系列生理生化过程, 形成细胞的逆境伤害适应机制, 从而使Ca2+ 和CaM在植物对逆境胁迫的感受、传递、响应和适应过程中起中心作用.国内外对花生Ca营养的研究较多, 主要涉及Ca对花生的生长发育、产量及产量构成因素、籽仁品质的影响, 以及形态解剖特征、生理生化特性和分子生物学机理, 初步明确了Ca对花生荚果发育、抗逆性、产量构成等方面的重要作用(Adams et al ., 1992 ; 周卫和林葆, 1996 , 2001 ; Chamlong et al ., 1999 ; 刘秀梅等, 2005 ; Rahman, 2006 ; 王才斌等, 2008 ; 李岳等, 2012 ).Ca对淀粉的水解与转运、蛋白质的合成等过程具有不可或缺的调控作用, 缺Ca可使蛋白质和RNA合成能力降低.植物产量与品质的形成主要是在一系列碳、氮代谢酶催化下进行的, 有关花生代谢酶活性与产量和品质关系的研究较多(张智猛等, 2008 ; 郭峰等, 2009 ; 崔光军等, 2010 ; 张佳蕾等, 2013 ), 较高的代谢酶活性有利于产量提高和品质改善. ...
调环酸钙对花生某些生理特性和产量的影响.
1
2008
... Ca是植物生长发育过程中必需的营养元素, 参与植物从种子萌发、生长分化、形态建成到开花结果等全过程.Ca2+ 信使参与调控植物抗逆反应, 如调节酶的活性、蛋白质磷酸化和基因表达等(Monroy & Dhindsa, 1995 ; Gong et al ., 1997 ; Yang & Poovaiah, 2003 ; Hetherington & Brownlee, 2004 ).环境胁迫下植物能通过提高胞内游离Ca2+ 浓度使其与钙调素(CaM)结合, 从而启动一系列生理生化过程, 形成细胞的逆境伤害适应机制, 从而使Ca2+ 和CaM在植物对逆境胁迫的感受、传递、响应和适应过程中起中心作用.国内外对花生Ca营养的研究较多, 主要涉及Ca对花生的生长发育、产量及产量构成因素、籽仁品质的影响, 以及形态解剖特征、生理生化特性和分子生物学机理, 初步明确了Ca对花生荚果发育、抗逆性、产量构成等方面的重要作用(Adams et al ., 1992 ; 周卫和林葆, 1996 , 2001 ; Chamlong et al ., 1999 ; 刘秀梅等, 2005 ; Rahman, 2006 ; 王才斌等, 2008 ; 李岳等, 2012 ).Ca对淀粉的水解与转运、蛋白质的合成等过程具有不可或缺的调控作用, 缺Ca可使蛋白质和RNA合成能力降低.植物产量与品质的形成主要是在一系列碳、氮代谢酶催化下进行的, 有关花生代谢酶活性与产量和品质关系的研究较多(张智猛等, 2008 ; 郭峰等, 2009 ; 崔光军等, 2010 ; 张佳蕾等, 2013 ), 较高的代谢酶活性有利于产量提高和品质改善. ...
钙肥不同用量对花生氮代谢的影响.
1
2014
... 高等植物体内绝大部分NH4 + 是通过谷氨酰胺合成酶/谷氨酸合成酶(GS/GOGAT)循环同化.而谷氨酸脱氢酶(GDH)主要在植物的衰老过程及逆境如高温和水分胁迫等状况下进行NH4 + 同化功能.施用钙肥提高了花生不同生育时期叶片中硝酸还原酶(NR)、GS和GOGAT等活性, 促进植株对氮素的吸收, 增加籽仁中蛋白质含量(王媛媛等, 2014 ).鲁翠涛等(2002)研究小麦(Triticum aestivum )培养液中加入Ca2+ , 在1 mmol·L-1 浓度时, 氮素代谢关键酶NR、GS等活性随着Ca2+ 浓度的增加而增大.本研究中增施钙肥显著提高了酸性土花生叶片的GS、GOGAT和GPT活性, 酶活性达到峰值的时间要早于不施钙处理, 且峰值持续时间较长, 有利于氮素积累.王志强等(2008)通过研究不同浓度的外源Ca2+ 对小麦幼苗氮素代谢的影响, 说明小麦幼苗不同氮同化途径对Ca2+ 的响应不同, GS途径比GDH途径对小麦氮素同化量的增加作用更大.酸性土花生不施钙肥处理的GDH活性要显著高于增施钙肥处理, 说明酸性土花生缺钙时主要是靠GDH途径来进行氮素的代谢和积累.值得一提的是, T2 处理的酸性土花生叶片GPT活性要显著高于T1 处理, 这也是前者籽仁的赖氨酸和总氨基酸含量高于后者的原因. ...
钙肥不同用量对花生氮代谢的影响.
1
2014
... 高等植物体内绝大部分NH4 + 是通过谷氨酰胺合成酶/谷氨酸合成酶(GS/GOGAT)循环同化.而谷氨酸脱氢酶(GDH)主要在植物的衰老过程及逆境如高温和水分胁迫等状况下进行NH4 + 同化功能.施用钙肥提高了花生不同生育时期叶片中硝酸还原酶(NR)、GS和GOGAT等活性, 促进植株对氮素的吸收, 增加籽仁中蛋白质含量(王媛媛等, 2014 ).鲁翠涛等(2002)研究小麦(Triticum aestivum )培养液中加入Ca2+ , 在1 mmol·L-1 浓度时, 氮素代谢关键酶NR、GS等活性随着Ca2+ 浓度的增加而增大.本研究中增施钙肥显著提高了酸性土花生叶片的GS、GOGAT和GPT活性, 酶活性达到峰值的时间要早于不施钙处理, 且峰值持续时间较长, 有利于氮素积累.王志强等(2008)通过研究不同浓度的外源Ca2+ 对小麦幼苗氮素代谢的影响, 说明小麦幼苗不同氮同化途径对Ca2+ 的响应不同, GS途径比GDH途径对小麦氮素同化量的增加作用更大.酸性土花生不施钙肥处理的GDH活性要显著高于增施钙肥处理, 说明酸性土花生缺钙时主要是靠GDH途径来进行氮素的代谢和积累.值得一提的是, T2 处理的酸性土花生叶片GPT活性要显著高于T1 处理, 这也是前者籽仁的赖氨酸和总氨基酸含量高于后者的原因. ...
外源钙离子对小麦幼苗氮素代谢的影响.
0
2008
外源钙离子对小麦幼苗氮素代谢的影响.
0
2008
植物转氨酶(GOT和GPT)活度比色测定方法及其应用.
0
1998
植物转氨酶(GOT和GPT)活度比色测定方法及其应用.
0
1998
钙、硼对花生生长、产量和品质的影响.
1
2001
... 近20年来, 过于重视增施增产效果明显的氮、磷、钾肥, 作为花生所需的大量元素的钙肥被忽视, 施用极少或不施, 一方面降低了土壤可交换钙的含量, 另一方面过量施肥使土壤酸化, 固态的钙得不到活化和释放, 严重影响了钙素的吸收利用.沿海酸性土壤由于钙流失严重, 花生因缺钙造成空秕率为9%-15%.施钙肥可明显增加花生荚果和籽仁产量, 主要是增加了单株结果数和出仁率、提高了果重(周录英等, 2008 ; 高芳等, 2011 ).本课题组前期研究表明酸性土花生增施钙肥后开花量高于对照, 施钙荚果产量明显提高.本研究结果表明钙肥不同用量均显著增加了酸性土花生的荚果产量, 两个试验点平均增产24.29%.增施钙肥显著提高了单株结果数和单株果重, 同时对提高双仁果率和出仁率效果显著.三庄试验点增施钙肥增产效果要好于文登试验点增施钙肥处理, 可能是试验点地力水平不同造成的, 也可能是选择的钙浓度梯度不是最合适的, 有待于进一步研究.有研究表明: 施钙肥花生的地上部营养生长比不施钙肥的差, 合理施用钙肥可以抑制花生主茎和侧枝旺长(吴文新等, 2001 ; 周录英等, 2008 ).本研究T1 处理的出仁率要高于T2 处理, 而荚果产量显著高于后者, 说明本试验条件下每667 m2 施14 kg CaO效果要好于每667 m2 施28 kg, 原因可能是高浓度钙抑制了花生的营养生长, 从而影响了产量. ...
钙、硼对花生生长、产量和品质的影响.
1
2001
... 近20年来, 过于重视增施增产效果明显的氮、磷、钾肥, 作为花生所需的大量元素的钙肥被忽视, 施用极少或不施, 一方面降低了土壤可交换钙的含量, 另一方面过量施肥使土壤酸化, 固态的钙得不到活化和释放, 严重影响了钙素的吸收利用.沿海酸性土壤由于钙流失严重, 花生因缺钙造成空秕率为9%-15%.施钙肥可明显增加花生荚果和籽仁产量, 主要是增加了单株结果数和出仁率、提高了果重(周录英等, 2008 ; 高芳等, 2011 ).本课题组前期研究表明酸性土花生增施钙肥后开花量高于对照, 施钙荚果产量明显提高.本研究结果表明钙肥不同用量均显著增加了酸性土花生的荚果产量, 两个试验点平均增产24.29%.增施钙肥显著提高了单株结果数和单株果重, 同时对提高双仁果率和出仁率效果显著.三庄试验点增施钙肥增产效果要好于文登试验点增施钙肥处理, 可能是试验点地力水平不同造成的, 也可能是选择的钙浓度梯度不是最合适的, 有待于进一步研究.有研究表明: 施钙肥花生的地上部营养生长比不施钙肥的差, 合理施用钙肥可以抑制花生主茎和侧枝旺长(吴文新等, 2001 ; 周录英等, 2008 ).本研究T1 处理的出仁率要高于T2 处理, 而荚果产量显著高于后者, 说明本试验条件下每667 m2 施14 kg CaO效果要好于每667 m2 施28 kg, 原因可能是高浓度钙抑制了花生的营养生长, 从而影响了产量. ...
地膜覆盖花生对钙、镁、硫吸收特性的研究.
1
2007
... 花生(Arachis hypogaea )是我国重要的经济作物, 总产量居油料作物之首, 是我国重要的食用油源、食品工业理想原料和出口创汇作物.花生是需钙量大的作物, 每形成100 kg荚果, 吸钙量达2.0-2.5 kg, 缺钙会导致花生空壳、烂果(张二全等, 1994 ; 吴旭银等, 2007 ).我国东南沿海花生主要种植于旱砂地, 该区域土壤盐基饱和度较低, 酸度较大, 土壤中钙大量流失, 严重影响花生籽粒的发育和产量的形成, 甚至造成绝产. ...
地膜覆盖花生对钙、镁、硫吸收特性的研究.
1
2007
... 花生(Arachis hypogaea )是我国重要的经济作物, 总产量居油料作物之首, 是我国重要的食用油源、食品工业理想原料和出口创汇作物.花生是需钙量大的作物, 每形成100 kg荚果, 吸钙量达2.0-2.5 kg, 缺钙会导致花生空壳、烂果(张二全等, 1994 ; 吴旭银等, 2007 ).我国东南沿海花生主要种植于旱砂地, 该区域土壤盐基饱和度较低, 酸度较大, 土壤中钙大量流失, 严重影响花生籽粒的发育和产量的形成, 甚至造成绝产. ...
Calcium/calmodulin-mediated signal network in plants.
1
2003
... Ca是植物生长发育过程中必需的营养元素, 参与植物从种子萌发、生长分化、形态建成到开花结果等全过程.Ca2+ 信使参与调控植物抗逆反应, 如调节酶的活性、蛋白质磷酸化和基因表达等(Monroy & Dhindsa, 1995 ; Gong et al ., 1997 ; Yang & Poovaiah, 2003 ; Hetherington & Brownlee, 2004 ).环境胁迫下植物能通过提高胞内游离Ca2+ 浓度使其与钙调素(CaM)结合, 从而启动一系列生理生化过程, 形成细胞的逆境伤害适应机制, 从而使Ca2+ 和CaM在植物对逆境胁迫的感受、传递、响应和适应过程中起中心作用.国内外对花生Ca营养的研究较多, 主要涉及Ca对花生的生长发育、产量及产量构成因素、籽仁品质的影响, 以及形态解剖特征、生理生化特性和分子生物学机理, 初步明确了Ca对花生荚果发育、抗逆性、产量构成等方面的重要作用(Adams et al ., 1992 ; 周卫和林葆, 1996 , 2001 ; Chamlong et al ., 1999 ; 刘秀梅等, 2005 ; Rahman, 2006 ; 王才斌等, 2008 ; 李岳等, 2012 ).Ca对淀粉的水解与转运、蛋白质的合成等过程具有不可或缺的调控作用, 缺Ca可使蛋白质和RNA合成能力降低.植物产量与品质的形成主要是在一系列碳、氮代谢酶催化下进行的, 有关花生代谢酶活性与产量和品质关系的研究较多(张智猛等, 2008 ; 郭峰等, 2009 ; 崔光军等, 2010 ; 张佳蕾等, 2013 ), 较高的代谢酶活性有利于产量提高和品质改善. ...
土壤钙素水平对花生施钙效果的影响
1
1994
... 花生(Arachis hypogaea )是我国重要的经济作物, 总产量居油料作物之首, 是我国重要的食用油源、食品工业理想原料和出口创汇作物.花生是需钙量大的作物, 每形成100 kg荚果, 吸钙量达2.0-2.5 kg, 缺钙会导致花生空壳、烂果(张二全等, 1994 ; 吴旭银等, 2007 ).我国东南沿海花生主要种植于旱砂地, 该区域土壤盐基饱和度较低, 酸度较大, 土壤中钙大量流失, 严重影响花生籽粒的发育和产量的形成, 甚至造成绝产. ...
土壤钙素水平对花生施钙效果的影响
1
1994
... 花生(Arachis hypogaea )是我国重要的经济作物, 总产量居油料作物之首, 是我国重要的食用油源、食品工业理想原料和出口创汇作物.花生是需钙量大的作物, 每形成100 kg荚果, 吸钙量达2.0-2.5 kg, 缺钙会导致花生空壳、烂果(张二全等, 1994 ; 吴旭银等, 2007 ).我国东南沿海花生主要种植于旱砂地, 该区域土壤盐基饱和度较低, 酸度较大, 土壤中钙大量流失, 严重影响花生籽粒的发育和产量的形成, 甚至造成绝产. ...
不同品质类型花生品质性状及相关酶活性差异.
1
2013
... Ca是植物生长发育过程中必需的营养元素, 参与植物从种子萌发、生长分化、形态建成到开花结果等全过程.Ca2+ 信使参与调控植物抗逆反应, 如调节酶的活性、蛋白质磷酸化和基因表达等(Monroy & Dhindsa, 1995 ; Gong et al ., 1997 ; Yang & Poovaiah, 2003 ; Hetherington & Brownlee, 2004 ).环境胁迫下植物能通过提高胞内游离Ca2+ 浓度使其与钙调素(CaM)结合, 从而启动一系列生理生化过程, 形成细胞的逆境伤害适应机制, 从而使Ca2+ 和CaM在植物对逆境胁迫的感受、传递、响应和适应过程中起中心作用.国内外对花生Ca营养的研究较多, 主要涉及Ca对花生的生长发育、产量及产量构成因素、籽仁品质的影响, 以及形态解剖特征、生理生化特性和分子生物学机理, 初步明确了Ca对花生荚果发育、抗逆性、产量构成等方面的重要作用(Adams et al ., 1992 ; 周卫和林葆, 1996 , 2001 ; Chamlong et al ., 1999 ; 刘秀梅等, 2005 ; Rahman, 2006 ; 王才斌等, 2008 ; 李岳等, 2012 ).Ca对淀粉的水解与转运、蛋白质的合成等过程具有不可或缺的调控作用, 缺Ca可使蛋白质和RNA合成能力降低.植物产量与品质的形成主要是在一系列碳、氮代谢酶催化下进行的, 有关花生代谢酶活性与产量和品质关系的研究较多(张智猛等, 2008 ; 郭峰等, 2009 ; 崔光军等, 2010 ; 张佳蕾等, 2013 ), 较高的代谢酶活性有利于产量提高和品质改善. ...
不同品质类型花生品质性状及相关酶活性差异.
1
2013
... Ca是植物生长发育过程中必需的营养元素, 参与植物从种子萌发、生长分化、形态建成到开花结果等全过程.Ca2+ 信使参与调控植物抗逆反应, 如调节酶的活性、蛋白质磷酸化和基因表达等(Monroy & Dhindsa, 1995 ; Gong et al ., 1997 ; Yang & Poovaiah, 2003 ; Hetherington & Brownlee, 2004 ).环境胁迫下植物能通过提高胞内游离Ca2+ 浓度使其与钙调素(CaM)结合, 从而启动一系列生理生化过程, 形成细胞的逆境伤害适应机制, 从而使Ca2+ 和CaM在植物对逆境胁迫的感受、传递、响应和适应过程中起中心作用.国内外对花生Ca营养的研究较多, 主要涉及Ca对花生的生长发育、产量及产量构成因素、籽仁品质的影响, 以及形态解剖特征、生理生化特性和分子生物学机理, 初步明确了Ca对花生荚果发育、抗逆性、产量构成等方面的重要作用(Adams et al ., 1992 ; 周卫和林葆, 1996 , 2001 ; Chamlong et al ., 1999 ; 刘秀梅等, 2005 ; Rahman, 2006 ; 王才斌等, 2008 ; 李岳等, 2012 ).Ca对淀粉的水解与转运、蛋白质的合成等过程具有不可或缺的调控作用, 缺Ca可使蛋白质和RNA合成能力降低.植物产量与品质的形成主要是在一系列碳、氮代谢酶催化下进行的, 有关花生代谢酶活性与产量和品质关系的研究较多(张智猛等, 2008 ; 郭峰等, 2009 ; 崔光军等, 2010 ; 张佳蕾等, 2013 ), 较高的代谢酶活性有利于产量提高和品质改善. ...
氮素水平对花生氮素代谢及相关酶活性的影响.
1
2008
... Ca是植物生长发育过程中必需的营养元素, 参与植物从种子萌发、生长分化、形态建成到开花结果等全过程.Ca2+ 信使参与调控植物抗逆反应, 如调节酶的活性、蛋白质磷酸化和基因表达等(Monroy & Dhindsa, 1995 ; Gong et al ., 1997 ; Yang & Poovaiah, 2003 ; Hetherington & Brownlee, 2004 ).环境胁迫下植物能通过提高胞内游离Ca2+ 浓度使其与钙调素(CaM)结合, 从而启动一系列生理生化过程, 形成细胞的逆境伤害适应机制, 从而使Ca2+ 和CaM在植物对逆境胁迫的感受、传递、响应和适应过程中起中心作用.国内外对花生Ca营养的研究较多, 主要涉及Ca对花生的生长发育、产量及产量构成因素、籽仁品质的影响, 以及形态解剖特征、生理生化特性和分子生物学机理, 初步明确了Ca对花生荚果发育、抗逆性、产量构成等方面的重要作用(Adams et al ., 1992 ; 周卫和林葆, 1996 , 2001 ; Chamlong et al ., 1999 ; 刘秀梅等, 2005 ; Rahman, 2006 ; 王才斌等, 2008 ; 李岳等, 2012 ).Ca对淀粉的水解与转运、蛋白质的合成等过程具有不可或缺的调控作用, 缺Ca可使蛋白质和RNA合成能力降低.植物产量与品质的形成主要是在一系列碳、氮代谢酶催化下进行的, 有关花生代谢酶活性与产量和品质关系的研究较多(张智猛等, 2008 ; 郭峰等, 2009 ; 崔光军等, 2010 ; 张佳蕾等, 2013 ), 较高的代谢酶活性有利于产量提高和品质改善. ...
氮素水平对花生氮素代谢及相关酶活性的影响.
1
2008
... Ca是植物生长发育过程中必需的营养元素, 参与植物从种子萌发、生长分化、形态建成到开花结果等全过程.Ca2+ 信使参与调控植物抗逆反应, 如调节酶的活性、蛋白质磷酸化和基因表达等(Monroy & Dhindsa, 1995 ; Gong et al ., 1997 ; Yang & Poovaiah, 2003 ; Hetherington & Brownlee, 2004 ).环境胁迫下植物能通过提高胞内游离Ca2+ 浓度使其与钙调素(CaM)结合, 从而启动一系列生理生化过程, 形成细胞的逆境伤害适应机制, 从而使Ca2+ 和CaM在植物对逆境胁迫的感受、传递、响应和适应过程中起中心作用.国内外对花生Ca营养的研究较多, 主要涉及Ca对花生的生长发育、产量及产量构成因素、籽仁品质的影响, 以及形态解剖特征、生理生化特性和分子生物学机理, 初步明确了Ca对花生荚果发育、抗逆性、产量构成等方面的重要作用(Adams et al ., 1992 ; 周卫和林葆, 1996 , 2001 ; Chamlong et al ., 1999 ; 刘秀梅等, 2005 ; Rahman, 2006 ; 王才斌等, 2008 ; 李岳等, 2012 ).Ca对淀粉的水解与转运、蛋白质的合成等过程具有不可或缺的调控作用, 缺Ca可使蛋白质和RNA合成能力降低.植物产量与品质的形成主要是在一系列碳、氮代谢酶催化下进行的, 有关花生代谢酶活性与产量和品质关系的研究较多(张智猛等, 2008 ; 郭峰等, 2009 ; 崔光军等, 2010 ; 张佳蕾等, 2013 ), 较高的代谢酶活性有利于产量提高和品质改善. ...
钙胁迫下花生荚果微区特征及植株生理生化反应变化.
1
2008
... 前人研究表明, 低浓度Ca2+ 明显抑制高粱(Sorghum bicolor )、马齿苋(Portulaca oleracea )及大叶伽兰菜(Kalanchoe laciniata )叶片PEPCase活性(焦进安和施教耐, 1987 ), 5 mmol·L-1 Ca2+ 抑制花生叶片磷酸烯醇式丙酮酸羧化酶、磷酸烯醇式丙酮酸磷酸酯酶和丙酮酸激酶活性(郭振飞等, 1997 ).本研究结果有所不同, 增施钙肥提高了酸性土花生花后45天之前的叶片PEPCase活性, T1 处理的活性要高于T2 处理.SS和SPS催化的蔗糖合成途径是植物叶片蔗糖合成的主要途径.增施钙肥显著提高了酸性土花生叶片SS和SPS活性, 花后45天之前SS提高幅度较大, 花后30天至花后60天SPS提高幅度较大, T1 处理的两种酶活性要高于T2 处理.花后60天之后施钙处理的PEPCase、SS和SPS活性显著低于不施钙处理, 原因可能是缺钙花生光合产物转化运输速率低(周恩生等, 2008 ), 进入成熟期后供钙, 结实正常的花生地上部逐渐枯黄, 而缺钙的花生仍还继续开花, 出现“老来青”现象造成的. ...
钙胁迫下花生荚果微区特征及植株生理生化反应变化.
1
2008
... 前人研究表明, 低浓度Ca2+ 明显抑制高粱(Sorghum bicolor )、马齿苋(Portulaca oleracea )及大叶伽兰菜(Kalanchoe laciniata )叶片PEPCase活性(焦进安和施教耐, 1987 ), 5 mmol·L-1 Ca2+ 抑制花生叶片磷酸烯醇式丙酮酸羧化酶、磷酸烯醇式丙酮酸磷酸酯酶和丙酮酸激酶活性(郭振飞等, 1997 ).本研究结果有所不同, 增施钙肥提高了酸性土花生花后45天之前的叶片PEPCase活性, T1 处理的活性要高于T2 处理.SS和SPS催化的蔗糖合成途径是植物叶片蔗糖合成的主要途径.增施钙肥显著提高了酸性土花生叶片SS和SPS活性, 花后45天之前SS提高幅度较大, 花后30天至花后60天SPS提高幅度较大, T1 处理的两种酶活性要高于T2 处理.花后60天之后施钙处理的PEPCase、SS和SPS活性显著低于不施钙处理, 原因可能是缺钙花生光合产物转化运输速率低(周恩生等, 2008 ), 进入成熟期后供钙, 结实正常的花生地上部逐渐枯黄, 而缺钙的花生仍还继续开花, 出现“老来青”现象造成的. ...
钙肥不同用量对花生生理特性及产量和品质的影响.
3
2008
... 近20年来, 过于重视增施增产效果明显的氮、磷、钾肥, 作为花生所需的大量元素的钙肥被忽视, 施用极少或不施, 一方面降低了土壤可交换钙的含量, 另一方面过量施肥使土壤酸化, 固态的钙得不到活化和释放, 严重影响了钙素的吸收利用.沿海酸性土壤由于钙流失严重, 花生因缺钙造成空秕率为9%-15%.施钙肥可明显增加花生荚果和籽仁产量, 主要是增加了单株结果数和出仁率、提高了果重(周录英等, 2008 ; 高芳等, 2011 ).本课题组前期研究表明酸性土花生增施钙肥后开花量高于对照, 施钙荚果产量明显提高.本研究结果表明钙肥不同用量均显著增加了酸性土花生的荚果产量, 两个试验点平均增产24.29%.增施钙肥显著提高了单株结果数和单株果重, 同时对提高双仁果率和出仁率效果显著.三庄试验点增施钙肥增产效果要好于文登试验点增施钙肥处理, 可能是试验点地力水平不同造成的, 也可能是选择的钙浓度梯度不是最合适的, 有待于进一步研究.有研究表明: 施钙肥花生的地上部营养生长比不施钙肥的差, 合理施用钙肥可以抑制花生主茎和侧枝旺长(吴文新等, 2001 ; 周录英等, 2008 ).本研究T1 处理的出仁率要高于T2 处理, 而荚果产量显著高于后者, 说明本试验条件下每667 m2 施14 kg CaO效果要好于每667 m2 施28 kg, 原因可能是高浓度钙抑制了花生的营养生长, 从而影响了产量. ...
... ; 周录英等, 2008 ).本研究T1 处理的出仁率要高于T2 处理, 而荚果产量显著高于后者, 说明本试验条件下每667 m2 施14 kg CaO效果要好于每667 m2 施28 kg, 原因可能是高浓度钙抑制了花生的营养生长, 从而影响了产量. ...
... 施钙不仅提高了花生籽仁中脂肪和蛋白质含量, 降低了籽仁中可溶性糖含量, 而且可提高脂肪的O/L值, 增加蛋白质组分中含量不足的赖氨酸和蛋氨酸含量, 从而延长花生制品货架寿命, 改善花生蛋白质品质(周录英等, 2008 ; 高芳等, 2011 ).吴文新等(2001)的研究表明, 施钙肥比对照花生种仁粗蛋白含量提高13.78%, 粗脂肪含量提高3.08%, 而粗淀粉降低14.17%.酸性土花生增施钙肥对品质的影响与上述研究结果一致, 增施钙肥均显著提高了两个试验点花生籽仁蛋白质和脂肪含量, 对赖氨酸和总氨基酸的提高幅度也较大, 说明钙肥对酸性土花生籽仁品质改善效果良好.T1 处理显著提高了文登试验点花生的油酸相对含量, 降低了亚油酸相对含量, 从而使O/L值显著提高, T2 处理的O/L值与T0 差异较小; 而T1 和T2 处理均显著提高了三庄试验点花生的油酸含量, 降低了亚油酸含量, 两者差异不显著. ...
钙肥不同用量对花生生理特性及产量和品质的影响.
3
2008
... 近20年来, 过于重视增施增产效果明显的氮、磷、钾肥, 作为花生所需的大量元素的钙肥被忽视, 施用极少或不施, 一方面降低了土壤可交换钙的含量, 另一方面过量施肥使土壤酸化, 固态的钙得不到活化和释放, 严重影响了钙素的吸收利用.沿海酸性土壤由于钙流失严重, 花生因缺钙造成空秕率为9%-15%.施钙肥可明显增加花生荚果和籽仁产量, 主要是增加了单株结果数和出仁率、提高了果重(周录英等, 2008 ; 高芳等, 2011 ).本课题组前期研究表明酸性土花生增施钙肥后开花量高于对照, 施钙荚果产量明显提高.本研究结果表明钙肥不同用量均显著增加了酸性土花生的荚果产量, 两个试验点平均增产24.29%.增施钙肥显著提高了单株结果数和单株果重, 同时对提高双仁果率和出仁率效果显著.三庄试验点增施钙肥增产效果要好于文登试验点增施钙肥处理, 可能是试验点地力水平不同造成的, 也可能是选择的钙浓度梯度不是最合适的, 有待于进一步研究.有研究表明: 施钙肥花生的地上部营养生长比不施钙肥的差, 合理施用钙肥可以抑制花生主茎和侧枝旺长(吴文新等, 2001 ; 周录英等, 2008 ).本研究T1 处理的出仁率要高于T2 处理, 而荚果产量显著高于后者, 说明本试验条件下每667 m2 施14 kg CaO效果要好于每667 m2 施28 kg, 原因可能是高浓度钙抑制了花生的营养生长, 从而影响了产量. ...
... ; 周录英等, 2008 ).本研究T1 处理的出仁率要高于T2 处理, 而荚果产量显著高于后者, 说明本试验条件下每667 m2 施14 kg CaO效果要好于每667 m2 施28 kg, 原因可能是高浓度钙抑制了花生的营养生长, 从而影响了产量. ...
... 施钙不仅提高了花生籽仁中脂肪和蛋白质含量, 降低了籽仁中可溶性糖含量, 而且可提高脂肪的O/L值, 增加蛋白质组分中含量不足的赖氨酸和蛋氨酸含量, 从而延长花生制品货架寿命, 改善花生蛋白质品质(周录英等, 2008 ; 高芳等, 2011 ).吴文新等(2001)的研究表明, 施钙肥比对照花生种仁粗蛋白含量提高13.78%, 粗脂肪含量提高3.08%, 而粗淀粉降低14.17%.酸性土花生增施钙肥对品质的影响与上述研究结果一致, 增施钙肥均显著提高了两个试验点花生籽仁蛋白质和脂肪含量, 对赖氨酸和总氨基酸的提高幅度也较大, 说明钙肥对酸性土花生籽仁品质改善效果良好.T1 处理显著提高了文登试验点花生的油酸相对含量, 降低了亚油酸相对含量, 从而使O/L值显著提高, T2 处理的O/L值与T0 差异较小; 而T1 和T2 处理均显著提高了三庄试验点花生的油酸含量, 降低了亚油酸含量, 两者差异不显著. ...
花生缺钙症状与超微结构特征的研究.
1
1996
... Ca是植物生长发育过程中必需的营养元素, 参与植物从种子萌发、生长分化、形态建成到开花结果等全过程.Ca2+ 信使参与调控植物抗逆反应, 如调节酶的活性、蛋白质磷酸化和基因表达等(Monroy & Dhindsa, 1995 ; Gong et al ., 1997 ; Yang & Poovaiah, 2003 ; Hetherington & Brownlee, 2004 ).环境胁迫下植物能通过提高胞内游离Ca2+ 浓度使其与钙调素(CaM)结合, 从而启动一系列生理生化过程, 形成细胞的逆境伤害适应机制, 从而使Ca2+ 和CaM在植物对逆境胁迫的感受、传递、响应和适应过程中起中心作用.国内外对花生Ca营养的研究较多, 主要涉及Ca对花生的生长发育、产量及产量构成因素、籽仁品质的影响, 以及形态解剖特征、生理生化特性和分子生物学机理, 初步明确了Ca对花生荚果发育、抗逆性、产量构成等方面的重要作用(Adams et al ., 1992 ; 周卫和林葆, 1996 , 2001 ; Chamlong et al ., 1999 ; 刘秀梅等, 2005 ; Rahman, 2006 ; 王才斌等, 2008 ; 李岳等, 2012 ).Ca对淀粉的水解与转运、蛋白质的合成等过程具有不可或缺的调控作用, 缺Ca可使蛋白质和RNA合成能力降低.植物产量与品质的形成主要是在一系列碳、氮代谢酶催化下进行的, 有关花生代谢酶活性与产量和品质关系的研究较多(张智猛等, 2008 ; 郭峰等, 2009 ; 崔光军等, 2010 ; 张佳蕾等, 2013 ), 较高的代谢酶活性有利于产量提高和品质改善. ...
花生缺钙症状与超微结构特征的研究.
1
1996
... Ca是植物生长发育过程中必需的营养元素, 参与植物从种子萌发、生长分化、形态建成到开花结果等全过程.Ca2+ 信使参与调控植物抗逆反应, 如调节酶的活性、蛋白质磷酸化和基因表达等(Monroy & Dhindsa, 1995 ; Gong et al ., 1997 ; Yang & Poovaiah, 2003 ; Hetherington & Brownlee, 2004 ).环境胁迫下植物能通过提高胞内游离Ca2+ 浓度使其与钙调素(CaM)结合, 从而启动一系列生理生化过程, 形成细胞的逆境伤害适应机制, 从而使Ca2+ 和CaM在植物对逆境胁迫的感受、传递、响应和适应过程中起中心作用.国内外对花生Ca营养的研究较多, 主要涉及Ca对花生的生长发育、产量及产量构成因素、籽仁品质的影响, 以及形态解剖特征、生理生化特性和分子生物学机理, 初步明确了Ca对花生荚果发育、抗逆性、产量构成等方面的重要作用(Adams et al ., 1992 ; 周卫和林葆, 1996 , 2001 ; Chamlong et al ., 1999 ; 刘秀梅等, 2005 ; Rahman, 2006 ; 王才斌等, 2008 ; 李岳等, 2012 ).Ca对淀粉的水解与转运、蛋白质的合成等过程具有不可或缺的调控作用, 缺Ca可使蛋白质和RNA合成能力降低.植物产量与品质的形成主要是在一系列碳、氮代谢酶催化下进行的, 有关花生代谢酶活性与产量和品质关系的研究较多(张智猛等, 2008 ; 郭峰等, 2009 ; 崔光军等, 2010 ; 张佳蕾等, 2013 ), 较高的代谢酶活性有利于产量提高和品质改善. ...
受钙影响的花生生殖生长及种子素质研究.
1
2001
... Ca是植物生长发育过程中必需的营养元素, 参与植物从种子萌发、生长分化、形态建成到开花结果等全过程.Ca2+ 信使参与调控植物抗逆反应, 如调节酶的活性、蛋白质磷酸化和基因表达等(Monroy & Dhindsa, 1995 ; Gong et al ., 1997 ; Yang & Poovaiah, 2003 ; Hetherington & Brownlee, 2004 ).环境胁迫下植物能通过提高胞内游离Ca2+ 浓度使其与钙调素(CaM)结合, 从而启动一系列生理生化过程, 形成细胞的逆境伤害适应机制, 从而使Ca2+ 和CaM在植物对逆境胁迫的感受、传递、响应和适应过程中起中心作用.国内外对花生Ca营养的研究较多, 主要涉及Ca对花生的生长发育、产量及产量构成因素、籽仁品质的影响, 以及形态解剖特征、生理生化特性和分子生物学机理, 初步明确了Ca对花生荚果发育、抗逆性、产量构成等方面的重要作用(Adams et al ., 1992 ; 周卫和林葆, 1996 , 2001 ; Chamlong et al ., 1999 ; 刘秀梅等, 2005 ; Rahman, 2006 ; 王才斌等, 2008 ; 李岳等, 2012 ).Ca对淀粉的水解与转运、蛋白质的合成等过程具有不可或缺的调控作用, 缺Ca可使蛋白质和RNA合成能力降低.植物产量与品质的形成主要是在一系列碳、氮代谢酶催化下进行的, 有关花生代谢酶活性与产量和品质关系的研究较多(张智猛等, 2008 ; 郭峰等, 2009 ; 崔光军等, 2010 ; 张佳蕾等, 2013 ), 较高的代谢酶活性有利于产量提高和品质改善. ...
受钙影响的花生生殖生长及种子素质研究.
1
2001
... Ca是植物生长发育过程中必需的营养元素, 参与植物从种子萌发、生长分化、形态建成到开花结果等全过程.Ca2+ 信使参与调控植物抗逆反应, 如调节酶的活性、蛋白质磷酸化和基因表达等(Monroy & Dhindsa, 1995 ; Gong et al ., 1997 ; Yang & Poovaiah, 2003 ; Hetherington & Brownlee, 2004 ).环境胁迫下植物能通过提高胞内游离Ca2+ 浓度使其与钙调素(CaM)结合, 从而启动一系列生理生化过程, 形成细胞的逆境伤害适应机制, 从而使Ca2+ 和CaM在植物对逆境胁迫的感受、传递、响应和适应过程中起中心作用.国内外对花生Ca营养的研究较多, 主要涉及Ca对花生的生长发育、产量及产量构成因素、籽仁品质的影响, 以及形态解剖特征、生理生化特性和分子生物学机理, 初步明确了Ca对花生荚果发育、抗逆性、产量构成等方面的重要作用(Adams et al ., 1992 ; 周卫和林葆, 1996 , 2001 ; Chamlong et al ., 1999 ; 刘秀梅等, 2005 ; Rahman, 2006 ; 王才斌等, 2008 ; 李岳等, 2012 ).Ca对淀粉的水解与转运、蛋白质的合成等过程具有不可或缺的调控作用, 缺Ca可使蛋白质和RNA合成能力降低.植物产量与品质的形成主要是在一系列碳、氮代谢酶催化下进行的, 有关花生代谢酶活性与产量和品质关系的研究较多(张智猛等, 2008 ; 郭峰等, 2009 ; 崔光军等, 2010 ; 张佳蕾等, 2013 ), 较高的代谢酶活性有利于产量提高和品质改善. ...
 , 万书波
, 万书波

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}