全世界盐碱地面积已经超过总陆地面积的6%和农业灌溉面积的20%, 预计到2050年, 将会有超过50%的耕地盐渍化(张建锋等, 2005 )。我国耕地中盐碱地约占15%, 另外还有2000多万hm2 的盐荒地(李彬等, 2005 )。根据我国土壤特性和所含盐分特点, 将其分为盐土和碱土两大类, 其中最主要的致害离子为Na+ 、Cl- 、HCO3 - 和CO3 2- , 这些离子对植物的胁迫作用, 除了包括直接的胁迫效应外, 还包括离子间复杂的相互作用(石德成等, 2002 )。
盐胁迫主要是渗透胁迫和离子毒害, 它影响各种离子在植物细胞内的分布, 破坏胞内离子平衡, 植物必须进行渗透调节并在细胞内重建离子稳态(Zhang et al ., 2010 ; Zhang & Shi, 2013 )。碱胁迫与盐胁迫相比, 除了渗透胁迫和离子毒害外, 还涉及高pH值伤害(Yang et al ., 2008 )。植物应对碱胁迫, 不仅要进行细胞内pH值调节以保持离子平衡, 同时还必须付出大量的物质和能量来调节根际微环境的pH值(Shi & Wang, 2005 )。所以渗透调节、细胞内离子平衡、根外pH值调节是植物抗碱胁迫的关键所在。
一直以来, 国内外抗盐方面的研究较多考虑单一盐(NaCl)对植物的影响, 主要涉及胁迫生理、表观遗传、离子转运、激素调节、基因表达等方面(孔令安等, 2000 ; Lu et al ., 2013 ; Wu et al ., 2013 )。而土壤碱胁迫这一严重的环境问题仍未得到研究者的充分重视, 仅有少量描述性报道涉及植物抗碱生理及分子机制(萨如拉等, 2014 ; 张一弓等, 2014 )。只有区别研究盐胁迫和碱胁迫, 才能真正揭示植物抗盐碱机制, 为最终培育出新的抗盐碱植物品种打下基础, 为盐碱化地的恢复和治理提供理论支持。
亚麻(Linum usitatissimum )不仅是重要的油料作物, 还是一种工业原料。亚麻油富含不饱和脂肪酸和对人体有益的多种营养物质, 另外, 亚麻的天然纤维具有吸汗、透气性良好和对人体无害等特点(McKenzie & Deyholos, 2011 )。目前, 有关亚麻盐胁迫下的生理响应研究不多, 且多集中在单一盐(NaCl)方面, 其对碱胁迫的响应及适应机制国内外未见相关报道(Ashraf & Fatima, 2004 ; Chemikosova et al ., 2006 )。
考虑到大多数内陆盐碱地的盐分组成特点, 本研究将2种中性盐(NaCl和Na2 SO4 )和2种碱性盐(NaHCO3 和Na2 CO3 )分别按1:1摩尔比混合, 分别模拟不同强度的盐和碱胁迫条件处理亚麻, 研究亚麻在盐、碱胁迫下的生理生态响应机制, 揭示盐胁迫和碱胁迫的异同点, 同时为亚麻抗盐碱品种的筛选和鉴别提供一定的理论依据。
1 材料和方法
1.1 实验材料
亚麻品种‘宁夏-15’。
1.2 模拟胁迫
根据我国内陆盐碱化土壤主要盐分组成以及盐度、碱度复杂多变的特点。选取NaCl、Na2 SO4 、NaHCO3 和Na2 CO3 , 模拟土壤盐碱胁迫环境, 其中NaCl、Na2 SO4 为中性盐, NaHCO3 、Na2 CO3 为碱性盐, 均按摩尔比1:1混合。利用便携式电导仪(DDG- 2080-S, 博取公司, 上海)、露点水势仪(Wescor Psypro, Wescor, Utah, USA)和数字pH计(PHS-3C, 虹益公司, 上海)分别测定各种浓度处理液的电导率、渗透势和pH值。
1.3 材料培养
挑选整齐一致的亚麻种子播种于直径17 cm的塑料花盆内, 每盆装2.5 kg洗净的细砂。共种植40盆, 每盆定苗5株, 出苗后每天用0.5倍Hoagland营养液进行浇灌, 培养2个月。整个实验在室外进行, 并进行人工遮雨, 白天温度为27.0-21.5 ℃, 夜晚为21.5- 18.5 ℃。4周后, 将40盆分为8组, 每组为5盆, 一盆为一个重复。有1组作为胁迫处理前基础生物量指标的测试样品。剩下的7组, 1组为对照组, 每天仍浇0.5倍Hoagland营养液; 其他6组按照设计的不同浓度对亚麻幼苗进行连续14天的胁迫处理。第15天对实验样品进行取样, 将根和地上部分从子叶痕处分离开来。称鲜质量后, 将植株置于80 ℃烘箱内杀青15 min, 杀青后将植株转置40 ℃的真空干燥器内烘干至恒质量, 称取干质量并记录。将样品磨成粉末, 用于测定各种溶质含量。
1.4 生理指标测定
1.4.1 生长指标的测定
相对生长率(RGR) = (ln处理后干质量-ln处理前干质量)/处理天数(Kingsbury et al ., 1984 )
绝对含水量(AWC) = (鲜质量-干质量)/鲜质量, 单位为mg·g-1 。
1.4.2 光合特性指标的测定
在取样前利用荧光仪(IMAGING-PAM, WALZ, Effeltrich, Germany)测定叶绿素荧光参数: 光系统II (PSII)原初光能转换效率(Φ PSII )、非光化学淬灭(NPQ )、光化学淬灭系数(q P ), 各参数数值均是在选定模式下系统自动计算生成。测定前, 叶片暗适应20 min, 每个处理测定5个叶片, 所有操作均在背景光强低于1 μmol·m-2 ·s-1 的环境中进行。仪器的饱和脉冲光的强度为2400 μmol·m-2 ·s-1 , 光化光强设为默认值185 μmol·m-2 ·s-1 , 测量光低于0.5 μmol·m-2 ·s-1 (Genty et al ., 1989 )。
待荧光测量完毕后, 将叶片采下, 洗净磨碎, 称取样品0.1 mg, 用体积比1:1的丙酮和无水乙醇作为萃取液进行萃取。在波长440、645和663 nm下测定吸光度, 每一个萃取实验要进行5次重复。应用Arnon (1949)方程计算叶绿素含量。
1.4.3 无机溶质指标的测定
使用离子色谱系统(DX-300, Thermo Fisher Sci- entific, Massachusetts, USA)、色谱柱(AS4A-SC, Dionex, Sunnyvale, USA)和电导检测器(CDM-II, Dionex, Sunnyvale, USA)测定。使用Na2 CO3 : NaHCO3 = 1.7:1.8 (mmol·L-1 )的混合溶液作为流动相。测量阳离子(K+ 、Na+ 、Ca2+ 和Mg2+ )和阴离子(NO3 - 、Cl- 、SO4 2- 和H2 PO4 - )含量。
1.4.4 有机溶质指标的测定
在70 ℃下利用70%酒精对样品进行萃取, 并参照鲍士旦(1981) 使用蒽酮方法测定可溶性糖含量。利用HCl (0.05 mol·L-1 )在100 ℃下提取2 h, 用高压液相色谱法分析测定有机酸组分及含量, 方法参照荆家海和丁钟荣(1981) 。脯氨酸和甜菜碱的提取和测定分别参考朱广廉等(1983)和Gorham等(1982)的方法。
1.5 数据分析
利用SPSS 17.0和Excel对数据进行统计、分析及绘图, 试验数据以平均值±标准误差表示, 每个数据均是5次重复的平均值, 采用Duncan检测植物各项生理参数的差异性, p < 0.05被视为存在显著性 差异。
2 结果
2.1 盐、碱胁迫对亚麻生长特性的影响
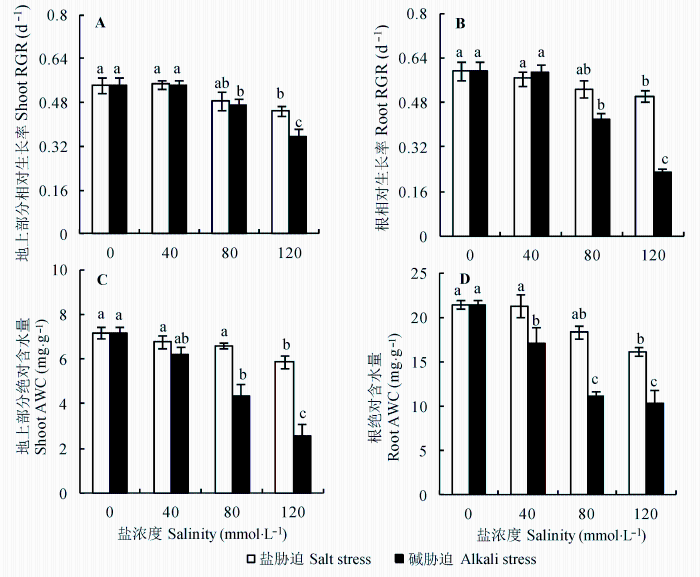
当盐浓度大于40 mmol·L-1 时, 亚麻地上部分和根RGR开始下降, 并随盐浓度增加而加速下降, 碱胁迫的下降幅度均大于相同浓度的盐胁迫; 高碱胁迫(120 mmol·L-1 )下亚麻根RGR降到很低的水平(图1A 、1B, p < 0.05)。盐、碱胁迫下亚麻地上部分、根含水量均随胁迫强度增加而下降, 而碱胁迫下含水量下降幅度明显大于盐胁迫下(图1C 、1D, p < 0.05)。
图1 盐、碱胁迫对亚麻地上部分和根相对生长率(RGR)和绝对含水量(WC)的影响(平均值±标准误差, n = 5)。不同小写字母表示不同处理间差异显著(p < 0.05)。
Fig. 1 Effects of salt and alkali stresses on shoot and root ground relative growth rate (RGR) and on absolute water content (AWC) (mean ± SE, n = 5). Different small letters indicate significant differences between different treatments (p < 0.05).
2.2 盐、碱胁迫对亚麻光合特性的影响
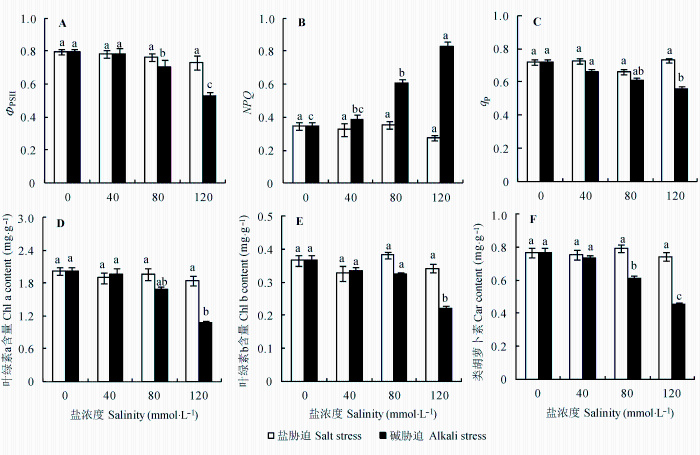
与对照相比, 盐胁迫对亚麻叶片各项荧光参数影响较小, 且差异不显著(图2A -2C, p < 0.05)。但是碱胁迫浓度大于80 mmol·L-1 时, 各项荧光参数变化明显, Φ PSII 和q P 随胁迫强度增加而下降, 与其相对的是NPQ 随胁迫强度增加而上升(图2A -2C, p < 0.05)。
图2 盐、碱胁迫对亚麻叶片光系统II原初光能转换效率(Φ PSII )、非光化学淬灭(NPQ )、光化学淬灭系数(q P )、叶绿素a (Chl a)、叶绿素b (Chl b)和类胡萝卜素(Car)含量的影响(平均值±标准误差, n = 5)。不同小写字母表示不同处理间差异显著(p < 0.05)。
Fig. 2 Effects of salt and alkali stresses on the maximum PSII quantum yield (Φ PSII ), non-photochemical quenching (NPQ ) and photochemical quenching (q P ) of chlorophyll a fluorescence, and chlorophyll a (Chl a), chlorophyll b (Chl b), carotenoids (Car) parameters in leaves of linseed seedlings (mean ± SE, n = 5). Different small letters indicate significant differences between different treatments (p < 0.05).
盐胁迫和中低度碱胁迫均没有影响亚麻叶绿素含量的变化, 但高强度碱胁迫(120 mmol·L-1 )导致3种叶绿素均急剧下降(图2D -2F, p < 0.05)。
2.3 盐、碱胁迫对亚麻离子平衡的影响
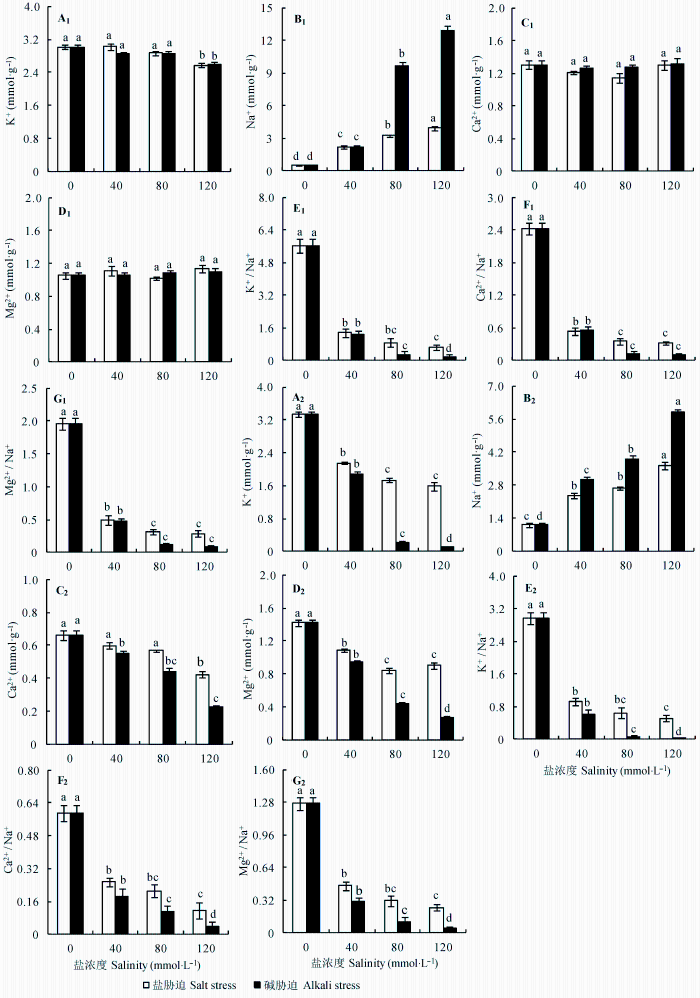
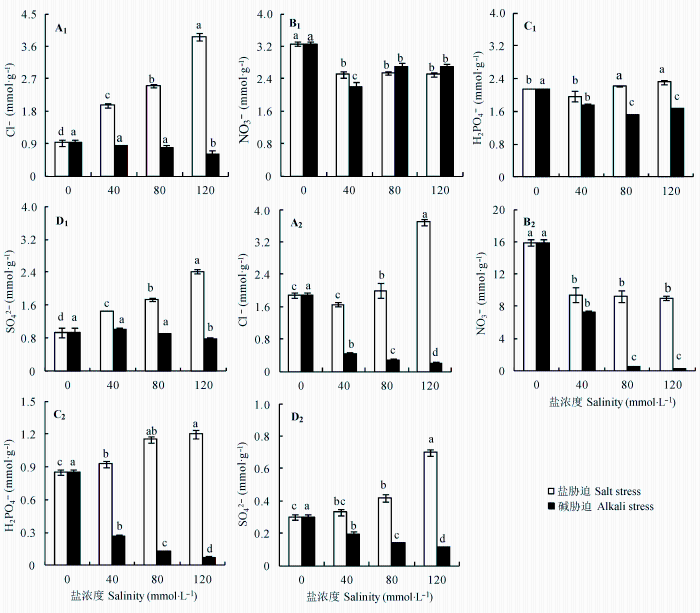
与对照相比, 盐、碱胁迫对地上部分中K+ 影响较小, 只有在盐浓度大于80 mmol·L-1 后K+ 含量才开始下降。但是根中K+ 含量下降明显, 且碱胁迫处理下降程度明显大于盐胁迫(图3A 1 、3A2 , p < 0.05)。地上部分和根中Na+ 含量均随盐度增加呈增加趋势, 且碱胁迫下增加幅度明显高于盐胁迫(图3B 1 、3B2 , p < 0.05)。如图3 所示, Ca2+ 和Mg2+ 变化趋势基本一致。地上部分中Ca2+ 和Mg2+ 含量在盐、碱胁迫下无显著变化, 在根中Ca2+ 和Mg2+ 含量则随胁迫强度增加而呈现下降趋势, 碱胁迫下降幅度明显大于盐胁迫(图3 C1 、3C2 , 3D1 、3D2 , p < 0.05)。与对照相比, 盐、碱胁迫使地上部分、根K+ /Na+ , Ca2+ /Na+ 和Mg2+ /Na+ 大幅度下降, 且随盐度增加缓慢降低, 碱胁迫下的下降幅度均大于盐胁迫(图3 E1 -3G2 , p < 0.05)。随着盐度增加, 盐胁迫下亚麻地上部分和根中的Cl- 含量均增加, 碱胁迫下Cl- 含量均下降, 根中的下降幅度明显大于地上部分(图4A 1 和A2 , p < 0.05)。盐、碱胁迫下地上部分和根中NO3 - 含量均低于对照, 根中NO3 - 含量在碱胁迫下明显低于盐胁迫(图4B 1 和B2 , p < 0.05)。随着胁迫强度增加, 盐胁迫均使地上部分和根中H2 PO4 - 和SO4 2- 含量增加, 但碱胁迫均导致其含量降低, 尤其在根中(图4C 1 -4D2 , p < 0.05)。
图3 盐、碱胁迫对亚麻地上部分和根中K+ , Na+ , Ca2+ , Mg2+ , K+ /Na+ , Ca2+ / Na+ 和Mg2+ /Na+ 含量的影响(平均值±标准误差, n = 5)。不同小写字母表示不同处理间差异显著(p < 0.05)。
Fig. 3 Effects of salt and alkali stresses on the contents of K+ , Na+ , Ca2+ , Mg2+ , K+ /Na+ , Ca2+ /Na+ and Mg2+ /Na+ in the shoots and roots of linseed seedlings (mean ± SE, n = 5). Different small letters indicate significant differences between different treatments (p < 0.05).
2.4 盐、碱胁迫对亚麻有机溶质的影响
如图5A 1 、5A2 所示, 随着胁迫强度的增加, 亚麻地上部分和根中可溶性糖含量在盐胁迫下呈现上升趋势, 而碱胁迫下呈现下降趋势(p < 0.05)。地上部分和根中有机酸在盐胁迫下或稍有减少或无明显变化, 但碱胁迫下有机酸含量不仅明显高于相同强度盐胁迫下, 而且均随盐度增加而明显上升(图5B 1 , 5B2 , p < 0.05)。另外, 盐、碱胁迫均使脯氨酸含量增加, 碱胁迫下增加幅度大于盐胁迫(5C1 , 5C2 , p < 0.05)。盐、碱胁迫下亚麻不但不积累甜菜碱, 反而抑制其合成, 导致含量降低, 根中含量降低幅度大于地上部分中(5D1 , 5D2 , p < 0.05)。
图4 盐、碱胁迫对亚麻地上部分和根干物质中Cl- 、NO3 - 、H2 PO4 - 和SO4 2- 含量的影响(平均值±标准误差, n = 5)。不同小写字母表示不同处理间差异显著(p < 0.05)。
Fig. 4 Effects of salt and alkali stresses on the contents of Cl- , NO3 - , H2 PO4 - and SO4 2- in the shoots and roots of linseed seedlings (mean ± SE, n = 5). Different small letters indicate significant differences between different treatments (p < 0.05).
图5 盐、碱胁迫对亚麻地上部分和根干物质中可溶性糖、有机酸、脯氨酸和甜菜碱含量的影响(平均值±标准误差, n = 5)。不同小写字母表示不同处理间差异显著(p < 0.05)。
Fig. 5 Effects of salt and alkali stresses on the contents of soluble sugars, organic acid, proline and betaine in the shoots and roots of linseed seedlings (mean ± SE, n = 5). Different small letters indicate significant differences between different treatments (p < 0.05).
3 讨论
3.1 盐、碱胁迫对亚麻生长特性的影响
RGR是生命活动的最终体现, 可以作为植物对外界环境胁迫的响应指标(Yang et al ., 2008 )。本实验结果表明在相同盐浓度下, 碱胁迫RGR降幅大于盐胁迫。碱胁迫对植物的伤害甚于盐胁迫的原因可能与两种胁迫的作用机制有关, 盐胁迫主要是渗透胁迫及离子毒害, 而碱胁迫除包括这两种胁迫作用外, 还包括高pH胁迫(石德成等, 2002 )。当胁迫强度大于40 mmol·L-1 时, 碱胁迫(高pH值)明显地抑制亚麻RGR, 尤其是根系生长(图1 )。该结果与抗碱植物明显不同, 例如地肤(Kochia scoparia )和虎尾草(Chl- oris virgata ) (Yang et al ., 2008 )。抗碱植物在高强度碱胁迫下仍然可以保持较强的根RGR, 这表明植物的根系对pH值的调节和适应能力, 可能是决定植物体自身抗碱性的关键生理特性。AWC的变化可以反映植物体受到外界环境胁迫时的基本生理反应。盐、碱胁迫下亚麻AWC的变化趋势和RGR相似。结果充分证明盐、碱胁迫属于不同胁迫类型, 碱胁迫对植物构成的危害明显大于盐胁迫。得到的结论与以往的一些报道相一致(颜宏等, 2000 ; 石德成等, 2002 )。高pH值会引起根系周围某些离子发生沉淀, 导致根系附近离子失衡, 破坏根系对营养元素的吸收; 另外, 高pH值可能会导致泡内质子减少, 进而破坏或抑制根细胞跨膜电化学势梯度, 最终影响根系的正常生理功能。
3.2 盐、碱胁迫对亚麻光合特性的影响
叶绿素荧光特性可以反映出外界胁迫环境对植物光合器官的伤害程度, 其中PSII变化可以清楚显示出外界环境对植物光合能力的影响程度(冯玉龙等, 2001 ; 梁红柱等, 2004 )。本研究发现, 盐胁迫下亚麻叶片各项荧光参数并没有明显的变化, 这表明盐胁迫对植物体自身的光合能力影响并不明显, 之前的一些报道(Everard et al ., 1994 ; Lu et al ., 2002 ) 也证明了这一点。但是, 碱胁迫下PSII活性下降明显, 表明碱胁迫引起了光抑制, 导致亚麻叶片的光合能力降低, 主要原因可能是大量Na+ 和高pH值共同作用的结果。NQP 属于光合系统的自我保护机制, 在碱胁迫下NQP 的增加有利于提高光能利用效率, 增强光电子传递速率。q P 反映了光化学能量转换的能力, 是反映光合速率高低的有效指标, 本实验结果表明碱胁迫严重抑制了亚麻的光合效率(Bolhàr- Nordenkampf et al ., 1989 )。叶绿素和类胡萝卜素是高等植物的主要光合色素。叶绿素和类胡萝卜素的积累在盐胁迫下变化不明显, 但是在碱胁迫下下降明显。这可能是碱胁迫引起Mg沉淀从而抑制了叶绿素和类胡萝卜素的合成(Reddy & Vora, 1986 ; Shi & Zhao, 1997 )。另外, 也有可能是高pH值促进了叶绿素分解酶的活性。
3.3 盐、碱胁迫对亚麻离子平衡的影响
Na+ 和K+ 代谢是植物适应盐渍环境最为重要的生理代谢活动, Na+ 对于大多数植物而言是有害的, 当环境中Na+ 含量增高时, 会拮抗植物对K+ 的吸收, 从而限制植物生长(Brugnoli & Lauteri, 1991 ; 王宁等, 2015 )。本研究发现, 盐、碱胁迫均造成亚麻地上部分和根中Na+ 含量升高, K+ 含量降低, 碱胁迫下的变化趋势明显大于盐胁迫(图3A 1 , 3A2 , 3B1 , 3B2 )。结果表明与盐胁迫相比, 碱胁迫的高pH值进一步干扰和破坏了根对Na+ 、K+ 吸收的选择性, 导致体内Na+ 、K+ 严重失衡, 这是碱胁迫危害大于盐胁迫的一个主要原因。另外, 碱胁迫下亚麻地上部分中积累大量Na+ , 致使叶绿体遭到破坏, 叶绿素含量急剧下降, 导致光合能力降低(图2 )。研究还发现, 与根相比, 地上部分中K+ 含量保持在较高水平, 这说明亚麻有一个将Na+ 控制在根部而将更多的K+ 运输到地上部分的控制适应过程(Munns & Tester, 2008 )。Ca2+ 对维持细胞膜的稳定性起着重要作用, Mg2+ 是叶绿素的主要组分, 研究发现某些植物的Ca2+ 、Mg2+ 积累受到盐胁迫抑制。本研究结果表明盐、碱胁迫对亚麻地上部分中Ca2+ 、Mg2+ 含量影响不大, 但碱胁迫明显降低其在根中的含量(图3C 1 , 3C2 , 3D1 , 3D2 )。Ca2+ 作为第二信使与盐超敏感性蛋白(SOS3)结合, 刺激SOS2的激酶活性, 排出有毒离子Na+ (Zhu, 2003 )。但是碱胁迫(高pH值)导致根周围Ca2+ 沉淀, Ca2+ 吸收明显受阻, 进而影响Na+ 外排, 加速了离子毒害作用。另外, Na+ /H+ 逆向转运蛋白依靠细胞膜两侧质子梯度排出Na+ , 高pH值导致质子亏缺, 质子梯度难于建立, Na+ 不能外排, 导致植物体内积累大量的Na+ (Tester & Davenport, 2003 ; 王宁等, 2015 )。植物体维持细胞内离子平衡是保障各种代谢过程正常进行的必要条件, 离子平衡实际上就是阴阳离子平衡的过程(杨春武等, 2008 )。植物吸收阳离子除了特异性离子通道外, 还有一类非选择性阳离子通道(Munns & Tester, 2008 ), 盐碱胁迫下Na+ 大量进入根部, 可能直接抑制根对其他营养离子的吸收, 导致植物体内离子比率失衡, 如图3 中可见盐、碱胁迫明显降低亚麻K+ /Na+ 、Ca2+ /Na+ 和Mg2+ /Na+ , 并且碱胁迫下的下降幅度均大于盐胁迫。研究还发现, 盐、碱胁迫造成NO3 - 含量降低, 表明大量Na+ 积累阻碍根对NO3 - 的吸收, 明显干扰氮吸收或代谢。碱胁迫强烈干扰无机阴离子积累和分布, 碱胁迫(高pH值)阻碍质子梯度的建立, 从而抑制亚麻根系对阴离子的吸收, 造成植物营养缺乏, 这可能是碱胁迫对植物伤害甚于盐胁迫的另一个重要原因。
3.4 盐、碱胁迫对亚麻有机溶质的影响
植物在遭受盐胁迫时通常会迅速积累一些有机溶质作为细胞质中的渗透调节物质, 除渗透调节作用外, 还可以保护生物大分子并清除自由基(王罗霞等, 2006 )。可溶性糖是植物生命活动最主要的能源物质, 而且它还具有渗透保护、碳贮存和自由基清除的功能(Hare et al ., 1998 )。本研究结果表明盐胁迫促进可溶性糖的合成和积累, 它不仅参与渗透调节, 还中和大量的Na+ 避免离子伤害, 因此可溶性糖的积累可以增强亚麻对盐渍环境的耐性(Gao et al ., 2001 )。但是, 碱胁迫尤其是高碱胁迫抑制了可溶性糖的积累, 这可能是大量的Na+ 和高pH值共同作用的结果, 碱胁迫破坏了光系统结构, 造成了光合能力降低, 影响碳同化过程(Guo et al ., 2011 )。脯氨酸和甜菜碱作为非毒性的渗透调节物质在植物抗逆生理中起非常重要的作用(彭志红等, 2002 ; 张立新和李生秀, 2004 )。盐、碱胁迫都明显刺激了脯氨酸的积累, 脯氨酸的积累可能是长期离子毒性的代谢反应, 其含量的增加可能出于维持渗透压和平衡细胞液泡内无机离子浓度的需要。甜菜碱在盐碱胁迫下呈现下降的趋势, 这可能是由于Na+ 的大量沉淀, 造成根对NO3 - 吸收降低, 阻碍了N代谢的正常进行, 影响了甜菜碱的合成。从有机酸实验结果可知, 亚麻可能通过促进有机酸的合成来弥补无机阴离子的缺失和维护细胞内pH值的稳定(汪建飞和沈其荣, 2006 )。根中合成的有机酸可能是用来调节根外的pH值, 增加根系对铁等矿质元素的吸收和利用, 进而缓解碱胁迫造成的营养缺失和离子失衡。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Arnon DI 1949 ). Copper enzymes in isolated chloroplasts phenoloxidases in Beta vulgaris.
Plant Physiology , 24 , 1 -15 .
[2]
Ashraf M Fatima H 2004 ). Intra-specific variation for salt tolerance in linseed (Linum usitatissimum L).
Journal of Agronomy and Crop Science , 173 , 193 -203 .
[本文引用: 1]
[3]
Bao SD 1981). Analytic Methods for Soil and Agriculture Chemistry . China Agriculture Press , Beijing . 150 -160 . (in Chinese)
[鲍士旦 (1981). 土壤农化分析. 中国农业出版社 , 北京 . 150 -160 .]
[4]
Bolhàr-Nordenkampf HR Long SP Baker NR Öquist G Schreiber U Lechner EG 1989 ). Chlorophyll fluorescence as a probe of the photosynthetic competence of leaves in the field: A review of current instrumentation.
Functional Ecology , 3 , 497 -514 .
[本文引用: 1]
[5]
Brugnoli E Lauteri M 1991 ). Effects of salinity on stomatal conductance, photosynthetic capacity, and carbon isotope discrimination of salt-tolerant (Gossypium hirsutum L.) and salt-sensitive (Phaseolus vulgaris L.) C3 non- halophytes.
Plant Physiology , 95 , 628 -635 .
[本文引用: 1]
[6]
Chemikosova SB Pavlencheva NV Gur’yanov OP Gorshkova TA 2006 ). The effect of soil drought on the phloem fiber development in long-fiber flax.
Russian Journal of Plant Physiology , 53 , 656 -662 .
[本文引用: 1]
[7]
Everard R Gucci SC Kann JA Loescher WH 1994 ). Gas exchange and carbon partitioning in the leaves of celery (Apium graveolens L.) at various levels of root zone salinity.
Plant Physiology , 106 , 281 -292 .
[本文引用: 1]
[8]
Feng YL Feng ZL Cao KF 2001 ). The protection against photodamage in Amomum villosum Lour.
Acta Phytophysiologica Sinica , 27 , 483 -488 . (in Chinese with English abstract)
[本文引用: 1]
[冯玉龙 , 冯志立 , 曹坤芳 (2001 ). 砂仁叶片光破坏的防御
. 植物生理学报 , 27 , 483 -488 .]
[本文引用: 1]
[9]
Gao ZH Xue YB Dai JR 2001 ). cDNA-AFLP analysis reveals that maize resistance to Bipolaris maydis is associated with the induction of multiple defense-related genes.
Chinese Science Bulletin , 46 , 1454 -1458 .
[本文引用: 1]
[10]
Genty B Briantais JM Baker NR 1989 ). The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence.
Biochimica et Biophysica Acta , 990 , 87 -92 .
[本文引用: 1]
[11]
Gorham J McDonnell E WynJones RG 1982 ). Determination of betaines as ultraviolet-absorbing esters.
Analytica Chimica Acta , 138 , 277 -283 .
[12]
Guo R Zhou J Hao WP Gong DJ 2011 ). Germination, growth, photosynthesis and ionic balance in Setaria viridis seedlings subjected to saline and alkaline stress.
Canadian Journal of Plant Science , 91 , 1077 -1088 .
[本文引用: 1]
[13]
Hare PD Cress WA Staden JV 1998 ). Dissecting the roles of osmolyte accumulation during stress.
Plant, Cell & Environment , 21 , 535 -553 .
[本文引用: 1]
[14]
Jing JH Ding ZR 1981). Analysis Method of Plant Biochemistry . Science Press , Beijing . 264 -267 . (in Chinese)
[荆家海 , 丁钟荣 (1981). 植物生物化学分析方法. 科学出版社 , 北京 . 264 -267 .]
[15]
Kingsbury RW Epstein E Peary RW 1984 ). Physiological responses to salinity in selected lines of wheat.
Plant Physiology , 74 , 417 -423 .
[本文引用: 1]
[16]
Kong LA Guo HH Dong XX 2000 ). A study on ultrastructure of hybrid rumex under salt stress.
Acta Prataculturae Sinica , 9 (2 ), 53 -57 . (in Chinese with English abstract)
[本文引用: 1]
[孔令安 , 郭洪海 , 董晓霞 (2000 ). 盐胁迫下杂交酸模超微结构的研究
. 草业学报 , 9 (2 ), 53 -57 .]
[本文引用: 1]
[17]
Li B Wang ZC Shun ZG Chen Y Yang F 2005 ). Resources and sustainable resource exploitation of salinized in China.
Agricultural Research in the Arid Areas , 23 (2 ), 152 -158 . (in Chinese with English abstract)
[本文引用: 1]
[李彬 , 王志春 , 孙志高 , 陈渊 , 杨福 (2005 ). 中国盐碱地资源与可持续利用研究
. 干旱地区农业研究 , 23 (2 ), 152 -158 .]
[本文引用: 1]
[18]
Liang HZ Dou DQ Feng YL 2004 ). Diurnal changes in photosynthesis and chlorophyll fluorescence parameters of Amomum villosum Lour. grown under tropical rainforest in rainy, and foggy and cool seasons at Xishuangbanna.
Acta Ecologica Sinica , 24 , 1421 -1429 . (in Chinese with English abstract)
[本文引用: 1]
[梁红柱 , 窦德泉 , 冯玉龙 (2004 ). 热带雨林下砂仁叶片光合作用和叶绿素荧光参数在雾凉季和雨季的日变化
. 生态学报 , 24 , 1421 -1429 .]
[本文引用: 1]
[19]
Lu CM Qiu NW Lu QT Wang BS Kuang TY 2002 ). Does saline stress lead to increased susceptibility of photosystem II to photoinhibition and changes in photosynthetic pigment composition in halophyte Suaeda salsa grown outdoors?
Plant Science , 163 , 1063 -1068 .
[本文引用: 1]
[20]
Lu YH Lam HM Pi EX Zhan QL Tsai S Wang CM Kwan YW Ngai SM 2013 ). Comparative metabolomics in Glycine max and Glycine soja under salt stress to reveal the phenotypes of their offspring.
Journal of Agricultural and Food Chemistry , 61 , 8711 -8721 .
[本文引用: 1]
[21]
McKenzie RR Deyholos MK 2011 ). Effects of plant growth regulator treatments on stem vascular tissue development in linseed (Linum usitatissimum L.).
Industrial Crops and Products , 34 , 1119 -1127 .
[本文引用: 1]
[22]
Munns R Tester M 2008 ). Mechanisms of salinity tolerance.
Annual Review of Plant Biology , 59 , 651 -681 .
[本文引用: 2]
[23]
Peng ZH Peng KQ Hu JJ Xiao LT 2002 ). Research progress on accumulation of proline under osmotic stress in plants.
Chinese Agriculture Science Bulletin , 18 (4 ), 80 -83 . (in Chinese with English abstract)
[本文引用: 1]
[彭志红 , 彭克勤 , 胡家金 , 萧浪涛 (2002 ). 渗透胁迫下植物脯氨酸积累的研究进展
. 中国农学通报 , 18 (4 ), 80 -83 .]
[本文引用: 1]
[24]
Reddy MP Vora AB 1986 ). Changes in pigment composition, Hill reaction activity and saccharides metabolism in bajra (Pennisetum typhoides S&H) leaves under NaCl salinity.
Photosynthetica , 20 , 50 -55 .
[本文引用: 1]
[25]
Sa RL Liu JH Liu W Bai JH Wang ZH 2014 ). Cation- responsive mechanisms of oats to alkali stress.
Acta Agronomica Sinica , 40 , 362 -368 . (in Chinese with English abstract)
[本文引用: 1]
[萨如拉 , 刘景辉 , 刘伟 , 白健慧 , 王占海 (2014 ). 燕麦对碱胁迫的阳离子响应机制
. 作物学报 , 40 , 362 -368 .]
[本文引用: 1]
[26]
Shi DC Zhao KF 1997 ). Effects of NaCl and Na2 CO3 on growth of Puccinellia tenuiflora and on present state of mineral elements in nutrient solution.
Acta Prataculturae Sinica , 6 , 51 -61 .
[本文引用: 1]
[27]
Shi DC Li YM Yang GH Li YD Zhao KF 2002 ). A simulation of salt and alkali stress mixed ecological conditions and analysis of their stress factors in the seedlings of Aneurolepidium chinense.
Acta Ecological Sinica , 22 , 1317 -1326 . (in Chinese with English abstract)
[本文引用: 3]
[石德成 , 李玉明 , 杨国会 , 李毅丹 , 赵可夫 (2002 ). 盐碱混合生态条件的人工模拟及其对羊草胁迫作用因素分析
. 生态学报 , 22 , 1317 -1326 .]
[本文引用: 3]
[28]
Shi DC Wang D 2005 ). Effects of various salt-alkaline mixed stresses on Aneurolepidium chinense (Trin.) Kitag.
Plant and Soil , 271 , 15 -26 .
[本文引用: 1]
[29]
Tester M Davenport RJ 2003 ). Na+ transport and Na+ tolerance in higher plants.
Annals of Botany , 91 , 503 -527 .
[本文引用: 1]
[30]
Wang LX Zhao ZG Wang SM 2006 ). Effect of nitric oxide on metabolism of reactive oxygen species and membrane lipid peroxidation in Triticum aestivum leaves under water stress.
Aata Prataculturae Sinica , 15 (4 ), 104 -108 . (in Chinese with English abstract)
[本文引用: 1]
[王罗霞 , 赵志光 , 王锁民 (2006 ). 一氧化氮对水分胁迫下小麦叶片活性氧代谢及膜脂过氧化的影响
. 草业学报 , 15 (4 ), 104 -108 .]
[本文引用: 1]
[31]
Wang N Yang J Huang Q Su GL Zhou H Xu QH Dong HL Yan GT 2015 ). Physiological salinity tolerance mechanism for transport of K+ and Na+ ions in cotton (Gossypium hirsutum L.) seedlings under salt stress.
Cotton Science , 27 , 208 -215 . (in Chinese with English abstract)
[本文引用: 2]
[王宁 , 杨杰 , 黄群 , 苏桂兰 , 周红 , 许庆华 , 董合林 , 严根土 (2015 ). 盐胁迫下棉花K+和Na+离子转运的耐盐性生理机制
. 棉花学报 , 27 , 208 -215 .]
[本文引用: 2]
[32]
Wang JF Shen QR 2006 ). Roles of organic metabolism in plant adaptation to nutrient deficiency and aluminum toxicity stress.
Chinese Journal of Applied Ecology , 17 , 2210 -2216 . (in Chinese with English abstract)
[本文引用: 1]
[汪建飞 , 沈其荣 (2006 ). 有机酸代谢在植物适应养分和铝毒胁迫中的作用
. 应用生态学报 , 17 , 2210 -2216 .]
[本文引用: 1]
[33]
Wu DZ Shen QF Cai SG Chen ZH Dai F Zhang GP 2013 ). Ionomic responses and correlations between elements and metabolites under salt stress in wild and cultivated barley.
Plant and Cell Physiology , 54 , 1976 -1988 .
[本文引用: 1]
[34]
Yan H Shi DC Yin SJ Zhao W 2000 ). Effects of saline- alkaline stress on the contents of nitrogen and several organisms of Aneurolepidium chinense. Journal of Northeast Normal University (
Natural Science ), 32 (3 ), 47 -52 . (in Chinese with English abstract)
[本文引用: 1]
[颜宏 , 石德成 , 尹尚军 , 赵伟 (2000 ). 盐碱胁迫对羊草体内N及几种有机代谢产物积累的影响
. 东北师范大学学报(自然科学版) , 32 (3 ), 47 -52 .]
[本文引用: 1]
[35]
Yang C Shi D Wang D 2008 ). Comparative effects of salt stress and alkali stress on growth, osmotic adjustment and ionic balance of an alkali resistant halophyte Suaeda glauca Bge.
Plant Growth Regulation , 56 , 179 -190 .
[本文引用: 3]
[36]
Yang CW Li CY Zhang ML Liu J Ju M Shi DC 2008 ). pH and ion balance in wheat-wheatgrass under salt or alkali stress.
Chinese Journal of Applied Ecology , 19 , 1000 -1005 . (in Chinese with English abstract)
[本文引用: 1]
[杨春武 , 李长有 , 张美丽 , 刘杰 , 鞠淼 , 石德成 (2008 ). 盐、碱胁迫下小冰麦体内的pH及离子平衡
. 应用生态学报 , 19 , 1000 -1005 .]
[本文引用: 1]
[37]
Zhang LX Li SX 2004 ). Research progress on relationships betain and drought/salt resistance of plants.
Acta Botanica Boreali-Occidentalia Sinica , 24 , 1765 -1771 . (in Chinese with English abstract)
[本文引用: 1]
[张立新 , 李生秀 (2004 ). 甜菜碱与植物抗旱/盐性研究进展
.西北植物学报 , 24 , 1765 -1771 .]
[本文引用: 1]
[38]
Zhang JF Zhang XD Zhou JX Liu GH Li DX 2005 ). World resources of saline soil and main amelioration measures.
Research of Soil and Water Conservation , 12 (6 ), 28 -30 . (in Chinese with English abstract)
[本文引用: 1]
[张建锋 , 张旭东 , 周金星 , 刘国华 , 李冬雪 (2005 ). 世界盐碱地资源及其改良利用的基本措施
. 水土保持研究 , 12 (6 ), 28 -30 .]
[本文引用: 1]
[39]
Zhang JL Flowers TJ Wang SM 2010 ). Mechanisms of sodium uptake by roots of higher plants.
Plant and Soil , 326 , 45 -60 .
[本文引用: 1]
[40]
Zhang JL Shi H 2013 ). Physiological and molecular mechanisms of plant salt tolerance.
Photosynthesis Research , 115 , 1 -22 .
[本文引用: 1]
[41]
Zhang YG Jianati , Li XS Zhang HH Shawulie , Ma HY Tang S 2014 ). Seed germination and physiological responses of Agropyron cristatum ‘Tawukumu’ under alkali stress.
Acta Agrestia Sinica , 22 , 783 -788 . (in Chinese with English abstract)
[本文引用: 1]
[张一弓 , 贾纳提 , 李学森 , 张荟荟 , 沙吾列 , 马海燕 , 唐森 (2014 ). 碱胁迫下塔乌库姆冰草的萌发及生理响应
. 草地学报 , 22 , 783 -788 .]
[本文引用: 1]
[42]
Zhu GL Deng XW Zuo WN 1983 ). Determination of free pro- line in plants.
Plant Physiology Communications , 1 , 35 -37 . (in Chinese)
[朱广廉 , 邓兴旺 , 左卫能 (1983 ). 植物体内游离脯氨酸的测定
. 植物生理学通讯 , 1 , 35 -37 .]
[43]
Zhu JK 2003 ). Regulation of ion homeostasis under salt stress.
Current Opinion in Plant Biology , 6 , 441 -445 .
[本文引用: 1]
Copper enzymes in isolated chloroplasts phenoloxidases in Beta vulgaris.
1949
Intra-specific variation for salt tolerance in linseed (Linum usitatissimum L).
1
2004
... 亚麻(Linum usitatissimum )不仅是重要的油料作物, 还是一种工业原料.亚麻油富含不饱和脂肪酸和对人体有益的多种营养物质, 另外, 亚麻的天然纤维具有吸汗、透气性良好和对人体无害等特点(McKenzie & Deyholos, 2011 ).目前, 有关亚麻盐胁迫下的生理响应研究不多, 且多集中在单一盐(NaCl)方面, 其对碱胁迫的响应及适应机制国内外未见相关报道(Ashraf & Fatima, 2004 ; Chemikosova et al ., 2006 ). ...
Chlorophyll fluorescence as a probe of the photosynthetic competence of leaves in the field: A review of current instrumentation.
1
1989
... 叶绿素荧光特性可以反映出外界胁迫环境对植物光合器官的伤害程度, 其中PSII变化可以清楚显示出外界环境对植物光合能力的影响程度(冯玉龙等, 2001 ; 梁红柱等, 2004 ).本研究发现, 盐胁迫下亚麻叶片各项荧光参数并没有明显的变化, 这表明盐胁迫对植物体自身的光合能力影响并不明显, 之前的一些报道(Everard et al ., 1994 ; Lu et al ., 2002 ) 也证明了这一点.但是, 碱胁迫下PSII活性下降明显, 表明碱胁迫引起了光抑制, 导致亚麻叶片的光合能力降低, 主要原因可能是大量Na+ 和高pH值共同作用的结果.NQP 属于光合系统的自我保护机制, 在碱胁迫下NQP 的增加有利于提高光能利用效率, 增强光电子传递速率.q P 反映了光化学能量转换的能力, 是反映光合速率高低的有效指标, 本实验结果表明碱胁迫严重抑制了亚麻的光合效率(Bolhàr- Nordenkampf et al ., 1989 ).叶绿素和类胡萝卜素是高等植物的主要光合色素.叶绿素和类胡萝卜素的积累在盐胁迫下变化不明显, 但是在碱胁迫下下降明显.这可能是碱胁迫引起Mg沉淀从而抑制了叶绿素和类胡萝卜素的合成(Reddy & Vora, 1986 ; Shi & Zhao, 1997 ).另外, 也有可能是高pH值促进了叶绿素分解酶的活性. ...
Effects of salinity on stomatal conductance, photosynthetic capacity, and carbon isotope discrimination of salt-tolerant (Gossypium hirsutum L.) and salt-sensitive (Phaseolus vulgaris L.) C3 non- halophytes.
1
1991
... Na+ 和K+ 代谢是植物适应盐渍环境最为重要的生理代谢活动, Na+ 对于大多数植物而言是有害的, 当环境中Na+ 含量增高时, 会拮抗植物对K+ 的吸收, 从而限制植物生长(Brugnoli & Lauteri, 1991 ; 王宁等, 2015 ).本研究发现, 盐、碱胁迫均造成亚麻地上部分和根中Na+ 含量升高, K+ 含量降低, 碱胁迫下的变化趋势明显大于盐胁迫(图3A 1 , 3A2 , 3B1 , 3B2 ).结果表明与盐胁迫相比, 碱胁迫的高pH值进一步干扰和破坏了根对Na+ 、K+ 吸收的选择性, 导致体内Na+ 、K+ 严重失衡, 这是碱胁迫危害大于盐胁迫的一个主要原因.另外, 碱胁迫下亚麻地上部分中积累大量Na+ , 致使叶绿体遭到破坏, 叶绿素含量急剧下降, 导致光合能力降低(图2 ).研究还发现, 与根相比, 地上部分中K+ 含量保持在较高水平, 这说明亚麻有一个将Na+ 控制在根部而将更多的K+ 运输到地上部分的控制适应过程(Munns & Tester, 2008 ).Ca2+ 对维持细胞膜的稳定性起着重要作用, Mg2+ 是叶绿素的主要组分, 研究发现某些植物的Ca2+ 、Mg2+ 积累受到盐胁迫抑制.本研究结果表明盐、碱胁迫对亚麻地上部分中Ca2+ 、Mg2+ 含量影响不大, 但碱胁迫明显降低其在根中的含量(图3C 1 , 3C2 , 3D1 , 3D2 ).Ca2+ 作为第二信使与盐超敏感性蛋白(SOS3)结合, 刺激SOS2的激酶活性, 排出有毒离子Na+ (Zhu, 2003 ).但是碱胁迫(高pH值)导致根周围Ca2+ 沉淀, Ca2+ 吸收明显受阻, 进而影响Na+ 外排, 加速了离子毒害作用.另外, Na+ /H+ 逆向转运蛋白依靠细胞膜两侧质子梯度排出Na+ , 高pH值导致质子亏缺, 质子梯度难于建立, Na+ 不能外排, 导致植物体内积累大量的Na+ (Tester & Davenport, 2003 ; 王宁等, 2015 ).植物体维持细胞内离子平衡是保障各种代谢过程正常进行的必要条件, 离子平衡实际上就是阴阳离子平衡的过程(杨春武等, 2008 ).植物吸收阳离子除了特异性离子通道外, 还有一类非选择性阳离子通道(Munns & Tester, 2008 ), 盐碱胁迫下Na+ 大量进入根部, 可能直接抑制根对其他营养离子的吸收, 导致植物体内离子比率失衡, 如图3 中可见盐、碱胁迫明显降低亚麻K+ /Na+ 、Ca2+ /Na+ 和Mg2+ /Na+ , 并且碱胁迫下的下降幅度均大于盐胁迫.研究还发现, 盐、碱胁迫造成NO3 - 含量降低, 表明大量Na+ 积累阻碍根对NO3 - 的吸收, 明显干扰氮吸收或代谢.碱胁迫强烈干扰无机阴离子积累和分布, 碱胁迫(高pH值)阻碍质子梯度的建立, 从而抑制亚麻根系对阴离子的吸收, 造成植物营养缺乏, 这可能是碱胁迫对植物伤害甚于盐胁迫的另一个重要原因. ...
The effect of soil drought on the phloem fiber development in long-fiber flax.
1
2006
... 亚麻(Linum usitatissimum )不仅是重要的油料作物, 还是一种工业原料.亚麻油富含不饱和脂肪酸和对人体有益的多种营养物质, 另外, 亚麻的天然纤维具有吸汗、透气性良好和对人体无害等特点(McKenzie & Deyholos, 2011 ).目前, 有关亚麻盐胁迫下的生理响应研究不多, 且多集中在单一盐(NaCl)方面, 其对碱胁迫的响应及适应机制国内外未见相关报道(Ashraf & Fatima, 2004 ; Chemikosova et al ., 2006 ). ...
Gas exchange and carbon partitioning in the leaves of celery (Apium graveolens L.) at various levels of root zone salinity.
1
1994
... 叶绿素荧光特性可以反映出外界胁迫环境对植物光合器官的伤害程度, 其中PSII变化可以清楚显示出外界环境对植物光合能力的影响程度(冯玉龙等, 2001 ; 梁红柱等, 2004 ).本研究发现, 盐胁迫下亚麻叶片各项荧光参数并没有明显的变化, 这表明盐胁迫对植物体自身的光合能力影响并不明显, 之前的一些报道(Everard et al ., 1994 ; Lu et al ., 2002 ) 也证明了这一点.但是, 碱胁迫下PSII活性下降明显, 表明碱胁迫引起了光抑制, 导致亚麻叶片的光合能力降低, 主要原因可能是大量Na+ 和高pH值共同作用的结果.NQP 属于光合系统的自我保护机制, 在碱胁迫下NQP 的增加有利于提高光能利用效率, 增强光电子传递速率.q P 反映了光化学能量转换的能力, 是反映光合速率高低的有效指标, 本实验结果表明碱胁迫严重抑制了亚麻的光合效率(Bolhàr- Nordenkampf et al ., 1989 ).叶绿素和类胡萝卜素是高等植物的主要光合色素.叶绿素和类胡萝卜素的积累在盐胁迫下变化不明显, 但是在碱胁迫下下降明显.这可能是碱胁迫引起Mg沉淀从而抑制了叶绿素和类胡萝卜素的合成(Reddy & Vora, 1986 ; Shi & Zhao, 1997 ).另外, 也有可能是高pH值促进了叶绿素分解酶的活性. ...
砂仁叶片光破坏的防御
1
2001
... 叶绿素荧光特性可以反映出外界胁迫环境对植物光合器官的伤害程度, 其中PSII变化可以清楚显示出外界环境对植物光合能力的影响程度(冯玉龙等, 2001 ; 梁红柱等, 2004 ).本研究发现, 盐胁迫下亚麻叶片各项荧光参数并没有明显的变化, 这表明盐胁迫对植物体自身的光合能力影响并不明显, 之前的一些报道(Everard et al ., 1994 ; Lu et al ., 2002 ) 也证明了这一点.但是, 碱胁迫下PSII活性下降明显, 表明碱胁迫引起了光抑制, 导致亚麻叶片的光合能力降低, 主要原因可能是大量Na+ 和高pH值共同作用的结果.NQP 属于光合系统的自我保护机制, 在碱胁迫下NQP 的增加有利于提高光能利用效率, 增强光电子传递速率.q P 反映了光化学能量转换的能力, 是反映光合速率高低的有效指标, 本实验结果表明碱胁迫严重抑制了亚麻的光合效率(Bolhàr- Nordenkampf et al ., 1989 ).叶绿素和类胡萝卜素是高等植物的主要光合色素.叶绿素和类胡萝卜素的积累在盐胁迫下变化不明显, 但是在碱胁迫下下降明显.这可能是碱胁迫引起Mg沉淀从而抑制了叶绿素和类胡萝卜素的合成(Reddy & Vora, 1986 ; Shi & Zhao, 1997 ).另外, 也有可能是高pH值促进了叶绿素分解酶的活性. ...
砂仁叶片光破坏的防御
1
2001
... 叶绿素荧光特性可以反映出外界胁迫环境对植物光合器官的伤害程度, 其中PSII变化可以清楚显示出外界环境对植物光合能力的影响程度(冯玉龙等, 2001 ; 梁红柱等, 2004 ).本研究发现, 盐胁迫下亚麻叶片各项荧光参数并没有明显的变化, 这表明盐胁迫对植物体自身的光合能力影响并不明显, 之前的一些报道(Everard et al ., 1994 ; Lu et al ., 2002 ) 也证明了这一点.但是, 碱胁迫下PSII活性下降明显, 表明碱胁迫引起了光抑制, 导致亚麻叶片的光合能力降低, 主要原因可能是大量Na+ 和高pH值共同作用的结果.NQP 属于光合系统的自我保护机制, 在碱胁迫下NQP 的增加有利于提高光能利用效率, 增强光电子传递速率.q P 反映了光化学能量转换的能力, 是反映光合速率高低的有效指标, 本实验结果表明碱胁迫严重抑制了亚麻的光合效率(Bolhàr- Nordenkampf et al ., 1989 ).叶绿素和类胡萝卜素是高等植物的主要光合色素.叶绿素和类胡萝卜素的积累在盐胁迫下变化不明显, 但是在碱胁迫下下降明显.这可能是碱胁迫引起Mg沉淀从而抑制了叶绿素和类胡萝卜素的合成(Reddy & Vora, 1986 ; Shi & Zhao, 1997 ).另外, 也有可能是高pH值促进了叶绿素分解酶的活性. ...
cDNA-AFLP analysis reveals that maize resistance to Bipolaris maydis is associated with the induction of multiple defense-related genes.
1
2001
... 植物在遭受盐胁迫时通常会迅速积累一些有机溶质作为细胞质中的渗透调节物质, 除渗透调节作用外, 还可以保护生物大分子并清除自由基(王罗霞等, 2006 ).可溶性糖是植物生命活动最主要的能源物质, 而且它还具有渗透保护、碳贮存和自由基清除的功能(Hare et al ., 1998 ).本研究结果表明盐胁迫促进可溶性糖的合成和积累, 它不仅参与渗透调节, 还中和大量的Na+ 避免离子伤害, 因此可溶性糖的积累可以增强亚麻对盐渍环境的耐性(Gao et al ., 2001 ).但是, 碱胁迫尤其是高碱胁迫抑制了可溶性糖的积累, 这可能是大量的Na+ 和高pH值共同作用的结果, 碱胁迫破坏了光系统结构, 造成了光合能力降低, 影响碳同化过程(Guo et al ., 2011 ).脯氨酸和甜菜碱作为非毒性的渗透调节物质在植物抗逆生理中起非常重要的作用(彭志红等, 2002 ; 张立新和李生秀, 2004 ).盐、碱胁迫都明显刺激了脯氨酸的积累, 脯氨酸的积累可能是长期离子毒性的代谢反应, 其含量的增加可能出于维持渗透压和平衡细胞液泡内无机离子浓度的需要.甜菜碱在盐碱胁迫下呈现下降的趋势, 这可能是由于Na+ 的大量沉淀, 造成根对NO3 - 吸收降低, 阻碍了N代谢的正常进行, 影响了甜菜碱的合成.从有机酸实验结果可知, 亚麻可能通过促进有机酸的合成来弥补无机阴离子的缺失和维护细胞内pH值的稳定(汪建飞和沈其荣, 2006 ).根中合成的有机酸可能是用来调节根外的pH值, 增加根系对铁等矿质元素的吸收和利用, 进而缓解碱胁迫造成的营养缺失和离子失衡. ...
The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence.
1
1989
... 在取样前利用荧光仪(IMAGING-PAM, WALZ, Effeltrich, Germany)测定叶绿素荧光参数: 光系统II (PSII)原初光能转换效率(Φ PSII )、非光化学淬灭(NPQ )、光化学淬灭系数(q P ), 各参数数值均是在选定模式下系统自动计算生成.测定前, 叶片暗适应20 min, 每个处理测定5个叶片, 所有操作均在背景光强低于1 μmol·m-2 ·s-1 的环境中进行.仪器的饱和脉冲光的强度为2400 μmol·m-2 ·s-1 , 光化光强设为默认值185 μmol·m-2 ·s-1 , 测量光低于0.5 μmol·m-2 ·s-1 (Genty et al ., 1989 ). ...
Determination of betaines as ultraviolet-absorbing esters.
1982
Germination, growth, photosynthesis and ionic balance in Setaria viridis seedlings subjected to saline and alkaline stress.
1
2011
... 植物在遭受盐胁迫时通常会迅速积累一些有机溶质作为细胞质中的渗透调节物质, 除渗透调节作用外, 还可以保护生物大分子并清除自由基(王罗霞等, 2006 ).可溶性糖是植物生命活动最主要的能源物质, 而且它还具有渗透保护、碳贮存和自由基清除的功能(Hare et al ., 1998 ).本研究结果表明盐胁迫促进可溶性糖的合成和积累, 它不仅参与渗透调节, 还中和大量的Na+ 避免离子伤害, 因此可溶性糖的积累可以增强亚麻对盐渍环境的耐性(Gao et al ., 2001 ).但是, 碱胁迫尤其是高碱胁迫抑制了可溶性糖的积累, 这可能是大量的Na+ 和高pH值共同作用的结果, 碱胁迫破坏了光系统结构, 造成了光合能力降低, 影响碳同化过程(Guo et al ., 2011 ).脯氨酸和甜菜碱作为非毒性的渗透调节物质在植物抗逆生理中起非常重要的作用(彭志红等, 2002 ; 张立新和李生秀, 2004 ).盐、碱胁迫都明显刺激了脯氨酸的积累, 脯氨酸的积累可能是长期离子毒性的代谢反应, 其含量的增加可能出于维持渗透压和平衡细胞液泡内无机离子浓度的需要.甜菜碱在盐碱胁迫下呈现下降的趋势, 这可能是由于Na+ 的大量沉淀, 造成根对NO3 - 吸收降低, 阻碍了N代谢的正常进行, 影响了甜菜碱的合成.从有机酸实验结果可知, 亚麻可能通过促进有机酸的合成来弥补无机阴离子的缺失和维护细胞内pH值的稳定(汪建飞和沈其荣, 2006 ).根中合成的有机酸可能是用来调节根外的pH值, 增加根系对铁等矿质元素的吸收和利用, 进而缓解碱胁迫造成的营养缺失和离子失衡. ...
Dissecting the roles of osmolyte accumulation during stress.
1
1998
... 植物在遭受盐胁迫时通常会迅速积累一些有机溶质作为细胞质中的渗透调节物质, 除渗透调节作用外, 还可以保护生物大分子并清除自由基(王罗霞等, 2006 ).可溶性糖是植物生命活动最主要的能源物质, 而且它还具有渗透保护、碳贮存和自由基清除的功能(Hare et al ., 1998 ).本研究结果表明盐胁迫促进可溶性糖的合成和积累, 它不仅参与渗透调节, 还中和大量的Na+ 避免离子伤害, 因此可溶性糖的积累可以增强亚麻对盐渍环境的耐性(Gao et al ., 2001 ).但是, 碱胁迫尤其是高碱胁迫抑制了可溶性糖的积累, 这可能是大量的Na+ 和高pH值共同作用的结果, 碱胁迫破坏了光系统结构, 造成了光合能力降低, 影响碳同化过程(Guo et al ., 2011 ).脯氨酸和甜菜碱作为非毒性的渗透调节物质在植物抗逆生理中起非常重要的作用(彭志红等, 2002 ; 张立新和李生秀, 2004 ).盐、碱胁迫都明显刺激了脯氨酸的积累, 脯氨酸的积累可能是长期离子毒性的代谢反应, 其含量的增加可能出于维持渗透压和平衡细胞液泡内无机离子浓度的需要.甜菜碱在盐碱胁迫下呈现下降的趋势, 这可能是由于Na+ 的大量沉淀, 造成根对NO3 - 吸收降低, 阻碍了N代谢的正常进行, 影响了甜菜碱的合成.从有机酸实验结果可知, 亚麻可能通过促进有机酸的合成来弥补无机阴离子的缺失和维护细胞内pH值的稳定(汪建飞和沈其荣, 2006 ).根中合成的有机酸可能是用来调节根外的pH值, 增加根系对铁等矿质元素的吸收和利用, 进而缓解碱胁迫造成的营养缺失和离子失衡. ...
Physiological responses to salinity in selected lines of wheat.
1
1984
... 相对生长率(RGR) = (ln处理后干质量-ln处理前干质量)/处理天数(Kingsbury et al ., 1984 ) ...
盐胁迫下杂交酸模超微结构的研究
1
2000
... 一直以来, 国内外抗盐方面的研究较多考虑单一盐(NaCl)对植物的影响, 主要涉及胁迫生理、表观遗传、离子转运、激素调节、基因表达等方面(孔令安等, 2000 ; Lu et al ., 2013 ; Wu et al ., 2013 ).而土壤碱胁迫这一严重的环境问题仍未得到研究者的充分重视, 仅有少量描述性报道涉及植物抗碱生理及分子机制(萨如拉等, 2014 ; 张一弓等, 2014 ).只有区别研究盐胁迫和碱胁迫, 才能真正揭示植物抗盐碱机制, 为最终培育出新的抗盐碱植物品种打下基础, 为盐碱化地的恢复和治理提供理论支持. ...
盐胁迫下杂交酸模超微结构的研究
1
2000
... 一直以来, 国内外抗盐方面的研究较多考虑单一盐(NaCl)对植物的影响, 主要涉及胁迫生理、表观遗传、离子转运、激素调节、基因表达等方面(孔令安等, 2000 ; Lu et al ., 2013 ; Wu et al ., 2013 ).而土壤碱胁迫这一严重的环境问题仍未得到研究者的充分重视, 仅有少量描述性报道涉及植物抗碱生理及分子机制(萨如拉等, 2014 ; 张一弓等, 2014 ).只有区别研究盐胁迫和碱胁迫, 才能真正揭示植物抗盐碱机制, 为最终培育出新的抗盐碱植物品种打下基础, 为盐碱化地的恢复和治理提供理论支持. ...
中国盐碱地资源与可持续利用研究
1
2005
... 全世界盐碱地面积已经超过总陆地面积的6%和农业灌溉面积的20%, 预计到2050年, 将会有超过50%的耕地盐渍化(张建锋等, 2005 ).我国耕地中盐碱地约占15%, 另外还有2000多万hm2 的盐荒地(李彬等, 2005 ).根据我国土壤特性和所含盐分特点, 将其分为盐土和碱土两大类, 其中最主要的致害离子为Na+ 、Cl- 、HCO3 - 和CO3 2- , 这些离子对植物的胁迫作用, 除了包括直接的胁迫效应外, 还包括离子间复杂的相互作用(石德成等, 2002 ). ...
中国盐碱地资源与可持续利用研究
1
2005
... 全世界盐碱地面积已经超过总陆地面积的6%和农业灌溉面积的20%, 预计到2050年, 将会有超过50%的耕地盐渍化(张建锋等, 2005 ).我国耕地中盐碱地约占15%, 另外还有2000多万hm2 的盐荒地(李彬等, 2005 ).根据我国土壤特性和所含盐分特点, 将其分为盐土和碱土两大类, 其中最主要的致害离子为Na+ 、Cl- 、HCO3 - 和CO3 2- , 这些离子对植物的胁迫作用, 除了包括直接的胁迫效应外, 还包括离子间复杂的相互作用(石德成等, 2002 ). ...
热带雨林下砂仁叶片光合作用和叶绿素荧光参数在雾凉季和雨季的日变化
1
2004
... 叶绿素荧光特性可以反映出外界胁迫环境对植物光合器官的伤害程度, 其中PSII变化可以清楚显示出外界环境对植物光合能力的影响程度(冯玉龙等, 2001 ; 梁红柱等, 2004 ).本研究发现, 盐胁迫下亚麻叶片各项荧光参数并没有明显的变化, 这表明盐胁迫对植物体自身的光合能力影响并不明显, 之前的一些报道(Everard et al ., 1994 ; Lu et al ., 2002 ) 也证明了这一点.但是, 碱胁迫下PSII活性下降明显, 表明碱胁迫引起了光抑制, 导致亚麻叶片的光合能力降低, 主要原因可能是大量Na+ 和高pH值共同作用的结果.NQP 属于光合系统的自我保护机制, 在碱胁迫下NQP 的增加有利于提高光能利用效率, 增强光电子传递速率.q P 反映了光化学能量转换的能力, 是反映光合速率高低的有效指标, 本实验结果表明碱胁迫严重抑制了亚麻的光合效率(Bolhàr- Nordenkampf et al ., 1989 ).叶绿素和类胡萝卜素是高等植物的主要光合色素.叶绿素和类胡萝卜素的积累在盐胁迫下变化不明显, 但是在碱胁迫下下降明显.这可能是碱胁迫引起Mg沉淀从而抑制了叶绿素和类胡萝卜素的合成(Reddy & Vora, 1986 ; Shi & Zhao, 1997 ).另外, 也有可能是高pH值促进了叶绿素分解酶的活性. ...
热带雨林下砂仁叶片光合作用和叶绿素荧光参数在雾凉季和雨季的日变化
1
2004
... 叶绿素荧光特性可以反映出外界胁迫环境对植物光合器官的伤害程度, 其中PSII变化可以清楚显示出外界环境对植物光合能力的影响程度(冯玉龙等, 2001 ; 梁红柱等, 2004 ).本研究发现, 盐胁迫下亚麻叶片各项荧光参数并没有明显的变化, 这表明盐胁迫对植物体自身的光合能力影响并不明显, 之前的一些报道(Everard et al ., 1994 ; Lu et al ., 2002 ) 也证明了这一点.但是, 碱胁迫下PSII活性下降明显, 表明碱胁迫引起了光抑制, 导致亚麻叶片的光合能力降低, 主要原因可能是大量Na+ 和高pH值共同作用的结果.NQP 属于光合系统的自我保护机制, 在碱胁迫下NQP 的增加有利于提高光能利用效率, 增强光电子传递速率.q P 反映了光化学能量转换的能力, 是反映光合速率高低的有效指标, 本实验结果表明碱胁迫严重抑制了亚麻的光合效率(Bolhàr- Nordenkampf et al ., 1989 ).叶绿素和类胡萝卜素是高等植物的主要光合色素.叶绿素和类胡萝卜素的积累在盐胁迫下变化不明显, 但是在碱胁迫下下降明显.这可能是碱胁迫引起Mg沉淀从而抑制了叶绿素和类胡萝卜素的合成(Reddy & Vora, 1986 ; Shi & Zhao, 1997 ).另外, 也有可能是高pH值促进了叶绿素分解酶的活性. ...
Does saline stress lead to increased susceptibility of photosystem II to photoinhibition and changes in photosynthetic pigment composition in halophyte Suaeda salsa grown outdoors?
1
2002
... 叶绿素荧光特性可以反映出外界胁迫环境对植物光合器官的伤害程度, 其中PSII变化可以清楚显示出外界环境对植物光合能力的影响程度(冯玉龙等, 2001 ; 梁红柱等, 2004 ).本研究发现, 盐胁迫下亚麻叶片各项荧光参数并没有明显的变化, 这表明盐胁迫对植物体自身的光合能力影响并不明显, 之前的一些报道(Everard et al ., 1994 ; Lu et al ., 2002 ) 也证明了这一点.但是, 碱胁迫下PSII活性下降明显, 表明碱胁迫引起了光抑制, 导致亚麻叶片的光合能力降低, 主要原因可能是大量Na+ 和高pH值共同作用的结果.NQP 属于光合系统的自我保护机制, 在碱胁迫下NQP 的增加有利于提高光能利用效率, 增强光电子传递速率.q P 反映了光化学能量转换的能力, 是反映光合速率高低的有效指标, 本实验结果表明碱胁迫严重抑制了亚麻的光合效率(Bolhàr- Nordenkampf et al ., 1989 ).叶绿素和类胡萝卜素是高等植物的主要光合色素.叶绿素和类胡萝卜素的积累在盐胁迫下变化不明显, 但是在碱胁迫下下降明显.这可能是碱胁迫引起Mg沉淀从而抑制了叶绿素和类胡萝卜素的合成(Reddy & Vora, 1986 ; Shi & Zhao, 1997 ).另外, 也有可能是高pH值促进了叶绿素分解酶的活性. ...
Comparative metabolomics in Glycine max and Glycine soja under salt stress to reveal the phenotypes of their offspring.
1
2013
... 一直以来, 国内外抗盐方面的研究较多考虑单一盐(NaCl)对植物的影响, 主要涉及胁迫生理、表观遗传、离子转运、激素调节、基因表达等方面(孔令安等, 2000 ; Lu et al ., 2013 ; Wu et al ., 2013 ).而土壤碱胁迫这一严重的环境问题仍未得到研究者的充分重视, 仅有少量描述性报道涉及植物抗碱生理及分子机制(萨如拉等, 2014 ; 张一弓等, 2014 ).只有区别研究盐胁迫和碱胁迫, 才能真正揭示植物抗盐碱机制, 为最终培育出新的抗盐碱植物品种打下基础, 为盐碱化地的恢复和治理提供理论支持. ...
Effects of plant growth regulator treatments on stem vascular tissue development in linseed (Linum usitatissimum L.).
1
2011
... 亚麻(Linum usitatissimum )不仅是重要的油料作物, 还是一种工业原料.亚麻油富含不饱和脂肪酸和对人体有益的多种营养物质, 另外, 亚麻的天然纤维具有吸汗、透气性良好和对人体无害等特点(McKenzie & Deyholos, 2011 ).目前, 有关亚麻盐胁迫下的生理响应研究不多, 且多集中在单一盐(NaCl)方面, 其对碱胁迫的响应及适应机制国内外未见相关报道(Ashraf & Fatima, 2004 ; Chemikosova et al ., 2006 ). ...
Mechanisms of salinity tolerance.
2
2008
... Na+ 和K+ 代谢是植物适应盐渍环境最为重要的生理代谢活动, Na+ 对于大多数植物而言是有害的, 当环境中Na+ 含量增高时, 会拮抗植物对K+ 的吸收, 从而限制植物生长(Brugnoli & Lauteri, 1991 ; 王宁等, 2015 ).本研究发现, 盐、碱胁迫均造成亚麻地上部分和根中Na+ 含量升高, K+ 含量降低, 碱胁迫下的变化趋势明显大于盐胁迫(图3A 1 , 3A2 , 3B1 , 3B2 ).结果表明与盐胁迫相比, 碱胁迫的高pH值进一步干扰和破坏了根对Na+ 、K+ 吸收的选择性, 导致体内Na+ 、K+ 严重失衡, 这是碱胁迫危害大于盐胁迫的一个主要原因.另外, 碱胁迫下亚麻地上部分中积累大量Na+ , 致使叶绿体遭到破坏, 叶绿素含量急剧下降, 导致光合能力降低(图2 ).研究还发现, 与根相比, 地上部分中K+ 含量保持在较高水平, 这说明亚麻有一个将Na+ 控制在根部而将更多的K+ 运输到地上部分的控制适应过程(Munns & Tester, 2008 ).Ca2+ 对维持细胞膜的稳定性起着重要作用, Mg2+ 是叶绿素的主要组分, 研究发现某些植物的Ca2+ 、Mg2+ 积累受到盐胁迫抑制.本研究结果表明盐、碱胁迫对亚麻地上部分中Ca2+ 、Mg2+ 含量影响不大, 但碱胁迫明显降低其在根中的含量(图3C 1 , 3C2 , 3D1 , 3D2 ).Ca2+ 作为第二信使与盐超敏感性蛋白(SOS3)结合, 刺激SOS2的激酶活性, 排出有毒离子Na+ (Zhu, 2003 ).但是碱胁迫(高pH值)导致根周围Ca2+ 沉淀, Ca2+ 吸收明显受阻, 进而影响Na+ 外排, 加速了离子毒害作用.另外, Na+ /H+ 逆向转运蛋白依靠细胞膜两侧质子梯度排出Na+ , 高pH值导致质子亏缺, 质子梯度难于建立, Na+ 不能外排, 导致植物体内积累大量的Na+ (Tester & Davenport, 2003 ; 王宁等, 2015 ).植物体维持细胞内离子平衡是保障各种代谢过程正常进行的必要条件, 离子平衡实际上就是阴阳离子平衡的过程(杨春武等, 2008 ).植物吸收阳离子除了特异性离子通道外, 还有一类非选择性阳离子通道(Munns & Tester, 2008 ), 盐碱胁迫下Na+ 大量进入根部, 可能直接抑制根对其他营养离子的吸收, 导致植物体内离子比率失衡, 如图3 中可见盐、碱胁迫明显降低亚麻K+ /Na+ 、Ca2+ /Na+ 和Mg2+ /Na+ , 并且碱胁迫下的下降幅度均大于盐胁迫.研究还发现, 盐、碱胁迫造成NO3 - 含量降低, 表明大量Na+ 积累阻碍根对NO3 - 的吸收, 明显干扰氮吸收或代谢.碱胁迫强烈干扰无机阴离子积累和分布, 碱胁迫(高pH值)阻碍质子梯度的建立, 从而抑制亚麻根系对阴离子的吸收, 造成植物营养缺乏, 这可能是碱胁迫对植物伤害甚于盐胁迫的另一个重要原因. ...
... ).植物吸收阳离子除了特异性离子通道外, 还有一类非选择性阳离子通道(Munns & Tester, 2008 ), 盐碱胁迫下Na+ 大量进入根部, 可能直接抑制根对其他营养离子的吸收, 导致植物体内离子比率失衡, 如图3 中可见盐、碱胁迫明显降低亚麻K+ /Na+ 、Ca2+ /Na+ 和Mg2+ /Na+ , 并且碱胁迫下的下降幅度均大于盐胁迫.研究还发现, 盐、碱胁迫造成NO3 - 含量降低, 表明大量Na+ 积累阻碍根对NO3 - 的吸收, 明显干扰氮吸收或代谢.碱胁迫强烈干扰无机阴离子积累和分布, 碱胁迫(高pH值)阻碍质子梯度的建立, 从而抑制亚麻根系对阴离子的吸收, 造成植物营养缺乏, 这可能是碱胁迫对植物伤害甚于盐胁迫的另一个重要原因. ...
渗透胁迫下植物脯氨酸积累的研究进展
1
2002
... 植物在遭受盐胁迫时通常会迅速积累一些有机溶质作为细胞质中的渗透调节物质, 除渗透调节作用外, 还可以保护生物大分子并清除自由基(王罗霞等, 2006 ).可溶性糖是植物生命活动最主要的能源物质, 而且它还具有渗透保护、碳贮存和自由基清除的功能(Hare et al ., 1998 ).本研究结果表明盐胁迫促进可溶性糖的合成和积累, 它不仅参与渗透调节, 还中和大量的Na+ 避免离子伤害, 因此可溶性糖的积累可以增强亚麻对盐渍环境的耐性(Gao et al ., 2001 ).但是, 碱胁迫尤其是高碱胁迫抑制了可溶性糖的积累, 这可能是大量的Na+ 和高pH值共同作用的结果, 碱胁迫破坏了光系统结构, 造成了光合能力降低, 影响碳同化过程(Guo et al ., 2011 ).脯氨酸和甜菜碱作为非毒性的渗透调节物质在植物抗逆生理中起非常重要的作用(彭志红等, 2002 ; 张立新和李生秀, 2004 ).盐、碱胁迫都明显刺激了脯氨酸的积累, 脯氨酸的积累可能是长期离子毒性的代谢反应, 其含量的增加可能出于维持渗透压和平衡细胞液泡内无机离子浓度的需要.甜菜碱在盐碱胁迫下呈现下降的趋势, 这可能是由于Na+ 的大量沉淀, 造成根对NO3 - 吸收降低, 阻碍了N代谢的正常进行, 影响了甜菜碱的合成.从有机酸实验结果可知, 亚麻可能通过促进有机酸的合成来弥补无机阴离子的缺失和维护细胞内pH值的稳定(汪建飞和沈其荣, 2006 ).根中合成的有机酸可能是用来调节根外的pH值, 增加根系对铁等矿质元素的吸收和利用, 进而缓解碱胁迫造成的营养缺失和离子失衡. ...
渗透胁迫下植物脯氨酸积累的研究进展
1
2002
... 植物在遭受盐胁迫时通常会迅速积累一些有机溶质作为细胞质中的渗透调节物质, 除渗透调节作用外, 还可以保护生物大分子并清除自由基(王罗霞等, 2006 ).可溶性糖是植物生命活动最主要的能源物质, 而且它还具有渗透保护、碳贮存和自由基清除的功能(Hare et al ., 1998 ).本研究结果表明盐胁迫促进可溶性糖的合成和积累, 它不仅参与渗透调节, 还中和大量的Na+ 避免离子伤害, 因此可溶性糖的积累可以增强亚麻对盐渍环境的耐性(Gao et al ., 2001 ).但是, 碱胁迫尤其是高碱胁迫抑制了可溶性糖的积累, 这可能是大量的Na+ 和高pH值共同作用的结果, 碱胁迫破坏了光系统结构, 造成了光合能力降低, 影响碳同化过程(Guo et al ., 2011 ).脯氨酸和甜菜碱作为非毒性的渗透调节物质在植物抗逆生理中起非常重要的作用(彭志红等, 2002 ; 张立新和李生秀, 2004 ).盐、碱胁迫都明显刺激了脯氨酸的积累, 脯氨酸的积累可能是长期离子毒性的代谢反应, 其含量的增加可能出于维持渗透压和平衡细胞液泡内无机离子浓度的需要.甜菜碱在盐碱胁迫下呈现下降的趋势, 这可能是由于Na+ 的大量沉淀, 造成根对NO3 - 吸收降低, 阻碍了N代谢的正常进行, 影响了甜菜碱的合成.从有机酸实验结果可知, 亚麻可能通过促进有机酸的合成来弥补无机阴离子的缺失和维护细胞内pH值的稳定(汪建飞和沈其荣, 2006 ).根中合成的有机酸可能是用来调节根外的pH值, 增加根系对铁等矿质元素的吸收和利用, 进而缓解碱胁迫造成的营养缺失和离子失衡. ...
Changes in pigment composition, Hill reaction activity and saccharides metabolism in bajra (Pennisetum typhoides S&H) leaves under NaCl salinity.
1
1986
... 叶绿素荧光特性可以反映出外界胁迫环境对植物光合器官的伤害程度, 其中PSII变化可以清楚显示出外界环境对植物光合能力的影响程度(冯玉龙等, 2001 ; 梁红柱等, 2004 ).本研究发现, 盐胁迫下亚麻叶片各项荧光参数并没有明显的变化, 这表明盐胁迫对植物体自身的光合能力影响并不明显, 之前的一些报道(Everard et al ., 1994 ; Lu et al ., 2002 ) 也证明了这一点.但是, 碱胁迫下PSII活性下降明显, 表明碱胁迫引起了光抑制, 导致亚麻叶片的光合能力降低, 主要原因可能是大量Na+ 和高pH值共同作用的结果.NQP 属于光合系统的自我保护机制, 在碱胁迫下NQP 的增加有利于提高光能利用效率, 增强光电子传递速率.q P 反映了光化学能量转换的能力, 是反映光合速率高低的有效指标, 本实验结果表明碱胁迫严重抑制了亚麻的光合效率(Bolhàr- Nordenkampf et al ., 1989 ).叶绿素和类胡萝卜素是高等植物的主要光合色素.叶绿素和类胡萝卜素的积累在盐胁迫下变化不明显, 但是在碱胁迫下下降明显.这可能是碱胁迫引起Mg沉淀从而抑制了叶绿素和类胡萝卜素的合成(Reddy & Vora, 1986 ; Shi & Zhao, 1997 ).另外, 也有可能是高pH值促进了叶绿素分解酶的活性. ...
燕麦对碱胁迫的阳离子响应机制
1
2014
... 一直以来, 国内外抗盐方面的研究较多考虑单一盐(NaCl)对植物的影响, 主要涉及胁迫生理、表观遗传、离子转运、激素调节、基因表达等方面(孔令安等, 2000 ; Lu et al ., 2013 ; Wu et al ., 2013 ).而土壤碱胁迫这一严重的环境问题仍未得到研究者的充分重视, 仅有少量描述性报道涉及植物抗碱生理及分子机制(萨如拉等, 2014 ; 张一弓等, 2014 ).只有区别研究盐胁迫和碱胁迫, 才能真正揭示植物抗盐碱机制, 为最终培育出新的抗盐碱植物品种打下基础, 为盐碱化地的恢复和治理提供理论支持. ...
燕麦对碱胁迫的阳离子响应机制
1
2014
... 一直以来, 国内外抗盐方面的研究较多考虑单一盐(NaCl)对植物的影响, 主要涉及胁迫生理、表观遗传、离子转运、激素调节、基因表达等方面(孔令安等, 2000 ; Lu et al ., 2013 ; Wu et al ., 2013 ).而土壤碱胁迫这一严重的环境问题仍未得到研究者的充分重视, 仅有少量描述性报道涉及植物抗碱生理及分子机制(萨如拉等, 2014 ; 张一弓等, 2014 ).只有区别研究盐胁迫和碱胁迫, 才能真正揭示植物抗盐碱机制, 为最终培育出新的抗盐碱植物品种打下基础, 为盐碱化地的恢复和治理提供理论支持. ...
Effects of NaCl and Na2 CO3 on growth of Puccinellia tenuiflora and on present state of mineral elements in nutrient solution.
1
1997
... 叶绿素荧光特性可以反映出外界胁迫环境对植物光合器官的伤害程度, 其中PSII变化可以清楚显示出外界环境对植物光合能力的影响程度(冯玉龙等, 2001 ; 梁红柱等, 2004 ).本研究发现, 盐胁迫下亚麻叶片各项荧光参数并没有明显的变化, 这表明盐胁迫对植物体自身的光合能力影响并不明显, 之前的一些报道(Everard et al ., 1994 ; Lu et al ., 2002 ) 也证明了这一点.但是, 碱胁迫下PSII活性下降明显, 表明碱胁迫引起了光抑制, 导致亚麻叶片的光合能力降低, 主要原因可能是大量Na+ 和高pH值共同作用的结果.NQP 属于光合系统的自我保护机制, 在碱胁迫下NQP 的增加有利于提高光能利用效率, 增强光电子传递速率.q P 反映了光化学能量转换的能力, 是反映光合速率高低的有效指标, 本实验结果表明碱胁迫严重抑制了亚麻的光合效率(Bolhàr- Nordenkampf et al ., 1989 ).叶绿素和类胡萝卜素是高等植物的主要光合色素.叶绿素和类胡萝卜素的积累在盐胁迫下变化不明显, 但是在碱胁迫下下降明显.这可能是碱胁迫引起Mg沉淀从而抑制了叶绿素和类胡萝卜素的合成(Reddy & Vora, 1986 ; Shi & Zhao, 1997 ).另外, 也有可能是高pH值促进了叶绿素分解酶的活性. ...
盐碱混合生态条件的人工模拟及其对羊草胁迫作用因素分析
3
2002
... 全世界盐碱地面积已经超过总陆地面积的6%和农业灌溉面积的20%, 预计到2050年, 将会有超过50%的耕地盐渍化(张建锋等, 2005 ).我国耕地中盐碱地约占15%, 另外还有2000多万hm2 的盐荒地(李彬等, 2005 ).根据我国土壤特性和所含盐分特点, 将其分为盐土和碱土两大类, 其中最主要的致害离子为Na+ 、Cl- 、HCO3 - 和CO3 2- , 这些离子对植物的胁迫作用, 除了包括直接的胁迫效应外, 还包括离子间复杂的相互作用(石德成等, 2002 ). ...
... RGR是生命活动的最终体现, 可以作为植物对外界环境胁迫的响应指标(Yang et al ., 2008 ).本实验结果表明在相同盐浓度下, 碱胁迫RGR降幅大于盐胁迫.碱胁迫对植物的伤害甚于盐胁迫的原因可能与两种胁迫的作用机制有关, 盐胁迫主要是渗透胁迫及离子毒害, 而碱胁迫除包括这两种胁迫作用外, 还包括高pH胁迫(石德成等, 2002 ).当胁迫强度大于40 mmol·L-1 时, 碱胁迫(高pH值)明显地抑制亚麻RGR, 尤其是根系生长(图1 ).该结果与抗碱植物明显不同, 例如地肤(Kochia scoparia )和虎尾草(Chl- oris virgata ) (Yang et al ., 2008 ).抗碱植物在高强度碱胁迫下仍然可以保持较强的根RGR, 这表明植物的根系对pH值的调节和适应能力, 可能是决定植物体自身抗碱性的关键生理特性.AWC的变化可以反映植物体受到外界环境胁迫时的基本生理反应.盐、碱胁迫下亚麻AWC的变化趋势和RGR相似.结果充分证明盐、碱胁迫属于不同胁迫类型, 碱胁迫对植物构成的危害明显大于盐胁迫.得到的结论与以往的一些报道相一致(颜宏等, 2000 ; 石德成等, 2002 ).高pH值会引起根系周围某些离子发生沉淀, 导致根系附近离子失衡, 破坏根系对营养元素的吸收; 另外, 高pH值可能会导致泡内质子减少, 进而破坏或抑制根细胞跨膜电化学势梯度, 最终影响根系的正常生理功能. ...
... ; 石德成等, 2002 ).高pH值会引起根系周围某些离子发生沉淀, 导致根系附近离子失衡, 破坏根系对营养元素的吸收; 另外, 高pH值可能会导致泡内质子减少, 进而破坏或抑制根细胞跨膜电化学势梯度, 最终影响根系的正常生理功能. ...
盐碱混合生态条件的人工模拟及其对羊草胁迫作用因素分析
3
2002
... 全世界盐碱地面积已经超过总陆地面积的6%和农业灌溉面积的20%, 预计到2050年, 将会有超过50%的耕地盐渍化(张建锋等, 2005 ).我国耕地中盐碱地约占15%, 另外还有2000多万hm2 的盐荒地(李彬等, 2005 ).根据我国土壤特性和所含盐分特点, 将其分为盐土和碱土两大类, 其中最主要的致害离子为Na+ 、Cl- 、HCO3 - 和CO3 2- , 这些离子对植物的胁迫作用, 除了包括直接的胁迫效应外, 还包括离子间复杂的相互作用(石德成等, 2002 ). ...
... RGR是生命活动的最终体现, 可以作为植物对外界环境胁迫的响应指标(Yang et al ., 2008 ).本实验结果表明在相同盐浓度下, 碱胁迫RGR降幅大于盐胁迫.碱胁迫对植物的伤害甚于盐胁迫的原因可能与两种胁迫的作用机制有关, 盐胁迫主要是渗透胁迫及离子毒害, 而碱胁迫除包括这两种胁迫作用外, 还包括高pH胁迫(石德成等, 2002 ).当胁迫强度大于40 mmol·L-1 时, 碱胁迫(高pH值)明显地抑制亚麻RGR, 尤其是根系生长(图1 ).该结果与抗碱植物明显不同, 例如地肤(Kochia scoparia )和虎尾草(Chl- oris virgata ) (Yang et al ., 2008 ).抗碱植物在高强度碱胁迫下仍然可以保持较强的根RGR, 这表明植物的根系对pH值的调节和适应能力, 可能是决定植物体自身抗碱性的关键生理特性.AWC的变化可以反映植物体受到外界环境胁迫时的基本生理反应.盐、碱胁迫下亚麻AWC的变化趋势和RGR相似.结果充分证明盐、碱胁迫属于不同胁迫类型, 碱胁迫对植物构成的危害明显大于盐胁迫.得到的结论与以往的一些报道相一致(颜宏等, 2000 ; 石德成等, 2002 ).高pH值会引起根系周围某些离子发生沉淀, 导致根系附近离子失衡, 破坏根系对营养元素的吸收; 另外, 高pH值可能会导致泡内质子减少, 进而破坏或抑制根细胞跨膜电化学势梯度, 最终影响根系的正常生理功能. ...
... ; 石德成等, 2002 ).高pH值会引起根系周围某些离子发生沉淀, 导致根系附近离子失衡, 破坏根系对营养元素的吸收; 另外, 高pH值可能会导致泡内质子减少, 进而破坏或抑制根细胞跨膜电化学势梯度, 最终影响根系的正常生理功能. ...
Effects of various salt-alkaline mixed stresses on Aneurolepidium chinense (Trin.) Kitag.
1
2005
... 盐胁迫主要是渗透胁迫和离子毒害, 它影响各种离子在植物细胞内的分布, 破坏胞内离子平衡, 植物必须进行渗透调节并在细胞内重建离子稳态(Zhang et al ., 2010 ; Zhang & Shi, 2013 ).碱胁迫与盐胁迫相比, 除了渗透胁迫和离子毒害外, 还涉及高pH值伤害(Yang et al ., 2008 ).植物应对碱胁迫, 不仅要进行细胞内pH值调节以保持离子平衡, 同时还必须付出大量的物质和能量来调节根际微环境的pH值(Shi & Wang, 2005 ).所以渗透调节、细胞内离子平衡、根外pH值调节是植物抗碱胁迫的关键所在. ...
Na+ transport and Na+ tolerance in higher plants.
1
2003
... Na+ 和K+ 代谢是植物适应盐渍环境最为重要的生理代谢活动, Na+ 对于大多数植物而言是有害的, 当环境中Na+ 含量增高时, 会拮抗植物对K+ 的吸收, 从而限制植物生长(Brugnoli & Lauteri, 1991 ; 王宁等, 2015 ).本研究发现, 盐、碱胁迫均造成亚麻地上部分和根中Na+ 含量升高, K+ 含量降低, 碱胁迫下的变化趋势明显大于盐胁迫(图3A 1 , 3A2 , 3B1 , 3B2 ).结果表明与盐胁迫相比, 碱胁迫的高pH值进一步干扰和破坏了根对Na+ 、K+ 吸收的选择性, 导致体内Na+ 、K+ 严重失衡, 这是碱胁迫危害大于盐胁迫的一个主要原因.另外, 碱胁迫下亚麻地上部分中积累大量Na+ , 致使叶绿体遭到破坏, 叶绿素含量急剧下降, 导致光合能力降低(图2 ).研究还发现, 与根相比, 地上部分中K+ 含量保持在较高水平, 这说明亚麻有一个将Na+ 控制在根部而将更多的K+ 运输到地上部分的控制适应过程(Munns & Tester, 2008 ).Ca2+ 对维持细胞膜的稳定性起着重要作用, Mg2+ 是叶绿素的主要组分, 研究发现某些植物的Ca2+ 、Mg2+ 积累受到盐胁迫抑制.本研究结果表明盐、碱胁迫对亚麻地上部分中Ca2+ 、Mg2+ 含量影响不大, 但碱胁迫明显降低其在根中的含量(图3C 1 , 3C2 , 3D1 , 3D2 ).Ca2+ 作为第二信使与盐超敏感性蛋白(SOS3)结合, 刺激SOS2的激酶活性, 排出有毒离子Na+ (Zhu, 2003 ).但是碱胁迫(高pH值)导致根周围Ca2+ 沉淀, Ca2+ 吸收明显受阻, 进而影响Na+ 外排, 加速了离子毒害作用.另外, Na+ /H+ 逆向转运蛋白依靠细胞膜两侧质子梯度排出Na+ , 高pH值导致质子亏缺, 质子梯度难于建立, Na+ 不能外排, 导致植物体内积累大量的Na+ (Tester & Davenport, 2003 ; 王宁等, 2015 ).植物体维持细胞内离子平衡是保障各种代谢过程正常进行的必要条件, 离子平衡实际上就是阴阳离子平衡的过程(杨春武等, 2008 ).植物吸收阳离子除了特异性离子通道外, 还有一类非选择性阳离子通道(Munns & Tester, 2008 ), 盐碱胁迫下Na+ 大量进入根部, 可能直接抑制根对其他营养离子的吸收, 导致植物体内离子比率失衡, 如图3 中可见盐、碱胁迫明显降低亚麻K+ /Na+ 、Ca2+ /Na+ 和Mg2+ /Na+ , 并且碱胁迫下的下降幅度均大于盐胁迫.研究还发现, 盐、碱胁迫造成NO3 - 含量降低, 表明大量Na+ 积累阻碍根对NO3 - 的吸收, 明显干扰氮吸收或代谢.碱胁迫强烈干扰无机阴离子积累和分布, 碱胁迫(高pH值)阻碍质子梯度的建立, 从而抑制亚麻根系对阴离子的吸收, 造成植物营养缺乏, 这可能是碱胁迫对植物伤害甚于盐胁迫的另一个重要原因. ...
一氧化氮对水分胁迫下小麦叶片活性氧代谢及膜脂过氧化的影响
1
2006
... 植物在遭受盐胁迫时通常会迅速积累一些有机溶质作为细胞质中的渗透调节物质, 除渗透调节作用外, 还可以保护生物大分子并清除自由基(王罗霞等, 2006 ).可溶性糖是植物生命活动最主要的能源物质, 而且它还具有渗透保护、碳贮存和自由基清除的功能(Hare et al ., 1998 ).本研究结果表明盐胁迫促进可溶性糖的合成和积累, 它不仅参与渗透调节, 还中和大量的Na+ 避免离子伤害, 因此可溶性糖的积累可以增强亚麻对盐渍环境的耐性(Gao et al ., 2001 ).但是, 碱胁迫尤其是高碱胁迫抑制了可溶性糖的积累, 这可能是大量的Na+ 和高pH值共同作用的结果, 碱胁迫破坏了光系统结构, 造成了光合能力降低, 影响碳同化过程(Guo et al ., 2011 ).脯氨酸和甜菜碱作为非毒性的渗透调节物质在植物抗逆生理中起非常重要的作用(彭志红等, 2002 ; 张立新和李生秀, 2004 ).盐、碱胁迫都明显刺激了脯氨酸的积累, 脯氨酸的积累可能是长期离子毒性的代谢反应, 其含量的增加可能出于维持渗透压和平衡细胞液泡内无机离子浓度的需要.甜菜碱在盐碱胁迫下呈现下降的趋势, 这可能是由于Na+ 的大量沉淀, 造成根对NO3 - 吸收降低, 阻碍了N代谢的正常进行, 影响了甜菜碱的合成.从有机酸实验结果可知, 亚麻可能通过促进有机酸的合成来弥补无机阴离子的缺失和维护细胞内pH值的稳定(汪建飞和沈其荣, 2006 ).根中合成的有机酸可能是用来调节根外的pH值, 增加根系对铁等矿质元素的吸收和利用, 进而缓解碱胁迫造成的营养缺失和离子失衡. ...
一氧化氮对水分胁迫下小麦叶片活性氧代谢及膜脂过氧化的影响
1
2006
... 植物在遭受盐胁迫时通常会迅速积累一些有机溶质作为细胞质中的渗透调节物质, 除渗透调节作用外, 还可以保护生物大分子并清除自由基(王罗霞等, 2006 ).可溶性糖是植物生命活动最主要的能源物质, 而且它还具有渗透保护、碳贮存和自由基清除的功能(Hare et al ., 1998 ).本研究结果表明盐胁迫促进可溶性糖的合成和积累, 它不仅参与渗透调节, 还中和大量的Na+ 避免离子伤害, 因此可溶性糖的积累可以增强亚麻对盐渍环境的耐性(Gao et al ., 2001 ).但是, 碱胁迫尤其是高碱胁迫抑制了可溶性糖的积累, 这可能是大量的Na+ 和高pH值共同作用的结果, 碱胁迫破坏了光系统结构, 造成了光合能力降低, 影响碳同化过程(Guo et al ., 2011 ).脯氨酸和甜菜碱作为非毒性的渗透调节物质在植物抗逆生理中起非常重要的作用(彭志红等, 2002 ; 张立新和李生秀, 2004 ).盐、碱胁迫都明显刺激了脯氨酸的积累, 脯氨酸的积累可能是长期离子毒性的代谢反应, 其含量的增加可能出于维持渗透压和平衡细胞液泡内无机离子浓度的需要.甜菜碱在盐碱胁迫下呈现下降的趋势, 这可能是由于Na+ 的大量沉淀, 造成根对NO3 - 吸收降低, 阻碍了N代谢的正常进行, 影响了甜菜碱的合成.从有机酸实验结果可知, 亚麻可能通过促进有机酸的合成来弥补无机阴离子的缺失和维护细胞内pH值的稳定(汪建飞和沈其荣, 2006 ).根中合成的有机酸可能是用来调节根外的pH值, 增加根系对铁等矿质元素的吸收和利用, 进而缓解碱胁迫造成的营养缺失和离子失衡. ...
盐胁迫下棉花K+和Na+离子转运的耐盐性生理机制
2
2015
... Na+ 和K+ 代谢是植物适应盐渍环境最为重要的生理代谢活动, Na+ 对于大多数植物而言是有害的, 当环境中Na+ 含量增高时, 会拮抗植物对K+ 的吸收, 从而限制植物生长(Brugnoli & Lauteri, 1991 ; 王宁等, 2015 ).本研究发现, 盐、碱胁迫均造成亚麻地上部分和根中Na+ 含量升高, K+ 含量降低, 碱胁迫下的变化趋势明显大于盐胁迫(图3A 1 , 3A2 , 3B1 , 3B2 ).结果表明与盐胁迫相比, 碱胁迫的高pH值进一步干扰和破坏了根对Na+ 、K+ 吸收的选择性, 导致体内Na+ 、K+ 严重失衡, 这是碱胁迫危害大于盐胁迫的一个主要原因.另外, 碱胁迫下亚麻地上部分中积累大量Na+ , 致使叶绿体遭到破坏, 叶绿素含量急剧下降, 导致光合能力降低(图2 ).研究还发现, 与根相比, 地上部分中K+ 含量保持在较高水平, 这说明亚麻有一个将Na+ 控制在根部而将更多的K+ 运输到地上部分的控制适应过程(Munns & Tester, 2008 ).Ca2+ 对维持细胞膜的稳定性起着重要作用, Mg2+ 是叶绿素的主要组分, 研究发现某些植物的Ca2+ 、Mg2+ 积累受到盐胁迫抑制.本研究结果表明盐、碱胁迫对亚麻地上部分中Ca2+ 、Mg2+ 含量影响不大, 但碱胁迫明显降低其在根中的含量(图3C 1 , 3C2 , 3D1 , 3D2 ).Ca2+ 作为第二信使与盐超敏感性蛋白(SOS3)结合, 刺激SOS2的激酶活性, 排出有毒离子Na+ (Zhu, 2003 ).但是碱胁迫(高pH值)导致根周围Ca2+ 沉淀, Ca2+ 吸收明显受阻, 进而影响Na+ 外排, 加速了离子毒害作用.另外, Na+ /H+ 逆向转运蛋白依靠细胞膜两侧质子梯度排出Na+ , 高pH值导致质子亏缺, 质子梯度难于建立, Na+ 不能外排, 导致植物体内积累大量的Na+ (Tester & Davenport, 2003 ; 王宁等, 2015 ).植物体维持细胞内离子平衡是保障各种代谢过程正常进行的必要条件, 离子平衡实际上就是阴阳离子平衡的过程(杨春武等, 2008 ).植物吸收阳离子除了特异性离子通道外, 还有一类非选择性阳离子通道(Munns & Tester, 2008 ), 盐碱胁迫下Na+ 大量进入根部, 可能直接抑制根对其他营养离子的吸收, 导致植物体内离子比率失衡, 如图3 中可见盐、碱胁迫明显降低亚麻K+ /Na+ 、Ca2+ /Na+ 和Mg2+ /Na+ , 并且碱胁迫下的下降幅度均大于盐胁迫.研究还发现, 盐、碱胁迫造成NO3 - 含量降低, 表明大量Na+ 积累阻碍根对NO3 - 的吸收, 明显干扰氮吸收或代谢.碱胁迫强烈干扰无机阴离子积累和分布, 碱胁迫(高pH值)阻碍质子梯度的建立, 从而抑制亚麻根系对阴离子的吸收, 造成植物营养缺乏, 这可能是碱胁迫对植物伤害甚于盐胁迫的另一个重要原因. ...
... ; 王宁等, 2015 ).植物体维持细胞内离子平衡是保障各种代谢过程正常进行的必要条件, 离子平衡实际上就是阴阳离子平衡的过程(杨春武等, 2008 ).植物吸收阳离子除了特异性离子通道外, 还有一类非选择性阳离子通道(Munns & Tester, 2008 ), 盐碱胁迫下Na+ 大量进入根部, 可能直接抑制根对其他营养离子的吸收, 导致植物体内离子比率失衡, 如图3 中可见盐、碱胁迫明显降低亚麻K+ /Na+ 、Ca2+ /Na+ 和Mg2+ /Na+ , 并且碱胁迫下的下降幅度均大于盐胁迫.研究还发现, 盐、碱胁迫造成NO3 - 含量降低, 表明大量Na+ 积累阻碍根对NO3 - 的吸收, 明显干扰氮吸收或代谢.碱胁迫强烈干扰无机阴离子积累和分布, 碱胁迫(高pH值)阻碍质子梯度的建立, 从而抑制亚麻根系对阴离子的吸收, 造成植物营养缺乏, 这可能是碱胁迫对植物伤害甚于盐胁迫的另一个重要原因. ...
盐胁迫下棉花K+和Na+离子转运的耐盐性生理机制
2
2015
... Na+ 和K+ 代谢是植物适应盐渍环境最为重要的生理代谢活动, Na+ 对于大多数植物而言是有害的, 当环境中Na+ 含量增高时, 会拮抗植物对K+ 的吸收, 从而限制植物生长(Brugnoli & Lauteri, 1991 ; 王宁等, 2015 ).本研究发现, 盐、碱胁迫均造成亚麻地上部分和根中Na+ 含量升高, K+ 含量降低, 碱胁迫下的变化趋势明显大于盐胁迫(图3A 1 , 3A2 , 3B1 , 3B2 ).结果表明与盐胁迫相比, 碱胁迫的高pH值进一步干扰和破坏了根对Na+ 、K+ 吸收的选择性, 导致体内Na+ 、K+ 严重失衡, 这是碱胁迫危害大于盐胁迫的一个主要原因.另外, 碱胁迫下亚麻地上部分中积累大量Na+ , 致使叶绿体遭到破坏, 叶绿素含量急剧下降, 导致光合能力降低(图2 ).研究还发现, 与根相比, 地上部分中K+ 含量保持在较高水平, 这说明亚麻有一个将Na+ 控制在根部而将更多的K+ 运输到地上部分的控制适应过程(Munns & Tester, 2008 ).Ca2+ 对维持细胞膜的稳定性起着重要作用, Mg2+ 是叶绿素的主要组分, 研究发现某些植物的Ca2+ 、Mg2+ 积累受到盐胁迫抑制.本研究结果表明盐、碱胁迫对亚麻地上部分中Ca2+ 、Mg2+ 含量影响不大, 但碱胁迫明显降低其在根中的含量(图3C 1 , 3C2 , 3D1 , 3D2 ).Ca2+ 作为第二信使与盐超敏感性蛋白(SOS3)结合, 刺激SOS2的激酶活性, 排出有毒离子Na+ (Zhu, 2003 ).但是碱胁迫(高pH值)导致根周围Ca2+ 沉淀, Ca2+ 吸收明显受阻, 进而影响Na+ 外排, 加速了离子毒害作用.另外, Na+ /H+ 逆向转运蛋白依靠细胞膜两侧质子梯度排出Na+ , 高pH值导致质子亏缺, 质子梯度难于建立, Na+ 不能外排, 导致植物体内积累大量的Na+ (Tester & Davenport, 2003 ; 王宁等, 2015 ).植物体维持细胞内离子平衡是保障各种代谢过程正常进行的必要条件, 离子平衡实际上就是阴阳离子平衡的过程(杨春武等, 2008 ).植物吸收阳离子除了特异性离子通道外, 还有一类非选择性阳离子通道(Munns & Tester, 2008 ), 盐碱胁迫下Na+ 大量进入根部, 可能直接抑制根对其他营养离子的吸收, 导致植物体内离子比率失衡, 如图3 中可见盐、碱胁迫明显降低亚麻K+ /Na+ 、Ca2+ /Na+ 和Mg2+ /Na+ , 并且碱胁迫下的下降幅度均大于盐胁迫.研究还发现, 盐、碱胁迫造成NO3 - 含量降低, 表明大量Na+ 积累阻碍根对NO3 - 的吸收, 明显干扰氮吸收或代谢.碱胁迫强烈干扰无机阴离子积累和分布, 碱胁迫(高pH值)阻碍质子梯度的建立, 从而抑制亚麻根系对阴离子的吸收, 造成植物营养缺乏, 这可能是碱胁迫对植物伤害甚于盐胁迫的另一个重要原因. ...
... ; 王宁等, 2015 ).植物体维持细胞内离子平衡是保障各种代谢过程正常进行的必要条件, 离子平衡实际上就是阴阳离子平衡的过程(杨春武等, 2008 ).植物吸收阳离子除了特异性离子通道外, 还有一类非选择性阳离子通道(Munns & Tester, 2008 ), 盐碱胁迫下Na+ 大量进入根部, 可能直接抑制根对其他营养离子的吸收, 导致植物体内离子比率失衡, 如图3 中可见盐、碱胁迫明显降低亚麻K+ /Na+ 、Ca2+ /Na+ 和Mg2+ /Na+ , 并且碱胁迫下的下降幅度均大于盐胁迫.研究还发现, 盐、碱胁迫造成NO3 - 含量降低, 表明大量Na+ 积累阻碍根对NO3 - 的吸收, 明显干扰氮吸收或代谢.碱胁迫强烈干扰无机阴离子积累和分布, 碱胁迫(高pH值)阻碍质子梯度的建立, 从而抑制亚麻根系对阴离子的吸收, 造成植物营养缺乏, 这可能是碱胁迫对植物伤害甚于盐胁迫的另一个重要原因. ...
有机酸代谢在植物适应养分和铝毒胁迫中的作用
1
2006
... 植物在遭受盐胁迫时通常会迅速积累一些有机溶质作为细胞质中的渗透调节物质, 除渗透调节作用外, 还可以保护生物大分子并清除自由基(王罗霞等, 2006 ).可溶性糖是植物生命活动最主要的能源物质, 而且它还具有渗透保护、碳贮存和自由基清除的功能(Hare et al ., 1998 ).本研究结果表明盐胁迫促进可溶性糖的合成和积累, 它不仅参与渗透调节, 还中和大量的Na+ 避免离子伤害, 因此可溶性糖的积累可以增强亚麻对盐渍环境的耐性(Gao et al ., 2001 ).但是, 碱胁迫尤其是高碱胁迫抑制了可溶性糖的积累, 这可能是大量的Na+ 和高pH值共同作用的结果, 碱胁迫破坏了光系统结构, 造成了光合能力降低, 影响碳同化过程(Guo et al ., 2011 ).脯氨酸和甜菜碱作为非毒性的渗透调节物质在植物抗逆生理中起非常重要的作用(彭志红等, 2002 ; 张立新和李生秀, 2004 ).盐、碱胁迫都明显刺激了脯氨酸的积累, 脯氨酸的积累可能是长期离子毒性的代谢反应, 其含量的增加可能出于维持渗透压和平衡细胞液泡内无机离子浓度的需要.甜菜碱在盐碱胁迫下呈现下降的趋势, 这可能是由于Na+ 的大量沉淀, 造成根对NO3 - 吸收降低, 阻碍了N代谢的正常进行, 影响了甜菜碱的合成.从有机酸实验结果可知, 亚麻可能通过促进有机酸的合成来弥补无机阴离子的缺失和维护细胞内pH值的稳定(汪建飞和沈其荣, 2006 ).根中合成的有机酸可能是用来调节根外的pH值, 增加根系对铁等矿质元素的吸收和利用, 进而缓解碱胁迫造成的营养缺失和离子失衡. ...
有机酸代谢在植物适应养分和铝毒胁迫中的作用
1
2006
... 植物在遭受盐胁迫时通常会迅速积累一些有机溶质作为细胞质中的渗透调节物质, 除渗透调节作用外, 还可以保护生物大分子并清除自由基(王罗霞等, 2006 ).可溶性糖是植物生命活动最主要的能源物质, 而且它还具有渗透保护、碳贮存和自由基清除的功能(Hare et al ., 1998 ).本研究结果表明盐胁迫促进可溶性糖的合成和积累, 它不仅参与渗透调节, 还中和大量的Na+ 避免离子伤害, 因此可溶性糖的积累可以增强亚麻对盐渍环境的耐性(Gao et al ., 2001 ).但是, 碱胁迫尤其是高碱胁迫抑制了可溶性糖的积累, 这可能是大量的Na+ 和高pH值共同作用的结果, 碱胁迫破坏了光系统结构, 造成了光合能力降低, 影响碳同化过程(Guo et al ., 2011 ).脯氨酸和甜菜碱作为非毒性的渗透调节物质在植物抗逆生理中起非常重要的作用(彭志红等, 2002 ; 张立新和李生秀, 2004 ).盐、碱胁迫都明显刺激了脯氨酸的积累, 脯氨酸的积累可能是长期离子毒性的代谢反应, 其含量的增加可能出于维持渗透压和平衡细胞液泡内无机离子浓度的需要.甜菜碱在盐碱胁迫下呈现下降的趋势, 这可能是由于Na+ 的大量沉淀, 造成根对NO3 - 吸收降低, 阻碍了N代谢的正常进行, 影响了甜菜碱的合成.从有机酸实验结果可知, 亚麻可能通过促进有机酸的合成来弥补无机阴离子的缺失和维护细胞内pH值的稳定(汪建飞和沈其荣, 2006 ).根中合成的有机酸可能是用来调节根外的pH值, 增加根系对铁等矿质元素的吸收和利用, 进而缓解碱胁迫造成的营养缺失和离子失衡. ...
Ionomic responses and correlations between elements and metabolites under salt stress in wild and cultivated barley.
1
2013
... 一直以来, 国内外抗盐方面的研究较多考虑单一盐(NaCl)对植物的影响, 主要涉及胁迫生理、表观遗传、离子转运、激素调节、基因表达等方面(孔令安等, 2000 ; Lu et al ., 2013 ; Wu et al ., 2013 ).而土壤碱胁迫这一严重的环境问题仍未得到研究者的充分重视, 仅有少量描述性报道涉及植物抗碱生理及分子机制(萨如拉等, 2014 ; 张一弓等, 2014 ).只有区别研究盐胁迫和碱胁迫, 才能真正揭示植物抗盐碱机制, 为最终培育出新的抗盐碱植物品种打下基础, 为盐碱化地的恢复和治理提供理论支持. ...
盐碱胁迫对羊草体内N及几种有机代谢产物积累的影响
1
2000
... RGR是生命活动的最终体现, 可以作为植物对外界环境胁迫的响应指标(Yang et al ., 2008 ).本实验结果表明在相同盐浓度下, 碱胁迫RGR降幅大于盐胁迫.碱胁迫对植物的伤害甚于盐胁迫的原因可能与两种胁迫的作用机制有关, 盐胁迫主要是渗透胁迫及离子毒害, 而碱胁迫除包括这两种胁迫作用外, 还包括高pH胁迫(石德成等, 2002 ).当胁迫强度大于40 mmol·L-1 时, 碱胁迫(高pH值)明显地抑制亚麻RGR, 尤其是根系生长(图1 ).该结果与抗碱植物明显不同, 例如地肤(Kochia scoparia )和虎尾草(Chl- oris virgata ) (Yang et al ., 2008 ).抗碱植物在高强度碱胁迫下仍然可以保持较强的根RGR, 这表明植物的根系对pH值的调节和适应能力, 可能是决定植物体自身抗碱性的关键生理特性.AWC的变化可以反映植物体受到外界环境胁迫时的基本生理反应.盐、碱胁迫下亚麻AWC的变化趋势和RGR相似.结果充分证明盐、碱胁迫属于不同胁迫类型, 碱胁迫对植物构成的危害明显大于盐胁迫.得到的结论与以往的一些报道相一致(颜宏等, 2000 ; 石德成等, 2002 ).高pH值会引起根系周围某些离子发生沉淀, 导致根系附近离子失衡, 破坏根系对营养元素的吸收; 另外, 高pH值可能会导致泡内质子减少, 进而破坏或抑制根细胞跨膜电化学势梯度, 最终影响根系的正常生理功能. ...
盐碱胁迫对羊草体内N及几种有机代谢产物积累的影响
1
2000
... RGR是生命活动的最终体现, 可以作为植物对外界环境胁迫的响应指标(Yang et al ., 2008 ).本实验结果表明在相同盐浓度下, 碱胁迫RGR降幅大于盐胁迫.碱胁迫对植物的伤害甚于盐胁迫的原因可能与两种胁迫的作用机制有关, 盐胁迫主要是渗透胁迫及离子毒害, 而碱胁迫除包括这两种胁迫作用外, 还包括高pH胁迫(石德成等, 2002 ).当胁迫强度大于40 mmol·L-1 时, 碱胁迫(高pH值)明显地抑制亚麻RGR, 尤其是根系生长(图1 ).该结果与抗碱植物明显不同, 例如地肤(Kochia scoparia )和虎尾草(Chl- oris virgata ) (Yang et al ., 2008 ).抗碱植物在高强度碱胁迫下仍然可以保持较强的根RGR, 这表明植物的根系对pH值的调节和适应能力, 可能是决定植物体自身抗碱性的关键生理特性.AWC的变化可以反映植物体受到外界环境胁迫时的基本生理反应.盐、碱胁迫下亚麻AWC的变化趋势和RGR相似.结果充分证明盐、碱胁迫属于不同胁迫类型, 碱胁迫对植物构成的危害明显大于盐胁迫.得到的结论与以往的一些报道相一致(颜宏等, 2000 ; 石德成等, 2002 ).高pH值会引起根系周围某些离子发生沉淀, 导致根系附近离子失衡, 破坏根系对营养元素的吸收; 另外, 高pH值可能会导致泡内质子减少, 进而破坏或抑制根细胞跨膜电化学势梯度, 最终影响根系的正常生理功能. ...
Comparative effects of salt stress and alkali stress on growth, osmotic adjustment and ionic balance of an alkali resistant halophyte Suaeda glauca Bge.
3
2008
... 盐胁迫主要是渗透胁迫和离子毒害, 它影响各种离子在植物细胞内的分布, 破坏胞内离子平衡, 植物必须进行渗透调节并在细胞内重建离子稳态(Zhang et al ., 2010 ; Zhang & Shi, 2013 ).碱胁迫与盐胁迫相比, 除了渗透胁迫和离子毒害外, 还涉及高pH值伤害(Yang et al ., 2008 ).植物应对碱胁迫, 不仅要进行细胞内pH值调节以保持离子平衡, 同时还必须付出大量的物质和能量来调节根际微环境的pH值(Shi & Wang, 2005 ).所以渗透调节、细胞内离子平衡、根外pH值调节是植物抗碱胁迫的关键所在. ...
... RGR是生命活动的最终体现, 可以作为植物对外界环境胁迫的响应指标(Yang et al ., 2008 ).本实验结果表明在相同盐浓度下, 碱胁迫RGR降幅大于盐胁迫.碱胁迫对植物的伤害甚于盐胁迫的原因可能与两种胁迫的作用机制有关, 盐胁迫主要是渗透胁迫及离子毒害, 而碱胁迫除包括这两种胁迫作用外, 还包括高pH胁迫(石德成等, 2002 ).当胁迫强度大于40 mmol·L-1 时, 碱胁迫(高pH值)明显地抑制亚麻RGR, 尤其是根系生长(图1 ).该结果与抗碱植物明显不同, 例如地肤(Kochia scoparia )和虎尾草(Chl- oris virgata ) (Yang et al ., 2008 ).抗碱植物在高强度碱胁迫下仍然可以保持较强的根RGR, 这表明植物的根系对pH值的调节和适应能力, 可能是决定植物体自身抗碱性的关键生理特性.AWC的变化可以反映植物体受到外界环境胁迫时的基本生理反应.盐、碱胁迫下亚麻AWC的变化趋势和RGR相似.结果充分证明盐、碱胁迫属于不同胁迫类型, 碱胁迫对植物构成的危害明显大于盐胁迫.得到的结论与以往的一些报道相一致(颜宏等, 2000 ; 石德成等, 2002 ).高pH值会引起根系周围某些离子发生沉淀, 导致根系附近离子失衡, 破坏根系对营养元素的吸收; 另外, 高pH值可能会导致泡内质子减少, 进而破坏或抑制根细胞跨膜电化学势梯度, 最终影响根系的正常生理功能. ...
... ) (Yang et al ., 2008 ).抗碱植物在高强度碱胁迫下仍然可以保持较强的根RGR, 这表明植物的根系对pH值的调节和适应能力, 可能是决定植物体自身抗碱性的关键生理特性.AWC的变化可以反映植物体受到外界环境胁迫时的基本生理反应.盐、碱胁迫下亚麻AWC的变化趋势和RGR相似.结果充分证明盐、碱胁迫属于不同胁迫类型, 碱胁迫对植物构成的危害明显大于盐胁迫.得到的结论与以往的一些报道相一致(颜宏等, 2000 ; 石德成等, 2002 ).高pH值会引起根系周围某些离子发生沉淀, 导致根系附近离子失衡, 破坏根系对营养元素的吸收; 另外, 高pH值可能会导致泡内质子减少, 进而破坏或抑制根细胞跨膜电化学势梯度, 最终影响根系的正常生理功能. ...
盐、碱胁迫下小冰麦体内的pH及离子平衡
1
2008
... Na+ 和K+ 代谢是植物适应盐渍环境最为重要的生理代谢活动, Na+ 对于大多数植物而言是有害的, 当环境中Na+ 含量增高时, 会拮抗植物对K+ 的吸收, 从而限制植物生长(Brugnoli & Lauteri, 1991 ; 王宁等, 2015 ).本研究发现, 盐、碱胁迫均造成亚麻地上部分和根中Na+ 含量升高, K+ 含量降低, 碱胁迫下的变化趋势明显大于盐胁迫(图3A 1 , 3A2 , 3B1 , 3B2 ).结果表明与盐胁迫相比, 碱胁迫的高pH值进一步干扰和破坏了根对Na+ 、K+ 吸收的选择性, 导致体内Na+ 、K+ 严重失衡, 这是碱胁迫危害大于盐胁迫的一个主要原因.另外, 碱胁迫下亚麻地上部分中积累大量Na+ , 致使叶绿体遭到破坏, 叶绿素含量急剧下降, 导致光合能力降低(图2 ).研究还发现, 与根相比, 地上部分中K+ 含量保持在较高水平, 这说明亚麻有一个将Na+ 控制在根部而将更多的K+ 运输到地上部分的控制适应过程(Munns & Tester, 2008 ).Ca2+ 对维持细胞膜的稳定性起着重要作用, Mg2+ 是叶绿素的主要组分, 研究发现某些植物的Ca2+ 、Mg2+ 积累受到盐胁迫抑制.本研究结果表明盐、碱胁迫对亚麻地上部分中Ca2+ 、Mg2+ 含量影响不大, 但碱胁迫明显降低其在根中的含量(图3C 1 , 3C2 , 3D1 , 3D2 ).Ca2+ 作为第二信使与盐超敏感性蛋白(SOS3)结合, 刺激SOS2的激酶活性, 排出有毒离子Na+ (Zhu, 2003 ).但是碱胁迫(高pH值)导致根周围Ca2+ 沉淀, Ca2+ 吸收明显受阻, 进而影响Na+ 外排, 加速了离子毒害作用.另外, Na+ /H+ 逆向转运蛋白依靠细胞膜两侧质子梯度排出Na+ , 高pH值导致质子亏缺, 质子梯度难于建立, Na+ 不能外排, 导致植物体内积累大量的Na+ (Tester & Davenport, 2003 ; 王宁等, 2015 ).植物体维持细胞内离子平衡是保障各种代谢过程正常进行的必要条件, 离子平衡实际上就是阴阳离子平衡的过程(杨春武等, 2008 ).植物吸收阳离子除了特异性离子通道外, 还有一类非选择性阳离子通道(Munns & Tester, 2008 ), 盐碱胁迫下Na+ 大量进入根部, 可能直接抑制根对其他营养离子的吸收, 导致植物体内离子比率失衡, 如图3 中可见盐、碱胁迫明显降低亚麻K+ /Na+ 、Ca2+ /Na+ 和Mg2+ /Na+ , 并且碱胁迫下的下降幅度均大于盐胁迫.研究还发现, 盐、碱胁迫造成NO3 - 含量降低, 表明大量Na+ 积累阻碍根对NO3 - 的吸收, 明显干扰氮吸收或代谢.碱胁迫强烈干扰无机阴离子积累和分布, 碱胁迫(高pH值)阻碍质子梯度的建立, 从而抑制亚麻根系对阴离子的吸收, 造成植物营养缺乏, 这可能是碱胁迫对植物伤害甚于盐胁迫的另一个重要原因. ...
盐、碱胁迫下小冰麦体内的pH及离子平衡
1
2008
... Na+ 和K+ 代谢是植物适应盐渍环境最为重要的生理代谢活动, Na+ 对于大多数植物而言是有害的, 当环境中Na+ 含量增高时, 会拮抗植物对K+ 的吸收, 从而限制植物生长(Brugnoli & Lauteri, 1991 ; 王宁等, 2015 ).本研究发现, 盐、碱胁迫均造成亚麻地上部分和根中Na+ 含量升高, K+ 含量降低, 碱胁迫下的变化趋势明显大于盐胁迫(图3A 1 , 3A2 , 3B1 , 3B2 ).结果表明与盐胁迫相比, 碱胁迫的高pH值进一步干扰和破坏了根对Na+ 、K+ 吸收的选择性, 导致体内Na+ 、K+ 严重失衡, 这是碱胁迫危害大于盐胁迫的一个主要原因.另外, 碱胁迫下亚麻地上部分中积累大量Na+ , 致使叶绿体遭到破坏, 叶绿素含量急剧下降, 导致光合能力降低(图2 ).研究还发现, 与根相比, 地上部分中K+ 含量保持在较高水平, 这说明亚麻有一个将Na+ 控制在根部而将更多的K+ 运输到地上部分的控制适应过程(Munns & Tester, 2008 ).Ca2+ 对维持细胞膜的稳定性起着重要作用, Mg2+ 是叶绿素的主要组分, 研究发现某些植物的Ca2+ 、Mg2+ 积累受到盐胁迫抑制.本研究结果表明盐、碱胁迫对亚麻地上部分中Ca2+ 、Mg2+ 含量影响不大, 但碱胁迫明显降低其在根中的含量(图3C 1 , 3C2 , 3D1 , 3D2 ).Ca2+ 作为第二信使与盐超敏感性蛋白(SOS3)结合, 刺激SOS2的激酶活性, 排出有毒离子Na+ (Zhu, 2003 ).但是碱胁迫(高pH值)导致根周围Ca2+ 沉淀, Ca2+ 吸收明显受阻, 进而影响Na+ 外排, 加速了离子毒害作用.另外, Na+ /H+ 逆向转运蛋白依靠细胞膜两侧质子梯度排出Na+ , 高pH值导致质子亏缺, 质子梯度难于建立, Na+ 不能外排, 导致植物体内积累大量的Na+ (Tester & Davenport, 2003 ; 王宁等, 2015 ).植物体维持细胞内离子平衡是保障各种代谢过程正常进行的必要条件, 离子平衡实际上就是阴阳离子平衡的过程(杨春武等, 2008 ).植物吸收阳离子除了特异性离子通道外, 还有一类非选择性阳离子通道(Munns & Tester, 2008 ), 盐碱胁迫下Na+ 大量进入根部, 可能直接抑制根对其他营养离子的吸收, 导致植物体内离子比率失衡, 如图3 中可见盐、碱胁迫明显降低亚麻K+ /Na+ 、Ca2+ /Na+ 和Mg2+ /Na+ , 并且碱胁迫下的下降幅度均大于盐胁迫.研究还发现, 盐、碱胁迫造成NO3 - 含量降低, 表明大量Na+ 积累阻碍根对NO3 - 的吸收, 明显干扰氮吸收或代谢.碱胁迫强烈干扰无机阴离子积累和分布, 碱胁迫(高pH值)阻碍质子梯度的建立, 从而抑制亚麻根系对阴离子的吸收, 造成植物营养缺乏, 这可能是碱胁迫对植物伤害甚于盐胁迫的另一个重要原因. ...
甜菜碱与植物抗旱/盐性研究进展
1
2004
... 植物在遭受盐胁迫时通常会迅速积累一些有机溶质作为细胞质中的渗透调节物质, 除渗透调节作用外, 还可以保护生物大分子并清除自由基(王罗霞等, 2006 ).可溶性糖是植物生命活动最主要的能源物质, 而且它还具有渗透保护、碳贮存和自由基清除的功能(Hare et al ., 1998 ).本研究结果表明盐胁迫促进可溶性糖的合成和积累, 它不仅参与渗透调节, 还中和大量的Na+ 避免离子伤害, 因此可溶性糖的积累可以增强亚麻对盐渍环境的耐性(Gao et al ., 2001 ).但是, 碱胁迫尤其是高碱胁迫抑制了可溶性糖的积累, 这可能是大量的Na+ 和高pH值共同作用的结果, 碱胁迫破坏了光系统结构, 造成了光合能力降低, 影响碳同化过程(Guo et al ., 2011 ).脯氨酸和甜菜碱作为非毒性的渗透调节物质在植物抗逆生理中起非常重要的作用(彭志红等, 2002 ; 张立新和李生秀, 2004 ).盐、碱胁迫都明显刺激了脯氨酸的积累, 脯氨酸的积累可能是长期离子毒性的代谢反应, 其含量的增加可能出于维持渗透压和平衡细胞液泡内无机离子浓度的需要.甜菜碱在盐碱胁迫下呈现下降的趋势, 这可能是由于Na+ 的大量沉淀, 造成根对NO3 - 吸收降低, 阻碍了N代谢的正常进行, 影响了甜菜碱的合成.从有机酸实验结果可知, 亚麻可能通过促进有机酸的合成来弥补无机阴离子的缺失和维护细胞内pH值的稳定(汪建飞和沈其荣, 2006 ).根中合成的有机酸可能是用来调节根外的pH值, 增加根系对铁等矿质元素的吸收和利用, 进而缓解碱胁迫造成的营养缺失和离子失衡. ...
甜菜碱与植物抗旱/盐性研究进展
1
2004
... 植物在遭受盐胁迫时通常会迅速积累一些有机溶质作为细胞质中的渗透调节物质, 除渗透调节作用外, 还可以保护生物大分子并清除自由基(王罗霞等, 2006 ).可溶性糖是植物生命活动最主要的能源物质, 而且它还具有渗透保护、碳贮存和自由基清除的功能(Hare et al ., 1998 ).本研究结果表明盐胁迫促进可溶性糖的合成和积累, 它不仅参与渗透调节, 还中和大量的Na+ 避免离子伤害, 因此可溶性糖的积累可以增强亚麻对盐渍环境的耐性(Gao et al ., 2001 ).但是, 碱胁迫尤其是高碱胁迫抑制了可溶性糖的积累, 这可能是大量的Na+ 和高pH值共同作用的结果, 碱胁迫破坏了光系统结构, 造成了光合能力降低, 影响碳同化过程(Guo et al ., 2011 ).脯氨酸和甜菜碱作为非毒性的渗透调节物质在植物抗逆生理中起非常重要的作用(彭志红等, 2002 ; 张立新和李生秀, 2004 ).盐、碱胁迫都明显刺激了脯氨酸的积累, 脯氨酸的积累可能是长期离子毒性的代谢反应, 其含量的增加可能出于维持渗透压和平衡细胞液泡内无机离子浓度的需要.甜菜碱在盐碱胁迫下呈现下降的趋势, 这可能是由于Na+ 的大量沉淀, 造成根对NO3 - 吸收降低, 阻碍了N代谢的正常进行, 影响了甜菜碱的合成.从有机酸实验结果可知, 亚麻可能通过促进有机酸的合成来弥补无机阴离子的缺失和维护细胞内pH值的稳定(汪建飞和沈其荣, 2006 ).根中合成的有机酸可能是用来调节根外的pH值, 增加根系对铁等矿质元素的吸收和利用, 进而缓解碱胁迫造成的营养缺失和离子失衡. ...
世界盐碱地资源及其改良利用的基本措施
1
2005
... 全世界盐碱地面积已经超过总陆地面积的6%和农业灌溉面积的20%, 预计到2050年, 将会有超过50%的耕地盐渍化(张建锋等, 2005 ).我国耕地中盐碱地约占15%, 另外还有2000多万hm2 的盐荒地(李彬等, 2005 ).根据我国土壤特性和所含盐分特点, 将其分为盐土和碱土两大类, 其中最主要的致害离子为Na+ 、Cl- 、HCO3 - 和CO3 2- , 这些离子对植物的胁迫作用, 除了包括直接的胁迫效应外, 还包括离子间复杂的相互作用(石德成等, 2002 ). ...
世界盐碱地资源及其改良利用的基本措施
1
2005
... 全世界盐碱地面积已经超过总陆地面积的6%和农业灌溉面积的20%, 预计到2050年, 将会有超过50%的耕地盐渍化(张建锋等, 2005 ).我国耕地中盐碱地约占15%, 另外还有2000多万hm2 的盐荒地(李彬等, 2005 ).根据我国土壤特性和所含盐分特点, 将其分为盐土和碱土两大类, 其中最主要的致害离子为Na+ 、Cl- 、HCO3 - 和CO3 2- , 这些离子对植物的胁迫作用, 除了包括直接的胁迫效应外, 还包括离子间复杂的相互作用(石德成等, 2002 ). ...
Mechanisms of sodium uptake by roots of higher plants.
1
2010
... 盐胁迫主要是渗透胁迫和离子毒害, 它影响各种离子在植物细胞内的分布, 破坏胞内离子平衡, 植物必须进行渗透调节并在细胞内重建离子稳态(Zhang et al ., 2010 ; Zhang & Shi, 2013 ).碱胁迫与盐胁迫相比, 除了渗透胁迫和离子毒害外, 还涉及高pH值伤害(Yang et al ., 2008 ).植物应对碱胁迫, 不仅要进行细胞内pH值调节以保持离子平衡, 同时还必须付出大量的物质和能量来调节根际微环境的pH值(Shi & Wang, 2005 ).所以渗透调节、细胞内离子平衡、根外pH值调节是植物抗碱胁迫的关键所在. ...
Physiological and molecular mechanisms of plant salt tolerance.
1
2013
... 盐胁迫主要是渗透胁迫和离子毒害, 它影响各种离子在植物细胞内的分布, 破坏胞内离子平衡, 植物必须进行渗透调节并在细胞内重建离子稳态(Zhang et al ., 2010 ; Zhang & Shi, 2013 ).碱胁迫与盐胁迫相比, 除了渗透胁迫和离子毒害外, 还涉及高pH值伤害(Yang et al ., 2008 ).植物应对碱胁迫, 不仅要进行细胞内pH值调节以保持离子平衡, 同时还必须付出大量的物质和能量来调节根际微环境的pH值(Shi & Wang, 2005 ).所以渗透调节、细胞内离子平衡、根外pH值调节是植物抗碱胁迫的关键所在. ...
碱胁迫下塔乌库姆冰草的萌发及生理响应
1
2014
... 一直以来, 国内外抗盐方面的研究较多考虑单一盐(NaCl)对植物的影响, 主要涉及胁迫生理、表观遗传、离子转运、激素调节、基因表达等方面(孔令安等, 2000 ; Lu et al ., 2013 ; Wu et al ., 2013 ).而土壤碱胁迫这一严重的环境问题仍未得到研究者的充分重视, 仅有少量描述性报道涉及植物抗碱生理及分子机制(萨如拉等, 2014 ; 张一弓等, 2014 ).只有区别研究盐胁迫和碱胁迫, 才能真正揭示植物抗盐碱机制, 为最终培育出新的抗盐碱植物品种打下基础, 为盐碱化地的恢复和治理提供理论支持. ...
碱胁迫下塔乌库姆冰草的萌发及生理响应
1
2014
... 一直以来, 国内外抗盐方面的研究较多考虑单一盐(NaCl)对植物的影响, 主要涉及胁迫生理、表观遗传、离子转运、激素调节、基因表达等方面(孔令安等, 2000 ; Lu et al ., 2013 ; Wu et al ., 2013 ).而土壤碱胁迫这一严重的环境问题仍未得到研究者的充分重视, 仅有少量描述性报道涉及植物抗碱生理及分子机制(萨如拉等, 2014 ; 张一弓等, 2014 ).只有区别研究盐胁迫和碱胁迫, 才能真正揭示植物抗盐碱机制, 为最终培育出新的抗盐碱植物品种打下基础, 为盐碱化地的恢复和治理提供理论支持. ...
Regulation of ion homeostasis under salt stress.
1
2003
... Na+ 和K+ 代谢是植物适应盐渍环境最为重要的生理代谢活动, Na+ 对于大多数植物而言是有害的, 当环境中Na+ 含量增高时, 会拮抗植物对K+ 的吸收, 从而限制植物生长(Brugnoli & Lauteri, 1991 ; 王宁等, 2015 ).本研究发现, 盐、碱胁迫均造成亚麻地上部分和根中Na+ 含量升高, K+ 含量降低, 碱胁迫下的变化趋势明显大于盐胁迫(图3A 1 , 3A2 , 3B1 , 3B2 ).结果表明与盐胁迫相比, 碱胁迫的高pH值进一步干扰和破坏了根对Na+ 、K+ 吸收的选择性, 导致体内Na+ 、K+ 严重失衡, 这是碱胁迫危害大于盐胁迫的一个主要原因.另外, 碱胁迫下亚麻地上部分中积累大量Na+ , 致使叶绿体遭到破坏, 叶绿素含量急剧下降, 导致光合能力降低(图2 ).研究还发现, 与根相比, 地上部分中K+ 含量保持在较高水平, 这说明亚麻有一个将Na+ 控制在根部而将更多的K+ 运输到地上部分的控制适应过程(Munns & Tester, 2008 ).Ca2+ 对维持细胞膜的稳定性起着重要作用, Mg2+ 是叶绿素的主要组分, 研究发现某些植物的Ca2+ 、Mg2+ 积累受到盐胁迫抑制.本研究结果表明盐、碱胁迫对亚麻地上部分中Ca2+ 、Mg2+ 含量影响不大, 但碱胁迫明显降低其在根中的含量(图3C 1 , 3C2 , 3D1 , 3D2 ).Ca2+ 作为第二信使与盐超敏感性蛋白(SOS3)结合, 刺激SOS2的激酶活性, 排出有毒离子Na+ (Zhu, 2003 ).但是碱胁迫(高pH值)导致根周围Ca2+ 沉淀, Ca2+ 吸收明显受阻, 进而影响Na+ 外排, 加速了离子毒害作用.另外, Na+ /H+ 逆向转运蛋白依靠细胞膜两侧质子梯度排出Na+ , 高pH值导致质子亏缺, 质子梯度难于建立, Na+ 不能外排, 导致植物体内积累大量的Na+ (Tester & Davenport, 2003 ; 王宁等, 2015 ).植物体维持细胞内离子平衡是保障各种代谢过程正常进行的必要条件, 离子平衡实际上就是阴阳离子平衡的过程(杨春武等, 2008 ).植物吸收阳离子除了特异性离子通道外, 还有一类非选择性阳离子通道(Munns & Tester, 2008 ), 盐碱胁迫下Na+ 大量进入根部, 可能直接抑制根对其他营养离子的吸收, 导致植物体内离子比率失衡, 如图3 中可见盐、碱胁迫明显降低亚麻K+ /Na+ 、Ca2+ /Na+ 和Mg2+ /Na+ , 并且碱胁迫下的下降幅度均大于盐胁迫.研究还发现, 盐、碱胁迫造成NO3 - 含量降低, 表明大量Na+ 积累阻碍根对NO3 - 的吸收, 明显干扰氮吸收或代谢.碱胁迫强烈干扰无机阴离子积累和分布, 碱胁迫(高pH值)阻碍质子梯度的建立, 从而抑制亚麻根系对阴离子的吸收, 造成植物营养缺乏, 这可能是碱胁迫对植物伤害甚于盐胁迫的另一个重要原因. ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}