
不同菌根类型树种的根系分泌物特征及其根际效应研究进展
收稿日期: 2024-06-12
录用日期: 2024-12-10
网络出版日期: 2024-12-10
基金资助
国家自然科学基金(32192433);国家自然科学基金(32071743);福建省科技厅公益类项目(2024R1002006)
Research progress on root exudates and rhizosphere effects of tree species associated with different mycorrhizal types
Received date: 2024-06-12
Accepted date: 2024-12-10
Online published: 2024-12-10
Supported by
National Natural Science Foundation of China(32192433);National Natural Science Foundation of China(32071743);Public Welfare Projects of Science and Technology Department of Fujian Province(2024R1002006)
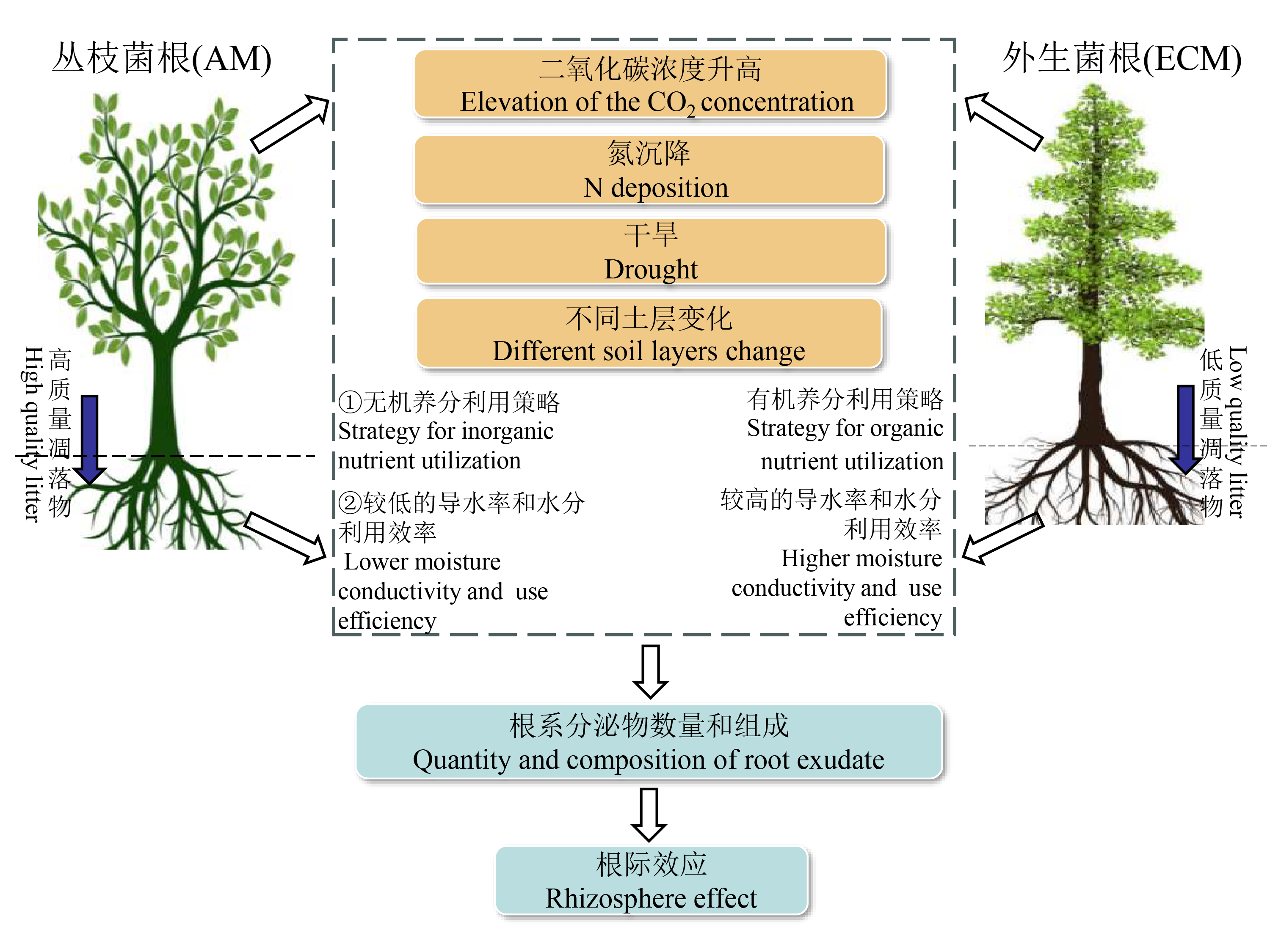
全球变化背景下, 地下生态过程的变化已成为生态学领域关注的热点问题。菌根是植物根系与菌根真菌形成的共生体。与树木相关的两种菌根真菌——丛枝菌根和外生菌根真菌广泛分布于陆地生态系统中, 且在形态和功能上存在显著差异。根系分泌物作为植物与土壤进行物质能量交换和信息传递的重要媒介, 在土壤碳动态变化中发挥着重要作用。不同菌根类型树种能够通过根系分泌物不断地调整有机质输入量和化学组成来积极响应环境变化, 影响森林生态系统养分动态及循环过程。而目前对不同菌根类型树种根系分泌物的组成和功能、变化规律及对植物与土壤的影响机制尚未明确。为此, 该文围绕目前该领域国内外的前沿动态, 针对不同菌根类型树种的根系分泌物特征、影响机制及其根际效应进行了综述, 以期为不同菌根类型树种的根系分泌物对全球变化的响应和适应机制研究提供参考。并提出了未来需要深入研究的方向: (1)加强对不同菌根类型树种根系分泌物的系统性研究; (2)加强菌根类型与其他环境因子耦合对根系分泌物影响机制的研究; (3)利用更加精准的技术手段, 全面深入地了解不同菌根类型树种根系分泌物特征的变化; (4)从植物生理和代谢角度深入揭示不同菌根类型树种根系分泌物的影响机理; (5)对不同菌根类型树种开展长期动态监测和模拟实验, 预测其根系分泌物对土壤生态过程的影响。
梁天豪 , 熊德成 , 刘源豪 , 杜旭龙 , 杨智杰 , 黄锦学 . 不同菌根类型树种的根系分泌物特征及其根际效应研究进展[J]. 植物生态学报, 2025 , 49(7) : 1038 -1052 . DOI: 10.17521/cjpe.2024.0196
Changes in belowground ecological processes in the context of global change have become one of the hot issues of concern in the field of ecology. Mycorrhiza are symbiotic relationships between plant roots and mycorrhizal fungi, widely distributed in terrestrial ecosystems. As two major types of mycorrhizal fungi related to trees, arbuscular mycorrhizal fungi and ectomycorrhizal fungi exhibit significant differences in morphology and function. Root exudates, as an important medium for exchange of matter, energy, and information between plants and soil, play a crucial role in soil carbon dynamics. The root exudates of tree species associated with different mycorrhizal types can actively respond to environmental changes by continuously adjusting their quantity and chemical composition, and influence the belowground carbon dynamics and cycling processes of forest ecosystems. Currently, the composition and function of root exudates from different types of mycorrhizal fungi, as well as the variation and impact on plant and soil, remain unclear. Therefore, this article combines current frontiers in the field both domestically and internationally, and summarizes the root exudate characteristics, influencing mechanisms, and rhizosphere effects of tree species associated with different mycorrhizal types. This review is expected to provide references for further research on the response and adaptation mechanism of root systems and exudates to global changes. In addition, it also proposes directions for future research on root exudates among different types of mycorrhizal fungi that require further investigation: (1) strengthening systematic research on root exudates among different types of mycorrhizal fungi; (2) studying the influence mechanism of mycorrhizal type on root exudates in combination with other environmental factors; (3) using more precise technological means to comprehensively understand the changes in root exudate characteristics among different types of mycorrhizal fungi; (4) deeply revealing the influencing mechanism of root exudates among different types of mycorrhizal fungi from the perspective of plant physiology and metabolism; (5) conducting long-term dynamic monitoring and simulation experiments on different types of mycorrhizal fungi to predict their impact on soil ecological processes.
| [1] | Anderson IC, Cairney JW (2007). Ectomycorrhizal fungi: exploring the mycelial frontier. FEMS Microbiology Reviews, 31, 388-406. |
| [2] | Asmar F, Eiland F, Nielsen NE (1994). Effect of extracellular enzyme activities on solubilization rate of soil organic nitrogen. Biology and Fertility of Soils, 17, 32-38. |
| [3] | Averill C, Turner BL, Finzi AC (2014). Mycorrhiza-mediated competition between plants and decomposers drives soil carbon storage. Nature, 505, 543-545. |
| [4] | Barceló M, van Bodegom PM, Tedersoo L, Olsson PA, Soudzilovskaia NA (2022). Mycorrhizal tree impacts on topsoil biogeochemical properties in tropical forests. Journal of Ecology, 110, 1271-1282. |
| [5] | Bastida F, Eldridge DJ, García C, Kenny Png G, Bardgett RD, Delgado-Baquerizo M (2021). Soil microbial diversity-biomass relationships are driven by soil carbon content across global biomes. The ISME Journal, 15, 2081-2091. |
| [6] | Brunner I, Herzog C, Dawes MA, Arend M, Sperisen C (2015). How tree roots respond to drought. Frontiers in Plant Science, 6, 547. DOI: 10.3389/fpls.2015.00547. |
| [7] | Brzostek ER, Dragoni D, Brown ZA, Phillips RP (2015). Mycorrhizal type determines the magnitude and direction of root-induced changes in decomposition in a temperate forest. New Phytologist, 206, 1274-1282. |
| [8] | Brzostek ER, Greco A, Drake JE, Finzi AC (2013). Root carbon inputs to the rhizosphere stimulate extracellular enzyme activity and increase nitrogen availability in temperate forest soils. Biogeochemistry, 115, 65-76. |
| [9] | Cao YT, Yu SQ, Shao HM, Tan R, Xu XY, Wang WF (2023). Effects of different mycorrhizal types in dominant activity. Acta Ecologica Sinica, 43, 1971-1980. |
| [曹雨婷, 于水强, 邵慧妹, 谭蕊, 徐新颖, 王维枫 (2023). 不同优势树种菌根类型差异对土壤胞外酶活性的影响. 生态学报, 43, 1971-1980.] | |
| [10] | Cheeke TE, Phillips RP, Brzostek ER, Rosling A, Bever JD, Fransson P (2017). Dominant mycorrhizal association of trees alters carbon and nutrient cycling by selecting for microbial groups with distinct enzyme function. New Phytologist, 214, 432-442. |
| [11] | Chen L, Cai YX, Lei HM, Qi XX, Lin JJ, Liao W, Huang ZX (2022). Comparison of soil nitrogen cycling between arbuscular mycorrhizal and ectomycorrhizal forests in a sub-tropical region. Chinese Journal of Ecology, 41, 218-226. |
| [陈亮, 蔡咏欣, 雷惠敏, 齐晓旭, 林俊均, 廖伟, 黄子玄 (2022). 亚热带丛枝菌根与外生菌根森林对土壤氮循环的影响. 生态学杂志, 41, 218-226.] | |
| [12] | Chen X, Ding ZJ, Tang M, Zhu B (2018). Greater variations of rhizosphere effects within mycorrhizal group than between mycorrhizal group in a temperate forest. Soil Biology & Biochemistry, 126, 237-246. |
| [13] | Cheng W, Parton WJ, Gonzalez-Meler MA, Phillips R, Asao S, McNickle GG, Brzostek E, Jastrow JD (2014). Synthesis and modeling perspectives of rhizosphere priming. New Phytologist, 201, 31-44. |
| [14] | Craig ME, Turner BL, Liang C, Clay K, Johnson DJ, Phillips RP (2018). Tree mycorrhizal type predicts within-site variability in the storage and distribution of soil organic matter. Global Change Biology, 24, 3317-3330. |
| [15] | Crim PM, Cumming JR (2020). Extracellular soil enzyme activities in high-elevation mixed red spruce forests in central Appalachia, USA. Forests, 11, 468. DOI: 10.3390/f11040468. |
| [16] | de Vries FT, Williams A, Stringer F, Willcocks R, McEwing R, Langridge H, Straathof AL (2019). Changes in root-exudate-induced respiration reveal a novel mechanism through which drought affects ecosystem carbon cycling. New Phytologist, 224, 132-145. |
| [17] | Fahey C, Winter K, Slot M, Kitajima K (2016). Influence of arbuscular mycorrhizal colonization on whole-plant respiration and thermal acclimation of tropical tree seedlings. Ecology and Evolution, 6, 859-870. |
| [18] | Gargallo-Garriga A, Preece C, Sardans J, Oravec M, Urban O, Pe?uelas J (2018). Root exudate metabolomes change under drought and show limited capacity for recovery. Scientific Reports, 8, 12696. DOI: 10.1038/s41598-018-30150-0. |
| [19] | Han MG, Sun LJ, Gan DY, Fu LC, Zhu B (2020). Root functional traits are key determinants of the rhizosphere effect on soil organic matter decomposition across 14 temperate hardwood species. Soil Biology & Biochemistry, 151, 108019. DOI: 10.1016/j.soilbio.2020.108019. |
| [20] | Hodge A, Helgason T, Fitter AH (2010). Nutritional ecology of arbuscular mycorrhizal fungi. Fungal Ecology, 3, 267-273. |
| [21] | Holz M, Zarebanadkouki M, Kuzyakov Y, Pausch J, Carminati A (2018). Root hairs increase rhizosphere extension and carbon input to soil. Annals of Botany, 121, 61-69. |
| [22] | Iversen CM, Keller JK, Garten Jr CT, Norby RJ (2012). Soil carbon and nitrogen cycling and storage throughout the soil profile in a sweetgum plantation after 11 years of CO2-enrichment. Global Change Biology, 18, 1684-1697. |
| [23] | Jiang Z, Fu YL, Zhou LY, He YH, Zhou GY, Dietrich P, Long JL, Wang XX, Jia SX, Ji YH, Jia Z, Song BQ, Liu RQ, Zhou XH (2023). Plant growth strategy determines the magnitude and direction of drought-induced changes in root exudates in subtropical forests. Global Change Biology, 31. 3476-3488. |
| [24] | Jiang Z, Thakur MP, Liu R, Zhou G, Zhou L, Fu Y, Zhang P, He Y, Shao J, Gao J, Li N, Wang X, Jia X, Chen Y, Zhang C, Zhou X (2022). Soil P availability and mycorrhizal type determine root exudation in subtropical forests. Soil Biology & Biochemistry, 171, 108722. DOI: 10.1016/j.soilbio.2022.108722. |
| [25] | Jiang ZY, Zou QQ, Yang L, Li WZ, Zhang HD, Chen XW, Wang XW (2021). Root exudation rate and rhizosphere effect of different mycorrhizal associations of tree species in typical black soil area. Chinese Journal of Ecology, 40, 2709-2718. |
| [蒋治岩, 邹青勤, 杨柳, 李汶倬, 张鹤东, 陈祥伟, 王秀伟 (2021). 典型黑土区不同菌根类型树种根系分泌速率及根际效应差异. 生态学杂志, 40, 2709-2718.] | |
| [26] | Karst J, Gaster J, Wiley E, Landh?usser SM (2017). Stress differentially causes roots of tree seedlings to exude carbon. Tree Physiology, 37, 154-164. |
| [27] | Kivlin SN, Emery SM, Rudgers JA (2013). Fungal symbionts alter plant responses to global change. American Journal of Botany, 100, 1445-1457. |
| [28] | Klein T, Siegwolf RTW, K?rner C (2016). Belowground carbon trade among tall trees in a temperate forest. Science, 352, 342-344. |
| [29] | Kuyper TW (2012). Ectomycorrhiza and the open nitrogen cycle in an afrotropical rainforest. New Phytologist, 195, 728-729. |
| [30] | Laliberté E (2017). Below-ground frontiers in trait-based plant ecology. New Phytologist, 213, 1597-1603. |
| [31] | Latef AAHA, Hashem A, Rasool S, Abd_Allah EF, Alqarawi AA, Egamberdieva D, Jan S, Anjum NA, Ahmad P (2016). Arbuscular mycorrhizal symbiosis and abiotic stress in plants: a review. Journal of Plant Biology, 59, 407-426. |
| [32] | Li F, Hao ZP, Chen BD (2019). Molecular mechanism for the adaption of arbuscular mycorrhizal symbiosis to phosphorus deficiency. Journal of Plant Nutrition and Fertilizers, 25, 1989-1997. |
| [李芳, 郝志鹏, 陈保冬 (2019). 菌根植物适应低磷胁迫的分子机制. 植物营养与肥料学报, 25, 1989-1997.] | |
| [33] | Li J, Xia SG, Shi XY, Sun QY, Zhao Q (2023). Soil acid buffering capacity of subtropical arbuscular and ectomycorrhizal forests and its main influencing. Chinese Journal of Ecology, 21, 3333-3340. |
| [李静, 夏尚光, 石晓芸, 孙庆业, 赵琼 (2024). 亚热带丛枝和外生菌根森林土壤酸缓冲性能及其主要影响因素. 生态学杂志, 21, 3333-3340.] | |
| [34] | Liese R, Alings K, Meier IC (2017). Root branching is a leading root trait of the plant economics spectrum in temperate trees. Frontiers in Plant Science, 8, 315. DOI: 10.3389/fpls.2017.00315. |
| [35] | Liese R, Lübbe T, Albers NW, Meier IC (2018). The mycorrhizal type governs root exudation and nitrogen uptake of temperate tree species. Tree Physiology, 38, 83-95. |
| [36] | Lin G, Craig ME, Jo I, Wang X, Zeng DH, Phillips RP (2022). Mycorrhizal associations of tree species influence soil nitrogen dynamics via effects on soil acid-base chemistry. Global Ecology and Biogeography, 31, 168-182. |
| [37] | Lin G, Guo D, Li L, Ma CG, Zeng DH (2018). Contrasting effects of ectomycorrhizal and arbuscular mycorrhizal tropical tree species on soil nitrogen cycling: the potential mechanisms and corresponding adaptive strategies. Oikos, 127, 518-530. |
| [38] | Lin G, McCormack ML, Ma C, Guo D (2017). Similar below-ground carbon cycling dynamics but contrasting modes of nitrogen cycling between arbuscular mycorrhizal and ectomycorrhizal forests. New Phytologist, 213, 1440-1451. |
| [39] | Lu XK, Mo JM, Zhang W, Mao QG, Liu RZ, Wang C, Wang SH, Zheng MH, Taiki M, Mao JH, Zhang YQ, Wang YF, Hang J (2019). Effects of simulated atmospheric nitrogen deposition on forest ecosystems in China: an overview. Journal of Tropical and Subtropical Botany, 27, 500-522. |
| [鲁显楷, 莫江明, 张炜, 毛庆功, 刘荣臻, 王聪, 王森浩, 郑棉海, Taiki M, 毛晋花, 张勇群, 王玉芳, 黄娟 (2022). 模拟大气氮沉降对中国森林生态系统影响的研究进展. 热带亚热带植物学报, 27, 500-522.] | |
| [40] | Martinière A, Gibrat R, Sentenac H, Dumont X, Gaillard I, Paris N (2018). Uncovering pH at both sides of the root plasma membrane interface using noninvasive imaging. Proceedings of the National Academy of Sciences of the United States of America, 115, 6488-6493. |
| [41] | McCormack ML, Dickie IA, Eissenstat DM, Fahey TJ, Fernandez CW, Guo D, Helmisaari HS, Hobbie EA, Iversen CM, Jackson RB, Lepp?lammi-Kujansuu J, Norby RJ, Phillips RP, Pregitzer KS, Pritchard SG, et al. (2015). Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processes. New Phytologist, 207, 505-518. |
| [42] | Midgley MG, Phillips RP (2016). Resource stoichiometry and the biogeochemical consequences of nitrogen deposition in a mixed deciduous forest. Ecology, 97, 3369-3378. |
| [43] | Mohan JE, Cowden CC, Baas P, Dawadi A, Frankson PT, Helmick K, Hughes E, Khan S, Lang A, Machmuller M, Taylor M, Witt CA (2014). Mycorrhizal fungi mediation of terrestrial ecosystem responses to global change: mini-review. Fungal Ecology, 10, 3-19. |
| [44] | Oburger E, Jones DL (2018). Sampling root exudates-mission impossible? Rhizosphere, 6, 116-133. |
| [45] | Phillips R, Brzostek ER, Midgley M, Keller AB (2016). Mycorrhizal associations as trait integrators for biogeochemical syndromes in forests//101st ESA Annual Meeting (August 7-12, 2016). ESA, Washington D.C. |
| [46] | Phillips RP, Brzostek E, Midgley MG (2013). The mycorrhizal-associated nutrient economy: a new framework for predicting carbon-nutrient couplings in temperate forests. New Phytologist, 199, 41-51. |
| [47] | Phillips RP, Fahey TJ (2006). Tree species and mycorrhizal associations influence the magnitude of rhizosphere effects. Ecology, 87, 1302-1313. |
| [48] | Phillips RP, Fahey TJ, Burton AJ (2008). Monitoring fine root growth and physiology in a mixed hardwood forest: implications for nitrogen deposition responses. Tree Physiology, 28, 1075-1085. |
| [49] | Phillips RP, Finzi AC, Bernhardt ES (2011). Enhanced root exudation induces microbial feedbacks to N cycling in a pine forest under long-term CO2fumigation. Ecology Letters, 14, 187-194. |
| [50] | Qi XX, Chen L, Zhu JA, Li Z, Lei HM, Shen Q, Wu HL, Quyang S, Zeng YL, H YT, Xiang WH (2022). Increase of soil phosphorus bioavailability with ectomycorrhizal tree dominance in subtropical secondary forests. Forest Ecology and Management, 521, 120435. DOI: 10.1016/j.foreco.2022.120435. |
| [51] | Qiang W, Gunina A, Kuzyakov Y, He L, Zhang Y, Liu B, Pang X (2023). Contributions of mycorrhizal fungi to soil aggregate formation during subalpine forest succession. Catena, 221, 106800. DOI: 10.1016/j.catena.2022.106800. |
| [52] | Querejeta JI, Egerton-Warburton LM, Allen MF (2007). Hydraulic lift may buffer rhizosphere hyphae against the negative effects of severe soil drying in a California Oak savanna. Soil Biology & Biochemistry, 39, 409-417. |
| [53] | Read DJ, Perez-Moreno J (2003). Mycorrhizas and nutrient cycling in ecosystems—A journey towards relevance? New Phytologist, 157, 475-492. |
| [54] | Rewald B, Rechenmacher A, Godbold DL (2014). It’s complicated: intraroot system variability of respiration and morphological traits in four deciduous tree species. Plant Physiology, 166, 736-745. |
| [55] | Rosling A, Midgley MG, Cheeke T, Urbina H, Fransson P, Phillips RP (2016). Phosphorus cycling in deciduous forest soil differs between stands dominated by ecto- and arbuscular mycorrhizal trees. New Phytologist, 209, 1184-1195. |
| [56] | Sawyer NA, Chambers SM, Cairney JWG (2003). Distribution of Amanita spp. genotypes under eastern Australian sclerophyll vegetation. Mycological Research, 107, 1157-1162. |
| [57] | Sinsabaugh RL (2010). Phenol oxidase, peroxidase and organic matter dynamics of soil. Soil Biology & Biochemistry, 42, 391-404. |
| [58] | Song FQ, Yang GT, Meng FR, Tian XJ, Dong AR (2004). The rhizospheric niche of seedlings of Populus ussruiensis colonized by arbuscular mycorrhizal (AM) fungi. Ecology and Environmental Sciences, 13, 211-216. |
| [宋福强, 杨国亭, 孟繁荣, 田兴军, 董爱荣 (2004). 丛枝菌根化大青杨苗木根际微域环境的研究. 生态环境, 13, 211-216.] | |
| [59] | Soudzilovskaia NA, van der Heijden MGA, Cornelissen JHC, Makarov MI, Onipchenko VG, Maslov MN, Akhmetzhanova AA, van Bodegom PM (2015). Quantitative assessment of the differential impacts of arbuscular and ectomycorrhiza on soil carbon cycling. New Phytologist, 208, 280-293. |
| [60] | Su YJ, Yang K, Zhang Q, Xu S, Yu LZ, Zhang JX (2024). Research progress on soil nitrogen and phosphorus availability characteristics of different mycorrhizal tree species. Chinese Journal of Ecology, 6, 1-17. |
| [苏颖佳, 杨凯, 张乾, 徐爽, 于立忠, 张金鑫 (2024). 不同菌根类型树种土壤氮、磷有效性特征及影响因素研究进展. 生态学杂志, 6, 1-17.] | |
| [61] | Sulman BN, Brzostek ER, Medici C, Shevliakova E, Menge D NL, Phillips RP (2017). Feedbacks between plant N demand and rhizosphere priming depend on type of mycorrhizal association. Ecology Letters, 20, 1043-1053. |
| [62] | Sun L, Ataka M, Han M, Han Y, Gan D, Xu T, Guo Y, Zhu B (2021). Root exudation as a major competitive fine-root functional trait of 18 coexisting species in a subtropical forest. New Phytologist, 229, 259-271. |
| [63] | Sun L, Kominami Y, Yoshimura K, Kitayama K (2017). Root-exudate flux variations among four co-existing canopy species in a temperate forest, Japan. Ecological Research, 32, 331-339. |
| [64] | Sun Y (2022). Comparative Study on Nutrient and Enzyme Activity in Rhizosphere Soil of Different Tree Species in Subtropical Zone. Master degree dissertation, Central South University of Forestry and Technology, Changsha. |
| [孙韵 (2022). 亚热带不同树种根际土壤养分及酶活性比较研究. 硕士学位论文, 中南林业科技大学, 长沙.] | |
| [65] | Talhelm AF, Pregitzer KS, Kubiske ME, Zak DR, Campany CE, Burton AJ, Dickson RE, Hendrey GR, Isebrands JG, Lewin KF, Nagy J, Karnosky DF (2014). Elevated carbon dioxide and ozone alter productivity and ecosystem carbon content in northern temperate forests. Global Change Biology, 20, 2492-2504. |
| [66] | Tan B, Li Y, Liu T, Tan X, He Y, You X, Leong KH, Liu C, Li L (2021). Response of plant rhizosphere microenvironment to water management in soil- and substrate-based controlled environment agriculture (CEA) systems: a review. Frontiers in Plant Science, 12, 691651. DOI: 10.3389/fpls.2021.691651. |
| [67] | Tekaya M, Dabbaghi O, Guesmi A, Attia F, Chehab H, Khezami L, Algathami FK, Hamadi NB, Hammami M, Prinsen E, Mechri B (2022). Arbuscular mycorrhizas modulate carbohydrate, phenolic compounds and hormonal metabolism to enhance water deficit tolerance of olive trees (Olea europaea). Agricultural Water Management, 274, 107947. DOI: 10.1016/j.agwat.2022.107947. |
| [68] | Terrer C, Vicca S, Hungate BA, Phillips RP, Prentice IC (2016). Mycorrhizal association as a primary control of the CO2 fertilization effect. Science, 353, 72-74. |
| [69] | Walker TS, Bais HP, Grotewold E, Vivanco JM (2003). Root exudation and rhizosphere biology. Plant Physiology, 132, 44-51. |
| [70] | Wallander H, Ekblad A, Godbold DL, Johnson D, Bahr A, Baldrian P, Bj?rk RG, Kieliszewska-Rokicka B, Kj?ller R, Kraigher H, Plassard C, Rudawska M (2013). Evaluation of methods to estimate production, biomass and turnover of ectomycorrhizal mycelium in forests soils—A review. Soil Biology & Biochemistry, 57, 1034-1047. |
| [71] | Wang QH, Gong YB, Zhang J (2006). Impact of forest ecosystems on atmospheric nitrogen deposition. Journal of Sichuan Forestry Science and Technology, 27(1), 19-25. |
| [王巧红, 宫渊波, 张君 (2006). 森林生态系统对大气氮沉降的响应. 四川林业科技, 27(1), 19-25.] | |
| [72] | Wang XQ, Wang CK (2019). Variations in topsoil carbon and nitrogen contents of five temperate plantations in Northeast China. Chinese Journal of Applied Ecology, 30, 1911-1918. |
| [王薪琪, 王传宽 (2019). 东北5种温带人工林表层土壤碳氮含量的分异. 应用生态学报, 30, 1911-1918.] | |
| [73] | Williams A, de Vries FT (2020). Plant root exudation under drought: implications for ecosystem functioning. New Phytologist, 225, 1899-1905. |
| [74] | Wu LK, Lin XM, Lin WX (2014). Advances and perspective in research on plant-soil-microbe interactions mediated by root exudates. Chinese Journal of Plant Ecology, 38, 298-310. |
| [吴林坤, 林向民, 林文雄 (2014). 根系分泌物介导下植物-土壤-微生物互作关系研究进展与展望. 植物生态学报, 38, 298-310.] | |
| [75] | Wurzburger N, Brookshrie ENJ (2017). Experimental evidence that mycorrhizal nitrogen strategies affect soil carbon. Ecology, 98, 1491-1497. |
| [76] | Yang K, Zhang Q, Zhu JJ, Wang QQ, Gao T, Wang GG (2023). Mycorrhizal type regulates trade-offs between plant and soil carbon in forests. Nature Climate Change, 14, 91-97. |
| [77] | Yang Y, Zhang X, Hartley IP, Dungait JAJ, Wen X, Li D, Guo Z, Quine TA (2022). Contrasting rhizosphere soil nutrient economy of plants associated with arbuscular mycorrhizal and ectomycorrhizal fungi in karst forests. Plant and Soil, 470, 81-93. |
| [78] | Yin HJ, Li YF, Xiao J, Xu ZF, Cheng XY, Liu Q (2013). Enhanced root exudation stimulates soil nitrogen transformations in a subalpine coniferous forest under experimental warming. Global Change Biology, 19, 2158-2167. |
| [79] | Yin HJ, Wheeler E, Phillips RP (2014). Root-induced changes in nutrient cycling in forests depend on exudation rates. Soil Biology & Biochemistry, 78, 213-221. |
| [80] | Yin HJ, Zhang ZL, Liu Q (2018). Root exudates and their ecological consequences in forest ecosystems: problems and perspective. Chinese Journal of Plant Ecology, 42, 1055-1070. |
| [尹华军, 张子良, 刘庆 (2018). 森林根系分泌物生态学研究: 问题与展望. 植物生态学报, 42, 1055-1070.] |
/
| 〈 |
|
〉 |
Copyright © 2026 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19
![]()
