|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
植物吸收根的增殖和生长与养分变异的关系——臭椿、翠菊、加拿大一枝黄花分根实验的启示
植物生态学报
2013, 37 (2):
93-103.
DOI: 10.3724/SP.J.1258.2013.00010
构件理论认为植物根可以相对独立地吸收养分和对所处环境的养分条件做出响应。根据成本-收益理论, 单个根(构件)的生死、生长发育与其吸收的养分收益和自身建造、维持的消耗有关。基于此, 该文提出两个关于吸收根生死条件的假设: 1)当可利用养分低于低临界值, 根死亡在一段时滞(数天到几周)后发生; 2)当可利用养分高于高临界值并持续一段时间, 新的侧根产生。为了检验这两个假设, 用臭椿(Ailanthus altissima)、翠菊(Callistephus chinensis)、加拿大一枝黄花(Solidago canadensis)作实验物种, 设计了温室分根实验。每株植物选3个一级根, 分别引入3个不同养分水平的斑块: 0、20、200 μg N·g-1。每4天将根暴露并拍照, 查数新根数并测量细根总长度和一级侧根长。由于高养分处理斑块内根的快速生长, 实验在开始后8天或12天结束。结果显示: 除臭椿在0养分处理外, 三物种在各养分处理下都有侧根产生, 总根长均有增加; 臭椿、翠菊、加拿大一枝黄花在不同观测时间和养分水平处理间的侧根数目和总根长差异显著, 而一级侧根长除臭椿外变异均较小; 整个过程中没有根死亡。研究结果部分支持两个假设。本研究还为进一步探究根模块构件增殖、生死过程机制提出新的建议, 即除需要更长的实验时间外, 还应该考虑: 1)多种资源各自及联合对根生长、生死过程的影响; 2)资源斑块和整个根系生长背景的资源丰度对比; 3)根构建和根维持的相对C消耗。
表2
臭椿、翠菊、加拿大一枝黄花侧根数、一级侧根长和总根长的重复测量方差分析结果
正文中引用本图/表的段落
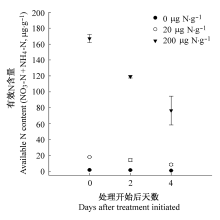
处理开始时, 引入养分斑块的根均为一级根, 且根长在不同物种、养分水平间没有显著差异(p > 0.05)。处理后, 三物种在三个养分水平下的侧根产生和根长生长程度很不相同。2.2.1 臭椿侧根数目、一级侧根长与总根长3次观测间臭椿的侧根数、一级侧根长和总根长差异显著(p < 0.001, 表2), 3次测量两两间(即第4与第8天, 第8与第12天间)的侧根数目、一级侧根长和总根长差异显著(p < 0.001, 表2)。3次观测的侧根数、一级侧根长和总根长在不同养分水平间也有显著差异(p < 0.001); 高养分处理的侧根数和总根长的平均值显著高于低养分(p < 0.001)和中养分(p < 0.05)处理, 而在中低养分处理之间差异不显著, 一级侧根长则在3个养分水平间均差异显著(p < 0.05)。养分水平与3次观测对侧根数、一级侧根长和总根长有显著交互作用(p < 0.001, 表2), 即3个养分水平下生长速率不同。
翠菊侧根数、一级侧根长和总根长在两次观测之间差异显著(p < 0.01, 表2)。养分处理对侧根数和根长影响显著(p < 0.05), 而对一级侧根长影响近显著(p < 0.087, 表2); 侧根数两次测量的平均值在低养分处理下显著低于中养分(p < 0.05)和高养分(p < 0.001)处理, 而在中、高养分处理间没有显著差异; 根长两次测量的平均值在低养分下显著低于高养分处理(p < 0.01), 但在中低养分间和中高养分间均差异不显著。养分水平与不同观测时间对侧根数和根长有显著交互作用(p < 0.01), 而对一级侧根长无显著影响(p < 0.1, 表2)。不同养分处理下侧根数与根长的两次观测值间差异显著(p < 0.05), 且在3个养分处理中格局一致, 即侧根数与根长在3个养分处理中第8天均显著大于第4天(p < 0.05, 图2)。
加拿大一枝黄花的侧根数与根长在两次观测间均差异显著(p < 0.001), 而对一级侧根长无显著影响(p = 0.55, 表2)。养分处理对侧根数和根长影响显著(p < 0.01), 对一级侧根长影响近显著 (p < 0.08, 表2); 高养分处理下侧根数与根长两次测量均值显著高于中养分(p < 0.05)与低养分(p < 0.001)处理, 而二者在中、低养分间没有显著差异。养分水平与不同观测时间对侧根数和根长有显著交互作用(p < 0.001), 而对一级侧根长无显著影响(p = 0.4, 表2)。不同养分处理下, 两次观测的侧根数间差异与根长之间差异均显著(p < 0.001), 且格局一致, 即侧根数与根长在低、中、高养分处理中均是第8天显著大于第4天(p < 0.001, 图2)。
一级侧根长平均值的变化格局与总根长和侧根数的变化格局不同, 表现在翠菊和加拿大一枝黄花对养分处理的低敏感性以及时间上的低变异性(图2)。这是由于一级侧根长的高变异性所致, 即新萌生的一级根很短, 而生长了一段时间的一级根则较长, 这也反映在一级侧根长平均值的较大标准误上(图2)。似乎一级侧根长的平均值对根生死过程的指示作用不是很明显, 也许平均值在这里并不是一个好的指标。
本实验结果还表明不同物种间对养分响应有显著差异, 即有效N浓度对不同物种的新根增殖和根生长影响不同, 这在本研究中得到充分显示(图2)。实验物种对低N浓度的耐受程度不同: 加拿大一枝黄花>翠菊>臭椿。三种氮水平处理对加拿大一枝黄花的侧根增殖和根长生长的影响似乎仅仅是浓度的影响(图2)。而翠菊的侧根生长和根长生长在高氮肥处理(200 μg N·g-1)初期(即第4天)受抑制可能与高N毒害有关(董佳和牟溥, 2012)。臭椿细根的生长速度缓慢也有可能与其物种的生物学特性有关(Hunt & Cornelissen, 1997; Einsmann et al., 1999)。
本文的其它图/表
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||