
|
|
||
|
高山垫状植物团状福禄草开花面积与方位随海拔的变化及其适应性
植物生态学报
2020, 44 (11):
1154-1163.
DOI: 10.17521/cjpe.2020.0124
作为高山生态系统中的奠基种(foundation species), 垫状植物自身种群的繁殖与扩张, 对高山生态系统功能稳定性起着关键作用。但是, 垫状植物如何在极端环境条件下实现资源的有效利用与分配, 达到繁殖最优化, 至今鲜为人知。该研究在滇西北白马雪山沿海拔梯度选择具有不同坡度及坡向的5个团状福禄草(Arenaria polytrichoides)种群, 调查、比较种群内、种群间以及具有不同性系统的植株个体之间的开花面积比、开花方位, 并分析不同生态因子对其开花特性的影响。结果表明: 随着海拔的升高, 团状福禄草个体变小, 其分配到开花的资源比例总体上随海拔上升呈现下降的趋势, 说明团状福禄草的繁殖分配受到由海拔所引起的生态因子的调控。但是, 部分低海拔种群内植物个体的繁殖分配显著低于部分高海拔种群, 说明海拔并非控制植物繁殖分配的唯一因素。此外, 植株开花总面积随植株个体增大而增加, 但开花面积比却随个体增大而变小, 说明植株分配到开花的资源增长速率可能低于植株个体的增长速率。在性别差异方面, 两性植株对开花的资源分配比例要显著高于雌性植株, 但是, 其差异程度受到海拔因素的影响。最后, 在同一种群内, 团状福禄草在冠层表面不同方位上的开花面积比存在显著差异性, 这种差异性在不同种群之间又具有不同的表现形式。 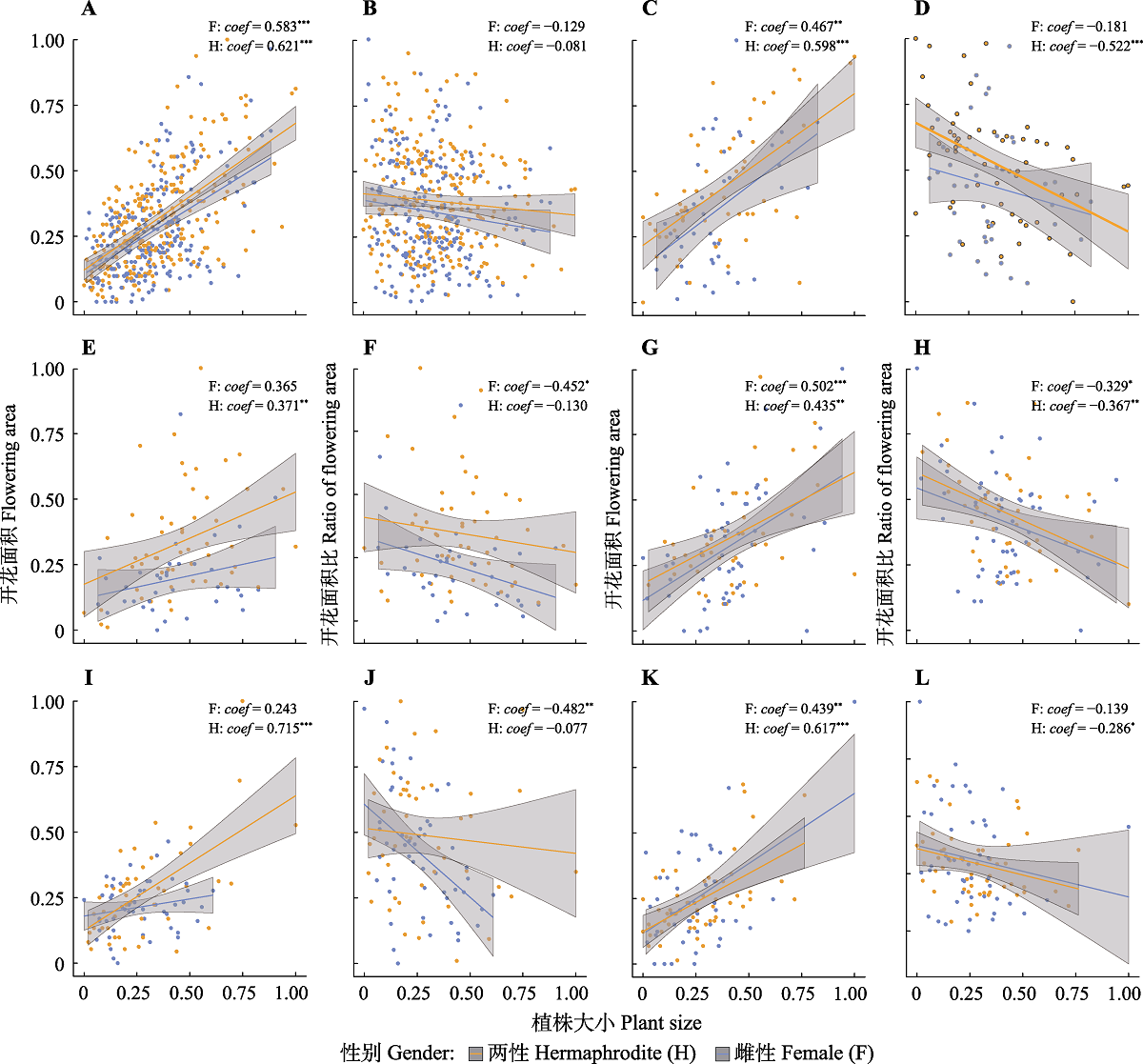 View image in article
图3
团状福禄草植株个体大小与其总开花面积及其面积比的关系。A、B, 所有种群。C、D, 种群1。E、F, 种群2。G、H, 种群3。I、J, 种群4。K、L, 种群5。回归线: 普通最小二乘线性回归线; Coef, 回归系数。所有数据被标准化至0-1之间。相关显著性: *, p ≤ 0.05; **, p ≤ 0.01; ***, p ≤ 0.001。
正文中引用本图/表的段落
首先, 将各方向上开花网格数相加, 得到各植株的总开花网格(面积), 其次, 分别将总开花网格数和每个方向的开花网格数都除以总网格数, 得到各个植株总的开花面积比(为了论述方便, 本研究中, 必要时将开花面积比称为“繁殖分配”)以及单个方位上的开花面积比, 并采用这些数据进行后续的比较分析。不同植株的大小(年龄)会对其资源分配策略产生影响, 即可能存在个体效应(St?cklin & Favre, 1994; Sakai & Sakai, 1995), 同时, 垫状植物冠层表面积大小通常与其年龄呈正相关关系(Chen et al., 2017a), 因此, 计数了每个调查个体的总网格数(即表面积大小), 将其作为植株大小的替代来进行后续分析。首先, 为了探究不同种群以及不同性别植株的总开花面积与其个体大小之间的关系, 对植株总开花面积与其个体大小做了回归分析。其次, 为了探究海拔对植株大小的影响, 采用混合效应模型比较了不同海拔种群间植株的大小, 植株大小为因变量, 海拔为主因素, 植株个体编号为随机因素。最后, 为了探究开花面积比(即繁殖分配)如何受到海拔、性别和植株大小的影响, 使用混合效应模型对上述关系进行拟合。模型中, 开花面积比为因变量, 海拔、性别和植株大小为主因素, 植株个体编号为随机因素。此外, 由于自变量与因变量间存在多样的组合形式, 基于赤池信息量准则(AIC)权重选择最优模型, 最优模型具有最大的AIC权重, 因子在每个模型的AIC权重被用来计算模型平均系数, 而每个因子在所有模型中的AIC值权重之和(w)被用来描述该因子的统计支持。同时, 评估了考虑交互作用的模型与上述模型的优劣, 校准AIC (AICc)结果显示两种模型没有差异(AICc = -402.1 vs. AICc = -401.4), 但是交互作用不显著, 因而以上述的模型结果为准。由于各组数据不完全符合正态分布及方差齐性原则, 使用Kruskal-Wallis非参数检验分别比较同一种群内不同性别、不同方位之间开花面积比以及不同种群之间开花面积比的差异。所有统计分析及画图都通过R 3.5.1 (R Core Team, 2018)实现, Kruskal-Wallis非参数检验使用“agricolae”包(de Felipe, 2019), 混合效应模型使用“coxme”包(Therneau, 2018), 模型选择使用“MuMIn”包(Bartoń, 2019)。
随着海拔升高, 种群内植物个体大小呈现减小的趋势(混合效应模型系数= -0.131, p < 0.001), 个体总开花面积比同样呈现下降趋势(图2), 最小值同时出现在种群2 (次低海拔)和种群5 (最高海拔), 说明种群2除了受到海拔影响外, 可能还受到其他环境因子的强烈影响。植株个体大小与其总开花面积之间呈显著正相关关系(图3A), 说明随着植株年龄增长, 植物分配到开花的资源量也随之增长。但是, 植株越大, 总开花面积比越小(表2; 图3B), 说明植株分配到开花的资源增长速率可能低于植株个体的增长速率。种群1和种群2坡向相同, 坡度相差达30° (表1), 个体总开花面积比差异显著(图2A)。种群3和种群4坡向、坡度均一致, 个体总开花面积比具有显著差异性(高海拔开花总比例低于低海)。此外, 种群5 (最高海拔)具有最低的总开花面积比, 说明海拔对个体繁殖分配具有强烈影响(表2)。
横断山高海拔地区绝大多数植物种群的更新与扩张都依赖有性繁殖(Peng et al., 2012)。植物对繁殖功能的资源分配, 在很大程度上决定了其种群未来的扩张速率。而作为植物实现繁殖功能的重要环节, 植物对开花的资源配置决定了植物的最终繁殖产出(Reekie & Bazzaz, 2005)。不同的环境因子在植物的每一个繁殖阶段都起着重要作用, 甚至一些因子(如资源量)直接决定着植物的繁殖分配及其产出(Delph & Carroll, 2001)。在我们的研究区域内, 环境胁迫程度(如土壤养分胁迫及低温胁迫)随海拔升高而增强(Chen et al., 2015b, 2019)。我们的研究结果显示, 随着海拔升高, 种群内植株个体呈现减小的趋势。植株大小对开花特性具有显著影响。一方面, 植株个体越大, 其总开花面积也越大, 但另一方面, 植株越大, 其总开花面积比越小(表2), 并且这种趋势在不同性别植株间具有高度一致性(图3), 说明资源限制是真实存在的, 即, 植物从环境中获取并分配给开花的资源量有限。此外, 该结果也说明植株分配到开花的资源增长速率可能低于植株个体的增长速率, 即繁殖分配的增加速率低于营养分配的增加速率。相应结果进一步暗示, 在面临资源限制或环境胁迫时, 团状福禄草的资源分配策略以保证植株存活(即增加营养分配)为首要目的。此外, 具有不同坡度及坡向的种群间开花特性也存在一定的变化,说明由坡向及坡度变化而引起的环境因子变化, 如太阳辐射(田辉等, 2007)和土壤养分(曹舰艇等, 2019), 可能也同时对其繁殖分配策略具有一定的影响。
本文的其它图/表
|




{kind=link}