|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
樟树幼苗机械损伤叶片对挥发性有机化合物及叶绿素荧光参数的影响
植物生态学报
2012, 36 (7):
671-680.
DOI: 10.3724/SP.J.1258.2012.00671
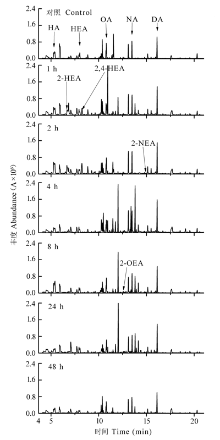
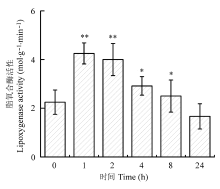
为探讨植物在机械损伤后C6-C10醛类化合物的释放机理, 及C6-C10醛类化合物对叶片光系统II (PSII)的影响, 以樟树(Cinnamomum camphora)为材料, 采用动态顶空气体循环法和热脱附/气相色谱/质谱联用技术(TDS-GC-MS), 对樟树幼苗叶片损伤后释放的C6-C10醛类化合物进行采集与分析, 并同步测定了脂氧合酶活性和损伤叶片的叶绿素荧光动力学参数。结果表明: 樟树幼苗叶片损伤后, 其挥发性有机化合物中己醛、庚醛、辛醛、壬醛和癸醛的释放量比损伤前分别增加了2.47、0.99、1.34、0.91和28.38倍(p < 0.01); 同时新增4种醛类化合物, 分别是: 2-己烯醛、2,4-己二烯醛、(E)-2-辛烯醛和(E)-2-壬烯醛。脂氧合酶活性比损伤前增加1.2倍(p < 0.01)。PSII单位反应中心复合体吸收的能量和被核心捕获的能量分别比损伤前下降12.8%和9.8% (p < 0.01)。单位面积反应中心的数量、电子传递量子产额、捕获激子能导致电子传递效率和叶片性能指数分别比损伤前增加23.3%、24.4%、22.6%和82.7% (p < 0.01)。损伤24 h后, 醛类化合物的种类、释放量、脂氧合酶活性及叶片叶绿素荧光动力学参数基本恢复到损伤前水平。说明机械损伤使PSII供体侧受损、脂氧合酶活性升高, 致使C6-C10醛类化合物大量释放, 樟树幼苗通过增加单位面积反应中心的数量来提高光合效率应对胁迫。
表2
机械损伤后樟树幼苗叶片主要叶绿素荧光参数的变化(平均值±标准误差)
正文中引用本图/表的段落
为探讨植物在机械损伤后C6-C10醛类化合物的释放机理, 及C6-C10醛类化合物对叶片光系统II (PSII)的影响, 以樟树(Cinnamomum camphora)为材料, 采用动态顶空气体循环法和热脱附/气相色谱/质谱联用技术(TDS-GC-MS), 对樟树幼苗叶片损伤后释放的C6-C10醛类化合物进行采集与分析, 并同步测定了脂氧合酶活性和损伤叶片的叶绿素荧光动力学参数。结果表明: 樟树幼苗叶片损伤后, 其挥发性有机化合物中己醛、庚醛、辛醛、壬醛和癸醛的释放量比损伤前分别增加了2.47、0.99、1.34、0.91和28.38倍(p < 0.01); 同时新增4种醛类化合物, 分别是: 2-己烯醛、2,4-己二烯醛、(E)-2-辛烯醛和(E)-2-壬烯醛。脂氧合酶活性比损伤前增加1.2倍(p < 0.01)。PSII单位反应中心复合体吸收的能量和被核心捕获的能量分别比损伤前下降12.8%和9.8% (p < 0.01)。单位面积反应中心的数量、电子传递量子产额、捕获激子能导致电子传递效率和叶片性能指数分别比损伤前增加23.3%、24.4%、22.6%和82.7% (p < 0.01)。损伤24 h后, 醛类化合物的种类、释放量、脂氧合酶活性及叶片叶绿素荧光动力学参数基本恢复到损伤前水平。说明机械损伤使PSII供体侧受损、脂氧合酶活性升高, 致使C6-C10醛类化合物大量释放, 樟树幼苗通过增加单位面积反应中心的数量来提高光合效率应对胁迫。
叶绿素荧光动力学参数的计算, 参考Strasser等 (2000)的计算方法。暗适应下PSⅡ最大量子产率Fv/Fm = (Fm - Fo)/Fm; 单位面积反应中心数目RC/ CS = (Fv/Fm) × (VJ/Mo) × Fo; 单位反应中心复合体吸收的能量ABS/RC = Mo/VJ/(Fv/Fm); 单位反应中心复合体中被核心捕获的能量TRo/RC = Mo/VJ; 捕获的激子将电子传递到电子传递链中QA-下游的其他电子受体的概率ψo = (1-VJ); 电子传递的量子产额ΦEo = (1-Fo/Fm)×ψo; 单位反应中心捕获的用于电子传递的能量ETo/RC = Mo × (1/VJ) × ψo; 叶片性能指数PIABS =1/ABS/RC × [Fv/Fm/(1-Fv/Fm)] × [ψo /(1- ψo)]; 荧光曲线的相对初始斜率Mo = 4 × (F300μs- Fo)/(Fm-Fo); 2 ms时的可变荧光VJ = (FJ-Fo)/(Fm- Fo)。上述公式中, Fm表示暗适应后的最大荧光强度; Fo表示暗适应后的最小荧光强度; F300μs表示在暗适应后照光300 μs时的荧光强度; FJ表示在暗适应后照光2 ms时的荧光强度。
损伤后1-2 h, 单位反应中心复合体吸收的能量ABS/RC和单位反应中心复合体中被核心捕获的能量TRo/RC均呈现下降趋势, 分别比损伤前下降了12.8%和9.8% (p < 0.01); 单位反应中心捕获的用于电子传递的能量ETo/RC略微上升。损伤24 h后, ABS/RC和TRo/RC恢复到损伤前水平; ETo/RC比损伤前增加19.6% (p < 0.01) (表2)。
损伤后1-2 h, 单位面积光合机构含有的反应中心数目RC/CS呈现增加趋势, 损伤后1 h和2 h的RC/CS分别比损伤前增加了11.6% (p < 0.05)和23.3% (p < 0.01); 电子传递的量子产额ΦEo和捕获的激子将电子传递到电子传递链中QA-下游的其他电子受体的概率ψo均呈现增加趋势, 分别比损伤 前增加了24.4%和22.6% (p < 0.01)。损伤24 h后, RC/CS、ΦEo和ψo均恢复到损伤前水平(表2)。
以损伤前的荧光参数值为基准, 依次计算损伤后不同时期荧光参数的升高或降低百分率, 可知损伤后樟树幼苗叶片PSII最大量子产率Fv/Fm无明显变化, 最大变化值出现在损伤2 h和4 h后, 4 h以后呈现逐渐下降趋势。损伤后1 h和2 h, 叶片性能指数PIABS呈显著增加趋势, 比损伤前分别增加48.39%和82.80% ( p < 0.01)。损伤24 h后, Fv/Fm略低于损伤前水平, PIABS恢复到损伤前水平(表2)。
机械损伤诱导植物启动多重响应机制, 引发的一个“外在性”现象是C6-C10醛类化合物的种类及释放量发生变化。研究表明热胁迫能诱导芦苇(Phrag- mites australis)叶片释放大量(E)-2-己烯醛(Loreto et al., 2006)。在明暗交替实验中, 暗适应下的Populus × canescens叶片能释放出大量乙醛和己醛挥发物(Graus et al., 2004), 而强光胁迫则会诱导饱和C6-C9醛类化合物和E-2-己烯醛大量释放(Loreto et al., 2006; Hu et al., 2009)。O3胁迫诱导玉米(Zea mays)、向日葵(Helianthus annuus)、烟草(Nicotiana tabacum)、红茄(Solanum integrifolium)和芸薹(Brassica campestris)大量释放己醛、庚醛、辛醛、壬醛和癸醛(Wildt et al., 2003)。高于常态浓度的O2能促使复叶槭(Acer negundo)释放更多醛类化合物, 低于常态浓度的CO2则会显著降低醛类化合物的释放量(Hu et al., 2009)。本研究发现机械损伤樟树幼苗释放的醛类化合物主要是己醛、(E,E) -2,4-己二烯醛、(E)-2-己烯醛等C6醛类化合物及庚醛、辛醛、壬醛和癸醛等。损伤1-8 h内上述醛类化合物先后达到释放高峰, 可知机械损伤与其他非生物胁迫一样均能促使植物释放C6-C10醛类化合物。实验中发现环状醛——糠醛的释放规律明显异于直链醛类化合物, 已知短链植物醛类的来源有两个: 植物存储池或特殊酶负责的合成路径(Wildt et al., 2003), 糠醛很可能只来自存储池。机械损伤叶片破坏糠醛的存储池, 致使糠醛释放量短暂升高(1 h); 之后胁迫效应显现, 作为副产品的糠醛合成受阻, 导致其存储量逐渐减少, 出现2-4 h的释放低值。当胁迫效应基本解除后, 糠醛的合成又恢复到常态(24-28 h)。相比之下, 损伤诱导的直链C6-C9醛类化合物更偏向于通过由LOX负责调控的脂肪酸/脂氧合酶途径, 因为实验中检测到LOX活性的时序性变化与直链C6-C9醛类化合物释放量变化相吻合, 并且这条途径已被证实负责合成短链的C6和C9醛类化合物(Loreto & Schnitzler, 2010; Feussner & Wasternack, 2002)。脂肪酸/脂氧合酶途径是利用脂酶把膜系统中的十八碳不饱和脂肪酸(如亚油酸△9,12和α-亚麻 酸△9,12,15)分离出来, 在LOX催化下形成13-(S)-羟基过氧化亚油酸, 之后, 13-(S)-氢过氧化物裂解酶(13-(S)-hydroperoxides, HPL)将其分解为n-己醛(由亚油酸产生)和(Z)-3-己烯醛(由α-亚麻酸产生), 然后再进一步形成其他衍生物(Hatanaka, 1993)。根据上述理论, 实验中C6-C10醛类化合物依次出现释放高峰的可能机理是: 胁迫初期, α-亚油酸12号碳原子双键断裂, 脱落的6碳链加氧形成己醛。α-亚麻酸12号碳原子双键断裂, 脱落的6碳链上原15号碳原子上的双键由于发生双键转移异构化反应, 同时受到醛基影响而迁移到14号碳原子上, 形成2-己烯醛, 进而形成衍生物2,4-己二烯醛; 随着胁迫程度加深, 同样存在于膜脂中的不饱和脂肪酸——油酸△99号碳原子双键也发生断裂形成壬醛, 同时, 亚油酸9号碳原子双键断裂形成2-壬烯醛, 壬醛和2-壬烯醛, 进一步反应形成辛醛、庚醛和2-辛烯醛。
机械损伤引起的一个“内在性”变化是对PSII反应中心产生影响。损伤后1-2 h, O点和P点的荧光强度及J、I两点间的相对可变荧光强度差值增加, 说明损伤加大叶片荧光散射。机械损伤合作杨叶片0.5 h后, Fv/Fm呈现下降趋势, 4 h内恢复到损伤前水平(沈应柏, 2008)。从机械损伤樟树幼苗叶片的研究同样发现, Fv/Fm在4 h内恢复到损伤前水平, 但损伤后1-2 h却呈现上升趋势, 推测可能与损伤程度及树种有关。天线色素吸收的能量除以热能和荧光的形式耗散掉外, 大部分被捕光复合体(light-harvesting complex II, LHC-II)捕获, LHC-II将捕获的光能依次传递给P680受体侧的去镁叶绿素(pheophytin, Pheo)→初级醌受体(QA)→次级醌受体(QB), 把吸收的能量用于固定CO2或其他用途(Strasser et al., 2000; 李鹏民等, 2005)。损伤后1-2 h, ABS/RC和TRo/RC均呈现下降趋势, 而ETo/RC略微上升(表2), 表征放氧复合体的参数OEC没有明显变化, 说明机械损伤胁迫很可能对天线色素造成损伤或形成抑制, 导致天线色素吸收的能量和反应中心捕获的能量减少, 表观上显示为ABS/RC和TRo/RC下降, 而ETo/RC呈略微上升, 表明PSII的电子受体侧接受电子的能力基本没受到影响。反映单位面积反应中心数量的参数RC/CS在损伤后快速上升(表2), 可能预示着机械损伤胁迫发生的同时, 樟树叶片通过提高PSII有活性反应中心的效率以弥补由于天线色素受损而导致的光能吸收不足, ΦEo、ψo两个反映电子传递到电子传递链中超过QA的概率参数以及叶片性能指数PIABS在损伤后1-2 h上升(表2), 表明这一防御机制取得了一定成效。反映叶片活力的性能指数PIABS由3个独立参数构成, 包括: Fv/Fm、比活性参数ABS/RC和捕获激子能导致电子传递效率ψo (李鹏民等, 2005), 被认为比Fv/Fm更能准确地反映植物光合机构的状态(Appenroth et al., 2001; 薛伟等, 2011)。从变化幅度看, 损伤2 h后PIABS比损伤前增加了82.8% (p < 0.01), 而Fv/Fm只增加了1.8%, 前者的变化幅度明显大于后者, 因此我们认为性能指数PIABS不仅能反映机械损伤胁迫对PSII反应中心Fv/Fm的影响, 还可以反映机械损伤对光合器官光能吸收和耗散的影响(Appenroth et al., 2001; van Heerden et al., 2003, 2004)。
本文的其它图/表
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||