
|
|
||
|
模拟酸沉降对南亚热带季风常绿阔叶林土壤微生物群落结构的长期影响
植物生态学报
2021, 45 (3):
298-308.
DOI: 10.17521/cjpe.2020.0217
土壤微生物是生态系统重要的组成成分, 尤其是在土壤风化严重, 养分贫瘠的热带和南亚热带森林生态系统中, 微生物在植物养分的获取、碳循环以及土壤的形成等生态过程中的作用尤为重要。该研究基于鼎湖山南亚热带季风常绿阔叶林长期(10年)的野外模拟酸沉降实验平台, 探究了土壤微生物群落结构对土壤酸化的响应。结果表明, 酸沉降处理显著降低土壤pH (即加剧酸化)。土壤酸化对微生物生物量碳(C)含量的影响不大, 但改变了土壤微生物生物量氮(N)和磷(P)的含量, 导致表层土壤(0-10 cm)微生物生物量C:P和N:P显著提高, 表明土壤酸化可能加剧了微生物P限制。土壤酸化还显著改变了土壤微生物群落结构, 导致次表层土壤(10-20 cm)真菌:细菌显著增加。进一步分析表明, 土壤pH和土壤有效P含量是影响土壤微生物群落最为主要的两个因素。 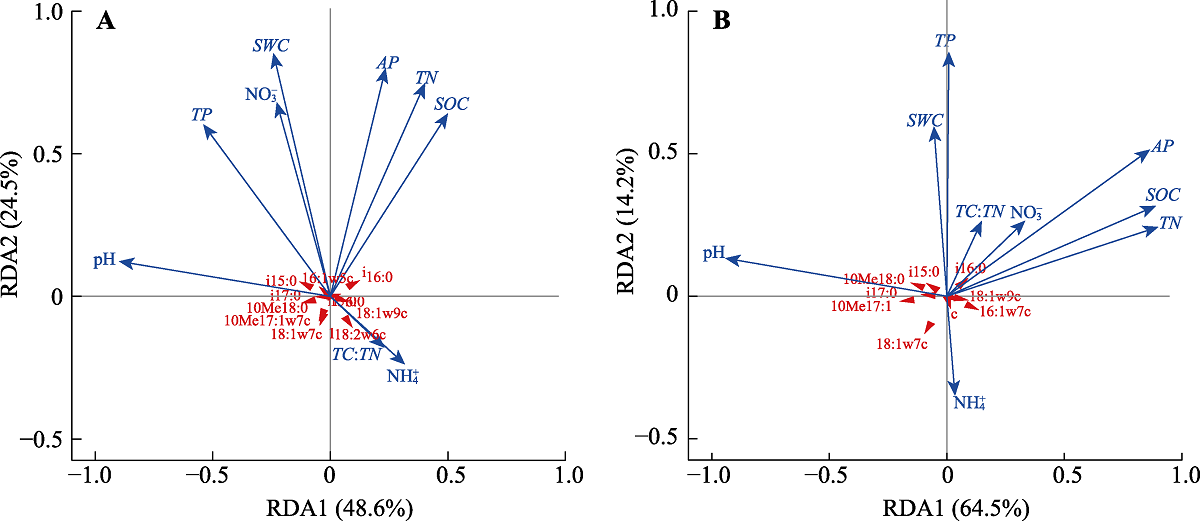 View image in article
图3
0-10 (A)和10-20 cm (B)土层中土壤理化性质与微生物群落结构间关系的冗余分析(RDA)。AP, 有效磷含量; NH4+, 土壤氨态氮含量; NO3-, 土壤硝态氮含量; SOC, 土壤有机碳含量; SWC, 土壤含水量; TC, 土壤总碳含量; TN, 土壤总氮含量; TP, 土壤总磷含量。
正文中引用本图/表的段落
将所有的土壤理化性质纳入RDA模型中发现, 在0-10 cm土层中(图3A), 除土壤铵态氮和硝态氮和TN含量以及土壤C:N外, 其他土壤理化性质均与微生物群落组成显著相关(p < 0.05), 所有土壤性质解释了土壤微生物群落结构变化的40.8%。第一主轴占全部解释方差的比例为46.8%, 与pH、SOC、TN和TP含量显著相关。第二主轴占全部解释方差的比例为24.5%, 与 SWC、SOC、TN、TP、NO3-和AP含量显著相关。使用双终止准则前向选择解释变量, 得到最主要的环境因子pH、SWC、SOC和AP含量, 解释土壤微生物群落结构33.7%的变化。同样, 在10-20 cm土层中环境因子解释了土壤微生物群落结构变化的47.8%, 其中最主要的环境因子pH、AP和TP含量, 共解释了土壤微生物群落结构变化的41.8% (图3B)。
本研究的对象为南亚热带季风常绿阔叶林, 其土壤高度风化, 并且在长期受到不平衡氮磷沉降后, 植物生产力和土壤微生物活性的主要限制性养分由氮转为了磷(Mo et al., 2008; Cleveland et al., 2013; Deng et al., 2017)。因此, 长达10年的酸沉降实验虽然显著改变了SWC、土壤硝态氮含量以及C:N (表2), 但这并未成为影响土壤微生物群落结构的主要因素(图3)。此外, 在酸化加剧背景下, 土壤AP、TP以及MBP含量均呈显著的下降趋势, 这意味着土壤酸化确实有可能导致热带森林磷限制的加剧, 从而减少微生物对磷的同化作用。
在本研究中, 土壤酸化的加剧导致土壤微生物群落结构发生显著变化(图2A), 导致10-20 cm土壤中F:B显著增加(表3), 其中土壤pH是导致微生物群落结构发生变化的主要原因(图3)。这些发现与先前亚热带森林生态系统中的研究结果相符(Liu et al., 2017; Li et al., 2018), 即在一定的pH范围内外源酸的输入对细菌和真菌有着不同的影响(Meng et al., 2019), 真菌对土壤pH的下降具有较强的适应性(B??th & Anderson,2003)。在亚热带阔叶落叶林中, Li等(2018)发现土壤酸化虽然对真菌和细菌均没有显著的影响, 但显著增加F:B。Liu等(2017)在亚热带人工林中则发现土壤酸化显著减少细菌的相对丰度, 但对真菌没有显著影响, 导致以细菌为主的微生物群落转变为以真菌为主的微生物群落。在北方森林生态系统中, 随着土壤pH的增加, 细菌相对丰度显著增加, 而真菌则在酸性土壤中表现出较高的活性(H?gberget al., 2007)。湿地森林生态系统中, 模拟氮沉降导致土壤pH显著下降也会使得土壤微生物群落结构发生显著的变化(Li et al., 2019)。而在土壤中性偏碱性的温带森林生态系统中, 两年的模拟酸雨处理导致土壤pH下降则并未显著改变土壤中的F:B (Zheng et al., 2018)。通常认为, 真菌将过剩的H+储存在液泡中的能力及其异于细菌的细胞壁结构是真菌和细菌具有不同的最适pH范围的主要原因(Ware et al., 1990)。因此相较于细菌, 真菌的最适pH范围更低且更广(Ware et al., 1990), 更能够适应未来的土壤酸化。
除土壤pH外, 土壤磷含量是导致本研究中土壤微生物群落结构发生变化的另一个主要原因(图3), 即土壤酸化导致的土壤磷限制加剧可能进一步促进土壤微生物群落向以真菌为主的微生物群落的转变(表2, 表3)。大量研究表明, 在土壤磷限制严重的热带森林生态系统中, 微生物真菌和细菌的比值显著高于农田和草地土壤, 且真菌丰度与土壤磷含量和C:N具有很强的相关关系(Lauber et al., 2008), 而这可能主要是真菌在养分贫瘠环境中较高的适应性, 以及在酸性条件下对磷具有更强的获取能力所导致(DeForest & Scott, 2010; Jones & Oburger, 2011)。例如, 在磷有效性过低的情况下, 为了维持生长所需的足够养分, 植物将更多的光合作用产物投入地下, 通过增加其根系上菌根真菌的定殖以扩大土壤磷的获取范围和增加吸收面积, 增加对养分的获取能力(van der Heijden et al., 1998; Wardle et al., 2004), 或者通过养分的重吸收从而减少养分的消耗等途径适应养分贫瘠的土壤, 而这些变化最终也将导致土壤中微生物的群落结构发生改变(Strickland & Rousk, 2010)。此外, 在热带森林土壤纤维素分解过程的研究中发现, 真菌和细菌生长的限制性养分存在差异, 细菌由于较大的比表面积对磷脂的需求较高, 使得土壤磷含量的改变也可能进一步抑制了细菌的活性(Nottingham et al., 2018)。不同的是, 温带森林生态系统中, 模拟酸沉降导致的微生物群落结构的变化主要是土壤中碳和氮的有效性改变所导致的(Killham et al., 1983), 在氮有效性较低的条件下, 氮的添加可以通过解除土壤中微生物的碳和氮的限制, 从而缓解氮添加导致的土壤pH下降对微生物生长和群落结构的影响(Treseder, 2008)。
本文的其它图/表
|


{kind=link}