|
|
||
|
川西王朗亚高山暗针叶林25.2 hm2动态监测样地物种组成与群落结构特征
植物生态学报
2022, 46 (9):
1005-1017.
DOI: 10.17521/cjpe.2022.0094
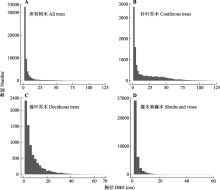
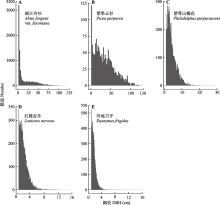
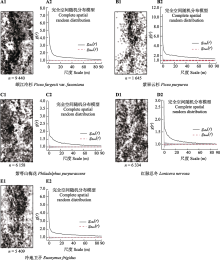
川西亚高山暗针叶林是中国寒温性针叶林的重要代表。以云杉(Picea spp.)和冷杉(Abies spp.)为优势树种的暗针叶林是青藏高原东缘亚高山森林的主体, 也是我国长江上游重要的生态屏障。该研究基于川西王朗保护区亚高山暗针叶林25.2 hm2动态监测样地的首轮清查数据, 分析样地木本植物组成和群落结构特征。依照全球森林观测网络(ForestGEO)的标准技术规范, 建立面积为25.2 hm2 (700 m × 360 m)的暗针叶林固定监测样地, 对胸径≥1 cm的木本植物进行挂牌标记、坐标定位、物种鉴定和检尺。主要结果有: 样地内共标记独立的木本植物个体56 574株, 分属15科27属46种, 其中针叶树4种, 常绿阔叶树6种, 落叶阔叶树36种; 每hm2个体数≤1的稀有种共13个, 占样地物种总数的28.26%, 占样地总个体数的0.16%。该群落以针叶树岷江冷杉(A. fargesii var. faxoniana, n = 9 440)和紫果云杉(P. purpurea, n = 1 645)为建群种和乔木层优势种, 重要值分别占总数的26.03%和10.69%, 其单位面积内的胸高断面积分别占总胸高断面积(37.87 m2·hm-2)的55.22%和24.84%。落叶阔叶树个体数占比最多(78.48%), 其中紫萼山梅花(Philadelphus purpurascens)、红脉忍冬(Lonicera nervosa)、冷地卫矛(Euonymus frigidus)为灌木层主要优势种。常绿阔叶树个体仅268株, 均为杜鹃属(Rhododendron)植物。木本植物的径级组成具有典型老龄林特征。单株最大胸径出现在紫果云杉, 达到127.07 cm。紫果云杉平均单株胸高断面积(0.14 ± 0.18) m2显著高于岷江冷杉(0.056 ± 0.11) m2。所有标记个体的径级分布呈倒“J”形, 表明群落处于稳定和正常生长状态。点格局分析的结果表明5个优势种(重要值占比>5%)均呈聚集分布, 但随着空间尺度的增加, 物种聚集程度变小, 趋向于随机分布。该研究的结果为深入研究川西亚高山暗针叶林的多样性维持与更新机制和该类型森林生态系统的保护与管理提供了基础信息。
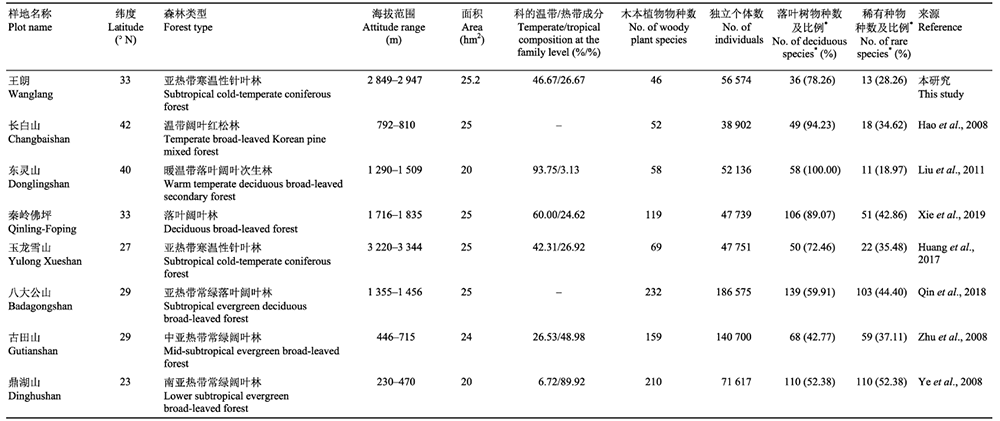
表1
王朗25.2hm2森林动态监测样地与其他7个不同纬度、不同植被类型样地特征的比较
正文中引用本图/表的段落
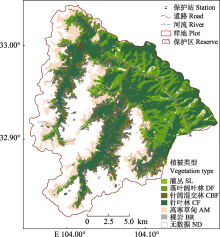
王朗样地中央平坦, 有河流穿过, 两侧阴坡和阳坡的海拔升高, 样地内海拔跨度为2 849-2 947 m (图2A)。首轮清查挂牌木本植物树种计46种, 隶属于15科27属, 包括裸子植物2科4属4种, 被子植物13科23属42种; 其中乔木14种, 灌木29种, 藤本3种(附录I)。总计挂牌56 574个独立植株, 55 814个分枝, 合112 388个, 独立植株平均密度为2 245株·hm-2。乔木、灌木、木质藤本独立植株分别为20 298、35 185和1 091株(附录I)。样地内包括针叶树4种, 常绿阔叶树6种(均为杜鹃属(Rhododendron)植物), 落叶阔叶树36种, 独立植株数分别为11 905、268和44 401株(附录I)。其他7个样地(长白山、东灵山、秦岭佛坪、玉龙雪山、八大公山、古田山和鼎湖山)的基本信息和数据特征见表1。已挂牌木本植物的空间分布见图2B。
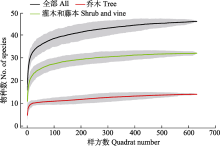
王朗25.2 hm2亚高山暗针叶林样地的物种组成具有典型的山地寒温性针叶林特征, 是北半球分布纬度较南的暗针叶林(蒋有绪, 1963)。王朗样地位于从东部亚热带向青藏高原过渡的川西峡谷山地区域, 其气候特征与北温带相似, 但其植物区系组成呈现出与北方暗针叶林区系组成不完全相似的特征。西南亚高山暗针叶林和北方暗针叶林的区系组成均以北温带成分为主、暗针叶树树种丰富, 王朗样地还包括了部分热带型物种, 如丰富的杜鹃属植物, 为典型的热带高山起源(蒋有绪, 1963)。在科水平上, 王朗样地木本植物热带成分组成与同为亚高山暗针叶林的玉龙雪山样地接近, 高于秦岭样地和东灵山样地, 低于古田山样地和鼎湖山样地(表1)。从样地物种累积曲线来看, 随样方数增加, 累积物种数快速增加。当样方数为全部数量一半时, 物种累计达43种, 占全部物种数的93.48%, 进一步增加样方数, 记录到的树种数增长缓慢。可见, 王朗样地对该区域亚高山针叶林木本植物多样性有较为充分的覆盖。
王朗样地中稀有种占样地个体数比例不大, 仅0.16%, 但占物种数比例较高(28.26%)。稀有种是木本植物多样性的重要组成部分, 根据已有样地报道, 不同森林类型中均有较高比例的稀有种存在(表1)。如长白山样地阔叶红松林和玉龙雪山样地寒温性针叶林中, 稀有种比例分别高达34.6%和35.48% (郝占庆等, 2008; 黄华等, 2017); 亚热带与热带森林如古田山亚热带常绿阔叶林样地和巴拿马Barro Colorado Island (BCI)热带雨林样地中, 稀有种比例分别为37.1%和36.8% (He et al., 1997; 祝燕等, 2008)。稀有种在生境中的个体数量虽少, 但其对生态系统功能和服务的贡献不完全依赖于个体数量(Lyons et al., 2005; Laura et al., 2019)。已有研究表明, 稀有种可能通过影响种间相互作用等间接过程增强生态系统功能(Laura et al., 2019)。如通过与微生物的互惠作用, 具有固氮作用的稀有种可以促进土壤肥力的增加, 从而使生态系统获得更高的生产力(Marsh et al., 2000)。另外, 根据Janzen-Connell理论, 稀有种的负密度制约强度较弱, 生长速率更快, 同时, 稀有种的存在可以稀释广布种的负密度制约强度, 从而减小其种内竞争强度, 最终促进整体生态系统服务(Janzen, 1970; Holt et al., 1994)。然而, 由于个体数量稀少、调查成本高、短期研究难以探测到其显著影响等原因, 对于稀有种的研究仍较不充分, 其对生态系统的影响普遍被低估(Lyons et al., 2005; Laura et al., 2019)。森林动态监测样地具有调查面积大、监测时间长等特点, 是深入研究稀有种在维持物种多样性及促进生态系统服务等作用的重要平台(Mi et al., 2012), 因此, 在王朗样地未来的研究中应更多地关注稀有种的生态功能。
本文的其它图/表
|