


紫茎泽兰(Eupatorium adenophorum)是我国外来入侵物种中危害最为严重的植物之一,属于菊科泽兰属,原产于中美洲的墨西哥和哥斯达黎加。作为一种有害的外来物种现已在夏威夷、澳大利亚、新西兰、泰国等地爆发式繁衍,广泛分布在世界热带、亚热带30多个国家和地区。此物种大约于20世纪40年代从中缅边境通过自然扩散传入我国云南省,约经半个世纪的扩散,现已在我国的云南、贵州、四川、广西、西藏、台湾等4省2区广泛分布和危害(强胜,1998;李振宇和谢焱,2002),并仍以每年大约60 km的速度,随西南风向东和北传播扩散(向业勋,1991)。在重庆和湖北的三峡地区亦发现紫茎泽兰,并采集到标本(段慧,2003)。
作为一种入侵性很强的外来物种,关于紫茎泽兰的研究无论在国外(Bess & Haramoto, 1959;Dodd, 1961; Auld, 1969a, 1969b, 1971; Auld & Martin, 1975; Arunachalam et al., 1999; Papes & Peterson, 2003),还是在国内(刘伦辉等,1985, 1989;赵国晶和马云萍,1989;强胜,1998;周俗和谢永良,1999;宋启示等,2000;强胜,2004) 都已经开展了很多。目前国内的研究热点主要集中在3个方面:1)分布范围及可能扩散的区域;2)紫茎泽兰侵入扩张的机理;3)防除方法。本文就邓外来入侵种的研究现状做一总结和展望,以期对我国正在进行和将要进行的有关研究有所裨益。
1 分布范围及可能扩散的区域
1.1 紫茎泽兰在中国的分布
在云南省,据调查,紫茎泽兰在10个地、州的98个县、市都有发生,面积达24.77万km2,其中以西南部和南部地区发生量最大(赵国晶和马云萍,1989)。在四川省,2002年对全省紫茎泽兰的分布进行了调查,发现自1978年在凉山州盐源县树河乡发现紫茎泽兰以来,已经在凉山州、攀枝花市、乐山市、宜宾市、雅安市、甘孜州6个市(州)共34个县(区)、400多个乡镇有紫茎泽兰分布,面积达86.478万hm2,成灾面积达82.277万hm2。其中凉山州和攀枝花市为全省的重灾区,占全省紫茎泽兰分布面积的99.15%(周俗等,2004)。在贵州省,1991年,已分布到黔西南、黔南、安顺、六盘水等4个地、州(市)的兴义、兴仁等14个县,其中侵入面积较大的有兴义、册享、望谟、安龙等4个县的南部地区。其分布范围已超越26°N,接近107°E,并有逐渐向贵州省东北方向发展蔓延的趋势(向业勋,1991)。在西藏自治区,在聂拉木县也有发现(陈开基,1989)。在广西壮族自治区,20世纪自70年代传入以来,到目前为止,最东达到横县,最北分布在乐业和南丹,在百色地区的田林、西林、隆林三地入侵最为严重,另外在南宁、靖西、平果、天蛾也有发生1)((1) 卢志军. 2004. 个人通讯)。紫茎泽兰在台湾省也有分布(李振宇和解焱,2002)。在重庆和湖北的三峡地区发现紫茎泽兰,并采集到其标本,这是目前我国分布纬度最北的记录(段惠,2003)。总之,目前紫茎泽兰在我国的大致分布范围如下:南起中缅、中越边境地区,北抵泸定-犍为-宜宾-重庆一线,西起贡山-冕宁-石棉,东到横县-南宁-南丹-贵阳-重庆一线。危害最严重的地区位于102°E左右,包括:云南省的临沧、思茅、昆明,广西省的百色,贵州省的黔西南、四川的攀枝花和凉山(卢志军和马克平,2004)。
图1
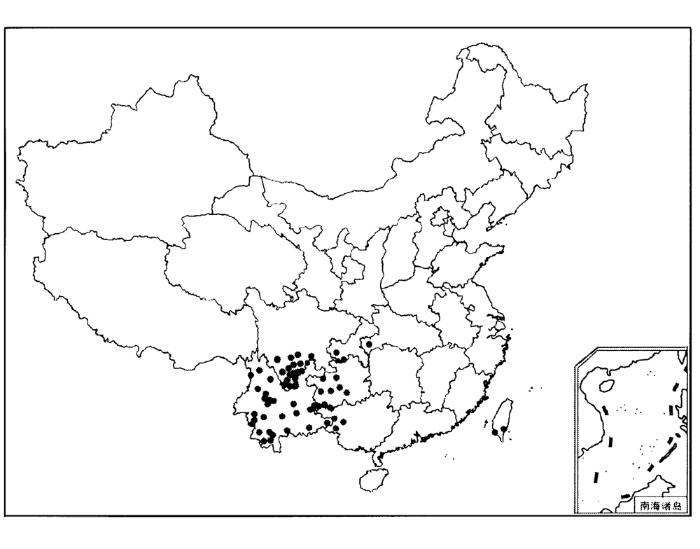
1.2 预测其可能扩散的区域
据调查数据测算,8年间紫茎泽兰向北纬推进了两度半,直线距离约280 km,平均每年在凉山州境内向北传播35 km(周俗和谢永良,1999)。由此可见,紫茎泽兰扩展速度相当惊人,因而,对其蔓延趋势做出预测,已成为迫切需要解决的问题。
最近,已经有学者应用生态位模拟的方法,通过收集其原产地墨西哥和中国的气象资料,来预测紫茎泽兰可能入侵的范围。预测的范围包括该植物在中国境内已分布的省份及未来在华中、华东易受入侵的区域。通过模型预测,目前紫茎泽兰分布的6个省区当中的4个省份(云南、贵州、四川、广西)与模型所预测的分布区十分吻合,而西藏自治区则包括在模型所预测分布区的边缘地带。预测的潜在分布区包括:甘肃、宁夏、陕西、山西、河南和湖北5省1区,而且模型还预测了和目前分布区不相连的两个潜在分布区,分别在我国的东北和东南,这些潜在分布区包括东北的黑龙江省、辽宁省和东南的福建和浙江两省(Papes & Peterson, 2003)。
卢志军和马克平(2004)通过调查其在我国的分布状况、比较其在原产地(墨西哥和哥斯达黎加)与我国分布区的地形因素和气候条件,也对其潜在分布区进行了推测:紫茎泽兰在不断地向东向北推进,但最终分布格局还将以云贵高原为主体,随着分布区扩大到云贵高原以外,它的扩展速度、危害程度都将逐渐降低。
2 紫茎泽兰侵入扩张的机理研究
紫茎泽兰作为一个成功的入侵种,兼备了许多入侵杂草所具有的显著特征,对其入侵机制和扩散机制的研究有助于其它有害入侵植物的研究,丰富和发展入侵生态学理论,推动我国外来入侵植物学研究的进步。
2.1 紫茎泽兰侵入扩张的分子机理研究
生物入侵为我们提供了很多快速适应与进化的经典模型。紫茎泽兰分布范围较广,生境条件多样,因此,从分子生态学的角度对不同侵入地区及不同生态环境中的紫茎泽兰进行遗传多样性分析,探索其遗传变异规律,开展紫茎泽兰分子生态适应机制的研究,对于解释紫茎泽兰的入侵机制有积极意义。紫茎泽兰作为一种入侵性极强的外来物种,目前其入侵原因尚不明了,从遗传学角度揭示其根源已经成为生物学界关注的热点(强胜,2004)。
强胜(2004)利用AFLP技术,对24个地区的紫茎泽兰种群的基因组DNA多态性进行了检测,并通过UPGMA聚类分析对结果进行了分析,结果表明:1)紫茎泽兰遗传多样性丰富,具有广泛的生态适应性;2)紫茎泽兰入侵时间长的云南省遗传多样性较高,入侵时间短的新入侵地区的遗传多样性则相对较低;3)风媒和水媒可能是紫茎泽兰传播的途径,因为可以在样点间观察到明显的地缘性亲缘关系。
2.2 紫茎泽兰侵入扩张的“新武器”——化感作用
近年来,Callaway和Aschehoud (2000)提出了“新武器”的假说,认为外来入侵种的入侵成功主要归因于其改变了入侵地的植物及微生物群落原有的种间关系,引入了新的相互作用机制,这种改变最初主要是通过生物化学物质加以实现。并通过实验证明,外来入侵种铺散矢车菊(Centaurea diffusa)所具有的化感作用,在入侵地对本地种发挥了比在原产地对原共生种更大的作用,是其在入侵地成功入侵和扩散的强大武器。
作为一种世界性的恶性杂草,紫茎泽兰已经被许多研究证明具有很强的化感作用(Tripathi et al., 1981;Angiras et al.,1988,1989;赵国晶和马云萍,1989;和爱军和刘伦辉,1990;Baruah et al.,1994;宋启示等,2000;于兴军等,2004)。紫茎泽兰的水浸提液可以对黑麦草(Lolium perenne)、白三叶(Trifolium repens)等植物的种子发芽和胚根与胚芽伸长产生抑制作用。 紫茎泽兰不同部位的化感作用也不同,叶的抑制作用远大于根茎(和爱军和刘伦辉,1990);最近于兴军等(2004)也通过实验发现,对于紫茎泽兰的入侵力来说,地上部分的化学作用比地下部分具有更大的贡献。不同提取方法也会影响紫茎泽兰的化感作用,赵国晶和马云萍(1989)研究表明:紫茎泽兰叶水提取液的化感作用>叶乙醚提取液的化感作用;而紫茎泽兰根的化感作用则相反,即根乙醚提取液的化感作用>根水提取液的化感作用。于兴军等(2004)通过对比紫茎泽兰入侵的不同生境发现,不同生境下紫茎泽兰茎和根的水浸提液的化感作用存在差异,即公路边>落叶阔叶林>常绿阔叶林,并通过关联分析显示出不同生境条件下的化感作用与本地植物的相对多度存在显著的相关关系,证明不同生境下化感作用的差异是造成紫茎泽兰入侵效果的原因之一。
对紫茎泽兰化学成分的研究可以揭示它与其它植物和动物相互作用的基础,阐明化感作用的化学机制,也是目前的研究热点之一。在国外,Bohlmann和 Rajindex(1981)从紫茎泽兰中分离得到4种化合物和6种倍半萜烯;Bordoloi 等(1985)从紫茎泽兰中分离得到5种具有昆虫拒食活性的倍半萜烯类化合物;Baruah等(1994)也分离鉴定了紫茎泽兰的化感物质为倍半萜烯化合物。在我国,许多学者开展了紫茎泽兰化感物质的研究。许云龙等(1988)从紫茎泽兰中分离得到β-谷甾醇,豆甾醇,蒲公英甾醇、棕榈酸酯及蒲公英甾醇乙酸酯等化合物;李蓉涛等(1997)从紫茎泽兰中分离得到8种化合物;丁智慧等(1999)从紫茎泽兰的花中分离到12个化合物,其中紫茎泽兰内酯为一新的杜松烯类倍半萜内酯化合物,另一种从紫茎泽兰中提取到的克拉维醇,是首次从泽兰属植物中分离得到,活性筛选显示这种化合物对小菜粉蝶幼虫(Pieris rapae)具有一定的拒食活性;宋启示等(2000)通过研究其地上部分的石油醚、乙酪和水提取物对豌豆(Pisum sativum)的种子萌发和幼苗生长的化感作用发现,它的化感物质主要集中于石油醚提取物及石油醚提取物吸附于活性炭后的甲酪洗脱物中。从石油醚提取物分离出的第二、第三和第四组分为主要的活性成分,且经鉴定,化感活性强的石油醚第二组分为9-酮-泽兰酮。提出了紫茎泽兰地上部分挥发性化感物质的释放以及地上部分和凋落物中的水溶性化感物质的淋溶等植物化感途径。
2.3 紫茎泽兰有利于入侵的生物学特性
紫茎泽兰具有很强的光合能力,其最大光合速率为23 μmol CO 2·m-2·s-1左右(王俊峰等,2004)。紫茎泽兰的光饱和点比较高, 接近阳性植物;紫茎泽兰对光照的适应范围较宽,光补偿点低,仅为13 μmol·m -2·s-1左右,而CO2补偿点为80 μmol·mol -1左右, 具有阳性偏阴的生态习性(刘文耀等,1988),适应大幅度的光环境很可能是其入侵成功的重要原因之一。另外,王俊峰和冯玉龙(2004)通过实验也发现,紫茎泽兰对光环境较强的适应能力可能是紫茎泽兰的入侵性强于兰花菊三七(Gynura sp.)的重要原因。紫茎泽兰通过叶片自遮荫来适应强光,高度自遮荫可以阻止其它植物在其冠内生长,而其自身的耐荫能力却很强,在相当于林下光强的弱光环境中仍能生长,并具有较强的光合能力(王俊峰等,2004)。刘文耀等(1988)发现从夏季到秋季,紫茎泽兰叶片的日平均光合速率不仅没有减少,而且略有增加,说明紫茎泽兰一直处于旺盛的生长状态,而在这期间,正是牧草及其它一些植物幼苗、幼树生长能力衰退的时期。紫茎泽兰所具有的这些光合特性都是其成功入侵的生理基础。
3 紫茎泽兰的防除
3.1 紫茎泽兰的人工防除
将紫茎泽兰在秋冬季进行挖除,晒干烧毁,而后立即种上适应性强、繁殖快的牧草和树种,如象草(Pennisetum purpureum)、狗芽根(Cynodon dactylon)、刚莠竹(Microstegium ciliatum)、白三叶以及云南松(Pinus yunnanensis)、兰桉(Eucalyptus globulus)、华山松(Pinus armandi)等,以减少和避免紫茎泽兰的侵占,最终实现植物替代(何大愚和梁家社,1988;向业勋,1991;孙劲,2003)。贵州等省区利用优质牧草皇竹草(Pennisetum sinese)来抑制紫茎泽兰种子的传播蔓延,是一种很有潜力的替代控制种(孙劲,2003)。
3.2 紫茎泽兰的化学防除
虽然化学防治不能根除紫茎泽兰,但对危害严重、面积大,在人工清除有困难的地方适当采用化学药剂进行防治,结合人工清除,在一定范围内是可行的。在使用时,要远离水源和人群,保护环境与生物安全。
澳大利亚的Auld就进行了一系列紫茎泽兰化学防除的研究。Auld (1972)根据除草剂的有效性、专一性及成本3个方面的综合考虑,筛选出3种化学除草剂:1)2,4-D胺;2)2,4-D和2,4,5-T的混合物;3)氯酸钠。并通过研究发现:0.6%的2,4-D胺在秋季施用具有很好的专一性,并且可以使紫茎泽兰达到很高的死亡率。
3.3 紫茎泽兰的生物防除
生物防除是控制外来种最有前景的途径,其主要方法包括用真菌和昆虫来控制外来种。Bess 和Haramoto(1959)将泽兰实蝇(Procecidochares utilǐs)从墨西哥引入夏威夷,对紫茎泽兰进行生物防治。研究发现,泽兰实蝇可以严重抑制紫茎泽兰的生长,主要是通过在其植株上产生虫瘿,使其开花数量减少,甚至导致死亡; 我国学者对于泽兰实蝇也开展了广泛的研究(何大愚和梁家社,1988;张智英等,1988;陈旭东和何大愚,1990a,1990b),获得了相似的结论(刘文耀等,1991)。提出了最佳释放虫量指标为每条虫占有10个枝条(陈旭东和何大愚,1990b),并从多方面探讨了泽兰实蝇对于紫茎泽兰的影响(郑征等,1989;陈旭东和何大愚,1990a;代聪等,1991),发现泽兰实蝇可以减弱紫茎泽兰的光合作用;影响紫茎泽兰的密度、种子的入侵能力以及种子的萌发率;而且,由于泽兰实蝇的寄生使营养流集中转向茎部,一部分形成膨大的虫瘿,累积了大量的生物量,另一部分提供给泽兰实蝇进行发育,致使根系发育不良,根冠比减小,影响根系的矿质营养及水分的吸收,严重影响紫茎泽兰的生长发育。但是就野外观察发现,这种天敌对于控制紫茎泽兰的效果并不理想(刘伦辉等,1985)。
链格孢菌(Alternaria alternata)菌株的菌丝体侵染紫茎泽兰的速度快、致病力强,能快速杀死该植物,被认为是控制紫茎泽兰很有潜力的新途径(强胜,1998)。万佐奎等(2001)就链格孢菌毒素对紫茎泽兰的致病机理进行了深入研究,结果发现,链格孢菌毒素使紫茎泽兰叶组织细胞膜透性上升,Na+和K+渗漏量增加,膜脂过氧化加强,丙二醛含量上升,说明致病毒素能够引起细胞膜伤害,造成膜功能的紊乱。链格孢菌毒素处理的紫茎泽兰叶片中过氧化物酶、抗坏血酸过氧化物和过氧化氢酶的活性均较对照降低,可能使活性氧清除系统中酶系统遭到破坏,活性氧过量积累,细胞因此受到伤害,最终使紫茎泽兰叶片表现出受害症状。但是致病毒素在细胞膜上的受体以及信导的传递等有待进一步研究。另外,链格孢菌是微生物除草剂开发中极其重要的一类真菌,有可能成为生物源的化学除草剂。
4 研究展望
4.1 紫茎泽兰长距离传播模型研究
4.2 紫茎泽兰化感作用
紫茎泽兰的化感作用在其入侵成功的过程中扮演着重要角色,但是还是缺乏在原产地与入侵地化感作用的对比研究,而这项研究应该有助于我们理解其入侵机制,了解其在原产地不能大面积扩散的原因。另外,对于赋予其具有化感作用的化学物质的结构、性质及作用机制尚需进一步研究。
4.3 紫茎泽兰生理生态适应性研究
紫茎泽兰作为一种恶性杂草,其生理生态研究还是了解的很少,特别是通过控制实验来开展对紫茎泽兰扩散过程的限制因子的研究和在不同水热条件和光照条件下的生理适应机制,以及它对这些限制因子的耐受范围的研究。所以,对于紫茎泽兰开展对模拟不同气候条件(包括平均温度、极端温度、大气湿度、光照强度等)下植物生理生态响应的研究,了解其在不同气候条件下的生理适应机制,将使我们能够洞悉外来入侵种在新入侵地的生态环境下的扩散机制,预测其潜在的入侵区域,发现其适应过程中的薄弱环节。
4.4 紫茎泽兰传粉过程及其生殖生态学特性
紫茎泽兰传粉过程及其生殖生态学的一些特性目前仍没有进行较系统的研究,而外来植物的繁殖能力大小对入侵是否成功意义很大,繁殖能力与植物的入侵能力呈正相关(Barret, 1983)。紫茎泽兰无融合生殖的基因调控机制,以及原产地和新入侵地的繁育系统的变化,都是将来研究的热点。
4.5 紫茎泽兰生物防除
参考文献
Allelopathic effects of weeds on germination and seeding growth of maize and soybean
Allelopathic effects of some weeds on germination and growth of chickpea (Cicer ariet)
Influence of soil properties on microbial populations, activity and biomass in humid subtropical mountainous ecosystems of India
Incidence of damage caused by organisms which attack crofton weed in the Richmond-Tweed region of New South Wales
The distribution of Eupatorium adenophorum Spreng. on the far north coast of New South Wales
Survey of weed problems on the north coast of New South Wales
Chemical control of Eupatorium adenophorum, crofton weed
The auteclogy of Eupatorium adenophorum Spreng. in Australia
The modes of origin of weeds
In: Baker HG, Stebbins GL eds. The Genetics of Colonizing Species.
Seed germination and growth inhibitory cadinenes from Eupatorium adenophorum Spreng

Chloroform extract of the aerial parts ofEupatorium adenophorum Spreng was fractionated and examined for growth inhibition. Bioassay-directed fractions of the plant materials afforded three known cadinenes and beta-sitosterol. The effects of different fractions as well as isolated cadinenes were determined usingAllium cepa, Raphanus sativus, andCucumis sativus seeds. Three-day exposure to these cadinenes significantly inhibited germination and seedling growth of all three assay seeds. The degree of inhibition was dependent upon seed species and the concentrations of the compounds tested. Cadinene(1) was found to be more inhibitory to the seeds tested and the activity of the cadinene(3) was less than that of(1) and(4).
Life history of colonizing plants: some demographic,genetic,and physiological features
In: Mooney HA, Drake JA eds. Ecolology of Biological Invasions of North America and Hawaii.
Biological control of Pamakani Eupatorium adenophorum in Hawii by a Tephritid gall fly, Procecidochares utilis. Ⅱ. Population studies of the weed, the fly and the parasites of the fly
Six cadinene derivatives from Ageratina adenophora
Absolute stereochemistry of the insect antifeedant cadinene from Eupatorium adenophorum
Long-distance seed dispersal in plant populations
Invasive plants versus their new and old neighbors: a mechanism for exotic invasion
Invading exotic plants are thought to succeed primarily because they have escaped their natural enemies, not because of novel interactions with their new neighbors. However, we find that Centaurea diffusa, a noxious weed in North America, has much stronger negative effects on grass species from North America than on closely related grass species from communities to which Centaurea is native. Centaurea's advantage against North American species appears to be due to differences in the effects of its root exudates and how these root exudates affect competition for resources. Our results may help to explain why some exotic species so successfully invade natural plant communities.
The potential threat of vicious weed Eupatorium adenophorum Spreng
Study on control effect of Procecidochares utilis on Eupatorium adenophorum and its evaluation
Study on the strategy of biological control of Eupatorium adenophorum by using Procecidochares utilis
Range extension of aquatic vascular plant species
A study on effect of Procecidochares utilis on control of Eupatorium adenophorum
Chemical constituents from the flowers of Eupatorium adenophorum
Biological control of Eupatorium adenophorum in Queensland
Eupatorium adenophorum Spreng
New leaf-spot disease of Eupatorium adenophorum,caused by Mycovellosiella eupatorii-odorati in China
Studies on the biological characteristics of Mycovellosiella eupatorii-odorati (Yen) Yen, a potential pathogen for the biological control of crofton weed,Eupatorium adenophorum
Effect of water extract of Eupatorium adenophorum on the germination of several plants
Advances in the control research of Eupatorium adenophorum
An integrated approach to the ecology and management of plant invasions
Chemical constituents from Eupatorium adenophorum
Studies on the distribution, harmfulness and control of Eupatorium adenophorum Spreng
The characteristic research of autecology ecology of Pamakani (Eupatorium adenophorum)
The effect of Procecidochares utilison growth and development, distribution of biomass of Eupatorium adenophorum
Photosynthetic characteristics of Eupatorium adenophorum and their ecological significance
Studies on some species of the genus Phaeoramularia in China
The influence of topographical factors on the invasion of the alien species, Eupatorium adenophorum
Mechanisms of long-distance dispersal of weed by wind
Ecological and physiological characteristics of invading species
In: Groves RH, Burdon JJ eds. Ecology of Biological Invasions: an Australian Perspective.
Attributes of invaders and invading process: terrestrial and vascular plants
In: Drake J, Mooney HA, Dicastri F,Groves RH,Kruger FJ, Rejmanek M, Willamson M eds. Biological Invasions: a Global Perspective.
Predicting the potential invasive distribution for Eupatorium adenophorum Spreng
The history and status of the study on crofton weed( Eupatorium adenophorum Spreng.) a worst worldwide weed
Genetic diversity in populations of Eupatorium adenophorum detected by AFLP marker
What attributes make some plant species more invasive?
Persistent soil seed bank of Eupatorium adenophorum
Allelopathic potential of Eupatorium adenophorum
Control of Eupatorium adenophorum, an overview
Introduction to the controlling and utilizing of Eupatorium adenophorum Spreng
Breeding systems of invasive alien plants in South Africa: does Baker's rule apply?
Seasonal variation in the seed banks of herbaceous species in ten contrasting habitats
The fuctional ecology of soil seed banks
In: Fenner M ed. Seeds: the Ecology of Regeneration in Plant Communities 2nd edn.
Allelopathic potential of Eupatorium adenophorum, a dominant ruderal weed of Meghalaya
Genetrically engineered fungi for weed control
The pathogenic mechanism of toxin of Alternaria alternata (Fr.) Keissler to Eupatorium adenophorum Spreng
The effect of light intensity on biomass allocation,leaf morphology and relative growth rate of two invasive plants
Acclimation of photosynthetic characteristics to growth light intensity in Eupatorium adenophorum Spreng
The distribution, harmfulness and elimination of Eupatorium adenophorum Spreng
The first report on the chemical constituents of Eupatorium adenophorum
Study on effect of Mycovellosiella eupatorii-odorati upon growth and physiological parameters of Eupatorium adenophorum
Relationships between allelopathy and invasiveness by Eupatorium adenophorum in different sites
Biology of a gall fly, Procecidochares utilǐs [Dip.:Tephritidae] and its impact on crofton weed, Eupatorium adenophorum
The investigation research on the distribution and harmfulness of Eupatorium adenophorum Spreng. in Yunnan Province
Advances in model construction of anemochoric seed long distance dispersal
Study on effect of Procecidochares utilǐs growth and photosynthesis of Eupatorium adenophorum
The damage situation and control countermeasures for Eupatorium adenophorum in Sichuan Province
The investigation report on the poisonous and injurious plant —— Eupatorium adenophorum Spreng. in Sichuan Province

{kind=link}
{kind=link}