


光合作用涉及光能的吸收、传递和转化, 有两大色素参与了这一过程: 叶绿素(chlorophyll)和类胡萝卜素(carotenoid)。叶绿素中有一部分叶绿素a在光反应中心负责将光能转变为化学能, 其余叶绿素则负责光能的捕获和传递。类胡萝卜素主要行使两个功能: 光能捕获(light-harvesting)和光破坏防御(photoprotection)。光照不足时, 捕光天线色素蛋白复合体(light-harvesting antenna complex)增大, 光能利用效率增加; 光照过剩时, 捕光天线色素蛋白复合体则会启动热能耗散(thermal energy dissipation), 来减轻光氧化的伤害(Demmig-Adams & Adams, 2000, 2006; Matsubara et al., 2008)。植物色素通过调整其构成和含量来适应波动的光照条件, 因此, 我们可以通过测定色素含量和构成的变化来衡量植物对不同光照条件的适应能力。当前, 已有大量研究报道了不同种类植株色素构成的差异(Murchie & Horton, 1997; Demmig-Adams & Adams, 2006), 然而对这种差异形成的原因还缺乏更深入的探讨。
遮阴成为城市中低矮植物生存所面临的一个严峻的问题。据调查, 城市中至少有1/4的植被处于遮阴生境中, 遮阴源包括建筑物、阔叶树、针叶树以及建筑用遮阴网等(Bell et al., 2000; Jiang et al., 2004)。遮阴对环境的改变包括光质和光量的变化、空气湿度和风力的变化、温度的变化以及落叶对植被的覆盖和两种植物之间的交感作用等(Bell et al., 2000; Koh et al., 2003; Jiang et al., 2004)。既然光照是植物生长所必需的, 那么在光照条件改变以后, 植物如何通过改变色素含量和构成来适应遮阴生境是一个值得关注的问题。植物生理学家们通过研究不同光照条件下植物的色素构成来探究不同色素在光合作用中所起的作用, 从而我们可以根据植物中色素的构成和分布特点, 推测植物对不同光照条件的适应程度, 以确定出不同光照环境中适合生长的植物类型。
近年来, 随着生物物理学与生物化学方法的完善, 科学家们应用X射线晶体学方法进一步确定了叶绿素和类胡萝卜素在叶绿体中三大蛋白复合体 ——光系统I (photosystem I, PSI)、光系统II (photosystem II, PSII)和捕光天线蛋白复合体中的具体结合位点及含量(Ferreira et al., 2004; Liu et al., 2004; Amunts et al., 2007), 应用飞秒时间分辨差异吸收光谱技术确定了部分色素的功能(Holt et al., 2005)。这些新发现为色素研究提供了进一步的理论依据。在本文中, 我们总结了由植物自身因素决定的光合色素含量和构成, 以及外部遮阴环境对色素构成的影响。指出现存工作中存在的不足和亟待解决的问题以及未来研究的方向, 以期望读者对色素研究有较全面的了解, 为开展进一步的研究工作提供思路。
1 植物体内的色素构成
叶绿素包括叶绿素a (chlorophyll a, Chl a)和叶绿素b (chlorophyll b, Chl b)。类胡萝卜素包括2大类: 一类为含氧的类胡萝卜素, 如紫黄质(violaxanthin, V)、花药黄质(antheraxanthin, A)、玉米黄质(zeax- anthin, Z)、黄体素(lutein, L)、环氧化黄体素(lutein epoxide, Lx)和新黄质(neoxanthin, N)等, 统称为叶黄质(xanthophylls, X); 另一类为不含氧的类胡萝卜素, 如β-胡萝卜素(β-carotene, β-Car)、α-胡萝卜素(α-carotene, α-Car)、八氢番茄红素和番茄红素等, 统称为胡萝卜素(carotene)。植物中发挥功能的6种主要的类胡萝卜素包括: 紫黄质、花药黄质、玉米黄质、黄体素、新黄质和β-胡萝卜素(Bell & Danneberger, 1999; McElroy et al., 2006)。
光合作用主要在叶绿体中进行, 叶绿体内是类囊体(thylakoid)和基质(stroma)。所有进行光合作用的色素均位于类囊体膜的蛋白复合体中, 而90%的色素结合在PSI、PSSII以及PSII的捕光色素蛋白复合体(light harvesting complex II, LHCII)中。
PSI和PSII的光反应中心复合体(reaction center complex)主要结合叶绿素a和β胡萝卜素, 还有一部分叶绿素a位于LHCII中, 叶绿素b则主要位于LHCII中。阳生植物叶绿素a/b的比值约为3/1, 阴生植物约为2.3/1, 表明阴生植物通过提高叶绿素b的含量来捕获更多的光能。结合于LHCII中的黄体素是植物体内的主要类胡萝卜素, 在阴生植物中可占整个类胡萝卜素比例的48%-51%, 阳生植物中占到35%-45% (林植芳等, 2006)。黄体素也是进行蛋白折叠所必需的(Dall’Osto et al., 2006), 黄体素既为吸收光能的辅助色素, 可以完全将激发能传递给叶绿素a (Liu et al., 2004), 又可在持续胁迫条件下与玉米黄质一起进行热能耗散(Demmig-Adams & Adams, 2006, Dall’Osto et al., 2007)。玉米黄质是植物体中进行热能耗散的主要类胡萝卜素, 它是由紫黄质在强光条件下转化而成的, 中间产物为花药黄质, 这三者之和统称为叶黄素库(xanthophyll cycle pool size, V + A + Z)。在短时期内, 植物叶黄素库的大小是维持不变的(Demmig-Adams et al., 1996)。玉米黄质可促进非光化学猝灭(non photochemical quenching, NPQ), 保护类囊体膜脂, 防止其过氧化, 减少PSII捕光天线的大小。紫黄质则在不存在非光化学猝灭时发生积累, 但又有报道证明紫黄质可以猝灭三线态的叶绿素(3Chl*), 紫黄质的积累对植株体内的光破坏防御是有利的(Davison et al., 2002), 从这两点看来紫黄质的作用是比较有争议性的(Dall’Osto et al., 2007)。新黄质主要在PSII的LHCII中存在, Hobe等(2006)曾指出位于LHCII三聚体上的新黄质其实可有可无, 其主要作用是固定LHCII三聚体, 而不是结构上必需的。新黄质在LHCII三聚体中存在时可以捕光, 将能量传递到叶绿素b (Liu et al., 2004); 光能过剩时新黄质从LHCII三聚体脱落, 可以猝灭单线态的氧(1O2*) (Liu et al., 2004)和超氧阴离子(O2-) (Dall’Osto et al., 2007), 最后生成脱落酸(Hobe et al., 2006)。β-胡萝卜素主要位于光反应中心, 在光系统I的捕光天线中也有存在, β-胡萝卜素也是重要的光破坏防御色素 (Munne-Bosch & Penuelas, 2003)。
2 阴生植物与阳生植物的色素含量和构成
Murchie和Horton (1998)曾依据植物最适生长的光照条件将植物分为3类: 阴生植物、阳生植物和中间型植物。然而大多数物种都不是绝对的阴生或阳生植物, 而是属于中间型植物。Rosevear等(2001)通过对23个物种的色素含量构成的分析指出影响色素构成的最主要的因素不是物种的生态来源, 而是其生长的条件。由此可见, 以前将植物简单分为阴生和阳生植物是不全面的。但在本文中为区分物种差异, 将植物暂且用阴生和阳生区分。
叶绿素含量和叶绿素a/b常常用来区分阴生和阳生植物。阴生植物单位叶面积上的叶绿素含量较高, 叶绿素a/b则较低。Johnson等(1993)通过对19个物种的测定, 报道了阳生植物的叶绿素a/b为2.91 ± 0.08, 而阴生植物的叶绿素a/b为2.59 ± 0.11。叶绿素含量是由叶片厚度和叶绿体数两个方面决定的, 弱光条件下叶绿素含量增多是为了捕获更多的光能, 然而是否所有植物的色素变化对光照的响应是一样的?答案是否定的。Johnson等(1993)在试验中没有发现色素含量与植物接受的光照条件的相关性, 指出不能以色素含量来确定植物来源, 这一结果与Thayer和Björkman (1990)对10种阳生植物和9种阴生植物的研究结果相一致。Murchie和Horton (1998)的试验结果表明绝对的阴生植物其色素含量和构成都是不随光照条件变化的, 阳生植物通过增加叶绿素含量来适应弱光条件, 中间型植物则是既改变叶绿素含量也改变叶绿素构成来适应光照变化。
与阴生植物相比, 阳生植物类胡萝卜素/叶绿素的比值更高, 叶黄素库也更大。Thayer 和Björk- man (1990)通过对10种阳生植物和9种阴生植物的比较指出, 无论是在叶面积还是叶绿素含量的基础上, 阳生植物的叶黄素库均为阴生植物的2-4倍。但正午时分玉米黄质的积累与叶黄素库的大小无关, 而是与光合速率相关的。Demmig- Adams和Adams (1992)指出玉米黄质的转化率与光合速率密切相关, 光合速率高的一年生农作物在正午将30%-50%的叶黄素库的色素转变为玉米黄质, 而光合速率较低的多年生灌木和攀援植物则将80%以上的叶黄素库色素转变为玉米黄质, 由此可见, 不是叶黄素库越大, 玉米黄质的转化率越高。光合速率最低的多年生灌木和攀缘植物在正午时分积累的玉米黄质最多, 积累的玉米黄质可以进行光的破坏防御。Demmig-Adams 和Adams (2006)还报道了光合速率高、快速生长的农作物将更多的太阳能用于光合作用和生长, 而利用叶黄素循环进行热能耗散的能力则不及光合速率低、生长慢的常绿植物。光合速率还影响其他色素的构成, 通常光合速率较高的阳生植物叶黄素库和β-胡萝卜素的含量均较高, 而黄体素的含量则偏低(Demmig-Adams & Adams, 1992)。
Johnson等(1993)通过对22种植物的比较发现黄体素和叶黄素库基本为负相关的关系, 黄体素在整个类胡萝卜素中的比例随着植物耐阴性的增加而增加, 而叶黄素库的相对含量随着耐阴性的增加而降低, 由此试以黄体素/叶黄素库来指示植物的耐阴性, 发现黄体素/叶黄素库与植物耐阴性呈高度正相关关系。阳生植物中β-胡萝卜素的含量变化与叶黄素库相似, 因为它们均为β, β-胡萝卜素, 而黄体素和α-胡萝卜素为β,ε-胡萝卜素(图1)。阴生植物中β-胡萝卜素与叶绿素的比值低于阳生植物(Hansen et al., 2002a)。阴生植物的β,ε-胡萝卜素含量较高, 有的含有较多α-胡萝卜素, 而阳生植物中则以β, β-胡萝卜素为主, 仅含少量的α-胡萝卜素, 或根本没有(Thayer & Björkman, 1990; Demmig-Adams & Adams, 1992; García-Plazaola & Becerril, 2000)。Matsubara等(2009)通过对86个新热带区物种的调查研究发现, 阴生叶和阳生叶中β, ε-胡萝卜素/叶绿素(a + b)的比值相似, 而β, β-胡萝卜素, 则是在阳生叶中含量较多。有报道证明新黄质的相对含量一般是不随光照强度而变化的(Johnson et al., 1993), 但Hansen等(2002b)发现在同样的光照条件下, 阴生植物松树中新黄质和黄体素与叶绿素的比值高于阳生植物橡树, 这一结果与Matsubara等(2009)的观察结果相似, 原因是新黄质和黄体素一样位于捕光色素天线复合体中, 行使光能捕获或光破坏防御的功能。可见阴生植物中含较多的新黄质以进行光能捕获。
图1
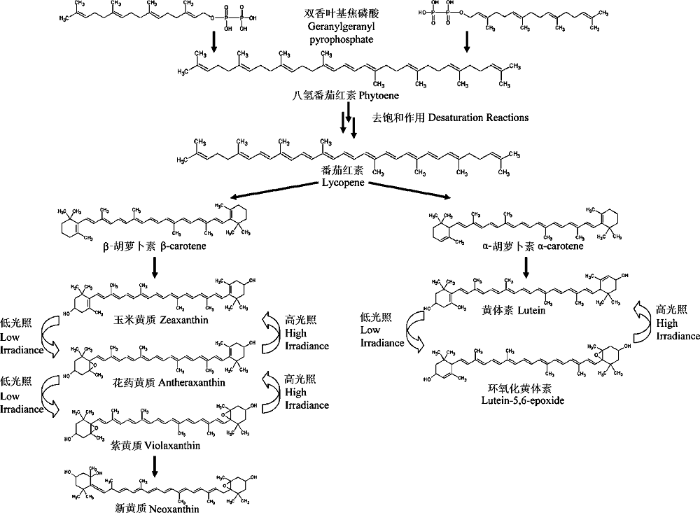
图1
参与光破坏防御与捕光作用的主要的类胡萝卜素的转化流程图(引自McElroy et al., 2006, 略有改动)。
Fig. 1
Schematic of conversion pathway of primary plant carotenoids responsible for photoprotection and light harvesting (adapted from McElroy et al., 2006).
3 遮阴对植株叶片色素含量和构成的影响
3.1 不同遮阴源对光强和光质的影响
遮阴不仅会降低光照强度, 也会改变光谱组成。
自然界中的可见光, 也就是有助于植物生长的光主要由直射、散射和反射的光线组成。遮阴一般会造成红光比例降低, 蓝光比例增高。这是因为红光主要来源于直射, 遮阴挡住了直射光源导致红光比例降低; 而蓝光的波长较短, 与大气中悬浮物的分子大小相似, 导致蓝光很容易被散射, 散射的蓝光可占到蓝光总量的25% (Bell et al., 2000), 遮阴使蓝光大量聚集到遮阴处。
Bell等(2000)对不同遮阴来源下的光谱构成做了6个月(春分至秋分)连续测定, 全面揭示了由遮阴源导致的光强和光质的差异。通常试验所用的遮阴网只能降低光照强度, 并不能改变光谱组成(Wherley et al., 2005)。在遮阴条件(树木和建筑物遮阴)下, 蓝光比例增高, 红光比例降低(图2)。而且随着遮阴程度的加重, 蓝光比例愈加增高, 红光比例愈加降低。建筑物下的远红光比例低于落叶树和针叶树(图2), 这是因为远红光能够穿透树冠, 却不能穿透建筑物。总体来说, 建筑物遮阴生境优于树木遮阴, 因为建筑物遮阴下的蓝光/远红光的比值最高, 红光/远红光的比值仅次于全日照条件。针叶树遮阴下的光谱组成是所有遮阴中对植物生长最不利的, 两大进行光合作用的有效光(蓝光和红光)在光谱组成中的比例均最低。
图2
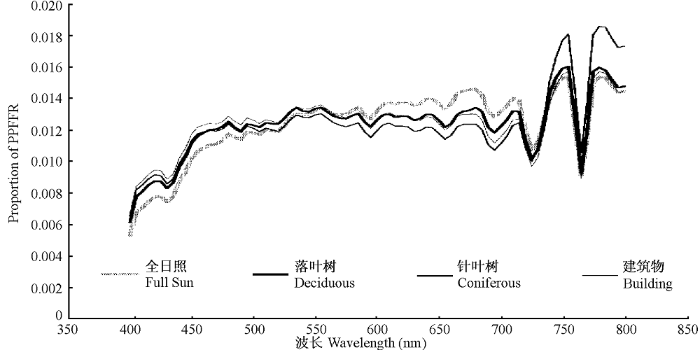
图2
落叶树遮阴、针叶树遮阴、建筑物遮阴和正常光照下不同波长的光占光合光量子与远红外光光量子之和的比例(测定地点: 美国俄亥俄州哥伦布市) (引自Bell et al., 2000, 略有改动)
Fig. 2
Proportion of photosynthetic photon flux plus far-red quanta (PPFFR; μmol·s-1·m-2) under deciduous shade, coniferous shade, building shade and full sun in Columbus, Ohio, USA (Adapted from Bell et al., 2000).
图3
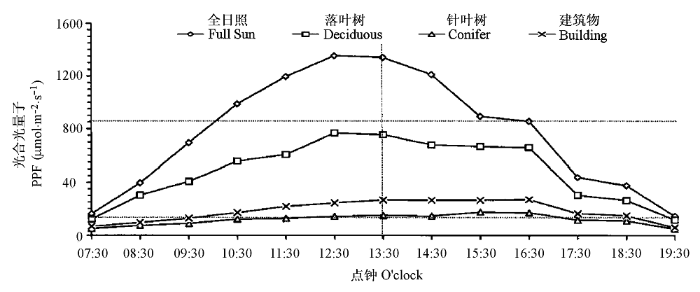
图3
落叶树遮阴、针叶树遮阴、建筑物遮阴和正常光照下的光合光量子的日变化动态图(测定地点: 美国俄亥俄州哥伦布市); 在150和867 μmol·m-2·s-1的横线代表多年生黑麦草的光补偿点和光饱和光强(引自Bell et al., 2000, 略有改动)。
Fig. 3
Diurnal photosynthetic photo flux (PPF) under deciduous shade, coniferous shade, building shade and full sun in Columbus, Ohio, USA. Horizontal lines at 150 and 867 μmol·m-2·s-1 represent the approximate light compensation and saturation points of perennial ryegrass (Adapted from Bell et al., 2000).
3.2 遮阴下光照强度的改变对色素含量和构成的影响
类胡萝卜素能够执行光能捕获和光破坏防御的功能主要是因为类胡萝卜素参与了两大循环, 也就是执行不同功能的类胡萝卜素会在不同的光照强度下进行相互转化。其中比较著名的, 也是所有高等植物中均存在的一个循环为叶黄素循环(xanthophyll cycle, V cycle), 另外一个循环仅在热带雨林的少数种属中存在, 为黄体素循环(Lx-L cycle)。强光照条件下, 当光能过剩时, 在低pH值(5.2), 紫黄质脱环氧化酶(violaxanthin de-epoxidase, VDE)的催化下, 紫黄质去环氧化经由花药黄质变为玉米黄质, 此转化只要几分钟即可完成; 弱光条件下, 当光能有限时, 玉米黄质在中性pH值(7.0- 7.5), 玉米黄质环氧化酶(zeaxanthin epoxidase, ZE)的催化下经花药黄质转变为紫黄质, 此转化需几小时或过夜才可完成。因此, 日出之前植物的去环氧化水平最高, 玉米黄质的含量也最低(Demmig- Adams & Adams, 1992)。叶黄素循环是20世纪50年代苏联科学家David Sapozhnikov最先发现的(Demmig-Adams et al., 1996), Thayer和Björkman (1990)与Gilmore和Yamamoto (1991)是首批使用高效液相色谱法对参与叶黄素循环的不同类胡萝卜素进行定量的科学家。鉴于Lx-L循环只在部分寄生植物、栎属(Quercus)、印加属(Inga)以及少数地衣中存在, 对此研究最多的科学家包括Matsubara等(2001, 2009, 2007, 2008)。光下, 环氧化黄体素去环氧化转变为黄体素, 此转化同紫黄质→玉米黄质的转化是同时进行的, 几分钟即可完成; 暗中, 黄体素转变为环氧化黄体素, 此转化很慢, 一般需要几天的时间, 不能像玉米黄质→紫黄质那样隔夜完成(day-night cycle), 因此正常的日夜交替是不会产生环氧化黄体素的。此循环被认为与叶黄素循环一样, 同样受紫黄质脱环氧化酶和玉米黄质环氧化酶的催化(García-Plazaola et al., 2007; Matsubara et al., 2008)。Matsubara等(2009)发现黄体素循环是印加属树木长寿命叶片适应不同光照条件的重要机制, 高的环氧化黄体素水平不仅可以帮助阴生叶捕获更多的光能, 也可以增强阳生叶的光破环防御功能。
对大多数植物来说, 低的光照强度引发的色素变化大致包括: 1)叶绿素含量上升; 类胡萝卜素/叶绿素的比值降低, 2)叶绿素a/b比值的降低, 3)叶黄素库变小, β-胡萝卜素在整个类胡萝卜素中的比例降低, 黄体素和新黄质的相对含量增加, 生长在持续弱光条件下的阴生植物中通常还具有很高的α-胡萝卜素, 可占到总量(α-胡萝卜素+ β-胡萝卜素)的23%-76%, 正常光照下植物体内的α-胡萝卜素含量则非常微小, 4)叶黄素库内紫黄质比例增加, 玉米黄质和花药黄质比例降低(Thayer & Björkma, 1990; Demmig-Adams & Adams, 1992; Johnson et al., 1993; Murchie & Horton, 1998; Atanasova et al., 2003)。
低的光照强度下叶绿素含量上升, 一方面是由于弱光减少了色素的光氧化伤害, 另一方面是由于叶绿体内的基粒变大, 基粒片层垛叠程度变高(Atanasova et al., 2003)。同一物种在不同的光照条件下色素含量及构成发生变化是有益还是无益?哪些变化是有助于适应光照变化的?哪些是不利的?例如遮阴条件下Chl a/b是否降低因品种而异(species-dependent)。一部分研究者认为Chl a/b降低是适应遮阴条件的表现(Baig et al., 2005), 而有的研究者则认为在不同的光照强度下, Chl a/b维持稳定是适应不同光照条件的表现(Thayer & Björkma, 1990; Johnson et al., 1993)。弱光条件下Chl a/b降低的解释通常为: 1)弱光条件导致LHCII的数量增加, LHCII中既有Chl a也有Chl b, 光反应中心则只有Chl a, 因此植株总的Chl a/b降低是为了在弱光下捕获更多的光能, 2)弱光条件下, PSI和PSII密度均降低, 但PSI降低的幅度远远大于PSII (Eichelmann et al., 2005); 同时垛叠类囊体的数量增加(Bertamini et al., 2006), PSII主要集中分布于垛叠的类囊体膜上, PSI则分布于非垛叠的类囊体膜上, 这些原因导致总的PSII/PSI比值增加, 而PSII中Chl a/b比值低于PSI, 因此总体Chl a/b比值降低。Chl a/b值维持不变的解释则为植物可以通过在弱光条件下降低反应中心的电子传递速率和Rubisco (核酮糖-1, 5-二磷酸羧化酶/加氧酶)活性来降低植物对光的需求, 这样无需增大捕光天线, Chl a/b比值可以维持不变, 同样可以适应弱光的条件(Murchie & Horton, 1998; Maxwell et al., 1999)。因此, 弱光条件下植物是通过调整色素比例捕获更多的光能来维持原有的光合速率, 还是通过降低光合速率来适应弱光, 哪一个效率更高?这是一个值得探讨的问题。有研究表明高羊茅(Festuca arundinacea)比早熟禾(Poa pratemsis)和黑麦草(Lolium perenne)更耐阴的原因在于高羊茅通过降低光合速率和呼吸速率来适应遮阴的条件, 而不是通过调整叶绿素含量来提高耐阴性(Jiang et al., 2004)。
3.3 遮阴下光质的改变对色素含量和构成的影响
图4
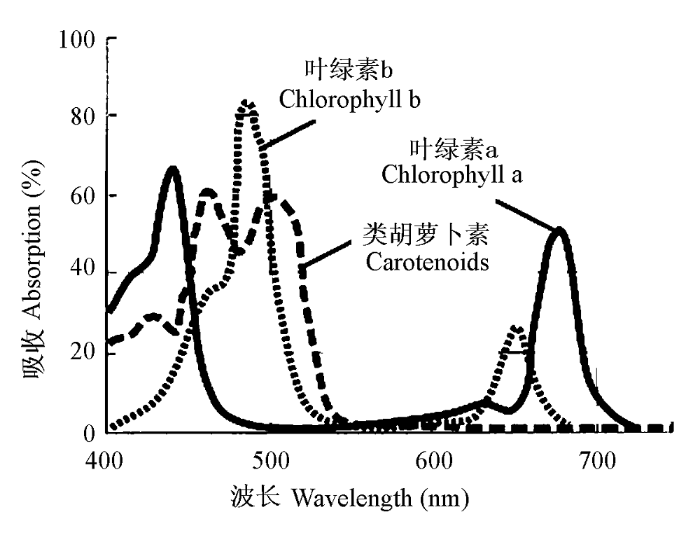
图4
叶绿素a、叶绿素b和类胡萝卜素的吸收波长。
来源:
Fig. 4
Absorption spectra of chlorophyll a, chlorophyll b and carotenoids.
Source:
传统观点认为蓝光培养的植株具有阳生植物的特点, 类胡萝卜素占的比例较高; 而红光培养的植株具有阴生植物的特点, 叶绿素含量较高, Chl a/b则较低(Leong et al., 1985; 储钟稀等, 1999; 许莉等, 2007)。白光为蓝光、绿光和红光的混合光, 其光质最接近自然光。马建忠等(1997)通过对黍(Panicum miliaceum)幼苗和储钟稀等(1999)对黄瓜(Cucumis sativus)的研究均证明白光处理的叶片叶绿素含量为最高, 其次为红光, 最后为蓝光。马建忠等(1997)还发现生长在远红外光下的黍, 稍有叶绿素积累, 但含量很低。对此现象的解释是虽然启动叶绿素合成的有效波长以蓝光为最优, 但在后续反应中, 红光处理的效果要高于蓝光, 可能是因为红光引发的作用范围更大。但储钟稀等(1999)发现Chl a/b以蓝光处理为最高, 红光处理为最低; 而马建忠等(1997)则发现红光和蓝光的Chl a/b均高于白光, 表明两种单色光在Chl a到b的转换方面没有白光有效。许莉等(2007)观测到叶用莴苣的叶绿素含量以红光处理为最高, 同时Chl a/b以红光处理为最低, 其次为白光, 而蓝光的叶绿素含量为最低, Chl a/b最高。Leong等(1985)则发现红光下生长的蕨类植物南洋山苏花(Asplenium australasicum)类囊体膜中叶绿素含量最低, Chl a/b最高, LHCII的量也比蓝光下少; 蓝光下的叶绿素含量最高, Chl a/b最低; 白光处理下的结果介于两者之间。由此可以看出1)叶绿素含量的增多通常伴随着Chl a/b的降低, 或者我们可以说, 叶绿素含量增多, 主要是Chl b含量增多的缘故(Atanasova et al., 2003)。2)作为混合光的白光, 其合成叶绿素的光质效应是介于蓝光和红光之间, 还是最优, 仍有异议。这是否与物种有关, 值得进一步探讨。3)就红光和蓝光的光质效应与物种的关系, Leong等(1985)曾对其进行了解释, 指出红光和蓝光的光质效应在一定的光照强度下相似, 超过这个光强, 蓝光光质效应高于红光; 低于这个光强, 蓝光光质效应低于红光。而对于不同的植物, 这个临界点是不同的, 特别是对于蕨类这种长期生长在遮阴环境下的植物, 此临界点较低。由此可见光质和光质效应产生的混合效果因物种而异。目前对于叶绿素和Chl a/b的变化趋势与红蓝光的关系还没有更完善的解释。
红光和蓝光等单色光对类胡萝卜素含量和构成的影响的研究还未见报道, 但因为PSI和PSII中的色素构成是有差异的, 因此可以根据以下原则进行推测: 红光照射下, PSII吸收的光能多, PSI吸收的光能少, 致使PSII/PSI的比值降低; 远红光照射下, PSI吸收的光能变多, 而PSII吸收的光能变少, 致使PSII/PSI的比值增加, 同时在远红外光照射下, 电子传递速率变高, PSII光反应中心叶绿素蛋白复合体的相对含量增加, 而LHCII与光反应中心蛋白复合体的比值降低(Glick et al., 1985)。Thayer和Björkman (1992)通过对棉花(Gossypium hirsutum)叶片和玉米(Zea mays)维管束鞘的类囊体色素蛋白复合体中PSII和PSI中色素构成的研究发现: 1) PSI中Chl a/b远远高于PSII, 棉花为8.4/1.9, 玉米为12.1/2.7; 2)叶绿素均匀分布于PSI和PSII中; 3)大约有2%的新黄质、18%的黄体素、30%的叶黄素库色素和76%的β胡萝卜素存在于PSI中。然而这种光系统之间的比例除受红光和远红光比例的影响, 还受其他因素的影响。Kim等(1993)证明在缺乏Chl b的大麦(Hordeum vulgare)突变体中, 这种光系统之间的转化是不存在的, 表明辅助色素(accessory pigment)在光系统中充当光质的感应分子将信号传达给光系统, 而光系统本身只是作为一个信号传达的途径(signal transduction pathway)。
红光/远红光的比值和红光/蓝光的比值也常用来作为光质区别的一个指标。Wherley等(2005)在同样的遮阴梯度下, 比较了遮阴网遮阴和落叶树遮阴的区别, 发现遮阴网遮阴下高羊茅植株体内的叶绿素含量更高, 于是作者认为是由于遮阴网遮阴下高的红光/远红光比值提高了高羊茅叶绿素的含量。然而Bradburne等(1989)则发现红光/远红光的比值越低, 叶绿素的含量越高, 捕光色素蛋白复合体浓度也越高, 而Chl a/b的比值越低。因此我们推测遮阴网遮阴下叶绿素含量高于落叶树遮阴的原因不是高的红光/远红光比值, 而可能是红光/蓝光的比值或是红光、蓝光和远红光共同作用的结果。
Ramalho等(2002)使用3种不同波长范围及不同红光/远红外光比值的灯源给植物提供光照, 结果发现, 以波长范围为425-650 nm, 红光/远红外光的比值为2左右的接近自然界中比值的HQI-BT灯下植物各项光合指标为最佳。由此我们可以看出光合作用或色素构成是不同比例的光质共同作用的结果, 而不是哪一种单色光所能替代的。
3.4 遮阴时间对色素含量和构成的影响
色素含量一般随遮阴时间的延长而出现由高到低, 或由低到高的变化, 而不会是持续增加或持续降低, 是因为色素对光照的变化需要一个适应的时间。刚刚置于遮阴生境中的草坪草叶绿素含量先降低, 而后逐渐升高(Bell & Danneberger, 1999)。在由强光(47.9 mol·m-2·d-1)转入弱光(4.7 mol·m-2·d-1)的过程中, 匍匐剪股颖(Agrostis stolonifera)体内的紫黄质、新黄质、黄体素和β-胡萝卜素在0-26 h内增加, 24-168 h后逐渐降低, 而玉米黄质和花药黄质的含量则持续降低; 在由弱光转入强光的过程中, 新黄质和黄体素的含量是先升高后降低的, 紫黄质和β-胡萝卜素的含量持续降低, 而玉米黄质和花药黄质含量持续上升(McElroy et al., 2006)。
每天至少保证4-5 h的光照时间对遮阴生境下植物的生长是很重要的。Bell和Danneberger (1999)发现每天接受6 h遮阴的匍匐剪股颖的草坪草质量与全日照条件没有明显差异。Jiang等(2004)也报道在90%光照的条件下每天对海滨雀稗(Paspalum vaginatum)施加5 h光照, 其质量与全日光条件下没有什么差别。日间的光量子分布梯度对色素变化也有很重要的影响, 自然光照下生长的植物的各项指标明显更优。Thayer和Björkman (1990)通过对棉花子叶施加人工光照和自然光照, 发现在人工光照下, (V + A + Z)/Chl (a + b)的比值是随着光强的增加几乎线性增长, 而在自然光照下, (V + A + Z)/Chl (a + b)的比值随着光强的增加而呈曲线增长, α胡萝卜素的含量也是在自然光照下减少得更快, 自然光照能够更有效地增大叶黄素库的大小, 更有效地降低α胡萝卜素的含量。这说明(V + A + Z)的库容量大小不仅与每天接收的光量子多少有关, 而且与光量子在一天的分布时间有关, 也就是色素含量是光照和温度协同作用的结果。
4 色素构成研究中存在的不足及展望
植物体内的色素构成的不同是多种因素共同影响的结果。本文在综述近20年有关色素构成的研究中发现许多研究者对于试验结果缺乏深入的原理解释, 对于试验中出现的与前人的研究结果不符的现象往往用物种差异一笔带过。物种差异固然存在, 但科学研究的目的在于将现象与理论相结合, 将宏观的植物科学研究中观测到的现象尽量与微观的理论研究结合起来。如近几年, 科学家们通过应用X射线晶体学的方法确定了色素在PSI和PSII及其外周天线色素蛋白复合体LHCII中的位置和含量, 高等植物豌豆(Pisum sativum var. alaska)的PSI中共有168个叶绿素, 其中光反应中心有100个左右, 外周LHCII中有56个(32 Chl a + 24 Chl b), 类胡萝卜素则至少应有20个(Amunts et al., 2007)。如此说来, PSI的Chl a/b应为5至6之间。蓝细菌PSII为一个二聚体, 每个单体的内反应天线蛋白CP43和CP47分别结合了13和16个叶绿素, 加之6个绑定于D1/D2蛋白的叶绿素, 共计35个Chl a, 还有11个β-胡萝卜素(Loll et al., 2005)。遗憾的是, 对高等植物中光系统II的色素构成还没有应用X射线晶体学的方法进行试验报道。至于PSII的色素天线蛋白复合体LHCII则是以三聚体形式存在的, 每个单体中含有8个Chl a, 6个Chl b, 也就是LHCII中Chl a/b比值应为1.3左右(Liu et al., 2004)。按照捕光色素蛋白复合体LHCII的数量庞大, 其所含Chl a可占到Chl a总量的一半, 大致可以推算植物Chl a/b比值变化与PSII和PSI比值之间的关系。对于与现存理论中不符的现象, 则应找出问题的根源。如传统上认为的蓝光培育的植物可类比于阳生植物, 而红光培育的植物可类比于阴生植物, 实际上红光照射下, PSII吸收的光能增多, 而PSI吸收的光能变少, 致使PSII/PSI降低, 而PSI中Chl a/b远远高于PSII (Thayer & Björkman, 1992), PSII/PSI降低实际也就伴随着Chl a/b的升高, 这与阴生植物的Chl a/b较低的事实明显不符。
叶绿素和类胡萝卜素在光合作用中起着不可替代的作用。遮阴引起遮阴小生境中光强、光质和光照时间的变化。光照条件的变化对植株内色素构成的影响主要是通过调控光反应中心和捕光色素蛋白复合体的比值以及PSII和PSI的比值来实现的。叶绿素和类胡萝卜素所处的位置不同, 功能也不同。我们可以根据不同的遮阴生境中的光质和光量特点, 结合植物自身的生长习性及其色素的含量和构成特点确定适合不同遮阴生境中生长的植物。这方面的研究不仅需要对同一物种在不同的光照条件下的色素构成进行比较, 也需要对大量物种在同一光照条件下的表现差异进行分析。因为色素含量和构成是多方面协调作用的结果, 从植物自身的遗传特征, 到叶龄差异, 再到光强和光质的双重影响。随着生物化学和生物物理学的迅猛发展, 我们需要结合这些微观上不断完善的基础理论知识, 分析在实际植物科学研究发现的问题; 同时也要将实际观测到的现象与已存的理论知识进行比较, 进一步完善微观理论科学研究中的结论。这两个方面需要共同发展, 互相补充。
致谢
国家科技支撑计划项目(2006BAD16B09-2)资助。
参考文献
The structure of a plant photosystem I supercomplex at 3.4 Å resolution
Comparative characteristics of growth and photosynthesis of sun and shade leaves from normal and pendulum walnut (Juglans regia L.) trees
Irradiance influences contents of photosynthetic pigments and proteins in tropical grasses and legumes
Temporal shade on creeping bentgrass turf
Spectral irradiance available for turfgrass growth in sun and shade
Shade effect alters leaf pigments and photosynthetic responses in Norway spruce (Picea abies L.) grown under field conditions
Reflected far-red light effects on chlorophyll and light-harvesting chlorophyll protein (LHC-II) contents under field conditions
Effect of different light quality on photosynthetic characteristics of cucumber leaves
Different roles of α- and β-branch xanthophylls in photosystem assembly and photoprotection
Lutein is needed for efficient chlorophyll triplet quenching in the major LHCII antenna complex of higher plants and effective photoprotection in vivo under strong light
Overexpression of [beta]-carotene hydroxylase enhances stress tolerance in Arabidopsis

Plant stress caused by extreme environmental conditions is already a principal reason for yield reduction in crops. The threat of global environment change makes it increasingly important to generate crop plants that will withstand such conditions. Stress, particularly stress caused by increased sunlight, leads to the production of reactive oxygen species that cause photo-oxidative cell damage. Carotenoids, which are present in the membranes of all photosynthetic organisms, help protect against such light-dependent oxidative damage. In plants, the xanthophyll cycle (the reversible interconversion of two carotenoids, violaxanthin and zeaxanthin) has a key photoprotective role and is therefore a promising target for genetic engineering to enhance stress tolerance. Here we show that in Arabidopsis thaliana overexpression of the chyB gene that encodes beta-carotene hydroxylase--an enzyme in the zeaxanthin biosynthetic pathway--causes a specific twofold increase in the size of the xanthophyll cycle pool. The plants are more tolerant to conditions of high light and high temperature, as shown by reduced leaf necrosis, reduced production of the stress indicator anthocyanin and reduced lipid peroxidation. Stress protection is probably due to the function of zeaxanthin in preventing oxidative damage of membranes.
Carotenoid composition in sun and shade leaves of plants with different life forms
Harvesting sunlight safely
Photoprotection in an ecological context: the remarkable complexity of thermal energy dissipation
Carotenoids 3: in vivo function of carotenoids in higher plants
Adjustment of leaf photosynthesis to shade in a natural canopy: reallocation of nitrogen
Architecture of the photosynthetic oxygen- evolving center
Photoprotection mechanisms in European beech (Fagus sylvatica L.) seedlings from diverse climatic origins
The lutein epoxide cycle in higher plants: its relationships to other xanthophyll cycles and possible functions
Several xanthophyll cycles have been described in photosynthetic organisms. Among them, only two are present in higher plants: the ubiquitous violaxanthin (V) cycle, and the taxonomically restricted lutein epoxide (Lx) cycle, whereas four cycles seem to occur in algae. Although V is synthesised through the beta-branch of the carotenoid biosynthetic pathway and Lx is the product of the alpha-branch; both are co-located in the same sites of the photosynthetic pigment-protein complexes isolated from thylakoids. Both xanthophylls are also de-epoxidised upon light exposure by the same enzyme, violaxanthin de-epoxidase (VDE) leading to the formation of zeaxanthin (Z) and lutein (L) at comparable rates. In contrast with VDE, the reverse reaction presumably catalysed by zeaxanthin epoxidase (ZE), is much slower (or even inactive) with L than with antheraxanthin (A) or Z. Consequently many species lack Lx altogether, and although the presence of Lx shows an irregular taxonomical distribution in unrelated taxa, it has a high fidelity at family level. In those plants which accumulate Lx, variations in ZE activity in vivo mean that a complete Lx-cycle occurs in some (with Lx pools being restored overnight), whereas in others a truncated cycle is observed in which VDE converts Lx into L, but regeneration of Lx by ZE is extremely slow. Accumulation of Lx to high concentrations is found most commonly in old leaves in deeply shaded canopies, and the Lx cycle in these leaves is usually truncated. This seemingly anomalous situation presumably arises because ZE has a low but finite affinity for L, and because deeply shaded leaves are not often exposed to light intensities strong enough to activate VDE. Notably, both in vitro and in vivo studies have recently shown that accumulation of Lx can increase the light harvesting efficiency in the antennae of PSII. We propose a model for the truncated Lx cycle in strong light in which VDE converts Lx to L which then occupies sites L2 and V1 in the light-harvesting antenna complex of PSII (Lhcb), displacing V and Z. There is correlative evidence that this photoconverted L facilitates energy dissipation via non-photochemical quenching and thereby converts a highly efficient light harvesting system to an energy dissipating system with improved capacity to engage photoprotection. Operation of the alpha- and beta-xanthophyll cycles with different L and Z epoxidation kinetics thus allows a combination of rapidly and slowly reversible modulation of light harvesting and photoprotection, with each cycle having distinct effects. Based on the patchy taxonomical distribution of Lx, we propose that the presence of Lx (and the Lx cycle) could be the result of a recurrent mutation in the epoxidase gene that increases its affinity for L, which is conserved whenever it confers an evolutionary advantage.
Resolution of lutein and zeaxanthin using a non-endcapped, lightly carbon-loaded C18 high-performance liquid chromatographic column
Effect of light quality on chloroplact-membrane organization and function in pea
Variation of pigment composition and antioxidative systems along the canopy light gradient in a mixed beech/oak forest: a comparative study on deciduous tree species differing in shade tolerance
Is the lower shade tolerance of Scots pine, relative to pedunculate oak, related to the composition of photosynthetic pigments?
Assembly of the major light-harvesting chlorophyll-a/b complex: thermodynamics and kinetics of neoxanthin binding
Carotenoid cation formation and the regulation of photosynthetic light harvesting
Assessment of low light tolerance of seashore paspalum and bermudagrass
Relationships between carotenoid composition and growth habit in British plant species
Dynamics of photosystem stoichiometry adjustment by light quality in chloroplasts
Shade and airflow restriction effects on creeping bentgrass golf greens
Effect of light quality on the composition, function, and structure of photosynthetic thylakoid membranes of Asplenium australasicum (Sm.) Hook
Lutein and lutein epoxide-lutein cycle in plants
Crystal structure of spinach major light-harvesting complex at 2.72Å resolution
The major light-harvesting complex of photosystem II (LHC-II) serves as the principal solar energy collector in the photosynthesis of green plants and presumably also functions in photoprotection under high-light conditions. Here we report the first X-ray structure of LHC-II in icosahedral proteoliposome assembly at atomic detail. One asymmetric unit of a large R32 unit cell contains ten LHC-II monomers. The 14 chlorophylls (Chl) in each monomer can be unambiguously distinguished as eight Chla and six Chlb molecules. Assignment of the orientation of the transition dipole moment of each chlorophyll has been achieved. All Chlb are located around the interface between adjacent monomers, and together with Chla they are the basis for efficient light harvesting. Four carotenoid-binding sites per monomer have been observed. The xanthophyll-cycle carotenoid at the monomer-monomer interface may be involved in the non-radiative dissipation of excessive energy, one of the photoprotective strategies that have evolved in plants.
Towards complete cofactor arrangement in the 3.0Å resolution structure of photosystem II
The effects of light quality on the development of chloroplast photosystems and the transcription of psbA gene of broomcorn millet (Panicum miliaceum)
Diurnal and acclimatory responses of violaxanthin and lutein epoxide in the Australian mistletoe Amyema miquelii
Sun-shade patterns of leaf carotenoid composition in 86 species of neotropical forest plants
Lutein epoxide cycle, light harvesting and photoprotection in species of the tropical tree genus Inga
Short- and long-term operation of the lutein-epoxide cycle in light-harvesting antenna complexes
Chloroplast acclimation in leaves of Guzmania monostachia in response to high light
Acclimation of leaves to high light (HL; 650 micromol m(-2) s(-1)) was investigated in the long-lived epiphytic bromeliad Guzmania monostachia and compared with plants maintained under low light (LL; 50 micromol m(-2) s(-1)). Despite a 60% decrease in total chlorophyll in HL-grown plants, the chlorophyll a/b ratio remained stable. Additionally, chloroplasts from HL-grown plants had a much lower thylakoid content and reduced granal stacking. Immunofluorescent labeling techniques were used to quantify the level of photosynthetic polypeptides. HL-grown plants had 30% to 40% of the content observed in LL-grown plants for the light-harvesting complex associated with photosystems I and II, the 33-kD photosystem II polypeptide, and Rubisco. These results were verified using conventional biochemical techniques, which revealed a comparable 60% decrease in Rubisco and total soluble protein. When expressed on a chlorophyll basis, the amount of protein and Rubisco was constant for HL- and LL-grown plants. Acclimation to HL involves a tightly coordinated adjustment of photosynthesis, indicating a highly regulated decrease in the number of photosynthetic units manifested at the level of the content of light-harvesting and electron transport components, the amount of Rubisco, and the induction of Crassulacean acid metabolism. This response occurs in mature leaves and may represent a strategy that is optimal for the resource-limited epiphytic niche.
Response of creeping bentgrass carotenoid composition to high and low irradiance
Photo-and antioxidative protection during summer leaf senescence in Pistacia lentvscus L. grown under mediterranean field conditions
Acclimation of photosynthesis to irradiance and spectral quality in British plant species: chlorophyll content, photosynthetic capacity and habitat preference
Contrasting patterns of photosynthetic acclimation to the light environment are dependent on the differential expression of the responses to altered irradiance and spectral quality
Photosynthetic performance and pigment composition of leaves from two tropical species is determined by light quality
Growth conditions are more important than species origin in determining leaf pigment content of British plant species
Leaf xanthophyll content and composition in sun and shade determined by HPLC
Carotenoid distribution and deepoxidation in thylakoid pigment-protein complexes from cotton leaves and bundle-sheath cells of maize
Tall fescue photomorphogenesis as influenced by changes in the spectral composition and light intensity
Effect of light quality on leaf lettuce photosynthesis and chlorophyll fluorescence

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}