


在被子植物中, 性比不仅受物种本身的遗传特性所制约(Bertin & Kerwin, 1998; Decker & Pilson, 2000; Volkova et al., 2007), 还与居群及个体大小、花在植株上的位置以及光照和温湿度等环境条件密切相关, 是遗传与环境因素互作的结果(Charnov, 1982; Lloyd & Bawa, 1984; Solomon, 1985; Bertin & Gwisc, 2002; Alonso, 2005)。在理想状态下, 居群性比的期望值为1﹕1 (Fisher, 1930), 但在环境非均质时, 性比会随环境梯度的变化而偏向于雄性或雌性(Lloyd, 1973; Charnov, 1982; Decker & Pilson, 2000)。相关研究表明, 在不同物种、居群和个体以及不同年份中, 植物的性比均可表现出一定程度的差异, 且这些差异可进一步影响其雌雄适合度和交配方式(Webb & Lloyd, 1980; Webb, 1981b; Thompson, 1987; Koul et al., 1989; Emms, 1993; Harder & Barrett, 1996; Decker & Pilson, 2000)。
伞形科是被子植物中性系统比较复杂的一个科, 具有雌雄同株和异株(Lloyd, 1973; Webb & Lloyd, 1980; Pickering, 2001)、雌全异株(Webb, 1981a; Caruso & Case, 2007)及雄全同株(Webb, 1979; Lovett Doust, 1980; Webb, 1981b)等性系统。由于受遗传、资源和环境等因素的影响, 其不同性系统的性比表现出多样化特点(Lloyd, 1973; Lovett Doust, 1980; Webb, 1979, 1980)。在具雄全同株性系统的伞形科植物中, 不同类群在性比上的差异对其交配方式及后代适合度具有重要影响(Webb, 1981b; Pickering, 2001; Davila & Wardle, 2007)。
簇花芹(Soranthus meyeri)隶属于伞形科簇花芹属, 主要分布于哈萨克斯坦和西西伯利亚地区, 在我国仅分布于新疆天山以北的沙漠地区, 是古尔班通古特沙漠中常见的多年生早春短命植物(秦雪梅和沈观冕, 1990; 钱亦兵等, 2007), 具有重要的固沙作用。据观察, 簇花芹植株上各伞形花序中雄花和两性花并存, 但不同大小的植株中以及植株不同位置伞形花序中的雄花和两性花数目均存在差异, 据此我们推测, 该物种可能存在大小依赖的性别分配。为了证实这一推测, 本文主要观测了簇花芹雄花和两性花的花部特征及其性比与植株大小的关系, 以及雄花和两性花着生位置与开花式样, 重点探讨了簇花芹花期大小依赖的性别资源分配, 以期为全面了解伞形科繁育系统的进化积累理论资料。
1 材料和方法
1.1 研究材料与地点
研究材料簇花芹主要生长在新疆北部古尔班通古特沙漠中的固定和半固定沙丘上。簇花芹株高30-70 cm, 基生叶呈莲座状, 茎上具有多个复伞形花序。每年3月中下旬积雪融化时, 进入生殖生长的植株地下芽开始萌动, 分化出多个复伞形花序, 且各伞形花序中均可分化出两性花和雄花, 至5月中下旬各伞形花序中的两性花和雄花陆续开放, 6月中下旬果实开始成熟。开花时, 其雄花和两性花均为辐射对称花, 萼齿5, 花瓣5枚离生, 外被短柔毛; 雄蕊5枚与花瓣互生, 花药为背着药, 纵向开裂。两性花雌蕊子房下位, 两室, 每室1枚胚珠; 花柱2, 基部常膨大为花柱基, 且与蜜腺盘愈合; 雄花无 雌蕊。
观测地点位于新疆克拉玛依市车牌子油田附近, 地理坐标为44°55′18.6″ N, 84°56′35.9″ E, 海拔300 m。此地属于古尔班通古特沙漠西南缘, 年平均气温6-10 ℃, 年积温3 000-3 500 ℃, 年降水量70-150 mm, 年蒸发量大于2 000 mm, 属于典型的温带大陆性干旱荒漠气候(钱亦兵等,2007)。本研究的所有实验均在2006-2008年进行。
1.2 研究方法
1.2.1 花部特征观测
在20个植株上分别选取即将开放的雄花和两性花各2朵, 将其分为两组(各20朵), 一组用于测量花部特征, 另一组以单花为单位将花药压碎, 用番红染液染色, 分别定容至1 mL制成花粉悬浮液, 取5 µL涂片, 并在Olympus BH-2型光学显微镜(Oly- mpus Corporation, Tokyo, Japan)下分别统计雄花和两性花的花粉数 n, 重复3次, 计算单花花粉数(n × 200)。同时, 在每个植株上随机选取即将开放的雄花和两性花各10朵(共计各200朵), 带回实验室置于80 ℃烘箱中烘干(48 h)至恒重, 每次随机取15朵用Sartorius BT25S电子天平(精度为0.01 mg)称重, 重复10次, 计算雄花与两性花的生物量。
1.2.2 性比与植株大小的关系
1.2.3 开花式样
1.2.4 数据分析
利用SPSS 11.0统计分析软件对数据进行分析。用单因素方差分析(one-way ANOVA)法分析不同植株上伞形花序数以及花序上雄花和两性花数目间的差异, 用Independent-Sample T Test对雄花和两性花的花部特征进行分析, 用Bivariate Correlations进行本实验中所有的相关性分析。曲线图使用Excel 2003软件绘制。
2 研究结果
2.1 花部特征
簇花芹的两性花大于雄花, 且二者在花瓣长、花丝、花药长、花粉量以及花生物量等花部特征上存在极显著差异(p< 0.01) (表1), 说明生产两性花所需资源大于生产雄花的。
表1 簇花芹雄花和两性花的花部特征(平均值±标准误差, n = 20)
Table 1
| 花类型 Floral morph | 花瓣长 Length of petal (mm) | 雄蕊 Stamen (mm) | 雌蕊 Pistil (mm) | 花粉数量 Number of pollen grains | 花生物量 Biomass per flower (mg) | ||
|---|---|---|---|---|---|---|---|
| 花丝长 Length of filament | 花药长 Length of anther | 子房长 Length of ovary | 花柱长 Length of style | ||||
| 两性花 Hermaphrodite flower | 3.15 ± 0.05 | 3.09 ± 0.05 | 0.92 ± 0.01 | 1.88 ± 0.43 | 0.72 ± 0.05 | 21 633 ± 634 | 12.10 ± 0.28 |
| 雄花 Staminate flower | 2.89 ± 0.04 | 2.83 ± 0.04 | 0.85 ± 0.01 | - | - | 18 312 ± 454 | 5.37 ± 0.16 |
| F | 16.99** | 14.43** | 27.26** | - | - | 13.06** | 425.51** |
*,p< 0.05; **,p< 0.01.
2.2 性比与植株大小的关系
2006-2008年, 簇花芹群体水平的性比分别为0.69 ± 0.03、0.62 ± 0.03和0.69 ± 0.02, 彼此间无显著性差异( p> 0.05), 说明不同年份中簇花芹群体水平的性比均接近于2:3, 是相对稳定的, 环境条件对其影响不大。
2007-2008年, 簇花芹的两性花数与植株生物量呈正相关关系(p < 0.01), 生物量大的植株两性花多, 但雄花数与植株生物量不具相关性( p > 0.05) ( 图1), 说明植株大小对两性花数影响较大。
图1
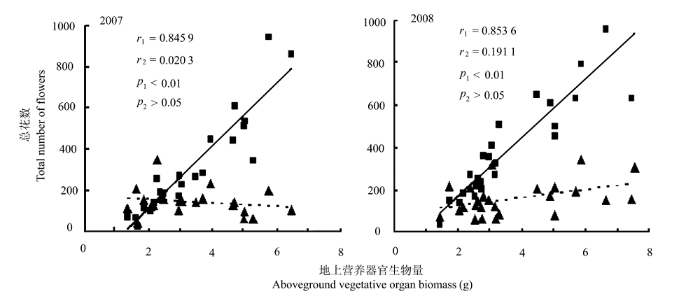
图1
簇花芹植株的总花数与其地上营养器官生物量的关系(2007-2008年)。r1, 两性花的相关系数; r2, 雄花的相关系数。■, 两性花; ▲, 雄花。实线表示两性花数随生物量的变化; 虚线表示雄花数随生物量的变化。
Fig. 1
Relationships between total number of flowers and aboveground vegetative organ biomass of individual plants of Soranthus meyeri in 2007 and 2008. r1, correlation coefficient of hermaphrodite flower; r2, correlation coefficient of staminate flower. ■, hermaphrodite flower; ▲, staminate flower. The solid lines represent the relationship between the number of hermaphrodite flowers and aboveground vegetative organ biomass of individual plant, and the dashed lines represent the relationship between the number of staminate flowers and aboveground vegetative organ biomass of individual plant.
2007-2008年簇花芹的性比与植株生物量呈正相关关系(p < 0.01), 生物量越大的植株性比越大( 图2), 说明其植株越大分配到雌性功能上的资源越多。
图2
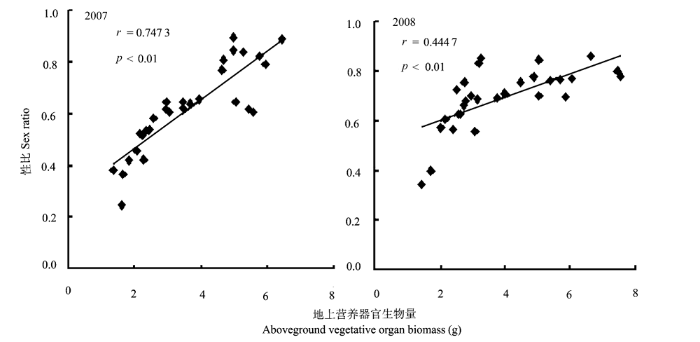
图2
簇花芹植株性比(两性花数/总花数)与其地上营养器官生物量的关系(2007-2008年)。
Fig. 2
Relationships between sex ratio (number of hermaphrodite flowers/total number of flowers) and aboveground vegetative organ biomass in individual of Soranthus meyeri in 2007 and 2008.
2.3 开花式样
图3
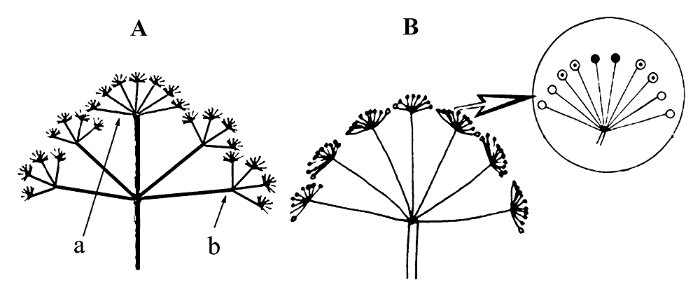
图3
簇花芹花序及花的分布。A, 植株上花序的分布。B, 伞形花序上雄花和两性花的分布。a, 一级复伞形花序; b, 二级复伞形花序; ○, 外围花; ⊙, 中间花; ●, 中央花。
Fig. 3
Arrangement of flowers and umbels within the inflorescence of Soranthus meyeri. A, Distribution of umbels on an individual plant. B, Distribution of staminate and hermaphrodite flowers in an umbellet. a, primary umbel; b, secondary umbel; ○, outer flower; ⊙, intermediate flower; ●, central flower.
表2 簇花芹植株的伞形花序数及花序上的雄花和两性花数(平均值±标准误差)
Table 2
| 观测指标 Items | 2006 | 2007 | 2008 | F | |
|---|---|---|---|---|---|
| 一级复伞形花序 Primary umbel | 伞形花序数 Number of umbellets | 15.4 ± 0.8 | 14.8 ± 0.8 | 16.2 ± 0.6 | 0.37 |
| 每伞形花序雄花数 Number of staminate flowers in an umbellet | 0.7 ± 0.3 | 4.0 ± 0.8 | 0.7 ± 0.2 | 15.51** | |
| 每伞形花序两性花数 Number of hermaphrodite flowers in an umbellet | 15.3 ± 0.5 | 13.3 ± 0.8 | 16.3 ± 0.6 | 5.78** | |
| 二级复伞形花序 Secondary umbel | 复伞形花序数 Number of umbels | 4.0 ± 0.3 | 3.0 ± 0.4 | 3.7 ± 0.4 | 2.05 |
| 每复伞形花序伞形花序数 Number of umbellets in an umbel | 4.4 ± 0.3 | 4.2 ± 0.3 | 4.3 ± 0.3 | 0.17 | |
| 每伞形花序雄花数 Number of staminate flowers in an umbellet | 7.4 ± 0.9 | 8.7 ± 0.9 | 6.7 ± 1.0 | 3.46* | |
| 每伞形花序两性花数 Number of hermaphrodite flowers in an umbellet | 9.0 ± 1.0 | 6.1 ± 0.9 | 9.3 ± 1.0 | 3.58* | |
*,p< 0.05; **,p< 0.01.
不同年份间植株的一、二级复伞形花序数和复伞形花序内伞形花序数无显著差异(p> 0.05), 而一级复伞形花序内各伞形花序间雄花和两性花数均存在极显著差异(p< 0.01), 二级复伞形花序内各伞形花序间雄花数和两性花数存在显著差异(p< 0.05)。
单株开花持续时间为9-11天, 一、二级复伞形花序开放时间均为5-6天。但一级复伞形花序开放约5天后, 二级复伞形花序才开始开放。一、二级复伞形花序中雄花和两性花的开花式样分别表现为: 雄花仅有雄性阶段, 在5枚花瓣序次开放的同时, 5枚花药亦序次开裂散粉, 整个过程持续(6.1 ± 0.6) h; 两性花雄性先熟, 开放过程持续3.5-6.0天, 且可划分为雄性、无性和雌性3个阶段。其中, 雄性阶段持续(6.1 ± 1.8) h, 花药开裂方式和持续时间与雄花的基本相同; 无性阶段表现为雌蕊花柱伸长但柱头不具可授性, 此阶段在花序外围花、中间花和中央花中的持续时间分别约为3、2和1天, 且彼此间存在不同程度的重叠; 雌性阶段以柱头具有可授性为标志, 持续时间为2.0-2.5天, 且花序不同位置的花完全重叠( 图4A)。
图4
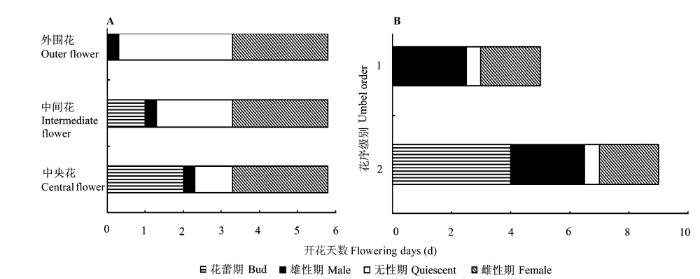
图4
簇花芹花序和复伞形花序水平的开花式样。A, 花序水平。B, 复伞形花序水平。
Fig. 4
Floral patterns within an umbellet and an umbel of Soranthus meyeri. A, At umbellet level. B, At umbel level.
复伞形花序内各伞形花序一般同时开放, 且花期持续时间基本相同。花序中花的开放是向心的, 表现为伞形花序的外围花先开, 然后依次向中央开放。一级复伞形花序中伞形花序的外围两性花散粉时, 中央花与中间花均处于蕾期; 当中间两性花进入雄性阶段散粉时, 外围花已进入无性阶段, 中央花仍处于蕾期; 而当中央两性花散粉时, 其外侧花均已进入无性阶段; 而当中央花进入无性阶段时, 其外侧花仍处在无性阶段。此阶段持续约1天后, 整个花序的两性花同时进入雌性阶段, 此阶段持续2.0-2.5天。因此, 从整个花序看, 其开花过程亦可分为雄性阶段、无性阶段和雌性阶段, 且其雌性功能和雄性功能被无性阶段完全分开; 不同位置两性花开花的雄性阶段和雌性阶段持续时间基本相同, 但无性阶段持续时间与其在花序中的位置密切相关, 具体表现为外围花>中间花>中央花(图4A)。
3 讨论
3.1 花期大小依赖的性别资源分配
植物有性繁殖过程中的性表达不仅受物种自身的遗传特性和资源状况影响, 而且还受环境条件(光照、温度和湿度等)的制约, 且不同类群中的制约因素可能有所不同(张大勇, 2004)。在雄全同株的伞形科植物Smyrnium olusatrum、Pastinaca sativa和峨参(Anthriscus sylvestris)中, 即使个体大小及其所处环境发生了变化, 但群体水平两性花与雄花比例始终保持为1:4, 说明其花期性比主要受遗传控制(Lovett Doust, 1980; Lovett Doust & Harper, 1980); 而在雄全同株的茄属(Solanum)植物中, 不少种类花期的雄花和两性花比率在不同居群和年份间存在显著差异, 说明其受环境的影响较大(Solomon, 1985; Diggle, 1993; Elle, 1999)。簇花芹植株上的两性花和雄花数虽然在不同年份间存在显著差异, 但其群体水平的性比均接近2:3, 彼此间无显著性差异, 该结果与Lovett Doust (1980)和Lovett Doust和Harper (1980)的研究结果基本一致, 说明簇花芹群体内的性比是相对稳定的, 可能受遗传因素控制。
资源分配是指植物将同化的资源用于生长与繁殖功能之间的比例(Charnov, 1982; Elle, 1999)。植物生长发育过程中可供利用的资源是有限的, 当分配给某一功能的资源增加时, 必然会导致分配给其他功能的资源减少, 彼此间存在着一种动态的权衡关系(Charnov, 1982; 张大勇, 2004)。在影响植物资源分配的各种因素中, 植株大小通常被认为是重要的制约因子, 它不仅影响资源在营养器官和繁殖器官之间的分配, 还影响着植株雌雄功能间的资源配置(Stöcklin & Faver,1994; Irwin, 2000)。簇花芹植株中雄花的花粉量及生物量等均比两性花的少, 说明其产生雄花比产生两性花所需资源少。在一、二级复伞形花序及其各伞形花序中, 两性花与雄花数及其生长位置与植株的资源分配也密切相关。从花序及花的开放顺序看, 其一级复伞形花序明显比二级复伞形花序开花早, 各花序外围花比内侧花开花早, 说明一级复伞形花序及各花序外围花发育早, 可从母株得到更多的资源; 而二级复伞形花序及各花序内侧花发育晚, 从母株获得的资源相对较少。为了能将个体有限的资源进行优化配置, 早发育的花序通过产生更多的两性花来增加其雌性适合度, 而晚发育的花序在雌性适合度达到饱和的前提下, 通过产生更多的雄花来避免其在雌性功能上投入的浪费, 增加其输出的总花粉量, 从而提高其整体适合度(Lovett Doust, 1980; Lovett Doust & Harper, 1980; Webb, 1981b; Bertin, 1982)。同时, 簇花芹植株将花粉包装到更多小的单位(雄花)比包装到一个大的单位(两性花)更能提高传粉成功率, 从而提高植株的雄性适合度, 这与Spalik (1991)对峨参的研究结果是一致的。此外, 在已报道的伞形科物种中, 性比(雄/雌)大多为恒定的4:1 (Cruden, 1976; Lloyd, 1979; Lovett Doust, 1980; Lovett Doust & Harper, 1980), 而在簇花芹中, 其两性花比例远大于该科其他植物, 究竟是遗传因素还是严酷的沙漠极端环境引起了其资源在雌雄功能间的分配差异, 从而造成了其性比的变化, 还有待于进一步探讨。
簇花芹植株水平的性比及两性花数与个体生物量均呈正相关关系(p< 0.01), 但雄花数与个体生物量不具相关性(p> 0.05), 说明其大小不同的个体对雌雄功能的资源投资也不相同, 较大的个体对雌性功能的投资大, 从而导致其性比发生变化(Charnov,1982; Thompson, 1987; Bertin & Kerwin, 1998)。这与Thompson (1987)对Lomatium grayi的研究结果一致, 根据Bateman (1948)的观点, 簇花芹的雌性繁殖成功可能受其资源限制较大, 而雄性繁殖成功更多地受制于其得到的交配机会, 植株通过对不同性功能的资源分配, 以取得最大的适合度收益(Charnov, 1982)。
3.2 开花式样的适应意义
雄全同株植物两性花和雄花的位置及其开放式样的差异会导致其在繁殖中所起的作用不同(张大勇, 2004)。生长在藜芦(Veratrum nigrum)顶生花序上的两性花比侧生花序上的雄花开放早, 传粉昆虫先访问早开放的两性花, 并将外源花粉沉降在柱头上以保证异交进行, 在其沿花序轴向下访问别的两性花过程中进行同株异花自交, 最后访问雄花以保障花粉输出并促进异交。由于藜芦的两性花具有延迟自交机制, 两性花和雄花的位置和开花顺序对其避免自交具有重要意义(廖万金等, 2003)。北美刺龙葵(Solanum carolinense)着生在花序顶端的雄花是为了更好地传粉, 而着生在花序基部的两性花是为了接受花粉提高结实率(Solomon, 1985)。由此可见, 在雄全同株植物中, 两性花和雄花不同的着生方式与开花式样对传粉昆虫的访花式样、交配方式以及后代适合度具有重要影响(Bertin, 1982; Solomon, 1985, 1986; 廖万金等, 2003; 张大勇, 2004)。
簇花芹一级复伞形花序的雄花数明显少于二级复伞形花序的, 这与许多雄全同株的伞形科植物随着复伞形花序分枝程度增加, 其雄性程度也显著提高的分布规律是一致的(Bell, 1971; Lovett Doust, 1980; Webb, 1981b), 其雄花数的增加有利于增大花展示以吸引更多传粉者进行传粉(Koul et al., 1989)。在开花顺序上, 簇花芹一级复伞形花序比二级复伞形花序早开放约5天, 但复伞形花序内各伞形花序几乎同时向心开放, 且花期持续时间基本相同。在开花过程中, 所有雄花仅有雄性阶段, 而两性花为雄性先熟, 且其雌雄阶段均被一无性阶段完全分离。由于花序内不同位置两性花的无性阶段持续时间表现不同, 因此, 虽然花序外围的两性花先开放, 但其花序中各花进入雌性阶段的时间是基本一致的, 花序的雌雄阶段被完全隔开(Koul et al., 1989; Davila & Wardle, 2002)。传粉昆虫访花时, 一级复伞形花序内所有花序几乎同时开放及其单花与花序雄性先熟且雌雄阶段完全分离的特性, 不仅有利于其两性花进行异株异花授粉, 增加后代的适合度, 还有效地避免了雌雄功能间的干扰; 而其一、二级复伞形花序花期雌雄阶段的重叠(1天左右), 可使传粉昆虫在植株内不同级别花序间进行同株异花传粉。这些特性与Davila和Wardle (2002)在伞形科植物Trachymene incisasubsp. incisa中的观察结果基本一致。二级复伞形花序与一级复伞形花序的开放式样相似, 但其雌雄性阶段的分离则有利于雄花花粉的输出, 增加其同株异花及异株异花传粉的机会(Koul et al., 1989; Davila & Wardle, 2007)。
致谢
国家自然科学基金(31060047)及新疆维吾尔自治区高技术研究发展计划项目(200810102)资助。美国Kentucky大学植物和土壤科学系的Carol C. Baskin教授和Jerry M. Baskin教授对本文的英文部分进行了修改, 在此表示衷心的感谢!
参考文献
Pollination success across an elevation and sex ratio gradient in gynodioecious Daphne laureola

Gynodioecy is a dimorphic breeding system in which hermaphrodite and female individuals coexist in populations. Sex ratio and gender-relative lifetime seed production determine the stability of gynodioecy, and both genetic and ecological factors may influence these parameters. I analyzed the consequences of variation in population sex ratio and site elevation for the relative pollination success of female and hermaphrodite individuals of Daphne laureola in southern Spain, where previous studies failed to detect female fecundity advantages at two mid-elevation sites. Pollination success, estimated as stigmatic pollen loads, number of pollen tubes per style, and percentage of fertilized flowers, was higher for hermaphrodites than females in populations with 20-56% females. Furthermore, female quantitative disadvantage in pollination success increased with elevation, suggesting that the higher availability of pollen due to the increased proportion of hermaphrodites could not mitigate the negative effect that other factors associated with elevation apparently had on pollination. Supplemental hand pollinations showed that female seed production was pollen limited in populations with a proportion of females >50%, although both pollination success and natural fruit set of females in these sites were the highest recorded.
Breeding systems and floral biology of the Umbelliferae or evidence for specialization in unspecialization flowers
In: Heywood VH ed. The Biology and Chemistry of the Umbelliferae.
The evolution and maintenance of andromonoecy
Floral sex ratios and gynomonoecy in Solidago (Asteraceae)
Floral sex ratios and gynomonoecy in Aster (Asteraceae)
Sex ratio variation in gynodioecious Lobelia siphilitica: effects of population size and geographic location
Variation in population sex ratio can be influenced by natural selection on alternate sex phenotypes as well as nonselective mechanisms, such as genetic drift and founder effects. If natural selection contributes to variation in population sex ratio, then sex ratio should covary with resource availability or herbivory. With nonselective mechanisms, sex ratio should covary with population size. We estimated sex ratio, resource availability, herbivory and size of 53 populations of gynodioecious Lobelia siphilitica. Females were more common in populations with higher annual temperatures, lower soil moisture and lower predation on female fruits, consistent with sex-specific selection. Females were also more common in small populations, consistent with drift, inbreeding or founder effects. However, small populations occurred in areas with higher temperatures than large populations, suggesting that female frequencies in small populations could be caused by sex-specific selection. Both selective and nonselective mechanisms likely affect sex ratio variation in this gynodioecious species.
Why be an hermaphrodite?
Sex ratio evolution in a variable environment
Intraspecific variation in pollen-ovule ratios and nectar secretion―preliminary evidence of ecotypic adaptation
Practical Pollination Biology
Reproductive ecology of the Australian herb Trachymene incisa subsp . incisa (Apiaceae)
Bee boys and fly girls: do pollinators prefer male or female umbels in protandrous parsnip
Biased sex ratios in the dioecious annual Croton texensis (Euphorbiaceae) are not due to environmental sex determination
Developmental plasticity, genetic variation, and the evolution of andromonoecy in Solanum hirtum (Solanaceae)
Sex allocation and reproductive success in the andromonoecious perennial Solanum carolinense (Solanaceae). I. Female success
Andromonoecy in Zigadenus paniculatus (Liliaceae): spatial and temporal patterns of sex allocation
On methodology of foraging behavior of pollinating insects
Pollen dispersal and mating patterns in animal-pollinated plants
In: Lloyd DG, Barrett SCH eds. Floral Biology: Studies on Floral Evolution in Animal-Pollinated Plants.
Morphological variation and female reproductive success in two sympatric Trillium species: evidence for phenotypic selection in Trillium erectum and Trillium grandiflorum (Liliaceae)
Reproductive biology of wild and cultivated carrot (Daucus carota L.)
A preliminary study on the reproductive features of Veratrum nigrum along an altitudinal gradient
Parental strategies of angiosperms
Modification of the gender of seed plants in varying conditions
BACKGROUND: Genes encoding proteins underlying host-pathogen co-evolution and which are selected for new resistance specificities frequently are under positive selection, a process that maintains diversity. Here, we tested the contribution of natural selection, recombination and transcriptional divergence to the evolutionary diversification of the plant defensins superfamily in three Arabidopsis species. The intracellular NOD-like receptor (NLR) family was used for comparison because positive selection has been well documented in its members. Similar to defensins, NLRs are encoded by a large and polymorphic gene family and many of their members are involved in the immune response. RESULTS: Gene trees of Arabidopsis defensins (DEFLs) show a high prevalence of clades containing orthologs. This indicates that their diversity dates back to a common ancestor and species-specific duplications did not significantly contribute to gene family expansion. DEFLs are characterized by a pervasive pattern of neutral evolution with infrequent positive and negative selection as well as recombination. In comparison, most NLR alignment groups are characterized by frequent occurrence of positive selection and recombination in their leucine-rich repeat (LRR) domain as well negative selection in their nucleotide-binding (NB-ARC) domain. While major NLR subgroups are expressed in pistils and leaves both in presence or absence of pathogen infection, the members of DEFL alignment groups are predominantly transcribed in pistils. Furthermore, conserved groups of NLRs and DEFLs are differentially expressed in response to Fusarium graminearum regardless of whether these genes are under positive selection or not. CONCLUSIONS: The present analyses of NLRs expands previous studies in Arabidopsis thaliana and highlights contrasting patterns of purifying and diversifying selection affecting different gene regions. DEFL genes show a different evolutionary trend, with fewer recombination events and significantly fewer instances of natural selection. Their heterogeneous expression pattern suggests that transcriptional divergence probably made the major contribution to functional diversification. In comparison to smaller families encoding pathogenesis-related (PR) proteins under positive selection, DEFLs are involved in a wide variety of processes that altogether might pose structural and functional trade-offs to their family-wide pattern of evolution.
Floral sex ratios in andromonoecious Umbelliferae
The resource costs of gender and maternal support in an andromonoecious umbellifer Smyrnium olusatrum L
Size and sex of floral displays affect insect visitation rates in the dioecious Australian alpine herb, Aciphylla glacialis (Apiaceae)
Vegetation-environment relationships in Gurbantunggut Desert
The taxonomic studies on Xinjiang Ferula and its close genera
Environmentally influenced changes in sex expression in an andromonoecious plant
Sexual allocation and andromonoecy: resource investment in male and hermaphrodite flowers of Solanum carolinense (Solanaceae)
On evolution of andromonoecy and ‘overproduction’ of flowers: a resource allocation model
Effects of plant size and morphological constraints on variation in reproductive components in two related species of Epilobium
Floral sex ratios, fruit-set, and resource allocation in plants
Effects of floral display on pollinator behavior and pollen dispersal
The ontogeny of flowering and sex expression in divergent populations of Lomatium grayi
Populations of Lomatium grayi (Umbelliferae), an andromonoecious, perennial herb, differe in growth rates, flowering frequency, and survivorship. Effects of these different life histories on the ontogeny of sex expression were analyzed for plants from two populations grown from seedlings in a common garden and monitored for six years. Plants from Smoot Hill, Washington grew faster, had a higher probability of flowering at each age and size after the first year of growth, and a higher probability of flowering repeatedly among years than did plants from Clarkston, Washington. The proportion of plants producing some hermaphroditic flowers increased with plant size in both populations. Smoot Hill plants, however, were more likely to begin flowering as small, staminate plants than Clarkston plants. Clarkston plants did not begin flowering until they were older and larger, and most of these plants produced some hermaphroditic flowers when they began reproduction. The positive association between production of hermaphroditic flowers and both plant size and age was consistent with the hypothesis that hermaphroditic flowers are more costly to produce than staminate flowers.Although the populations did not differ in the total number of flowers per plant produced at any age or size, Smoot Hill plants consistently produced a lower percentage of hermaphroditic flowers than Clarkston plants at larger sizes and later ages. Consequently, selection for faster growth rates and higher flowering frequency at small sizes and early ages may have favored the more staminate-biased sex ratios in the Smoot Hill population.
Sex ratios in populations of Geranium sylvaticum in European Russia
Breeding systems and the evolution of dioecy in New Zealand apioid Umbelliferae
Gynodioecy in Gingidia flabellata (Umbelliferae)
Andromonoecism, protandy and sexual selection in Umbelliferae
Sex ratios in New Zealand apioid Umbelliferae

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}