


高山林线是山地森林最显著的生态界限(Holt- meier, 2003)。由于极端的环境条件, 林线生物类群均处于相对复杂的临界状态, 是典型的生态脆弱区(Butler et al., 1994)和气候变化敏感地带(Körner & Paulsen, 2004)。高山森林-苔原交错带(alpine forest- tundra ecotone, AFTE)涵盖了从暗针叶林到矮曲林、高山灌丛及草甸等各种植被类型, 包括了高山林线(timberline)和树线(treeline)等重要的生态界限(Holtmeier, 2003)。高山森林-苔原交错带是全球高山地区普遍存在的植被过渡区。由于短距离内不同植被类型的交错分布, 使得高山森林与苔原之间基因交流、个体迁移和种替代等过程对气候变化的反应可能更加敏感而迅速(刘洋等, 2009)。因此, 高山森林-苔原交错带成为指示气候变化、能量交换、物质循环及生物多样性预警方面的热点区域(Kupfer & Cairns, 1996; Alftine et al., 2003; Walther et al., 2005; Díaz-Varela et al., 2010)。
植被在高山的受限分布可能导致相应的土壤生态过程的变化, 对环境敏感的土壤微生物可以作为高山气候变化的预警及敏感指标。土壤微生物的群落结构通常受到植被(Balser & Firestone, 2004)和季节动态(Bardgett et al., 1999; Waldrop & Firestone, 2006; Björk et al., 2008)的影响。近年来, 有关高山生态系统土壤微生物的研究逐渐增多, 涉及高山苔原土壤微生物群落的季节动态(Björk et al., 2008), 高山苔原微生物群落结构及功能群(Lipson et al., 2002), 苔原不同植被类型下土壤微生物生物量、养分有效性及氮矿化潜力(Chu & Grogan, 2010), 高山林线土壤微生物生物量及生理类群数量(齐泽民和王开运, 2009), 高山区土壤微生物数量及其与生态因子的相关性(邵宝林等, 2006)等。但由于高山林线所处的海拔太高, 且开展研究工作难度较大, 对青藏高原东缘高山土壤微生物及其相关的土壤生态过程的研究还十分欠缺。
高山/亚高山是研究气候变化的关键区域, 监控这些敏感和脆弱的生态系统对反映气候变化有重要价值。高山生态系统受季节性雪被的作用明显, 存在明显的生长季节和非生长季节(Schadt et al., 2003; Björk et al., 2008)。在川西高山地区, 冬季土壤冻结期长达5-6个月, 每年11月至次年4月形成季节性雪被(Wu et al., 2010), 从5月到10月是植物开始生长到生长结束的季节。研究发现, 气候变化可能改变高山生态系统的季节性雪被状况和土壤冻融循环过程, 影响高山生态系统碳氮循环(ACIA Secretariat, 2005), 进而显著影响土壤微生物生物量和群落结构(Lipson & Monson, 1998; Koponen et al., 2006)。地处青藏高原东缘和长江上游的川西林区分布着世界上海拔较高的高山森林-苔原交错带, 研究该区域的土壤微生物的季节动态及其影响因素对预测和理解全球气候变化对高山生态系统的影响具有重要的意义。因此, 本文以青藏高原东缘的米亚罗鹧鸪山高山森林-苔原交错带(原始针叶林、林线、树线、密灌丛、疏灌丛和高山草甸)的土壤微生物为研究对象, 研究了微生物生物量碳氮和可培养微生物数量的季节动态, 以期深入了解全球气候变化背景下高山生态系统的土壤微生物特征。
1 研究地概况和样地描述
四川省理县米亚罗林区(31°43′-31°51′ N, 102°40′-103°02′ E), 位于青藏高原东缘褶皱带最外缘, 岷江上游主要支流杂谷脑河上游, 海拔2200-5500 m, 年平均气温6-12 ℃, 1月平均气温-8 ℃, 7月平均气温12.6 ℃, 年积温为1200-1400 ℃, 年降水量为600-1100 mm, 年蒸发量为1000-1900 mm, 具有降水次数多、强度小的特点。地形以石质山地为主, 土壤具有粗骨(石砾含量高)和薄层等特征。从低海拔到高海拔, 土壤类型依次为山地暗棕壤、棕色针叶林土、高山草甸土。杂谷脑河北侧的鹧鸪山山体海拔3200-4800 m, 这是大渡河与岷江的分水岭, 为重要的江河源区。山地的垂直地带性明显, 自河谷至山顶依次分布有针阔混交林、暗针叶林、高山灌丛、高山草甸等。
本研究范围在米亚罗鹧鸪山(31°51′428′′ N、102°41′230′′ E)海拔3900-4200 m的高山森林-苔原交错区。从低海拔到高海拔, 植被类型由暗针叶林过渡到高山苔原, 地被物厚度逐渐降低, 土壤类型由棕色针叶林土过渡到高山草甸土。这一区域包括了典型的自然条件下形成的林线和树线, 能很好地反映川西高山森林-苔原交错带植被、土壤及地被物的生态特征。采用GPS定位, 沿垂直于等高线方向, 在高山森林-苔原区域设置海拔高差300 m、宽30 m的3条样带, 样带与样带之间距离约500 m。在样带上以海拔50 m间距设置样地(样地面积为30 m × 30 m, 约1000 m2), 从低海拔到高海拔, 样地分别经过暗针叶林、林线、树线、密灌丛、疏灌丛和高山草甸6个植被群落类型。于2009年8月植被生长旺季, 对样地的基本特征进行了调查, 结果见表1。
表1 高山森林-苔原交错带基本特征
Table 1
| 样带 Belt transect | 暗针叶林Dark-conifer forest | 林线Timberline | 树线 Treeline | 密灌丛 Dense shrub | 疏灌丛 Sparse shrub | 高山草甸 Alpine meadow |
|---|---|---|---|---|---|---|
| 海拔 Elevation (m) | 3 900 | 3 975 | 4 050 | 4 105 | 4 135 | 4 200 |
| 坡向 Slope aspect (°) | EN 5 | EN 4 | EN 5 | EN 5 | EN 3 | EN 3 |
| 坡度 Slope degree (°) | 32 | 34 | 40 | 36 | 38 | 34 |
| 木本植物平均高度 Wood average height (m) | 22.0 | 14.0 | 8.0 | 5.0 | 1.5 | – |
| 草本植物平均高度 Herbage average height (cm) | 10 | 20 | 50 | 35 | 20 | 10 |
| 郁闭度 Crown density (%) | 80 | 75 | 70 | 65 | 20 | – |
| 草本植物盖度 Cover degree of herb (%) | 20 | 30 | 40 | 60 | 90 | 85 |
| 苔藓厚度 Thickness of moss (cm) | 10.0 | 4.5 | 4.0 | 1.8 | 0.5 | 0.6 |
| 枯落物厚度 Thickness of litter (cm) | 9.0 | 8.0 | 5.8 | 4.5 | 2.5 | 1.2 |
高山森林-苔原交错带暗针叶林优势树种是岷江冷杉(Abies faxoniana), 灌木优势种有大理杜鹃(Rhododendron taliense)、皱皮杜鹃(R. wiltonii)、鹧鸪杜鹃(R. zheguense)、冰川茶藨(Ribes glaciale)、华西小檗(Berberis silva-taroucana)、红毛花楸(Sorbus rufopilosa)、 细枝绣线菊(Spiraea myrtilloides)、窄叶鲜卑花(Sibiraea angustata)、柳叶忍冬(Lonicera lanceolata)、刚毛忍冬(L. hispida)、越橘叶忍冬(L. myrtillus)、叉柱柳(Salix divergentistyla)、金露梅(Potentilla fruticosa), 草本优势种为野黄韭(Allium rude)、圆穂蓼(Polygonum macrophyllum)、东方草莓(Fragaria orientalis)、肾叶龙胆(Gentiana crassuloides)、箭叶橐吾(Ligularia sagitta)等。苔藓种类有塔藓(Hylocomium splendens)、锦丝藓(Actinot- huidium hookeri)、大羽藓(Thuidium cymbifolium)、万年藓(Clinacium dendroides)、地钱(Marchantiaceae spp.)等。
2 材料和方法
2009年5月4日-11月2日, 在林内距地面1.5 m和土壤深度10 cm处分别安置一个纽扣式温度记录器(iButton DS1923-F5, Dallas Semiconducotr, Dallas, USA), 同步监测空气温度和土壤温度, 每2 h自动记录1次数据。土壤样品采集于2009年5月4号、8月2号和11月2号。各标准地中以“S”形布点, 去除表层的苔藓、枯枝落叶后, 用直径为5 cm的土钻采集样地内0-10 cm表层的土壤, 10个样点的土壤混合后作为一个土样。采集的土壤放入无菌塑料袋内, 置于有冰袋的保鲜盒中运回实验室。去掉可见植物残根和石块后, 用四分法取舍, 过2 mm筛后, 混合均匀, 保存于4 ℃冰箱中备用。土壤中的细菌、真菌和放线菌数量分别采用牛肉蛋白胨培养方法、马铃薯葡萄糖琼脂培养基和高氏1号培养基法测定(中国科学院南京土壤研究所微生物室, 1985), 细菌、真菌、放线菌均采用平板计数法计数。土壤微生物生物量碳和氮采用改进的氯仿熏蒸-K2SO4浸提法测定(吴金水, 2006), 提取液中的碳和氮采用有机碳自动分析仪(Total organic carbon analyzer-2500, Shimadzu Corporation, Japan)测定。
采用SPSS 11.5 for Windows统计软件作统计分析, 对每个植被及每个季节的土壤微生物, 采用单因素方差分析(one-way ANOVA)方法, 检验土壤生物参数在植被类型和季节动态上的差异。用双因素方差分析(two-way ANOVA)检验植被类型和季节动态及其交互作用对土壤微生物生物量碳、氮和微生物数量的影响, 微生物生物量碳、氮与细菌、真菌、放线菌数量的关系采用Pearson相关分析, 用Excel做图表。
3 研究结果
3.1 高山森林-苔原交错带大气温度和土壤温度动态
根据温度记录器每2 h自动监测的数据, 可以看出, 高山森林-苔原交错带5-10月的日平均气温基本都在0 ℃以上(图1A), 土壤月平均温度也都高于0 ℃ (图1B), 这个阶段可作为高山植被的生长季节。生长季节月平均气温和土壤温度分别为: 5月4.12 ℃和3.33 ℃、6月6.74 ℃和6.08 ℃、7月8.95 ℃和8.86 ℃、8月8.71 ℃和9.16 ℃、9月7.57 ℃和8.30 ℃、10月1.46 ℃和3.22 ℃。其中, 5月和10月的平均气温和土壤温度最低, 8月的土壤温度最高。因此, 将5月作为高山植物的生长季初期, 8月作为植物生长季中期, 10月作为生长季末期。不同植被类型之间土壤温度有差异, 从暗针叶林到高山草甸, 5-10月的土壤平均温度为暗针叶林5.25 ℃、林线5.17 ℃、树线5.69 ℃、密灌丛6.81 ℃、疏灌丛8.16 ℃、高山草甸7.86 ℃。各植被类型中, 林线土壤月平均温度最低, 而疏灌丛和高山草甸的土壤月平均温度高于其他植被类型。
图1
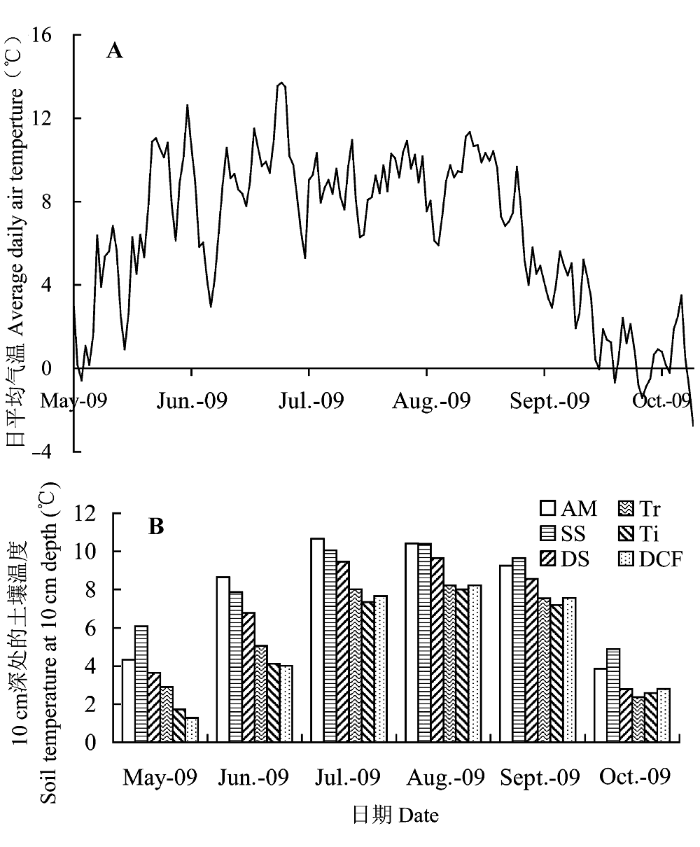
图1
2009年5月至10月高山森林-苔原交错带日平均气温(A)和土壤月平均温度(B)动态。AM, 高山草甸; DCF, 暗针叶林; DS, 密灌丛; SS, 疏灌丛; Ti, 林线; Tr, 树线。
Fig. 1
Dynamics of average daily air temperature (A) and soil temperature at 10 cm depth by month (B) from May to October, 2009 in the forest-alpine tundra ecotone. AM, alpine meadow; DCF, dark-conifer forest; DS, dense shrub; SS, sparse shrub; Ti, timberline; Tr, treeline.
3.2 土壤微生物生物量和数量动态
对比交错带不同植被类型的土壤微生物生物量碳氮分布及其比值(图2A、2B、2C)和可培养细菌、真菌、放线菌数量(图2D、2E、2F)发现, 细菌数量占绝对优势, 占整个微生物总数的91.61%-99.81%, 放线菌数次之, 放线菌的比例介于0.16%-8.31%, 真菌数量最少, 不到微生物总数的0.1%。生长季初期, 林线的土壤微生物生物量碳显著低于高山草甸、疏灌丛和密灌丛, 且林线土壤微生物生物量氮也显著低于其他植被; 树线以下细菌、真菌、放线菌数量显著低于树线以上。在生长季中期, 草甸的微生物生物量碳、氮最高, 草甸和密灌丛微生物生物量碳显著大于其他植被, 密灌丛的微生物生物量氮显著高于其他植被, 林线的土壤微生物生物量氮显著低于其他植被; 树线以下土壤细菌数量显著低于树线以上, 草甸土壤真菌数量显著高于其他植被, 密灌丛土壤放线菌数量显著高于其他植被。生长季末期, 密灌丛土壤的微生物生物量碳显著低于其他植被, 树线以下微生物生物量氮、细菌及放线菌数量显著低于树线以上, 林线和暗针叶林土壤的真菌数量显著低于其他植被。暗针叶林、林线和树线的土壤MBC/MBN显著高于其他植被类型。
图2
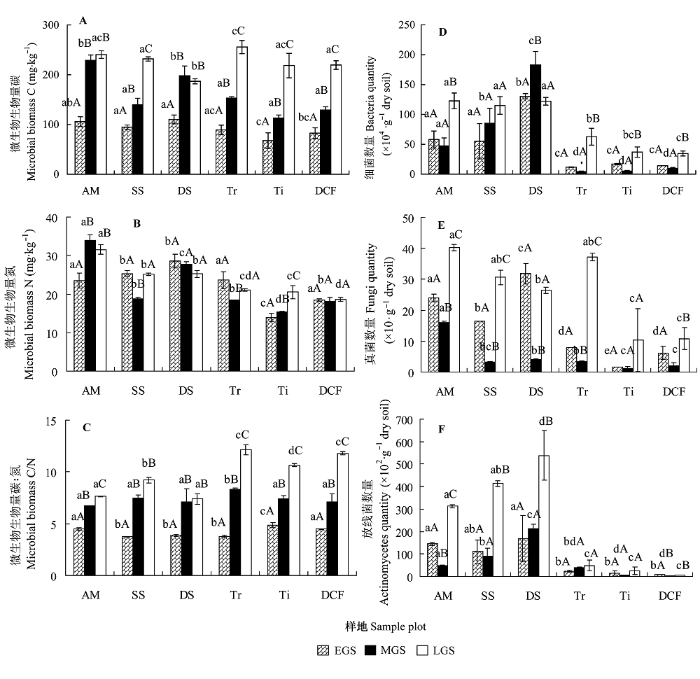
图2
高山森林-苔原交错带土壤微生物生物量碳(A)、氮(B)和碳氮比(C)和可培养细菌(D)、真菌(E)、放线菌(F)数量的季节变化(平均值±标准偏差)。不同小写字母表示相同季节里植被之间差异显著(p < 0.05); 不同大写字母表示相同植被在不同季节之间差异显著(p < 0.05)。EGS, 生长季初期; LGS, 生长季末期; MGS, 生长季中期。AM, 高山草甸; DCF, 暗针叶林; DS, 密灌丛; SS, 疏灌丛; Ti, 林线; Tr, 树线。
Fig. 2
Seasonal changes of soil microbial biomass carbon (A), nitrogen (B) and C: N (C), as well as quantity of cultivate bacteria (D), fungi (E) and actinomycetes (F) of soil in forest-alpine tundra ecotone (mean ± SD). Lower case letters indicate significant differences among different vegetations within same season (p < 0.05); upper case letters indicate significant differences among different seasons within same vegetation (p < 0.05). EGS, early in the growing season; LGS, late in the growing season; MGS, middle in the growing season. AM, alpine meadow; DCF, dark-conifer forest; DS, dense shrub; SS, sparse shrub; Ti, timberline; Tr, treeline.
对比交错带不同季节微生物的变化, 各植被类型微生物生物量碳(MBC)初期显著低于中期和末期, 疏灌丛、树线、林线和暗针叶林三个季节间MBC差异显著。各植被类型的微生物生物量氮(MBN)的季节变化趋势不一致, 但林线MBN具有显著的季节变化。暗针叶林、林线、树线和高山草甸末期的土壤细菌数量显著高于生长季初期和中期, 密灌丛中期的土壤细菌数量显著高于初期和末期。树线、疏灌丛和高山草甸末期的土壤真菌数量显著高于初期和中期, 林线季节之间土壤真菌数量差异不显著。树线以上的土壤放线菌数量在植物生长季末期显著大于生长季初期和生长季中期, 树线以下土壤放线菌数量在季节之间差异不显著。MBC/MBN的趋势为生长季末期>生长季中期>生长季初期。
以植被类型和季节变异作为处理因子, 对土壤微生物生物量碳、氮及其比值, 以及细菌、真菌、放线菌数量进行双因素方差分析, 结果见表2。总的来看, 植被类型、季节变化及其两者的交互效应对土壤微生物生物量碳、氮及其比值, 以及细菌、真菌、放线菌数量都有显著作用(p < 0.001或p = 0.004 4)。
表2 植被类型(从暗针叶林到高山草甸)和季节(5-10月三次采样)及其交互作用对高山森林-苔原交错带10 cm深处土壤微生物生物量碳、氮和微生物数量的双因素方差分析结果
Table 2
| 变量 Variable | 植被 Vegetation | 季节 Season | 植被×季节 Vegetation×season | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| df | F | p | df | F | p | df | F | p | |||
| 微生物生物量碳 MBC | 5 | 20.64 | <0.001 | 2 | 481.75 | <0.001 | 10 | 14.40 | <0.001 | ||
| 微生物生物量氮 MBN | 5 | 120.48 | <0.001 | 2 | 7.44 | 0.004 | 10 | 20.21 | <0.001 | ||
| 微生物生物量碳氮比 MBC/MBN | 5 | 26.53 | <0.001 | 2 | 541.92 | <0.001 | 10 | 17.07 | <0.001 | ||
| 细菌 Bacteria | 5 | 86.20 | <0.001 | 2 | 22.59 | <0.001 | 10 | 7.16 | <0.001 | ||
| 真菌 Fungi | 5 | 55.36 | <0.001 | 2 | 166.69 | <0.001 | 10 | 14.13 | <0.001 | ||
| 放线菌 Actinomycetes | 5 | 51.18 | <0.001 | 2 | 58.46 | <0.001 | 10 | 14.37 | <0.001 | ||
MBC, microbial biomass carbon; MBN, microbial biomass nitrogen.
3.3 土壤微生物生物量碳、氮和微生物数量的关系
三个季节的微生物数据的相关分析结果(图3) 表明, 土壤微生物生物量碳、氮与细菌、真菌、放线菌数量存在一定的线性正相关关系。其中, MBC与真菌数量呈显著正相关关系(p < 0.05), MBC与细菌、放线菌数量相关性不显著(p > 0.05); MBN与细菌、真菌数量呈极显著正相关关系(p < 0.01), MBN与放线菌数量呈显著正相关关系(p < 0.05)。
图3
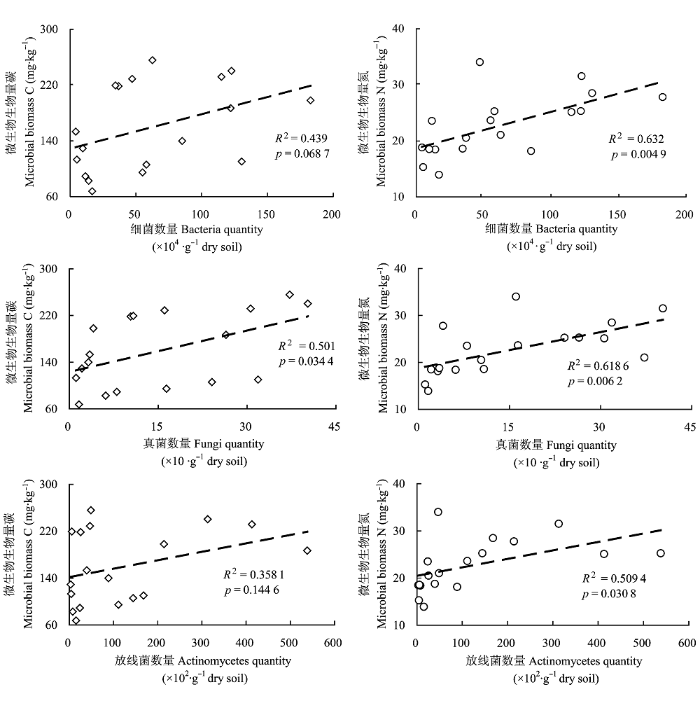
图3
土壤微生物生物量碳、氮与细菌、真菌、放线菌数量的相关关系。
Fig. 3
Relationships between soil microbial biomass carbon, nitrogen and bacteria, fungi, actinomycetes quantity.
4 讨论
土壤微生物生物量和数量的分布受气候条件、土壤质地、植被组成和覆盖度的综合影响(Nielsen et al., 2009)。许多研究发现, 土壤微生物与地上植被的关系密切(Lipson & Monson, 1998; Rajaniemi & Allison, 2009), 植被类型会影响土壤微生物群落结构和数量。Imberger和Chiu (2001)的研究表明, 从高山针叶林过渡到草甸, 10 cm土层的微生物生物量显著下降。也有相反的结论: 武夷山中亚热带常绿阔叶林、针叶林、亚高山矮林, 以及高山草甸土壤微生物生物量随海拔升高而增大(何容等, 2009)。本研究发现, 不同时期土壤微生物在各植被类型间分布有差异。植物生长季初期和生长季中期, 树线以上群落的MBC高于树线下的群落, 而到生长季末期则恰恰相反, 暗针叶林、林线和树线MBC显著升高, 各植被间MBC的差异减小, 可能是生长季末期森林凋落物的大量输入和雪被覆盖所致。土壤微生物数量基本上也是以树线为界, 树线以下群落土壤微生物数量显著低于树线以上群落, 其中密灌丛的土壤细菌数量最多。可能由于草本植物根系发达, 密集于表层, 根系分泌物和脱落物是微生物丰富的能源物质, 有利于土壤养分的积累和微生物的生长繁殖(丁玲玲等, 2007; 何容等, 2009)。
同时, 植被也会影响土壤温度和水分等环境特征, 这都是影响微生物群落结构和功能的潜在因素。低温和短的生长季是高山生态系统的两个主要特征。温度控制假说认为, 生长季节平均土壤温度(10 cm深处) (6.7 ± 0.8) ℃是全球林线分布的控制因子(Körner & Paulsen, 2004)。从生长季节土壤月平均温度来看, 并不是随海拔升高土壤温度呈降低趋势, 疏灌丛和高山草甸的土壤月平均温度高于其他植被类型, 林线所处的土壤月平均温度最低。鹧鸪山林线海拔接近4000 m, 而王朗自然保护区林线海拔3300 m, 海拔、温度、植被等因素可能导致鹧鸪山林线交错带土壤微生物总数少于王朗林线交错带的土壤微生物数量(齐泽民和王开运, 2009)。一些研究表明, 交错带物质交换和能量流动比相邻的群落更频繁、物种多样性更高(Risser, 1995; Callaway et al., 2002), 如王朗自然保护区林线过渡带比邻近植被土壤具有相对较强的生物学活性, 表层土壤细菌、真菌、放线菌及微生物总量高于邻近植被上缘流石滩草甸与下缘冷杉林(齐泽民等, 2009)。但本研究结果发现, 林线的土壤微生物生物量和数量在整个森林-苔原交错带中并不是最高的。
土壤微生物群落的季节动态对于植物吸收利用土壤中的氮有重要意义。植物和微生物在季节动态上尽量避免对氮的竞争, 植物生长季初期对氮吸收和微生物在生长期末对氮的固定增强了高山生态系统对氮的固定和利用(Jaeger et al., 1999; Miller et al., 2009)。微生物生物量的周转是植物生长季节吸收有机氮和无机氮的最大来源(Schmidt et al., 2007)。在科罗拉多州的高山草甸的土壤, 已观察到微生物生物量在秋季和冬季增长而在融雪的春季迅速下降(Lipson et al., 2002), 这些微生物生物量的季节变化与高山植物对有效性氮的利用相关。融雪造成大量的蛋白质从冬季微生物中释放出来, 这些蛋白质转化为生长期可供植物吸收的氨基酸。Lipson和Monson (1998)的研究表明, 植物在生长季前期大量吸收氮用于植物生长, 微生物对氮的固定主要发生在植物凋落后。土壤微生物生物量和数量的季节波动与植物生长季末期大量的凋落物输入有关(Liu et al., 2003; Schadt et al., 2003; Devi & Yadava, 2006; Waldrop & Firestone, 2006; Nielsen et al., 2009), 因为10月正是高山植物凋落的高峰期。研究结果发现, 植物生长季初期的MBC显著低于生长季中期和末期, 可能是因为积雪融化后大量的真菌降解转化, 导致MBC降低。MBN的季节变化趋势各植被之间有差异, 并不是所有的植被都表现为生长季末期高于生长季初期和中期, 只有高山草甸和林线的季节变化与MBC一致, 可能是Lipson和Monson (1998)与Lipson等(2002)的研究只限于高山嵩草草甸, 并没有包括针叶林和灌丛等植被类型, 植被类型和微环境的差异可能造成MBN的季节变化不同。
相关研究也证明: 不同植被类型、季节变化和年际动态对土壤微生物群落组成和数量都有显著影响(Kieft et al., 1998; 刘满强等, 2003; Waldrop & Firestone, 2006)。土壤微生物主要以植物残体为营养源, 森林-苔原交错带凋落物的质和量的变化必然导致土壤微生物在各植物群落中分布不均一(Lipson & Monson, 1998; Sun et al., 2009)。地上/地下凋落物调节着土壤碳氮库大小及周转速率, 进而影响了土壤微生物的群落构成和生物量(Bardgett et al., 1999; Sun et al., 2009)。Waldrop和Firestone (2006)也研究证实, 植被或季节的变化实质上导致了土壤生态系统养分和碳循环的改变, 从而影响了微生物群落的结构。由于本研究缺乏对各植被类型凋落物输入量、土壤有效养分的监测, 引起土壤微生物在植被类型和季节动态上的变异机制还有待于进一步研究。
通常真菌和细菌在土壤环境中占据不同的生态位, 细菌在土壤氮转化过程中起着重要作用, 而真菌在土壤碳和能源循环过程中作用巨大(Imberger & Chiu, 2001; 杨万勤等, 2006)。本研究结果也证实了这一点, MBC与土壤真菌数量呈显著正相关关系, MBN与土壤细菌数量呈极显著正相关关系。定量真菌/细菌的比例很重要(West, 1986)。土壤微生物生物量碳氮比反映了土壤中真菌和细菌的比例(Fauci & Dick, 1994)。本项研究结果表明, 生长季末期MBC/MBN显著增加, 说明土壤真菌数量增多。虽然对细菌和真菌各自的生物量还无法判断, 但从可培养的微生物数量来看, 生长季末期真菌数量较生长季初期和中期显著增多。秋冬季土壤生态系统中真菌占优势, 主要是真菌比细菌更善于消化纤维素, 有利于分解凋落物, 而在夏季, 植物生长分泌出的淀粉和糖能为土壤细菌提供食物来源(Krajick, 2006)。Pennisi (2003)研究发现, 真菌是冬季土壤微生物的优势类群, 真菌生物量冬季是夏季的3倍, 冬季真菌的生物量是细菌的15倍。Lipson等(2002)对落基山干旱草甸的研究表明, 土壤微生物生物量在秋冬季升高, 春季雪被融化后迅速降低, 冬季真菌与细菌的比值更高。苔原土壤微生物生物量在冬季积雪达到高峰期, 真菌生物量占大多数(Schadt et al., 2003)。Imberger和Chiu (2001)研究认为, 从亚高山针叶林过渡到高山草甸, 0-10 cm表层土壤的真菌/细菌比例降低, 说明随着植被变化土壤中的木质素和纤维素含量逐渐降低。我们研究发现, 生长季中期和生长季末期的MBC/MBN在树线区域最高, 且可培养的土壤真菌/细菌比值在整个生长季节也是树线区域最高, 可能是树线区域土壤中木质素和纤维素含量偏高。但本实验没有测定土壤中木质素和纤维素的含量, 还需进一步研究。
高山生态系统是受气候变化影响最明显的区域(IPCC, 2007)。Pauli等(2007)的研究表明, 过去50 年中, 欧洲阿尔卑斯山的升温是全球平均气温升高的2倍以上, 青藏高原升温速度相当于全球变暖速率的3倍。全球变暖导致高山冰川消融加速, 山地冰川和积雪面积呈退缩趋势, 雪线和林线位置上升。1957-2003年观测发现, 气候变化使阿尔卑斯山中部山地生态交错带向上移动, 平均每10年林线、树线和苔原线分布向高海拔迁移25 m、13 m、11 m (Díaz-Varela et al., 2010)。目前发现, 相对于非生长季节, 生长季节岷江冷杉凋落物具有最高的MBC和MBN (周晓庆等, 2011)。美国西部和欧洲山地生态系统的冬季积雪大幅减少, 使雪被下的土壤微生物群落呈指数增加, 有机物质分解速率加快(Monson et al., 2006)。气候变暖使许多亚高山/高山积雪消融时间提前(Walther et al., 2002), 这可能延长高山植物的生长季, 使植物获得更多的碳, 并增加生物量。由此推测, 气候变暖可能会使高山土壤微生物生物量增加, 加速土壤有机质的分解, 使土壤中更多的碳释放到大气中, 进而加剧了气候变暖的 趋势。
5 结论
综上所述, 高山森林-草甸交错带植被类型和季节动态对土壤微生物生物量碳、氮和微生物群落结构都有显著作用。交错带MBC在植物生长季末期显著高于生长季中期和初期, MBC的季节动态变异大于MBN。可培养微生物数量季节动态为生长季末期>生长季初期>生长季中期。生长季末期交错带土壤真菌数量显著增加, 且MBC/MBN在三个季节中最高。在植物生长季初期和中期, 林线的微生物生物量处于低谷, 树线可以作为交错带微生物数量的分界线, 树线以下群落的土壤微生物数量显著低于树线以上群落。MBN和细菌、真菌和放线菌存在显著的相关关系, 而MBC仅和真菌数量存在显著的相关关系。导致这些结果出现的机制还有待于深入研究, 如细菌和真菌各自的生物量及不同季节土壤微生物多样性可作为下一步研究的内容。本研究结果为深入了解高山森林-苔原交错带土壤微生物群落结构及相应的土壤生态学过程提供了重要的科学依据, 对于理解全球气候变化对陆地生态系统的影响具有重要的意义。
致谢
国家自然科学基金项目(31170423和31000- 213)、教育部博士点基金项目(20115103120003和20105103110002)和博士后基金项目(20110491732)、四川省教育厅重点项目(2011ZA079)、四川省科技支撑计划项目(2010NZ0051)、四川省杰出青年学术技术带头人培育计划项目(2011JQ0035)资助。感谢四川农业大学林学院苏江峰、李勇、刘凯、钟军、李泽波、赖刚、李瑞久、柯巍等同学在野外群落调查和采样过程中的帮助。
参考文献
Feedback-driven response to multidecadal climatic variability at an Alpine treeline
Linking microbial community composition and soil processes in a California annual grassland and mixed-conifer forest
Seasonal changes in soil microbial communities along a fertility gradient of temperate grasslands
The measurement of soil fungal: bacterial biomass ratios as an indicator of ecosystem self-regulation in temperate meadow grasslands
Temporal variation in soil microbial communities in Alpine tundra
Stability of alpine treeline in Northern Montana, USA
Posi- tive interactions among alpine plants increase with stress
Soil microbial biomass, nutrient availability and nitrogen mineralization potential among vegetation-types in a low arctic tundra landscape
Seasonal dynamics in soil microbial biomass C, N and P in a mixed-oak forest ecosystem of Manipur, North-east India
Spatio-temporal analysis of alpine ecotones: a spatial explicit model targeting altitudinal vegetation shifts
The characteristics of soil microorganism quantity under different alpine grasslands in eastern Qilian Mountain
Microbial biomass as an indicator of soil quality: effects of long-term management and recent soil amendments. In: Doran JW, Coleman DC, Bezdicek DF, Stewart BA eds. Defining Soil Quality for a Sustainable Environment
Variations of soil microbial biomass across four different plant communities along an elevation gradient in Wuyi Mountains, China
Mountain Timberlines: Ecology, Patchiness, and Dynamics
Spatial changes of soil fungal and bacterial biomass from a sub-alpine coniferous forest to grassland in a humid, sub-tropical region
Contribution of working group III to the fourth assessment report of the intergovernmental panel on climate change. In: Metz B, Davidson OR, Bosch PR, Dave R, Meyer LA eds
Seasonal partitioning of nitrogen by plants and soil microorganisms in an alpine ecosystem
Microbial communities, biomass, and activities in soils as affected by freeze thaw cycles
A world-wide study of high altitude treeline temperatures
Living the high life: the mountaintop environment of the Andes harbors a Noah’s ark of previously undocumented species
The suitability of montane ecotones as indicators of global climatic change
Plant-microbe competition for soil amino acids in the alpine tundra: effects of freeze- thaw and dry-rewet events

Amino acids have been shown to be a potentially significant N source for the alpine sedge, Kobresia myosuroides. We hypothesised that freeze-thaw and dry-rewet events allow this plant species increased access to amino acids by disrupting microbial cells, which decreases the size of competing microbial populations, but increases soil amino acid concentrations. To test this hypothesis, we characterized freeze-thaw and dry-rewet events in the field and simulated them in laboratory experiments on plant-soil microcosms. In one experiment, (15)N,(13)C-[2]-glycine was added to microcosms that had previously been subjected to a freeze-thaw or dry-rewet event, and isotopic concentrations in the plant and microbial fractions were compared to non-stressed controls. Microbial biomass and uptake of the labeled glycine were unaffected by the freezing and drying treatments, but microbial uptake of (15)N was lower in the two warmer treatments (dry-rewet and summer control) then in the two colder treatments (freeze-thaw and fall control). Plant uptake of glycine-(15)N was decreased by climatic disturbance, and uptake in plants that had been frozen appeared to be dependent on the severity of the freeze. The fact that intact glycine was absorbed by the plants was confirmed by near equal enrichment of plant tissues in (13)C and (15)N. Plants under optimal conditions recovered 3.5% of the added (15)N and microbes recovered 5.0%. The majority of the (13)C and (15)N label remained in a non-extractable fraction in the bulk soil. To better understand the isolated influences of environmental perturbations on soil amino acid pools and population sizes of amino-acid utilizing microbes, separate experiments were performed in which soils, alone, were subjected to drying and rewetting or freezing and thawing. Potential respiration of glycine and glutamate (substrate-induced respiration; SIR) by the soil microbial communities was unaffected by a single freeze-thaw event. Glycine SIR was decreased slightly ( approximately 10%) by the most extreme drying treatment, but glutamate SIR was not significantly affected. Freezing lowered the concentration of water-extractable amino acids while drying increased their concentration. We interpret the surprising former result as either a decrease in proteolytic activity in frozen soils relative to amino acid uptake, or a stimulation in microbial uptake by physical nutrient release from the soil. We conclude that climatic disturbance does not provide opportunities for increased amino acid uptake by K. myosuroides, but that this plant competes well for amino acid N under non-stressed conditions, especially when soils are warm. We also note that this alpine tundra microbial community's high resistance to freeze-thaw and dry-rewet events is novel and contrasts with studies in other ecosystems.
Changes in soil microbial community structure and function in an alpine dry meadow following spring snow melt
Seasonal dynamics of soil microbial biomass and its significance to indicate soil quality under different vegetations restored on degraded red soils
Responses of alpine biodiversity to climate change
Seasonal variation in nitrogen uptake and turnover in two high-elevation soils: mineralization responses are site-dependent
Winter forest soil respiration controlled by climate and microbial community composition
Seasonal variations and effects of nutrient applications on N and P and microbial biomass under two temperate heathland plants
Signals of range expansions and contractions of vascular plants in the high Alps: observations (1994-2004) at the GLORIA master site Schrankogel, Tyrol, Austria
Neither cold nor snow stops tundra fungi
Soil properties of the subalpine timberline ecotone in Western Sichuan
The soil properties of the subalpine timberline ecotone and adjacent vegetations in Western Sichuan
Abiotic conditions and plant cover differentially affect microbial biomass and community composition on dune gradients
Seasonal dynamics of previously unknown fungal lineages in tundra soils
Biogeochemical consequences of rapid microbial turnover and seasonal succession in soil
Soil microbial quantity and its relations with ecological factors in northern alp region of Hengduan Mountains
Stellera chamaejasme L. increases soil N availability, turnover rates and microbial biomass in an alpine meadow ecosystem on the eastern Tibetan Plateau of China
Seasonal dynamics of microbial community composition and function in oak canopy and open grassland soils
Soil microbial communities are closely associated with aboveground plant communities, with multiple potential drivers of this relationship. Plants can affect available soil carbon, temperature, and water content, which each have the potential to affect microbial community composition and function. These same variables change seasonally, and thus plant control on microbial community composition may be modulated or overshadowed by annual climatic patterns. We examined microbial community composition, C cycling processes, and environmental data in California annual grassland soils from beneath oak canopies and in open grassland areas to distinguish factors controlling microbial community composition and function seasonally and in association with the two plant overstory communities. Every 3 months for up to 2 years, we monitored microbial community composition using phospholipid fatty acid (PLFA) analysis, microbial biomass, respiration rates, microbial enzyme activities, and the activity of microbial groups using isotope labeling of PLFA biomarkers (13C-PLFA). Distinct microbial communities were associated with oak canopy soils and open grassland soils and microbial communities displayed seasonal patterns from year to year. The effects of plant species and seasonal climate on microbial community composition were similar in magnitude. In this Mediterranean ecosystem, plant control of microbial community composition was primarily due to effects on soil water content, whereas the changes in microbial community composition seasonally appeared to be due, in large part, to soil temperature. Available soil carbon was not a significant control on microbial community composition. Microbial community composition (PLFA) and 13C-PLFA ordination values were strongly related to intra-annual variability in soil enzyme activities and soil respiration, but microbial biomass was not. In this Mediterranean climate, soil microclimate appeared to be the master variable controlling microbial community composition and function.
Climate change and high mountain vegetation shifts. In: Broll G, Keplin B eds. Mountain Ecosystems, Studies in Treeline Ecology
Ecological responses to recent climate change
There is now ample evidence of the ecological impacts of recent climate change, from polar terrestrial to tropical marine environments. The responses of both flora and fauna span an array of ecosystems and organizational hierarchies, from the species to the community levels. Despite continued uncertainty as to community and ecosystem trajectories under global change, our review exposes a coherent pattern of ecological change across systems. Although we are only at an early stage in the projected trends of global warming, ecological responses to recent climate change are already clearly visible.
Improvement of the selective respiratory inhibition technique to measure eukaryote: prokaryote ratios in soils
Litter decomposition in two subalpine forests during the freeze-thaw season
Dynamics of microbial biomass during litter decomposition in the alpine forest

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}