


生态化学计量学综合生物学、物理学和化学等学科的基本原理, 通过分析比较生态过程中多重化学元素的平衡关系, 为研究碳(C)、氮(N)、磷(P)等元素在生态系统过程中的耦合关系提供了一种综合方法(Elser et al., 2000)。植物能够主动地调整养分需求, 从而调整体内各元素的相对丰度,灵活地适应周围生长环境的变化, 这些过程将导致植物组织的C:N:P化学计量比值发生变化, 因此植物C、N、P生态化学计量特征研究为探索植物的养分利用状况提供了一种重要的手段(曾德慧和陈广生, 2005)。N和P是植物的最基本营养元素, 并且是自然陆地生态系统的主要限制性元素, 在植物生长过程中的各种生理代谢活动中发挥了十分重要的作用, 相互独立而又相互影响(Han et al., 2005),最终对植物叶片C的固定产生影响。植物体内任何一种元素含量发生变化时都会引起C:N:P化学计量比值的变化, 根据植物N:P的变化, 可以判断植物受N或P哪种营养元素的相对限制, 环境中N、P的缺乏或过量将导致植物体内C:N和C:P的上升或下降(Tjoelker et al., 2005)。
叶是植物通过光合作用获取能源和合成光合产物的主要器官。研究表明, 叶的化学计量学特征可能与功能群或环境梯度有关(Han et al., 2005), 同时植物叶片的N:P值能够反映植物在种群(Gallardo & Covelo, 2005)、群落(Koerselman & Meuleman, 1996; Güsewell, 2004)、生态系统(Tessier & Raynal, 2003)乃至全球(Reich & Oleksyn, 2004)尺度上生长受养分限制的情况。
群落优势种通常对生态群落的组成结构和生态环境的形成具有明显的控制作用。N添加直接影响到植物N营养的供应与循环, 并在一定程度上会改变群落组成结构(Lü et al., 2011)。本文研究了青藏高原高寒草甸6种群落优势种植物叶片C、N、P含量随不同N施入量的变化特征, 探索C:N:P化学计量特征与N、P养分吸收特征的内在联系, 为草地的科学管理提供依据, 加深科学上理解高寒草甸施氮肥的作用。
1 材料和方法
1.1 研究区自然概况
研究样地位于青藏高原东缘甘南藏族自治州玛曲县境内兰州大学高寒草甸与湿地生态系统定位研究站(阿孜分站)。地理坐标为33°39′ N, 101°53′ E, 海拔3 650 m, 年平均气温为2.2 ℃, 年降水量为672 mm (Chu et al., 2008), 属于高寒半湿润半干旱气候, 降雨集中在短暂的夏季7-8月份。年日照时数约2580 h, 平均年霜期大于270天。植被属于高寒草甸类。植被类型为莎草类+禾草类+双子叶杂草类群系, 优势种主要有莎草科的嵩草(Kobresia myosuroides)和矮藨草(Scirpus pumilus); 禾本科的垂穗披碱草(Elymus nutans)、甘青剪股颖(Agrostis hugoniana)和草地早熟禾(Poa pratensis); 双子叶杂草类包括菊科的长毛风毛菊(Saussurea hieracioides)、星状风毛菊(Saussurea stella)和瑞苓草(Saussurea nigrescens); 毛茛科的高原毛茛(Ranunculus tanguticus)、钝裂银莲花(Anemone obtusiloba)、草玉梅(Anemone rivularis); 玄参科的甘肃马先蒿(Pedicularis kansuensis)、中国马先蒿(Pedicularis chinensis)和短腺小米草(Euphrasia regelii); 蔷薇科的莓叶委陵菜(Poten- tilla fragarioides)和鹅绒委陵菜(Potentilla anserina);豆科植物多枝黄耆(Astragalus polycladus)、少花米口袋(Gueldenstaedtia verna)和黄花棘豆(Oxytropis ochrocephala)等。土壤类型为亚高山草甸土, 施氮肥处理前试验地的土壤养分状况见表1。
表1 氮素添加前土壤的养分状况
Table 1
| 土层 Soil layer (cm) | 有机质 Organic matter (g·kg-1) | 全氮 Total nitrogen (g·kg-1) | 全磷 Total phosphorus (g·kg-1) | pH |
|---|---|---|---|---|
| 0-15 | 70.5 | 3.72 | 0.984 | 6.33 |
| 15-30 | 36.3 | 1.95 | 0.712 | 6.44 |
| 30-45 | 24.0 | 1.23 | 0.618 | 6.63 |
1.2 实验设计与样品采集
实验地四周用围栏保护以防止牛羊踩踏和取食, 以减少扰动对实验结果可靠性的影响。5月生长季初期进行施肥实验。根据以往在该地长期施肥试验研究(施氮肥量10 g·m-2 (Ren et al., 2010))、当地最佳施氮肥量(15 g·m-2 (纪亚君, 2006))、氮肥添加对群落物种的影响(当N施加量为25.2 g·m-2时, 一些杂草如西藏大戟(Euphorbia tibetica)消失(仁青吉 等, 2004))等研究资料, 本文设置4个施肥梯度, N0(0 g·m-2)、N1 (20 g·m-2)、N2 (40 g·m-2)、N3 (60 g·m-2), 折合施N量分别为0、7、14和21 g·m-2, 并且将氮肥添加的最大量确定为21 g·m-2。每一处理6个重复, 共计24个样方, 分别编号1-24, 每个样方四角用PVC管标记, 完全随机排列。氮肥选用硝酸铵,溶解于5 L自来水中, 于2012年5月中旬用洒水壶均匀地喷洒在小区内, N0处理只浇水。
选取以上样地6种群落优势种作为研究对象, 分别为: 草玉梅、甘肃马先蒿、莓叶委陵菜、黄花棘豆、垂穗披碱草和嵩草。8月下旬采集每个样方内以上6个物种的叶片装袋, 70 ℃烘干至恒量。
1.3 室内测定
烘干植物样品粉碎后过0.149 mm筛, 用于养分分析。用重铬酸钾容量法(外加热法)测样品C的含量, 采用HClO4-H2SO4消煮法消煮, 消煮后的溶液经定容、沉淀和稀释后, 用SmartChem 200化学分析仪(WestCo Scientific Instrument, Brookfielg, USA)对消煮液进行样品全N和全P分析测定。
1.4 统计分析
植物叶片中C、N、P采用g·kg-1表示, C:N、C:P、N:P化学计量比中的C、N、P采用质量百分比表示。采用Microsoft Excel 2007整理和计算数据, 用SPSS 17.0软件对实验数据进行统计分析, 用LSD多重比较进行显著性检验。采用Origin 10.0作图。
2 结果和分析
2.1 氮肥添加对植物叶片C、N、P的影响
N0处理时, 6种群落优势种植物叶片C含量存在显著的差异(p < 0.0041), 叶片C含量最高的是黄花棘豆, 其次为甘肃马先蒿和嵩草, 再依次为垂穗披碱草、莓叶委陵菜和草玉梅(表2)。黄花棘豆叶片C含量是草玉梅的106%。N0处理时, 6种群落优势种植物叶片P含量也存在显著的差异(p < 0.0031), 叶片P含量最高的也是黄花棘豆, 其余依次为草玉梅、莓叶委陵菜、甘肃马先蒿、嵩草和垂穗披碱草。垂穗披碱草叶片P含量是黄花棘豆叶片的60%。N0处理时, 6种群落优势种叶片N含量从高到低依次为黄花棘豆、莓叶委陵菜、甘肃马先蒿、垂穗披碱草、草玉梅和嵩草, 这些物种之间也存在显著性的差异(p < 0.0028)。
表2 氮肥添加对6种群落优势种植物叶片C、N、P含量(g·kg-1)的影响(平均值±标准误差, n = 6)
Table 2
| 元素 Element | 处理 Treatment | 草玉梅 Anemone rivularis | 甘肃马先蒿Pedicularis kansuensis | 莓叶委陵菜 Potentilla ragarioides | 黄花棘豆 Oxytropis ochrocephala | 垂穗披碱草 Elymus nutans | 嵩草 Kobresia myosuroides |
|---|---|---|---|---|---|---|---|
| C | N0 | 431 ± 14.931a | 464 ± 2.480a | 442 ± 3.724a | 467 ± 1.424a | 459 ± 5.314a | 461 ± 4.993a |
| N1 | 438 ± 15.271a | 466 ± 3.954a | 451 ± 6.083a | 468 ± 3.143a | 460 ± 10.902a | 453 ± 3.444a | |
| N2 | 440 ± 12.064a | 465 ± 6.991a | 456 ± 11.410a | 471 ± 4.913a | 461 ± 11.675a | 463 ± 3.652a | |
| N3 | 445 ± 10.442a | 467 ± 3.992a | 452 ± 13.700a | 461 ± 4.393a | 463 ± 10.904a | 467 ± 6.454a | |
| N | N0 | 16.5 ± 1.094a | 17.3 ± 2.087a | 18.1 ± 1.131a | 24.5 ± 1.113a | 16.7 ± 0.036a | 11.5 ± 0.188a |
| N1 | 18.9 ± 1.298b | 21.1 ± 2.094b | 25.7 ± 2.74b | 22.6 ± 1.190a | 21.2 ± 0.166b | 15.8 ± 0.211b | |
| N2 | 24.1 ± 2.127c | 26.0 ± 1.158c | 29.7 ± 2.144c | 26.8 ± 1.114a | 26.3 ± 0.122c | 18.8 ± 0.212c | |
| N3 | 32.5 ± 2.169d | 32.2 ± 2.087d | 36.7 ± 1.081d | 23.1 ± 1.124a | 31.0 ± 0.366d | 22.8 ± 0.211d | |
| P | N0 | 1.716 ± 0.077a | 1.636 ± 0.053a | 1.672 ± 0.083a | 2.509 ± 0.089a | 1.488 ± 0.070a | 1.591 ± 0.077a |
| N1 | 1.713 ± 0.065a | 1.640 ± 0.070a | 1.682 ± 0.095a | 2.488 ± 0.080a | 1.490 ± 0.081a | 1.575 ± 0.042a | |
| N2 | 1.718 ± 0.076a | 1.638 ± 0.063a | 1.668 ± 0.064a | 2.506 ± 0.114a | 1.478 ± 0.067a | 1.555 ± 0.064a | |
| N3 | 1.721 ± 0.090a | 1.628 ± 0.075a | 1.679 ± 0.071a | 2.511 ± 0.109a | 1.502 ± 0.070a | 1.585 ± 0.073a |
N0, N1, N2, and N3 represent the nitrogen addition treatment of 0, 7, 14, and 21 g·m-2, respectively. Different lower-case letters within species indicate significant differences between treatments (p < 0.05).
N0、N1、N2和N3指氮素添加量分别为0、7、14和21 g·m-2。同一植物不同小写字母表示处理间差异显著(p < 0.05)。
氮肥添加对6种植物叶片C和P含量没有显著影响(p > 0.92), 对黄花棘豆叶片N含量也没有显著影响(p > 0.85), 而显著增加了其他5个物种叶片的N含量, N3处理叶片N含量最高, N0处理叶片N含量最低; N3处理是N0处理的1.85-2倍(表2)。
2.2 氮肥添加对植物叶片C:N、C:P以及N:P的影响
2.2.1 氮肥添加对植物叶片C:N的影响
N0处理时, 6种群落优势种叶片C:N存在显著的差异(p < 0.003)。C:N最高的是嵩草(图1F), 其次依次为垂穗披碱草(图1E)、甘肃马先蒿(图1B)、草玉梅(图1A)、莓叶委陵菜(图1C)和黄花棘豆(图1D)。嵩草叶片C:N是黄花棘豆的2.11倍。氮肥添加对黄花棘豆叶片C:N无显著影响(图1D)(p > 0.073), 4种处理其叶片C:N都保持在19左右。氮肥添加显著降低了其他5种植物叶片C:N (p < 0.0029)。降低程度随N施加量的增加而增加, N0处理的最高, N3处理的最低; 降低幅度最大的为嵩草, 从N0处理的40.2降低到N3处理的20.3, 降低到原来的50%, 降低幅度最小的为草玉梅, 从N0处理的26.8降低到N3处理的14.5, 降低到原来的54%。其余3个物种的C:N也有大幅度的下降。N3处理后6个物种C:N从高到低依次为嵩草、黄花棘豆、垂穗披碱草、甘肃马先蒿、草玉梅和莓叶委陵菜(图1)。
图1
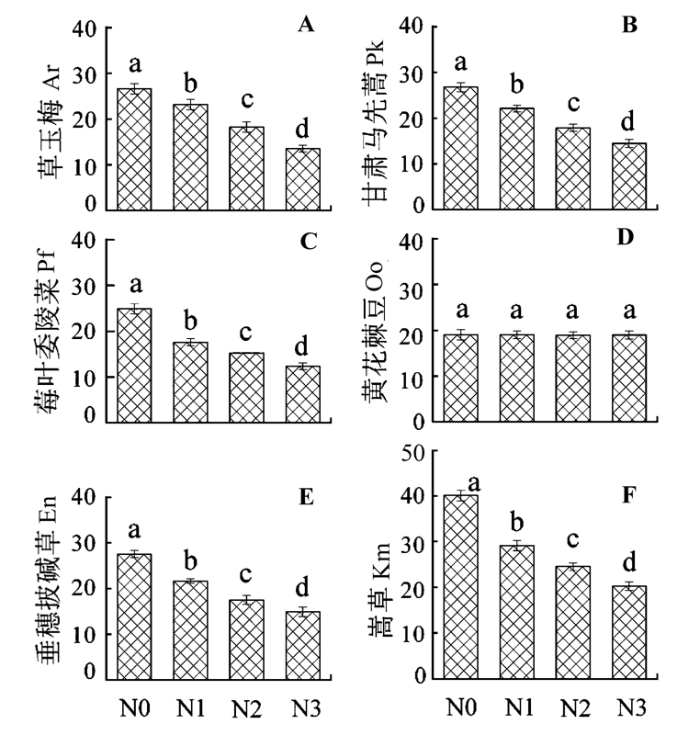
图1
氮肥添加对6种植物叶片C:N的影响(平均值±标准误差, n = 6)。N0、N1、N2和N3指氮素添加量分别为0、7、14和21 g·m-2。不同字母表示处理间差异显著(p < 0.05)。
Fig. 1
Effects of nitrogen (N) addition on leaf C:N in six plant species (mean ± SE, n = 6). N0, N1, N2, and N3 represent the nitrogen addition treatment of 0, 7, 14, and 21 g·m-2, respectively. Different letters indicate significant differences between treatments (p < 0.05). Ar, Anemone rivularis; En, Elymus nutans; Km, Kobresia myosuroides; Oo, Oxytropis ochoocephala; Pk, Pedicularis kansuensis; Pf, Potentilla fragarioides.
2.2.2 氮肥添加对植物叶片C:P的影响
图2
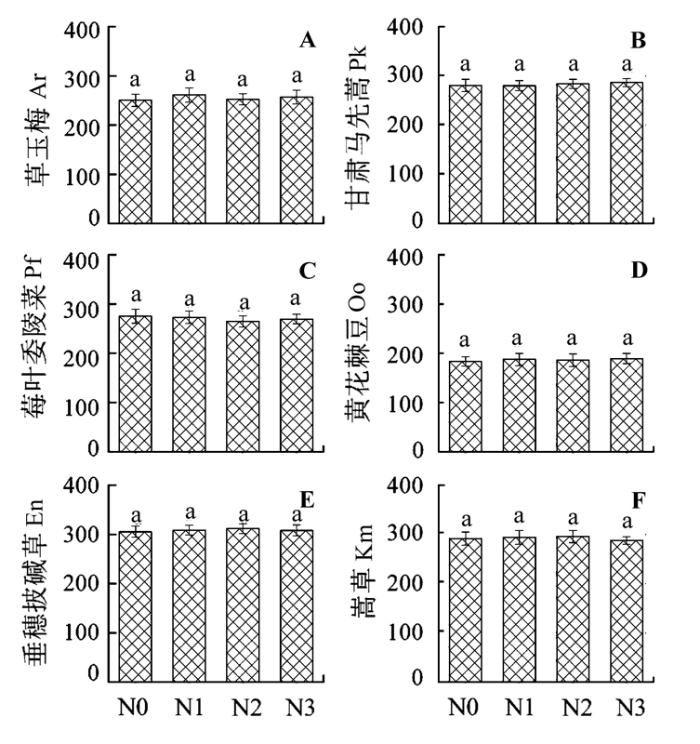
图2
氮肥添加对6种植物叶片C:P的影响(平均值±标准误差, n = 6)。N0、N1、N2和N3指氮素添加量分别为0、7、14和21 g·m-2。不同字母表示处理间差异显著(p < 0.05)。
Fig. 2
Effects of nitrogen (N) addition on leaf C:P in six plant species (mean ± SE, n = 6). N0, N1, N2, and N3 represent the nitrogen addition treatment of 0, 7, 14, and 21 g·m-2, respectively. Different letters indicate significant differences between treatments (p < 0.05). Ar, Anemone rivularis; En, Elymus nutans; Km, Kobresia myosuroides; Oo, Oxytropis ochoocephala; Pk, Pedicularis kansuensis; Pf, Potentilla fragarioides.
2.2.3 氮肥添加对植物叶片N:P的影响
图3
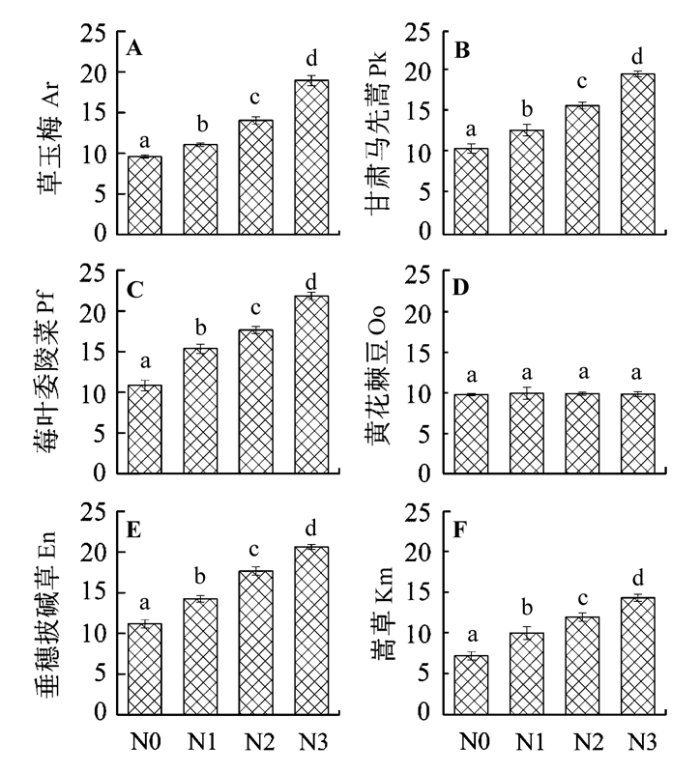
图3
氮肥添加对6种植物叶片N:P的影响(平均值±标准误差, n = 6)。N0、N1、N2和N3指氮素添加量分别为0、7、14和21 g·m-2。不同字母表示处理间差异显著(p < 0.05)。
Fig. 3
Effects of nitrogen (N) addition on leaf N:P in six plant species (mean ± SE, n = 6). N0, N1, N2, and N3 represent the nitrogen addition treatment of 0, 7, 14, and 21 g·m-2, respectively. Different letters indicate significant differences between treatments (p < 0.05). Ar, Anemone rivularis; En, Elymus nutans; Km, Kobresia myosuroides; Oo, Oxytropis ochoocephala; Pk, Pedicularis kansuensis; Pf, Potentilla fragarioides.
3 讨论
3.1 N添加对植物C、N、P营养元素的影响
青藏高原高寒草甸6种群落优势种植物叶片N、P质量浓度分别为11.5-24.5和1.49-2.51 g·kg-1, 除黄花棘豆外, 其他5种群落优势种植物叶片N、P质量浓度为11.5-18.5和1.49-1.72 g·kg-1。我国草地植物叶片N、P质量浓度的平均值为28.6和1.9 g·kg-1 (Han et al., 2005), 青藏高原高寒草甸其他5种群落优势种植物叶片N和P质量浓度含量低于我国草地植物叶片N和P的平均值。豆科植物黄花棘豆叶片N、P质量浓度分别为24.5和2.51 g·kg-1, 我国草地豆科植物叶片N、P的平均值为30.6和2.0 g·kg-1 (He et al., 2008), 其叶片N质量浓度低于我国其他地区豆科植物, 而叶片P质量浓度高于我国其他地区豆科植物。He等(2008)研究表明, 植物叶片P浓度与气候因子在区域尺度上的相关性很小, 区域植物叶片P的变化情况可以通过土壤资源和植物系统发育的变化更好地说明。土壤P含量与土壤母质的理化性质相关, 土壤中的P主要源于岩石的风化和淋洗。在相同土壤P含量条件下, 植物叶片中的P含量差异很大, 这可能与植物系统发育的变化有关。
组成植物体组织的物质可分为结构性、功能性和贮藏性3类。草地生态系统中, C是组成植物体的结构性物质, N和P则为生物体的功能性物质, 其分布和贮量直接关系到草地生态系统功能的正常发挥。对于同一植物种而言, 功能性和贮藏性物质的含量受环境影响变化较大, 结构性物质受环境的影响较小, 含量相对较稳定(Sterner & Elser, 2002), 这可能是在N素添加过程中6种群落优势种叶片C保持不变的原因。本研究中, 随N添加量的增大, 6种群落优势种草玉梅、甘肃马先蒿、莓叶委陵菜、黄花棘豆、垂穗披碱草和嵩草叶片的C和P含量保持不变。而随着N含量显著增加, 5种群落优势种草玉梅、甘肃马先蒿、莓叶委陵菜、垂穗披碱草和嵩草叶片N含量也显著增加, 与安卓等(2011)的N添加对长芒草(Stipa bungeana) N和P含量影响的研究结果一致。
本研究所选6个物种分属4个功能群, 莎草类的嵩草, 禾草类的垂穗披碱草, 草玉梅、甘肃马先蒿、莓叶委陵菜为杂草类, 黄花棘豆为豆科植物。本研究中6个物种叶片N和P含量最高的是豆科的黄花棘豆, 叶片N含量最低的是莎草类的嵩草, 其他4个物种之间没有显著差异; 叶片P含量最低的是禾草类的垂穗披碱草。对于不同的功能群, 植物叶片的N、P含量都表现为: 豆科>非豆科; 与非莎草相比, 莎草叶片具有较低的N含量; 与非莎草和非禾草相比, 莎草和禾草叶片具有较低的N含量和P含量。植物功能性状的变异反映了组织结构和功能的关键平衡(Tilman et al., 1997), 从而在组织水平上, 禾草比非禾草、非豆科植物比豆科植物具有更高的养分利用效率(Tjoelker et al., 2005)。该研究结果与宋彦涛等(2012)的研究结果一致。
本研究结果表明, 氮肥添加对豆科植物黄花棘豆叶片全N含量无显著影响。大多数研究认为豆科植物通过生物固N作用一般能满足其自身N的需求, 因此豆科植物生长过程N的供应与否对其影响并不大。相反, 在植物生长早期N的过量施用会抑制根瘤的形成, 进而降低生物固N效率(Jenkins & Bottomley, 1984)。也有研究表明, 给豆科植物施N肥, 虽然不能提高植物叶片全N的含量, 但能促进地上部分的生长、新叶片的萌生和通过叶片的光合作用为植物根瘤的生物固N提供能量, 同时提高作物产量(Hannaway & Shuler, 1993)。本文结果表明, 虽然N肥添加未增加豆科植物黄花棘豆叶片全N含量, 但N添加提高了植物的光合作用, 增加了地上生物量的积累。
3.2 N添加对植物C、N、P生态化学计量特征的 影响
生态化学计量学应用的一个重要方面是可根据植物叶片的N:P值判断环境对植物生长养分供应的状况(曾德慧和陈广生, 2005)。Koerselman和Meuleman (1996)在欧洲40个地点的湿地生态系统的施肥实验得出, N:P小于14表示生态系统是受N限制, N:P大于16表示生态系统是受P限制, N:P在14-16时, 生态系统同时受N和P的限制或者同时不受二者限制。Güsewell (2004)则认为N:P < 10时, 植被会受N肥的限制;N:P > 20时, 植被会受P肥的限制, 而在这两者之间时, 施肥效应和N:P之间没有明确的关系。然而, 由于研究区域、植物的生长阶段、植物的组织及植物种类存在差异, 这些因素都会影响N:P化学计量比值临界值的变化(Güsewell, 2004; 曾德慧和陈广生, 2005)。He等(2008)对中国草原植被C:N、N:P化学计量比值进行了分析, 指出中国植物缺P导致其N:P值高于全球其他地区草地生态系统的水平。Han等(2005)对中国的753种高等陆地植物进行了大尺度水平上的化学计量特征研究, 认为中国陆地植物生长普遍受到P的限制, 而青藏草原高寒草甸6种群落优势种植物的叶片N含量低于中国其他草原区相应群落, 其N:P为7.3-11.2, 综合判断青藏高原高寒草甸相对于中国其他草原区来说更多是受到N元素限制。
生态化学计量学应用的一个重要方面是可根据植物叶片的N:P值判断环境对植物生长养分供应的状况(曾德慧和陈广生, 2005)。Koerselman和Meuleman (1996)在欧洲40个地点的湿地生态系统的施肥实验得出, N:P小于14表示生态系统是受N限制, N:P大于16表示生态系统是受P限制, N:P在14-16时, 生态系统同时受N和P的限制或者同时不受二者限制。Güsewell (2004)则认为N:P < 10时, 植被会受N肥的限制;N:P > 20时, 植被会受P肥的限制, 而在这两者之间时, 施肥效应和N:P之间没有明确的关系。然而, 由于研究区域、植物的生长阶段、植物的组织及植物种类存在差异, 这些因素都会影响N:P化学计量比值临界值的变化(Güsewell, 2004; 曾德慧和陈广生, 2005)。He等(2008)对中国草原植被C:N、N:P化学计量比值进行了分析, 指出中国植物缺P导致其N:P值高于全球其他地区草地生态系统的水平。Han等(2005)对中国的753种高等陆地植物进行了大尺度水平上的化学计量特征研究, 认为中国陆地植物生长普遍受到P的限制, 而青藏草原高寒草甸6种群落优势种植物的叶片N含量低于中国其他草原区相应群落, 其N:P为7.3-11.2, 综合判断青藏高原高寒草甸相对于中国其他草原区来说更多是受到N元素限制。
生态化学计量学应用的一个重要方面是可根据植物叶片的N:P值判断环境对植物生长养分供应的状况(曾德慧和陈广生, 2005)。Koerselman和Meuleman (1996)在欧洲40个地点的湿地生态系统的施肥实验得出, N:P小于14表示生态系统是受N限制, N:P大于16表示生态系统是受P限制, N:P在14-16时, 生态系统同时受N和P的限制或者同时不受二者限制。Güsewell (2004)则认为N:P < 10时, 植被会受N肥的限制;N:P > 20时, 植被会受P肥的限制, 而在这两者之间时, 施肥效应和N:P之间没有明确的关系。然而, 由于研究区域、植物的生长阶段、植物的组织及植物种类存在差异, 这些因素都会影响N:P化学计量比值临界值的变化(Güsewell, 2004; 曾德慧和陈广生, 2005)。He等(2008)对中国草原植被C:N、N:P化学计量比值进行了分析, 指出中国植物缺P导致其N:P值高于全球其他地区草地生态系统的水平。Han等(2005)对中国的753种高等陆地植物进行了大尺度水平上的化学计量特征研究, 认为中国陆地植物生长普遍受到P的限制, 而青藏草原高寒草甸6种群落优势种植物的叶片N含量低于中国其他草原区相应群落, 其N:P为7.3-11.2, 综合判断青藏高原高寒草甸相对于中国其他草原区来说更多是受到N元素限制。
土壤养分组成是植物的外界环境中非常重要的因子, 植物光合作用和矿质代谢等过程与土壤养分供应状况有显著相关性。在植物体内C、N、P都是相互作用的, 植物体的C:N和C:P通常能够反映出植物对N和P的利用效率高低, 可以在一定程度上判断环境对植物生长的N和P养分供应的状况。叶片N含量是植物光合能力的决定性因素, 光合作用时碳水化合物的形成(C)过程中需要大量蛋白酶(N)的参与, 而蛋白酶的装配形成前提是需要大量遗传物质核酸的复制和转录(P)(Sterner & Elser, 2002), 因此植物体的C与N、P含量具有明显的相关性, 植物体内光合作用和矿质代谢之间存在一定的内在联系。光合作用C的同化和植物营养元素的吸收是属于不同途径。通常C不是植物生长的限制元素, C在大多数植物体内含量很高且变异较小。因此影响C:N和C:P的主要因素是N和P含量的变化(Hedin, 2004)。本研究结果表明, 除黄花棘豆外, 其他5种群落优势种草玉梅、甘肃马先蒿、莓叶委陵菜、垂穗披碱草和嵩草叶片的C:P没有明显改变而C:N随N素添加量的增加而逐渐降低。由于叶片的N、P含量反映了土壤N、P的有效性, 研究结果表明外源N素添加增加了土壤的有效N供应, 随N素添加量的增加, 土壤中的有效N供应也变得相对充足, 植物叶片中的N也迅速增加, N:P也由未添加氮素前的7.3-11.2增加到添加后的大于16, 说明随着N素的增加, 植物受P元素的限制增加。杨阔等(2010)对青藏高原草地植物群落冠层叶片N、P化学计量学的研究发现, N:P值随着温度的升高而增大, 解释了青藏高原高寒草甸群落优势种N:P值低的原因, 这一结果与Reich和Oleksyn (2004)在全球尺度上的研究结果完全一致。
综合分析表明, 青藏高原高寒草甸群落优势种豆科植物黄花棘豆叶片全N含量低于我国其他草地豆科植物, P含量高于我国其他草地豆科植物; 其他5个优势物种叶片N、P含量都低于我国其他草地非豆科植物, 说明该区豆科植物生长主要是受N元素的限制, 而非豆科植物的生长受N、P两种元素的限制; 未添加N肥时, 6个物种N:P在7.3-11.2之间, 与我国其他地区草地植物叶片N:P相比较, 植物叶片的N:P都很低, 说明该地区植物生长更受N元素的限制。N肥添加未改变6个植物叶片的C和P含量, 也未改变黄花棘豆叶片N含量, 说明豆科植物黄花棘豆叶片N含量对N元素添加不敏感, 其他5个物种叶片N含量随着N添加量的增加而增加, 随着N添加量的增加, 植物生长受P元素限制。该研究结果可为该区合理施肥和科学管理提供科学依据。
致谢
感谢兰州大学高寒草甸与湿地生态系统定位研究站杜国祯教授以及李宏林、刘旭东、周小龙、张鹏飞、郭治和王静等同学在野外工作中给予的帮助。
参考文献
Stipa bungeana of steppe grasslands in the Loess Plateau, China.
氮素添加对黄土高原典型草原长芒草氮磷重吸收率及C:N:P化学计量特征的影响
Balance between facilitation and resource competition determines biomass-density relationships in plant populations
Biological stoichiometry from genes to ecosystems
Spatial pattern and scale of leaf N and P concentration in a Quereus robur population
N:P ratios in terrestrial plants: variation and functional significance
Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China
Nitrogen fertilization in alfalfa production
Leaf nitrogen: phosphorus stoichiometry across Chinese grassland biomes
Global organization of terrestrial plant-nutrient interactions
Seasonal response of uninoculated alfalfa to N fertilizer: soil N, nodule turnover, and symbiotic effectiveness of Rhizobium meliloti
Effects of nitrogen on the grass yield of Qinghai alpine meadow
青海高寒草甸施氮肥增产效应浅析
The vegetation N:P ratio: a new tool to detect the nature of nutrient limitation
Nutrient resorption response to fire and nitrogen addition in a semi-arid grassland
Global patterns of plant leaf N and P in relation to temperature and latitude
Restoration of degraded typical alpine meadowland on the Qinghai-Tibetan Plateau―effect of fertilizing and cutting on grassland quality
青藏高原典型高寒草甸退化草地的恢复——施肥刈割对草地质量的影响
Effects of resource additions on species richness and ANPP in an alpine meadow community
Leaf nitrogen and phosphorus stoichiometry in 80 herbaceous plant species of Songnen grassland in Northeast China
松嫩草地80种草本植物叶片氮磷化学计量特征
Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation
The influence of functional diversity and composition on ecosystem processes
Linking leaf and root trait syndromes among 39 grassland and savannah species
Canopy leaf N and P stoichiometry in grassland communities of Qinghai- Tibetan Plateau, China
青藏高原草地植物群落冠层叶片氮磷化学计量学分析

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}