


有花植物的花对称性与传粉系统密切相关。典型的两侧对称花有融合的管状花冠, 并且只有一个对称平面。这种结构使传粉者只有采用正确的定位或姿势才能附在花上进行有效地传粉。这就造成昆虫的取食和传粉变得更加困难, 要求有更高的精确性。辐射对称的花则相反, 拥有两个或更多的对称平面(Luo et al., 1996), 使得传粉昆虫更容易附着, 完成传粉过程。所以, 相对有更多的传粉昆虫类群可以对辐射对称的花进行有效地传粉(Cronk & Möller, 1997; Richards, 1997)。在有花植物1.4亿年的历史中, 从辐射对称花到两侧对称花的进化转变是十分重要的一步(Stebbins, 1974)。因为花的结构和对称性影响着传粉者的行为, 所以当较原始的辐射对称的被子植物的花发生适应性进化时, 往往会影响植物和相应的传粉昆虫的对应关系(Stebbins, 1950)。研究表明: 两侧对称类型是多次独立出现的, 同时伴随着传粉系统的特化, 比如兰科和玄参科植物表现出非常特化的传粉系统(Neal et al., 1998)。
花对称性和传粉昆虫的特异性能保证花粉传递有更高的效率(Faegri & van der Pijl, 1979; Ingrouille,1992; 李交昆和唐璐璐, 2012)。因为花粉传递的效率依赖于花和特定传粉者的身体吻合(Schemske & Horvitz, 1989; Wolfe & Barrett, 1989; Harder & Barrett, 1993), 对那些大于或小于种群平均值的花, 其花粉的输入和输出会受到影响, 进而影响植物的适合度(Berg, 1960; Fenster, 1991)。花特征的一些细微的改变甚至都可能影响传粉过程(Campbell et al., 1991; Harder & Barrett, 1993)。因此, 对两侧对称的花, 自然选择应该维持花的大小不变, 因为稳定选择和定向选择会减少花特征的变异(Lande & Arnold, 1983)。与之相反, 假如辐射对称的花由许多泛化的传粉者来传粉(Richards, 1997), 那些偏离平均大小的花不应有较多的适合度的损失。因此, 传粉者施加的稳定选择使两侧对称的花更加稳定, 变异性更低(Berg, 1960)。相对来说, 植物营养性状, 比如叶片的大小更可能依赖于环境因素(比如光照条件、环境温度或土壤条件等), 因此可能表现出更高的可塑性(Berg, 1960; Bradshaw, 1965)。而且, 在传粉昆虫的选择作用下, 花特征之间的整合性要高于营养性状间的整合性, 花功能上的稳定性导致花性状和营养器官性状间的相关性也会减弱(Berg, 1960)。目前对Berg假说进行检验的证据并不完全一致(Armbruster et al., 1999; Wolfe & Krstolic, 1999; Meng et al., 2008; Gong & Huang, 2009; Pélabon et al., 2013)。本研究中, 我们对青藏高原东部高寒草甸植物群落常见开花植物进行了研究, 通过测定花特征和叶片特征及控制系统发育关系上的比较, 主要检验Berg的两个假设: 1)在一个植物群落中, 具有两侧对称花的物种其花大小的变异程度应该比辐射对称植物的更小; 2)特化的两侧对称植物花大小的变异比叶片大小的变异更低。
1 材料和方法
1.1 研究地点
研究地点位于青藏高原东部的甘肃省玛曲县, 研究植物群落属于典型高寒草甸植物群落。该研究地点(101.88° E, 35.97° N)地处青藏高原东北缘, 平均海拔3500 m, 年平均气温1.2 ℃, 最冷的12月到翌年2月平均气温-9.3 ℃, 最热的6到8月平均气温10.0 ℃, 年降水量620 mm。
1.2 研究材料及方法
2010年5-8月, 我们对植物群落内的开花植物进行了测定, 每个物种测定20株, 共测定了50个物种(约占该群落内开花植物的90%以上), 包括31种辐射对称植物和19种两侧对称植物。花和叶片的测定选择在每个物种的盛花期进行, 有多个花的物种(指产生花序)每个个体选择至少3朵处于花中期的花, 用数显游标卡尺测量花的性状(精确到0.1 mm), 同时测定植物茎部从基部往上第二片叶的长和宽。对于辐射对称的花, 一般测量花冠直径、花瓣或花萼的长和宽, 或花冠筒长; 对两侧对称的花, 除了测量花冠直径, 花瓣长和宽, 还要测量相对特化的结构, 如距长、盔高、花冠筒长、上下唇瓣间距等, 这些都作为衡量植物花大小的指标(表1)。
表1 50个物种花及叶特征的变异系数*
Table 1
| 科 Family | 特征 Trait | 变异系数 Mean CV | |
|---|---|---|---|
| 辐射对称物种 Radial species | |||
| 喉毛花 Comastoma pulmonarium4 | 龙胆科 Gentianaceae | 叶大小(长、宽) Leaf size (length, width) | 0.242 |
| 花大小(花直径、瓣长、瓣宽) Flower size (diameter, length, width) | 0.144 | ||
| 湿生扁蕾 Gentianopsis paludosa4 | 龙胆科 Gentianaceae | 叶大小(长、宽) Leaf size (length, width) | 0.355 |
| 花大小(花直径、瓣长、瓣宽) Flower size (diameter, length, width) | 0.134 | ||
| 瞿麦 Dianthus superbus6 | 石竹科 Caryophyllaceae | 叶大小(长、宽) Leaf size (length, width) | 0.262 |
| 花大小(花直径、瓣长、瓣宽) Flower size (diameter, length, width) | 0.118 | ||
| 沼生柳叶菜 Epilobium palustre6 | 柳叶菜科 Onagraceae | 叶大小(长、宽) Leaf size (length, width) | 0.280 |
| 花大小(花直径、瓣长、瓣宽) Flower size (diameter, length, width) | 0.136 | ||
| 柳兰 Chamerion angustifolium6 | 柳叶菜科 Onagraceae | 叶大小(长、宽) Leaf size (length, width) | 0.200 |
| 花大小(花直径、瓣长、瓣宽) Flower size (diameter, length, width) | 0.055 | ||
| 花荵 Polemonium caeruleum6 | 花荵科 Polemoniaceae | 叶大小(长、宽) Leaf size (length, width) | 0.239 |
| 花大小(花直径、瓣长、瓣宽) Flower size (diameter, length, width) | 0.082 | ||
| 莓叶委陵菜 Potentilla fragarioides2 | 蔷薇科 Rosaceae | 叶大小(长、宽) Leaf size (length, width) | 0.198 |
| 花大小(花直径、瓣长、瓣宽) Flower size (diameter, length, width) | 0.119 | ||
| 银露梅 Potentilla glabra2 | 蔷薇科 Rosaceae | 叶大小(长、宽) Leaf size (length, width) | 0.185 |
| 花大小(花直径、瓣长、瓣宽) Flower size (diameter, length, width) | 0.119 | ||
| 金露梅 Potentilla fruticosa2 | 蔷薇科 Rosaceae | 叶大小(长、宽) Leaf size (length, width) | 0.222 |
| 花大小(花直径、瓣长、瓣宽) Flower size (diameter, length, width) | 0.064 | ||
| 丝叶毛茛 Ranunculus tanguticus var. capillaceus1 | 毛茛科 Ranunculaceae | 叶大小(长、宽) Leaf size (length, width) | 0.575 |
| 花大小(花直径、瓣长、瓣宽) Flower size (diameter, length, width) | 0.136 | ||
| 钝裂银莲花 Anemone obtusiloba1 | 毛茛科 Ranunculaceae | 叶大小(长、宽) Leaf size (length, width) | 0.464 |
| 花大小(花直径、瓣长、瓣宽) Flower size (diameter, length, width) | 0.140 | ||
| 高山唐松草 Thalictrum alpinum1 | 毛茛科 Ranunculaceae | 叶大小(长、宽) Leaf size (length, width) | 0.113 |
| 花大小(花直径、瓣长、瓣宽) Flower size (diameter, length, width) | 0.154 | ||
| 小花草玉梅 Anemone rivularis var. flore-minore1 | 毛茛科 Ranunculaceae | 叶大小(长、宽) Leaf size (length, width) | 0.223 |
| 花大小(花直径、瓣长、瓣宽) Flower size (diameter, length, width) | 0.143 | ||
| 毛茛状金莲花 Trollius ranunculoides1 | 毛茛科 Ranunculaceae | 花大小(花直径、瓣长、瓣宽) Flower size (diameter, length, width) | 0.137 |
| 甘青老鹳草 Geranium pylzowianum6 | 牻牛儿苗科 Geraniaceae | 叶大小(长、宽) Leaf size (length, width) | 0.127 |
| 花大小(花直径、瓣长、瓣宽) Flower size (diameter, length, width) | 0.096 | ||
| 鼠掌老鹳草 Geranium sibiricum6 | 牻牛儿苗科 Geraniaceae | 叶大小(长、宽) Leaf size (length, width) | 0.281 |
| 花大小(花直径、瓣长、瓣宽) Flower size (diameter, length, width) | 0.155 | ||
| 水团花 Adina pilulifera3 | 茜草科 Rubiaceae | 叶大小(长、宽) Leaf size (length, width) | 0.236 |
| 花大小(花直径、瓣长、瓣宽) Flower size (diameter, length, width) | 0.114 | ||
| 直立点地梅 Androsace erecta6 | 报春花科 Primulaceae | 花大小(花直径、瓣长、瓣宽) Flower size (diameter, length, width) | 0.067 |
| 黄帚橐吾 Ligularia virgaurea5 | 菊科 Asteraceae | 叶大小(长、宽) Leaf size (length, width) | 0.233 |
| 花大小(花直径、瓣长、瓣宽) Flower size (diameter, length, width) | 0.161 | ||
| 毛连菜 Picris hieracioides6 | 菊科 Asteraceae | 叶大小(长、宽) Leaf size (length, width) | 0.223 |
| 花大小(花直径、瓣长、瓣宽) Flower size (diameter, length, width) | 0.153 | ||
| 沼生苦苣菜 Sonchus palustris6 | 菊科 Asteraceae | 花大小(花直径、瓣长、瓣宽) Flower size (diameter, length, width) | 0.100 |
| 黑褐千里光 Senecio atrofuscus6 | 菊科 Asteraceae | 花大小(花直径、瓣长、瓣宽) Flower size (diameter, length, width) | 0.127 |
| 缘毛紫菀 Aster souliei6 | 菊科 Asteraceae | 花大小(花直径、瓣长、瓣宽) Flower size (diameter, length, width) | 0.132 |
| 高山紫菀 Aster alpinus6 | 菊科 Asteraceae | 叶大小(长、宽) Leaf size (length, width) | 0.172 |
| 花大小(花直径、瓣长、瓣宽) Flower size (diameter, length, width) | 0.172 | ||
| 科 Family | 特征 Trait | 变异系数 Mean CV | |
| 甘青微孔草 Microula pseudotrichocarpa3 | 紫草科 Boraginaceae | 花大小(花直径、瓣长、瓣宽) Flower size (diameter, length, width) | 0.105 |
| 刺芒龙胆 Gentiana aristata4 | 龙胆科 Gentianaceae | 花大小(花冠筒长、花冠直径、花冠裂片长、花冠裂片宽、花冠筒直径) Flower size (corolla tube length, corolla diameter, corolla lobe length, corolla lobe width, corolla tube diameter) | 0.093 |
| 鳞叶龙胆 Gentiana squarrosa4 | 龙胆科 Gentianaceae | 叶大小(长、宽) Leaf size (length, width) | 0.223 |
| 花大小(花冠筒长、花冠直径、花冠裂片长、花冠裂片宽、花冠筒直径) Flower size (corolla tube length, corolla diameter, corolla lobe length, corolla lobe width, corolla tube diameter) | 0.240 | ||
| 狼毒 Stellera chamaejasme6 | 瑞香科 Thymelaeaceae | 花大小(花直径、瓣长、瓣宽) Flower size (diameter, length, width) | 0.128 |
| 夏河缬草 Valeriana xiaheensis5 | 败酱科 Valerianaceae | 花大小(花冠筒长、花冠筒直径、花直径、花瓣长、花瓣宽) Flower size (corolla tube length, corolla tube diameter, corolla diameter, petal length and width) | 0.220 |
| 高山韭 Allium sikkimense | 百合科 Liliaceae | 花大小(花瓣长、宽) Flower size (length and width) | 0.115 |
| 高原鸢尾 Iris collettii | 鸢尾科 Iridaceae | 花大小(外花被直径、内花被直径、外花瓣长、外花瓣宽、内花瓣长、内花瓣宽) Flower size (outside perianth diameter, inside perianth diameter, outside petal length and width, inside petal length and width ) | 0.080 |
| 两侧对称物种 Bilateral species | |||
| 碎米蕨叶马先蒿 Pedicularis cheilanthifolia5 | 列当科 Orobanchaceae | 叶大小(长、宽) Leaf size (length and width) | 0.248 |
| 花大小(花冠筒长、下唇长、下唇宽、花冠与下唇间距) Flower size (corolla tube length, Lower lip length and width, distance between corolla and lower lip) | 0.102 | ||
| 甘肃马先蒿 Pedicularis kansuensis5 | 列当科 Orobanchaceae | 叶大小(长、宽) Leaf size (length and width) | 0.101 |
| 花大小(花冠筒长、下唇长、下唇宽、花冠与下唇间距) Flower size (corolla tube length, Lower lip length and width, distance between corolla and lower lip) | 0.077 | ||
| 四川马先蒿 Pedicularis szetschuanica5 | 列当科 Orobanchaceae | 叶大小(长、宽) Leaf size (length and width) | 0.286 |
| 花大小(花冠筒长、下唇长、下唇宽、花冠与下唇间距) Flower size (corolla tube length, Lower lip length and width, distance between corolla and lower lip) | 0.124 | ||
| 露蕊乌头 Gymnaconitum gymnandrum1 | 毛茛科 Ranunculaceae | 叶大小(长、宽) Leaf size (length and width) | 0.259 |
| 花大小(盔高、萼片直径、侧萼片宽、侧萼片高、侧萼片间距) Flower size (galea height, sepal diameter, Lateral sepal width and length, distance between lateral sepals) | 0.122 | ||
| 展毛翠雀花 Delphinium kamaonense1 | 毛茛科 Ranunculaceae | 叶大小(长、宽) Leaf size (length and width) | 0.124 |
| 花大小(花冠直径、距长、上萼片宽、下萼片宽) Flower size (corolla diameter, spur length, upper sepal width, under sepal width) | 0.068 | ||
| 黄花棘豆 Oxytropis ochrocephala2 | 豆科 Fabaceae | 叶大小(长、宽) Leaf size (length and width) | 0.247 |
| 花大小(翼瓣长、龙骨瓣长、旗瓣长) Flower size (wing petal length, keel length, flag petal length) | 0.158 | ||
| 甘肃米口袋 Gueldenstaedtia gansuensis2 | 豆科 Fabaceae | 叶大小(长、宽) Leaf size (length and width) | 0.092 |
| 花大小(龙骨瓣长、旗瓣长、龙骨瓣宽、翼瓣长、旗瓣宽) Flower size (keel length and width, wing petal length, flag petal length and width) | 0.199 | ||
| 青海苜蓿 Medicago archiducis-nicolai2 | 豆科 Fabaceae | 叶大小(长、宽) Leaf size (length and width) | 0.292 |
| 花大小(龙骨瓣长、旗瓣长、龙骨瓣宽、翼瓣长、旗瓣宽) Flower size (keel length and width, wing petal length and width, flag petal length and width) | 0.09 | ||
| 歪头菜 Vicia unijuga2 | 豆科 Fabaceae | 叶大小(长、宽) Leaf size (length and width) | 0.198 |
| 花大小(翼瓣长、龙骨瓣长、旗瓣长) Flower size (wing petal length, keel length, flag petal length) | 0.208 | ||
| 苦马豆 Sphaerophysa salsula2 | 豆科 Fabaceae | 叶大小(长、宽) Leaf size (length and width) | 0.064 |
| 花大小(翼瓣长、龙骨瓣长、旗瓣长) Flower size (wing petal length, keel length, flag petal length) | 0.05 | ||
| 山黧豆 Lathyrus quinquenervius2 | 豆科 Fabaceae | 叶大小(长、宽) Leaf size (length and width) | 0.276 |
| 花大小(翼瓣长、龙骨瓣长和宽、旗瓣长) Flower size (wing petal length, keel length and width, flag petal length) | 0.124 | ||
| 科 Family | 特征 Trait | 变异系数 Mean CV | |
| 救荒野豌豆 Vicia sativa2 | 豆科 Fabaceae | 花大小(翼瓣长、龙骨瓣长、旗瓣长) Flower size (wing petal length, keel length, flag petal length) | 0.060 |
| 大叶鼠尾草 `Salvia grandifolia4 | 唇形科 Lamiaceae | 叶大小(长、宽) Leaf size (length and width) | 0.181 |
| 花大小(花冠筒长、下唇长、下唇宽、上下唇间距) Flower size (corolla tube length, lower lip length and width, distance between lips) | 0.099 | ||
| 藏荆芥 Nepeta angustifolia4 | 唇形科 Lamiaceae | 叶大小(长、宽) Leaf size (length and width) | 0.187 |
| 花大小(花萼筒长、花萼筒直径、上唇长、上唇宽、下唇长、下唇宽) Flower size (corolla tube length, corolla tube diameter, upper lip length and width, lower lip length and width) | 0.076 | ||
| 异色黄芩 Scutellaria discolor4 | 唇形科 Lamiaceae | 叶大小(长、宽) Leaf size (length and width) | 0.225 |
| 花大小(花萼筒长、花萼筒直径、上唇长、上唇宽、下唇长、下唇宽) Flower size (corolla tube length, corolla tube diameter, upper lip length and width, lower lip length and width) | 0.080 | ||
| 毛果婆婆纳 Veronica eriogyne3 | 玄参科 Scrophulariaceae | 叶大小(长、宽) Leaf size (length and width) | 0.111 |
| 花大小(花萼筒直径、上唇宽、下唇宽) Flower size (corolla tube diameter, upper lip width, lower lip width) | 0.129 | ||
| 短腺小米草 Euphrasia regelii3 | 玄参科 Scrophulariaceae | 叶大小(长、宽) Leaf size (length and width) | 0.204 |
| 花大小(上唇长、上唇宽、下唇长、下唇宽、上下唇间距) Flower size (upper lip length and width, lower lip length and width, distance between lips) | 0.151 | ||
| 灯笼草 Clinopodium polycephalum4 | 唇形科 Lamiaceae | 叶大小(长、宽) Leaf size (length and width) | 0.167 |
| 花大小(花萼筒长、上唇长、上唇宽、下唇长、下唇宽) Flower size (corolla tube length, upper lip length and width, lower lip length and width) | 0.118 | ||
| 陕西紫堇 Corydalis shensiana6 | 罂粟科 Papaveraceae | 叶大小(长、宽)Leaf size (length and width) | 0.264 |
| 花大小(上花瓣长、距长、下花瓣长、下花瓣宽、上下瓣间距) Flower size (upper petal length, lower petal length and width, distance between petals, spur length) | 0.180 | ||
* 系统发育独立比较共分为6个分类组, 拉丁名右上角数字表明该物种所在分类组; 高山韭和高原鸢尾没有相匹配的两侧对称物种, 因此并没有归入任何分类组。
* the species studied were categorized into six groups of taxonomic pairs, the superscript numbers following species names indicate the corresponding taxonomic group; Allium sikkimense and Iris collettii were excluded due to lack of matching bilateral species.
1.3 数据处理与分析
计算每个物种所测量花特征及叶片大小(长和宽)的变异系数(CV)。对于测量了多个花的个体, 估计个体每个特征的平均值, 用不同花特征CV的平均值代表个体花大小的平均CV, 物种花大小和叶片大小的CV (或平均大小)为个体水平CV (或大小)的平均值。在此基础上, 两侧对称物种和辐射对称物种之间的CV (和平均花大小)的差异用非参数Mann-Whitney U检验。
种间的比较会因为不同的物种在系统发生上不独立而造成误差(Ridley, 1983; Harvey & Pagel, 1991), 所以我们采用系统发生上成对的比较来减少起源差异(Healy & Guilford, 1990; Read, 1991)。 根据Chase等(1993)和The Angiosperm Phylogeny Group (2009)的系统发生树, 我们用成对比较方法(Møller & Birkhead, 1992)比较亲缘关系最近且花对称性不同的物种对。所有测定的50个物种代表了6对独立的分类组(表1), 每组都包含辐射对称物种和两侧对称物种, 然后对这6组的两种对称类型的物种进行非参数Mann-Whitney U检验。
2 结果
图1
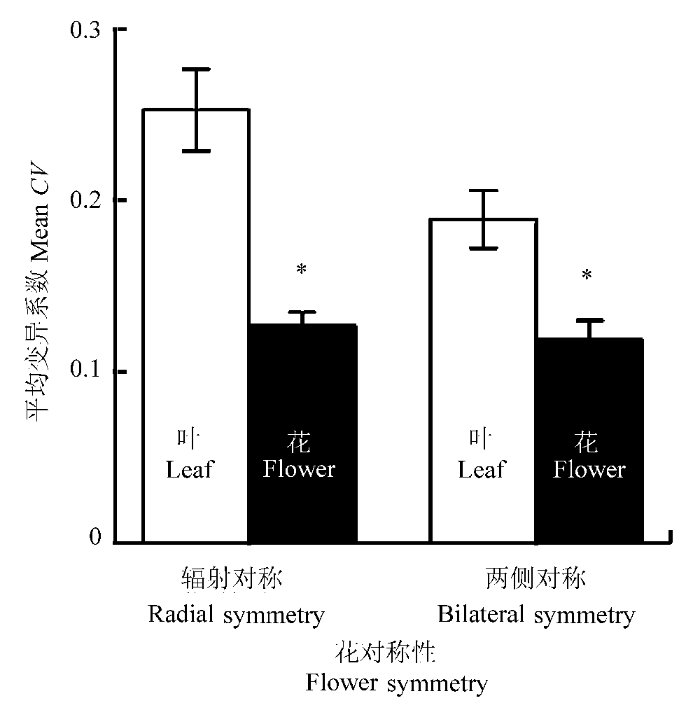
图1
辐射对称与两侧对称物种花和叶大小变异系数的比较(平均值±标准误差)。*, 花和叶大小变异系数差异显著(p < 0.05)。
Fig. 1
Comparisons of the coefficient of variation (CV) in flower size and leaf size between species with radial and bilateral flowers (mean ± SE). *, there is a significant difference between the CV of flower size and that of leaf size (p < 0.05).
用系统发育独立对比分析(6组独立的对比)同样没有发现两种对称性之间花大小的变异程度有显著的差异(图2, Mann-Whitney U, Z = - 0.964, p = 0.394)。
图2
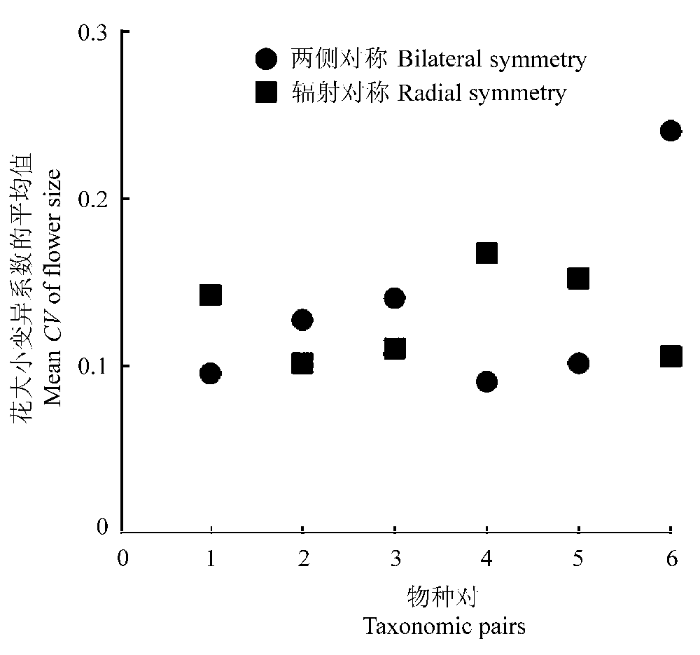
图2
辐射对称与两侧对称物种花大小变异的系统发育独立比较。共6个分类组, 每一组包括两侧对称和辐射对称的物种对, 分别估计变异系数的平均值, 然后对6组值进行非参数检验: Mann-Whitney U, Z = - 0.964, p = 0.394。
Fig. 2
Pairwise phylogenetic comparisons of flower size in bilaterally and radially symmetrical species. The paired points represent family means or means of higher taxonomic groups for six independent cases where the phylogeny branches dichotomously into a bilateral and radial pair. A non-parametric test was used to examine the differences in mean coefficient of variation (CV) of the six pairs, Mann-Whitney U, Z = - 0.964, p = 0.394.
3 讨论
因为具有两侧对称花的植物的繁殖成功对传粉者传粉活动的精确性要求非常高, 所以在植物-传粉者的相互作用下, 两侧对称花的结构与其高度特化的传粉昆虫的传粉部位(头、胸、背、腹或口器等)有非常高的吻合度(比如兰科和玄参科植物的花与其特有传粉者)。而具有辐射对称花的植物往往由多种传粉昆虫类群传粉, 其花结构与传粉者没有特别强的专一性。因此, Berg假说认为传粉者的选择导致两侧对称植物花大小的变异比辐射对称植物的更低。而且, 植物营养特征更易受环境影响, 比花特征有更高的变异性(Berg, 1960; Armbruster et al., 1999; Meng et al., 2008; Pélabon et al., 2013)。我们对高山草甸植物群落50个物种的研究结果表明, 不论是两侧对称植物还是辐射对称植物, 花大小的变异都显著低于叶片大小的变异。但是, 辐射对称物种花大小的变异程度和两侧对称物种的相似, 即使控制系统发育的影响后, 也没有发现显著的差异。
花的形态和大小一直以来被认为是保守的性状(Stebbins, 1950; Bradshaw, 1965)。实际上, 由于花形态的稳定性, 使得花成为非常有效的分类依据, 尽管一些物种的花大小仍然表现出显著的变异(Campbell et al., 1991; Wolfe, 1992; Conner & Rush, 1996)。人们预测两侧对称花大小的变异比辐射对称花的更低(Berg, 1960), 此观点得到了一些实验的证实(Wolfe & Krstolic, 1999; Ushimaru & Hyodo, 2005; Herrera et al., 2008; van Kleunen et al., 2008; Gong & Huang, 2009)。Fenster (1991)对10种蜂鸟传粉植物的研究也证实花冠筒深度的表型方差和花冠筒的平均深度是负相关的, 这说明花冠深度是受选择作用的, 选择使其变异减小。Wolfe和Krstolic (1999)对31个物种(17个辐射对称, 14个两侧对称)的研究证实了两侧对称花大小的变异比辐射对称花的更低。Gong和Huang (2009)对中国横断山脉高山草甸植物群落27种开花植物的调查也支持Berg假说——两侧对称植物的花展示的变异显著低于辐射对称植物, 而且主要与植物传粉系统的特化程度有关。Armbruster等(1999)同样发现特化的花两侧对称植物的花的变异低于辐射对称花植物或风媒植物; 而且, 不论花两侧对称或辐射对称或风媒植物, 花大小的变异显著低于叶大小的变异, 我们的结果与之相似。不论花的对称性或特化程度, 植物花特征可能总是处于传粉昆虫施加的强的选择压力之下, 以保证有效地吸引和传粉(Møller & Eriksson, 1994; Armbruster et al., 1999), 从而表现出比营养特征更低的变异性。例如, Pérez-Barrales等(2007)对Narcissus papyraceus不同种群的研究表明: 由蛾类传粉、更为特化的种群花特征的变异性显著低于营养特征的, 而且, 这种差异明显高于由蝇类传粉、特化程度较低的种群。
我们的结果显然不支持Berg假设中的“在传粉者作用下, 两侧对称花的大小的变异程度低于辐射对称花”。因为传粉昆虫的传粉活动通常依赖于植物群落的物种组成和局域气候环境, 所以研究地点植被和环境的差异可能对开花植物-传粉昆虫相互作用的强度有影响。比如Gong和Huang (2009)研究了中国横断山脉高山草甸植物群落, 该地区生物多样性(包括植物及传粉昆虫种类)远高于我们研究的青藏高原地区, 传粉昆虫与开花植物的相互作用可能更强。而青藏高原地区由于海拔高, 气候环境波动较大, 有效传粉昆虫类群相对单一(以熊蜂和蝇类为主), 可能会减弱传粉昆虫的选择作用。另外, 因为植物花对称性是从花结构及对称面的多少和传粉行为的精确性来考虑, 简单地将植物分为两侧对称和辐射对称类型可能掩盖了真实的传粉者类群的特化程度。比如很多管状花的植物(如柳叶菜科植物)往往传粉特化程度非常高, 仅由熊蜂或蛾类传粉, 这类传粉昆虫可能对花特征有更强的选择作用, 导致花的变异性降低。如果从花对称性考虑, 这类植物是辐射对称型, 但其传粉昆虫类群的特化程度高, 与辐射对称植物泛化的传粉系统并不一致。从植物传粉类群的特化程度(Gong & Huang, 2009)与花对称性综合考虑, 可能会有更准确的结果和更合理的解释。
致谢
感谢兰州大学生命科学学院宋海坤、李婵鹤、马建超在野外植物特征测定过程中给予的帮助。
参考文献
Covariance and decoupling of floral and vegetative traits in nine neotropical plants: a re-evaluation of Berg’s correlation-pleiades concept
Nearly forty years ago R. L. Berg proposed that plants with specialized pollination ecology evolve genetic and developmental systems that decouple floral morphology from phenotypic variation in vegetative traits. These species evolve separate floral and vegetative trait clusters, or as she termed them,
The ecological significance of correlation pleiades
Evolutionary significance of phenotypic plasticity in plants
Components of phenotypic selection: pollen export and flower corrolla width in Ipomopsis aggregata
DOI:10.1111/j.1558-5646.1991.tb02648.x
URL
PMID:28563828
[本文引用: 2]

In the hummingbird-pollinated herb Ipomopsis aggregata, selection through male function during pollination favors wide corolla tubes. We explored the mechanisms behind this selection, using phenotypic selection analysis to compare effects of corolla width on two components of male pollination success, pollinator visit rate and pollen exported per visit. During single visits by captive hummingbirds, flowers with wider corollas exported more pollen, and more dye used as a pollen analogue, to stigmas of recipient flowers. Corolla width was less strongly related to visit rate in the field, and had no direct effect on visit rate after nectar production and corolla length were controlled for. Moreover, the phenotypic selection differential was 80% higher for the effect on pollen exported per visit, suggesting that this is the more important mechanism of selection.
Phylogenetics of seed plants: an analysis of nucleotide sequences from the plastid gene rbcL
Effects of flower size and number on pollinator visitation to wild radish, Raphanus raphanistrum
DOI:10.1007/BF00330014
URL
PMID:28307144
[本文引用: 1]
Plant traits that increase pollinator visitation should be under strong selection. However, few studies have demonstrated a causal link between natural variation in attractive traits and natural variation in visitation to whole plants. Here we examine the effects of flower number and size on visitation to wild radish by two taxa of pollinators over 3 years, using a combination of multiple regression and experimental reductions in both traits. We found strong, consistent evidence that increases in both flower number and size cause increased visitation by syrphid flies. The results for small bees were harder to interpret, because the multiple regression and experimental manipulation results did not agree. It is likely that increased flower size causes a weak increase in small-bee visitation, but strong relationships between flower number and small-bee visitation seen in 2 years of observational studies were not corroborated by experimental manipulation of this trait. Small bees may actually have responded to an unmeasured trait correlated with flower number, or lower small-bee abundances when the flower number manipulation was conducted may have reduced our ability to detect a causal relationship. We conclude that studies using only 1 year, one method, or measuring only one trait may not provide an adequate understanding of the effects of plant traits on pollinator attraction.
Genetics of floral symmetry revealed
DOI:10.1016/s0169-5347(97)01028-8 URL PMID:21237985 [本文引用: 1]
Selection on floral morphology by hummingbirds
DOI:10.2307/2388696 URL [本文引用: 2]
Floral symmetry: pollinator- mediated stabilizing selection on flower size in bilateral species
DOI:10.1098/rspb.2009.1254
URL
PMID:19710062
[本文引用: 5]
Pollinator-mediated stabilizing selection (PMSS) has been proposed as the driver of the evolutionary shift from radial to bilateral symmetry of flowers. Studies have shown that variation in flower size is lower in bilateral than in radial species, but whether bilateral flowers experience more stabilizing selection pressures by employing fewer, more specialized pollinators than radial flowers remains unclear. To test the PMSS hypothesis, we investigate plant-pollinator interactions from a whole community in an alpine meadow in Hengduan Mountains, China, to examine: (i) variance in flower size and level of ecological generalization (pollinator diversity calculated using functional groups) in 14 bilateral and 13 radial species and (ii) the role pollinator diversity played in explaining the difference of variance in flower size between bilateral and radial species. Our data showed that bilateral species had less variance in flower size and were visited by fewer pollinator groups. Pollinator diversity accounted for up to 40 per cent of the difference in variance in flower size between bilateral and radial species. The mediator effect of pollinator diversity on the relationship between floral symmetry and variance in flower size in the community is consistent with the PMSS hypothesis.
Pollen removal from tristylous Pontederia cordata: effects of anther position and pollinator specialization
The Comparative Method in Evolutionary Biology
Olfactory-bulb size and nocturnality in birds
Perianth organization and intra-specific floral variability
DOI:10.1111/j.1438-8677.2008.00091.x
URL
PMID:18950427
[本文引用: 1]
Floral symmetry and fusion of perianth parts are factors that contribute to fine-tune the match between flowers and their animal pollination vectors. In the present study, we investigated whether the possession of a sympetalous (fused) corolla and bilateral symmetry of flowers translate into decreased intra-specific variability as a result of natural stabilizing selection exerted by pollinators. Average size of the corolla and intra-specific variability were determined in two sets of southern Spanish entomophilous plant species. In the first set, taxa were paired by family to control for the effect of phylogeny (phylogenetically independent contrasts), whereas in the second set species were selected at random. Flower size data from a previous study (with different species) were also used to test the hypothesis that petal fusion contributes to decrease intra-specific variability. In the phylogenetically independent contrasts, floral symmetry was a significant correlate of intra-specific variation, with bilaterally symmetrical flowers showing more constancy than radially symmetrical flowers (i.e. unsophisticated from a functional perspective). As regards petal fusion, species with fused petals were on average more constant than choripetalous species, but the difference was not statistically significant. The reanalysis of data from a previous study yielded largely similar results, with a distinct effect of symmetry on variability, but no effect of petal fusion. The randomly-chosen species sample, on the other hand, failed to reveal any significant effect of either symmetry or petal fusion on intra-specific variation. The problem of low-statistical power in this kind of analysis, and the difficulty of testing an evolutionary hypothesis that involves phenotypic traits with a high degree of morphological correlation is discussed.
The measurement of selection on correlated characters
DOI:10.1111/j.1558-5646.1983.tb00236.x URL PMID:28556011 [本文引用: 1]
Progress in the study of floral symmetry
花对称性的研究进展
Origin of floral asymmetry in Antirrhinum
DOI:10.1038/383794a0
URL
PMID:8893002
[本文引用: 1]
Dorsoventral asymmetry in flowers is thought to have evolved many times from a radially symmetrical ancestral condition. The first gene controlling floral asymmetry, cycloidea in Antirrhinum, has been isolated. The cycloidea gene is expressed at a very early stage in dorsal regions of floral meristems, where it affects growth rate and primordium initiation. Expression continues through to later stages in dorsal primordia to affect the asymmetry, size and cell types of petals and stamens.
Covariance of floral and vegetative traits in four species of Ranunculaceae: a comparison between specialized and generalized pollination systems
DOI:10.1111/j.1744-7909.2008.00722.x
URL
PMID:18924281
[本文引用: 2]
Theory predicts that tighter correlation between floral traits and weaker relationship between floral and vegetative traits more likely occur in specialized flowers than generalized flowers, favoring by precise fit with pollinators. However, traits and trait correlations frequently vary under different environments. Through detecting spatiotemporal variation in phenotypic traits (floral organ size and vegetative size) and trait correlations in four Ranunculaceae species, we examined four predictions. Overall, our results supported these predictions to a certain degree. The mean coefficient of variation (CV) of floral traits in two specialized species (Delphinium kamaonense and Aconitum gymnandrum) was marginally significantly lower than that of another two generalized species (Trollius ranunculoides and Anemone obtusiloba). The two specialized species also showed marginally significantly smaller CV in floral traits than vegetative size across the two species. The absolute mean correlation between floral and vegetative traits, or that between floral traits in species with specialized flowers was not significantly lower, or higher than that in generalized plants, weakly supporting the predictions. Furthermore, we documented a large variation in trait correlations of four species among different seasons and populations. Study of covariance of floral and vegetative traits will benefit from the contrast of results obtained from generalized and specialized pollination systems.
A pairwise comparative method as illustrated by copulation frequency in birds
Patterns of fluctuating asymmetry in flowers: implications for sexual selection in plants
Floral symmetry and its role in plant-pollinator systems: terminology, distribution, and hypotheses
Decoupled phenotypic variation between floral and vegetative traits: distinguishing between developmental and environmental correlations
Differences in pollinator faunas may generate geographic differences in floral morphology and integration in Narcissus papyraceus(Amaryllidaceae)
Passerine polygyny: a role for parasites?
Temporal variation in selection on a floral character
DOI:10.1111/j.1558-5646.1989.tb04240.x URL PMID:28568553 [本文引用: 1]
An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III
Why do bilaterally symmetrical flowers orient vertically? Flower orientation influences pollinator landing behaviour
Support for the predictions of the pollinator-mediated stabilizing selection hypothesis
Why does the size of reproductive structures decline through time in Hydrophyllum appendiculatum(Hydrophyllaceae)? Developmental constraints vs. resource limitation
Patterns of pollen removal and deposition in tristylous Pontederia cordata L.(Pontederiaceae)

{kind=link}
{kind=link}
{kind=link}
{kind=link}