Responses of the functional traits in Cleistogenes squarrosa to nitrogen addition and drought
YANG Hao1, *, , LUO Ya-Chen1, 2
1Key Laboratory of Ecosystem Network Observation and Modeling, Institute of Geographical Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, Chinaand 2School of Geographical Sciences, Southwest University, Chongqing 400715, China
Plant functional traits have been widely used to study the responses of plant to environmental change. Cleistogenes squarrosa is an important C4 species in Nei Mongol grassland. How its functional traits would respond to varied nitrogen and water conditions have rarely been studied. Our subject was to examine the responses of the whole-plant traits and leaf morphological and physiological traits to nitrogen addition and drought in this species.
Methods
We conducted a pot experiment with a gradient of N addition (0, 10.5, 35.0, and 56.0 g·m-2·a-1) and water treatments (natural precipitation vs. 70% of mean monthly precipitation) in 2013. The whole-plant traits (e.g., root depth and stem-leaf biomass ratio), leaf morphological (e.g., leaf area and specific leaf area) and physiological traits (e.g., photosynthetic rate, water use efficiency, and leaf N content) were investigated.
Important findings
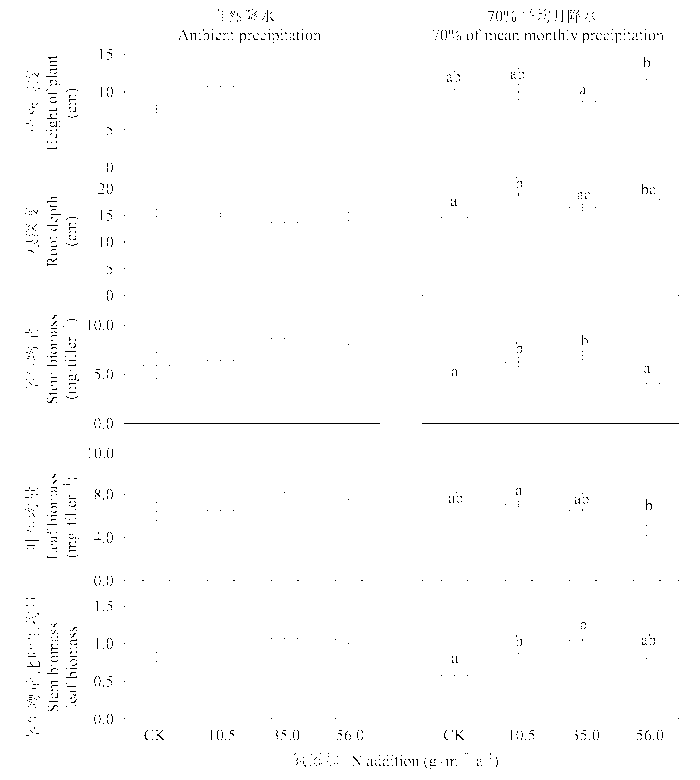
N addition had a significant effect on the whole-plant traits in C. squarrosa. The effects of N addition, water treatments, and an interaction between N addition and water treatments on leaf morphological and physiological traits were highly significant in most cases. The patterns of functional traits in response to N addition differed between plants under natural precipitation and with reduced mean monthly precipitation. Root depth, stem biomass, and stem-leaf biomass ratio were increased in treatments with low and intermediate N additions under reduced precipitation, but not changed under natural precipitation. Specific leaf area increased along the N addition gradient under drought, but did not change under natural precipitation. High N addition stimulated photosynthetic rate and transpiration rate and increased water use efficiency under natural precipitation, but had no effect on under reduced precipitation. Leaf N content on area basis increased slightly with the increases in N addition under natural precipitation, but decreased significantly under reduced precipitation. N addition influenced mainly the leaf morphological and physiological traits under natural precipitation and the whole-plant traits and leaf morphological traits under reduced precipitation. In conclusion, our results indicate that the functional traits in C. squarrosa respond to N addition and the patterns of responses differ under different water conditions, reflecting the adaptation to changes in N and water availability.
Keywords:C4 plant
;
leaf nitrogen content
;
morphological traits
;
Nei Mongol grassland
;
photosynthetic rate
;
physiological traits
YANGHao, LUOYa-Chen. Responses of the functional traits in Cleistogenes squarrosa to nitrogen addition and drought. Chinese Journal of Plant Ecology, 2015, 39(1): 32-42 https://doi.org/10.17521/cjpe.2015.0004
盆栽实验于2013年设置在中国科学院内蒙古草原生态系统定位研究站(116.68° E, 43.55° N, 海拔1250 m)。该区域气候属于温带半干旱大陆性气候, 年降水量为346 mm, 其中80%降水集中在生长季(5-8月); 年平均气温0.3 ℃。1982-2008年期间, 6月和7月平均月降水量分别为55.5 mm和92.9 mm, 两个月总降水量为148.4 mm。2013年6月和8月降水量分别为87.4 mm和74.4 mm, 总降水量为161.8 mm, 接近于多年平均值。土壤为暗栗钙土。典型群落类型为羊草草原群落, 以羊草、大针茅和糙隐子草等为优势种。
1.2 盆栽实验设计
盆栽实验地处于定位站外围, 包括4个氮添加梯度和2个降水处理, 每个处理4个重复。氮添加梯度为对照、低氮添加(10.5 g·m-2·a-1)、中氮添加(35.0 g·m-2·a-1)和高氮添加(56.0 g·m-2·a-1), 氮肥采用分析纯尿素。内蒙古典型草原区的氮沉降量为1.0-1.4 g·m-2·a-1 (Lü & Tian, 2007), 因此, 低氮、中氮和高氮处理的氮添加量相当于内蒙古典型草原区氮沉降量的9倍、29倍和47倍。降水处理为当年自然降水和干旱处理, 干旱处理中浇水量为70%平均月降水量(1982-2008年)。减少降水的方法是搭建遮雨棚。遮雨棚南面高50 cm, 北面高90 cm。晴天时收起遮雨棚, 下雨时进行遮挡以防止盆栽接收自然降水。5月6-8日, 在遮雨棚及旁边放置盆栽用的盆。每个盆直径30 cm, 装实验地附近的0-20 cm过筛土至盆边缘5 cm, 后将盆埋入土中, 盆边露出地面约5 cm。盆离遮雨棚的边在1 m左右。盆放置好后, 进行播种。定期浇水, 剔除杂草, 保证种子萌发及植株生长。6月初进行剔苗, 保证每盆在20丛左右。6月4日, 进行施肥。6月10日至8月3日, 每5天浇1次地下水(氮、磷、钾的浓度都在检测线以下)。总计6月(10-30号)浇水量为26 mm (每次460 mL, 约为6.5 mm降水), 7月为65 mm (每次760 mL, 约为10.8 mm降水)。8月3-6日进行指标测定, 并收获各器官。
Table 1 Results of two-way ANOVAs for the effects of N addition, water treatments and their interaction on whole-plant traits and leaf morphological traits in Cleistogenes squarrosa
图2 施肥和干旱对糙隐子草整株性状的影响(平均值±标准误差, n = 4)。不同的字母表示处理间有显著的差异(LSD检验, p < 0.05)。
Fig. 2 Effects of N addition and drought on whole-plant traits in Cleistogenes squarrosa (mean ± SE, n = 4). Difference letters indicate significant differences among treatments (LSD test, p < 0.05).
Table 2
表2
表2 氮添加、水处理和氮水交互作用对糙隐子草叶生理性状和叶氮含量的影响
Table 2 Results of two-way ANOVAs for the effects of N addition, water treatments and their interaction on leaf photosynthetic traits and leaf nitrogen content in Cleistogenes squarrosa
叶生理形状 Leaf photosynthetic traits
叶片氮含量 Leaf nitrogen content
光合速率 Pn (μmol CO2·m-2·s-1)
蒸腾速率 Tr (mmol H2O·m-2·s-1)
气孔导度 Gs (mol H2O·m-2·s-1)
胞间CO2浓度 Ci (μmol CO2·mol-1)
水分利用效率 WUE (μmol CO2·mol-1 H2O)
气孔限制 Ls
单位质量叶片 氮含量 Nmass (%)
单位面积叶片 氮含量 Narea (g·m-2)
F
p
F
p
F
p
F
p
F
p
F
p
F
p
F
p
氮添加 N addition
3.91
0.024*
1.86
0.169
1.39
0.274
11.28
0.000**
2.44
0.095
11.49
0.000**
6.71
0.003**
1.34
0.290
水处理 Water treatment
9.19
0.007**
12.89
0.002**
10.85
0.004**
6.29
0.021*
0.44
0.513
6.50
0.019*
7.55
0.012*
15.05
0.001**
氮添加×水处理 N addition × Water treatment
7.82
0.001**
2.70
0.073
4.14
0.020*
10.03
0.000**
3.71
0.029*
9.91
0.000**
0.45
0.720
5.16
0.008**
Ci, intercellular CO2 concentration; Gs, stomatal conductance; Ls, stomatal limitation; Narea, leaf nitrogen content based on leaf area; Nmass, leaf nitrogen content based on leaf mass; Pn, net photosynthetic rate; Tr, Transpiration rate; WUE, water use efficiency. *, p < 0.05; **, p < 0.01.
图3 施肥和干旱对糙隐子草叶片形态性状的影响(平均值±标准误差, n = 4)。不同的字母表示处理间有显著的差异(LSD检验, p < 0.05)。
Fig. 3 Effects of N addition and drought on leaf morphological traits in Cleistogenes squarrosa (mean ± SE, n = 4). Difference letters indicate significant differences among treatments (LSD test, p < 0.05).
图4 施肥和干旱对糙隐子草叶片光合性状的影响(平均值±标准误差, n = 4)。Ci, 胞间CO2浓度; Gs, 气孔导度; Ls, 气孔限制值; Pn, 光合速率; Tr, 蒸腾速率; WUE, 水分利用效率。不同的字母表示处理间有显著的差异(LSD test, p < 0.05)。
Fig. 4 Effects of N addition and drought on leaf photosynthetic traits in Cleistogenes squarrosa (mean ± SE, n = 4). Ci, intercellular CO2 concentration; Gs, stomatal conductance; Ls, stomatal limitation; Pn, net photosynthetic rate; Tr, transpiration rate; WUE, water use efficiency. Difference letters indicate significant differences among treatments (LSD test, p < 0.05).
图5 施肥和干旱对糙隐子草叶片单位质量氮含量和单位面积氮含量的影响(平均值±标准误差, n = 4)。不同的字母表示处理间有显著的差异(LSD test, p < 0.05)。
Fig. 5 Effects of N addition and drought on leaf nitrogen content based on leaf mass (Nmass) and leaf nitrogen content based on leaf area (Narea) in Cleistogenes squarrosa (mean ± SE, n = 4). Difference letters indicate significant differences among treatments (LSD test, p < 0.05).
Fig. 6 The cluster dendrogram of plant traits. Ci, intercellular CO2 concentration; Gs, stomatal conductance; Ls, stomatal limitation; Narea, leaf nitrogen content based on leaf area; Nmass, leaf nitrogen content based on leaf mass; Pn, net photosynthetic rate; Tr, transpiration rate; WUE, water use efficiency.
3 讨论
氮素和水分是干旱和半干旱草地生态系统植物生长和生物量产生的主要限制因子(Bai et al., 2004, 2008)。我们的研究表明, 不同的氮添加量显著影响了糙隐子草的功能性状, 并且在自然降水和干旱处理下功能性状的响应格局是不同的。在自然降水下, 氮添加主要影响糙隐子草的叶形态和光合生理性状, 而在干旱条件下, 氮添加主要影响糙隐子草的整株性状和形态性状。
Differential responses of plant functional trait to grazing between two contrasting dominant C3 and C4 species in a typical steppe of Inner Mongolia, China
Plant trait responses to grazing — A global synthesis
1
2007
... 植物功能性状是指能够通过影响植物个体的生长、繁殖和生存而间接影响植物适应度的植物性状, 包括形态、生理和物候等性状(Violle et al., 2007).植物功能性状能够反映植物个体对环境的响应和适应, 将环境变化同植物群落的结构和功能联系起来(Díaz et al., 2007; Violle et al., 2007; Lavorel et al., 2011).关于全球变化和植物功能性状之间关系的研究是当前植物生态学研究的热点问题之一. ...
Changes in nitrogen resorption traits of six temperate grassland species along a multi-level N addition gradient
2
2008
... 糙隐子草(Cleistogenes squarrosa)是一种多年生丛生小禾草, 属C4植物, 广泛分布于内蒙古高原的草甸草原、典型草原和荒漠草原, 是针茅草原和羊草草原的下层优势种(中国科学院内蒙古宁夏综合考察队, 1985), 也是C4植物中生物量比例最高的物种.相比于共存的C3植物, 糙隐子草具有高光合能力、高水分利用效率、抗干旱和耐牧等特点(汪诗平和王艳芬, 2001; Chen et al., 2002; 汪诗平等, 2003).在中度放牧条件下或退化草场恢复一段时间后, 其在群落中可处于优势地位(王鑫厅等, 2014).全球变化和人为干扰下, 糙隐子草功能性状的改变可能会影响草原生态系统的结构和功能.近年来, 糙隐子草功能性状的相关研究, 主要集中于氮添加、放牧等野外控制实验以及不同生境下的形态和生理功能性状(崔骁勇等, 2001; Chen et al., 2002, 2005; 万宏伟等, 2008; Zheng et al., 2011; 秦洁等, 2014).羊草草原长期氮添加实验结果表明, 糙隐子草叶片氮含量、叶绿素含量随施氮量的增加而增加, 叶绿素a和b的比值随施氮量的增加而降低(Huang et al., 2008; 万宏伟等, 2008), 说明氮添加不仅能够提高糙隐子草的光合能力, 还可能通过改变群落结构而改变糙隐子草生长的光环境.不同的放牧强度对糙隐子草生理和形态功能特征也有显著的影响(Zheng et al., 2011).放牧降低了糙隐子草叶和茎干质量、叶面积, 增加了叶片氮含量和胞间CO2浓度; 光合速率、蒸腾速率和气孔导度等光合生理性状在湿润年份和干旱年份对放牧的响应有所不同.原因可能是放牧改变了土壤养分和水分条件(Milchunas & Lauenroth, 1993), 进而导致了这些功能性状的变化(Adler et al., 2004, 2005).在不同的植物群落中, 糙隐子草叶片含水量在芨芨草(Achnatherum splendens)和冷蒿(Artemisia frigida)群落中高于线叶菊(Filifolium sibiricum)、羊草(Leymus chinensis)、大针茅(Stipa grandis)和小叶锦鸡儿(Caragana microphylla)群落(Chen et al., 2002).光合利用率在半干旱生境中较高, 水分利用率在干旱生境中较高(崔骁勇等, 2001). ...
... 糙隐子草(Cleistogenes squarrosa)是一种多年生丛生小禾草, 属C4植物, 广泛分布于内蒙古高原的草甸草原、典型草原和荒漠草原, 是针茅草原和羊草草原的下层优势种(中国科学院内蒙古宁夏综合考察队, 1985), 也是C4植物中生物量比例最高的物种.相比于共存的C3植物, 糙隐子草具有高光合能力、高水分利用效率、抗干旱和耐牧等特点(汪诗平和王艳芬, 2001; Chen et al., 2002; 汪诗平等, 2003).在中度放牧条件下或退化草场恢复一段时间后, 其在群落中可处于优势地位(王鑫厅等, 2014).全球变化和人为干扰下, 糙隐子草功能性状的改变可能会影响草原生态系统的结构和功能.近年来, 糙隐子草功能性状的相关研究, 主要集中于氮添加、放牧等野外控制实验以及不同生境下的形态和生理功能性状(崔骁勇等, 2001; Chen et al., 2002, 2005; 万宏伟等, 2008; Zheng et al., 2011; 秦洁等, 2014).羊草草原长期氮添加实验结果表明, 糙隐子草叶片氮含量、叶绿素含量随施氮量的增加而增加, 叶绿素a和b的比值随施氮量的增加而降低(Huang et al., 2008; 万宏伟等, 2008), 说明氮添加不仅能够提高糙隐子草的光合能力, 还可能通过改变群落结构而改变糙隐子草生长的光环境.不同的放牧强度对糙隐子草生理和形态功能特征也有显著的影响(Zheng et al., 2011).放牧降低了糙隐子草叶和茎干质量、叶面积, 增加了叶片氮含量和胞间CO2浓度; 光合速率、蒸腾速率和气孔导度等光合生理性状在湿润年份和干旱年份对放牧的响应有所不同.原因可能是放牧改变了土壤养分和水分条件(Milchunas & Lauenroth, 1993), 进而导致了这些功能性状的变化(Adler et al., 2004, 2005).在不同的植物群落中, 糙隐子草叶片含水量在芨芨草(Achnatherum splendens)和冷蒿(Artemisia frigida)群落中高于线叶菊(Filifolium sibiricum)、羊草(Leymus chinensis)、大针茅(Stipa grandis)和小叶锦鸡儿(Caragana microphylla)群落(Chen et al., 2002).光合利用率在半干旱生境中较高, 水分利用率在干旱生境中较高(崔骁勇等, 2001). ...
1
... 糙隐子草(Cleistogenes squarrosa)是一种多年生丛生小禾草, 属C4植物, 广泛分布于内蒙古高原的草甸草原、典型草原和荒漠草原, 是针茅草原和羊草草原的下层优势种(中国科学院内蒙古宁夏综合考察队, 1985), 也是C4植物中生物量比例最高的物种.相比于共存的C3植物, 糙隐子草具有高光合能力、高水分利用效率、抗干旱和耐牧等特点(汪诗平和王艳芬, 2001; Chen et al., 2002; 汪诗平等, 2003).在中度放牧条件下或退化草场恢复一段时间后, 其在群落中可处于优势地位(王鑫厅等, 2014).全球变化和人为干扰下, 糙隐子草功能性状的改变可能会影响草原生态系统的结构和功能.近年来, 糙隐子草功能性状的相关研究, 主要集中于氮添加、放牧等野外控制实验以及不同生境下的形态和生理功能性状(崔骁勇等, 2001; Chen et al., 2002, 2005; 万宏伟等, 2008; Zheng et al., 2011; 秦洁等, 2014).羊草草原长期氮添加实验结果表明, 糙隐子草叶片氮含量、叶绿素含量随施氮量的增加而增加, 叶绿素a和b的比值随施氮量的增加而降低(Huang et al., 2008; 万宏伟等, 2008), 说明氮添加不仅能够提高糙隐子草的光合能力, 还可能通过改变群落结构而改变糙隐子草生长的光环境.不同的放牧强度对糙隐子草生理和形态功能特征也有显著的影响(Zheng et al., 2011).放牧降低了糙隐子草叶和茎干质量、叶面积, 增加了叶片氮含量和胞间CO2浓度; 光合速率、蒸腾速率和气孔导度等光合生理性状在湿润年份和干旱年份对放牧的响应有所不同.原因可能是放牧改变了土壤养分和水分条件(Milchunas & Lauenroth, 1993), 进而导致了这些功能性状的变化(Adler et al., 2004, 2005).在不同的植物群落中, 糙隐子草叶片含水量在芨芨草(Achnatherum splendens)和冷蒿(Artemisia frigida)群落中高于线叶菊(Filifolium sibiricum)、羊草(Leymus chinensis)、大针茅(Stipa grandis)和小叶锦鸡儿(Caragana microphylla)群落(Chen et al., 2002).光合利用率在半干旱生境中较高, 水分利用率在干旱生境中较高(崔骁勇等, 2001). ...
Using plant functional traits to understand the landscape distribution of multiple ecosystem services
1
2011
... 植物功能性状是指能够通过影响植物个体的生长、繁殖和生存而间接影响植物适应度的植物性状, 包括形态、生理和物候等性状(Violle et al., 2007).植物功能性状能够反映植物个体对环境的响应和适应, 将环境变化同植物群落的结构和功能联系起来(Díaz et al., 2007; Violle et al., 2007; Lavorel et al., 2011).关于全球变化和植物功能性状之间关系的研究是当前植物生态学研究的热点问题之一. ...
Spatial and temporal patterns of nitrogen deposition in China: Synthesis of observational data. Journal of Geophysical Research: Atmospheres, 112,
1
2007
... 盆栽实验地处于定位站外围, 包括4个氮添加梯度和2个降水处理, 每个处理4个重复.氮添加梯度为对照、低氮添加(10.5 g·m-2·a-1)、中氮添加(35.0 g·m-2·a-1)和高氮添加(56.0 g·m-2·a-1), 氮肥采用分析纯尿素.内蒙古典型草原区的氮沉降量为1.0-1.4 g·m-2·a-1 (Lü & Tian, 2007), 因此, 低氮、中氮和高氮处理的氮添加量相当于内蒙古典型草原区氮沉降量的9倍、29倍和47倍.降水处理为当年自然降水和干旱处理, 干旱处理中浇水量为70%平均月降水量(1982-2008年).减少降水的方法是搭建遮雨棚.遮雨棚南面高50 cm, 北面高90 cm.晴天时收起遮雨棚, 下雨时进行遮挡以防止盆栽接收自然降水.5月6-8日, 在遮雨棚及旁边放置盆栽用的盆.每个盆直径30 cm, 装实验地附近的0-20 cm过筛土至盆边缘5 cm, 后将盆埋入土中, 盆边露出地面约5 cm.盆离遮雨棚的边在1 m左右.盆放置好后, 进行播种.定期浇水, 剔除杂草, 保证种子萌发及植株生长.6月初进行剔苗, 保证每盆在20丛左右.6月4日, 进行施肥.6月10日至8月3日, 每5天浇1次地下水(氮、磷、钾的浓度都在检测线以下).总计6月(10-30号)浇水量为26 mm (每次460 mL, 约为6.5 mm降水), 7月为65 mm (每次760 mL, 约为10.8 mm降水).8月3-6日进行指标测定, 并收获各器官. ...
Quantitative effects of grazing on vegetation and soils over a global range of environments
1
1993
... 糙隐子草(Cleistogenes squarrosa)是一种多年生丛生小禾草, 属C4植物, 广泛分布于内蒙古高原的草甸草原、典型草原和荒漠草原, 是针茅草原和羊草草原的下层优势种(中国科学院内蒙古宁夏综合考察队, 1985), 也是C4植物中生物量比例最高的物种.相比于共存的C3植物, 糙隐子草具有高光合能力、高水分利用效率、抗干旱和耐牧等特点(汪诗平和王艳芬, 2001; Chen et al., 2002; 汪诗平等, 2003).在中度放牧条件下或退化草场恢复一段时间后, 其在群落中可处于优势地位(王鑫厅等, 2014).全球变化和人为干扰下, 糙隐子草功能性状的改变可能会影响草原生态系统的结构和功能.近年来, 糙隐子草功能性状的相关研究, 主要集中于氮添加、放牧等野外控制实验以及不同生境下的形态和生理功能性状(崔骁勇等, 2001; Chen et al., 2002, 2005; 万宏伟等, 2008; Zheng et al., 2011; 秦洁等, 2014).羊草草原长期氮添加实验结果表明, 糙隐子草叶片氮含量、叶绿素含量随施氮量的增加而增加, 叶绿素a和b的比值随施氮量的增加而降低(Huang et al., 2008; 万宏伟等, 2008), 说明氮添加不仅能够提高糙隐子草的光合能力, 还可能通过改变群落结构而改变糙隐子草生长的光环境.不同的放牧强度对糙隐子草生理和形态功能特征也有显著的影响(Zheng et al., 2011).放牧降低了糙隐子草叶和茎干质量、叶面积, 增加了叶片氮含量和胞间CO2浓度; 光合速率、蒸腾速率和气孔导度等光合生理性状在湿润年份和干旱年份对放牧的响应有所不同.原因可能是放牧改变了土壤养分和水分条件(Milchunas & Lauenroth, 1993), 进而导致了这些功能性状的变化(Adler et al., 2004, 2005).在不同的植物群落中, 糙隐子草叶片含水量在芨芨草(Achnatherum splendens)和冷蒿(Artemisia frigida)群落中高于线叶菊(Filifolium sibiricum)、羊草(Leymus chinensis)、大针茅(Stipa grandis)和小叶锦鸡儿(Caragana microphylla)群落(Chen et al., 2002).光合利用率在半干旱生境中较高, 水分利用率在干旱生境中较高(崔骁勇等, 2001). ...
The scaling of leaf area and mass: the cost of light interception increases with leaf size
... 植物功能性状是指能够通过影响植物个体的生长、繁殖和生存而间接影响植物适应度的植物性状, 包括形态、生理和物候等性状(Violle et al., 2007).植物功能性状能够反映植物个体对环境的响应和适应, 将环境变化同植物群落的结构和功能联系起来(Díaz et al., 2007; Violle et al., 2007; Lavorel et al., 2011).关于全球变化和植物功能性状之间关系的研究是当前植物生态学研究的热点问题之一. ...
... ; Violle et al., 2007; Lavorel et al., 2011).关于全球变化和植物功能性状之间关系的研究是当前植物生态学研究的热点问题之一. ...
羊草草原群落6种植物叶片功能特性对氮素添加的响应
4
2008
... 糙隐子草(Cleistogenes squarrosa)是一种多年生丛生小禾草, 属C4植物, 广泛分布于内蒙古高原的草甸草原、典型草原和荒漠草原, 是针茅草原和羊草草原的下层优势种(中国科学院内蒙古宁夏综合考察队, 1985), 也是C4植物中生物量比例最高的物种.相比于共存的C3植物, 糙隐子草具有高光合能力、高水分利用效率、抗干旱和耐牧等特点(汪诗平和王艳芬, 2001; Chen et al., 2002; 汪诗平等, 2003).在中度放牧条件下或退化草场恢复一段时间后, 其在群落中可处于优势地位(王鑫厅等, 2014).全球变化和人为干扰下, 糙隐子草功能性状的改变可能会影响草原生态系统的结构和功能.近年来, 糙隐子草功能性状的相关研究, 主要集中于氮添加、放牧等野外控制实验以及不同生境下的形态和生理功能性状(崔骁勇等, 2001; Chen et al., 2002, 2005; 万宏伟等, 2008; Zheng et al., 2011; 秦洁等, 2014).羊草草原长期氮添加实验结果表明, 糙隐子草叶片氮含量、叶绿素含量随施氮量的增加而增加, 叶绿素a和b的比值随施氮量的增加而降低(Huang et al., 2008; 万宏伟等, 2008), 说明氮添加不仅能够提高糙隐子草的光合能力, 还可能通过改变群落结构而改变糙隐子草生长的光环境.不同的放牧强度对糙隐子草生理和形态功能特征也有显著的影响(Zheng et al., 2011).放牧降低了糙隐子草叶和茎干质量、叶面积, 增加了叶片氮含量和胞间CO2浓度; 光合速率、蒸腾速率和气孔导度等光合生理性状在湿润年份和干旱年份对放牧的响应有所不同.原因可能是放牧改变了土壤养分和水分条件(Milchunas & Lauenroth, 1993), 进而导致了这些功能性状的变化(Adler et al., 2004, 2005).在不同的植物群落中, 糙隐子草叶片含水量在芨芨草(Achnatherum splendens)和冷蒿(Artemisia frigida)群落中高于线叶菊(Filifolium sibiricum)、羊草(Leymus chinensis)、大针茅(Stipa grandis)和小叶锦鸡儿(Caragana microphylla)群落(Chen et al., 2002).光合利用率在半干旱生境中较高, 水分利用率在干旱生境中较高(崔骁勇等, 2001). ...
... 糙隐子草(Cleistogenes squarrosa)是一种多年生丛生小禾草, 属C4植物, 广泛分布于内蒙古高原的草甸草原、典型草原和荒漠草原, 是针茅草原和羊草草原的下层优势种(中国科学院内蒙古宁夏综合考察队, 1985), 也是C4植物中生物量比例最高的物种.相比于共存的C3植物, 糙隐子草具有高光合能力、高水分利用效率、抗干旱和耐牧等特点(汪诗平和王艳芬, 2001; Chen et al., 2002; 汪诗平等, 2003).在中度放牧条件下或退化草场恢复一段时间后, 其在群落中可处于优势地位(王鑫厅等, 2014).全球变化和人为干扰下, 糙隐子草功能性状的改变可能会影响草原生态系统的结构和功能.近年来, 糙隐子草功能性状的相关研究, 主要集中于氮添加、放牧等野外控制实验以及不同生境下的形态和生理功能性状(崔骁勇等, 2001; Chen et al., 2002, 2005; 万宏伟等, 2008; Zheng et al., 2011; 秦洁等, 2014).羊草草原长期氮添加实验结果表明, 糙隐子草叶片氮含量、叶绿素含量随施氮量的增加而增加, 叶绿素a和b的比值随施氮量的增加而降低(Huang et al., 2008; 万宏伟等, 2008), 说明氮添加不仅能够提高糙隐子草的光合能力, 还可能通过改变群落结构而改变糙隐子草生长的光环境.不同的放牧强度对糙隐子草生理和形态功能特征也有显著的影响(Zheng et al., 2011).放牧降低了糙隐子草叶和茎干质量、叶面积, 增加了叶片氮含量和胞间CO2浓度; 光合速率、蒸腾速率和气孔导度等光合生理性状在湿润年份和干旱年份对放牧的响应有所不同.原因可能是放牧改变了土壤养分和水分条件(Milchunas & Lauenroth, 1993), 进而导致了这些功能性状的变化(Adler et al., 2004, 2005).在不同的植物群落中, 糙隐子草叶片含水量在芨芨草(Achnatherum splendens)和冷蒿(Artemisia frigida)群落中高于线叶菊(Filifolium sibiricum)、羊草(Leymus chinensis)、大针茅(Stipa grandis)和小叶锦鸡儿(Caragana microphylla)群落(Chen et al., 2002).光合利用率在半干旱生境中较高, 水分利用率在干旱生境中较高(崔骁勇等, 2001). ...
不同放牧率下糙隐子草种群补偿性生长的研究
1
2001
... 糙隐子草(Cleistogenes squarrosa)是一种多年生丛生小禾草, 属C4植物, 广泛分布于内蒙古高原的草甸草原、典型草原和荒漠草原, 是针茅草原和羊草草原的下层优势种(中国科学院内蒙古宁夏综合考察队, 1985), 也是C4植物中生物量比例最高的物种.相比于共存的C3植物, 糙隐子草具有高光合能力、高水分利用效率、抗干旱和耐牧等特点(汪诗平和王艳芬, 2001; Chen et al., 2002; 汪诗平等, 2003).在中度放牧条件下或退化草场恢复一段时间后, 其在群落中可处于优势地位(王鑫厅等, 2014).全球变化和人为干扰下, 糙隐子草功能性状的改变可能会影响草原生态系统的结构和功能.近年来, 糙隐子草功能性状的相关研究, 主要集中于氮添加、放牧等野外控制实验以及不同生境下的形态和生理功能性状(崔骁勇等, 2001; Chen et al., 2002, 2005; 万宏伟等, 2008; Zheng et al., 2011; 秦洁等, 2014).羊草草原长期氮添加实验结果表明, 糙隐子草叶片氮含量、叶绿素含量随施氮量的增加而增加, 叶绿素a和b的比值随施氮量的增加而降低(Huang et al., 2008; 万宏伟等, 2008), 说明氮添加不仅能够提高糙隐子草的光合能力, 还可能通过改变群落结构而改变糙隐子草生长的光环境.不同的放牧强度对糙隐子草生理和形态功能特征也有显著的影响(Zheng et al., 2011).放牧降低了糙隐子草叶和茎干质量、叶面积, 增加了叶片氮含量和胞间CO2浓度; 光合速率、蒸腾速率和气孔导度等光合生理性状在湿润年份和干旱年份对放牧的响应有所不同.原因可能是放牧改变了土壤养分和水分条件(Milchunas & Lauenroth, 1993), 进而导致了这些功能性状的变化(Adler et al., 2004, 2005).在不同的植物群落中, 糙隐子草叶片含水量在芨芨草(Achnatherum splendens)和冷蒿(Artemisia frigida)群落中高于线叶菊(Filifolium sibiricum)、羊草(Leymus chinensis)、大针茅(Stipa grandis)和小叶锦鸡儿(Caragana microphylla)群落(Chen et al., 2002).光合利用率在半干旱生境中较高, 水分利用率在干旱生境中较高(崔骁勇等, 2001). ...
气候变化和放牧活动对糙隐子草种群的影响
1
2003
... 糙隐子草(Cleistogenes squarrosa)是一种多年生丛生小禾草, 属C4植物, 广泛分布于内蒙古高原的草甸草原、典型草原和荒漠草原, 是针茅草原和羊草草原的下层优势种(中国科学院内蒙古宁夏综合考察队, 1985), 也是C4植物中生物量比例最高的物种.相比于共存的C3植物, 糙隐子草具有高光合能力、高水分利用效率、抗干旱和耐牧等特点(汪诗平和王艳芬, 2001; Chen et al., 2002; 汪诗平等, 2003).在中度放牧条件下或退化草场恢复一段时间后, 其在群落中可处于优势地位(王鑫厅等, 2014).全球变化和人为干扰下, 糙隐子草功能性状的改变可能会影响草原生态系统的结构和功能.近年来, 糙隐子草功能性状的相关研究, 主要集中于氮添加、放牧等野外控制实验以及不同生境下的形态和生理功能性状(崔骁勇等, 2001; Chen et al., 2002, 2005; 万宏伟等, 2008; Zheng et al., 2011; 秦洁等, 2014).羊草草原长期氮添加实验结果表明, 糙隐子草叶片氮含量、叶绿素含量随施氮量的增加而增加, 叶绿素a和b的比值随施氮量的增加而降低(Huang et al., 2008; 万宏伟等, 2008), 说明氮添加不仅能够提高糙隐子草的光合能力, 还可能通过改变群落结构而改变糙隐子草生长的光环境.不同的放牧强度对糙隐子草生理和形态功能特征也有显著的影响(Zheng et al., 2011).放牧降低了糙隐子草叶和茎干质量、叶面积, 增加了叶片氮含量和胞间CO2浓度; 光合速率、蒸腾速率和气孔导度等光合生理性状在湿润年份和干旱年份对放牧的响应有所不同.原因可能是放牧改变了土壤养分和水分条件(Milchunas & Lauenroth, 1993), 进而导致了这些功能性状的变化(Adler et al., 2004, 2005).在不同的植物群落中, 糙隐子草叶片含水量在芨芨草(Achnatherum splendens)和冷蒿(Artemisia frigida)群落中高于线叶菊(Filifolium sibiricum)、羊草(Leymus chinensis)、大针茅(Stipa grandis)和小叶锦鸡儿(Caragana microphylla)群落(Chen et al., 2002).光合利用率在半干旱生境中较高, 水分利用率在干旱生境中较高(崔骁勇等, 2001). ...
气候变化和放牧活动对糙隐子草种群的影响
1
2003
... 糙隐子草(Cleistogenes squarrosa)是一种多年生丛生小禾草, 属C4植物, 广泛分布于内蒙古高原的草甸草原、典型草原和荒漠草原, 是针茅草原和羊草草原的下层优势种(中国科学院内蒙古宁夏综合考察队, 1985), 也是C4植物中生物量比例最高的物种.相比于共存的C3植物, 糙隐子草具有高光合能力、高水分利用效率、抗干旱和耐牧等特点(汪诗平和王艳芬, 2001; Chen et al., 2002; 汪诗平等, 2003).在中度放牧条件下或退化草场恢复一段时间后, 其在群落中可处于优势地位(王鑫厅等, 2014).全球变化和人为干扰下, 糙隐子草功能性状的改变可能会影响草原生态系统的结构和功能.近年来, 糙隐子草功能性状的相关研究, 主要集中于氮添加、放牧等野外控制实验以及不同生境下的形态和生理功能性状(崔骁勇等, 2001; Chen et al., 2002, 2005; 万宏伟等, 2008; Zheng et al., 2011; 秦洁等, 2014).羊草草原长期氮添加实验结果表明, 糙隐子草叶片氮含量、叶绿素含量随施氮量的增加而增加, 叶绿素a和b的比值随施氮量的增加而降低(Huang et al., 2008; 万宏伟等, 2008), 说明氮添加不仅能够提高糙隐子草的光合能力, 还可能通过改变群落结构而改变糙隐子草生长的光环境.不同的放牧强度对糙隐子草生理和形态功能特征也有显著的影响(Zheng et al., 2011).放牧降低了糙隐子草叶和茎干质量、叶面积, 增加了叶片氮含量和胞间CO2浓度; 光合速率、蒸腾速率和气孔导度等光合生理性状在湿润年份和干旱年份对放牧的响应有所不同.原因可能是放牧改变了土壤养分和水分条件(Milchunas & Lauenroth, 1993), 进而导致了这些功能性状的变化(Adler et al., 2004, 2005).在不同的植物群落中, 糙隐子草叶片含水量在芨芨草(Achnatherum splendens)和冷蒿(Artemisia frigida)群落中高于线叶菊(Filifolium sibiricum)、羊草(Leymus chinensis)、大针茅(Stipa grandis)和小叶锦鸡儿(Caragana microphylla)群落(Chen et al., 2002).光合利用率在半干旱生境中较高, 水分利用率在干旱生境中较高(崔骁勇等, 2001). ...
不同恢复演替阶段糙隐子草种群的点格局分析
1
2014
... 糙隐子草(Cleistogenes squarrosa)是一种多年生丛生小禾草, 属C4植物, 广泛分布于内蒙古高原的草甸草原、典型草原和荒漠草原, 是针茅草原和羊草草原的下层优势种(中国科学院内蒙古宁夏综合考察队, 1985), 也是C4植物中生物量比例最高的物种.相比于共存的C3植物, 糙隐子草具有高光合能力、高水分利用效率、抗干旱和耐牧等特点(汪诗平和王艳芬, 2001; Chen et al., 2002; 汪诗平等, 2003).在中度放牧条件下或退化草场恢复一段时间后, 其在群落中可处于优势地位(王鑫厅等, 2014).全球变化和人为干扰下, 糙隐子草功能性状的改变可能会影响草原生态系统的结构和功能.近年来, 糙隐子草功能性状的相关研究, 主要集中于氮添加、放牧等野外控制实验以及不同生境下的形态和生理功能性状(崔骁勇等, 2001; Chen et al., 2002, 2005; 万宏伟等, 2008; Zheng et al., 2011; 秦洁等, 2014).羊草草原长期氮添加实验结果表明, 糙隐子草叶片氮含量、叶绿素含量随施氮量的增加而增加, 叶绿素a和b的比值随施氮量的增加而降低(Huang et al., 2008; 万宏伟等, 2008), 说明氮添加不仅能够提高糙隐子草的光合能力, 还可能通过改变群落结构而改变糙隐子草生长的光环境.不同的放牧强度对糙隐子草生理和形态功能特征也有显著的影响(Zheng et al., 2011).放牧降低了糙隐子草叶和茎干质量、叶面积, 增加了叶片氮含量和胞间CO2浓度; 光合速率、蒸腾速率和气孔导度等光合生理性状在湿润年份和干旱年份对放牧的响应有所不同.原因可能是放牧改变了土壤养分和水分条件(Milchunas & Lauenroth, 1993), 进而导致了这些功能性状的变化(Adler et al., 2004, 2005).在不同的植物群落中, 糙隐子草叶片含水量在芨芨草(Achnatherum splendens)和冷蒿(Artemisia frigida)群落中高于线叶菊(Filifolium sibiricum)、羊草(Leymus chinensis)、大针茅(Stipa grandis)和小叶锦鸡儿(Caragana microphylla)群落(Chen et al., 2002).光合利用率在半干旱生境中较高, 水分利用率在干旱生境中较高(崔骁勇等, 2001). ...
不同恢复演替阶段糙隐子草种群的点格局分析
1
2014
... 糙隐子草(Cleistogenes squarrosa)是一种多年生丛生小禾草, 属C4植物, 广泛分布于内蒙古高原的草甸草原、典型草原和荒漠草原, 是针茅草原和羊草草原的下层优势种(中国科学院内蒙古宁夏综合考察队, 1985), 也是C4植物中生物量比例最高的物种.相比于共存的C3植物, 糙隐子草具有高光合能力、高水分利用效率、抗干旱和耐牧等特点(汪诗平和王艳芬, 2001; Chen et al., 2002; 汪诗平等, 2003).在中度放牧条件下或退化草场恢复一段时间后, 其在群落中可处于优势地位(王鑫厅等, 2014).全球变化和人为干扰下, 糙隐子草功能性状的改变可能会影响草原生态系统的结构和功能.近年来, 糙隐子草功能性状的相关研究, 主要集中于氮添加、放牧等野外控制实验以及不同生境下的形态和生理功能性状(崔骁勇等, 2001; Chen et al., 2002, 2005; 万宏伟等, 2008; Zheng et al., 2011; 秦洁等, 2014).羊草草原长期氮添加实验结果表明, 糙隐子草叶片氮含量、叶绿素含量随施氮量的增加而增加, 叶绿素a和b的比值随施氮量的增加而降低(Huang et al., 2008; 万宏伟等, 2008), 说明氮添加不仅能够提高糙隐子草的光合能力, 还可能通过改变群落结构而改变糙隐子草生长的光环境.不同的放牧强度对糙隐子草生理和形态功能特征也有显著的影响(Zheng et al., 2011).放牧降低了糙隐子草叶和茎干质量、叶面积, 增加了叶片氮含量和胞间CO2浓度; 光合速率、蒸腾速率和气孔导度等光合生理性状在湿润年份和干旱年份对放牧的响应有所不同.原因可能是放牧改变了土壤养分和水分条件(Milchunas & Lauenroth, 1993), 进而导致了这些功能性状的变化(Adler et al., 2004, 2005).在不同的植物群落中, 糙隐子草叶片含水量在芨芨草(Achnatherum splendens)和冷蒿(Artemisia frigida)群落中高于线叶菊(Filifolium sibiricum)、羊草(Leymus chinensis)、大针茅(Stipa grandis)和小叶锦鸡儿(Caragana microphylla)群落(Chen et al., 2002).光合利用率在半干旱生境中较高, 水分利用率在干旱生境中较高(崔骁勇等, 2001). ...
Plant ecological strategies: Some leading dimensions of variation between species
1
2002
... 典型相关分析也表明, 叶片的形态性状和生理性状是高度相关的.单位面积的叶氮含量, 作为联系叶形态和生理功能的桥梁, 发挥着重要的作用.在糙隐子草生存环境的氮水条件发生改变时, 首先通过改变形态功能性状以适应环境的改变.当极端条件(例如高氮添加)发生时, 转向生理性状的改变.糙隐子草功能性状应对环境压力的不同响应能使其在波动的环境中或与其他优势种竞争时, 拥有更强的生存能力和竞争能力(Reich et al., 1997; Westoby et al., 2002; Reich et al., 2007). ...
Differential responses of plant functional trait to grazing between two contrasting dominant C3 and C4 species in a typical steppe of Inner Mongolia, China
3
2011
... 糙隐子草(Cleistogenes squarrosa)是一种多年生丛生小禾草, 属C4植物, 广泛分布于内蒙古高原的草甸草原、典型草原和荒漠草原, 是针茅草原和羊草草原的下层优势种(中国科学院内蒙古宁夏综合考察队, 1985), 也是C4植物中生物量比例最高的物种.相比于共存的C3植物, 糙隐子草具有高光合能力、高水分利用效率、抗干旱和耐牧等特点(汪诗平和王艳芬, 2001; Chen et al., 2002; 汪诗平等, 2003).在中度放牧条件下或退化草场恢复一段时间后, 其在群落中可处于优势地位(王鑫厅等, 2014).全球变化和人为干扰下, 糙隐子草功能性状的改变可能会影响草原生态系统的结构和功能.近年来, 糙隐子草功能性状的相关研究, 主要集中于氮添加、放牧等野外控制实验以及不同生境下的形态和生理功能性状(崔骁勇等, 2001; Chen et al., 2002, 2005; 万宏伟等, 2008; Zheng et al., 2011; 秦洁等, 2014).羊草草原长期氮添加实验结果表明, 糙隐子草叶片氮含量、叶绿素含量随施氮量的增加而增加, 叶绿素a和b的比值随施氮量的增加而降低(Huang et al., 2008; 万宏伟等, 2008), 说明氮添加不仅能够提高糙隐子草的光合能力, 还可能通过改变群落结构而改变糙隐子草生长的光环境.不同的放牧强度对糙隐子草生理和形态功能特征也有显著的影响(Zheng et al., 2011).放牧降低了糙隐子草叶和茎干质量、叶面积, 增加了叶片氮含量和胞间CO2浓度; 光合速率、蒸腾速率和气孔导度等光合生理性状在湿润年份和干旱年份对放牧的响应有所不同.原因可能是放牧改变了土壤养分和水分条件(Milchunas & Lauenroth, 1993), 进而导致了这些功能性状的变化(Adler et al., 2004, 2005).在不同的植物群落中, 糙隐子草叶片含水量在芨芨草(Achnatherum splendens)和冷蒿(Artemisia frigida)群落中高于线叶菊(Filifolium sibiricum)、羊草(Leymus chinensis)、大针茅(Stipa grandis)和小叶锦鸡儿(Caragana microphylla)群落(Chen et al., 2002).光合利用率在半干旱生境中较高, 水分利用率在干旱生境中较高(崔骁勇等, 2001). ...
... ), 说明氮添加不仅能够提高糙隐子草的光合能力, 还可能通过改变群落结构而改变糙隐子草生长的光环境.不同的放牧强度对糙隐子草生理和形态功能特征也有显著的影响(Zheng et al., 2011).放牧降低了糙隐子草叶和茎干质量、叶面积, 增加了叶片氮含量和胞间CO2浓度; 光合速率、蒸腾速率和气孔导度等光合生理性状在湿润年份和干旱年份对放牧的响应有所不同.原因可能是放牧改变了土壤养分和水分条件(Milchunas & Lauenroth, 1993), 进而导致了这些功能性状的变化(Adler et al., 2004, 2005).在不同的植物群落中, 糙隐子草叶片含水量在芨芨草(Achnatherum splendens)和冷蒿(Artemisia frigida)群落中高于线叶菊(Filifolium sibiricum)、羊草(Leymus chinensis)、大针茅(Stipa grandis)和小叶锦鸡儿(Caragana microphylla)群落(Chen et al., 2002).光合利用率在半干旱生境中较高, 水分利用率在干旱生境中较高(崔骁勇等, 2001). ...
 , 罗亚晨
, 罗亚晨

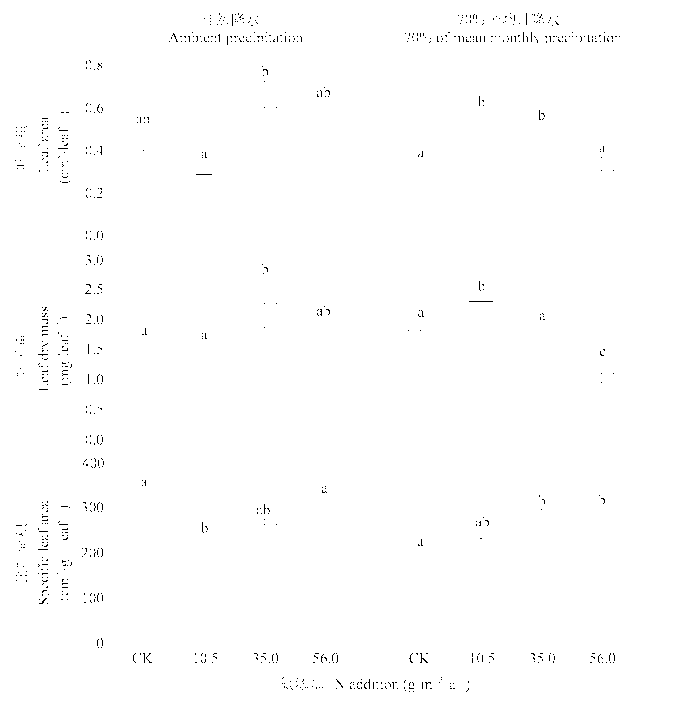

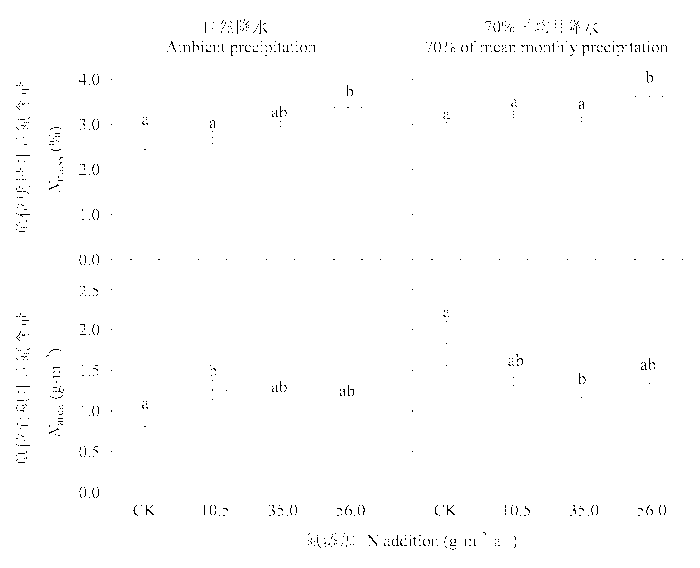
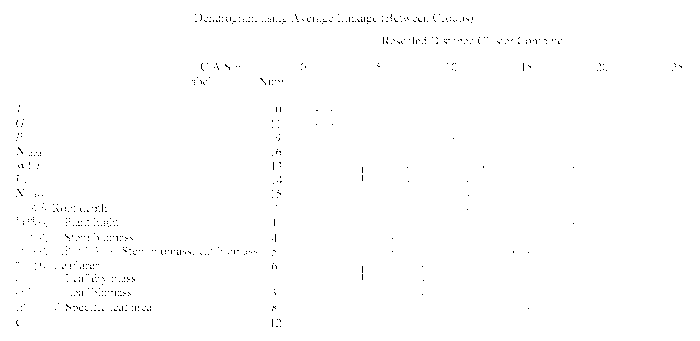
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}