子囊菌纲(Ascomycetes)麦角菌科(Clavicipitac- eae)的无性型Neotyphodium 属及有性型Epichloë 属真菌, 普遍分布于冷季型的禾本科植物中, 在植物体内完成其部分或全部生活史(Schardl et al ., 2004 )。Neotyphodium 属内生真菌与宿主植物的关系具有代表性的研究意义, 通过垂直方式传播, 在高羊茅(Festuca elata )、多年生黑麦草(Lolium perenne )、Achnatherum robustum 和羽茅(Achnatherum sibiricum )上都存在有这类内生真菌(Wei et al ., 2007 )。Neotyphodium 内生真菌的全部生活周期都在植物体内完成, 菌丝在宿主禾草开花期进入子房, 植物产生种子后, 内生真菌便存在于种子中, 从而可以通过种子进行传播, 其间不形成任何子座或孢子。目前所研究的禾草植物中的内生真菌大部分属于此类传播方式。Epichloë 属真菌能够感染7个族16个属以上的禾本科植物, 在世界范围内分布广泛(Moon et al ., 2004 ; Schardl et al ., 2004 )。 Epichloë 真菌通过水平方式传播, 偶尔能在植物的茎秆和花序上形成特征性的子座, 内生真菌形成有性孢子, 通过虫媒作用, 与其他植株上配型相反的有性孢子结合, 并产生子囊孢子, 侵染新的个体(Bultman & Leuchtm- ann, 2003 ), 因此宿主的有性繁殖被完全抑制。Leuchtmann和Clay发现大多数感染Epichloë typhina 的披碱草属(Elymus )植株的同一个个体上, 往往同时存在带有子座和产生种子的两类生殖枝, 内生真菌只在产生子座的生殖枝营有性生殖, 进行水平传播, 而内生真菌在多数产生种子的生殖枝随着宿主的无性繁殖进行垂直传播, 这种既存在水平传播又有垂直传播的方式称为混合传播方式(White, 1988 )。
中国21个地区披碱草(Elymus dahuricus )的染菌率调查表明, 有6个地区的披碱草种子中Neotypho- dium 的感染率在4.4%-100.0%, 绝大多数染菌的披碱草比不染菌的多产生一种生物碱——波胺, 波胺的浓度水平随着种群的位置和季节而变化(Zhang & Nan, 2007 )。在南京发现了Neotyphodium uncinatum 与苇状羊茅(Festuca arundinacea )的共生体(纪燕玲等, 2003 )。还有研究发现, Epichloë yangzii 是共生于鹅观草属(Roegneria )植物的产子座有性型内生真菌(申靖等, 2009 )。Wei等(2007) 对中国北方地区172个禾草地理种群的内生真菌感染状况进行广泛调查后发现, 在所有检测的41种禾草中, 25种禾草感染了内生真菌, 其中羽茅(Achnatherum sibiricum )的感染率最高, 这表明内生真菌广泛存在于天然禾草种群中, 而且内生真菌与禾草的相互作用可能对中国北方草原生态系统和畜牧业产生影响。
对高羊茅和黑麦草的研究发现, Neotyphodium 能提高宿主的生长(Marks et al ., 1991 )、耐旱性(Elmi & West, 1995 )、对养分的吸收和利用(Malinowski & Belesky, 2000 ), 以及宿主对动物的拒食能力(Clay, 1990 )。然而, 很多研究发现天然禾草的情况与栽培禾草的不一致甚至相反, 且Epichloë 与天然禾草共生对宿主的有益影响可能小于Neotyphodium, 原因可能是Epichloë 能在宿主部分或全部生殖枝上产生子座, 抑制宿主的开花和结实(Leuchtmann et al ., 1994 ); 也可能是多数Epichloë 不产生生物碱或生物碱的产生水平很低(Leuchtmann et al ., 2000 )。大部分研究都集中在Neotyphodium 对宿主的影响, 以及影响Epichloë 产子座的条件。关于Epichloë 对宿主的生理生态特性影响研究较少。有报道指出: 由于Epichloë typhina 在宿主细弱剪股颖(Agrostis tenuis )叶鞘上产生子座抑制宿主抽穗、开花, 从而增强了宿主地上、地下营养生长, 宿主分蘖数显著增多(de Battista et al ., 1990 )。在本研究中, 通过对感染不同传播方式的羽茅植株进行叶鞘分离, 我们得到Neotyphodium 和Epichloë 内生真菌, 从内生真菌的传播方式入手, 通过田间栽培试验, 探讨不同传播方式的Neotyphodium 和Epichloë 属内生真菌对羽茅生长与生理特性的影响。
1 材料和方法
1.1 实验材料
2009年4月, 在南开大学的网室样地中, 在8块1 m × 1 m的样方中播撒采自中国农业科学院呼伦贝尔草原生态系统实验站的饱满羽茅种子, 每块样方间距50 cm, 为了保证垂直传播的内生真菌Neotyphodium 和水平传播的内生真菌Epichloë 不感染到不染菌的样方, 我们每年在羽茅抽穗结种之前, 将刚抽穗但还未成熟的羽茅的穗剪掉, 防止羽茅种子成熟后感染到其他不染菌样方, 以确保在第二年的实验中不染菌样方中的羽茅植株仍是未受内生真菌感染的羽茅。随机分布选择4块样方播撒染菌种子, 相互间隔开的另外4块播撒不染菌种子, 每个样方播撒100-120粒。不染菌种子通过60 ℃高温处理后获得, 处理时间为30天。所有检测幼苗均不感染内生真菌。高温处理对羽茅的种子发芽率、发芽势和发芽指数均无显著影响(李夏等, 2010 )。2011年5月, 发现2块染菌样方中, 羽茅植株的生殖枝上产生子座, 另外2块染菌的样方中, 染菌羽茅植株并没有形成子座。我们分别于2011年和2012年测定标记好长子座的植株的生理生态特性, 测定指标前, 通过苯胺兰染色法在显微镜下检测植株的染菌状况, 并且在产生子座的植株中, 用叶鞘分离内生真菌的方法, 分离和纯化出与羽茅共生的通过水平传播的Epichloë, 从未产生子座的植株中分离得到垂直传播的Neotyphodium, 分离纯化得到的两个属内生真菌形态不同。我们将不同传播方式的羽茅分别标记为: Ne——感染Neotyphodium 的植株; Ep——感染Epichloë 的植株; EF——不染菌植株。EF、Ne、Ep所在样方土壤成分分别为: N%分别是0.033%、0.057%和0.097%; C%分别是1.19%、1.507%和2.16%。每块样方中选取3-5个重复进行羽茅生理生态指标的测定。
1.2 生长与生理指标测定
2011年5月和2012年5月, 测量植株的生长指标: 株高、叶片长和叶片宽、叶面积和比叶面积。选取晴天9:00-12:00, 利用植株新叶完全展开的第一片叶, 使用LI-6400便携式光合作用测定仪(LI-COR, Lincoln, USA)测定植株的气体交换特征参数。由LI-6400-02BLED红蓝光源自动控制光强, 叶温控制在环境温度(25 ± 0.5) ℃, CO2 浓度为自然状态下大气的浓度(大约为400 μmol·mol-1 ), 在光合有效辐射通量密度为1200 µmol·m-2 ·s-1 下, 测定叶片最大净光合速率(P max )等光合生理生态参数。
测定光合色素含量时采用乙醇-丙酮混合液法(无水乙醇:丙酮=1:1)浸泡提取色素, 分别测定浸泡液在470 nm、646 nm和663 nm处的吸光值。用测定的吸光度值计算羽茅叶片的叶绿素a、叶绿素b及类胡萝卜素的含量。叶片全氮含量的测定采用凯氏定氮法。有机质含量测定采用重铬酸钾氧化法, 指示剂采用邻菲罗啉, 用0.2 mol·L-1 FeSO4 溶液进行滴定。可溶性糖含量采用蒽酮比色法进行测定。总酚含量的测定参照Malinowski等(1998) 的方法, 采用无水乙醇:水(1:1)混合液浸提总酚, FeCl3 (0.9%)与铁氰化钾(0.6%)处理后, 在720 nm处测定吸光度得到总酚含量。非结构性碳水化合物(TNC, %)参照酶解法进行测定(da Silveira et al ., 1978 )。
1.3 数据分析
应用SPSS 13.0软件和SYSTAT 13.0软件进行统计分析, 对数据采用单因素方差分析(one-way ANO- VA), 用最小显著差异法(LSD )(p < 0.05)进行比较。
2 结果
2.1 不同传播方式的内生真菌感染对羽茅生长的影响
由图1 可知, 不同传播方式的内生真菌对羽茅的生长指标影响不同。实验测定的第一年, 垂直传播的内生真菌显著增加了羽茅叶长与叶宽之比(Ne: (56.62 ± 4.15), EF: (37.02 ± 2.71), Ep: (42.58 ± 3.88)) (图1 )。然而, 2011和2012年两年都测得水平传播的内生真菌显著降低了羽茅的株高(图1 , 图2 )。不同传播方式的内生真菌感染羽茅后, 其叶片长度具有显著差异(Ne: (50.60 ± 2.28) cm, EF: (42.82 ± 1.92) cm, Ep: (23.24 ± 0.75) cm), 在2011年5月测得的变化趋势为: Ne > EF > Ep (图1 )。
图1 2011年感染不同传播方式内生真菌Epichloë 和Neotyphodium 对羽茅生长状况的影响(平均值±标准误差)。不同小写字母表示p 值在0.05的水平上差异显著。EF, 不染菌植株; Ep, 感染Epichloë 的植株; Ne, 感染Neotyphodium 的植株。
Fig. 1 Effects of Epichloë and Neotyphodium infection on growth of Achnatherum sibiricum in 2011 (mean ± SE ). Different lowercase letters indicate significant differences at the 0.05 level. EF, endophyte-free plant; Ep, plant infected by Epichloë; Ne, plant infected by Neotyphodium .
从图2 看出, 实验测定的第二年受到垂直传播内生真菌感染的植株和不染菌植株的叶面积都显著高于感染水平传播内生真菌的植株(图2 )。感染Epichloë 的植株比叶面积(SLA )显著高于不染菌植株和感染Neotyphodium 的植株, 但这一变化在第一年的观测中没有表现出来(Ne: (25.36 ± 0.49) m2 ·kg-1 , EF: (23.70 ± 1.30) m2 ·kg-1 , Ep: (25.05 ± 1.07) m2 ·kg-1 ) (图3 )。
图2 2012年感染不同传播方式内生真菌Epichloë 和Neotyphodium 对羽茅生长状况的影响(平均值±标准偏差)。不同小写字母表示p 值在0.05的水平上差异显著。EF, 不染菌植株; Ep, 感染Epichloë 的植株; Ne, 感染Neotyphodium 的植株。
Fig. 2 Effects of Epichloë and Neotyphodium infection on growth of Achnatherum sibiricum in 2012 (mean ± SD ). Different lowercase letters indicate significant differences at the 0.05 level. EF, endophyte-free plant; Ep, plant infected by Epichloë; Ne, plant infected by Neotyphodium .
图3 感染不同传播方式内生真菌Epichloë 和Neotyphodium 对羽茅比叶面积的影响(平均值±标准偏差)。不同小写字母表示p 值在0.05的水平上差异显著。EF, 不染菌植株; Ep, 感染Epichloë 的植株; Ne, 感染Neotyphodium 的植株。
Fig. 3 Effects of Epichloë and Neotyphodium infection on specific leaf area in Achnatherum sibiricum (mean ± SD ). Different lowercase letters indicate significant differences at the 0.05 level. EF, endophyte-free plant; Ep, plant infected by Epichloë; Ne, plant infected by Neotyphodium .
2.2 不同传播方式的内生真菌感染对羽茅光合色素含量与光合特性的影响
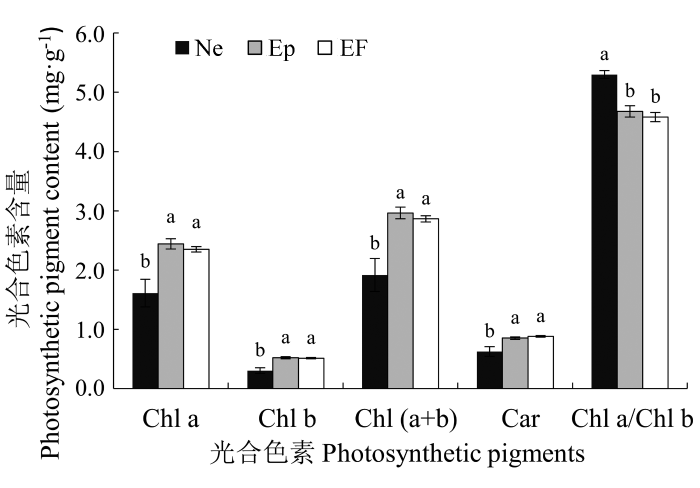
从图4 可以看出, 实验进行的第一年测得不同传播方式的内生真菌对羽茅光合色素含量的影响不同。不染菌植株的类胡萝卜素含量显著高于感染垂直传播内生真菌的植株(EF: (0.88 ± 0.01) mg·g-1 ; Ne: (0.63 ± 0.08) mg·g-1 ) (图4 ), 感染Neotyphodium 属内生真菌的羽茅叶绿素a/b显著高于Ep和EF (Ne: (5.30 ± 0.06), EF: (4.58 ± 0.08), Ep: (4.68 ± 0.10))(图4 )。感染Epichloë 内生真菌有促进羽茅光合色素积累的趋势, 但是差异不显著。较高的光合色素含量导致不染菌植株具有较大的最大净光合速率。内生真菌感染显著影响了羽茅的最大净光合速率, 在两年的测定中都得到相同的结果: 不染菌植株的最大净光合显著高于感染Epichloë 的植株和感染Neotypho- dium 的植株, 感染Epichloë 的羽茅最大净光合速率显著低于感染Neotyphodium 的羽茅(图5 )。
图4 2011年感染不同传播方式内生真菌Epichloë 和Neotyphodium 对羽茅光合色素的影响(平均值±标准误差)。不同小写字母表示p 值在0.05的水平上差异显著。EF, 不染菌植株; Ep, 感染Epichloë 的植株; Ne, 感染Neotyphodium 的植株。Car, 胡萝卜素; Chl a, 叶绿素a; Chl b, 叶绿素b。
Fig. 4 The influence of Epichloë and Neotyphodium infection on photosynthetic pigments in Achnatherum sibiricum in 2011 (mean ± SE ). Different lowercase letters indicate significant differences at the 0.05 level. EF, endophyte-free plant; Ep, plant infected by Epichloë; Ne, plant infected by Neotyphodium . Car, carotene; Chl a, chlorophyll a; Chl b, chlorophyll b.
图5 感染不同传播方式内生真菌Epichloë 和Neotyphodium 对羽茅最大净光合速率的影响(平均值±标准误差)。不同字母表示p 值在0.05的水平上差异显著。EF, 不染菌植株; Ep, 感染Epichloë 的植株; Ne, 感染Neotyphodium 的植株。
Fig. 5 The influence of Epichloë and Neotyphodium infection on maximum net photosynthetic rate in Achnatherum sibiricum (mean ± SE ). Different lowercase letters indicate significant differences at the 0.05 level. EF, endophyte-free plant; Ep, plant infected by Epichloë; Ne, plant infected by Neotyphodium .
在羽茅的光合特性中, 实验进行的第一年测得感染Neotyphodium 的内生真菌的羽茅具有最大的水分利用效率, 并且是不染菌植株的水分利用效率的1.14倍, 实验进行的第二年我们发现, 两种传播方式感染内生真菌的羽茅水分利用效率都显著高于不染菌植株(表1 )。不染菌植株的光能利用效率在两年的观测中得到一致的结果, 都显著高于分别感染两种不同传播方式的内生真菌的羽茅植株。在第一年的测定结果中, 感染Epichloë 的羽茅的气孔导度和水分利用效率显著低于感染Neotyphodium 的羽茅和未染菌的羽茅, 并且水平传播的内生真菌对羽茅的光合指标(除了胞间CO2 浓度外)的影响均为负效应。
2.3 不同传播方式的内生真菌感染对羽茅可溶性糖和总酚含量的影响
实验进行的第一年测得: Ne可溶性糖的含量显著增加了, 与EF相比, 染菌植株更趋向于在叶片中积累更多的可溶性糖。Ne与Ep可溶性糖含量无显著差异(图6A )。Ne总酚含量最高, 但不同传播方式内生真菌的羽茅之间无显著差异(图6B ), 同样地, 实验进行的第二年测得植物中非结构性碳水化合物的含量也不存在显著差异(图6C )。
图6 感染不同传播方式内生真菌Epichloë 和Neotyphodium 对2011年羽茅可溶性糖和总酚含量的影响以及对2012年羽茅的非结构性碳含量的影响(平均值±标准偏差)。A, 可溶性糖含量。B, 总酚含量。C, 非结构性碳水化合物含量。不同小写字母表示p 值在0.05的水平上差异显著。EF, 不染菌植株; Ep, 感染Epichloë 的植株; Ne, 感染Neotyphodium 的植株。
Fig. 6 Effects of Epichloë and Neotyphodium infection on soluble sugar and total phenolic contents in 2011 and the total nonstructural carbohydrate in 2012 in Achnatherum sibiricum (mean ± SD ). A, Soluble sugar content. B, Total phenolic content. C, Total nonstructural carbohydrate content. Different lowercase letters indicate significant differences at the 0.05 level. EF, endophyte-free plant; Ep, plant infected by Epichloë; Ne, plant infected by Neotyphodium .
2.4 不同传播方式的内生真菌感染对羽茅碳氮含量的影响
感染不同传播方式的内生真菌对羽茅氮含量和碳含量的影响不同。感染Epichloë 的羽茅氮含量显著高于感染Neotyphodium 的羽茅(图7 )。感染Epichloë 的羽茅碳含量显著高于感染Neotyphodium 的羽茅和未染菌羽茅。感染Neotyphodium 的羽茅碳氮比C:N显著高于感染Epichloë 的羽茅(图7 )。
图7 2012年感染不同传播方式内生真菌Epichloë 和Neotyphodium 对羽茅的碳、氮含量以及碳氮比的影响(平均值±标准偏差)。不同小写字母表示p 值在0.05的水平上差异显著。EF, 不染菌植株; Ep, 感染Epichloë 的植株; Ne, 感染Neotyphodium 的植株。
Fig. 7 Effects of Epichloë and Neotyphodium infection on nitrogen content, carbon content and C:N in Achnatherum sibiricum in 2012 (mean ± SD ). Different lowercase letters indicate significant differences at the 0.05 level. EF, endophyte-free plant; Ep, plant infected by Epichloë; Ne, plant infected by Neotyphodium .
3 讨论
Bradshaw和Snaydon (1959) 最早报道内生真菌可以影响宿主植物的生长。但关于不同传播方式的内生真菌, 尤其是水平传播方式的内生真菌对宿主植物的影响相对较少, 大部分研究集中在垂直传播的内生真菌Neotyphodium 对宿主植物的影响。有研究认为Neotyphodium 与宿主植物在长期的共同进化过程中形成了一种互利共生的关系, 可促进植物生长和分蘖形成, 增强植物抗病害、抗动物采食及抗恶劣环境等能力(Moon et al ., 2004 )。已有研究表明: 通过和宿主植物及其生长环境的相互作用, Epichloë 在宿主植物旗叶的叶鞘或茎秆上形成子座, 未成熟的子座由一层致密的菌丝层组成, 表面密布分生孢子, 但关于这些真菌影响植物的生长和生理指标的研究较少。本研究不仅从生有子座的羽茅中分离出有性型内生真菌, 还得出垂直传播的内生真菌显著促进了植物的叶片长宽比, 而水平传播的内生真菌对羽茅的株高和叶宽有显著的负效应。Groppe等(1999) 也得出类似的结果, 认为植物生长能力较强有助于植物中内生真菌的传播。垂直传播的内生真菌对宿主植物生长的改变可能与其产生吲哚乙酸有关(de Battista et al ., 1990 ), 这也可能与植物中内生真菌的传播方式有关(Tintjer et al ., 2008 )。
对于感染不同传播方式内生真菌的羽茅而言, 染菌植株的蒸腾速率、气孔导度和光能利用效率都显著高于不染菌植株。这表明内生真菌对植株的光合作用有一定的促进作用。这可能对宿主植物的传播方式也有一定的影响(Cheplick & Faeth, 2009 )。我们发现无性型内生真菌可以提高羽茅的气孔限制值和水分利用效率, 但有性型内生真菌的作用却复杂多变。Morse等(2002) 研究发现, 不染菌的亚利桑那羊茅(Festuca arizonica )表现出更高的光合速率和气孔导度。可见, 羽茅-内生真菌之间的共生关系在光合特性方面不仅与羽茅的染菌状况有关, 也与感染内生真菌的传播方式有一定的关系。
碳水化合物是植物中碳库的储存(Donaghy & Fulkerson, 1997 , 1998 ; Cheplick & Chui, 2001 )。已有研究表明内生真菌可以增加多年生黑麦草中碳水化合物的含量(Cheplick & Cho, 2003 ; Hunt et al ., 2005 ; Rasmussen et al ., 2007 )。本研究中, 水平传播的内生真菌促使宿主具有更高的碳含量, 但垂直传播的内生真菌使宿主体内具有更高的碳氮比。可溶性糖是一种重要的渗透调节物质, 在干旱胁迫过程中, 植物体内可溶性糖含量的变化在一定程度上能反映其对不良环境的适应能力(Sánchez et al ., 1998 )。我们还发现感染垂直传播的内生真菌的羽茅比不染菌植株和感染有性型内生真菌的植株中积累更多的可溶性糖。本研究中垂直传播的内生真菌使宿主中储存更多的可溶性糖, 这对植物是有益处的, 而水平传播的内生真菌并没有这一表现, 并且垂直传播的内生真菌使宿主产生的总酚含量高于水平传播的内生真菌, 由此我们推测, 当感染不同传播方式的内生真菌的宿主受到生物或非生物胁迫时, 通过垂直传播方式感染内生真菌的植株防御能力很可能强于通过水平传播方式感染内生真菌的植株, 这还有待进一步的实验研究。
宿主种群间不同的传播方式也可能是由于内生真菌或者宿主植物的基因型不同(Bucheli & Leuchtmann, 1996 ; Kover & Clay, 1998 )。不同传播方式的内生真菌对羽茅生长与生理特性的影响不同, 还可能是由于大部分Epichloë 并不是每年都能在宿主植物上产生子座, 子座的产生与否, 不仅与内生真菌种类以及宿主植物的基因型有关, 而且与植物的生长环境、营养状态等有关, 这还有待于进一步研究。
致谢 感谢南开大学生态学实验室尹立佳、魏茂英、王欣禹在实验工作中给予的帮助。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Bradshaw AD Snaydon RW 1959 ). Population differentiation within plant species in response to soil factors
. Nature , 183 , 129 -130 .
[本文引用: 1]
[2]
Bucheli E Leuchtmann A 1996 ). Evidence for genetic
[本文引用: 1]
[3]
differentiation between choke-inducing and asymptomtic strains of the Epichloë grass endophyte from Brachy- podium sylvaticum.Evolution , 50 , 1879 -1887 .
[4]
Bultman TL Leuchtmann A 2003 ). A test of host specialization by insect vectors as a mechanism for reproductive isolation among entomophilous fungal species
. Oikos , 103 , 681 -687 .
[本文引用: 1]
[5]
Cheplick GP Cho R 2003 ). Interactive effects of fungal endophyte infection and host genotype on growth and storage in Lolium perenne
. New Phytologist , 158 , 183 -191 .
[本文引用: 1]
[6]
Cheplick GP Chui T 2001 ). Effects of competitive stress on vegetative growth, storage, and regrowth after defoliation in Phleum pratense
. Oikos , 95 , 291 -299 .
[本文引用: 1]
[7]
Cheplick GP Faeth S 2009). Ecology and Evolution of the Grass-endophyte Symbiosis . Oxford University Press, Oxford, UK .
[本文引用: 2]
[8]
Clay K 1990 ). Fungal endophytes of grasses
. Annual Review of Ecology and Systematics , 21 , 275 -297 .
[9]
da Silveira AJ Feitosa TFF Stull JW 1978 ). A rapid technique for total nonstructural carbohydrate determination of plant tissue
. Journal of Agricutural and Food Chemistry , 26 , 770 -772 .
[本文引用: 1]
[10]
de Battista JP Bacon CW Severson R Plattner RD Bouton JH 1990 ). Indole acetic acid production by the fungal endophyte of tall fescue
. Agronomy Journal , 82 , 878 -880 .
[本文引用: 2]
[11]
Donaghy DJ Fulkerson WJ 1997 ). The importance of water-soluble carbohydrate reserves on regrowth and root growth of Lolium perenne (L.)
. Grass and Forage Science , 52 , 401 -407 .
[本文引用: 1]
[12]
Donaghy DJ Fulkerson WJ 1998 ). Priority for allocation of water-soluble carbohydrate reserves during regrowth of Lolium perenne
. Grass and Forage Science , 53 , 211 -218 .
[本文引用: 1]
[13]
Elmi AA West CP 1995 ). Endophyte infection effects on stomatal conductance, osmotic adjustment and drought recovery of tall fescue
. New Phytologist , 131 , 61 -67 .
[本文引用: 1]
[14]
Groppe K Steinger T Sanders I Schmid B Wiemken A Boller T 1999 ). Interaction between the endophytic fungus Epichloe bromicola and the grass Bromus erectus: Effects of endophyte infection, fungal concentration and environment on grass growth and flowering
. Molecular Ecology , 8 , 1827 -1835 .
[本文引用: 2]
[15]
Hunt MG Rasmussen S Newton PCD Parsons AJ Newman JA 2005 ). Near-term impacts of elevated CO2 , nitrogen and fungal endophyte-infection on Lolium perenne L. growth, chemical composition and alkaloid production
. Plant, Cell & Environment , 28 , 1345 -1354 .
[16]
Ji YL Wang ZW Yu HS Wang SM 2003 ). Neotyphodium uncinatum, an endophytic fungus obtained from Festuca arundinacea Schreb
. Journal of Nanjing Agricultural University , 26 (2 ), 47 -50 .
[本文引用: 1]
(in Chinese with English abstract) [纪燕玲 , 王志伟 , 于汉寿 , 王世梅 (2003 ). 分离自苇状羊茅(Festucaa rundinacea Schreb.)的内生真菌Neotyphodium uncinatum
. 南京农业大学学报 , 26 (2 ), 47 -50 .]
[本文引用: 1]
[17]
Kover PX Clay K 1998 ). Trade-off between virulence and vertical transmission and the maintenance of a virulent plant pathogen
. The American Naturalist , 152 , 165 -175 .
[本文引用: 2]
[18]
Leuchtmann A Schardl CL Siegel MR 1994 ). Sexual compatibility and taxonomy of a new species of Epichloe symbiotic with fine Fescue grasses
. Mycologia , 86 , 802 -812 .
[本文引用: 1]
[19]
Leuchtmann A Schmidt D Bush LP 2000 ). Different levels of protective alkaloids in grasses with stroma-forming and seed-transmitted Epichloë/Neotyphodium endophytes
. Jou- rnal of Chemical Ecology , 26 , 1025 -1036 .
[20]
Li X Han R Ren AZ Gao YB 2010 ). Using high-temperature treatment to construct endophyte-free Achnatherum sibiricum
. Microbiology China , 37 , 1395 -1400 .
[本文引用: 1]
[李夏 , 韩荣 , 任安芝 , 高玉葆 (2010 ). 高温处理构建不感染内生真菌羽茅种群的方法探讨
. 微生物学通报 , 37 , 1395 -1400 .]
[本文引用: 1]
[21]
Malinowski DP Alloush GA Belesky DP 1998 ). Evidence for chemical changes on the root surface of tall fescue in response to infection with the fungal endophyte Neotyphodium coenophialum
. Plant and Soil , 205 , 1 -12 .
[本文引用: 1]
[22]
Malinowski DP Belesky DP 2000 ). Adaptations of endophyte-infected cool-season grasses to environmental stresses: Mechanisms of drought and mineral stress tolerance
. Crop Science , 40 , 923 -940 .
[本文引用: 2]
[23]
Marks S Clay K Cheplick GP 1991 ). Effects of fungal endophytes on interspecific and intraspecific competition in the grasses Festuca arundinacea and Lolium perenne
. Journal of Applied Ecology , 28 , 194 -204 .
[本文引用: 2]
[24]
Moon CD Craven KD Leuchtmann A Clement SL Schardl CL 2004 ). Prevalence of interspecific hybrids amongst asexual fungal endophytes of grasses
. Molecular Ecology , 13 , 1455 -1467 .
[25]
Morse LJ Day TA Faeth SH 2002 ). Effect of Neotyphodium endophyte infection on growth and leaf gas exchange of Arizona fescue under contrasting water availability regimes
. Environmental and Experimental Botany , 48 , 257 -268 .
[本文引用: 2]
[26]
Rasmussen S Parsons AJ Bassett S Christensen MJ Hume DE Johnson LJ Johnson RD Simpson WR Stacke C Voisey CR Xue H Newman JA 2007 ). High nitrogen supply and carbohydrate content reduce fungal endophyte and alkaloid concentration in Lolium perenne
. New Phytologist , 173 , 787 -797 .
[27]
Sánchez FJ Manzanares M de Andres EF Tenorio JL Ayerbe L 1998 ). Turgor maintenance, osmotic adjustment and soluble sugar and proline accumulation in 49 pea cultivars in response to water stress
. Field Crops Research , 59 , 225 -235 .
[本文引用: 2]
[28]
Schardl CL Leuchtmann A Spiering MJ 2004 ). Symbioses of grasses with seedborne fungal endophytes
. Annual Review of Plant Biology , 55 , 315 -340 .
[本文引用: 1]
[29]
Shen J Tao WW Chen C Chen YG Wang ZW 2009 ). Review on the grass endophyte research in China 9―Seed transmission characteristics and distribution in host plants of Epichloё yangzii
. Pratacultural Science , 26 , 146 -151 .
[本文引用: 2]
(in Chinese with English abstract) [申靖 , 陶文文 , 陈昌 , 陈永敢 , 王志伟 (2009 ). 禾本科植物内生真菌研究9——Epichloё yangzii的种传特性及其在宿主体内的分布
. 草业科学 , 26 , 146 -151 .]
[本文引用: 2]
[30]
Tintjer T Leuchtmann A Clay K 2008 ). Variation in horizontal and vertical transmission of the endophyte Epichloë elymi infecting the grass Elymus hystrix
. New Phytologist , 179 , 236 -246 .
[本文引用: 1]
[31]
Wei YK Gao YB Zhang X Su D Wang YH Xu H Lin F Ren AZ Chen L Nie LY 2007 ). Distribution and diversity of Epichloë/Neotyphodium fungal endophytes from different populations of Achnatherum sibiricum (Poaceae) in the Inner Mongolia Steppe, China
. Fungal Diversity , 24 , 329 -345 .
[本文引用: 2]
[32]
White JF 1988 ). Endophyte-host associations in forage grasses. XI. A proposal concerning origin and evolution
. Mycologia , 80 , 442 -446 .
[33]
Zhang YP Nan ZB 2007 ). Distribution of Epichloë endophytes in Chinese populations of Elymus dahuricus and variation in peramine levels
. Symbiosis , 43 , 13 -19 .
[本文引用: 1]
Population differentiation within plant species in response to soil factors
1
1959
... Bradshaw和Snaydon (1959) 最早报道内生真菌可以影响宿主植物的生长.但关于不同传播方式的内生真菌, 尤其是水平传播方式的内生真菌对宿主植物的影响相对较少, 大部分研究集中在垂直传播的内生真菌Neotyphodium 对宿主植物的影响.有研究认为Neotyphodium 与宿主植物在长期的共同进化过程中形成了一种互利共生的关系, 可促进植物生长和分蘖形成, 增强植物抗病害、抗动物采食及抗恶劣环境等能力(Moon et al ., 2004 ).已有研究表明: 通过和宿主植物及其生长环境的相互作用, Epichloë 在宿主植物旗叶的叶鞘或茎秆上形成子座, 未成熟的子座由一层致密的菌丝层组成, 表面密布分生孢子, 但关于这些真菌影响植物的生长和生理指标的研究较少.本研究不仅从生有子座的羽茅中分离出有性型内生真菌, 还得出垂直传播的内生真菌显著促进了植物的叶片长宽比, 而水平传播的内生真菌对羽茅的株高和叶宽有显著的负效应.Groppe等(1999) 也得出类似的结果, 认为植物生长能力较强有助于植物中内生真菌的传播.垂直传播的内生真菌对宿主植物生长的改变可能与其产生吲哚乙酸有关(de Battista et al ., 1990 ), 这也可能与植物中内生真菌的传播方式有关(Tintjer et al ., 2008 ). ...
Evidence for genetic
1
1996
... 宿主种群间不同的传播方式也可能是由于内生真菌或者宿主植物的基因型不同(Bucheli & Leuchtmann, 1996 ; Kover & Clay, 1998 ).不同传播方式的内生真菌对羽茅生长与生理特性的影响不同, 还可能是由于大部分Epichloë 并不是每年都能在宿主植物上产生子座, 子座的产生与否, 不仅与内生真菌种类以及宿主植物的基因型有关, 而且与植物的生长环境、营养状态等有关, 这还有待于进一步研究. ...
A test of host specialization by insect vectors as a mechanism for reproductive isolation among entomophilous fungal species
1
2003
... 子囊菌纲(Ascomycetes)麦角菌科(Clavicipitac- eae)的无性型Neotyphodium 属及有性型Epichloë 属真菌, 普遍分布于冷季型的禾本科植物中, 在植物体内完成其部分或全部生活史(Schardl et al ., 2004 ).Neotyphodium 属内生真菌与宿主植物的关系具有代表性的研究意义, 通过垂直方式传播, 在高羊茅(Festuca elata )、多年生黑麦草(Lolium perenne )、Achnatherum robustum 和羽茅(Achnatherum sibiricum )上都存在有这类内生真菌(Wei et al ., 2007 ).Neotyphodium 内生真菌的全部生活周期都在植物体内完成, 菌丝在宿主禾草开花期进入子房, 植物产生种子后, 内生真菌便存在于种子中, 从而可以通过种子进行传播, 其间不形成任何子座或孢子.目前所研究的禾草植物中的内生真菌大部分属于此类传播方式.Epichloë 属真菌能够感染7个族16个属以上的禾本科植物, 在世界范围内分布广泛(Moon et al ., 2004 ; Schardl et al ., 2004 ). Epichloë 真菌通过水平方式传播, 偶尔能在植物的茎秆和花序上形成特征性的子座, 内生真菌形成有性孢子, 通过虫媒作用, 与其他植株上配型相反的有性孢子结合, 并产生子囊孢子, 侵染新的个体(Bultman & Leuchtm- ann, 2003 ), 因此宿主的有性繁殖被完全抑制.Leuchtmann和Clay发现大多数感染Epichloë typhina 的披碱草属(Elymus )植株的同一个个体上, 往往同时存在带有子座和产生种子的两类生殖枝, 内生真菌只在产生子座的生殖枝营有性生殖, 进行水平传播, 而内生真菌在多数产生种子的生殖枝随着宿主的无性繁殖进行垂直传播, 这种既存在水平传播又有垂直传播的方式称为混合传播方式(White, 1988 ). ...
Interactive effects of fungal endophyte infection and host genotype on growth and storage in Lolium perenne
1
2003
... 碳水化合物是植物中碳库的储存(Donaghy & Fulkerson, 1997 , 1998 ; Cheplick & Chui, 2001 ).已有研究表明内生真菌可以增加多年生黑麦草中碳水化合物的含量(Cheplick & Cho, 2003 ; Hunt et al ., 2005 ; Rasmussen et al ., 2007 ).本研究中, 水平传播的内生真菌促使宿主具有更高的碳含量, 但垂直传播的内生真菌使宿主体内具有更高的碳氮比.可溶性糖是一种重要的渗透调节物质, 在干旱胁迫过程中, 植物体内可溶性糖含量的变化在一定程度上能反映其对不良环境的适应能力(Sánchez et al ., 1998 ).我们还发现感染垂直传播的内生真菌的羽茅比不染菌植株和感染有性型内生真菌的植株中积累更多的可溶性糖.本研究中垂直传播的内生真菌使宿主中储存更多的可溶性糖, 这对植物是有益处的, 而水平传播的内生真菌并没有这一表现, 并且垂直传播的内生真菌使宿主产生的总酚含量高于水平传播的内生真菌, 由此我们推测, 当感染不同传播方式的内生真菌的宿主受到生物或非生物胁迫时, 通过垂直传播方式感染内生真菌的植株防御能力很可能强于通过水平传播方式感染内生真菌的植株, 这还有待进一步的实验研究. ...
Effects of competitive stress on vegetative growth, storage, and regrowth after defoliation in Phleum pratense
1
2001
... 碳水化合物是植物中碳库的储存(Donaghy & Fulkerson, 1997 , 1998 ; Cheplick & Chui, 2001 ).已有研究表明内生真菌可以增加多年生黑麦草中碳水化合物的含量(Cheplick & Cho, 2003 ; Hunt et al ., 2005 ; Rasmussen et al ., 2007 ).本研究中, 水平传播的内生真菌促使宿主具有更高的碳含量, 但垂直传播的内生真菌使宿主体内具有更高的碳氮比.可溶性糖是一种重要的渗透调节物质, 在干旱胁迫过程中, 植物体内可溶性糖含量的变化在一定程度上能反映其对不良环境的适应能力(Sánchez et al ., 1998 ).我们还发现感染垂直传播的内生真菌的羽茅比不染菌植株和感染有性型内生真菌的植株中积累更多的可溶性糖.本研究中垂直传播的内生真菌使宿主中储存更多的可溶性糖, 这对植物是有益处的, 而水平传播的内生真菌并没有这一表现, 并且垂直传播的内生真菌使宿主产生的总酚含量高于水平传播的内生真菌, 由此我们推测, 当感染不同传播方式的内生真菌的宿主受到生物或非生物胁迫时, 通过垂直传播方式感染内生真菌的植株防御能力很可能强于通过水平传播方式感染内生真菌的植株, 这还有待进一步的实验研究. ...
2
... 对高羊茅和黑麦草的研究发现, Neotyphodium 能提高宿主的生长(Marks et al ., 1991 )、耐旱性(Elmi & West, 1995 )、对养分的吸收和利用(Malinowski & Belesky, 2000 ), 以及宿主对动物的拒食能力(Clay, 1990 ).然而, 很多研究发现天然禾草的情况与栽培禾草的不一致甚至相反, 且Epichloë 与天然禾草共生对宿主的有益影响可能小于Neotyphodium, 原因可能是Epichloë 能在宿主部分或全部生殖枝上产生子座, 抑制宿主的开花和结实(Leuchtmann et al ., 1994 ); 也可能是多数Epichloë 不产生生物碱或生物碱的产生水平很低(Leuchtmann et al ., 2000 ).大部分研究都集中在Neotyphodium 对宿主的影响, 以及影响Epichloë 产子座的条件.关于Epichloë 对宿主的生理生态特性影响研究较少.有报道指出: 由于Epichloë typhina 在宿主细弱剪股颖(Agrostis tenuis )叶鞘上产生子座抑制宿主抽穗、开花, 从而增强了宿主地上、地下营养生长, 宿主分蘖数显著增多(de Battista et al ., 1990 ).在本研究中, 通过对感染不同传播方式的羽茅植株进行叶鞘分离, 我们得到Neotyphodium 和Epichloë 内生真菌, 从内生真菌的传播方式入手, 通过田间栽培试验, 探讨不同传播方式的Neotyphodium 和Epichloë 属内生真菌对羽茅生长与生理特性的影响. ...
... 对于感染不同传播方式内生真菌的羽茅而言, 染菌植株的蒸腾速率、气孔导度和光能利用效率都显著高于不染菌植株.这表明内生真菌对植株的光合作用有一定的促进作用.这可能对宿主植物的传播方式也有一定的影响(Cheplick & Faeth, 2009 ).我们发现无性型内生真菌可以提高羽茅的气孔限制值和水分利用效率, 但有性型内生真菌的作用却复杂多变.Morse等(2002) 研究发现, 不染菌的亚利桑那羊茅(Festuca arizonica )表现出更高的光合速率和气孔导度.可见, 羽茅-内生真菌之间的共生关系在光合特性方面不仅与羽茅的染菌状况有关, 也与感染内生真菌的传播方式有一定的关系. ...
Fungal endophytes of grasses
0
1990
A rapid technique for total nonstructural carbohydrate determination of plant tissue
1
1978
... 测定光合色素含量时采用乙醇-丙酮混合液法(无水乙醇:丙酮=1:1)浸泡提取色素, 分别测定浸泡液在470 nm、646 nm和663 nm处的吸光值.用测定的吸光度值计算羽茅叶片的叶绿素a、叶绿素b及类胡萝卜素的含量.叶片全氮含量的测定采用凯氏定氮法.有机质含量测定采用重铬酸钾氧化法, 指示剂采用邻菲罗啉, 用0.2 mol·L-1 FeSO4 溶液进行滴定.可溶性糖含量采用蒽酮比色法进行测定.总酚含量的测定参照Malinowski等(1998) 的方法, 采用无水乙醇:水(1:1)混合液浸提总酚, FeCl3 (0.9%)与铁氰化钾(0.6%)处理后, 在720 nm处测定吸光度得到总酚含量.非结构性碳水化合物(TNC, %)参照酶解法进行测定(da Silveira et al ., 1978 ). ...
Indole acetic acid production by the fungal endophyte of tall fescue
2
1990
... 对高羊茅和黑麦草的研究发现, Neotyphodium 能提高宿主的生长(Marks et al ., 1991 )、耐旱性(Elmi & West, 1995 )、对养分的吸收和利用(Malinowski & Belesky, 2000 ), 以及宿主对动物的拒食能力(Clay, 1990 ).然而, 很多研究发现天然禾草的情况与栽培禾草的不一致甚至相反, 且Epichloë 与天然禾草共生对宿主的有益影响可能小于Neotyphodium, 原因可能是Epichloë 能在宿主部分或全部生殖枝上产生子座, 抑制宿主的开花和结实(Leuchtmann et al ., 1994 ); 也可能是多数Epichloë 不产生生物碱或生物碱的产生水平很低(Leuchtmann et al ., 2000 ).大部分研究都集中在Neotyphodium 对宿主的影响, 以及影响Epichloë 产子座的条件.关于Epichloë 对宿主的生理生态特性影响研究较少.有报道指出: 由于Epichloë typhina 在宿主细弱剪股颖(Agrostis tenuis )叶鞘上产生子座抑制宿主抽穗、开花, 从而增强了宿主地上、地下营养生长, 宿主分蘖数显著增多(de Battista et al ., 1990 ).在本研究中, 通过对感染不同传播方式的羽茅植株进行叶鞘分离, 我们得到Neotyphodium 和Epichloë 内生真菌, 从内生真菌的传播方式入手, 通过田间栽培试验, 探讨不同传播方式的Neotyphodium 和Epichloë 属内生真菌对羽茅生长与生理特性的影响. ...
... Bradshaw和Snaydon (1959) 最早报道内生真菌可以影响宿主植物的生长.但关于不同传播方式的内生真菌, 尤其是水平传播方式的内生真菌对宿主植物的影响相对较少, 大部分研究集中在垂直传播的内生真菌Neotyphodium 对宿主植物的影响.有研究认为Neotyphodium 与宿主植物在长期的共同进化过程中形成了一种互利共生的关系, 可促进植物生长和分蘖形成, 增强植物抗病害、抗动物采食及抗恶劣环境等能力(Moon et al ., 2004 ).已有研究表明: 通过和宿主植物及其生长环境的相互作用, Epichloë 在宿主植物旗叶的叶鞘或茎秆上形成子座, 未成熟的子座由一层致密的菌丝层组成, 表面密布分生孢子, 但关于这些真菌影响植物的生长和生理指标的研究较少.本研究不仅从生有子座的羽茅中分离出有性型内生真菌, 还得出垂直传播的内生真菌显著促进了植物的叶片长宽比, 而水平传播的内生真菌对羽茅的株高和叶宽有显著的负效应.Groppe等(1999) 也得出类似的结果, 认为植物生长能力较强有助于植物中内生真菌的传播.垂直传播的内生真菌对宿主植物生长的改变可能与其产生吲哚乙酸有关(de Battista et al ., 1990 ), 这也可能与植物中内生真菌的传播方式有关(Tintjer et al ., 2008 ). ...
The importance of water-soluble carbohydrate reserves on regrowth and root growth of Lolium perenne (L.)
1
1997
... 碳水化合物是植物中碳库的储存(Donaghy & Fulkerson, 1997 , 1998 ; Cheplick & Chui, 2001 ).已有研究表明内生真菌可以增加多年生黑麦草中碳水化合物的含量(Cheplick & Cho, 2003 ; Hunt et al ., 2005 ; Rasmussen et al ., 2007 ).本研究中, 水平传播的内生真菌促使宿主具有更高的碳含量, 但垂直传播的内生真菌使宿主体内具有更高的碳氮比.可溶性糖是一种重要的渗透调节物质, 在干旱胁迫过程中, 植物体内可溶性糖含量的变化在一定程度上能反映其对不良环境的适应能力(Sánchez et al ., 1998 ).我们还发现感染垂直传播的内生真菌的羽茅比不染菌植株和感染有性型内生真菌的植株中积累更多的可溶性糖.本研究中垂直传播的内生真菌使宿主中储存更多的可溶性糖, 这对植物是有益处的, 而水平传播的内生真菌并没有这一表现, 并且垂直传播的内生真菌使宿主产生的总酚含量高于水平传播的内生真菌, 由此我们推测, 当感染不同传播方式的内生真菌的宿主受到生物或非生物胁迫时, 通过垂直传播方式感染内生真菌的植株防御能力很可能强于通过水平传播方式感染内生真菌的植株, 这还有待进一步的实验研究. ...
Priority for allocation of water-soluble carbohydrate reserves during regrowth of Lolium perenne
1
1998
... 碳水化合物是植物中碳库的储存(Donaghy & Fulkerson, 1997 , 1998 ; Cheplick & Chui, 2001 ).已有研究表明内生真菌可以增加多年生黑麦草中碳水化合物的含量(Cheplick & Cho, 2003 ; Hunt et al ., 2005 ; Rasmussen et al ., 2007 ).本研究中, 水平传播的内生真菌促使宿主具有更高的碳含量, 但垂直传播的内生真菌使宿主体内具有更高的碳氮比.可溶性糖是一种重要的渗透调节物质, 在干旱胁迫过程中, 植物体内可溶性糖含量的变化在一定程度上能反映其对不良环境的适应能力(Sánchez et al ., 1998 ).我们还发现感染垂直传播的内生真菌的羽茅比不染菌植株和感染有性型内生真菌的植株中积累更多的可溶性糖.本研究中垂直传播的内生真菌使宿主中储存更多的可溶性糖, 这对植物是有益处的, 而水平传播的内生真菌并没有这一表现, 并且垂直传播的内生真菌使宿主产生的总酚含量高于水平传播的内生真菌, 由此我们推测, 当感染不同传播方式的内生真菌的宿主受到生物或非生物胁迫时, 通过垂直传播方式感染内生真菌的植株防御能力很可能强于通过水平传播方式感染内生真菌的植株, 这还有待进一步的实验研究. ...
Endophyte infection effects on stomatal conductance, osmotic adjustment and drought recovery of tall fescue
1
1995
... 对高羊茅和黑麦草的研究发现, Neotyphodium 能提高宿主的生长(Marks et al ., 1991 )、耐旱性(Elmi & West, 1995 )、对养分的吸收和利用(Malinowski & Belesky, 2000 ), 以及宿主对动物的拒食能力(Clay, 1990 ).然而, 很多研究发现天然禾草的情况与栽培禾草的不一致甚至相反, 且Epichloë 与天然禾草共生对宿主的有益影响可能小于Neotyphodium, 原因可能是Epichloë 能在宿主部分或全部生殖枝上产生子座, 抑制宿主的开花和结实(Leuchtmann et al ., 1994 ); 也可能是多数Epichloë 不产生生物碱或生物碱的产生水平很低(Leuchtmann et al ., 2000 ).大部分研究都集中在Neotyphodium 对宿主的影响, 以及影响Epichloë 产子座的条件.关于Epichloë 对宿主的生理生态特性影响研究较少.有报道指出: 由于Epichloë typhina 在宿主细弱剪股颖(Agrostis tenuis )叶鞘上产生子座抑制宿主抽穗、开花, 从而增强了宿主地上、地下营养生长, 宿主分蘖数显著增多(de Battista et al ., 1990 ).在本研究中, 通过对感染不同传播方式的羽茅植株进行叶鞘分离, 我们得到Neotyphodium 和Epichloë 内生真菌, 从内生真菌的传播方式入手, 通过田间栽培试验, 探讨不同传播方式的Neotyphodium 和Epichloë 属内生真菌对羽茅生长与生理特性的影响. ...
Interaction between the endophytic fungus Epichloe bromicola and the grass Bromus erectus: Effects of endophyte infection, fungal concentration and environment on grass growth and flowering
2
1999
... Bradshaw和Snaydon (1959) 最早报道内生真菌可以影响宿主植物的生长.但关于不同传播方式的内生真菌, 尤其是水平传播方式的内生真菌对宿主植物的影响相对较少, 大部分研究集中在垂直传播的内生真菌Neotyphodium 对宿主植物的影响.有研究认为Neotyphodium 与宿主植物在长期的共同进化过程中形成了一种互利共生的关系, 可促进植物生长和分蘖形成, 增强植物抗病害、抗动物采食及抗恶劣环境等能力(Moon et al ., 2004 ).已有研究表明: 通过和宿主植物及其生长环境的相互作用, Epichloë 在宿主植物旗叶的叶鞘或茎秆上形成子座, 未成熟的子座由一层致密的菌丝层组成, 表面密布分生孢子, 但关于这些真菌影响植物的生长和生理指标的研究较少.本研究不仅从生有子座的羽茅中分离出有性型内生真菌, 还得出垂直传播的内生真菌显著促进了植物的叶片长宽比, 而水平传播的内生真菌对羽茅的株高和叶宽有显著的负效应.Groppe等(1999) 也得出类似的结果, 认为植物生长能力较强有助于植物中内生真菌的传播.垂直传播的内生真菌对宿主植物生长的改变可能与其产生吲哚乙酸有关(de Battista et al ., 1990 ), 这也可能与植物中内生真菌的传播方式有关(Tintjer et al ., 2008 ). ...
... 碳水化合物是植物中碳库的储存(Donaghy & Fulkerson, 1997 , 1998 ; Cheplick & Chui, 2001 ).已有研究表明内生真菌可以增加多年生黑麦草中碳水化合物的含量(Cheplick & Cho, 2003 ; Hunt et al ., 2005 ; Rasmussen et al ., 2007 ).本研究中, 水平传播的内生真菌促使宿主具有更高的碳含量, 但垂直传播的内生真菌使宿主体内具有更高的碳氮比.可溶性糖是一种重要的渗透调节物质, 在干旱胁迫过程中, 植物体内可溶性糖含量的变化在一定程度上能反映其对不良环境的适应能力(Sánchez et al ., 1998 ).我们还发现感染垂直传播的内生真菌的羽茅比不染菌植株和感染有性型内生真菌的植株中积累更多的可溶性糖.本研究中垂直传播的内生真菌使宿主中储存更多的可溶性糖, 这对植物是有益处的, 而水平传播的内生真菌并没有这一表现, 并且垂直传播的内生真菌使宿主产生的总酚含量高于水平传播的内生真菌, 由此我们推测, 当感染不同传播方式的内生真菌的宿主受到生物或非生物胁迫时, 通过垂直传播方式感染内生真菌的植株防御能力很可能强于通过水平传播方式感染内生真菌的植株, 这还有待进一步的实验研究. ...
Near-term impacts of elevated CO2 , nitrogen and fungal endophyte-infection on Lolium perenne L. growth, chemical composition and alkaloid production
0
2005
分离自苇状羊茅(Festucaa rundinacea Schreb.)的内生真菌Neotyphodium uncinatum
1
2003
... 中国21个地区披碱草(Elymus dahuricus )的染菌率调查表明, 有6个地区的披碱草种子中Neotypho- dium 的感染率在4.4%-100.0%, 绝大多数染菌的披碱草比不染菌的多产生一种生物碱——波胺, 波胺的浓度水平随着种群的位置和季节而变化(Zhang & Nan, 2007 ).在南京发现了Neotyphodium uncinatum 与苇状羊茅(Festuca arundinacea )的共生体(纪燕玲等, 2003 ).还有研究发现, Epichloë yangzii 是共生于鹅观草属(Roegneria )植物的产子座有性型内生真菌(申靖等, 2009 ).Wei等(2007) 对中国北方地区172个禾草地理种群的内生真菌感染状况进行广泛调查后发现, 在所有检测的41种禾草中, 25种禾草感染了内生真菌, 其中羽茅(Achnatherum sibiricum )的感染率最高, 这表明内生真菌广泛存在于天然禾草种群中, 而且内生真菌与禾草的相互作用可能对中国北方草原生态系统和畜牧业产生影响. ...
分离自苇状羊茅(Festucaa rundinacea Schreb.)的内生真菌Neotyphodium uncinatum
1
2003
... 中国21个地区披碱草(Elymus dahuricus )的染菌率调查表明, 有6个地区的披碱草种子中Neotypho- dium 的感染率在4.4%-100.0%, 绝大多数染菌的披碱草比不染菌的多产生一种生物碱——波胺, 波胺的浓度水平随着种群的位置和季节而变化(Zhang & Nan, 2007 ).在南京发现了Neotyphodium uncinatum 与苇状羊茅(Festuca arundinacea )的共生体(纪燕玲等, 2003 ).还有研究发现, Epichloë yangzii 是共生于鹅观草属(Roegneria )植物的产子座有性型内生真菌(申靖等, 2009 ).Wei等(2007) 对中国北方地区172个禾草地理种群的内生真菌感染状况进行广泛调查后发现, 在所有检测的41种禾草中, 25种禾草感染了内生真菌, 其中羽茅(Achnatherum sibiricum )的感染率最高, 这表明内生真菌广泛存在于天然禾草种群中, 而且内生真菌与禾草的相互作用可能对中国北方草原生态系统和畜牧业产生影响. ...
Trade-off between virulence and vertical transmission and the maintenance of a virulent plant pathogen
2
1998
... 对高羊茅和黑麦草的研究发现, Neotyphodium 能提高宿主的生长(Marks et al ., 1991 )、耐旱性(Elmi & West, 1995 )、对养分的吸收和利用(Malinowski & Belesky, 2000 ), 以及宿主对动物的拒食能力(Clay, 1990 ).然而, 很多研究发现天然禾草的情况与栽培禾草的不一致甚至相反, 且Epichloë 与天然禾草共生对宿主的有益影响可能小于Neotyphodium, 原因可能是Epichloë 能在宿主部分或全部生殖枝上产生子座, 抑制宿主的开花和结实(Leuchtmann et al ., 1994 ); 也可能是多数Epichloë 不产生生物碱或生物碱的产生水平很低(Leuchtmann et al ., 2000 ).大部分研究都集中在Neotyphodium 对宿主的影响, 以及影响Epichloë 产子座的条件.关于Epichloë 对宿主的生理生态特性影响研究较少.有报道指出: 由于Epichloë typhina 在宿主细弱剪股颖(Agrostis tenuis )叶鞘上产生子座抑制宿主抽穗、开花, 从而增强了宿主地上、地下营养生长, 宿主分蘖数显著增多(de Battista et al ., 1990 ).在本研究中, 通过对感染不同传播方式的羽茅植株进行叶鞘分离, 我们得到Neotyphodium 和Epichloë 内生真菌, 从内生真菌的传播方式入手, 通过田间栽培试验, 探讨不同传播方式的Neotyphodium 和Epichloë 属内生真菌对羽茅生长与生理特性的影响. ...
... 宿主种群间不同的传播方式也可能是由于内生真菌或者宿主植物的基因型不同(Bucheli & Leuchtmann, 1996 ; Kover & Clay, 1998 ).不同传播方式的内生真菌对羽茅生长与生理特性的影响不同, 还可能是由于大部分Epichloë 并不是每年都能在宿主植物上产生子座, 子座的产生与否, 不仅与内生真菌种类以及宿主植物的基因型有关, 而且与植物的生长环境、营养状态等有关, 这还有待于进一步研究. ...
Sexual compatibility and taxonomy of a new species of Epichloe symbiotic with fine Fescue grasses
1
1994
... 对高羊茅和黑麦草的研究发现, Neotyphodium 能提高宿主的生长(Marks et al ., 1991 )、耐旱性(Elmi & West, 1995 )、对养分的吸收和利用(Malinowski & Belesky, 2000 ), 以及宿主对动物的拒食能力(Clay, 1990 ).然而, 很多研究发现天然禾草的情况与栽培禾草的不一致甚至相反, 且Epichloë 与天然禾草共生对宿主的有益影响可能小于Neotyphodium, 原因可能是Epichloë 能在宿主部分或全部生殖枝上产生子座, 抑制宿主的开花和结实(Leuchtmann et al ., 1994 ); 也可能是多数Epichloë 不产生生物碱或生物碱的产生水平很低(Leuchtmann et al ., 2000 ).大部分研究都集中在Neotyphodium 对宿主的影响, 以及影响Epichloë 产子座的条件.关于Epichloë 对宿主的生理生态特性影响研究较少.有报道指出: 由于Epichloë typhina 在宿主细弱剪股颖(Agrostis tenuis )叶鞘上产生子座抑制宿主抽穗、开花, 从而增强了宿主地上、地下营养生长, 宿主分蘖数显著增多(de Battista et al ., 1990 ).在本研究中, 通过对感染不同传播方式的羽茅植株进行叶鞘分离, 我们得到Neotyphodium 和Epichloë 内生真菌, 从内生真菌的传播方式入手, 通过田间栽培试验, 探讨不同传播方式的Neotyphodium 和Epichloë 属内生真菌对羽茅生长与生理特性的影响. ...
Different levels of protective alkaloids in grasses with stroma-forming and seed-transmitted Epichloë/Neotyphodium endophytes
0
2000
高温处理构建不感染内生真菌羽茅种群的方法探讨
1
2010
... 2009年4月, 在南开大学的网室样地中, 在8块1 m × 1 m的样方中播撒采自中国农业科学院呼伦贝尔草原生态系统实验站的饱满羽茅种子, 每块样方间距50 cm, 为了保证垂直传播的内生真菌Neotyphodium 和水平传播的内生真菌Epichloë 不感染到不染菌的样方, 我们每年在羽茅抽穗结种之前, 将刚抽穗但还未成熟的羽茅的穗剪掉, 防止羽茅种子成熟后感染到其他不染菌样方, 以确保在第二年的实验中不染菌样方中的羽茅植株仍是未受内生真菌感染的羽茅.随机分布选择4块样方播撒染菌种子, 相互间隔开的另外4块播撒不染菌种子, 每个样方播撒100-120粒.不染菌种子通过60 ℃高温处理后获得, 处理时间为30天.所有检测幼苗均不感染内生真菌.高温处理对羽茅的种子发芽率、发芽势和发芽指数均无显著影响(李夏等, 2010 ).2011年5月, 发现2块染菌样方中, 羽茅植株的生殖枝上产生子座, 另外2块染菌的样方中, 染菌羽茅植株并没有形成子座.我们分别于2011年和2012年测定标记好长子座的植株的生理生态特性, 测定指标前, 通过苯胺兰染色法在显微镜下检测植株的染菌状况, 并且在产生子座的植株中, 用叶鞘分离内生真菌的方法, 分离和纯化出与羽茅共生的通过水平传播的Epichloë, 从未产生子座的植株中分离得到垂直传播的Neotyphodium, 分离纯化得到的两个属内生真菌形态不同.我们将不同传播方式的羽茅分别标记为: Ne——感染Neotyphodium 的植株; Ep——感染Epichloë 的植株; EF——不染菌植株.EF、Ne、Ep所在样方土壤成分分别为: N%分别是0.033%、0.057%和0.097%; C%分别是1.19%、1.507%和2.16%.每块样方中选取3-5个重复进行羽茅生理生态指标的测定. ...
高温处理构建不感染内生真菌羽茅种群的方法探讨
1
2010
... 2009年4月, 在南开大学的网室样地中, 在8块1 m × 1 m的样方中播撒采自中国农业科学院呼伦贝尔草原生态系统实验站的饱满羽茅种子, 每块样方间距50 cm, 为了保证垂直传播的内生真菌Neotyphodium 和水平传播的内生真菌Epichloë 不感染到不染菌的样方, 我们每年在羽茅抽穗结种之前, 将刚抽穗但还未成熟的羽茅的穗剪掉, 防止羽茅种子成熟后感染到其他不染菌样方, 以确保在第二年的实验中不染菌样方中的羽茅植株仍是未受内生真菌感染的羽茅.随机分布选择4块样方播撒染菌种子, 相互间隔开的另外4块播撒不染菌种子, 每个样方播撒100-120粒.不染菌种子通过60 ℃高温处理后获得, 处理时间为30天.所有检测幼苗均不感染内生真菌.高温处理对羽茅的种子发芽率、发芽势和发芽指数均无显著影响(李夏等, 2010 ).2011年5月, 发现2块染菌样方中, 羽茅植株的生殖枝上产生子座, 另外2块染菌的样方中, 染菌羽茅植株并没有形成子座.我们分别于2011年和2012年测定标记好长子座的植株的生理生态特性, 测定指标前, 通过苯胺兰染色法在显微镜下检测植株的染菌状况, 并且在产生子座的植株中, 用叶鞘分离内生真菌的方法, 分离和纯化出与羽茅共生的通过水平传播的Epichloë, 从未产生子座的植株中分离得到垂直传播的Neotyphodium, 分离纯化得到的两个属内生真菌形态不同.我们将不同传播方式的羽茅分别标记为: Ne——感染Neotyphodium 的植株; Ep——感染Epichloë 的植株; EF——不染菌植株.EF、Ne、Ep所在样方土壤成分分别为: N%分别是0.033%、0.057%和0.097%; C%分别是1.19%、1.507%和2.16%.每块样方中选取3-5个重复进行羽茅生理生态指标的测定. ...
Evidence for chemical changes on the root surface of tall fescue in response to infection with the fungal endophyte Neotyphodium coenophialum
1
1998
... 测定光合色素含量时采用乙醇-丙酮混合液法(无水乙醇:丙酮=1:1)浸泡提取色素, 分别测定浸泡液在470 nm、646 nm和663 nm处的吸光值.用测定的吸光度值计算羽茅叶片的叶绿素a、叶绿素b及类胡萝卜素的含量.叶片全氮含量的测定采用凯氏定氮法.有机质含量测定采用重铬酸钾氧化法, 指示剂采用邻菲罗啉, 用0.2 mol·L-1 FeSO4 溶液进行滴定.可溶性糖含量采用蒽酮比色法进行测定.总酚含量的测定参照Malinowski等(1998) 的方法, 采用无水乙醇:水(1:1)混合液浸提总酚, FeCl3 (0.9%)与铁氰化钾(0.6%)处理后, 在720 nm处测定吸光度得到总酚含量.非结构性碳水化合物(TNC, %)参照酶解法进行测定(da Silveira et al ., 1978 ). ...
Adaptations of endophyte-infected cool-season grasses to environmental stresses: Mechanisms of drought and mineral stress tolerance
2
2000
... 对高羊茅和黑麦草的研究发现, Neotyphodium 能提高宿主的生长(Marks et al ., 1991 )、耐旱性(Elmi & West, 1995 )、对养分的吸收和利用(Malinowski & Belesky, 2000 ), 以及宿主对动物的拒食能力(Clay, 1990 ).然而, 很多研究发现天然禾草的情况与栽培禾草的不一致甚至相反, 且Epichloë 与天然禾草共生对宿主的有益影响可能小于Neotyphodium, 原因可能是Epichloë 能在宿主部分或全部生殖枝上产生子座, 抑制宿主的开花和结实(Leuchtmann et al ., 1994 ); 也可能是多数Epichloë 不产生生物碱或生物碱的产生水平很低(Leuchtmann et al ., 2000 ).大部分研究都集中在Neotyphodium 对宿主的影响, 以及影响Epichloë 产子座的条件.关于Epichloë 对宿主的生理生态特性影响研究较少.有报道指出: 由于Epichloë typhina 在宿主细弱剪股颖(Agrostis tenuis )叶鞘上产生子座抑制宿主抽穗、开花, 从而增强了宿主地上、地下营养生长, 宿主分蘖数显著增多(de Battista et al ., 1990 ).在本研究中, 通过对感染不同传播方式的羽茅植株进行叶鞘分离, 我们得到Neotyphodium 和Epichloë 内生真菌, 从内生真菌的传播方式入手, 通过田间栽培试验, 探讨不同传播方式的Neotyphodium 和Epichloë 属内生真菌对羽茅生长与生理特性的影响. ...
... )、对养分的吸收和利用(Malinowski & Belesky, 2000 ), 以及宿主对动物的拒食能力(Clay, 1990 ).然而, 很多研究发现天然禾草的情况与栽培禾草的不一致甚至相反, 且Epichloë 与天然禾草共生对宿主的有益影响可能小于Neotyphodium, 原因可能是Epichloë 能在宿主部分或全部生殖枝上产生子座, 抑制宿主的开花和结实(Leuchtmann et al ., 1994 ); 也可能是多数Epichloë 不产生生物碱或生物碱的产生水平很低(Leuchtmann et al ., 2000 ).大部分研究都集中在Neotyphodium 对宿主的影响, 以及影响Epichloë 产子座的条件.关于Epichloë 对宿主的生理生态特性影响研究较少.有报道指出: 由于Epichloë typhina 在宿主细弱剪股颖(Agrostis tenuis )叶鞘上产生子座抑制宿主抽穗、开花, 从而增强了宿主地上、地下营养生长, 宿主分蘖数显著增多(de Battista et al ., 1990 ).在本研究中, 通过对感染不同传播方式的羽茅植株进行叶鞘分离, 我们得到Neotyphodium 和Epichloë 内生真菌, 从内生真菌的传播方式入手, 通过田间栽培试验, 探讨不同传播方式的Neotyphodium 和Epichloë 属内生真菌对羽茅生长与生理特性的影响. ...
Effects of fungal endophytes on interspecific and intraspecific competition in the grasses Festuca arundinacea and Lolium perenne
2
1991
... 子囊菌纲(Ascomycetes)麦角菌科(Clavicipitac- eae)的无性型Neotyphodium 属及有性型Epichloë 属真菌, 普遍分布于冷季型的禾本科植物中, 在植物体内完成其部分或全部生活史(Schardl et al ., 2004 ).Neotyphodium 属内生真菌与宿主植物的关系具有代表性的研究意义, 通过垂直方式传播, 在高羊茅(Festuca elata )、多年生黑麦草(Lolium perenne )、Achnatherum robustum 和羽茅(Achnatherum sibiricum )上都存在有这类内生真菌(Wei et al ., 2007 ).Neotyphodium 内生真菌的全部生活周期都在植物体内完成, 菌丝在宿主禾草开花期进入子房, 植物产生种子后, 内生真菌便存在于种子中, 从而可以通过种子进行传播, 其间不形成任何子座或孢子.目前所研究的禾草植物中的内生真菌大部分属于此类传播方式.Epichloë 属真菌能够感染7个族16个属以上的禾本科植物, 在世界范围内分布广泛(Moon et al ., 2004 ; Schardl et al ., 2004 ). Epichloë 真菌通过水平方式传播, 偶尔能在植物的茎秆和花序上形成特征性的子座, 内生真菌形成有性孢子, 通过虫媒作用, 与其他植株上配型相反的有性孢子结合, 并产生子囊孢子, 侵染新的个体(Bultman & Leuchtm- ann, 2003 ), 因此宿主的有性繁殖被完全抑制.Leuchtmann和Clay发现大多数感染Epichloë typhina 的披碱草属(Elymus )植株的同一个个体上, 往往同时存在带有子座和产生种子的两类生殖枝, 内生真菌只在产生子座的生殖枝营有性生殖, 进行水平传播, 而内生真菌在多数产生种子的生殖枝随着宿主的无性繁殖进行垂直传播, 这种既存在水平传播又有垂直传播的方式称为混合传播方式(White, 1988 ). ...
... Bradshaw和Snaydon (1959) 最早报道内生真菌可以影响宿主植物的生长.但关于不同传播方式的内生真菌, 尤其是水平传播方式的内生真菌对宿主植物的影响相对较少, 大部分研究集中在垂直传播的内生真菌Neotyphodium 对宿主植物的影响.有研究认为Neotyphodium 与宿主植物在长期的共同进化过程中形成了一种互利共生的关系, 可促进植物生长和分蘖形成, 增强植物抗病害、抗动物采食及抗恶劣环境等能力(Moon et al ., 2004 ).已有研究表明: 通过和宿主植物及其生长环境的相互作用, Epichloë 在宿主植物旗叶的叶鞘或茎秆上形成子座, 未成熟的子座由一层致密的菌丝层组成, 表面密布分生孢子, 但关于这些真菌影响植物的生长和生理指标的研究较少.本研究不仅从生有子座的羽茅中分离出有性型内生真菌, 还得出垂直传播的内生真菌显著促进了植物的叶片长宽比, 而水平传播的内生真菌对羽茅的株高和叶宽有显著的负效应.Groppe等(1999) 也得出类似的结果, 认为植物生长能力较强有助于植物中内生真菌的传播.垂直传播的内生真菌对宿主植物生长的改变可能与其产生吲哚乙酸有关(de Battista et al ., 1990 ), 这也可能与植物中内生真菌的传播方式有关(Tintjer et al ., 2008 ). ...
Prevalence of interspecific hybrids amongst asexual fungal endophytes of grasses
0
2004
Effect of Neotyphodium endophyte infection on growth and leaf gas exchange of Arizona fescue under contrasting water availability regimes
2
2002
... 对于感染不同传播方式内生真菌的羽茅而言, 染菌植株的蒸腾速率、气孔导度和光能利用效率都显著高于不染菌植株.这表明内生真菌对植株的光合作用有一定的促进作用.这可能对宿主植物的传播方式也有一定的影响(Cheplick & Faeth, 2009 ).我们发现无性型内生真菌可以提高羽茅的气孔限制值和水分利用效率, 但有性型内生真菌的作用却复杂多变.Morse等(2002) 研究发现, 不染菌的亚利桑那羊茅(Festuca arizonica )表现出更高的光合速率和气孔导度.可见, 羽茅-内生真菌之间的共生关系在光合特性方面不仅与羽茅的染菌状况有关, 也与感染内生真菌的传播方式有一定的关系. ...
... 碳水化合物是植物中碳库的储存(Donaghy & Fulkerson, 1997 , 1998 ; Cheplick & Chui, 2001 ).已有研究表明内生真菌可以增加多年生黑麦草中碳水化合物的含量(Cheplick & Cho, 2003 ; Hunt et al ., 2005 ; Rasmussen et al ., 2007 ).本研究中, 水平传播的内生真菌促使宿主具有更高的碳含量, 但垂直传播的内生真菌使宿主体内具有更高的碳氮比.可溶性糖是一种重要的渗透调节物质, 在干旱胁迫过程中, 植物体内可溶性糖含量的变化在一定程度上能反映其对不良环境的适应能力(Sánchez et al ., 1998 ).我们还发现感染垂直传播的内生真菌的羽茅比不染菌植株和感染有性型内生真菌的植株中积累更多的可溶性糖.本研究中垂直传播的内生真菌使宿主中储存更多的可溶性糖, 这对植物是有益处的, 而水平传播的内生真菌并没有这一表现, 并且垂直传播的内生真菌使宿主产生的总酚含量高于水平传播的内生真菌, 由此我们推测, 当感染不同传播方式的内生真菌的宿主受到生物或非生物胁迫时, 通过垂直传播方式感染内生真菌的植株防御能力很可能强于通过水平传播方式感染内生真菌的植株, 这还有待进一步的实验研究. ...
High nitrogen supply and carbohydrate content reduce fungal endophyte and alkaloid concentration in Lolium perenne
0
2007
Turgor maintenance, osmotic adjustment and soluble sugar and proline accumulation in 49 pea cultivars in response to water stress
2
1998
... 子囊菌纲(Ascomycetes)麦角菌科(Clavicipitac- eae)的无性型Neotyphodium 属及有性型Epichloë 属真菌, 普遍分布于冷季型的禾本科植物中, 在植物体内完成其部分或全部生活史(Schardl et al ., 2004 ).Neotyphodium 属内生真菌与宿主植物的关系具有代表性的研究意义, 通过垂直方式传播, 在高羊茅(Festuca elata )、多年生黑麦草(Lolium perenne )、Achnatherum robustum 和羽茅(Achnatherum sibiricum )上都存在有这类内生真菌(Wei et al ., 2007 ).Neotyphodium 内生真菌的全部生活周期都在植物体内完成, 菌丝在宿主禾草开花期进入子房, 植物产生种子后, 内生真菌便存在于种子中, 从而可以通过种子进行传播, 其间不形成任何子座或孢子.目前所研究的禾草植物中的内生真菌大部分属于此类传播方式.Epichloë 属真菌能够感染7个族16个属以上的禾本科植物, 在世界范围内分布广泛(Moon et al ., 2004 ; Schardl et al ., 2004 ). Epichloë 真菌通过水平方式传播, 偶尔能在植物的茎秆和花序上形成特征性的子座, 内生真菌形成有性孢子, 通过虫媒作用, 与其他植株上配型相反的有性孢子结合, 并产生子囊孢子, 侵染新的个体(Bultman & Leuchtm- ann, 2003 ), 因此宿主的有性繁殖被完全抑制.Leuchtmann和Clay发现大多数感染Epichloë typhina 的披碱草属(Elymus )植株的同一个个体上, 往往同时存在带有子座和产生种子的两类生殖枝, 内生真菌只在产生子座的生殖枝营有性生殖, 进行水平传播, 而内生真菌在多数产生种子的生殖枝随着宿主的无性繁殖进行垂直传播, 这种既存在水平传播又有垂直传播的方式称为混合传播方式(White, 1988 ). ...
... 碳水化合物是植物中碳库的储存(Donaghy & Fulkerson, 1997 , 1998 ; Cheplick & Chui, 2001 ).已有研究表明内生真菌可以增加多年生黑麦草中碳水化合物的含量(Cheplick & Cho, 2003 ; Hunt et al ., 2005 ; Rasmussen et al ., 2007 ).本研究中, 水平传播的内生真菌促使宿主具有更高的碳含量, 但垂直传播的内生真菌使宿主体内具有更高的碳氮比.可溶性糖是一种重要的渗透调节物质, 在干旱胁迫过程中, 植物体内可溶性糖含量的变化在一定程度上能反映其对不良环境的适应能力(Sánchez et al ., 1998 ).我们还发现感染垂直传播的内生真菌的羽茅比不染菌植株和感染有性型内生真菌的植株中积累更多的可溶性糖.本研究中垂直传播的内生真菌使宿主中储存更多的可溶性糖, 这对植物是有益处的, 而水平传播的内生真菌并没有这一表现, 并且垂直传播的内生真菌使宿主产生的总酚含量高于水平传播的内生真菌, 由此我们推测, 当感染不同传播方式的内生真菌的宿主受到生物或非生物胁迫时, 通过垂直传播方式感染内生真菌的植株防御能力很可能强于通过水平传播方式感染内生真菌的植株, 这还有待进一步的实验研究. ...
Symbioses of grasses with seedborne fungal endophytes
1
2004
... 子囊菌纲(Ascomycetes)麦角菌科(Clavicipitac- eae)的无性型Neotyphodium 属及有性型Epichloë 属真菌, 普遍分布于冷季型的禾本科植物中, 在植物体内完成其部分或全部生活史(Schardl et al ., 2004 ).Neotyphodium 属内生真菌与宿主植物的关系具有代表性的研究意义, 通过垂直方式传播, 在高羊茅(Festuca elata )、多年生黑麦草(Lolium perenne )、Achnatherum robustum 和羽茅(Achnatherum sibiricum )上都存在有这类内生真菌(Wei et al ., 2007 ).Neotyphodium 内生真菌的全部生活周期都在植物体内完成, 菌丝在宿主禾草开花期进入子房, 植物产生种子后, 内生真菌便存在于种子中, 从而可以通过种子进行传播, 其间不形成任何子座或孢子.目前所研究的禾草植物中的内生真菌大部分属于此类传播方式.Epichloë 属真菌能够感染7个族16个属以上的禾本科植物, 在世界范围内分布广泛(Moon et al ., 2004 ; Schardl et al ., 2004 ). Epichloë 真菌通过水平方式传播, 偶尔能在植物的茎秆和花序上形成特征性的子座, 内生真菌形成有性孢子, 通过虫媒作用, 与其他植株上配型相反的有性孢子结合, 并产生子囊孢子, 侵染新的个体(Bultman & Leuchtm- ann, 2003 ), 因此宿主的有性繁殖被完全抑制.Leuchtmann和Clay发现大多数感染Epichloë typhina 的披碱草属(Elymus )植株的同一个个体上, 往往同时存在带有子座和产生种子的两类生殖枝, 内生真菌只在产生子座的生殖枝营有性生殖, 进行水平传播, 而内生真菌在多数产生种子的生殖枝随着宿主的无性繁殖进行垂直传播, 这种既存在水平传播又有垂直传播的方式称为混合传播方式(White, 1988 ). ...
禾本科植物内生真菌研究9——Epichloё yangzii的种传特性及其在宿主体内的分布
2
2009
... 中国21个地区披碱草(Elymus dahuricus )的染菌率调查表明, 有6个地区的披碱草种子中Neotypho- dium 的感染率在4.4%-100.0%, 绝大多数染菌的披碱草比不染菌的多产生一种生物碱——波胺, 波胺的浓度水平随着种群的位置和季节而变化(Zhang & Nan, 2007 ).在南京发现了Neotyphodium uncinatum 与苇状羊茅(Festuca arundinacea )的共生体(纪燕玲等, 2003 ).还有研究发现, Epichloë yangzii 是共生于鹅观草属(Roegneria )植物的产子座有性型内生真菌(申靖等, 2009 ).Wei等(2007) 对中国北方地区172个禾草地理种群的内生真菌感染状况进行广泛调查后发现, 在所有检测的41种禾草中, 25种禾草感染了内生真菌, 其中羽茅(Achnatherum sibiricum )的感染率最高, 这表明内生真菌广泛存在于天然禾草种群中, 而且内生真菌与禾草的相互作用可能对中国北方草原生态系统和畜牧业产生影响. ...
... Bradshaw和Snaydon (1959) 最早报道内生真菌可以影响宿主植物的生长.但关于不同传播方式的内生真菌, 尤其是水平传播方式的内生真菌对宿主植物的影响相对较少, 大部分研究集中在垂直传播的内生真菌Neotyphodium 对宿主植物的影响.有研究认为Neotyphodium 与宿主植物在长期的共同进化过程中形成了一种互利共生的关系, 可促进植物生长和分蘖形成, 增强植物抗病害、抗动物采食及抗恶劣环境等能力(Moon et al ., 2004 ).已有研究表明: 通过和宿主植物及其生长环境的相互作用, Epichloë 在宿主植物旗叶的叶鞘或茎秆上形成子座, 未成熟的子座由一层致密的菌丝层组成, 表面密布分生孢子, 但关于这些真菌影响植物的生长和生理指标的研究较少.本研究不仅从生有子座的羽茅中分离出有性型内生真菌, 还得出垂直传播的内生真菌显著促进了植物的叶片长宽比, 而水平传播的内生真菌对羽茅的株高和叶宽有显著的负效应.Groppe等(1999) 也得出类似的结果, 认为植物生长能力较强有助于植物中内生真菌的传播.垂直传播的内生真菌对宿主植物生长的改变可能与其产生吲哚乙酸有关(de Battista et al ., 1990 ), 这也可能与植物中内生真菌的传播方式有关(Tintjer et al ., 2008 ). ...
禾本科植物内生真菌研究9——Epichloё yangzii的种传特性及其在宿主体内的分布
2
2009
... 中国21个地区披碱草(Elymus dahuricus )的染菌率调查表明, 有6个地区的披碱草种子中Neotypho- dium 的感染率在4.4%-100.0%, 绝大多数染菌的披碱草比不染菌的多产生一种生物碱——波胺, 波胺的浓度水平随着种群的位置和季节而变化(Zhang & Nan, 2007 ).在南京发现了Neotyphodium uncinatum 与苇状羊茅(Festuca arundinacea )的共生体(纪燕玲等, 2003 ).还有研究发现, Epichloë yangzii 是共生于鹅观草属(Roegneria )植物的产子座有性型内生真菌(申靖等, 2009 ).Wei等(2007) 对中国北方地区172个禾草地理种群的内生真菌感染状况进行广泛调查后发现, 在所有检测的41种禾草中, 25种禾草感染了内生真菌, 其中羽茅(Achnatherum sibiricum )的感染率最高, 这表明内生真菌广泛存在于天然禾草种群中, 而且内生真菌与禾草的相互作用可能对中国北方草原生态系统和畜牧业产生影响. ...
... Bradshaw和Snaydon (1959) 最早报道内生真菌可以影响宿主植物的生长.但关于不同传播方式的内生真菌, 尤其是水平传播方式的内生真菌对宿主植物的影响相对较少, 大部分研究集中在垂直传播的内生真菌Neotyphodium 对宿主植物的影响.有研究认为Neotyphodium 与宿主植物在长期的共同进化过程中形成了一种互利共生的关系, 可促进植物生长和分蘖形成, 增强植物抗病害、抗动物采食及抗恶劣环境等能力(Moon et al ., 2004 ).已有研究表明: 通过和宿主植物及其生长环境的相互作用, Epichloë 在宿主植物旗叶的叶鞘或茎秆上形成子座, 未成熟的子座由一层致密的菌丝层组成, 表面密布分生孢子, 但关于这些真菌影响植物的生长和生理指标的研究较少.本研究不仅从生有子座的羽茅中分离出有性型内生真菌, 还得出垂直传播的内生真菌显著促进了植物的叶片长宽比, 而水平传播的内生真菌对羽茅的株高和叶宽有显著的负效应.Groppe等(1999) 也得出类似的结果, 认为植物生长能力较强有助于植物中内生真菌的传播.垂直传播的内生真菌对宿主植物生长的改变可能与其产生吲哚乙酸有关(de Battista et al ., 1990 ), 这也可能与植物中内生真菌的传播方式有关(Tintjer et al ., 2008 ). ...
Variation in horizontal and vertical transmission of the endophyte Epichloë elymi infecting the grass Elymus hystrix
1
2008
... 子囊菌纲(Ascomycetes)麦角菌科(Clavicipitac- eae)的无性型Neotyphodium 属及有性型Epichloë 属真菌, 普遍分布于冷季型的禾本科植物中, 在植物体内完成其部分或全部生活史(Schardl et al ., 2004 ).Neotyphodium 属内生真菌与宿主植物的关系具有代表性的研究意义, 通过垂直方式传播, 在高羊茅(Festuca elata )、多年生黑麦草(Lolium perenne )、Achnatherum robustum 和羽茅(Achnatherum sibiricum )上都存在有这类内生真菌(Wei et al ., 2007 ).Neotyphodium 内生真菌的全部生活周期都在植物体内完成, 菌丝在宿主禾草开花期进入子房, 植物产生种子后, 内生真菌便存在于种子中, 从而可以通过种子进行传播, 其间不形成任何子座或孢子.目前所研究的禾草植物中的内生真菌大部分属于此类传播方式.Epichloë 属真菌能够感染7个族16个属以上的禾本科植物, 在世界范围内分布广泛(Moon et al ., 2004 ; Schardl et al ., 2004 ). Epichloë 真菌通过水平方式传播, 偶尔能在植物的茎秆和花序上形成特征性的子座, 内生真菌形成有性孢子, 通过虫媒作用, 与其他植株上配型相反的有性孢子结合, 并产生子囊孢子, 侵染新的个体(Bultman & Leuchtm- ann, 2003 ), 因此宿主的有性繁殖被完全抑制.Leuchtmann和Clay发现大多数感染Epichloë typhina 的披碱草属(Elymus )植株的同一个个体上, 往往同时存在带有子座和产生种子的两类生殖枝, 内生真菌只在产生子座的生殖枝营有性生殖, 进行水平传播, 而内生真菌在多数产生种子的生殖枝随着宿主的无性繁殖进行垂直传播, 这种既存在水平传播又有垂直传播的方式称为混合传播方式(White, 1988 ). ...
Distribution and diversity of Epichloë/Neotyphodium fungal endophytes from different populations of Achnatherum sibiricum (Poaceae) in the Inner Mongolia Steppe, China
2
2007
... 子囊菌纲(Ascomycetes)麦角菌科(Clavicipitac- eae)的无性型Neotyphodium 属及有性型Epichloë 属真菌, 普遍分布于冷季型的禾本科植物中, 在植物体内完成其部分或全部生活史(Schardl et al ., 2004 ).Neotyphodium 属内生真菌与宿主植物的关系具有代表性的研究意义, 通过垂直方式传播, 在高羊茅(Festuca elata )、多年生黑麦草(Lolium perenne )、Achnatherum robustum 和羽茅(Achnatherum sibiricum )上都存在有这类内生真菌(Wei et al ., 2007 ).Neotyphodium 内生真菌的全部生活周期都在植物体内完成, 菌丝在宿主禾草开花期进入子房, 植物产生种子后, 内生真菌便存在于种子中, 从而可以通过种子进行传播, 其间不形成任何子座或孢子.目前所研究的禾草植物中的内生真菌大部分属于此类传播方式.Epichloë 属真菌能够感染7个族16个属以上的禾本科植物, 在世界范围内分布广泛(Moon et al ., 2004 ; Schardl et al ., 2004 ). Epichloë 真菌通过水平方式传播, 偶尔能在植物的茎秆和花序上形成特征性的子座, 内生真菌形成有性孢子, 通过虫媒作用, 与其他植株上配型相反的有性孢子结合, 并产生子囊孢子, 侵染新的个体(Bultman & Leuchtm- ann, 2003 ), 因此宿主的有性繁殖被完全抑制.Leuchtmann和Clay发现大多数感染Epichloë typhina 的披碱草属(Elymus )植株的同一个个体上, 往往同时存在带有子座和产生种子的两类生殖枝, 内生真菌只在产生子座的生殖枝营有性生殖, 进行水平传播, 而内生真菌在多数产生种子的生殖枝随着宿主的无性繁殖进行垂直传播, 这种既存在水平传播又有垂直传播的方式称为混合传播方式(White, 1988 ). ...
... 中国21个地区披碱草(Elymus dahuricus )的染菌率调查表明, 有6个地区的披碱草种子中Neotypho- dium 的感染率在4.4%-100.0%, 绝大多数染菌的披碱草比不染菌的多产生一种生物碱——波胺, 波胺的浓度水平随着种群的位置和季节而变化(Zhang & Nan, 2007 ).在南京发现了Neotyphodium uncinatum 与苇状羊茅(Festuca arundinacea )的共生体(纪燕玲等, 2003 ).还有研究发现, Epichloë yangzii 是共生于鹅观草属(Roegneria )植物的产子座有性型内生真菌(申靖等, 2009 ).Wei等(2007) 对中国北方地区172个禾草地理种群的内生真菌感染状况进行广泛调查后发现, 在所有检测的41种禾草中, 25种禾草感染了内生真菌, 其中羽茅(Achnatherum sibiricum )的感染率最高, 这表明内生真菌广泛存在于天然禾草种群中, 而且内生真菌与禾草的相互作用可能对中国北方草原生态系统和畜牧业产生影响. ...
Endophyte-host associations in forage grasses. XI. A proposal concerning origin and evolution
0
1988
Distribution of Epichloë endophytes in Chinese populations of Elymus dahuricus and variation in peramine levels
1
2007
... 中国21个地区披碱草(Elymus dahuricus )的染菌率调查表明, 有6个地区的披碱草种子中Neotypho- dium 的感染率在4.4%-100.0%, 绝大多数染菌的披碱草比不染菌的多产生一种生物碱——波胺, 波胺的浓度水平随着种群的位置和季节而变化(Zhang & Nan, 2007 ).在南京发现了Neotyphodium uncinatum 与苇状羊茅(Festuca arundinacea )的共生体(纪燕玲等, 2003 ).还有研究发现, Epichloë yangzii 是共生于鹅观草属(Roegneria )植物的产子座有性型内生真菌(申靖等, 2009 ).Wei等(2007) 对中国北方地区172个禾草地理种群的内生真菌感染状况进行广泛调查后发现, 在所有检测的41种禾草中, 25种禾草感染了内生真菌, 其中羽茅(Achnatherum sibiricum )的感染率最高, 这表明内生真菌广泛存在于天然禾草种群中, 而且内生真菌与禾草的相互作用可能对中国北方草原生态系统和畜牧业产生影响. ...



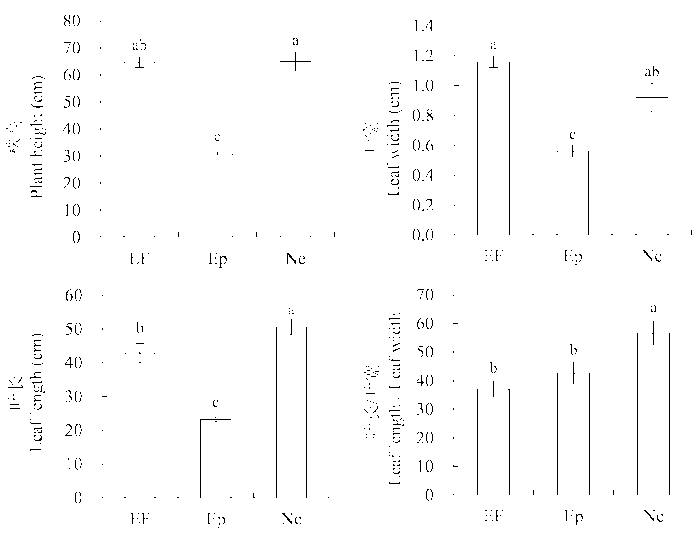
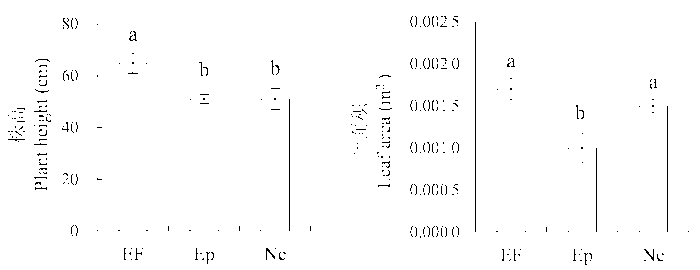
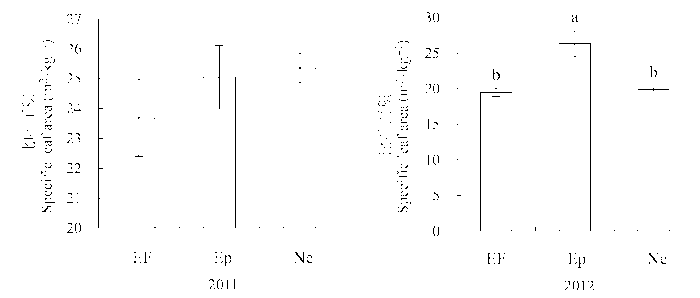

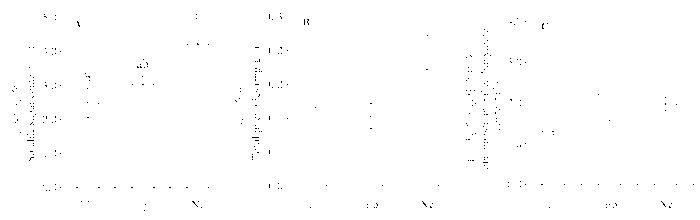
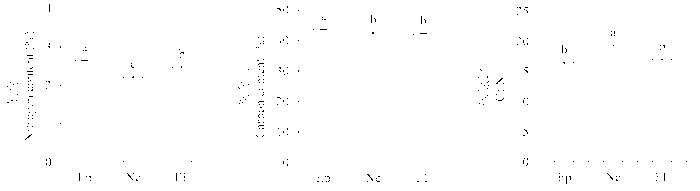
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}