Soil salinity is a major limiting factor for plant establishment, development and productivity in arid environment. This study was conducted to determine the responses and adaptation of xylem hydraulic conductivity to salt stress in Populus euphratica in order to understand the mechanisms of stress resistance and restoration strategy of this species.
Methods
The responses and adaptation of hydraulic conductivity to different levels of salt stress (NaCl concentrations of 0, 0.05, 0.15 and 0.30 mol·L-1) were investigated in P. euphratica seedlings. The testing plants were subjected to salt stresses for three months, and the stomatal conductance of leaves, hydraulic conductivity and vulnerability to cavitation of roots and stem xylem, anatomical structure of xylem vessels and root morphology and distribution were measured. The resulting data were analyzed to determine the relationships of salt stress with root uptake, hydraulic conductivity of xylem and foliar transpiration by using ANOVA, LSD and Pearson correlations.
Important findings
The roots of P. euphratica seedlings were more responsive to salt stress than stem and leaves. Root length and root tips were significantly inhibited by the salt stresses imposed. Under the salt stress, root hydraulic conductivity was significantly reduced and root xylem was more vulnerable to cavitation. The responses of stem xylem conductivity to salt stresses varied with the level of salt stress. Under a mild (0.05 mol·L-1 NaCl) and moderate (0.15 mol·L-1 NaCl) salt stress, Populus euphratica seedlings adjusted hydraulic conductivity in stem xylem by increasing the wall thickness of conduit and between conduits as well the wall mechanical strength to maintain norm growth. Under a severe salt stress (0.30 mol·L-1 NaCl), hydraulic conductivity and safety and efficacy of water transportation of stem xylem in P. euphratica seedlings significantly decreased accompanied by reduced stomatal conductance of leaves, which eventually inhibited the plant growth.
ZHOUHong-Hua, LIWei-Hong. Responses and adaptation of xylem hydraulic conductivity to salt stress in Populus euphratica. Chinese Journal of Plant Ecology, 2015, 39(1): 81-91 https://doi.org/10.17521/cjpe.2015.0009
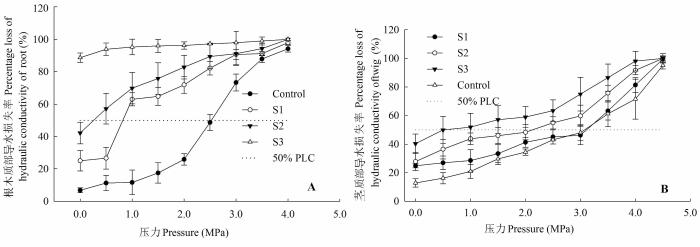
Fig. 3 Curves of xylem vulnerability to cavitation in roots and stems of Populus euphratica seedlings under different concentration salt stresses (mean ± SE). PLC, percentage loss of hydraulic conductivity. Control, 0 mol·L-1 NaCl; S1, 0.05 mol·L-1 NaCl; S2, 0.15 mol·L-1 NaCl; S3, 0.30 mol·L-1 NaCl.
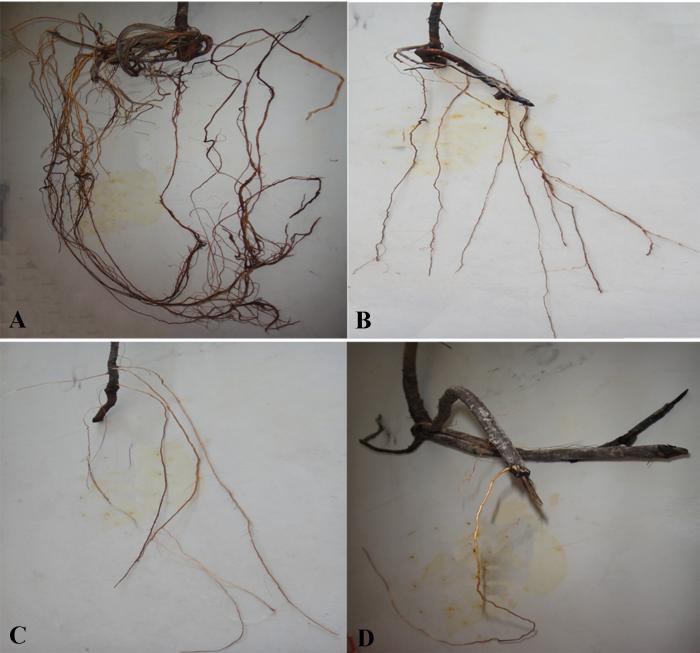
Fig. 5 Root morphology and distribution of Populus euphratica seedlings under different concentration salt stresses. A, control level (0 mol·L-1 NaCl solution); B, S1 level (0.05 mol·L-1 NaCl solution); C, S2 level (0.15 mol·L-1 NaCl solution); D, S3 level (0.30 mol·L-1 NaCl solution).
Stimulus-induced downregulation of root water transport involves reactive oxygen species- activated cell signalling and plasma membrane intrinsic protein internalization
CowanIR (1982). Regulation of water use in relation to carbon gain in higher plants. In: Lange OL, Nobel PS, Osmond CB, Ziegler H eds. Physiological Plant Ecology Ⅱ Water Relations and Carbon Assimilation. Springer-Verlag, Berlin. 589-613.
MansourMMF (1997). Cell permeability under salt stress. In: Jaiwal PK, Singh RP, Gulati A eds. Strategies for Improving Salt Tolerance in Higher Plants. Science Publishers, Boca Raton, USA. 87-110.
Effects of tree harvesting, forest floor removal, and compaction on soil microbial biomass, microbial respiration, and N availability in a boreal aspen forest in British Columbia
Boric acid and salinity effects on maize roots. Response of aquaporins ZmPIP1 and Zm PIP2, and plasm a membrane H+-ATPase, in relation to water and nutrient uptake
Regulation by arbuscular mycorrhizae of the integrated physiological response to salinity in plants: New challenges in physiological and molecular studies
Effects of NaCl and Na2CO3 stresses on photosynthesis and parameters of chlorophyll fluorescence in Helianthus tuberosus seedlings. Journal of Plant Ecology (Chinese Version)
Hypoxia affects root sodium and chloride concentrations and alters water conductance in salt-treated jack pine (Pinus banksiana) seedlings
1
2003
... 全世界盐渍化土壤面积约有10 × 108 hm2, 占世界陆地面积的7.6%, 而中国是世界盐碱地大国之一, 盐渍土面积约0.27 × 108 hm2, 大多分布在中国西北干旱区, 且这一面积随着人类的不合理灌溉正不断地扩大(高光林等, 2003).土壤盐渍化已成为影响植物生长发育的重要环境限制因素(Evelin et al., 2009).一方面, 土壤盐胁迫会降低土壤水势, 发生渗透胁迫(Munns, 2002), 促使根系产生脱落酸传递到叶片(Kang & Zhang, 2004), 诱导植物发生水分胁迫, 使叶片气孔部分关闭(Rahnama et al., 2010), 减少蒸腾(Schachtman & Goodger, 2008), 降低根系吸水能力(Navarro et al., 2007; Aroca et al., 2012; Calvo-Polanco et al., 2014), 从而导致植物叶片同化能力的降低和植物总光合面积的下降(Nastou et al., 1999; 杨启良等, 2009), 最终抑制植物生长.另一方面, 土壤盐胁迫也会通过离子毒害来影响酶活性、脂质代谢以及蛋白质合成等植物的代谢过程来抑制植物生长(Munns & Tester, 2008).盐胁迫下, 植物根系会缺氧(Haeussler & Kabzems, 2006; Mariani et al., 2006), 造成更多NaCl进入根系(Redfield et al., 2003), 促使根系和冠层中的Na+、Cl-浓度不断积累 (Mansour, 1997; Apostol & Zwiazek, 2003).高浓度Na+可置换质膜和细胞内膜系统所结合的Ca2+, 使膜结构完整性及膜功能受到破坏(Boursiac et al., 2008), 从而影响一系列生理代谢功能.另外, 高浓度Na+还会阻碍植物对K+和Ca2+的吸收和运输, 高浓度Cl-会抑制植物对NO-3和HPO42-的吸收, 造成植物营养吸收不平衡, 使碳水化合物不能顺利地被运输(张新春等, 2002; Apostol et al., 2004; López- Pérez et al., 2007), 从而抑制植物生长发育. ...
Naphthenic acids affect plant water conductance but do not alter shoot Na+ and Cl- concentrations in jack pine (Pinus banksiana) seedlings
2
2004
... 全世界盐渍化土壤面积约有10 × 108 hm2, 占世界陆地面积的7.6%, 而中国是世界盐碱地大国之一, 盐渍土面积约0.27 × 108 hm2, 大多分布在中国西北干旱区, 且这一面积随着人类的不合理灌溉正不断地扩大(高光林等, 2003).土壤盐渍化已成为影响植物生长发育的重要环境限制因素(Evelin et al., 2009).一方面, 土壤盐胁迫会降低土壤水势, 发生渗透胁迫(Munns, 2002), 促使根系产生脱落酸传递到叶片(Kang & Zhang, 2004), 诱导植物发生水分胁迫, 使叶片气孔部分关闭(Rahnama et al., 2010), 减少蒸腾(Schachtman & Goodger, 2008), 降低根系吸水能力(Navarro et al., 2007; Aroca et al., 2012; Calvo-Polanco et al., 2014), 从而导致植物叶片同化能力的降低和植物总光合面积的下降(Nastou et al., 1999; 杨启良等, 2009), 最终抑制植物生长.另一方面, 土壤盐胁迫也会通过离子毒害来影响酶活性、脂质代谢以及蛋白质合成等植物的代谢过程来抑制植物生长(Munns & Tester, 2008).盐胁迫下, 植物根系会缺氧(Haeussler & Kabzems, 2006; Mariani et al., 2006), 造成更多NaCl进入根系(Redfield et al., 2003), 促使根系和冠层中的Na+、Cl-浓度不断积累 (Mansour, 1997; Apostol & Zwiazek, 2003).高浓度Na+可置换质膜和细胞内膜系统所结合的Ca2+, 使膜结构完整性及膜功能受到破坏(Boursiac et al., 2008), 从而影响一系列生理代谢功能.另外, 高浓度Na+还会阻碍植物对K+和Ca2+的吸收和运输, 高浓度Cl-会抑制植物对NO-3和HPO42-的吸收, 造成植物营养吸收不平衡, 使碳水化合物不能顺利地被运输(张新春等, 2002; Apostol et al., 2004; López- Pérez et al., 2007), 从而抑制植物生长发育. ...
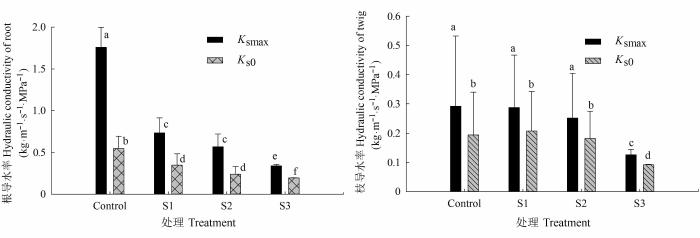
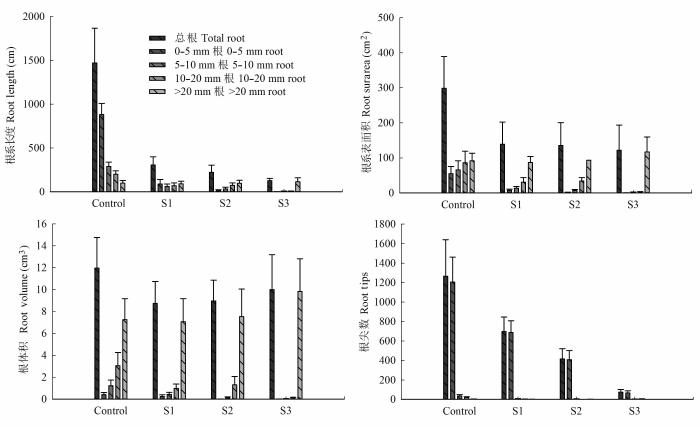
... 细根和根尖的数量与胡杨根系导水效率成正相关(表1).根系生长受限降低了根系吸收面积, 加之根系存储Na+相对较多, 木质部汁液中积累了较高浓度的Cl-, 导致离子毒害的发生, 此外土壤盐胁迫也降低了土壤水势, 这将进一步减弱根系的吸水和传输能力(Martínez-Ballesta et al., 2000; Apostol et al., 2004; Navarro et al., 2007; 杨启良等, 2009; 姚立民等, 2011; Calvo-Polanco et al., 2014).植物水分传输有两种途径, 一种是细胞-细胞间的水分传输途径, 一种是木质部内水分传输途径.大量研究证实, 盐胁迫下根细胞内通道蛋白活性和数量以及渗透物质积累增加(Qi et al., 2009; Aroca et al., 2012; Ruiz-Lozano et al., 2012; Calvo-Polanco et al., 2014), 但也有研究表明盐胁迫下根细胞内水通道蛋白显著受抑, 根系导水率与根系水通道蛋白的含量出现一致下调(Martínez-Ballesta et al., 2003, 2008).我们的研究结果表明, 盐胁迫显著降低了根木质部初始导水率和潜在最大导水率, 且随着盐分浓度的增加, 潜在导水率与初始导水率间的差异减小(图2), 即盐胁迫缩小了根木质部对水分传导率的调节范围.盐胁迫使根更易发生栓塞, 显著加剧了木质部栓塞脆弱性(图3), 减弱了木质部对逆境的抵御能力.这表明盐胁迫可能抑制了胡杨幼株根细胞内的水通道蛋白的活动(数量和活性)和渗透物质的积累, 并限制了胡杨幼株根系生长, 导致根木质部导水速率的 降低. ...
Regulation of root water uptake under abiotic stress conditions
4
2012
... 全世界盐渍化土壤面积约有10 × 108 hm2, 占世界陆地面积的7.6%, 而中国是世界盐碱地大国之一, 盐渍土面积约0.27 × 108 hm2, 大多分布在中国西北干旱区, 且这一面积随着人类的不合理灌溉正不断地扩大(高光林等, 2003).土壤盐渍化已成为影响植物生长发育的重要环境限制因素(Evelin et al., 2009).一方面, 土壤盐胁迫会降低土壤水势, 发生渗透胁迫(Munns, 2002), 促使根系产生脱落酸传递到叶片(Kang & Zhang, 2004), 诱导植物发生水分胁迫, 使叶片气孔部分关闭(Rahnama et al., 2010), 减少蒸腾(Schachtman & Goodger, 2008), 降低根系吸水能力(Navarro et al., 2007; Aroca et al., 2012; Calvo-Polanco et al., 2014), 从而导致植物叶片同化能力的降低和植物总光合面积的下降(Nastou et al., 1999; 杨启良等, 2009), 最终抑制植物生长.另一方面, 土壤盐胁迫也会通过离子毒害来影响酶活性、脂质代谢以及蛋白质合成等植物的代谢过程来抑制植物生长(Munns & Tester, 2008).盐胁迫下, 植物根系会缺氧(Haeussler & Kabzems, 2006; Mariani et al., 2006), 造成更多NaCl进入根系(Redfield et al., 2003), 促使根系和冠层中的Na+、Cl-浓度不断积累 (Mansour, 1997; Apostol & Zwiazek, 2003).高浓度Na+可置换质膜和细胞内膜系统所结合的Ca2+, 使膜结构完整性及膜功能受到破坏(Boursiac et al., 2008), 从而影响一系列生理代谢功能.另外, 高浓度Na+还会阻碍植物对K+和Ca2+的吸收和运输, 高浓度Cl-会抑制植物对NO-3和HPO42-的吸收, 造成植物营养吸收不平衡, 使碳水化合物不能顺利地被运输(张新春等, 2002; Apostol et al., 2004; López- Pérez et al., 2007), 从而抑制植物生长发育. ...
... 研究表明, 植物根可以通过改变细胞内渗透性来抵御一定程度的土壤盐胁迫.一种途径是增强质膜内水通道蛋白的数量和活性(Qi et al., 2009; Calvo-Polanco et al., 2014), 从而增强细胞膜渗透性, 调节根系的水分吸收能力(García-Sánchez et al., 2000; Postaire et al., 2010).另一种途径是通过在植物体内累积脯氨酸、可溶性糖、甘氨酸、甜菜碱、甘露醇等代谢物的含量来平衡细胞质的水势, 减缓渗透胁迫(Aroca et al., 2012; Ruiz-Lozano et al., 2012; Calvo-Polanco et al., 2014), 从而让植物适应盐胁迫.尽管有很多学者就NaCl胁迫对植物根水力导度的影响进行了研究, 但大多数的研究都集中在短期的轻度盐胁迫实验处理(从几分钟到24 h或几天), 且都是针对根和根内细胞-细胞间的水分传导特征(细胞内物质积累和基因表达)研究(Guo et al., 2006; Horie et al., 2011; Muries et al., 2011; Sutka et al., 2011; Calvo-Polanco et al., 2014).对于长期盐胁迫, 尤其是高浓度盐胁迫下, 植物的根、茎、叶整个水分传导通道(木质部)的响应和形态适应研究较少.本研究以中国西北干旱、盐渍化区荒漠河岸林建群种胡杨(Populus euphratica) 2-3年生幼株为研究对象, 系统研究了轻度、中度和重度盐胁迫下胡杨根、茎、叶水分传输通道(木质部)的响应与适应, 以期为盐渍化区域的荒漠河岸林保护、恢复与重建提供理论基础. ...
... 细根和根尖的数量与胡杨根系导水效率成正相关(表1).根系生长受限降低了根系吸收面积, 加之根系存储Na+相对较多, 木质部汁液中积累了较高浓度的Cl-, 导致离子毒害的发生, 此外土壤盐胁迫也降低了土壤水势, 这将进一步减弱根系的吸水和传输能力(Martínez-Ballesta et al., 2000; Apostol et al., 2004; Navarro et al., 2007; 杨启良等, 2009; 姚立民等, 2011; Calvo-Polanco et al., 2014).植物水分传输有两种途径, 一种是细胞-细胞间的水分传输途径, 一种是木质部内水分传输途径.大量研究证实, 盐胁迫下根细胞内通道蛋白活性和数量以及渗透物质积累增加(Qi et al., 2009; Aroca et al., 2012; Ruiz-Lozano et al., 2012; Calvo-Polanco et al., 2014), 但也有研究表明盐胁迫下根细胞内水通道蛋白显著受抑, 根系导水率与根系水通道蛋白的含量出现一致下调(Martínez-Ballesta et al., 2003, 2008).我们的研究结果表明, 盐胁迫显著降低了根木质部初始导水率和潜在最大导水率, 且随着盐分浓度的增加, 潜在导水率与初始导水率间的差异减小(图2), 即盐胁迫缩小了根木质部对水分传导率的调节范围.盐胁迫使根更易发生栓塞, 显著加剧了木质部栓塞脆弱性(图3), 减弱了木质部对逆境的抵御能力.这表明盐胁迫可能抑制了胡杨幼株根细胞内的水通道蛋白的活动(数量和活性)和渗透物质的积累, 并限制了胡杨幼株根系生长, 导致根木质部导水速率的 降低. ...
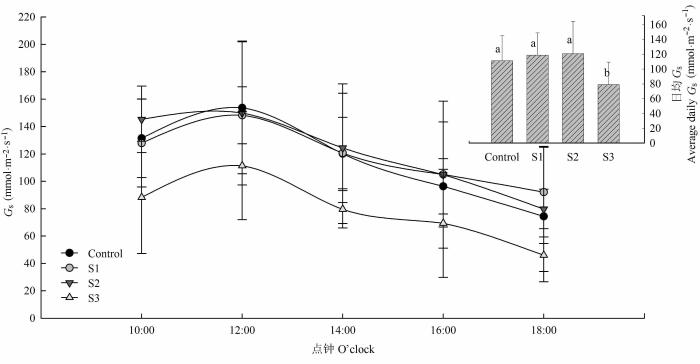
... 茎木质部是植物将根部水分运输至叶片的重要通道.三种浓度土壤盐胁迫处理下胡杨幼株茎木质部栓塞程度均受到明显影响, 但胡杨幼株的水分传导速率仅在重度盐胁迫下显著下降, 在轻度和中度盐胁迫下与对照相比却并没有发生显著变化.这表明, 在轻度和中度盐胁迫下, 胡杨茎木质部具有一定的自我调节能力.这可能与细胞内水通道蛋白数量和活性或渗透物质积累有关(Maurel, 1997; Zhang et al., 2003; Qi et al., 2009; Aroca et al., 2012; Ruiz-Lozano et al., 2012; Calvo-Polanco et al., 2014).但也有研究表明, 盐胁迫还可以通过改变细胞形态来调节水分传导(López-Pérez et al., 2007; Navarro et al., 2007; Silva et al., 2008).木质部导管是影响水分运输的重要参数.根据Hagen-Poiseuille定律, 在其他条件一致时, 水在导管中的导水速率与导管直径的4次方成正比, 即导管直径越大, 导管运输水分的能力越强, 导水率越大(Bass, 1982).大导管高流率体现了导管输水的“有效性”, 但却降低了水分输导的“安全性”, 因为大导管细胞壁薄, 机械强度低, 更容易引发气穴化, 发生阻塞 (Zimmermann, 1983).本研究分析导管细胞的解剖结构发现, 轻度和中度盐胁迫下, 与对照相比, 胡杨幼株导管直径并未显著减小, 而导管密度、管壁厚度、管壁间厚度以及管壁机械强度显著提高; 重度盐胁迫下, 茎导管直径较对照显著下降, 导管密度、管壁厚度、管壁间厚度和管壁机械强度与S2处理差异并不显著.这表明轻度和中度盐胁迫下胡杨茎木质部可能是通过增加导管密度来消除了导管直径变小的弊端, 保持了木质部的输水效率, 同时通过增加管壁厚度和机械强度提高了水分输导的安全性, 从而保证了胡杨的正常生长; 但重度盐胁迫下胡杨幼株木质部栓塞明显增强, 输水有效性和安全性显著下降, 木质部水分传导的自我适应和调控能力显著减弱, 导致木质部水分传导速率急速下降. ...
New Perspectives in Wood Anatomy. The Hague,
1
1982
... 茎木质部是植物将根部水分运输至叶片的重要通道.三种浓度土壤盐胁迫处理下胡杨幼株茎木质部栓塞程度均受到明显影响, 但胡杨幼株的水分传导速率仅在重度盐胁迫下显著下降, 在轻度和中度盐胁迫下与对照相比却并没有发生显著变化.这表明, 在轻度和中度盐胁迫下, 胡杨茎木质部具有一定的自我调节能力.这可能与细胞内水通道蛋白数量和活性或渗透物质积累有关(Maurel, 1997; Zhang et al., 2003; Qi et al., 2009; Aroca et al., 2012; Ruiz-Lozano et al., 2012; Calvo-Polanco et al., 2014).但也有研究表明, 盐胁迫还可以通过改变细胞形态来调节水分传导(López-Pérez et al., 2007; Navarro et al., 2007; Silva et al., 2008).木质部导管是影响水分运输的重要参数.根据Hagen-Poiseuille定律, 在其他条件一致时, 水在导管中的导水速率与导管直径的4次方成正比, 即导管直径越大, 导管运输水分的能力越强, 导水率越大(Bass, 1982).大导管高流率体现了导管输水的“有效性”, 但却降低了水分输导的“安全性”, 因为大导管细胞壁薄, 机械强度低, 更容易引发气穴化, 发生阻塞 (Zimmermann, 1983).本研究分析导管细胞的解剖结构发现, 轻度和中度盐胁迫下, 与对照相比, 胡杨幼株导管直径并未显著减小, 而导管密度、管壁厚度、管壁间厚度以及管壁机械强度显著提高; 重度盐胁迫下, 茎导管直径较对照显著下降, 导管密度、管壁厚度、管壁间厚度和管壁机械强度与S2处理差异并不显著.这表明轻度和中度盐胁迫下胡杨茎木质部可能是通过增加导管密度来消除了导管直径变小的弊端, 保持了木质部的输水效率, 同时通过增加管壁厚度和机械强度提高了水分输导的安全性, 从而保证了胡杨的正常生长; 但重度盐胁迫下胡杨幼株木质部栓塞明显增强, 输水有效性和安全性显著下降, 木质部水分传导的自我适应和调控能力显著减弱, 导致木质部水分传导速率急速下降. ...
Stimulus-induced downregulation of root water transport involves reactive oxygen species- activated cell signalling and plasma membrane intrinsic protein internalization
1
2008
... 全世界盐渍化土壤面积约有10 × 108 hm2, 占世界陆地面积的7.6%, 而中国是世界盐碱地大国之一, 盐渍土面积约0.27 × 108 hm2, 大多分布在中国西北干旱区, 且这一面积随着人类的不合理灌溉正不断地扩大(高光林等, 2003).土壤盐渍化已成为影响植物生长发育的重要环境限制因素(Evelin et al., 2009).一方面, 土壤盐胁迫会降低土壤水势, 发生渗透胁迫(Munns, 2002), 促使根系产生脱落酸传递到叶片(Kang & Zhang, 2004), 诱导植物发生水分胁迫, 使叶片气孔部分关闭(Rahnama et al., 2010), 减少蒸腾(Schachtman & Goodger, 2008), 降低根系吸水能力(Navarro et al., 2007; Aroca et al., 2012; Calvo-Polanco et al., 2014), 从而导致植物叶片同化能力的降低和植物总光合面积的下降(Nastou et al., 1999; 杨启良等, 2009), 最终抑制植物生长.另一方面, 土壤盐胁迫也会通过离子毒害来影响酶活性、脂质代谢以及蛋白质合成等植物的代谢过程来抑制植物生长(Munns & Tester, 2008).盐胁迫下, 植物根系会缺氧(Haeussler & Kabzems, 2006; Mariani et al., 2006), 造成更多NaCl进入根系(Redfield et al., 2003), 促使根系和冠层中的Na+、Cl-浓度不断积累 (Mansour, 1997; Apostol & Zwiazek, 2003).高浓度Na+可置换质膜和细胞内膜系统所结合的Ca2+, 使膜结构完整性及膜功能受到破坏(Boursiac et al., 2008), 从而影响一系列生理代谢功能.另外, 高浓度Na+还会阻碍植物对K+和Ca2+的吸收和运输, 高浓度Cl-会抑制植物对NO-3和HPO42-的吸收, 造成植物营养吸收不平衡, 使碳水化合物不能顺利地被运输(张新春等, 2002; Apostol et al., 2004; López- Pérez et al., 2007), 从而抑制植物生长发育. ...
Mild salt stress conditions induce different responses in root hydraulic conductivity of Phaseolus vulgaris over-time
7
2014
... 全世界盐渍化土壤面积约有10 × 108 hm2, 占世界陆地面积的7.6%, 而中国是世界盐碱地大国之一, 盐渍土面积约0.27 × 108 hm2, 大多分布在中国西北干旱区, 且这一面积随着人类的不合理灌溉正不断地扩大(高光林等, 2003).土壤盐渍化已成为影响植物生长发育的重要环境限制因素(Evelin et al., 2009).一方面, 土壤盐胁迫会降低土壤水势, 发生渗透胁迫(Munns, 2002), 促使根系产生脱落酸传递到叶片(Kang & Zhang, 2004), 诱导植物发生水分胁迫, 使叶片气孔部分关闭(Rahnama et al., 2010), 减少蒸腾(Schachtman & Goodger, 2008), 降低根系吸水能力(Navarro et al., 2007; Aroca et al., 2012; Calvo-Polanco et al., 2014), 从而导致植物叶片同化能力的降低和植物总光合面积的下降(Nastou et al., 1999; 杨启良等, 2009), 最终抑制植物生长.另一方面, 土壤盐胁迫也会通过离子毒害来影响酶活性、脂质代谢以及蛋白质合成等植物的代谢过程来抑制植物生长(Munns & Tester, 2008).盐胁迫下, 植物根系会缺氧(Haeussler & Kabzems, 2006; Mariani et al., 2006), 造成更多NaCl进入根系(Redfield et al., 2003), 促使根系和冠层中的Na+、Cl-浓度不断积累 (Mansour, 1997; Apostol & Zwiazek, 2003).高浓度Na+可置换质膜和细胞内膜系统所结合的Ca2+, 使膜结构完整性及膜功能受到破坏(Boursiac et al., 2008), 从而影响一系列生理代谢功能.另外, 高浓度Na+还会阻碍植物对K+和Ca2+的吸收和运输, 高浓度Cl-会抑制植物对NO-3和HPO42-的吸收, 造成植物营养吸收不平衡, 使碳水化合物不能顺利地被运输(张新春等, 2002; Apostol et al., 2004; López- Pérez et al., 2007), 从而抑制植物生长发育. ...
... 研究表明, 植物根可以通过改变细胞内渗透性来抵御一定程度的土壤盐胁迫.一种途径是增强质膜内水通道蛋白的数量和活性(Qi et al., 2009; Calvo-Polanco et al., 2014), 从而增强细胞膜渗透性, 调节根系的水分吸收能力(García-Sánchez et al., 2000; Postaire et al., 2010).另一种途径是通过在植物体内累积脯氨酸、可溶性糖、甘氨酸、甜菜碱、甘露醇等代谢物的含量来平衡细胞质的水势, 减缓渗透胁迫(Aroca et al., 2012; Ruiz-Lozano et al., 2012; Calvo-Polanco et al., 2014), 从而让植物适应盐胁迫.尽管有很多学者就NaCl胁迫对植物根水力导度的影响进行了研究, 但大多数的研究都集中在短期的轻度盐胁迫实验处理(从几分钟到24 h或几天), 且都是针对根和根内细胞-细胞间的水分传导特征(细胞内物质积累和基因表达)研究(Guo et al., 2006; Horie et al., 2011; Muries et al., 2011; Sutka et al., 2011; Calvo-Polanco et al., 2014).对于长期盐胁迫, 尤其是高浓度盐胁迫下, 植物的根、茎、叶整个水分传导通道(木质部)的响应和形态适应研究较少.本研究以中国西北干旱、盐渍化区荒漠河岸林建群种胡杨(Populus euphratica) 2-3年生幼株为研究对象, 系统研究了轻度、中度和重度盐胁迫下胡杨根、茎、叶水分传输通道(木质部)的响应与适应, 以期为盐渍化区域的荒漠河岸林保护、恢复与重建提供理论基础. ...
... ; Calvo-Polanco et al., 2014), 从而让植物适应盐胁迫.尽管有很多学者就NaCl胁迫对植物根水力导度的影响进行了研究, 但大多数的研究都集中在短期的轻度盐胁迫实验处理(从几分钟到24 h或几天), 且都是针对根和根内细胞-细胞间的水分传导特征(细胞内物质积累和基因表达)研究(Guo et al., 2006; Horie et al., 2011; Muries et al., 2011; Sutka et al., 2011; Calvo-Polanco et al., 2014).对于长期盐胁迫, 尤其是高浓度盐胁迫下, 植物的根、茎、叶整个水分传导通道(木质部)的响应和形态适应研究较少.本研究以中国西北干旱、盐渍化区荒漠河岸林建群种胡杨(Populus euphratica) 2-3年生幼株为研究对象, 系统研究了轻度、中度和重度盐胁迫下胡杨根、茎、叶水分传输通道(木质部)的响应与适应, 以期为盐渍化区域的荒漠河岸林保护、恢复与重建提供理论基础. ...
... 全世界盐渍化土壤面积约有10 × 108 hm2, 占世界陆地面积的7.6%, 而中国是世界盐碱地大国之一, 盐渍土面积约0.27 × 108 hm2, 大多分布在中国西北干旱区, 且这一面积随着人类的不合理灌溉正不断地扩大(高光林等, 2003).土壤盐渍化已成为影响植物生长发育的重要环境限制因素(Evelin et al., 2009).一方面, 土壤盐胁迫会降低土壤水势, 发生渗透胁迫(Munns, 2002), 促使根系产生脱落酸传递到叶片(Kang & Zhang, 2004), 诱导植物发生水分胁迫, 使叶片气孔部分关闭(Rahnama et al., 2010), 减少蒸腾(Schachtman & Goodger, 2008), 降低根系吸水能力(Navarro et al., 2007; Aroca et al., 2012; Calvo-Polanco et al., 2014), 从而导致植物叶片同化能力的降低和植物总光合面积的下降(Nastou et al., 1999; 杨启良等, 2009), 最终抑制植物生长.另一方面, 土壤盐胁迫也会通过离子毒害来影响酶活性、脂质代谢以及蛋白质合成等植物的代谢过程来抑制植物生长(Munns & Tester, 2008).盐胁迫下, 植物根系会缺氧(Haeussler & Kabzems, 2006; Mariani et al., 2006), 造成更多NaCl进入根系(Redfield et al., 2003), 促使根系和冠层中的Na+、Cl-浓度不断积累 (Mansour, 1997; Apostol & Zwiazek, 2003).高浓度Na+可置换质膜和细胞内膜系统所结合的Ca2+, 使膜结构完整性及膜功能受到破坏(Boursiac et al., 2008), 从而影响一系列生理代谢功能.另外, 高浓度Na+还会阻碍植物对K+和Ca2+的吸收和运输, 高浓度Cl-会抑制植物对NO-3和HPO42-的吸收, 造成植物营养吸收不平衡, 使碳水化合物不能顺利地被运输(张新春等, 2002; Apostol et al., 2004; López- Pérez et al., 2007), 从而抑制植物生长发育. ...
Effects of tree harvesting, forest floor removal, and compaction on soil microbial biomass, microbial respiration, and N availability in a boreal aspen forest in British Columbia
1
2006
... 全世界盐渍化土壤面积约有10 × 108 hm2, 占世界陆地面积的7.6%, 而中国是世界盐碱地大国之一, 盐渍土面积约0.27 × 108 hm2, 大多分布在中国西北干旱区, 且这一面积随着人类的不合理灌溉正不断地扩大(高光林等, 2003).土壤盐渍化已成为影响植物生长发育的重要环境限制因素(Evelin et al., 2009).一方面, 土壤盐胁迫会降低土壤水势, 发生渗透胁迫(Munns, 2002), 促使根系产生脱落酸传递到叶片(Kang & Zhang, 2004), 诱导植物发生水分胁迫, 使叶片气孔部分关闭(Rahnama et al., 2010), 减少蒸腾(Schachtman & Goodger, 2008), 降低根系吸水能力(Navarro et al., 2007; Aroca et al., 2012; Calvo-Polanco et al., 2014), 从而导致植物叶片同化能力的降低和植物总光合面积的下降(Nastou et al., 1999; 杨启良等, 2009), 最终抑制植物生长.另一方面, 土壤盐胁迫也会通过离子毒害来影响酶活性、脂质代谢以及蛋白质合成等植物的代谢过程来抑制植物生长(Munns & Tester, 2008).盐胁迫下, 植物根系会缺氧(Haeussler & Kabzems, 2006; Mariani et al., 2006), 造成更多NaCl进入根系(Redfield et al., 2003), 促使根系和冠层中的Na+、Cl-浓度不断积累 (Mansour, 1997; Apostol & Zwiazek, 2003).高浓度Na+可置换质膜和细胞内膜系统所结合的Ca2+, 使膜结构完整性及膜功能受到破坏(Boursiac et al., 2008), 从而影响一系列生理代谢功能.另外, 高浓度Na+还会阻碍植物对K+和Ca2+的吸收和运输, 高浓度Cl-会抑制植物对NO-3和HPO42-的吸收, 造成植物营养吸收不平衡, 使碳水化合物不能顺利地被运输(张新春等, 2002; Apostol et al., 2004; López- Pérez et al., 2007), 从而抑制植物生长发育. ...
Influence of saline stress on root hydraulic conductance and PIP expression in Arabidopsis
1
2003
... 细根和根尖的数量与胡杨根系导水效率成正相关(表1).根系生长受限降低了根系吸收面积, 加之根系存储Na+相对较多, 木质部汁液中积累了较高浓度的Cl-, 导致离子毒害的发生, 此外土壤盐胁迫也降低了土壤水势, 这将进一步减弱根系的吸水和传输能力(Martínez-Ballesta et al., 2000; Apostol et al., 2004; Navarro et al., 2007; 杨启良等, 2009; 姚立民等, 2011; Calvo-Polanco et al., 2014).植物水分传输有两种途径, 一种是细胞-细胞间的水分传输途径, 一种是木质部内水分传输途径.大量研究证实, 盐胁迫下根细胞内通道蛋白活性和数量以及渗透物质积累增加(Qi et al., 2009; Aroca et al., 2012; Ruiz-Lozano et al., 2012; Calvo-Polanco et al., 2014), 但也有研究表明盐胁迫下根细胞内水通道蛋白显著受抑, 根系导水率与根系水通道蛋白的含量出现一致下调(Martínez-Ballesta et al., 2003, 2008).我们的研究结果表明, 盐胁迫显著降低了根木质部初始导水率和潜在最大导水率, 且随着盐分浓度的增加, 潜在导水率与初始导水率间的差异减小(图2), 即盐胁迫缩小了根木质部对水分传导率的调节范围.盐胁迫使根更易发生栓塞, 显著加剧了木质部栓塞脆弱性(图3), 减弱了木质部对逆境的抵御能力.这表明盐胁迫可能抑制了胡杨幼株根细胞内的水通道蛋白的活动(数量和活性)和渗透物质的积累, 并限制了胡杨幼株根系生长, 导致根木质部导水速率的 降低. ...
Boric acid and salinity effects on maize roots. Response of aquaporins ZmPIP1 and Zm PIP2, and plasm a membrane H+-ATPase, in relation to water and nutrient uptake
1
2008
... 细根和根尖的数量与胡杨根系导水效率成正相关(表1).根系生长受限降低了根系吸收面积, 加之根系存储Na+相对较多, 木质部汁液中积累了较高浓度的Cl-, 导致离子毒害的发生, 此外土壤盐胁迫也降低了土壤水势, 这将进一步减弱根系的吸水和传输能力(Martínez-Ballesta et al., 2000; Apostol et al., 2004; Navarro et al., 2007; 杨启良等, 2009; 姚立民等, 2011; Calvo-Polanco et al., 2014).植物水分传输有两种途径, 一种是细胞-细胞间的水分传输途径, 一种是木质部内水分传输途径.大量研究证实, 盐胁迫下根细胞内通道蛋白活性和数量以及渗透物质积累增加(Qi et al., 2009; Aroca et al., 2012; Ruiz-Lozano et al., 2012; Calvo-Polanco et al., 2014), 但也有研究表明盐胁迫下根细胞内水通道蛋白显著受抑, 根系导水率与根系水通道蛋白的含量出现一致下调(Martínez-Ballesta et al., 2003, 2008).我们的研究结果表明, 盐胁迫显著降低了根木质部初始导水率和潜在最大导水率, 且随着盐分浓度的增加, 潜在导水率与初始导水率间的差异减小(图2), 即盐胁迫缩小了根木质部对水分传导率的调节范围.盐胁迫使根更易发生栓塞, 显著加剧了木质部栓塞脆弱性(图3), 减弱了木质部对逆境的抵御能力.这表明盐胁迫可能抑制了胡杨幼株根细胞内的水通道蛋白的活动(数量和活性)和渗透物质的积累, 并限制了胡杨幼株根系生长, 导致根木质部导水速率的 降低. ...
Regulation of water channel activity in whole roots and in protoplasts from roots of melon plants grown under saline conditions
1
2000
... 细根和根尖的数量与胡杨根系导水效率成正相关(表1).根系生长受限降低了根系吸收面积, 加之根系存储Na+相对较多, 木质部汁液中积累了较高浓度的Cl-, 导致离子毒害的发生, 此外土壤盐胁迫也降低了土壤水势, 这将进一步减弱根系的吸水和传输能力(Martínez-Ballesta et al., 2000; Apostol et al., 2004; Navarro et al., 2007; 杨启良等, 2009; 姚立民等, 2011; Calvo-Polanco et al., 2014).植物水分传输有两种途径, 一种是细胞-细胞间的水分传输途径, 一种是木质部内水分传输途径.大量研究证实, 盐胁迫下根细胞内通道蛋白活性和数量以及渗透物质积累增加(Qi et al., 2009; Aroca et al., 2012; Ruiz-Lozano et al., 2012; Calvo-Polanco et al., 2014), 但也有研究表明盐胁迫下根细胞内水通道蛋白显著受抑, 根系导水率与根系水通道蛋白的含量出现一致下调(Martínez-Ballesta et al., 2003, 2008).我们的研究结果表明, 盐胁迫显著降低了根木质部初始导水率和潜在最大导水率, 且随着盐分浓度的增加, 潜在导水率与初始导水率间的差异减小(图2), 即盐胁迫缩小了根木质部对水分传导率的调节范围.盐胁迫使根更易发生栓塞, 显著加剧了木质部栓塞脆弱性(图3), 减弱了木质部对逆境的抵御能力.这表明盐胁迫可能抑制了胡杨幼株根细胞内的水通道蛋白的活动(数量和活性)和渗透物质的积累, 并限制了胡杨幼株根系生长, 导致根木质部导水速率的 降低. ...
Aquaporins and water permeability of plant membranes
1
1997
... 茎木质部是植物将根部水分运输至叶片的重要通道.三种浓度土壤盐胁迫处理下胡杨幼株茎木质部栓塞程度均受到明显影响, 但胡杨幼株的水分传导速率仅在重度盐胁迫下显著下降, 在轻度和中度盐胁迫下与对照相比却并没有发生显著变化.这表明, 在轻度和中度盐胁迫下, 胡杨茎木质部具有一定的自我调节能力.这可能与细胞内水通道蛋白数量和活性或渗透物质积累有关(Maurel, 1997; Zhang et al., 2003; Qi et al., 2009; Aroca et al., 2012; Ruiz-Lozano et al., 2012; Calvo-Polanco et al., 2014).但也有研究表明, 盐胁迫还可以通过改变细胞形态来调节水分传导(López-Pérez et al., 2007; Navarro et al., 2007; Silva et al., 2008).木质部导管是影响水分运输的重要参数.根据Hagen-Poiseuille定律, 在其他条件一致时, 水在导管中的导水速率与导管直径的4次方成正比, 即导管直径越大, 导管运输水分的能力越强, 导水率越大(Bass, 1982).大导管高流率体现了导管输水的“有效性”, 但却降低了水分输导的“安全性”, 因为大导管细胞壁薄, 机械强度低, 更容易引发气穴化, 发生阻塞 (Zimmermann, 1983).本研究分析导管细胞的解剖结构发现, 轻度和中度盐胁迫下, 与对照相比, 胡杨幼株导管直径并未显著减小, 而导管密度、管壁厚度、管壁间厚度以及管壁机械强度显著提高; 重度盐胁迫下, 茎导管直径较对照显著下降, 导管密度、管壁厚度、管壁间厚度和管壁机械强度与S2处理差异并不显著.这表明轻度和中度盐胁迫下胡杨茎木质部可能是通过增加导管密度来消除了导管直径变小的弊端, 保持了木质部的输水效率, 同时通过增加管壁厚度和机械强度提高了水分输导的安全性, 从而保证了胡杨的正常生长; 但重度盐胁迫下胡杨幼株木质部栓塞明显增强, 输水有效性和安全性显著下降, 木质部水分传导的自我适应和调控能力显著减弱, 导致木质部水分传导速率急速下降. ...
Comparative physiology of salt and water stress
1
2002
... 全世界盐渍化土壤面积约有10 × 108 hm2, 占世界陆地面积的7.6%, 而中国是世界盐碱地大国之一, 盐渍土面积约0.27 × 108 hm2, 大多分布在中国西北干旱区, 且这一面积随着人类的不合理灌溉正不断地扩大(高光林等, 2003).土壤盐渍化已成为影响植物生长发育的重要环境限制因素(Evelin et al., 2009).一方面, 土壤盐胁迫会降低土壤水势, 发生渗透胁迫(Munns, 2002), 促使根系产生脱落酸传递到叶片(Kang & Zhang, 2004), 诱导植物发生水分胁迫, 使叶片气孔部分关闭(Rahnama et al., 2010), 减少蒸腾(Schachtman & Goodger, 2008), 降低根系吸水能力(Navarro et al., 2007; Aroca et al., 2012; Calvo-Polanco et al., 2014), 从而导致植物叶片同化能力的降低和植物总光合面积的下降(Nastou et al., 1999; 杨启良等, 2009), 最终抑制植物生长.另一方面, 土壤盐胁迫也会通过离子毒害来影响酶活性、脂质代谢以及蛋白质合成等植物的代谢过程来抑制植物生长(Munns & Tester, 2008).盐胁迫下, 植物根系会缺氧(Haeussler & Kabzems, 2006; Mariani et al., 2006), 造成更多NaCl进入根系(Redfield et al., 2003), 促使根系和冠层中的Na+、Cl-浓度不断积累 (Mansour, 1997; Apostol & Zwiazek, 2003).高浓度Na+可置换质膜和细胞内膜系统所结合的Ca2+, 使膜结构完整性及膜功能受到破坏(Boursiac et al., 2008), 从而影响一系列生理代谢功能.另外, 高浓度Na+还会阻碍植物对K+和Ca2+的吸收和运输, 高浓度Cl-会抑制植物对NO-3和HPO42-的吸收, 造成植物营养吸收不平衡, 使碳水化合物不能顺利地被运输(张新春等, 2002; Apostol et al., 2004; López- Pérez et al., 2007), 从而抑制植物生长发育. ...
Mechanisms of salinity tolerance
1
2008
... 全世界盐渍化土壤面积约有10 × 108 hm2, 占世界陆地面积的7.6%, 而中国是世界盐碱地大国之一, 盐渍土面积约0.27 × 108 hm2, 大多分布在中国西北干旱区, 且这一面积随着人类的不合理灌溉正不断地扩大(高光林等, 2003).土壤盐渍化已成为影响植物生长发育的重要环境限制因素(Evelin et al., 2009).一方面, 土壤盐胁迫会降低土壤水势, 发生渗透胁迫(Munns, 2002), 促使根系产生脱落酸传递到叶片(Kang & Zhang, 2004), 诱导植物发生水分胁迫, 使叶片气孔部分关闭(Rahnama et al., 2010), 减少蒸腾(Schachtman & Goodger, 2008), 降低根系吸水能力(Navarro et al., 2007; Aroca et al., 2012; Calvo-Polanco et al., 2014), 从而导致植物叶片同化能力的降低和植物总光合面积的下降(Nastou et al., 1999; 杨启良等, 2009), 最终抑制植物生长.另一方面, 土壤盐胁迫也会通过离子毒害来影响酶活性、脂质代谢以及蛋白质合成等植物的代谢过程来抑制植物生长(Munns & Tester, 2008).盐胁迫下, 植物根系会缺氧(Haeussler & Kabzems, 2006; Mariani et al., 2006), 造成更多NaCl进入根系(Redfield et al., 2003), 促使根系和冠层中的Na+、Cl-浓度不断积累 (Mansour, 1997; Apostol & Zwiazek, 2003).高浓度Na+可置换质膜和细胞内膜系统所结合的Ca2+, 使膜结构完整性及膜功能受到破坏(Boursiac et al., 2008), 从而影响一系列生理代谢功能.另外, 高浓度Na+还会阻碍植物对K+和Ca2+的吸收和运输, 高浓度Cl-会抑制植物对NO-3和HPO42-的吸收, 造成植物营养吸收不平衡, 使碳水化合物不能顺利地被运输(张新春等, 2002; Apostol et al., 2004; López- Pérez et al., 2007), 从而抑制植物生长发育. ...
Identification and differential induction of the expression of aquaporins by salinity in broccoli plants
1
2011
... 研究表明, 植物根可以通过改变细胞内渗透性来抵御一定程度的土壤盐胁迫.一种途径是增强质膜内水通道蛋白的数量和活性(Qi et al., 2009; Calvo-Polanco et al., 2014), 从而增强细胞膜渗透性, 调节根系的水分吸收能力(García-Sánchez et al., 2000; Postaire et al., 2010).另一种途径是通过在植物体内累积脯氨酸、可溶性糖、甘氨酸、甜菜碱、甘露醇等代谢物的含量来平衡细胞质的水势, 减缓渗透胁迫(Aroca et al., 2012; Ruiz-Lozano et al., 2012; Calvo-Polanco et al., 2014), 从而让植物适应盐胁迫.尽管有很多学者就NaCl胁迫对植物根水力导度的影响进行了研究, 但大多数的研究都集中在短期的轻度盐胁迫实验处理(从几分钟到24 h或几天), 且都是针对根和根内细胞-细胞间的水分传导特征(细胞内物质积累和基因表达)研究(Guo et al., 2006; Horie et al., 2011; Muries et al., 2011; Sutka et al., 2011; Calvo-Polanco et al., 2014).对于长期盐胁迫, 尤其是高浓度盐胁迫下, 植物的根、茎、叶整个水分传导通道(木质部)的响应和形态适应研究较少.本研究以中国西北干旱、盐渍化区荒漠河岸林建群种胡杨(Populus euphratica) 2-3年生幼株为研究对象, 系统研究了轻度、中度和重度盐胁迫下胡杨根、茎、叶水分传输通道(木质部)的响应与适应, 以期为盐渍化区域的荒漠河岸林保护、恢复与重建提供理论基础. ...
Leaf anatomical responses, ion content and CO2 assimilation in three lemon cultivars under NaCl salinity
1
1999
... 全世界盐渍化土壤面积约有10 × 108 hm2, 占世界陆地面积的7.6%, 而中国是世界盐碱地大国之一, 盐渍土面积约0.27 × 108 hm2, 大多分布在中国西北干旱区, 且这一面积随着人类的不合理灌溉正不断地扩大(高光林等, 2003).土壤盐渍化已成为影响植物生长发育的重要环境限制因素(Evelin et al., 2009).一方面, 土壤盐胁迫会降低土壤水势, 发生渗透胁迫(Munns, 2002), 促使根系产生脱落酸传递到叶片(Kang & Zhang, 2004), 诱导植物发生水分胁迫, 使叶片气孔部分关闭(Rahnama et al., 2010), 减少蒸腾(Schachtman & Goodger, 2008), 降低根系吸水能力(Navarro et al., 2007; Aroca et al., 2012; Calvo-Polanco et al., 2014), 从而导致植物叶片同化能力的降低和植物总光合面积的下降(Nastou et al., 1999; 杨启良等, 2009), 最终抑制植物生长.另一方面, 土壤盐胁迫也会通过离子毒害来影响酶活性、脂质代谢以及蛋白质合成等植物的代谢过程来抑制植物生长(Munns & Tester, 2008).盐胁迫下, 植物根系会缺氧(Haeussler & Kabzems, 2006; Mariani et al., 2006), 造成更多NaCl进入根系(Redfield et al., 2003), 促使根系和冠层中的Na+、Cl-浓度不断积累 (Mansour, 1997; Apostol & Zwiazek, 2003).高浓度Na+可置换质膜和细胞内膜系统所结合的Ca2+, 使膜结构完整性及膜功能受到破坏(Boursiac et al., 2008), 从而影响一系列生理代谢功能.另外, 高浓度Na+还会阻碍植物对K+和Ca2+的吸收和运输, 高浓度Cl-会抑制植物对NO-3和HPO42-的吸收, 造成植物营养吸收不平衡, 使碳水化合物不能顺利地被运输(张新春等, 2002; Apostol et al., 2004; López- Pérez et al., 2007), 从而抑制植物生长发育. ...
Effects of sodium chloride on water potential components, hydraulic conductivity, gas exchange and leaf ultrastructure of Arbutus unedo plants
4
2007
... 全世界盐渍化土壤面积约有10 × 108 hm2, 占世界陆地面积的7.6%, 而中国是世界盐碱地大国之一, 盐渍土面积约0.27 × 108 hm2, 大多分布在中国西北干旱区, 且这一面积随着人类的不合理灌溉正不断地扩大(高光林等, 2003).土壤盐渍化已成为影响植物生长发育的重要环境限制因素(Evelin et al., 2009).一方面, 土壤盐胁迫会降低土壤水势, 发生渗透胁迫(Munns, 2002), 促使根系产生脱落酸传递到叶片(Kang & Zhang, 2004), 诱导植物发生水分胁迫, 使叶片气孔部分关闭(Rahnama et al., 2010), 减少蒸腾(Schachtman & Goodger, 2008), 降低根系吸水能力(Navarro et al., 2007; Aroca et al., 2012; Calvo-Polanco et al., 2014), 从而导致植物叶片同化能力的降低和植物总光合面积的下降(Nastou et al., 1999; 杨启良等, 2009), 最终抑制植物生长.另一方面, 土壤盐胁迫也会通过离子毒害来影响酶活性、脂质代谢以及蛋白质合成等植物的代谢过程来抑制植物生长(Munns & Tester, 2008).盐胁迫下, 植物根系会缺氧(Haeussler & Kabzems, 2006; Mariani et al., 2006), 造成更多NaCl进入根系(Redfield et al., 2003), 促使根系和冠层中的Na+、Cl-浓度不断积累 (Mansour, 1997; Apostol & Zwiazek, 2003).高浓度Na+可置换质膜和细胞内膜系统所结合的Ca2+, 使膜结构完整性及膜功能受到破坏(Boursiac et al., 2008), 从而影响一系列生理代谢功能.另外, 高浓度Na+还会阻碍植物对K+和Ca2+的吸收和运输, 高浓度Cl-会抑制植物对NO-3和HPO42-的吸收, 造成植物营养吸收不平衡, 使碳水化合物不能顺利地被运输(张新春等, 2002; Apostol et al., 2004; López- Pérez et al., 2007), 从而抑制植物生长发育. ...
... 细根和根尖的数量与胡杨根系导水效率成正相关(表1).根系生长受限降低了根系吸收面积, 加之根系存储Na+相对较多, 木质部汁液中积累了较高浓度的Cl-, 导致离子毒害的发生, 此外土壤盐胁迫也降低了土壤水势, 这将进一步减弱根系的吸水和传输能力(Martínez-Ballesta et al., 2000; Apostol et al., 2004; Navarro et al., 2007; 杨启良等, 2009; 姚立民等, 2011; Calvo-Polanco et al., 2014).植物水分传输有两种途径, 一种是细胞-细胞间的水分传输途径, 一种是木质部内水分传输途径.大量研究证实, 盐胁迫下根细胞内通道蛋白活性和数量以及渗透物质积累增加(Qi et al., 2009; Aroca et al., 2012; Ruiz-Lozano et al., 2012; Calvo-Polanco et al., 2014), 但也有研究表明盐胁迫下根细胞内水通道蛋白显著受抑, 根系导水率与根系水通道蛋白的含量出现一致下调(Martínez-Ballesta et al., 2003, 2008).我们的研究结果表明, 盐胁迫显著降低了根木质部初始导水率和潜在最大导水率, 且随着盐分浓度的增加, 潜在导水率与初始导水率间的差异减小(图2), 即盐胁迫缩小了根木质部对水分传导率的调节范围.盐胁迫使根更易发生栓塞, 显著加剧了木质部栓塞脆弱性(图3), 减弱了木质部对逆境的抵御能力.这表明盐胁迫可能抑制了胡杨幼株根细胞内的水通道蛋白的活动(数量和活性)和渗透物质的积累, 并限制了胡杨幼株根系生长, 导致根木质部导水速率的 降低. ...
... 茎木质部是植物将根部水分运输至叶片的重要通道.三种浓度土壤盐胁迫处理下胡杨幼株茎木质部栓塞程度均受到明显影响, 但胡杨幼株的水分传导速率仅在重度盐胁迫下显著下降, 在轻度和中度盐胁迫下与对照相比却并没有发生显著变化.这表明, 在轻度和中度盐胁迫下, 胡杨茎木质部具有一定的自我调节能力.这可能与细胞内水通道蛋白数量和活性或渗透物质积累有关(Maurel, 1997; Zhang et al., 2003; Qi et al., 2009; Aroca et al., 2012; Ruiz-Lozano et al., 2012; Calvo-Polanco et al., 2014).但也有研究表明, 盐胁迫还可以通过改变细胞形态来调节水分传导(López-Pérez et al., 2007; Navarro et al., 2007; Silva et al., 2008).木质部导管是影响水分运输的重要参数.根据Hagen-Poiseuille定律, 在其他条件一致时, 水在导管中的导水速率与导管直径的4次方成正比, 即导管直径越大, 导管运输水分的能力越强, 导水率越大(Bass, 1982).大导管高流率体现了导管输水的“有效性”, 但却降低了水分输导的“安全性”, 因为大导管细胞壁薄, 机械强度低, 更容易引发气穴化, 发生阻塞 (Zimmermann, 1983).本研究分析导管细胞的解剖结构发现, 轻度和中度盐胁迫下, 与对照相比, 胡杨幼株导管直径并未显著减小, 而导管密度、管壁厚度、管壁间厚度以及管壁机械强度显著提高; 重度盐胁迫下, 茎导管直径较对照显著下降, 导管密度、管壁厚度、管壁间厚度和管壁机械强度与S2处理差异并不显著.这表明轻度和中度盐胁迫下胡杨茎木质部可能是通过增加导管密度来消除了导管直径变小的弊端, 保持了木质部的输水效率, 同时通过增加管壁厚度和机械强度提高了水分输导的安全性, 从而保证了胡杨的正常生长; 但重度盐胁迫下胡杨幼株木质部栓塞明显增强, 输水有效性和安全性显著下降, 木质部水分传导的自我适应和调控能力显著减弱, 导致木质部水分传导速率急速下降. ...
A PIPı aquaporin contributes to hydrostatic pressure-induced water transport in both the root and rosette of
1
2010
... 研究表明, 植物根可以通过改变细胞内渗透性来抵御一定程度的土壤盐胁迫.一种途径是增强质膜内水通道蛋白的数量和活性(Qi et al., 2009; Calvo-Polanco et al., 2014), 从而增强细胞膜渗透性, 调节根系的水分吸收能力(García-Sánchez et al., 2000; Postaire et al., 2010).另一种途径是通过在植物体内累积脯氨酸、可溶性糖、甘氨酸、甜菜碱、甘露醇等代谢物的含量来平衡细胞质的水势, 减缓渗透胁迫(Aroca et al., 2012; Ruiz-Lozano et al., 2012; Calvo-Polanco et al., 2014), 从而让植物适应盐胁迫.尽管有很多学者就NaCl胁迫对植物根水力导度的影响进行了研究, 但大多数的研究都集中在短期的轻度盐胁迫实验处理(从几分钟到24 h或几天), 且都是针对根和根内细胞-细胞间的水分传导特征(细胞内物质积累和基因表达)研究(Guo et al., 2006; Horie et al., 2011; Muries et al., 2011; Sutka et al., 2011; Calvo-Polanco et al., 2014).对于长期盐胁迫, 尤其是高浓度盐胁迫下, 植物的根、茎、叶整个水分传导通道(木质部)的响应和形态适应研究较少.本研究以中国西北干旱、盐渍化区荒漠河岸林建群种胡杨(Populus euphratica) 2-3年生幼株为研究对象, 系统研究了轻度、中度和重度盐胁迫下胡杨根、茎、叶水分传输通道(木质部)的响应与适应, 以期为盐渍化区域的荒漠河岸林保护、恢复与重建提供理论基础. ...
Increase in aquaporin activity is involved in leaf succulence of the euhalophyte Suaeda salsa, under salinity
3
2009
... 研究表明, 植物根可以通过改变细胞内渗透性来抵御一定程度的土壤盐胁迫.一种途径是增强质膜内水通道蛋白的数量和活性(Qi et al., 2009; Calvo-Polanco et al., 2014), 从而增强细胞膜渗透性, 调节根系的水分吸收能力(García-Sánchez et al., 2000; Postaire et al., 2010).另一种途径是通过在植物体内累积脯氨酸、可溶性糖、甘氨酸、甜菜碱、甘露醇等代谢物的含量来平衡细胞质的水势, 减缓渗透胁迫(Aroca et al., 2012; Ruiz-Lozano et al., 2012; Calvo-Polanco et al., 2014), 从而让植物适应盐胁迫.尽管有很多学者就NaCl胁迫对植物根水力导度的影响进行了研究, 但大多数的研究都集中在短期的轻度盐胁迫实验处理(从几分钟到24 h或几天), 且都是针对根和根内细胞-细胞间的水分传导特征(细胞内物质积累和基因表达)研究(Guo et al., 2006; Horie et al., 2011; Muries et al., 2011; Sutka et al., 2011; Calvo-Polanco et al., 2014).对于长期盐胁迫, 尤其是高浓度盐胁迫下, 植物的根、茎、叶整个水分传导通道(木质部)的响应和形态适应研究较少.本研究以中国西北干旱、盐渍化区荒漠河岸林建群种胡杨(Populus euphratica) 2-3年生幼株为研究对象, 系统研究了轻度、中度和重度盐胁迫下胡杨根、茎、叶水分传输通道(木质部)的响应与适应, 以期为盐渍化区域的荒漠河岸林保护、恢复与重建提供理论基础. ...
... 细根和根尖的数量与胡杨根系导水效率成正相关(表1).根系生长受限降低了根系吸收面积, 加之根系存储Na+相对较多, 木质部汁液中积累了较高浓度的Cl-, 导致离子毒害的发生, 此外土壤盐胁迫也降低了土壤水势, 这将进一步减弱根系的吸水和传输能力(Martínez-Ballesta et al., 2000; Apostol et al., 2004; Navarro et al., 2007; 杨启良等, 2009; 姚立民等, 2011; Calvo-Polanco et al., 2014).植物水分传输有两种途径, 一种是细胞-细胞间的水分传输途径, 一种是木质部内水分传输途径.大量研究证实, 盐胁迫下根细胞内通道蛋白活性和数量以及渗透物质积累增加(Qi et al., 2009; Aroca et al., 2012; Ruiz-Lozano et al., 2012; Calvo-Polanco et al., 2014), 但也有研究表明盐胁迫下根细胞内水通道蛋白显著受抑, 根系导水率与根系水通道蛋白的含量出现一致下调(Martínez-Ballesta et al., 2003, 2008).我们的研究结果表明, 盐胁迫显著降低了根木质部初始导水率和潜在最大导水率, 且随着盐分浓度的增加, 潜在导水率与初始导水率间的差异减小(图2), 即盐胁迫缩小了根木质部对水分传导率的调节范围.盐胁迫使根更易发生栓塞, 显著加剧了木质部栓塞脆弱性(图3), 减弱了木质部对逆境的抵御能力.这表明盐胁迫可能抑制了胡杨幼株根细胞内的水通道蛋白的活动(数量和活性)和渗透物质的积累, 并限制了胡杨幼株根系生长, 导致根木质部导水速率的 降低. ...
... 茎木质部是植物将根部水分运输至叶片的重要通道.三种浓度土壤盐胁迫处理下胡杨幼株茎木质部栓塞程度均受到明显影响, 但胡杨幼株的水分传导速率仅在重度盐胁迫下显著下降, 在轻度和中度盐胁迫下与对照相比却并没有发生显著变化.这表明, 在轻度和中度盐胁迫下, 胡杨茎木质部具有一定的自我调节能力.这可能与细胞内水通道蛋白数量和活性或渗透物质积累有关(Maurel, 1997; Zhang et al., 2003; Qi et al., 2009; Aroca et al., 2012; Ruiz-Lozano et al., 2012; Calvo-Polanco et al., 2014).但也有研究表明, 盐胁迫还可以通过改变细胞形态来调节水分传导(López-Pérez et al., 2007; Navarro et al., 2007; Silva et al., 2008).木质部导管是影响水分运输的重要参数.根据Hagen-Poiseuille定律, 在其他条件一致时, 水在导管中的导水速率与导管直径的4次方成正比, 即导管直径越大, 导管运输水分的能力越强, 导水率越大(Bass, 1982).大导管高流率体现了导管输水的“有效性”, 但却降低了水分输导的“安全性”, 因为大导管细胞壁薄, 机械强度低, 更容易引发气穴化, 发生阻塞 (Zimmermann, 1983).本研究分析导管细胞的解剖结构发现, 轻度和中度盐胁迫下, 与对照相比, 胡杨幼株导管直径并未显著减小, 而导管密度、管壁厚度、管壁间厚度以及管壁机械强度显著提高; 重度盐胁迫下, 茎导管直径较对照显著下降, 导管密度、管壁厚度、管壁间厚度和管壁机械强度与S2处理差异并不显著.这表明轻度和中度盐胁迫下胡杨茎木质部可能是通过增加导管密度来消除了导管直径变小的弊端, 保持了木质部的输水效率, 同时通过增加管壁厚度和机械强度提高了水分输导的安全性, 从而保证了胡杨的正常生长; 但重度盐胁迫下胡杨幼株木质部栓塞明显增强, 输水有效性和安全性显著下降, 木质部水分传导的自我适应和调控能力显著减弱, 导致木质部水分传导速率急速下降. ...
Stomatal conductance as a screen for osmotic stress tolerance in durum wheat growing in saline soil
1
2010
... 全世界盐渍化土壤面积约有10 × 108 hm2, 占世界陆地面积的7.6%, 而中国是世界盐碱地大国之一, 盐渍土面积约0.27 × 108 hm2, 大多分布在中国西北干旱区, 且这一面积随着人类的不合理灌溉正不断地扩大(高光林等, 2003).土壤盐渍化已成为影响植物生长发育的重要环境限制因素(Evelin et al., 2009).一方面, 土壤盐胁迫会降低土壤水势, 发生渗透胁迫(Munns, 2002), 促使根系产生脱落酸传递到叶片(Kang & Zhang, 2004), 诱导植物发生水分胁迫, 使叶片气孔部分关闭(Rahnama et al., 2010), 减少蒸腾(Schachtman & Goodger, 2008), 降低根系吸水能力(Navarro et al., 2007; Aroca et al., 2012; Calvo-Polanco et al., 2014), 从而导致植物叶片同化能力的降低和植物总光合面积的下降(Nastou et al., 1999; 杨启良等, 2009), 最终抑制植物生长.另一方面, 土壤盐胁迫也会通过离子毒害来影响酶活性、脂质代谢以及蛋白质合成等植物的代谢过程来抑制植物生长(Munns & Tester, 2008).盐胁迫下, 植物根系会缺氧(Haeussler & Kabzems, 2006; Mariani et al., 2006), 造成更多NaCl进入根系(Redfield et al., 2003), 促使根系和冠层中的Na+、Cl-浓度不断积累 (Mansour, 1997; Apostol & Zwiazek, 2003).高浓度Na+可置换质膜和细胞内膜系统所结合的Ca2+, 使膜结构完整性及膜功能受到破坏(Boursiac et al., 2008), 从而影响一系列生理代谢功能.另外, 高浓度Na+还会阻碍植物对K+和Ca2+的吸收和运输, 高浓度Cl-会抑制植物对NO-3和HPO42-的吸收, 造成植物营养吸收不平衡, 使碳水化合物不能顺利地被运输(张新春等, 2002; Apostol et al., 2004; López- Pérez et al., 2007), 从而抑制植物生长发育. ...
Responses of red-osier dogwood to oil sands tailings treated with gypsum or alum
1
2003
... 全世界盐渍化土壤面积约有10 × 108 hm2, 占世界陆地面积的7.6%, 而中国是世界盐碱地大国之一, 盐渍土面积约0.27 × 108 hm2, 大多分布在中国西北干旱区, 且这一面积随着人类的不合理灌溉正不断地扩大(高光林等, 2003).土壤盐渍化已成为影响植物生长发育的重要环境限制因素(Evelin et al., 2009).一方面, 土壤盐胁迫会降低土壤水势, 发生渗透胁迫(Munns, 2002), 促使根系产生脱落酸传递到叶片(Kang & Zhang, 2004), 诱导植物发生水分胁迫, 使叶片气孔部分关闭(Rahnama et al., 2010), 减少蒸腾(Schachtman & Goodger, 2008), 降低根系吸水能力(Navarro et al., 2007; Aroca et al., 2012; Calvo-Polanco et al., 2014), 从而导致植物叶片同化能力的降低和植物总光合面积的下降(Nastou et al., 1999; 杨启良等, 2009), 最终抑制植物生长.另一方面, 土壤盐胁迫也会通过离子毒害来影响酶活性、脂质代谢以及蛋白质合成等植物的代谢过程来抑制植物生长(Munns & Tester, 2008).盐胁迫下, 植物根系会缺氧(Haeussler & Kabzems, 2006; Mariani et al., 2006), 造成更多NaCl进入根系(Redfield et al., 2003), 促使根系和冠层中的Na+、Cl-浓度不断积累 (Mansour, 1997; Apostol & Zwiazek, 2003).高浓度Na+可置换质膜和细胞内膜系统所结合的Ca2+, 使膜结构完整性及膜功能受到破坏(Boursiac et al., 2008), 从而影响一系列生理代谢功能.另外, 高浓度Na+还会阻碍植物对K+和Ca2+的吸收和运输, 高浓度Cl-会抑制植物对NO-3和HPO42-的吸收, 造成植物营养吸收不平衡, 使碳水化合物不能顺利地被运输(张新春等, 2002; Apostol et al., 2004; López- Pérez et al., 2007), 从而抑制植物生长发育. ...
Regulation by arbuscular mycorrhizae of the integrated physiological response to salinity in plants: New challenges in physiological and molecular studies
3
2012
... 研究表明, 植物根可以通过改变细胞内渗透性来抵御一定程度的土壤盐胁迫.一种途径是增强质膜内水通道蛋白的数量和活性(Qi et al., 2009; Calvo-Polanco et al., 2014), 从而增强细胞膜渗透性, 调节根系的水分吸收能力(García-Sánchez et al., 2000; Postaire et al., 2010).另一种途径是通过在植物体内累积脯氨酸、可溶性糖、甘氨酸、甜菜碱、甘露醇等代谢物的含量来平衡细胞质的水势, 减缓渗透胁迫(Aroca et al., 2012; Ruiz-Lozano et al., 2012; Calvo-Polanco et al., 2014), 从而让植物适应盐胁迫.尽管有很多学者就NaCl胁迫对植物根水力导度的影响进行了研究, 但大多数的研究都集中在短期的轻度盐胁迫实验处理(从几分钟到24 h或几天), 且都是针对根和根内细胞-细胞间的水分传导特征(细胞内物质积累和基因表达)研究(Guo et al., 2006; Horie et al., 2011; Muries et al., 2011; Sutka et al., 2011; Calvo-Polanco et al., 2014).对于长期盐胁迫, 尤其是高浓度盐胁迫下, 植物的根、茎、叶整个水分传导通道(木质部)的响应和形态适应研究较少.本研究以中国西北干旱、盐渍化区荒漠河岸林建群种胡杨(Populus euphratica) 2-3年生幼株为研究对象, 系统研究了轻度、中度和重度盐胁迫下胡杨根、茎、叶水分传输通道(木质部)的响应与适应, 以期为盐渍化区域的荒漠河岸林保护、恢复与重建提供理论基础. ...
... 细根和根尖的数量与胡杨根系导水效率成正相关(表1).根系生长受限降低了根系吸收面积, 加之根系存储Na+相对较多, 木质部汁液中积累了较高浓度的Cl-, 导致离子毒害的发生, 此外土壤盐胁迫也降低了土壤水势, 这将进一步减弱根系的吸水和传输能力(Martínez-Ballesta et al., 2000; Apostol et al., 2004; Navarro et al., 2007; 杨启良等, 2009; 姚立民等, 2011; Calvo-Polanco et al., 2014).植物水分传输有两种途径, 一种是细胞-细胞间的水分传输途径, 一种是木质部内水分传输途径.大量研究证实, 盐胁迫下根细胞内通道蛋白活性和数量以及渗透物质积累增加(Qi et al., 2009; Aroca et al., 2012; Ruiz-Lozano et al., 2012; Calvo-Polanco et al., 2014), 但也有研究表明盐胁迫下根细胞内水通道蛋白显著受抑, 根系导水率与根系水通道蛋白的含量出现一致下调(Martínez-Ballesta et al., 2003, 2008).我们的研究结果表明, 盐胁迫显著降低了根木质部初始导水率和潜在最大导水率, 且随着盐分浓度的增加, 潜在导水率与初始导水率间的差异减小(图2), 即盐胁迫缩小了根木质部对水分传导率的调节范围.盐胁迫使根更易发生栓塞, 显著加剧了木质部栓塞脆弱性(图3), 减弱了木质部对逆境的抵御能力.这表明盐胁迫可能抑制了胡杨幼株根细胞内的水通道蛋白的活动(数量和活性)和渗透物质的积累, 并限制了胡杨幼株根系生长, 导致根木质部导水速率的 降低. ...
... 茎木质部是植物将根部水分运输至叶片的重要通道.三种浓度土壤盐胁迫处理下胡杨幼株茎木质部栓塞程度均受到明显影响, 但胡杨幼株的水分传导速率仅在重度盐胁迫下显著下降, 在轻度和中度盐胁迫下与对照相比却并没有发生显著变化.这表明, 在轻度和中度盐胁迫下, 胡杨茎木质部具有一定的自我调节能力.这可能与细胞内水通道蛋白数量和活性或渗透物质积累有关(Maurel, 1997; Zhang et al., 2003; Qi et al., 2009; Aroca et al., 2012; Ruiz-Lozano et al., 2012; Calvo-Polanco et al., 2014).但也有研究表明, 盐胁迫还可以通过改变细胞形态来调节水分传导(López-Pérez et al., 2007; Navarro et al., 2007; Silva et al., 2008).木质部导管是影响水分运输的重要参数.根据Hagen-Poiseuille定律, 在其他条件一致时, 水在导管中的导水速率与导管直径的4次方成正比, 即导管直径越大, 导管运输水分的能力越强, 导水率越大(Bass, 1982).大导管高流率体现了导管输水的“有效性”, 但却降低了水分输导的“安全性”, 因为大导管细胞壁薄, 机械强度低, 更容易引发气穴化, 发生阻塞 (Zimmermann, 1983).本研究分析导管细胞的解剖结构发现, 轻度和中度盐胁迫下, 与对照相比, 胡杨幼株导管直径并未显著减小, 而导管密度、管壁厚度、管壁间厚度以及管壁机械强度显著提高; 重度盐胁迫下, 茎导管直径较对照显著下降, 导管密度、管壁厚度、管壁间厚度和管壁机械强度与S2处理差异并不显著.这表明轻度和中度盐胁迫下胡杨茎木质部可能是通过增加导管密度来消除了导管直径变小的弊端, 保持了木质部的输水效率, 同时通过增加管壁厚度和机械强度提高了水分输导的安全性, 从而保证了胡杨的正常生长; 但重度盐胁迫下胡杨幼株木质部栓塞明显增强, 输水有效性和安全性显著下降, 木质部水分传导的自我适应和调控能力显著减弱, 导致木质部水分传导速率急速下降. ...
Chemical root to shoot signaling under drought
1
2008
... 全世界盐渍化土壤面积约有10 × 108 hm2, 占世界陆地面积的7.6%, 而中国是世界盐碱地大国之一, 盐渍土面积约0.27 × 108 hm2, 大多分布在中国西北干旱区, 且这一面积随着人类的不合理灌溉正不断地扩大(高光林等, 2003).土壤盐渍化已成为影响植物生长发育的重要环境限制因素(Evelin et al., 2009).一方面, 土壤盐胁迫会降低土壤水势, 发生渗透胁迫(Munns, 2002), 促使根系产生脱落酸传递到叶片(Kang & Zhang, 2004), 诱导植物发生水分胁迫, 使叶片气孔部分关闭(Rahnama et al., 2010), 减少蒸腾(Schachtman & Goodger, 2008), 降低根系吸水能力(Navarro et al., 2007; Aroca et al., 2012; Calvo-Polanco et al., 2014), 从而导致植物叶片同化能力的降低和植物总光合面积的下降(Nastou et al., 1999; 杨启良等, 2009), 最终抑制植物生长.另一方面, 土壤盐胁迫也会通过离子毒害来影响酶活性、脂质代谢以及蛋白质合成等植物的代谢过程来抑制植物生长(Munns & Tester, 2008).盐胁迫下, 植物根系会缺氧(Haeussler & Kabzems, 2006; Mariani et al., 2006), 造成更多NaCl进入根系(Redfield et al., 2003), 促使根系和冠层中的Na+、Cl-浓度不断积累 (Mansour, 1997; Apostol & Zwiazek, 2003).高浓度Na+可置换质膜和细胞内膜系统所结合的Ca2+, 使膜结构完整性及膜功能受到破坏(Boursiac et al., 2008), 从而影响一系列生理代谢功能.另外, 高浓度Na+还会阻碍植物对K+和Ca2+的吸收和运输, 高浓度Cl-会抑制植物对NO-3和HPO42-的吸收, 造成植物营养吸收不平衡, 使碳水化合物不能顺利地被运输(张新春等, 2002; Apostol et al., 2004; López- Pérez et al., 2007), 从而抑制植物生长发育. ...
Osmotic versus toxic effects of NaCl on pepper plants
1
2008
... 茎木质部是植物将根部水分运输至叶片的重要通道.三种浓度土壤盐胁迫处理下胡杨幼株茎木质部栓塞程度均受到明显影响, 但胡杨幼株的水分传导速率仅在重度盐胁迫下显著下降, 在轻度和中度盐胁迫下与对照相比却并没有发生显著变化.这表明, 在轻度和中度盐胁迫下, 胡杨茎木质部具有一定的自我调节能力.这可能与细胞内水通道蛋白数量和活性或渗透物质积累有关(Maurel, 1997; Zhang et al., 2003; Qi et al., 2009; Aroca et al., 2012; Ruiz-Lozano et al., 2012; Calvo-Polanco et al., 2014).但也有研究表明, 盐胁迫还可以通过改变细胞形态来调节水分传导(López-Pérez et al., 2007; Navarro et al., 2007; Silva et al., 2008).木质部导管是影响水分运输的重要参数.根据Hagen-Poiseuille定律, 在其他条件一致时, 水在导管中的导水速率与导管直径的4次方成正比, 即导管直径越大, 导管运输水分的能力越强, 导水率越大(Bass, 1982).大导管高流率体现了导管输水的“有效性”, 但却降低了水分输导的“安全性”, 因为大导管细胞壁薄, 机械强度低, 更容易引发气穴化, 发生阻塞 (Zimmermann, 1983).本研究分析导管细胞的解剖结构发现, 轻度和中度盐胁迫下, 与对照相比, 胡杨幼株导管直径并未显著减小, 而导管密度、管壁厚度、管壁间厚度以及管壁机械强度显著提高; 重度盐胁迫下, 茎导管直径较对照显著下降, 导管密度、管壁厚度、管壁间厚度和管壁机械强度与S2处理差异并不显著.这表明轻度和中度盐胁迫下胡杨茎木质部可能是通过增加导管密度来消除了导管直径变小的弊端, 保持了木质部的输水效率, 同时通过增加管壁厚度和机械强度提高了水分输导的安全性, 从而保证了胡杨的正常生长; 但重度盐胁迫下胡杨幼株木质部栓塞明显增强, 输水有效性和安全性显著下降, 木质部水分传导的自我适应和调控能力显著减弱, 导致木质部水分传导速率急速下降. ...
Hydraulic properties of a mangrove Avicennia germinans as affected by NaCl
 , 李卫红
, 李卫红


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}