Aims Forest trees alter litter inputs, turnover and rhizospheric activities, modify soil physical, chemical and biological properties, and consequently affect soil organic carbon (SOC) storage and carbon sink strength. That how to select appropriate tree species in afforestation, reforestation and management practices is critical to enhancing forest carbon sequestration. The objective of this study was to determine the effects of tree species on SOC density and vertical distributions.Methods A common garden experiment with the same climate, soil, and management history was established in Maoershan Forest Ecosystem Station, Northeast China, in 2004. The experimental design was a completely randomized arrangement with twenty 25 m × 25 m plots, consisting of monocultures of five tree species, including white birch (Betula platyphylla), Manchurian walnut (Juglans mandshurica), Manchurian ash (Fraxinus mandshurica), Dahurian larch (Larix gmelinii), and Mongolian pine (Pinus sylvestris var. mongolica), each with four replicated plots. A decade after the establishment (2013-2014), we measured carbon density and related factors (i.e., bulk density, total nitrogen concentration, microbial biomass carbon, microbial biomass nitrogen, pH value) in soils of the 0-40 cm depth for these monocultures. Important findings Results showed that tree species significantly influenced the SOC density in the 0-40 cm depth (p < 0.05). SOC density in the 0-10 cm depth varied from 2.79 to 3.08 kg·m-2, in the order of walnut > ash> birch > larch > pine, in the 10-20 cm depth from 1.56 to 2.19 kg·m-2, in the order of pine > walnut > ash > birch > larch, in the 20-30 cm depth from 1.17 to 2.10 kg·m-2, and in the 20-40 cm depth from 0.84 to 1.43 kg·m-2. The greatest SOC density occurred in the birch stands in the 20-40 cm depth. The vertical distributions of SOC density varied with tree species. The percentage of SOC in the 0-10 cm depth over the total SOC in the soil profile was significantly higher in the walnut and larch stands than in others, while the percentage of SOC in the 20-40 cm depth over the total SOC was highest in the birch stands. SOC concentration and soil bulk density differed significantly among the stands of different tree species, and were negatively correlated. SOC density was positively correlated with soil microbial biomass and soil pH in the walnut, ash, and larch stands, and with total nitrogen density in all the stands. We conclude that tree species modifies soil properties and microbial activity, thereby influencing SOC density, and that different patterns of vertical distributions of SOC density among monocultures of different tree species may be attributed to varying SOC controls at each soil depth.
Keywords:tree species
;
soil property
;
soil organic carbon
;
soil microbe
;
vertical distribution
WANGXin-Qi, WANGChuan-Kuan, HANYi. Effects of tree species on soil organic carbon density: A common garden experiment of five temperate tree species. Chinese Journal of Plant Ecology, 2015, 39(11): 1033-1043 https://doi.org/10.17521/cjpe.2015.0100
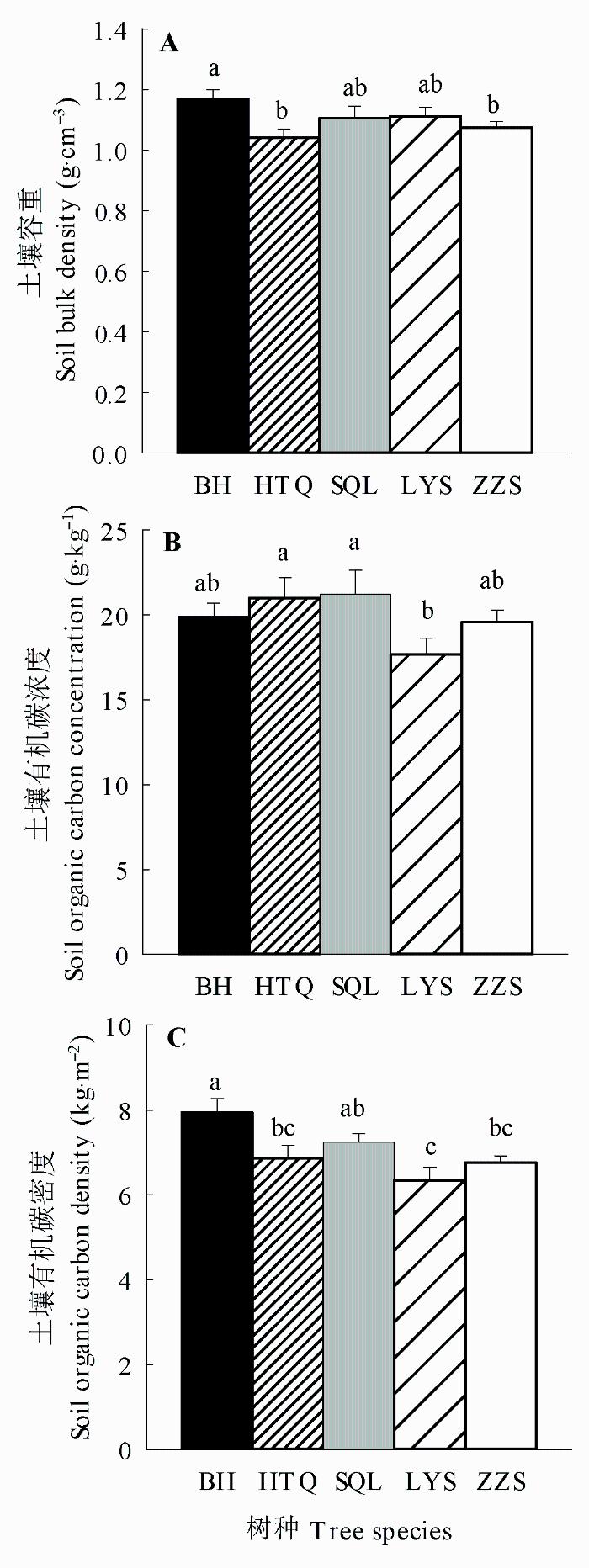
Fig. 1 Comparisons of soil bulk density (A), soil organic carbon concentration (B), and soil organic carbon density (C) among the monocultures of five tree species for 0-40 cm soil layer (mean ± SE). BH, HTQ, LYS, SQL, and ZZS represent Betula platyphylla, Juglans mandshurica, Larix gmelinii, Fraxinus mandshurica, and Pinus sylvestris var. mongolica, respectively. Different lowercase letters indicate significant differences among tree species.
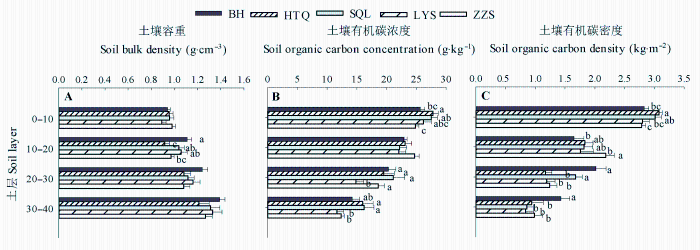
Fig. 2 Comparisons of the vertical changes in soil bulk density (A), soil organic carbon concentration (B), and soil organic carbon density (C) among the monocultures of five tree species (mean ± SE). BH, HTQ, LYS, SQL, and ZZS represent Betula platyphylla, Juglans mandshurica, Larix gmelinii, Fraxinus mandshurica, and Pinus sylvestris var. mongolica, respectively. Different lowercase letters within the same soil layers indicate significant differences among tree species, and the soil layers without designation of letters are not significantly different among groups.
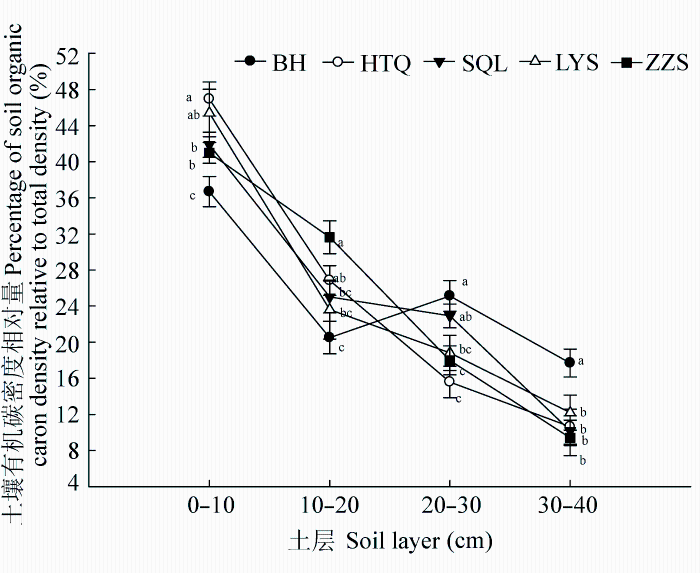
Fig. 3 Vertical changes in the percentage of soil organic carbon in each soil layer over the total soil organic carbon in the monocultures of five tree species (mean ± SE). BH, HTQ, LYS, SQL, and ZZS represent Betula platyphylla, Juglans mandshurica, Larix gmelinii, Fraxinus mandshurica, and Pinus sylvestris var. mongolica, respectively. Different lowercase letters within the same soil layers indicate significant differences among tree species.
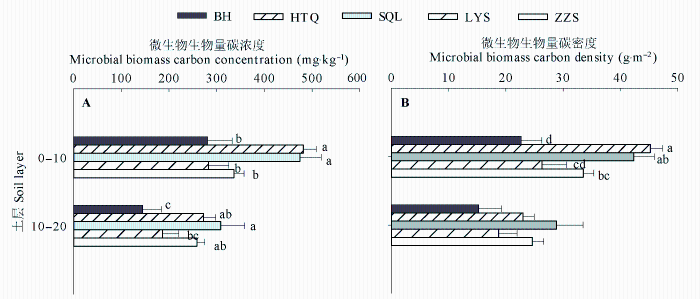
Fig. 4 Comparisons of the vertical changes in soil microbial biomass carbon concentration (A) and density (B) among the monocultures of five tree species (mean ± SE). BH, HTQ, LYS, SQL, and ZZS represent Betula platyphylla, Juglans mandshurica, Larix gmelinii, Fraxinus mandshurica, and Pinus sylvestris var. mongolica, respectively. Different lowercase letters within the same soil layers indicate significant differences among tree species, and the soil layers without designation of letters are not significantly different among groups.
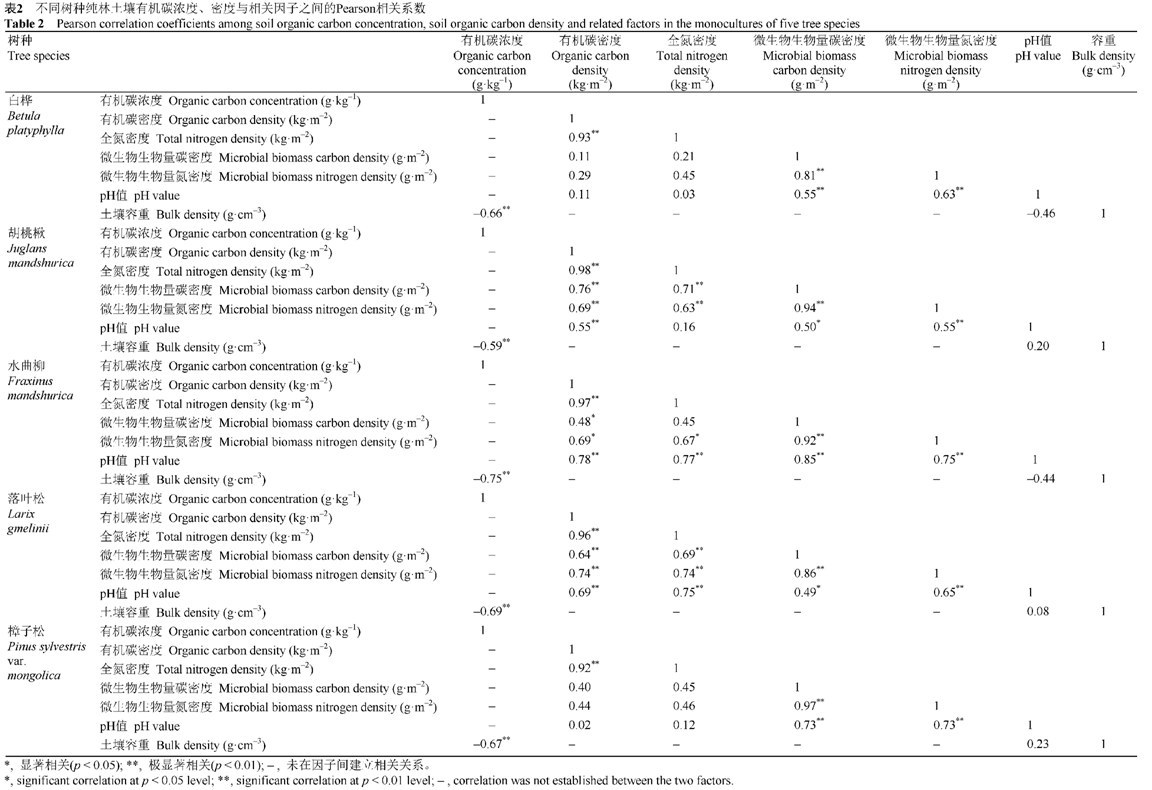
Table2 Pearson correlation coefficients among soil organic carbon concentration,soil organic carbon density and related factors in the monocultures of five tree species
Impact of Tilia platyphyllos Scop., Fraxinus excelsior L., Acer pseudoplatanus L., Quercus robur L. and Fagus sylvatica L. on earthworm biomass and physico-chemical properties of a loamy topsoil.
Studies of the development of woodland conditions under different trees. II. The forest floor.
Journal of Ecology, 42, 71-80.
[26]
OvingtonJD (1956).
Studies of the development of woodland conditions under different trees. IV. The ignition loss, water, carbon and nitrogen content of the mineral soil.
Journal of Ecology, 44, 171-179.
[27]
ParryML, CanzianOF, PalutikofJP, vander Linden P, HansonCE (2007). Climate Change 2007: Impacts, adaptation and vulnerability: Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change Cambridge University Press, Cambridge, UK Impacts, adaptation and vulnerability: Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, UK. 976.
... 我们发现, 阔叶树种0-10 cm土层SOC密度显著高于针叶树种, 树种间SOC密度大小顺序与SOC浓度顺序基本一致(图2B和2C), 说明树种主要通过影响SOC浓度进而造成其SOC密度间的差异.产生这种差异可能与森林凋落物、根系周转代谢产物等有关(Mueller et al., 2012).阔叶树种凋落物质量较高、分解速率快, 从而有更多的腐殖质进入土壤形成SOC (郭忠玲等, 2006; Vesterdal et al., 2008).此外, 阔叶树种土壤中蚯蚓生物量显著高于针叶树种(Reich et al., 2005), 蚯蚓能够将凋落物整合进入矿质土层, 从而提高SOC储量.树种间10-20 cm土层SOC浓度的差异显著性微弱(p = 0.09), 土壤容重差异显著, 但SOC密度大小顺序仍与SOC浓度基本一致(图2B, 2C), 说明树种通过改变SOC浓度和容重进而影响SOC密度.随着土层加深, 地上凋落物对深层土壤的影响逐渐减小, 而根系凋落物对SOC的影响逐渐增大(Rasse et al., 2005).樟子松林10-20 cm土层SOC密度最高, 可能与樟子松林外生菌根主要分布于该土层范围有关(朱教君等, 2007).外生菌根不仅可促进根系对营养物质的吸收, 还可以降低根系的分解速率(Adam Langley et al., 2006), 从而促进与根系有关的SOC储存.白桦林20-40 cm土层SOC密度显著高于其他树种纯林(图2C), 可能是由于先锋树种幼龄期生长力旺盛、对土壤空间利用充分(杨慧等, 2007), 因而积存了较多深层土壤有机质. ...
Effects of tree species, stand age and soil type on soil microbial biomass and its activity in a southern boreal forest.
1
1998
... 土壤酸度增强会限制微生物活性, 从而降低微生物数量(Bauhus et al., 1998).本研究结果显示, SOC密度与土壤微生物生物量呈显著正相关的树种纯林, 其SOC密度与土壤pH值也呈正相关; 否则, 两者之间没有显著的相关关系(表2).因此, 土壤pH值可能是通过微生物而间接影响SOC密度, 而不是通过释放出较多的水解阳离子(铁离子、铝离子)而促进SOC的化学稳定性(Mueller et al., 2012).然而, Mueller等(2012)报道: 土壤总酸度与土壤碳储量显著正相关.产生这种相悖结论的原因可能是, 土壤pH值仅表示土壤总酸度的一部分, 而不代表土壤整体酸度(Thomas, 1996).另外, 本研究中各树种纯林SOC密度与总氮密度均显著正相关(表2), 表明森林土壤碳氮通过土壤有机质紧密联系在一起, 经历相似的机理过程(Gurmesa et al., 2013). ...
Nitrogen-total. In: Page AL, Miller RH, Keeney DR eds.
0
1982
Does tree species composition control soil organic carbon pools in Mediterranean mountain forests?
3
2011
... 森林是陆地生态系统的重要组分, 其植被碳库约占全球植被碳库的80%, 其土壤碳库约占全球土壤碳库的70% (Jandl et al., 2007).土壤碳库因储量大而对森林碳循环过程影响显著, 进而影响生态系统的结构和功能(Poeplau et al., 2011).树种通过改变凋落物的输入(Díaz-Pinés et al., 2011)与输出(Vesterdal et al., 2012)、根系活动(Hansson et al., 2013)等深刻地影响土壤的物理、化学和生物学性质及固碳功能(Mueller et al., 2012).因此, 合理选择树种是造林、再造林、森林经营等碳汇林业中一个亟待解决的理论和实践问题(Parry et al., 2007). ...
... 森林土壤有机碳(SOC)由植物、动物和微生物等各降解阶段的残留混合物组成, 是碳输入和输出的动态平衡结果(Vesterdal et al., 2013).不同树种地上凋落物的数量与质量、地下根系及根系共生体、分泌物等不同, 会改变土壤养分状况、酸度、土壤动物微生物的数量和活性等理化和生物学性质, 从而影响SOC的输入和输出(Hobbie et al., 2007; Díaz-Pinés et al., 2011; Vesterdal et al., 2012).土壤养分状况的改善能提高森林净初级生产量以及凋落物产量, 进而促进SOC的积累(Gurmesa et al., 2013).增加土壤微生物的数量及其活性能提高SOC的稳定性, 促进SOC的积累(Xu et al., 2013).但是, 不同树种通过这些过程对SOC的综合影响程度及这些过程的主要影响因子尚不清晰. ...
... 树种通过地上和地下凋落物的输入量及质量(Hagen-Thorn et al., 2004; Díaz-Pinés et al., 2011)、树木根系周转、分泌物等代谢产物的产生综合影响SOC密度(Hansson et al., 2013).虽然本研究未能测定地上凋落物以及根系碳浓度, 但上述结果表明, 不同凋落物质量导致SOC浓度产生差异, 进而可能直接导致树种间SOC密度差异, 土壤微生物生物量以及土壤pH值则可能间接影响SOC密度. ...
Aluminum solubility and mobility in relation to organic carbon in surface soils affected by six tree species of the northeastern United States.
1
2003
... 以往有关树种对土壤性质影响的研究多采用选择立地相邻、树龄相近、基质相似的样方(Neirynck et al., 2000; Laganière et al., 2012), 或在天然混交林中就各个树种的单独个体进行测定(Dijkstra & Fitzhugh, 2003).因受到林龄、立地条件等相关因素的影响, 这类研究很难阐明树种对土壤理化和生物学性质的效应(Vesterdal et al., 2013).例如, Mareschal等(2010)报道温带阔叶林SOC储量高于针叶林, 而Gurmesa等(2013)则发现针叶林SOC储量高于阔叶林.同样, 我国亚热带地区主要树种对SOC的影响(李强等, 2008; Wang et al., 2010)的研究结论也不一致.其中, 研究方法存在缺陷是导致这些结果不一致的重要原因之一. ...
Calculation of organic matter and nutrients stored in soils under contrasting management regimes.
Soil carbon accumulation and nitrogen retention traits of four tree species grown in common gardens.
2
2013
... 森林土壤有机碳(SOC)由植物、动物和微生物等各降解阶段的残留混合物组成, 是碳输入和输出的动态平衡结果(Vesterdal et al., 2013).不同树种地上凋落物的数量与质量、地下根系及根系共生体、分泌物等不同, 会改变土壤养分状况、酸度、土壤动物微生物的数量和活性等理化和生物学性质, 从而影响SOC的输入和输出(Hobbie et al., 2007; Díaz-Pinés et al., 2011; Vesterdal et al., 2012).土壤养分状况的改善能提高森林净初级生产量以及凋落物产量, 进而促进SOC的积累(Gurmesa et al., 2013).增加土壤微生物的数量及其活性能提高SOC的稳定性, 促进SOC的积累(Xu et al., 2013).但是, 不同树种通过这些过程对SOC的综合影响程度及这些过程的主要影响因子尚不清晰. ...
... 土壤酸度增强会限制微生物活性, 从而降低微生物数量(Bauhus et al., 1998).本研究结果显示, SOC密度与土壤微生物生物量呈显著正相关的树种纯林, 其SOC密度与土壤pH值也呈正相关; 否则, 两者之间没有显著的相关关系(表2).因此, 土壤pH值可能是通过微生物而间接影响SOC密度, 而不是通过释放出较多的水解阳离子(铁离子、铝离子)而促进SOC的化学稳定性(Mueller et al., 2012).然而, Mueller等(2012)报道: 土壤总酸度与土壤碳储量显著正相关.产生这种相悖结论的原因可能是, 土壤pH值仅表示土壤总酸度的一部分, 而不代表土壤整体酸度(Thomas, 1996).另外, 本研究中各树种纯林SOC密度与总氮密度均显著正相关(表2), 表明森林土壤碳氮通过土壤有机质紧密联系在一起, 经历相似的机理过程(Gurmesa et al., 2013). ...
The impact of six European tree species on the chemistry of mineral topsoil in forest plantations on former agricultural land.
1
2004
... 树种通过地上和地下凋落物的输入量及质量(Hagen-Thorn et al., 2004; Díaz-Pinés et al., 2011)、树木根系周转、分泌物等代谢产物的产生综合影响SOC密度(Hansson et al., 2013).虽然本研究未能测定地上凋落物以及根系碳浓度, 但上述结果表明, 不同凋落物质量导致SOC浓度产生差异, 进而可能直接导致树种间SOC密度差异, 土壤微生物生物量以及土壤pH值则可能间接影响SOC密度. ...
Fine root production and turnover of tree and understorey vegetation in Scots pine, silver birch and Norway spruce stands in SW Sweden.
2
2013
... 森林是陆地生态系统的重要组分, 其植被碳库约占全球植被碳库的80%, 其土壤碳库约占全球土壤碳库的70% (Jandl et al., 2007).土壤碳库因储量大而对森林碳循环过程影响显著, 进而影响生态系统的结构和功能(Poeplau et al., 2011).树种通过改变凋落物的输入(Díaz-Pinés et al., 2011)与输出(Vesterdal et al., 2012)、根系活动(Hansson et al., 2013)等深刻地影响土壤的物理、化学和生物学性质及固碳功能(Mueller et al., 2012).因此, 合理选择树种是造林、再造林、森林经营等碳汇林业中一个亟待解决的理论和实践问题(Parry et al., 2007). ...
... 树种通过地上和地下凋落物的输入量及质量(Hagen-Thorn et al., 2004; Díaz-Pinés et al., 2011)、树木根系周转、分泌物等代谢产物的产生综合影响SOC密度(Hansson et al., 2013).虽然本研究未能测定地上凋落物以及根系碳浓度, 但上述结果表明, 不同凋落物质量导致SOC浓度产生差异, 进而可能直接导致树种间SOC密度差异, 土壤微生物生物量以及土壤pH值则可能间接影响SOC密度. ...
Tree species effects on soil organic matter dynamics: The role of soil cation composition.
2
2007
... 森林土壤有机碳(SOC)由植物、动物和微生物等各降解阶段的残留混合物组成, 是碳输入和输出的动态平衡结果(Vesterdal et al., 2013).不同树种地上凋落物的数量与质量、地下根系及根系共生体、分泌物等不同, 会改变土壤养分状况、酸度、土壤动物微生物的数量和活性等理化和生物学性质, 从而影响SOC的输入和输出(Hobbie et al., 2007; Díaz-Pinés et al., 2011; Vesterdal et al., 2012).土壤养分状况的改善能提高森林净初级生产量以及凋落物产量, 进而促进SOC的积累(Gurmesa et al., 2013).增加土壤微生物的数量及其活性能提高SOC的稳定性, 促进SOC的积累(Xu et al., 2013).但是, 不同树种通过这些过程对SOC的综合影响程度及这些过程的主要影响因子尚不清晰. ...
... 早在20世纪50年代, Ovington (1954, 1956)就采用同质园(common garden)试验方法研究不同树种对SOC的影响.之后, 在北美、欧洲等地的温带和北方森林中相继采用同质园试验探讨树种对土壤碳储量、土壤呼吸、土壤养分循环以及土壤理化性质的影响(Hobbie et al., 2007; Mueller et al., 2012; Vesterdal et al., 2012).通过在相同林龄、相同经营历史、相同土壤基质的立地条件下建立同质园试验, 可以最大程度地降低气候和立地条件差异带来的影响(Vesterdal et al., 2013).然而, 同质园试验的建立对立地条件要求高、成本大、试验期限长, 因此, 目前因试验重复量有限、林龄小等问题而难以形成一致的结论. ...
How strongly can forest management influence soil carbon sequestration?
1
2007
... 森林是陆地生态系统的重要组分, 其植被碳库约占全球植被碳库的80%, 其土壤碳库约占全球土壤碳库的70% (Jandl et al., 2007).土壤碳库因储量大而对森林碳循环过程影响显著, 进而影响生态系统的结构和功能(Poeplau et al., 2011).树种通过改变凋落物的输入(Díaz-Pinés et al., 2011)与输出(Vesterdal et al., 2012)、根系活动(Hansson et al., 2013)等深刻地影响土壤的物理、化学和生物学性质及固碳功能(Mueller et al., 2012).因此, 合理选择树种是造林、再造林、森林经营等碳汇林业中一个亟待解决的理论和实践问题(Parry et al., 2007). ...
Humic substances in soils: Are they really chemically distinct?
Tree species effects on coupled cycles of carbon, nitrogen, and acidity in mineral soils at a common garden experiment.
4
2012
... 森林是陆地生态系统的重要组分, 其植被碳库约占全球植被碳库的80%, 其土壤碳库约占全球土壤碳库的70% (Jandl et al., 2007).土壤碳库因储量大而对森林碳循环过程影响显著, 进而影响生态系统的结构和功能(Poeplau et al., 2011).树种通过改变凋落物的输入(Díaz-Pinés et al., 2011)与输出(Vesterdal et al., 2012)、根系活动(Hansson et al., 2013)等深刻地影响土壤的物理、化学和生物学性质及固碳功能(Mueller et al., 2012).因此, 合理选择树种是造林、再造林、森林经营等碳汇林业中一个亟待解决的理论和实践问题(Parry et al., 2007). ...
... 早在20世纪50年代, Ovington (1954, 1956)就采用同质园(common garden)试验方法研究不同树种对SOC的影响.之后, 在北美、欧洲等地的温带和北方森林中相继采用同质园试验探讨树种对土壤碳储量、土壤呼吸、土壤养分循环以及土壤理化性质的影响(Hobbie et al., 2007; Mueller et al., 2012; Vesterdal et al., 2012).通过在相同林龄、相同经营历史、相同土壤基质的立地条件下建立同质园试验, 可以最大程度地降低气候和立地条件差异带来的影响(Vesterdal et al., 2013).然而, 同质园试验的建立对立地条件要求高、成本大、试验期限长, 因此, 目前因试验重复量有限、林龄小等问题而难以形成一致的结论. ...
... 我们发现, 阔叶树种0-10 cm土层SOC密度显著高于针叶树种, 树种间SOC密度大小顺序与SOC浓度顺序基本一致(图2B和2C), 说明树种主要通过影响SOC浓度进而造成其SOC密度间的差异.产生这种差异可能与森林凋落物、根系周转代谢产物等有关(Mueller et al., 2012).阔叶树种凋落物质量较高、分解速率快, 从而有更多的腐殖质进入土壤形成SOC (郭忠玲等, 2006; Vesterdal et al., 2008).此外, 阔叶树种土壤中蚯蚓生物量显著高于针叶树种(Reich et al., 2005), 蚯蚓能够将凋落物整合进入矿质土层, 从而提高SOC储量.树种间10-20 cm土层SOC浓度的差异显著性微弱(p = 0.09), 土壤容重差异显著, 但SOC密度大小顺序仍与SOC浓度基本一致(图2B, 2C), 说明树种通过改变SOC浓度和容重进而影响SOC密度.随着土层加深, 地上凋落物对深层土壤的影响逐渐减小, 而根系凋落物对SOC的影响逐渐增大(Rasse et al., 2005).樟子松林10-20 cm土层SOC密度最高, 可能与樟子松林外生菌根主要分布于该土层范围有关(朱教君等, 2007).外生菌根不仅可促进根系对营养物质的吸收, 还可以降低根系的分解速率(Adam Langley et al., 2006), 从而促进与根系有关的SOC储存.白桦林20-40 cm土层SOC密度显著高于其他树种纯林(图2C), 可能是由于先锋树种幼龄期生长力旺盛、对土壤空间利用充分(杨慧等, 2007), 因而积存了较多深层土壤有机质. ...
... 土壤酸度增强会限制微生物活性, 从而降低微生物数量(Bauhus et al., 1998).本研究结果显示, SOC密度与土壤微生物生物量呈显著正相关的树种纯林, 其SOC密度与土壤pH值也呈正相关; 否则, 两者之间没有显著的相关关系(表2).因此, 土壤pH值可能是通过微生物而间接影响SOC密度, 而不是通过释放出较多的水解阳离子(铁离子、铝离子)而促进SOC的化学稳定性(Mueller et al., 2012).然而, Mueller等(2012)报道: 土壤总酸度与土壤碳储量显著正相关.产生这种相悖结论的原因可能是, 土壤pH值仅表示土壤总酸度的一部分, 而不代表土壤整体酸度(Thomas, 1996).另外, 本研究中各树种纯林SOC密度与总氮密度均显著正相关(表2), 表明森林土壤碳氮通过土壤有机质紧密联系在一起, 经历相似的机理过程(Gurmesa et al., 2013). ...
Impact of Tilia platyphyllos Scop., Fraxinus excelsior L., Acer pseudoplatanus L., Quercus robur L. and Fagus sylvatica L. on earthworm biomass and physico-chemical properties of a loamy topsoil.
1
2000
... 以往有关树种对土壤性质影响的研究多采用选择立地相邻、树龄相近、基质相似的样方(Neirynck et al., 2000; Laganière et al., 2012), 或在天然混交林中就各个树种的单独个体进行测定(Dijkstra & Fitzhugh, 2003).因受到林龄、立地条件等相关因素的影响, 这类研究很难阐明树种对土壤理化和生物学性质的效应(Vesterdal et al., 2013).例如, Mareschal等(2010)报道温带阔叶林SOC储量高于针叶林, 而Gurmesa等(2013)则发现针叶林SOC储量高于阔叶林.同样, 我国亚热带地区主要树种对SOC的影响(李强等, 2008; Wang et al., 2010)的研究结论也不一致.其中, 研究方法存在缺陷是导致这些结果不一致的重要原因之一. ...
Studies of the development of woodland conditions under different trees. II. The forest floor.
0
1954
Studies of the development of woodland conditions under different trees. IV. The ignition loss, water, carbon and nitrogen content of the mineral soil.
0
1956
1
2007
... 森林是陆地生态系统的重要组分, 其植被碳库约占全球植被碳库的80%, 其土壤碳库约占全球土壤碳库的70% (Jandl et al., 2007).土壤碳库因储量大而对森林碳循环过程影响显著, 进而影响生态系统的结构和功能(Poeplau et al., 2011).树种通过改变凋落物的输入(Díaz-Pinés et al., 2011)与输出(Vesterdal et al., 2012)、根系活动(Hansson et al., 2013)等深刻地影响土壤的物理、化学和生物学性质及固碳功能(Mueller et al., 2012).因此, 合理选择树种是造林、再造林、森林经营等碳汇林业中一个亟待解决的理论和实践问题(Parry et al., 2007). ...
Temporal dynamics of soil organic carbon after land-use change in the temperate zone—Carbon response functions as a model approach.
2
2011
... 森林是陆地生态系统的重要组分, 其植被碳库约占全球植被碳库的80%, 其土壤碳库约占全球土壤碳库的70% (Jandl et al., 2007).土壤碳库因储量大而对森林碳循环过程影响显著, 进而影响生态系统的结构和功能(Poeplau et al., 2011).树种通过改变凋落物的输入(Díaz-Pinés et al., 2011)与输出(Vesterdal et al., 2012)、根系活动(Hansson et al., 2013)等深刻地影响土壤的物理、化学和生物学性质及固碳功能(Mueller et al., 2012).因此, 合理选择树种是造林、再造林、森林经营等碳汇林业中一个亟待解决的理论和实践问题(Parry et al., 2007). ...
Do tree species influence soil carbon stocks in temperate and boreal forests?
3
2013
... 森林土壤有机碳(SOC)由植物、动物和微生物等各降解阶段的残留混合物组成, 是碳输入和输出的动态平衡结果(Vesterdal et al., 2013).不同树种地上凋落物的数量与质量、地下根系及根系共生体、分泌物等不同, 会改变土壤养分状况、酸度、土壤动物微生物的数量和活性等理化和生物学性质, 从而影响SOC的输入和输出(Hobbie et al., 2007; Díaz-Pinés et al., 2011; Vesterdal et al., 2012).土壤养分状况的改善能提高森林净初级生产量以及凋落物产量, 进而促进SOC的积累(Gurmesa et al., 2013).增加土壤微生物的数量及其活性能提高SOC的稳定性, 促进SOC的积累(Xu et al., 2013).但是, 不同树种通过这些过程对SOC的综合影响程度及这些过程的主要影响因子尚不清晰. ...
... 以往有关树种对土壤性质影响的研究多采用选择立地相邻、树龄相近、基质相似的样方(Neirynck et al., 2000; Laganière et al., 2012), 或在天然混交林中就各个树种的单独个体进行测定(Dijkstra & Fitzhugh, 2003).因受到林龄、立地条件等相关因素的影响, 这类研究很难阐明树种对土壤理化和生物学性质的效应(Vesterdal et al., 2013).例如, Mareschal等(2010)报道温带阔叶林SOC储量高于针叶林, 而Gurmesa等(2013)则发现针叶林SOC储量高于阔叶林.同样, 我国亚热带地区主要树种对SOC的影响(李强等, 2008; Wang et al., 2010)的研究结论也不一致.其中, 研究方法存在缺陷是导致这些结果不一致的重要原因之一. ...
... 早在20世纪50年代, Ovington (1954, 1956)就采用同质园(common garden)试验方法研究不同树种对SOC的影响.之后, 在北美、欧洲等地的温带和北方森林中相继采用同质园试验探讨树种对土壤碳储量、土壤呼吸、土壤养分循环以及土壤理化性质的影响(Hobbie et al., 2007; Mueller et al., 2012; Vesterdal et al., 2012).通过在相同林龄、相同经营历史、相同土壤基质的立地条件下建立同质园试验, 可以最大程度地降低气候和立地条件差异带来的影响(Vesterdal et al., 2013).然而, 同质园试验的建立对立地条件要求高、成本大、试验期限长, 因此, 目前因试验重复量有限、林龄小等问题而难以形成一致的结论. ...
Soil respiration and rates of soil carbon turnover differ among six common European tree species.
3
2012
... 森林是陆地生态系统的重要组分, 其植被碳库约占全球植被碳库的80%, 其土壤碳库约占全球土壤碳库的70% (Jandl et al., 2007).土壤碳库因储量大而对森林碳循环过程影响显著, 进而影响生态系统的结构和功能(Poeplau et al., 2011).树种通过改变凋落物的输入(Díaz-Pinés et al., 2011)与输出(Vesterdal et al., 2012)、根系活动(Hansson et al., 2013)等深刻地影响土壤的物理、化学和生物学性质及固碳功能(Mueller et al., 2012).因此, 合理选择树种是造林、再造林、森林经营等碳汇林业中一个亟待解决的理论和实践问题(Parry et al., 2007). ...
... 森林土壤有机碳(SOC)由植物、动物和微生物等各降解阶段的残留混合物组成, 是碳输入和输出的动态平衡结果(Vesterdal et al., 2013).不同树种地上凋落物的数量与质量、地下根系及根系共生体、分泌物等不同, 会改变土壤养分状况、酸度、土壤动物微生物的数量和活性等理化和生物学性质, 从而影响SOC的输入和输出(Hobbie et al., 2007; Díaz-Pinés et al., 2011; Vesterdal et al., 2012).土壤养分状况的改善能提高森林净初级生产量以及凋落物产量, 进而促进SOC的积累(Gurmesa et al., 2013).增加土壤微生物的数量及其活性能提高SOC的稳定性, 促进SOC的积累(Xu et al., 2013).但是, 不同树种通过这些过程对SOC的综合影响程度及这些过程的主要影响因子尚不清晰. ...
... 早在20世纪50年代, Ovington (1954, 1956)就采用同质园(common garden)试验方法研究不同树种对SOC的影响.之后, 在北美、欧洲等地的温带和北方森林中相继采用同质园试验探讨树种对土壤碳储量、土壤呼吸、土壤养分循环以及土壤理化性质的影响(Hobbie et al., 2007; Mueller et al., 2012; Vesterdal et al., 2012).通过在相同林龄、相同经营历史、相同土壤基质的立地条件下建立同质园试验, 可以最大程度地降低气候和立地条件差异带来的影响(Vesterdal et al., 2013).然而, 同质园试验的建立对立地条件要求高、成本大、试验期限长, 因此, 目前因试验重复量有限、林龄小等问题而难以形成一致的结论. ...
Carbon and nitrogen in forest floor and mineral soil under six common European tree species.
1
2008
... 我们发现, 阔叶树种0-10 cm土层SOC密度显著高于针叶树种, 树种间SOC密度大小顺序与SOC浓度顺序基本一致(图2B和2C), 说明树种主要通过影响SOC浓度进而造成其SOC密度间的差异.产生这种差异可能与森林凋落物、根系周转代谢产物等有关(Mueller et al., 2012).阔叶树种凋落物质量较高、分解速率快, 从而有更多的腐殖质进入土壤形成SOC (郭忠玲等, 2006; Vesterdal et al., 2008).此外, 阔叶树种土壤中蚯蚓生物量显著高于针叶树种(Reich et al., 2005), 蚯蚓能够将凋落物整合进入矿质土层, 从而提高SOC储量.树种间10-20 cm土层SOC浓度的差异显著性微弱(p = 0.09), 土壤容重差异显著, 但SOC密度大小顺序仍与SOC浓度基本一致(图2B, 2C), 说明树种通过改变SOC浓度和容重进而影响SOC密度.随着土层加深, 地上凋落物对深层土壤的影响逐渐减小, 而根系凋落物对SOC的影响逐渐增大(Rasse et al., 2005).樟子松林10-20 cm土层SOC密度最高, 可能与樟子松林外生菌根主要分布于该土层范围有关(朱教君等, 2007).外生菌根不仅可促进根系对营养物质的吸收, 还可以降低根系的分解速率(Adam Langley et al., 2006), 从而促进与根系有关的SOC储存.白桦林20-40 cm土层SOC密度显著高于其他树种纯林(图2C), 可能是由于先锋树种幼龄期生长力旺盛、对土壤空间利用充分(杨慧等, 2007), 因而积存了较多深层土壤有机质. ...
Soil organic carbon stock and chemical composition in four plantations of indigenous tree species in subtropical China.
1
2010
... 以往有关树种对土壤性质影响的研究多采用选择立地相邻、树龄相近、基质相似的样方(Neirynck et al., 2000; Laganière et al., 2012), 或在天然混交林中就各个树种的单独个体进行测定(Dijkstra & Fitzhugh, 2003).因受到林龄、立地条件等相关因素的影响, 这类研究很难阐明树种对土壤理化和生物学性质的效应(Vesterdal et al., 2013).例如, Mareschal等(2010)报道温带阔叶林SOC储量高于针叶林, 而Gurmesa等(2013)则发现针叶林SOC储量高于阔叶林.同样, 我国亚热带地区主要树种对SOC的影响(李强等, 2008; Wang et al., 2010)的研究结论也不一致.其中, 研究方法存在缺陷是导致这些结果不一致的重要原因之一. ...
What is the impact of afforestation on the carbon stocks of Irish mineral soils?
A global analysis of soil microbial biomass carbon, nitrogen and phosphorus in terrestrial ecosystems.
3
2013
... 森林土壤有机碳(SOC)由植物、动物和微生物等各降解阶段的残留混合物组成, 是碳输入和输出的动态平衡结果(Vesterdal et al., 2013).不同树种地上凋落物的数量与质量、地下根系及根系共生体、分泌物等不同, 会改变土壤养分状况、酸度、土壤动物微生物的数量和活性等理化和生物学性质, 从而影响SOC的输入和输出(Hobbie et al., 2007; Díaz-Pinés et al., 2011; Vesterdal et al., 2012).土壤养分状况的改善能提高森林净初级生产量以及凋落物产量, 进而促进SOC的积累(Gurmesa et al., 2013).增加土壤微生物的数量及其活性能提高SOC的稳定性, 促进SOC的积累(Xu et al., 2013).但是, 不同树种通过这些过程对SOC的综合影响程度及这些过程的主要影响因子尚不清晰. ...
 , 韩轶
, 韩轶


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}