植物根系作为植被与土壤之间物质、能量交换的主要通道(黄晶晶, 2013 ), 在植物水分和养分吸收、运移过程中有着不可替代的作用(Kong et al ., 2014 ), 影响着地上部分以及整个植株的生长和发育, 其中根系的分支状况和构型对营养物质的吸收起着关键作用(Dannowski & Block, 2005 )。根系构型是植物与环境异质性资源相互适应的结果(Malamy, 2005 ), 其对环境的适应是通过根系分支结构、空间分布格局等构型特征的改变表现出来的(Tracy et al ., 2015 )。为了应对地理环境和资源供给水平造成的胁迫和干扰, 植物根系会根据所处生境的资源条件适时调整和转化根系分叉数和分支角度等构型特征, 实现植物功能性状间的资源权衡(陈莹婷和许振柱, 2014 )。根系分叉数影响植物根系对资源物质的觅食能力和植株的固定作用(孙浩燕等, 2014 ), 根系分支角度与根系的分布范围和拓展能力紧密相关(黄晶晶, 2013 ), 二者在很大程度上决定了根系在土壤中的位置和对土体的觅养效率(Malamy, 2005 ), 反映了不同生境中植物根系在物质吸收、利用和循环中的作用。地形作为生境条件的综合指示, 控制水热因子的重新分配, 修饰植被空间分布格局, 此自然法则具有普适性(常学礼等, 2015 )。坡向是地形重要的影响因素之一, 影响了太阳辐射和降水的空间再分配, 往往导致局部小气候的产生(Cantón et al ., 2004 ), 从而改变光照、温度及土壤水分的分布, 导致群落环境和植物的生存条件产生差异(侯兆疆等, 2014 ), 进而改变植物碳资源分配格局和根系构型构建策略(Oppelt et al ., 2001 )。为了提高植株生存适合度和竞争能力, 植物根系通过分支角度和分叉数等性状的响应, 促使根系构型模式进行适应性调整, 构建与环境梯度相匹配的根系营养吸收和传输系统。因此, 从不同坡向资源条件变化的角度研究植物根系构型的空间分异格局和资源配置模式, 对于理解植物在异质生境中的环境适应机制具有重要的理论意义。
红砂(Reaumuria songarica )是一种广泛分布于我国西北荒漠地区的柽柳科小灌木, 是荒漠植被的常见建群种和优势种, 具有极强的抗干旱能力。在全球变化背景下, 红砂等荒漠植物面对日益严酷的环境压力, 需要不断调整主要功能性状构建的资源策略, 适时改变植物碳资源分配格局和根系构型构建策略(单立山等, 2013 ), 这对维持荒漠生态系统的稳定性具有重要意义。目前, 许多学者围绕植物根系分支模式的竞争能力(Glimskär, 2000 )、根系构型的拓扑结构(马新明等, 2006 )、多枝柽柳(Tamarix ramosissima )和骆驼刺(Alhagi sparsifolia )幼苗生物量分配及根系分布特征(曾凡江等, 2010 )、不同树种间细根结构和形态特征(熊德成等, 2012 )、骆驼刺群落在干旱生境下细根生产与周转(冀卫萍等, 2013 )、植物根系构型的生态适应性(杜建会等, 2014 )以及根系功能性状的可塑性与空间异质性(Druille et al ., 2015 )等方面对植物根系构型进行了系统研究, 但是有关生境梯度下植物根系构型构建的资源配置权衡机制等研究仍显不足, 限制了对异质生境中植物表型可塑性机制的解析。本文通过研究祁连山北坡荒漠草地红砂根系分叉数和分支角度的分布格局, 试图解析: (1)不同坡向红砂根系分叉数和分支角度的变化规律; (2)造成红砂根系分叉数和分支角度异速生长关系的原因, 旨在从植物根系功能性状建成的角度阐明红砂种群的环境适应性规律, 为进一步了解荒漠植物的表型可塑性机制提供理论依据。
1 材料和方法
1.1 研究区域和样地概况
研究区位于祁连山北坡中山区的甘肃省肃南县皂矾沟地区, 属于山地荒漠草原, 地理位置为38.91°-38.98° N, 99.81°-99.87° E, 海拔1834-2118 m, 年平均气温6-7 ℃, ≥0 ℃积温为2933-3300 ℃, 年降水量173.1 mm, 年蒸发量2013.7 mm, 是年降水量的11.6倍。土壤为山地灰漠土, 由于气候干燥和长期剥蚀的作用, 形成砾石倾斜和裸岩广布的低山地貌景观。植物以旱生小半灌木为主, 红砂属(Reaumuria )植物占相当比例。主要植物有红砂、合头草(Sympegma regelii )、驼绒黎(Krascheninnikovia ceratoides )、盐爪爪(Kalidium foliatum )、珍珠猪毛菜(Salsola passerina )、霸王(Zygophyllum xanthoxylon ), 伴生植物有灌木亚菊(Ajania fruticulosa )、中亚紫菀木(Asterothamnus centraliasiaticus )、碱韭(Allium polyrhizum )、黄花瓦松(Orostachys spinosus )、黄花补血草(Limonium aureum )、三芒草(Aristida adscensionis )等。供试草地为灌木和旱生草本的混生群落。受干旱气候的影响, 南坡植被稀疏、生长状况差, 北坡植被郁闭度较高。
1.2 实验设置
在地形图上将研究区划分为4个坡向, 每个坡向沿垂直海拔梯度(1940-2115 m)间隔30 m布置1条长140 m、宽10 m的水平样带, 总计20条水平样带。在多次现场踏勘的基础上, 在每条水平样带上间隔20 m设置1个5 m × 5 m的样方, 共计120个样方, GPS测量每个样方的经纬度和海拔。第一, 进行草地群落学特征调查, 测定样方内所有植物的盖度、高度、密度和地上生物量。第二, 每一样方内随机选择1株生长良好的红砂, 每个坡向共计30株红砂用于测定植株的生物量和根系性状, 首先将每株红砂的地上枝条刈割并装入自封袋, 然后采用壕沟法和根系跟踪相结合的方法挖掘整株根系, 在离植株2 m处挖一条2 m × 0.6 m × 1.6 m壕沟, 然后利用小铁铲慢慢清除植株周围的泥沙到壕沟并不断清理沟中层积的沙土, 直至直径>3 mm的整株根系完全暴露, 因为直径<3 mm根系构型的重建在研究中很难实现(Oppelt et al ., 2000 , 2005 ), 尽量保持根系在自然空间中的位置。根系完全暴露后, 采用50 cm × 50 cm的网格确定根系的分布位置(直到没有根系出现为止), 按照1:50的比例在35 cm × 25 cm的坐标纸上准确绘制根系构型图, 然后把植株根系装入尼龙袋带回实验室。在室内将根系置于细筛之上用水冲洗, 同时去除其他杂质, 仅留下红砂的根系; 用游标卡尺、卷尺和量角器等工具, 分别测定所有样点红砂的根长、分叉数和分支夹角(°); 之后把植株根系和地上部分分别装入信封, 在80 ℃烘箱里烘24 h后用电子天平(0.0001 g)称质量并记录。第三, 进行土壤水分采样, 每个样方内随机选取3个样点, 用土钻(直径4 cm)采集0-100 cm土层范围分5层间隔20 cm取土壤样品, 钻取的土壤先按土层分取, 之后将各层土壤进行混合, 重复3次, 剔除样品中明显的植物根段和枯落物等杂质, 装入编号的铝盒中, 带回实验室, 在105 ℃的烘箱内烘12 h取出称质量, 计算出各样地0-100 cm土层土壤质量含水量。
1.3 空间数据采集
利用ArcGIS对研究区1:5万地形图进行数字化, 得到研究区域10 m分辨率的数字高程模型(DEM)。在DEM基础上利用Spatial Analyst模块中的Surface Analysis命令提取坡向, 它表示地表面上一点的切平面的法线矢量n 在水平面的投影nxoy 与过该点的正北方向的夹角, 其中坡向组别以国际划分法确定(陈瑶等, 2006 ), 具体方法参见汤国安等(2010) 文献。
1.4 数据处理
将坡向划分为北坡(315°-360°和0°-45°)、东坡(45°-135°)、南坡(135°-225°)和西坡(225°-315°) 4个坡向组。根据游标卡尺、卷尺和量角器等工具测定获得的根长、分叉数和分支角度等, 计算出各坡向红砂根系的比根长。比根长表示单位生物量的细根长度, 它们反映投入到根系的生物量中用于吸收养分的效率, 其值越大, 表明投入相同的成本有越高的养分利用效率(Bauhus et al ., 2000 )。计算公式为: 比根长(cm·g-1 ) =根长(cm)/生物量(g)。根系分叉数和分支角度数据分析主要采用标准化主轴估计(SMA)的方法(Warton et al ., 2006 ), 由软件(S)MATR Version 2.0 (Falster et al ., 2006 )计算完成。对每个样方红砂根系分叉数和分支角度进行对数(以10为底)转换后再进行分析, 对数转换使之符合正态分布。采用y = axb , 线性转换成log(y ) = log(a ) + b log(x ), 式中x 和y 表示两个特征参数, b 为斜率, 即异速生长参数或相对生长的指数, 当|b | = 1时, 表示两者是等速变化关系; 当|b| 显著偏离1时, 两者间为异速变化关系(Harvey & Pagel, 1991 )。实验所用数据采用SPSS 16.0软件进行处理分析, 用SigmaPlot 10.0和Excel软件绘图。
2 结果分析
2.1 不同坡向红砂根系分叉数与分支角度的变化
单因素方差分析表明, 不同坡向草地红砂的根系分叉数与分支角度均存在显著差异(p < 0.05, 图1 )。随着坡向由南坡转向西坡、东坡和北坡, 红砂根系分叉数呈逐渐增加趋势, 从南坡的15.34个增加至北坡的31.17个, 增加了1.03倍; 而红砂根系分支角度呈相反的变化趋势, 从南坡的86°减小至北坡的62°, 减小了28%。
图1 红砂根系分叉数和分支角度沿坡向梯度的变化(平均值±标准误差)。不同小写字母表示坡向间差异显著(p < 0.05)。
Fig. 1 Changes in root forks and branch angle of Reaumuria songarica along the aspect gradient (mean ± SE ). Different lowercase letters indicate significant differences among slope aspects (p < 0.05).
2.2 不同坡向红砂根系分叉数与分支角度之间的关系
Pearson相关性分析表明, 南坡和北坡甘肃红砂根系分叉数和分支角度均存在极显著负相关(p < 0.01), 而东坡和西坡存在显著负相关(p < 0.05, 图2 )。在坡向梯度上随着红砂根系分叉数的增加, 根系分支角度均呈减小趋势, 表明红砂根系分叉数和分支角度之间存在权衡关系。北坡和南坡红砂根系分叉数和分支角度回归方程的SMA斜率逐渐减小(p < 0.05), 而东坡、西坡甘肃臭草根系分叉数和分支角度回归方程的SMA斜率之间无显著差异(p > 0.05), 各个坡向的SMA斜率的绝对值显著小于1 (p < 0.05), 表明根系分叉数的增长速率大于根系分支角度的生长速率, 红砂根系具有较强的分支能力, 有利于发挥根系吸收代谢及固土蓄水能力作用。
图2 不同坡向红砂根系分叉数和分支角度的关系。A, 南坡; B, 西坡; C, 东坡; D, 北坡。
Fig. 2 The relationship between root forks and branch angle of Reaumuria songarica in different aspects of slopes. A, South slope; B, West slope; C, East slope; D, North slope.
2.3 不同坡向红砂的生长特征
不同坡向梯度分布的红砂生长特征差异均显著(p < 0.05, 表1 )。随着坡向由南坡转向西坡、东坡和北坡, 红砂种群的根冠比呈逐渐减小趋势, 从南坡到北坡减小了48.08%; 密度、高度、根深和比根长呈相反的变化趋势逐渐增加, 从南坡到北坡分别增大了1.5倍、0.53倍、1.8倍和0.57倍。这些生长特征值在东坡和西坡之间无显著差异(p > 0.05)。
2.4 不同坡向红砂草地群落及土壤水分特征
不同坡向红砂草地群落和土壤水分特征存在差异(p < 0.05, 表2 )。随着坡向由南坡向西坡、东坡和北坡转变, 土壤含水量呈递增趋势, 由南坡的6.1%增加到北坡的11.5%, 土壤水分在南坡具有最小值, 北坡具有最大值; 从南坡到北坡, 草地群落的盖度、密度、高度、地上生物量和地下生物量呈递增趋势, 群落盖度增加了2.61倍, 群落密度增加了2.13倍, 群落高度增加了31%, 地上生物量增加了67%, 地下生物量增加了93%, 且在东坡和西坡之间无显著差异(p > 0.05)。表明在4个坡向中, 北坡生境条件最适合植株的生长。
3 讨论
植物在长期的进化过程中, 通过生理和发育调节形成与环境相适应的表型特征, 进而具备了提高植物水分、养分等资源利用效率的功能(李西良等, 2014 )。在异质性生境中, 植物根系面对水分和养分吸收、运移和存储的环境压迫, 其分叉数和分支角度可能会发生适应性变化, 并通过物质能量吸收、固着格局的调整反作用于植被生态系统的稳定和种群更新机制, 体现了植物根系进行空间探索以适应环境胁迫的能力(Zhao et al ., 2013 )。本研究发现, 随着坡向由南坡向西坡、东坡和北坡的转变, 红砂根系分支角度逐渐减小, 而根系分叉数呈相反的变化趋势(图1 ), 在有限的资源环境中, 红砂分配给根系分叉数和分支角度资源间存在着“此消彼长”的权衡关系。坡向梯度上红砂根系分叉数和分支角度关系的差异性是植物根系构建的资源配置模式对生境异质性的适应对策。
植物外部形态具有高度的环境可塑性特征, 在长期生长发育的过程中, 生境差异性使其主要构件采取了不同的生态对策, 能够协调各功能性状间的资源分配模式以规避环境风险(史元春等, 2015 )。植物根系构型构建与植物自身根系碳投入和土壤水分养分的收益权衡相契合, 是植物提高自身资源获取能力的一个重要策略(杨小林等, 2008 )。研究区位于祁连山与河西走廊过渡带的荒漠草原区, 山地南坡的植物生存环境比较严酷, 红砂的生物量水平低(表1 , 表2 ), 为了提高自身的生存和繁衍能力, 红砂将更多的同化产物分配给地下根系, 着力构建强大的物质吸收和存储系统, 从而限制了地上生物量的分配比例(表1 ), 形成了相对较小的、有助于降低植株总体蒸腾耗水量的枝叶性状体系(表3 )。南坡植被稀疏(表2 )、大气温度较高, 结构松散的沙土毛细管吸收作用力弱。在土壤含水量偏低(表2 )和无地下水补给的条件下, 荒漠植物往往需要较大集水面积才能满足植株最低水平的生理耗水需求, 红砂选择了增加根系分支角度、减小分叉数的根系构型构建模式, 原因如下: 1)红砂属于浅根系植物, 绝大部分根系与地面平行伸展, 形成相对较大的根系分支角度(图1 ), 更有利于侧根趋向于水平分布、扩大根系的有效觅养空间, 同样单位(长度)的根系拥有的吸水土壤体积可能较大, 从而具有比同一生境中相邻植物更强的水资源竞争能力, 可以获得更多的生存机会(蒋礼学和李彦, 2008 )。这符合相关学者“分支角度越大, 根系越趋向于水平分布, 向根更广的范围扩展以寻求资源”的研究结论(黄晶晶, 2013 )。2)植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 保证自身的生存和生长的策略(杜建会等, 2014 )。生长在南坡的红砂减小其根系分叉数的根系构型(图1 ), 在一定程度上降低了根系内部的交叠重复, 可以避免细根间资源空间的竞争, 从而提高根系对营养物质的吸收效率来加快对土壤资源的利用, 以最佳的收益/代价关系适应贫瘠的资源环境, 与相关学者得出一致的结论, 即“当植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 减少根系分叉数的策略以降低根系内部的交叠重复, 避免细根营养物质的竞争” (单立山等, 2012 ; 杜建会等, 2014 )。因此, 分布于南坡的红砂根系分叉数和分支角度间呈极显著负相关关系(p < 0.01, 图2 ), 实现了光合产物在构件水平的优化配置。
植物构件的资源分配是与栖息环境相对应的种群特征的反映方式, 它反映了种群特定的生态适应策略(张茜等, 2014 )。根系构型会对周围环境产生适应性的改变, 表现出不一样的适应特性(Oppelt et al ., 2005 )。相对于南坡, 山地北坡草地群落的盖度、密度均较大, 红砂在群落中没有高度优势(表1 , 表2 ), 相对拥挤的群落环境使植物面临着严重的邻体干扰, 红砂将较多的生物量分配给地上枝叶以利于植株获得高度优势, 增强枝叶的捕光能力, 无形中限制了分配给根系的生物量比例(表1 )。在根冠比较低的情况下, 红砂需要将有限的资源用于构建根系的吸收面积, 增强根系的“觅食”能力(董鸣, 1996 ), 以最大限度保证植株在生境竞争中占据优势, 红砂选择了增加根系分叉数、减小分支角度的根系构型。原因如下: 1)在土壤含水量相对较高的北坡(表2 ), 红砂以根系分叉数增加速度大于分支角度减小速度的资源配置模式(图2 ), 通过增大根系分叉数(图1 ), 有效增加了根系分支能力和单位土体中根系的拥挤度, 构建了密集的根系网络系统, 降低了根系传输系统的碳需求量, 实现了单位根系土壤拓展体积较小状态下的植株生存的水分供给需求, 这与单立山等(2013)对红砂和白刺(Nitraria tangutorum )、杜建会等(2014)对塔克拉玛干柽柳(Tamarix taklamakanensis )的研究结果相一致。2)植物根系在深层土壤的分布有利于其吸收利用深层土壤水分, 促进植株的生长发育(席海洋等, 2011 )。北坡群落密度高(表2 )、植物之间拥挤度大, 种内和种间容易造成根系重叠竞争, 空间上无法满足大范围根系的构型塑造。红砂采取缩小根系水平范围, 将根系分支角度减小, 扩大根系在深层土壤中的垂直分布(表1 ), 避免与群落中其他植株根系的剧烈竞争。同时红砂植物通过增大比根长提高对养分和水分的利用效率来加快对土壤资源的利用(表1 ), 提高自身的竞争力以维持生存。因此, 红砂根系分叉数和分支角度间呈极显著负相关关系(p < 0.01, 图2 ), 即以资源权衡分配机制进化响应诸多竞争压力, 维持植物生长的水分条件, 从而有利于生物多样性的群落结构, 这符合相关学者“分叉数较大的植物能够迅速占据大量土壤空间快速生长, 适宜生长在土壤资源丰富的环境中”的结论(周艳松和王立群, 2011 ; 孙浩燕等, 2014 )。
植物个体间的相互作用实际上是可获得性资源的有限性导致的构件水平的竞争(马维玲等, 2010 ), 根系是植物营养物质摄取、运输、贮存和合成场所, 也是寻觅新的生境和营养的繁殖器官(黄晶晶等, 2013 ), 影响植株个体的生长、发育和植物种群的生活史对策(Dannowski & Block, 2005 )。东坡和西坡属于半阴和半阳坡, 是植被类型的交错带, 植被多样性较高, 群落组成较为复杂, 草地群落密度分别为17株·m-2 和15株·m-2 , 盖度为36%和34%, 高度为21.32 cm和21.70 cm, 都介于南坡和北坡之间, 植物之间的邻体干扰较大。为了增强资源竞争能力和生境适合度, 红砂选择了折中的地上、地下生物量配置模式, 兼顾了地上植株空间拓展和地下根系水分、养分吸收传输的物质需求。受地下生物量有限性的约束, 在权衡根系吸水面积和吸水效率等生理生态功能的基础上, 红砂选择了既有利于提高根系分布密度, 又能增强根系空间拓展能力的根系分叉数和分支角度优化配置模式。因此生长在东坡和西坡植株的二者的相关性仅达到显著水平(p < 0.05, 图1 ), 是根系分支角度由大到小、根系分叉数由小到大转变的区域。
4 结论
根系分叉数和分支角度关系不仅是植物个体发育过程的表现, 也体现植物在特定环境的构型塑造和对特定生境的适应性。本研究发现, 生境对根系分叉数和分支角度关系有显著的影响, 不同坡向中红砂根系功能性状存在差异。生长在南坡的红砂选择了增加根系分支角度、减小分叉数的扩散型根系构型, 是其在逆境中扩展根系的分布范围及降低细根营养物质的竞争的生长策略; 北坡的红砂通过增大根系分叉数、减小分支角度, 趋向于构建紧缩型的根系构型, 是其提高根系吸收利用效率和减弱荒漠植物“大根系”冗余消耗养分的弊端的生活史对策。这种结果较好地验证了植物通过调整表型可塑性的自我补偿、更新策略。本文仅从坡向角度分析了根系分叉数-分支角度的变化特征, 它们的关系可能还受到坡度等其他微地形因子以及物种遗传特性的影响, 还需要进一步的实验验证。
发表在2015年(39卷)第1期81-91页周洪华等的文章“胡杨木质部水分传导对盐分胁迫的响应与适应”中, 89页基金项目: “国家自然科学基金(41271061)和新疆维吾尔自治区自然基金(2014211A077)。”应为“国家自然科学基金(41271006)、新疆维吾尔自治区自然基金(2014211A077)和新疆青年科技创新人才工程(2014721035)。” 特此更正。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Bauhus J Khanna PK Menden N 2000 ). Aboveground and belowground interactions in mixed plantations of Eucalyptus globulus and Acacia mearnsii.
Canadian Journal of Forest Research , 30 , 1886 -1894 .
[本文引用: 1]
[2]
Cantón Y Del Barrio G Solé-Benet A Lázaro R 2004 ). Topographic controls on the spatial distribution of ground cover in the Tabernas badlands of SE Spain.
Catena , 55 , 341 -365 .
[本文引用: 1]
[3]
Chang XL Lü SH Feng ZY Ye SX 2015 ). Impact of topography on the spatial distribution pattern of net primary productivity in a meadow.
Acta Ecologica Sinica , 35 , 3339 -3348 .
[本文引用: 1]
(in Chinese with English abstract) [常学礼 , 吕世海 , 冯朝阳 , 叶生星 (2015 ). 地形对草甸草原植被生产力分布格局的影响.
生态学报 , 35 , 3339 -3348 .]
[本文引用: 1]
[4]
Chen Y Xu X Zhang DR Wei Y 2006 ). Correlations between vegetation distribution and topographical factors in the northwest of Longmen Mountain, Sichuan Province.
Chinese Journal of Ecology , 25 , 1052 -1055 .
[本文引用: 1]
(in Chinese with English abstract) [陈瑶 , 胥晓 , 张德然 , 魏勇 (2006 ). 四川龙门山西北部植被分布与地形因子的相关性.
生态学杂志 , 25 , 1052 -1055 .]
[本文引用: 1]
[5]
Chen YT Xu ZZ 2014 ). Review on research of leaf economics spectrum.
Chinese Journal of Plant Ecology , 38 , 1135 -1153 .
[本文引用: 1]
(in Chinese with English abstract) [陈莹婷 , 许振柱 (2014 ). 植物叶经济谱的研究进展.
植物生态学报 , 38 , 1135 -1153 .]
[本文引用: 1]
[6]
Dannowski M Block A 2005 ). Fractal geometry and root system structures of heterogeneous plant communities.
Plant and Soil , 272 , 61 -76 .
[本文引用: 2]
[7]
Dong M 1996 ). Clonal growth in plants in relation to resource heterogeneity: Foraging behavior.
Acta Botanica Sinica , 38 , 828 -835 .
[本文引用: 1]
(in Chinese with English abstract) [董鸣 (1996 ). 资源异质性环境中的植物克隆生长: 觅食行为.
植物学报 , 38 , 828 -835 .]
[本文引用: 1]
[8]
Druille M Cabello MN García Parisi PA Golluscio RA Omacini M 2015 ). Glyphosate vulnerability explains changes in root-symbionts propagules viability in pampean grasslands.
Agriculture, Ecosystems and Environment , 202 , 48 -55 .
[本文引用: 1]
[9]
Du JH Liu AL Dong YX Hu MY Liang J Li W 2014 ). Architectural characteristics of roots in typical coastal psammophytes of South China.
Chinese Journal of Plant Ecology , 38 , 888 -895 .
[本文引用: 3]
(in Chinese with English abstract) [杜建会 , 刘安隆 , 董玉祥 , 胡绵友 , 梁杰 , 李薇 (2014 ). 华南海岸典型沙生植物根系构型特征.
植物生态学报 ,38 , 888 -895 .]
[本文引用: 3]
[10]
Falster DS Warton DI Wright IJ
URL
[本文引用: 1]
[11]
Glimskär A 2000 ). Estimates of root system topology of five plant species grown at steady-state nutrition.
Plant and Soil , 227 , 249 -256 .
[12]
Harvey PH Pagel MD 1991 ). The Comparative Method in Evolutionary Biology . Oxford University Press , Oxford, UK .
[本文引用: 1]
[13]
Hou ZJ Zhao CZ Li Y Zhang Q Ma XL 2014 ). Trade-off between height and branch numbers in Stellera chamaejasme on slopes of different aspects in a degraded alpine grassland.
Chinese Journal of Plant Ecology , 38 , 281 -288 .
[本文引用: 4]
(in Chinese with English abstract) [侯兆疆 , 赵成章 , 李钰 , 张茜 , 马小丽 (2014 ). 不同坡向高寒退化草地狼毒株高和枝条数的权衡关系.
植物生态学报 , 38 , 281 -288 .]
[本文引用: 4]
[14]
Huang JJ 2013 ). Cloning Root System Distribution and Architecture of Populus euphratica in Ejina Oasis.
Master degree dissertation, Beijing Forestry University, Beijing.
(in Chinese with English abstract) [黄晶晶 (2013 ). 额济纳荒漠绿洲胡杨根系特征的研究
. 硕士学位论文, 北京林业大学, 北京.]
[15]
Huang JJ Jing JL Cao DC Zhang N Li JW Xia YG Lü S 2013 ). Cloning root system distribution and architecture of different forest age Populus euphratica in Ejina Oasis.
Acta Ecologica Sinica , 33 , 4331 -4342 .
[本文引用: 1]
(in Chinese with English abstract) [黄晶晶 , 井家林 , 曹德昌 , 张楠 , 李景文 , 夏延国 , 吕爽 (2013 ). 不同林龄胡杨克隆繁殖根系分布特征及其构型.
生态学报 , 33 , 4331 -4342 .]
[本文引用: 1]
[16]
Ji WP Wang JJ Zhao XC Lai LM Wang YJ Ma YJ Zhao CQ Zheng YR 2013 ). Fine root production and turnover of Alhagi sparsifolia community in arid area of Xinjiang, Northwest China.
Chinese Journal of Ecology , 32 , 2635 -2640 .
[本文引用: 1]
(in Chinese with English abstract) [冀卫萍 , 王健健 , 赵学春 , 来利明 , 王永吉 , 马远见 , 赵春强 , 郑元润 (2013 ). 干旱区骆驼刺群落细根生产与周转.
生态学杂志 , 32 , 2635 -2640 .]
[本文引用: 1]
[17]
Jiang LX Li Y 2008 ). Comparison on architecture characteristics of root systems and leaf traits for three desert shrubs adapted to arid habitat.
Journal of Desert Research , 28 , 1118 -1124 .
[本文引用: 1]
(in Chinese with English abstract) [蒋礼学 , 李彦 (2008 ). 三种荒漠灌木根系的构形特征与叶性因子对干旱生境的适应性比较.
中国沙漠 , 28 , 1118 -1124 .]
[本文引用: 1]
[18]
Kong XP Zhang ML de Smet I Ding ZJ 2014 ). Designer crops: Optimal root system architecture for nutrient acquisition.
Trends in Biotechnology , 32 , 597 -598 .
[本文引用: 1]
[19]
Li XL Hou XY Wu XH Sa RL Ji L Chen HJ Liu ZY Ding Y 2014 ). Plastic responses of stem and leaf functional traits in Leymus chinensis to long-term grazing in a meadow steppe.
Chinese Journal of Plant Ecology , 38 , 440 -451 .
[本文引用: 1]
(in Chinese with English abstract) [李西良 , 侯向阳 , 吴新宏 , 萨茹拉 , 纪磊 , 陈海军 , 刘志英 , 丁勇 (2014 ). 草甸草原羊草茎叶功能性状对长期过度放牧的可塑性响应.
植物生态学报 , 38 , 440 -451 .]
[本文引用: 1]
[20]
Ma WL Shi PL Li WH He YT Zhang XZ Shen ZX Chai SY 2010 ). Changes in individual plant traits and biomass allocation in alpine meadow with elevation variation on the Qinghai-Tibetan Plateau.
Science China: Life Sciences , 53 , 1142 -1151 .
[本文引用: 1]
[马维玲 , 石培礼 , 李文华 , 何永涛 , 张宪洲 , 沈振西 (2010 ). 青藏高原高寒草甸植株性状和生物量分配的海拔梯度变异
. 中国科学: 生命科学 , 40 , 533 -543 .]
[本文引用: 1]
[21]
Ma XM Xi L Xiong SP Yang J 2006 ). Dynamic changes of morphological parameters of tobacco root in field.
Chinese Journal of Applied Ecology , 17 , 373 -376 .
[本文引用: 2]
(in Chinese with English abstract) [马新明 , 席磊 , 熊淑萍 , 杨娟 (2006 ). 大田期烟草根系构型参数的动态变化.
应用生态学报 , 17 , 373 -376 .]
[本文引用: 2]
[22]
Malamy JE 2005 ). Intrinsic and environmental response pathways that regulate root system architecture.
Plant, Cell & Environment , 28 , 67 -77 .
[本文引用: 1]
[23]
Oppelt AL Kurth W Dzierzon H Jentschke G Godbold DL 2000 ). Structure and fractal dimensions of root systems of four co-occurring fruit tree species from Botswana.
Annals of Forest Science , 57 , 463 -475 .
[本文引用: 1]
[24]
Oppelt AL Kurth W Godbold DL 2001 ). Topology, scaling relations and Leonardo’s rule in root systems from African tree species.
Tree Physiology , 21 , 117 -128 .
[本文引用: 2]
[25]
Oppelt AL Kurth W Godbold DL 2005 ). Contrasting rooting patterns of some arid-zone fruit tree species from Botswana-II. Coarse root distribution.
Agroforestry Systems , 64 , 13 -24 .
[本文引用: 1]
[26]
Shan LS Li Y Dong QL Geng DM 2012 ). Ecological adaptation of Reaumuria soongorica root system architecture to arid environment.
Journal of Desert Research , 32 , 1283 -1290 .
[本文引用: 1]
(in Chinese with English abstract) [单立山 , 李毅 , 董秋莲 , 耿东梅 (2012 ). 红砂根系构型对干旱的生态适应.
中国沙漠 , 32 , 1283 -1290 .]
[本文引用: 1]
[27]
Shan LS Li Y Ren W Su SP Dong QL Geng DM 2013 ). Root architecture of two desert plants in central Hexi Corridor of Northwest China.
Chinese Journal of Applied Ecology , 24 , 25 -31 .
[本文引用: 1]
(in Chinese with English abstract) [单立山 , 李毅 , 任伟 , 苏世平 , 董秋莲 , 耿东梅 (2013 ). 河西走廊中部两种荒漠植物根系构型特征.
应用生态学报 , 24 , 25 -31 .]
[本文引用: 1]
[28]
Shi YC Zhao CZ Song QH Du J Chen J Wang JW 2015 ). Slope-related variations in twig and leaf traits of Robinia pseudoacacia in the northern mountains of Lanzhou.
Chinese Journal of Plant Ecology , 39 , 362 -370 .
[本文引用: 2]
(in Chinese with English abstract) [史元春 , 赵成章 , 宋清华 , 杜晶 , 陈静 , 王继伟 (2015 ). 兰州北山刺槐枝叶性状的坡向差异性.
植物生态学报 , 39 , 362 -370 .]
[本文引用: 2]
[29]
Sun HY Li XK Ren T Cong RH Lu JW 2014 ). Effects of fertilizer in shallow soils on growth and distribution of rice roots at seedling stage.
Scientia Agricultura Sinica , 47 , 2476 -2484 .
(in Chinese with English abstract) [孙浩燕 , 李小坤 , 任涛 , 丛日环 , 鲁剑巍 (2014 ). 浅层施肥对水稻苗期根系生长及分布的影响.
中国农业科学 ,47 , 2476 -2484 .]
[30]
Tang GA Li FY Liu XJ 2010 ). Tutorial of Digital Elevation Model. 2nd edn . Science Press , Beijing . 149 .
[本文引用: 1]
(in Chinese) [汤国安 , 李发源 , 刘学军 (2010 ). 数字高程模型教程. 第二版 . 科学出版社 , 北京 . 149 .]
[本文引用: 1]
[31]
Tracy SR Black CR Roberts JA Dodd IC Mooney SJ 2015 ). Using X-ray Computed Tomography to explore the role of abscisic acid in moderating the impact of soil compaction on root system architecture.
Environmental and Experimental Botany , 110 , 11 -18 .
[本文引用: 1]
[32]
Warton DI Wright IJ Falster DS Westoby M 2006 ). Bivariate line-fitting methods for allometry.
Biological Reviews , 81 , 259 -291 .
[本文引用: 1]
[33]
Xi HY Feng Q Si JH Chang ZQ Su YH Guo R 2011 ). Spatio-temporal characteristics of soil in Ejina Oasis.
Journal of Desert Research , 31 , 68 -75 .
[本文引用: 1]
(in Chinese with English abstract) [席海洋 , 冯起 , 司建华 , 常宗强 , 苏永红 , 郭瑞 (2011 ). 额济纳绿洲不同植被覆盖下土壤特性的时空变化
. 中国沙漠 , 31 , 68 -75 ].
[本文引用: 1]
[34]
Xiong DC Huang JX Yang ZJ Lu ZL Cheng GS Yang YS 2012 ). Fine root architecture and morphology among different branch orders of six subtropical tree species.
Acta Ecologica Sinica , 32 , 1888 -1897 .
[本文引用: 1]
(in Chinese with English abstract) [熊德成 , 黄锦学 , 杨智杰 , 卢正立 , 陈光水 , 杨玉盛 (2012 ). 亚热带6种树种细根序级结构和形态特征.
生态学报 , 32 , 1888 -1897 .]
[本文引用: 1]
[35]
Yang XL Zhang XM Li YL Li SC Sun HL 2008 ). Analysis of root architecture and root adaptive strategy in the Taklimakan desert area of China.
Journal of Plant Ecology (Chinese Version) , 32 , 1268 -1276 .
[本文引用: 1]
(in Chinese with English abstract) [杨小林 , 张希明 , 李义玲 , 李绍才 , 孙海龙 (2008 ). 塔克拉玛干沙漠腹地3种植物根系构型及其生境适应策略.
植物生态学报 , 32 , 1268 -1276 .]
[本文引用: 1]
[36]
Zeng FJ Guo HF Liu B Zeng J Xing WJ Zhang XL 2010 ). Characteristics of biomass allocation and root distribution of Tamarix ramosissima Ledeb. and Alhagi sparsifolia Shap. seedlings.
Arid Land Geography , 33 , 59 -64 .
[本文引用: 1]
(in Chinese with English abstract) [曾凡江 , 郭海峰 . 刘波 , 曾杰 , 邢文娟 , 张晓蕾 (2010 ). 多枝柽柳和疏叶骆驼刺幼苗生物量分配及根系分布特征.
干旱区地理 , 33 , 59 -64 .]
[本文引用: 1]
[37]
Zhang Q Zhao CZ Dong XG Ma XL Hou ZJ Li Y 2014 ). Trade-off between the biomass and number of flowers in Stellera chamaejasme along an elevation gradient in a degraded alpine grassland.
Chinese Journal of Plant Ecology , 38 , 452 -459 .
[本文引用: 1]
(in Chinese with English abstract) [张茜 , 赵成章 , 董小刚 , 马小丽 , 侯兆疆 , 李钰 (2014 ). 高寒退化草地狼毒种群不同海拔花大小-数量的权衡关系.
植物生态学报 , 38 , 452 -459 .]
[本文引用: 1]
[38]
Zhao CZ Sheng YP Ren H Gao FY Zhang J 2013 ). A study on the root competitive pattern of annual pasture in mixed grassland in alpine region.
Acta Ecologica Sinica , 33 , 145 -149 .
[本文引用: 1]
[39]
Zhou YS Wang LQ 2011 ). Ecological adaptation of root architecture to grassland degradation in
Potentilla acaulis. Chinese Journal of Plant Ecology , 35 , 490 -499 .
(in Chinese with English abstract) [周艳松 , 王立群 (2011 ). 星毛委陵菜根系构型对草原退化的生态适应.
植物生态学报 , 35 , 490 -499 .]
Aboveground and belowground interactions in mixed plantations of Eucalyptus globulus and Acacia mearnsii.
1
2000
... 将坡向划分为北坡(315°-360°和0°-45°)、东坡(45°-135°)、南坡(135°-225°)和西坡(225°-315°) 4个坡向组.根据游标卡尺、卷尺和量角器等工具测定获得的根长、分叉数和分支角度等, 计算出各坡向红砂根系的比根长.比根长表示单位生物量的细根长度, 它们反映投入到根系的生物量中用于吸收养分的效率, 其值越大, 表明投入相同的成本有越高的养分利用效率(Bauhus et al ., 2000 ).计算公式为: 比根长(cm·g-1 ) =根长(cm)/生物量(g).根系分叉数和分支角度数据分析主要采用标准化主轴估计(SMA)的方法(Warton et al ., 2006 ), 由软件(S)MATR Version 2.0 (Falster et al ., 2006 )计算完成.对每个样方红砂根系分叉数和分支角度进行对数(以10为底)转换后再进行分析, 对数转换使之符合正态分布.采用y = axb , 线性转换成log(y ) = log(a ) + b log(x ), 式中x 和y 表示两个特征参数, b 为斜率, 即异速生长参数或相对生长的指数, 当|b | = 1时, 表示两者是等速变化关系; 当|b| 显著偏离1时, 两者间为异速变化关系(Harvey & Pagel, 1991 ).实验所用数据采用SPSS 16.0软件进行处理分析, 用SigmaPlot 10.0和Excel软件绘图. ...
Topographic controls on the spatial distribution of ground cover in the Tabernas badlands of SE Spain.
1
2004
... 植物根系作为植被与土壤之间物质、能量交换的主要通道(黄晶晶, 2013 ), 在植物水分和养分吸收、运移过程中有着不可替代的作用(Kong et al ., 2014 ), 影响着地上部分以及整个植株的生长和发育, 其中根系的分支状况和构型对营养物质的吸收起着关键作用(Dannowski & Block, 2005 ).根系构型是植物与环境异质性资源相互适应的结果(Malamy, 2005 ), 其对环境的适应是通过根系分支结构、空间分布格局等构型特征的改变表现出来的(Tracy et al ., 2015 ).为了应对地理环境和资源供给水平造成的胁迫和干扰, 植物根系会根据所处生境的资源条件适时调整和转化根系分叉数和分支角度等构型特征, 实现植物功能性状间的资源权衡(陈莹婷和许振柱, 2014 ).根系分叉数影响植物根系对资源物质的觅食能力和植株的固定作用(孙浩燕等, 2014 ), 根系分支角度与根系的分布范围和拓展能力紧密相关(黄晶晶, 2013 ), 二者在很大程度上决定了根系在土壤中的位置和对土体的觅养效率(Malamy, 2005 ), 反映了不同生境中植物根系在物质吸收、利用和循环中的作用.地形作为生境条件的综合指示, 控制水热因子的重新分配, 修饰植被空间分布格局, 此自然法则具有普适性(常学礼等, 2015 ).坡向是地形重要的影响因素之一, 影响了太阳辐射和降水的空间再分配, 往往导致局部小气候的产生(Cantón et al ., 2004 ), 从而改变光照、温度及土壤水分的分布, 导致群落环境和植物的生存条件产生差异(侯兆疆等, 2014 ), 进而改变植物碳资源分配格局和根系构型构建策略(Oppelt et al ., 2001 ).为了提高植株生存适合度和竞争能力, 植物根系通过分支角度和分叉数等性状的响应, 促使根系构型模式进行适应性调整, 构建与环境梯度相匹配的根系营养吸收和传输系统.因此, 从不同坡向资源条件变化的角度研究植物根系构型的空间分异格局和资源配置模式, 对于理解植物在异质生境中的环境适应机制具有重要的理论意义. ...
地形对草甸草原植被生产力分布格局的影响.
1
2015
... 植物根系作为植被与土壤之间物质、能量交换的主要通道(黄晶晶, 2013 ), 在植物水分和养分吸收、运移过程中有着不可替代的作用(Kong et al ., 2014 ), 影响着地上部分以及整个植株的生长和发育, 其中根系的分支状况和构型对营养物质的吸收起着关键作用(Dannowski & Block, 2005 ).根系构型是植物与环境异质性资源相互适应的结果(Malamy, 2005 ), 其对环境的适应是通过根系分支结构、空间分布格局等构型特征的改变表现出来的(Tracy et al ., 2015 ).为了应对地理环境和资源供给水平造成的胁迫和干扰, 植物根系会根据所处生境的资源条件适时调整和转化根系分叉数和分支角度等构型特征, 实现植物功能性状间的资源权衡(陈莹婷和许振柱, 2014 ).根系分叉数影响植物根系对资源物质的觅食能力和植株的固定作用(孙浩燕等, 2014 ), 根系分支角度与根系的分布范围和拓展能力紧密相关(黄晶晶, 2013 ), 二者在很大程度上决定了根系在土壤中的位置和对土体的觅养效率(Malamy, 2005 ), 反映了不同生境中植物根系在物质吸收、利用和循环中的作用.地形作为生境条件的综合指示, 控制水热因子的重新分配, 修饰植被空间分布格局, 此自然法则具有普适性(常学礼等, 2015 ).坡向是地形重要的影响因素之一, 影响了太阳辐射和降水的空间再分配, 往往导致局部小气候的产生(Cantón et al ., 2004 ), 从而改变光照、温度及土壤水分的分布, 导致群落环境和植物的生存条件产生差异(侯兆疆等, 2014 ), 进而改变植物碳资源分配格局和根系构型构建策略(Oppelt et al ., 2001 ).为了提高植株生存适合度和竞争能力, 植物根系通过分支角度和分叉数等性状的响应, 促使根系构型模式进行适应性调整, 构建与环境梯度相匹配的根系营养吸收和传输系统.因此, 从不同坡向资源条件变化的角度研究植物根系构型的空间分异格局和资源配置模式, 对于理解植物在异质生境中的环境适应机制具有重要的理论意义. ...
地形对草甸草原植被生产力分布格局的影响.
1
2015
... 植物根系作为植被与土壤之间物质、能量交换的主要通道(黄晶晶, 2013 ), 在植物水分和养分吸收、运移过程中有着不可替代的作用(Kong et al ., 2014 ), 影响着地上部分以及整个植株的生长和发育, 其中根系的分支状况和构型对营养物质的吸收起着关键作用(Dannowski & Block, 2005 ).根系构型是植物与环境异质性资源相互适应的结果(Malamy, 2005 ), 其对环境的适应是通过根系分支结构、空间分布格局等构型特征的改变表现出来的(Tracy et al ., 2015 ).为了应对地理环境和资源供给水平造成的胁迫和干扰, 植物根系会根据所处生境的资源条件适时调整和转化根系分叉数和分支角度等构型特征, 实现植物功能性状间的资源权衡(陈莹婷和许振柱, 2014 ).根系分叉数影响植物根系对资源物质的觅食能力和植株的固定作用(孙浩燕等, 2014 ), 根系分支角度与根系的分布范围和拓展能力紧密相关(黄晶晶, 2013 ), 二者在很大程度上决定了根系在土壤中的位置和对土体的觅养效率(Malamy, 2005 ), 反映了不同生境中植物根系在物质吸收、利用和循环中的作用.地形作为生境条件的综合指示, 控制水热因子的重新分配, 修饰植被空间分布格局, 此自然法则具有普适性(常学礼等, 2015 ).坡向是地形重要的影响因素之一, 影响了太阳辐射和降水的空间再分配, 往往导致局部小气候的产生(Cantón et al ., 2004 ), 从而改变光照、温度及土壤水分的分布, 导致群落环境和植物的生存条件产生差异(侯兆疆等, 2014 ), 进而改变植物碳资源分配格局和根系构型构建策略(Oppelt et al ., 2001 ).为了提高植株生存适合度和竞争能力, 植物根系通过分支角度和分叉数等性状的响应, 促使根系构型模式进行适应性调整, 构建与环境梯度相匹配的根系营养吸收和传输系统.因此, 从不同坡向资源条件变化的角度研究植物根系构型的空间分异格局和资源配置模式, 对于理解植物在异质生境中的环境适应机制具有重要的理论意义. ...
四川龙门山西北部植被分布与地形因子的相关性.
1
2006
... 利用ArcGIS对研究区1:5万地形图进行数字化, 得到研究区域10 m分辨率的数字高程模型(DEM).在DEM基础上利用Spatial Analyst模块中的Surface Analysis命令提取坡向, 它表示地表面上一点的切平面的法线矢量n 在水平面的投影nxoy 与过该点的正北方向的夹角, 其中坡向组别以国际划分法确定(陈瑶等, 2006 ), 具体方法参见汤国安等(2010) 文献. ...
四川龙门山西北部植被分布与地形因子的相关性.
1
2006
... 利用ArcGIS对研究区1:5万地形图进行数字化, 得到研究区域10 m分辨率的数字高程模型(DEM).在DEM基础上利用Spatial Analyst模块中的Surface Analysis命令提取坡向, 它表示地表面上一点的切平面的法线矢量n 在水平面的投影nxoy 与过该点的正北方向的夹角, 其中坡向组别以国际划分法确定(陈瑶等, 2006 ), 具体方法参见汤国安等(2010) 文献. ...
植物叶经济谱的研究进展.
1
2014
... 植物根系作为植被与土壤之间物质、能量交换的主要通道(黄晶晶, 2013 ), 在植物水分和养分吸收、运移过程中有着不可替代的作用(Kong et al ., 2014 ), 影响着地上部分以及整个植株的生长和发育, 其中根系的分支状况和构型对营养物质的吸收起着关键作用(Dannowski & Block, 2005 ).根系构型是植物与环境异质性资源相互适应的结果(Malamy, 2005 ), 其对环境的适应是通过根系分支结构、空间分布格局等构型特征的改变表现出来的(Tracy et al ., 2015 ).为了应对地理环境和资源供给水平造成的胁迫和干扰, 植物根系会根据所处生境的资源条件适时调整和转化根系分叉数和分支角度等构型特征, 实现植物功能性状间的资源权衡(陈莹婷和许振柱, 2014 ).根系分叉数影响植物根系对资源物质的觅食能力和植株的固定作用(孙浩燕等, 2014 ), 根系分支角度与根系的分布范围和拓展能力紧密相关(黄晶晶, 2013 ), 二者在很大程度上决定了根系在土壤中的位置和对土体的觅养效率(Malamy, 2005 ), 反映了不同生境中植物根系在物质吸收、利用和循环中的作用.地形作为生境条件的综合指示, 控制水热因子的重新分配, 修饰植被空间分布格局, 此自然法则具有普适性(常学礼等, 2015 ).坡向是地形重要的影响因素之一, 影响了太阳辐射和降水的空间再分配, 往往导致局部小气候的产生(Cantón et al ., 2004 ), 从而改变光照、温度及土壤水分的分布, 导致群落环境和植物的生存条件产生差异(侯兆疆等, 2014 ), 进而改变植物碳资源分配格局和根系构型构建策略(Oppelt et al ., 2001 ).为了提高植株生存适合度和竞争能力, 植物根系通过分支角度和分叉数等性状的响应, 促使根系构型模式进行适应性调整, 构建与环境梯度相匹配的根系营养吸收和传输系统.因此, 从不同坡向资源条件变化的角度研究植物根系构型的空间分异格局和资源配置模式, 对于理解植物在异质生境中的环境适应机制具有重要的理论意义. ...
植物叶经济谱的研究进展.
1
2014
... 植物根系作为植被与土壤之间物质、能量交换的主要通道(黄晶晶, 2013 ), 在植物水分和养分吸收、运移过程中有着不可替代的作用(Kong et al ., 2014 ), 影响着地上部分以及整个植株的生长和发育, 其中根系的分支状况和构型对营养物质的吸收起着关键作用(Dannowski & Block, 2005 ).根系构型是植物与环境异质性资源相互适应的结果(Malamy, 2005 ), 其对环境的适应是通过根系分支结构、空间分布格局等构型特征的改变表现出来的(Tracy et al ., 2015 ).为了应对地理环境和资源供给水平造成的胁迫和干扰, 植物根系会根据所处生境的资源条件适时调整和转化根系分叉数和分支角度等构型特征, 实现植物功能性状间的资源权衡(陈莹婷和许振柱, 2014 ).根系分叉数影响植物根系对资源物质的觅食能力和植株的固定作用(孙浩燕等, 2014 ), 根系分支角度与根系的分布范围和拓展能力紧密相关(黄晶晶, 2013 ), 二者在很大程度上决定了根系在土壤中的位置和对土体的觅养效率(Malamy, 2005 ), 反映了不同生境中植物根系在物质吸收、利用和循环中的作用.地形作为生境条件的综合指示, 控制水热因子的重新分配, 修饰植被空间分布格局, 此自然法则具有普适性(常学礼等, 2015 ).坡向是地形重要的影响因素之一, 影响了太阳辐射和降水的空间再分配, 往往导致局部小气候的产生(Cantón et al ., 2004 ), 从而改变光照、温度及土壤水分的分布, 导致群落环境和植物的生存条件产生差异(侯兆疆等, 2014 ), 进而改变植物碳资源分配格局和根系构型构建策略(Oppelt et al ., 2001 ).为了提高植株生存适合度和竞争能力, 植物根系通过分支角度和分叉数等性状的响应, 促使根系构型模式进行适应性调整, 构建与环境梯度相匹配的根系营养吸收和传输系统.因此, 从不同坡向资源条件变化的角度研究植物根系构型的空间分异格局和资源配置模式, 对于理解植物在异质生境中的环境适应机制具有重要的理论意义. ...
Fractal geometry and root system structures of heterogeneous plant communities.
2
2005
... 植物根系作为植被与土壤之间物质、能量交换的主要通道(黄晶晶, 2013 ), 在植物水分和养分吸收、运移过程中有着不可替代的作用(Kong et al ., 2014 ), 影响着地上部分以及整个植株的生长和发育, 其中根系的分支状况和构型对营养物质的吸收起着关键作用(Dannowski & Block, 2005 ).根系构型是植物与环境异质性资源相互适应的结果(Malamy, 2005 ), 其对环境的适应是通过根系分支结构、空间分布格局等构型特征的改变表现出来的(Tracy et al ., 2015 ).为了应对地理环境和资源供给水平造成的胁迫和干扰, 植物根系会根据所处生境的资源条件适时调整和转化根系分叉数和分支角度等构型特征, 实现植物功能性状间的资源权衡(陈莹婷和许振柱, 2014 ).根系分叉数影响植物根系对资源物质的觅食能力和植株的固定作用(孙浩燕等, 2014 ), 根系分支角度与根系的分布范围和拓展能力紧密相关(黄晶晶, 2013 ), 二者在很大程度上决定了根系在土壤中的位置和对土体的觅养效率(Malamy, 2005 ), 反映了不同生境中植物根系在物质吸收、利用和循环中的作用.地形作为生境条件的综合指示, 控制水热因子的重新分配, 修饰植被空间分布格局, 此自然法则具有普适性(常学礼等, 2015 ).坡向是地形重要的影响因素之一, 影响了太阳辐射和降水的空间再分配, 往往导致局部小气候的产生(Cantón et al ., 2004 ), 从而改变光照、温度及土壤水分的分布, 导致群落环境和植物的生存条件产生差异(侯兆疆等, 2014 ), 进而改变植物碳资源分配格局和根系构型构建策略(Oppelt et al ., 2001 ).为了提高植株生存适合度和竞争能力, 植物根系通过分支角度和分叉数等性状的响应, 促使根系构型模式进行适应性调整, 构建与环境梯度相匹配的根系营养吸收和传输系统.因此, 从不同坡向资源条件变化的角度研究植物根系构型的空间分异格局和资源配置模式, 对于理解植物在异质生境中的环境适应机制具有重要的理论意义. ...
... 植物个体间的相互作用实际上是可获得性资源的有限性导致的构件水平的竞争(马维玲等, 2010 ), 根系是植物营养物质摄取、运输、贮存和合成场所, 也是寻觅新的生境和营养的繁殖器官(黄晶晶等, 2013 ), 影响植株个体的生长、发育和植物种群的生活史对策(Dannowski & Block, 2005 ).东坡和西坡属于半阴和半阳坡, 是植被类型的交错带, 植被多样性较高, 群落组成较为复杂, 草地群落密度分别为17株·m-2 和15株·m-2 , 盖度为36%和34%, 高度为21.32 cm和21.70 cm, 都介于南坡和北坡之间, 植物之间的邻体干扰较大.为了增强资源竞争能力和生境适合度, 红砂选择了折中的地上、地下生物量配置模式, 兼顾了地上植株空间拓展和地下根系水分、养分吸收传输的物质需求.受地下生物量有限性的约束, 在权衡根系吸水面积和吸水效率等生理生态功能的基础上, 红砂选择了既有利于提高根系分布密度, 又能增强根系空间拓展能力的根系分叉数和分支角度优化配置模式.因此生长在东坡和西坡植株的二者的相关性仅达到显著水平(p < 0.05, 图1 ), 是根系分支角度由大到小、根系分叉数由小到大转变的区域. ...
资源异质性环境中的植物克隆生长: 觅食行为.
1
1996
... 植物构件的资源分配是与栖息环境相对应的种群特征的反映方式, 它反映了种群特定的生态适应策略(张茜等, 2014 ).根系构型会对周围环境产生适应性的改变, 表现出不一样的适应特性(Oppelt et al ., 2005 ).相对于南坡, 山地北坡草地群落的盖度、密度均较大, 红砂在群落中没有高度优势(表1 , 表2 ), 相对拥挤的群落环境使植物面临着严重的邻体干扰, 红砂将较多的生物量分配给地上枝叶以利于植株获得高度优势, 增强枝叶的捕光能力, 无形中限制了分配给根系的生物量比例(表1 ).在根冠比较低的情况下, 红砂需要将有限的资源用于构建根系的吸收面积, 增强根系的“觅食”能力(董鸣, 1996 ), 以最大限度保证植株在生境竞争中占据优势, 红砂选择了增加根系分叉数、减小分支角度的根系构型.原因如下: 1)在土壤含水量相对较高的北坡(表2 ), 红砂以根系分叉数增加速度大于分支角度减小速度的资源配置模式(图2 ), 通过增大根系分叉数(图1 ), 有效增加了根系分支能力和单位土体中根系的拥挤度, 构建了密集的根系网络系统, 降低了根系传输系统的碳需求量, 实现了单位根系土壤拓展体积较小状态下的植株生存的水分供给需求, 这与单立山等(2013)对红砂和白刺(Nitraria tangutorum )、杜建会等(2014)对塔克拉玛干柽柳(Tamarix taklamakanensis )的研究结果相一致.2)植物根系在深层土壤的分布有利于其吸收利用深层土壤水分, 促进植株的生长发育(席海洋等, 2011 ).北坡群落密度高(表2 )、植物之间拥挤度大, 种内和种间容易造成根系重叠竞争, 空间上无法满足大范围根系的构型塑造.红砂采取缩小根系水平范围, 将根系分支角度减小, 扩大根系在深层土壤中的垂直分布(表1 ), 避免与群落中其他植株根系的剧烈竞争.同时红砂植物通过增大比根长提高对养分和水分的利用效率来加快对土壤资源的利用(表1 ), 提高自身的竞争力以维持生存.因此, 红砂根系分叉数和分支角度间呈极显著负相关关系(p < 0.01, 图2 ), 即以资源权衡分配机制进化响应诸多竞争压力, 维持植物生长的水分条件, 从而有利于生物多样性的群落结构, 这符合相关学者“分叉数较大的植物能够迅速占据大量土壤空间快速生长, 适宜生长在土壤资源丰富的环境中”的结论(周艳松和王立群, 2011 ; 孙浩燕等, 2014 ). ...
资源异质性环境中的植物克隆生长: 觅食行为.
1
1996
... 植物构件的资源分配是与栖息环境相对应的种群特征的反映方式, 它反映了种群特定的生态适应策略(张茜等, 2014 ).根系构型会对周围环境产生适应性的改变, 表现出不一样的适应特性(Oppelt et al ., 2005 ).相对于南坡, 山地北坡草地群落的盖度、密度均较大, 红砂在群落中没有高度优势(表1 , 表2 ), 相对拥挤的群落环境使植物面临着严重的邻体干扰, 红砂将较多的生物量分配给地上枝叶以利于植株获得高度优势, 增强枝叶的捕光能力, 无形中限制了分配给根系的生物量比例(表1 ).在根冠比较低的情况下, 红砂需要将有限的资源用于构建根系的吸收面积, 增强根系的“觅食”能力(董鸣, 1996 ), 以最大限度保证植株在生境竞争中占据优势, 红砂选择了增加根系分叉数、减小分支角度的根系构型.原因如下: 1)在土壤含水量相对较高的北坡(表2 ), 红砂以根系分叉数增加速度大于分支角度减小速度的资源配置模式(图2 ), 通过增大根系分叉数(图1 ), 有效增加了根系分支能力和单位土体中根系的拥挤度, 构建了密集的根系网络系统, 降低了根系传输系统的碳需求量, 实现了单位根系土壤拓展体积较小状态下的植株生存的水分供给需求, 这与单立山等(2013)对红砂和白刺(Nitraria tangutorum )、杜建会等(2014)对塔克拉玛干柽柳(Tamarix taklamakanensis )的研究结果相一致.2)植物根系在深层土壤的分布有利于其吸收利用深层土壤水分, 促进植株的生长发育(席海洋等, 2011 ).北坡群落密度高(表2 )、植物之间拥挤度大, 种内和种间容易造成根系重叠竞争, 空间上无法满足大范围根系的构型塑造.红砂采取缩小根系水平范围, 将根系分支角度减小, 扩大根系在深层土壤中的垂直分布(表1 ), 避免与群落中其他植株根系的剧烈竞争.同时红砂植物通过增大比根长提高对养分和水分的利用效率来加快对土壤资源的利用(表1 ), 提高自身的竞争力以维持生存.因此, 红砂根系分叉数和分支角度间呈极显著负相关关系(p < 0.01, 图2 ), 即以资源权衡分配机制进化响应诸多竞争压力, 维持植物生长的水分条件, 从而有利于生物多样性的群落结构, 这符合相关学者“分叉数较大的植物能够迅速占据大量土壤空间快速生长, 适宜生长在土壤资源丰富的环境中”的结论(周艳松和王立群, 2011 ; 孙浩燕等, 2014 ). ...
Glyphosate vulnerability explains changes in root-symbionts propagules viability in pampean grasslands.
1
2015
... 红砂(Reaumuria songarica )是一种广泛分布于我国西北荒漠地区的柽柳科小灌木, 是荒漠植被的常见建群种和优势种, 具有极强的抗干旱能力.在全球变化背景下, 红砂等荒漠植物面对日益严酷的环境压力, 需要不断调整主要功能性状构建的资源策略, 适时改变植物碳资源分配格局和根系构型构建策略(单立山等, 2013 ), 这对维持荒漠生态系统的稳定性具有重要意义.目前, 许多学者围绕植物根系分支模式的竞争能力(Glimskär, 2000 )、根系构型的拓扑结构(马新明等, 2006 )、多枝柽柳(Tamarix ramosissima )和骆驼刺(Alhagi sparsifolia )幼苗生物量分配及根系分布特征(曾凡江等, 2010 )、不同树种间细根结构和形态特征(熊德成等, 2012 )、骆驼刺群落在干旱生境下细根生产与周转(冀卫萍等, 2013 )、植物根系构型的生态适应性(杜建会等, 2014 )以及根系功能性状的可塑性与空间异质性(Druille et al ., 2015 )等方面对植物根系构型进行了系统研究, 但是有关生境梯度下植物根系构型构建的资源配置权衡机制等研究仍显不足, 限制了对异质生境中植物表型可塑性机制的解析.本文通过研究祁连山北坡荒漠草地红砂根系分叉数和分支角度的分布格局, 试图解析: (1)不同坡向红砂根系分叉数和分支角度的变化规律; (2)造成红砂根系分叉数和分支角度异速生长关系的原因, 旨在从植物根系功能性状建成的角度阐明红砂种群的环境适应性规律, 为进一步了解荒漠植物的表型可塑性机制提供理论依据. ...
华南海岸典型沙生植物根系构型特征.
3
2014
... 红砂(Reaumuria songarica )是一种广泛分布于我国西北荒漠地区的柽柳科小灌木, 是荒漠植被的常见建群种和优势种, 具有极强的抗干旱能力.在全球变化背景下, 红砂等荒漠植物面对日益严酷的环境压力, 需要不断调整主要功能性状构建的资源策略, 适时改变植物碳资源分配格局和根系构型构建策略(单立山等, 2013 ), 这对维持荒漠生态系统的稳定性具有重要意义.目前, 许多学者围绕植物根系分支模式的竞争能力(Glimskär, 2000 )、根系构型的拓扑结构(马新明等, 2006 )、多枝柽柳(Tamarix ramosissima )和骆驼刺(Alhagi sparsifolia )幼苗生物量分配及根系分布特征(曾凡江等, 2010 )、不同树种间细根结构和形态特征(熊德成等, 2012 )、骆驼刺群落在干旱生境下细根生产与周转(冀卫萍等, 2013 )、植物根系构型的生态适应性(杜建会等, 2014 )以及根系功能性状的可塑性与空间异质性(Druille et al ., 2015 )等方面对植物根系构型进行了系统研究, 但是有关生境梯度下植物根系构型构建的资源配置权衡机制等研究仍显不足, 限制了对异质生境中植物表型可塑性机制的解析.本文通过研究祁连山北坡荒漠草地红砂根系分叉数和分支角度的分布格局, 试图解析: (1)不同坡向红砂根系分叉数和分支角度的变化规律; (2)造成红砂根系分叉数和分支角度异速生长关系的原因, 旨在从植物根系功能性状建成的角度阐明红砂种群的环境适应性规律, 为进一步了解荒漠植物的表型可塑性机制提供理论依据. ...
... 植物外部形态具有高度的环境可塑性特征, 在长期生长发育的过程中, 生境差异性使其主要构件采取了不同的生态对策, 能够协调各功能性状间的资源分配模式以规避环境风险(史元春等, 2015 ).植物根系构型构建与植物自身根系碳投入和土壤水分养分的收益权衡相契合, 是植物提高自身资源获取能力的一个重要策略(杨小林等, 2008 ).研究区位于祁连山与河西走廊过渡带的荒漠草原区, 山地南坡的植物生存环境比较严酷, 红砂的生物量水平低(表1 , 表2 ), 为了提高自身的生存和繁衍能力, 红砂将更多的同化产物分配给地下根系, 着力构建强大的物质吸收和存储系统, 从而限制了地上生物量的分配比例(表1 ), 形成了相对较小的、有助于降低植株总体蒸腾耗水量的枝叶性状体系(表3 ).南坡植被稀疏(表2 )、大气温度较高, 结构松散的沙土毛细管吸收作用力弱.在土壤含水量偏低(表2 )和无地下水补给的条件下, 荒漠植物往往需要较大集水面积才能满足植株最低水平的生理耗水需求, 红砂选择了增加根系分支角度、减小分叉数的根系构型构建模式, 原因如下: 1)红砂属于浅根系植物, 绝大部分根系与地面平行伸展, 形成相对较大的根系分支角度(图1 ), 更有利于侧根趋向于水平分布、扩大根系的有效觅养空间, 同样单位(长度)的根系拥有的吸水土壤体积可能较大, 从而具有比同一生境中相邻植物更强的水资源竞争能力, 可以获得更多的生存机会(蒋礼学和李彦, 2008 ).这符合相关学者“分支角度越大, 根系越趋向于水平分布, 向根更广的范围扩展以寻求资源”的研究结论(黄晶晶, 2013 ).2)植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 保证自身的生存和生长的策略(杜建会等, 2014 ).生长在南坡的红砂减小其根系分叉数的根系构型(图1 ), 在一定程度上降低了根系内部的交叠重复, 可以避免细根间资源空间的竞争, 从而提高根系对营养物质的吸收效率来加快对土壤资源的利用, 以最佳的收益/代价关系适应贫瘠的资源环境, 与相关学者得出一致的结论, 即“当植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 减少根系分叉数的策略以降低根系内部的交叠重复, 避免细根营养物质的竞争” (单立山等, 2012 ; 杜建会等, 2014 ).因此, 分布于南坡的红砂根系分叉数和分支角度间呈极显著负相关关系(p < 0.01, 图2 ), 实现了光合产物在构件水平的优化配置. ...
... ; 杜建会等, 2014 ).因此, 分布于南坡的红砂根系分叉数和分支角度间呈极显著负相关关系(p < 0.01, 图2 ), 实现了光合产物在构件水平的优化配置. ...
华南海岸典型沙生植物根系构型特征.
3
2014
... 红砂(Reaumuria songarica )是一种广泛分布于我国西北荒漠地区的柽柳科小灌木, 是荒漠植被的常见建群种和优势种, 具有极强的抗干旱能力.在全球变化背景下, 红砂等荒漠植物面对日益严酷的环境压力, 需要不断调整主要功能性状构建的资源策略, 适时改变植物碳资源分配格局和根系构型构建策略(单立山等, 2013 ), 这对维持荒漠生态系统的稳定性具有重要意义.目前, 许多学者围绕植物根系分支模式的竞争能力(Glimskär, 2000 )、根系构型的拓扑结构(马新明等, 2006 )、多枝柽柳(Tamarix ramosissima )和骆驼刺(Alhagi sparsifolia )幼苗生物量分配及根系分布特征(曾凡江等, 2010 )、不同树种间细根结构和形态特征(熊德成等, 2012 )、骆驼刺群落在干旱生境下细根生产与周转(冀卫萍等, 2013 )、植物根系构型的生态适应性(杜建会等, 2014 )以及根系功能性状的可塑性与空间异质性(Druille et al ., 2015 )等方面对植物根系构型进行了系统研究, 但是有关生境梯度下植物根系构型构建的资源配置权衡机制等研究仍显不足, 限制了对异质生境中植物表型可塑性机制的解析.本文通过研究祁连山北坡荒漠草地红砂根系分叉数和分支角度的分布格局, 试图解析: (1)不同坡向红砂根系分叉数和分支角度的变化规律; (2)造成红砂根系分叉数和分支角度异速生长关系的原因, 旨在从植物根系功能性状建成的角度阐明红砂种群的环境适应性规律, 为进一步了解荒漠植物的表型可塑性机制提供理论依据. ...
... 植物外部形态具有高度的环境可塑性特征, 在长期生长发育的过程中, 生境差异性使其主要构件采取了不同的生态对策, 能够协调各功能性状间的资源分配模式以规避环境风险(史元春等, 2015 ).植物根系构型构建与植物自身根系碳投入和土壤水分养分的收益权衡相契合, 是植物提高自身资源获取能力的一个重要策略(杨小林等, 2008 ).研究区位于祁连山与河西走廊过渡带的荒漠草原区, 山地南坡的植物生存环境比较严酷, 红砂的生物量水平低(表1 , 表2 ), 为了提高自身的生存和繁衍能力, 红砂将更多的同化产物分配给地下根系, 着力构建强大的物质吸收和存储系统, 从而限制了地上生物量的分配比例(表1 ), 形成了相对较小的、有助于降低植株总体蒸腾耗水量的枝叶性状体系(表3 ).南坡植被稀疏(表2 )、大气温度较高, 结构松散的沙土毛细管吸收作用力弱.在土壤含水量偏低(表2 )和无地下水补给的条件下, 荒漠植物往往需要较大集水面积才能满足植株最低水平的生理耗水需求, 红砂选择了增加根系分支角度、减小分叉数的根系构型构建模式, 原因如下: 1)红砂属于浅根系植物, 绝大部分根系与地面平行伸展, 形成相对较大的根系分支角度(图1 ), 更有利于侧根趋向于水平分布、扩大根系的有效觅养空间, 同样单位(长度)的根系拥有的吸水土壤体积可能较大, 从而具有比同一生境中相邻植物更强的水资源竞争能力, 可以获得更多的生存机会(蒋礼学和李彦, 2008 ).这符合相关学者“分支角度越大, 根系越趋向于水平分布, 向根更广的范围扩展以寻求资源”的研究结论(黄晶晶, 2013 ).2)植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 保证自身的生存和生长的策略(杜建会等, 2014 ).生长在南坡的红砂减小其根系分叉数的根系构型(图1 ), 在一定程度上降低了根系内部的交叠重复, 可以避免细根间资源空间的竞争, 从而提高根系对营养物质的吸收效率来加快对土壤资源的利用, 以最佳的收益/代价关系适应贫瘠的资源环境, 与相关学者得出一致的结论, 即“当植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 减少根系分叉数的策略以降低根系内部的交叠重复, 避免细根营养物质的竞争” (单立山等, 2012 ; 杜建会等, 2014 ).因此, 分布于南坡的红砂根系分叉数和分支角度间呈极显著负相关关系(p < 0.01, 图2 ), 实现了光合产物在构件水平的优化配置. ...
... ; 杜建会等, 2014 ).因此, 分布于南坡的红砂根系分叉数和分支角度间呈极显著负相关关系(p < 0.01, 图2 ), 实现了光合产物在构件水平的优化配置. ...
1
... 红砂(Reaumuria songarica )是一种广泛分布于我国西北荒漠地区的柽柳科小灌木, 是荒漠植被的常见建群种和优势种, 具有极强的抗干旱能力.在全球变化背景下, 红砂等荒漠植物面对日益严酷的环境压力, 需要不断调整主要功能性状构建的资源策略, 适时改变植物碳资源分配格局和根系构型构建策略(单立山等, 2013 ), 这对维持荒漠生态系统的稳定性具有重要意义.目前, 许多学者围绕植物根系分支模式的竞争能力(Glimskär, 2000 )、根系构型的拓扑结构(马新明等, 2006 )、多枝柽柳(Tamarix ramosissima )和骆驼刺(Alhagi sparsifolia )幼苗生物量分配及根系分布特征(曾凡江等, 2010 )、不同树种间细根结构和形态特征(熊德成等, 2012 )、骆驼刺群落在干旱生境下细根生产与周转(冀卫萍等, 2013 )、植物根系构型的生态适应性(杜建会等, 2014 )以及根系功能性状的可塑性与空间异质性(Druille et al ., 2015 )等方面对植物根系构型进行了系统研究, 但是有关生境梯度下植物根系构型构建的资源配置权衡机制等研究仍显不足, 限制了对异质生境中植物表型可塑性机制的解析.本文通过研究祁连山北坡荒漠草地红砂根系分叉数和分支角度的分布格局, 试图解析: (1)不同坡向红砂根系分叉数和分支角度的变化规律; (2)造成红砂根系分叉数和分支角度异速生长关系的原因, 旨在从植物根系功能性状建成的角度阐明红砂种群的环境适应性规律, 为进一步了解荒漠植物的表型可塑性机制提供理论依据. ...
Estimates of root system topology of five plant species grown at steady-state nutrition.
0
2000
1
1991
... 植物根系作为植被与土壤之间物质、能量交换的主要通道(黄晶晶, 2013 ), 在植物水分和养分吸收、运移过程中有着不可替代的作用(Kong et al ., 2014 ), 影响着地上部分以及整个植株的生长和发育, 其中根系的分支状况和构型对营养物质的吸收起着关键作用(Dannowski & Block, 2005 ).根系构型是植物与环境异质性资源相互适应的结果(Malamy, 2005 ), 其对环境的适应是通过根系分支结构、空间分布格局等构型特征的改变表现出来的(Tracy et al ., 2015 ).为了应对地理环境和资源供给水平造成的胁迫和干扰, 植物根系会根据所处生境的资源条件适时调整和转化根系分叉数和分支角度等构型特征, 实现植物功能性状间的资源权衡(陈莹婷和许振柱, 2014 ).根系分叉数影响植物根系对资源物质的觅食能力和植株的固定作用(孙浩燕等, 2014 ), 根系分支角度与根系的分布范围和拓展能力紧密相关(黄晶晶, 2013 ), 二者在很大程度上决定了根系在土壤中的位置和对土体的觅养效率(Malamy, 2005 ), 反映了不同生境中植物根系在物质吸收、利用和循环中的作用.地形作为生境条件的综合指示, 控制水热因子的重新分配, 修饰植被空间分布格局, 此自然法则具有普适性(常学礼等, 2015 ).坡向是地形重要的影响因素之一, 影响了太阳辐射和降水的空间再分配, 往往导致局部小气候的产生(Cantón et al ., 2004 ), 从而改变光照、温度及土壤水分的分布, 导致群落环境和植物的生存条件产生差异(侯兆疆等, 2014 ), 进而改变植物碳资源分配格局和根系构型构建策略(Oppelt et al ., 2001 ).为了提高植株生存适合度和竞争能力, 植物根系通过分支角度和分叉数等性状的响应, 促使根系构型模式进行适应性调整, 构建与环境梯度相匹配的根系营养吸收和传输系统.因此, 从不同坡向资源条件变化的角度研究植物根系构型的空间分异格局和资源配置模式, 对于理解植物在异质生境中的环境适应机制具有重要的理论意义. ...
不同坡向高寒退化草地狼毒株高和枝条数的权衡关系.
4
2014
... 植物根系作为植被与土壤之间物质、能量交换的主要通道(黄晶晶, 2013 ), 在植物水分和养分吸收、运移过程中有着不可替代的作用(Kong et al ., 2014 ), 影响着地上部分以及整个植株的生长和发育, 其中根系的分支状况和构型对营养物质的吸收起着关键作用(Dannowski & Block, 2005 ).根系构型是植物与环境异质性资源相互适应的结果(Malamy, 2005 ), 其对环境的适应是通过根系分支结构、空间分布格局等构型特征的改变表现出来的(Tracy et al ., 2015 ).为了应对地理环境和资源供给水平造成的胁迫和干扰, 植物根系会根据所处生境的资源条件适时调整和转化根系分叉数和分支角度等构型特征, 实现植物功能性状间的资源权衡(陈莹婷和许振柱, 2014 ).根系分叉数影响植物根系对资源物质的觅食能力和植株的固定作用(孙浩燕等, 2014 ), 根系分支角度与根系的分布范围和拓展能力紧密相关(黄晶晶, 2013 ), 二者在很大程度上决定了根系在土壤中的位置和对土体的觅养效率(Malamy, 2005 ), 反映了不同生境中植物根系在物质吸收、利用和循环中的作用.地形作为生境条件的综合指示, 控制水热因子的重新分配, 修饰植被空间分布格局, 此自然法则具有普适性(常学礼等, 2015 ).坡向是地形重要的影响因素之一, 影响了太阳辐射和降水的空间再分配, 往往导致局部小气候的产生(Cantón et al ., 2004 ), 从而改变光照、温度及土壤水分的分布, 导致群落环境和植物的生存条件产生差异(侯兆疆等, 2014 ), 进而改变植物碳资源分配格局和根系构型构建策略(Oppelt et al ., 2001 ).为了提高植株生存适合度和竞争能力, 植物根系通过分支角度和分叉数等性状的响应, 促使根系构型模式进行适应性调整, 构建与环境梯度相匹配的根系营养吸收和传输系统.因此, 从不同坡向资源条件变化的角度研究植物根系构型的空间分异格局和资源配置模式, 对于理解植物在异质生境中的环境适应机制具有重要的理论意义. ...
... ), 根系分支角度与根系的分布范围和拓展能力紧密相关(黄晶晶, 2013 ), 二者在很大程度上决定了根系在土壤中的位置和对土体的觅养效率(Malamy, 2005 ), 反映了不同生境中植物根系在物质吸收、利用和循环中的作用.地形作为生境条件的综合指示, 控制水热因子的重新分配, 修饰植被空间分布格局, 此自然法则具有普适性(常学礼等, 2015 ).坡向是地形重要的影响因素之一, 影响了太阳辐射和降水的空间再分配, 往往导致局部小气候的产生(Cantón et al ., 2004 ), 从而改变光照、温度及土壤水分的分布, 导致群落环境和植物的生存条件产生差异(侯兆疆等, 2014 ), 进而改变植物碳资源分配格局和根系构型构建策略(Oppelt et al ., 2001 ).为了提高植株生存适合度和竞争能力, 植物根系通过分支角度和分叉数等性状的响应, 促使根系构型模式进行适应性调整, 构建与环境梯度相匹配的根系营养吸收和传输系统.因此, 从不同坡向资源条件变化的角度研究植物根系构型的空间分异格局和资源配置模式, 对于理解植物在异质生境中的环境适应机制具有重要的理论意义. ...
... 植物外部形态具有高度的环境可塑性特征, 在长期生长发育的过程中, 生境差异性使其主要构件采取了不同的生态对策, 能够协调各功能性状间的资源分配模式以规避环境风险(史元春等, 2015 ).植物根系构型构建与植物自身根系碳投入和土壤水分养分的收益权衡相契合, 是植物提高自身资源获取能力的一个重要策略(杨小林等, 2008 ).研究区位于祁连山与河西走廊过渡带的荒漠草原区, 山地南坡的植物生存环境比较严酷, 红砂的生物量水平低(表1 , 表2 ), 为了提高自身的生存和繁衍能力, 红砂将更多的同化产物分配给地下根系, 着力构建强大的物质吸收和存储系统, 从而限制了地上生物量的分配比例(表1 ), 形成了相对较小的、有助于降低植株总体蒸腾耗水量的枝叶性状体系(表3 ).南坡植被稀疏(表2 )、大气温度较高, 结构松散的沙土毛细管吸收作用力弱.在土壤含水量偏低(表2 )和无地下水补给的条件下, 荒漠植物往往需要较大集水面积才能满足植株最低水平的生理耗水需求, 红砂选择了增加根系分支角度、减小分叉数的根系构型构建模式, 原因如下: 1)红砂属于浅根系植物, 绝大部分根系与地面平行伸展, 形成相对较大的根系分支角度(图1 ), 更有利于侧根趋向于水平分布、扩大根系的有效觅养空间, 同样单位(长度)的根系拥有的吸水土壤体积可能较大, 从而具有比同一生境中相邻植物更强的水资源竞争能力, 可以获得更多的生存机会(蒋礼学和李彦, 2008 ).这符合相关学者“分支角度越大, 根系越趋向于水平分布, 向根更广的范围扩展以寻求资源”的研究结论(黄晶晶, 2013 ).2)植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 保证自身的生存和生长的策略(杜建会等, 2014 ).生长在南坡的红砂减小其根系分叉数的根系构型(图1 ), 在一定程度上降低了根系内部的交叠重复, 可以避免细根间资源空间的竞争, 从而提高根系对营养物质的吸收效率来加快对土壤资源的利用, 以最佳的收益/代价关系适应贫瘠的资源环境, 与相关学者得出一致的结论, 即“当植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 减少根系分叉数的策略以降低根系内部的交叠重复, 避免细根营养物质的竞争” (单立山等, 2012 ; 杜建会等, 2014 ).因此, 分布于南坡的红砂根系分叉数和分支角度间呈极显著负相关关系(p < 0.01, 图2 ), 实现了光合产物在构件水平的优化配置. ...
... 植物个体间的相互作用实际上是可获得性资源的有限性导致的构件水平的竞争(马维玲等, 2010 ), 根系是植物营养物质摄取、运输、贮存和合成场所, 也是寻觅新的生境和营养的繁殖器官(黄晶晶等, 2013 ), 影响植株个体的生长、发育和植物种群的生活史对策(Dannowski & Block, 2005 ).东坡和西坡属于半阴和半阳坡, 是植被类型的交错带, 植被多样性较高, 群落组成较为复杂, 草地群落密度分别为17株·m-2 和15株·m-2 , 盖度为36%和34%, 高度为21.32 cm和21.70 cm, 都介于南坡和北坡之间, 植物之间的邻体干扰较大.为了增强资源竞争能力和生境适合度, 红砂选择了折中的地上、地下生物量配置模式, 兼顾了地上植株空间拓展和地下根系水分、养分吸收传输的物质需求.受地下生物量有限性的约束, 在权衡根系吸水面积和吸水效率等生理生态功能的基础上, 红砂选择了既有利于提高根系分布密度, 又能增强根系空间拓展能力的根系分叉数和分支角度优化配置模式.因此生长在东坡和西坡植株的二者的相关性仅达到显著水平(p < 0.05, 图1 ), 是根系分支角度由大到小、根系分叉数由小到大转变的区域. ...
不同坡向高寒退化草地狼毒株高和枝条数的权衡关系.
4
2014
... 植物根系作为植被与土壤之间物质、能量交换的主要通道(黄晶晶, 2013 ), 在植物水分和养分吸收、运移过程中有着不可替代的作用(Kong et al ., 2014 ), 影响着地上部分以及整个植株的生长和发育, 其中根系的分支状况和构型对营养物质的吸收起着关键作用(Dannowski & Block, 2005 ).根系构型是植物与环境异质性资源相互适应的结果(Malamy, 2005 ), 其对环境的适应是通过根系分支结构、空间分布格局等构型特征的改变表现出来的(Tracy et al ., 2015 ).为了应对地理环境和资源供给水平造成的胁迫和干扰, 植物根系会根据所处生境的资源条件适时调整和转化根系分叉数和分支角度等构型特征, 实现植物功能性状间的资源权衡(陈莹婷和许振柱, 2014 ).根系分叉数影响植物根系对资源物质的觅食能力和植株的固定作用(孙浩燕等, 2014 ), 根系分支角度与根系的分布范围和拓展能力紧密相关(黄晶晶, 2013 ), 二者在很大程度上决定了根系在土壤中的位置和对土体的觅养效率(Malamy, 2005 ), 反映了不同生境中植物根系在物质吸收、利用和循环中的作用.地形作为生境条件的综合指示, 控制水热因子的重新分配, 修饰植被空间分布格局, 此自然法则具有普适性(常学礼等, 2015 ).坡向是地形重要的影响因素之一, 影响了太阳辐射和降水的空间再分配, 往往导致局部小气候的产生(Cantón et al ., 2004 ), 从而改变光照、温度及土壤水分的分布, 导致群落环境和植物的生存条件产生差异(侯兆疆等, 2014 ), 进而改变植物碳资源分配格局和根系构型构建策略(Oppelt et al ., 2001 ).为了提高植株生存适合度和竞争能力, 植物根系通过分支角度和分叉数等性状的响应, 促使根系构型模式进行适应性调整, 构建与环境梯度相匹配的根系营养吸收和传输系统.因此, 从不同坡向资源条件变化的角度研究植物根系构型的空间分异格局和资源配置模式, 对于理解植物在异质生境中的环境适应机制具有重要的理论意义. ...
... ), 根系分支角度与根系的分布范围和拓展能力紧密相关(黄晶晶, 2013 ), 二者在很大程度上决定了根系在土壤中的位置和对土体的觅养效率(Malamy, 2005 ), 反映了不同生境中植物根系在物质吸收、利用和循环中的作用.地形作为生境条件的综合指示, 控制水热因子的重新分配, 修饰植被空间分布格局, 此自然法则具有普适性(常学礼等, 2015 ).坡向是地形重要的影响因素之一, 影响了太阳辐射和降水的空间再分配, 往往导致局部小气候的产生(Cantón et al ., 2004 ), 从而改变光照、温度及土壤水分的分布, 导致群落环境和植物的生存条件产生差异(侯兆疆等, 2014 ), 进而改变植物碳资源分配格局和根系构型构建策略(Oppelt et al ., 2001 ).为了提高植株生存适合度和竞争能力, 植物根系通过分支角度和分叉数等性状的响应, 促使根系构型模式进行适应性调整, 构建与环境梯度相匹配的根系营养吸收和传输系统.因此, 从不同坡向资源条件变化的角度研究植物根系构型的空间分异格局和资源配置模式, 对于理解植物在异质生境中的环境适应机制具有重要的理论意义. ...
... 植物外部形态具有高度的环境可塑性特征, 在长期生长发育的过程中, 生境差异性使其主要构件采取了不同的生态对策, 能够协调各功能性状间的资源分配模式以规避环境风险(史元春等, 2015 ).植物根系构型构建与植物自身根系碳投入和土壤水分养分的收益权衡相契合, 是植物提高自身资源获取能力的一个重要策略(杨小林等, 2008 ).研究区位于祁连山与河西走廊过渡带的荒漠草原区, 山地南坡的植物生存环境比较严酷, 红砂的生物量水平低(表1 , 表2 ), 为了提高自身的生存和繁衍能力, 红砂将更多的同化产物分配给地下根系, 着力构建强大的物质吸收和存储系统, 从而限制了地上生物量的分配比例(表1 ), 形成了相对较小的、有助于降低植株总体蒸腾耗水量的枝叶性状体系(表3 ).南坡植被稀疏(表2 )、大气温度较高, 结构松散的沙土毛细管吸收作用力弱.在土壤含水量偏低(表2 )和无地下水补给的条件下, 荒漠植物往往需要较大集水面积才能满足植株最低水平的生理耗水需求, 红砂选择了增加根系分支角度、减小分叉数的根系构型构建模式, 原因如下: 1)红砂属于浅根系植物, 绝大部分根系与地面平行伸展, 形成相对较大的根系分支角度(图1 ), 更有利于侧根趋向于水平分布、扩大根系的有效觅养空间, 同样单位(长度)的根系拥有的吸水土壤体积可能较大, 从而具有比同一生境中相邻植物更强的水资源竞争能力, 可以获得更多的生存机会(蒋礼学和李彦, 2008 ).这符合相关学者“分支角度越大, 根系越趋向于水平分布, 向根更广的范围扩展以寻求资源”的研究结论(黄晶晶, 2013 ).2)植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 保证自身的生存和生长的策略(杜建会等, 2014 ).生长在南坡的红砂减小其根系分叉数的根系构型(图1 ), 在一定程度上降低了根系内部的交叠重复, 可以避免细根间资源空间的竞争, 从而提高根系对营养物质的吸收效率来加快对土壤资源的利用, 以最佳的收益/代价关系适应贫瘠的资源环境, 与相关学者得出一致的结论, 即“当植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 减少根系分叉数的策略以降低根系内部的交叠重复, 避免细根营养物质的竞争” (单立山等, 2012 ; 杜建会等, 2014 ).因此, 分布于南坡的红砂根系分叉数和分支角度间呈极显著负相关关系(p < 0.01, 图2 ), 实现了光合产物在构件水平的优化配置. ...
... 植物个体间的相互作用实际上是可获得性资源的有限性导致的构件水平的竞争(马维玲等, 2010 ), 根系是植物营养物质摄取、运输、贮存和合成场所, 也是寻觅新的生境和营养的繁殖器官(黄晶晶等, 2013 ), 影响植株个体的生长、发育和植物种群的生活史对策(Dannowski & Block, 2005 ).东坡和西坡属于半阴和半阳坡, 是植被类型的交错带, 植被多样性较高, 群落组成较为复杂, 草地群落密度分别为17株·m-2 和15株·m-2 , 盖度为36%和34%, 高度为21.32 cm和21.70 cm, 都介于南坡和北坡之间, 植物之间的邻体干扰较大.为了增强资源竞争能力和生境适合度, 红砂选择了折中的地上、地下生物量配置模式, 兼顾了地上植株空间拓展和地下根系水分、养分吸收传输的物质需求.受地下生物量有限性的约束, 在权衡根系吸水面积和吸水效率等生理生态功能的基础上, 红砂选择了既有利于提高根系分布密度, 又能增强根系空间拓展能力的根系分叉数和分支角度优化配置模式.因此生长在东坡和西坡植株的二者的相关性仅达到显著水平(p < 0.05, 图1 ), 是根系分支角度由大到小、根系分叉数由小到大转变的区域. ...
不同林龄胡杨克隆繁殖根系分布特征及其构型.
1
2013
... 红砂(Reaumuria songarica )是一种广泛分布于我国西北荒漠地区的柽柳科小灌木, 是荒漠植被的常见建群种和优势种, 具有极强的抗干旱能力.在全球变化背景下, 红砂等荒漠植物面对日益严酷的环境压力, 需要不断调整主要功能性状构建的资源策略, 适时改变植物碳资源分配格局和根系构型构建策略(单立山等, 2013 ), 这对维持荒漠生态系统的稳定性具有重要意义.目前, 许多学者围绕植物根系分支模式的竞争能力(Glimskär, 2000 )、根系构型的拓扑结构(马新明等, 2006 )、多枝柽柳(Tamarix ramosissima )和骆驼刺(Alhagi sparsifolia )幼苗生物量分配及根系分布特征(曾凡江等, 2010 )、不同树种间细根结构和形态特征(熊德成等, 2012 )、骆驼刺群落在干旱生境下细根生产与周转(冀卫萍等, 2013 )、植物根系构型的生态适应性(杜建会等, 2014 )以及根系功能性状的可塑性与空间异质性(Druille et al ., 2015 )等方面对植物根系构型进行了系统研究, 但是有关生境梯度下植物根系构型构建的资源配置权衡机制等研究仍显不足, 限制了对异质生境中植物表型可塑性机制的解析.本文通过研究祁连山北坡荒漠草地红砂根系分叉数和分支角度的分布格局, 试图解析: (1)不同坡向红砂根系分叉数和分支角度的变化规律; (2)造成红砂根系分叉数和分支角度异速生长关系的原因, 旨在从植物根系功能性状建成的角度阐明红砂种群的环境适应性规律, 为进一步了解荒漠植物的表型可塑性机制提供理论依据. ...
不同林龄胡杨克隆繁殖根系分布特征及其构型.
1
2013
... 红砂(Reaumuria songarica )是一种广泛分布于我国西北荒漠地区的柽柳科小灌木, 是荒漠植被的常见建群种和优势种, 具有极强的抗干旱能力.在全球变化背景下, 红砂等荒漠植物面对日益严酷的环境压力, 需要不断调整主要功能性状构建的资源策略, 适时改变植物碳资源分配格局和根系构型构建策略(单立山等, 2013 ), 这对维持荒漠生态系统的稳定性具有重要意义.目前, 许多学者围绕植物根系分支模式的竞争能力(Glimskär, 2000 )、根系构型的拓扑结构(马新明等, 2006 )、多枝柽柳(Tamarix ramosissima )和骆驼刺(Alhagi sparsifolia )幼苗生物量分配及根系分布特征(曾凡江等, 2010 )、不同树种间细根结构和形态特征(熊德成等, 2012 )、骆驼刺群落在干旱生境下细根生产与周转(冀卫萍等, 2013 )、植物根系构型的生态适应性(杜建会等, 2014 )以及根系功能性状的可塑性与空间异质性(Druille et al ., 2015 )等方面对植物根系构型进行了系统研究, 但是有关生境梯度下植物根系构型构建的资源配置权衡机制等研究仍显不足, 限制了对异质生境中植物表型可塑性机制的解析.本文通过研究祁连山北坡荒漠草地红砂根系分叉数和分支角度的分布格局, 试图解析: (1)不同坡向红砂根系分叉数和分支角度的变化规律; (2)造成红砂根系分叉数和分支角度异速生长关系的原因, 旨在从植物根系功能性状建成的角度阐明红砂种群的环境适应性规律, 为进一步了解荒漠植物的表型可塑性机制提供理论依据. ...
干旱区骆驼刺群落细根生产与周转.
1
2013
... 植物外部形态具有高度的环境可塑性特征, 在长期生长发育的过程中, 生境差异性使其主要构件采取了不同的生态对策, 能够协调各功能性状间的资源分配模式以规避环境风险(史元春等, 2015 ).植物根系构型构建与植物自身根系碳投入和土壤水分养分的收益权衡相契合, 是植物提高自身资源获取能力的一个重要策略(杨小林等, 2008 ).研究区位于祁连山与河西走廊过渡带的荒漠草原区, 山地南坡的植物生存环境比较严酷, 红砂的生物量水平低(表1 , 表2 ), 为了提高自身的生存和繁衍能力, 红砂将更多的同化产物分配给地下根系, 着力构建强大的物质吸收和存储系统, 从而限制了地上生物量的分配比例(表1 ), 形成了相对较小的、有助于降低植株总体蒸腾耗水量的枝叶性状体系(表3 ).南坡植被稀疏(表2 )、大气温度较高, 结构松散的沙土毛细管吸收作用力弱.在土壤含水量偏低(表2 )和无地下水补给的条件下, 荒漠植物往往需要较大集水面积才能满足植株最低水平的生理耗水需求, 红砂选择了增加根系分支角度、减小分叉数的根系构型构建模式, 原因如下: 1)红砂属于浅根系植物, 绝大部分根系与地面平行伸展, 形成相对较大的根系分支角度(图1 ), 更有利于侧根趋向于水平分布、扩大根系的有效觅养空间, 同样单位(长度)的根系拥有的吸水土壤体积可能较大, 从而具有比同一生境中相邻植物更强的水资源竞争能力, 可以获得更多的生存机会(蒋礼学和李彦, 2008 ).这符合相关学者“分支角度越大, 根系越趋向于水平分布, 向根更广的范围扩展以寻求资源”的研究结论(黄晶晶, 2013 ).2)植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 保证自身的生存和生长的策略(杜建会等, 2014 ).生长在南坡的红砂减小其根系分叉数的根系构型(图1 ), 在一定程度上降低了根系内部的交叠重复, 可以避免细根间资源空间的竞争, 从而提高根系对营养物质的吸收效率来加快对土壤资源的利用, 以最佳的收益/代价关系适应贫瘠的资源环境, 与相关学者得出一致的结论, 即“当植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 减少根系分叉数的策略以降低根系内部的交叠重复, 避免细根营养物质的竞争” (单立山等, 2012 ; 杜建会等, 2014 ).因此, 分布于南坡的红砂根系分叉数和分支角度间呈极显著负相关关系(p < 0.01, 图2 ), 实现了光合产物在构件水平的优化配置. ...
干旱区骆驼刺群落细根生产与周转.
1
2013
... 植物外部形态具有高度的环境可塑性特征, 在长期生长发育的过程中, 生境差异性使其主要构件采取了不同的生态对策, 能够协调各功能性状间的资源分配模式以规避环境风险(史元春等, 2015 ).植物根系构型构建与植物自身根系碳投入和土壤水分养分的收益权衡相契合, 是植物提高自身资源获取能力的一个重要策略(杨小林等, 2008 ).研究区位于祁连山与河西走廊过渡带的荒漠草原区, 山地南坡的植物生存环境比较严酷, 红砂的生物量水平低(表1 , 表2 ), 为了提高自身的生存和繁衍能力, 红砂将更多的同化产物分配给地下根系, 着力构建强大的物质吸收和存储系统, 从而限制了地上生物量的分配比例(表1 ), 形成了相对较小的、有助于降低植株总体蒸腾耗水量的枝叶性状体系(表3 ).南坡植被稀疏(表2 )、大气温度较高, 结构松散的沙土毛细管吸收作用力弱.在土壤含水量偏低(表2 )和无地下水补给的条件下, 荒漠植物往往需要较大集水面积才能满足植株最低水平的生理耗水需求, 红砂选择了增加根系分支角度、减小分叉数的根系构型构建模式, 原因如下: 1)红砂属于浅根系植物, 绝大部分根系与地面平行伸展, 形成相对较大的根系分支角度(图1 ), 更有利于侧根趋向于水平分布、扩大根系的有效觅养空间, 同样单位(长度)的根系拥有的吸水土壤体积可能较大, 从而具有比同一生境中相邻植物更强的水资源竞争能力, 可以获得更多的生存机会(蒋礼学和李彦, 2008 ).这符合相关学者“分支角度越大, 根系越趋向于水平分布, 向根更广的范围扩展以寻求资源”的研究结论(黄晶晶, 2013 ).2)植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 保证自身的生存和生长的策略(杜建会等, 2014 ).生长在南坡的红砂减小其根系分叉数的根系构型(图1 ), 在一定程度上降低了根系内部的交叠重复, 可以避免细根间资源空间的竞争, 从而提高根系对营养物质的吸收效率来加快对土壤资源的利用, 以最佳的收益/代价关系适应贫瘠的资源环境, 与相关学者得出一致的结论, 即“当植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 减少根系分叉数的策略以降低根系内部的交叠重复, 避免细根营养物质的竞争” (单立山等, 2012 ; 杜建会等, 2014 ).因此, 分布于南坡的红砂根系分叉数和分支角度间呈极显著负相关关系(p < 0.01, 图2 ), 实现了光合产物在构件水平的优化配置. ...
三种荒漠灌木根系的构形特征与叶性因子对干旱生境的适应性比较.
1
2008
... 植物根系作为植被与土壤之间物质、能量交换的主要通道(黄晶晶, 2013 ), 在植物水分和养分吸收、运移过程中有着不可替代的作用(Kong et al ., 2014 ), 影响着地上部分以及整个植株的生长和发育, 其中根系的分支状况和构型对营养物质的吸收起着关键作用(Dannowski & Block, 2005 ).根系构型是植物与环境异质性资源相互适应的结果(Malamy, 2005 ), 其对环境的适应是通过根系分支结构、空间分布格局等构型特征的改变表现出来的(Tracy et al ., 2015 ).为了应对地理环境和资源供给水平造成的胁迫和干扰, 植物根系会根据所处生境的资源条件适时调整和转化根系分叉数和分支角度等构型特征, 实现植物功能性状间的资源权衡(陈莹婷和许振柱, 2014 ).根系分叉数影响植物根系对资源物质的觅食能力和植株的固定作用(孙浩燕等, 2014 ), 根系分支角度与根系的分布范围和拓展能力紧密相关(黄晶晶, 2013 ), 二者在很大程度上决定了根系在土壤中的位置和对土体的觅养效率(Malamy, 2005 ), 反映了不同生境中植物根系在物质吸收、利用和循环中的作用.地形作为生境条件的综合指示, 控制水热因子的重新分配, 修饰植被空间分布格局, 此自然法则具有普适性(常学礼等, 2015 ).坡向是地形重要的影响因素之一, 影响了太阳辐射和降水的空间再分配, 往往导致局部小气候的产生(Cantón et al ., 2004 ), 从而改变光照、温度及土壤水分的分布, 导致群落环境和植物的生存条件产生差异(侯兆疆等, 2014 ), 进而改变植物碳资源分配格局和根系构型构建策略(Oppelt et al ., 2001 ).为了提高植株生存适合度和竞争能力, 植物根系通过分支角度和分叉数等性状的响应, 促使根系构型模式进行适应性调整, 构建与环境梯度相匹配的根系营养吸收和传输系统.因此, 从不同坡向资源条件变化的角度研究植物根系构型的空间分异格局和资源配置模式, 对于理解植物在异质生境中的环境适应机制具有重要的理论意义. ...
三种荒漠灌木根系的构形特征与叶性因子对干旱生境的适应性比较.
1
2008
... 植物根系作为植被与土壤之间物质、能量交换的主要通道(黄晶晶, 2013 ), 在植物水分和养分吸收、运移过程中有着不可替代的作用(Kong et al ., 2014 ), 影响着地上部分以及整个植株的生长和发育, 其中根系的分支状况和构型对营养物质的吸收起着关键作用(Dannowski & Block, 2005 ).根系构型是植物与环境异质性资源相互适应的结果(Malamy, 2005 ), 其对环境的适应是通过根系分支结构、空间分布格局等构型特征的改变表现出来的(Tracy et al ., 2015 ).为了应对地理环境和资源供给水平造成的胁迫和干扰, 植物根系会根据所处生境的资源条件适时调整和转化根系分叉数和分支角度等构型特征, 实现植物功能性状间的资源权衡(陈莹婷和许振柱, 2014 ).根系分叉数影响植物根系对资源物质的觅食能力和植株的固定作用(孙浩燕等, 2014 ), 根系分支角度与根系的分布范围和拓展能力紧密相关(黄晶晶, 2013 ), 二者在很大程度上决定了根系在土壤中的位置和对土体的觅养效率(Malamy, 2005 ), 反映了不同生境中植物根系在物质吸收、利用和循环中的作用.地形作为生境条件的综合指示, 控制水热因子的重新分配, 修饰植被空间分布格局, 此自然法则具有普适性(常学礼等, 2015 ).坡向是地形重要的影响因素之一, 影响了太阳辐射和降水的空间再分配, 往往导致局部小气候的产生(Cantón et al ., 2004 ), 从而改变光照、温度及土壤水分的分布, 导致群落环境和植物的生存条件产生差异(侯兆疆等, 2014 ), 进而改变植物碳资源分配格局和根系构型构建策略(Oppelt et al ., 2001 ).为了提高植株生存适合度和竞争能力, 植物根系通过分支角度和分叉数等性状的响应, 促使根系构型模式进行适应性调整, 构建与环境梯度相匹配的根系营养吸收和传输系统.因此, 从不同坡向资源条件变化的角度研究植物根系构型的空间分异格局和资源配置模式, 对于理解植物在异质生境中的环境适应机制具有重要的理论意义. ...
Designer crops: Optimal root system architecture for nutrient acquisition.
1
2014
... 植物在长期的进化过程中, 通过生理和发育调节形成与环境相适应的表型特征, 进而具备了提高植物水分、养分等资源利用效率的功能(李西良等, 2014 ).在异质性生境中, 植物根系面对水分和养分吸收、运移和存储的环境压迫, 其分叉数和分支角度可能会发生适应性变化, 并通过物质能量吸收、固着格局的调整反作用于植被生态系统的稳定和种群更新机制, 体现了植物根系进行空间探索以适应环境胁迫的能力(Zhao et al ., 2013 ).本研究发现, 随着坡向由南坡向西坡、东坡和北坡的转变, 红砂根系分支角度逐渐减小, 而根系分叉数呈相反的变化趋势(图1 ), 在有限的资源环境中, 红砂分配给根系分叉数和分支角度资源间存在着“此消彼长”的权衡关系.坡向梯度上红砂根系分叉数和分支角度关系的差异性是植物根系构建的资源配置模式对生境异质性的适应对策. ...
草甸草原羊草茎叶功能性状对长期过度放牧的可塑性响应.
1
2014
... 植物个体间的相互作用实际上是可获得性资源的有限性导致的构件水平的竞争(马维玲等, 2010 ), 根系是植物营养物质摄取、运输、贮存和合成场所, 也是寻觅新的生境和营养的繁殖器官(黄晶晶等, 2013 ), 影响植株个体的生长、发育和植物种群的生活史对策(Dannowski & Block, 2005 ).东坡和西坡属于半阴和半阳坡, 是植被类型的交错带, 植被多样性较高, 群落组成较为复杂, 草地群落密度分别为17株·m-2 和15株·m-2 , 盖度为36%和34%, 高度为21.32 cm和21.70 cm, 都介于南坡和北坡之间, 植物之间的邻体干扰较大.为了增强资源竞争能力和生境适合度, 红砂选择了折中的地上、地下生物量配置模式, 兼顾了地上植株空间拓展和地下根系水分、养分吸收传输的物质需求.受地下生物量有限性的约束, 在权衡根系吸水面积和吸水效率等生理生态功能的基础上, 红砂选择了既有利于提高根系分布密度, 又能增强根系空间拓展能力的根系分叉数和分支角度优化配置模式.因此生长在东坡和西坡植株的二者的相关性仅达到显著水平(p < 0.05, 图1 ), 是根系分支角度由大到小、根系分叉数由小到大转变的区域. ...
草甸草原羊草茎叶功能性状对长期过度放牧的可塑性响应.
1
2014
... 植物个体间的相互作用实际上是可获得性资源的有限性导致的构件水平的竞争(马维玲等, 2010 ), 根系是植物营养物质摄取、运输、贮存和合成场所, 也是寻觅新的生境和营养的繁殖器官(黄晶晶等, 2013 ), 影响植株个体的生长、发育和植物种群的生活史对策(Dannowski & Block, 2005 ).东坡和西坡属于半阴和半阳坡, 是植被类型的交错带, 植被多样性较高, 群落组成较为复杂, 草地群落密度分别为17株·m-2 和15株·m-2 , 盖度为36%和34%, 高度为21.32 cm和21.70 cm, 都介于南坡和北坡之间, 植物之间的邻体干扰较大.为了增强资源竞争能力和生境适合度, 红砂选择了折中的地上、地下生物量配置模式, 兼顾了地上植株空间拓展和地下根系水分、养分吸收传输的物质需求.受地下生物量有限性的约束, 在权衡根系吸水面积和吸水效率等生理生态功能的基础上, 红砂选择了既有利于提高根系分布密度, 又能增强根系空间拓展能力的根系分叉数和分支角度优化配置模式.因此生长在东坡和西坡植株的二者的相关性仅达到显著水平(p < 0.05, 图1 ), 是根系分支角度由大到小、根系分叉数由小到大转变的区域. ...
青藏高原高寒草甸植株性状和生物量分配的海拔梯度变异
1
2010
... 红砂(Reaumuria songarica )是一种广泛分布于我国西北荒漠地区的柽柳科小灌木, 是荒漠植被的常见建群种和优势种, 具有极强的抗干旱能力.在全球变化背景下, 红砂等荒漠植物面对日益严酷的环境压力, 需要不断调整主要功能性状构建的资源策略, 适时改变植物碳资源分配格局和根系构型构建策略(单立山等, 2013 ), 这对维持荒漠生态系统的稳定性具有重要意义.目前, 许多学者围绕植物根系分支模式的竞争能力(Glimskär, 2000 )、根系构型的拓扑结构(马新明等, 2006 )、多枝柽柳(Tamarix ramosissima )和骆驼刺(Alhagi sparsifolia )幼苗生物量分配及根系分布特征(曾凡江等, 2010 )、不同树种间细根结构和形态特征(熊德成等, 2012 )、骆驼刺群落在干旱生境下细根生产与周转(冀卫萍等, 2013 )、植物根系构型的生态适应性(杜建会等, 2014 )以及根系功能性状的可塑性与空间异质性(Druille et al ., 2015 )等方面对植物根系构型进行了系统研究, 但是有关生境梯度下植物根系构型构建的资源配置权衡机制等研究仍显不足, 限制了对异质生境中植物表型可塑性机制的解析.本文通过研究祁连山北坡荒漠草地红砂根系分叉数和分支角度的分布格局, 试图解析: (1)不同坡向红砂根系分叉数和分支角度的变化规律; (2)造成红砂根系分叉数和分支角度异速生长关系的原因, 旨在从植物根系功能性状建成的角度阐明红砂种群的环境适应性规律, 为进一步了解荒漠植物的表型可塑性机制提供理论依据. ...
青藏高原高寒草甸植株性状和生物量分配的海拔梯度变异
1
2010
... 红砂(Reaumuria songarica )是一种广泛分布于我国西北荒漠地区的柽柳科小灌木, 是荒漠植被的常见建群种和优势种, 具有极强的抗干旱能力.在全球变化背景下, 红砂等荒漠植物面对日益严酷的环境压力, 需要不断调整主要功能性状构建的资源策略, 适时改变植物碳资源分配格局和根系构型构建策略(单立山等, 2013 ), 这对维持荒漠生态系统的稳定性具有重要意义.目前, 许多学者围绕植物根系分支模式的竞争能力(Glimskär, 2000 )、根系构型的拓扑结构(马新明等, 2006 )、多枝柽柳(Tamarix ramosissima )和骆驼刺(Alhagi sparsifolia )幼苗生物量分配及根系分布特征(曾凡江等, 2010 )、不同树种间细根结构和形态特征(熊德成等, 2012 )、骆驼刺群落在干旱生境下细根生产与周转(冀卫萍等, 2013 )、植物根系构型的生态适应性(杜建会等, 2014 )以及根系功能性状的可塑性与空间异质性(Druille et al ., 2015 )等方面对植物根系构型进行了系统研究, 但是有关生境梯度下植物根系构型构建的资源配置权衡机制等研究仍显不足, 限制了对异质生境中植物表型可塑性机制的解析.本文通过研究祁连山北坡荒漠草地红砂根系分叉数和分支角度的分布格局, 试图解析: (1)不同坡向红砂根系分叉数和分支角度的变化规律; (2)造成红砂根系分叉数和分支角度异速生长关系的原因, 旨在从植物根系功能性状建成的角度阐明红砂种群的环境适应性规律, 为进一步了解荒漠植物的表型可塑性机制提供理论依据. ...
大田期烟草根系构型参数的动态变化.
2
2006
... 植物根系作为植被与土壤之间物质、能量交换的主要通道(黄晶晶, 2013 ), 在植物水分和养分吸收、运移过程中有着不可替代的作用(Kong et al ., 2014 ), 影响着地上部分以及整个植株的生长和发育, 其中根系的分支状况和构型对营养物质的吸收起着关键作用(Dannowski & Block, 2005 ).根系构型是植物与环境异质性资源相互适应的结果(Malamy, 2005 ), 其对环境的适应是通过根系分支结构、空间分布格局等构型特征的改变表现出来的(Tracy et al ., 2015 ).为了应对地理环境和资源供给水平造成的胁迫和干扰, 植物根系会根据所处生境的资源条件适时调整和转化根系分叉数和分支角度等构型特征, 实现植物功能性状间的资源权衡(陈莹婷和许振柱, 2014 ).根系分叉数影响植物根系对资源物质的觅食能力和植株的固定作用(孙浩燕等, 2014 ), 根系分支角度与根系的分布范围和拓展能力紧密相关(黄晶晶, 2013 ), 二者在很大程度上决定了根系在土壤中的位置和对土体的觅养效率(Malamy, 2005 ), 反映了不同生境中植物根系在物质吸收、利用和循环中的作用.地形作为生境条件的综合指示, 控制水热因子的重新分配, 修饰植被空间分布格局, 此自然法则具有普适性(常学礼等, 2015 ).坡向是地形重要的影响因素之一, 影响了太阳辐射和降水的空间再分配, 往往导致局部小气候的产生(Cantón et al ., 2004 ), 从而改变光照、温度及土壤水分的分布, 导致群落环境和植物的生存条件产生差异(侯兆疆等, 2014 ), 进而改变植物碳资源分配格局和根系构型构建策略(Oppelt et al ., 2001 ).为了提高植株生存适合度和竞争能力, 植物根系通过分支角度和分叉数等性状的响应, 促使根系构型模式进行适应性调整, 构建与环境梯度相匹配的根系营养吸收和传输系统.因此, 从不同坡向资源条件变化的角度研究植物根系构型的空间分异格局和资源配置模式, 对于理解植物在异质生境中的环境适应机制具有重要的理论意义. ...
... ), 二者在很大程度上决定了根系在土壤中的位置和对土体的觅养效率(Malamy, 2005 ), 反映了不同生境中植物根系在物质吸收、利用和循环中的作用.地形作为生境条件的综合指示, 控制水热因子的重新分配, 修饰植被空间分布格局, 此自然法则具有普适性(常学礼等, 2015 ).坡向是地形重要的影响因素之一, 影响了太阳辐射和降水的空间再分配, 往往导致局部小气候的产生(Cantón et al ., 2004 ), 从而改变光照、温度及土壤水分的分布, 导致群落环境和植物的生存条件产生差异(侯兆疆等, 2014 ), 进而改变植物碳资源分配格局和根系构型构建策略(Oppelt et al ., 2001 ).为了提高植株生存适合度和竞争能力, 植物根系通过分支角度和分叉数等性状的响应, 促使根系构型模式进行适应性调整, 构建与环境梯度相匹配的根系营养吸收和传输系统.因此, 从不同坡向资源条件变化的角度研究植物根系构型的空间分异格局和资源配置模式, 对于理解植物在异质生境中的环境适应机制具有重要的理论意义. ...
大田期烟草根系构型参数的动态变化.
2
2006
... 植物根系作为植被与土壤之间物质、能量交换的主要通道(黄晶晶, 2013 ), 在植物水分和养分吸收、运移过程中有着不可替代的作用(Kong et al ., 2014 ), 影响着地上部分以及整个植株的生长和发育, 其中根系的分支状况和构型对营养物质的吸收起着关键作用(Dannowski & Block, 2005 ).根系构型是植物与环境异质性资源相互适应的结果(Malamy, 2005 ), 其对环境的适应是通过根系分支结构、空间分布格局等构型特征的改变表现出来的(Tracy et al ., 2015 ).为了应对地理环境和资源供给水平造成的胁迫和干扰, 植物根系会根据所处生境的资源条件适时调整和转化根系分叉数和分支角度等构型特征, 实现植物功能性状间的资源权衡(陈莹婷和许振柱, 2014 ).根系分叉数影响植物根系对资源物质的觅食能力和植株的固定作用(孙浩燕等, 2014 ), 根系分支角度与根系的分布范围和拓展能力紧密相关(黄晶晶, 2013 ), 二者在很大程度上决定了根系在土壤中的位置和对土体的觅养效率(Malamy, 2005 ), 反映了不同生境中植物根系在物质吸收、利用和循环中的作用.地形作为生境条件的综合指示, 控制水热因子的重新分配, 修饰植被空间分布格局, 此自然法则具有普适性(常学礼等, 2015 ).坡向是地形重要的影响因素之一, 影响了太阳辐射和降水的空间再分配, 往往导致局部小气候的产生(Cantón et al ., 2004 ), 从而改变光照、温度及土壤水分的分布, 导致群落环境和植物的生存条件产生差异(侯兆疆等, 2014 ), 进而改变植物碳资源分配格局和根系构型构建策略(Oppelt et al ., 2001 ).为了提高植株生存适合度和竞争能力, 植物根系通过分支角度和分叉数等性状的响应, 促使根系构型模式进行适应性调整, 构建与环境梯度相匹配的根系营养吸收和传输系统.因此, 从不同坡向资源条件变化的角度研究植物根系构型的空间分异格局和资源配置模式, 对于理解植物在异质生境中的环境适应机制具有重要的理论意义. ...
... ), 二者在很大程度上决定了根系在土壤中的位置和对土体的觅养效率(Malamy, 2005 ), 反映了不同生境中植物根系在物质吸收、利用和循环中的作用.地形作为生境条件的综合指示, 控制水热因子的重新分配, 修饰植被空间分布格局, 此自然法则具有普适性(常学礼等, 2015 ).坡向是地形重要的影响因素之一, 影响了太阳辐射和降水的空间再分配, 往往导致局部小气候的产生(Cantón et al ., 2004 ), 从而改变光照、温度及土壤水分的分布, 导致群落环境和植物的生存条件产生差异(侯兆疆等, 2014 ), 进而改变植物碳资源分配格局和根系构型构建策略(Oppelt et al ., 2001 ).为了提高植株生存适合度和竞争能力, 植物根系通过分支角度和分叉数等性状的响应, 促使根系构型模式进行适应性调整, 构建与环境梯度相匹配的根系营养吸收和传输系统.因此, 从不同坡向资源条件变化的角度研究植物根系构型的空间分异格局和资源配置模式, 对于理解植物在异质生境中的环境适应机制具有重要的理论意义. ...
Intrinsic and environmental response pathways that regulate root system architecture.
1
2005
... 在地形图上将研究区划分为4个坡向, 每个坡向沿垂直海拔梯度(1940-2115 m)间隔30 m布置1条长140 m、宽10 m的水平样带, 总计20条水平样带.在多次现场踏勘的基础上, 在每条水平样带上间隔20 m设置1个5 m × 5 m的样方, 共计120个样方, GPS测量每个样方的经纬度和海拔.第一, 进行草地群落学特征调查, 测定样方内所有植物的盖度、高度、密度和地上生物量.第二, 每一样方内随机选择1株生长良好的红砂, 每个坡向共计30株红砂用于测定植株的生物量和根系性状, 首先将每株红砂的地上枝条刈割并装入自封袋, 然后采用壕沟法和根系跟踪相结合的方法挖掘整株根系, 在离植株2 m处挖一条2 m × 0.6 m × 1.6 m壕沟, 然后利用小铁铲慢慢清除植株周围的泥沙到壕沟并不断清理沟中层积的沙土, 直至直径>3 mm的整株根系完全暴露, 因为直径<3 mm根系构型的重建在研究中很难实现(Oppelt et al ., 2000 , 2005 ), 尽量保持根系在自然空间中的位置.根系完全暴露后, 采用50 cm × 50 cm的网格确定根系的分布位置(直到没有根系出现为止), 按照1:50的比例在35 cm × 25 cm的坐标纸上准确绘制根系构型图, 然后把植株根系装入尼龙袋带回实验室.在室内将根系置于细筛之上用水冲洗, 同时去除其他杂质, 仅留下红砂的根系; 用游标卡尺、卷尺和量角器等工具, 分别测定所有样点红砂的根长、分叉数和分支夹角(°); 之后把植株根系和地上部分分别装入信封, 在80 ℃烘箱里烘24 h后用电子天平(0.0001 g)称质量并记录.第三, 进行土壤水分采样, 每个样方内随机选取3个样点, 用土钻(直径4 cm)采集0-100 cm土层范围分5层间隔20 cm取土壤样品, 钻取的土壤先按土层分取, 之后将各层土壤进行混合, 重复3次, 剔除样品中明显的植物根段和枯落物等杂质, 装入编号的铝盒中, 带回实验室, 在105 ℃的烘箱内烘12 h取出称质量, 计算出各样地0-100 cm土层土壤质量含水量. ...
Structure and fractal dimensions of root systems of four co-occurring fruit tree species from Botswana.
1
2000
... 植物根系作为植被与土壤之间物质、能量交换的主要通道(黄晶晶, 2013 ), 在植物水分和养分吸收、运移过程中有着不可替代的作用(Kong et al ., 2014 ), 影响着地上部分以及整个植株的生长和发育, 其中根系的分支状况和构型对营养物质的吸收起着关键作用(Dannowski & Block, 2005 ).根系构型是植物与环境异质性资源相互适应的结果(Malamy, 2005 ), 其对环境的适应是通过根系分支结构、空间分布格局等构型特征的改变表现出来的(Tracy et al ., 2015 ).为了应对地理环境和资源供给水平造成的胁迫和干扰, 植物根系会根据所处生境的资源条件适时调整和转化根系分叉数和分支角度等构型特征, 实现植物功能性状间的资源权衡(陈莹婷和许振柱, 2014 ).根系分叉数影响植物根系对资源物质的觅食能力和植株的固定作用(孙浩燕等, 2014 ), 根系分支角度与根系的分布范围和拓展能力紧密相关(黄晶晶, 2013 ), 二者在很大程度上决定了根系在土壤中的位置和对土体的觅养效率(Malamy, 2005 ), 反映了不同生境中植物根系在物质吸收、利用和循环中的作用.地形作为生境条件的综合指示, 控制水热因子的重新分配, 修饰植被空间分布格局, 此自然法则具有普适性(常学礼等, 2015 ).坡向是地形重要的影响因素之一, 影响了太阳辐射和降水的空间再分配, 往往导致局部小气候的产生(Cantón et al ., 2004 ), 从而改变光照、温度及土壤水分的分布, 导致群落环境和植物的生存条件产生差异(侯兆疆等, 2014 ), 进而改变植物碳资源分配格局和根系构型构建策略(Oppelt et al ., 2001 ).为了提高植株生存适合度和竞争能力, 植物根系通过分支角度和分叉数等性状的响应, 促使根系构型模式进行适应性调整, 构建与环境梯度相匹配的根系营养吸收和传输系统.因此, 从不同坡向资源条件变化的角度研究植物根系构型的空间分异格局和资源配置模式, 对于理解植物在异质生境中的环境适应机制具有重要的理论意义. ...
Topology, scaling relations and Leonardo’s rule in root systems from African tree species.
2
2001
... 在地形图上将研究区划分为4个坡向, 每个坡向沿垂直海拔梯度(1940-2115 m)间隔30 m布置1条长140 m、宽10 m的水平样带, 总计20条水平样带.在多次现场踏勘的基础上, 在每条水平样带上间隔20 m设置1个5 m × 5 m的样方, 共计120个样方, GPS测量每个样方的经纬度和海拔.第一, 进行草地群落学特征调查, 测定样方内所有植物的盖度、高度、密度和地上生物量.第二, 每一样方内随机选择1株生长良好的红砂, 每个坡向共计30株红砂用于测定植株的生物量和根系性状, 首先将每株红砂的地上枝条刈割并装入自封袋, 然后采用壕沟法和根系跟踪相结合的方法挖掘整株根系, 在离植株2 m处挖一条2 m × 0.6 m × 1.6 m壕沟, 然后利用小铁铲慢慢清除植株周围的泥沙到壕沟并不断清理沟中层积的沙土, 直至直径>3 mm的整株根系完全暴露, 因为直径<3 mm根系构型的重建在研究中很难实现(Oppelt et al ., 2000 , 2005 ), 尽量保持根系在自然空间中的位置.根系完全暴露后, 采用50 cm × 50 cm的网格确定根系的分布位置(直到没有根系出现为止), 按照1:50的比例在35 cm × 25 cm的坐标纸上准确绘制根系构型图, 然后把植株根系装入尼龙袋带回实验室.在室内将根系置于细筛之上用水冲洗, 同时去除其他杂质, 仅留下红砂的根系; 用游标卡尺、卷尺和量角器等工具, 分别测定所有样点红砂的根长、分叉数和分支夹角(°); 之后把植株根系和地上部分分别装入信封, 在80 ℃烘箱里烘24 h后用电子天平(0.0001 g)称质量并记录.第三, 进行土壤水分采样, 每个样方内随机选取3个样点, 用土钻(直径4 cm)采集0-100 cm土层范围分5层间隔20 cm取土壤样品, 钻取的土壤先按土层分取, 之后将各层土壤进行混合, 重复3次, 剔除样品中明显的植物根段和枯落物等杂质, 装入编号的铝盒中, 带回实验室, 在105 ℃的烘箱内烘12 h取出称质量, 计算出各样地0-100 cm土层土壤质量含水量. ...
... 植物构件的资源分配是与栖息环境相对应的种群特征的反映方式, 它反映了种群特定的生态适应策略(张茜等, 2014 ).根系构型会对周围环境产生适应性的改变, 表现出不一样的适应特性(Oppelt et al ., 2005 ).相对于南坡, 山地北坡草地群落的盖度、密度均较大, 红砂在群落中没有高度优势(表1 , 表2 ), 相对拥挤的群落环境使植物面临着严重的邻体干扰, 红砂将较多的生物量分配给地上枝叶以利于植株获得高度优势, 增强枝叶的捕光能力, 无形中限制了分配给根系的生物量比例(表1 ).在根冠比较低的情况下, 红砂需要将有限的资源用于构建根系的吸收面积, 增强根系的“觅食”能力(董鸣, 1996 ), 以最大限度保证植株在生境竞争中占据优势, 红砂选择了增加根系分叉数、减小分支角度的根系构型.原因如下: 1)在土壤含水量相对较高的北坡(表2 ), 红砂以根系分叉数增加速度大于分支角度减小速度的资源配置模式(图2 ), 通过增大根系分叉数(图1 ), 有效增加了根系分支能力和单位土体中根系的拥挤度, 构建了密集的根系网络系统, 降低了根系传输系统的碳需求量, 实现了单位根系土壤拓展体积较小状态下的植株生存的水分供给需求, 这与单立山等(2013)对红砂和白刺(Nitraria tangutorum )、杜建会等(2014)对塔克拉玛干柽柳(Tamarix taklamakanensis )的研究结果相一致.2)植物根系在深层土壤的分布有利于其吸收利用深层土壤水分, 促进植株的生长发育(席海洋等, 2011 ).北坡群落密度高(表2 )、植物之间拥挤度大, 种内和种间容易造成根系重叠竞争, 空间上无法满足大范围根系的构型塑造.红砂采取缩小根系水平范围, 将根系分支角度减小, 扩大根系在深层土壤中的垂直分布(表1 ), 避免与群落中其他植株根系的剧烈竞争.同时红砂植物通过增大比根长提高对养分和水分的利用效率来加快对土壤资源的利用(表1 ), 提高自身的竞争力以维持生存.因此, 红砂根系分叉数和分支角度间呈极显著负相关关系(p < 0.01, 图2 ), 即以资源权衡分配机制进化响应诸多竞争压力, 维持植物生长的水分条件, 从而有利于生物多样性的群落结构, 这符合相关学者“分叉数较大的植物能够迅速占据大量土壤空间快速生长, 适宜生长在土壤资源丰富的环境中”的结论(周艳松和王立群, 2011 ; 孙浩燕等, 2014 ). ...
Contrasting rooting patterns of some arid-zone fruit tree species from Botswana-II. Coarse root distribution.
1
2005
... 植物外部形态具有高度的环境可塑性特征, 在长期生长发育的过程中, 生境差异性使其主要构件采取了不同的生态对策, 能够协调各功能性状间的资源分配模式以规避环境风险(史元春等, 2015 ).植物根系构型构建与植物自身根系碳投入和土壤水分养分的收益权衡相契合, 是植物提高自身资源获取能力的一个重要策略(杨小林等, 2008 ).研究区位于祁连山与河西走廊过渡带的荒漠草原区, 山地南坡的植物生存环境比较严酷, 红砂的生物量水平低(表1 , 表2 ), 为了提高自身的生存和繁衍能力, 红砂将更多的同化产物分配给地下根系, 着力构建强大的物质吸收和存储系统, 从而限制了地上生物量的分配比例(表1 ), 形成了相对较小的、有助于降低植株总体蒸腾耗水量的枝叶性状体系(表3 ).南坡植被稀疏(表2 )、大气温度较高, 结构松散的沙土毛细管吸收作用力弱.在土壤含水量偏低(表2 )和无地下水补给的条件下, 荒漠植物往往需要较大集水面积才能满足植株最低水平的生理耗水需求, 红砂选择了增加根系分支角度、减小分叉数的根系构型构建模式, 原因如下: 1)红砂属于浅根系植物, 绝大部分根系与地面平行伸展, 形成相对较大的根系分支角度(图1 ), 更有利于侧根趋向于水平分布、扩大根系的有效觅养空间, 同样单位(长度)的根系拥有的吸水土壤体积可能较大, 从而具有比同一生境中相邻植物更强的水资源竞争能力, 可以获得更多的生存机会(蒋礼学和李彦, 2008 ).这符合相关学者“分支角度越大, 根系越趋向于水平分布, 向根更广的范围扩展以寻求资源”的研究结论(黄晶晶, 2013 ).2)植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 保证自身的生存和生长的策略(杜建会等, 2014 ).生长在南坡的红砂减小其根系分叉数的根系构型(图1 ), 在一定程度上降低了根系内部的交叠重复, 可以避免细根间资源空间的竞争, 从而提高根系对营养物质的吸收效率来加快对土壤资源的利用, 以最佳的收益/代价关系适应贫瘠的资源环境, 与相关学者得出一致的结论, 即“当植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 减少根系分叉数的策略以降低根系内部的交叠重复, 避免细根营养物质的竞争” (单立山等, 2012 ; 杜建会等, 2014 ).因此, 分布于南坡的红砂根系分叉数和分支角度间呈极显著负相关关系(p < 0.01, 图2 ), 实现了光合产物在构件水平的优化配置. ...
红砂根系构型对干旱的生态适应.
1
2012
... 红砂(Reaumuria songarica )是一种广泛分布于我国西北荒漠地区的柽柳科小灌木, 是荒漠植被的常见建群种和优势种, 具有极强的抗干旱能力.在全球变化背景下, 红砂等荒漠植物面对日益严酷的环境压力, 需要不断调整主要功能性状构建的资源策略, 适时改变植物碳资源分配格局和根系构型构建策略(单立山等, 2013 ), 这对维持荒漠生态系统的稳定性具有重要意义.目前, 许多学者围绕植物根系分支模式的竞争能力(Glimskär, 2000 )、根系构型的拓扑结构(马新明等, 2006 )、多枝柽柳(Tamarix ramosissima )和骆驼刺(Alhagi sparsifolia )幼苗生物量分配及根系分布特征(曾凡江等, 2010 )、不同树种间细根结构和形态特征(熊德成等, 2012 )、骆驼刺群落在干旱生境下细根生产与周转(冀卫萍等, 2013 )、植物根系构型的生态适应性(杜建会等, 2014 )以及根系功能性状的可塑性与空间异质性(Druille et al ., 2015 )等方面对植物根系构型进行了系统研究, 但是有关生境梯度下植物根系构型构建的资源配置权衡机制等研究仍显不足, 限制了对异质生境中植物表型可塑性机制的解析.本文通过研究祁连山北坡荒漠草地红砂根系分叉数和分支角度的分布格局, 试图解析: (1)不同坡向红砂根系分叉数和分支角度的变化规律; (2)造成红砂根系分叉数和分支角度异速生长关系的原因, 旨在从植物根系功能性状建成的角度阐明红砂种群的环境适应性规律, 为进一步了解荒漠植物的表型可塑性机制提供理论依据. ...
红砂根系构型对干旱的生态适应.
1
2012
... 红砂(Reaumuria songarica )是一种广泛分布于我国西北荒漠地区的柽柳科小灌木, 是荒漠植被的常见建群种和优势种, 具有极强的抗干旱能力.在全球变化背景下, 红砂等荒漠植物面对日益严酷的环境压力, 需要不断调整主要功能性状构建的资源策略, 适时改变植物碳资源分配格局和根系构型构建策略(单立山等, 2013 ), 这对维持荒漠生态系统的稳定性具有重要意义.目前, 许多学者围绕植物根系分支模式的竞争能力(Glimskär, 2000 )、根系构型的拓扑结构(马新明等, 2006 )、多枝柽柳(Tamarix ramosissima )和骆驼刺(Alhagi sparsifolia )幼苗生物量分配及根系分布特征(曾凡江等, 2010 )、不同树种间细根结构和形态特征(熊德成等, 2012 )、骆驼刺群落在干旱生境下细根生产与周转(冀卫萍等, 2013 )、植物根系构型的生态适应性(杜建会等, 2014 )以及根系功能性状的可塑性与空间异质性(Druille et al ., 2015 )等方面对植物根系构型进行了系统研究, 但是有关生境梯度下植物根系构型构建的资源配置权衡机制等研究仍显不足, 限制了对异质生境中植物表型可塑性机制的解析.本文通过研究祁连山北坡荒漠草地红砂根系分叉数和分支角度的分布格局, 试图解析: (1)不同坡向红砂根系分叉数和分支角度的变化规律; (2)造成红砂根系分叉数和分支角度异速生长关系的原因, 旨在从植物根系功能性状建成的角度阐明红砂种群的环境适应性规律, 为进一步了解荒漠植物的表型可塑性机制提供理论依据. ...
河西走廊中部两种荒漠植物根系构型特征.
1
2013
... 植物外部形态具有高度的环境可塑性特征, 在长期生长发育的过程中, 生境差异性使其主要构件采取了不同的生态对策, 能够协调各功能性状间的资源分配模式以规避环境风险(史元春等, 2015 ).植物根系构型构建与植物自身根系碳投入和土壤水分养分的收益权衡相契合, 是植物提高自身资源获取能力的一个重要策略(杨小林等, 2008 ).研究区位于祁连山与河西走廊过渡带的荒漠草原区, 山地南坡的植物生存环境比较严酷, 红砂的生物量水平低(表1 , 表2 ), 为了提高自身的生存和繁衍能力, 红砂将更多的同化产物分配给地下根系, 着力构建强大的物质吸收和存储系统, 从而限制了地上生物量的分配比例(表1 ), 形成了相对较小的、有助于降低植株总体蒸腾耗水量的枝叶性状体系(表3 ).南坡植被稀疏(表2 )、大气温度较高, 结构松散的沙土毛细管吸收作用力弱.在土壤含水量偏低(表2 )和无地下水补给的条件下, 荒漠植物往往需要较大集水面积才能满足植株最低水平的生理耗水需求, 红砂选择了增加根系分支角度、减小分叉数的根系构型构建模式, 原因如下: 1)红砂属于浅根系植物, 绝大部分根系与地面平行伸展, 形成相对较大的根系分支角度(图1 ), 更有利于侧根趋向于水平分布、扩大根系的有效觅养空间, 同样单位(长度)的根系拥有的吸水土壤体积可能较大, 从而具有比同一生境中相邻植物更强的水资源竞争能力, 可以获得更多的生存机会(蒋礼学和李彦, 2008 ).这符合相关学者“分支角度越大, 根系越趋向于水平分布, 向根更广的范围扩展以寻求资源”的研究结论(黄晶晶, 2013 ).2)植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 保证自身的生存和生长的策略(杜建会等, 2014 ).生长在南坡的红砂减小其根系分叉数的根系构型(图1 ), 在一定程度上降低了根系内部的交叠重复, 可以避免细根间资源空间的竞争, 从而提高根系对营养物质的吸收效率来加快对土壤资源的利用, 以最佳的收益/代价关系适应贫瘠的资源环境, 与相关学者得出一致的结论, 即“当植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 减少根系分叉数的策略以降低根系内部的交叠重复, 避免细根营养物质的竞争” (单立山等, 2012 ; 杜建会等, 2014 ).因此, 分布于南坡的红砂根系分叉数和分支角度间呈极显著负相关关系(p < 0.01, 图2 ), 实现了光合产物在构件水平的优化配置. ...
河西走廊中部两种荒漠植物根系构型特征.
1
2013
... 植物外部形态具有高度的环境可塑性特征, 在长期生长发育的过程中, 生境差异性使其主要构件采取了不同的生态对策, 能够协调各功能性状间的资源分配模式以规避环境风险(史元春等, 2015 ).植物根系构型构建与植物自身根系碳投入和土壤水分养分的收益权衡相契合, 是植物提高自身资源获取能力的一个重要策略(杨小林等, 2008 ).研究区位于祁连山与河西走廊过渡带的荒漠草原区, 山地南坡的植物生存环境比较严酷, 红砂的生物量水平低(表1 , 表2 ), 为了提高自身的生存和繁衍能力, 红砂将更多的同化产物分配给地下根系, 着力构建强大的物质吸收和存储系统, 从而限制了地上生物量的分配比例(表1 ), 形成了相对较小的、有助于降低植株总体蒸腾耗水量的枝叶性状体系(表3 ).南坡植被稀疏(表2 )、大气温度较高, 结构松散的沙土毛细管吸收作用力弱.在土壤含水量偏低(表2 )和无地下水补给的条件下, 荒漠植物往往需要较大集水面积才能满足植株最低水平的生理耗水需求, 红砂选择了增加根系分支角度、减小分叉数的根系构型构建模式, 原因如下: 1)红砂属于浅根系植物, 绝大部分根系与地面平行伸展, 形成相对较大的根系分支角度(图1 ), 更有利于侧根趋向于水平分布、扩大根系的有效觅养空间, 同样单位(长度)的根系拥有的吸水土壤体积可能较大, 从而具有比同一生境中相邻植物更强的水资源竞争能力, 可以获得更多的生存机会(蒋礼学和李彦, 2008 ).这符合相关学者“分支角度越大, 根系越趋向于水平分布, 向根更广的范围扩展以寻求资源”的研究结论(黄晶晶, 2013 ).2)植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 保证自身的生存和生长的策略(杜建会等, 2014 ).生长在南坡的红砂减小其根系分叉数的根系构型(图1 ), 在一定程度上降低了根系内部的交叠重复, 可以避免细根间资源空间的竞争, 从而提高根系对营养物质的吸收效率来加快对土壤资源的利用, 以最佳的收益/代价关系适应贫瘠的资源环境, 与相关学者得出一致的结论, 即“当植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 减少根系分叉数的策略以降低根系内部的交叠重复, 避免细根营养物质的竞争” (单立山等, 2012 ; 杜建会等, 2014 ).因此, 分布于南坡的红砂根系分叉数和分支角度间呈极显著负相关关系(p < 0.01, 图2 ), 实现了光合产物在构件水平的优化配置. ...
兰州北山刺槐枝叶性状的坡向差异性.
2
2015
... 植物根系作为植被与土壤之间物质、能量交换的主要通道(黄晶晶, 2013 ), 在植物水分和养分吸收、运移过程中有着不可替代的作用(Kong et al ., 2014 ), 影响着地上部分以及整个植株的生长和发育, 其中根系的分支状况和构型对营养物质的吸收起着关键作用(Dannowski & Block, 2005 ).根系构型是植物与环境异质性资源相互适应的结果(Malamy, 2005 ), 其对环境的适应是通过根系分支结构、空间分布格局等构型特征的改变表现出来的(Tracy et al ., 2015 ).为了应对地理环境和资源供给水平造成的胁迫和干扰, 植物根系会根据所处生境的资源条件适时调整和转化根系分叉数和分支角度等构型特征, 实现植物功能性状间的资源权衡(陈莹婷和许振柱, 2014 ).根系分叉数影响植物根系对资源物质的觅食能力和植株的固定作用(孙浩燕等, 2014 ), 根系分支角度与根系的分布范围和拓展能力紧密相关(黄晶晶, 2013 ), 二者在很大程度上决定了根系在土壤中的位置和对土体的觅养效率(Malamy, 2005 ), 反映了不同生境中植物根系在物质吸收、利用和循环中的作用.地形作为生境条件的综合指示, 控制水热因子的重新分配, 修饰植被空间分布格局, 此自然法则具有普适性(常学礼等, 2015 ).坡向是地形重要的影响因素之一, 影响了太阳辐射和降水的空间再分配, 往往导致局部小气候的产生(Cantón et al ., 2004 ), 从而改变光照、温度及土壤水分的分布, 导致群落环境和植物的生存条件产生差异(侯兆疆等, 2014 ), 进而改变植物碳资源分配格局和根系构型构建策略(Oppelt et al ., 2001 ).为了提高植株生存适合度和竞争能力, 植物根系通过分支角度和分叉数等性状的响应, 促使根系构型模式进行适应性调整, 构建与环境梯度相匹配的根系营养吸收和传输系统.因此, 从不同坡向资源条件变化的角度研究植物根系构型的空间分异格局和资源配置模式, 对于理解植物在异质生境中的环境适应机制具有重要的理论意义. ...
... 植物构件的资源分配是与栖息环境相对应的种群特征的反映方式, 它反映了种群特定的生态适应策略(张茜等, 2014 ).根系构型会对周围环境产生适应性的改变, 表现出不一样的适应特性(Oppelt et al ., 2005 ).相对于南坡, 山地北坡草地群落的盖度、密度均较大, 红砂在群落中没有高度优势(表1 , 表2 ), 相对拥挤的群落环境使植物面临着严重的邻体干扰, 红砂将较多的生物量分配给地上枝叶以利于植株获得高度优势, 增强枝叶的捕光能力, 无形中限制了分配给根系的生物量比例(表1 ).在根冠比较低的情况下, 红砂需要将有限的资源用于构建根系的吸收面积, 增强根系的“觅食”能力(董鸣, 1996 ), 以最大限度保证植株在生境竞争中占据优势, 红砂选择了增加根系分叉数、减小分支角度的根系构型.原因如下: 1)在土壤含水量相对较高的北坡(表2 ), 红砂以根系分叉数增加速度大于分支角度减小速度的资源配置模式(图2 ), 通过增大根系分叉数(图1 ), 有效增加了根系分支能力和单位土体中根系的拥挤度, 构建了密集的根系网络系统, 降低了根系传输系统的碳需求量, 实现了单位根系土壤拓展体积较小状态下的植株生存的水分供给需求, 这与单立山等(2013)对红砂和白刺(Nitraria tangutorum )、杜建会等(2014)对塔克拉玛干柽柳(Tamarix taklamakanensis )的研究结果相一致.2)植物根系在深层土壤的分布有利于其吸收利用深层土壤水分, 促进植株的生长发育(席海洋等, 2011 ).北坡群落密度高(表2 )、植物之间拥挤度大, 种内和种间容易造成根系重叠竞争, 空间上无法满足大范围根系的构型塑造.红砂采取缩小根系水平范围, 将根系分支角度减小, 扩大根系在深层土壤中的垂直分布(表1 ), 避免与群落中其他植株根系的剧烈竞争.同时红砂植物通过增大比根长提高对养分和水分的利用效率来加快对土壤资源的利用(表1 ), 提高自身的竞争力以维持生存.因此, 红砂根系分叉数和分支角度间呈极显著负相关关系(p < 0.01, 图2 ), 即以资源权衡分配机制进化响应诸多竞争压力, 维持植物生长的水分条件, 从而有利于生物多样性的群落结构, 这符合相关学者“分叉数较大的植物能够迅速占据大量土壤空间快速生长, 适宜生长在土壤资源丰富的环境中”的结论(周艳松和王立群, 2011 ; 孙浩燕等, 2014 ). ...
兰州北山刺槐枝叶性状的坡向差异性.
2
2015
... 植物根系作为植被与土壤之间物质、能量交换的主要通道(黄晶晶, 2013 ), 在植物水分和养分吸收、运移过程中有着不可替代的作用(Kong et al ., 2014 ), 影响着地上部分以及整个植株的生长和发育, 其中根系的分支状况和构型对营养物质的吸收起着关键作用(Dannowski & Block, 2005 ).根系构型是植物与环境异质性资源相互适应的结果(Malamy, 2005 ), 其对环境的适应是通过根系分支结构、空间分布格局等构型特征的改变表现出来的(Tracy et al ., 2015 ).为了应对地理环境和资源供给水平造成的胁迫和干扰, 植物根系会根据所处生境的资源条件适时调整和转化根系分叉数和分支角度等构型特征, 实现植物功能性状间的资源权衡(陈莹婷和许振柱, 2014 ).根系分叉数影响植物根系对资源物质的觅食能力和植株的固定作用(孙浩燕等, 2014 ), 根系分支角度与根系的分布范围和拓展能力紧密相关(黄晶晶, 2013 ), 二者在很大程度上决定了根系在土壤中的位置和对土体的觅养效率(Malamy, 2005 ), 反映了不同生境中植物根系在物质吸收、利用和循环中的作用.地形作为生境条件的综合指示, 控制水热因子的重新分配, 修饰植被空间分布格局, 此自然法则具有普适性(常学礼等, 2015 ).坡向是地形重要的影响因素之一, 影响了太阳辐射和降水的空间再分配, 往往导致局部小气候的产生(Cantón et al ., 2004 ), 从而改变光照、温度及土壤水分的分布, 导致群落环境和植物的生存条件产生差异(侯兆疆等, 2014 ), 进而改变植物碳资源分配格局和根系构型构建策略(Oppelt et al ., 2001 ).为了提高植株生存适合度和竞争能力, 植物根系通过分支角度和分叉数等性状的响应, 促使根系构型模式进行适应性调整, 构建与环境梯度相匹配的根系营养吸收和传输系统.因此, 从不同坡向资源条件变化的角度研究植物根系构型的空间分异格局和资源配置模式, 对于理解植物在异质生境中的环境适应机制具有重要的理论意义. ...
... 植物构件的资源分配是与栖息环境相对应的种群特征的反映方式, 它反映了种群特定的生态适应策略(张茜等, 2014 ).根系构型会对周围环境产生适应性的改变, 表现出不一样的适应特性(Oppelt et al ., 2005 ).相对于南坡, 山地北坡草地群落的盖度、密度均较大, 红砂在群落中没有高度优势(表1 , 表2 ), 相对拥挤的群落环境使植物面临着严重的邻体干扰, 红砂将较多的生物量分配给地上枝叶以利于植株获得高度优势, 增强枝叶的捕光能力, 无形中限制了分配给根系的生物量比例(表1 ).在根冠比较低的情况下, 红砂需要将有限的资源用于构建根系的吸收面积, 增强根系的“觅食”能力(董鸣, 1996 ), 以最大限度保证植株在生境竞争中占据优势, 红砂选择了增加根系分叉数、减小分支角度的根系构型.原因如下: 1)在土壤含水量相对较高的北坡(表2 ), 红砂以根系分叉数增加速度大于分支角度减小速度的资源配置模式(图2 ), 通过增大根系分叉数(图1 ), 有效增加了根系分支能力和单位土体中根系的拥挤度, 构建了密集的根系网络系统, 降低了根系传输系统的碳需求量, 实现了单位根系土壤拓展体积较小状态下的植株生存的水分供给需求, 这与单立山等(2013)对红砂和白刺(Nitraria tangutorum )、杜建会等(2014)对塔克拉玛干柽柳(Tamarix taklamakanensis )的研究结果相一致.2)植物根系在深层土壤的分布有利于其吸收利用深层土壤水分, 促进植株的生长发育(席海洋等, 2011 ).北坡群落密度高(表2 )、植物之间拥挤度大, 种内和种间容易造成根系重叠竞争, 空间上无法满足大范围根系的构型塑造.红砂采取缩小根系水平范围, 将根系分支角度减小, 扩大根系在深层土壤中的垂直分布(表1 ), 避免与群落中其他植株根系的剧烈竞争.同时红砂植物通过增大比根长提高对养分和水分的利用效率来加快对土壤资源的利用(表1 ), 提高自身的竞争力以维持生存.因此, 红砂根系分叉数和分支角度间呈极显著负相关关系(p < 0.01, 图2 ), 即以资源权衡分配机制进化响应诸多竞争压力, 维持植物生长的水分条件, 从而有利于生物多样性的群落结构, 这符合相关学者“分叉数较大的植物能够迅速占据大量土壤空间快速生长, 适宜生长在土壤资源丰富的环境中”的结论(周艳松和王立群, 2011 ; 孙浩燕等, 2014 ). ...
浅层施肥对水稻苗期根系生长及分布的影响.
0
2014
浅层施肥对水稻苗期根系生长及分布的影响.
0
2014
1
2010
... 植物根系作为植被与土壤之间物质、能量交换的主要通道(黄晶晶, 2013 ), 在植物水分和养分吸收、运移过程中有着不可替代的作用(Kong et al ., 2014 ), 影响着地上部分以及整个植株的生长和发育, 其中根系的分支状况和构型对营养物质的吸收起着关键作用(Dannowski & Block, 2005 ).根系构型是植物与环境异质性资源相互适应的结果(Malamy, 2005 ), 其对环境的适应是通过根系分支结构、空间分布格局等构型特征的改变表现出来的(Tracy et al ., 2015 ).为了应对地理环境和资源供给水平造成的胁迫和干扰, 植物根系会根据所处生境的资源条件适时调整和转化根系分叉数和分支角度等构型特征, 实现植物功能性状间的资源权衡(陈莹婷和许振柱, 2014 ).根系分叉数影响植物根系对资源物质的觅食能力和植株的固定作用(孙浩燕等, 2014 ), 根系分支角度与根系的分布范围和拓展能力紧密相关(黄晶晶, 2013 ), 二者在很大程度上决定了根系在土壤中的位置和对土体的觅养效率(Malamy, 2005 ), 反映了不同生境中植物根系在物质吸收、利用和循环中的作用.地形作为生境条件的综合指示, 控制水热因子的重新分配, 修饰植被空间分布格局, 此自然法则具有普适性(常学礼等, 2015 ).坡向是地形重要的影响因素之一, 影响了太阳辐射和降水的空间再分配, 往往导致局部小气候的产生(Cantón et al ., 2004 ), 从而改变光照、温度及土壤水分的分布, 导致群落环境和植物的生存条件产生差异(侯兆疆等, 2014 ), 进而改变植物碳资源分配格局和根系构型构建策略(Oppelt et al ., 2001 ).为了提高植株生存适合度和竞争能力, 植物根系通过分支角度和分叉数等性状的响应, 促使根系构型模式进行适应性调整, 构建与环境梯度相匹配的根系营养吸收和传输系统.因此, 从不同坡向资源条件变化的角度研究植物根系构型的空间分异格局和资源配置模式, 对于理解植物在异质生境中的环境适应机制具有重要的理论意义. ...
1
2010
... 植物根系作为植被与土壤之间物质、能量交换的主要通道(黄晶晶, 2013 ), 在植物水分和养分吸收、运移过程中有着不可替代的作用(Kong et al ., 2014 ), 影响着地上部分以及整个植株的生长和发育, 其中根系的分支状况和构型对营养物质的吸收起着关键作用(Dannowski & Block, 2005 ).根系构型是植物与环境异质性资源相互适应的结果(Malamy, 2005 ), 其对环境的适应是通过根系分支结构、空间分布格局等构型特征的改变表现出来的(Tracy et al ., 2015 ).为了应对地理环境和资源供给水平造成的胁迫和干扰, 植物根系会根据所处生境的资源条件适时调整和转化根系分叉数和分支角度等构型特征, 实现植物功能性状间的资源权衡(陈莹婷和许振柱, 2014 ).根系分叉数影响植物根系对资源物质的觅食能力和植株的固定作用(孙浩燕等, 2014 ), 根系分支角度与根系的分布范围和拓展能力紧密相关(黄晶晶, 2013 ), 二者在很大程度上决定了根系在土壤中的位置和对土体的觅养效率(Malamy, 2005 ), 反映了不同生境中植物根系在物质吸收、利用和循环中的作用.地形作为生境条件的综合指示, 控制水热因子的重新分配, 修饰植被空间分布格局, 此自然法则具有普适性(常学礼等, 2015 ).坡向是地形重要的影响因素之一, 影响了太阳辐射和降水的空间再分配, 往往导致局部小气候的产生(Cantón et al ., 2004 ), 从而改变光照、温度及土壤水分的分布, 导致群落环境和植物的生存条件产生差异(侯兆疆等, 2014 ), 进而改变植物碳资源分配格局和根系构型构建策略(Oppelt et al ., 2001 ).为了提高植株生存适合度和竞争能力, 植物根系通过分支角度和分叉数等性状的响应, 促使根系构型模式进行适应性调整, 构建与环境梯度相匹配的根系营养吸收和传输系统.因此, 从不同坡向资源条件变化的角度研究植物根系构型的空间分异格局和资源配置模式, 对于理解植物在异质生境中的环境适应机制具有重要的理论意义. ...
Using X-ray Computed Tomography to explore the role of abscisic acid in moderating the impact of soil compaction on root system architecture.
1
2015
... 将坡向划分为北坡(315°-360°和0°-45°)、东坡(45°-135°)、南坡(135°-225°)和西坡(225°-315°) 4个坡向组.根据游标卡尺、卷尺和量角器等工具测定获得的根长、分叉数和分支角度等, 计算出各坡向红砂根系的比根长.比根长表示单位生物量的细根长度, 它们反映投入到根系的生物量中用于吸收养分的效率, 其值越大, 表明投入相同的成本有越高的养分利用效率(Bauhus et al ., 2000 ).计算公式为: 比根长(cm·g-1 ) =根长(cm)/生物量(g).根系分叉数和分支角度数据分析主要采用标准化主轴估计(SMA)的方法(Warton et al ., 2006 ), 由软件(S)MATR Version 2.0 (Falster et al ., 2006 )计算完成.对每个样方红砂根系分叉数和分支角度进行对数(以10为底)转换后再进行分析, 对数转换使之符合正态分布.采用y = axb , 线性转换成log(y ) = log(a ) + b log(x ), 式中x 和y 表示两个特征参数, b 为斜率, 即异速生长参数或相对生长的指数, 当|b | = 1时, 表示两者是等速变化关系; 当|b| 显著偏离1时, 两者间为异速变化关系(Harvey & Pagel, 1991 ).实验所用数据采用SPSS 16.0软件进行处理分析, 用SigmaPlot 10.0和Excel软件绘图. ...
Bivariate line-fitting methods for allometry.
1
2006
... 植物构件的资源分配是与栖息环境相对应的种群特征的反映方式, 它反映了种群特定的生态适应策略(张茜等, 2014 ).根系构型会对周围环境产生适应性的改变, 表现出不一样的适应特性(Oppelt et al ., 2005 ).相对于南坡, 山地北坡草地群落的盖度、密度均较大, 红砂在群落中没有高度优势(表1 , 表2 ), 相对拥挤的群落环境使植物面临着严重的邻体干扰, 红砂将较多的生物量分配给地上枝叶以利于植株获得高度优势, 增强枝叶的捕光能力, 无形中限制了分配给根系的生物量比例(表1 ).在根冠比较低的情况下, 红砂需要将有限的资源用于构建根系的吸收面积, 增强根系的“觅食”能力(董鸣, 1996 ), 以最大限度保证植株在生境竞争中占据优势, 红砂选择了增加根系分叉数、减小分支角度的根系构型.原因如下: 1)在土壤含水量相对较高的北坡(表2 ), 红砂以根系分叉数增加速度大于分支角度减小速度的资源配置模式(图2 ), 通过增大根系分叉数(图1 ), 有效增加了根系分支能力和单位土体中根系的拥挤度, 构建了密集的根系网络系统, 降低了根系传输系统的碳需求量, 实现了单位根系土壤拓展体积较小状态下的植株生存的水分供给需求, 这与单立山等(2013)对红砂和白刺(Nitraria tangutorum )、杜建会等(2014)对塔克拉玛干柽柳(Tamarix taklamakanensis )的研究结果相一致.2)植物根系在深层土壤的分布有利于其吸收利用深层土壤水分, 促进植株的生长发育(席海洋等, 2011 ).北坡群落密度高(表2 )、植物之间拥挤度大, 种内和种间容易造成根系重叠竞争, 空间上无法满足大范围根系的构型塑造.红砂采取缩小根系水平范围, 将根系分支角度减小, 扩大根系在深层土壤中的垂直分布(表1 ), 避免与群落中其他植株根系的剧烈竞争.同时红砂植物通过增大比根长提高对养分和水分的利用效率来加快对土壤资源的利用(表1 ), 提高自身的竞争力以维持生存.因此, 红砂根系分叉数和分支角度间呈极显著负相关关系(p < 0.01, 图2 ), 即以资源权衡分配机制进化响应诸多竞争压力, 维持植物生长的水分条件, 从而有利于生物多样性的群落结构, 这符合相关学者“分叉数较大的植物能够迅速占据大量土壤空间快速生长, 适宜生长在土壤资源丰富的环境中”的结论(周艳松和王立群, 2011 ; 孙浩燕等, 2014 ). ...
额济纳绿洲不同植被覆盖下土壤特性的时空变化
1
2011
... 红砂(Reaumuria songarica )是一种广泛分布于我国西北荒漠地区的柽柳科小灌木, 是荒漠植被的常见建群种和优势种, 具有极强的抗干旱能力.在全球变化背景下, 红砂等荒漠植物面对日益严酷的环境压力, 需要不断调整主要功能性状构建的资源策略, 适时改变植物碳资源分配格局和根系构型构建策略(单立山等, 2013 ), 这对维持荒漠生态系统的稳定性具有重要意义.目前, 许多学者围绕植物根系分支模式的竞争能力(Glimskär, 2000 )、根系构型的拓扑结构(马新明等, 2006 )、多枝柽柳(Tamarix ramosissima )和骆驼刺(Alhagi sparsifolia )幼苗生物量分配及根系分布特征(曾凡江等, 2010 )、不同树种间细根结构和形态特征(熊德成等, 2012 )、骆驼刺群落在干旱生境下细根生产与周转(冀卫萍等, 2013 )、植物根系构型的生态适应性(杜建会等, 2014 )以及根系功能性状的可塑性与空间异质性(Druille et al ., 2015 )等方面对植物根系构型进行了系统研究, 但是有关生境梯度下植物根系构型构建的资源配置权衡机制等研究仍显不足, 限制了对异质生境中植物表型可塑性机制的解析.本文通过研究祁连山北坡荒漠草地红砂根系分叉数和分支角度的分布格局, 试图解析: (1)不同坡向红砂根系分叉数和分支角度的变化规律; (2)造成红砂根系分叉数和分支角度异速生长关系的原因, 旨在从植物根系功能性状建成的角度阐明红砂种群的环境适应性规律, 为进一步了解荒漠植物的表型可塑性机制提供理论依据. ...
额济纳绿洲不同植被覆盖下土壤特性的时空变化
1
2011
... 红砂(Reaumuria songarica )是一种广泛分布于我国西北荒漠地区的柽柳科小灌木, 是荒漠植被的常见建群种和优势种, 具有极强的抗干旱能力.在全球变化背景下, 红砂等荒漠植物面对日益严酷的环境压力, 需要不断调整主要功能性状构建的资源策略, 适时改变植物碳资源分配格局和根系构型构建策略(单立山等, 2013 ), 这对维持荒漠生态系统的稳定性具有重要意义.目前, 许多学者围绕植物根系分支模式的竞争能力(Glimskär, 2000 )、根系构型的拓扑结构(马新明等, 2006 )、多枝柽柳(Tamarix ramosissima )和骆驼刺(Alhagi sparsifolia )幼苗生物量分配及根系分布特征(曾凡江等, 2010 )、不同树种间细根结构和形态特征(熊德成等, 2012 )、骆驼刺群落在干旱生境下细根生产与周转(冀卫萍等, 2013 )、植物根系构型的生态适应性(杜建会等, 2014 )以及根系功能性状的可塑性与空间异质性(Druille et al ., 2015 )等方面对植物根系构型进行了系统研究, 但是有关生境梯度下植物根系构型构建的资源配置权衡机制等研究仍显不足, 限制了对异质生境中植物表型可塑性机制的解析.本文通过研究祁连山北坡荒漠草地红砂根系分叉数和分支角度的分布格局, 试图解析: (1)不同坡向红砂根系分叉数和分支角度的变化规律; (2)造成红砂根系分叉数和分支角度异速生长关系的原因, 旨在从植物根系功能性状建成的角度阐明红砂种群的环境适应性规律, 为进一步了解荒漠植物的表型可塑性机制提供理论依据. ...
亚热带6种树种细根序级结构和形态特征.
1
2012
... 植物外部形态具有高度的环境可塑性特征, 在长期生长发育的过程中, 生境差异性使其主要构件采取了不同的生态对策, 能够协调各功能性状间的资源分配模式以规避环境风险(史元春等, 2015 ).植物根系构型构建与植物自身根系碳投入和土壤水分养分的收益权衡相契合, 是植物提高自身资源获取能力的一个重要策略(杨小林等, 2008 ).研究区位于祁连山与河西走廊过渡带的荒漠草原区, 山地南坡的植物生存环境比较严酷, 红砂的生物量水平低(表1 , 表2 ), 为了提高自身的生存和繁衍能力, 红砂将更多的同化产物分配给地下根系, 着力构建强大的物质吸收和存储系统, 从而限制了地上生物量的分配比例(表1 ), 形成了相对较小的、有助于降低植株总体蒸腾耗水量的枝叶性状体系(表3 ).南坡植被稀疏(表2 )、大气温度较高, 结构松散的沙土毛细管吸收作用力弱.在土壤含水量偏低(表2 )和无地下水补给的条件下, 荒漠植物往往需要较大集水面积才能满足植株最低水平的生理耗水需求, 红砂选择了增加根系分支角度、减小分叉数的根系构型构建模式, 原因如下: 1)红砂属于浅根系植物, 绝大部分根系与地面平行伸展, 形成相对较大的根系分支角度(图1 ), 更有利于侧根趋向于水平分布、扩大根系的有效觅养空间, 同样单位(长度)的根系拥有的吸水土壤体积可能较大, 从而具有比同一生境中相邻植物更强的水资源竞争能力, 可以获得更多的生存机会(蒋礼学和李彦, 2008 ).这符合相关学者“分支角度越大, 根系越趋向于水平分布, 向根更广的范围扩展以寻求资源”的研究结论(黄晶晶, 2013 ).2)植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 保证自身的生存和生长的策略(杜建会等, 2014 ).生长在南坡的红砂减小其根系分叉数的根系构型(图1 ), 在一定程度上降低了根系内部的交叠重复, 可以避免细根间资源空间的竞争, 从而提高根系对营养物质的吸收效率来加快对土壤资源的利用, 以最佳的收益/代价关系适应贫瘠的资源环境, 与相关学者得出一致的结论, 即“当植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 减少根系分叉数的策略以降低根系内部的交叠重复, 避免细根营养物质的竞争” (单立山等, 2012 ; 杜建会等, 2014 ).因此, 分布于南坡的红砂根系分叉数和分支角度间呈极显著负相关关系(p < 0.01, 图2 ), 实现了光合产物在构件水平的优化配置. ...
亚热带6种树种细根序级结构和形态特征.
1
2012
... 植物外部形态具有高度的环境可塑性特征, 在长期生长发育的过程中, 生境差异性使其主要构件采取了不同的生态对策, 能够协调各功能性状间的资源分配模式以规避环境风险(史元春等, 2015 ).植物根系构型构建与植物自身根系碳投入和土壤水分养分的收益权衡相契合, 是植物提高自身资源获取能力的一个重要策略(杨小林等, 2008 ).研究区位于祁连山与河西走廊过渡带的荒漠草原区, 山地南坡的植物生存环境比较严酷, 红砂的生物量水平低(表1 , 表2 ), 为了提高自身的生存和繁衍能力, 红砂将更多的同化产物分配给地下根系, 着力构建强大的物质吸收和存储系统, 从而限制了地上生物量的分配比例(表1 ), 形成了相对较小的、有助于降低植株总体蒸腾耗水量的枝叶性状体系(表3 ).南坡植被稀疏(表2 )、大气温度较高, 结构松散的沙土毛细管吸收作用力弱.在土壤含水量偏低(表2 )和无地下水补给的条件下, 荒漠植物往往需要较大集水面积才能满足植株最低水平的生理耗水需求, 红砂选择了增加根系分支角度、减小分叉数的根系构型构建模式, 原因如下: 1)红砂属于浅根系植物, 绝大部分根系与地面平行伸展, 形成相对较大的根系分支角度(图1 ), 更有利于侧根趋向于水平分布、扩大根系的有效觅养空间, 同样单位(长度)的根系拥有的吸水土壤体积可能较大, 从而具有比同一生境中相邻植物更强的水资源竞争能力, 可以获得更多的生存机会(蒋礼学和李彦, 2008 ).这符合相关学者“分支角度越大, 根系越趋向于水平分布, 向根更广的范围扩展以寻求资源”的研究结论(黄晶晶, 2013 ).2)植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 保证自身的生存和生长的策略(杜建会等, 2014 ).生长在南坡的红砂减小其根系分叉数的根系构型(图1 ), 在一定程度上降低了根系内部的交叠重复, 可以避免细根间资源空间的竞争, 从而提高根系对营养物质的吸收效率来加快对土壤资源的利用, 以最佳的收益/代价关系适应贫瘠的资源环境, 与相关学者得出一致的结论, 即“当植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 减少根系分叉数的策略以降低根系内部的交叠重复, 避免细根营养物质的竞争” (单立山等, 2012 ; 杜建会等, 2014 ).因此, 分布于南坡的红砂根系分叉数和分支角度间呈极显著负相关关系(p < 0.01, 图2 ), 实现了光合产物在构件水平的优化配置. ...
塔克拉玛干沙漠腹地3种植物根系构型及其生境适应策略.
1
2008
... 红砂(Reaumuria songarica )是一种广泛分布于我国西北荒漠地区的柽柳科小灌木, 是荒漠植被的常见建群种和优势种, 具有极强的抗干旱能力.在全球变化背景下, 红砂等荒漠植物面对日益严酷的环境压力, 需要不断调整主要功能性状构建的资源策略, 适时改变植物碳资源分配格局和根系构型构建策略(单立山等, 2013 ), 这对维持荒漠生态系统的稳定性具有重要意义.目前, 许多学者围绕植物根系分支模式的竞争能力(Glimskär, 2000 )、根系构型的拓扑结构(马新明等, 2006 )、多枝柽柳(Tamarix ramosissima )和骆驼刺(Alhagi sparsifolia )幼苗生物量分配及根系分布特征(曾凡江等, 2010 )、不同树种间细根结构和形态特征(熊德成等, 2012 )、骆驼刺群落在干旱生境下细根生产与周转(冀卫萍等, 2013 )、植物根系构型的生态适应性(杜建会等, 2014 )以及根系功能性状的可塑性与空间异质性(Druille et al ., 2015 )等方面对植物根系构型进行了系统研究, 但是有关生境梯度下植物根系构型构建的资源配置权衡机制等研究仍显不足, 限制了对异质生境中植物表型可塑性机制的解析.本文通过研究祁连山北坡荒漠草地红砂根系分叉数和分支角度的分布格局, 试图解析: (1)不同坡向红砂根系分叉数和分支角度的变化规律; (2)造成红砂根系分叉数和分支角度异速生长关系的原因, 旨在从植物根系功能性状建成的角度阐明红砂种群的环境适应性规律, 为进一步了解荒漠植物的表型可塑性机制提供理论依据. ...
塔克拉玛干沙漠腹地3种植物根系构型及其生境适应策略.
1
2008
... 红砂(Reaumuria songarica )是一种广泛分布于我国西北荒漠地区的柽柳科小灌木, 是荒漠植被的常见建群种和优势种, 具有极强的抗干旱能力.在全球变化背景下, 红砂等荒漠植物面对日益严酷的环境压力, 需要不断调整主要功能性状构建的资源策略, 适时改变植物碳资源分配格局和根系构型构建策略(单立山等, 2013 ), 这对维持荒漠生态系统的稳定性具有重要意义.目前, 许多学者围绕植物根系分支模式的竞争能力(Glimskär, 2000 )、根系构型的拓扑结构(马新明等, 2006 )、多枝柽柳(Tamarix ramosissima )和骆驼刺(Alhagi sparsifolia )幼苗生物量分配及根系分布特征(曾凡江等, 2010 )、不同树种间细根结构和形态特征(熊德成等, 2012 )、骆驼刺群落在干旱生境下细根生产与周转(冀卫萍等, 2013 )、植物根系构型的生态适应性(杜建会等, 2014 )以及根系功能性状的可塑性与空间异质性(Druille et al ., 2015 )等方面对植物根系构型进行了系统研究, 但是有关生境梯度下植物根系构型构建的资源配置权衡机制等研究仍显不足, 限制了对异质生境中植物表型可塑性机制的解析.本文通过研究祁连山北坡荒漠草地红砂根系分叉数和分支角度的分布格局, 试图解析: (1)不同坡向红砂根系分叉数和分支角度的变化规律; (2)造成红砂根系分叉数和分支角度异速生长关系的原因, 旨在从植物根系功能性状建成的角度阐明红砂种群的环境适应性规律, 为进一步了解荒漠植物的表型可塑性机制提供理论依据. ...
多枝柽柳和疏叶骆驼刺幼苗生物量分配及根系分布特征.
1
2010
... 植物构件的资源分配是与栖息环境相对应的种群特征的反映方式, 它反映了种群特定的生态适应策略(张茜等, 2014 ).根系构型会对周围环境产生适应性的改变, 表现出不一样的适应特性(Oppelt et al ., 2005 ).相对于南坡, 山地北坡草地群落的盖度、密度均较大, 红砂在群落中没有高度优势(表1 , 表2 ), 相对拥挤的群落环境使植物面临着严重的邻体干扰, 红砂将较多的生物量分配给地上枝叶以利于植株获得高度优势, 增强枝叶的捕光能力, 无形中限制了分配给根系的生物量比例(表1 ).在根冠比较低的情况下, 红砂需要将有限的资源用于构建根系的吸收面积, 增强根系的“觅食”能力(董鸣, 1996 ), 以最大限度保证植株在生境竞争中占据优势, 红砂选择了增加根系分叉数、减小分支角度的根系构型.原因如下: 1)在土壤含水量相对较高的北坡(表2 ), 红砂以根系分叉数增加速度大于分支角度减小速度的资源配置模式(图2 ), 通过增大根系分叉数(图1 ), 有效增加了根系分支能力和单位土体中根系的拥挤度, 构建了密集的根系网络系统, 降低了根系传输系统的碳需求量, 实现了单位根系土壤拓展体积较小状态下的植株生存的水分供给需求, 这与单立山等(2013)对红砂和白刺(Nitraria tangutorum )、杜建会等(2014)对塔克拉玛干柽柳(Tamarix taklamakanensis )的研究结果相一致.2)植物根系在深层土壤的分布有利于其吸收利用深层土壤水分, 促进植株的生长发育(席海洋等, 2011 ).北坡群落密度高(表2 )、植物之间拥挤度大, 种内和种间容易造成根系重叠竞争, 空间上无法满足大范围根系的构型塑造.红砂采取缩小根系水平范围, 将根系分支角度减小, 扩大根系在深层土壤中的垂直分布(表1 ), 避免与群落中其他植株根系的剧烈竞争.同时红砂植物通过增大比根长提高对养分和水分的利用效率来加快对土壤资源的利用(表1 ), 提高自身的竞争力以维持生存.因此, 红砂根系分叉数和分支角度间呈极显著负相关关系(p < 0.01, 图2 ), 即以资源权衡分配机制进化响应诸多竞争压力, 维持植物生长的水分条件, 从而有利于生物多样性的群落结构, 这符合相关学者“分叉数较大的植物能够迅速占据大量土壤空间快速生长, 适宜生长在土壤资源丰富的环境中”的结论(周艳松和王立群, 2011 ; 孙浩燕等, 2014 ). ...
多枝柽柳和疏叶骆驼刺幼苗生物量分配及根系分布特征.
1
2010
... 植物构件的资源分配是与栖息环境相对应的种群特征的反映方式, 它反映了种群特定的生态适应策略(张茜等, 2014 ).根系构型会对周围环境产生适应性的改变, 表现出不一样的适应特性(Oppelt et al ., 2005 ).相对于南坡, 山地北坡草地群落的盖度、密度均较大, 红砂在群落中没有高度优势(表1 , 表2 ), 相对拥挤的群落环境使植物面临着严重的邻体干扰, 红砂将较多的生物量分配给地上枝叶以利于植株获得高度优势, 增强枝叶的捕光能力, 无形中限制了分配给根系的生物量比例(表1 ).在根冠比较低的情况下, 红砂需要将有限的资源用于构建根系的吸收面积, 增强根系的“觅食”能力(董鸣, 1996 ), 以最大限度保证植株在生境竞争中占据优势, 红砂选择了增加根系分叉数、减小分支角度的根系构型.原因如下: 1)在土壤含水量相对较高的北坡(表2 ), 红砂以根系分叉数增加速度大于分支角度减小速度的资源配置模式(图2 ), 通过增大根系分叉数(图1 ), 有效增加了根系分支能力和单位土体中根系的拥挤度, 构建了密集的根系网络系统, 降低了根系传输系统的碳需求量, 实现了单位根系土壤拓展体积较小状态下的植株生存的水分供给需求, 这与单立山等(2013)对红砂和白刺(Nitraria tangutorum )、杜建会等(2014)对塔克拉玛干柽柳(Tamarix taklamakanensis )的研究结果相一致.2)植物根系在深层土壤的分布有利于其吸收利用深层土壤水分, 促进植株的生长发育(席海洋等, 2011 ).北坡群落密度高(表2 )、植物之间拥挤度大, 种内和种间容易造成根系重叠竞争, 空间上无法满足大范围根系的构型塑造.红砂采取缩小根系水平范围, 将根系分支角度减小, 扩大根系在深层土壤中的垂直分布(表1 ), 避免与群落中其他植株根系的剧烈竞争.同时红砂植物通过增大比根长提高对养分和水分的利用效率来加快对土壤资源的利用(表1 ), 提高自身的竞争力以维持生存.因此, 红砂根系分叉数和分支角度间呈极显著负相关关系(p < 0.01, 图2 ), 即以资源权衡分配机制进化响应诸多竞争压力, 维持植物生长的水分条件, 从而有利于生物多样性的群落结构, 这符合相关学者“分叉数较大的植物能够迅速占据大量土壤空间快速生长, 适宜生长在土壤资源丰富的环境中”的结论(周艳松和王立群, 2011 ; 孙浩燕等, 2014 ). ...
高寒退化草地狼毒种群不同海拔花大小-数量的权衡关系.
1
2014
... 植物在长期的进化过程中, 通过生理和发育调节形成与环境相适应的表型特征, 进而具备了提高植物水分、养分等资源利用效率的功能(李西良等, 2014 ).在异质性生境中, 植物根系面对水分和养分吸收、运移和存储的环境压迫, 其分叉数和分支角度可能会发生适应性变化, 并通过物质能量吸收、固着格局的调整反作用于植被生态系统的稳定和种群更新机制, 体现了植物根系进行空间探索以适应环境胁迫的能力(Zhao et al ., 2013 ).本研究发现, 随着坡向由南坡向西坡、东坡和北坡的转变, 红砂根系分支角度逐渐减小, 而根系分叉数呈相反的变化趋势(图1 ), 在有限的资源环境中, 红砂分配给根系分叉数和分支角度资源间存在着“此消彼长”的权衡关系.坡向梯度上红砂根系分叉数和分支角度关系的差异性是植物根系构建的资源配置模式对生境异质性的适应对策. ...
高寒退化草地狼毒种群不同海拔花大小-数量的权衡关系.
1
2014
... 植物在长期的进化过程中, 通过生理和发育调节形成与环境相适应的表型特征, 进而具备了提高植物水分、养分等资源利用效率的功能(李西良等, 2014 ).在异质性生境中, 植物根系面对水分和养分吸收、运移和存储的环境压迫, 其分叉数和分支角度可能会发生适应性变化, 并通过物质能量吸收、固着格局的调整反作用于植被生态系统的稳定和种群更新机制, 体现了植物根系进行空间探索以适应环境胁迫的能力(Zhao et al ., 2013 ).本研究发现, 随着坡向由南坡向西坡、东坡和北坡的转变, 红砂根系分支角度逐渐减小, 而根系分叉数呈相反的变化趋势(图1 ), 在有限的资源环境中, 红砂分配给根系分叉数和分支角度资源间存在着“此消彼长”的权衡关系.坡向梯度上红砂根系分叉数和分支角度关系的差异性是植物根系构建的资源配置模式对生境异质性的适应对策. ...
A study on the root competitive pattern of annual pasture in mixed grassland in alpine region.
1
2013
... 植物构件的资源分配是与栖息环境相对应的种群特征的反映方式, 它反映了种群特定的生态适应策略(张茜等, 2014 ).根系构型会对周围环境产生适应性的改变, 表现出不一样的适应特性(Oppelt et al ., 2005 ).相对于南坡, 山地北坡草地群落的盖度、密度均较大, 红砂在群落中没有高度优势(表1 , 表2 ), 相对拥挤的群落环境使植物面临着严重的邻体干扰, 红砂将较多的生物量分配给地上枝叶以利于植株获得高度优势, 增强枝叶的捕光能力, 无形中限制了分配给根系的生物量比例(表1 ).在根冠比较低的情况下, 红砂需要将有限的资源用于构建根系的吸收面积, 增强根系的“觅食”能力(董鸣, 1996 ), 以最大限度保证植株在生境竞争中占据优势, 红砂选择了增加根系分叉数、减小分支角度的根系构型.原因如下: 1)在土壤含水量相对较高的北坡(表2 ), 红砂以根系分叉数增加速度大于分支角度减小速度的资源配置模式(图2 ), 通过增大根系分叉数(图1 ), 有效增加了根系分支能力和单位土体中根系的拥挤度, 构建了密集的根系网络系统, 降低了根系传输系统的碳需求量, 实现了单位根系土壤拓展体积较小状态下的植株生存的水分供给需求, 这与单立山等(2013)对红砂和白刺(Nitraria tangutorum )、杜建会等(2014)对塔克拉玛干柽柳(Tamarix taklamakanensis )的研究结果相一致.2)植物根系在深层土壤的分布有利于其吸收利用深层土壤水分, 促进植株的生长发育(席海洋等, 2011 ).北坡群落密度高(表2 )、植物之间拥挤度大, 种内和种间容易造成根系重叠竞争, 空间上无法满足大范围根系的构型塑造.红砂采取缩小根系水平范围, 将根系分支角度减小, 扩大根系在深层土壤中的垂直分布(表1 ), 避免与群落中其他植株根系的剧烈竞争.同时红砂植物通过增大比根长提高对养分和水分的利用效率来加快对土壤资源的利用(表1 ), 提高自身的竞争力以维持生存.因此, 红砂根系分叉数和分支角度间呈极显著负相关关系(p < 0.01, 图2 ), 即以资源权衡分配机制进化响应诸多竞争压力, 维持植物生长的水分条件, 从而有利于生物多样性的群落结构, 这符合相关学者“分叉数较大的植物能够迅速占据大量土壤空间快速生长, 适宜生长在土壤资源丰富的环境中”的结论(周艳松和王立群, 2011 ; 孙浩燕等, 2014 ). ...
星毛委陵菜根系构型对草原退化的生态适应.
0
2011
星毛委陵菜根系构型对草原退化的生态适应.
0
2011
 , 徐婷, 段贝贝, 韩玲, 冯威
, 徐婷, 段贝贝, 韩玲, 冯威


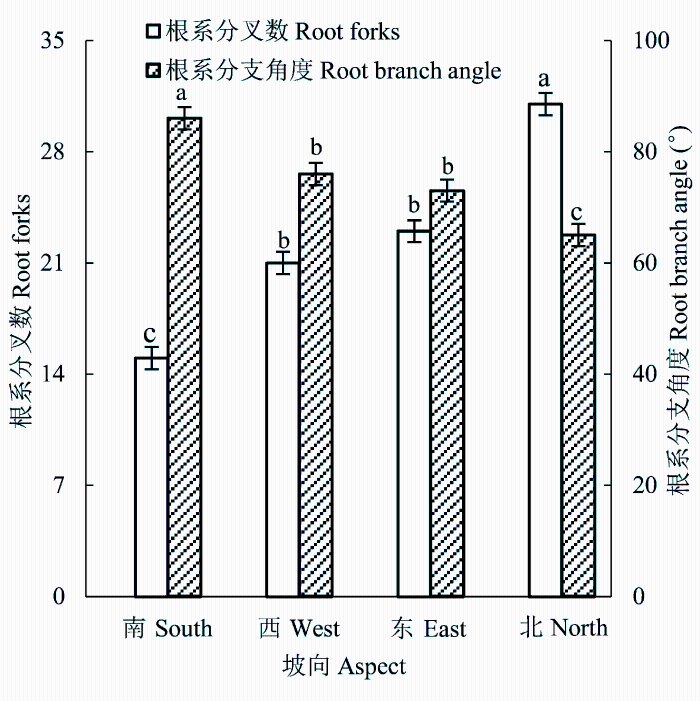
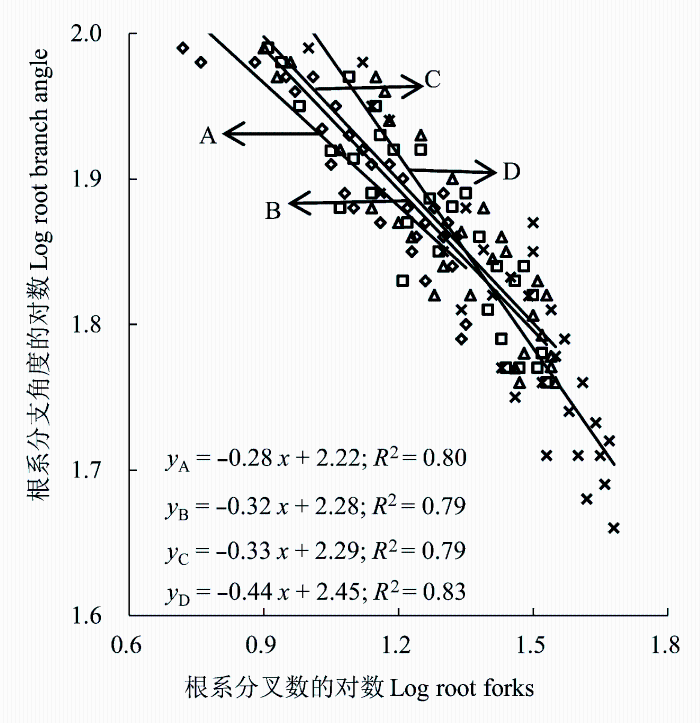
{kind=link}
{kind=link}
{kind=link}
{kind=link}