林木细根生产量约占全球净初级生产力的30%, 在森林生态系统物质和能量循环过程中具有十分重要的作用(Gill & Jackson, 2000 )。不同树种的特性存在差别, 了解不同树种间根系生产动态及其与环境因子间关系的差异, 对于评估不同森林生态系统生产力及其对全球环境变化响应的差别具有重要的作用(Coleman et al ., 2000 )。
不同演替阶段的树种生理和形态特征有所不同(Givnish, 1988 ; Messier, 1999 ; Ellis et al ., 2000 ), 细根特性也可能因所处演替阶段不同而存在差异。Paz (2003)研究了55种树种的根形态特征后发现, 演替晚期树种的根总体上要比演替早期树种的根更粗。Coll等(2008)的研究表明, 演替晚期树种的主根生物量所占比例更大。受生境条件等因素的影响, 不同演替阶段树种的细根生长速度可能也存在差异。在演替早期, 群落结构简单且立地条件相对较差, 植被更倾向于分配更多的生物量给根系(屠玉麟和杨军, 1995 ), 并通过迅速和广泛地占据土壤空间获取资源(Brassard et al ., 2009 ), 具有觅食的广布性; 而演替晚期群落的物种多样性较丰富且结构复杂, 土壤有机质和养分含量较高, 种间竞争激烈, 为了提高竞争力, 细根会迅速识别富养斑块并大量增生(Brassard et al ., 2009 ), 养分利用精细, 利用效率提高。Zangaro等(2012)对几个热带树种的研究发现, 细根生物量、细根直径随演替的进行而增加, 而根总长、比根长、根毛长度、根毛发生率、内生菌根菌侵染率等降低, 表明与演替后期阶段树种相比, 演替早期阶段的树种细根具有更强的养分获取能力, 生长速度可能更快。同时, 不同演替阶段的树种受栖息地环境的影响程度可能不同。如演替早期的热带树种生长受开阔栖息地的局限较大(Zangaro et al ., 2012 ), 而一些演替晚期的树种具有耐阴性(Kitajima, 1994 ; Walters & Reich, 1996 ; Khurana & Singh, 2006 ; Poorter & Rozendaal, 2008 ), 林冠更加稠密, 透光度更小。Du和Fang (2014)对中国东北的白桦(Betula platyphylla )-中国山杨(Populus davidiana )林和落叶松(Larix gmelinii )林的研究也曾发现, 演替早期的先锋树种白桦和中国山杨, 根呼吸对温度的敏感性要高于演替晚期的优势树种落叶松。此外, 不同演替阶段树种的物候情况在长期森林动态研究中十分重要, 且是预测未来气候变化环境下的森林变化所必须考虑的问题(Du & Fang, 2014 )。然而目前有关森林不同演替阶段的树种细根生产动态和细根物候方面差异的研究仍十分缺乏。分析研究不同演替阶段不同树种林分的细根生产量、季节动态及其所受环境因子影响的差异, 将有助于增进我们了解不同演替阶段的树种差异对细根生产可能造成的影响。
马尾松(Pinus massoniana )是我国南方重要的速生用材树种, 为亚热带群落演替早期优势树种(王伯荪和彭少麟, 1985 )。米槠(Castanopsis carlesii )是我国中亚热带地区最典型的地带性植被常绿阔叶林的代表性树种, 为演替后期顶极种(刘浩凤, 2006 )。本文通过对马尾松和米槠两个树种的人工林进行为期两年(2013年1月至2014年12月)的微根管观测, 对两种人工林细根的生产量动态及其径级和土层的分布进行研究, 并分析了两年的细根生产量动态变化与气温、降水、土壤10 cm温度和土壤10 cm含水率等环境因子间的相关性, 进一步明确了不同演替阶段树种的细根生产量变化与环境因子间的相关关系, 为预测全球气候变化对中亚热带不同演替阶段树种的根系生产力影响提供了基础数据。本研究试图解决的科学问题为: 演替早期的马尾松林分和演替晚期的米槠林分根系生产量及其动态差异、季节分布如何? 这两种不同演替阶段树种林分根系生产量动态与各环境因子间的相关关系有何差异?两种不同演替阶段树种林分的根系生产量径级和土层分配情况怎样?
1 试验地概况和研究方法
1.1 试验地概况
试验地位于福建省三明市格氏栲自然保护区(26.32° N, 117.60° E)和三明市金丝湾森林公园陈大林业采育场(26.32° N, 117.60° E)内, 其中研究林分马尾松人工林分布于三明市金丝湾森林公园陈大林业采育场内, 米槠人工林分布于三明格氏栲自然保护区内。试验地所在区域属于中亚热带季风气候, 年平均气温19.1 ℃, 年降水量1749 mm, 年蒸发量1585 mm, 相对湿度81%, 平均海拔300 m, 平均坡度25°-35°, 土壤以花岗岩发育的红壤和黄壤为主, 还分布有红黄壤、黄棕壤等, 多呈酸性, 土壤厚度超过1 m (王超, 2012 )。马尾松人工林是经1976年皆伐次生林后营造人工纯林形成; 林分密度为1500株·hm-2 , 平均树高18.3 m, 平均胸径18.3 cm; 林下植被覆盖度约为90%, 灌木较少, 主要以芒萁(Dicranopteris dichotoma )、芒(Miscanthus sinensis )和蕨等草本为主(王超, 2012 ); 林分表层(0-20 cm)的土壤容重为1.33 g·cm-3 , 土壤全N和全P含量分别为0.8 g·kg-1 和0.35 g·kg-1 。米槠人工林则是米槠次生林经皆伐、火烧, 再通过挖穴造林和幼林抚育形成; 林分密度为2042株·hm-2 , 平均树高14.2 m, 平均胸径16.6 cm; 林分内分布的植物主要以毛冬青(Ilex pubescens )、乌饭树(Vaccinium bracteatum )、薄叶山矾(Symplocos anomala )、桂北木姜子( Litsea subcoriacea )、山姜(Alpinia japonica )等为主, 草本以芒萁为主; 林分表层(0-20 cm)的土壤容重为1.18 g·cm-3 , 土壤全N和全P含量分别为1.02 g·kg-1 和0.37 g·kg-1 。
1.2 研究方法
1.2.1 微根管法
2011年4月, 在每种人工林中各建立3块20 m × 20 m的固定标准地, 在每个标准地各安装6-7根PVC材料制成的透明圆形管(即微根管), 每个林分共20根。微根管长度为90 cm, 内径为5 cm, 微根管安装成与地面呈45°角插入土壤中, 换算成垂直深度约为40 cm。外露地表的微根管部分用定制的橡胶塞子塞好, 再用黑色胶布缠绕, 然后用浅色胶布覆盖。从2011年11月开始图像拍摄, 每月一次; 本研究所采用的数据为2013年1月至2014年12月所拍摄图像数据。
在每月下旬使用微根管影像成像系统(ET-100, Bartz Technology, Santa Barbara, USA)采集同一位置根系生长过程的影像, 每片林分根系影像的收集工作均在一天内完成。观测窗面积为1.8 cm × 1.35 cm (长×宽), 每管收集图片40张左右。收集的影像图片带回室内, 使用根系分析系统WinRHIZO TRON MF 2005a图像分析软件进行分析处理, 得到各影像中细根长度、直径等数据。在影像处理过程中, 根据影像采集的时间、林分、标准地号、管号、图像位置、个体根编号等建立细根数据库。依据试验地中亚热带气候的特征, 将3-5月划为春季、6-8月划为夏季、9-11月划为秋季、12-2月划为冬季。将观测到的细根按直径划分为4个径级, 即0-0.3 mm、0.3-0.6 mm、0.6-1.0 mm和1-2 mm。根据微根管垂直观测深度, 将细根出生深度划分为0-10 cm、10-20 cm、20-30 cm和30-40 cm 4个土层。
1.2.2 微根管法细根根长生产量的计算
微根管能够观测到每个拍摄间隔期内新长的细根根长和老根伸长的增加量, 参考Burton等(2000) 文献中的方法, 本研究中每个间隔期内细根的根长生产量等于新长的根长与老根伸长的增加量之和除以总观测窗面积(mm·cm-2 )。林分细根的日生产量等于该间隔期内细根总生产量除以该间隔期的天数(mm·cm-2 ·d-1 ), 所求得的日生产量即代表了这一间隔期内细根生产的平均水平; 再根据当月天数, 由日生产量计算出每月的细根生产量(mm·cm-2 ·month-1 )。
1.2.3 环境因子的测定
在微根管拍摄时, 同时测定土壤10 cm深处的温度数据, 并利用TDR测定土壤10 cm深处的土壤含水率。每月的大气温度、降水量数据来自前期安装的自动小型气象站。
1.2.4 数据处理方法
采用重复测量方差分析方法, 比较两林分细根月根长生产量的差异(以月份为重复测量因素, 林分为处理因素); 采用偏相关分析方法分析细根月生产量与月平均气温、月降水量及测定时刻土壤10 cm深处的温度和含水量等环境因子之间的相关性; 采用一元线性回归分析方法分析两林分细根的月根长生产量与月平均气温和土壤10 cm深处温度之间的线性回归关系; 采用χ 2 检验, 对两林分的细根生产量径级、土层的分布进行差异显著性检验。所有数据的统计分析均用SPSS 17.0软件进行, 用Origin 8.0软件作图。
2 结果分析
2.1 两种林分细根生产量及时间动态
对两林分细根月根长生产量作重复测量方差分析表明(表1 ), 两种林分的细根根长生产量呈显著的月变化(月份: p < 0.001), 且两种林分间细根根长生产量月变化模式有显著的差异(月份×林分: p = 0.004)。同时, 两种林分间的细根根长生产量呈显著的差异(林分: p = 0.005)。
在观测的2年内, 两种林分细根生产量的年际波动均较大。2013年和2014年, 马尾松人工林细根年根长生产量分别为3.25和2.79 mm·cm-2 ·a-1 ; 米槠人工林细根年根长生产量分别为0.90和0.69 mm·cm-2 ·a-1 , 马尾松人工林分别约为米槠人工林的3.6倍和4.0倍。比较两林分2013和2014两年细根生产量的月动态模式发现, 演替后期树种米槠林分的月动态模式更为稳定, 而演替早期马尾松林分的月动态模式稳定性更差。观测期内, 米槠人工林细根月根长生产量的最大值均是出现在6月; 而马尾松人工林细根月根长生产量的每年最大值分别出现在8月和6月; 与2014年相比, 2013年5月和6月的细根根长生产量出现明显的低值。
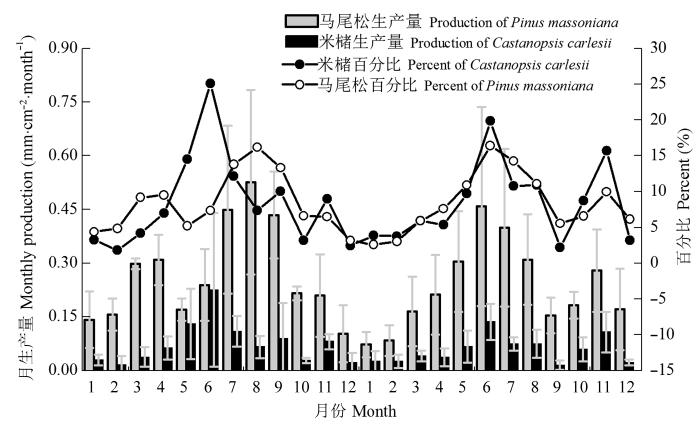
两林分各月细根生产量占当年总生产量的比例如图1 中的折线所示。若以每月细根生产量占年总生产量5%作为评判细根活跃活动的阈值, 观察图1 可知, 马尾松人工林细根月生产量占当年总生产量5%以下的月份集中在冬季(2013年1、2、12月以及2014年的1、2月); 米槠人工林的则有2013年1、2、3、10、12月以及2014年的1、2、9、12月。可以发现, 与演替后期树种米槠人工林相比, 演替早期马尾松人工林的细根生长开始时间可能更早, 而结束时间更晚, 且细根生长季时间更长。
图1 马尾松及米槠人工林2013年1月至2014年12月的细根月根长生产量动态及各月所占当年总生产量的百分比(平均值±标准误差)。
Fig. 1 Monthly fine root production and proportion of Pinus massoniana and Castanopsis carlesii plantation from January 2013 to December 2014 (mean ± SE ).
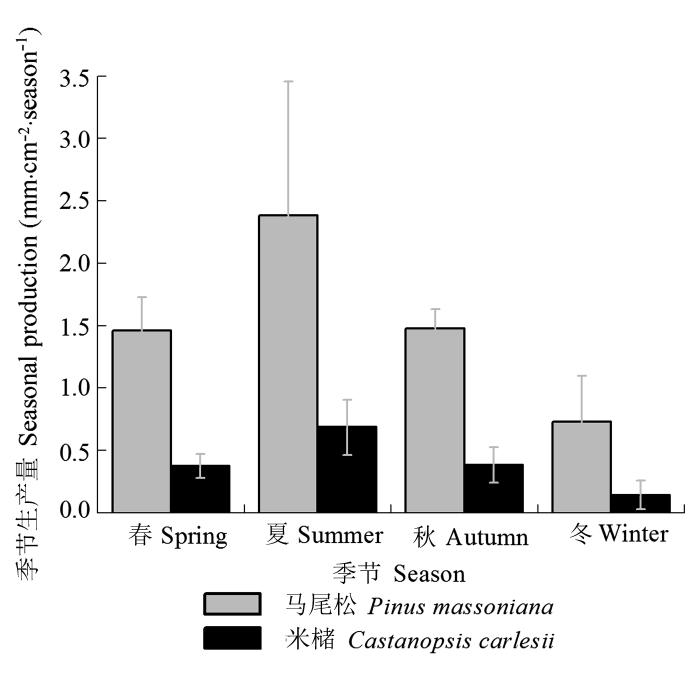
从细根根长生产量的季节分布来看(图2 ), 马尾松和米槠人工林间十分相似, 均以夏季生产量最高, 分别为2.38 mm·cm-2 (39.4%)和0.69 mm·cm-2 (43.3%); 其次是秋季, 分别为1.48 mm·cm-2 (24.4%)和0.38 mm·cm-2 (24.1%); 再次是春季, 分别为1.46 mm·cm-2 (24.1%)和0.37 mm·cm-2 (23.6%); 季节生产量最低的均是冬季, 分别为0.73 mm·cm-2 (12.1%)和0.14 mm·cm-2 (9.0%)。
图2 马尾松及米槠人工林2013-2014年细根的季节生产量(平均值±标准误差)。
Fig. 2 Seasonal distribution of total fine root production in Pinus massoniana and Castanopsis carlesii plantations during the two-year experiment period (mean ± SE ).
2.2 两种林分细根月生产量动态与环境因子的关系
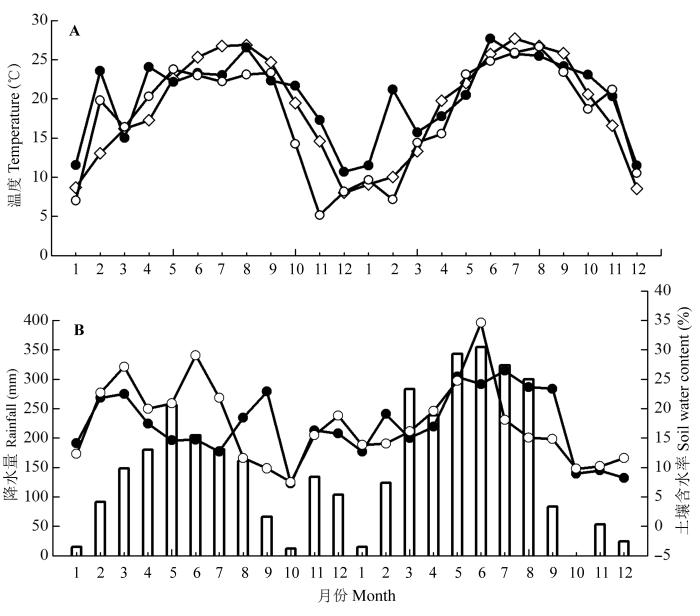
试验地月平均气温、降水量, 以及两种林分测定时刻的土壤温度、土壤含水量如图3 所示。
图3 试验地2013年1月至2014年12月的月平均气温、降水量及土壤温度、土壤含水率分布图。A, 试验地月平均气温(◇)和马尾松人工林(●)和米槠人工林(○)的土壤温度。B, 试验地月降水量(□)和马尾松人工林(●)和米槠人工林(○)的土壤含水率。
Fig. 3 Monthly mean air temperature, rainfall, soil temperature and soil water content of the experimental sites from January 2013 to December 2014. A, Monthly mean air temperature (◇) of experimental field, soil temperature of Pinus massoniana plantation (●) and Castanopsis carlesii plantation (○). B, Monthly rainfall of experimental field (□), soil water content of Pinus massoniana plantation (●) and Castanopsis carlesii plantation (○).
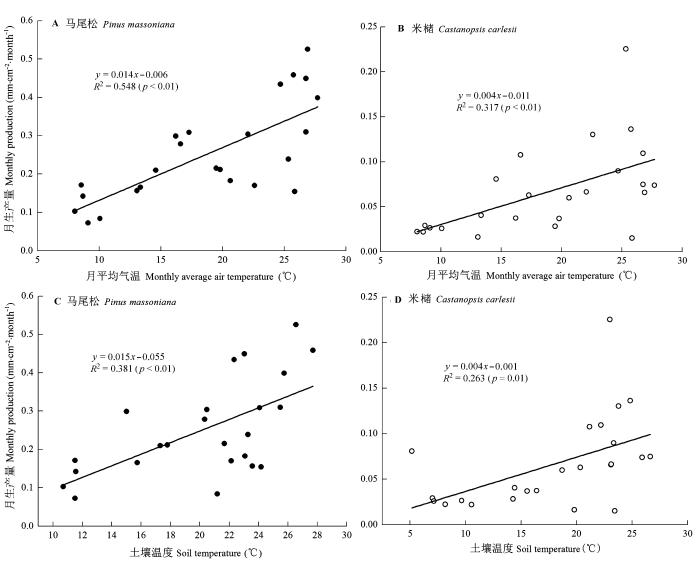
对两种人工林的细根月生产量与各环境因子作偏相关分析, 结果显示, 马尾松人工林两年间的细根月生产量与气温(控制降水)、土壤温度(控制土壤含水率)均呈极显著正偏相关(p < 0.01), 而米槠人工林的细根月生产量与气温(控制降水)、土壤温度(控制土壤含水率)均呈显著正偏相关(p < 0.05); 两林分的细根月生产量与降水(控制气温)、土壤含水率(控制土壤温度)的偏相关均不显著。分别对两林分的细根月生产量动态与气温、土壤温度进行一元线性回归分析, 结果如图4 所示。马尾松人工林细根月生产量与气温、土壤温度的线性回归直线的斜率均远高于米槠人工林, 同时决定系数R 2 亦大于米槠人工林。这表明两林分的细根月生产量主要受温度(气温和土壤温度)的影响; 且马尾松人工林细根月生产量与温度间的相关性高于米槠人工林。
图4 两林分月生产量分别与温度因子的相关图及回归方程。
Fig. 4 Correlation diagrams and regression equations of fine roots monthly production and environmental factors of the two forests.
2.3 两种人工林细根生产量的径级和土层分布
2.3.1 两种人工林细根生产量的径级分布
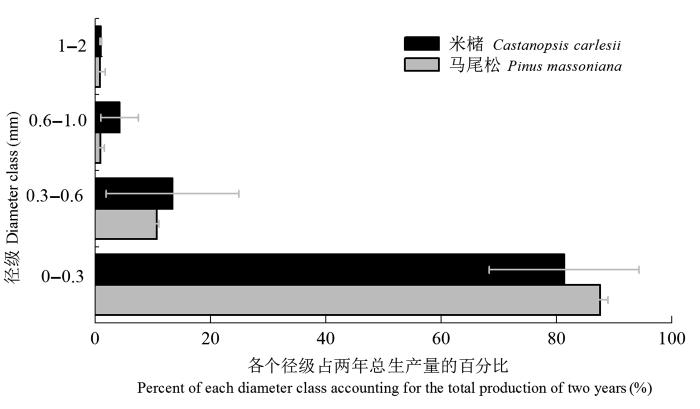
马尾松和米槠人工林在观测期2年内的细根根长生产量均呈现出随着径级增大而减少的趋势(图5 ), 且这种趋势在前3个径级表现尤为明显。两种人工林均是0-0.3 mm径级细根根长生产量所占比例最大, 分别占87.5%和81.3%; 其次是0.3-0.6 mm径级, 马尾松和米槠人工林分别为10.7%和13.4%; 而0.6-1.0 mm和1-2 mm径级所占的比例均比较小。表明两种人工林细根根长生产量均绝大部分分布在0-0.3 mm径级内。经χ 2 检验, 两林分细根根长生产量的径级分布并无显著差异(p > 0.05)。
图5 马尾松及米槠人工林细根2年总生产量的径级分布(平均值±标准误差)。
Fig. 5 Distribution among diameter classes for total fine root production of Pinus massoniana and Castanopsis carlesii plantations during the two-year experiment period (mean ± SE ).
2.3.2 两种人工林细根生产量的土层分布
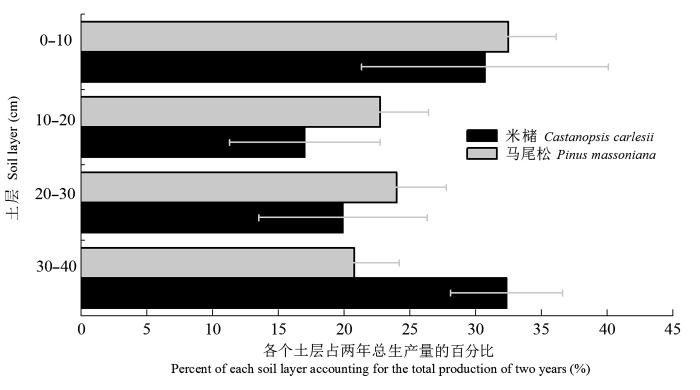
马尾松和米槠人工林细根2年总根长生产量的土层分配如图6 所示。马尾松人工林0-10 cm土层所占细根根长生产量的比例为32.5%, 明显高于其他三个土层; 而其他3个土层所占比例相差不大, 10-20 cm土层、20-30 cm土层和30-40 cm土层分别为22.7%、24.0%和20.8%。而米槠人工林30-40 cm土层所占细根根长生产量的比例最高, 达32.4%; 0-10 cm土层所占比例略低于30-40 cm土层, 为30.7%; 20-30 cm土层占20.0%; 而10-20 cm土层所占比例最小, 为17.0%。经χ 2 检验, 两林分细根根长生产量在土层分布上并无显著差异(p > 0.05)。
图6 马尾松及米槠人工林细根2年总生产量的土层分布(平均值±标准误差)。
Fig. 6 Distribution among soil depths for total fine root production of Pinus massoniana and Castanopsis carlesii plantations during the two-year experiment period (mean ± SE ).
3 讨论
3.1 不同演替阶段树种林分的细根生产量及时间动态差异
本研究中, 马尾松人工林在观测期2年间的细根年根长生产量均明显高于米槠人工林, 不同演替阶段的树种在遗传特性方面存在差异, 可能是造成两种人工林细根生产量相差较大的一个主要原因。不同演替阶段的树种生长速率差异较大(Zangaro et al ., 2012 ), 已有研究表明, 演替早期树种比演替晚期树种具有更高的光合作用能力(Reich et al ., 1998 ; Poorter & Rozendaal, 2008 ), 而这种更高的光合作用能力又将提高可输出为根生长所用的光合产物的数量(Nielsen et al ., 1998 ; Lynch et al ., 2005 ), 进而使得演替早期树种的根生长速率更快。另外, 马尾松为演替早期树种, 对生境适应能力较强, 因而细根的生产量更高; 而米槠属于演替后期树种, 其细根的生产量也较低。Zangaro等(2012)对几个热带树种的研究也发现, 细根生物量、细根直径随演替进行而增加, 而根总长、比根长、根毛长度、根毛发生率、内生菌根菌侵染率等降低, 表明与演替后期阶段树种相比, 演替早期阶段的树种细根可能具有更强的养分获取能力。虽然本研究区域在亚热带, 不同演替阶段树种的细根生产也可能呈现相似的差异特征。两种人工林观测第一年细根生产量均比第二年明显增大, 可能与细根生产的年际波动有关, 也可能与微根管安装过程对细根生产的刺激作用有关。有研究表明, 微根管安装切断了原有根系, 导致初期根系的大量增生与分枝, 这种刺激作用可能随时间的推移而减弱(Joslin & Wolfe, 1997 )。
我们研究的两种人工林, 在观测的两年间细根月生产量的最高值均出现在夏季月份(6-8月), 且从生产量季节分布看, 两种人工林均是夏季生产量最高, 此结果与Steele等(1997) 以及姜红英等(2010)在温带森林开展的细根生产动态研究的结论相一致。然而, 在黄锦学等(2012) 、郑金兴等(2012)以及胡双成(2015)这几个同处亚热带的微根管研究中, 细根生产高峰出现在春季, 这可能与计算生产量的方式不同(干质量生产量或根长生产量)、观测年份的气候差异、不同树种的细根生产存在差异或者林龄等因素有关。
3.2 环境因子对不同演替阶段林分细根月生产量动态影响的差异
Fukuzawa等(2007)对日本北部一个温带森林的研究证实: 温度是驱使细根生产量呈现季节性特征的重要因素。另有研究表明大气温度是影响细根生产的最重要环境因子(Tierney & Fahey, 2001 )。Steele等(1997)研究发现土壤温度与细根生产呈正相关。本研究中, 马尾松和米槠人工林的细根生产动态与温度因子存在正相关, 也说明了温度对细根生产有较大的影响。图4D 中, 有两点偏离较明显(2013年6月和2013年11月), 6月和11月应该都是米槠人工林细根生产量的峰值(夏季和秋季), 可能因为受本身的树种特性(遗传)的影响更大, 而受温度的影响可能较小, 造成这两个月林分土壤温度虽然偏低, 但细根生产量特别大。与温度相比, 本研究中水分对细根生产的影响较小, 这可能是因为试验地所在地区雨热同期, 常年降水量和土壤水分较为充足, 使得细根生产量对水分因子的依赖性相对更弱。米槠人工林细根生产与温度的相关性较弱, 这可能与其是演替后期树种有关。有研究证实一些演替晚期的树种具有耐阴性(Kitajima, 1994 ; Walters & Reich, 1996 ; Khurana & Singh, 2006 ; Poorter & Rozendaal, 2008 ), 林冠更为稠密, 透光度减小, 因而对外界温度、光照的变化不敏感, 使得细根生长也受温度影响较小。Du和Fang (2014)对中国东北的白桦-中国山杨林和落叶松林研究发现, 演替早期的先锋树种白桦和中国山杨, 根呼吸对温度的敏感性要高于演替晚期的优势树种落叶松, 表明演替早期树种根系对温度的响应可能更敏感。这也说明演替早期树种的细根生产可能更易受外界气候变化的影响, 而演替晚期树种则可能由于长期处于较稳定的环境条件下, 细根生产对气候变化的响应较小。这从米槠人工林两年间的细根生产月动态模式比马尾松人工林更为稳定亦可反映出来。这可能也与演替晚期的林分更加郁闭, 对外界环境变化更不敏感, 而演替早期的林分更加稀疏, 受环境变化影响更大有关。同时, 我们还发现演替早期马尾松林分的细根物候与演替晚期米槠林分存在较大差异, 这可能与马尾松为阳性先锋树种, 林分稀疏, 土壤温度一般较米槠人工林的高有关(图3 )。较高的土壤温度使春季细根生产开始时间提前、而冬季结束时间延迟。Alvarez- Uria和Körner (2007)研究证实, 欧洲桦(Betula pubescens )根系显著生长所需的最低温度是6 ℃。Schenker等(2014)研究表明: 不考虑树种差异, 土壤温度低于7 ℃会使温带阔叶树的根系生物量明显 减少。
3.3 两种人工林细根生产量的径级和土层分布特征
两种人工林两年的观测期内总的细根生产量在径级分配上, 各径级所占总生产量的比例均是随着径级的增大而减少, 且细根生产量绝大部分均是分布在0-0.3 mm直径范围内。胡双成(2015)的研究中, 米槠人工幼林的细根生产在0-0.3 mm范围内所占比重最大, 约为32.8%, 本研究结论与之较一致。然而, 这与已有的部分微根管法研究中的结果有所差异。黄锦学等(2012)对细柄阿丁枫(Altingia gracilipes )和米槠天然林的研究中, 0.3-0.6 mm直径的细根生产所占比例最大, 细柄阿丁枫和米槠天然林分别占43.8%和44.7%。在Tierney和Fahey (2001)对温带阔叶林的研究中, 也是直径在0.3-0.6 mm范围内的细根生产占总细根生产量的比例最大。这可能与生产量表示的方法不同有关, 本研究用的是细根的根长生产量, 而前述的两个研究用的是干物质生产量; 此外, 树种特性差异以及不同温度带在环境因子上存在差别也可能是研究结果不同的原因。
本研究中, 马尾松人工林的表层细根生产量占观测期总细根生产量的比例最大, Fukuzawa等(2007)对日本北部一处主要被Quercus crispula 和Sasa senanensis 覆盖的寒温带森林的研究也显示表层土壤(0-15 cm)细根生产量所占比例最高。史建伟等(2007)对落叶松和水曲柳(Fraxinus mandshurica )的研究表明, 两种林分的细根表层年生产量均最高, 底层最低。然而本研究中两种人工林细根生产量并未随着土层加深而递减, 马尾松人工林20-30 cm土层细根生产量所占比例也大于10-20 cm土层; 米槠人工林的底层细根生产量所占比例甚至略高于表层, 并超过了中间两个土层细根生产量所占的比例。可能是演替后期米槠林分的根系在土壤资源利用策略上竞争更激烈, 导致根系更加充分地利用底层土壤资源, 使得底层根系生产量略高于表层。而演替早期马尾松则是表层0-10 cm所占比例最高, 这可能与其根系对土壤资源的竞争强度较小, 而更集中在表层有关。王韦韦等(2015)采用土芯法对马尾松和米槠人工林的研究表明, 0-10 cm细根生物量明显多于其他土层。而本研究中细根生产并未有类似特征, 这可能与研究方法不同有关。与土芯法等比较, 许多研究均证明微根管法低估了表土层细根生产量, 而夸大了深土层细根生产量(Bragg et al ., 1983 ; Upchurch & Ritchie, 1983 ; Parker et al ., 1991 ; de Ruijter et al ., 1996 )。表土层根系生长的低估可能与管-土界面缝隙或压实(Dixon et al ., 1994 )和土壤温度上升(Hansson et al ., 1995 )有关; 而底层土壤根系生长的高估则可能与根系倾向于沿管壁生长有关(Parker et al ., 1991 ; Heeraman & Juma, 1993 )。我们的其他微根管研究也得出相似的结论, 如黄锦学等(2012)的研究中, 细根生产均表现为20-40 cm土层的细根生产量所占比重最大; 胡双成(2015)的研究中也是20-40 cm土层细根所占生产量的比例最大, 可能也是由于微根管法低估表层细根生产造成的。 我们未能检测出两个林分的垂直分布差异, 主要还是因为细根变异大、样本量不足。
4 结论
4.1 本文通过研究马尾松人工林和米槠人工林2013至2014两年的细根月根长生产量及其动态后发现, 两种林分的细根生产量呈现显著差异, 马尾松人工林细根年根长生产量约为米槠人工林的4倍; 两种林分的细根生产量呈现显著的月变化, 峰值均出现在夏季, 且两年内总细根生产量以夏季出生的细根生产量最大。
4.2 两林分细根月生产量均与气温、土壤温度极显著或显著正偏相关, 与降水量、土壤含水率的偏相关均不显著; 演替早期马尾松人工林细根月生产量与气温、土壤温度的相关性明显高于米槠人工林。
4.3 两林分均是直径0-0.3 mm细根所占细根生产量比例最大; 土层分布上, 马尾松人工林0-10 cm土层细根所占生产量的比例最大, 米槠人工林30-40 cm土层细根所占生产量比例最大。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Alvarez-Uria P Körner C 2007 ). Low temperature limits of root growth in deciduous and evergreen temperate tree species.
Functional Ecology , 21 , 211 -218 .
[2]
Bragg PL Govi G Cannell RQ 1983 ). A comparison of methods, including angled and vertical minirhizotrons, for studying root growth and distribution in a spring oat crop.
Plant and Soil , 73 , 435 -440 .
[本文引用: 1]
[3]
Brassard BW Chen HYH Bergeron Y 2009 ). Influence of environmental variability on root dynamics in northern forests.
Critical Reviews in Plant Sciences , 28 , 179 -197 .
[本文引用: 2]
[4]
Burton AJ Pregitzer KS Hendrick RL 2000 ). Relationships between fine root dynamics and nitrogen availability in Michigan northern hardwood forests.
Oecologia , 125 , 389 -399 .
[本文引用: 1]
[5]
Coleman MD Dickson RE Isebrands JG 2000 ). Contrasting fine-root production, survival and soil CO2 efflux in pine and poplar plantations.
Plant and Soil , 225 , 129 -139 .
[本文引用: 1]
[6]
Coll L Potvin C Messier C Delagrange S 2008 ). Root architecture and allocation patterns of eight native tropical species with different successional status used in open-grown mixed plantations in Panama.
Trees , 22 , 585 -596 .
[7]
de Ruijter FJ Veen BW van Oijen M 1996 ). A comparison of soil core sampling and minirhizotrons to quantify root development of field-grown potatoes.
Plant and Soil , 182 , 301 -312 .
[本文引用: 1]
[8]
Dixon RK Solomon AM Brown S Houghton RA Trexier MC Wisniewski J 1994 ). Carbon pools and flux of global forest ecosystems.
Science , 263 , 185 -190 .
[本文引用: 1]
[9]
Du EZ Fang JY 2014 ). Linking belowground and aboveground phenology in two boreal forests in Northeast China.
Oecologia , 176 , 883 -892 .
[本文引用: 1]
[10]
Ellis AR Hubbell SP Potvin C 2000 ). In situ field measurements of photosynthetic rates of tropical tree species: A test of the functional group hypothesis.
Canadian Journal of Botany , 78 , 1336 -1347 .
[本文引用: 1]
[11]
Fukuzawa K Shibata H Takagi K Satoh F Koike T Sasa K 2007 ). Vertical distribution and seasonal pattern of fine-root dynamics in a cool-temperate forest in northern Japan: Implication of the understory vegetation, Sasa dwarf bamboo.
Ecological Research , 22 , 485 -495 .
[12]
Gill RA Jackson RB 2000 ). Global patterns of root turnover for terrestrial ecosystems.
New Phytologist , 147 , 13 -31 .
[本文引用: 1]
[13]
Givnish TJ 1988 ). Adaptation to sun and shade: A whole-plant perspective.
Functional Plant Biology , 15 , 63 -92 .
[本文引用: 1]
[14]
Hansson AC Zhao AF Andrén O 1995 ). Fine-root production and mortality in degraded vegetation in Horqin Sandy Rangeland in Inner Mongolia, China.
Arid Soil Research and Rehabilitation , 9 , 1 -13 .
[本文引用: 1]
[15]
Heeraman DA Juma NG 1993 ). A comparison of minirhizotron, core and monolith methods for quantifying barley (Hordeum vulgare L.) and fababean (Vicia faba L.) root distribution.
Plant and Soil , 148 , 29 -41 .
[本文引用: 1]
[16]
Hu SC 2015 ). Fine Root Production, Mortality and Biomass at Early Stages of Different Regeneration Methods in Subtropical Forests. Master degree dissertation,
Fujian Normal University , Fuzhou . 1 -48 .
(in Chinese with English abstract) [胡双成 (2015 ). 亚热带森林不同更新方式初期细根生产、死亡和现存量研究
. 硕士学位论文, 福建师范大学, 福州 . 1 -48 .]
[17]
Huang JX Ling H Yang ZJ Lu ZL Xiong DC Chen GS Yang YS Xie JS 2012 ). Estimating fine root production and mortality in subtropical Altingia grlilipes and Castanopsis carlesii forests.
Acta Ecologica Sinica , 32 , 4472 -4480 .
(in Chinese with English abstract) [黄锦学 , 凌华 , 杨智杰 , 卢正立 , 熊德成 , 陈光水 , 杨玉盛 , 谢锦升 (2012 ). 中亚热带细柄阿丁枫和米槠群落细根的生产和死亡动态.
生态学报 , 32 , 4472 -4480 .]
[18]
Jiang HY Gu JC Qiu J Wang ZQ 2010 ). Seasonal variations of fine root production and mortality in Larix gmelinii plantation in 2004-2008.
Chinese Journal of Applied Ecology , 21 , 2465 -2471 .
(in Chinese with English abstract) [姜红英 , 谷加存 , 邱俊 , 王政权 (2010 ). 2004- 2008年落叶松人工林细根生产和死亡的季节动态.
应用生态学报 , 21 , 2465 -2471 .]
[19]
Joslin JD Wolfe MH 1997 ). Disturbances during minirhiz- otron installation can affect root observation data.
Soil Science Society of America Journal , 63 , 218 -221 .
[本文引用: 1]
[20]
Khurana E Singh JS 2006 ). Impact of life-history traits on response of seedlings of five tree species of tropical dry forest to shade.
Journal of Tropical Ecology , 22 , 653 -661 .
[本文引用: 2]
[21]
Kitajima K 1994 ). Relative importance of photosynthetic traits and allocation patterns as correlates of seedling shade tolerance of 13 tropical trees.
Oecologia , 98 , 419 -428 .
[本文引用: 2]
[22]
Liu HF 2006 ). Studies on the Intraspecific Competition in Restoration Phase of Castanopsis carlesii Community of the Tropical Zone. Master degree dissertation,
Fujian Agriculture and Forestry University , Fuzhou . 1 -56 .
[本文引用: 1]
(in Chinese with English abstract) [刘浩凤 (2006 ). 亚热带米槠群落生态恢复中种间竞争研究
. 硕士学位论文, 福建农林大学, 福州 . 1 -56 .]
[本文引用: 1]
[23]
Lynch JP Ho MD Phosphorus L 2005 ). Rhizoeconomics: Carbon costs of phosphorus acquisition.
Plant and Soil , 269 , 45 -56 .
[本文引用: 1]
[24]
Messier C Doucet R Ruel JC Claveau Y Kelly C Lechowicz MJ 1999 ). Functional ecology of advance regeneration in relation to light in boreal forests.
Canadian Journal of Forest Research , 29 , 812 -823 .
[本文引用: 1]
[25]
Nielsen KL Bouma TJ Lynch JP Eissenstat DM 1998 ). Effects of phosphorus availability and vesicular-arbuscular mycorrhizas on the carbon budget of common bean (Phaseolus vulgaris).
New Phytologist , 139 , 647 -656 .
[本文引用: 1]
[26]
Parker CJ Carr MKV Jarvis NJ Puplampu BO Lee VH 1991 ). An evaluation of the minirhizotron technique for estimating root distribution in potatoes.
Journal of Agricultural Science , 116 , 341 -350 .
[本文引用: 2]
[27]
Paz H 2003 ). Root/shoot allocation and root architecture in seedlings: Variation among forest sites, microhabitats, and ecological groups.
Biotropica , 35 , 318 -332 .
[28]
Poorter L Rozendaal DMA 2008 ). Leaf size and leaf display of thirty-eight tropical tree species.
Oecologia , 158 , 35 -46 .
[本文引用: 3]
[29]
Reich PB Tjoelker MG Walters MB Vanderklein DW Buschena C 1998 ). Close association of RGR, leaf and root morphology, seed mass and shade tolerance in seedlings of nine boreal tree species grown in high and low light.
Functional Ecology , 12 , 327 -338 .
[本文引用: 1]
[30]
Schenker G Lenz A Körner C Hoch G 2014 ). Physiological minimum temperatures for root growth in seven common European broad-leaved tree species.
Tree Physiology , 34 , 302 -313 .
[31]
Shi JW Wang ZQ Yu SQ Quan XK Sun Y Jia SX Mei L 2007 ). Estimating fine root production, mortality and turnover with minirhizotrons in Larix gmelinii and Fraxinus mandshurica plantations. Journal of Plant Ecology (
Chinese Version ), 31 , 333 -342 .
(in Chinese with English abstract) [史建伟 , 王政权 , 于水强 , 全先奎 , 孙玥 , 贾淑霞 , 梅莉 (2007 ). 落叶松和水曲柳人工林细根生长、死亡和周转.
植物生态学报 , 31 , 333 -342 .]
[32]
Steele SJ Gower ST Vogel JG Norman JM 1997 ). Root mass, net primary production and turnover in aspen, jack pine and black spruce forests in Saskatchewan and Manitoba, Canada.
Tree Physiology , 17 , 577 -587 .
[33]
Tierney GL Fahey TJ 2001 ). Evaluating minirhizotron estimates of fine root longevity and production in the forest floor of a temperate broadleaf forest.
Plant and Soil , 229 , 167 -176 .
[本文引用: 1]
[34]
Tu YL Yang J 1995 ). Community biomass research of karst scrub in central Guizhou.
Carsologica Sinica , 14 , 199 -208 .
[本文引用: 1]
(in Chinese) [屠玉麟 , 杨军 (1995 ). 贵州中部喀斯特灌丛群落生物量研究.
中国岩溶 , 14 , 199 -208 .]
[本文引用: 1]
[35]
Upchurch DR Ritchie JT 1983 ). Root observations using a video recording system in mini-rhizotrons.
Agronomy Journal , 75 , 1009 -1015 .
[本文引用: 1]
[36]
Walters MB Reich PB 1996 ). Are shade tolerance, survival, and growth linked? Low light and nitrogen effects on hardwood seedlings.
Ecology , 77 , 841 -853 .
[本文引用: 2]
[37]
Wang BS Peng SL 1985 ). Analysis on the forest communities of Dinghushan V. Linear system of the community succession.
Acta Scifntiarum Naturalium Universitatis Sunyaatseni , (4 ), 75 -80 .
[本文引用: 1]
(in Chinese with English abstract) [王伯荪 , 彭少麟 (1985 ). 鼎湖山森林群落分析V. 群落演替的线性系统与预测
. 中山大学学报 , (4 ), 75 -80 .]
[本文引用: 1]
[38]
Wang C 2012 ). The Dynamic of Soil Respiration and Its Mechanism to Various Regeneration of Broad-Leaved Evergreen Forests. Master degree dissertation,
Fujian Normal University, Fuzhou . 1 -48 .
[本文引用: 2]
(in Chinese) [王超 (2012 ). 常绿阔叶林不同更新方式下土壤呼吸的动态及机制
. 硕士学位论文, 福建师范大学, 福州 . 1 -48 .]
[本文引用: 2]
[39]
Wang WW Xiong DC Huang JX Huang CC Yang ZJ Hu SC Lin CF Chen GS 2015 ). Comparison of fine-root traits between two subtropical tree species Pinus massoniana and Castanopsis carlesii differing in succession stages.
Acta Ecologica Sinica , 35 , 5813 -5821 .
(in Chinese with English abstract) [王韦韦 , 熊德成 , 黄锦学 , 黄超超 , 杨智杰 , 胡双成 , 林成芳 , 陈光水 (2015 ). 亚热带不同演替树种米槠和马尾松细根性状对比研究.
生态学报 , 35 , 5813 -5821 .]
[40]
Zangaro W Alves RA Lescano LE Ansanelo AP Nogueira MA 2012 ). Investment in fine roots and arbuscular mycorrhizal fungi decrease during succession in three Brazilian ecosystems.
Biotropica , 44 , 141 -150 .
[本文引用: 2]
[41]
Zheng JX Huang JX Wang ZZ Xiong DC Yang ZJ Chen GS 2012 ). Fine root longevity and controlling factors in a Phoebe bournei plantation.
Acta Ecologica Sinica , 32 , 7532 -7539 .
(in Chinese with English abstract) [郑金兴 , 黄锦学 , 王珍珍 , 熊德成 , 杨智杰 , 陈光水 (2012 ). 闽楠人工林细根寿命及其影响因素.
生态学报 , 32 , 7532 -7539 .]
Low temperature limits of root growth in deciduous and evergreen temperate tree species.
0
2007
A comparison of methods, including angled and vertical minirhizotrons, for studying root growth and distribution in a spring oat crop.
1
1983
... 本研究中, 马尾松人工林的表层细根生产量占观测期总细根生产量的比例最大, Fukuzawa等(2007)对日本北部一处主要被Quercus crispula 和Sasa senanensis 覆盖的寒温带森林的研究也显示表层土壤(0-15 cm)细根生产量所占比例最高.史建伟等(2007)对落叶松和水曲柳(Fraxinus mandshurica )的研究表明, 两种林分的细根表层年生产量均最高, 底层最低.然而本研究中两种人工林细根生产量并未随着土层加深而递减, 马尾松人工林20-30 cm土层细根生产量所占比例也大于10-20 cm土层; 米槠人工林的底层细根生产量所占比例甚至略高于表层, 并超过了中间两个土层细根生产量所占的比例.可能是演替后期米槠林分的根系在土壤资源利用策略上竞争更激烈, 导致根系更加充分地利用底层土壤资源, 使得底层根系生产量略高于表层.而演替早期马尾松则是表层0-10 cm所占比例最高, 这可能与其根系对土壤资源的竞争强度较小, 而更集中在表层有关.王韦韦等(2015)采用土芯法对马尾松和米槠人工林的研究表明, 0-10 cm细根生物量明显多于其他土层.而本研究中细根生产并未有类似特征, 这可能与研究方法不同有关.与土芯法等比较, 许多研究均证明微根管法低估了表土层细根生产量, 而夸大了深土层细根生产量(Bragg et al ., 1983 ; Upchurch & Ritchie, 1983 ; Parker et al ., 1991 ; de Ruijter et al ., 1996 ).表土层根系生长的低估可能与管-土界面缝隙或压实(Dixon et al ., 1994 )和土壤温度上升(Hansson et al ., 1995 )有关; 而底层土壤根系生长的高估则可能与根系倾向于沿管壁生长有关(Parker et al ., 1991 ; Heeraman & Juma, 1993 ).我们的其他微根管研究也得出相似的结论, 如黄锦学等(2012)的研究中, 细根生产均表现为20-40 cm土层的细根生产量所占比重最大; 胡双成(2015)的研究中也是20-40 cm土层细根所占生产量的比例最大, 可能也是由于微根管法低估表层细根生产造成的. 我们未能检测出两个林分的垂直分布差异, 主要还是因为细根变异大、样本量不足. ...
Influence of environmental variability on root dynamics in northern forests.
2
2009
... 不同演替阶段的树种生理和形态特征有所不同(Givnish, 1988 ; Messier, 1999 ; Ellis et al ., 2000 ), 细根特性也可能因所处演替阶段不同而存在差异.Paz (2003)研究了55种树种的根形态特征后发现, 演替晚期树种的根总体上要比演替早期树种的根更粗.Coll等(2008)的研究表明, 演替晚期树种的主根生物量所占比例更大.受生境条件等因素的影响, 不同演替阶段树种的细根生长速度可能也存在差异.在演替早期, 群落结构简单且立地条件相对较差, 植被更倾向于分配更多的生物量给根系(屠玉麟和杨军, 1995 ), 并通过迅速和广泛地占据土壤空间获取资源(Brassard et al ., 2009 ), 具有觅食的广布性; 而演替晚期群落的物种多样性较丰富且结构复杂, 土壤有机质和养分含量较高, 种间竞争激烈, 为了提高竞争力, 细根会迅速识别富养斑块并大量增生(Brassard et al ., 2009 ), 养分利用精细, 利用效率提高.Zangaro等(2012)对几个热带树种的研究发现, 细根生物量、细根直径随演替的进行而增加, 而根总长、比根长、根毛长度、根毛发生率、内生菌根菌侵染率等降低, 表明与演替后期阶段树种相比, 演替早期阶段的树种细根具有更强的养分获取能力, 生长速度可能更快.同时, 不同演替阶段的树种受栖息地环境的影响程度可能不同.如演替早期的热带树种生长受开阔栖息地的局限较大(Zangaro et al ., 2012 ), 而一些演替晚期的树种具有耐阴性(Kitajima, 1994 ; Walters & Reich, 1996 ; Khurana & Singh, 2006 ; Poorter & Rozendaal, 2008 ), 林冠更加稠密, 透光度更小.Du和Fang (2014)对中国东北的白桦(Betula platyphylla )-中国山杨(Populus davidiana )林和落叶松(Larix gmelinii )林的研究也曾发现, 演替早期的先锋树种白桦和中国山杨, 根呼吸对温度的敏感性要高于演替晚期的优势树种落叶松.此外, 不同演替阶段树种的物候情况在长期森林动态研究中十分重要, 且是预测未来气候变化环境下的森林变化所必须考虑的问题(Du & Fang, 2014 ).然而目前有关森林不同演替阶段的树种细根生产动态和细根物候方面差异的研究仍十分缺乏.分析研究不同演替阶段不同树种林分的细根生产量、季节动态及其所受环境因子影响的差异, 将有助于增进我们了解不同演替阶段的树种差异对细根生产可能造成的影响. ...
... ), 具有觅食的广布性; 而演替晚期群落的物种多样性较丰富且结构复杂, 土壤有机质和养分含量较高, 种间竞争激烈, 为了提高竞争力, 细根会迅速识别富养斑块并大量增生(Brassard et al ., 2009 ), 养分利用精细, 利用效率提高.Zangaro等(2012)对几个热带树种的研究发现, 细根生物量、细根直径随演替的进行而增加, 而根总长、比根长、根毛长度、根毛发生率、内生菌根菌侵染率等降低, 表明与演替后期阶段树种相比, 演替早期阶段的树种细根具有更强的养分获取能力, 生长速度可能更快.同时, 不同演替阶段的树种受栖息地环境的影响程度可能不同.如演替早期的热带树种生长受开阔栖息地的局限较大(Zangaro et al ., 2012 ), 而一些演替晚期的树种具有耐阴性(Kitajima, 1994 ; Walters & Reich, 1996 ; Khurana & Singh, 2006 ; Poorter & Rozendaal, 2008 ), 林冠更加稠密, 透光度更小.Du和Fang (2014)对中国东北的白桦(Betula platyphylla )-中国山杨(Populus davidiana )林和落叶松(Larix gmelinii )林的研究也曾发现, 演替早期的先锋树种白桦和中国山杨, 根呼吸对温度的敏感性要高于演替晚期的优势树种落叶松.此外, 不同演替阶段树种的物候情况在长期森林动态研究中十分重要, 且是预测未来气候变化环境下的森林变化所必须考虑的问题(Du & Fang, 2014 ).然而目前有关森林不同演替阶段的树种细根生产动态和细根物候方面差异的研究仍十分缺乏.分析研究不同演替阶段不同树种林分的细根生产量、季节动态及其所受环境因子影响的差异, 将有助于增进我们了解不同演替阶段的树种差异对细根生产可能造成的影响. ...
Relationships between fine root dynamics and nitrogen availability in Michigan northern hardwood forests.
1
2000
... 微根管能够观测到每个拍摄间隔期内新长的细根根长和老根伸长的增加量, 参考Burton等(2000) 文献中的方法, 本研究中每个间隔期内细根的根长生产量等于新长的根长与老根伸长的增加量之和除以总观测窗面积(mm·cm-2 ).林分细根的日生产量等于该间隔期内细根总生产量除以该间隔期的天数(mm·cm-2 ·d-1 ), 所求得的日生产量即代表了这一间隔期内细根生产的平均水平; 再根据当月天数, 由日生产量计算出每月的细根生产量(mm·cm-2 ·month-1 ). ...
Contrasting fine-root production, survival and soil CO2 efflux in pine and poplar plantations.
1
2000
... 林木细根生产量约占全球净初级生产力的30%, 在森林生态系统物质和能量循环过程中具有十分重要的作用(Gill & Jackson, 2000 ).不同树种的特性存在差别, 了解不同树种间根系生产动态及其与环境因子间关系的差异, 对于评估不同森林生态系统生产力及其对全球环境变化响应的差别具有重要的作用(Coleman et al ., 2000 ). ...
Root architecture and allocation patterns of eight native tropical species with different successional status used in open-grown mixed plantations in Panama.
0
2008
A comparison of soil core sampling and minirhizotrons to quantify root development of field-grown potatoes.
1
1996
... 本研究中, 马尾松人工林的表层细根生产量占观测期总细根生产量的比例最大, Fukuzawa等(2007)对日本北部一处主要被Quercus crispula 和Sasa senanensis 覆盖的寒温带森林的研究也显示表层土壤(0-15 cm)细根生产量所占比例最高.史建伟等(2007)对落叶松和水曲柳(Fraxinus mandshurica )的研究表明, 两种林分的细根表层年生产量均最高, 底层最低.然而本研究中两种人工林细根生产量并未随着土层加深而递减, 马尾松人工林20-30 cm土层细根生产量所占比例也大于10-20 cm土层; 米槠人工林的底层细根生产量所占比例甚至略高于表层, 并超过了中间两个土层细根生产量所占的比例.可能是演替后期米槠林分的根系在土壤资源利用策略上竞争更激烈, 导致根系更加充分地利用底层土壤资源, 使得底层根系生产量略高于表层.而演替早期马尾松则是表层0-10 cm所占比例最高, 这可能与其根系对土壤资源的竞争强度较小, 而更集中在表层有关.王韦韦等(2015)采用土芯法对马尾松和米槠人工林的研究表明, 0-10 cm细根生物量明显多于其他土层.而本研究中细根生产并未有类似特征, 这可能与研究方法不同有关.与土芯法等比较, 许多研究均证明微根管法低估了表土层细根生产量, 而夸大了深土层细根生产量(Bragg et al ., 1983 ; Upchurch & Ritchie, 1983 ; Parker et al ., 1991 ; de Ruijter et al ., 1996 ).表土层根系生长的低估可能与管-土界面缝隙或压实(Dixon et al ., 1994 )和土壤温度上升(Hansson et al ., 1995 )有关; 而底层土壤根系生长的高估则可能与根系倾向于沿管壁生长有关(Parker et al ., 1991 ; Heeraman & Juma, 1993 ).我们的其他微根管研究也得出相似的结论, 如黄锦学等(2012)的研究中, 细根生产均表现为20-40 cm土层的细根生产量所占比重最大; 胡双成(2015)的研究中也是20-40 cm土层细根所占生产量的比例最大, 可能也是由于微根管法低估表层细根生产造成的. 我们未能检测出两个林分的垂直分布差异, 主要还是因为细根变异大、样本量不足. ...
Carbon pools and flux of global forest ecosystems.
1
1994
... 本研究中, 马尾松人工林的表层细根生产量占观测期总细根生产量的比例最大, Fukuzawa等(2007)对日本北部一处主要被Quercus crispula 和Sasa senanensis 覆盖的寒温带森林的研究也显示表层土壤(0-15 cm)细根生产量所占比例最高.史建伟等(2007)对落叶松和水曲柳(Fraxinus mandshurica )的研究表明, 两种林分的细根表层年生产量均最高, 底层最低.然而本研究中两种人工林细根生产量并未随着土层加深而递减, 马尾松人工林20-30 cm土层细根生产量所占比例也大于10-20 cm土层; 米槠人工林的底层细根生产量所占比例甚至略高于表层, 并超过了中间两个土层细根生产量所占的比例.可能是演替后期米槠林分的根系在土壤资源利用策略上竞争更激烈, 导致根系更加充分地利用底层土壤资源, 使得底层根系生产量略高于表层.而演替早期马尾松则是表层0-10 cm所占比例最高, 这可能与其根系对土壤资源的竞争强度较小, 而更集中在表层有关.王韦韦等(2015)采用土芯法对马尾松和米槠人工林的研究表明, 0-10 cm细根生物量明显多于其他土层.而本研究中细根生产并未有类似特征, 这可能与研究方法不同有关.与土芯法等比较, 许多研究均证明微根管法低估了表土层细根生产量, 而夸大了深土层细根生产量(Bragg et al ., 1983 ; Upchurch & Ritchie, 1983 ; Parker et al ., 1991 ; de Ruijter et al ., 1996 ).表土层根系生长的低估可能与管-土界面缝隙或压实(Dixon et al ., 1994 )和土壤温度上升(Hansson et al ., 1995 )有关; 而底层土壤根系生长的高估则可能与根系倾向于沿管壁生长有关(Parker et al ., 1991 ; Heeraman & Juma, 1993 ).我们的其他微根管研究也得出相似的结论, 如黄锦学等(2012)的研究中, 细根生产均表现为20-40 cm土层的细根生产量所占比重最大; 胡双成(2015)的研究中也是20-40 cm土层细根所占生产量的比例最大, 可能也是由于微根管法低估表层细根生产造成的. 我们未能检测出两个林分的垂直分布差异, 主要还是因为细根变异大、样本量不足. ...
Linking belowground and aboveground phenology in two boreal forests in Northeast China.
1
2014
... 不同演替阶段的树种生理和形态特征有所不同(Givnish, 1988 ; Messier, 1999 ; Ellis et al ., 2000 ), 细根特性也可能因所处演替阶段不同而存在差异.Paz (2003)研究了55种树种的根形态特征后发现, 演替晚期树种的根总体上要比演替早期树种的根更粗.Coll等(2008)的研究表明, 演替晚期树种的主根生物量所占比例更大.受生境条件等因素的影响, 不同演替阶段树种的细根生长速度可能也存在差异.在演替早期, 群落结构简单且立地条件相对较差, 植被更倾向于分配更多的生物量给根系(屠玉麟和杨军, 1995 ), 并通过迅速和广泛地占据土壤空间获取资源(Brassard et al ., 2009 ), 具有觅食的广布性; 而演替晚期群落的物种多样性较丰富且结构复杂, 土壤有机质和养分含量较高, 种间竞争激烈, 为了提高竞争力, 细根会迅速识别富养斑块并大量增生(Brassard et al ., 2009 ), 养分利用精细, 利用效率提高.Zangaro等(2012)对几个热带树种的研究发现, 细根生物量、细根直径随演替的进行而增加, 而根总长、比根长、根毛长度、根毛发生率、内生菌根菌侵染率等降低, 表明与演替后期阶段树种相比, 演替早期阶段的树种细根具有更强的养分获取能力, 生长速度可能更快.同时, 不同演替阶段的树种受栖息地环境的影响程度可能不同.如演替早期的热带树种生长受开阔栖息地的局限较大(Zangaro et al ., 2012 ), 而一些演替晚期的树种具有耐阴性(Kitajima, 1994 ; Walters & Reich, 1996 ; Khurana & Singh, 2006 ; Poorter & Rozendaal, 2008 ), 林冠更加稠密, 透光度更小.Du和Fang (2014)对中国东北的白桦(Betula platyphylla )-中国山杨(Populus davidiana )林和落叶松(Larix gmelinii )林的研究也曾发现, 演替早期的先锋树种白桦和中国山杨, 根呼吸对温度的敏感性要高于演替晚期的优势树种落叶松.此外, 不同演替阶段树种的物候情况在长期森林动态研究中十分重要, 且是预测未来气候变化环境下的森林变化所必须考虑的问题(Du & Fang, 2014 ).然而目前有关森林不同演替阶段的树种细根生产动态和细根物候方面差异的研究仍十分缺乏.分析研究不同演替阶段不同树种林分的细根生产量、季节动态及其所受环境因子影响的差异, 将有助于增进我们了解不同演替阶段的树种差异对细根生产可能造成的影响. ...
In situ field measurements of photosynthetic rates of tropical tree species: A test of the functional group hypothesis.
1
2000
... 不同演替阶段的树种生理和形态特征有所不同(Givnish, 1988 ; Messier, 1999 ; Ellis et al ., 2000 ), 细根特性也可能因所处演替阶段不同而存在差异.Paz (2003)研究了55种树种的根形态特征后发现, 演替晚期树种的根总体上要比演替早期树种的根更粗.Coll等(2008)的研究表明, 演替晚期树种的主根生物量所占比例更大.受生境条件等因素的影响, 不同演替阶段树种的细根生长速度可能也存在差异.在演替早期, 群落结构简单且立地条件相对较差, 植被更倾向于分配更多的生物量给根系(屠玉麟和杨军, 1995 ), 并通过迅速和广泛地占据土壤空间获取资源(Brassard et al ., 2009 ), 具有觅食的广布性; 而演替晚期群落的物种多样性较丰富且结构复杂, 土壤有机质和养分含量较高, 种间竞争激烈, 为了提高竞争力, 细根会迅速识别富养斑块并大量增生(Brassard et al ., 2009 ), 养分利用精细, 利用效率提高.Zangaro等(2012)对几个热带树种的研究发现, 细根生物量、细根直径随演替的进行而增加, 而根总长、比根长、根毛长度、根毛发生率、内生菌根菌侵染率等降低, 表明与演替后期阶段树种相比, 演替早期阶段的树种细根具有更强的养分获取能力, 生长速度可能更快.同时, 不同演替阶段的树种受栖息地环境的影响程度可能不同.如演替早期的热带树种生长受开阔栖息地的局限较大(Zangaro et al ., 2012 ), 而一些演替晚期的树种具有耐阴性(Kitajima, 1994 ; Walters & Reich, 1996 ; Khurana & Singh, 2006 ; Poorter & Rozendaal, 2008 ), 林冠更加稠密, 透光度更小.Du和Fang (2014)对中国东北的白桦(Betula platyphylla )-中国山杨(Populus davidiana )林和落叶松(Larix gmelinii )林的研究也曾发现, 演替早期的先锋树种白桦和中国山杨, 根呼吸对温度的敏感性要高于演替晚期的优势树种落叶松.此外, 不同演替阶段树种的物候情况在长期森林动态研究中十分重要, 且是预测未来气候变化环境下的森林变化所必须考虑的问题(Du & Fang, 2014 ).然而目前有关森林不同演替阶段的树种细根生产动态和细根物候方面差异的研究仍十分缺乏.分析研究不同演替阶段不同树种林分的细根生产量、季节动态及其所受环境因子影响的差异, 将有助于增进我们了解不同演替阶段的树种差异对细根生产可能造成的影响. ...
Vertical distribution and seasonal pattern of fine-root dynamics in a cool-temperate forest in northern Japan: Implication of the understory vegetation, Sasa dwarf bamboo.
0
2007
Global patterns of root turnover for terrestrial ecosystems.
1
2000
... 林木细根生产量约占全球净初级生产力的30%, 在森林生态系统物质和能量循环过程中具有十分重要的作用(Gill & Jackson, 2000 ).不同树种的特性存在差别, 了解不同树种间根系生产动态及其与环境因子间关系的差异, 对于评估不同森林生态系统生产力及其对全球环境变化响应的差别具有重要的作用(Coleman et al ., 2000 ). ...
Adaptation to sun and shade: A whole-plant perspective.
1
1988
... 不同演替阶段的树种生理和形态特征有所不同(Givnish, 1988 ; Messier, 1999 ; Ellis et al ., 2000 ), 细根特性也可能因所处演替阶段不同而存在差异.Paz (2003)研究了55种树种的根形态特征后发现, 演替晚期树种的根总体上要比演替早期树种的根更粗.Coll等(2008)的研究表明, 演替晚期树种的主根生物量所占比例更大.受生境条件等因素的影响, 不同演替阶段树种的细根生长速度可能也存在差异.在演替早期, 群落结构简单且立地条件相对较差, 植被更倾向于分配更多的生物量给根系(屠玉麟和杨军, 1995 ), 并通过迅速和广泛地占据土壤空间获取资源(Brassard et al ., 2009 ), 具有觅食的广布性; 而演替晚期群落的物种多样性较丰富且结构复杂, 土壤有机质和养分含量较高, 种间竞争激烈, 为了提高竞争力, 细根会迅速识别富养斑块并大量增生(Brassard et al ., 2009 ), 养分利用精细, 利用效率提高.Zangaro等(2012)对几个热带树种的研究发现, 细根生物量、细根直径随演替的进行而增加, 而根总长、比根长、根毛长度、根毛发生率、内生菌根菌侵染率等降低, 表明与演替后期阶段树种相比, 演替早期阶段的树种细根具有更强的养分获取能力, 生长速度可能更快.同时, 不同演替阶段的树种受栖息地环境的影响程度可能不同.如演替早期的热带树种生长受开阔栖息地的局限较大(Zangaro et al ., 2012 ), 而一些演替晚期的树种具有耐阴性(Kitajima, 1994 ; Walters & Reich, 1996 ; Khurana & Singh, 2006 ; Poorter & Rozendaal, 2008 ), 林冠更加稠密, 透光度更小.Du和Fang (2014)对中国东北的白桦(Betula platyphylla )-中国山杨(Populus davidiana )林和落叶松(Larix gmelinii )林的研究也曾发现, 演替早期的先锋树种白桦和中国山杨, 根呼吸对温度的敏感性要高于演替晚期的优势树种落叶松.此外, 不同演替阶段树种的物候情况在长期森林动态研究中十分重要, 且是预测未来气候变化环境下的森林变化所必须考虑的问题(Du & Fang, 2014 ).然而目前有关森林不同演替阶段的树种细根生产动态和细根物候方面差异的研究仍十分缺乏.分析研究不同演替阶段不同树种林分的细根生产量、季节动态及其所受环境因子影响的差异, 将有助于增进我们了解不同演替阶段的树种差异对细根生产可能造成的影响. ...
Fine-root production and mortality in degraded vegetation in Horqin Sandy Rangeland in Inner Mongolia, China.
1
1995
... 本研究中, 马尾松人工林的表层细根生产量占观测期总细根生产量的比例最大, Fukuzawa等(2007)对日本北部一处主要被Quercus crispula 和Sasa senanensis 覆盖的寒温带森林的研究也显示表层土壤(0-15 cm)细根生产量所占比例最高.史建伟等(2007)对落叶松和水曲柳(Fraxinus mandshurica )的研究表明, 两种林分的细根表层年生产量均最高, 底层最低.然而本研究中两种人工林细根生产量并未随着土层加深而递减, 马尾松人工林20-30 cm土层细根生产量所占比例也大于10-20 cm土层; 米槠人工林的底层细根生产量所占比例甚至略高于表层, 并超过了中间两个土层细根生产量所占的比例.可能是演替后期米槠林分的根系在土壤资源利用策略上竞争更激烈, 导致根系更加充分地利用底层土壤资源, 使得底层根系生产量略高于表层.而演替早期马尾松则是表层0-10 cm所占比例最高, 这可能与其根系对土壤资源的竞争强度较小, 而更集中在表层有关.王韦韦等(2015)采用土芯法对马尾松和米槠人工林的研究表明, 0-10 cm细根生物量明显多于其他土层.而本研究中细根生产并未有类似特征, 这可能与研究方法不同有关.与土芯法等比较, 许多研究均证明微根管法低估了表土层细根生产量, 而夸大了深土层细根生产量(Bragg et al ., 1983 ; Upchurch & Ritchie, 1983 ; Parker et al ., 1991 ; de Ruijter et al ., 1996 ).表土层根系生长的低估可能与管-土界面缝隙或压实(Dixon et al ., 1994 )和土壤温度上升(Hansson et al ., 1995 )有关; 而底层土壤根系生长的高估则可能与根系倾向于沿管壁生长有关(Parker et al ., 1991 ; Heeraman & Juma, 1993 ).我们的其他微根管研究也得出相似的结论, 如黄锦学等(2012)的研究中, 细根生产均表现为20-40 cm土层的细根生产量所占比重最大; 胡双成(2015)的研究中也是20-40 cm土层细根所占生产量的比例最大, 可能也是由于微根管法低估表层细根生产造成的. 我们未能检测出两个林分的垂直分布差异, 主要还是因为细根变异大、样本量不足. ...
A comparison of minirhizotron, core and monolith methods for quantifying barley (Hordeum vulgare L.) and fababean (Vicia faba L.) root distribution.
1
1993
... 本研究中, 马尾松人工林的表层细根生产量占观测期总细根生产量的比例最大, Fukuzawa等(2007)对日本北部一处主要被Quercus crispula 和Sasa senanensis 覆盖的寒温带森林的研究也显示表层土壤(0-15 cm)细根生产量所占比例最高.史建伟等(2007)对落叶松和水曲柳(Fraxinus mandshurica )的研究表明, 两种林分的细根表层年生产量均最高, 底层最低.然而本研究中两种人工林细根生产量并未随着土层加深而递减, 马尾松人工林20-30 cm土层细根生产量所占比例也大于10-20 cm土层; 米槠人工林的底层细根生产量所占比例甚至略高于表层, 并超过了中间两个土层细根生产量所占的比例.可能是演替后期米槠林分的根系在土壤资源利用策略上竞争更激烈, 导致根系更加充分地利用底层土壤资源, 使得底层根系生产量略高于表层.而演替早期马尾松则是表层0-10 cm所占比例最高, 这可能与其根系对土壤资源的竞争强度较小, 而更集中在表层有关.王韦韦等(2015)采用土芯法对马尾松和米槠人工林的研究表明, 0-10 cm细根生物量明显多于其他土层.而本研究中细根生产并未有类似特征, 这可能与研究方法不同有关.与土芯法等比较, 许多研究均证明微根管法低估了表土层细根生产量, 而夸大了深土层细根生产量(Bragg et al ., 1983 ; Upchurch & Ritchie, 1983 ; Parker et al ., 1991 ; de Ruijter et al ., 1996 ).表土层根系生长的低估可能与管-土界面缝隙或压实(Dixon et al ., 1994 )和土壤温度上升(Hansson et al ., 1995 )有关; 而底层土壤根系生长的高估则可能与根系倾向于沿管壁生长有关(Parker et al ., 1991 ; Heeraman & Juma, 1993 ).我们的其他微根管研究也得出相似的结论, 如黄锦学等(2012)的研究中, 细根生产均表现为20-40 cm土层的细根生产量所占比重最大; 胡双成(2015)的研究中也是20-40 cm土层细根所占生产量的比例最大, 可能也是由于微根管法低估表层细根生产造成的. 我们未能检测出两个林分的垂直分布差异, 主要还是因为细根变异大、样本量不足. ...
亚热带森林不同更新方式初期细根生产、死亡和现存量研究
0
2015
亚热带森林不同更新方式初期细根生产、死亡和现存量研究
0
2015
中亚热带细柄阿丁枫和米槠群落细根的生产和死亡动态.
0
2012
中亚热带细柄阿丁枫和米槠群落细根的生产和死亡动态.
0
2012
2004- 2008年落叶松人工林细根生产和死亡的季节动态.
0
2010
2004- 2008年落叶松人工林细根生产和死亡的季节动态.
0
2010
Disturbances during minirhiz- otron installation can affect root observation data.
1
1997
... 本研究中, 马尾松人工林在观测期2年间的细根年根长生产量均明显高于米槠人工林, 不同演替阶段的树种在遗传特性方面存在差异, 可能是造成两种人工林细根生产量相差较大的一个主要原因.不同演替阶段的树种生长速率差异较大(Zangaro et al ., 2012 ), 已有研究表明, 演替早期树种比演替晚期树种具有更高的光合作用能力(Reich et al ., 1998 ; Poorter & Rozendaal, 2008 ), 而这种更高的光合作用能力又将提高可输出为根生长所用的光合产物的数量(Nielsen et al ., 1998 ; Lynch et al ., 2005 ), 进而使得演替早期树种的根生长速率更快.另外, 马尾松为演替早期树种, 对生境适应能力较强, 因而细根的生产量更高; 而米槠属于演替后期树种, 其细根的生产量也较低.Zangaro等(2012)对几个热带树种的研究也发现, 细根生物量、细根直径随演替进行而增加, 而根总长、比根长、根毛长度、根毛发生率、内生菌根菌侵染率等降低, 表明与演替后期阶段树种相比, 演替早期阶段的树种细根可能具有更强的养分获取能力.虽然本研究区域在亚热带, 不同演替阶段树种的细根生产也可能呈现相似的差异特征.两种人工林观测第一年细根生产量均比第二年明显增大, 可能与细根生产的年际波动有关, 也可能与微根管安装过程对细根生产的刺激作用有关.有研究表明, 微根管安装切断了原有根系, 导致初期根系的大量增生与分枝, 这种刺激作用可能随时间的推移而减弱(Joslin & Wolfe, 1997 ). ...
Impact of life-history traits on response of seedlings of five tree species of tropical dry forest to shade.
2
2006
... 不同演替阶段的树种生理和形态特征有所不同(Givnish, 1988 ; Messier, 1999 ; Ellis et al ., 2000 ), 细根特性也可能因所处演替阶段不同而存在差异.Paz (2003)研究了55种树种的根形态特征后发现, 演替晚期树种的根总体上要比演替早期树种的根更粗.Coll等(2008)的研究表明, 演替晚期树种的主根生物量所占比例更大.受生境条件等因素的影响, 不同演替阶段树种的细根生长速度可能也存在差异.在演替早期, 群落结构简单且立地条件相对较差, 植被更倾向于分配更多的生物量给根系(屠玉麟和杨军, 1995 ), 并通过迅速和广泛地占据土壤空间获取资源(Brassard et al ., 2009 ), 具有觅食的广布性; 而演替晚期群落的物种多样性较丰富且结构复杂, 土壤有机质和养分含量较高, 种间竞争激烈, 为了提高竞争力, 细根会迅速识别富养斑块并大量增生(Brassard et al ., 2009 ), 养分利用精细, 利用效率提高.Zangaro等(2012)对几个热带树种的研究发现, 细根生物量、细根直径随演替的进行而增加, 而根总长、比根长、根毛长度、根毛发生率、内生菌根菌侵染率等降低, 表明与演替后期阶段树种相比, 演替早期阶段的树种细根具有更强的养分获取能力, 生长速度可能更快.同时, 不同演替阶段的树种受栖息地环境的影响程度可能不同.如演替早期的热带树种生长受开阔栖息地的局限较大(Zangaro et al ., 2012 ), 而一些演替晚期的树种具有耐阴性(Kitajima, 1994 ; Walters & Reich, 1996 ; Khurana & Singh, 2006 ; Poorter & Rozendaal, 2008 ), 林冠更加稠密, 透光度更小.Du和Fang (2014)对中国东北的白桦(Betula platyphylla )-中国山杨(Populus davidiana )林和落叶松(Larix gmelinii )林的研究也曾发现, 演替早期的先锋树种白桦和中国山杨, 根呼吸对温度的敏感性要高于演替晚期的优势树种落叶松.此外, 不同演替阶段树种的物候情况在长期森林动态研究中十分重要, 且是预测未来气候变化环境下的森林变化所必须考虑的问题(Du & Fang, 2014 ).然而目前有关森林不同演替阶段的树种细根生产动态和细根物候方面差异的研究仍十分缺乏.分析研究不同演替阶段不同树种林分的细根生产量、季节动态及其所受环境因子影响的差异, 将有助于增进我们了解不同演替阶段的树种差异对细根生产可能造成的影响. ...
... Fukuzawa等(2007)对日本北部一个温带森林的研究证实: 温度是驱使细根生产量呈现季节性特征的重要因素.另有研究表明大气温度是影响细根生产的最重要环境因子(Tierney & Fahey, 2001 ).Steele等(1997)研究发现土壤温度与细根生产呈正相关.本研究中, 马尾松和米槠人工林的细根生产动态与温度因子存在正相关, 也说明了温度对细根生产有较大的影响.图4D 中, 有两点偏离较明显(2013年6月和2013年11月), 6月和11月应该都是米槠人工林细根生产量的峰值(夏季和秋季), 可能因为受本身的树种特性(遗传)的影响更大, 而受温度的影响可能较小, 造成这两个月林分土壤温度虽然偏低, 但细根生产量特别大.与温度相比, 本研究中水分对细根生产的影响较小, 这可能是因为试验地所在地区雨热同期, 常年降水量和土壤水分较为充足, 使得细根生产量对水分因子的依赖性相对更弱.米槠人工林细根生产与温度的相关性较弱, 这可能与其是演替后期树种有关.有研究证实一些演替晚期的树种具有耐阴性(Kitajima, 1994 ; Walters & Reich, 1996 ; Khurana & Singh, 2006 ; Poorter & Rozendaal, 2008 ), 林冠更为稠密, 透光度减小, 因而对外界温度、光照的变化不敏感, 使得细根生长也受温度影响较小.Du和Fang (2014)对中国东北的白桦-中国山杨林和落叶松林研究发现, 演替早期的先锋树种白桦和中国山杨, 根呼吸对温度的敏感性要高于演替晚期的优势树种落叶松, 表明演替早期树种根系对温度的响应可能更敏感.这也说明演替早期树种的细根生产可能更易受外界气候变化的影响, 而演替晚期树种则可能由于长期处于较稳定的环境条件下, 细根生产对气候变化的响应较小.这从米槠人工林两年间的细根生产月动态模式比马尾松人工林更为稳定亦可反映出来.这可能也与演替晚期的林分更加郁闭, 对外界环境变化更不敏感, 而演替早期的林分更加稀疏, 受环境变化影响更大有关.同时, 我们还发现演替早期马尾松林分的细根物候与演替晚期米槠林分存在较大差异, 这可能与马尾松为阳性先锋树种, 林分稀疏, 土壤温度一般较米槠人工林的高有关(图3 ).较高的土壤温度使春季细根生产开始时间提前、而冬季结束时间延迟.Alvarez- Uria和Körner (2007)研究证实, 欧洲桦(Betula pubescens )根系显著生长所需的最低温度是6 ℃.Schenker等(2014)研究表明: 不考虑树种差异, 土壤温度低于7 ℃会使温带阔叶树的根系生物量明显 减少. ...
Relative importance of photosynthetic traits and allocation patterns as correlates of seedling shade tolerance of 13 tropical trees.
2
1994
... 不同演替阶段的树种生理和形态特征有所不同(Givnish, 1988 ; Messier, 1999 ; Ellis et al ., 2000 ), 细根特性也可能因所处演替阶段不同而存在差异.Paz (2003)研究了55种树种的根形态特征后发现, 演替晚期树种的根总体上要比演替早期树种的根更粗.Coll等(2008)的研究表明, 演替晚期树种的主根生物量所占比例更大.受生境条件等因素的影响, 不同演替阶段树种的细根生长速度可能也存在差异.在演替早期, 群落结构简单且立地条件相对较差, 植被更倾向于分配更多的生物量给根系(屠玉麟和杨军, 1995 ), 并通过迅速和广泛地占据土壤空间获取资源(Brassard et al ., 2009 ), 具有觅食的广布性; 而演替晚期群落的物种多样性较丰富且结构复杂, 土壤有机质和养分含量较高, 种间竞争激烈, 为了提高竞争力, 细根会迅速识别富养斑块并大量增生(Brassard et al ., 2009 ), 养分利用精细, 利用效率提高.Zangaro等(2012)对几个热带树种的研究发现, 细根生物量、细根直径随演替的进行而增加, 而根总长、比根长、根毛长度、根毛发生率、内生菌根菌侵染率等降低, 表明与演替后期阶段树种相比, 演替早期阶段的树种细根具有更强的养分获取能力, 生长速度可能更快.同时, 不同演替阶段的树种受栖息地环境的影响程度可能不同.如演替早期的热带树种生长受开阔栖息地的局限较大(Zangaro et al ., 2012 ), 而一些演替晚期的树种具有耐阴性(Kitajima, 1994 ; Walters & Reich, 1996 ; Khurana & Singh, 2006 ; Poorter & Rozendaal, 2008 ), 林冠更加稠密, 透光度更小.Du和Fang (2014)对中国东北的白桦(Betula platyphylla )-中国山杨(Populus davidiana )林和落叶松(Larix gmelinii )林的研究也曾发现, 演替早期的先锋树种白桦和中国山杨, 根呼吸对温度的敏感性要高于演替晚期的优势树种落叶松.此外, 不同演替阶段树种的物候情况在长期森林动态研究中十分重要, 且是预测未来气候变化环境下的森林变化所必须考虑的问题(Du & Fang, 2014 ).然而目前有关森林不同演替阶段的树种细根生产动态和细根物候方面差异的研究仍十分缺乏.分析研究不同演替阶段不同树种林分的细根生产量、季节动态及其所受环境因子影响的差异, 将有助于增进我们了解不同演替阶段的树种差异对细根生产可能造成的影响. ...
... Fukuzawa等(2007)对日本北部一个温带森林的研究证实: 温度是驱使细根生产量呈现季节性特征的重要因素.另有研究表明大气温度是影响细根生产的最重要环境因子(Tierney & Fahey, 2001 ).Steele等(1997)研究发现土壤温度与细根生产呈正相关.本研究中, 马尾松和米槠人工林的细根生产动态与温度因子存在正相关, 也说明了温度对细根生产有较大的影响.图4D 中, 有两点偏离较明显(2013年6月和2013年11月), 6月和11月应该都是米槠人工林细根生产量的峰值(夏季和秋季), 可能因为受本身的树种特性(遗传)的影响更大, 而受温度的影响可能较小, 造成这两个月林分土壤温度虽然偏低, 但细根生产量特别大.与温度相比, 本研究中水分对细根生产的影响较小, 这可能是因为试验地所在地区雨热同期, 常年降水量和土壤水分较为充足, 使得细根生产量对水分因子的依赖性相对更弱.米槠人工林细根生产与温度的相关性较弱, 这可能与其是演替后期树种有关.有研究证实一些演替晚期的树种具有耐阴性(Kitajima, 1994 ; Walters & Reich, 1996 ; Khurana & Singh, 2006 ; Poorter & Rozendaal, 2008 ), 林冠更为稠密, 透光度减小, 因而对外界温度、光照的变化不敏感, 使得细根生长也受温度影响较小.Du和Fang (2014)对中国东北的白桦-中国山杨林和落叶松林研究发现, 演替早期的先锋树种白桦和中国山杨, 根呼吸对温度的敏感性要高于演替晚期的优势树种落叶松, 表明演替早期树种根系对温度的响应可能更敏感.这也说明演替早期树种的细根生产可能更易受外界气候变化的影响, 而演替晚期树种则可能由于长期处于较稳定的环境条件下, 细根生产对气候变化的响应较小.这从米槠人工林两年间的细根生产月动态模式比马尾松人工林更为稳定亦可反映出来.这可能也与演替晚期的林分更加郁闭, 对外界环境变化更不敏感, 而演替早期的林分更加稀疏, 受环境变化影响更大有关.同时, 我们还发现演替早期马尾松林分的细根物候与演替晚期米槠林分存在较大差异, 这可能与马尾松为阳性先锋树种, 林分稀疏, 土壤温度一般较米槠人工林的高有关(图3 ).较高的土壤温度使春季细根生产开始时间提前、而冬季结束时间延迟.Alvarez- Uria和Körner (2007)研究证实, 欧洲桦(Betula pubescens )根系显著生长所需的最低温度是6 ℃.Schenker等(2014)研究表明: 不考虑树种差异, 土壤温度低于7 ℃会使温带阔叶树的根系生物量明显 减少. ...
亚热带米槠群落生态恢复中种间竞争研究
1
2006
... 马尾松(Pinus massoniana )是我国南方重要的速生用材树种, 为亚热带群落演替早期优势树种(王伯荪和彭少麟, 1985 ).米槠(Castanopsis carlesii )是我国中亚热带地区最典型的地带性植被常绿阔叶林的代表性树种, 为演替后期顶极种(刘浩凤, 2006 ).本文通过对马尾松和米槠两个树种的人工林进行为期两年(2013年1月至2014年12月)的微根管观测, 对两种人工林细根的生产量动态及其径级和土层的分布进行研究, 并分析了两年的细根生产量动态变化与气温、降水、土壤10 cm温度和土壤10 cm含水率等环境因子间的相关性, 进一步明确了不同演替阶段树种的细根生产量变化与环境因子间的相关关系, 为预测全球气候变化对中亚热带不同演替阶段树种的根系生产力影响提供了基础数据.本研究试图解决的科学问题为: 演替早期的马尾松林分和演替晚期的米槠林分根系生产量及其动态差异、季节分布如何? 这两种不同演替阶段树种林分根系生产量动态与各环境因子间的相关关系有何差异?两种不同演替阶段树种林分的根系生产量径级和土层分配情况怎样? ...
亚热带米槠群落生态恢复中种间竞争研究
1
2006
... 马尾松(Pinus massoniana )是我国南方重要的速生用材树种, 为亚热带群落演替早期优势树种(王伯荪和彭少麟, 1985 ).米槠(Castanopsis carlesii )是我国中亚热带地区最典型的地带性植被常绿阔叶林的代表性树种, 为演替后期顶极种(刘浩凤, 2006 ).本文通过对马尾松和米槠两个树种的人工林进行为期两年(2013年1月至2014年12月)的微根管观测, 对两种人工林细根的生产量动态及其径级和土层的分布进行研究, 并分析了两年的细根生产量动态变化与气温、降水、土壤10 cm温度和土壤10 cm含水率等环境因子间的相关性, 进一步明确了不同演替阶段树种的细根生产量变化与环境因子间的相关关系, 为预测全球气候变化对中亚热带不同演替阶段树种的根系生产力影响提供了基础数据.本研究试图解决的科学问题为: 演替早期的马尾松林分和演替晚期的米槠林分根系生产量及其动态差异、季节分布如何? 这两种不同演替阶段树种林分根系生产量动态与各环境因子间的相关关系有何差异?两种不同演替阶段树种林分的根系生产量径级和土层分配情况怎样? ...
Rhizoeconomics: Carbon costs of phosphorus acquisition.
1
2005
... 本研究中, 马尾松人工林在观测期2年间的细根年根长生产量均明显高于米槠人工林, 不同演替阶段的树种在遗传特性方面存在差异, 可能是造成两种人工林细根生产量相差较大的一个主要原因.不同演替阶段的树种生长速率差异较大(Zangaro et al ., 2012 ), 已有研究表明, 演替早期树种比演替晚期树种具有更高的光合作用能力(Reich et al ., 1998 ; Poorter & Rozendaal, 2008 ), 而这种更高的光合作用能力又将提高可输出为根生长所用的光合产物的数量(Nielsen et al ., 1998 ; Lynch et al ., 2005 ), 进而使得演替早期树种的根生长速率更快.另外, 马尾松为演替早期树种, 对生境适应能力较强, 因而细根的生产量更高; 而米槠属于演替后期树种, 其细根的生产量也较低.Zangaro等(2012)对几个热带树种的研究也发现, 细根生物量、细根直径随演替进行而增加, 而根总长、比根长、根毛长度、根毛发生率、内生菌根菌侵染率等降低, 表明与演替后期阶段树种相比, 演替早期阶段的树种细根可能具有更强的养分获取能力.虽然本研究区域在亚热带, 不同演替阶段树种的细根生产也可能呈现相似的差异特征.两种人工林观测第一年细根生产量均比第二年明显增大, 可能与细根生产的年际波动有关, 也可能与微根管安装过程对细根生产的刺激作用有关.有研究表明, 微根管安装切断了原有根系, 导致初期根系的大量增生与分枝, 这种刺激作用可能随时间的推移而减弱(Joslin & Wolfe, 1997 ). ...
Functional ecology of advance regeneration in relation to light in boreal forests.
1
1999
... 不同演替阶段的树种生理和形态特征有所不同(Givnish, 1988 ; Messier, 1999 ; Ellis et al ., 2000 ), 细根特性也可能因所处演替阶段不同而存在差异.Paz (2003)研究了55种树种的根形态特征后发现, 演替晚期树种的根总体上要比演替早期树种的根更粗.Coll等(2008)的研究表明, 演替晚期树种的主根生物量所占比例更大.受生境条件等因素的影响, 不同演替阶段树种的细根生长速度可能也存在差异.在演替早期, 群落结构简单且立地条件相对较差, 植被更倾向于分配更多的生物量给根系(屠玉麟和杨军, 1995 ), 并通过迅速和广泛地占据土壤空间获取资源(Brassard et al ., 2009 ), 具有觅食的广布性; 而演替晚期群落的物种多样性较丰富且结构复杂, 土壤有机质和养分含量较高, 种间竞争激烈, 为了提高竞争力, 细根会迅速识别富养斑块并大量增生(Brassard et al ., 2009 ), 养分利用精细, 利用效率提高.Zangaro等(2012)对几个热带树种的研究发现, 细根生物量、细根直径随演替的进行而增加, 而根总长、比根长、根毛长度、根毛发生率、内生菌根菌侵染率等降低, 表明与演替后期阶段树种相比, 演替早期阶段的树种细根具有更强的养分获取能力, 生长速度可能更快.同时, 不同演替阶段的树种受栖息地环境的影响程度可能不同.如演替早期的热带树种生长受开阔栖息地的局限较大(Zangaro et al ., 2012 ), 而一些演替晚期的树种具有耐阴性(Kitajima, 1994 ; Walters & Reich, 1996 ; Khurana & Singh, 2006 ; Poorter & Rozendaal, 2008 ), 林冠更加稠密, 透光度更小.Du和Fang (2014)对中国东北的白桦(Betula platyphylla )-中国山杨(Populus davidiana )林和落叶松(Larix gmelinii )林的研究也曾发现, 演替早期的先锋树种白桦和中国山杨, 根呼吸对温度的敏感性要高于演替晚期的优势树种落叶松.此外, 不同演替阶段树种的物候情况在长期森林动态研究中十分重要, 且是预测未来气候变化环境下的森林变化所必须考虑的问题(Du & Fang, 2014 ).然而目前有关森林不同演替阶段的树种细根生产动态和细根物候方面差异的研究仍十分缺乏.分析研究不同演替阶段不同树种林分的细根生产量、季节动态及其所受环境因子影响的差异, 将有助于增进我们了解不同演替阶段的树种差异对细根生产可能造成的影响. ...
Effects of phosphorus availability and vesicular-arbuscular mycorrhizas on the carbon budget of common bean (Phaseolus vulgaris).
1
1998
... 本研究中, 马尾松人工林在观测期2年间的细根年根长生产量均明显高于米槠人工林, 不同演替阶段的树种在遗传特性方面存在差异, 可能是造成两种人工林细根生产量相差较大的一个主要原因.不同演替阶段的树种生长速率差异较大(Zangaro et al ., 2012 ), 已有研究表明, 演替早期树种比演替晚期树种具有更高的光合作用能力(Reich et al ., 1998 ; Poorter & Rozendaal, 2008 ), 而这种更高的光合作用能力又将提高可输出为根生长所用的光合产物的数量(Nielsen et al ., 1998 ; Lynch et al ., 2005 ), 进而使得演替早期树种的根生长速率更快.另外, 马尾松为演替早期树种, 对生境适应能力较强, 因而细根的生产量更高; 而米槠属于演替后期树种, 其细根的生产量也较低.Zangaro等(2012)对几个热带树种的研究也发现, 细根生物量、细根直径随演替进行而增加, 而根总长、比根长、根毛长度、根毛发生率、内生菌根菌侵染率等降低, 表明与演替后期阶段树种相比, 演替早期阶段的树种细根可能具有更强的养分获取能力.虽然本研究区域在亚热带, 不同演替阶段树种的细根生产也可能呈现相似的差异特征.两种人工林观测第一年细根生产量均比第二年明显增大, 可能与细根生产的年际波动有关, 也可能与微根管安装过程对细根生产的刺激作用有关.有研究表明, 微根管安装切断了原有根系, 导致初期根系的大量增生与分枝, 这种刺激作用可能随时间的推移而减弱(Joslin & Wolfe, 1997 ). ...
An evaluation of the minirhizotron technique for estimating root distribution in potatoes.
2
1991
... 本研究中, 马尾松人工林的表层细根生产量占观测期总细根生产量的比例最大, Fukuzawa等(2007)对日本北部一处主要被Quercus crispula 和Sasa senanensis 覆盖的寒温带森林的研究也显示表层土壤(0-15 cm)细根生产量所占比例最高.史建伟等(2007)对落叶松和水曲柳(Fraxinus mandshurica )的研究表明, 两种林分的细根表层年生产量均最高, 底层最低.然而本研究中两种人工林细根生产量并未随着土层加深而递减, 马尾松人工林20-30 cm土层细根生产量所占比例也大于10-20 cm土层; 米槠人工林的底层细根生产量所占比例甚至略高于表层, 并超过了中间两个土层细根生产量所占的比例.可能是演替后期米槠林分的根系在土壤资源利用策略上竞争更激烈, 导致根系更加充分地利用底层土壤资源, 使得底层根系生产量略高于表层.而演替早期马尾松则是表层0-10 cm所占比例最高, 这可能与其根系对土壤资源的竞争强度较小, 而更集中在表层有关.王韦韦等(2015)采用土芯法对马尾松和米槠人工林的研究表明, 0-10 cm细根生物量明显多于其他土层.而本研究中细根生产并未有类似特征, 这可能与研究方法不同有关.与土芯法等比较, 许多研究均证明微根管法低估了表土层细根生产量, 而夸大了深土层细根生产量(Bragg et al ., 1983 ; Upchurch & Ritchie, 1983 ; Parker et al ., 1991 ; de Ruijter et al ., 1996 ).表土层根系生长的低估可能与管-土界面缝隙或压实(Dixon et al ., 1994 )和土壤温度上升(Hansson et al ., 1995 )有关; 而底层土壤根系生长的高估则可能与根系倾向于沿管壁生长有关(Parker et al ., 1991 ; Heeraman & Juma, 1993 ).我们的其他微根管研究也得出相似的结论, 如黄锦学等(2012)的研究中, 细根生产均表现为20-40 cm土层的细根生产量所占比重最大; 胡双成(2015)的研究中也是20-40 cm土层细根所占生产量的比例最大, 可能也是由于微根管法低估表层细根生产造成的. 我们未能检测出两个林分的垂直分布差异, 主要还是因为细根变异大、样本量不足. ...
... )有关; 而底层土壤根系生长的高估则可能与根系倾向于沿管壁生长有关(Parker et al ., 1991 ; Heeraman & Juma, 1993 ).我们的其他微根管研究也得出相似的结论, 如黄锦学等(2012)的研究中, 细根生产均表现为20-40 cm土层的细根生产量所占比重最大; 胡双成(2015)的研究中也是20-40 cm土层细根所占生产量的比例最大, 可能也是由于微根管法低估表层细根生产造成的. 我们未能检测出两个林分的垂直分布差异, 主要还是因为细根变异大、样本量不足. ...
Root/shoot allocation and root architecture in seedlings: Variation among forest sites, microhabitats, and ecological groups.
0
2003
Leaf size and leaf display of thirty-eight tropical tree species.
3
2008
... 不同演替阶段的树种生理和形态特征有所不同(Givnish, 1988 ; Messier, 1999 ; Ellis et al ., 2000 ), 细根特性也可能因所处演替阶段不同而存在差异.Paz (2003)研究了55种树种的根形态特征后发现, 演替晚期树种的根总体上要比演替早期树种的根更粗.Coll等(2008)的研究表明, 演替晚期树种的主根生物量所占比例更大.受生境条件等因素的影响, 不同演替阶段树种的细根生长速度可能也存在差异.在演替早期, 群落结构简单且立地条件相对较差, 植被更倾向于分配更多的生物量给根系(屠玉麟和杨军, 1995 ), 并通过迅速和广泛地占据土壤空间获取资源(Brassard et al ., 2009 ), 具有觅食的广布性; 而演替晚期群落的物种多样性较丰富且结构复杂, 土壤有机质和养分含量较高, 种间竞争激烈, 为了提高竞争力, 细根会迅速识别富养斑块并大量增生(Brassard et al ., 2009 ), 养分利用精细, 利用效率提高.Zangaro等(2012)对几个热带树种的研究发现, 细根生物量、细根直径随演替的进行而增加, 而根总长、比根长、根毛长度、根毛发生率、内生菌根菌侵染率等降低, 表明与演替后期阶段树种相比, 演替早期阶段的树种细根具有更强的养分获取能力, 生长速度可能更快.同时, 不同演替阶段的树种受栖息地环境的影响程度可能不同.如演替早期的热带树种生长受开阔栖息地的局限较大(Zangaro et al ., 2012 ), 而一些演替晚期的树种具有耐阴性(Kitajima, 1994 ; Walters & Reich, 1996 ; Khurana & Singh, 2006 ; Poorter & Rozendaal, 2008 ), 林冠更加稠密, 透光度更小.Du和Fang (2014)对中国东北的白桦(Betula platyphylla )-中国山杨(Populus davidiana )林和落叶松(Larix gmelinii )林的研究也曾发现, 演替早期的先锋树种白桦和中国山杨, 根呼吸对温度的敏感性要高于演替晚期的优势树种落叶松.此外, 不同演替阶段树种的物候情况在长期森林动态研究中十分重要, 且是预测未来气候变化环境下的森林变化所必须考虑的问题(Du & Fang, 2014 ).然而目前有关森林不同演替阶段的树种细根生产动态和细根物候方面差异的研究仍十分缺乏.分析研究不同演替阶段不同树种林分的细根生产量、季节动态及其所受环境因子影响的差异, 将有助于增进我们了解不同演替阶段的树种差异对细根生产可能造成的影响. ...
... 本研究中, 马尾松人工林在观测期2年间的细根年根长生产量均明显高于米槠人工林, 不同演替阶段的树种在遗传特性方面存在差异, 可能是造成两种人工林细根生产量相差较大的一个主要原因.不同演替阶段的树种生长速率差异较大(Zangaro et al ., 2012 ), 已有研究表明, 演替早期树种比演替晚期树种具有更高的光合作用能力(Reich et al ., 1998 ; Poorter & Rozendaal, 2008 ), 而这种更高的光合作用能力又将提高可输出为根生长所用的光合产物的数量(Nielsen et al ., 1998 ; Lynch et al ., 2005 ), 进而使得演替早期树种的根生长速率更快.另外, 马尾松为演替早期树种, 对生境适应能力较强, 因而细根的生产量更高; 而米槠属于演替后期树种, 其细根的生产量也较低.Zangaro等(2012)对几个热带树种的研究也发现, 细根生物量、细根直径随演替进行而增加, 而根总长、比根长、根毛长度、根毛发生率、内生菌根菌侵染率等降低, 表明与演替后期阶段树种相比, 演替早期阶段的树种细根可能具有更强的养分获取能力.虽然本研究区域在亚热带, 不同演替阶段树种的细根生产也可能呈现相似的差异特征.两种人工林观测第一年细根生产量均比第二年明显增大, 可能与细根生产的年际波动有关, 也可能与微根管安装过程对细根生产的刺激作用有关.有研究表明, 微根管安装切断了原有根系, 导致初期根系的大量增生与分枝, 这种刺激作用可能随时间的推移而减弱(Joslin & Wolfe, 1997 ). ...
... Fukuzawa等(2007)对日本北部一个温带森林的研究证实: 温度是驱使细根生产量呈现季节性特征的重要因素.另有研究表明大气温度是影响细根生产的最重要环境因子(Tierney & Fahey, 2001 ).Steele等(1997)研究发现土壤温度与细根生产呈正相关.本研究中, 马尾松和米槠人工林的细根生产动态与温度因子存在正相关, 也说明了温度对细根生产有较大的影响.图4D 中, 有两点偏离较明显(2013年6月和2013年11月), 6月和11月应该都是米槠人工林细根生产量的峰值(夏季和秋季), 可能因为受本身的树种特性(遗传)的影响更大, 而受温度的影响可能较小, 造成这两个月林分土壤温度虽然偏低, 但细根生产量特别大.与温度相比, 本研究中水分对细根生产的影响较小, 这可能是因为试验地所在地区雨热同期, 常年降水量和土壤水分较为充足, 使得细根生产量对水分因子的依赖性相对更弱.米槠人工林细根生产与温度的相关性较弱, 这可能与其是演替后期树种有关.有研究证实一些演替晚期的树种具有耐阴性(Kitajima, 1994 ; Walters & Reich, 1996 ; Khurana & Singh, 2006 ; Poorter & Rozendaal, 2008 ), 林冠更为稠密, 透光度减小, 因而对外界温度、光照的变化不敏感, 使得细根生长也受温度影响较小.Du和Fang (2014)对中国东北的白桦-中国山杨林和落叶松林研究发现, 演替早期的先锋树种白桦和中国山杨, 根呼吸对温度的敏感性要高于演替晚期的优势树种落叶松, 表明演替早期树种根系对温度的响应可能更敏感.这也说明演替早期树种的细根生产可能更易受外界气候变化的影响, 而演替晚期树种则可能由于长期处于较稳定的环境条件下, 细根生产对气候变化的响应较小.这从米槠人工林两年间的细根生产月动态模式比马尾松人工林更为稳定亦可反映出来.这可能也与演替晚期的林分更加郁闭, 对外界环境变化更不敏感, 而演替早期的林分更加稀疏, 受环境变化影响更大有关.同时, 我们还发现演替早期马尾松林分的细根物候与演替晚期米槠林分存在较大差异, 这可能与马尾松为阳性先锋树种, 林分稀疏, 土壤温度一般较米槠人工林的高有关(图3 ).较高的土壤温度使春季细根生产开始时间提前、而冬季结束时间延迟.Alvarez- Uria和Körner (2007)研究证实, 欧洲桦(Betula pubescens )根系显著生长所需的最低温度是6 ℃.Schenker等(2014)研究表明: 不考虑树种差异, 土壤温度低于7 ℃会使温带阔叶树的根系生物量明显 减少. ...
Close association of RGR, leaf and root morphology, seed mass and shade tolerance in seedlings of nine boreal tree species grown in high and low light.
1
1998
... 本研究中, 马尾松人工林在观测期2年间的细根年根长生产量均明显高于米槠人工林, 不同演替阶段的树种在遗传特性方面存在差异, 可能是造成两种人工林细根生产量相差较大的一个主要原因.不同演替阶段的树种生长速率差异较大(Zangaro et al ., 2012 ), 已有研究表明, 演替早期树种比演替晚期树种具有更高的光合作用能力(Reich et al ., 1998 ; Poorter & Rozendaal, 2008 ), 而这种更高的光合作用能力又将提高可输出为根生长所用的光合产物的数量(Nielsen et al ., 1998 ; Lynch et al ., 2005 ), 进而使得演替早期树种的根生长速率更快.另外, 马尾松为演替早期树种, 对生境适应能力较强, 因而细根的生产量更高; 而米槠属于演替后期树种, 其细根的生产量也较低.Zangaro等(2012)对几个热带树种的研究也发现, 细根生物量、细根直径随演替进行而增加, 而根总长、比根长、根毛长度、根毛发生率、内生菌根菌侵染率等降低, 表明与演替后期阶段树种相比, 演替早期阶段的树种细根可能具有更强的养分获取能力.虽然本研究区域在亚热带, 不同演替阶段树种的细根生产也可能呈现相似的差异特征.两种人工林观测第一年细根生产量均比第二年明显增大, 可能与细根生产的年际波动有关, 也可能与微根管安装过程对细根生产的刺激作用有关.有研究表明, 微根管安装切断了原有根系, 导致初期根系的大量增生与分枝, 这种刺激作用可能随时间的推移而减弱(Joslin & Wolfe, 1997 ). ...
Physiological minimum temperatures for root growth in seven common European broad-leaved tree species.
0
2014
落叶松和水曲柳人工林细根生长、死亡和周转.
0
2007
落叶松和水曲柳人工林细根生长、死亡和周转.
0
2007
Root mass, net primary production and turnover in aspen, jack pine and black spruce forests in Saskatchewan and Manitoba, Canada.
0
1997
Evaluating minirhizotron estimates of fine root longevity and production in the forest floor of a temperate broadleaf forest.
1
2001
... Fukuzawa等(2007)对日本北部一个温带森林的研究证实: 温度是驱使细根生产量呈现季节性特征的重要因素.另有研究表明大气温度是影响细根生产的最重要环境因子(Tierney & Fahey, 2001 ).Steele等(1997)研究发现土壤温度与细根生产呈正相关.本研究中, 马尾松和米槠人工林的细根生产动态与温度因子存在正相关, 也说明了温度对细根生产有较大的影响.图4D 中, 有两点偏离较明显(2013年6月和2013年11月), 6月和11月应该都是米槠人工林细根生产量的峰值(夏季和秋季), 可能因为受本身的树种特性(遗传)的影响更大, 而受温度的影响可能较小, 造成这两个月林分土壤温度虽然偏低, 但细根生产量特别大.与温度相比, 本研究中水分对细根生产的影响较小, 这可能是因为试验地所在地区雨热同期, 常年降水量和土壤水分较为充足, 使得细根生产量对水分因子的依赖性相对更弱.米槠人工林细根生产与温度的相关性较弱, 这可能与其是演替后期树种有关.有研究证实一些演替晚期的树种具有耐阴性(Kitajima, 1994 ; Walters & Reich, 1996 ; Khurana & Singh, 2006 ; Poorter & Rozendaal, 2008 ), 林冠更为稠密, 透光度减小, 因而对外界温度、光照的变化不敏感, 使得细根生长也受温度影响较小.Du和Fang (2014)对中国东北的白桦-中国山杨林和落叶松林研究发现, 演替早期的先锋树种白桦和中国山杨, 根呼吸对温度的敏感性要高于演替晚期的优势树种落叶松, 表明演替早期树种根系对温度的响应可能更敏感.这也说明演替早期树种的细根生产可能更易受外界气候变化的影响, 而演替晚期树种则可能由于长期处于较稳定的环境条件下, 细根生产对气候变化的响应较小.这从米槠人工林两年间的细根生产月动态模式比马尾松人工林更为稳定亦可反映出来.这可能也与演替晚期的林分更加郁闭, 对外界环境变化更不敏感, 而演替早期的林分更加稀疏, 受环境变化影响更大有关.同时, 我们还发现演替早期马尾松林分的细根物候与演替晚期米槠林分存在较大差异, 这可能与马尾松为阳性先锋树种, 林分稀疏, 土壤温度一般较米槠人工林的高有关(图3 ).较高的土壤温度使春季细根生产开始时间提前、而冬季结束时间延迟.Alvarez- Uria和Körner (2007)研究证实, 欧洲桦(Betula pubescens )根系显著生长所需的最低温度是6 ℃.Schenker等(2014)研究表明: 不考虑树种差异, 土壤温度低于7 ℃会使温带阔叶树的根系生物量明显 减少. ...
贵州中部喀斯特灌丛群落生物量研究.
1
1995
... 不同演替阶段的树种生理和形态特征有所不同(Givnish, 1988 ; Messier, 1999 ; Ellis et al ., 2000 ), 细根特性也可能因所处演替阶段不同而存在差异.Paz (2003)研究了55种树种的根形态特征后发现, 演替晚期树种的根总体上要比演替早期树种的根更粗.Coll等(2008)的研究表明, 演替晚期树种的主根生物量所占比例更大.受生境条件等因素的影响, 不同演替阶段树种的细根生长速度可能也存在差异.在演替早期, 群落结构简单且立地条件相对较差, 植被更倾向于分配更多的生物量给根系(屠玉麟和杨军, 1995 ), 并通过迅速和广泛地占据土壤空间获取资源(Brassard et al ., 2009 ), 具有觅食的广布性; 而演替晚期群落的物种多样性较丰富且结构复杂, 土壤有机质和养分含量较高, 种间竞争激烈, 为了提高竞争力, 细根会迅速识别富养斑块并大量增生(Brassard et al ., 2009 ), 养分利用精细, 利用效率提高.Zangaro等(2012)对几个热带树种的研究发现, 细根生物量、细根直径随演替的进行而增加, 而根总长、比根长、根毛长度、根毛发生率、内生菌根菌侵染率等降低, 表明与演替后期阶段树种相比, 演替早期阶段的树种细根具有更强的养分获取能力, 生长速度可能更快.同时, 不同演替阶段的树种受栖息地环境的影响程度可能不同.如演替早期的热带树种生长受开阔栖息地的局限较大(Zangaro et al ., 2012 ), 而一些演替晚期的树种具有耐阴性(Kitajima, 1994 ; Walters & Reich, 1996 ; Khurana & Singh, 2006 ; Poorter & Rozendaal, 2008 ), 林冠更加稠密, 透光度更小.Du和Fang (2014)对中国东北的白桦(Betula platyphylla )-中国山杨(Populus davidiana )林和落叶松(Larix gmelinii )林的研究也曾发现, 演替早期的先锋树种白桦和中国山杨, 根呼吸对温度的敏感性要高于演替晚期的优势树种落叶松.此外, 不同演替阶段树种的物候情况在长期森林动态研究中十分重要, 且是预测未来气候变化环境下的森林变化所必须考虑的问题(Du & Fang, 2014 ).然而目前有关森林不同演替阶段的树种细根生产动态和细根物候方面差异的研究仍十分缺乏.分析研究不同演替阶段不同树种林分的细根生产量、季节动态及其所受环境因子影响的差异, 将有助于增进我们了解不同演替阶段的树种差异对细根生产可能造成的影响. ...
贵州中部喀斯特灌丛群落生物量研究.
1
1995
... 不同演替阶段的树种生理和形态特征有所不同(Givnish, 1988 ; Messier, 1999 ; Ellis et al ., 2000 ), 细根特性也可能因所处演替阶段不同而存在差异.Paz (2003)研究了55种树种的根形态特征后发现, 演替晚期树种的根总体上要比演替早期树种的根更粗.Coll等(2008)的研究表明, 演替晚期树种的主根生物量所占比例更大.受生境条件等因素的影响, 不同演替阶段树种的细根生长速度可能也存在差异.在演替早期, 群落结构简单且立地条件相对较差, 植被更倾向于分配更多的生物量给根系(屠玉麟和杨军, 1995 ), 并通过迅速和广泛地占据土壤空间获取资源(Brassard et al ., 2009 ), 具有觅食的广布性; 而演替晚期群落的物种多样性较丰富且结构复杂, 土壤有机质和养分含量较高, 种间竞争激烈, 为了提高竞争力, 细根会迅速识别富养斑块并大量增生(Brassard et al ., 2009 ), 养分利用精细, 利用效率提高.Zangaro等(2012)对几个热带树种的研究发现, 细根生物量、细根直径随演替的进行而增加, 而根总长、比根长、根毛长度、根毛发生率、内生菌根菌侵染率等降低, 表明与演替后期阶段树种相比, 演替早期阶段的树种细根具有更强的养分获取能力, 生长速度可能更快.同时, 不同演替阶段的树种受栖息地环境的影响程度可能不同.如演替早期的热带树种生长受开阔栖息地的局限较大(Zangaro et al ., 2012 ), 而一些演替晚期的树种具有耐阴性(Kitajima, 1994 ; Walters & Reich, 1996 ; Khurana & Singh, 2006 ; Poorter & Rozendaal, 2008 ), 林冠更加稠密, 透光度更小.Du和Fang (2014)对中国东北的白桦(Betula platyphylla )-中国山杨(Populus davidiana )林和落叶松(Larix gmelinii )林的研究也曾发现, 演替早期的先锋树种白桦和中国山杨, 根呼吸对温度的敏感性要高于演替晚期的优势树种落叶松.此外, 不同演替阶段树种的物候情况在长期森林动态研究中十分重要, 且是预测未来气候变化环境下的森林变化所必须考虑的问题(Du & Fang, 2014 ).然而目前有关森林不同演替阶段的树种细根生产动态和细根物候方面差异的研究仍十分缺乏.分析研究不同演替阶段不同树种林分的细根生产量、季节动态及其所受环境因子影响的差异, 将有助于增进我们了解不同演替阶段的树种差异对细根生产可能造成的影响. ...
Root observations using a video recording system in mini-rhizotrons.
1
1983
... 本研究中, 马尾松人工林的表层细根生产量占观测期总细根生产量的比例最大, Fukuzawa等(2007)对日本北部一处主要被Quercus crispula 和Sasa senanensis 覆盖的寒温带森林的研究也显示表层土壤(0-15 cm)细根生产量所占比例最高.史建伟等(2007)对落叶松和水曲柳(Fraxinus mandshurica )的研究表明, 两种林分的细根表层年生产量均最高, 底层最低.然而本研究中两种人工林细根生产量并未随着土层加深而递减, 马尾松人工林20-30 cm土层细根生产量所占比例也大于10-20 cm土层; 米槠人工林的底层细根生产量所占比例甚至略高于表层, 并超过了中间两个土层细根生产量所占的比例.可能是演替后期米槠林分的根系在土壤资源利用策略上竞争更激烈, 导致根系更加充分地利用底层土壤资源, 使得底层根系生产量略高于表层.而演替早期马尾松则是表层0-10 cm所占比例最高, 这可能与其根系对土壤资源的竞争强度较小, 而更集中在表层有关.王韦韦等(2015)采用土芯法对马尾松和米槠人工林的研究表明, 0-10 cm细根生物量明显多于其他土层.而本研究中细根生产并未有类似特征, 这可能与研究方法不同有关.与土芯法等比较, 许多研究均证明微根管法低估了表土层细根生产量, 而夸大了深土层细根生产量(Bragg et al ., 1983 ; Upchurch & Ritchie, 1983 ; Parker et al ., 1991 ; de Ruijter et al ., 1996 ).表土层根系生长的低估可能与管-土界面缝隙或压实(Dixon et al ., 1994 )和土壤温度上升(Hansson et al ., 1995 )有关; 而底层土壤根系生长的高估则可能与根系倾向于沿管壁生长有关(Parker et al ., 1991 ; Heeraman & Juma, 1993 ).我们的其他微根管研究也得出相似的结论, 如黄锦学等(2012)的研究中, 细根生产均表现为20-40 cm土层的细根生产量所占比重最大; 胡双成(2015)的研究中也是20-40 cm土层细根所占生产量的比例最大, 可能也是由于微根管法低估表层细根生产造成的. 我们未能检测出两个林分的垂直分布差异, 主要还是因为细根变异大、样本量不足. ...
Are shade tolerance, survival, and growth linked? Low light and nitrogen effects on hardwood seedlings.
2
1996
... 不同演替阶段的树种生理和形态特征有所不同(Givnish, 1988 ; Messier, 1999 ; Ellis et al ., 2000 ), 细根特性也可能因所处演替阶段不同而存在差异.Paz (2003)研究了55种树种的根形态特征后发现, 演替晚期树种的根总体上要比演替早期树种的根更粗.Coll等(2008)的研究表明, 演替晚期树种的主根生物量所占比例更大.受生境条件等因素的影响, 不同演替阶段树种的细根生长速度可能也存在差异.在演替早期, 群落结构简单且立地条件相对较差, 植被更倾向于分配更多的生物量给根系(屠玉麟和杨军, 1995 ), 并通过迅速和广泛地占据土壤空间获取资源(Brassard et al ., 2009 ), 具有觅食的广布性; 而演替晚期群落的物种多样性较丰富且结构复杂, 土壤有机质和养分含量较高, 种间竞争激烈, 为了提高竞争力, 细根会迅速识别富养斑块并大量增生(Brassard et al ., 2009 ), 养分利用精细, 利用效率提高.Zangaro等(2012)对几个热带树种的研究发现, 细根生物量、细根直径随演替的进行而增加, 而根总长、比根长、根毛长度、根毛发生率、内生菌根菌侵染率等降低, 表明与演替后期阶段树种相比, 演替早期阶段的树种细根具有更强的养分获取能力, 生长速度可能更快.同时, 不同演替阶段的树种受栖息地环境的影响程度可能不同.如演替早期的热带树种生长受开阔栖息地的局限较大(Zangaro et al ., 2012 ), 而一些演替晚期的树种具有耐阴性(Kitajima, 1994 ; Walters & Reich, 1996 ; Khurana & Singh, 2006 ; Poorter & Rozendaal, 2008 ), 林冠更加稠密, 透光度更小.Du和Fang (2014)对中国东北的白桦(Betula platyphylla )-中国山杨(Populus davidiana )林和落叶松(Larix gmelinii )林的研究也曾发现, 演替早期的先锋树种白桦和中国山杨, 根呼吸对温度的敏感性要高于演替晚期的优势树种落叶松.此外, 不同演替阶段树种的物候情况在长期森林动态研究中十分重要, 且是预测未来气候变化环境下的森林变化所必须考虑的问题(Du & Fang, 2014 ).然而目前有关森林不同演替阶段的树种细根生产动态和细根物候方面差异的研究仍十分缺乏.分析研究不同演替阶段不同树种林分的细根生产量、季节动态及其所受环境因子影响的差异, 将有助于增进我们了解不同演替阶段的树种差异对细根生产可能造成的影响. ...
... Fukuzawa等(2007)对日本北部一个温带森林的研究证实: 温度是驱使细根生产量呈现季节性特征的重要因素.另有研究表明大气温度是影响细根生产的最重要环境因子(Tierney & Fahey, 2001 ).Steele等(1997)研究发现土壤温度与细根生产呈正相关.本研究中, 马尾松和米槠人工林的细根生产动态与温度因子存在正相关, 也说明了温度对细根生产有较大的影响.图4D 中, 有两点偏离较明显(2013年6月和2013年11月), 6月和11月应该都是米槠人工林细根生产量的峰值(夏季和秋季), 可能因为受本身的树种特性(遗传)的影响更大, 而受温度的影响可能较小, 造成这两个月林分土壤温度虽然偏低, 但细根生产量特别大.与温度相比, 本研究中水分对细根生产的影响较小, 这可能是因为试验地所在地区雨热同期, 常年降水量和土壤水分较为充足, 使得细根生产量对水分因子的依赖性相对更弱.米槠人工林细根生产与温度的相关性较弱, 这可能与其是演替后期树种有关.有研究证实一些演替晚期的树种具有耐阴性(Kitajima, 1994 ; Walters & Reich, 1996 ; Khurana & Singh, 2006 ; Poorter & Rozendaal, 2008 ), 林冠更为稠密, 透光度减小, 因而对外界温度、光照的变化不敏感, 使得细根生长也受温度影响较小.Du和Fang (2014)对中国东北的白桦-中国山杨林和落叶松林研究发现, 演替早期的先锋树种白桦和中国山杨, 根呼吸对温度的敏感性要高于演替晚期的优势树种落叶松, 表明演替早期树种根系对温度的响应可能更敏感.这也说明演替早期树种的细根生产可能更易受外界气候变化的影响, 而演替晚期树种则可能由于长期处于较稳定的环境条件下, 细根生产对气候变化的响应较小.这从米槠人工林两年间的细根生产月动态模式比马尾松人工林更为稳定亦可反映出来.这可能也与演替晚期的林分更加郁闭, 对外界环境变化更不敏感, 而演替早期的林分更加稀疏, 受环境变化影响更大有关.同时, 我们还发现演替早期马尾松林分的细根物候与演替晚期米槠林分存在较大差异, 这可能与马尾松为阳性先锋树种, 林分稀疏, 土壤温度一般较米槠人工林的高有关(图3 ).较高的土壤温度使春季细根生产开始时间提前、而冬季结束时间延迟.Alvarez- Uria和Körner (2007)研究证实, 欧洲桦(Betula pubescens )根系显著生长所需的最低温度是6 ℃.Schenker等(2014)研究表明: 不考虑树种差异, 土壤温度低于7 ℃会使温带阔叶树的根系生物量明显 减少. ...
鼎湖山森林群落分析V. 群落演替的线性系统与预测
1
1985
... 马尾松(Pinus massoniana )是我国南方重要的速生用材树种, 为亚热带群落演替早期优势树种(王伯荪和彭少麟, 1985 ).米槠(Castanopsis carlesii )是我国中亚热带地区最典型的地带性植被常绿阔叶林的代表性树种, 为演替后期顶极种(刘浩凤, 2006 ).本文通过对马尾松和米槠两个树种的人工林进行为期两年(2013年1月至2014年12月)的微根管观测, 对两种人工林细根的生产量动态及其径级和土层的分布进行研究, 并分析了两年的细根生产量动态变化与气温、降水、土壤10 cm温度和土壤10 cm含水率等环境因子间的相关性, 进一步明确了不同演替阶段树种的细根生产量变化与环境因子间的相关关系, 为预测全球气候变化对中亚热带不同演替阶段树种的根系生产力影响提供了基础数据.本研究试图解决的科学问题为: 演替早期的马尾松林分和演替晚期的米槠林分根系生产量及其动态差异、季节分布如何? 这两种不同演替阶段树种林分根系生产量动态与各环境因子间的相关关系有何差异?两种不同演替阶段树种林分的根系生产量径级和土层分配情况怎样? ...
鼎湖山森林群落分析V. 群落演替的线性系统与预测
1
1985
... 马尾松(Pinus massoniana )是我国南方重要的速生用材树种, 为亚热带群落演替早期优势树种(王伯荪和彭少麟, 1985 ).米槠(Castanopsis carlesii )是我国中亚热带地区最典型的地带性植被常绿阔叶林的代表性树种, 为演替后期顶极种(刘浩凤, 2006 ).本文通过对马尾松和米槠两个树种的人工林进行为期两年(2013年1月至2014年12月)的微根管观测, 对两种人工林细根的生产量动态及其径级和土层的分布进行研究, 并分析了两年的细根生产量动态变化与气温、降水、土壤10 cm温度和土壤10 cm含水率等环境因子间的相关性, 进一步明确了不同演替阶段树种的细根生产量变化与环境因子间的相关关系, 为预测全球气候变化对中亚热带不同演替阶段树种的根系生产力影响提供了基础数据.本研究试图解决的科学问题为: 演替早期的马尾松林分和演替晚期的米槠林分根系生产量及其动态差异、季节分布如何? 这两种不同演替阶段树种林分根系生产量动态与各环境因子间的相关关系有何差异?两种不同演替阶段树种林分的根系生产量径级和土层分配情况怎样? ...
常绿阔叶林不同更新方式下土壤呼吸的动态及机制
2
2012
... 试验地位于福建省三明市格氏栲自然保护区(26.32° N, 117.60° E)和三明市金丝湾森林公园陈大林业采育场(26.32° N, 117.60° E)内, 其中研究林分马尾松人工林分布于三明市金丝湾森林公园陈大林业采育场内, 米槠人工林分布于三明格氏栲自然保护区内.试验地所在区域属于中亚热带季风气候, 年平均气温19.1 ℃, 年降水量1749 mm, 年蒸发量1585 mm, 相对湿度81%, 平均海拔300 m, 平均坡度25°-35°, 土壤以花岗岩发育的红壤和黄壤为主, 还分布有红黄壤、黄棕壤等, 多呈酸性, 土壤厚度超过1 m (王超, 2012 ).马尾松人工林是经1976年皆伐次生林后营造人工纯林形成; 林分密度为1500株·hm-2 , 平均树高18.3 m, 平均胸径18.3 cm; 林下植被覆盖度约为90%, 灌木较少, 主要以芒萁(Dicranopteris dichotoma )、芒(Miscanthus sinensis )和蕨等草本为主(王超, 2012 ); 林分表层(0-20 cm)的土壤容重为1.33 g·cm-3 , 土壤全N和全P含量分别为0.8 g·kg-1 和0.35 g·kg-1 .米槠人工林则是米槠次生林经皆伐、火烧, 再通过挖穴造林和幼林抚育形成; 林分密度为2042株·hm-2 , 平均树高14.2 m, 平均胸径16.6 cm; 林分内分布的植物主要以毛冬青(Ilex pubescens )、乌饭树(Vaccinium bracteatum )、薄叶山矾(Symplocos anomala )、桂北木姜子( Litsea subcoriacea )、山姜(Alpinia japonica )等为主, 草本以芒萁为主; 林分表层(0-20 cm)的土壤容重为1.18 g·cm-3 , 土壤全N和全P含量分别为1.02 g·kg-1 和0.37 g·kg-1 . ...
... )和蕨等草本为主(王超, 2012 ); 林分表层(0-20 cm)的土壤容重为1.33 g·cm-3 , 土壤全N和全P含量分别为0.8 g·kg-1 和0.35 g·kg-1 .米槠人工林则是米槠次生林经皆伐、火烧, 再通过挖穴造林和幼林抚育形成; 林分密度为2042株·hm-2 , 平均树高14.2 m, 平均胸径16.6 cm; 林分内分布的植物主要以毛冬青(Ilex pubescens )、乌饭树(Vaccinium bracteatum )、薄叶山矾(Symplocos anomala )、桂北木姜子( Litsea subcoriacea )、山姜(Alpinia japonica )等为主, 草本以芒萁为主; 林分表层(0-20 cm)的土壤容重为1.18 g·cm-3 , 土壤全N和全P含量分别为1.02 g·kg-1 和0.37 g·kg-1 . ...
常绿阔叶林不同更新方式下土壤呼吸的动态及机制
2
2012
... 试验地位于福建省三明市格氏栲自然保护区(26.32° N, 117.60° E)和三明市金丝湾森林公园陈大林业采育场(26.32° N, 117.60° E)内, 其中研究林分马尾松人工林分布于三明市金丝湾森林公园陈大林业采育场内, 米槠人工林分布于三明格氏栲自然保护区内.试验地所在区域属于中亚热带季风气候, 年平均气温19.1 ℃, 年降水量1749 mm, 年蒸发量1585 mm, 相对湿度81%, 平均海拔300 m, 平均坡度25°-35°, 土壤以花岗岩发育的红壤和黄壤为主, 还分布有红黄壤、黄棕壤等, 多呈酸性, 土壤厚度超过1 m (王超, 2012 ).马尾松人工林是经1976年皆伐次生林后营造人工纯林形成; 林分密度为1500株·hm-2 , 平均树高18.3 m, 平均胸径18.3 cm; 林下植被覆盖度约为90%, 灌木较少, 主要以芒萁(Dicranopteris dichotoma )、芒(Miscanthus sinensis )和蕨等草本为主(王超, 2012 ); 林分表层(0-20 cm)的土壤容重为1.33 g·cm-3 , 土壤全N和全P含量分别为0.8 g·kg-1 和0.35 g·kg-1 .米槠人工林则是米槠次生林经皆伐、火烧, 再通过挖穴造林和幼林抚育形成; 林分密度为2042株·hm-2 , 平均树高14.2 m, 平均胸径16.6 cm; 林分内分布的植物主要以毛冬青(Ilex pubescens )、乌饭树(Vaccinium bracteatum )、薄叶山矾(Symplocos anomala )、桂北木姜子( Litsea subcoriacea )、山姜(Alpinia japonica )等为主, 草本以芒萁为主; 林分表层(0-20 cm)的土壤容重为1.18 g·cm-3 , 土壤全N和全P含量分别为1.02 g·kg-1 和0.37 g·kg-1 . ...
... )和蕨等草本为主(王超, 2012 ); 林分表层(0-20 cm)的土壤容重为1.33 g·cm-3 , 土壤全N和全P含量分别为0.8 g·kg-1 和0.35 g·kg-1 .米槠人工林则是米槠次生林经皆伐、火烧, 再通过挖穴造林和幼林抚育形成; 林分密度为2042株·hm-2 , 平均树高14.2 m, 平均胸径16.6 cm; 林分内分布的植物主要以毛冬青(Ilex pubescens )、乌饭树(Vaccinium bracteatum )、薄叶山矾(Symplocos anomala )、桂北木姜子( Litsea subcoriacea )、山姜(Alpinia japonica )等为主, 草本以芒萁为主; 林分表层(0-20 cm)的土壤容重为1.18 g·cm-3 , 土壤全N和全P含量分别为1.02 g·kg-1 和0.37 g·kg-1 . ...
亚热带不同演替树种米槠和马尾松细根性状对比研究.
0
2015
亚热带不同演替树种米槠和马尾松细根性状对比研究.
0
2015
Investment in fine roots and arbuscular mycorrhizal fungi decrease during succession in three Brazilian ecosystems.
2
2012
... 不同演替阶段的树种生理和形态特征有所不同(Givnish, 1988 ; Messier, 1999 ; Ellis et al ., 2000 ), 细根特性也可能因所处演替阶段不同而存在差异.Paz (2003)研究了55种树种的根形态特征后发现, 演替晚期树种的根总体上要比演替早期树种的根更粗.Coll等(2008)的研究表明, 演替晚期树种的主根生物量所占比例更大.受生境条件等因素的影响, 不同演替阶段树种的细根生长速度可能也存在差异.在演替早期, 群落结构简单且立地条件相对较差, 植被更倾向于分配更多的生物量给根系(屠玉麟和杨军, 1995 ), 并通过迅速和广泛地占据土壤空间获取资源(Brassard et al ., 2009 ), 具有觅食的广布性; 而演替晚期群落的物种多样性较丰富且结构复杂, 土壤有机质和养分含量较高, 种间竞争激烈, 为了提高竞争力, 细根会迅速识别富养斑块并大量增生(Brassard et al ., 2009 ), 养分利用精细, 利用效率提高.Zangaro等(2012)对几个热带树种的研究发现, 细根生物量、细根直径随演替的进行而增加, 而根总长、比根长、根毛长度、根毛发生率、内生菌根菌侵染率等降低, 表明与演替后期阶段树种相比, 演替早期阶段的树种细根具有更强的养分获取能力, 生长速度可能更快.同时, 不同演替阶段的树种受栖息地环境的影响程度可能不同.如演替早期的热带树种生长受开阔栖息地的局限较大(Zangaro et al ., 2012 ), 而一些演替晚期的树种具有耐阴性(Kitajima, 1994 ; Walters & Reich, 1996 ; Khurana & Singh, 2006 ; Poorter & Rozendaal, 2008 ), 林冠更加稠密, 透光度更小.Du和Fang (2014)对中国东北的白桦(Betula platyphylla )-中国山杨(Populus davidiana )林和落叶松(Larix gmelinii )林的研究也曾发现, 演替早期的先锋树种白桦和中国山杨, 根呼吸对温度的敏感性要高于演替晚期的优势树种落叶松.此外, 不同演替阶段树种的物候情况在长期森林动态研究中十分重要, 且是预测未来气候变化环境下的森林变化所必须考虑的问题(Du & Fang, 2014 ).然而目前有关森林不同演替阶段的树种细根生产动态和细根物候方面差异的研究仍十分缺乏.分析研究不同演替阶段不同树种林分的细根生产量、季节动态及其所受环境因子影响的差异, 将有助于增进我们了解不同演替阶段的树种差异对细根生产可能造成的影响. ...
... 本研究中, 马尾松人工林在观测期2年间的细根年根长生产量均明显高于米槠人工林, 不同演替阶段的树种在遗传特性方面存在差异, 可能是造成两种人工林细根生产量相差较大的一个主要原因.不同演替阶段的树种生长速率差异较大(Zangaro et al ., 2012 ), 已有研究表明, 演替早期树种比演替晚期树种具有更高的光合作用能力(Reich et al ., 1998 ; Poorter & Rozendaal, 2008 ), 而这种更高的光合作用能力又将提高可输出为根生长所用的光合产物的数量(Nielsen et al ., 1998 ; Lynch et al ., 2005 ), 进而使得演替早期树种的根生长速率更快.另外, 马尾松为演替早期树种, 对生境适应能力较强, 因而细根的生产量更高; 而米槠属于演替后期树种, 其细根的生产量也较低.Zangaro等(2012)对几个热带树种的研究也发现, 细根生物量、细根直径随演替进行而增加, 而根总长、比根长、根毛长度、根毛发生率、内生菌根菌侵染率等降低, 表明与演替后期阶段树种相比, 演替早期阶段的树种细根可能具有更强的养分获取能力.虽然本研究区域在亚热带, 不同演替阶段树种的细根生产也可能呈现相似的差异特征.两种人工林观测第一年细根生产量均比第二年明显增大, 可能与细根生产的年际波动有关, 也可能与微根管安装过程对细根生产的刺激作用有关.有研究表明, 微根管安装切断了原有根系, 导致初期根系的大量增生与分枝, 这种刺激作用可能随时间的推移而减弱(Joslin & Wolfe, 1997 ). ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}