2007年, 加拿大科学家Dudley发表了一篇题为“Kin recognize in an annual plant”的文章(Dudley & File, 2007 )。该研究发现, 一年生植物Cakile edentula 能够识别周边其他同类个体与自己的亲缘关系, 并根据邻居亲缘关系的远近, 调整自身形态策略: 与近亲缘个体共存时, 侧根较多, 而与远亲缘个体共存时侧根较少。Dudley以此推测植物能够识别周边植物的亲缘关系, 当遇到近亲缘个体时, 采取较为缓和的竞争策略; 而当面临远亲缘个体或者非亲缘个体时, 则采用激烈的竞争策略。该文快速吸引了大量科学家的关注, 后续的许多研究也陆续发表在《Ecology Letters》、《New Phytologist》、《Journal of Experimental Botany》、《Functional Ecology》、《BMC Plant Biology》、《Planta》等国际主流刊物上。
其实, 植物亲缘识别的概念并非Dudley首创(Hodge, 2012 )。关于植物亲缘识别最早可以追溯到19世纪关于植物的自交不亲和(self-incompatibility)的研究, 近则可以追溯到1946年Muller等对灰白银胶菊(Parthenium argentatum )面对不同亲缘个体根系分布的研究(Muller, 1946 )。而在后续的30年间, 并没有提出所谓植物亲缘识别的概念。直到1980年, Nakamura在《Evolution Theory》上发表了题为“Plant kin selection”的文章, 首次提出了植物亲缘识别的概念。 而Dudley的文章再次唤起了科学家对植物亲缘识别研究的关注。
本文通过回顾国际上植物亲缘识别已有的研究结果, 结合最新的研究发现, 对植物亲缘识别的研究现状进行综述, 对现存的问题进行分析, 以期为后续的研究提供相关参考建议。
1 植物亲缘识别的提出
在动物界中, 已发现许多生物亲缘识别的现象, 而且动物的亲缘识别能力能够被自然所选择, 并一代又一代地遗传下来, 那么这种能力势必对生物的生存和繁殖具有重大的意义, 科学家们一直试图揭示这种行为所蕴含的自然奥秘。
关于亲缘识别的意义, 必须关联到一个重要的进化理论——亲缘选择理论。达尔文进化学说认为, 生物生存的最终目的是繁殖后代, 在恶劣的环境中生存下来, 实现最大的生殖效益。因此, 生物的一切行为就必须是自私自利的。但是, 自然界中存在一个“反常”现象困扰着达尔文: 在白蚁等社会性昆虫群体中, 存在工蚁为了保全整个蚁群, 不惜牺牲自己的行为, 称之为利他行为(altruism)。直至1963年, 剑桥大学动物学家Hamilton W. D.提出利他行为的亲缘选择学说(kin selection) (Hamilton, 1964 ), 并运用广义适合度(inclusive fitness)解释动物的利他行为。该理论认为, 达尔文理论中生物繁殖的目的实质上就是自身基因的延续, 而近亲缘个体间共享更多的基因, 因此帮助近亲缘个体更好地生存并繁殖更多的后代, 同样达到了延续共有基因的目的。
根据Hamilton规则, 亲缘关系越近的个体, 其共享的基因越多, 这种基于亲缘关系的利他倾向越强。Hamilton的学生兼好友Dawkins C. R.则进一步深化了Hamilton亲缘选择理论, 并在其传世之作《自私的基因》(Selfish Gene)中明确提出生物利他行为, 实质上是基因的自私行为, 明确了亲缘选择理论中自然选择基本单位为基因的观点, 并且阐明基因不但是自然界中自我复制的基本单位, 更是维护自我利益的基本单位的观点(Dawkins, 2006 )。有机体只是基因的载体, 被基因所控制的机器, 个体的行为, 不管是利他的或者自私的, 都是由基因控制的。个体在行为上表现可以利他, 但基因上必须利己——表面利他, 实则利己。个体是基因的载体, 离开了载体, 基因无法存留也无意义。基因的保留靠的是个体的繁殖, 为了使自己的复制品能够保存下去, 基因控制近亲个体更加表现为利他, 以保存和繁衍更大量的基因后代(Dawkins, 2006 )。噬菌体, 一团蛋白质包裹的基因, 其没有细胞器, 没有细胞, 没有组织与器官, 其生存的实质就是使基因不断地延续下去, 而作为自然界的其他生物, 我们虽具有形形色色的组织、器官之类, 但我们与噬菌体生存的实质并无区别, 都是基因延续的机器。
许多研究表明, 在多种生物种中存在亲缘选择现象, 如海葵(Anthopleura elegant ) (Ayre & Grosberg, 1995 )、美新亮丽鲷(Neolamprologus pulcher ) (Dierkes et al ., 2005 )、蝾螈(Ambystoma tigrinum nebulosum ) (Pfennig & Collins, 1993 ), 白额食蜂鸟(Merops bullockoides ) (Emlen et al ., 1995 )、拜氏黄鼠(Spermophilus beldingi ) (Mateo, 2003 )、黑猩猩(Chimpanzee ) (Morin et al ., 1994 )。更有趣的是, 在对盘基网柄菌(Dictyostelium discoideum ) (Ostrowski et al ., 2008 )、酵母菌(Saccharomyces cerevisiae ) (Smukalla et al ., 2008 )等微生物的研究中, 同样发现类似亲缘选择现象。以上研究表明, 亲缘选择行为覆盖了肠腔类、鱼类、两栖类、鸟类、哺乳类, 乃至微生物等生物界的大部分类群。
亲缘选择理论的前提, 就是生物首先必须能够识别周边个体与自己的亲缘关系, 然后根据彼此间的关系调整自身生活史策略, 以促进近亲缘个体的生存与繁衍。因此, 亲缘识别是亲缘选择的前提, 没有亲缘识别, 亲缘选择理论则无法成立。
目前, 已在动物界、微生物界中发现了个体的亲缘选择行为, 但对于植物界是否存在亲缘选择现象, 相关的研究仍很少(Hodge, 2012 )。
2 植物亲缘识别的表征
自1980年, 植物亲缘识别概念提出至今, 科学家们已在包括Lycopersicon lycopersicum (Willison et al ., 1987 )、长叶车前(Plantago lanceolata ) (Tonsor, 1989 )、Cakile edentula (Donohue, 2003 ; Dudley & File, 2007 ; Bhatt et al ., 2011 )、大麦(Hordeum vulgare ) (Ninkovic, 2003 )、狭叶羽扇豆(Lupinus angustifolius ) (Milla et al ., 2012 )、拟南芥(Arabidopsis thaliana ) (Goodnight, 1985 ; Biedrzycki et al ., 2010 ; Masclaux et al ., 2010 ; Caffaro et al ., 2013 ; Crepy & Casal, 2015 )、水稻(Oryza sativa ) (Fang et al ., 2013 )、豚草(Ambrosia artemisiifolia ) (File et al ., 2012 )等20种植物中进行验证, 其中有许多研究结果支持植物存在亲缘识别 (表1 ), 也有一些研究得到否定的结果(表2 )。
目前研究的主流思路是将目标植物与不同亲缘关系的个体(同物种)共同种植, 测定目标植物个体的形态学特征(株高、茎粗、叶面积、根系长度、花序数量、种子数量、叶片重叠度、根系交叉程度等)或者生理代谢特征(光合速率, 呼吸速率、植株次级代谢物质含量、叶片挥发物、根系分泌物等), 还有少数从植物响应胁迫应答的角度进行研究。通过以上指标来检测目标个体是否存在表型或者代谢差异, 从而间接地推断植物是否能够识别周边植物与自己的亲缘关系, 并调整自身的生理生态策略(图1 )。
图1 植物亲缘识别的作用方式。参照Bais, 2015。
Fig. 1 Possible interaction of kin recognition in plants. Modified from Bais, 2015.
2.1 生长发育特征
Willison等(1987)对垂序商陆(Phytolacca americana )、苘麻(Abutilon theophrasti )、Solanum mauritanianum 研究发现, 垂序商陆与近亲缘个体(来自同一母体)共存时, 具有更高的株高和种子数量, 而苘麻具有更高的幼苗存活率。Tonsor (1989)研究表明, 长叶车前(Plantago lanceolata )与近亲缘(克隆体)共处时, 形成更多的花序及坐果数。对Impatiens capensis 的研究也表明, 与近亲缘个体共处时, 目标个体提早了开花时间(Argyres & Schmitt, 1992 )。以上这些早期的研究认为, 与近亲缘个体共存时, 植物个体具有较高的繁殖效益, 恰好符合达尔文关于生物生存的最终目的是获得最大生殖效益的原则, 从正面证实了植物亲缘识别可能带来的好处。
相比于早期研究侧重繁殖性状, 近期研究则更侧重于对生长性状的探讨, 主要通过生物量、株高、叶面积、根冠比、根系长度等指标表征植物与不同亲缘个体共存时, 目标个体的生存特征, 从侧面表征植物的亲缘识别行为。对Cakile edentula 、大麦、拟南芥、Distichlis spicata 等的研究均表明, 植物与近亲缘个体共存时, 提高了自身生物量(Donohue, 2003 ; Bhatt et al ., 2011 ; Biernaskie, 2011 ; Mercer & Eppley, 2014 )。但也有研究表明与近亲缘个体共处时, 目标个体生物量反而可能降低(Cheplick & Kane, 2004 ; Milla et al ., 2009 ), 或者没有影响(Ninkovic, 2003 )。此外, Tonsor (1989)研究发现, 当长叶车前与近亲缘个体共存时, 干质量虽然下降, 但产生更多的子代, 表明与近亲缘个体共存时虽然生存效益下降, 但是繁殖效益提高。因此, 目前而言, 用生物量的大小来判断植物是否存在亲缘识别或者亲缘利他存在着一定的局限性。
此外, 部分研究也采用以目标植物的根系生长情况来表征植物面临不同亲缘的生长状态, 目前有以下6种主要结果(图2 ): (1)与近亲缘个体共存, 根系生长无差异; (2)与近亲缘个体共存, 形成较少侧根; (3)与近亲缘个体共存, 形成较小根系; (4)与近亲缘个体共存, 形成较大根系; (5)根系趋向近亲缘个体生长; (6)根系回避近亲缘个体生长。
图2 植物近亲缘个体共存根系生长模式。参照Depuydt, 2014。
Fig. 2 Root growth model of target plants responded to kin neighbors. Modified from Depuydt (2014) .
其中大多数支持植物存在亲缘识别的研究 (表1 )认为, 目标植物与远亲缘共存时具有较大的根系, 从而加强竞争(Ninkovic, 2003 ; Dudley & File, 2007 ; Biedrzycki et al ., 2010 ), 而反对的结果(表2 )则表明, 与近亲缘个体共存时形成更多根系(Murphy & Dudley, 2009 )或者两者间并无差异(Milla et al ., 2012 ; Semchenko et al ., 2014 )。Semchenko等(2014)的研究结果则提供了另一种新的思路, 目标植物与不同亲缘个体共存时, 根系长度并无差异, 但显著改变根系的构型。由于根系长度无法准确地表征植物的亲缘识别特征, 因此, 有学者提出从根系生长趋性的角度进行分析(趋向近亲缘或者远亲缘)。Caffaro等(2013)和Fang等(2013)的研究均表明, 目标植物根系更趋向于近亲缘个体方向生长。另外还有部分研究采用目标植物根冠比来表征其与周边亲缘邻居的竞争强度(Bhatt et al ., 2011 ; Mercer & Eppley, 2014 )。首先, 当目标植物面对不同亲缘邻居, 其总生物量的形成可能并不受影响, 但其可能通过改变地上部地下部分配(根冠比)而改变竞争策略; 其次, 目标植物根系减少, 并不一定意味着个体间竞争减弱, 其可能通过将更多能量投入到地上部生产或者加大次级代谢产物的合成, 增加了个体间的竞争压力。因此, 采用根冠比能在一定程度上弥补采用根系生物量和根系长度较片面表征个体间竞争强度的不足。
种子(后代)数量、种子萌发率、幼苗成活率等指标也被许多研究所应用(Willison et al ., 1987 ; Tonsor, 1989 ; Cheplick & Kane, 2004 ; Milla et al ., 2009 ; Masclaux et al ., 2010 ), 因其能够最直观地表征植物能否因亲缘识别而带来直接效益, 即繁殖效益。Willison等(1987)及Tonsor (1989)的研究均表明, 植物个体与近亲缘共存时具有更高的繁殖收益。
据Cheplick和Kane (2004) 对Triplasis purpurea 的研究发现, 与近亲缘个体共存时, 其种子数量反而显著减少。Milla等(2009)对狭叶羽扇豆(Lupinus angustifolius )所进行的全生命周期实验发现, 与近亲缘个体共存时, 目标植物的开花数、结果数均小于远亲缘个体共存。Masclaux等(2010)对拟南芥的研究也得到类似的结果。从理论上, 繁殖指标能够最直观地表征植物的亲缘识别效益——繁殖效益, 但目前研究仍然无法得到统一的结论。
从支持植物亲缘识别存在(表1 )和反对的研究结果(表2 )来看, 大多支持植物亲缘识别的研究多以生长指标为证据, 且多为植物苗期的数据, 而否定植物亲缘识别的研究侧重测定繁殖指标。相对而言, 两者均存在一定的不足。首先, 苗期的形态学数据仅能表明植物生长初期的竞争特征, 并无法反映个体到生长后期, 特别是繁殖期面对不同亲缘邻居的策略; 其次, 繁殖指标能够直接地表征植物个体的繁殖效益, 但也仅是这一特定时期的表征。因此, 目前的研究存在一定的缺陷, 均采用某特定时间的离散数据, 片面判断植物是否存在亲缘识别(选择), 而缺乏对指标进行长周期的、持续的、全生育时期的测定。
2.2 生理代谢特征
前述的诸多研究表明: 植物与不同亲缘个体共存能够显著改变自身某些生长发育策略, 但也有部分研究表明这种差异并不显著。那么生长发育指标无差异是否意味着植物亲缘识别就不存在了呢?为了回答这个问题, 少数研究人员从更为微观的角度, 从植物的生理代谢层面来探讨该现象。
研究表明, 当植物与同种/异种个体共存时, 能够改变自身初、次级代谢平衡。Ormeño等(2007)发现, 当迷迭香(Rosmarinus officinalis )、Pinus halepensis 与同种个体共存时, 叶片中酚类物质显著提高。Broz等(2010)通过代谢组学方法发现, 当Centaurea maculosa 与同种共存时, 提高了叶片酚类合成、蛋白酶抑制剂合成等次级代谢过程, 而降低了糖酵解、三羧酸循环、脂类代谢、氨基酸代谢等初级代谢过程。以上结果证明, 植物与同种个体共存时, 可能将更多的能量投入到次级代谢中, 在植株中形成了更多的酚类、抗氧化酶等物质, 以抵御外来植食者和病菌的侵害; 而减少叶片及根系形成等初级代谢的投入, 以减少同种个体对外部养分的竞争, 增强对外界环境胁迫的抵御能力。
相关研究表明, 当植物与不同亲缘个体共存时, 也存在类似的现象。Biedrzyck等(2011)研究发现, 拟南芥与远亲缘个体共存时, 根系病害相关基因的表达量上调。Badri等(2012)进一步研究发现, 其根系分泌物中病害防御类的蛋白也显著增加, 以上结果暗示植物或许能够识别周边邻居的亲缘关系, 并调节自身的某些生理代谢过程。
综上所述, 目前从植物生理代谢角度探讨植物亲缘识别的研究相对较少, 但现存对同种/异种植物个体共存时的代谢研究的思路方法, 将可供我们参考借鉴。
2.3 环境胁迫应答
一般认为, 同种或近亲缘植物共存, 因其生理生态特性高度相似, 所构成聚落物种单一, 更容易受到病虫害的危害(Schmitt & Antonovics, 1986 ; Weiblen et al ., 2006 ; Burns & Strauss, 2011 ; Liu et al ., 2012 )。但据Karban和Shiojiri (2009)、Karban等(2013)对Artemisia tridentata 的研究发现, 当植株受到机械损伤或者昆虫取食后, 能够向周边释放挥发物, 这些物质能够诱导周边植物的预警抗性, 而且近亲缘个体受诱导后, 抗性优于远亲缘个体, 表明亲缘识别可能影响植物的通讯与防御。
对龙葵(Solanum nigrum ) (Schmidt & Balswin, 2006 )、Pinus halepensis (Ormeño et al ., 2007 )、Centaurea maculosa (Broz et al ., 2010 )、拟南芥(Badri et al ., 2012 )等的研究也发现同种个体共存植株体内抗性物质提高的现象, 这有力地支持了Karban等的观点。
2.4 亲缘识别的途径和机制
现有研究表明植物存在亲缘识别的能力, 但对于植物究竟是通过何种途径进行通讯和识别则仍不完全清楚, 而植物亲缘识别机制也是近10年才缓慢开始, 仅有少量文献进行了报导。已有研究初步表明, 植物可能通过根系分泌物、叶片挥发物、感光载体等途径进行亲缘识别。
2.4.1 根系分泌物
研究表明, 植物能够识别土壤环境的养分条件, 将根系向资源丰富区域集中, 而减少向资源匮乏地区的增殖, 表明植物根系的生长和分布是由土壤中的营养物质浓度所驱动的(Hodge, 2004 )。但近年研究发现, 一些植物的根系生长和分布并不是受土壤营养水平驱动的, 而是更多受到邻近植物根系的影响(Hodge, 2009 ; Mommer et al ., 2010 ), 植物的亲缘识别可能正属于这种非养分驱动行为。
Biedrzycki等(2010)将拟南芥浸泡于含有不同亲缘个体根系分泌物的培养液中, 发现当暴露在近亲缘个体的根系分泌物时, 目标个体形成了较少的侧根, 而暴露在远亲缘个体根系分泌物中则形成较多侧根(图3A ), 这种现象在添加根系分泌物抑制剂后消失(图3B )。由此推测, 根系分泌物可能介导了植物亲缘识别, 但文中并没有提到具体的分泌物。此外, 这一选择性响应很有可能是由两个拟南芥品种根系分泌物中的某些特定的化学成分缺失导致的。Biedrzycki等(2011)后续进行的转录组分析发现, 目标植物暴露在远缘个体的根系分泌物后, 根系中ATP/GST转运载体、次级代谢合成、病程相关基因表达量上调。更有趣的是, 除了根系病程相关基因表达量上调外, 根系分泌物中的病程相关蛋白也相应增多(Badri et al ., 2012 )。这些结果是否暗示着植物亲缘识别和植物对于病原体的识别可能属于同一个途径, 植物将远亲缘个体按照病原体进行识别和对待, 该假设有待进一步验证。此外, 已有研究表明, 当目标植物与远缘个体共存时, 促进了目标个体根系的生长(初级代谢), 转录组分析表明, 根系中初级代谢相关基因的表达量并无明显上调(Biedrzycki et al ., 2011 ), 但次级代谢产物合成量增加, 这两个似乎是相互矛盾的结果。
图3 不同亲缘个体根系分泌物对拟南芥侧根生长的影响。A, 未添加根系分泌物抑制剂。B, 添加根系分泌物抑制剂。
Fig. 3 Lateral roots in Arabidopsis thaliana plants grown solitary (own) or in sibling (kin) and non-sibling (stranger) exudates from the secretion inhibitor experiment. A, Without secretion inhibitor of root. B, With secretion inhibitor of root.
2.4.2 叶片挥发物
许多植物受到虫害胁迫或者物理伤害后, 都能够向周边释放挥发物, 且这些挥发物能够诱导周边个体的防御能力(Wu & Baldwin, 2009 )。那么, 对于同一警报挥发物, 不同亲缘个体的响应效果是否有差别呢?
Karban和Shiojiri (2009)、Karban等(2013)的研究有利地支持了上述观点, 研究结果表明, 当Arte- misia tridentate 受到机械损伤或者昆虫取食后所释放的警报挥发物能够提高周边个体的抗虫性, 而且近亲缘个体抗虫性的提高程度显著高于远亲缘个体, 作者猜测这可能与挥发物的组成有关。其进一步分析发现, Artemisia tridentate 挥发物的组成与其亲缘关系显著相关, 即亲缘关系越接近, 其叶片挥发物的组成越相似。因此, 其认为来源于近亲缘个体的警报信号彼此间的成分更加相似, 因此受体植物更容易识别这些信息(Karban et al ., 2013 )。
在胁迫条件下, 植物个体能够从亲缘识别中获益。那么, 在非胁迫条件下又是怎样的情况呢?Ninkovic (2003)将大麦暴露在不同基因型大麦的挥发物中后发现, 当Kara大麦暴露在Alva大麦的挥发物中时, Kara大麦形成了更多的根系, 而当Kara暴露在同种大麦的挥发物中或者空气中则没有这种现象。该研究表明, 大麦能够对不同来源的挥发物进行差异响应, 并调整自身的生长策略。当识别到远缘个体挥发物时, 感知到周边可能存在的竞争, 故而加大对根系的投入, 加强自身的竞争能力, 当感知到近亲缘个体挥发物时, 则减少根系分布, 减少近亲缘个体间的竞争。
2.4.3 感光受体
植物光合色素对光吸收具有选择性, 投射到叶片上的红光和蓝光大部分均被吸收, 仅有少量的红光、红外光和蓝光能够穿透叶片, 透射到冠层下(Smith, 2000 ; Casal, 2013 ); 而冠层下的植物或者叶片进化出一种有效的机制, 能够通过冠层下红光与远红外光的比例, 判断周边的光照条件, 调整叶片的生长方向, 向光照充分的地方生长, 并减少叶片间的相互遮蔽(Casal, 2013 )。已有研究表明, 植物主要通过光敏色素B (phyB)和隐花色素(cry1)识别这两种光的微弱变化(Sellaro et al ., 2010 ; Keller et al ., 2011 )。
Crep和Casal (2015) 的最新研究表明, 拟南芥与近亲缘共存时, 能够改变叶面的生长方向, 避免与近亲缘个体的叶片交叉, 而与远缘个体共同生长时, 叶片则相互交叉。他们推测植物可能通过感知红光与红外光的比值, 来判别周边的植物。因此, 其选用光敏色素B、隐花色素1、隐花色素2 (cry2)、向光素1 (phto1)、向光素2 (phto2)感受载体缺失突变体进行试验, 发现近亲缘个体叶片避免交叉的现象消失, 该研究首次证明光可能也是植物亲缘识别的重要途径, 并且证明感光载体参与了植物亲缘识别过程。
综上所述, 目前对于亲缘识别机制的研究非常匮乏, 虽然已初步探明根系分泌物、叶片挥发物、受光载体可能介导了这一行为, 但究竟是何种具体物质介导了这种行为仍不清楚; 此外, 植物体内是否存在某种载体接受识别信息, 亲缘识别物质是否存在浓度效应等问题仍待进一步研究。
3 亲缘识别研究的不足及改进策略
总之, 目前对于植物亲缘识别(选择)是否真实存在, 仍存在较大的争议(表1 , 表2 ), 且目前的研究主要集中在目标植物与不同亲缘邻居共存时表型的差异。综合前人研究, 出现这种相矛盾的结果可能与实验材料的选择、亲缘关系的界定标准、环境条件及测定的指标有关。
3.1 实验材料的选择
现有研究已在包括拟南芥等至少20种植物上进行验证(表1 , 表2 )。实验采用的研究对象不一样, 不同植物本身亲缘识别的强度、途径、方式可能存在较大差异, 因此容易造成研究结果的差异(Biedrzycki & Bais, 2010 )。从植物分类学的角度看, 目前所报道的20个种分别来自2纲9目10科20属(表3 )。按照恩格勒植物分类系统, 这些种所覆盖的目, 诸如桔梗目、车前目、管状花目隶属于后生花被亚纲, 蔷薇目、罂粟目、无患子目、锦葵目隶属于异被花类, 中央子目和禾本目隶属于同被花类, 均属于被子植物, 而相对原始的裸子植物目前仍未见相关报道。从品种来源上看, 目前多数实验采用野生植物, 部分实验采用模式植物(拟南芥、水稻), 而经长期人工筛选的作物研究相对较少, 仅在大麦(Ninkovic, 2003 )、玉米(Zea may ) (Lin, 2013 )、大豆(Glycine max ) (肖桃艳等, 2015 )上有少量报道。因此, 建议在后续试验中加强向裸子植物和人工长期筛选作物两个方向延伸, 验证植物亲缘识别现象在原始的裸子植物以及长期人工选育作物(部分原始基因丢失)中是否仍然存在, 从而确定这种行为是否具有普遍性。此外, 目前的研究对象除Artemisia tridentate 外均为草本植物, 建议在后续研究中多关注灌木及木本植物。
3.2 亲缘关系的界定
不同研究对实验材料亲缘关系的界定方式和亲缘程度的选择也存在较大差异, 目前主要包括以下3种界定方式: 第一种以是否来自同一亲本为区分标准(Argyres & Schmitt, 1992 ; Dudley & File, 2007 ; Bhatt et al ., 2011 ; Semchenko et al ., 2014 ); 第二种以基因型为区分标准(Ninkovic, 2003 ; Biedrzycki et al ., 2010 ; Crepy & Casal, 2015 ); 第三种以人为克隆体为区分标准(Tonsor, 1989 ; Karban & Shiojiri, 2009 ; Mercer & Eppley, 2014 )。不同亲缘关系的界定方式, 也是导致目前研究结果差异较大的最主要原因。关于何种界定方式最为合适, 目前仍没有统一的定论。参考其他生物亲缘识别的相关研究, 动物主要以个体间血缘关系远近来界定个体间的亲缘关系(Emlen et al ., 1995 ), 而微生物主要以分裂方式进行繁殖, 故研究中主要以基因型或遗传距离来区分个体(菌群)之间的亲缘关系(Ostrowski et al ., 2008 )。目前植物亲缘识别研究主要以亲本关系及基因型(遗传距离)两种亲缘界定方式为主流, 且使用这两种方式的研究中, 均有支持和否定亲缘识别的结果, 所以这两种方式都可以得到相对准确的结果。因此, 选取何种亲缘界定方式最为合适仍很难判断, 在短期内可能仍会呈现多种界别方式共存的局面。为了尽量减少因采用不同亲缘界别方式所可能带来的误差, 建议后续实验中, 仅在采用同种亲缘界别方式的实验间进行结果比较。
3.3 测定指标的选定
指标选定方面主要存在两个问题。第一个是选用的指标范围多局限于形态学和生长发育指标。目前大部分研究主要采用诸如株高、叶面积、生物量、开花数、开花时间、坐果数、种子数量、种子萌发率、种子存活率、根系长度、根冠比、叶片重叠度、根系交叉度等形态学指标, 间接表征植物是否存在亲缘识别现象。还有少数研究采用植物代谢酶活性(肖桃艳等, 2015 )、植物光合作用和呼吸作用速率(Meier et al ., 2013 )、根系分泌物(Badri et al ., 2012 )等生理代谢指标, 菌根数量(File et al ., 2012 )等环境指标, 基因表达模式(Biedrzycki et al ., 2011 ; Masclaux et al ., 2010 )等指标, 来研究目标植物与不同亲缘个体共存时的表现。
研究表明, 植物面对周边个体, 不但能够产生一定的形态学响应, 而且在更深层次上还有许多“看不见”的生理生化策略的变化(Karban et al ., 2000 ; Baldwin et al ., 2006 )。对白车轴草(Trifolium repens ) (Lepik et al ., 2012 )、狭叶羽扇豆(Milla et al ., 2012 )的研究发现: 这两种植物分别与不同亲缘个体共存时, 其叶面积、株高、生物量、根长均无差异, 但未测定生理学指标, 因此还不能完全否定植物亲缘识别的存在。另外一个问题是指标上的以偏概全。部分研究采用某单一指标, 如侧根长(Dudley & File, 2007 , Biedrzycki et al ., 2010 )、生物量(Argyres & Schmitt, 1992 )、根冠比(Bhatt et al ., 2011 )存在的差异(其他指标可能没有差异)为标准, 判断植物会否存在亲缘识别。但是, 单一指标的差异并不能代表所有指标的趋势, 因此利用单一指标的差异表征亲缘识别现象的存在是以偏概全。因此, 建议在后续的实验中, 除了考察形态指标外, 还应该加大对植物生理生化指标的研究, 例如植株的养分利用能力, 次级代谢产物合成, 根系分泌物, 叶片挥发物, 由目前的以表观形态观察为主, 逐步转变到以植物形态变化与生理变化相结合的复合途径, 能够更加全面和直接地表征植物亲缘识别特征以及辅助对植物亲缘识别机理的研究。
3.4 环境因素的影响
外界环境的优劣和资源的丰贫对植物生长具有重要的影响, 植物能够感知这些差异并通过一定的形态和生理策略响应这些外界因素(例如根系的向肥性和叶片的向光性等)。目前, 大部分植物亲缘识别的研究侧重于植物亲缘识别本身, 较少关注环境因素的影响。Andalo等(2001)研究发现, 在低CO2 浓度下, 近亲缘个体共存形成较多的果实和种子, 而CO2 浓度提高后, 远亲缘组形成更多的果实和种子。这表明环境因子对植物亲缘识别的影响可能要比原来想象的更加复杂。另外, 根据Hamilton (1964)定律, 亲缘识别(选择)在一定的条件下才能发生。在外界环境因素优越的条件下, 个体生存得到满足的情况下, 才表现亲缘合作趋向, 而在资源短缺时, 个体生存压力巨大, 则可能呈现竞争趋向(File et al ., 2011 )。因此, 不同实验, 对于水分、养分、光照、湿度设定差异, 也是造成相互矛盾结果的原因之一。鉴于目前植物亲缘识别研究仍处于初级阶段, 且大部分研究主要采用室内盆栽实验, 环境因素可控, 可能采用富余养分的实验设计相对合适。后续还可以开展相关环境因素对植物亲缘识别影响的研究, 以及探索野外条件下环境限制因子相对较多条件下植物的亲缘识别行为。
4 展望
前人已从不同角度, 对植物是否存在亲缘识别进行了很多有益的探索, 但目前对植物亲缘识别研究暂无统一模式, 不同研究对于亲缘关系的界定标准、实验材料、测定指标、环境因素均存在较大差异, 得出的结果也大相径庭, 不同科学家对植物亲缘识别是否存在也存在较大争议, 不过, 正是这种争议和矛盾, 不断地推动着该领域研究的发展, 使我们不断向真理靠近(Biedrzycki & Bais, 2010 )。
未来的研究, 应对亲缘关系的界定、研究材料的选择及实验设计有一个相对统一的标准。除了阐明不同亲缘物种共存的形态学和表型性状外, 从植物生理生化、分子、蛋白质组和代谢组学水平上解析植物亲缘识别的机制将是下一阶段研究的重点, 揭示植物亲缘识别的主要途径、介导物质以及植物接收信息的载体将是研究其机理的主要工作(Crepy & Casal, 2015 ); 同时应该考虑不同环境条件对亲缘识别的影响。希望通过广大科研工作者的努力, 更加完整地阐明植物亲缘识别行为, 揭示这一有趣的自然进化奥秘。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Andalo C Goldringer I Godelle B 2001 ). Inter- and intragenotypic competition under elevated carbon dioxide in Arabidopsis thaliana.
Ecology , 82 , 157 -164 .
[2]
Argyres AZ Schmitt J 1992 ). Neighbor relatedness and competitive performance in Impatiens capensis (Balsaminaceae): A test of the resource partitioning hypothesis.
American Journal of Botany , 79 , 181 -185 .
[本文引用: 3]
[3]
Ayre DJ Grosberg RK 1995 ). Aggression, habituation, and clonal coexistence in the sea anemone Anthopleura elegantissima.
American Naturalist , 146 , 427 -453 .
[本文引用: 1]
[4]
Badri DV De-la-Peña C Lei ZT Manter DK Chaparro JM Guimarães RL Sumner LW Vivanco JM 2012 ). Root secreted metabolites and proteins are involved in the early events of plant-plant recognition prior to competition.
PLoS ONE , 7 , e46640 .
[5]
Bais HP 2015 ). Shedding light on kin recognition response in plants.
New Phytologist , 205 , 4 -6 .
[6]
Baldwin IT Halitschke R Paschold A Von Dahl CC Preston AC 2006 ). Volatile signaling in plant-plant interactions: “Talking trees” in the genomics era.
Science , 311 , 812 -815 .
[本文引用: 1]
[7]
Bhatt MV Khandelwal A Dudley SA 2011 ). Kin recognition, not competitive interactions, predicts root allocation in young Cakile edentula seedling pairs.
New Phytologist , 189 , 1135 -1142 .
[本文引用: 5]
[8]
Biedrzycki ML Bais HP 2010 ). Kin recognition in plants: A mysterious behaviour unsolved.
Journal of Experimental Botany , 61 , 4123 -4128 .
[本文引用: 6]
[9]
Biedrzycki ML Jilany TA Dudley SA Bais HP 2010 ). Root exudates mediate kin recognition in plants.
Communicative & Integrative Biology , 3 , 28 -35 .
[10]
Biedrzycki ML Venkatachalam L Bais HP 2011 ). Transcriptome analysis of Arabidopsis thaliana plants in response to kin and stranger recognition.
Plant Signaling & Behavior , 6 , 1515 -1524 .
[本文引用: 2]
[11]
Biernaskie JM 2011 ). Evidence for competition and cooperation among climbing plants.
Proceedings of the Royal Society B: Biological Sciences , 278 , 1989 -1996 .
[本文引用: 1]
[12]
Broz AK Broeckling CD De-La-Peña C Lewis MR Greene E Callaway RM Sumner LW Vivanco JM 2010 ). Plant neighbor identity influences plant biochemistry and physiology related to defense.
BMC Plant Biology , 10 , 115 .
[本文引用: 1]
[13]
Burns JH Strauss SY 2011 ). More closely related species are more ecologically similar in an experimental test.
Proceedings of the National Academy of Sciences of the United States of America , 108 , 5302 -5307 .
[本文引用: 1]
[14]
Caffaro MM Vivanco JM Botto J Rubio G 2013 ). Root architecture of Arabidopsis is affected by competition with neighbouring plants.
Plant Growth Regulation , 70 , 141 -147 .
[本文引用: 1]
[15]
Casal JJ 2013 ). Photoreceptor signaling networks in plant responses to shade.
Annual Review of Plant Biology , 64 , 403 -427 .
[本文引用: 2]
[16]
Cheplick GP Kane KH 2004 ). Genetic relatedness and competition in Triplasis purpurea (Poaceae): Resource partitioning or kin selection?
International Journal of Plant Sciences , 165 , 623 -630 .
[本文引用: 2]
[17]
Crepy MA Casal JJ 2015 ). Photoreceptor-mediated kin recognition in plants.
New Phytologist , 205 , 329 -338 .
[本文引用: 3]
[18]
Dawkins R 2006 ). The Selfish Gene (30th Anniversary edition) . Oxford University Press , Oxford .
[19]
Depuydt S 2014 ). Arguments for and against self and non-self root recognition in plants.
Frontiers in Plant Science , 5 , 614 .
[本文引用: 1]
[20]
Dierkes P Heg D Taborsky M Skubic E Achmann R 2005 ). Genetic relatedness in groups is sex-specific and declines with age of helpers in a cooperatively breeding cichlid.
Ecology Letters , 8 , 968 -975 .
[本文引用: 1]
[21]
Donohue K 2003 ). The influence of neighbor relatedness on multilevel selection in the Great Lakes sea rocket.
The American Naturalist , 162 , 77 -92 .
[本文引用: 2]
[22]
Dudley SA File AL 2007 ). Kin recognition in an annual plant.
Biology Letters , 3 , 435 -438 .
[本文引用: 5]
[23]
Emlen ST Wrege PH Demong NJ 1995 ). Making decisions in the family: An evolutionary perspective.
American Scientist , 83 , 148 -157 .
[本文引用: 2]
[24]
Fang SQ Clark RT Zheng Y Iyer-Pascuzzi AS Weitz JS Kochian LV Edelsbrunner H Liao H Benfey PN 2013 ). Genotypic recognition and spatial responses by rice roots.
Proceedings of the National Academy of Sciences of the United States of America , 110 , 2670 -2675 .
[本文引用: 1]
[25]
File AL Murphy GP Dudley SA 2011 ). Fitness consequences of plants growing with siblings: Reconciling kin selection, niche partitioning and competitive ability.
Proceedings of the Royal Society B: Biological Sciences , 279 , 209 -218 .
[本文引用: 1]
[26]
File AL Klironomos J Maherali H Dudley SA 2012 ). Plant kin recognition enhances abundance of symbiotic microbial partner.
PLoS ONE , 7 : e45648 .
[本文引用: 2]
[27]
Goodnight CJ 1985 ). The influence of environmental variation on group and individual selection in a cress.
Evolution , 39 , 545 -558 .
[本文引用: 1]
[28]
Hamilton WD 1964 ). The genetical evolution of social behaviour. II.
Journal of Theoretical Biology , 7 , 17 -52 .
[本文引用: 1]
[29]
Hodge A 2004 ). The plastic plant: Root responses to heterogeneous supplies of nutrients.
New Phytologist , 162 , 9 -24 .
[本文引用: 1]
[30]
Hodge A 2009 ). Root decisions.
Plant, Cell & Environment , 32 , 628 -640 .
[本文引用: 1]
[31]
Hodge A 2012 ). Plant root interactions. In: Witzany G, Baluska F eds. Biocommunication of Plants . Springer, Berlin . 157 -169 .
[32]
Karban R Baldwin IT Baxter KJ Laue G Felton GW 2000 ). Communication between plants: Induced resistance in wild tobacco plants following clipping of neighboring sagebrush.
Oecologia , 125 , 66 -71 .
[本文引用: 1]
[33]
Karban R Shiojiri K 2009 ). Self-recognition affects plant communication and defense.
Ecology Letters , 12 , 502 -506 .
[本文引用: 1]
[34]
Karban R Shiojiri K Ishizaki S Wetzel WC Evans RY 2013 ). Kin recognition affects plant communication and defence.
Proceedings of the Royal Society B: Biological Sciences , 280 , 20123062 .
[本文引用: 1]
[35]
Keller MM Jaillais Y Pedmale UV Moreno1 JE Chory J Ballaré CL 2011 ). Cryptochrome 1 and phytochrome B control shade-avoidance responses in Arabidopsis via partially independent hormonal cascades.
The Plant Journal , 67 , 195 -207 .
[本文引用: 1]
[36]
Lepik A Abakumova M Zobel K Semchenko M 2012 ). Kin recognition is density-dependent and uncommon among temperate grassland plants.
Functional Ecology , 26 , 1214 -1220 .
[本文引用: 1]
[37]
Lin WP 2013 ). A Preliminary Inverstigations of Kin Altruism in Maize. Master degree dissertation, College of Agriculture, South China Agricultural University
, Guangzhou.
[本文引用: 1]
(in Chinese with English abstract). [林威鹏 (2013 ). 利用玉米检验植物亲缘利他假说的初探
. 硕士学位论文, 华南农业大学, 广州 .]
[本文引用: 1]
[38]
Liu XB Liang MX Etienne RS Wang YF Staehelin C Yu SX 2012 ). Experimental evidence for a phylogenetic Janzen—Connell effect in a subtropical forest.
Ecology Letters , 15 , 111 -118 .
[本文引用: 1]
[39]
Masclaux F Hammond RL Meunier J Gouhier-Darimont C Keller L Reymond P 2010 ). Competitive ability not kinship affects growth of Arabidopsis thaliana accessions.
New Phytologist , 185 , 322 -331 .
[本文引用: 3]
[40]
Mateo JM 2003 ). Kin recognition in ground squirrels and other rodents.
Journal of Mammalogy , 84 , 1163 -1181 .
[本文引用: 1]
[41]
Meier IC Angert A Falik O Shelef O Rachmilevitch S 2013 ). Increased root oxygen uptake in pea plants responding to non-self neighbors.
Planta , 238 , 577 -586 .
[本文引用: 1]
[42]
Mercer CA Eppley SM 2014 ). Kin and sex recognition in a dioecious grass.
Plant Ecology , 215 , 845 -852 .
[本文引用: 3]
[43]
Milla R Del Burgo AV Escudero A Iriondo JM 2012 ). Kinship rivalry does not trigger specific allocation strategies in Lupinus angustifolius.
Annals of Botany , 110 , 165 -175 .
[本文引用: 3]
[44]
Milla R Forero DM Escudero A Iriondo JM 2009 ). Growing with siblings: A common ground for cooperation or for fiercer competition among plants?
Proceedings of the Royal Society B: Biological Sciences , 276 , 2531 -2540 .
[本文引用: 2]
[45]
Mommer L van Ruijven JV de Caluwe H Smit-Tiekstra AE Wagemaker CAM Ouborg NJ Bögemann GM van der Weerden GM Berendse F de Kroon H 2010 ). Unveiling below-ground species abundance in a biodiversity experiment: A test of vertical niche differentiation among grassland species.
Journal of Ecology , 98 , 1117 -1127 .
[本文引用: 1]
[46]
Morin PA Moore JJ Chakraborty R Jin L Goodall J Woodruff DS 1994 ). Kin selection, social structure, gene flow, and the evolution of chimpanzees.
Science , 265 , 1193 -1201 .
[本文引用: 1]
[47]
Muller CH 1946 ). Root Development and Ecological Relations of Guayule. United States Department of Agriculture
, Economic Research Service. Washington.
[本文引用: 1]
[48]
Murphy GP Dudley SA 2009 ). Kin recognition: Competition and cooperation in Impatiens (Balsaminaceae).
American Journal of Botany , 96 , 1990 -1996 .
[本文引用: 1]
[49]
Nakamura RR 1980 ). Plant kin selection.
Evolutionary Theory , 5 , 113 -117 .
[50]
Ninkovic V 2003 ). Volatile communication between barley plants affects biomass allocation.
Journal of Experimental Botany , 54 , 1931 -1939 .
[本文引用: 5]
[51]
Ormeño E Fernandez C Mévy J 2007 ). Plant coexistence alters terpene emission and content of Mediterranean species.
Phytochemistry , 68 , 840 -852 .
[本文引用: 1]
[52]
Ostrowski EA Katoh M Shaulsky G Queller DC Strassmann JE Barton NH 2008 ). Kin discrimination increases with genetic distance in a social amoeba.
PLoS Biology , 6 , e287 .
[本文引用: 2]
[53]
Pfennig DW Collins JP 1993 ). Kinship affects morphogenesis in cannibalistic salamanders.
Nature , 362 , 836 -838 .
[本文引用: 1]
[54]
Schmidt DD Baldwin IT 2006 ). Transcriptional responses of Solanum nigrum to methyl jasmonate and competition: A glasshouse and field study.
Functional Ecology , 20 , 500 -508 .
[55]
Schmitt J Antonovics J 1986 ). Experimental studies of the evolutionary significance of sexual reproduction. IV. Effect of neighbor relatedness and aphid infestation on seedling performance.
Evolution , 40 , 830 -836 .
[本文引用: 1]
[56]
Sellaro R Crepy M Trupkin SA Karayekov E Buchovsky AS Rossi C Casal JJ 2010 ). Cryptochrome as a sensor of the blue/green ratio of natural radiation in Arabidopsis.
Plant Physiology , 154 , 401 -409 .
[本文引用: 1]
[57]
Semchenko M Saar S Lepik A 2014 ). Plant root exudates mediate neighbour recognition and trigger complex behavioural changes.
New Phytologist , 204 , 631 -637 .
[本文引用: 2]
[58]
Smith H 2000 ). Phytochromes and light signal perception by plants—An emerging synthesis.
Nature , 407 , 585 -591 .
[本文引用: 1]
[59]
Smukalla S Caldara M Pochet N Beauvais A Guadagnini S Yan C Vinces MD Jansen A Prevost MC Latgé JP Fink GR Foster KR Verstrepen KJ 2008 ). FLO1 is a variable green beard gene that drives biofilm-like cooperation in budding yeast.
Cell , 135 , 726 -737 .
[本文引用: 1]
[60]
Tonsor SJ 1989 ). Relatedness and intraspecific competition in Plantago lanceolata.
The American Naturalist , 134 , 897 -906 .
[本文引用: 3]
[61]
Weiblen GD Webb CO Novotny V Basset Y Miller SE 2006 ). Phylogenetic dispersion of host use in a tropical insect herbivore community.
Ecology , 87 (sp7 ), S62 -S75 .
[本文引用: 1]
[62]
Willson MF Hoppes WG Goldman DA Thomas PA Katusic- Malmborg PL Bothwell JL 1987 ). Sibling competition in plants: an experimental study.
American Naturalist , 129 , 304 -311
[63]
Wu JQ Baldwin IT 2009 ). Herbivory-induced signalling in plants: Perception and action.
Plant, Cell & Environment , 32 , 1161 -1174 .
[本文引用: 1]
[64]
Xiao TY Lin WP Cai KZ 2015 ). Effects of intercropping soybean varieties with different kin relationships on the growth and nitrogen utilization efficiency.
Chinese Journal of Ecology , 34 , 2140 -2148 .
(in Chinese with English abstract) [肖桃艳 , 林威鹏 , 蔡昆争 (2013 ). 不同亲缘关系大豆品种间种对生长及氮素利用效率的影响.
生态学杂志 , 34 , 2140 -2148 .]
Inter- and intragenotypic competition under elevated carbon dioxide in Arabidopsis thaliana.
0
2001
Neighbor relatedness and competitive performance in Impatiens capensis (Balsaminaceae): A test of the resource partitioning hypothesis.
3
1992
... Willison等(1987)对垂序商陆(Phytolacca americana )、苘麻(Abutilon theophrasti )、Solanum mauritanianum 研究发现, 垂序商陆与近亲缘个体(来自同一母体)共存时, 具有更高的株高和种子数量, 而苘麻具有更高的幼苗存活率.Tonsor (1989)研究表明, 长叶车前(Plantago lanceolata )与近亲缘(克隆体)共处时, 形成更多的花序及坐果数.对Impatiens capensis 的研究也表明, 与近亲缘个体共处时, 目标个体提早了开花时间(Argyres & Schmitt, 1992 ).以上这些早期的研究认为, 与近亲缘个体共存时, 植物个体具有较高的繁殖效益, 恰好符合达尔文关于生物生存的最终目的是获得最大生殖效益的原则, 从正面证实了植物亲缘识别可能带来的好处. ...
... 不同研究对实验材料亲缘关系的界定方式和亲缘程度的选择也存在较大差异, 目前主要包括以下3种界定方式: 第一种以是否来自同一亲本为区分标准(Argyres & Schmitt, 1992 ; Dudley & File, 2007 ; Bhatt et al ., 2011 ; Semchenko et al ., 2014 ); 第二种以基因型为区分标准(Ninkovic, 2003 ; Biedrzycki et al ., 2010 ; Crepy & Casal, 2015 ); 第三种以人为克隆体为区分标准(Tonsor, 1989 ; Karban & Shiojiri, 2009 ; Mercer & Eppley, 2014 ).不同亲缘关系的界定方式, 也是导致目前研究结果差异较大的最主要原因.关于何种界定方式最为合适, 目前仍没有统一的定论.参考其他生物亲缘识别的相关研究, 动物主要以个体间血缘关系远近来界定个体间的亲缘关系(Emlen et al ., 1995 ), 而微生物主要以分裂方式进行繁殖, 故研究中主要以基因型或遗传距离来区分个体(菌群)之间的亲缘关系(Ostrowski et al ., 2008 ).目前植物亲缘识别研究主要以亲本关系及基因型(遗传距离)两种亲缘界定方式为主流, 且使用这两种方式的研究中, 均有支持和否定亲缘识别的结果, 所以这两种方式都可以得到相对准确的结果.因此, 选取何种亲缘界定方式最为合适仍很难判断, 在短期内可能仍会呈现多种界别方式共存的局面.为了尽量减少因采用不同亲缘界别方式所可能带来的误差, 建议后续实验中, 仅在采用同种亲缘界别方式的实验间进行结果比较. ...
... 研究表明, 植物面对周边个体, 不但能够产生一定的形态学响应, 而且在更深层次上还有许多“看不见”的生理生化策略的变化(Karban et al ., 2000 ; Baldwin et al ., 2006 ).对白车轴草(Trifolium repens ) (Lepik et al ., 2012 )、狭叶羽扇豆(Milla et al ., 2012 )的研究发现: 这两种植物分别与不同亲缘个体共存时, 其叶面积、株高、生物量、根长均无差异, 但未测定生理学指标, 因此还不能完全否定植物亲缘识别的存在.另外一个问题是指标上的以偏概全.部分研究采用某单一指标, 如侧根长(Dudley & File, 2007 , Biedrzycki et al ., 2010 )、生物量(Argyres & Schmitt, 1992 )、根冠比(Bhatt et al ., 2011 )存在的差异(其他指标可能没有差异)为标准, 判断植物会否存在亲缘识别.但是, 单一指标的差异并不能代表所有指标的趋势, 因此利用单一指标的差异表征亲缘识别现象的存在是以偏概全.因此, 建议在后续的实验中, 除了考察形态指标外, 还应该加大对植物生理生化指标的研究, 例如植株的养分利用能力, 次级代谢产物合成, 根系分泌物, 叶片挥发物, 由目前的以表观形态观察为主, 逐步转变到以植物形态变化与生理变化相结合的复合途径, 能够更加全面和直接地表征植物亲缘识别特征以及辅助对植物亲缘识别机理的研究. ...
Aggression, habituation, and clonal coexistence in the sea anemone Anthopleura elegantissima.
1
1995
... 许多研究表明, 在多种生物种中存在亲缘选择现象, 如海葵(Anthopleura elegant ) (Ayre & Grosberg, 1995 )、美新亮丽鲷(Neolamprologus pulcher ) (Dierkes et al ., 2005 )、蝾螈(Ambystoma tigrinum nebulosum ) (Pfennig & Collins, 1993 ), 白额食蜂鸟(Merops bullockoides ) (Emlen et al ., 1995 )、拜氏黄鼠(Spermophilus beldingi ) (Mateo, 2003 )、黑猩猩(Chimpanzee ) (Morin et al ., 1994 ).更有趣的是, 在对盘基网柄菌(Dictyostelium discoideum ) (Ostrowski et al ., 2008 )、酵母菌(Saccharomyces cerevisiae ) (Smukalla et al ., 2008 )等微生物的研究中, 同样发现类似亲缘选择现象.以上研究表明, 亲缘选择行为覆盖了肠腔类、鱼类、两栖类、鸟类、哺乳类, 乃至微生物等生物界的大部分类群. ...
Root secreted metabolites and proteins are involved in the early events of plant-plant recognition prior to competition.
0
2012
Shedding light on kin recognition response in plants.
0
2015
Volatile signaling in plant-plant interactions: “Talking trees” in the genomics era.
1
2006
... 研究表明, 植物面对周边个体, 不但能够产生一定的形态学响应, 而且在更深层次上还有许多“看不见”的生理生化策略的变化(Karban et al ., 2000 ; Baldwin et al ., 2006 ).对白车轴草(Trifolium repens ) (Lepik et al ., 2012 )、狭叶羽扇豆(Milla et al ., 2012 )的研究发现: 这两种植物分别与不同亲缘个体共存时, 其叶面积、株高、生物量、根长均无差异, 但未测定生理学指标, 因此还不能完全否定植物亲缘识别的存在.另外一个问题是指标上的以偏概全.部分研究采用某单一指标, 如侧根长(Dudley & File, 2007 , Biedrzycki et al ., 2010 )、生物量(Argyres & Schmitt, 1992 )、根冠比(Bhatt et al ., 2011 )存在的差异(其他指标可能没有差异)为标准, 判断植物会否存在亲缘识别.但是, 单一指标的差异并不能代表所有指标的趋势, 因此利用单一指标的差异表征亲缘识别现象的存在是以偏概全.因此, 建议在后续的实验中, 除了考察形态指标外, 还应该加大对植物生理生化指标的研究, 例如植株的养分利用能力, 次级代谢产物合成, 根系分泌物, 叶片挥发物, 由目前的以表观形态观察为主, 逐步转变到以植物形态变化与生理变化相结合的复合途径, 能够更加全面和直接地表征植物亲缘识别特征以及辅助对植物亲缘识别机理的研究. ...
Kin recognition, not competitive interactions, predicts root allocation in young Cakile edentula seedling pairs.
5
2011
... 自1980年, 植物亲缘识别概念提出至今, 科学家们已在包括Lycopersicon lycopersicum (Willison et al ., 1987 )、长叶车前(Plantago lanceolata ) (Tonsor, 1989 )、Cakile edentula (Donohue, 2003 ; Dudley & File, 2007 ; Bhatt et al ., 2011 )、大麦(Hordeum vulgare ) (Ninkovic, 2003 )、狭叶羽扇豆(Lupinus angustifolius ) (Milla et al ., 2012 )、拟南芥(Arabidopsis thaliana ) (Goodnight, 1985 ; Biedrzycki et al ., 2010 ; Masclaux et al ., 2010 ; Caffaro et al ., 2013 ; Crepy & Casal, 2015 )、水稻(Oryza sativa ) (Fang et al ., 2013 )、豚草(Ambrosia artemisiifolia ) (File et al ., 2012 )等20种植物中进行验证, 其中有许多研究结果支持植物存在亲缘识别 (表1 ), 也有一些研究得到否定的结果(表2 ). ...
... 相比于早期研究侧重繁殖性状, 近期研究则更侧重于对生长性状的探讨, 主要通过生物量、株高、叶面积、根冠比、根系长度等指标表征植物与不同亲缘个体共存时, 目标个体的生存特征, 从侧面表征植物的亲缘识别行为.对Cakile edentula 、大麦、拟南芥、Distichlis spicata 等的研究均表明, 植物与近亲缘个体共存时, 提高了自身生物量(Donohue, 2003 ; Bhatt et al ., 2011 ; Biernaskie, 2011 ; Mercer & Eppley, 2014 ).但也有研究表明与近亲缘个体共处时, 目标个体生物量反而可能降低(Cheplick & Kane, 2004 ; Milla et al ., 2009 ), 或者没有影响(Ninkovic, 2003 ).此外, Tonsor (1989)研究发现, 当长叶车前与近亲缘个体共存时, 干质量虽然下降, 但产生更多的子代, 表明与近亲缘个体共存时虽然生存效益下降, 但是繁殖效益提高.因此, 目前而言, 用生物量的大小来判断植物是否存在亲缘识别或者亲缘利他存在着一定的局限性. ...
... 其中大多数支持植物存在亲缘识别的研究 (表1 )认为, 目标植物与远亲缘共存时具有较大的根系, 从而加强竞争(Ninkovic, 2003 ; Dudley & File, 2007 ; Biedrzycki et al ., 2010 ), 而反对的结果(表2 )则表明, 与近亲缘个体共存时形成更多根系(Murphy & Dudley, 2009 )或者两者间并无差异(Milla et al ., 2012 ; Semchenko et al ., 2014 ).Semchenko等(2014)的研究结果则提供了另一种新的思路, 目标植物与不同亲缘个体共存时, 根系长度并无差异, 但显著改变根系的构型.由于根系长度无法准确地表征植物的亲缘识别特征, 因此, 有学者提出从根系生长趋性的角度进行分析(趋向近亲缘或者远亲缘).Caffaro等(2013)和Fang等(2013)的研究均表明, 目标植物根系更趋向于近亲缘个体方向生长.另外还有部分研究采用目标植物根冠比来表征其与周边亲缘邻居的竞争强度(Bhatt et al ., 2011 ; Mercer & Eppley, 2014 ).首先, 当目标植物面对不同亲缘邻居, 其总生物量的形成可能并不受影响, 但其可能通过改变地上部地下部分配(根冠比)而改变竞争策略; 其次, 目标植物根系减少, 并不一定意味着个体间竞争减弱, 其可能通过将更多能量投入到地上部生产或者加大次级代谢产物的合成, 增加了个体间的竞争压力.因此, 采用根冠比能在一定程度上弥补采用根系生物量和根系长度较片面表征个体间竞争强度的不足. ...
... 不同研究对实验材料亲缘关系的界定方式和亲缘程度的选择也存在较大差异, 目前主要包括以下3种界定方式: 第一种以是否来自同一亲本为区分标准(Argyres & Schmitt, 1992 ; Dudley & File, 2007 ; Bhatt et al ., 2011 ; Semchenko et al ., 2014 ); 第二种以基因型为区分标准(Ninkovic, 2003 ; Biedrzycki et al ., 2010 ; Crepy & Casal, 2015 ); 第三种以人为克隆体为区分标准(Tonsor, 1989 ; Karban & Shiojiri, 2009 ; Mercer & Eppley, 2014 ).不同亲缘关系的界定方式, 也是导致目前研究结果差异较大的最主要原因.关于何种界定方式最为合适, 目前仍没有统一的定论.参考其他生物亲缘识别的相关研究, 动物主要以个体间血缘关系远近来界定个体间的亲缘关系(Emlen et al ., 1995 ), 而微生物主要以分裂方式进行繁殖, 故研究中主要以基因型或遗传距离来区分个体(菌群)之间的亲缘关系(Ostrowski et al ., 2008 ).目前植物亲缘识别研究主要以亲本关系及基因型(遗传距离)两种亲缘界定方式为主流, 且使用这两种方式的研究中, 均有支持和否定亲缘识别的结果, 所以这两种方式都可以得到相对准确的结果.因此, 选取何种亲缘界定方式最为合适仍很难判断, 在短期内可能仍会呈现多种界别方式共存的局面.为了尽量减少因采用不同亲缘界别方式所可能带来的误差, 建议后续实验中, 仅在采用同种亲缘界别方式的实验间进行结果比较. ...
... 研究表明, 植物面对周边个体, 不但能够产生一定的形态学响应, 而且在更深层次上还有许多“看不见”的生理生化策略的变化(Karban et al ., 2000 ; Baldwin et al ., 2006 ).对白车轴草(Trifolium repens ) (Lepik et al ., 2012 )、狭叶羽扇豆(Milla et al ., 2012 )的研究发现: 这两种植物分别与不同亲缘个体共存时, 其叶面积、株高、生物量、根长均无差异, 但未测定生理学指标, 因此还不能完全否定植物亲缘识别的存在.另外一个问题是指标上的以偏概全.部分研究采用某单一指标, 如侧根长(Dudley & File, 2007 , Biedrzycki et al ., 2010 )、生物量(Argyres & Schmitt, 1992 )、根冠比(Bhatt et al ., 2011 )存在的差异(其他指标可能没有差异)为标准, 判断植物会否存在亲缘识别.但是, 单一指标的差异并不能代表所有指标的趋势, 因此利用单一指标的差异表征亲缘识别现象的存在是以偏概全.因此, 建议在后续的实验中, 除了考察形态指标外, 还应该加大对植物生理生化指标的研究, 例如植株的养分利用能力, 次级代谢产物合成, 根系分泌物, 叶片挥发物, 由目前的以表观形态观察为主, 逐步转变到以植物形态变化与生理变化相结合的复合途径, 能够更加全面和直接地表征植物亲缘识别特征以及辅助对植物亲缘识别机理的研究. ...
Kin recognition in plants: A mysterious behaviour unsolved.
6
2010
... 自1980年, 植物亲缘识别概念提出至今, 科学家们已在包括Lycopersicon lycopersicum (Willison et al ., 1987 )、长叶车前(Plantago lanceolata ) (Tonsor, 1989 )、Cakile edentula (Donohue, 2003 ; Dudley & File, 2007 ; Bhatt et al ., 2011 )、大麦(Hordeum vulgare ) (Ninkovic, 2003 )、狭叶羽扇豆(Lupinus angustifolius ) (Milla et al ., 2012 )、拟南芥(Arabidopsis thaliana ) (Goodnight, 1985 ; Biedrzycki et al ., 2010 ; Masclaux et al ., 2010 ; Caffaro et al ., 2013 ; Crepy & Casal, 2015 )、水稻(Oryza sativa ) (Fang et al ., 2013 )、豚草(Ambrosia artemisiifolia ) (File et al ., 2012 )等20种植物中进行验证, 其中有许多研究结果支持植物存在亲缘识别 (表1 ), 也有一些研究得到否定的结果(表2 ). ...
... 其中大多数支持植物存在亲缘识别的研究 (表1 )认为, 目标植物与远亲缘共存时具有较大的根系, 从而加强竞争(Ninkovic, 2003 ; Dudley & File, 2007 ; Biedrzycki et al ., 2010 ), 而反对的结果(表2 )则表明, 与近亲缘个体共存时形成更多根系(Murphy & Dudley, 2009 )或者两者间并无差异(Milla et al ., 2012 ; Semchenko et al ., 2014 ).Semchenko等(2014)的研究结果则提供了另一种新的思路, 目标植物与不同亲缘个体共存时, 根系长度并无差异, 但显著改变根系的构型.由于根系长度无法准确地表征植物的亲缘识别特征, 因此, 有学者提出从根系生长趋性的角度进行分析(趋向近亲缘或者远亲缘).Caffaro等(2013)和Fang等(2013)的研究均表明, 目标植物根系更趋向于近亲缘个体方向生长.另外还有部分研究采用目标植物根冠比来表征其与周边亲缘邻居的竞争强度(Bhatt et al ., 2011 ; Mercer & Eppley, 2014 ).首先, 当目标植物面对不同亲缘邻居, 其总生物量的形成可能并不受影响, 但其可能通过改变地上部地下部分配(根冠比)而改变竞争策略; 其次, 目标植物根系减少, 并不一定意味着个体间竞争减弱, 其可能通过将更多能量投入到地上部生产或者加大次级代谢产物的合成, 增加了个体间的竞争压力.因此, 采用根冠比能在一定程度上弥补采用根系生物量和根系长度较片面表征个体间竞争强度的不足. ...
... 现有研究已在包括拟南芥等至少20种植物上进行验证(表1 , 表2 ).实验采用的研究对象不一样, 不同植物本身亲缘识别的强度、途径、方式可能存在较大差异, 因此容易造成研究结果的差异(Biedrzycki & Bais, 2010 ).从植物分类学的角度看, 目前所报道的20个种分别来自2纲9目10科20属(表3 ).按照恩格勒植物分类系统, 这些种所覆盖的目, 诸如桔梗目、车前目、管状花目隶属于后生花被亚纲, 蔷薇目、罂粟目、无患子目、锦葵目隶属于异被花类, 中央子目和禾本目隶属于同被花类, 均属于被子植物, 而相对原始的裸子植物目前仍未见相关报道.从品种来源上看, 目前多数实验采用野生植物, 部分实验采用模式植物(拟南芥、水稻), 而经长期人工筛选的作物研究相对较少, 仅在大麦(Ninkovic, 2003 )、玉米(Zea may ) (Lin, 2013 )、大豆(Glycine max ) (肖桃艳等, 2015 )上有少量报道.因此, 建议在后续试验中加强向裸子植物和人工长期筛选作物两个方向延伸, 验证植物亲缘识别现象在原始的裸子植物以及长期人工选育作物(部分原始基因丢失)中是否仍然存在, 从而确定这种行为是否具有普遍性.此外, 目前的研究对象除Artemisia tridentate 外均为草本植物, 建议在后续研究中多关注灌木及木本植物. ...
... 不同研究对实验材料亲缘关系的界定方式和亲缘程度的选择也存在较大差异, 目前主要包括以下3种界定方式: 第一种以是否来自同一亲本为区分标准(Argyres & Schmitt, 1992 ; Dudley & File, 2007 ; Bhatt et al ., 2011 ; Semchenko et al ., 2014 ); 第二种以基因型为区分标准(Ninkovic, 2003 ; Biedrzycki et al ., 2010 ; Crepy & Casal, 2015 ); 第三种以人为克隆体为区分标准(Tonsor, 1989 ; Karban & Shiojiri, 2009 ; Mercer & Eppley, 2014 ).不同亲缘关系的界定方式, 也是导致目前研究结果差异较大的最主要原因.关于何种界定方式最为合适, 目前仍没有统一的定论.参考其他生物亲缘识别的相关研究, 动物主要以个体间血缘关系远近来界定个体间的亲缘关系(Emlen et al ., 1995 ), 而微生物主要以分裂方式进行繁殖, 故研究中主要以基因型或遗传距离来区分个体(菌群)之间的亲缘关系(Ostrowski et al ., 2008 ).目前植物亲缘识别研究主要以亲本关系及基因型(遗传距离)两种亲缘界定方式为主流, 且使用这两种方式的研究中, 均有支持和否定亲缘识别的结果, 所以这两种方式都可以得到相对准确的结果.因此, 选取何种亲缘界定方式最为合适仍很难判断, 在短期内可能仍会呈现多种界别方式共存的局面.为了尽量减少因采用不同亲缘界别方式所可能带来的误差, 建议后续实验中, 仅在采用同种亲缘界别方式的实验间进行结果比较. ...
... 研究表明, 植物面对周边个体, 不但能够产生一定的形态学响应, 而且在更深层次上还有许多“看不见”的生理生化策略的变化(Karban et al ., 2000 ; Baldwin et al ., 2006 ).对白车轴草(Trifolium repens ) (Lepik et al ., 2012 )、狭叶羽扇豆(Milla et al ., 2012 )的研究发现: 这两种植物分别与不同亲缘个体共存时, 其叶面积、株高、生物量、根长均无差异, 但未测定生理学指标, 因此还不能完全否定植物亲缘识别的存在.另外一个问题是指标上的以偏概全.部分研究采用某单一指标, 如侧根长(Dudley & File, 2007 , Biedrzycki et al ., 2010 )、生物量(Argyres & Schmitt, 1992 )、根冠比(Bhatt et al ., 2011 )存在的差异(其他指标可能没有差异)为标准, 判断植物会否存在亲缘识别.但是, 单一指标的差异并不能代表所有指标的趋势, 因此利用单一指标的差异表征亲缘识别现象的存在是以偏概全.因此, 建议在后续的实验中, 除了考察形态指标外, 还应该加大对植物生理生化指标的研究, 例如植株的养分利用能力, 次级代谢产物合成, 根系分泌物, 叶片挥发物, 由目前的以表观形态观察为主, 逐步转变到以植物形态变化与生理变化相结合的复合途径, 能够更加全面和直接地表征植物亲缘识别特征以及辅助对植物亲缘识别机理的研究. ...
... 前人已从不同角度, 对植物是否存在亲缘识别进行了很多有益的探索, 但目前对植物亲缘识别研究暂无统一模式, 不同研究对于亲缘关系的界定标准、实验材料、测定指标、环境因素均存在较大差异, 得出的结果也大相径庭, 不同科学家对植物亲缘识别是否存在也存在较大争议, 不过, 正是这种争议和矛盾, 不断地推动着该领域研究的发展, 使我们不断向真理靠近(Biedrzycki & Bais, 2010 ). ...
Root exudates mediate kin recognition in plants.
0
2010
Transcriptome analysis of Arabidopsis thaliana plants in response to kin and stranger recognition.
2
2011
... Biedrzycki等(2010)将拟南芥浸泡于含有不同亲缘个体根系分泌物的培养液中, 发现当暴露在近亲缘个体的根系分泌物时, 目标个体形成了较少的侧根, 而暴露在远亲缘个体根系分泌物中则形成较多侧根(图3A ), 这种现象在添加根系分泌物抑制剂后消失(图3B ).由此推测, 根系分泌物可能介导了植物亲缘识别, 但文中并没有提到具体的分泌物.此外, 这一选择性响应很有可能是由两个拟南芥品种根系分泌物中的某些特定的化学成分缺失导致的.Biedrzycki等(2011)后续进行的转录组分析发现, 目标植物暴露在远缘个体的根系分泌物后, 根系中ATP/GST转运载体、次级代谢合成、病程相关基因表达量上调.更有趣的是, 除了根系病程相关基因表达量上调外, 根系分泌物中的病程相关蛋白也相应增多(Badri et al ., 2012 ).这些结果是否暗示着植物亲缘识别和植物对于病原体的识别可能属于同一个途径, 植物将远亲缘个体按照病原体进行识别和对待, 该假设有待进一步验证.此外, 已有研究表明, 当目标植物与远缘个体共存时, 促进了目标个体根系的生长(初级代谢), 转录组分析表明, 根系中初级代谢相关基因的表达量并无明显上调(Biedrzycki et al ., 2011 ), 但次级代谢产物合成量增加, 这两个似乎是相互矛盾的结果. ...
... 指标选定方面主要存在两个问题.第一个是选用的指标范围多局限于形态学和生长发育指标.目前大部分研究主要采用诸如株高、叶面积、生物量、开花数、开花时间、坐果数、种子数量、种子萌发率、种子存活率、根系长度、根冠比、叶片重叠度、根系交叉度等形态学指标, 间接表征植物是否存在亲缘识别现象.还有少数研究采用植物代谢酶活性(肖桃艳等, 2015 )、植物光合作用和呼吸作用速率(Meier et al ., 2013 )、根系分泌物(Badri et al ., 2012 )等生理代谢指标, 菌根数量(File et al ., 2012 )等环境指标, 基因表达模式(Biedrzycki et al ., 2011 ; Masclaux et al ., 2010 )等指标, 来研究目标植物与不同亲缘个体共存时的表现. ...
Evidence for competition and cooperation among climbing plants.
1
2011
... 相比于早期研究侧重繁殖性状, 近期研究则更侧重于对生长性状的探讨, 主要通过生物量、株高、叶面积、根冠比、根系长度等指标表征植物与不同亲缘个体共存时, 目标个体的生存特征, 从侧面表征植物的亲缘识别行为.对Cakile edentula 、大麦、拟南芥、Distichlis spicata 等的研究均表明, 植物与近亲缘个体共存时, 提高了自身生物量(Donohue, 2003 ; Bhatt et al ., 2011 ; Biernaskie, 2011 ; Mercer & Eppley, 2014 ).但也有研究表明与近亲缘个体共处时, 目标个体生物量反而可能降低(Cheplick & Kane, 2004 ; Milla et al ., 2009 ), 或者没有影响(Ninkovic, 2003 ).此外, Tonsor (1989)研究发现, 当长叶车前与近亲缘个体共存时, 干质量虽然下降, 但产生更多的子代, 表明与近亲缘个体共存时虽然生存效益下降, 但是繁殖效益提高.因此, 目前而言, 用生物量的大小来判断植物是否存在亲缘识别或者亲缘利他存在着一定的局限性. ...
Plant neighbor identity influences plant biochemistry and physiology related to defense.
1
2010
... 对龙葵(Solanum nigrum ) (Schmidt & Balswin, 2006 )、Pinus halepensis (Ormeño et al ., 2007 )、Centaurea maculosa (Broz et al ., 2010 )、拟南芥(Badri et al ., 2012 )等的研究也发现同种个体共存植株体内抗性物质提高的现象, 这有力地支持了Karban等的观点. ...
More closely related species are more ecologically similar in an experimental test.
1
2011
... 一般认为, 同种或近亲缘植物共存, 因其生理生态特性高度相似, 所构成聚落物种单一, 更容易受到病虫害的危害(Schmitt & Antonovics, 1986 ; Weiblen et al ., 2006 ; Burns & Strauss, 2011 ; Liu et al ., 2012 ).但据Karban和Shiojiri (2009)、Karban等(2013)对Artemisia tridentata 的研究发现, 当植株受到机械损伤或者昆虫取食后, 能够向周边释放挥发物, 这些物质能够诱导周边植物的预警抗性, 而且近亲缘个体受诱导后, 抗性优于远亲缘个体, 表明亲缘识别可能影响植物的通讯与防御. ...
Root architecture of Arabidopsis is affected by competition with neighbouring plants.
1
2013
... 自1980年, 植物亲缘识别概念提出至今, 科学家们已在包括Lycopersicon lycopersicum (Willison et al ., 1987 )、长叶车前(Plantago lanceolata ) (Tonsor, 1989 )、Cakile edentula (Donohue, 2003 ; Dudley & File, 2007 ; Bhatt et al ., 2011 )、大麦(Hordeum vulgare ) (Ninkovic, 2003 )、狭叶羽扇豆(Lupinus angustifolius ) (Milla et al ., 2012 )、拟南芥(Arabidopsis thaliana ) (Goodnight, 1985 ; Biedrzycki et al ., 2010 ; Masclaux et al ., 2010 ; Caffaro et al ., 2013 ; Crepy & Casal, 2015 )、水稻(Oryza sativa ) (Fang et al ., 2013 )、豚草(Ambrosia artemisiifolia ) (File et al ., 2012 )等20种植物中进行验证, 其中有许多研究结果支持植物存在亲缘识别 (表1 ), 也有一些研究得到否定的结果(表2 ). ...
Photoreceptor signaling networks in plant responses to shade.
2
2013
... 植物光合色素对光吸收具有选择性, 投射到叶片上的红光和蓝光大部分均被吸收, 仅有少量的红光、红外光和蓝光能够穿透叶片, 透射到冠层下(Smith, 2000 ; Casal, 2013 ); 而冠层下的植物或者叶片进化出一种有效的机制, 能够通过冠层下红光与远红外光的比例, 判断周边的光照条件, 调整叶片的生长方向, 向光照充分的地方生长, 并减少叶片间的相互遮蔽(Casal, 2013 ).已有研究表明, 植物主要通过光敏色素B (phyB)和隐花色素(cry1)识别这两种光的微弱变化(Sellaro et al ., 2010 ; Keller et al ., 2011 ). ...
... ); 而冠层下的植物或者叶片进化出一种有效的机制, 能够通过冠层下红光与远红外光的比例, 判断周边的光照条件, 调整叶片的生长方向, 向光照充分的地方生长, 并减少叶片间的相互遮蔽(Casal, 2013 ).已有研究表明, 植物主要通过光敏色素B (phyB)和隐花色素(cry1)识别这两种光的微弱变化(Sellaro et al ., 2010 ; Keller et al ., 2011 ). ...
Genetic relatedness and competition in Triplasis purpurea (Poaceae): Resource partitioning or kin selection?
2
2004
... 相比于早期研究侧重繁殖性状, 近期研究则更侧重于对生长性状的探讨, 主要通过生物量、株高、叶面积、根冠比、根系长度等指标表征植物与不同亲缘个体共存时, 目标个体的生存特征, 从侧面表征植物的亲缘识别行为.对Cakile edentula 、大麦、拟南芥、Distichlis spicata 等的研究均表明, 植物与近亲缘个体共存时, 提高了自身生物量(Donohue, 2003 ; Bhatt et al ., 2011 ; Biernaskie, 2011 ; Mercer & Eppley, 2014 ).但也有研究表明与近亲缘个体共处时, 目标个体生物量反而可能降低(Cheplick & Kane, 2004 ; Milla et al ., 2009 ), 或者没有影响(Ninkovic, 2003 ).此外, Tonsor (1989)研究发现, 当长叶车前与近亲缘个体共存时, 干质量虽然下降, 但产生更多的子代, 表明与近亲缘个体共存时虽然生存效益下降, 但是繁殖效益提高.因此, 目前而言, 用生物量的大小来判断植物是否存在亲缘识别或者亲缘利他存在着一定的局限性. ...
... 种子(后代)数量、种子萌发率、幼苗成活率等指标也被许多研究所应用(Willison et al ., 1987 ; Tonsor, 1989 ; Cheplick & Kane, 2004 ; Milla et al ., 2009 ; Masclaux et al ., 2010 ), 因其能够最直观地表征植物能否因亲缘识别而带来直接效益, 即繁殖效益.Willison等(1987)及Tonsor (1989)的研究均表明, 植物个体与近亲缘共存时具有更高的繁殖收益. ...
Photoreceptor-mediated kin recognition in plants.
3
2015
... 自1980年, 植物亲缘识别概念提出至今, 科学家们已在包括Lycopersicon lycopersicum (Willison et al ., 1987 )、长叶车前(Plantago lanceolata ) (Tonsor, 1989 )、Cakile edentula (Donohue, 2003 ; Dudley & File, 2007 ; Bhatt et al ., 2011 )、大麦(Hordeum vulgare ) (Ninkovic, 2003 )、狭叶羽扇豆(Lupinus angustifolius ) (Milla et al ., 2012 )、拟南芥(Arabidopsis thaliana ) (Goodnight, 1985 ; Biedrzycki et al ., 2010 ; Masclaux et al ., 2010 ; Caffaro et al ., 2013 ; Crepy & Casal, 2015 )、水稻(Oryza sativa ) (Fang et al ., 2013 )、豚草(Ambrosia artemisiifolia ) (File et al ., 2012 )等20种植物中进行验证, 其中有许多研究结果支持植物存在亲缘识别 (表1 ), 也有一些研究得到否定的结果(表2 ). ...
... 不同研究对实验材料亲缘关系的界定方式和亲缘程度的选择也存在较大差异, 目前主要包括以下3种界定方式: 第一种以是否来自同一亲本为区分标准(Argyres & Schmitt, 1992 ; Dudley & File, 2007 ; Bhatt et al ., 2011 ; Semchenko et al ., 2014 ); 第二种以基因型为区分标准(Ninkovic, 2003 ; Biedrzycki et al ., 2010 ; Crepy & Casal, 2015 ); 第三种以人为克隆体为区分标准(Tonsor, 1989 ; Karban & Shiojiri, 2009 ; Mercer & Eppley, 2014 ).不同亲缘关系的界定方式, 也是导致目前研究结果差异较大的最主要原因.关于何种界定方式最为合适, 目前仍没有统一的定论.参考其他生物亲缘识别的相关研究, 动物主要以个体间血缘关系远近来界定个体间的亲缘关系(Emlen et al ., 1995 ), 而微生物主要以分裂方式进行繁殖, 故研究中主要以基因型或遗传距离来区分个体(菌群)之间的亲缘关系(Ostrowski et al ., 2008 ).目前植物亲缘识别研究主要以亲本关系及基因型(遗传距离)两种亲缘界定方式为主流, 且使用这两种方式的研究中, 均有支持和否定亲缘识别的结果, 所以这两种方式都可以得到相对准确的结果.因此, 选取何种亲缘界定方式最为合适仍很难判断, 在短期内可能仍会呈现多种界别方式共存的局面.为了尽量减少因采用不同亲缘界别方式所可能带来的误差, 建议后续实验中, 仅在采用同种亲缘界别方式的实验间进行结果比较. ...
... 未来的研究, 应对亲缘关系的界定、研究材料的选择及实验设计有一个相对统一的标准.除了阐明不同亲缘物种共存的形态学和表型性状外, 从植物生理生化、分子、蛋白质组和代谢组学水平上解析植物亲缘识别的机制将是下一阶段研究的重点, 揭示植物亲缘识别的主要途径、介导物质以及植物接收信息的载体将是研究其机理的主要工作(Crepy & Casal, 2015 ); 同时应该考虑不同环境条件对亲缘识别的影响.希望通过广大科研工作者的努力, 更加完整地阐明植物亲缘识别行为, 揭示这一有趣的自然进化奥秘. ...
Arguments for and against self and non-self root recognition in plants.
1
2014
... Root growth model of target plants responded to kin neighbors. Modified from Depuydt (2014) . ...
Genetic relatedness in groups is sex-specific and declines with age of helpers in a cooperatively breeding cichlid.
1
2005
... 许多研究表明, 在多种生物种中存在亲缘选择现象, 如海葵(Anthopleura elegant ) (Ayre & Grosberg, 1995 )、美新亮丽鲷(Neolamprologus pulcher ) (Dierkes et al ., 2005 )、蝾螈(Ambystoma tigrinum nebulosum ) (Pfennig & Collins, 1993 ), 白额食蜂鸟(Merops bullockoides ) (Emlen et al ., 1995 )、拜氏黄鼠(Spermophilus beldingi ) (Mateo, 2003 )、黑猩猩(Chimpanzee ) (Morin et al ., 1994 ).更有趣的是, 在对盘基网柄菌(Dictyostelium discoideum ) (Ostrowski et al ., 2008 )、酵母菌(Saccharomyces cerevisiae ) (Smukalla et al ., 2008 )等微生物的研究中, 同样发现类似亲缘选择现象.以上研究表明, 亲缘选择行为覆盖了肠腔类、鱼类、两栖类、鸟类、哺乳类, 乃至微生物等生物界的大部分类群. ...
The influence of neighbor relatedness on multilevel selection in the Great Lakes sea rocket.
2
2003
... 自1980年, 植物亲缘识别概念提出至今, 科学家们已在包括Lycopersicon lycopersicum (Willison et al ., 1987 )、长叶车前(Plantago lanceolata ) (Tonsor, 1989 )、Cakile edentula (Donohue, 2003 ; Dudley & File, 2007 ; Bhatt et al ., 2011 )、大麦(Hordeum vulgare ) (Ninkovic, 2003 )、狭叶羽扇豆(Lupinus angustifolius ) (Milla et al ., 2012 )、拟南芥(Arabidopsis thaliana ) (Goodnight, 1985 ; Biedrzycki et al ., 2010 ; Masclaux et al ., 2010 ; Caffaro et al ., 2013 ; Crepy & Casal, 2015 )、水稻(Oryza sativa ) (Fang et al ., 2013 )、豚草(Ambrosia artemisiifolia ) (File et al ., 2012 )等20种植物中进行验证, 其中有许多研究结果支持植物存在亲缘识别 (表1 ), 也有一些研究得到否定的结果(表2 ). ...
... 相比于早期研究侧重繁殖性状, 近期研究则更侧重于对生长性状的探讨, 主要通过生物量、株高、叶面积、根冠比、根系长度等指标表征植物与不同亲缘个体共存时, 目标个体的生存特征, 从侧面表征植物的亲缘识别行为.对Cakile edentula 、大麦、拟南芥、Distichlis spicata 等的研究均表明, 植物与近亲缘个体共存时, 提高了自身生物量(Donohue, 2003 ; Bhatt et al ., 2011 ; Biernaskie, 2011 ; Mercer & Eppley, 2014 ).但也有研究表明与近亲缘个体共处时, 目标个体生物量反而可能降低(Cheplick & Kane, 2004 ; Milla et al ., 2009 ), 或者没有影响(Ninkovic, 2003 ).此外, Tonsor (1989)研究发现, 当长叶车前与近亲缘个体共存时, 干质量虽然下降, 但产生更多的子代, 表明与近亲缘个体共存时虽然生存效益下降, 但是繁殖效益提高.因此, 目前而言, 用生物量的大小来判断植物是否存在亲缘识别或者亲缘利他存在着一定的局限性. ...
Kin recognition in an annual plant.
5
2007
... 2007年, 加拿大科学家Dudley发表了一篇题为“Kin recognize in an annual plant”的文章(Dudley & File, 2007 ).该研究发现, 一年生植物Cakile edentula 能够识别周边其他同类个体与自己的亲缘关系, 并根据邻居亲缘关系的远近, 调整自身形态策略: 与近亲缘个体共存时, 侧根较多, 而与远亲缘个体共存时侧根较少.Dudley以此推测植物能够识别周边植物的亲缘关系, 当遇到近亲缘个体时, 采取较为缓和的竞争策略; 而当面临远亲缘个体或者非亲缘个体时, 则采用激烈的竞争策略.该文快速吸引了大量科学家的关注, 后续的许多研究也陆续发表在《Ecology Letters》、《New Phytologist》、《Journal of Experimental Botany》、《Functional Ecology》、《BMC Plant Biology》、《Planta》等国际主流刊物上. ...
... 自1980年, 植物亲缘识别概念提出至今, 科学家们已在包括Lycopersicon lycopersicum (Willison et al ., 1987 )、长叶车前(Plantago lanceolata ) (Tonsor, 1989 )、Cakile edentula (Donohue, 2003 ; Dudley & File, 2007 ; Bhatt et al ., 2011 )、大麦(Hordeum vulgare ) (Ninkovic, 2003 )、狭叶羽扇豆(Lupinus angustifolius ) (Milla et al ., 2012 )、拟南芥(Arabidopsis thaliana ) (Goodnight, 1985 ; Biedrzycki et al ., 2010 ; Masclaux et al ., 2010 ; Caffaro et al ., 2013 ; Crepy & Casal, 2015 )、水稻(Oryza sativa ) (Fang et al ., 2013 )、豚草(Ambrosia artemisiifolia ) (File et al ., 2012 )等20种植物中进行验证, 其中有许多研究结果支持植物存在亲缘识别 (表1 ), 也有一些研究得到否定的结果(表2 ). ...
... 其中大多数支持植物存在亲缘识别的研究 (表1 )认为, 目标植物与远亲缘共存时具有较大的根系, 从而加强竞争(Ninkovic, 2003 ; Dudley & File, 2007 ; Biedrzycki et al ., 2010 ), 而反对的结果(表2 )则表明, 与近亲缘个体共存时形成更多根系(Murphy & Dudley, 2009 )或者两者间并无差异(Milla et al ., 2012 ; Semchenko et al ., 2014 ).Semchenko等(2014)的研究结果则提供了另一种新的思路, 目标植物与不同亲缘个体共存时, 根系长度并无差异, 但显著改变根系的构型.由于根系长度无法准确地表征植物的亲缘识别特征, 因此, 有学者提出从根系生长趋性的角度进行分析(趋向近亲缘或者远亲缘).Caffaro等(2013)和Fang等(2013)的研究均表明, 目标植物根系更趋向于近亲缘个体方向生长.另外还有部分研究采用目标植物根冠比来表征其与周边亲缘邻居的竞争强度(Bhatt et al ., 2011 ; Mercer & Eppley, 2014 ).首先, 当目标植物面对不同亲缘邻居, 其总生物量的形成可能并不受影响, 但其可能通过改变地上部地下部分配(根冠比)而改变竞争策略; 其次, 目标植物根系减少, 并不一定意味着个体间竞争减弱, 其可能通过将更多能量投入到地上部生产或者加大次级代谢产物的合成, 增加了个体间的竞争压力.因此, 采用根冠比能在一定程度上弥补采用根系生物量和根系长度较片面表征个体间竞争强度的不足. ...
... 不同研究对实验材料亲缘关系的界定方式和亲缘程度的选择也存在较大差异, 目前主要包括以下3种界定方式: 第一种以是否来自同一亲本为区分标准(Argyres & Schmitt, 1992 ; Dudley & File, 2007 ; Bhatt et al ., 2011 ; Semchenko et al ., 2014 ); 第二种以基因型为区分标准(Ninkovic, 2003 ; Biedrzycki et al ., 2010 ; Crepy & Casal, 2015 ); 第三种以人为克隆体为区分标准(Tonsor, 1989 ; Karban & Shiojiri, 2009 ; Mercer & Eppley, 2014 ).不同亲缘关系的界定方式, 也是导致目前研究结果差异较大的最主要原因.关于何种界定方式最为合适, 目前仍没有统一的定论.参考其他生物亲缘识别的相关研究, 动物主要以个体间血缘关系远近来界定个体间的亲缘关系(Emlen et al ., 1995 ), 而微生物主要以分裂方式进行繁殖, 故研究中主要以基因型或遗传距离来区分个体(菌群)之间的亲缘关系(Ostrowski et al ., 2008 ).目前植物亲缘识别研究主要以亲本关系及基因型(遗传距离)两种亲缘界定方式为主流, 且使用这两种方式的研究中, 均有支持和否定亲缘识别的结果, 所以这两种方式都可以得到相对准确的结果.因此, 选取何种亲缘界定方式最为合适仍很难判断, 在短期内可能仍会呈现多种界别方式共存的局面.为了尽量减少因采用不同亲缘界别方式所可能带来的误差, 建议后续实验中, 仅在采用同种亲缘界别方式的实验间进行结果比较. ...
... 研究表明, 植物面对周边个体, 不但能够产生一定的形态学响应, 而且在更深层次上还有许多“看不见”的生理生化策略的变化(Karban et al ., 2000 ; Baldwin et al ., 2006 ).对白车轴草(Trifolium repens ) (Lepik et al ., 2012 )、狭叶羽扇豆(Milla et al ., 2012 )的研究发现: 这两种植物分别与不同亲缘个体共存时, 其叶面积、株高、生物量、根长均无差异, 但未测定生理学指标, 因此还不能完全否定植物亲缘识别的存在.另外一个问题是指标上的以偏概全.部分研究采用某单一指标, 如侧根长(Dudley & File, 2007 , Biedrzycki et al ., 2010 )、生物量(Argyres & Schmitt, 1992 )、根冠比(Bhatt et al ., 2011 )存在的差异(其他指标可能没有差异)为标准, 判断植物会否存在亲缘识别.但是, 单一指标的差异并不能代表所有指标的趋势, 因此利用单一指标的差异表征亲缘识别现象的存在是以偏概全.因此, 建议在后续的实验中, 除了考察形态指标外, 还应该加大对植物生理生化指标的研究, 例如植株的养分利用能力, 次级代谢产物合成, 根系分泌物, 叶片挥发物, 由目前的以表观形态观察为主, 逐步转变到以植物形态变化与生理变化相结合的复合途径, 能够更加全面和直接地表征植物亲缘识别特征以及辅助对植物亲缘识别机理的研究. ...
Making decisions in the family: An evolutionary perspective.
2
1995
... 许多研究表明, 在多种生物种中存在亲缘选择现象, 如海葵(Anthopleura elegant ) (Ayre & Grosberg, 1995 )、美新亮丽鲷(Neolamprologus pulcher ) (Dierkes et al ., 2005 )、蝾螈(Ambystoma tigrinum nebulosum ) (Pfennig & Collins, 1993 ), 白额食蜂鸟(Merops bullockoides ) (Emlen et al ., 1995 )、拜氏黄鼠(Spermophilus beldingi ) (Mateo, 2003 )、黑猩猩(Chimpanzee ) (Morin et al ., 1994 ).更有趣的是, 在对盘基网柄菌(Dictyostelium discoideum ) (Ostrowski et al ., 2008 )、酵母菌(Saccharomyces cerevisiae ) (Smukalla et al ., 2008 )等微生物的研究中, 同样发现类似亲缘选择现象.以上研究表明, 亲缘选择行为覆盖了肠腔类、鱼类、两栖类、鸟类、哺乳类, 乃至微生物等生物界的大部分类群. ...
... 不同研究对实验材料亲缘关系的界定方式和亲缘程度的选择也存在较大差异, 目前主要包括以下3种界定方式: 第一种以是否来自同一亲本为区分标准(Argyres & Schmitt, 1992 ; Dudley & File, 2007 ; Bhatt et al ., 2011 ; Semchenko et al ., 2014 ); 第二种以基因型为区分标准(Ninkovic, 2003 ; Biedrzycki et al ., 2010 ; Crepy & Casal, 2015 ); 第三种以人为克隆体为区分标准(Tonsor, 1989 ; Karban & Shiojiri, 2009 ; Mercer & Eppley, 2014 ).不同亲缘关系的界定方式, 也是导致目前研究结果差异较大的最主要原因.关于何种界定方式最为合适, 目前仍没有统一的定论.参考其他生物亲缘识别的相关研究, 动物主要以个体间血缘关系远近来界定个体间的亲缘关系(Emlen et al ., 1995 ), 而微生物主要以分裂方式进行繁殖, 故研究中主要以基因型或遗传距离来区分个体(菌群)之间的亲缘关系(Ostrowski et al ., 2008 ).目前植物亲缘识别研究主要以亲本关系及基因型(遗传距离)两种亲缘界定方式为主流, 且使用这两种方式的研究中, 均有支持和否定亲缘识别的结果, 所以这两种方式都可以得到相对准确的结果.因此, 选取何种亲缘界定方式最为合适仍很难判断, 在短期内可能仍会呈现多种界别方式共存的局面.为了尽量减少因采用不同亲缘界别方式所可能带来的误差, 建议后续实验中, 仅在采用同种亲缘界别方式的实验间进行结果比较. ...
Genotypic recognition and spatial responses by rice roots.
1
2013
... 自1980年, 植物亲缘识别概念提出至今, 科学家们已在包括Lycopersicon lycopersicum (Willison et al ., 1987 )、长叶车前(Plantago lanceolata ) (Tonsor, 1989 )、Cakile edentula (Donohue, 2003 ; Dudley & File, 2007 ; Bhatt et al ., 2011 )、大麦(Hordeum vulgare ) (Ninkovic, 2003 )、狭叶羽扇豆(Lupinus angustifolius ) (Milla et al ., 2012 )、拟南芥(Arabidopsis thaliana ) (Goodnight, 1985 ; Biedrzycki et al ., 2010 ; Masclaux et al ., 2010 ; Caffaro et al ., 2013 ; Crepy & Casal, 2015 )、水稻(Oryza sativa ) (Fang et al ., 2013 )、豚草(Ambrosia artemisiifolia ) (File et al ., 2012 )等20种植物中进行验证, 其中有许多研究结果支持植物存在亲缘识别 (表1 ), 也有一些研究得到否定的结果(表2 ). ...
Fitness consequences of plants growing with siblings: Reconciling kin selection, niche partitioning and competitive ability.
1
2011
... 外界环境的优劣和资源的丰贫对植物生长具有重要的影响, 植物能够感知这些差异并通过一定的形态和生理策略响应这些外界因素(例如根系的向肥性和叶片的向光性等).目前, 大部分植物亲缘识别的研究侧重于植物亲缘识别本身, 较少关注环境因素的影响.Andalo等(2001)研究发现, 在低CO2 浓度下, 近亲缘个体共存形成较多的果实和种子, 而CO2 浓度提高后, 远亲缘组形成更多的果实和种子.这表明环境因子对植物亲缘识别的影响可能要比原来想象的更加复杂.另外, 根据Hamilton (1964)定律, 亲缘识别(选择)在一定的条件下才能发生.在外界环境因素优越的条件下, 个体生存得到满足的情况下, 才表现亲缘合作趋向, 而在资源短缺时, 个体生存压力巨大, 则可能呈现竞争趋向(File et al ., 2011 ).因此, 不同实验, 对于水分、养分、光照、湿度设定差异, 也是造成相互矛盾结果的原因之一.鉴于目前植物亲缘识别研究仍处于初级阶段, 且大部分研究主要采用室内盆栽实验, 环境因素可控, 可能采用富余养分的实验设计相对合适.后续还可以开展相关环境因素对植物亲缘识别影响的研究, 以及探索野外条件下环境限制因子相对较多条件下植物的亲缘识别行为. ...
Plant kin recognition enhances abundance of symbiotic microbial partner.
2
2012
... 自1980年, 植物亲缘识别概念提出至今, 科学家们已在包括Lycopersicon lycopersicum (Willison et al ., 1987 )、长叶车前(Plantago lanceolata ) (Tonsor, 1989 )、Cakile edentula (Donohue, 2003 ; Dudley & File, 2007 ; Bhatt et al ., 2011 )、大麦(Hordeum vulgare ) (Ninkovic, 2003 )、狭叶羽扇豆(Lupinus angustifolius ) (Milla et al ., 2012 )、拟南芥(Arabidopsis thaliana ) (Goodnight, 1985 ; Biedrzycki et al ., 2010 ; Masclaux et al ., 2010 ; Caffaro et al ., 2013 ; Crepy & Casal, 2015 )、水稻(Oryza sativa ) (Fang et al ., 2013 )、豚草(Ambrosia artemisiifolia ) (File et al ., 2012 )等20种植物中进行验证, 其中有许多研究结果支持植物存在亲缘识别 (表1 ), 也有一些研究得到否定的结果(表2 ). ...
... 指标选定方面主要存在两个问题.第一个是选用的指标范围多局限于形态学和生长发育指标.目前大部分研究主要采用诸如株高、叶面积、生物量、开花数、开花时间、坐果数、种子数量、种子萌发率、种子存活率、根系长度、根冠比、叶片重叠度、根系交叉度等形态学指标, 间接表征植物是否存在亲缘识别现象.还有少数研究采用植物代谢酶活性(肖桃艳等, 2015 )、植物光合作用和呼吸作用速率(Meier et al ., 2013 )、根系分泌物(Badri et al ., 2012 )等生理代谢指标, 菌根数量(File et al ., 2012 )等环境指标, 基因表达模式(Biedrzycki et al ., 2011 ; Masclaux et al ., 2010 )等指标, 来研究目标植物与不同亲缘个体共存时的表现. ...
The influence of environmental variation on group and individual selection in a cress.
1
1985
... 自1980年, 植物亲缘识别概念提出至今, 科学家们已在包括Lycopersicon lycopersicum (Willison et al ., 1987 )、长叶车前(Plantago lanceolata ) (Tonsor, 1989 )、Cakile edentula (Donohue, 2003 ; Dudley & File, 2007 ; Bhatt et al ., 2011 )、大麦(Hordeum vulgare ) (Ninkovic, 2003 )、狭叶羽扇豆(Lupinus angustifolius ) (Milla et al ., 2012 )、拟南芥(Arabidopsis thaliana ) (Goodnight, 1985 ; Biedrzycki et al ., 2010 ; Masclaux et al ., 2010 ; Caffaro et al ., 2013 ; Crepy & Casal, 2015 )、水稻(Oryza sativa ) (Fang et al ., 2013 )、豚草(Ambrosia artemisiifolia ) (File et al ., 2012 )等20种植物中进行验证, 其中有许多研究结果支持植物存在亲缘识别 (表1 ), 也有一些研究得到否定的结果(表2 ). ...
The genetical evolution of social behaviour. II.
1
1964
... 关于亲缘识别的意义, 必须关联到一个重要的进化理论——亲缘选择理论.达尔文进化学说认为, 生物生存的最终目的是繁殖后代, 在恶劣的环境中生存下来, 实现最大的生殖效益.因此, 生物的一切行为就必须是自私自利的.但是, 自然界中存在一个“反常”现象困扰着达尔文: 在白蚁等社会性昆虫群体中, 存在工蚁为了保全整个蚁群, 不惜牺牲自己的行为, 称之为利他行为(altruism).直至1963年, 剑桥大学动物学家Hamilton W. D.提出利他行为的亲缘选择学说(kin selection) (Hamilton, 1964 ), 并运用广义适合度(inclusive fitness)解释动物的利他行为.该理论认为, 达尔文理论中生物繁殖的目的实质上就是自身基因的延续, 而近亲缘个体间共享更多的基因, 因此帮助近亲缘个体更好地生存并繁殖更多的后代, 同样达到了延续共有基因的目的. ...
The plastic plant: Root responses to heterogeneous supplies of nutrients.
1
2004
... 研究表明, 植物能够识别土壤环境的养分条件, 将根系向资源丰富区域集中, 而减少向资源匮乏地区的增殖, 表明植物根系的生长和分布是由土壤中的营养物质浓度所驱动的(Hodge, 2004 ).但近年研究发现, 一些植物的根系生长和分布并不是受土壤营养水平驱动的, 而是更多受到邻近植物根系的影响(Hodge, 2009 ; Mommer et al ., 2010 ), 植物的亲缘识别可能正属于这种非养分驱动行为. ...
Root decisions.
1
2009
... 研究表明, 植物能够识别土壤环境的养分条件, 将根系向资源丰富区域集中, 而减少向资源匮乏地区的增殖, 表明植物根系的生长和分布是由土壤中的营养物质浓度所驱动的(Hodge, 2004 ).但近年研究发现, 一些植物的根系生长和分布并不是受土壤营养水平驱动的, 而是更多受到邻近植物根系的影响(Hodge, 2009 ; Mommer et al ., 2010 ), 植物的亲缘识别可能正属于这种非养分驱动行为. ...
Communication between plants: Induced resistance in wild tobacco plants following clipping of neighboring sagebrush.
1
2000
... 研究表明, 植物面对周边个体, 不但能够产生一定的形态学响应, 而且在更深层次上还有许多“看不见”的生理生化策略的变化(Karban et al ., 2000 ; Baldwin et al ., 2006 ).对白车轴草(Trifolium repens ) (Lepik et al ., 2012 )、狭叶羽扇豆(Milla et al ., 2012 )的研究发现: 这两种植物分别与不同亲缘个体共存时, 其叶面积、株高、生物量、根长均无差异, 但未测定生理学指标, 因此还不能完全否定植物亲缘识别的存在.另外一个问题是指标上的以偏概全.部分研究采用某单一指标, 如侧根长(Dudley & File, 2007 , Biedrzycki et al ., 2010 )、生物量(Argyres & Schmitt, 1992 )、根冠比(Bhatt et al ., 2011 )存在的差异(其他指标可能没有差异)为标准, 判断植物会否存在亲缘识别.但是, 单一指标的差异并不能代表所有指标的趋势, 因此利用单一指标的差异表征亲缘识别现象的存在是以偏概全.因此, 建议在后续的实验中, 除了考察形态指标外, 还应该加大对植物生理生化指标的研究, 例如植株的养分利用能力, 次级代谢产物合成, 根系分泌物, 叶片挥发物, 由目前的以表观形态观察为主, 逐步转变到以植物形态变化与生理变化相结合的复合途径, 能够更加全面和直接地表征植物亲缘识别特征以及辅助对植物亲缘识别机理的研究. ...
Self-recognition affects plant communication and defense.
1
2009
... 不同研究对实验材料亲缘关系的界定方式和亲缘程度的选择也存在较大差异, 目前主要包括以下3种界定方式: 第一种以是否来自同一亲本为区分标准(Argyres & Schmitt, 1992 ; Dudley & File, 2007 ; Bhatt et al ., 2011 ; Semchenko et al ., 2014 ); 第二种以基因型为区分标准(Ninkovic, 2003 ; Biedrzycki et al ., 2010 ; Crepy & Casal, 2015 ); 第三种以人为克隆体为区分标准(Tonsor, 1989 ; Karban & Shiojiri, 2009 ; Mercer & Eppley, 2014 ).不同亲缘关系的界定方式, 也是导致目前研究结果差异较大的最主要原因.关于何种界定方式最为合适, 目前仍没有统一的定论.参考其他生物亲缘识别的相关研究, 动物主要以个体间血缘关系远近来界定个体间的亲缘关系(Emlen et al ., 1995 ), 而微生物主要以分裂方式进行繁殖, 故研究中主要以基因型或遗传距离来区分个体(菌群)之间的亲缘关系(Ostrowski et al ., 2008 ).目前植物亲缘识别研究主要以亲本关系及基因型(遗传距离)两种亲缘界定方式为主流, 且使用这两种方式的研究中, 均有支持和否定亲缘识别的结果, 所以这两种方式都可以得到相对准确的结果.因此, 选取何种亲缘界定方式最为合适仍很难判断, 在短期内可能仍会呈现多种界别方式共存的局面.为了尽量减少因采用不同亲缘界别方式所可能带来的误差, 建议后续实验中, 仅在采用同种亲缘界别方式的实验间进行结果比较. ...
Kin recognition affects plant communication and defence.
1
2013
... Karban和Shiojiri (2009)、Karban等(2013)的研究有利地支持了上述观点, 研究结果表明, 当Arte- misia tridentate 受到机械损伤或者昆虫取食后所释放的警报挥发物能够提高周边个体的抗虫性, 而且近亲缘个体抗虫性的提高程度显著高于远亲缘个体, 作者猜测这可能与挥发物的组成有关.其进一步分析发现, Artemisia tridentate 挥发物的组成与其亲缘关系显著相关, 即亲缘关系越接近, 其叶片挥发物的组成越相似.因此, 其认为来源于近亲缘个体的警报信号彼此间的成分更加相似, 因此受体植物更容易识别这些信息(Karban et al ., 2013 ). ...
Cryptochrome 1 and phytochrome B control shade-avoidance responses in Arabidopsis via partially independent hormonal cascades.
1
2011
... 植物光合色素对光吸收具有选择性, 投射到叶片上的红光和蓝光大部分均被吸收, 仅有少量的红光、红外光和蓝光能够穿透叶片, 透射到冠层下(Smith, 2000 ; Casal, 2013 ); 而冠层下的植物或者叶片进化出一种有效的机制, 能够通过冠层下红光与远红外光的比例, 判断周边的光照条件, 调整叶片的生长方向, 向光照充分的地方生长, 并减少叶片间的相互遮蔽(Casal, 2013 ).已有研究表明, 植物主要通过光敏色素B (phyB)和隐花色素(cry1)识别这两种光的微弱变化(Sellaro et al ., 2010 ; Keller et al ., 2011 ). ...
Kin recognition is density-dependent and uncommon among temperate grassland plants.
1
2012
... 研究表明, 植物面对周边个体, 不但能够产生一定的形态学响应, 而且在更深层次上还有许多“看不见”的生理生化策略的变化(Karban et al ., 2000 ; Baldwin et al ., 2006 ).对白车轴草(Trifolium repens ) (Lepik et al ., 2012 )、狭叶羽扇豆(Milla et al ., 2012 )的研究发现: 这两种植物分别与不同亲缘个体共存时, 其叶面积、株高、生物量、根长均无差异, 但未测定生理学指标, 因此还不能完全否定植物亲缘识别的存在.另外一个问题是指标上的以偏概全.部分研究采用某单一指标, 如侧根长(Dudley & File, 2007 , Biedrzycki et al ., 2010 )、生物量(Argyres & Schmitt, 1992 )、根冠比(Bhatt et al ., 2011 )存在的差异(其他指标可能没有差异)为标准, 判断植物会否存在亲缘识别.但是, 单一指标的差异并不能代表所有指标的趋势, 因此利用单一指标的差异表征亲缘识别现象的存在是以偏概全.因此, 建议在后续的实验中, 除了考察形态指标外, 还应该加大对植物生理生化指标的研究, 例如植株的养分利用能力, 次级代谢产物合成, 根系分泌物, 叶片挥发物, 由目前的以表观形态观察为主, 逐步转变到以植物形态变化与生理变化相结合的复合途径, 能够更加全面和直接地表征植物亲缘识别特征以及辅助对植物亲缘识别机理的研究. ...
利用玉米检验植物亲缘利他假说的初探
1
2013
... 现有研究已在包括拟南芥等至少20种植物上进行验证(表1 , 表2 ).实验采用的研究对象不一样, 不同植物本身亲缘识别的强度、途径、方式可能存在较大差异, 因此容易造成研究结果的差异(Biedrzycki & Bais, 2010 ).从植物分类学的角度看, 目前所报道的20个种分别来自2纲9目10科20属(表3 ).按照恩格勒植物分类系统, 这些种所覆盖的目, 诸如桔梗目、车前目、管状花目隶属于后生花被亚纲, 蔷薇目、罂粟目、无患子目、锦葵目隶属于异被花类, 中央子目和禾本目隶属于同被花类, 均属于被子植物, 而相对原始的裸子植物目前仍未见相关报道.从品种来源上看, 目前多数实验采用野生植物, 部分实验采用模式植物(拟南芥、水稻), 而经长期人工筛选的作物研究相对较少, 仅在大麦(Ninkovic, 2003 )、玉米(Zea may ) (Lin, 2013 )、大豆(Glycine max ) (肖桃艳等, 2015 )上有少量报道.因此, 建议在后续试验中加强向裸子植物和人工长期筛选作物两个方向延伸, 验证植物亲缘识别现象在原始的裸子植物以及长期人工选育作物(部分原始基因丢失)中是否仍然存在, 从而确定这种行为是否具有普遍性.此外, 目前的研究对象除Artemisia tridentate 外均为草本植物, 建议在后续研究中多关注灌木及木本植物. ...
利用玉米检验植物亲缘利他假说的初探
1
2013
... 现有研究已在包括拟南芥等至少20种植物上进行验证(表1 , 表2 ).实验采用的研究对象不一样, 不同植物本身亲缘识别的强度、途径、方式可能存在较大差异, 因此容易造成研究结果的差异(Biedrzycki & Bais, 2010 ).从植物分类学的角度看, 目前所报道的20个种分别来自2纲9目10科20属(表3 ).按照恩格勒植物分类系统, 这些种所覆盖的目, 诸如桔梗目、车前目、管状花目隶属于后生花被亚纲, 蔷薇目、罂粟目、无患子目、锦葵目隶属于异被花类, 中央子目和禾本目隶属于同被花类, 均属于被子植物, 而相对原始的裸子植物目前仍未见相关报道.从品种来源上看, 目前多数实验采用野生植物, 部分实验采用模式植物(拟南芥、水稻), 而经长期人工筛选的作物研究相对较少, 仅在大麦(Ninkovic, 2003 )、玉米(Zea may ) (Lin, 2013 )、大豆(Glycine max ) (肖桃艳等, 2015 )上有少量报道.因此, 建议在后续试验中加强向裸子植物和人工长期筛选作物两个方向延伸, 验证植物亲缘识别现象在原始的裸子植物以及长期人工选育作物(部分原始基因丢失)中是否仍然存在, 从而确定这种行为是否具有普遍性.此外, 目前的研究对象除Artemisia tridentate 外均为草本植物, 建议在后续研究中多关注灌木及木本植物. ...
Experimental evidence for a phylogenetic Janzen—Connell effect in a subtropical forest.
1
2012
... 一般认为, 同种或近亲缘植物共存, 因其生理生态特性高度相似, 所构成聚落物种单一, 更容易受到病虫害的危害(Schmitt & Antonovics, 1986 ; Weiblen et al ., 2006 ; Burns & Strauss, 2011 ; Liu et al ., 2012 ).但据Karban和Shiojiri (2009)、Karban等(2013)对Artemisia tridentata 的研究发现, 当植株受到机械损伤或者昆虫取食后, 能够向周边释放挥发物, 这些物质能够诱导周边植物的预警抗性, 而且近亲缘个体受诱导后, 抗性优于远亲缘个体, 表明亲缘识别可能影响植物的通讯与防御. ...
Competitive ability not kinship affects growth of Arabidopsis thaliana accessions.
3
2010
... 自1980年, 植物亲缘识别概念提出至今, 科学家们已在包括Lycopersicon lycopersicum (Willison et al ., 1987 )、长叶车前(Plantago lanceolata ) (Tonsor, 1989 )、Cakile edentula (Donohue, 2003 ; Dudley & File, 2007 ; Bhatt et al ., 2011 )、大麦(Hordeum vulgare ) (Ninkovic, 2003 )、狭叶羽扇豆(Lupinus angustifolius ) (Milla et al ., 2012 )、拟南芥(Arabidopsis thaliana ) (Goodnight, 1985 ; Biedrzycki et al ., 2010 ; Masclaux et al ., 2010 ; Caffaro et al ., 2013 ; Crepy & Casal, 2015 )、水稻(Oryza sativa ) (Fang et al ., 2013 )、豚草(Ambrosia artemisiifolia ) (File et al ., 2012 )等20种植物中进行验证, 其中有许多研究结果支持植物存在亲缘识别 (表1 ), 也有一些研究得到否定的结果(表2 ). ...
... 种子(后代)数量、种子萌发率、幼苗成活率等指标也被许多研究所应用(Willison et al ., 1987 ; Tonsor, 1989 ; Cheplick & Kane, 2004 ; Milla et al ., 2009 ; Masclaux et al ., 2010 ), 因其能够最直观地表征植物能否因亲缘识别而带来直接效益, 即繁殖效益.Willison等(1987)及Tonsor (1989)的研究均表明, 植物个体与近亲缘共存时具有更高的繁殖收益. ...
... 指标选定方面主要存在两个问题.第一个是选用的指标范围多局限于形态学和生长发育指标.目前大部分研究主要采用诸如株高、叶面积、生物量、开花数、开花时间、坐果数、种子数量、种子萌发率、种子存活率、根系长度、根冠比、叶片重叠度、根系交叉度等形态学指标, 间接表征植物是否存在亲缘识别现象.还有少数研究采用植物代谢酶活性(肖桃艳等, 2015 )、植物光合作用和呼吸作用速率(Meier et al ., 2013 )、根系分泌物(Badri et al ., 2012 )等生理代谢指标, 菌根数量(File et al ., 2012 )等环境指标, 基因表达模式(Biedrzycki et al ., 2011 ; Masclaux et al ., 2010 )等指标, 来研究目标植物与不同亲缘个体共存时的表现. ...
Kin recognition in ground squirrels and other rodents.
1
2003
... 许多研究表明, 在多种生物种中存在亲缘选择现象, 如海葵(Anthopleura elegant ) (Ayre & Grosberg, 1995 )、美新亮丽鲷(Neolamprologus pulcher ) (Dierkes et al ., 2005 )、蝾螈(Ambystoma tigrinum nebulosum ) (Pfennig & Collins, 1993 ), 白额食蜂鸟(Merops bullockoides ) (Emlen et al ., 1995 )、拜氏黄鼠(Spermophilus beldingi ) (Mateo, 2003 )、黑猩猩(Chimpanzee ) (Morin et al ., 1994 ).更有趣的是, 在对盘基网柄菌(Dictyostelium discoideum ) (Ostrowski et al ., 2008 )、酵母菌(Saccharomyces cerevisiae ) (Smukalla et al ., 2008 )等微生物的研究中, 同样发现类似亲缘选择现象.以上研究表明, 亲缘选择行为覆盖了肠腔类、鱼类、两栖类、鸟类、哺乳类, 乃至微生物等生物界的大部分类群. ...
Increased root oxygen uptake in pea plants responding to non-self neighbors.
1
2013
... 指标选定方面主要存在两个问题.第一个是选用的指标范围多局限于形态学和生长发育指标.目前大部分研究主要采用诸如株高、叶面积、生物量、开花数、开花时间、坐果数、种子数量、种子萌发率、种子存活率、根系长度、根冠比、叶片重叠度、根系交叉度等形态学指标, 间接表征植物是否存在亲缘识别现象.还有少数研究采用植物代谢酶活性(肖桃艳等, 2015 )、植物光合作用和呼吸作用速率(Meier et al ., 2013 )、根系分泌物(Badri et al ., 2012 )等生理代谢指标, 菌根数量(File et al ., 2012 )等环境指标, 基因表达模式(Biedrzycki et al ., 2011 ; Masclaux et al ., 2010 )等指标, 来研究目标植物与不同亲缘个体共存时的表现. ...
Kin and sex recognition in a dioecious grass.
3
2014
... 相比于早期研究侧重繁殖性状, 近期研究则更侧重于对生长性状的探讨, 主要通过生物量、株高、叶面积、根冠比、根系长度等指标表征植物与不同亲缘个体共存时, 目标个体的生存特征, 从侧面表征植物的亲缘识别行为.对Cakile edentula 、大麦、拟南芥、Distichlis spicata 等的研究均表明, 植物与近亲缘个体共存时, 提高了自身生物量(Donohue, 2003 ; Bhatt et al ., 2011 ; Biernaskie, 2011 ; Mercer & Eppley, 2014 ).但也有研究表明与近亲缘个体共处时, 目标个体生物量反而可能降低(Cheplick & Kane, 2004 ; Milla et al ., 2009 ), 或者没有影响(Ninkovic, 2003 ).此外, Tonsor (1989)研究发现, 当长叶车前与近亲缘个体共存时, 干质量虽然下降, 但产生更多的子代, 表明与近亲缘个体共存时虽然生存效益下降, 但是繁殖效益提高.因此, 目前而言, 用生物量的大小来判断植物是否存在亲缘识别或者亲缘利他存在着一定的局限性. ...
... 其中大多数支持植物存在亲缘识别的研究 (表1 )认为, 目标植物与远亲缘共存时具有较大的根系, 从而加强竞争(Ninkovic, 2003 ; Dudley & File, 2007 ; Biedrzycki et al ., 2010 ), 而反对的结果(表2 )则表明, 与近亲缘个体共存时形成更多根系(Murphy & Dudley, 2009 )或者两者间并无差异(Milla et al ., 2012 ; Semchenko et al ., 2014 ).Semchenko等(2014)的研究结果则提供了另一种新的思路, 目标植物与不同亲缘个体共存时, 根系长度并无差异, 但显著改变根系的构型.由于根系长度无法准确地表征植物的亲缘识别特征, 因此, 有学者提出从根系生长趋性的角度进行分析(趋向近亲缘或者远亲缘).Caffaro等(2013)和Fang等(2013)的研究均表明, 目标植物根系更趋向于近亲缘个体方向生长.另外还有部分研究采用目标植物根冠比来表征其与周边亲缘邻居的竞争强度(Bhatt et al ., 2011 ; Mercer & Eppley, 2014 ).首先, 当目标植物面对不同亲缘邻居, 其总生物量的形成可能并不受影响, 但其可能通过改变地上部地下部分配(根冠比)而改变竞争策略; 其次, 目标植物根系减少, 并不一定意味着个体间竞争减弱, 其可能通过将更多能量投入到地上部生产或者加大次级代谢产物的合成, 增加了个体间的竞争压力.因此, 采用根冠比能在一定程度上弥补采用根系生物量和根系长度较片面表征个体间竞争强度的不足. ...
... 不同研究对实验材料亲缘关系的界定方式和亲缘程度的选择也存在较大差异, 目前主要包括以下3种界定方式: 第一种以是否来自同一亲本为区分标准(Argyres & Schmitt, 1992 ; Dudley & File, 2007 ; Bhatt et al ., 2011 ; Semchenko et al ., 2014 ); 第二种以基因型为区分标准(Ninkovic, 2003 ; Biedrzycki et al ., 2010 ; Crepy & Casal, 2015 ); 第三种以人为克隆体为区分标准(Tonsor, 1989 ; Karban & Shiojiri, 2009 ; Mercer & Eppley, 2014 ).不同亲缘关系的界定方式, 也是导致目前研究结果差异较大的最主要原因.关于何种界定方式最为合适, 目前仍没有统一的定论.参考其他生物亲缘识别的相关研究, 动物主要以个体间血缘关系远近来界定个体间的亲缘关系(Emlen et al ., 1995 ), 而微生物主要以分裂方式进行繁殖, 故研究中主要以基因型或遗传距离来区分个体(菌群)之间的亲缘关系(Ostrowski et al ., 2008 ).目前植物亲缘识别研究主要以亲本关系及基因型(遗传距离)两种亲缘界定方式为主流, 且使用这两种方式的研究中, 均有支持和否定亲缘识别的结果, 所以这两种方式都可以得到相对准确的结果.因此, 选取何种亲缘界定方式最为合适仍很难判断, 在短期内可能仍会呈现多种界别方式共存的局面.为了尽量减少因采用不同亲缘界别方式所可能带来的误差, 建议后续实验中, 仅在采用同种亲缘界别方式的实验间进行结果比较. ...
Kinship rivalry does not trigger specific allocation strategies in Lupinus angustifolius.
3
2012
... 自1980年, 植物亲缘识别概念提出至今, 科学家们已在包括Lycopersicon lycopersicum (Willison et al ., 1987 )、长叶车前(Plantago lanceolata ) (Tonsor, 1989 )、Cakile edentula (Donohue, 2003 ; Dudley & File, 2007 ; Bhatt et al ., 2011 )、大麦(Hordeum vulgare ) (Ninkovic, 2003 )、狭叶羽扇豆(Lupinus angustifolius ) (Milla et al ., 2012 )、拟南芥(Arabidopsis thaliana ) (Goodnight, 1985 ; Biedrzycki et al ., 2010 ; Masclaux et al ., 2010 ; Caffaro et al ., 2013 ; Crepy & Casal, 2015 )、水稻(Oryza sativa ) (Fang et al ., 2013 )、豚草(Ambrosia artemisiifolia ) (File et al ., 2012 )等20种植物中进行验证, 其中有许多研究结果支持植物存在亲缘识别 (表1 ), 也有一些研究得到否定的结果(表2 ). ...
... 其中大多数支持植物存在亲缘识别的研究 (表1 )认为, 目标植物与远亲缘共存时具有较大的根系, 从而加强竞争(Ninkovic, 2003 ; Dudley & File, 2007 ; Biedrzycki et al ., 2010 ), 而反对的结果(表2 )则表明, 与近亲缘个体共存时形成更多根系(Murphy & Dudley, 2009 )或者两者间并无差异(Milla et al ., 2012 ; Semchenko et al ., 2014 ).Semchenko等(2014)的研究结果则提供了另一种新的思路, 目标植物与不同亲缘个体共存时, 根系长度并无差异, 但显著改变根系的构型.由于根系长度无法准确地表征植物的亲缘识别特征, 因此, 有学者提出从根系生长趋性的角度进行分析(趋向近亲缘或者远亲缘).Caffaro等(2013)和Fang等(2013)的研究均表明, 目标植物根系更趋向于近亲缘个体方向生长.另外还有部分研究采用目标植物根冠比来表征其与周边亲缘邻居的竞争强度(Bhatt et al ., 2011 ; Mercer & Eppley, 2014 ).首先, 当目标植物面对不同亲缘邻居, 其总生物量的形成可能并不受影响, 但其可能通过改变地上部地下部分配(根冠比)而改变竞争策略; 其次, 目标植物根系减少, 并不一定意味着个体间竞争减弱, 其可能通过将更多能量投入到地上部生产或者加大次级代谢产物的合成, 增加了个体间的竞争压力.因此, 采用根冠比能在一定程度上弥补采用根系生物量和根系长度较片面表征个体间竞争强度的不足. ...
... 研究表明, 植物面对周边个体, 不但能够产生一定的形态学响应, 而且在更深层次上还有许多“看不见”的生理生化策略的变化(Karban et al ., 2000 ; Baldwin et al ., 2006 ).对白车轴草(Trifolium repens ) (Lepik et al ., 2012 )、狭叶羽扇豆(Milla et al ., 2012 )的研究发现: 这两种植物分别与不同亲缘个体共存时, 其叶面积、株高、生物量、根长均无差异, 但未测定生理学指标, 因此还不能完全否定植物亲缘识别的存在.另外一个问题是指标上的以偏概全.部分研究采用某单一指标, 如侧根长(Dudley & File, 2007 , Biedrzycki et al ., 2010 )、生物量(Argyres & Schmitt, 1992 )、根冠比(Bhatt et al ., 2011 )存在的差异(其他指标可能没有差异)为标准, 判断植物会否存在亲缘识别.但是, 单一指标的差异并不能代表所有指标的趋势, 因此利用单一指标的差异表征亲缘识别现象的存在是以偏概全.因此, 建议在后续的实验中, 除了考察形态指标外, 还应该加大对植物生理生化指标的研究, 例如植株的养分利用能力, 次级代谢产物合成, 根系分泌物, 叶片挥发物, 由目前的以表观形态观察为主, 逐步转变到以植物形态变化与生理变化相结合的复合途径, 能够更加全面和直接地表征植物亲缘识别特征以及辅助对植物亲缘识别机理的研究. ...
Growing with siblings: A common ground for cooperation or for fiercer competition among plants?
2
2009
... 相比于早期研究侧重繁殖性状, 近期研究则更侧重于对生长性状的探讨, 主要通过生物量、株高、叶面积、根冠比、根系长度等指标表征植物与不同亲缘个体共存时, 目标个体的生存特征, 从侧面表征植物的亲缘识别行为.对Cakile edentula 、大麦、拟南芥、Distichlis spicata 等的研究均表明, 植物与近亲缘个体共存时, 提高了自身生物量(Donohue, 2003 ; Bhatt et al ., 2011 ; Biernaskie, 2011 ; Mercer & Eppley, 2014 ).但也有研究表明与近亲缘个体共处时, 目标个体生物量反而可能降低(Cheplick & Kane, 2004 ; Milla et al ., 2009 ), 或者没有影响(Ninkovic, 2003 ).此外, Tonsor (1989)研究发现, 当长叶车前与近亲缘个体共存时, 干质量虽然下降, 但产生更多的子代, 表明与近亲缘个体共存时虽然生存效益下降, 但是繁殖效益提高.因此, 目前而言, 用生物量的大小来判断植物是否存在亲缘识别或者亲缘利他存在着一定的局限性. ...
... 种子(后代)数量、种子萌发率、幼苗成活率等指标也被许多研究所应用(Willison et al ., 1987 ; Tonsor, 1989 ; Cheplick & Kane, 2004 ; Milla et al ., 2009 ; Masclaux et al ., 2010 ), 因其能够最直观地表征植物能否因亲缘识别而带来直接效益, 即繁殖效益.Willison等(1987)及Tonsor (1989)的研究均表明, 植物个体与近亲缘共存时具有更高的繁殖收益. ...
Unveiling below-ground species abundance in a biodiversity experiment: A test of vertical niche differentiation among grassland species.
1
2010
... 研究表明, 植物能够识别土壤环境的养分条件, 将根系向资源丰富区域集中, 而减少向资源匮乏地区的增殖, 表明植物根系的生长和分布是由土壤中的营养物质浓度所驱动的(Hodge, 2004 ).但近年研究发现, 一些植物的根系生长和分布并不是受土壤营养水平驱动的, 而是更多受到邻近植物根系的影响(Hodge, 2009 ; Mommer et al ., 2010 ), 植物的亲缘识别可能正属于这种非养分驱动行为. ...
Kin selection, social structure, gene flow, and the evolution of chimpanzees.
1
1994
... 许多研究表明, 在多种生物种中存在亲缘选择现象, 如海葵(Anthopleura elegant ) (Ayre & Grosberg, 1995 )、美新亮丽鲷(Neolamprologus pulcher ) (Dierkes et al ., 2005 )、蝾螈(Ambystoma tigrinum nebulosum ) (Pfennig & Collins, 1993 ), 白额食蜂鸟(Merops bullockoides ) (Emlen et al ., 1995 )、拜氏黄鼠(Spermophilus beldingi ) (Mateo, 2003 )、黑猩猩(Chimpanzee ) (Morin et al ., 1994 ).更有趣的是, 在对盘基网柄菌(Dictyostelium discoideum ) (Ostrowski et al ., 2008 )、酵母菌(Saccharomyces cerevisiae ) (Smukalla et al ., 2008 )等微生物的研究中, 同样发现类似亲缘选择现象.以上研究表明, 亲缘选择行为覆盖了肠腔类、鱼类、两栖类、鸟类、哺乳类, 乃至微生物等生物界的大部分类群. ...
Root Development and Ecological Relations of Guayule. United States Department of Agriculture
1
1946
... 其实, 植物亲缘识别的概念并非Dudley首创(Hodge, 2012 ).关于植物亲缘识别最早可以追溯到19世纪关于植物的自交不亲和(self-incompatibility)的研究, 近则可以追溯到1946年Muller等对灰白银胶菊(Parthenium argentatum )面对不同亲缘个体根系分布的研究(Muller, 1946 ).而在后续的30年间, 并没有提出所谓植物亲缘识别的概念.直到1980年, Nakamura在《Evolution Theory》上发表了题为“Plant kin selection”的文章, 首次提出了植物亲缘识别的概念. 而Dudley的文章再次唤起了科学家对植物亲缘识别研究的关注. ...
Kin recognition: Competition and cooperation in Impatiens (Balsaminaceae).
1
2009
... 其中大多数支持植物存在亲缘识别的研究 (表1 )认为, 目标植物与远亲缘共存时具有较大的根系, 从而加强竞争(Ninkovic, 2003 ; Dudley & File, 2007 ; Biedrzycki et al ., 2010 ), 而反对的结果(表2 )则表明, 与近亲缘个体共存时形成更多根系(Murphy & Dudley, 2009 )或者两者间并无差异(Milla et al ., 2012 ; Semchenko et al ., 2014 ).Semchenko等(2014)的研究结果则提供了另一种新的思路, 目标植物与不同亲缘个体共存时, 根系长度并无差异, 但显著改变根系的构型.由于根系长度无法准确地表征植物的亲缘识别特征, 因此, 有学者提出从根系生长趋性的角度进行分析(趋向近亲缘或者远亲缘).Caffaro等(2013)和Fang等(2013)的研究均表明, 目标植物根系更趋向于近亲缘个体方向生长.另外还有部分研究采用目标植物根冠比来表征其与周边亲缘邻居的竞争强度(Bhatt et al ., 2011 ; Mercer & Eppley, 2014 ).首先, 当目标植物面对不同亲缘邻居, 其总生物量的形成可能并不受影响, 但其可能通过改变地上部地下部分配(根冠比)而改变竞争策略; 其次, 目标植物根系减少, 并不一定意味着个体间竞争减弱, 其可能通过将更多能量投入到地上部生产或者加大次级代谢产物的合成, 增加了个体间的竞争压力.因此, 采用根冠比能在一定程度上弥补采用根系生物量和根系长度较片面表征个体间竞争强度的不足. ...
Plant kin selection.
0
1980
Volatile communication between barley plants affects biomass allocation.
5
2003
... 自1980年, 植物亲缘识别概念提出至今, 科学家们已在包括Lycopersicon lycopersicum (Willison et al ., 1987 )、长叶车前(Plantago lanceolata ) (Tonsor, 1989 )、Cakile edentula (Donohue, 2003 ; Dudley & File, 2007 ; Bhatt et al ., 2011 )、大麦(Hordeum vulgare ) (Ninkovic, 2003 )、狭叶羽扇豆(Lupinus angustifolius ) (Milla et al ., 2012 )、拟南芥(Arabidopsis thaliana ) (Goodnight, 1985 ; Biedrzycki et al ., 2010 ; Masclaux et al ., 2010 ; Caffaro et al ., 2013 ; Crepy & Casal, 2015 )、水稻(Oryza sativa ) (Fang et al ., 2013 )、豚草(Ambrosia artemisiifolia ) (File et al ., 2012 )等20种植物中进行验证, 其中有许多研究结果支持植物存在亲缘识别 (表1 ), 也有一些研究得到否定的结果(表2 ). ...
... 相比于早期研究侧重繁殖性状, 近期研究则更侧重于对生长性状的探讨, 主要通过生物量、株高、叶面积、根冠比、根系长度等指标表征植物与不同亲缘个体共存时, 目标个体的生存特征, 从侧面表征植物的亲缘识别行为.对Cakile edentula 、大麦、拟南芥、Distichlis spicata 等的研究均表明, 植物与近亲缘个体共存时, 提高了自身生物量(Donohue, 2003 ; Bhatt et al ., 2011 ; Biernaskie, 2011 ; Mercer & Eppley, 2014 ).但也有研究表明与近亲缘个体共处时, 目标个体生物量反而可能降低(Cheplick & Kane, 2004 ; Milla et al ., 2009 ), 或者没有影响(Ninkovic, 2003 ).此外, Tonsor (1989)研究发现, 当长叶车前与近亲缘个体共存时, 干质量虽然下降, 但产生更多的子代, 表明与近亲缘个体共存时虽然生存效益下降, 但是繁殖效益提高.因此, 目前而言, 用生物量的大小来判断植物是否存在亲缘识别或者亲缘利他存在着一定的局限性. ...
... 其中大多数支持植物存在亲缘识别的研究 (表1 )认为, 目标植物与远亲缘共存时具有较大的根系, 从而加强竞争(Ninkovic, 2003 ; Dudley & File, 2007 ; Biedrzycki et al ., 2010 ), 而反对的结果(表2 )则表明, 与近亲缘个体共存时形成更多根系(Murphy & Dudley, 2009 )或者两者间并无差异(Milla et al ., 2012 ; Semchenko et al ., 2014 ).Semchenko等(2014)的研究结果则提供了另一种新的思路, 目标植物与不同亲缘个体共存时, 根系长度并无差异, 但显著改变根系的构型.由于根系长度无法准确地表征植物的亲缘识别特征, 因此, 有学者提出从根系生长趋性的角度进行分析(趋向近亲缘或者远亲缘).Caffaro等(2013)和Fang等(2013)的研究均表明, 目标植物根系更趋向于近亲缘个体方向生长.另外还有部分研究采用目标植物根冠比来表征其与周边亲缘邻居的竞争强度(Bhatt et al ., 2011 ; Mercer & Eppley, 2014 ).首先, 当目标植物面对不同亲缘邻居, 其总生物量的形成可能并不受影响, 但其可能通过改变地上部地下部分配(根冠比)而改变竞争策略; 其次, 目标植物根系减少, 并不一定意味着个体间竞争减弱, 其可能通过将更多能量投入到地上部生产或者加大次级代谢产物的合成, 增加了个体间的竞争压力.因此, 采用根冠比能在一定程度上弥补采用根系生物量和根系长度较片面表征个体间竞争强度的不足. ...
... 现有研究已在包括拟南芥等至少20种植物上进行验证(表1 , 表2 ).实验采用的研究对象不一样, 不同植物本身亲缘识别的强度、途径、方式可能存在较大差异, 因此容易造成研究结果的差异(Biedrzycki & Bais, 2010 ).从植物分类学的角度看, 目前所报道的20个种分别来自2纲9目10科20属(表3 ).按照恩格勒植物分类系统, 这些种所覆盖的目, 诸如桔梗目、车前目、管状花目隶属于后生花被亚纲, 蔷薇目、罂粟目、无患子目、锦葵目隶属于异被花类, 中央子目和禾本目隶属于同被花类, 均属于被子植物, 而相对原始的裸子植物目前仍未见相关报道.从品种来源上看, 目前多数实验采用野生植物, 部分实验采用模式植物(拟南芥、水稻), 而经长期人工筛选的作物研究相对较少, 仅在大麦(Ninkovic, 2003 )、玉米(Zea may ) (Lin, 2013 )、大豆(Glycine max ) (肖桃艳等, 2015 )上有少量报道.因此, 建议在后续试验中加强向裸子植物和人工长期筛选作物两个方向延伸, 验证植物亲缘识别现象在原始的裸子植物以及长期人工选育作物(部分原始基因丢失)中是否仍然存在, 从而确定这种行为是否具有普遍性.此外, 目前的研究对象除Artemisia tridentate 外均为草本植物, 建议在后续研究中多关注灌木及木本植物. ...
... 不同研究对实验材料亲缘关系的界定方式和亲缘程度的选择也存在较大差异, 目前主要包括以下3种界定方式: 第一种以是否来自同一亲本为区分标准(Argyres & Schmitt, 1992 ; Dudley & File, 2007 ; Bhatt et al ., 2011 ; Semchenko et al ., 2014 ); 第二种以基因型为区分标准(Ninkovic, 2003 ; Biedrzycki et al ., 2010 ; Crepy & Casal, 2015 ); 第三种以人为克隆体为区分标准(Tonsor, 1989 ; Karban & Shiojiri, 2009 ; Mercer & Eppley, 2014 ).不同亲缘关系的界定方式, 也是导致目前研究结果差异较大的最主要原因.关于何种界定方式最为合适, 目前仍没有统一的定论.参考其他生物亲缘识别的相关研究, 动物主要以个体间血缘关系远近来界定个体间的亲缘关系(Emlen et al ., 1995 ), 而微生物主要以分裂方式进行繁殖, 故研究中主要以基因型或遗传距离来区分个体(菌群)之间的亲缘关系(Ostrowski et al ., 2008 ).目前植物亲缘识别研究主要以亲本关系及基因型(遗传距离)两种亲缘界定方式为主流, 且使用这两种方式的研究中, 均有支持和否定亲缘识别的结果, 所以这两种方式都可以得到相对准确的结果.因此, 选取何种亲缘界定方式最为合适仍很难判断, 在短期内可能仍会呈现多种界别方式共存的局面.为了尽量减少因采用不同亲缘界别方式所可能带来的误差, 建议后续实验中, 仅在采用同种亲缘界别方式的实验间进行结果比较. ...
Plant coexistence alters terpene emission and content of Mediterranean species.
1
2007
... 对龙葵(Solanum nigrum ) (Schmidt & Balswin, 2006 )、Pinus halepensis (Ormeño et al ., 2007 )、Centaurea maculosa (Broz et al ., 2010 )、拟南芥(Badri et al ., 2012 )等的研究也发现同种个体共存植株体内抗性物质提高的现象, 这有力地支持了Karban等的观点. ...
Kin discrimination increases with genetic distance in a social amoeba.
2
2008
... 许多研究表明, 在多种生物种中存在亲缘选择现象, 如海葵(Anthopleura elegant ) (Ayre & Grosberg, 1995 )、美新亮丽鲷(Neolamprologus pulcher ) (Dierkes et al ., 2005 )、蝾螈(Ambystoma tigrinum nebulosum ) (Pfennig & Collins, 1993 ), 白额食蜂鸟(Merops bullockoides ) (Emlen et al ., 1995 )、拜氏黄鼠(Spermophilus beldingi ) (Mateo, 2003 )、黑猩猩(Chimpanzee ) (Morin et al ., 1994 ).更有趣的是, 在对盘基网柄菌(Dictyostelium discoideum ) (Ostrowski et al ., 2008 )、酵母菌(Saccharomyces cerevisiae ) (Smukalla et al ., 2008 )等微生物的研究中, 同样发现类似亲缘选择现象.以上研究表明, 亲缘选择行为覆盖了肠腔类、鱼类、两栖类、鸟类、哺乳类, 乃至微生物等生物界的大部分类群. ...
... 不同研究对实验材料亲缘关系的界定方式和亲缘程度的选择也存在较大差异, 目前主要包括以下3种界定方式: 第一种以是否来自同一亲本为区分标准(Argyres & Schmitt, 1992 ; Dudley & File, 2007 ; Bhatt et al ., 2011 ; Semchenko et al ., 2014 ); 第二种以基因型为区分标准(Ninkovic, 2003 ; Biedrzycki et al ., 2010 ; Crepy & Casal, 2015 ); 第三种以人为克隆体为区分标准(Tonsor, 1989 ; Karban & Shiojiri, 2009 ; Mercer & Eppley, 2014 ).不同亲缘关系的界定方式, 也是导致目前研究结果差异较大的最主要原因.关于何种界定方式最为合适, 目前仍没有统一的定论.参考其他生物亲缘识别的相关研究, 动物主要以个体间血缘关系远近来界定个体间的亲缘关系(Emlen et al ., 1995 ), 而微生物主要以分裂方式进行繁殖, 故研究中主要以基因型或遗传距离来区分个体(菌群)之间的亲缘关系(Ostrowski et al ., 2008 ).目前植物亲缘识别研究主要以亲本关系及基因型(遗传距离)两种亲缘界定方式为主流, 且使用这两种方式的研究中, 均有支持和否定亲缘识别的结果, 所以这两种方式都可以得到相对准确的结果.因此, 选取何种亲缘界定方式最为合适仍很难判断, 在短期内可能仍会呈现多种界别方式共存的局面.为了尽量减少因采用不同亲缘界别方式所可能带来的误差, 建议后续实验中, 仅在采用同种亲缘界别方式的实验间进行结果比较. ...
Kinship affects morphogenesis in cannibalistic salamanders.
1
1993
... 许多研究表明, 在多种生物种中存在亲缘选择现象, 如海葵(Anthopleura elegant ) (Ayre & Grosberg, 1995 )、美新亮丽鲷(Neolamprologus pulcher ) (Dierkes et al ., 2005 )、蝾螈(Ambystoma tigrinum nebulosum ) (Pfennig & Collins, 1993 ), 白额食蜂鸟(Merops bullockoides ) (Emlen et al ., 1995 )、拜氏黄鼠(Spermophilus beldingi ) (Mateo, 2003 )、黑猩猩(Chimpanzee ) (Morin et al ., 1994 ).更有趣的是, 在对盘基网柄菌(Dictyostelium discoideum ) (Ostrowski et al ., 2008 )、酵母菌(Saccharomyces cerevisiae ) (Smukalla et al ., 2008 )等微生物的研究中, 同样发现类似亲缘选择现象.以上研究表明, 亲缘选择行为覆盖了肠腔类、鱼类、两栖类、鸟类、哺乳类, 乃至微生物等生物界的大部分类群. ...
Transcriptional responses of Solanum nigrum to methyl jasmonate and competition: A glasshouse and field study.
0
2006
Experimental studies of the evolutionary significance of sexual reproduction. IV. Effect of neighbor relatedness and aphid infestation on seedling performance.
1
1986
... 一般认为, 同种或近亲缘植物共存, 因其生理生态特性高度相似, 所构成聚落物种单一, 更容易受到病虫害的危害(Schmitt & Antonovics, 1986 ; Weiblen et al ., 2006 ; Burns & Strauss, 2011 ; Liu et al ., 2012 ).但据Karban和Shiojiri (2009)、Karban等(2013)对Artemisia tridentata 的研究发现, 当植株受到机械损伤或者昆虫取食后, 能够向周边释放挥发物, 这些物质能够诱导周边植物的预警抗性, 而且近亲缘个体受诱导后, 抗性优于远亲缘个体, 表明亲缘识别可能影响植物的通讯与防御. ...
Cryptochrome as a sensor of the blue/green ratio of natural radiation in Arabidopsis.
1
2010
... 植物光合色素对光吸收具有选择性, 投射到叶片上的红光和蓝光大部分均被吸收, 仅有少量的红光、红外光和蓝光能够穿透叶片, 透射到冠层下(Smith, 2000 ; Casal, 2013 ); 而冠层下的植物或者叶片进化出一种有效的机制, 能够通过冠层下红光与远红外光的比例, 判断周边的光照条件, 调整叶片的生长方向, 向光照充分的地方生长, 并减少叶片间的相互遮蔽(Casal, 2013 ).已有研究表明, 植物主要通过光敏色素B (phyB)和隐花色素(cry1)识别这两种光的微弱变化(Sellaro et al ., 2010 ; Keller et al ., 2011 ). ...
Plant root exudates mediate neighbour recognition and trigger complex behavioural changes.
2
2014
... 其中大多数支持植物存在亲缘识别的研究 (表1 )认为, 目标植物与远亲缘共存时具有较大的根系, 从而加强竞争(Ninkovic, 2003 ; Dudley & File, 2007 ; Biedrzycki et al ., 2010 ), 而反对的结果(表2 )则表明, 与近亲缘个体共存时形成更多根系(Murphy & Dudley, 2009 )或者两者间并无差异(Milla et al ., 2012 ; Semchenko et al ., 2014 ).Semchenko等(2014)的研究结果则提供了另一种新的思路, 目标植物与不同亲缘个体共存时, 根系长度并无差异, 但显著改变根系的构型.由于根系长度无法准确地表征植物的亲缘识别特征, 因此, 有学者提出从根系生长趋性的角度进行分析(趋向近亲缘或者远亲缘).Caffaro等(2013)和Fang等(2013)的研究均表明, 目标植物根系更趋向于近亲缘个体方向生长.另外还有部分研究采用目标植物根冠比来表征其与周边亲缘邻居的竞争强度(Bhatt et al ., 2011 ; Mercer & Eppley, 2014 ).首先, 当目标植物面对不同亲缘邻居, 其总生物量的形成可能并不受影响, 但其可能通过改变地上部地下部分配(根冠比)而改变竞争策略; 其次, 目标植物根系减少, 并不一定意味着个体间竞争减弱, 其可能通过将更多能量投入到地上部生产或者加大次级代谢产物的合成, 增加了个体间的竞争压力.因此, 采用根冠比能在一定程度上弥补采用根系生物量和根系长度较片面表征个体间竞争强度的不足. ...
... 不同研究对实验材料亲缘关系的界定方式和亲缘程度的选择也存在较大差异, 目前主要包括以下3种界定方式: 第一种以是否来自同一亲本为区分标准(Argyres & Schmitt, 1992 ; Dudley & File, 2007 ; Bhatt et al ., 2011 ; Semchenko et al ., 2014 ); 第二种以基因型为区分标准(Ninkovic, 2003 ; Biedrzycki et al ., 2010 ; Crepy & Casal, 2015 ); 第三种以人为克隆体为区分标准(Tonsor, 1989 ; Karban & Shiojiri, 2009 ; Mercer & Eppley, 2014 ).不同亲缘关系的界定方式, 也是导致目前研究结果差异较大的最主要原因.关于何种界定方式最为合适, 目前仍没有统一的定论.参考其他生物亲缘识别的相关研究, 动物主要以个体间血缘关系远近来界定个体间的亲缘关系(Emlen et al ., 1995 ), 而微生物主要以分裂方式进行繁殖, 故研究中主要以基因型或遗传距离来区分个体(菌群)之间的亲缘关系(Ostrowski et al ., 2008 ).目前植物亲缘识别研究主要以亲本关系及基因型(遗传距离)两种亲缘界定方式为主流, 且使用这两种方式的研究中, 均有支持和否定亲缘识别的结果, 所以这两种方式都可以得到相对准确的结果.因此, 选取何种亲缘界定方式最为合适仍很难判断, 在短期内可能仍会呈现多种界别方式共存的局面.为了尽量减少因采用不同亲缘界别方式所可能带来的误差, 建议后续实验中, 仅在采用同种亲缘界别方式的实验间进行结果比较. ...
Phytochromes and light signal perception by plants—An emerging synthesis.
1
2000
... 植物光合色素对光吸收具有选择性, 投射到叶片上的红光和蓝光大部分均被吸收, 仅有少量的红光、红外光和蓝光能够穿透叶片, 透射到冠层下(Smith, 2000 ; Casal, 2013 ); 而冠层下的植物或者叶片进化出一种有效的机制, 能够通过冠层下红光与远红外光的比例, 判断周边的光照条件, 调整叶片的生长方向, 向光照充分的地方生长, 并减少叶片间的相互遮蔽(Casal, 2013 ).已有研究表明, 植物主要通过光敏色素B (phyB)和隐花色素(cry1)识别这两种光的微弱变化(Sellaro et al ., 2010 ; Keller et al ., 2011 ). ...
FLO1 is a variable green beard gene that drives biofilm-like cooperation in budding yeast.
1
2008
... 许多研究表明, 在多种生物种中存在亲缘选择现象, 如海葵(Anthopleura elegant ) (Ayre & Grosberg, 1995 )、美新亮丽鲷(Neolamprologus pulcher ) (Dierkes et al ., 2005 )、蝾螈(Ambystoma tigrinum nebulosum ) (Pfennig & Collins, 1993 ), 白额食蜂鸟(Merops bullockoides ) (Emlen et al ., 1995 )、拜氏黄鼠(Spermophilus beldingi ) (Mateo, 2003 )、黑猩猩(Chimpanzee ) (Morin et al ., 1994 ).更有趣的是, 在对盘基网柄菌(Dictyostelium discoideum ) (Ostrowski et al ., 2008 )、酵母菌(Saccharomyces cerevisiae ) (Smukalla et al ., 2008 )等微生物的研究中, 同样发现类似亲缘选择现象.以上研究表明, 亲缘选择行为覆盖了肠腔类、鱼类、两栖类、鸟类、哺乳类, 乃至微生物等生物界的大部分类群. ...
Relatedness and intraspecific competition in Plantago lanceolata.
3
1989
... 自1980年, 植物亲缘识别概念提出至今, 科学家们已在包括Lycopersicon lycopersicum (Willison et al ., 1987 )、长叶车前(Plantago lanceolata ) (Tonsor, 1989 )、Cakile edentula (Donohue, 2003 ; Dudley & File, 2007 ; Bhatt et al ., 2011 )、大麦(Hordeum vulgare ) (Ninkovic, 2003 )、狭叶羽扇豆(Lupinus angustifolius ) (Milla et al ., 2012 )、拟南芥(Arabidopsis thaliana ) (Goodnight, 1985 ; Biedrzycki et al ., 2010 ; Masclaux et al ., 2010 ; Caffaro et al ., 2013 ; Crepy & Casal, 2015 )、水稻(Oryza sativa ) (Fang et al ., 2013 )、豚草(Ambrosia artemisiifolia ) (File et al ., 2012 )等20种植物中进行验证, 其中有许多研究结果支持植物存在亲缘识别 (表1 ), 也有一些研究得到否定的结果(表2 ). ...
... 种子(后代)数量、种子萌发率、幼苗成活率等指标也被许多研究所应用(Willison et al ., 1987 ; Tonsor, 1989 ; Cheplick & Kane, 2004 ; Milla et al ., 2009 ; Masclaux et al ., 2010 ), 因其能够最直观地表征植物能否因亲缘识别而带来直接效益, 即繁殖效益.Willison等(1987)及Tonsor (1989)的研究均表明, 植物个体与近亲缘共存时具有更高的繁殖收益. ...
... 不同研究对实验材料亲缘关系的界定方式和亲缘程度的选择也存在较大差异, 目前主要包括以下3种界定方式: 第一种以是否来自同一亲本为区分标准(Argyres & Schmitt, 1992 ; Dudley & File, 2007 ; Bhatt et al ., 2011 ; Semchenko et al ., 2014 ); 第二种以基因型为区分标准(Ninkovic, 2003 ; Biedrzycki et al ., 2010 ; Crepy & Casal, 2015 ); 第三种以人为克隆体为区分标准(Tonsor, 1989 ; Karban & Shiojiri, 2009 ; Mercer & Eppley, 2014 ).不同亲缘关系的界定方式, 也是导致目前研究结果差异较大的最主要原因.关于何种界定方式最为合适, 目前仍没有统一的定论.参考其他生物亲缘识别的相关研究, 动物主要以个体间血缘关系远近来界定个体间的亲缘关系(Emlen et al ., 1995 ), 而微生物主要以分裂方式进行繁殖, 故研究中主要以基因型或遗传距离来区分个体(菌群)之间的亲缘关系(Ostrowski et al ., 2008 ).目前植物亲缘识别研究主要以亲本关系及基因型(遗传距离)两种亲缘界定方式为主流, 且使用这两种方式的研究中, 均有支持和否定亲缘识别的结果, 所以这两种方式都可以得到相对准确的结果.因此, 选取何种亲缘界定方式最为合适仍很难判断, 在短期内可能仍会呈现多种界别方式共存的局面.为了尽量减少因采用不同亲缘界别方式所可能带来的误差, 建议后续实验中, 仅在采用同种亲缘界别方式的实验间进行结果比较. ...
Phylogenetic dispersion of host use in a tropical insect herbivore community.
1
2006
... 一般认为, 同种或近亲缘植物共存, 因其生理生态特性高度相似, 所构成聚落物种单一, 更容易受到病虫害的危害(Schmitt & Antonovics, 1986 ; Weiblen et al ., 2006 ; Burns & Strauss, 2011 ; Liu et al ., 2012 ).但据Karban和Shiojiri (2009)、Karban等(2013)对Artemisia tridentata 的研究发现, 当植株受到机械损伤或者昆虫取食后, 能够向周边释放挥发物, 这些物质能够诱导周边植物的预警抗性, 而且近亲缘个体受诱导后, 抗性优于远亲缘个体, 表明亲缘识别可能影响植物的通讯与防御. ...
Sibling competition in plants: an experimental study.
0
1987
Herbivory-induced signalling in plants: Perception and action.
1
2009
... 许多植物受到虫害胁迫或者物理伤害后, 都能够向周边释放挥发物, 且这些挥发物能够诱导周边个体的防御能力(Wu & Baldwin, 2009 ).那么, 对于同一警报挥发物, 不同亲缘个体的响应效果是否有差别呢? ...
不同亲缘关系大豆品种间种对生长及氮素利用效率的影响.
0
2013
不同亲缘关系大豆品种间种对生长及氮素利用效率的影响.
0
2013



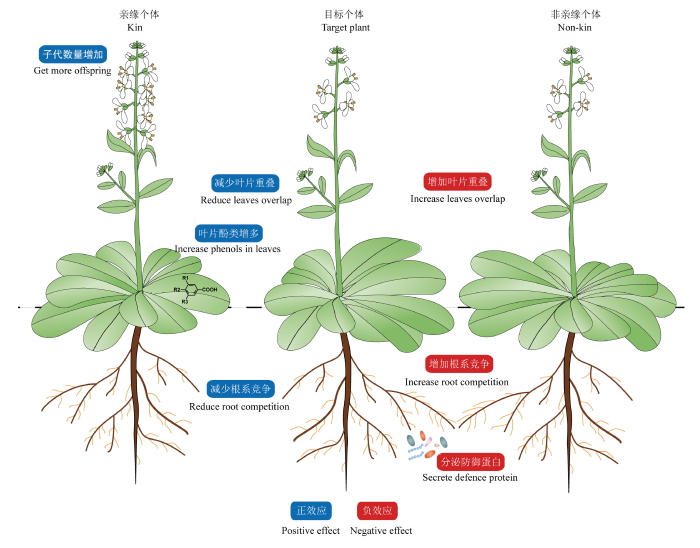
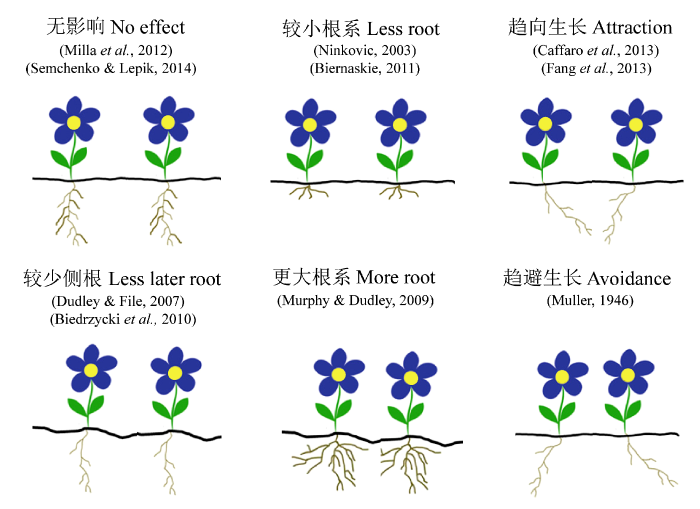
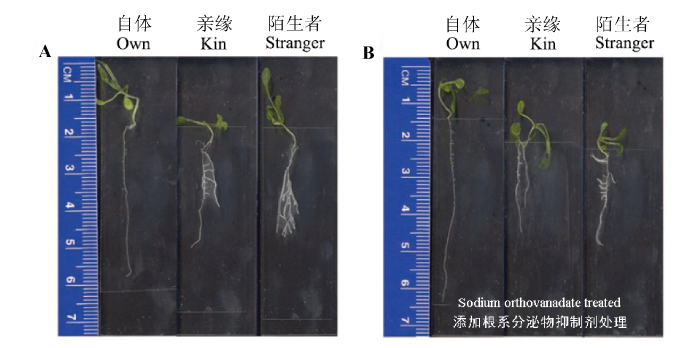
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}