第八次全国森林资源清查结果显示, 我国人工林面积已达6900多万hm2 , 约占全球人工林面积的1/4, 在我国木材生产和生态安全保障中起着至关重要的作用(徐济德, 2014 )。在我国实行天然林保护的背景下, 人工林, 尤其是用材林在提供林产品方面的作用愈加重要, 并且其面积还在持续增加(徐济德, 2014 )。但是, 我国人工林管理粗放、经营水平低, 导致其生产力不高, 平均蓄积量仅为52.8 m3 ·hm-2 , 不到发达国家的1/3 (徐济德, 2014 )。要提高人工林生产和生态服务功能, 亟需改变过去传统的粗放式管理模式, 实行高度集约式经营。在大部分区域, 土壤氮素缺乏是森林生态系统生产力最重要的限制因子(Deluca et al ., 2002 ; Zhou et al ., 2003 ; 于明坚等, 2005 )。为解决氮素的限制, 施氮肥已经成为提高人工林生产力的一种高度集约式经营管理措施(Fox, 2000 ; Lee & Jose, 2003 )。但是, 施肥在提高林地生产力的同时, 还会对土壤呼吸等地下生态过程产生深刻影响。
土壤呼吸是陆地生态系统碳循环的重要环节, 是在土壤新陈代谢功能过程中, 土壤产生大量的CO2 , 并向大气中释放CO2 的过程(杨晶和李凌浩, 2003 )。据估计, 每年从土壤释放到大气中的碳高达98 Pg·a-1 (Bond-Lamberty & Thompson, 2010 ), 相当于化石燃料燃烧所释放碳的10倍多(Zhu & Cheng, 2011 )。因此, 土壤呼吸速率的微小变化都可能会对大气中CO2 浓度产生巨大影响。在森林生态系统中, 施氮肥对土壤呼吸的影响国内外已有大量研究报道, 但不同树种的森林生态系统土壤呼吸对施氮肥的响应之间存在差异, 主要表现为促进(Cleveland & Townsend, 2006 ; Tu et al ., 2013 )、抑制(Bowden et al ., 2004 ; Olsson et al ., 2005 ; 贾淑霞等, 2007 ; Mo et al ., 2008 ; Janssens et al ., 2010 )或无影响(Samuelson et al ., 2009 )。例如, 施氮肥使亚热带竹(Pleioblastus amarus )林生态系统的土壤呼吸增加26%-52% (Tu et al ., 2013), 使北方森林生态系统的土壤呼吸降低40% (Olsson et al ., 2005 )。施氮肥在不同森林和区域对土壤呼吸的影响存在差异, 主要与树种特性和土壤性质有关, 也与各实验所施用氮肥的类型和施用时间长短有关(Tu et al ., 2013)。
土壤呼吸可以分为自养呼吸和异养呼吸。在我们的研究中自养呼吸主要是指植物根系呼吸, 而异养呼吸主要是指土壤微生物呼吸。这些组分对施肥的响应至关重要, 关系到生态系统碳平衡(Tyree et al ., 2006 )。目前主要采用挖壕沟-切根法、树干环割等方法来区分植物根系呼吸和土壤微生物呼吸(Högberg et al ., 2001 ; Tu et al ., 2013 )。但是, 目前大部分实验只是研究了土壤总呼吸对施氮肥的响应, 而关于根系呼吸和微生物呼吸对施氮肥的响应还研究得比较少, 为数不多的研究所取得的结果还相互矛盾(Tyree et al ., 2006 ; Tu et al ., 2013 )。例如, Tu等(2013)在竹林中发现施氮肥促进了根系呼吸和微生物呼吸; 而Janssens等(2010)通过整合分析发现施氮肥后土壤微生物呼吸降低15%。在亚热带地区, 关于施肥对人工林土壤呼吸过程影响的研究, 尤其是对土壤呼吸不同组分的研究还极其有限, 这严重地限制了我们对施肥措施对土壤碳循环过程影响的深入理解, 不利于人工林的可持续经营管理。
杉木(Cunninghamia lanceolata )人工林在我国亚热带地区的种植面积已接近900万hm2 , 在木材生产和生态安全保障等方面均具有举足轻重的作用(国家林业局森林资源管理司, 2010 )。为探讨施氮肥对杉木人工林土壤呼吸的影响, 我们在湖南会同林区的杉木幼林生态系统中开展了施肥试验, 以研究施氮肥对杉木幼林土壤呼吸的影响。相对于土壤总呼吸的变化, 了解土壤呼吸不同组分对施氮肥的响应更有助于认知土壤碳释放过程和对其进行有效的调控。因此, 为了明确土壤呼吸中不同组分(即根系呼吸和微生物呼吸)对施加氮肥的响应, 我们采用挖壕沟-切根法来区分土壤根系呼吸和微生物呼吸。在该研究中, 我们测定了土壤呼吸、根系生物量、微生物群落结构及相关环境因子的变化, 以期明确在杉木幼林生态系统中施氮肥对土壤总呼吸、根系呼吸和微生物呼吸的影响及其机理, 为杉木人工林土壤固碳增汇管理提供基础数据和理论依据, 该研究结果也将有助于我们了解氮沉降增加对中亚热带森林生态系统土壤碳释放的影响。
1 研究地区和研究方法
1.1 研究区概况
该实验样地位于湖南省会同县广坪镇, 地处云贵高原, 属于低山丘陵地貌。该区域年降水量为1200-1300 mm, 主要分布在4-8月份; 年平均气温为16.5 ℃, 最高气温和最低气温分别出现在7月份和1月份。研究区的土壤属于起源于板岩和灰岩的红黄壤, 施肥前各处理间土壤基本性质差异不显著(表1 )。施肥实验在5年生杉木幼林中进行, 种植密度为2000株·hm-2 。施肥前杉木幼林的平均树高和胸径见表1 , 林下植被主要是一些蕨类和茅草。
1.2 试验设计与指标测定
实验样地设在坡向、坡度基本一致的5年生杉木幼林, 面积约5 hm2 , 在其内设置3个50 m × 20 m的小区, 每个小区之间的距离不低于20 m。然后将每个小区划分为2个20 m × 20 m的样方, 样方之间有10 m的缓冲区。该实验包括2个处理, 每个处理重复3次, 采用随机区组设计。在每个小区的2个样方中随机布置施肥实验的2个处理, 即对照(CK)和添加氮肥(N)。同时, 在每个样方内随机设置2个1 m × 1 m切根亚样方, 通过与非切根小区比较, 用来区分根系呼吸和土壤微生物呼吸, 以研究土壤呼吸不同组分(根系呼吸和微生物呼吸)对施氮肥的响应。切根采用挖壕沟的方法, 即在1 m × 1 m切根亚样方的四周挖60 cm深、10 cm宽的沟, 并将PVC板放入壕沟内侧, 用来阻止切根小区外的树木根系进入切根小区土壤中, 然后将土壤回填到壕沟内并压实。为保证切根小区内没有植物根系进入, 每月采用手工方式及时清除切根小区内的所有灌木和草本植物。因杉木的根系主要分布在0-40 cm土层中, 挖60 cm的深度足以切断杉木根系并阻止新根系进入切根小区内。氮肥为尿素, 施肥量为200 kg N·hm-2 ·a-1 。施肥从2012年4月份开始, 采用均匀播撒的方式, 施肥时间选择在每年4月中下旬的降雨前夕。
在每个亚样方内随机设置8个固定的PVC土壤环, 其中4个设置在2个1 m × 1 m的切根亚样方内, 将土壤环插入土壤中至2 cm深, 以确保密闭。土壤呼吸采用便携式呼吸测定系统(LI-8100, LI-COR, Lincoln, USA)进行测定。为减少切根处理过程对土壤的扰动和切根所产生的死亡根系的分解对土壤呼吸影响, 土壤呼吸在切根处理14个月后即2013年6月份开始进行测定, 以确保切根小区内的细根完全分解。除受天气等因素的影响无法测定时, 土壤呼吸每月测定一次, 测定时间选择在每月的中旬, 并且在测定前2-3天无明显降水。在测定土壤呼吸的同时, 利用LI-8100便携式呼吸测定系统(LI-COR, Lin- coln, USA)所带的土壤温度和含水量探针测定0-5 cm土层的土壤温度和含水量。土壤根系呼吸采用差值法计算, 为有根系小区的土壤呼吸减去切根小区的土壤呼吸; 而微生物呼吸为切根小区的土壤呼吸。土壤呼吸年通量采用月通量(月通量为每月实测土壤呼吸速率乘以每月的实际天数)累加法计算(范跃新等, 2014 )。
在2013年6月份采集细根和土壤样品。细根生物量采用土钻法测定, 在每个小区中根据距离树干的远近选择9个采样点, 采集10 cm × 10 cm × 10 cm的土块, 将直径小于2 mm的根全部挑出, 用水洗净后烘干称定质量。土壤矿质氮(铵态氮与硝态氮之和)采用2 mol·L-1 的KCl溶液进行提取, 然后采用比色法测定, 其中铵态氮采用靛酚蓝比色法, 利用紫外分光光度计在625 nm处进行比色, 硝态氮利用双波长比色法, 将KCl提取液在220 nm和275 nm处直接比色; 有效磷采用0.025 mol·L-1 的HCl和0.03 mol·L-1 的NH4 F混合样进行提取, 采用钼锑抗比色法测定, 具体操作过程见鲁如坤(2000)。土壤pH值采用1: 2.5的水溶液电位法测定。土壤微生物量碳采用氯仿熏蒸-0.5 mol·L-1 K2 SO4 提取法(鲁如坤, 2000 )测定; 土壤微生物群落结构利用磷脂脂肪酸法测定, 具体操作过程见Wang等(2013)。
1.3 数据统计分析
利用SPSS 17.0软件分析不同施肥处理的土壤呼吸及其环境因子(温度、土壤含水量、有效养分、pH值、细根生物量及养分含量、微生物量碳、微生物群落结构等)、土壤呼吸温度敏感性和呼吸年通量等数据, 不同处理之间影响的差异性采用配对样本t 检验进行比较分析。土壤温度与土壤呼吸的关系采用模型进行分析, 土壤呼吸温度敏感性(Q 10 )利用Fang和Moncrieff (2001)所提出的公式Q 10 = e10 b b 为温度反应系数。差异显著性和相关显著性以α = 0.05为基准。
2 结果和分析
2.1 土壤环境因子
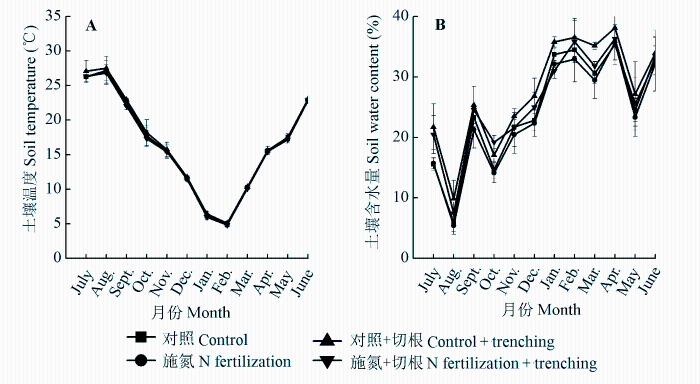
在杉木幼林中, 不同处理之间0-5 cm土层的土壤温度不存在差异, 而且无论是切根小区还是非切根小区, 施氮肥均没有改变土壤温度的月变化动态(图1A )。土壤温度最低值出现在2月份, 而最大值出现在8月份, 分别比大气温度推迟1个月。土壤含水量的峰值出现在8月份和4月份, 但施氮肥对土壤含水量影响不显著(图1B )。
施氮肥改变了土壤养分有效性, 使矿质氮(铵态氮和硝态氮)含量增加了1.66倍, 同时也显著降低了土壤pH值(表2 )。施肥后细根的生物量显著降低, 比对照的细根生物量降低了32.1%。并且在施肥小区, 细根的氮含量也显著增加, 并导致细根C:N显著降低。在施氮肥小区中, 土壤微生物生物量碳的含量仅为对照小区的66.2% (表3 )。土壤细菌和真菌浓度在施氮肥后也分别降低了24.0%和36.3%, 并且微生物的群落结构组成发生了改变, 表现为细菌:真菌从对照的0.42降低为0.34。在细菌中, 革兰氏阳性细菌在施肥后显著降低。
图1 施氮肥和切根对杉木幼林土壤(0-5 cm)温度与含水量的影响(平均值±标准偏差)。
Fig. 1 Influence of N fertilization and root trenching on soil temperature and water content (0-5 cm) in a young Cunninghamia lanceolata plantation (mean ± SD ).
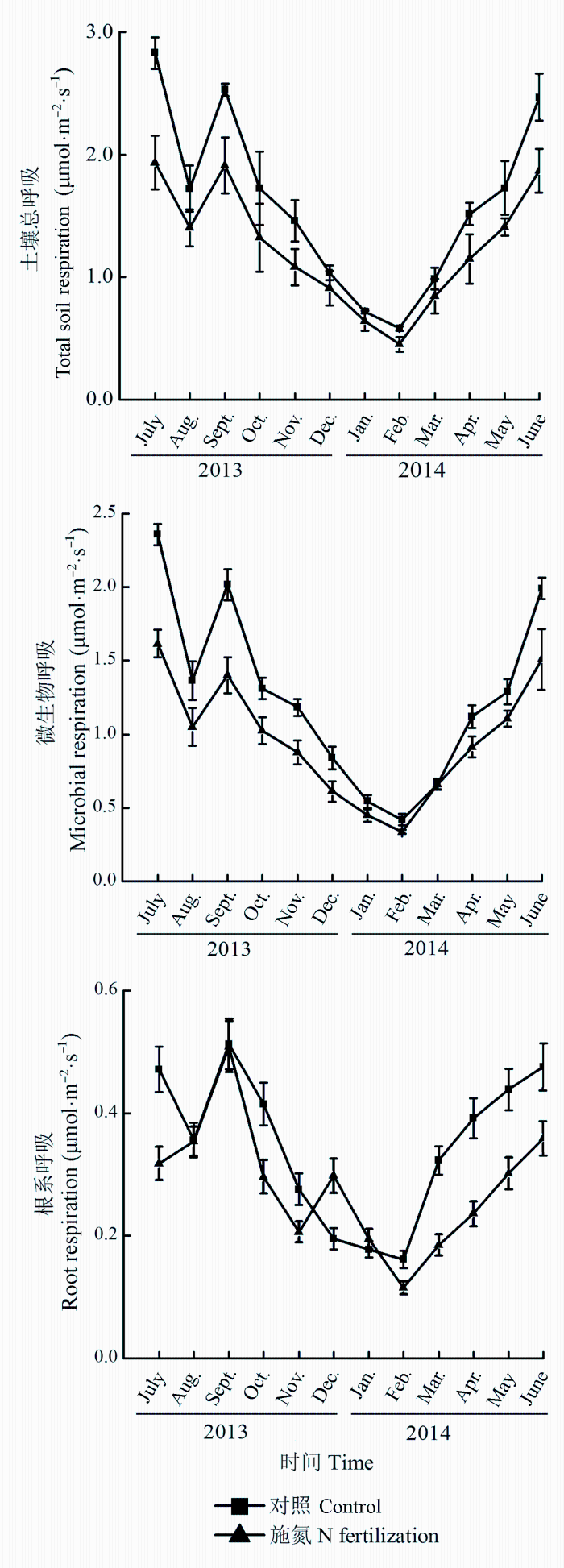
图2 施氮肥对杉木幼林土壤总呼吸、微生物呼吸和根系呼吸的影响(平均值±标准偏差)。
Fig. 2 Influence of N fertilization on total soil, microbial and root respiration in a young Cunninghamia lanceolata plantation (mean ± SD ).
2.2 土壤呼吸速率与年通量
与对照相比, 施氮肥使土壤总呼吸速率降低了11.0%-31.4%, 其中7月份降低幅度最大, 1月份降低幅度最小, 平均下降了22.7% (图2 )。但是施氮肥没有改变土壤总呼吸的变化动态。土壤总呼吸速率的月变化在0.451-2.829 μmol·m-2 ·s-1 之间, 其中分别在2月份和7月份出现最低值和最高值。施氮肥也影响了土壤异养呼吸和自养呼吸, 分别比对照降低了23.5%和19.6%。另外, 土壤异养呼吸和自养呼吸的月变化动态与土壤总呼吸一致, 其变化范围分别在0.336-2.358 μmol·m-2 ·s-1 之间和0.115-0.513 μmol·
m-2 ·s-1 之间(图2 )。
因土壤根系呼吸是采用间接法计算得出的, 没有对应的温度和含水量, 无法通过模型进行模拟。因此, 表4 中只列出了土壤总呼吸和微生物呼吸的模型参数。通过指数模型、线性模型和双因素模型拟合土壤温度、含水量及其交互作用对土壤呼吸的影响(表4 ), 发现土壤呼吸与土壤温度极显著相关, 与土壤含水量的相关性虽低于与土壤温度的相关性, 但也达到了显著或极显著水平。双因素模型 (R s = a ebT Wc T 为气温(℃), a 为温度为0 ℃时的土壤呼吸速率, b 为温度反应系数, c 为水分反应系数, W 为土壤含水量)的模拟效果与单因素指数模型基本没有差别。在指数模型中, 土壤温度可以解释土壤呼吸月动态变化的69.4%-79.5%, 在双因素模型中, 土壤温度和含水量共同解释土壤呼吸月动态变化的69.9%-79.7%, 而在线性模型中, 土壤含水量对土壤呼吸月动态变化的解释仅为1.0%-10.1%。这表明土壤温度是土壤呼吸月动态变化的主要影响因素, 土壤含水量的贡献较低。但是在土壤含水量较低的8月份, 土壤呼吸主要受土壤含水量控制。
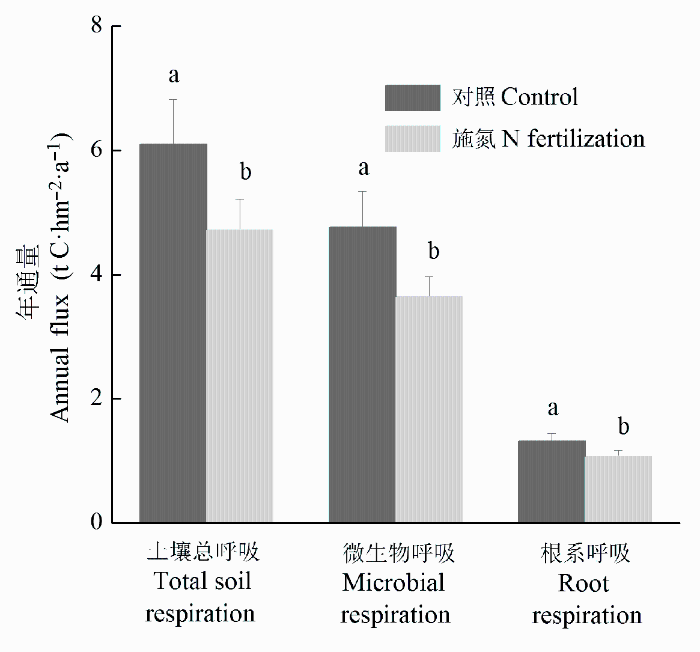
施氮肥后杉木幼林土壤呼吸的年通量也发生了变化(图3 )。施氮肥后土壤总呼吸、微生物呼吸和根系呼吸的年通量是4.71 C·hm-2 、3.64 t C·hm-2 和1.06 t C·hm-2 , 分别比对照降低了22.7%、23.5%和19.6%。施氮肥没有改变根系呼吸和微生物呼吸对土壤总呼吸的贡献。在施氮肥前土壤根系和微生物呼吸年通量对土壤总呼吸年通量的贡献分别为21.7%和78.3%, 施氮肥后其贡献分别为22.6%和77.4%。
图3 杉木幼林土壤总呼吸、微生物呼吸和根系呼吸年通量(平均值±标准偏差)。
Fig. 3 Annual flux of total soil, microbial and root respiration in a young Cunninghamia lanceolata plantation (mean ± SD ).
2.3 土壤呼吸的温度敏感性
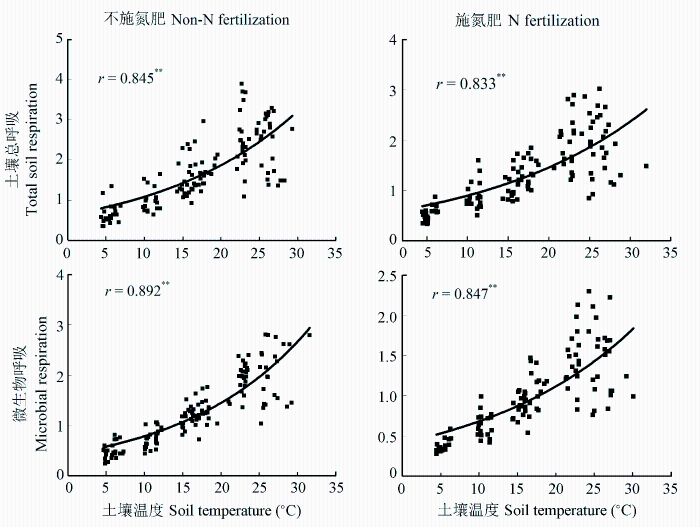
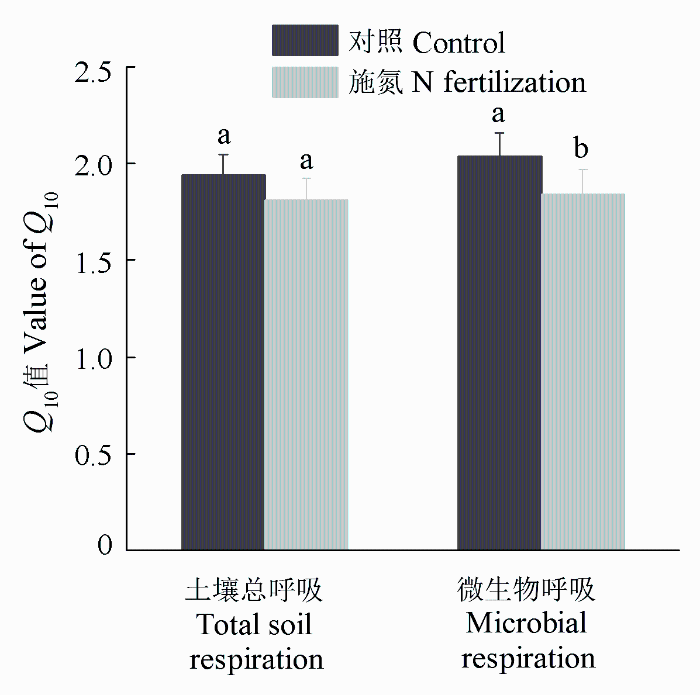
土壤总呼吸速率和微生物呼吸速率均与土壤温度呈很好的指数关系(图4 ), 并且无论是否施氮肥均表现为微生物呼吸与土壤温度的相关系数略大于土壤总呼吸与土壤温度的相关系数。杉木人工幼林土壤总呼吸的温度敏感性(1.944)与微生物呼吸相比差异不显著(图5 )。施氮肥后土壤微生物呼吸的温度敏感性显著降低, 从对照的2.038下降为1.844。施氮肥后土壤总呼吸的温度敏感性为1.809, 与对照相比无显著变化。
3 讨论
施氮肥降低了中亚热带地区杉木幼林土壤呼吸。这一结果与其他地区以前的部分研究结果是一致的(Persson et al ., 1998 ; Olsson et al ., 2005 ; 贾淑霞等, 2007 ; Mo et al ., 2008 ; Janssens et al ., 2010 )。在该研究中, 施氮肥使杉木幼林土壤总呼吸平均降低了22.7%, 这一降幅在已有研究结果的范围之内。例如, 在东北林业大学帽儿山实验林场尖砬沟森林培育实验站, 贾淑霞等(2007)发现外加氮2年后落叶松(Larix gmelinii )和水曲柳(Fraxinus mandshurica )人工林土壤呼吸速率分别降低了34.9%和25.8%。施肥对土壤总呼吸的降低作用主要来自于微生物呼吸(异养呼吸)和根系呼吸(自养呼吸)的降低。但是上述实验没有区分土壤呼吸的不同组分, 无法研究施肥对微生物呼吸和根系呼吸的影响。在该研究中, 我们采用切根法将土壤总呼吸区分为根系呼吸和微生物呼吸, 并发现施氮肥后土壤微生物呼吸和根系呼吸分别降低了23.5%和19.6%。虽然在森林生态系统中挖壕沟-切根法常用来区分和量化根系呼吸与微生物呼吸, 但也存在一些不足。第一, 切根处理后多长时间可以准确量化根系呼吸和土壤微生物呼吸难以确定, 不同树种之间也存在差异。在切根处理后, 切根处理样方内产生了死亡根系, 这些根系完全分解后才能准确量化根系呼吸与微生物呼吸。这是因为: 在死亡根系完全分解前测定土壤呼吸会高估土壤微生物呼吸, 从而使根系呼吸被低估(Tu et al ., 2013 )。在本研究中, 我们虽然在切根处理后14个月才开始测定土壤呼吸, 但也有可能切根处理样方内的死亡细根没有完全分解, 使得根系呼吸被低估。第二, 切根处理可能会影响土壤微环境, 如温度和含水量。土壤环境因子影响土壤呼吸强度(Rustad et al ., 2000 ), 因此也会影响根系呼吸和微生物呼吸测定结果的准确性。在本研究中, 切根处理对土壤温度和含水量基本没有影响。
施氮肥对土壤根系呼吸的降低作用可能是因为氮肥降低了植物光合产物向地下根系的分配, 使得根系生物量和活性降低(Janssens et al ., 2010 )。此外, 光合产物向地下根系分配的减少还可能会对根际土壤产生负激发效应, 从而使土壤呼吸降低(Phillips & Fahey, 2007 )。另一方面, 有研究表明根系的寿命随土壤肥力的提高而降低, 而在低氮的环境中植物可以通过降低根系的周转来减少氮的损失(Pregitzer et al ., 1995 )。因此, 施氮肥后土壤氮素有效性提高, 导致根系死亡率增加和根系现存生物量降低。在挪威云杉(Picea abies )林中, Persson等(1998)发现施氮降低了挪威云杉的根细生物量, 抑制了植物根系活性。在该研究中, 我们也发现施氮肥后0-10 cm土层中杉木细根生物量下降了32.1% (表2 )。在我国东北的尖砬沟森林培育实验站, 梅莉等(2008)也发现了施氮肥后0-10 cm土层中水曲柳活细根生物量下降了13.4%-23.4%。因此, 根系生物量的下降是土壤呼吸降低的重要原因之一。
图4 施氮肥和不施氮肥条件下土壤总呼吸和微生物与土壤温度的关系。
Fig. 4 Relationship of total soil and microbial respiration with soil temperature in the fertilized and unfertilized plots.
图5 杉木幼林施氮肥对土壤总呼吸和微生物呼吸的温度敏感性(Q 10 )的影响(平均值±标准偏差)。
Fig. 5 Effect of N fertilization on the temperature sensitivity (Q 10 ) of total soil and microbial respiration in a young Cunninghamia lanceolata plantation (mean ± SD ).
微生物呼吸主要体现为微生物对土壤有机质的分解而释放CO2 的过程, 微生物群落结构的变化会影响土壤有机质的分解(Janssens et al ., 2010 )。施氮肥改变了土壤微环境, 使土壤微生物群落结构和活性发生了变化, 进而会影响微生物对土壤有机质的分解过程(Treseder, 2008 ; Frey et al ., 2014 )。我们的研究发现了施氮肥后土壤微生物量碳含量以及细菌的浓度显著降低, 细菌:真菌比降低(表3 ), 表明施氮肥改变了土壤微生物群落结构。因此, 施氮肥后杉木幼林土壤微生物活性的下降和群落结构的改变是土壤微生物呼吸降低的另一个重要因素。与我们的研究结果相似, 通过对82个氮添加对微生物量影响的野外研究结果的整合分析, Treseder (2008)发现施氮肥后土壤微生物量平均降低了15%, 并且微生物量对氮肥的响应与土壤CO2 释放的响应显著相关。我们还发现施氮肥降低了土壤真菌的生物量, 这与Frey等(2014)在温带森林生态系统中发现的施肥土壤中的真菌生物量和相对丰度低于不施肥土壤一致。一方面, 由于施氮肥降低了根系生物量(表2 ), 使根系输入到土壤中的碳减少, 抑制了土壤微生物生长, 导致土壤微生物呼吸降低。另一方面, 由于该施肥实验是在南方酸性土壤进行的, 较低的土壤pH值表示土壤可能存在铝毒。虽然该研究没有测定土壤中溶解态铝的含量, 但有研究表明土壤pH值越低, 土壤中溶解态铝含量越高, 这会降低植物对营养元素的吸收(俞元春和丁爱芳, 2001 )。施氮肥降低了土壤pH值(表2 ), 可能导致土壤中溶解态铝含量增加, 使杉木根系生物量和土壤微生物活性降低, 最终导致土壤微生物呼吸作用降低。
土壤温度和含水量是影响土壤呼吸的重要非生物因素(Rustad et al ., 2000 ; Wang et al ., 2004 )。在杉木幼林中, 我们发现土壤呼吸与土壤温度的相关性大于土壤呼吸与土壤含水量的相关性。目前, 在亚热带其他森林中所进行的研究也发现了土壤呼吸与土壤温度存在很好的指数关系, 而与土壤含水量相关性较弱或不相关(Sheng et al ., 2010 ; 韩天丰等, 2011 ; 吴君君等, 2014 ; 范跃新等, 2014 )。除8月份外, 土壤呼吸的季节性变化(图2 )与土壤温度(图1 )高度一致, 这说明在大部分时间内土壤温度是土壤呼吸的主要控制因子, 而土壤含水量不是限制因子。指数模型和双因素模型的模拟结果也充分证实了这一观点。在8月份, 土壤含量低于10%, 限制了土壤呼吸, 导致该月份土壤呼吸降低, 成为土壤呼吸的主要控制因子。这也说明在水热充足的亚热带地区确实存在夏季干旱现象。目前, 已有一些学者研究了夏季干旱对土壤呼吸过程的影响(Welp et al ., 2007 ; Schindlbacher et al ., 2012 )。例如, 在温带云杉林中, Schindlbacher等(2012)认为夏季干旱降低了土壤呼吸, 并在一定程度上可以抵消升温对土壤呼吸的增加效应; Borken等(2006)发现长期的夏季干旱主要降低了土壤的微生物呼吸, 即抑制了土壤有机碳的分解。
土壤呼吸的温度敏感性反映土壤呼吸速率对温度变化的敏感程度, 即温度变化10 ℃时土壤呼吸的相对变化, 常用Q 10 表示(Pregitzer et al ., 2000 )。在杉木幼林生态系统中, 土壤呼吸的Q 10 值在1.81- 2.04之间, 与王传华等(2011)得出的马尾松(Pinus massoniana )林和枫香树(Liquidambar formosana )林(0-5 cm土层)土壤呼吸的Q 10 值分别为1.86和2.10, 范跃新等(2014)得出的中亚热带地区不同演替阶段常绿阔叶林土壤呼吸的Q 10 值在1.79-2.64之间相符。但是, 这些亚热带森林的研究结果低于贾淑霞等(2007)在温带落叶松和水曲柳林中所得的土壤呼吸Q 10 值(2.45-3.29)。在杉木幼林中, 施氮肥虽然对土壤总呼吸温度敏感性的影响不显著, 但土壤总呼吸温度敏感性有降低趋势。土壤呼吸与光合产物供应紧密相关, 施氮降低了光合产物向根系的分配是导致土壤总呼吸温度敏感性呈下降趋势的一个原因(Sun et al ., 2014 )。另一方面, 施氮增加了土壤有机碳的稳定性, 形成更多难分解的有机质(Janssens et al ., 2010 ), 这也会导致土壤总呼吸温度敏感性的降低。同样, 在我国东北的落叶松和水曲柳人工林中, 贾淑霞等(2007)也发现施氮肥后土壤呼吸的Q 10 值没有发生显著变化。他们认为施氮肥没有影响Q 10 值是因为根系组织中氮含量保持不变, 根系呼吸也没有发生变化。这是因为根细胞中维持蛋白质周转和溶质梯度所消耗的能量占根系维持呼吸的绝大部分, 如果根系组织中的氮含量保持不变, 根系呼吸也不会发生变化(Burton et al ., 2002 )。更重要的是, 我们发现施氮肥显著降低了土壤微生物呼吸的Q 10 值。其实, 造成土壤总呼吸温度敏感性降低的因素也可以导致土壤微生物呼吸温度敏感性的降低。另外, 施氮肥可以抑制土壤酶的活性, 对其产生副作用(Aber et al ., 1998 )。这一变化也可以导致土壤微生物呼吸温度敏感性的降低(Sun et al ., 2014 )。这一结果表明施氮肥能减弱全球变暖对土壤有机碳分解的影响。
综上所述, 在杉木幼林中, 施氮肥抑制了土壤根系呼吸和微生物呼吸, 进而导致土壤总呼吸降低。土壤微生物呼吸的降低说明微生物对土壤有机质的分解能力下降, 使土壤有机碳的释放减少。根系生物量和微生物量及活性的降低是施氮肥降低土壤呼吸的主要原因。
致谢 感谢中国科学院会同森林实验站张秀永、于小军和申正其在实验布置工作中给予的帮助,以及黄珂在土壤呼吸测定和于小军在土壤性质分析中所给予的帮助。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Aber J McDowell W Nadelhoffer K Magill A Berntson G Kamakea M McNulty S Currie W Rustad L Fernandez I 1998 ). Nitrogen saturation in temperate forest ecosystems.
BioScience , 48 , 921 -934 .
[本文引用: 1]
[2]
Bond-Lamberty B Thompson A 2010 ). Temperature- associated increases in the global soil respiration record.
Nature , 464 , 579 -582 .
[本文引用: 1]
[3]
Borken W Savage K Davidson EA Trumbore SE 2006 ). Effects of experimental drought on soil respiration and radiocarbon efflux from a temperate forest soil.
Global Change Biology , 12 , 177 -193 .
[4]
Bowden RD Davidson E Savage K Arabia C Steudler P 2004 ). Chronic nitrogen additions reduce total soil respiration and microbial respiration in temperate forest soils at the Harvard Forest.
Forest Ecology and Management , 196 , 43 -56 .
[本文引用: 1]
[5]
Burton AJ Pregitzer KS Ruess RW Hendrick RL Allen MF 2002 ). Root respiration in North American forests: Effects of nitrogen concentration and temperature across biomes.
Oecologia , 131 , 559 -568 .
[本文引用: 1]
[6]
Cleveland CC Townsend AR 2006 ). Nutrient additions to a tropical rain forest drive substantial soil carbon dioxide losses to the atmosphere.
Proceedings of the National Academy of Sciences of the United States of America , 103 , 10316 -10321 .
[本文引用: 1]
[7]
Deluca TH Nilsson MC Zackrisson O 2002 ). Nitrogen mineralization and phenol accumulation along a fire chronosequence in northern Sweden.
Oecologia , 133 , 206 -214 .
[本文引用: 1]
[8]
Department of Forest Resources Management , State Forestry Administration 2010 ). The 7th national forest inventory and status of forest resources.
Forest Resources Management , (1 ), 1 -8 .
[本文引用: 1]
(in Chinese with English abstract) [国家林业局森林资源管理司 (2010 ). 第七次全国森林资源清查及森林资源状况. 林业资源管理,
(1 ), 1 -8 .]
[本文引用: 1]
[9]
Fan YX Yang YS Guo JF Yang ZJ Chen GS Xie JS Zhong XJ Xu LL 2014 ). Changes in soil respiration and its temperature sensitivity at different successional stages of evergreen broadleaved forests in mid-subtropical China.
Chinese Journal of Plant Ecology , 38 , 1155 -1165 .
[本文引用: 2]
(in Chinese with English abstract) [范跃新 , 杨玉盛 , 郭剑芬 , 杨智杰 , 陈光水 , 谢锦升 , 钟小剑 , 徐玲琳 (2014 ). 中亚热带常绿阔叶林不同演替阶段土壤呼吸及其温度敏感性的变化. 植物生态学报,
38 , 1155 -1165 .]
[本文引用: 2]
[10]
Fang C Moncrieff JB 2001 ). The dependence of soil CO2 efflux on temperature.
Soil Biology & Biochemistry , 33 , 155 -165 .
[11]
Fox TR 2000 ). Sustained productivity in intensively managed forest plantations.
Forest Ecology and Management , 138 , 187 -202 .
[本文引用: 1]
[12]
Frey SD Ollinger S Nadelhoffer K Bowden R Brzostek E Burton A Caldwell BA Crow S Goodale CL Grandy AS Finzi A Kramer MG Lajtha K LeMoine J Marin M McDowell WH Minocha R Sadowsky JJ Templer PH Wickings K 2014 ). Chronic nitrogen additions suppress decomposition and sequester soil carbon in temperate forests.
Biogeochemistry , 121 , 305 -316 .
[本文引用: 1]
[13]
Han TF Zhou GY Li YL Liu JX Zhang DQ 2011 ). Partitioning soil respiration in lower subtropical forests at different successional stages in southern China.
Chinese Journal of Plant Ecology , 35 , 946 -954 .
[本文引用: 1]
(in Chinese with English abstract) [韩天丰 , 周国逸 , 李跃林 , 刘菊秀 , 张德强 (2011 ). 中国南亚热带森林不同演替阶段土壤呼吸的分离量化. 植物生态学报,
35 , 946 -954 .]
[本文引用: 1]
[14]
Högberg P Nordgren A Buchmann N Taylor AFS Ekblad A Högberg MN Nyberg G Ottosson-Löfvenius M Read DJ 2001 ). Large-scale forest girdling shows that current photosynthesis drives soil respiration.
Nature , 411 , 789 -792 .
[本文引用: 1]
[15]
Janssens IA Dieleman W Luyssaert S Subke J-A Reichstein M Ceulemans R Ciais P Dolman AJ Grace J Matteucci G Papale D Piao SL Schulze E-D Tang J Law BE 2010 ). Reduction of forest soil respiration in response to nitrogen deposition.
Nature Geoscience , 3 , 315 -322 .
[本文引用: 5]
[16]
Jia SX Wang ZQ Mei L Sun Y Quan XK Shi JW Yu SQ Sun HL Gu JC 2007 ). Effect of nitrogen fertilization on soil respiration in Larix gmelinii and Fraxinus mandshurica plantations in China. Journal of Plant Ecology (
Chinese Version ), 31 , 372 -379 .
[本文引用: 2]
(in Chinese with English abstract) [贾淑霞 , 王政权 , 梅莉 , 孙玥 , 全先奎 , 史建伟 , 于水强 , 孙海龙 , 谷加存 (2007 ). 施肥对落叶松和水曲柳人工林土壤呼吸的影响. 植物生态学报,
31 , 372 -379 .]
[本文引用: 2]
[17]
Lee KH Jose S 2003 ). Soil respiration, fine root production, and microbial biomass in cottonwood and loblolly pine plantations along a nitrogen fertilization gradient.
Forest Ecology and Management , 185 , 263 -273 .
[本文引用: 1]
[18]
Lu RK 2000). Methods of Soil Agricultural Chemistry Analysis . Chinese Agricultural Science and Technology Press, Beijing .
[本文引用: 1]
(in Chinese) [鲁如坤 (2000). 土壤农业化学分析方法. 中国农业科技出版社 , 北京.]
[本文引用: 1]
[19]
Mei L Wang ZQ Zhang XJ Yu LZ Du Y 2008 ). Effects of nitrogen fertilization on fine root biomass production and turnover of Fraxinus mandshurica plantation.
Chinese Journal of Ecology , 27 , 1663 -1668 .
(in Chinese with English abstract) [梅莉 , 王政权 , 张秀娟 , 于立忠 , 杜英 (2008 ). 施氮肥对水曲柳人工林细根生产和周转的影响. 生态学杂志,
27 , 1663 -1668 .]
[20]
Mo JM Zhang W Zhu WX Gundersen P Fang YT Li DJ Wang H 2008 ). Nitrogen addition reduces soil respiration in a mature tropical forest in southern China.
Global Change Biology , 14 , 403 -412 .
[本文引用: 2]
[21]
Olsson P Linder S Giesler R Högberg P 2005 ). Fertilization of boreal forest reduces both autotrophic and heterotrophic soil respiration.
Global Change Biology , 11 , 1745 -1753 .
[本文引用: 3]
[22]
Persson H Ahlström K Clemensson-Lindell A 1998 ). Nitrogen addition and removal at Gårdsjön-effects on fine-root growth and fine-root chemistry.
Forest Ecology and Management , 101 , 199 -205 .
[本文引用: 1]
[23]
Phillips RP Fahey TJ 2007 ). Fertilization effects on fineroot biomass, rhizosphere microbes and respiratory fluxes in hardwood forest soils.
New Phytologist , 176 , 655 -664 .
[本文引用: 1]
[24]
Pregitzer KS King JS Burton AJ Brown SE 2000 ). Responses of tree fine roots to temperature.
New Phytologist , 147 , 105 -115 .
[本文引用: 1]
[25]
Pregitzer KS Zak DR Curtis PS Kubiske M Teeri JA Vogel CS 1995 ). Atmospheric CO2 , soil nitrogen and turnover of fine roots.
New Phytologist , 129 , 579 -585 .
[本文引用: 1]
[26]
Rustad LE Huntington TG Boone RD 2000 ). Controls on soil respiration: Implications for climate change.
Biogeochemistry , 48 , 1 -6 .
[本文引用: 2]
[27]
Samuelson L Mathew R Stokes T Feng YC Aubrey D Coleman M 2009 ). Soil and microbial respiration in a loblolly pine plantation in response to seven years of irrigation and fertilization.
Forest Ecology and Management , 258 , 2431 -2438 .
[本文引用: 1]
[28]
Schindlbacher A Wunderlich S Borken W Kitzler B Zechmeister-Boltenstern S Jandl R 2012 ). Soil respiration under climate change: Prolonged summer drought offsets soil warming effects.
Global Change Biology , 18 , 2270 -2279 .
[本文引用: 1]
[29]
Sheng H Yang YS Yang ZJ Chen GS Xie JS Guo JF Zou SQ 2010 ). The dynamic response of soil respiration to land-use changes in subtropical China.
Global Change Biology , 16 , 1107 -1121 .
[本文引用: 1]
[30]
Sun ZZ Liu LL Ma YC Yin GD Zhao C Zhang Y Piao SL 2014 ). The effect of nitrogen addition on soil respiration from a nitrogen-limited forest soil.
Agricultural and Forest Meteorology , 197 , 103 -110 .
[本文引用: 2]
[31]
Treseder KK 2008 ). Nitrogen additions and microbial biomass: A meta-analysis of ecosystem studies.
Ecology Letters , 11 , 1111 -1120 .
[本文引用: 1]
[32]
Tu LH Hu TX Zhang J Li XW Hu HL Liu L Xiao YL 2013 ). Nitrogen addition stimulates different components of soil respiration in a subtropical bamboo ecosystem.
Soil Biology & Biochemistry , 58 , 255 -264 .
[本文引用: 4]
[33]
Tyree MC Seiler JR Aust WM Sampson DA Fox TR 2006 ). Long-term effects of site preparation and fertilization on total soil CO2 efflux and heterotrophic respiration in a 33-year-old Pinus taeda L. plantation on the wet flats of the Virginia Lower Coastal Plain.
Forest Ecology and Management , 234 , 363 -369 .
[本文引用: 2]
[34]
Wang CH Chen FQ Wang Y Li JQ 2011 ). Soil heterotrophic respiration and its sensitivity to soil temperature and moisture in Liquidambar formosana and Pinus massoniana forests in hilly areas of southeast Hubei Province, China.
Chinese Journal of Applied Ecology , 22 , 600 -606 .
(in Chinese with English abstract) [王传华 , 陈芳清 , 王愿 , 李俊清 (2011 ). 鄂东南低丘马尾松林和枫香林土壤异养呼吸及温湿度敏感性. 应用生态学报,
22 , 600 -606 .]
[35]
Wang M Li QR Xiao DM Dong BL 2004 ). Effects of soil temperature and soil water content on soil respiration in three forest types in Changbai Mountain.
Journal of Forestry Research , 15 , 113 -118 .
[本文引用: 1]
[36]
Wang QK He TX Wang SL Liu L 2013 ). Carbon input manipulation affects soil respiration and microbial community composition in a subtropical coniferous forest.
Agricultural and Forest Meteorology, 178- 179 , 152 -160 .
[37]
Welp LR Randerson JT Liu HP 2007 ). The sensitivity of carbon fluxes to spring warming and summer drought depends on plant functional type in boreal forest ecosystems.
Agricultural and Forest Meteorology , 147 , 172 -185 .
[本文引用: 1]
[38]
Wu JJ Yang ZJ Liu XF Xiong DC Lin WS Chen CQ Wang XH 2014 ). Analysis of soil respiration and components in Castanopsis carlesii and Cunninghamia lanceolata plantations.
Chinese Journal of Plant Ecology , 38 , 45 -53 .
[本文引用: 1]
(in Chinese with English abstract) [吴君君 , 杨智杰 , 刘小飞 , 熊德成 , 林伟盛 , 陈朝琪 , 王小红 (2014 ). 米槠和杉木人工林土壤呼吸及其组分分析. 植物生态学报,
38 , 45 -53 .]
[本文引用: 1]
[39]
Xu JD 2014 ). The 8th forest resources inventory results and analysis in China.
Forestry Economics , (3 ), 6 -8 .
[本文引用: 3]
(in Chinese with English abstract) [徐济德 (2014 ). 我国第八次森林资源清查结果及分析. 林业经济,
(3 ), 6 -8 .]
[本文引用: 3]
[40]
Yang J Li LH 2003 ). Soil respiration and its measuring method.
Chinese Journal of Plant , (5 ), 36 -37 .
[本文引用: 1]
(in Chinese) [杨晶 , 李凌浩 (2003 ). 土壤呼吸及其测定法. 植物杂志,
(5 ), 36 -37 .]
[本文引用: 1]
[41]
Yu MJ Xu XH Li MH Fu HL 2005 ). Biocycle of nitrogen in a Cyclobalanopsis glauca dominated evergreen broad leaved forest in East China.
Acta Ecologica Sinica , 25 , 740 -748 .
[本文引用: 1]
(in Chinese with English abstract) [于明坚 , 徐学红 , 李铭红 , 付海龙 (2005 ). 青冈常绿阔叶林氮的生物循环. 生态学报,
25 , 740 -748 .]
[本文引用: 1]
[42]
Yu YC Ding AF 2001 ). Effects of simulated acid rain on dissolution and transformation of aluminum in acid soils of south China.
Soil and Environmental Sciences , 10 (2 ), 87 -90 .
[本文引用: 1]
(in Chinese with English abstract) [俞元春 , 丁爱芳 (2001 ). 模拟酸雨对酸性土壤铝溶出及其形态转化的影响. 土壤与环境,
10(2) , 87 -90 .]
[本文引用: 1]
[43]
Zhou CP Ouyang H Pei ZY Xu XL 2003 ). Net soil nitrogen mineralization in Chinese forest ecosystems.
Acta Phytoecologica Sinica , 27 , 170 -176 .
[本文引用: 1]
[44]
Zhu B Cheng WX 2011 ). Rhizosphere priming effect increases the temperature sensitivity of soil organic matter decomposition.
Global Change Biology , 17 , 2172 -2183 .
[本文引用: 1]
Nitrogen saturation in temperate forest ecosystems.
1
1998
... 土壤呼吸的温度敏感性反映土壤呼吸速率对温度变化的敏感程度, 即温度变化10 ℃时土壤呼吸的相对变化, 常用Q 10 表示(Pregitzer et al ., 2000 ).在杉木幼林生态系统中, 土壤呼吸的Q 10 值在1.81- 2.04之间, 与王传华等(2011)得出的马尾松(Pinus massoniana )林和枫香树(Liquidambar formosana )林(0-5 cm土层)土壤呼吸的Q 10 值分别为1.86和2.10, 范跃新等(2014)得出的中亚热带地区不同演替阶段常绿阔叶林土壤呼吸的Q 10 值在1.79-2.64之间相符.但是, 这些亚热带森林的研究结果低于贾淑霞等(2007)在温带落叶松和水曲柳林中所得的土壤呼吸Q 10 值(2.45-3.29).在杉木幼林中, 施氮肥虽然对土壤总呼吸温度敏感性的影响不显著, 但土壤总呼吸温度敏感性有降低趋势.土壤呼吸与光合产物供应紧密相关, 施氮降低了光合产物向根系的分配是导致土壤总呼吸温度敏感性呈下降趋势的一个原因(Sun et al ., 2014 ).另一方面, 施氮增加了土壤有机碳的稳定性, 形成更多难分解的有机质(Janssens et al ., 2010 ), 这也会导致土壤总呼吸温度敏感性的降低.同样, 在我国东北的落叶松和水曲柳人工林中, 贾淑霞等(2007)也发现施氮肥后土壤呼吸的Q 10 值没有发生显著变化.他们认为施氮肥没有影响Q 10 值是因为根系组织中氮含量保持不变, 根系呼吸也没有发生变化.这是因为根细胞中维持蛋白质周转和溶质梯度所消耗的能量占根系维持呼吸的绝大部分, 如果根系组织中的氮含量保持不变, 根系呼吸也不会发生变化(Burton et al ., 2002 ).更重要的是, 我们发现施氮肥显著降低了土壤微生物呼吸的Q 10 值.其实, 造成土壤总呼吸温度敏感性降低的因素也可以导致土壤微生物呼吸温度敏感性的降低.另外, 施氮肥可以抑制土壤酶的活性, 对其产生副作用(Aber et al ., 1998 ).这一变化也可以导致土壤微生物呼吸温度敏感性的降低(Sun et al ., 2014 ).这一结果表明施氮肥能减弱全球变暖对土壤有机碳分解的影响. ...
Temperature- associated increases in the global soil respiration record.
1
2010
... 土壤呼吸是陆地生态系统碳循环的重要环节, 是在土壤新陈代谢功能过程中, 土壤产生大量的CO2 , 并向大气中释放CO2 的过程(杨晶和李凌浩, 2003 ).据估计, 每年从土壤释放到大气中的碳高达98 Pg·a-1 (Bond-Lamberty & Thompson, 2010 ), 相当于化石燃料燃烧所释放碳的10倍多(Zhu & Cheng, 2011 ).因此, 土壤呼吸速率的微小变化都可能会对大气中CO2 浓度产生巨大影响.在森林生态系统中, 施氮肥对土壤呼吸的影响国内外已有大量研究报道, 但不同树种的森林生态系统土壤呼吸对施氮肥的响应之间存在差异, 主要表现为促进(Cleveland & Townsend, 2006 ; Tu et al ., 2013 )、抑制(Bowden et al ., 2004 ; Olsson et al ., 2005 ; 贾淑霞等, 2007 ; Mo et al ., 2008 ; Janssens et al ., 2010 )或无影响(Samuelson et al ., 2009 ).例如, 施氮肥使亚热带竹(Pleioblastus amarus )林生态系统的土壤呼吸增加26%-52% (Tu et al ., 2013), 使北方森林生态系统的土壤呼吸降低40% (Olsson et al ., 2005 ).施氮肥在不同森林和区域对土壤呼吸的影响存在差异, 主要与树种特性和土壤性质有关, 也与各实验所施用氮肥的类型和施用时间长短有关(Tu et al ., 2013). ...
Effects of experimental drought on soil respiration and radiocarbon efflux from a temperate forest soil.
0
2006
Chronic nitrogen additions reduce total soil respiration and microbial respiration in temperate forest soils at the Harvard Forest.
1
2004
... 土壤呼吸是陆地生态系统碳循环的重要环节, 是在土壤新陈代谢功能过程中, 土壤产生大量的CO2 , 并向大气中释放CO2 的过程(杨晶和李凌浩, 2003 ).据估计, 每年从土壤释放到大气中的碳高达98 Pg·a-1 (Bond-Lamberty & Thompson, 2010 ), 相当于化石燃料燃烧所释放碳的10倍多(Zhu & Cheng, 2011 ).因此, 土壤呼吸速率的微小变化都可能会对大气中CO2 浓度产生巨大影响.在森林生态系统中, 施氮肥对土壤呼吸的影响国内外已有大量研究报道, 但不同树种的森林生态系统土壤呼吸对施氮肥的响应之间存在差异, 主要表现为促进(Cleveland & Townsend, 2006 ; Tu et al ., 2013 )、抑制(Bowden et al ., 2004 ; Olsson et al ., 2005 ; 贾淑霞等, 2007 ; Mo et al ., 2008 ; Janssens et al ., 2010 )或无影响(Samuelson et al ., 2009 ).例如, 施氮肥使亚热带竹(Pleioblastus amarus )林生态系统的土壤呼吸增加26%-52% (Tu et al ., 2013), 使北方森林生态系统的土壤呼吸降低40% (Olsson et al ., 2005 ).施氮肥在不同森林和区域对土壤呼吸的影响存在差异, 主要与树种特性和土壤性质有关, 也与各实验所施用氮肥的类型和施用时间长短有关(Tu et al ., 2013). ...
Root respiration in North American forests: Effects of nitrogen concentration and temperature across biomes.
1
2002
... 土壤呼吸的温度敏感性反映土壤呼吸速率对温度变化的敏感程度, 即温度变化10 ℃时土壤呼吸的相对变化, 常用Q 10 表示(Pregitzer et al ., 2000 ).在杉木幼林生态系统中, 土壤呼吸的Q 10 值在1.81- 2.04之间, 与王传华等(2011)得出的马尾松(Pinus massoniana )林和枫香树(Liquidambar formosana )林(0-5 cm土层)土壤呼吸的Q 10 值分别为1.86和2.10, 范跃新等(2014)得出的中亚热带地区不同演替阶段常绿阔叶林土壤呼吸的Q 10 值在1.79-2.64之间相符.但是, 这些亚热带森林的研究结果低于贾淑霞等(2007)在温带落叶松和水曲柳林中所得的土壤呼吸Q 10 值(2.45-3.29).在杉木幼林中, 施氮肥虽然对土壤总呼吸温度敏感性的影响不显著, 但土壤总呼吸温度敏感性有降低趋势.土壤呼吸与光合产物供应紧密相关, 施氮降低了光合产物向根系的分配是导致土壤总呼吸温度敏感性呈下降趋势的一个原因(Sun et al ., 2014 ).另一方面, 施氮增加了土壤有机碳的稳定性, 形成更多难分解的有机质(Janssens et al ., 2010 ), 这也会导致土壤总呼吸温度敏感性的降低.同样, 在我国东北的落叶松和水曲柳人工林中, 贾淑霞等(2007)也发现施氮肥后土壤呼吸的Q 10 值没有发生显著变化.他们认为施氮肥没有影响Q 10 值是因为根系组织中氮含量保持不变, 根系呼吸也没有发生变化.这是因为根细胞中维持蛋白质周转和溶质梯度所消耗的能量占根系维持呼吸的绝大部分, 如果根系组织中的氮含量保持不变, 根系呼吸也不会发生变化(Burton et al ., 2002 ).更重要的是, 我们发现施氮肥显著降低了土壤微生物呼吸的Q 10 值.其实, 造成土壤总呼吸温度敏感性降低的因素也可以导致土壤微生物呼吸温度敏感性的降低.另外, 施氮肥可以抑制土壤酶的活性, 对其产生副作用(Aber et al ., 1998 ).这一变化也可以导致土壤微生物呼吸温度敏感性的降低(Sun et al ., 2014 ).这一结果表明施氮肥能减弱全球变暖对土壤有机碳分解的影响. ...
Nutrient additions to a tropical rain forest drive substantial soil carbon dioxide losses to the atmosphere.
1
2006
... 土壤呼吸是陆地生态系统碳循环的重要环节, 是在土壤新陈代谢功能过程中, 土壤产生大量的CO2 , 并向大气中释放CO2 的过程(杨晶和李凌浩, 2003 ).据估计, 每年从土壤释放到大气中的碳高达98 Pg·a-1 (Bond-Lamberty & Thompson, 2010 ), 相当于化石燃料燃烧所释放碳的10倍多(Zhu & Cheng, 2011 ).因此, 土壤呼吸速率的微小变化都可能会对大气中CO2 浓度产生巨大影响.在森林生态系统中, 施氮肥对土壤呼吸的影响国内外已有大量研究报道, 但不同树种的森林生态系统土壤呼吸对施氮肥的响应之间存在差异, 主要表现为促进(Cleveland & Townsend, 2006 ; Tu et al ., 2013 )、抑制(Bowden et al ., 2004 ; Olsson et al ., 2005 ; 贾淑霞等, 2007 ; Mo et al ., 2008 ; Janssens et al ., 2010 )或无影响(Samuelson et al ., 2009 ).例如, 施氮肥使亚热带竹(Pleioblastus amarus )林生态系统的土壤呼吸增加26%-52% (Tu et al ., 2013), 使北方森林生态系统的土壤呼吸降低40% (Olsson et al ., 2005 ).施氮肥在不同森林和区域对土壤呼吸的影响存在差异, 主要与树种特性和土壤性质有关, 也与各实验所施用氮肥的类型和施用时间长短有关(Tu et al ., 2013). ...
Nitrogen mineralization and phenol accumulation along a fire chronosequence in northern Sweden.
1
2002
... 第八次全国森林资源清查结果显示, 我国人工林面积已达6900多万hm2 , 约占全球人工林面积的1/4, 在我国木材生产和生态安全保障中起着至关重要的作用(徐济德, 2014 ).在我国实行天然林保护的背景下, 人工林, 尤其是用材林在提供林产品方面的作用愈加重要, 并且其面积还在持续增加(徐济德, 2014 ).但是, 我国人工林管理粗放、经营水平低, 导致其生产力不高, 平均蓄积量仅为52.8 m3 ·hm-2 , 不到发达国家的1/3 (徐济德, 2014 ).要提高人工林生产和生态服务功能, 亟需改变过去传统的粗放式管理模式, 实行高度集约式经营.在大部分区域, 土壤氮素缺乏是森林生态系统生产力最重要的限制因子(Deluca et al ., 2002 ; Zhou et al ., 2003 ; 于明坚等, 2005 ).为解决氮素的限制, 施氮肥已经成为提高人工林生产力的一种高度集约式经营管理措施(Fox, 2000 ; Lee & Jose, 2003 ).但是, 施肥在提高林地生产力的同时, 还会对土壤呼吸等地下生态过程产生深刻影响. ...
第七次全国森林资源清查及森林资源状况. 林业资源管理,
1
2010
... 杉木(Cunninghamia lanceolata )人工林在我国亚热带地区的种植面积已接近900万hm2 , 在木材生产和生态安全保障等方面均具有举足轻重的作用(国家林业局森林资源管理司, 2010 ).为探讨施氮肥对杉木人工林土壤呼吸的影响, 我们在湖南会同林区的杉木幼林生态系统中开展了施肥试验, 以研究施氮肥对杉木幼林土壤呼吸的影响.相对于土壤总呼吸的变化, 了解土壤呼吸不同组分对施氮肥的响应更有助于认知土壤碳释放过程和对其进行有效的调控.因此, 为了明确土壤呼吸中不同组分(即根系呼吸和微生物呼吸)对施加氮肥的响应, 我们采用挖壕沟-切根法来区分土壤根系呼吸和微生物呼吸.在该研究中, 我们测定了土壤呼吸、根系生物量、微生物群落结构及相关环境因子的变化, 以期明确在杉木幼林生态系统中施氮肥对土壤总呼吸、根系呼吸和微生物呼吸的影响及其机理, 为杉木人工林土壤固碳增汇管理提供基础数据和理论依据, 该研究结果也将有助于我们了解氮沉降增加对中亚热带森林生态系统土壤碳释放的影响. ...
第七次全国森林资源清查及森林资源状况. 林业资源管理,
1
2010
... 杉木(Cunninghamia lanceolata )人工林在我国亚热带地区的种植面积已接近900万hm2 , 在木材生产和生态安全保障等方面均具有举足轻重的作用(国家林业局森林资源管理司, 2010 ).为探讨施氮肥对杉木人工林土壤呼吸的影响, 我们在湖南会同林区的杉木幼林生态系统中开展了施肥试验, 以研究施氮肥对杉木幼林土壤呼吸的影响.相对于土壤总呼吸的变化, 了解土壤呼吸不同组分对施氮肥的响应更有助于认知土壤碳释放过程和对其进行有效的调控.因此, 为了明确土壤呼吸中不同组分(即根系呼吸和微生物呼吸)对施加氮肥的响应, 我们采用挖壕沟-切根法来区分土壤根系呼吸和微生物呼吸.在该研究中, 我们测定了土壤呼吸、根系生物量、微生物群落结构及相关环境因子的变化, 以期明确在杉木幼林生态系统中施氮肥对土壤总呼吸、根系呼吸和微生物呼吸的影响及其机理, 为杉木人工林土壤固碳增汇管理提供基础数据和理论依据, 该研究结果也将有助于我们了解氮沉降增加对中亚热带森林生态系统土壤碳释放的影响. ...
中亚热带常绿阔叶林不同演替阶段土壤呼吸及其温度敏感性的变化. 植物生态学报,
2
2014
... 在每个亚样方内随机设置8个固定的PVC土壤环, 其中4个设置在2个1 m × 1 m的切根亚样方内, 将土壤环插入土壤中至2 cm深, 以确保密闭.土壤呼吸采用便携式呼吸测定系统(LI-8100, LI-COR, Lincoln, USA)进行测定.为减少切根处理过程对土壤的扰动和切根所产生的死亡根系的分解对土壤呼吸影响, 土壤呼吸在切根处理14个月后即2013年6月份开始进行测定, 以确保切根小区内的细根完全分解.除受天气等因素的影响无法测定时, 土壤呼吸每月测定一次, 测定时间选择在每月的中旬, 并且在测定前2-3天无明显降水.在测定土壤呼吸的同时, 利用LI-8100便携式呼吸测定系统(LI-COR, Lin- coln, USA)所带的土壤温度和含水量探针测定0-5 cm土层的土壤温度和含水量.土壤根系呼吸采用差值法计算, 为有根系小区的土壤呼吸减去切根小区的土壤呼吸; 而微生物呼吸为切根小区的土壤呼吸.土壤呼吸年通量采用月通量(月通量为每月实测土壤呼吸速率乘以每月的实际天数)累加法计算(范跃新等, 2014 ). ...
... 土壤温度和含水量是影响土壤呼吸的重要非生物因素(Rustad et al ., 2000 ; Wang et al ., 2004 ).在杉木幼林中, 我们发现土壤呼吸与土壤温度的相关性大于土壤呼吸与土壤含水量的相关性.目前, 在亚热带其他森林中所进行的研究也发现了土壤呼吸与土壤温度存在很好的指数关系, 而与土壤含水量相关性较弱或不相关(Sheng et al ., 2010 ; 韩天丰等, 2011 ; 吴君君等, 2014 ; 范跃新等, 2014 ).除8月份外, 土壤呼吸的季节性变化(图2 )与土壤温度(图1 )高度一致, 这说明在大部分时间内土壤温度是土壤呼吸的主要控制因子, 而土壤含水量不是限制因子.指数模型和双因素模型的模拟结果也充分证实了这一观点.在8月份, 土壤含量低于10%, 限制了土壤呼吸, 导致该月份土壤呼吸降低, 成为土壤呼吸的主要控制因子.这也说明在水热充足的亚热带地区确实存在夏季干旱现象.目前, 已有一些学者研究了夏季干旱对土壤呼吸过程的影响(Welp et al ., 2007 ; Schindlbacher et al ., 2012 ).例如, 在温带云杉林中, Schindlbacher等(2012)认为夏季干旱降低了土壤呼吸, 并在一定程度上可以抵消升温对土壤呼吸的增加效应; Borken等(2006)发现长期的夏季干旱主要降低了土壤的微生物呼吸, 即抑制了土壤有机碳的分解. ...
中亚热带常绿阔叶林不同演替阶段土壤呼吸及其温度敏感性的变化. 植物生态学报,
2
2014
... 在每个亚样方内随机设置8个固定的PVC土壤环, 其中4个设置在2个1 m × 1 m的切根亚样方内, 将土壤环插入土壤中至2 cm深, 以确保密闭.土壤呼吸采用便携式呼吸测定系统(LI-8100, LI-COR, Lincoln, USA)进行测定.为减少切根处理过程对土壤的扰动和切根所产生的死亡根系的分解对土壤呼吸影响, 土壤呼吸在切根处理14个月后即2013年6月份开始进行测定, 以确保切根小区内的细根完全分解.除受天气等因素的影响无法测定时, 土壤呼吸每月测定一次, 测定时间选择在每月的中旬, 并且在测定前2-3天无明显降水.在测定土壤呼吸的同时, 利用LI-8100便携式呼吸测定系统(LI-COR, Lin- coln, USA)所带的土壤温度和含水量探针测定0-5 cm土层的土壤温度和含水量.土壤根系呼吸采用差值法计算, 为有根系小区的土壤呼吸减去切根小区的土壤呼吸; 而微生物呼吸为切根小区的土壤呼吸.土壤呼吸年通量采用月通量(月通量为每月实测土壤呼吸速率乘以每月的实际天数)累加法计算(范跃新等, 2014 ). ...
... 土壤温度和含水量是影响土壤呼吸的重要非生物因素(Rustad et al ., 2000 ; Wang et al ., 2004 ).在杉木幼林中, 我们发现土壤呼吸与土壤温度的相关性大于土壤呼吸与土壤含水量的相关性.目前, 在亚热带其他森林中所进行的研究也发现了土壤呼吸与土壤温度存在很好的指数关系, 而与土壤含水量相关性较弱或不相关(Sheng et al ., 2010 ; 韩天丰等, 2011 ; 吴君君等, 2014 ; 范跃新等, 2014 ).除8月份外, 土壤呼吸的季节性变化(图2 )与土壤温度(图1 )高度一致, 这说明在大部分时间内土壤温度是土壤呼吸的主要控制因子, 而土壤含水量不是限制因子.指数模型和双因素模型的模拟结果也充分证实了这一观点.在8月份, 土壤含量低于10%, 限制了土壤呼吸, 导致该月份土壤呼吸降低, 成为土壤呼吸的主要控制因子.这也说明在水热充足的亚热带地区确实存在夏季干旱现象.目前, 已有一些学者研究了夏季干旱对土壤呼吸过程的影响(Welp et al ., 2007 ; Schindlbacher et al ., 2012 ).例如, 在温带云杉林中, Schindlbacher等(2012)认为夏季干旱降低了土壤呼吸, 并在一定程度上可以抵消升温对土壤呼吸的增加效应; Borken等(2006)发现长期的夏季干旱主要降低了土壤的微生物呼吸, 即抑制了土壤有机碳的分解. ...
The dependence of soil CO2 efflux on temperature.
0
2001
Sustained productivity in intensively managed forest plantations.
1
2000
... 第八次全国森林资源清查结果显示, 我国人工林面积已达6900多万hm2 , 约占全球人工林面积的1/4, 在我国木材生产和生态安全保障中起着至关重要的作用(徐济德, 2014 ).在我国实行天然林保护的背景下, 人工林, 尤其是用材林在提供林产品方面的作用愈加重要, 并且其面积还在持续增加(徐济德, 2014 ).但是, 我国人工林管理粗放、经营水平低, 导致其生产力不高, 平均蓄积量仅为52.8 m3 ·hm-2 , 不到发达国家的1/3 (徐济德, 2014 ).要提高人工林生产和生态服务功能, 亟需改变过去传统的粗放式管理模式, 实行高度集约式经营.在大部分区域, 土壤氮素缺乏是森林生态系统生产力最重要的限制因子(Deluca et al ., 2002 ; Zhou et al ., 2003 ; 于明坚等, 2005 ).为解决氮素的限制, 施氮肥已经成为提高人工林生产力的一种高度集约式经营管理措施(Fox, 2000 ; Lee & Jose, 2003 ).但是, 施肥在提高林地生产力的同时, 还会对土壤呼吸等地下生态过程产生深刻影响. ...
Chronic nitrogen additions suppress decomposition and sequester soil carbon in temperate forests.
1
2014
... 微生物呼吸主要体现为微生物对土壤有机质的分解而释放CO2 的过程, 微生物群落结构的变化会影响土壤有机质的分解(Janssens et al ., 2010 ).施氮肥改变了土壤微环境, 使土壤微生物群落结构和活性发生了变化, 进而会影响微生物对土壤有机质的分解过程(Treseder, 2008 ; Frey et al ., 2014 ).我们的研究发现了施氮肥后土壤微生物量碳含量以及细菌的浓度显著降低, 细菌:真菌比降低(表3 ), 表明施氮肥改变了土壤微生物群落结构.因此, 施氮肥后杉木幼林土壤微生物活性的下降和群落结构的改变是土壤微生物呼吸降低的另一个重要因素.与我们的研究结果相似, 通过对82个氮添加对微生物量影响的野外研究结果的整合分析, Treseder (2008)发现施氮肥后土壤微生物量平均降低了15%, 并且微生物量对氮肥的响应与土壤CO2 释放的响应显著相关.我们还发现施氮肥降低了土壤真菌的生物量, 这与Frey等(2014)在温带森林生态系统中发现的施肥土壤中的真菌生物量和相对丰度低于不施肥土壤一致.一方面, 由于施氮肥降低了根系生物量(表2 ), 使根系输入到土壤中的碳减少, 抑制了土壤微生物生长, 导致土壤微生物呼吸降低.另一方面, 由于该施肥实验是在南方酸性土壤进行的, 较低的土壤pH值表示土壤可能存在铝毒.虽然该研究没有测定土壤中溶解态铝的含量, 但有研究表明土壤pH值越低, 土壤中溶解态铝含量越高, 这会降低植物对营养元素的吸收(俞元春和丁爱芳, 2001 ).施氮肥降低了土壤pH值(表2 ), 可能导致土壤中溶解态铝含量增加, 使杉木根系生物量和土壤微生物活性降低, 最终导致土壤微生物呼吸作用降低. ...
中国南亚热带森林不同演替阶段土壤呼吸的分离量化. 植物生态学报,
1
2011
... 土壤温度和含水量是影响土壤呼吸的重要非生物因素(Rustad et al ., 2000 ; Wang et al ., 2004 ).在杉木幼林中, 我们发现土壤呼吸与土壤温度的相关性大于土壤呼吸与土壤含水量的相关性.目前, 在亚热带其他森林中所进行的研究也发现了土壤呼吸与土壤温度存在很好的指数关系, 而与土壤含水量相关性较弱或不相关(Sheng et al ., 2010 ; 韩天丰等, 2011 ; 吴君君等, 2014 ; 范跃新等, 2014 ).除8月份外, 土壤呼吸的季节性变化(图2 )与土壤温度(图1 )高度一致, 这说明在大部分时间内土壤温度是土壤呼吸的主要控制因子, 而土壤含水量不是限制因子.指数模型和双因素模型的模拟结果也充分证实了这一观点.在8月份, 土壤含量低于10%, 限制了土壤呼吸, 导致该月份土壤呼吸降低, 成为土壤呼吸的主要控制因子.这也说明在水热充足的亚热带地区确实存在夏季干旱现象.目前, 已有一些学者研究了夏季干旱对土壤呼吸过程的影响(Welp et al ., 2007 ; Schindlbacher et al ., 2012 ).例如, 在温带云杉林中, Schindlbacher等(2012)认为夏季干旱降低了土壤呼吸, 并在一定程度上可以抵消升温对土壤呼吸的增加效应; Borken等(2006)发现长期的夏季干旱主要降低了土壤的微生物呼吸, 即抑制了土壤有机碳的分解. ...
中国南亚热带森林不同演替阶段土壤呼吸的分离量化. 植物生态学报,
1
2011
... 土壤温度和含水量是影响土壤呼吸的重要非生物因素(Rustad et al ., 2000 ; Wang et al ., 2004 ).在杉木幼林中, 我们发现土壤呼吸与土壤温度的相关性大于土壤呼吸与土壤含水量的相关性.目前, 在亚热带其他森林中所进行的研究也发现了土壤呼吸与土壤温度存在很好的指数关系, 而与土壤含水量相关性较弱或不相关(Sheng et al ., 2010 ; 韩天丰等, 2011 ; 吴君君等, 2014 ; 范跃新等, 2014 ).除8月份外, 土壤呼吸的季节性变化(图2 )与土壤温度(图1 )高度一致, 这说明在大部分时间内土壤温度是土壤呼吸的主要控制因子, 而土壤含水量不是限制因子.指数模型和双因素模型的模拟结果也充分证实了这一观点.在8月份, 土壤含量低于10%, 限制了土壤呼吸, 导致该月份土壤呼吸降低, 成为土壤呼吸的主要控制因子.这也说明在水热充足的亚热带地区确实存在夏季干旱现象.目前, 已有一些学者研究了夏季干旱对土壤呼吸过程的影响(Welp et al ., 2007 ; Schindlbacher et al ., 2012 ).例如, 在温带云杉林中, Schindlbacher等(2012)认为夏季干旱降低了土壤呼吸, 并在一定程度上可以抵消升温对土壤呼吸的增加效应; Borken等(2006)发现长期的夏季干旱主要降低了土壤的微生物呼吸, 即抑制了土壤有机碳的分解. ...
Large-scale forest girdling shows that current photosynthesis drives soil respiration.
1
2001
... 土壤呼吸可以分为自养呼吸和异养呼吸.在我们的研究中自养呼吸主要是指植物根系呼吸, 而异养呼吸主要是指土壤微生物呼吸.这些组分对施肥的响应至关重要, 关系到生态系统碳平衡(Tyree et al ., 2006 ).目前主要采用挖壕沟-切根法、树干环割等方法来区分植物根系呼吸和土壤微生物呼吸(Högberg et al ., 2001 ; Tu et al ., 2013 ).但是, 目前大部分实验只是研究了土壤总呼吸对施氮肥的响应, 而关于根系呼吸和微生物呼吸对施氮肥的响应还研究得比较少, 为数不多的研究所取得的结果还相互矛盾(Tyree et al ., 2006 ; Tu et al ., 2013 ).例如, Tu等(2013)在竹林中发现施氮肥促进了根系呼吸和微生物呼吸; 而Janssens等(2010)通过整合分析发现施氮肥后土壤微生物呼吸降低15%.在亚热带地区, 关于施肥对人工林土壤呼吸过程影响的研究, 尤其是对土壤呼吸不同组分的研究还极其有限, 这严重地限制了我们对施肥措施对土壤碳循环过程影响的深入理解, 不利于人工林的可持续经营管理. ...
Reduction of forest soil respiration in response to nitrogen deposition.
5
2010
... 土壤呼吸是陆地生态系统碳循环的重要环节, 是在土壤新陈代谢功能过程中, 土壤产生大量的CO2 , 并向大气中释放CO2 的过程(杨晶和李凌浩, 2003 ).据估计, 每年从土壤释放到大气中的碳高达98 Pg·a-1 (Bond-Lamberty & Thompson, 2010 ), 相当于化石燃料燃烧所释放碳的10倍多(Zhu & Cheng, 2011 ).因此, 土壤呼吸速率的微小变化都可能会对大气中CO2 浓度产生巨大影响.在森林生态系统中, 施氮肥对土壤呼吸的影响国内外已有大量研究报道, 但不同树种的森林生态系统土壤呼吸对施氮肥的响应之间存在差异, 主要表现为促进(Cleveland & Townsend, 2006 ; Tu et al ., 2013 )、抑制(Bowden et al ., 2004 ; Olsson et al ., 2005 ; 贾淑霞等, 2007 ; Mo et al ., 2008 ; Janssens et al ., 2010 )或无影响(Samuelson et al ., 2009 ).例如, 施氮肥使亚热带竹(Pleioblastus amarus )林生态系统的土壤呼吸增加26%-52% (Tu et al ., 2013), 使北方森林生态系统的土壤呼吸降低40% (Olsson et al ., 2005 ).施氮肥在不同森林和区域对土壤呼吸的影响存在差异, 主要与树种特性和土壤性质有关, 也与各实验所施用氮肥的类型和施用时间长短有关(Tu et al ., 2013). ...
... 施氮肥降低了中亚热带地区杉木幼林土壤呼吸.这一结果与其他地区以前的部分研究结果是一致的(Persson et al ., 1998 ; Olsson et al ., 2005 ; 贾淑霞等, 2007 ; Mo et al ., 2008 ; Janssens et al ., 2010 ).在该研究中, 施氮肥使杉木幼林土壤总呼吸平均降低了22.7%, 这一降幅在已有研究结果的范围之内.例如, 在东北林业大学帽儿山实验林场尖砬沟森林培育实验站, 贾淑霞等(2007)发现外加氮2年后落叶松(Larix gmelinii )和水曲柳(Fraxinus mandshurica )人工林土壤呼吸速率分别降低了34.9%和25.8%.施肥对土壤总呼吸的降低作用主要来自于微生物呼吸(异养呼吸)和根系呼吸(自养呼吸)的降低.但是上述实验没有区分土壤呼吸的不同组分, 无法研究施肥对微生物呼吸和根系呼吸的影响.在该研究中, 我们采用切根法将土壤总呼吸区分为根系呼吸和微生物呼吸, 并发现施氮肥后土壤微生物呼吸和根系呼吸分别降低了23.5%和19.6%.虽然在森林生态系统中挖壕沟-切根法常用来区分和量化根系呼吸与微生物呼吸, 但也存在一些不足.第一, 切根处理后多长时间可以准确量化根系呼吸和土壤微生物呼吸难以确定, 不同树种之间也存在差异.在切根处理后, 切根处理样方内产生了死亡根系, 这些根系完全分解后才能准确量化根系呼吸与微生物呼吸.这是因为: 在死亡根系完全分解前测定土壤呼吸会高估土壤微生物呼吸, 从而使根系呼吸被低估(Tu et al ., 2013 ).在本研究中, 我们虽然在切根处理后14个月才开始测定土壤呼吸, 但也有可能切根处理样方内的死亡细根没有完全分解, 使得根系呼吸被低估.第二, 切根处理可能会影响土壤微环境, 如温度和含水量.土壤环境因子影响土壤呼吸强度(Rustad et al ., 2000 ), 因此也会影响根系呼吸和微生物呼吸测定结果的准确性.在本研究中, 切根处理对土壤温度和含水量基本没有影响. ...
... 施氮肥对土壤根系呼吸的降低作用可能是因为氮肥降低了植物光合产物向地下根系的分配, 使得根系生物量和活性降低(Janssens et al ., 2010 ).此外, 光合产物向地下根系分配的减少还可能会对根际土壤产生负激发效应, 从而使土壤呼吸降低(Phillips & Fahey, 2007 ).另一方面, 有研究表明根系的寿命随土壤肥力的提高而降低, 而在低氮的环境中植物可以通过降低根系的周转来减少氮的损失(Pregitzer et al ., 1995 ).因此, 施氮肥后土壤氮素有效性提高, 导致根系死亡率增加和根系现存生物量降低.在挪威云杉(Picea abies )林中, Persson等(1998)发现施氮降低了挪威云杉的根细生物量, 抑制了植物根系活性.在该研究中, 我们也发现施氮肥后0-10 cm土层中杉木细根生物量下降了32.1% (表2 ).在我国东北的尖砬沟森林培育实验站, 梅莉等(2008)也发现了施氮肥后0-10 cm土层中水曲柳活细根生物量下降了13.4%-23.4%.因此, 根系生物量的下降是土壤呼吸降低的重要原因之一. ...
... 微生物呼吸主要体现为微生物对土壤有机质的分解而释放CO2 的过程, 微生物群落结构的变化会影响土壤有机质的分解(Janssens et al ., 2010 ).施氮肥改变了土壤微环境, 使土壤微生物群落结构和活性发生了变化, 进而会影响微生物对土壤有机质的分解过程(Treseder, 2008 ; Frey et al ., 2014 ).我们的研究发现了施氮肥后土壤微生物量碳含量以及细菌的浓度显著降低, 细菌:真菌比降低(表3 ), 表明施氮肥改变了土壤微生物群落结构.因此, 施氮肥后杉木幼林土壤微生物活性的下降和群落结构的改变是土壤微生物呼吸降低的另一个重要因素.与我们的研究结果相似, 通过对82个氮添加对微生物量影响的野外研究结果的整合分析, Treseder (2008)发现施氮肥后土壤微生物量平均降低了15%, 并且微生物量对氮肥的响应与土壤CO2 释放的响应显著相关.我们还发现施氮肥降低了土壤真菌的生物量, 这与Frey等(2014)在温带森林生态系统中发现的施肥土壤中的真菌生物量和相对丰度低于不施肥土壤一致.一方面, 由于施氮肥降低了根系生物量(表2 ), 使根系输入到土壤中的碳减少, 抑制了土壤微生物生长, 导致土壤微生物呼吸降低.另一方面, 由于该施肥实验是在南方酸性土壤进行的, 较低的土壤pH值表示土壤可能存在铝毒.虽然该研究没有测定土壤中溶解态铝的含量, 但有研究表明土壤pH值越低, 土壤中溶解态铝含量越高, 这会降低植物对营养元素的吸收(俞元春和丁爱芳, 2001 ).施氮肥降低了土壤pH值(表2 ), 可能导致土壤中溶解态铝含量增加, 使杉木根系生物量和土壤微生物活性降低, 最终导致土壤微生物呼吸作用降低. ...
... 土壤呼吸的温度敏感性反映土壤呼吸速率对温度变化的敏感程度, 即温度变化10 ℃时土壤呼吸的相对变化, 常用Q 10 表示(Pregitzer et al ., 2000 ).在杉木幼林生态系统中, 土壤呼吸的Q 10 值在1.81- 2.04之间, 与王传华等(2011)得出的马尾松(Pinus massoniana )林和枫香树(Liquidambar formosana )林(0-5 cm土层)土壤呼吸的Q 10 值分别为1.86和2.10, 范跃新等(2014)得出的中亚热带地区不同演替阶段常绿阔叶林土壤呼吸的Q 10 值在1.79-2.64之间相符.但是, 这些亚热带森林的研究结果低于贾淑霞等(2007)在温带落叶松和水曲柳林中所得的土壤呼吸Q 10 值(2.45-3.29).在杉木幼林中, 施氮肥虽然对土壤总呼吸温度敏感性的影响不显著, 但土壤总呼吸温度敏感性有降低趋势.土壤呼吸与光合产物供应紧密相关, 施氮降低了光合产物向根系的分配是导致土壤总呼吸温度敏感性呈下降趋势的一个原因(Sun et al ., 2014 ).另一方面, 施氮增加了土壤有机碳的稳定性, 形成更多难分解的有机质(Janssens et al ., 2010 ), 这也会导致土壤总呼吸温度敏感性的降低.同样, 在我国东北的落叶松和水曲柳人工林中, 贾淑霞等(2007)也发现施氮肥后土壤呼吸的Q 10 值没有发生显著变化.他们认为施氮肥没有影响Q 10 值是因为根系组织中氮含量保持不变, 根系呼吸也没有发生变化.这是因为根细胞中维持蛋白质周转和溶质梯度所消耗的能量占根系维持呼吸的绝大部分, 如果根系组织中的氮含量保持不变, 根系呼吸也不会发生变化(Burton et al ., 2002 ).更重要的是, 我们发现施氮肥显著降低了土壤微生物呼吸的Q 10 值.其实, 造成土壤总呼吸温度敏感性降低的因素也可以导致土壤微生物呼吸温度敏感性的降低.另外, 施氮肥可以抑制土壤酶的活性, 对其产生副作用(Aber et al ., 1998 ).这一变化也可以导致土壤微生物呼吸温度敏感性的降低(Sun et al ., 2014 ).这一结果表明施氮肥能减弱全球变暖对土壤有机碳分解的影响. ...
施肥对落叶松和水曲柳人工林土壤呼吸的影响. 植物生态学报,
2
2007
... 土壤呼吸是陆地生态系统碳循环的重要环节, 是在土壤新陈代谢功能过程中, 土壤产生大量的CO2 , 并向大气中释放CO2 的过程(杨晶和李凌浩, 2003 ).据估计, 每年从土壤释放到大气中的碳高达98 Pg·a-1 (Bond-Lamberty & Thompson, 2010 ), 相当于化石燃料燃烧所释放碳的10倍多(Zhu & Cheng, 2011 ).因此, 土壤呼吸速率的微小变化都可能会对大气中CO2 浓度产生巨大影响.在森林生态系统中, 施氮肥对土壤呼吸的影响国内外已有大量研究报道, 但不同树种的森林生态系统土壤呼吸对施氮肥的响应之间存在差异, 主要表现为促进(Cleveland & Townsend, 2006 ; Tu et al ., 2013 )、抑制(Bowden et al ., 2004 ; Olsson et al ., 2005 ; 贾淑霞等, 2007 ; Mo et al ., 2008 ; Janssens et al ., 2010 )或无影响(Samuelson et al ., 2009 ).例如, 施氮肥使亚热带竹(Pleioblastus amarus )林生态系统的土壤呼吸增加26%-52% (Tu et al ., 2013), 使北方森林生态系统的土壤呼吸降低40% (Olsson et al ., 2005 ).施氮肥在不同森林和区域对土壤呼吸的影响存在差异, 主要与树种特性和土壤性质有关, 也与各实验所施用氮肥的类型和施用时间长短有关(Tu et al ., 2013). ...
... 施氮肥降低了中亚热带地区杉木幼林土壤呼吸.这一结果与其他地区以前的部分研究结果是一致的(Persson et al ., 1998 ; Olsson et al ., 2005 ; 贾淑霞等, 2007 ; Mo et al ., 2008 ; Janssens et al ., 2010 ).在该研究中, 施氮肥使杉木幼林土壤总呼吸平均降低了22.7%, 这一降幅在已有研究结果的范围之内.例如, 在东北林业大学帽儿山实验林场尖砬沟森林培育实验站, 贾淑霞等(2007)发现外加氮2年后落叶松(Larix gmelinii )和水曲柳(Fraxinus mandshurica )人工林土壤呼吸速率分别降低了34.9%和25.8%.施肥对土壤总呼吸的降低作用主要来自于微生物呼吸(异养呼吸)和根系呼吸(自养呼吸)的降低.但是上述实验没有区分土壤呼吸的不同组分, 无法研究施肥对微生物呼吸和根系呼吸的影响.在该研究中, 我们采用切根法将土壤总呼吸区分为根系呼吸和微生物呼吸, 并发现施氮肥后土壤微生物呼吸和根系呼吸分别降低了23.5%和19.6%.虽然在森林生态系统中挖壕沟-切根法常用来区分和量化根系呼吸与微生物呼吸, 但也存在一些不足.第一, 切根处理后多长时间可以准确量化根系呼吸和土壤微生物呼吸难以确定, 不同树种之间也存在差异.在切根处理后, 切根处理样方内产生了死亡根系, 这些根系完全分解后才能准确量化根系呼吸与微生物呼吸.这是因为: 在死亡根系完全分解前测定土壤呼吸会高估土壤微生物呼吸, 从而使根系呼吸被低估(Tu et al ., 2013 ).在本研究中, 我们虽然在切根处理后14个月才开始测定土壤呼吸, 但也有可能切根处理样方内的死亡细根没有完全分解, 使得根系呼吸被低估.第二, 切根处理可能会影响土壤微环境, 如温度和含水量.土壤环境因子影响土壤呼吸强度(Rustad et al ., 2000 ), 因此也会影响根系呼吸和微生物呼吸测定结果的准确性.在本研究中, 切根处理对土壤温度和含水量基本没有影响. ...
施肥对落叶松和水曲柳人工林土壤呼吸的影响. 植物生态学报,
2
2007
... 土壤呼吸是陆地生态系统碳循环的重要环节, 是在土壤新陈代谢功能过程中, 土壤产生大量的CO2 , 并向大气中释放CO2 的过程(杨晶和李凌浩, 2003 ).据估计, 每年从土壤释放到大气中的碳高达98 Pg·a-1 (Bond-Lamberty & Thompson, 2010 ), 相当于化石燃料燃烧所释放碳的10倍多(Zhu & Cheng, 2011 ).因此, 土壤呼吸速率的微小变化都可能会对大气中CO2 浓度产生巨大影响.在森林生态系统中, 施氮肥对土壤呼吸的影响国内外已有大量研究报道, 但不同树种的森林生态系统土壤呼吸对施氮肥的响应之间存在差异, 主要表现为促进(Cleveland & Townsend, 2006 ; Tu et al ., 2013 )、抑制(Bowden et al ., 2004 ; Olsson et al ., 2005 ; 贾淑霞等, 2007 ; Mo et al ., 2008 ; Janssens et al ., 2010 )或无影响(Samuelson et al ., 2009 ).例如, 施氮肥使亚热带竹(Pleioblastus amarus )林生态系统的土壤呼吸增加26%-52% (Tu et al ., 2013), 使北方森林生态系统的土壤呼吸降低40% (Olsson et al ., 2005 ).施氮肥在不同森林和区域对土壤呼吸的影响存在差异, 主要与树种特性和土壤性质有关, 也与各实验所施用氮肥的类型和施用时间长短有关(Tu et al ., 2013). ...
... 施氮肥降低了中亚热带地区杉木幼林土壤呼吸.这一结果与其他地区以前的部分研究结果是一致的(Persson et al ., 1998 ; Olsson et al ., 2005 ; 贾淑霞等, 2007 ; Mo et al ., 2008 ; Janssens et al ., 2010 ).在该研究中, 施氮肥使杉木幼林土壤总呼吸平均降低了22.7%, 这一降幅在已有研究结果的范围之内.例如, 在东北林业大学帽儿山实验林场尖砬沟森林培育实验站, 贾淑霞等(2007)发现外加氮2年后落叶松(Larix gmelinii )和水曲柳(Fraxinus mandshurica )人工林土壤呼吸速率分别降低了34.9%和25.8%.施肥对土壤总呼吸的降低作用主要来自于微生物呼吸(异养呼吸)和根系呼吸(自养呼吸)的降低.但是上述实验没有区分土壤呼吸的不同组分, 无法研究施肥对微生物呼吸和根系呼吸的影响.在该研究中, 我们采用切根法将土壤总呼吸区分为根系呼吸和微生物呼吸, 并发现施氮肥后土壤微生物呼吸和根系呼吸分别降低了23.5%和19.6%.虽然在森林生态系统中挖壕沟-切根法常用来区分和量化根系呼吸与微生物呼吸, 但也存在一些不足.第一, 切根处理后多长时间可以准确量化根系呼吸和土壤微生物呼吸难以确定, 不同树种之间也存在差异.在切根处理后, 切根处理样方内产生了死亡根系, 这些根系完全分解后才能准确量化根系呼吸与微生物呼吸.这是因为: 在死亡根系完全分解前测定土壤呼吸会高估土壤微生物呼吸, 从而使根系呼吸被低估(Tu et al ., 2013 ).在本研究中, 我们虽然在切根处理后14个月才开始测定土壤呼吸, 但也有可能切根处理样方内的死亡细根没有完全分解, 使得根系呼吸被低估.第二, 切根处理可能会影响土壤微环境, 如温度和含水量.土壤环境因子影响土壤呼吸强度(Rustad et al ., 2000 ), 因此也会影响根系呼吸和微生物呼吸测定结果的准确性.在本研究中, 切根处理对土壤温度和含水量基本没有影响. ...
Soil respiration, fine root production, and microbial biomass in cottonwood and loblolly pine plantations along a nitrogen fertilization gradient.
1
2003
... 第八次全国森林资源清查结果显示, 我国人工林面积已达6900多万hm2 , 约占全球人工林面积的1/4, 在我国木材生产和生态安全保障中起着至关重要的作用(徐济德, 2014 ).在我国实行天然林保护的背景下, 人工林, 尤其是用材林在提供林产品方面的作用愈加重要, 并且其面积还在持续增加(徐济德, 2014 ).但是, 我国人工林管理粗放、经营水平低, 导致其生产力不高, 平均蓄积量仅为52.8 m3 ·hm-2 , 不到发达国家的1/3 (徐济德, 2014 ).要提高人工林生产和生态服务功能, 亟需改变过去传统的粗放式管理模式, 实行高度集约式经营.在大部分区域, 土壤氮素缺乏是森林生态系统生产力最重要的限制因子(Deluca et al ., 2002 ; Zhou et al ., 2003 ; 于明坚等, 2005 ).为解决氮素的限制, 施氮肥已经成为提高人工林生产力的一种高度集约式经营管理措施(Fox, 2000 ; Lee & Jose, 2003 ).但是, 施肥在提高林地生产力的同时, 还会对土壤呼吸等地下生态过程产生深刻影响. ...
1
... 在2013年6月份采集细根和土壤样品.细根生物量采用土钻法测定, 在每个小区中根据距离树干的远近选择9个采样点, 采集10 cm × 10 cm × 10 cm的土块, 将直径小于2 mm的根全部挑出, 用水洗净后烘干称定质量.土壤矿质氮(铵态氮与硝态氮之和)采用2 mol·L-1 的KCl溶液进行提取, 然后采用比色法测定, 其中铵态氮采用靛酚蓝比色法, 利用紫外分光光度计在625 nm处进行比色, 硝态氮利用双波长比色法, 将KCl提取液在220 nm和275 nm处直接比色; 有效磷采用0.025 mol·L-1 的HCl和0.03 mol·L-1 的NH4 F混合样进行提取, 采用钼锑抗比色法测定, 具体操作过程见鲁如坤(2000).土壤pH值采用1: 2.5的水溶液电位法测定.土壤微生物量碳采用氯仿熏蒸-0.5 mol·L-1 K2 SO4 提取法(鲁如坤, 2000 )测定; 土壤微生物群落结构利用磷脂脂肪酸法测定, 具体操作过程见Wang等(2013). ...
1
... 在2013年6月份采集细根和土壤样品.细根生物量采用土钻法测定, 在每个小区中根据距离树干的远近选择9个采样点, 采集10 cm × 10 cm × 10 cm的土块, 将直径小于2 mm的根全部挑出, 用水洗净后烘干称定质量.土壤矿质氮(铵态氮与硝态氮之和)采用2 mol·L-1 的KCl溶液进行提取, 然后采用比色法测定, 其中铵态氮采用靛酚蓝比色法, 利用紫外分光光度计在625 nm处进行比色, 硝态氮利用双波长比色法, 将KCl提取液在220 nm和275 nm处直接比色; 有效磷采用0.025 mol·L-1 的HCl和0.03 mol·L-1 的NH4 F混合样进行提取, 采用钼锑抗比色法测定, 具体操作过程见鲁如坤(2000).土壤pH值采用1: 2.5的水溶液电位法测定.土壤微生物量碳采用氯仿熏蒸-0.5 mol·L-1 K2 SO4 提取法(鲁如坤, 2000 )测定; 土壤微生物群落结构利用磷脂脂肪酸法测定, 具体操作过程见Wang等(2013). ...
施氮肥对水曲柳人工林细根生产和周转的影响. 生态学杂志,
0
2008
施氮肥对水曲柳人工林细根生产和周转的影响. 生态学杂志,
0
2008
Nitrogen addition reduces soil respiration in a mature tropical forest in southern China.
2
2008
... 土壤呼吸是陆地生态系统碳循环的重要环节, 是在土壤新陈代谢功能过程中, 土壤产生大量的CO2 , 并向大气中释放CO2 的过程(杨晶和李凌浩, 2003 ).据估计, 每年从土壤释放到大气中的碳高达98 Pg·a-1 (Bond-Lamberty & Thompson, 2010 ), 相当于化石燃料燃烧所释放碳的10倍多(Zhu & Cheng, 2011 ).因此, 土壤呼吸速率的微小变化都可能会对大气中CO2 浓度产生巨大影响.在森林生态系统中, 施氮肥对土壤呼吸的影响国内外已有大量研究报道, 但不同树种的森林生态系统土壤呼吸对施氮肥的响应之间存在差异, 主要表现为促进(Cleveland & Townsend, 2006 ; Tu et al ., 2013 )、抑制(Bowden et al ., 2004 ; Olsson et al ., 2005 ; 贾淑霞等, 2007 ; Mo et al ., 2008 ; Janssens et al ., 2010 )或无影响(Samuelson et al ., 2009 ).例如, 施氮肥使亚热带竹(Pleioblastus amarus )林生态系统的土壤呼吸增加26%-52% (Tu et al ., 2013), 使北方森林生态系统的土壤呼吸降低40% (Olsson et al ., 2005 ).施氮肥在不同森林和区域对土壤呼吸的影响存在差异, 主要与树种特性和土壤性质有关, 也与各实验所施用氮肥的类型和施用时间长短有关(Tu et al ., 2013). ...
... 施氮肥降低了中亚热带地区杉木幼林土壤呼吸.这一结果与其他地区以前的部分研究结果是一致的(Persson et al ., 1998 ; Olsson et al ., 2005 ; 贾淑霞等, 2007 ; Mo et al ., 2008 ; Janssens et al ., 2010 ).在该研究中, 施氮肥使杉木幼林土壤总呼吸平均降低了22.7%, 这一降幅在已有研究结果的范围之内.例如, 在东北林业大学帽儿山实验林场尖砬沟森林培育实验站, 贾淑霞等(2007)发现外加氮2年后落叶松(Larix gmelinii )和水曲柳(Fraxinus mandshurica )人工林土壤呼吸速率分别降低了34.9%和25.8%.施肥对土壤总呼吸的降低作用主要来自于微生物呼吸(异养呼吸)和根系呼吸(自养呼吸)的降低.但是上述实验没有区分土壤呼吸的不同组分, 无法研究施肥对微生物呼吸和根系呼吸的影响.在该研究中, 我们采用切根法将土壤总呼吸区分为根系呼吸和微生物呼吸, 并发现施氮肥后土壤微生物呼吸和根系呼吸分别降低了23.5%和19.6%.虽然在森林生态系统中挖壕沟-切根法常用来区分和量化根系呼吸与微生物呼吸, 但也存在一些不足.第一, 切根处理后多长时间可以准确量化根系呼吸和土壤微生物呼吸难以确定, 不同树种之间也存在差异.在切根处理后, 切根处理样方内产生了死亡根系, 这些根系完全分解后才能准确量化根系呼吸与微生物呼吸.这是因为: 在死亡根系完全分解前测定土壤呼吸会高估土壤微生物呼吸, 从而使根系呼吸被低估(Tu et al ., 2013 ).在本研究中, 我们虽然在切根处理后14个月才开始测定土壤呼吸, 但也有可能切根处理样方内的死亡细根没有完全分解, 使得根系呼吸被低估.第二, 切根处理可能会影响土壤微环境, 如温度和含水量.土壤环境因子影响土壤呼吸强度(Rustad et al ., 2000 ), 因此也会影响根系呼吸和微生物呼吸测定结果的准确性.在本研究中, 切根处理对土壤温度和含水量基本没有影响. ...
Fertilization of boreal forest reduces both autotrophic and heterotrophic soil respiration.
3
2005
... 土壤呼吸是陆地生态系统碳循环的重要环节, 是在土壤新陈代谢功能过程中, 土壤产生大量的CO2 , 并向大气中释放CO2 的过程(杨晶和李凌浩, 2003 ).据估计, 每年从土壤释放到大气中的碳高达98 Pg·a-1 (Bond-Lamberty & Thompson, 2010 ), 相当于化石燃料燃烧所释放碳的10倍多(Zhu & Cheng, 2011 ).因此, 土壤呼吸速率的微小变化都可能会对大气中CO2 浓度产生巨大影响.在森林生态系统中, 施氮肥对土壤呼吸的影响国内外已有大量研究报道, 但不同树种的森林生态系统土壤呼吸对施氮肥的响应之间存在差异, 主要表现为促进(Cleveland & Townsend, 2006 ; Tu et al ., 2013 )、抑制(Bowden et al ., 2004 ; Olsson et al ., 2005 ; 贾淑霞等, 2007 ; Mo et al ., 2008 ; Janssens et al ., 2010 )或无影响(Samuelson et al ., 2009 ).例如, 施氮肥使亚热带竹(Pleioblastus amarus )林生态系统的土壤呼吸增加26%-52% (Tu et al ., 2013), 使北方森林生态系统的土壤呼吸降低40% (Olsson et al ., 2005 ).施氮肥在不同森林和区域对土壤呼吸的影响存在差异, 主要与树种特性和土壤性质有关, 也与各实验所施用氮肥的类型和施用时间长短有关(Tu et al ., 2013). ...
... ., 2013), 使北方森林生态系统的土壤呼吸降低40% (Olsson et al ., 2005 ).施氮肥在不同森林和区域对土壤呼吸的影响存在差异, 主要与树种特性和土壤性质有关, 也与各实验所施用氮肥的类型和施用时间长短有关(Tu et al ., 2013). ...
... 施氮肥降低了中亚热带地区杉木幼林土壤呼吸.这一结果与其他地区以前的部分研究结果是一致的(Persson et al ., 1998 ; Olsson et al ., 2005 ; 贾淑霞等, 2007 ; Mo et al ., 2008 ; Janssens et al ., 2010 ).在该研究中, 施氮肥使杉木幼林土壤总呼吸平均降低了22.7%, 这一降幅在已有研究结果的范围之内.例如, 在东北林业大学帽儿山实验林场尖砬沟森林培育实验站, 贾淑霞等(2007)发现外加氮2年后落叶松(Larix gmelinii )和水曲柳(Fraxinus mandshurica )人工林土壤呼吸速率分别降低了34.9%和25.8%.施肥对土壤总呼吸的降低作用主要来自于微生物呼吸(异养呼吸)和根系呼吸(自养呼吸)的降低.但是上述实验没有区分土壤呼吸的不同组分, 无法研究施肥对微生物呼吸和根系呼吸的影响.在该研究中, 我们采用切根法将土壤总呼吸区分为根系呼吸和微生物呼吸, 并发现施氮肥后土壤微生物呼吸和根系呼吸分别降低了23.5%和19.6%.虽然在森林生态系统中挖壕沟-切根法常用来区分和量化根系呼吸与微生物呼吸, 但也存在一些不足.第一, 切根处理后多长时间可以准确量化根系呼吸和土壤微生物呼吸难以确定, 不同树种之间也存在差异.在切根处理后, 切根处理样方内产生了死亡根系, 这些根系完全分解后才能准确量化根系呼吸与微生物呼吸.这是因为: 在死亡根系完全分解前测定土壤呼吸会高估土壤微生物呼吸, 从而使根系呼吸被低估(Tu et al ., 2013 ).在本研究中, 我们虽然在切根处理后14个月才开始测定土壤呼吸, 但也有可能切根处理样方内的死亡细根没有完全分解, 使得根系呼吸被低估.第二, 切根处理可能会影响土壤微环境, 如温度和含水量.土壤环境因子影响土壤呼吸强度(Rustad et al ., 2000 ), 因此也会影响根系呼吸和微生物呼吸测定结果的准确性.在本研究中, 切根处理对土壤温度和含水量基本没有影响. ...
Nitrogen addition and removal at Gårdsjön-effects on fine-root growth and fine-root chemistry.
1
1998
... 施氮肥降低了中亚热带地区杉木幼林土壤呼吸.这一结果与其他地区以前的部分研究结果是一致的(Persson et al ., 1998 ; Olsson et al ., 2005 ; 贾淑霞等, 2007 ; Mo et al ., 2008 ; Janssens et al ., 2010 ).在该研究中, 施氮肥使杉木幼林土壤总呼吸平均降低了22.7%, 这一降幅在已有研究结果的范围之内.例如, 在东北林业大学帽儿山实验林场尖砬沟森林培育实验站, 贾淑霞等(2007)发现外加氮2年后落叶松(Larix gmelinii )和水曲柳(Fraxinus mandshurica )人工林土壤呼吸速率分别降低了34.9%和25.8%.施肥对土壤总呼吸的降低作用主要来自于微生物呼吸(异养呼吸)和根系呼吸(自养呼吸)的降低.但是上述实验没有区分土壤呼吸的不同组分, 无法研究施肥对微生物呼吸和根系呼吸的影响.在该研究中, 我们采用切根法将土壤总呼吸区分为根系呼吸和微生物呼吸, 并发现施氮肥后土壤微生物呼吸和根系呼吸分别降低了23.5%和19.6%.虽然在森林生态系统中挖壕沟-切根法常用来区分和量化根系呼吸与微生物呼吸, 但也存在一些不足.第一, 切根处理后多长时间可以准确量化根系呼吸和土壤微生物呼吸难以确定, 不同树种之间也存在差异.在切根处理后, 切根处理样方内产生了死亡根系, 这些根系完全分解后才能准确量化根系呼吸与微生物呼吸.这是因为: 在死亡根系完全分解前测定土壤呼吸会高估土壤微生物呼吸, 从而使根系呼吸被低估(Tu et al ., 2013 ).在本研究中, 我们虽然在切根处理后14个月才开始测定土壤呼吸, 但也有可能切根处理样方内的死亡细根没有完全分解, 使得根系呼吸被低估.第二, 切根处理可能会影响土壤微环境, 如温度和含水量.土壤环境因子影响土壤呼吸强度(Rustad et al ., 2000 ), 因此也会影响根系呼吸和微生物呼吸测定结果的准确性.在本研究中, 切根处理对土壤温度和含水量基本没有影响. ...
Fertilization effects on fineroot biomass, rhizosphere microbes and respiratory fluxes in hardwood forest soils.
1
2007
... 施氮肥对土壤根系呼吸的降低作用可能是因为氮肥降低了植物光合产物向地下根系的分配, 使得根系生物量和活性降低(Janssens et al ., 2010 ).此外, 光合产物向地下根系分配的减少还可能会对根际土壤产生负激发效应, 从而使土壤呼吸降低(Phillips & Fahey, 2007 ).另一方面, 有研究表明根系的寿命随土壤肥力的提高而降低, 而在低氮的环境中植物可以通过降低根系的周转来减少氮的损失(Pregitzer et al ., 1995 ).因此, 施氮肥后土壤氮素有效性提高, 导致根系死亡率增加和根系现存生物量降低.在挪威云杉(Picea abies )林中, Persson等(1998)发现施氮降低了挪威云杉的根细生物量, 抑制了植物根系活性.在该研究中, 我们也发现施氮肥后0-10 cm土层中杉木细根生物量下降了32.1% (表2 ).在我国东北的尖砬沟森林培育实验站, 梅莉等(2008)也发现了施氮肥后0-10 cm土层中水曲柳活细根生物量下降了13.4%-23.4%.因此, 根系生物量的下降是土壤呼吸降低的重要原因之一. ...
Responses of tree fine roots to temperature.
1
2000
... 土壤呼吸的温度敏感性反映土壤呼吸速率对温度变化的敏感程度, 即温度变化10 ℃时土壤呼吸的相对变化, 常用Q 10 表示(Pregitzer et al ., 2000 ).在杉木幼林生态系统中, 土壤呼吸的Q 10 值在1.81- 2.04之间, 与王传华等(2011)得出的马尾松(Pinus massoniana )林和枫香树(Liquidambar formosana )林(0-5 cm土层)土壤呼吸的Q 10 值分别为1.86和2.10, 范跃新等(2014)得出的中亚热带地区不同演替阶段常绿阔叶林土壤呼吸的Q 10 值在1.79-2.64之间相符.但是, 这些亚热带森林的研究结果低于贾淑霞等(2007)在温带落叶松和水曲柳林中所得的土壤呼吸Q 10 值(2.45-3.29).在杉木幼林中, 施氮肥虽然对土壤总呼吸温度敏感性的影响不显著, 但土壤总呼吸温度敏感性有降低趋势.土壤呼吸与光合产物供应紧密相关, 施氮降低了光合产物向根系的分配是导致土壤总呼吸温度敏感性呈下降趋势的一个原因(Sun et al ., 2014 ).另一方面, 施氮增加了土壤有机碳的稳定性, 形成更多难分解的有机质(Janssens et al ., 2010 ), 这也会导致土壤总呼吸温度敏感性的降低.同样, 在我国东北的落叶松和水曲柳人工林中, 贾淑霞等(2007)也发现施氮肥后土壤呼吸的Q 10 值没有发生显著变化.他们认为施氮肥没有影响Q 10 值是因为根系组织中氮含量保持不变, 根系呼吸也没有发生变化.这是因为根细胞中维持蛋白质周转和溶质梯度所消耗的能量占根系维持呼吸的绝大部分, 如果根系组织中的氮含量保持不变, 根系呼吸也不会发生变化(Burton et al ., 2002 ).更重要的是, 我们发现施氮肥显著降低了土壤微生物呼吸的Q 10 值.其实, 造成土壤总呼吸温度敏感性降低的因素也可以导致土壤微生物呼吸温度敏感性的降低.另外, 施氮肥可以抑制土壤酶的活性, 对其产生副作用(Aber et al ., 1998 ).这一变化也可以导致土壤微生物呼吸温度敏感性的降低(Sun et al ., 2014 ).这一结果表明施氮肥能减弱全球变暖对土壤有机碳分解的影响. ...
Atmospheric CO2 , soil nitrogen and turnover of fine roots.
1
1995
... 施氮肥对土壤根系呼吸的降低作用可能是因为氮肥降低了植物光合产物向地下根系的分配, 使得根系生物量和活性降低(Janssens et al ., 2010 ).此外, 光合产物向地下根系分配的减少还可能会对根际土壤产生负激发效应, 从而使土壤呼吸降低(Phillips & Fahey, 2007 ).另一方面, 有研究表明根系的寿命随土壤肥力的提高而降低, 而在低氮的环境中植物可以通过降低根系的周转来减少氮的损失(Pregitzer et al ., 1995 ).因此, 施氮肥后土壤氮素有效性提高, 导致根系死亡率增加和根系现存生物量降低.在挪威云杉(Picea abies )林中, Persson等(1998)发现施氮降低了挪威云杉的根细生物量, 抑制了植物根系活性.在该研究中, 我们也发现施氮肥后0-10 cm土层中杉木细根生物量下降了32.1% (表2 ).在我国东北的尖砬沟森林培育实验站, 梅莉等(2008)也发现了施氮肥后0-10 cm土层中水曲柳活细根生物量下降了13.4%-23.4%.因此, 根系生物量的下降是土壤呼吸降低的重要原因之一. ...
Controls on soil respiration: Implications for climate change.
2
2000
... 施氮肥降低了中亚热带地区杉木幼林土壤呼吸.这一结果与其他地区以前的部分研究结果是一致的(Persson et al ., 1998 ; Olsson et al ., 2005 ; 贾淑霞等, 2007 ; Mo et al ., 2008 ; Janssens et al ., 2010 ).在该研究中, 施氮肥使杉木幼林土壤总呼吸平均降低了22.7%, 这一降幅在已有研究结果的范围之内.例如, 在东北林业大学帽儿山实验林场尖砬沟森林培育实验站, 贾淑霞等(2007)发现外加氮2年后落叶松(Larix gmelinii )和水曲柳(Fraxinus mandshurica )人工林土壤呼吸速率分别降低了34.9%和25.8%.施肥对土壤总呼吸的降低作用主要来自于微生物呼吸(异养呼吸)和根系呼吸(自养呼吸)的降低.但是上述实验没有区分土壤呼吸的不同组分, 无法研究施肥对微生物呼吸和根系呼吸的影响.在该研究中, 我们采用切根法将土壤总呼吸区分为根系呼吸和微生物呼吸, 并发现施氮肥后土壤微生物呼吸和根系呼吸分别降低了23.5%和19.6%.虽然在森林生态系统中挖壕沟-切根法常用来区分和量化根系呼吸与微生物呼吸, 但也存在一些不足.第一, 切根处理后多长时间可以准确量化根系呼吸和土壤微生物呼吸难以确定, 不同树种之间也存在差异.在切根处理后, 切根处理样方内产生了死亡根系, 这些根系完全分解后才能准确量化根系呼吸与微生物呼吸.这是因为: 在死亡根系完全分解前测定土壤呼吸会高估土壤微生物呼吸, 从而使根系呼吸被低估(Tu et al ., 2013 ).在本研究中, 我们虽然在切根处理后14个月才开始测定土壤呼吸, 但也有可能切根处理样方内的死亡细根没有完全分解, 使得根系呼吸被低估.第二, 切根处理可能会影响土壤微环境, 如温度和含水量.土壤环境因子影响土壤呼吸强度(Rustad et al ., 2000 ), 因此也会影响根系呼吸和微生物呼吸测定结果的准确性.在本研究中, 切根处理对土壤温度和含水量基本没有影响. ...
... 土壤温度和含水量是影响土壤呼吸的重要非生物因素(Rustad et al ., 2000 ; Wang et al ., 2004 ).在杉木幼林中, 我们发现土壤呼吸与土壤温度的相关性大于土壤呼吸与土壤含水量的相关性.目前, 在亚热带其他森林中所进行的研究也发现了土壤呼吸与土壤温度存在很好的指数关系, 而与土壤含水量相关性较弱或不相关(Sheng et al ., 2010 ; 韩天丰等, 2011 ; 吴君君等, 2014 ; 范跃新等, 2014 ).除8月份外, 土壤呼吸的季节性变化(图2 )与土壤温度(图1 )高度一致, 这说明在大部分时间内土壤温度是土壤呼吸的主要控制因子, 而土壤含水量不是限制因子.指数模型和双因素模型的模拟结果也充分证实了这一观点.在8月份, 土壤含量低于10%, 限制了土壤呼吸, 导致该月份土壤呼吸降低, 成为土壤呼吸的主要控制因子.这也说明在水热充足的亚热带地区确实存在夏季干旱现象.目前, 已有一些学者研究了夏季干旱对土壤呼吸过程的影响(Welp et al ., 2007 ; Schindlbacher et al ., 2012 ).例如, 在温带云杉林中, Schindlbacher等(2012)认为夏季干旱降低了土壤呼吸, 并在一定程度上可以抵消升温对土壤呼吸的增加效应; Borken等(2006)发现长期的夏季干旱主要降低了土壤的微生物呼吸, 即抑制了土壤有机碳的分解. ...
Soil and microbial respiration in a loblolly pine plantation in response to seven years of irrigation and fertilization.
1
2009
... 土壤呼吸是陆地生态系统碳循环的重要环节, 是在土壤新陈代谢功能过程中, 土壤产生大量的CO2 , 并向大气中释放CO2 的过程(杨晶和李凌浩, 2003 ).据估计, 每年从土壤释放到大气中的碳高达98 Pg·a-1 (Bond-Lamberty & Thompson, 2010 ), 相当于化石燃料燃烧所释放碳的10倍多(Zhu & Cheng, 2011 ).因此, 土壤呼吸速率的微小变化都可能会对大气中CO2 浓度产生巨大影响.在森林生态系统中, 施氮肥对土壤呼吸的影响国内外已有大量研究报道, 但不同树种的森林生态系统土壤呼吸对施氮肥的响应之间存在差异, 主要表现为促进(Cleveland & Townsend, 2006 ; Tu et al ., 2013 )、抑制(Bowden et al ., 2004 ; Olsson et al ., 2005 ; 贾淑霞等, 2007 ; Mo et al ., 2008 ; Janssens et al ., 2010 )或无影响(Samuelson et al ., 2009 ).例如, 施氮肥使亚热带竹(Pleioblastus amarus )林生态系统的土壤呼吸增加26%-52% (Tu et al ., 2013), 使北方森林生态系统的土壤呼吸降低40% (Olsson et al ., 2005 ).施氮肥在不同森林和区域对土壤呼吸的影响存在差异, 主要与树种特性和土壤性质有关, 也与各实验所施用氮肥的类型和施用时间长短有关(Tu et al ., 2013). ...
Soil respiration under climate change: Prolonged summer drought offsets soil warming effects.
1
2012
... 土壤温度和含水量是影响土壤呼吸的重要非生物因素(Rustad et al ., 2000 ; Wang et al ., 2004 ).在杉木幼林中, 我们发现土壤呼吸与土壤温度的相关性大于土壤呼吸与土壤含水量的相关性.目前, 在亚热带其他森林中所进行的研究也发现了土壤呼吸与土壤温度存在很好的指数关系, 而与土壤含水量相关性较弱或不相关(Sheng et al ., 2010 ; 韩天丰等, 2011 ; 吴君君等, 2014 ; 范跃新等, 2014 ).除8月份外, 土壤呼吸的季节性变化(图2 )与土壤温度(图1 )高度一致, 这说明在大部分时间内土壤温度是土壤呼吸的主要控制因子, 而土壤含水量不是限制因子.指数模型和双因素模型的模拟结果也充分证实了这一观点.在8月份, 土壤含量低于10%, 限制了土壤呼吸, 导致该月份土壤呼吸降低, 成为土壤呼吸的主要控制因子.这也说明在水热充足的亚热带地区确实存在夏季干旱现象.目前, 已有一些学者研究了夏季干旱对土壤呼吸过程的影响(Welp et al ., 2007 ; Schindlbacher et al ., 2012 ).例如, 在温带云杉林中, Schindlbacher等(2012)认为夏季干旱降低了土壤呼吸, 并在一定程度上可以抵消升温对土壤呼吸的增加效应; Borken等(2006)发现长期的夏季干旱主要降低了土壤的微生物呼吸, 即抑制了土壤有机碳的分解. ...
The dynamic response of soil respiration to land-use changes in subtropical China.
1
2010
... 土壤温度和含水量是影响土壤呼吸的重要非生物因素(Rustad et al ., 2000 ; Wang et al ., 2004 ).在杉木幼林中, 我们发现土壤呼吸与土壤温度的相关性大于土壤呼吸与土壤含水量的相关性.目前, 在亚热带其他森林中所进行的研究也发现了土壤呼吸与土壤温度存在很好的指数关系, 而与土壤含水量相关性较弱或不相关(Sheng et al ., 2010 ; 韩天丰等, 2011 ; 吴君君等, 2014 ; 范跃新等, 2014 ).除8月份外, 土壤呼吸的季节性变化(图2 )与土壤温度(图1 )高度一致, 这说明在大部分时间内土壤温度是土壤呼吸的主要控制因子, 而土壤含水量不是限制因子.指数模型和双因素模型的模拟结果也充分证实了这一观点.在8月份, 土壤含量低于10%, 限制了土壤呼吸, 导致该月份土壤呼吸降低, 成为土壤呼吸的主要控制因子.这也说明在水热充足的亚热带地区确实存在夏季干旱现象.目前, 已有一些学者研究了夏季干旱对土壤呼吸过程的影响(Welp et al ., 2007 ; Schindlbacher et al ., 2012 ).例如, 在温带云杉林中, Schindlbacher等(2012)认为夏季干旱降低了土壤呼吸, 并在一定程度上可以抵消升温对土壤呼吸的增加效应; Borken等(2006)发现长期的夏季干旱主要降低了土壤的微生物呼吸, 即抑制了土壤有机碳的分解. ...
The effect of nitrogen addition on soil respiration from a nitrogen-limited forest soil.
2
2014
... 土壤呼吸的温度敏感性反映土壤呼吸速率对温度变化的敏感程度, 即温度变化10 ℃时土壤呼吸的相对变化, 常用Q 10 表示(Pregitzer et al ., 2000 ).在杉木幼林生态系统中, 土壤呼吸的Q 10 值在1.81- 2.04之间, 与王传华等(2011)得出的马尾松(Pinus massoniana )林和枫香树(Liquidambar formosana )林(0-5 cm土层)土壤呼吸的Q 10 值分别为1.86和2.10, 范跃新等(2014)得出的中亚热带地区不同演替阶段常绿阔叶林土壤呼吸的Q 10 值在1.79-2.64之间相符.但是, 这些亚热带森林的研究结果低于贾淑霞等(2007)在温带落叶松和水曲柳林中所得的土壤呼吸Q 10 值(2.45-3.29).在杉木幼林中, 施氮肥虽然对土壤总呼吸温度敏感性的影响不显著, 但土壤总呼吸温度敏感性有降低趋势.土壤呼吸与光合产物供应紧密相关, 施氮降低了光合产物向根系的分配是导致土壤总呼吸温度敏感性呈下降趋势的一个原因(Sun et al ., 2014 ).另一方面, 施氮增加了土壤有机碳的稳定性, 形成更多难分解的有机质(Janssens et al ., 2010 ), 这也会导致土壤总呼吸温度敏感性的降低.同样, 在我国东北的落叶松和水曲柳人工林中, 贾淑霞等(2007)也发现施氮肥后土壤呼吸的Q 10 值没有发生显著变化.他们认为施氮肥没有影响Q 10 值是因为根系组织中氮含量保持不变, 根系呼吸也没有发生变化.这是因为根细胞中维持蛋白质周转和溶质梯度所消耗的能量占根系维持呼吸的绝大部分, 如果根系组织中的氮含量保持不变, 根系呼吸也不会发生变化(Burton et al ., 2002 ).更重要的是, 我们发现施氮肥显著降低了土壤微生物呼吸的Q 10 值.其实, 造成土壤总呼吸温度敏感性降低的因素也可以导致土壤微生物呼吸温度敏感性的降低.另外, 施氮肥可以抑制土壤酶的活性, 对其产生副作用(Aber et al ., 1998 ).这一变化也可以导致土壤微生物呼吸温度敏感性的降低(Sun et al ., 2014 ).这一结果表明施氮肥能减弱全球变暖对土壤有机碳分解的影响. ...
... ).这一变化也可以导致土壤微生物呼吸温度敏感性的降低(Sun et al ., 2014 ).这一结果表明施氮肥能减弱全球变暖对土壤有机碳分解的影响. ...
Nitrogen additions and microbial biomass: A meta-analysis of ecosystem studies.
1
2008
... 微生物呼吸主要体现为微生物对土壤有机质的分解而释放CO2 的过程, 微生物群落结构的变化会影响土壤有机质的分解(Janssens et al ., 2010 ).施氮肥改变了土壤微环境, 使土壤微生物群落结构和活性发生了变化, 进而会影响微生物对土壤有机质的分解过程(Treseder, 2008 ; Frey et al ., 2014 ).我们的研究发现了施氮肥后土壤微生物量碳含量以及细菌的浓度显著降低, 细菌:真菌比降低(表3 ), 表明施氮肥改变了土壤微生物群落结构.因此, 施氮肥后杉木幼林土壤微生物活性的下降和群落结构的改变是土壤微生物呼吸降低的另一个重要因素.与我们的研究结果相似, 通过对82个氮添加对微生物量影响的野外研究结果的整合分析, Treseder (2008)发现施氮肥后土壤微生物量平均降低了15%, 并且微生物量对氮肥的响应与土壤CO2 释放的响应显著相关.我们还发现施氮肥降低了土壤真菌的生物量, 这与Frey等(2014)在温带森林生态系统中发现的施肥土壤中的真菌生物量和相对丰度低于不施肥土壤一致.一方面, 由于施氮肥降低了根系生物量(表2 ), 使根系输入到土壤中的碳减少, 抑制了土壤微生物生长, 导致土壤微生物呼吸降低.另一方面, 由于该施肥实验是在南方酸性土壤进行的, 较低的土壤pH值表示土壤可能存在铝毒.虽然该研究没有测定土壤中溶解态铝的含量, 但有研究表明土壤pH值越低, 土壤中溶解态铝含量越高, 这会降低植物对营养元素的吸收(俞元春和丁爱芳, 2001 ).施氮肥降低了土壤pH值(表2 ), 可能导致土壤中溶解态铝含量增加, 使杉木根系生物量和土壤微生物活性降低, 最终导致土壤微生物呼吸作用降低. ...
Nitrogen addition stimulates different components of soil respiration in a subtropical bamboo ecosystem.
4
2013
... 土壤呼吸是陆地生态系统碳循环的重要环节, 是在土壤新陈代谢功能过程中, 土壤产生大量的CO2 , 并向大气中释放CO2 的过程(杨晶和李凌浩, 2003 ).据估计, 每年从土壤释放到大气中的碳高达98 Pg·a-1 (Bond-Lamberty & Thompson, 2010 ), 相当于化石燃料燃烧所释放碳的10倍多(Zhu & Cheng, 2011 ).因此, 土壤呼吸速率的微小变化都可能会对大气中CO2 浓度产生巨大影响.在森林生态系统中, 施氮肥对土壤呼吸的影响国内外已有大量研究报道, 但不同树种的森林生态系统土壤呼吸对施氮肥的响应之间存在差异, 主要表现为促进(Cleveland & Townsend, 2006 ; Tu et al ., 2013 )、抑制(Bowden et al ., 2004 ; Olsson et al ., 2005 ; 贾淑霞等, 2007 ; Mo et al ., 2008 ; Janssens et al ., 2010 )或无影响(Samuelson et al ., 2009 ).例如, 施氮肥使亚热带竹(Pleioblastus amarus )林生态系统的土壤呼吸增加26%-52% (Tu et al ., 2013), 使北方森林生态系统的土壤呼吸降低40% (Olsson et al ., 2005 ).施氮肥在不同森林和区域对土壤呼吸的影响存在差异, 主要与树种特性和土壤性质有关, 也与各实验所施用氮肥的类型和施用时间长短有关(Tu et al ., 2013). ...
... 土壤呼吸可以分为自养呼吸和异养呼吸.在我们的研究中自养呼吸主要是指植物根系呼吸, 而异养呼吸主要是指土壤微生物呼吸.这些组分对施肥的响应至关重要, 关系到生态系统碳平衡(Tyree et al ., 2006 ).目前主要采用挖壕沟-切根法、树干环割等方法来区分植物根系呼吸和土壤微生物呼吸(Högberg et al ., 2001 ; Tu et al ., 2013 ).但是, 目前大部分实验只是研究了土壤总呼吸对施氮肥的响应, 而关于根系呼吸和微生物呼吸对施氮肥的响应还研究得比较少, 为数不多的研究所取得的结果还相互矛盾(Tyree et al ., 2006 ; Tu et al ., 2013 ).例如, Tu等(2013)在竹林中发现施氮肥促进了根系呼吸和微生物呼吸; 而Janssens等(2010)通过整合分析发现施氮肥后土壤微生物呼吸降低15%.在亚热带地区, 关于施肥对人工林土壤呼吸过程影响的研究, 尤其是对土壤呼吸不同组分的研究还极其有限, 这严重地限制了我们对施肥措施对土壤碳循环过程影响的深入理解, 不利于人工林的可持续经营管理. ...
... ; Tu et al ., 2013 ).例如, Tu等(2013)在竹林中发现施氮肥促进了根系呼吸和微生物呼吸; 而Janssens等(2010)通过整合分析发现施氮肥后土壤微生物呼吸降低15%.在亚热带地区, 关于施肥对人工林土壤呼吸过程影响的研究, 尤其是对土壤呼吸不同组分的研究还极其有限, 这严重地限制了我们对施肥措施对土壤碳循环过程影响的深入理解, 不利于人工林的可持续经营管理. ...
... 施氮肥降低了中亚热带地区杉木幼林土壤呼吸.这一结果与其他地区以前的部分研究结果是一致的(Persson et al ., 1998 ; Olsson et al ., 2005 ; 贾淑霞等, 2007 ; Mo et al ., 2008 ; Janssens et al ., 2010 ).在该研究中, 施氮肥使杉木幼林土壤总呼吸平均降低了22.7%, 这一降幅在已有研究结果的范围之内.例如, 在东北林业大学帽儿山实验林场尖砬沟森林培育实验站, 贾淑霞等(2007)发现外加氮2年后落叶松(Larix gmelinii )和水曲柳(Fraxinus mandshurica )人工林土壤呼吸速率分别降低了34.9%和25.8%.施肥对土壤总呼吸的降低作用主要来自于微生物呼吸(异养呼吸)和根系呼吸(自养呼吸)的降低.但是上述实验没有区分土壤呼吸的不同组分, 无法研究施肥对微生物呼吸和根系呼吸的影响.在该研究中, 我们采用切根法将土壤总呼吸区分为根系呼吸和微生物呼吸, 并发现施氮肥后土壤微生物呼吸和根系呼吸分别降低了23.5%和19.6%.虽然在森林生态系统中挖壕沟-切根法常用来区分和量化根系呼吸与微生物呼吸, 但也存在一些不足.第一, 切根处理后多长时间可以准确量化根系呼吸和土壤微生物呼吸难以确定, 不同树种之间也存在差异.在切根处理后, 切根处理样方内产生了死亡根系, 这些根系完全分解后才能准确量化根系呼吸与微生物呼吸.这是因为: 在死亡根系完全分解前测定土壤呼吸会高估土壤微生物呼吸, 从而使根系呼吸被低估(Tu et al ., 2013 ).在本研究中, 我们虽然在切根处理后14个月才开始测定土壤呼吸, 但也有可能切根处理样方内的死亡细根没有完全分解, 使得根系呼吸被低估.第二, 切根处理可能会影响土壤微环境, 如温度和含水量.土壤环境因子影响土壤呼吸强度(Rustad et al ., 2000 ), 因此也会影响根系呼吸和微生物呼吸测定结果的准确性.在本研究中, 切根处理对土壤温度和含水量基本没有影响. ...
Long-term effects of site preparation and fertilization on total soil CO2 efflux and heterotrophic respiration in a 33-year-old Pinus taeda L. plantation on the wet flats of the Virginia Lower Coastal Plain.
2
2006
... 土壤呼吸可以分为自养呼吸和异养呼吸.在我们的研究中自养呼吸主要是指植物根系呼吸, 而异养呼吸主要是指土壤微生物呼吸.这些组分对施肥的响应至关重要, 关系到生态系统碳平衡(Tyree et al ., 2006 ).目前主要采用挖壕沟-切根法、树干环割等方法来区分植物根系呼吸和土壤微生物呼吸(Högberg et al ., 2001 ; Tu et al ., 2013 ).但是, 目前大部分实验只是研究了土壤总呼吸对施氮肥的响应, 而关于根系呼吸和微生物呼吸对施氮肥的响应还研究得比较少, 为数不多的研究所取得的结果还相互矛盾(Tyree et al ., 2006 ; Tu et al ., 2013 ).例如, Tu等(2013)在竹林中发现施氮肥促进了根系呼吸和微生物呼吸; 而Janssens等(2010)通过整合分析发现施氮肥后土壤微生物呼吸降低15%.在亚热带地区, 关于施肥对人工林土壤呼吸过程影响的研究, 尤其是对土壤呼吸不同组分的研究还极其有限, 这严重地限制了我们对施肥措施对土壤碳循环过程影响的深入理解, 不利于人工林的可持续经营管理. ...
... ).但是, 目前大部分实验只是研究了土壤总呼吸对施氮肥的响应, 而关于根系呼吸和微生物呼吸对施氮肥的响应还研究得比较少, 为数不多的研究所取得的结果还相互矛盾(Tyree et al ., 2006 ; Tu et al ., 2013 ).例如, Tu等(2013)在竹林中发现施氮肥促进了根系呼吸和微生物呼吸; 而Janssens等(2010)通过整合分析发现施氮肥后土壤微生物呼吸降低15%.在亚热带地区, 关于施肥对人工林土壤呼吸过程影响的研究, 尤其是对土壤呼吸不同组分的研究还极其有限, 这严重地限制了我们对施肥措施对土壤碳循环过程影响的深入理解, 不利于人工林的可持续经营管理. ...
鄂东南低丘马尾松林和枫香林土壤异养呼吸及温湿度敏感性. 应用生态学报,
0
2011
鄂东南低丘马尾松林和枫香林土壤异养呼吸及温湿度敏感性. 应用生态学报,
0
2011
Effects of soil temperature and soil water content on soil respiration in three forest types in Changbai Mountain.
1
2004
... 土壤温度和含水量是影响土壤呼吸的重要非生物因素(Rustad et al ., 2000 ; Wang et al ., 2004 ).在杉木幼林中, 我们发现土壤呼吸与土壤温度的相关性大于土壤呼吸与土壤含水量的相关性.目前, 在亚热带其他森林中所进行的研究也发现了土壤呼吸与土壤温度存在很好的指数关系, 而与土壤含水量相关性较弱或不相关(Sheng et al ., 2010 ; 韩天丰等, 2011 ; 吴君君等, 2014 ; 范跃新等, 2014 ).除8月份外, 土壤呼吸的季节性变化(图2 )与土壤温度(图1 )高度一致, 这说明在大部分时间内土壤温度是土壤呼吸的主要控制因子, 而土壤含水量不是限制因子.指数模型和双因素模型的模拟结果也充分证实了这一观点.在8月份, 土壤含量低于10%, 限制了土壤呼吸, 导致该月份土壤呼吸降低, 成为土壤呼吸的主要控制因子.这也说明在水热充足的亚热带地区确实存在夏季干旱现象.目前, 已有一些学者研究了夏季干旱对土壤呼吸过程的影响(Welp et al ., 2007 ; Schindlbacher et al ., 2012 ).例如, 在温带云杉林中, Schindlbacher等(2012)认为夏季干旱降低了土壤呼吸, 并在一定程度上可以抵消升温对土壤呼吸的增加效应; Borken等(2006)发现长期的夏季干旱主要降低了土壤的微生物呼吸, 即抑制了土壤有机碳的分解. ...
Carbon input manipulation affects soil respiration and microbial community composition in a subtropical coniferous forest.
0
2013
The sensitivity of carbon fluxes to spring warming and summer drought depends on plant functional type in boreal forest ecosystems.
1
2007
... 土壤温度和含水量是影响土壤呼吸的重要非生物因素(Rustad et al ., 2000 ; Wang et al ., 2004 ).在杉木幼林中, 我们发现土壤呼吸与土壤温度的相关性大于土壤呼吸与土壤含水量的相关性.目前, 在亚热带其他森林中所进行的研究也发现了土壤呼吸与土壤温度存在很好的指数关系, 而与土壤含水量相关性较弱或不相关(Sheng et al ., 2010 ; 韩天丰等, 2011 ; 吴君君等, 2014 ; 范跃新等, 2014 ).除8月份外, 土壤呼吸的季节性变化(图2 )与土壤温度(图1 )高度一致, 这说明在大部分时间内土壤温度是土壤呼吸的主要控制因子, 而土壤含水量不是限制因子.指数模型和双因素模型的模拟结果也充分证实了这一观点.在8月份, 土壤含量低于10%, 限制了土壤呼吸, 导致该月份土壤呼吸降低, 成为土壤呼吸的主要控制因子.这也说明在水热充足的亚热带地区确实存在夏季干旱现象.目前, 已有一些学者研究了夏季干旱对土壤呼吸过程的影响(Welp et al ., 2007 ; Schindlbacher et al ., 2012 ).例如, 在温带云杉林中, Schindlbacher等(2012)认为夏季干旱降低了土壤呼吸, 并在一定程度上可以抵消升温对土壤呼吸的增加效应; Borken等(2006)发现长期的夏季干旱主要降低了土壤的微生物呼吸, 即抑制了土壤有机碳的分解. ...
米槠和杉木人工林土壤呼吸及其组分分析. 植物生态学报,
1
2014
... 土壤温度和含水量是影响土壤呼吸的重要非生物因素(Rustad et al ., 2000 ; Wang et al ., 2004 ).在杉木幼林中, 我们发现土壤呼吸与土壤温度的相关性大于土壤呼吸与土壤含水量的相关性.目前, 在亚热带其他森林中所进行的研究也发现了土壤呼吸与土壤温度存在很好的指数关系, 而与土壤含水量相关性较弱或不相关(Sheng et al ., 2010 ; 韩天丰等, 2011 ; 吴君君等, 2014 ; 范跃新等, 2014 ).除8月份外, 土壤呼吸的季节性变化(图2 )与土壤温度(图1 )高度一致, 这说明在大部分时间内土壤温度是土壤呼吸的主要控制因子, 而土壤含水量不是限制因子.指数模型和双因素模型的模拟结果也充分证实了这一观点.在8月份, 土壤含量低于10%, 限制了土壤呼吸, 导致该月份土壤呼吸降低, 成为土壤呼吸的主要控制因子.这也说明在水热充足的亚热带地区确实存在夏季干旱现象.目前, 已有一些学者研究了夏季干旱对土壤呼吸过程的影响(Welp et al ., 2007 ; Schindlbacher et al ., 2012 ).例如, 在温带云杉林中, Schindlbacher等(2012)认为夏季干旱降低了土壤呼吸, 并在一定程度上可以抵消升温对土壤呼吸的增加效应; Borken等(2006)发现长期的夏季干旱主要降低了土壤的微生物呼吸, 即抑制了土壤有机碳的分解. ...
米槠和杉木人工林土壤呼吸及其组分分析. 植物生态学报,
1
2014
... 土壤温度和含水量是影响土壤呼吸的重要非生物因素(Rustad et al ., 2000 ; Wang et al ., 2004 ).在杉木幼林中, 我们发现土壤呼吸与土壤温度的相关性大于土壤呼吸与土壤含水量的相关性.目前, 在亚热带其他森林中所进行的研究也发现了土壤呼吸与土壤温度存在很好的指数关系, 而与土壤含水量相关性较弱或不相关(Sheng et al ., 2010 ; 韩天丰等, 2011 ; 吴君君等, 2014 ; 范跃新等, 2014 ).除8月份外, 土壤呼吸的季节性变化(图2 )与土壤温度(图1 )高度一致, 这说明在大部分时间内土壤温度是土壤呼吸的主要控制因子, 而土壤含水量不是限制因子.指数模型和双因素模型的模拟结果也充分证实了这一观点.在8月份, 土壤含量低于10%, 限制了土壤呼吸, 导致该月份土壤呼吸降低, 成为土壤呼吸的主要控制因子.这也说明在水热充足的亚热带地区确实存在夏季干旱现象.目前, 已有一些学者研究了夏季干旱对土壤呼吸过程的影响(Welp et al ., 2007 ; Schindlbacher et al ., 2012 ).例如, 在温带云杉林中, Schindlbacher等(2012)认为夏季干旱降低了土壤呼吸, 并在一定程度上可以抵消升温对土壤呼吸的增加效应; Borken等(2006)发现长期的夏季干旱主要降低了土壤的微生物呼吸, 即抑制了土壤有机碳的分解. ...
我国第八次森林资源清查结果及分析. 林业经济,
3
2014
... 第八次全国森林资源清查结果显示, 我国人工林面积已达6900多万hm2 , 约占全球人工林面积的1/4, 在我国木材生产和生态安全保障中起着至关重要的作用(徐济德, 2014 ).在我国实行天然林保护的背景下, 人工林, 尤其是用材林在提供林产品方面的作用愈加重要, 并且其面积还在持续增加(徐济德, 2014 ).但是, 我国人工林管理粗放、经营水平低, 导致其生产力不高, 平均蓄积量仅为52.8 m3 ·hm-2 , 不到发达国家的1/3 (徐济德, 2014 ).要提高人工林生产和生态服务功能, 亟需改变过去传统的粗放式管理模式, 实行高度集约式经营.在大部分区域, 土壤氮素缺乏是森林生态系统生产力最重要的限制因子(Deluca et al ., 2002 ; Zhou et al ., 2003 ; 于明坚等, 2005 ).为解决氮素的限制, 施氮肥已经成为提高人工林生产力的一种高度集约式经营管理措施(Fox, 2000 ; Lee & Jose, 2003 ).但是, 施肥在提高林地生产力的同时, 还会对土壤呼吸等地下生态过程产生深刻影响. ...
... ).在我国实行天然林保护的背景下, 人工林, 尤其是用材林在提供林产品方面的作用愈加重要, 并且其面积还在持续增加(徐济德, 2014 ).但是, 我国人工林管理粗放、经营水平低, 导致其生产力不高, 平均蓄积量仅为52.8 m3 ·hm-2 , 不到发达国家的1/3 (徐济德, 2014 ).要提高人工林生产和生态服务功能, 亟需改变过去传统的粗放式管理模式, 实行高度集约式经营.在大部分区域, 土壤氮素缺乏是森林生态系统生产力最重要的限制因子(Deluca et al ., 2002 ; Zhou et al ., 2003 ; 于明坚等, 2005 ).为解决氮素的限制, 施氮肥已经成为提高人工林生产力的一种高度集约式经营管理措施(Fox, 2000 ; Lee & Jose, 2003 ).但是, 施肥在提高林地生产力的同时, 还会对土壤呼吸等地下生态过程产生深刻影响. ...
... , 不到发达国家的1/3 (徐济德, 2014 ).要提高人工林生产和生态服务功能, 亟需改变过去传统的粗放式管理模式, 实行高度集约式经营.在大部分区域, 土壤氮素缺乏是森林生态系统生产力最重要的限制因子(Deluca et al ., 2002 ; Zhou et al ., 2003 ; 于明坚等, 2005 ).为解决氮素的限制, 施氮肥已经成为提高人工林生产力的一种高度集约式经营管理措施(Fox, 2000 ; Lee & Jose, 2003 ).但是, 施肥在提高林地生产力的同时, 还会对土壤呼吸等地下生态过程产生深刻影响. ...
我国第八次森林资源清查结果及分析. 林业经济,
3
2014
... 第八次全国森林资源清查结果显示, 我国人工林面积已达6900多万hm2 , 约占全球人工林面积的1/4, 在我国木材生产和生态安全保障中起着至关重要的作用(徐济德, 2014 ).在我国实行天然林保护的背景下, 人工林, 尤其是用材林在提供林产品方面的作用愈加重要, 并且其面积还在持续增加(徐济德, 2014 ).但是, 我国人工林管理粗放、经营水平低, 导致其生产力不高, 平均蓄积量仅为52.8 m3 ·hm-2 , 不到发达国家的1/3 (徐济德, 2014 ).要提高人工林生产和生态服务功能, 亟需改变过去传统的粗放式管理模式, 实行高度集约式经营.在大部分区域, 土壤氮素缺乏是森林生态系统生产力最重要的限制因子(Deluca et al ., 2002 ; Zhou et al ., 2003 ; 于明坚等, 2005 ).为解决氮素的限制, 施氮肥已经成为提高人工林生产力的一种高度集约式经营管理措施(Fox, 2000 ; Lee & Jose, 2003 ).但是, 施肥在提高林地生产力的同时, 还会对土壤呼吸等地下生态过程产生深刻影响. ...
... ).在我国实行天然林保护的背景下, 人工林, 尤其是用材林在提供林产品方面的作用愈加重要, 并且其面积还在持续增加(徐济德, 2014 ).但是, 我国人工林管理粗放、经营水平低, 导致其生产力不高, 平均蓄积量仅为52.8 m3 ·hm-2 , 不到发达国家的1/3 (徐济德, 2014 ).要提高人工林生产和生态服务功能, 亟需改变过去传统的粗放式管理模式, 实行高度集约式经营.在大部分区域, 土壤氮素缺乏是森林生态系统生产力最重要的限制因子(Deluca et al ., 2002 ; Zhou et al ., 2003 ; 于明坚等, 2005 ).为解决氮素的限制, 施氮肥已经成为提高人工林生产力的一种高度集约式经营管理措施(Fox, 2000 ; Lee & Jose, 2003 ).但是, 施肥在提高林地生产力的同时, 还会对土壤呼吸等地下生态过程产生深刻影响. ...
... , 不到发达国家的1/3 (徐济德, 2014 ).要提高人工林生产和生态服务功能, 亟需改变过去传统的粗放式管理模式, 实行高度集约式经营.在大部分区域, 土壤氮素缺乏是森林生态系统生产力最重要的限制因子(Deluca et al ., 2002 ; Zhou et al ., 2003 ; 于明坚等, 2005 ).为解决氮素的限制, 施氮肥已经成为提高人工林生产力的一种高度集约式经营管理措施(Fox, 2000 ; Lee & Jose, 2003 ).但是, 施肥在提高林地生产力的同时, 还会对土壤呼吸等地下生态过程产生深刻影响. ...
土壤呼吸及其测定法. 植物杂志,
1
2003
... 土壤呼吸是陆地生态系统碳循环的重要环节, 是在土壤新陈代谢功能过程中, 土壤产生大量的CO2 , 并向大气中释放CO2 的过程(杨晶和李凌浩, 2003 ).据估计, 每年从土壤释放到大气中的碳高达98 Pg·a-1 (Bond-Lamberty & Thompson, 2010 ), 相当于化石燃料燃烧所释放碳的10倍多(Zhu & Cheng, 2011 ).因此, 土壤呼吸速率的微小变化都可能会对大气中CO2 浓度产生巨大影响.在森林生态系统中, 施氮肥对土壤呼吸的影响国内外已有大量研究报道, 但不同树种的森林生态系统土壤呼吸对施氮肥的响应之间存在差异, 主要表现为促进(Cleveland & Townsend, 2006 ; Tu et al ., 2013 )、抑制(Bowden et al ., 2004 ; Olsson et al ., 2005 ; 贾淑霞等, 2007 ; Mo et al ., 2008 ; Janssens et al ., 2010 )或无影响(Samuelson et al ., 2009 ).例如, 施氮肥使亚热带竹(Pleioblastus amarus )林生态系统的土壤呼吸增加26%-52% (Tu et al ., 2013), 使北方森林生态系统的土壤呼吸降低40% (Olsson et al ., 2005 ).施氮肥在不同森林和区域对土壤呼吸的影响存在差异, 主要与树种特性和土壤性质有关, 也与各实验所施用氮肥的类型和施用时间长短有关(Tu et al ., 2013). ...
土壤呼吸及其测定法. 植物杂志,
1
2003
... 土壤呼吸是陆地生态系统碳循环的重要环节, 是在土壤新陈代谢功能过程中, 土壤产生大量的CO2 , 并向大气中释放CO2 的过程(杨晶和李凌浩, 2003 ).据估计, 每年从土壤释放到大气中的碳高达98 Pg·a-1 (Bond-Lamberty & Thompson, 2010 ), 相当于化石燃料燃烧所释放碳的10倍多(Zhu & Cheng, 2011 ).因此, 土壤呼吸速率的微小变化都可能会对大气中CO2 浓度产生巨大影响.在森林生态系统中, 施氮肥对土壤呼吸的影响国内外已有大量研究报道, 但不同树种的森林生态系统土壤呼吸对施氮肥的响应之间存在差异, 主要表现为促进(Cleveland & Townsend, 2006 ; Tu et al ., 2013 )、抑制(Bowden et al ., 2004 ; Olsson et al ., 2005 ; 贾淑霞等, 2007 ; Mo et al ., 2008 ; Janssens et al ., 2010 )或无影响(Samuelson et al ., 2009 ).例如, 施氮肥使亚热带竹(Pleioblastus amarus )林生态系统的土壤呼吸增加26%-52% (Tu et al ., 2013), 使北方森林生态系统的土壤呼吸降低40% (Olsson et al ., 2005 ).施氮肥在不同森林和区域对土壤呼吸的影响存在差异, 主要与树种特性和土壤性质有关, 也与各实验所施用氮肥的类型和施用时间长短有关(Tu et al ., 2013). ...
青冈常绿阔叶林氮的生物循环. 生态学报,
1
2005
... 第八次全国森林资源清查结果显示, 我国人工林面积已达6900多万hm2 , 约占全球人工林面积的1/4, 在我国木材生产和生态安全保障中起着至关重要的作用(徐济德, 2014 ).在我国实行天然林保护的背景下, 人工林, 尤其是用材林在提供林产品方面的作用愈加重要, 并且其面积还在持续增加(徐济德, 2014 ).但是, 我国人工林管理粗放、经营水平低, 导致其生产力不高, 平均蓄积量仅为52.8 m3 ·hm-2 , 不到发达国家的1/3 (徐济德, 2014 ).要提高人工林生产和生态服务功能, 亟需改变过去传统的粗放式管理模式, 实行高度集约式经营.在大部分区域, 土壤氮素缺乏是森林生态系统生产力最重要的限制因子(Deluca et al ., 2002 ; Zhou et al ., 2003 ; 于明坚等, 2005 ).为解决氮素的限制, 施氮肥已经成为提高人工林生产力的一种高度集约式经营管理措施(Fox, 2000 ; Lee & Jose, 2003 ).但是, 施肥在提高林地生产力的同时, 还会对土壤呼吸等地下生态过程产生深刻影响. ...
青冈常绿阔叶林氮的生物循环. 生态学报,
1
2005
... 第八次全国森林资源清查结果显示, 我国人工林面积已达6900多万hm2 , 约占全球人工林面积的1/4, 在我国木材生产和生态安全保障中起着至关重要的作用(徐济德, 2014 ).在我国实行天然林保护的背景下, 人工林, 尤其是用材林在提供林产品方面的作用愈加重要, 并且其面积还在持续增加(徐济德, 2014 ).但是, 我国人工林管理粗放、经营水平低, 导致其生产力不高, 平均蓄积量仅为52.8 m3 ·hm-2 , 不到发达国家的1/3 (徐济德, 2014 ).要提高人工林生产和生态服务功能, 亟需改变过去传统的粗放式管理模式, 实行高度集约式经营.在大部分区域, 土壤氮素缺乏是森林生态系统生产力最重要的限制因子(Deluca et al ., 2002 ; Zhou et al ., 2003 ; 于明坚等, 2005 ).为解决氮素的限制, 施氮肥已经成为提高人工林生产力的一种高度集约式经营管理措施(Fox, 2000 ; Lee & Jose, 2003 ).但是, 施肥在提高林地生产力的同时, 还会对土壤呼吸等地下生态过程产生深刻影响. ...
模拟酸雨对酸性土壤铝溶出及其形态转化的影响. 土壤与环境,
1
2001
... 微生物呼吸主要体现为微生物对土壤有机质的分解而释放CO2 的过程, 微生物群落结构的变化会影响土壤有机质的分解(Janssens et al ., 2010 ).施氮肥改变了土壤微环境, 使土壤微生物群落结构和活性发生了变化, 进而会影响微生物对土壤有机质的分解过程(Treseder, 2008 ; Frey et al ., 2014 ).我们的研究发现了施氮肥后土壤微生物量碳含量以及细菌的浓度显著降低, 细菌:真菌比降低(表3 ), 表明施氮肥改变了土壤微生物群落结构.因此, 施氮肥后杉木幼林土壤微生物活性的下降和群落结构的改变是土壤微生物呼吸降低的另一个重要因素.与我们的研究结果相似, 通过对82个氮添加对微生物量影响的野外研究结果的整合分析, Treseder (2008)发现施氮肥后土壤微生物量平均降低了15%, 并且微生物量对氮肥的响应与土壤CO2 释放的响应显著相关.我们还发现施氮肥降低了土壤真菌的生物量, 这与Frey等(2014)在温带森林生态系统中发现的施肥土壤中的真菌生物量和相对丰度低于不施肥土壤一致.一方面, 由于施氮肥降低了根系生物量(表2 ), 使根系输入到土壤中的碳减少, 抑制了土壤微生物生长, 导致土壤微生物呼吸降低.另一方面, 由于该施肥实验是在南方酸性土壤进行的, 较低的土壤pH值表示土壤可能存在铝毒.虽然该研究没有测定土壤中溶解态铝的含量, 但有研究表明土壤pH值越低, 土壤中溶解态铝含量越高, 这会降低植物对营养元素的吸收(俞元春和丁爱芳, 2001 ).施氮肥降低了土壤pH值(表2 ), 可能导致土壤中溶解态铝含量增加, 使杉木根系生物量和土壤微生物活性降低, 最终导致土壤微生物呼吸作用降低. ...
模拟酸雨对酸性土壤铝溶出及其形态转化的影响. 土壤与环境,
1
2001
... 微生物呼吸主要体现为微生物对土壤有机质的分解而释放CO2 的过程, 微生物群落结构的变化会影响土壤有机质的分解(Janssens et al ., 2010 ).施氮肥改变了土壤微环境, 使土壤微生物群落结构和活性发生了变化, 进而会影响微生物对土壤有机质的分解过程(Treseder, 2008 ; Frey et al ., 2014 ).我们的研究发现了施氮肥后土壤微生物量碳含量以及细菌的浓度显著降低, 细菌:真菌比降低(表3 ), 表明施氮肥改变了土壤微生物群落结构.因此, 施氮肥后杉木幼林土壤微生物活性的下降和群落结构的改变是土壤微生物呼吸降低的另一个重要因素.与我们的研究结果相似, 通过对82个氮添加对微生物量影响的野外研究结果的整合分析, Treseder (2008)发现施氮肥后土壤微生物量平均降低了15%, 并且微生物量对氮肥的响应与土壤CO2 释放的响应显著相关.我们还发现施氮肥降低了土壤真菌的生物量, 这与Frey等(2014)在温带森林生态系统中发现的施肥土壤中的真菌生物量和相对丰度低于不施肥土壤一致.一方面, 由于施氮肥降低了根系生物量(表2 ), 使根系输入到土壤中的碳减少, 抑制了土壤微生物生长, 导致土壤微生物呼吸降低.另一方面, 由于该施肥实验是在南方酸性土壤进行的, 较低的土壤pH值表示土壤可能存在铝毒.虽然该研究没有测定土壤中溶解态铝的含量, 但有研究表明土壤pH值越低, 土壤中溶解态铝含量越高, 这会降低植物对营养元素的吸收(俞元春和丁爱芳, 2001 ).施氮肥降低了土壤pH值(表2 ), 可能导致土壤中溶解态铝含量增加, 使杉木根系生物量和土壤微生物活性降低, 最终导致土壤微生物呼吸作用降低. ...
Net soil nitrogen mineralization in Chinese forest ecosystems.
1
2003
... 第八次全国森林资源清查结果显示, 我国人工林面积已达6900多万hm2 , 约占全球人工林面积的1/4, 在我国木材生产和生态安全保障中起着至关重要的作用(徐济德, 2014 ).在我国实行天然林保护的背景下, 人工林, 尤其是用材林在提供林产品方面的作用愈加重要, 并且其面积还在持续增加(徐济德, 2014 ).但是, 我国人工林管理粗放、经营水平低, 导致其生产力不高, 平均蓄积量仅为52.8 m3 ·hm-2 , 不到发达国家的1/3 (徐济德, 2014 ).要提高人工林生产和生态服务功能, 亟需改变过去传统的粗放式管理模式, 实行高度集约式经营.在大部分区域, 土壤氮素缺乏是森林生态系统生产力最重要的限制因子(Deluca et al ., 2002 ; Zhou et al ., 2003 ; 于明坚等, 2005 ).为解决氮素的限制, 施氮肥已经成为提高人工林生产力的一种高度集约式经营管理措施(Fox, 2000 ; Lee & Jose, 2003 ).但是, 施肥在提高林地生产力的同时, 还会对土壤呼吸等地下生态过程产生深刻影响. ...
Rhizosphere priming effect increases the temperature sensitivity of soil organic matter decomposition.
1
2011
... 土壤呼吸是陆地生态系统碳循环的重要环节, 是在土壤新陈代谢功能过程中, 土壤产生大量的CO2 , 并向大气中释放CO2 的过程(杨晶和李凌浩, 2003 ).据估计, 每年从土壤释放到大气中的碳高达98 Pg·a-1 (Bond-Lamberty & Thompson, 2010 ), 相当于化石燃料燃烧所释放碳的10倍多(Zhu & Cheng, 2011 ).因此, 土壤呼吸速率的微小变化都可能会对大气中CO2 浓度产生巨大影响.在森林生态系统中, 施氮肥对土壤呼吸的影响国内外已有大量研究报道, 但不同树种的森林生态系统土壤呼吸对施氮肥的响应之间存在差异, 主要表现为促进(Cleveland & Townsend, 2006 ; Tu et al ., 2013 )、抑制(Bowden et al ., 2004 ; Olsson et al ., 2005 ; 贾淑霞等, 2007 ; Mo et al ., 2008 ; Janssens et al ., 2010 )或无影响(Samuelson et al ., 2009 ).例如, 施氮肥使亚热带竹(Pleioblastus amarus )林生态系统的土壤呼吸增加26%-52% (Tu et al ., 2013), 使北方森林生态系统的土壤呼吸降低40% (Olsson et al ., 2005 ).施氮肥在不同森林和区域对土壤呼吸的影响存在差异, 主要与树种特性和土壤性质有关, 也与各实验所施用氮肥的类型和施用时间长短有关(Tu et al ., 2013). ...


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}