孟德云 , 万书波
, 万书波
MENG De-Yun, WAN Shu-Bo
通讯作者:
版权声明: 2015 植物生态学报编辑部 本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。
基金资助:
展开
摘要
为探讨外源多胺对花生(Arachis hypogaea)抗盐性的影响, 以盆栽花生‘花育22’为试验材料, 通过叶面喷施1 mmol·L-1腐胺(Put)、1 mmol·L-1亚精胺(Spd)、1 mmol·L-1精胺(Spm)的方法, 研究多胺对150 mmol·L-1 NaCl胁迫下盆栽花生的缓解作用。结果表明, 与对照(CK)相比, 盐胁迫显著抑制了花生植株的生长与荚果产量, 降低了叶绿素含量和抗氧化酶活性, 丙二醛(MDA)含量、叶片相对电导率增加; 在盐胁迫下, 叶面喷施Put、Spd、Spm处理均可有效促进花生植株的生长, 提高了超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)的活性, 增强植株抗氧化能力, 显著降低了花生叶片相对电导率和MDA的积累量, 从而缓解盐胁迫对质膜的过氧化伤害; 提高了叶绿素含量, 促进了植株高度的生长与分支数增多, 增加了干物质积累量, 从而提高了花生荚果产量; 其中, Spm处理引起的变化幅度大于Spd和Put处理。研究结果说明, 多胺有利于花生幼苗在盐胁迫下活性氧代谢和光合色素含量的提高, 促进花生植株的生长, 降低盐胁迫对花生植株的抑制作用, 且Spm处理的效果最好。
关键词:
Abstract
Aims Soil salinity is a major limiting factor for plant establishment, development and productivity. In recent years, the contradiction between oil crops and food crops for land is increasingly prominent. In order not to take up the land for food, peanut planting on saline-alkali land could be a promising option. However, peanuts have been rarely grown in saline-alkali land, which may be due to the reduction of peanut yield caused by salt stress. Therefore, research of peanut salt resistance has important practical significance.Methods In order to investigate the effects of exogenous polyamines on peanut (Arachis hypogaea) grown in pots under salt stress, ‘Huayu 22’, one of the peanut cultivars, was used as materials by being foliar-sprayed with 1 mmol·L-1 putrescine (Put), 1 mmol·L-1 spermidine (Spd) and 1 mmol·L-1 spermine (Spm) to elucidate the role of exogenous polyamines on peanuts under 150 mmol·L-1NaCl. Important findingsResults showed that growth, yield, chlorophyll contents and antioxidant enzyme activities of peanut seedling decreased, however, malondialdehyde (MDA) content and relative electrolytic leakage increased under salt stress. Meanwhile, exogenous polyamines significantly improved the activities of superoxide dismutase (SOD), peroxidase (POD) and catalase (CAT), and reduced the relative electrolytic leakage and MDA content in peanut leaves under salt stress and thus alleviating the oxidative damage of salt stress on plasma membrane. It is obvious that exogenous polyamines could improve chlorophyll contents, plant height, number of branch and the amount of dry matter accumulation, even pod yield under salt stress. Among these three polyamines, the effects of exogenous Spm on alleviating salt stress were most effective These results showed that exogenous polyamines, especially Spm, were favorable for the seedlings to increase reactive oxygen metabolism and photosynthesis, which improved peanut growth and reduced the inhibitory effects of salt stress on peanuts.
Keywords:
土壤盐渍化是影响农业生产和生态环境的严重问题, 据统计, 全球有20%的耕地受到不同程度的盐渍化(李建国等, 2012), 严重影响作物生长, 降低作物产量。花生(Arachis hypogaea)是我国重要的油料经济作物, 具有重要的营养价值和经济价值。近年来油料作物与粮食作物争地的矛盾日益突出, 为了不与粮争地, 将花生种植在盐碱地上是不错的选择(万书波, 2009)。但是在中国, 花生很少种植在盐碱地, 这可能与盐胁迫影响花生产量有关。因此, 提高花生耐盐性研究具有重要的现实意义。
多胺是一类广泛存在于动植物体内具有强烈的生物活性的多聚阳离子、低分子量的脂肪族含氮碱。植物细胞中的多胺主要包括腐胺(Put)、亚精胺(Spd)和精胺(Spm)。研究发现多胺与植物体的生长、发育以及对环境胁迫的响应等多种生理过程相关, 并且参与植物体中的信号转导(Galston & Sauheny, 1990)。研究发现多胺可以通过多种机制响应非生物胁迫, 如影响清除活性氧的酶系统(徐仰仓等, 2001)、非酶系统(Maccarrone et al., 1998)以及作为第二信使参与植物的逆境胁迫信号传递(Liu et al., 2000; Sudha & Ravishankar, 2003)等。
研究表明, 盐胁迫下植物体内源多胺含量发生剧烈变化, 且因植物种类不同、组织部位、胁迫时间的不同而表现出差异(Liu et al., 2007)。外施多胺可提高植物耐盐性, 但不同植物种类及不同种类多胺的施用效应差异较大。例如, 外源Put和Spd对减轻盐敏感型水稻(Oryza sativa)所受的胁迫伤害有一定的作用, 而施用外源Spm却没有任何明显效果(Ndayiragije & Lutts, 2007)。施用外源Spm能够通过调节叶绿体抗氧化系统显著缓解黄瓜(Cucumis sativus)幼苗受到的盐胁迫损伤(Shu et al., 2013)。在花生中, 关于多胺的研究甚少, 只有极少数关于多胺氧化酶及衰老的研究, 缺乏对多胺在花生抗逆性方面的研究。本文以‘花育22’为试验材料, 重点研究盐胁迫下, 喷施外源多胺Put、Spd、Spm对盆栽花生植株生长及产量的影响, 以期为盐碱地花生种植提供理论支持。
试验于山东省农业科学院饮马泉试验农场进行, 供试材料为花生‘花育22’ (种子由山东省农业科学院提供)。土壤为该农场0-20 cm土层土壤, 所用栽培盆为内径30 cm、高40 cm的瓷花盆。试验于5月16日播种, 每盆三穴, 每穴两粒, 出苗后每穴保留1株健壮苗。于6月3日与7月3日两次浇150 mmol·L-1 NaCl溶液, 每盆用2 L浇透(土壤最终盐浓度为0.2%), 以浇纯净水处理为对照(CK)。于始花期、下针期、结荚期分别叶面喷施Put、Spd、Spm溶液, 浓度均为1 mmol·L-1, 每盆喷施30 mL (刘俊等, 2006; Roychoudhury et al., 2011; 王晓云等, 1999), CK与单纯盐处理叶面喷施等量纯净水, 共3次喷施处理, 均在无风傍晚进行。处理7天后取样测定相关指标, 每个处理5个重复。
分别于7月14日(始花期, 记为FE)、7月29日(下针期, 记为FP)、8月20日(结荚期, 记为PP)、9月26日(收获期, 记为PF)取样测定相关指标。每次选取5株完整植株, 根部深入土壤30 cm进行挖掘。将总叶片、枝茎、根系和荚果样品分开置于105 ℃的烘箱内杀青30 min, 85 ℃烘干至恒质量后称量; 收获期分别进行植株高度、侧枝数、第一对侧枝平均长度的统计与测定; 各时期取功能叶倒三叶鲜样测定叶绿素含量、相对电导率, 液氮速冻倒三叶用于测定活性氧清除酶活性和丙二醛(MDA)含量。
乙醇浸提法测定叶绿素含量(李合生, 2000); 相对电导率用DDS-11A型电导率仪(上海大普仪器有限公司, 上海)测定; MDA含量采用硫代巴比妥酸(TBA)法测定(赵世杰等, 1994); 超氧化物歧化酶(SOD)活性采用氮蓝四唑法测定(Giannopolitis & Ries, 1977); 过氧化物酶(POD)活性采用愈创木酚法测定(曾韶西等, 1997); 过氧化氢酶(CAT)活性采用过氧化氢紫外线法测定(张治安和陈展宇, 2008)。每个指标重复测定5次, 取平均值。
采用SigmaPlot 10.0进行数据处理和绘图, SPSS 16.0进行显著性分析。
由表1可以看出, 盐胁迫显著抑制了花生的主茎高、侧枝长和分枝数, 与CK相比, 主茎高度降低了15.42%, 侧枝长度下降了16.07%, 分枝数减少了32.52%。外施多胺可明显改善花生营养生长, 部分缓解盐胁迫造成的抑制作用, 花生主茎高、侧枝长、分枝数均较单一盐胁迫显著增加。其中Spm的施用对花生主茎高和侧枝长影响最显著, 较单一盐胁迫分别增加了11.96%和16.67%; Put和Spm对分枝数的增加幅度相同, 均比单一盐胁迫植株增加了20.68%, Spd对分枝数的增加最显著, 增加达31.02%。可见, 外源多胺能有效地缓解盐胁迫对花生生长的抑制作用。
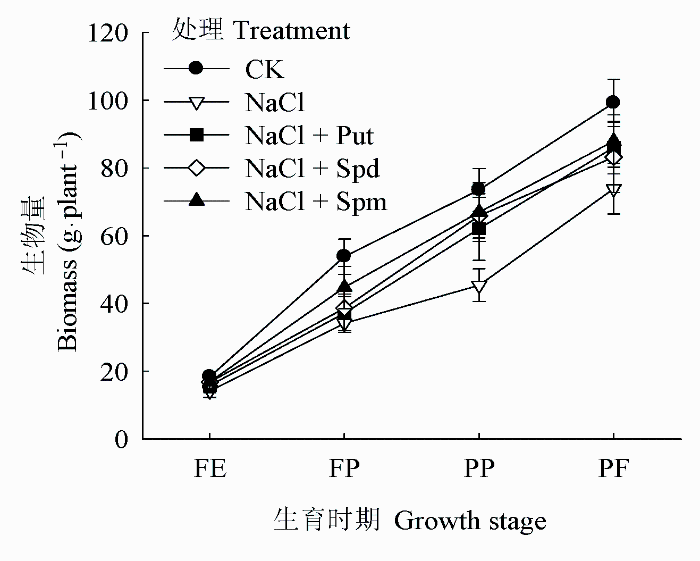
盐胁迫降低了各时期花生总生物量的积累(图1), 较CK的FE、FP、PP和PF时期分别下降了22.86%、36.58%、38.22%和25.54%。外源多胺能提高花生植株总生物量, 其中Spm对总生物量的贡献大于Spd和Put处理, 各时期内相对于单纯盐胁迫处理分别增加了19.15%、30.96%、47.48%和18.99%, 其中在FP和PP两时期的增加幅度尤为明显。
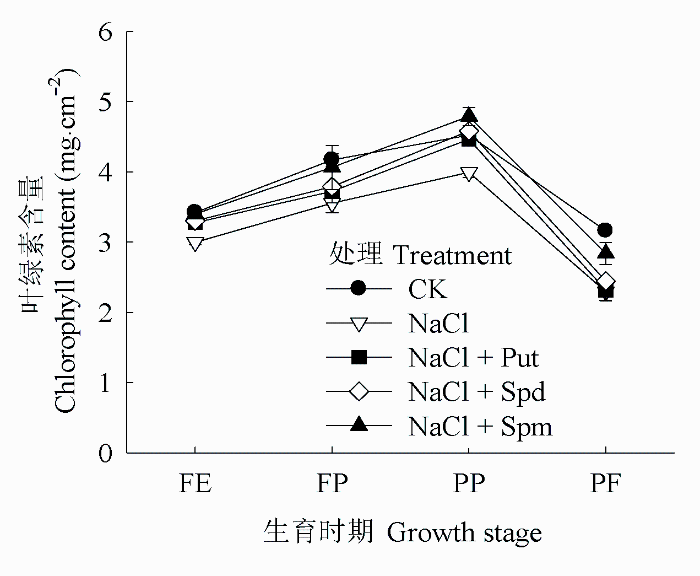
由图2可知, 花生叶片叶绿素含量在结荚期达到峰值, 随后逐渐进入衰老进程, 叶绿素含量下降迅速。与CK相比, 盐胁迫降低了各时期的叶绿素含量。叶面喷施外源多胺能够减缓叶绿素含量的下降。其中Spm效果最明显, 在FE、FP、PP和PF等时期叶绿素含量分别比单纯盐胁迫处理高出13.45%、14.38%、19.95%、23.58%。外源Put和Spd两处理也能提高叶绿素含量, 但两个处理间差异不明显。
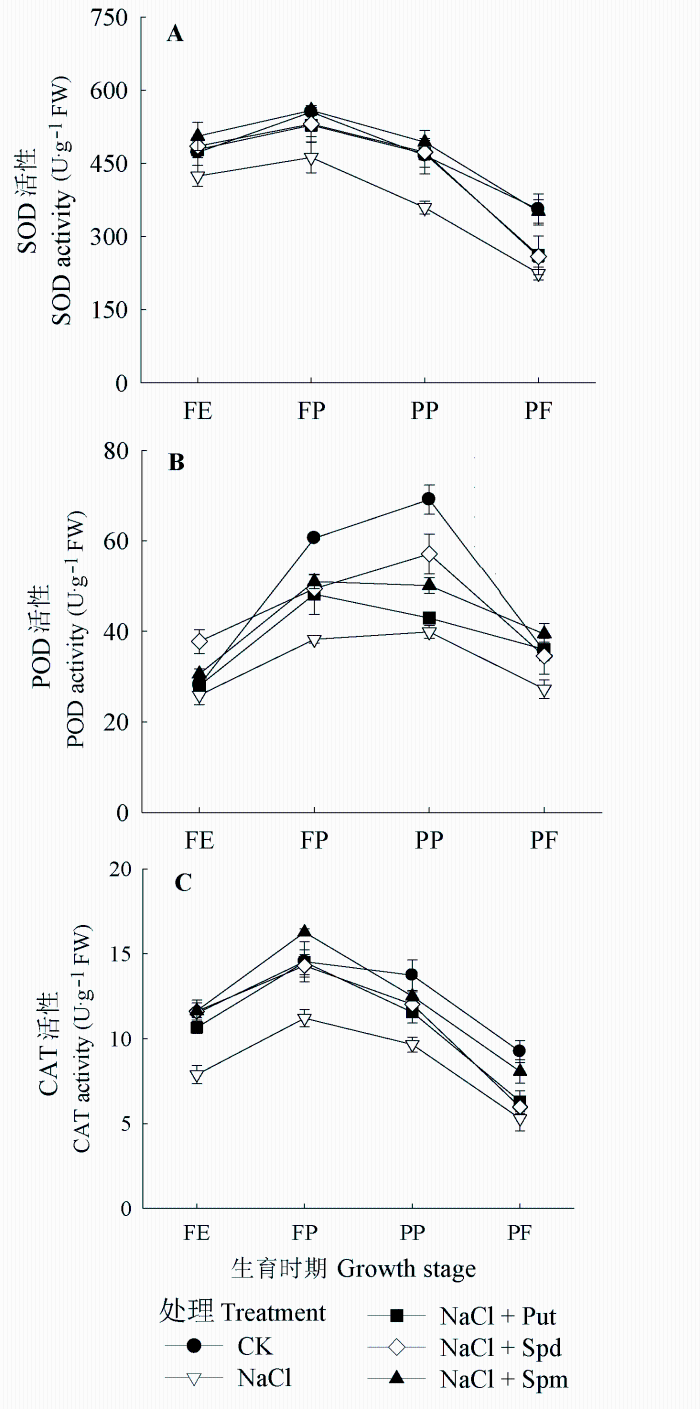
在整个生育时期内花生叶片活性氧清除酶SOD、POD、CAT活性整体呈先增加后下降的趋势。盐胁迫处理后各时期SOD、POD、CAT活性都明显低于CK, 外源喷施多胺均提高了活性氧清除酶活性(图3)。其中以喷施Spm缓解效果最好, 与单纯盐胁迫相比, FE、FP、PP和PF 4个时期的SOD活性分别增加了19.20%、20.87%、37.35%和56.73%, POD活性分别增加了18.05%、33.46、25.66%和44.63%, CAT活性分别增加了47.67%、45.22%、29.49%和52.52%。喷施Put处理和Spd处理对活性氧清除酶活性的影响整体上小于Spm处理, 其中Put和Spd两处理对SOD活性的影响没有明显差异, Spd处理对POD活性和CAT活性的影响略强于Put处理。说明施外源多胺可以提高花生叶片活性氧清除酶活性, 有利于对活性氧的清除, 缓解盐胁迫下因活性氧积累对花生植株造成的伤害, 且Spm和Spd处理的作用效果好于Put。
表1 盐胁迫下外源多胺对花生植株农艺性状的影响(平均值±标准偏差)
Table 1 Effects of exogenous polyamines on agronomic characters of Arachis hypogaea seedlings under salt stress (mean ± SD)
| 处理 Treatment | 主茎高 Plant height (cm) | 侧枝长 Branch length (cm) | 分枝数 Branch number |
|---|---|---|---|
| CK | 31.33 ± 0.63a | 32.17 ± 0.31a | 14.33 ± 0.24a |
| NaCl | 26.50 ± 0.25c | 27.00 ± 0.48b | 9.67 ± 0.24c |
| NaCl + Put | 27.67 ± 0.29bc | 30.50 ± 0.85ab | 11.67 ± 0.47b |
| NaCl + Spd | 29.33 ± 0.52b | 29.25 ± 0.64ab | 12.67 ± 0.24ab |
| NaCl + Spm | 29.67 ± 0.58b | 31.50 ± 0.38a | 11.67 ± 0.24b |
图1 盐胁迫下外源多胺对花生植株总生物量的影响(平均值±标准偏差)。FE、FP、PP和PF分别代表始花期、下针期、结荚期和收获期。Put、Spd、Spm同
Fig. 1 Effects of exogenous polyamines on total biomass of Arachis hypogaea under salt stress (mean ± SD). FE, FP, PP and PF represent flower early stage, flower-pegging stage, pod-setting stage and pod filling stage, respectively. Put, Spd, Spm see
图2 盐胁迫下外源多胺对花生叶片叶绿素含量的影响(平均值±标准偏差)。FE、FP、PP和PF分别代表始花期、下针期、结荚期和收获期。Put、Spd、Spm同
Fig. 2 Effects of exogenous polyamines on leaf chlorophyll content of Arachis hypogaea under salt stress (mean ± SD). FE, FP, PP and PF represent flower early stage, flower-pegging stage, pod-setting stage and pod filling stage, respectively. Put, Spd, Spm see
图3 盐胁迫下外源多胺对花生叶片超氧化物歧化酶(SOD)活性(A)、过氧化物酶(POD)活性(B)、过氧化氢酶(CAT)活性(C)的影响(平均值±标准偏差)。FE、FP、PP和PF分别代表始花期、下针期、结荚期和收获期。Put、Spd、Spm同
Fig. 3 Effects of exogenous polyamines on superoxide dismutase (SOD) activity (A), peroxidase (POD) activity (B) and catalse (CAT) activity (C) of Arachis hypogaea leaves under salt stress (mean ± SD). FE, FP, PP and PF represent flower early stage, flower-pegging stage, pod-setting stage and pod filling stage, respectively. Put, Spd, Spm see
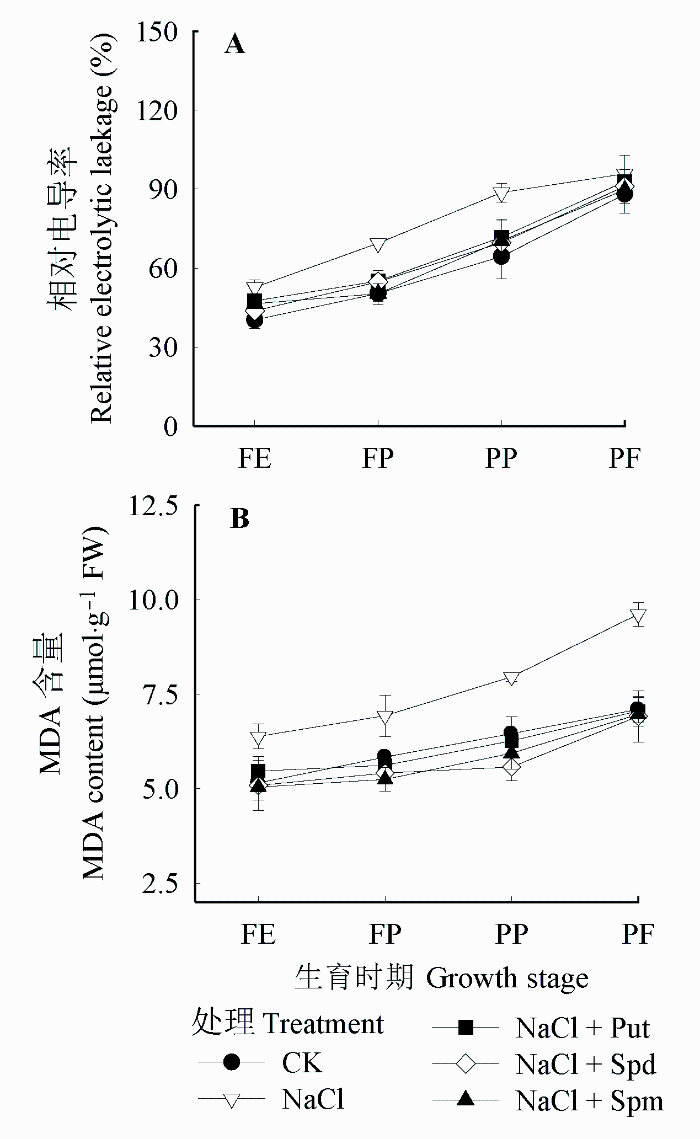
植物细胞受胁迫损伤程度可通过细胞膜相对电导率和MDA含量的变化来反映。随着植株的生长发育, 花生叶片细胞膜相对电导率和MDA含量都逐渐增加。由图4可知, 与CK相比, 盐胁迫增大了生物膜损伤程度, 各生育时期的相对电导率和MDA含量均高于对照。外源喷施多胺可有效地降低盐胁迫对花生生物膜的损伤。喷施Put、Spd、Spm与单纯盐胁迫相比, 相对电导率在FE期分别下降了9.98%、17.08%和11.80%, 在FP期分别增加下降了20.80%、21.15%和27.63%, 而在PP期与PF期, 3种处理之间没有明显差异。喷施Put、Spd、Spm与单纯盐胁迫相比, MDA含量在FE期分别下降了14.41%、20.34%和21.02%, 在FP期分别下降了18.93%、21.95%和24.34%, 在PP期分别下降了21.26%、29.95%和25.42%, 在PF期3种处理之间没有明显差异。由此可见, 盐胁迫使花生叶片生物膜透性增大, 可能与活性氧过量积累引起的膜脂过氧化程度加剧有关, 外源喷施多胺, 尤其是Spd和Spm, 能有效地降低叶片相对电导率和MDA含量, 减轻盐胁迫对花生植株的伤害。
图4 盐胁迫下外源多胺对花生叶片相对电导率(A)和丙二醛(MDA)含量(B)的影响(平均值±标准偏差)。FE、FP、PP和PF分别代表始花期、下针期、结荚期和收获期。Put、Spd、Spm同
Fig. 4 Effects of exogenous polyamines on relative electrolytic leakage (A) and malondialdehyde (MDA) content (B) of Arachis hypogaea leaves under salt stress (mean ± SD). FE, FP, PP and PF represent flower early stage, flower-pegging stage, pod-setting stage and pod filling stage, respectively. Put, Spd, Spm see
从表2可以看出, 盐胁迫显著降低了花生单株果针数、单株总果数和饱果数, 从而降低了单株产量, 外源喷施多胺能够在一定程度上提高盐胁迫下的花生产量, Put、Spd、Spm处理相对于单纯盐胁迫产量分别提高了10.34%、13.38%和16.90%。3种多胺对果针数及饱果数均有明显影响, 其中Spm的作
用最显著, 从而能够显著提高单株产量。
盐胁迫主要通过抑制或诱导多种酶系统以及气孔导度等途径来降低光合作用和能量代谢, 抑制植物组织的生长与发育, 从而降低植物干物质积累与植物产量(Parida & Das, 2005)。用Put和Spd浸种均可缓解盐胁迫对大麦(Hordeum vulgare)幼苗的盐害, 促进生长和干物质积累(Sun et al., 2002)。Krishnam- urthy和Bhagwat (1989)的研究表明, 高浓度NaCl胁迫下, 水稻种子萌发和生长受阻, 但外源Put可恢复种子萌发和生长, 促进盐胁迫下水稻禾苗的生长, 增加稻谷产量。
本实验中叶面喷施外源多胺能够缓解盐胁迫对植株生长的抑制, 提高花生主茎高度和侧枝长度的生长, 分枝数增多(表1), 干物质积累量增加(图1), 荚果产量提高(表2)。
光合作用为植物生长发育提供物质与能量, 是植物生长发育的基础。而叶绿素是光合作用中光反应阶段色素蛋白复合体的重要组成部分, 叶绿素含量的高低直接影响植物的光合作用能力(Maslenkova et al., 1993)。盐胁迫不仅能提高叶绿素酶的活性, 加速叶绿素的降解, 也能抑制叶绿素的合成(刁丰秋等, 1997)。本实验中盐胁迫处理显著降低了叶绿素含量, 喷施3种多胺在生育前期能够有效地提高叶绿素含量的积累, 在生育后期能有效地缓解叶绿素的降解速率(图2)。有研究表明, 外源多胺能够延缓黑暗诱导下离体叶片的衰老, 特别是Spd和Spm具有明显的保绿和抑制蛋白质降解的作用(蒋琳等, 1993), 王晓云等(1999)也发现大田抗衰老能力强的花生品种连体叶片内具有较高的Spd和Spm含量, 从而可减缓叶绿素的降解。
胁迫条件下叶绿素的降解与活性氧有关。Shioi等(1991)认为活性氧攻击叶绿素4吡咯环而导致卟啉大环裂解, 加速了叶绿素的降解和含量降低。本实验结果表明: 3种多胺不同程度地提高了叶片活性氧清除酶(SOD、POD、CAT)的活性(图3), 从而抑制了活性氧的生成速率, 延缓了叶绿素的降解。其中外源Spm对活性氧清除酶活性的作用效果最显著。徐仰仓等(2001)对春小麦的研究表明: 外源Spm既可直接作用于活性氧清除酶分子而提高活性, 也可以诱导活性氧清除酶分子的合成。刘俊等(2006)研究表明外施多胺通过提高玉米幼苗叶绿体结合态多胺含量而增加叶片PSII光化学效率、净光合速率和活性氧清除酶活性而缓解盐害, 其中Put的效果最好。这与本实验花生结果略有差异, 可能是因为植物种类的不同(Gill & Tuteja, 2010)。
盐胁迫对质膜的伤害被认为是盐对植物的直接原初伤害, 过剩的活性氧自由基引发或加剧膜脂过氧化, 造成细胞膜系统的损伤, 导致细胞膜的透性增大(Larkindale & Huang, 2004)。外源多胺的喷施使得活性氧清除酶活性大幅提高, 从而有效地降低活性氧, 减轻膜脂过氧化(图4)。
表2 盐胁迫下外源多胺对花生产量的影响(平均值±标准偏差)
Table 2 Effects of exogenous polyamines on the yield of Arachis hypogaea under salt stress (mean ± SD)
| 处理 Treatment | 产量 Pods mass per plant (g) | 饱果数 Full fruits number per plant | 总果数 Total pod number per plant | 果针数 Gynophores number per plant |
|---|---|---|---|---|
| CK | 47.57 ± 1.59a | 29.67 ± 0.67a | 39.70 ± 1.33a | 67.00 ± 2.00a |
| NaCl | 35.50 ± 0.93c | 16.21 ± 0.33c | 30.67 ± 2.67b | 40.67 ± 1.48d |
| NaCl + Put | 39.17 ± 0.87bc | 17.67 ± 0.33bc | 31.00 ± 0.67b | 43.00 ± 2.67cd |
| NaCl + Spd | 40.25 ± 1.13b | 21.00 ± 1.24b | 31.67 ± 0.48b | 45.00 ± 1.67c |
| NaCl + Spm | 41.50 ± 1.32b | 20.67 ± 0.67b | 39.33 ± 1.24a | 57.00 ± 3.33b |
本实验结果表明: 整体而言Spm对花生盐胁迫的缓解作用效果最好, 其原因可能在于多胺作用机制的复杂性。多胺可以作用于植物的离子通道, 阻断一种液泡膜上快速激活的离子通道, 从而阻断Na+渗透到细胞质基质中(Bruggemann et al., 1998; Kusano et al., 2007a), 并且不同多胺的阻断强度可能取决于多胺所带净电荷量, 即3种常见多胺对离子通道的阻断效果Spm > Spd >> Put (Kusano et al., 2007b)。Yamaguchi等(2007)对缺失Spm拟南芥突变体的研究则表明Spm可能通过控制钙渗透性通道调节气孔关闭, 从而提高干旱胁迫下拟南芥的光合作用。Jouve等(2004)、Ghosh等(2011)、Roychoudhury等(2008)都发现植物耐盐性的提高与植物体内源Spm含量的增加有关。内源多胺尤其是Spm提高植物抗盐性可能与多胺调节质膜H+-ATPase的活性有关。Roy等(2005)发现耐盐植物根部的质膜处富含Spd和Spm, 而盐敏感型植物此处富含Put。
综上所述, 不同植物中多胺对盐胁迫的调节作用机制可能在大田盆栽花生上也适用, 外源多胺的施用, 尤其是Spm, 减轻了盐胁迫对花生生长的危害, 增加了盐胁迫下花生叶片叶绿素含量和抗氧化酶活性, 有利于光合作用的进行、干物质的积累。在生产上, 花生的产量主要取决于花生植株的光合作用能力及干物质积累速率。因此, 本研究为盐碱地上花生的种植提供了参考。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
| [1] |
Long term exogenous putrescine application improves grain yield of a salt-sensitive rice cultivar exposed to NaCl. |
| [2] |
Cytoplasmic polyamines block the fast-activating vacuolar cation channel. |
| [3] |
Changes in composition and function of thylakoid membrane isolated from barley seedling leaves under salt stress. 盐胁迫对大麦叶片类囊体膜脂组成和功能的影响. |
| [4] |
Polyamines in plant physiology. |
| [5] |
Differential responses of two rice varieties to salt stress.
|
| [6] |
Superoxide dismutases II. Purification and quantitative relationship with water- soluble protein in seedling. |
| [7] |
Polyamines and abiotic stress tolerance in plants. |
| [8] |
The effect of polyamines on metabolism of active oxygen in detached leaves of Hordeum vulgare var. nudum Hook. f. 多胺对裸大麦离体叶片活性氧代谢的影响. |
| [9] |
Polyamine, carbohydrate, and proline content changes during salt stress exposure of aspen (Populus tremula L.): Involvement of oxidation and osmoregulation metabolism.
|
| [10] |
a). The polyamine spermine rescues Arabidopsis from salinity and drought stresses. |
| [11] |
b). Advances in polyamine research in 2007. |
| [12] |
Polyamines as modulators of salt tolerance in rice cultivars.
|
| [13] |
Changes of lipid composition and saturation level in leaves and roots for heat-stressed and heat-acclimated creeping bentgrass (Agrostis stolonifera). |
| [14] |
|
| [15] |
The present situation and hot issues in the salt-affected soil research. 土壤盐渍化研究现状及未来研究热点. |
| [16] |
Effects of exogenous polyamines on chloroplast-bound polyamines content and photosynthesis of corn suffering salt stress. 外源多胺对盐胁迫下玉米叶绿体结合态多胺水平和光合作用的影响. |
| [17] |
Polyamines and their ability to provide environmental stress tolerance to plants. |
| [18] |
Inward potassium channel in guard cells as a target for polyamine regulation of stomatal movements. |
| [19] |
Natural polyamines inhibit soybean (Glycine max) lipoxygenase-1, but not the lipoxygenase-2 isozyme. |
| [20] |
Adaptation to salinity as monitored by PSII oxygen evolving reactions in barley thylakoids. |
| [21] |
Salt tolerance and salinity effects on plants: A review. |
| [22] |
Spermidine treatment to rice seedlings recovers salinity stress-induced damage of plasma membrane and PM-bound H+-ATPase in salt-tolerant and salt-sensitive rice cultivars.
|
| [23] |
Comparative physiological and molecular responses of a common aromatic indica rice cultivar to high salinity with non-aromatic indica rice cultivars.
|
| [24] |
Amelioration of salinity stress by exogenously applied spermidine or spermine in three varieties of indica rice differing in their level of salt tolerance. |
| [25] |
Effects of exogenous spermine on chlorophyll fluorescence, antioxidant system and ultrastructure of chloroplasts in Cucumis sativus L. under salt stress. |
| [26] |
Putrescine facilitated enhancement of capsaicin production in cell suspension cultures of Capsicum frutescens. |
| [27] |
Mechanism of the effect of polyamine on the activity of tonoplasts of barley roots under salt stress. |
| [28] |
Opportunities facing peanut industry in china and strategies for its science and technology development. 我国花生产业面临的机遇与科技发展战略. |
| [29] |
Research on changes of polyamines during leaf senescence of different type peanut varieties. 花生不同衰老型品种叶片衰老过程中多胺变化规律的研究. |
| [30] |
Promoting effect of exogenous spermine on anti-oxidative enzyme activity in wheat seedlings. 外源精胺对小麦幼苗抗氧化酶活性的促进作用. |
| [31] |
A protective role for the polyamine spermine against drought stress in Arabidopsis.
|
| [32] |
Enzymatic degradation of chlorophyll in Chenopodium album. Plant Cell Physiology ,
|
| [33] |
Comparison of the changes of membrane protective system in rice seedlings during enhancement of chilling resistance by different stress pretreatment. 不同胁迫预处理提高水稻幼苗抗寒性期间膜保护系统的变化比较. |
| [34] |
|
| [35] |
Improvements of method for measurement of malondialdehyde in plant tissue. 植物组织中丙二醛测定方法的改进. |

/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}