Response of tree growth to nitrogen addition in a Larix gmelinii primitive forest
LIU Xiu-Yuan1, 2, DU En-Zai3, XU Long-Chao1, 2, SHEN Hai-Hua1, FANG Jing-Yun1, HU Hui-Feng1, *,
1State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China2University of Chinese Academy of Sciences, Beijing 100049, Chinaand 3College of Urban and Environmental Sciences, and Key Laboratory for Earth Surface Processes of the Ministry of Education, Peking University, Beijing 100871, China
氮沉降对树木生长的影响是全球变化研究的一个核心问题。该文通过设置4种氮添加水平(对照(0)、低氮(20 kg N·hm-2·a-1)、中氮(50 kg N·hm-2·a-1)和高氮(100 kg N·hm-2·a-1)), 研究了模拟氮沉降对落叶松(Larix gmelinii)原始林树木胸径生长的影响。结果表明: 中氮和高氮添加对落叶松胸径相对生长率有显著影响, 而且这种影响随施氮年限的增加而增强。不同高度的树木对氮添加的响应有很大差异, 较低树木(树高<16.5 m)的生长对氮添加无显著响应, 而较高(树高>16.5 m)的树木在中氮和高氮处理下胸径生长有显著加速(胸径相对生长率增幅>79.5%), 但随着树木高度的进一步增加, 这种加速作用明显下降。研究结果显示氮添加会促进落叶松胸径生长, 这种促进作用主要发生在较高的落叶松个体中。
关键词:北方森林
;
胸径
;
落叶松
;
氮沉降
;
树木生长
Abstract
Aims
Global nitrogen (N) deposition has been consistently enhanced and significantly influenced the carbon (C) cycle of forest ecosystem. However, studies on the quantitive response of C sequestration to N deposition in China’s boreal forest ecosystem still remains insufficient. Tree growth is one of important components of C sequestration in forest ecosystem. In order to accurately evaluate the influence of N deposition on C sequestration in China’s boreal forest ecosystem, This study was designed to investigate how N deposition influenced the tree diameter at breast height (DBH) growth of a Larix gmelinii primitive forest.
Methods
We carried out a N enrichment experiment in an L. gmelinii primitive forest in Greater Khingan Mountains with four N addition levels (control (0), low N (20 kg N·hm-2·a-1), medium N (50 kg N·hm-2·a-1) and high N (100 kg N·hm-2·a-1)). DBH was measured from 2012-2014 within all treatments.
Important findings
Medium N and high N additions significantly influenced the relative DBH growth of L. gmelinii, and the influence became more pronounced through time. N deposition significantly affected the DBH growth among different tree height classes: low height class (tree height <16.5 m) did not show significant response to N deposition, while high height class (tree height >16.5 m) showed significantly accelerated growth under medium and high N additions (the relevant DBH growth rate >79.5%), and this acceleration effect decreased as the tree height increased. This study indicated that N addition did enhance the DBH growth of L. gmelinii, but this enhancement mainly affected taller L. gmelinii trees.
Keywords:boreal forest
;
diameter at breast height
;
Larix gmelinii
;
nitrogen deposition
;
tree growth
LIUXiu-Yuan, DUEn-Zai, XULong-Chao, SHENHai-Hua, FANGJing-Yun, HUHui-Feng. Response of tree growth to nitrogen addition in a Larix gmelinii primitive forest. Chinese Journal of Plant Ecology, 2015, 39(5): 433-441 https://doi.org/10.17521/cjpe.2015.0042
为此, 本研究利用北京大学生态学系牵头建立的中国森林生态系统养分添加实验网络(Nutrient Enrichment Experiments in Chinese Forests, NEECF)平台(Du et al., 2013), 对平台最北端内蒙古根河站点中的落叶松原始林进行了连续3年的乔木胸径监测, 试图回答以下两个问题: 1)落叶松胸径生长对氮添加是如何响应的, 这种响应在不同氮添加水平及不同年份下是否有差异?2)原始林中不同树高级的落叶松胸径生长对氮添加的响应有何差异?
1 材料和方法
1.1 样地概况
本试验地为NEECF试验网络平台中最北端的根河寒温带针叶林样地, 样地位于大兴安岭西北坡的内蒙古大兴安岭森林生态系统国家野外科学观测研究站(50.93° N, 121.50° E), 海拔780-1142 m, 地貌以丘陵为主, 平均坡度12°。年平均气温-5.4 ℃, 最低气温-55 ℃ (2009), >10 ℃积温1403 ℃, 年日照时间2594 h。年降水量450-550 mm, 60%集中在7、8月份, 降雪厚度20-40 cm, 地表蒸散量800- 1200 mm, 无霜期80天。冬季(候平均气温<10 ℃)长达9个月, 夏季(候平均气温>22 ℃)不到一个月。受太平洋暖湿空气影响, 降水集中在7、8月份, 占全年降水量的50%-70%, 而冬季的11月至次年4月受蒙古高压的影响, 降水量仅占全年降水量的12%左右。典型土壤类型为棕色针叶林土, 土层平均厚度20-40 cm, 土壤呈酸性pH为4.5-6.5 (杜恩在, 2013)。研究区域基本没有受到污染, 背景氮沉降水平仅为0.2 g N ·m-2·a-1 (Sheng et al., 2013)。
所选样地为落叶松纯林, 总体平均林分密度为(840 ± 72)株·hm-2, 平均胸径为(17.1 ± 2.1) cm, 平均树高(15.6 ± 1.4) m, 最大树龄大于300年(图1) (Du & Fang, 2014)。样地不同处理组间落叶松的起始株数、胸径及树高均无显著差异(起始株数: p = 0.84; 胸径: p = 0.84; 树高: p = 0.87)。样地垂直结构分为3层: (1)乔木层(>3 m), 以落叶松为优势种; (2)灌木层(0.3-3.0 m), 物种组成包括: 白桦(Betula platyphylla)、柴桦(B. fruticosa)、杜香(Ledum palustre)、兴安杜鹃(Rhododendron dauricum)、笃斯越桔(Vaccinium uliginosum); (3)地被层(< 0.3 m), 包括红花鹿蹄草(Pyrola asarifolia subsp. incarnate)、舞鹤草(Maianthemum bifolium)、越桔(V. vitis-idaea)及垂枝藓(Rhytidium rugosum)等(杜恩在, 2013)。
Fig. 1 Diameter at breast height (DBH) and tree height frequency distribution in Larix gmelinii primitive forest stand (Du, 2013).
1.2 研究方法
1.2.1 试验方案
模拟氮沉降试验基于NEECF试验平台而建立, 试验始于2010年, 采用随机区组设计, 分别设对照 (0)、低氮(20 kg N·hm-2·a-1)、中氮(50 kg N·hm-2·a-1)和高氮(100 kg N·hm-2·a-1)共4个氮添加水平的处理, 每种处理设3个重复, 样方面积为20 m × 20 m。相邻样方之间留有10 m宽的缓冲带, 每块样方中心15 m × 15 m的面积划分成9个5 m × 5 m的小区, 用于观测和采样。氮添加梯度的设置考虑到与以往同类实验的可比性设置了50和100 kg N·hm-2·a-1的氮添加梯度, 同时考虑到敏感森林生态系统较低的氮沉降临界负荷(Bobbink et al., 2010), 设置了20 kg N·hm-2·a-1的氮添加梯度。氮添加处理平均分5次在每年5月至9月每月上旬进行, 将相应量NH4NO3溶于30 L水混合均匀, 用背式喷雾器均匀地喷洒到对应的样方林下, 对照组喷洒相同量的水。非生长季有雪覆盖期间不作任何处理。详细的实验方案见文献(杜恩在, 2013)。
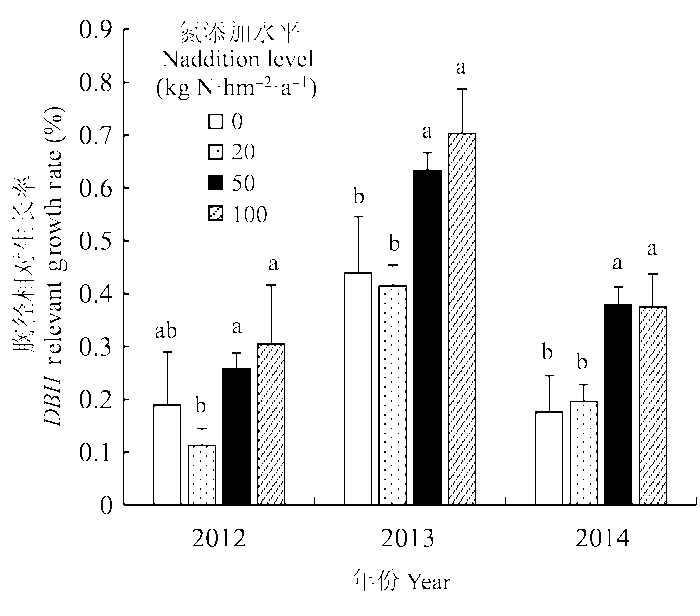
Fig. 3 Responses of Larix gmelinii diameter at breast height (DBH) relative growth rate under different N treatments in different years. a and b represent the significant level of multiple comparisons of mean DBH relevant growth rate under different N treatments in each year (p < 0.05).
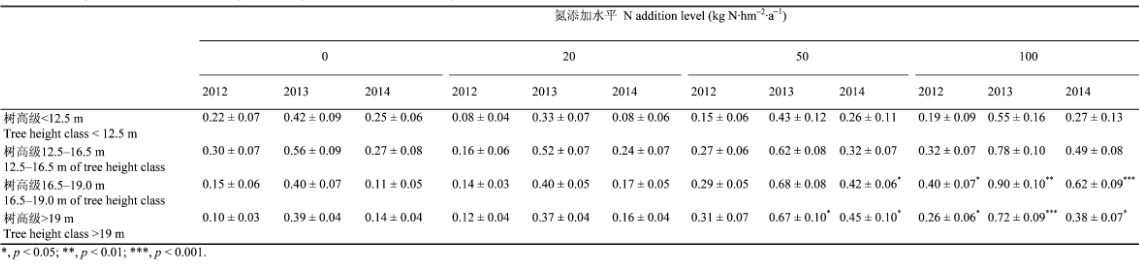
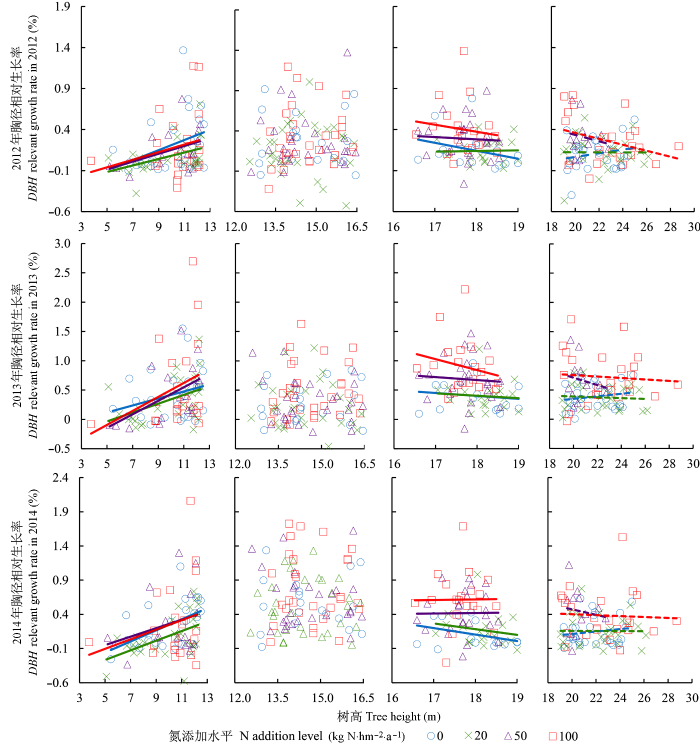
在<12.5 m树高级下, 落叶松胸径相对生长率均与树高呈显著正相关关系(p < 0.01), 而不同氮处理间则均无显著差异(表2; 图4)。12.5-16.5 m树高级落叶松胸径相对生长率与树高在3年间均无显著相关关系(2012: p = 0.55; 2013: p = 0.9; 2014: p = 0.99), 且其对氮添加并无显著响应(表2; 图4)。
Table 2
表2
表2 2012-2014年各树高级落叶松胸径相对生长率(%)(平均值±标准误差)
Table 2 Larix gmelinii diameter at breast height relevant growth rate(%) of all tree height classes from 2012 to 2014(mean±SE)
随着树高的增加, 落叶松胸径相对生长率对氮添加开始产生响应。三年间16.5-19.0 m树高级落叶松胸径相对生长率与树高的关系均呈显著负相关关系(2012: p = 0.03; 2013: p = 0.01; 2014: p = 0.02), 而氮添加对其胸径相对生长率具有促进作用(图4), 其中3年高氮处理和2014年中氮处理的促进作用均显著, 胸径相对生长率增幅大于124.2% (表2)。>19 m树高级胸径相对生长率与树高均无显著相关关系(2012: p = 0.3; 2013: p = 0.92; 2014: p = 0.89), 但对氮添加表现正向响应(图4), 3年中氮以及后两年高氮处理下胸径相对生长率均显著增加, 增幅大于79.5% (表2)。
3 讨论
3.1 落叶松胸径生长对氮添加的响应及年际间 变化
总体而言, 氮素添加增加了落叶松原始林的胸径相对生长速率, 说明氮是研究区域内森林生态系统中的一个限制因素, 这与已有对加拿大北方森林短期氮添加研究的结果一致(Newton & Amponsah, 2006; Jung & Chang, 2012)。然而, 研究区域内落叶松乔木在不同氮添加水平下的响应却是不一样的。低水平氮添加(20 kg N·hm-2·a-1)在短期内对树木相对生长影响不显著, 而在中氮(50 kg N·hm-2·a-1)和
高氮(100 kg N·hm-2·a-1)处理下, 落叶松胸径相对生长率对氮添加存在显著响应(图3), 这与Houle和Moore (2008)对加拿大北方森林为期3年的研究结果一致。事实上, 北方森林乔木在高水平氮添加与自然情况下低水平氮沉降的响应机制确实存在差异。例如, 研究表明在自然情况下较低水平的氮沉降对北方森林乔木的影响受到苔藓层的抑制, 而在高氮添加水平下, 活性氮将绕过苔藓层直接作用于乔木(Gundale et al., 2011; Du et al., 2014)。但是, Högberg等(2006)和Magnani等(2007)的长期研究结果则表明, 低水平氮添加也会促进氮限制森林乔木的生长。因此, 还需要继续对落叶松进行低水平、长时期的氮添加试验, 以模拟真实氮沉降情况和预测乔木的响应(Högberg et al., 2006)。
Fig. 4 The analysis of covariance (ANCOVA) between Larix gmelinii diameter at breast height (DBH) relevant growth rate and tree height and N addition. The solid line represents there exists significant correlativity between DBH relevant growth rate and tree height (p < 0.05). The dash line represents there does not exist significant correlativity between DBH relevant growth rate and tree height, but the ANOVA shows significant difference between different nitrogen treatments (p < 0.01). The figure with only point represents there does not exist significant correlativity between DBH relevant growth rate and tree height, and the ANOVA shows no significant difference between different nitrogen treatments.
Four years of simulated N and S depositions did not cause N saturation in a mixedwood boreal forest ecosystem in the oil sands region in northern Alberta, Canada.
... 树木生长是北方森林生态系统固碳过程的重要组分之一.乔木树干木质组织C:N高达500:1 (Magnani et al., 2007), 而且随氮沉降增加, 土壤可利用氮水平提高, 越来越多的碳将被分配至地上木质组织中(Litton et al., 2007).已有研究表明: 模拟氮沉降会促进树木生长(Magnani et al., 2007; Ciais et al., 2008), 但对氮添加的响应程度受到研究地点、施肥量、肥料种类、施肥年限等多种因素的影响.一般来说, 研究地点背景氮沉降水平越高, 施肥量越小, 施肥年限越长, 树龄越大, 树木生长对氮添加的响应越弱(Gundersen et al., 1998; 周利勋等, 2004; Rappe-George et al., 2013).此外, 由于不同树种对氮的利用和分配不同, 导致对氮添加的响应也不相同(Hyvönen et al., 2008).除了上述因素之外, 树高也是影响树木对氮添加响应的重要因素, 尤其在树种单一的群落中.树高不仅表征了树木对地上空间和光照的竞争能力, 还与树木根系面积及侧根长度等存在联系.研究表明, 树木越高, 树木根系面积越大, 其对养分的竞争能力越强, 对氮添加的响应程度可能越高(Kajimoto et al., 1999, 2003).此外, 树高与树龄也存在较好的正相关关系(O’Brien et al., 1995).因此, 研究不同高度树木对氮添加的响应, 将有助于树木种群发展趋势的预测以及生态系统碳汇功能的准确评估. ...
Short and long-term impacts of nitrogen deposition on carbon sequestration by forest ecosystems.
1
2014
... 自工业革命以来, 人类活动产生的活性氮不断增加, 全球氮沉降水平不断上升, 并且氮沉降速率在达到峰值前还会增加2到3倍(Lamarque et al., 2005; Galloway et al., 2008).氮沉降的持续增加已经对森林生态系统结构和功能造成了多方面的影响(Magill et al., 2000; de Vries et al., 2003; Janssens & Luyssaert, 2009; Bobbink et al., 2010), 其中对森林生态系统碳固存的影响是近年来生物地球化学循环领域的热点之一.作为全球最大的森林生物群区和重要的碳库组成之一, 北方森林总碳库约为272 Pg C (占全球森林碳库的32%), 且具有稳定的碳汇功能(碳汇强度为0.5 Pg C·a-1, 占全球森林碳汇的20%左右)(Pan et al., 2011).此外, 由于北方森林长期以来一直处于氮限制状态(Vitousek & Howarth, 1991), 其碳固存功能在氮沉降逐渐增加的情况下将如何响应引起了广泛的关注.目前, 欧洲及美国已对此进行了较为系统的监测研究, 发现氮沉降会显著增加北方森林的碳汇功能(de Vries et al., 2014).然而, 不同研究的量化结果存在较大差异(Magnani et al., 2007; de Vries et al., 2008; Sutton et al., 2008), 这些差异的主要来源之一是森林生态系统中树木生长对氮沉降响应的不确定性(Janssens & Luyssaert, 2009). ...
Intensive monitoring of forest ecosystems in Europe: 2: Atmospheric deposition and its impacts on soil solution chemistry.
1
2003
... 自工业革命以来, 人类活动产生的活性氮不断增加, 全球氮沉降水平不断上升, 并且氮沉降速率在达到峰值前还会增加2到3倍(Lamarque et al., 2005; Galloway et al., 2008).氮沉降的持续增加已经对森林生态系统结构和功能造成了多方面的影响(Magill et al., 2000; de Vries et al., 2003; Janssens & Luyssaert, 2009; Bobbink et al., 2010), 其中对森林生态系统碳固存的影响是近年来生物地球化学循环领域的热点之一.作为全球最大的森林生物群区和重要的碳库组成之一, 北方森林总碳库约为272 Pg C (占全球森林碳库的32%), 且具有稳定的碳汇功能(碳汇强度为0.5 Pg C·a-1, 占全球森林碳汇的20%左右)(Pan et al., 2011).此外, 由于北方森林长期以来一直处于氮限制状态(Vitousek & Howarth, 1991), 其碳固存功能在氮沉降逐渐增加的情况下将如何响应引起了广泛的关注.目前, 欧洲及美国已对此进行了较为系统的监测研究, 发现氮沉降会显著增加北方森林的碳汇功能(de Vries et al., 2014).然而, 不同研究的量化结果存在较大差异(Magnani et al., 2007; de Vries et al., 2008; Sutton et al., 2008), 这些差异的主要来源之一是森林生态系统中树木生长对氮沉降响应的不确定性(Janssens & Luyssaert, 2009). ...
Ecologically implausible carbon response?
1
2008
... 自工业革命以来, 人类活动产生的活性氮不断增加, 全球氮沉降水平不断上升, 并且氮沉降速率在达到峰值前还会增加2到3倍(Lamarque et al., 2005; Galloway et al., 2008).氮沉降的持续增加已经对森林生态系统结构和功能造成了多方面的影响(Magill et al., 2000; de Vries et al., 2003; Janssens & Luyssaert, 2009; Bobbink et al., 2010), 其中对森林生态系统碳固存的影响是近年来生物地球化学循环领域的热点之一.作为全球最大的森林生物群区和重要的碳库组成之一, 北方森林总碳库约为272 Pg C (占全球森林碳库的32%), 且具有稳定的碳汇功能(碳汇强度为0.5 Pg C·a-1, 占全球森林碳汇的20%左右)(Pan et al., 2011).此外, 由于北方森林长期以来一直处于氮限制状态(Vitousek & Howarth, 1991), 其碳固存功能在氮沉降逐渐增加的情况下将如何响应引起了广泛的关注.目前, 欧洲及美国已对此进行了较为系统的监测研究, 发现氮沉降会显著增加北方森林的碳汇功能(de Vries et al., 2014).然而, 不同研究的量化结果存在较大差异(Magnani et al., 2007; de Vries et al., 2008; Sutton et al., 2008), 这些差异的主要来源之一是森林生态系统中树木生长对氮沉降响应的不确定性(Janssens & Luyssaert, 2009). ...
NEECF: A project of nutrient enrichment experiments in China’s forests.
1
2013
... 为此, 本研究利用北京大学生态学系牵头建立的中国森林生态系统养分添加实验网络(Nutrient Enrichment Experiments in Chinese Forests, NEECF)平台(Du et al., 2013), 对平台最北端内蒙古根河站点中的落叶松原始林进行了连续3年的乔木胸径监测, 试图回答以下两个问题: 1)落叶松胸径生长对氮添加是如何响应的, 这种响应在不同氮添加水平及不同年份下是否有差异?2)原始林中不同树高级的落叶松胸径生长对氮添加的响应有何差异? ...
氮添加对兴安落叶松原始林碳收支主要过程的影响
2
2013
... 所选样地为落叶松纯林, 总体平均林分密度为(840 ± 72)株·hm-2, 平均胸径为(17.1 ± 2.1) cm, 平均树高(15.6 ± 1.4) m, 最大树龄大于300年(图1) (Du & Fang, 2014).样地不同处理组间落叶松的起始株数、胸径及树高均无显著差异(起始株数: p = 0.84; 胸径: p = 0.84; 树高: p = 0.87).样地垂直结构分为3层: (1)乔木层(>3 m), 以落叶松为优势种; (2)灌木层(0.3-3.0 m), 物种组成包括: 白桦(Betula platyphylla)、柴桦(B. fruticosa)、杜香(Ledum palustre)、兴安杜鹃(Rhododendron dauricum)、笃斯越桔(Vaccinium uliginosum); (3)地被层(< 0.3 m), 包括红花鹿蹄草(Pyrola asarifolia subsp. incarnate)、舞鹤草(Maianthemum bifolium)、越桔(V. vitis-idaea)及垂枝藓(Rhytidium rugosum)等(杜恩在, 2013). ...
... 模拟氮沉降试验基于NEECF试验平台而建立, 试验始于2010年, 采用随机区组设计, 分别设对照 (0)、低氮(20 kg N·hm-2·a-1)、中氮(50 kg N·hm-2·a-1)和高氮(100 kg N·hm-2·a-1)共4个氮添加水平的处理, 每种处理设3个重复, 样方面积为20 m × 20 m.相邻样方之间留有10 m宽的缓冲带, 每块样方中心15 m × 15 m的面积划分成9个5 m × 5 m的小区, 用于观测和采样.氮添加梯度的设置考虑到与以往同类实验的可比性设置了50和100 kg N·hm-2·a-1的氮添加梯度, 同时考虑到敏感森林生态系统较低的氮沉降临界负荷(Bobbink et al., 2010), 设置了20 kg N·hm-2·a-1的氮添加梯度.氮添加处理平均分5次在每年5月至9月每月上旬进行, 将相应量NH4NO3溶于30 L水混合均匀, 用背式喷雾器均匀地喷洒到对应的样方林下, 对照组喷洒相同量的水.非生长季有雪覆盖期间不作任何处理.详细的实验方案见文献(杜恩在, 2013). ...
氮添加对兴安落叶松原始林碳收支主要过程的影响
2
2013
... 所选样地为落叶松纯林, 总体平均林分密度为(840 ± 72)株·hm-2, 平均胸径为(17.1 ± 2.1) cm, 平均树高(15.6 ± 1.4) m, 最大树龄大于300年(图1) (Du & Fang, 2014).样地不同处理组间落叶松的起始株数、胸径及树高均无显著差异(起始株数: p = 0.84; 胸径: p = 0.84; 树高: p = 0.87).样地垂直结构分为3层: (1)乔木层(>3 m), 以落叶松为优势种; (2)灌木层(0.3-3.0 m), 物种组成包括: 白桦(Betula platyphylla)、柴桦(B. fruticosa)、杜香(Ledum palustre)、兴安杜鹃(Rhododendron dauricum)、笃斯越桔(Vaccinium uliginosum); (3)地被层(< 0.3 m), 包括红花鹿蹄草(Pyrola asarifolia subsp. incarnate)、舞鹤草(Maianthemum bifolium)、越桔(V. vitis-idaea)及垂枝藓(Rhytidium rugosum)等(杜恩在, 2013). ...
... 模拟氮沉降试验基于NEECF试验平台而建立, 试验始于2010年, 采用随机区组设计, 分别设对照 (0)、低氮(20 kg N·hm-2·a-1)、中氮(50 kg N·hm-2·a-1)和高氮(100 kg N·hm-2·a-1)共4个氮添加水平的处理, 每种处理设3个重复, 样方面积为20 m × 20 m.相邻样方之间留有10 m宽的缓冲带, 每块样方中心15 m × 15 m的面积划分成9个5 m × 5 m的小区, 用于观测和采样.氮添加梯度的设置考虑到与以往同类实验的可比性设置了50和100 kg N·hm-2·a-1的氮添加梯度, 同时考虑到敏感森林生态系统较低的氮沉降临界负荷(Bobbink et al., 2010), 设置了20 kg N·hm-2·a-1的氮添加梯度.氮添加处理平均分5次在每年5月至9月每月上旬进行, 将相应量NH4NO3溶于30 L水混合均匀, 用背式喷雾器均匀地喷洒到对应的样方林下, 对照组喷洒相同量的水.非生长季有雪覆盖期间不作任何处理.详细的实验方案见文献(杜恩在, 2013). ...
Weak growth response to nitrogen deposition in an old-growth boreal forest.
Four years of simulated N and S depositions did not cause N saturation in a mixedwood boreal forest ecosystem in the oil sands region in northern Alberta, Canada.
1
2012
... 总体而言, 氮素添加增加了落叶松原始林的胸径相对生长速率, 说明氮是研究区域内森林生态系统中的一个限制因素, 这与已有对加拿大北方森林短期氮添加研究的结果一致(Newton & Amponsah, 2006; Jung & Chang, 2012).然而, 研究区域内落叶松乔木在不同氮添加水平下的响应却是不一样的.低水平氮添加(20 kg N·hm-2·a-1)在短期内对树木相对生长影响不显著, 而在中氮(50 kg N·hm-2·a-1)和 ...
Root system development of Larix gmelinii trees affected by micro-scale conditions of permafrost soils in central Siberia.
3
2003
... 树木生长是北方森林生态系统固碳过程的重要组分之一.乔木树干木质组织C:N高达500:1 (Magnani et al., 2007), 而且随氮沉降增加, 土壤可利用氮水平提高, 越来越多的碳将被分配至地上木质组织中(Litton et al., 2007).已有研究表明: 模拟氮沉降会促进树木生长(Magnani et al., 2007; Ciais et al., 2008), 但对氮添加的响应程度受到研究地点、施肥量、肥料种类、施肥年限等多种因素的影响.一般来说, 研究地点背景氮沉降水平越高, 施肥量越小, 施肥年限越长, 树龄越大, 树木生长对氮添加的响应越弱(Gundersen et al., 1998; 周利勋等, 2004; Rappe-George et al., 2013).此外, 由于不同树种对氮的利用和分配不同, 导致对氮添加的响应也不相同(Hyvönen et al., 2008).除了上述因素之外, 树高也是影响树木对氮添加响应的重要因素, 尤其在树种单一的群落中.树高不仅表征了树木对地上空间和光照的竞争能力, 还与树木根系面积及侧根长度等存在联系.研究表明, 树木越高, 树木根系面积越大, 其对养分的竞争能力越强, 对氮添加的响应程度可能越高(Kajimoto et al., 1999, 2003).此外, 树高与树龄也存在较好的正相关关系(O’Brien et al., 1995).因此, 研究不同高度树木对氮添加的响应, 将有助于树木种群发展趋势的预测以及生态系统碳汇功能的准确评估. ...



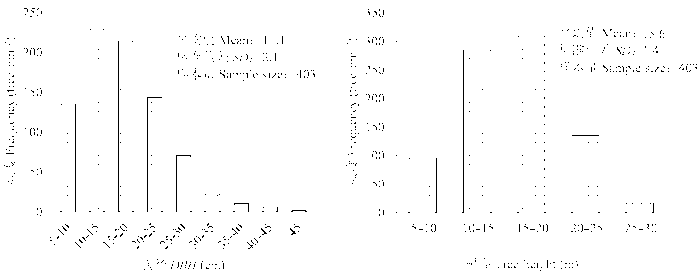
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}