近年来, 随着对森林根系活动和根际过程研究的不断深入, 根系在调控土壤功能和养分代谢过程中的重要作用逐渐被人们所认知和关注(Chapin et al ., 2009 ; Cheng et al ., 2014 )。植物除了通过细根周转向土壤输入碳(C)和养分外, 还可通过根系主动或被动地向周围土壤释放一系列化合物, 即狭义的根系分泌物, 主要包括一些低分子(有机酸、糖类、酚类和各种氨基酸等)和高分子(蛋白质、黏液等)有机化合物(Dijkstra & Cheng, 2007 )。研究表明, 通常森林根系分泌物量占植物光合同化产物的1%-5% (Grayston et al ., 1997 ), 并受森林类型、树种、环境条件以及土壤养分有效性等诸多因素影响Fransson & Johansson, 2010 ; Shi et al ., 2011 )。由于根系分泌物大多为一些易于土壤微生物直接利用的含碳有机物, 可为土壤微生物系统提供重要且丰富的C源和氮(N)源, 有效地改变土壤微生物数量和活性, 而深刻地影响根际土壤有机质分解和养分代谢等过程(Phillips et al ., 2009 ; Yin et al ., 2013 ), 从而导致根系分泌物在调控土壤C养分转化过程中发挥与其含量不成比例的重要作用和功能。植物根系分泌物输入及其介导的根系-土壤-微生物互作效应已成为陆地生态系统碳循环过程研究中一个非常重要但又最缺乏了解的关键环节(Jones et al ., 2004 ; 吴林坤等, 2014 )。
尽管当前对根系分泌物及其诱导的生态学效应研究已取得一定的进展, 但是对根系分泌物在森林根际生态过程中的作用及调控机理等方面我们依然缺乏足够的了解(Phillips et al ., 2011 ; Cheng et al ., 2014 )。目前有关森林根系分泌物研究大都仅关注了根系C源输入, 而忽略了根系分泌物中N成分变化及其伴随的C:N化学计量特征对土壤C养分循环过程的激发效应(priming effect), 这种忽略将极大地限制对森林根系-土壤-微生物互作机制的深入认识(Espinosa-Urgel & Ramos, 2001 ; Yin et al ., 2014 )。植物根系分泌物主要为一系列含碳化合物, 其C:N比值通常高于根际微生物的C:N (Cleveland & Liptzin, 2007 )。另外, 由于根系和微生物活动对根际有效N素的获取和激烈竞争, 使得根际区通常成为C过剩而N受限制强烈的区域(Kuzyakov, 2002 )。因此, 根系分泌物N含量或C:N化学计量特征成为驱动根际微生物群落组成和活性的重要调控因子。相应地, 根际微生物利用根系分泌物生长和合成胞外酶的能力严重地受根系分泌物N输入通量的制约, 从而反过来调控土壤生物地球化学过程及其对森林结构和功能的生态反馈效应(Drake et al ., 2013 )。此外, 根系分泌物对森林土壤C养分所诱导的激发效应与供试土壤养分状况(主要指土壤N的有效性)密切相关, 因为后者数量和质量是决定微生物对其生长和胞外酶产生之间能量分配的关键要素(Sullivan & Hart, 2013 ; Chen et al ., 2014 )。然而, 目前该方面研究的直接试验证据几乎还未见报道, 因此开展根系分泌物C:N化学计量特征对土壤C养分转化过程的影响成为一个十分重要但又极度缺乏的研究课题, 尤其在叠加不同土壤养分有效性的条件下。
西南亚高山针叶林(subalpine coniferous forest)是青藏高原东部高寒林区的重要组成部分, 对于维持我国西部地区区域生态安全和促进区域可持续发展具有十分重要的“生态屏障(ecological barrier)”作用(刘庆, 2002 )。随着该区土地利用方式和管理措施的巨大变化, 自20世纪初期该区森林被大规模采伐, 随后在皆伐迹地营造了大面积的亚高山人工针叶林, 成为该区森林的重要组成部分。前期研究发现, 亚高山天然针叶林和云杉人工林具有截然不同的土壤有机质和养分含量(Xu et al ., 2010 )。因此, 本研究以川西亚高山天然林和云杉人工林两种森林林型土壤为试验对象, 通过配制不同C:N化学计量特征的模拟根系分泌物溶液进行人工添加试验, 测定不同处理下两种土壤碳含量动态及其微生物特征变化, 以探究根系分泌物C:N化学计量特征对不同森林类型土壤碳循环过程的影响差异, 为深入揭示森林根系分泌物与土壤C养分循环过程的耦合机制提供理论支持。
1 材料和方法
1.1 研究区概况
试验样地位于四川省理县米亚罗自然保护区, 地理位置为103.21° E、31.40° N, 海拔2620 m。本区域属季风性山地气候, 夏季温凉多雨, 冬季寒冷干燥, 1月平均气温为-8 ℃, 7月平均气温为12.6 ℃, ≥10 ℃的年积温为1200-1400 ℃, 年降水量为600-1100 mm, 年蒸发量为1000-1900 mm。成土母岩主要为千枚岩、板岩和白云岩等残坡积风化物, 土壤主要为暗棕壤土。天然林主要乔木有云杉(Picea asperata )、鳞皮冷杉(Abies squamata )、川西云杉(Picea likiangensis var. balfouriana )和红桦(Betula albosinensis )等; 主要灌木有疏花槭(Acer laxiflorum )、红毛五加(Acanthopanax giraldii )、散生栒子(Cotoneaster divaricatus )、扁刺蔷薇(Rosa sweginzowii )、紫花卫茅(Euonymus porphyreus )和川滇高山栎(Quercus aquifolioides )等; 主要草本植物有钩柱唐松草(Thalictrum uncatum )、蛛毛蟹甲草(Cacalia roborowskii )、歪头菜(Vicia unijuga )、冷蕨(Cystopteris fragilis )、东方草莓(Fragaria orientalis )和禾本科植物等, 而研究的人工针叶林主要是在原始林采伐迹地上更新而成的云杉纯林, 林下基本无灌木层和草本层。
1.2 供试土壤采集
试验土壤来源于位于四川省理县米亚罗镇夹壁沟的天然针叶林(约160年)和云杉人工林(约75年), 两个林地相距约300 m。2013年10月20日在两个林分下各设3个随机样方, 按照“S”随机取样方法取0-20 cm深的土壤样品, 每个样方分别取5个重复土样, 然后将同一样方的5个土样充分混合, 再将3个样方的混合土样混合。用冰袋保存混合后的土样带回实验室, 过2 mm筛。每份样品分为两份, 一份用于土壤本底理化性质测定(表1 ), 另一份则用于模拟根系分泌物添加试验。
1.3 试验处理与分析方法
1.3.1 模拟根系分泌物的配制和添加
文献调研结果表明: 森林根系分泌物常见的低分子化合物主要包括糖类、氨基酸和有机酸。因此, 在本试验中我们分别选取葡萄糖、柠檬酸、谷氨酸3种物质进行不同组合, 并配制具有不同化学计量特征(即溶液中纯C:N质量比, 简称C:N)的模拟根系分泌物溶液, 并用NH4 Cl配制仅添加N (N-only)处理溶液, 共进行5种处理: N-only、C:N = 10、C:N = 50、C:N = 100和仅添加C (C-only), 并用去离子水作为对照处理(CK), 每种处理的化学组成和含量见表2 。基于我们前期的实验结果, 上述几种处理模拟根系分泌物溶液的纯C浓度均配制为60 mg·L-1 , 除N-only处理纯N浓度为6 mg·L-1 之外, 其他几种处理纯N浓度则根据其化学计量特征比而变化。为了保证配制溶液的质量和有效性, 每隔5天我们重新配制新鲜溶液进行添加处理。我们前期的预试验表明: 每日添加5 mL水能使土壤含水量与处理前土壤水分含量基本保持一致。人工添加试验从2013年10月27日开始, 每日各处理添加量为5 mL, 整个试验周期持续50天。每种处理设置3个重复, 每个重复分别取200 g试验土壤装入500 mL烧杯中, 每天溶液添加后用玻璃棒将试验土壤搅拌均匀, 所有烧杯置于室内环境条件下进行培养, 直至试验结束。
1.3.2 测定方法
土壤pH值采用电位法测定; 土壤含水量采用烘干法测定; 土壤总碳(TC)采用元素分析仪(MACRO cube, Elementar, Hanau, Germany)测定; 土壤活性和惰性碳库采用化学酸提法进行测定(Rovira & Vallejo, 2002 )。具体步骤如下: 称量过筛土壤0.5 g,加20 mL 2.5 mol·L-1 H2 SO4 溶液, 然后放在105 ℃下水解30 min, 离心, 把上清液转移到三角瓶中, 并用20 mL去离子水小心、缓慢地冲洗离心管, 上清液也加入三角瓶中, 这部分浸提液体中的土壤C含量被定义为活性碳库I (labile pool I carbon, LPI-C)。剩余土壤残渣用2 mL13 mol·L-1 H2 SO4 溶液在室温下(不停摇动)浸提12 h, 然后把浸提液用去离子水稀释到1 mol·L-1 , 在105 ℃下水解3 h, 这部分浸提液中的C含量被定义为活性碳库II (labile pool II carbon, LPII-C)。土壤惰性C库(recalcitrant pool carbon, RP-C)为土壤TC与活性C库(LPI-C与LPII-C之和)差值。土壤微生物生物量碳(MBC)和微生物生物量氮(MBN)采用氯仿熏蒸法测定。
土壤微生物活细胞生物量和群落结构采用磷脂脂肪酸(PLFA)方法测定, 它是一种快速、可靠的分析土壤微生物群落结构的方法(Bossio et al ., 1998 )。测定方法如下: 8 g鲜土经提取、层析、甲酯化后, 溶于正己烷的磷酯用氮气吹干, -20 ℃保存。样品用100 μL 0.025 mg·mL-1 酯化C 19:0溶液溶解, 并用气相色谱仪(Agilent 6850, Agilent, Santa Clara, USA)进行PLFA的鉴定和量化。其中色谱柱为Agilent 19091B-102E毛细管柱(25 m × 0.2 mm × 0.33 μm), 载气为H2 。进样量为2 μL, 分流比100: 1。进样口温度250 ℃, 检测器温度300 ℃。二阶升温程序如下: 起始温度170 ℃, 以5 ℃·min-1 升至260 ℃, 再以40 ℃·min-1 升至310 ℃。气相色谱全过程由MIDI软件(MIDI Microbial ID, Newark, USA)自动控制, 其中Sherlock 6.0 (MIDI Inc.)用于PLFA鉴定。用峰面积和内标曲线法定量测定PLFA, PLFA含量用nmol·g-1 表示。分别计算土壤总PLFA含量和细菌(Bacteria)、真菌(Fungi)、放线菌(Actinomycetes) PLFA含量, 以及表征微生物群落结构的重要指标细菌:真菌(bacteria:fungi)。
1.3.3 数据分析
利用SPSS 20.0软件进行统计分析, 分别对不同处理下(CK、N-only、C:N = 10、C:N = 50、C:N = 100和C-only)各土壤指标进行单因素方差分析(one-way ANOVA), 并采用Duncan法进行多重比较。所有图形均利用Microsoft Office Excel 2003软件绘制。
2 研究结果
2.1 不同模拟根系分泌物添加处理下两种林分土壤pH变化
研究结果表明: 模拟根系分泌物处理均显著增加了两种林分的土壤pH值。其中, 对于天然林而言, 根系分泌物C:N = 50添加处理下土壤pH值最大, 而人工林则以C:N = 100处理下土壤pH值最大。但需要说明的是, 虽然几种根系分泌模拟添加处理对土壤pH有显著性影响, 但各处理间土壤pH差异绝对值极小, 因而我们认为根系分泌物添加导致的pH变化对土壤生态过程的影响有限(表3 )。
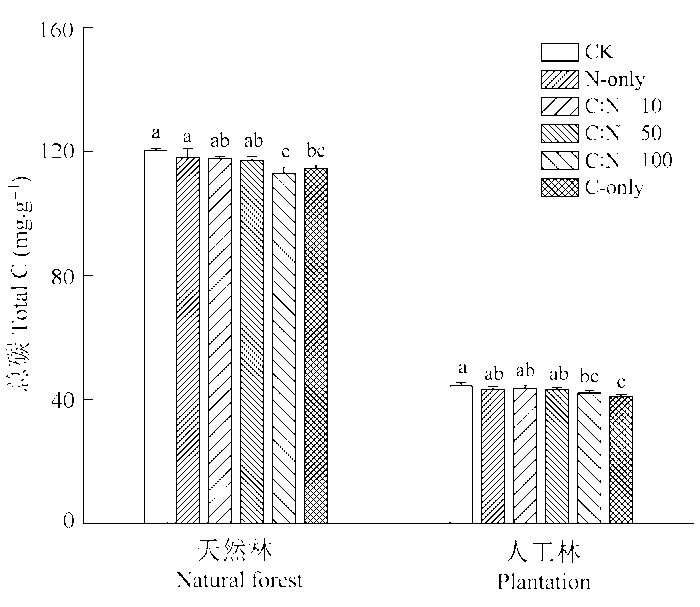
2.2 根系分泌物不同C:N化学计量特征对两种林分土壤TC的影响
天然林和人工林土壤TC含量对模拟根系分泌物添加处理表现出相似的响应趋势(图1 )。与对照相比, 几种根系分泌物添加处理均降低了两种林分土壤TC含量, 但仅C:N = 100和C-only两种处理与对照之间存在显著性差异, 而N-only、C:N = 10和C:N = 50三种添加处理下两种林分土壤TC含量与对照之间均无显著性差异, 表明根系分泌物N源输入在一定程度上缓和了外源C添加后所导致的两种林分土壤TC含量降低幅度。此外, 与对照相比, 根系分泌物C添加导致的人工林土壤TC含量的降低幅度比天然林更大, 其中C:N = 100和C-only两种处理导致天然林TC含量分别降低了6.35%和4.94%, 而使人工林土壤TC含量分别降低了10.05%和5.32% (图1 )。
图1 不同C:N化学计量特征的根系分泌物模拟添加对天然林和人工林土壤总碳含量的影响(平均值±标准偏差)。CK, 对照; C-only, 仅添加C; N-only, 仅添加N。不同小写字母表示同一林型各处理间在p < 0.05水平上差异显著。
Fig. 1 Effects of simulated root exudates under different C:N stoichiometry on total carbon content in natural forest and the plantation (mean ± SD ). CK, control. Different lowercase letters mean significant differences (p < 0.05) among treatments at a given forest type.
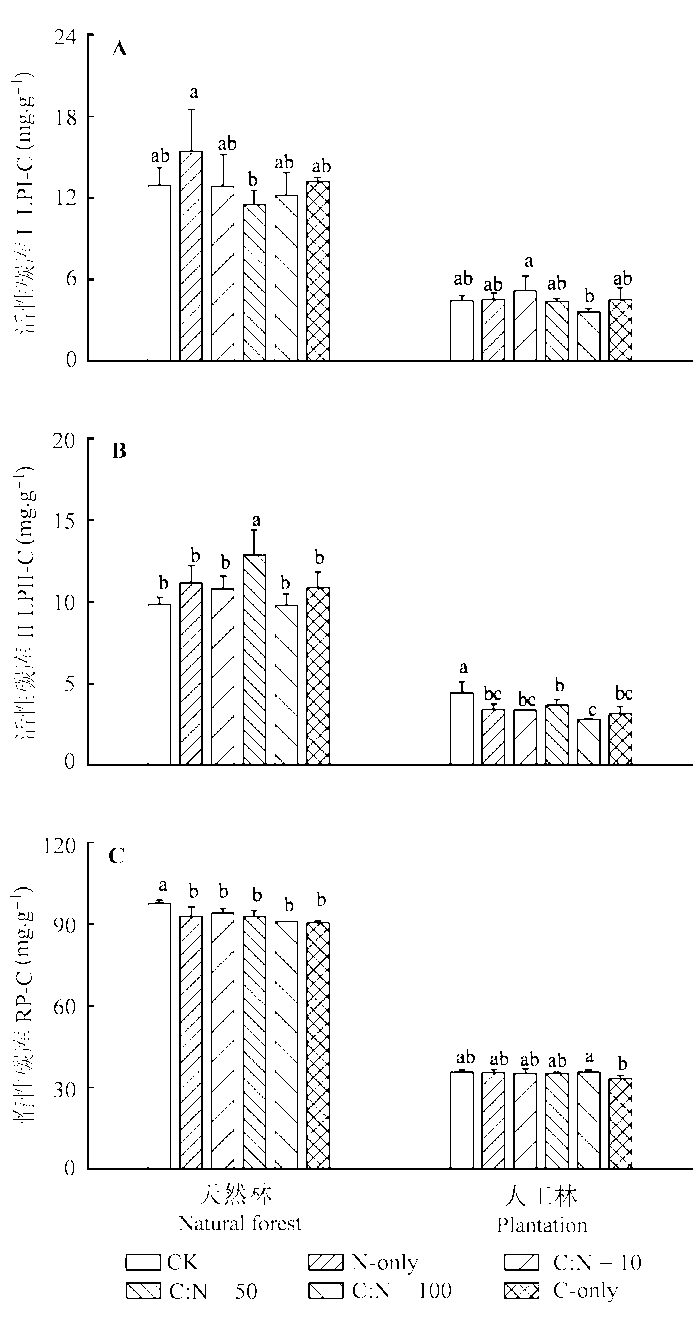
2.3 根系分泌物不同C:N化学计量特征对两种林分土壤活性和惰性碳库的影响
模拟根系分泌物添加处理对天然林和人工林土壤活性和惰性碳库的影响具有较大的差异, 且与土壤C库组分形式有关(图2 )。对LPI-C而言, 根系分泌物添加处理对两种林分土壤LPI-C的影响无明显规律。天然林土壤LPII-C对根系分泌物添加处理的响应表现出一致的增加趋势, 但仅C:N = 50处理与对照之间差异显著; 相反, N-only、C:N = 10、C:N = 50、C:N = 100和C-only 5种模拟分泌物添加处理则使人工林土壤LPII-C含量均显著降低, 分别下降了22.85%、24.25%、17.43%、36.84%和29.17% (图2 B)。N-only、C:N = 10、C:N = 50、C:N = 100和C-only 5种模拟分泌物添加处理使天然林土壤RP-C分别下降了4.93%、3.75%、5.01%、6.99%和7.41%, 而几种根系分泌物添加处理对人工林土壤RP-C则无显著影响(图2 C)。
图2 不同C:N化学计量特征的根系分泌物模拟添加对天然林和人工林土壤活性和惰性碳库的影响(平均值±标准偏差)。A, 活性碳库I。B, 活性碳库II。C, 惰性碳库。CK, 对照; C-only, 仅添加C; N-only, 仅添加N。不同小写字母表示同一林型各处理间在p < 0.05水平上差异显著。
Fig. 2 Effects of simulated root exudates under different C:N stoichiometry on soil labile and recalcitrant carbon pool in natural forest and the plantation (mean ± SD ). A, labile carbon pool I (LPI-C). B, labile carbon pool II (LPII-C). C, recalcitrant carbon pool (RP-C). CK, control. Lowercase letters mean significant differences (p < 0.05) among treatments at a given forest type.
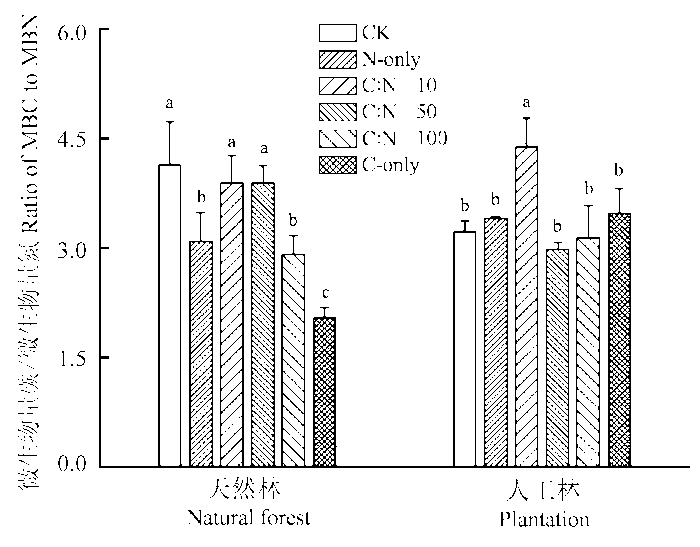
2.4 根系分泌物不同C:N化学计量特征对土壤微生物生物量碳氮比(MBC:MBN)的影响
两种林分MBC:MBN对根系分泌物添加处理总体表现出不同的响应趋势(图3 )。对天然林而言, 几种根系分泌物添加处理总体降低了土壤MBC:MBN值, 但仅N-only、C:N = 100和C-only三种处理与对照之间存在显著差异。对人工林而言, 与对照相比, 除C:N = 10处理使人工林土壤MBC:MBN比值显著增加外, 其他几种处理对土壤MBC:MBN则无明显影响(图3 )。
图3 不同C:N化学计量特征的根系分泌物添加对天然林和人工林不同C:N化学计量特征根系分泌物模拟溶液对天然林和人工林土壤微生物生物量碳氮比的影响(平均值±标准偏差)。CK, 对照; C-only, 仅添加C; N-only, 仅添加N。不同小写字母表示同一林型各处理间在p < 0.05水平上差异显著。
Fig. 3 Effects of simulated root exudates under different C:N stoichiometry on ratio of (microbial biomass C : microbial biomass N) MBC:MBN in natural forest and the plantation (mean ± SD ). CK, control. Lowercase letters mean significant differences (p < 0.05) among treatments at a given forest type.
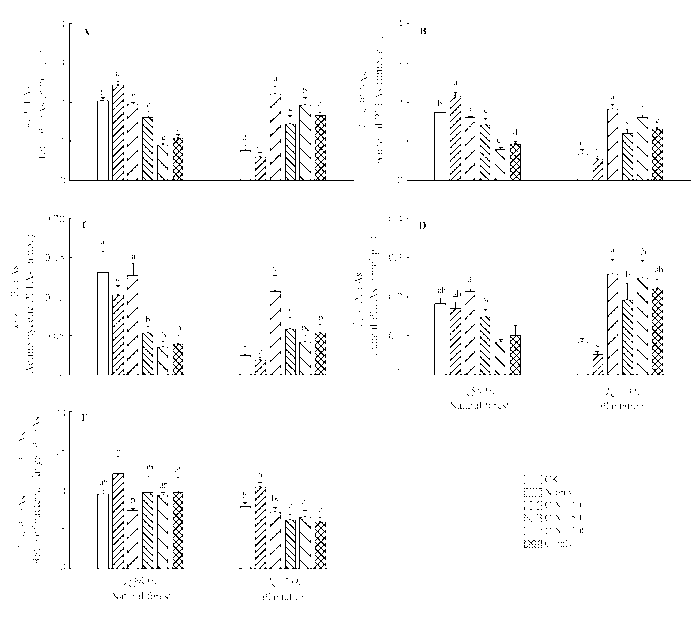
2.5 根系分泌物不同C:N化学计量特征对土壤微生物群落结构的影响
天然林和人工林土壤微生物群落结构对模拟根系分泌物添加处理总体表现出相反的响应趋势(图4 )。对于天然林而言, 除N-only处理导致土壤微生物PLFA总量显著高于对照之外, 其他几种处理下土壤微生物PLFA总量均低于对照, 但仅C:N = 50、C:N = 100和N-only三种处理与对照之间差异显著; 根系分泌物添加处理对人工林土壤微生物PLFA总量的影响趋势则与天然林刚好相反, 除N-only处理导致人工林土壤PLFA总量略低于对照外, 其他几种处理导致人工林土壤微生物PLFA总量显著高于对照(图4 A)。
类似地, 根系分泌物添加处理对两种林分土壤细菌、放线菌和真菌PLFA含量的影响基本与土壤微生物PLFA总量的变化趋势一致, 即与对照相比, 除N-only处理对上述三大类群微生物PLFA总量影响不明显外, 其他几种处理导致天然林土壤细菌、放线菌和真菌PLFA含量显著降低, 而使人工林三大类群微生物PLFA含量显著增加(图4B -4D)。
模拟根系分泌物添加处理对两种林分土壤细菌:真菌相对优势度(bacterial:fungal dominance)的影响也具有较大差异。N-only处理使得两种土壤细菌:真菌比值明显高于对照, 但仅人工林表现出显著性差异(图4E ), 表明N添加降低了两种林分土壤真菌的相对丰度。C添加对人工林土壤细菌:真菌比值的影响效应则与N-only处理刚好相反, 除C:N = 10处理与对照差异不显著之外, 其他3种含C分泌物添加处理(C:N = 50、C:N = 100和C-only)均导致人工林土壤细菌:真菌比值显著低于对照。然而, 对于天然林而言, 除C:N = 50处理使得土壤细菌:真菌比值显著低于对照之外, 其他几种含碳化合物添加处理对天然林土壤细菌:真菌比值影响均不明显(图4E )。
图4 不同C:N化学计量特征的根系分泌物添加对天然林和人工林土壤微生物磷酸脂肪酸(PLFA)含量的影响(平均值±标准偏差)。A, 微生物总PLFA。B, 细菌PLFA。C, 放线菌PLFA。D, 真菌PLFA。E, 细菌PLFA/真菌PLFA。CK, 对照; C-only, 仅添加C; N-only, 仅添加N。不同小写字母表示同一林型各处理间在p < 0.05水平上差异显著。
Fig. 4 Effects of simulated root exudates under different C:N stoichiometry on contents of soil microbial phospholipid fatty acid (PLFA) in natural forest and the plantation (mean ± SD ). A, total PLFA in microbia. B, bacterial PLFA. C, actinomycetic PLFA. D, fungal PLFA. E, ratio of bacterial/fungal PLFA. CK, control. Lowercase letters mean significant differences (p < 0.05) among treatments at a given forest type.
3 讨论
3.1 两种林分土壤C过程对模拟根系分泌物C:N化学计量特征的响应差异
土壤是陆地生态系统最大的有机碳库, 外源活性C添加通过土壤激发效应而引起土壤有机质矿化分解过程在短期内发生强烈的改变(Fontaine et al ., 2004 ), 并影响生态系统多个养分循环过程。此外, 由于土壤C-N转化过程密切的偶联效应, 土壤有机质分解和C库动态受根系N源输入以及土壤N养分状况控制。然而, 目前关于N养分有效性对土壤有机质分解的影响效应还未取得一致性的结论, 主要存在截然不同的两种假说: (1)微生物氮矿化(microbial nitrogen mining)假说。该假说认为土壤获N (N-acquiring)微生物通常利用活性外源C作为能源提高其生长和活性, 进而刺激土壤惰性有机质分解和养分转化过程, 以获取微生物生长所需各种养分(Moorhead & Sinsabaugh, 2006 ; 孙悦等, 2014 ); (2)化学计量分解理论(stoichiometric decomposition)假说, 根据该理论假说, 当外源C和N输入与土壤微生物C:N化学计量特征高度匹配时, 土壤微生物才具有最高的活性, 此时土壤有机质分解速率最大(Hessen et al ., 2004 ; Chen et al ., 2014 )。上述两种假说赋予了N养分有效性对土壤有机质稳定性相反的影响效应。第一种假说表明N受限条件(nutrient-poor condition)更有利于土壤有机质分解, 而第二种假说意味着N富集条件(nutrient-rich condition)更有利于土壤有机质分解(Chen et al ., 2014 )。
在本研究中, C-only和C:N = 100 (后一种处理N浓度仅为0.6 mg·L-1 , 因此其N含量基本可忽略)两种处理显著降低了两种林分土壤TC含量, 而适量的N添加(N-only、C:N = 50和C:N = 10)则在一定程度上缓和了土壤TC含量的降低(图1 )。土壤N限制是西南亚高山针叶林主要特征之一(Xu et al ., 2010 ), 人工林更是如此(表1 ), 根系分泌物外源C添加进一步诱导了该区两种林分土壤N限制, 土壤微生物利用输入C源并刺激微生物生长和活性, 导致土壤有效N被快速消耗(Bengtson et al ., 2012 ; Burns et al ., 2013 )。低的有效N促使微生物分泌更多的胞外酶以加速土壤有机质分解, 释放无机N来满足微生物生长所需的养分, 从而促进土壤有机质分解速率和CO2 释放, 使两种林分土壤TC含量显著降低。然而, 添加一定量的N组分在一定程度上缓和了两种林分土壤N养分限制, 使得微生物不需要通过分解难以利用的土壤有机质获取N, 而是偏好利用易于分解的有机化合物, 相应地减少土壤胞外酶的分泌, 从而降低土壤有机质分解和CO2 释放, 并缓和了两种针叶林土壤TC含量的降低幅度。上述研究结果极好地印证了前人所提出的N矿化假说理论, 即N养分有效性受限会促进土壤有机质分解。
此外, 研究结果表明模拟根系分泌物添加处理所导致的人工林土壤TC含量降低幅度总体比天然林大(图1 ), 我们推测这主要与两种针叶林各自不同的土壤化学特性(土壤总N和C:N)有关。对于两种林分土壤而言, 人工林土壤总N含量远低于天然林(表1 ), 而人工林土壤C:N明显高于天然林土壤C:N (表1 ), 表明人工林土壤N养分限制程度比天然林更为严重。因此, 相对于天然林土壤而言, 人工林土壤微生物更大的养分需求导致活性C源输入后对其土壤有机质分解所诱导的激发效应更强, 相应地增加了人工林土壤有机质矿化速率和CO2 排放, 从而使得人工林土壤TC含量降低幅度比天然林明显。Zhang等(2013)通过外源C添加对土壤有机质分解速率影响的元分析(meta-analysis)也表明, 外源C添加对那些具有低总N含量或高C:N特性的土壤类型所诱导的有机质分解激发效应更强, 这与我们的研究结果高度一致。需要说明的是, 在本研究中, 虽然不同根系分泌物模拟处理对两种林分土壤活性碳库(LPI-C, LPII-C)和惰性碳库(RP-C)具有一定影响, 但各C组分在根系分泌物添加处理之间和林分之间并无明显规律; 加之我们在研究中并没有同步监测与不同C组分动态相关的特异性微生物代谢酶活性变化, 因此无法阐明两种林分土壤C组分在各处理之间的变化规律及其潜在机制, 这需要我们进一步深入研究。
3.2 模拟根系分泌C:N化学计量特征对两种林分土壤微生物群落结构的影响
土壤有机质矿化及其伴随的碳库动态变化实质上是土壤微生物综合作用的结果, 而微生物组成和群落结构变化将深刻影响土壤生物地球化学循环过程(Schmidt et al ., 2011 )。在本研究中, 虽然模拟根系分泌物添加总体降低了天然林和人工林土壤TC含量, 但两种林分土壤对模拟根系分泌物C:N化学计量特征表现出相反的微生物反馈机制(图4 )。除N-only处理之外, 其他几种根系分泌物处理导致人工林土壤微生物PLFA总量和细菌、放线菌、真菌PLFA含量明显增加, 并伴随着更强的土壤有机质(SOM)分解激发效应和TC降低。这与Blagodatskaya 和Kuzyakov (2008)的报道一致, 其研究结果表明土壤微生物生物量大小调控外源活性C添加对土壤激发效应的幅度和方向。与人工林相反, 根系分泌物C源添加处理导致天然林土壤微生物总PLFA含量和细菌、放线菌、真菌PLFA含量明显下降, 这与C源添加后土壤TC含量下降这一结果并不吻合, 我们推测这可能与几种处理下土壤微生物生物量形成和胞外酶分配权衡(Fontaine et al ., 2011 )或特异性微生物代谢酶活性增强有关(de Graaff et al ., 2010)。但遗憾的是, 由于试验设计限制, 在本研究中我们并没有同步监测与土壤C变化相关的微生物代谢酶活性变化, 使得无法准确提供不同处理下土壤碳动态变化的机理性证据和解释。
此外, 根系分泌物处理诱导两种林分土壤微生物群落结构也产生了不同的改变。虽然两种林分土壤微生物群落结构变化在C:N = 10、C:N = 50、C:N = 100和C-only几种处理之间无明显差异, 但通过PLFA分析结果总体表明N添加倾向增加两种林分土壤细菌:真菌比值, 而C添加显著降低人工林土壤细菌:真菌比值, 但对天然林土壤细菌:真菌的影响并不明显。土壤微生物群落结构及其细菌:真菌相对优势度改变深刻地影响和调控土壤C养分循环过程(Strickland & Rousk, 2010 )。目前有关根系分泌物输入对森林土壤微生物群落结构影响的具体机理并不清楚, 基于文献调研和本文的试验结果, 我们提出如下两种假说解释不同化学计量特征根系分泌物处理后人工林土壤微生物群落结构变化的原因。
(1)微生物竞争性假说。正如前面所讨论的, 不同C:N化学计量特征根系分泌物添加所导致的微生物养分需求大小变化是诱导人工林土壤微生物群落结构(细菌/真菌相对优势度)改变的主要原因之一。在本研究中, 除N-only处理之外, 其他几种模拟处理均不同程度地进一步加剧了人工林土壤N限制, 更强的微生物N需求迫使微生物去矿化惰性有机质获取所需要的养分, 并改变微生物活性和微生物群落结构组成, 结果导致那些主要以分解惰性有机质的慢生长k策略功能微生物群(主要为真菌)获得更强的竞争性优势(Fontaine et al ., 2003 ; Ramirez et al ., 2012 ), 从而降低土壤细菌:真菌比值; 相反地, N添加处理则在一定程度上缓和了人工林土壤N限制程度, 从而减轻了微生物通过矿化惰性有机质库以获取N素的需求度(Craine et al ., 2007 ), 并使得那些以分解活性有机质的快生长r策略功能微生物群(主要为细菌)获得更强的竞争性优势(Dorodnikov et al ., 2009 ; Chen et al ., 2014 ), 从而导致N-only处理下人工林土壤细菌:真菌比值显著高于其他几种处理。
(2)细菌和真菌化学计量特征差异。导致人工林土壤细菌:真菌比值不同变化的另外一个原因可能与细菌和真菌本身微生物化学计量特征差异有关。研究表明, 真菌微生物量C:N通常高于细菌, 前者C:N一般介于5-15之间, 而后者C:N大多在3-6之间(McGill & Cole, 1981 ), 这表明真菌对养分需求比细菌低(Güsewell & Gessner, 2009 )。在本研究中, 除N-only之外, 其他几种模拟处理其C:N均明显高于细菌C:N比值, 相应地, 使得土壤微生物真菌相对于细菌而言获得更大的竞争性优势, 从而明显降低土壤细菌:真菌比值; 相反, N添加处理则在一定程度上缓和了人工林土壤N限制乃至土壤变为C限制, 土壤C:N降低, 相应地导致土壤细菌优势度更明显, 从而使得N-only处理下人工林土壤细菌:真菌显著高于其他几种处理(Carney et al ., 2007 )。此外, 土壤MBC:MBN在一定程度上是表征土壤细菌:真菌相对丰度的重要参数, 其中MBC:MBN比值越大, 表明真菌相对比例越高, 土壤细菌:真菌比值则越小(Zhao et al ., 2010 )。然而, 在本研究中, 我们发现基于土壤MBC:MBN比值所反馈的两种林分微生物群落结构变化趋势与通过PLFA方法所获得的微生物群落结构变化趋势并不完全一致。例如, N-only、C:N = 100和C-only几种处理导致天然林土壤MBC:MBN比值显著降低, 从而增加了土壤细菌:真菌比值; 而人工林除C:N = 10之外, 其他几种添加处理对土壤MBC:MBN无明显影响, 具体原因还有待进一步深入探究。虽然两种方法表征的土壤微生物群落结构变化信息并不完全吻合, 但二者从不同角度均强有力地证实了根系分泌物添加处理诱导天然林和人工林微生物群落结构发生了截然不同的改变。
3.3 结论
根系分泌物处理C添加促进了两种林分土壤激发效应而降低了土壤TC含量, 而同时添加N在一定程度上缓和了两种林分土壤TC含量的降低幅度, 且C添加处理导致的含有较高有效性N的天然林土壤TC含量降低幅度明显低于土壤N限制更严重的人工林, 表明根系N源输入和土壤N养分有效性共同调控根系活性C源输入对土壤过程和功能的生态学效应。此外, 人工林和天然林土壤微生物群落结构变化对模拟根系分泌物添加也表现出不同的微生物反馈机制, 可能与两种林分土壤化学特性不同有关。但需要说明的是, 我们必须承认试验本身有一定的局限性。比如, 在本研究中为了区别外源C和N添加对土壤过程的影响差异, 溶液配制时我们利用NH4 Cl来调节不同处理中N的相对浓度, 这与根系分泌物N组分输入通常以氨基酸形式而非NH4 + 形式输入不相符; 同时, 我们在根系分泌物中常见的三大类低分子化合物(氨基酸、有机酸和糖类)中各选择了一种物质进行模拟配制, 加之本研究中采用每天一次性的溶液添加方式, 这与野外森林根系分泌物实际输入状况(即持续渗入到根际土壤)有所差异, 这些因素都可能对本文研究结果产生潜在的影响。此外, 由于本研究中模拟根系分泌物添加处理时间仅持续50天, 我们不能确定所获得的结果是土壤的短期反应还是长期持续的真实状况, 因而在用本文研究结果解释和外推森林根系分泌物化学计量特征与土壤C循环过程的偶联机制时, 必须足够谨慎。这需要我们进一步深入开展根系分泌物对土壤生物地球化学过程的长期野外原位研究和数据积累, 并在未来实验研究和C循环模型构建中充分重视N素对土壤C激发效应的调控作用。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Bengtson P Barker J Grayston SJ 2012 ). Evidence of a strong coupling between root exudation, C and N availability, and stimulated SOM decomposition caused by rhizosphere priming effects.
Ecology and Evolution , 2 , 1843 -1852 .
[本文引用: 1]
[2]
Blagodatskaya Е Kuzyakov Y 2008 ). Mechanisms of real and apparent priming effects and their dependence on soil microbial biomass and community structure: Critical review.
Biology and Fertility of Soils , 45 , 115 -131 .
[3]
Bossio DA Scow KM Gunapala N Graham KJ 1998 ). Determinants of soil microbial communities: Effects of agricultural management, season, and soil type on phospholipid fatty acid profiles.
Microbial Ecology , 36 , 1 -12 .
[本文引用: 1]
[4]
Burns RG DeForest JL Marxsen J Sinsabaugh RL Stromberger ME Wallenstein MD Weintraub MN Zoppini A 2013 ). Soil enzymes in a changing environment: Current knowledge and future directions.
Soil Biology & Biochemistry , 58 , 216 -234 .
[本文引用: 1]
[5]
Carney KM Hungate BA Drake BG Megonigal JP 2007 ). Altered soil microbial community at elevated CO2 leads to loss of soil carbon.
Proceedings of the National Academy of Sciences of the United States of America , 104 , 4990 -4995 .
[本文引用: 1]
[6]
Chapin FS III McFarland J David McGuire A Euskirchen ES Ruess RW Kielland K 2009 ). The changing global carbon cycle: Linking plant-soil carbon dynamics to global consequences.
Journal of Ecology , 97 , 840 -850 .
[本文引用: 1]
[7]
Chen RR Senbayram M Blagodatsky S Myachina O Dittert K Lin XG Blagodatskaya E Kuzyakov Y 2014 ). Soil C and N availability determine the priming effect: Microbial N mining and stoichiometric decomposition theories.
Global Change Biology , 20 , 2356 -2367 .
[本文引用: 4]
[8]
Cheng WX Parton WJ Gonzalez-Meler MA Phillips R Asao S McNickle GG Brzostek E Jastrow JD 2014 ). Synthesis and modeling perspectives of rhizosphere priming.
New Phytologist , 201 , 31 -44 .
[本文引用: 2]
[9]
Cleveland CC Liptzin D 2007 ). C:N:P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass?
Biogeochemistry , 85 , 235 -252 .
[本文引用: 1]
[10]
Craine JM Morrow C Fierer N 2007 ). Microbial nitrogen limitation increases decomposition.
Ecology , 88 , 2105 -2113 .
[本文引用: 1]
[11]
de Graaff MA Classen AT Castro HF Schadt CW 2010 ). Labile soil carbon inputs mediate the soil microbial community composition and plant residue decomposition rates.
New Phytologist , 188 , 1055 -1064 .
[12]
Dijkstra FA Cheng WX 2007 ). Interactions between soil and tree roots accelerate long-term soil carbon decomposition.
Ecology Letters , 10 , 1046 -1053 .
[本文引用: 1]
[13]
Dorodnikov M Blagodatskaya E Blagodatsky S Marhan S Fangmeier A Kuzyakov Y 2009 ). Stimulation of microbial extracellular enzyme activities by elevated CO2 depends on soil aggregate size.
Global Change Biology , 15 , 1603 -1614 .
[本文引用: 1]
[14]
Drake J Darby B Giasson MA Kramer M Phillips R Finzi A 2013 ). Stoichiometry constrains microbial response to root exudation-insights from a model and a field experiment in a temperate forest.
Biogeosciences , 10 , 821 -838 .
[本文引用: 1]
[15]
Espinosa-Urgel M Ramos JL 2001 ). Expression of a Pseudomonas putida aminotransferase involved in lysine catabolism is induced in the rhizosphere.
Applied and Environmental Microbiology , 67 , 5219 -5224 .
[本文引用: 1]
[16]
Fontaine S Bardoux G Abbadie L Mariotti A 2004 ). Carbon input to soil may decrease soil carbon content.
Ecology Letters , 7 , 314 -320 .
[本文引用: 1]
[17]
Fontaine S Henault C Aamor A Bdioui N Bloor JMG Maire V Mary B Revaillot S Maron PA 2011 ). Fungi mediate long term sequestration of carbon and nitrogen in soil through their priming effect.
Soil Biology & Biochemistry , 43 , 86 -96 .
[本文引用: 1]
[18]
Fontaine S Mariotti A Abbadie L 2003 ). The priming effect of organic matter: A question of microbial competition?
Soil Biology & Biochemistry , 35 , 837 -843 .
[本文引用: 1]
[19]
Fransson P Johansson EM 2010 ). Elevated CO2 and nitrogen influence exudation of soluble organic compounds by ectomycorrhizal root systems.
FEMS Microbiology Ecology , 71 , 186 -196 .
[本文引用: 1]
[20]
Güsewell S Gessner MO 2009 ). N:P ratios influence litter decomposition and colonization by fungi and bacteria in microcosms.
Functional Ecology , 23 , 211 -219 .
[本文引用: 1]
[21]
Grayston SJ Vaughan D Jones D 1997 ). Rhizosphere carbon flow in trees, in comparison with annual plants: The importance of root exudation and its impact on microbial activity and nutrient availability.
Applied Soil Ecology , 5 , 29 -56 .
[本文引用: 1]
[22]
Hessen DO Ågren GI Anderson TR Elser JJ de Ruiter PC 2004 ). Carbon sequestration in ecosystems: The role of stoichiometry.
Ecology , 85 , 1179 -1192 .
[本文引用: 1]
[23]
Jones DL Hodge A Kuzyakov Y 2004 ). Plant and mycorrhizal regulation of rhizodeposition.
New Phytologist , 163 , 459 -480 .
[本文引用: 1]
[24]
Kuzyakov Y 2002 ). Review: Factors affecting rhizosphere priming effects.
Journal of Plant Nutrition and Soil Science , 165 , 382 -396 .
[本文引用: 1]
[25]
Liu Q 2002 ). Ecological Research on Subalpine Coniferous Forests in China . Sichuan University Press , Chengdu . 142 .(in Chinese)
[本文引用: 1]
[刘庆 (2002 ). 亚高山针叶林生态学研究 . 四川大学出版社 , 成都 . 142 .]
[本文引用: 1]
[26]
McGill WB Cole CV 1981 ). Comparative aspects of cycling of organic C, N, S and P through soil organic matter.
Geoderma , 26 , 267 -286 .
[本文引用: 1]
[27]
Moorhead DL Sinsabaugh RL 2006 ). A theoretical model of litter decay and microbial interaction.
Ecological Monographs , 76 , 151 -174 .
[本文引用: 1]
[28]
Phillips RP Bernhardt ES Schlesinger WH 2009 ). Elevated CO2 increases root exudation from loblolly pine (Pinus taeda) seedlings as an N-mediated response.
Tree Physiology , 29 , 1513 -1523 .
[本文引用: 1]
[29]
Phillips RP Finzi AC Bernhardt ES 2011 ). Enhanced root exudation induces microbial feedbacks to N cycling in a pine forest under long-term CO2 fumigation.
Ecology Letters , 14 , 187 -194 .
[本文引用: 1]
[30]
Ramirez KS Craine JM Fierer N 2012 ). Consistent effects of nitrogen amendments on soil microbial communities and processes across biomes.
Global Change Biology , 18 , 1918 -1927 .
[本文引用: 1]
[31]
Rovira P Vallejo VR 2002 ). Labile and recalcitrant pools of carbon and nitrogen in organic matter decomposing at different depths in soil: An acid hydrolysis approach.
Geoderma , 107 , 109 -141 .
[本文引用: 1]
[32]
Schmidt MW Torn MS Abiven S Dittmar T Guggenberger G Janssens IA Kleber M Kögel-Knabner I Lehmann J Manning DA Nannipieri P Rasse DP Weiner S Trumbore SE 2011 ). Persistence of soil organic matter as an ecosystem property.
Nature , 478 , 49 -56 .
[本文引用: 1]
[33]
Shi SJ Condron L Larsen S Richardson AE Jones E Jiao J O’Callaghan M Stewart A 2011 ). In situ sampling of low molecular weight organic anions from rhizosphere of radiata pine (Pinus radiata) grown in a rhizotron system.
Environmental and Experimental Botany , 70 , 131 -142 .
[本文引用: 1]
[34]
Strickland MS Rousk J 2010 ). Considering fungal: Bacterial dominance in soils-Methods, controls, and ecosystem implications.
Soil Biology & Biochemistry , 42 , 1385 -1395 .
[本文引用: 1]
[35]
Sullivan BW Hart SC 2013 ). Evaluation of mechanisms controlling the priming of soil carbon along a substrate age gradient.
Soil Biology & Biochemistry , 58 , 293 -301 .
[本文引用: 1]
[36]
Sun Y Xu XL Kuzyakov Y 2014 ). Mechanisms of rhizos- phere priming effects and their ecological significance.
Chinese Journal of Plant Ecology , 38 , 62 -75 .(in Chinese with English abstract)
[本文引用: 1]
[孙悦 , 徐兴良 , Kuzyakov Y 2014 ). 根际激发效应的发生机制及其生态重要性
. 植物生态学报 , 38 , 62 -75 .]
[本文引用: 1]
[37]
Wu LK Lin XM Lin WX 2014 ). Advances and perspective in research on plant-soil-microbe interactions mediated by root exudates.
Chinese Journal of Plant Ecology , 38 , 298 -310 .(in Chinese with English abstract)
[本文引用: 1]
[吴林坤 , 林向民 , 林文雄 (2014 ). 根系分泌物介导下植物-土壤-微生物互作关系研究进展与展望
. 植物生态学报 , 38 , 298 -310 .]
[本文引用: 1]
[38]
Xu ZF Wan C Xiong P Tang Z Hu R Cao G Liu Q 2010 ). Initial responses of soil CO2 efflux and C, N pools to experimental warming in two contrasting forest ecosys- tems, Eastern Tibetan Plateau, China.
Plant and Soil , 336 , 183 -195 .
[本文引用: 2]
[39]
Yin HJ Li YF Xiao J Xu ZF Cheng XY Liu Q 2013 ). Enhanced root exudation stimulates soil nitrogen transformations in a subalpine coniferous forest under experimental warming.
Global Change Biology , 19 , 2158 -2167 .
[本文引用: 1]
[40]
Yin HJ Wheeler E Phillips RP 2014 ). Root-induced changes in nutrient cycling in forests depend on exudation rates.
Soil Biology & Biochemistry , 78 , 213 -221 .
[本文引用: 1]
[41]
Zhang WD Wang XF Wang SL 2013 ). Addition of external organic carbon and native soil organic carbon decomposi- tion: A meta-analysis.
PLoS ONE , 8 , e54779 .
[42]
Zhao Q Zeng DH Fan ZP 2010 ). Nitrogen and phosphorus transformations in the rhizospheres of three tree species in a nutrient-poor sandy soil.
Applied Soil Ecology , 46 , 341 -346 .
[本文引用: 1]
Evidence of a strong coupling between root exudation, C and N availability, and stimulated SOM decomposition caused by rhizosphere priming effects.
1
2012
... 在本研究中, C-only和C:N = 100 (后一种处理N浓度仅为0.6 mg·L-1 , 因此其N含量基本可忽略)两种处理显著降低了两种林分土壤TC含量, 而适量的N添加(N-only、C:N = 50和C:N = 10)则在一定程度上缓和了土壤TC含量的降低(图1 ).土壤N限制是西南亚高山针叶林主要特征之一(Xu et al ., 2010 ), 人工林更是如此(表1 ), 根系分泌物外源C添加进一步诱导了该区两种林分土壤N限制, 土壤微生物利用输入C源并刺激微生物生长和活性, 导致土壤有效N被快速消耗(Bengtson et al ., 2012 ; Burns et al ., 2013 ).低的有效N促使微生物分泌更多的胞外酶以加速土壤有机质分解, 释放无机N来满足微生物生长所需的养分, 从而促进土壤有机质分解速率和CO2 释放, 使两种林分土壤TC含量显著降低.然而, 添加一定量的N组分在一定程度上缓和了两种林分土壤N养分限制, 使得微生物不需要通过分解难以利用的土壤有机质获取N, 而是偏好利用易于分解的有机化合物, 相应地减少土壤胞外酶的分泌, 从而降低土壤有机质分解和CO2 释放, 并缓和了两种针叶林土壤TC含量的降低幅度.上述研究结果极好地印证了前人所提出的N矿化假说理论, 即N养分有效性受限会促进土壤有机质分解. ...
Mechanisms of real and apparent priming effects and their dependence on soil microbial biomass and community structure: Critical review.
0
2008
Determinants of soil microbial communities: Effects of agricultural management, season, and soil type on phospholipid fatty acid profiles.
1
1998
... 土壤微生物活细胞生物量和群落结构采用磷脂脂肪酸(PLFA)方法测定, 它是一种快速、可靠的分析土壤微生物群落结构的方法(Bossio et al ., 1998 ).测定方法如下: 8 g鲜土经提取、层析、甲酯化后, 溶于正己烷的磷酯用氮气吹干, -20 ℃保存.样品用100 μL 0.025 mg·mL-1 酯化C 19:0溶液溶解, 并用气相色谱仪(Agilent 6850, Agilent, Santa Clara, USA)进行PLFA的鉴定和量化.其中色谱柱为Agilent 19091B-102E毛细管柱(25 m × 0.2 mm × 0.33 μm), 载气为H2 .进样量为2 μL, 分流比100: 1.进样口温度250 ℃, 检测器温度300 ℃.二阶升温程序如下: 起始温度170 ℃, 以5 ℃·min-1 升至260 ℃, 再以40 ℃·min-1 升至310 ℃.气相色谱全过程由MIDI软件(MIDI Microbial ID, Newark, USA)自动控制, 其中Sherlock 6.0 (MIDI Inc.)用于PLFA鉴定.用峰面积和内标曲线法定量测定PLFA, PLFA含量用nmol·g-1 表示.分别计算土壤总PLFA含量和细菌(Bacteria)、真菌(Fungi)、放线菌(Actinomycetes) PLFA含量, 以及表征微生物群落结构的重要指标细菌:真菌(bacteria:fungi). ...
Soil enzymes in a changing environment: Current knowledge and future directions.
1
2013
... 在本研究中, C-only和C:N = 100 (后一种处理N浓度仅为0.6 mg·L-1 , 因此其N含量基本可忽略)两种处理显著降低了两种林分土壤TC含量, 而适量的N添加(N-only、C:N = 50和C:N = 10)则在一定程度上缓和了土壤TC含量的降低(图1 ).土壤N限制是西南亚高山针叶林主要特征之一(Xu et al ., 2010 ), 人工林更是如此(表1 ), 根系分泌物外源C添加进一步诱导了该区两种林分土壤N限制, 土壤微生物利用输入C源并刺激微生物生长和活性, 导致土壤有效N被快速消耗(Bengtson et al ., 2012 ; Burns et al ., 2013 ).低的有效N促使微生物分泌更多的胞外酶以加速土壤有机质分解, 释放无机N来满足微生物生长所需的养分, 从而促进土壤有机质分解速率和CO2 释放, 使两种林分土壤TC含量显著降低.然而, 添加一定量的N组分在一定程度上缓和了两种林分土壤N养分限制, 使得微生物不需要通过分解难以利用的土壤有机质获取N, 而是偏好利用易于分解的有机化合物, 相应地减少土壤胞外酶的分泌, 从而降低土壤有机质分解和CO2 释放, 并缓和了两种针叶林土壤TC含量的降低幅度.上述研究结果极好地印证了前人所提出的N矿化假说理论, 即N养分有效性受限会促进土壤有机质分解. ...
Altered soil microbial community at elevated CO2 leads to loss of soil carbon.
1
2007
... (2)细菌和真菌化学计量特征差异.导致人工林土壤细菌:真菌比值不同变化的另外一个原因可能与细菌和真菌本身微生物化学计量特征差异有关.研究表明, 真菌微生物量C:N通常高于细菌, 前者C:N一般介于5-15之间, 而后者C:N大多在3-6之间(McGill & Cole, 1981 ), 这表明真菌对养分需求比细菌低(Güsewell & Gessner, 2009 ).在本研究中, 除N-only之外, 其他几种模拟处理其C:N均明显高于细菌C:N比值, 相应地, 使得土壤微生物真菌相对于细菌而言获得更大的竞争性优势, 从而明显降低土壤细菌:真菌比值; 相反, N添加处理则在一定程度上缓和了人工林土壤N限制乃至土壤变为C限制, 土壤C:N降低, 相应地导致土壤细菌优势度更明显, 从而使得N-only处理下人工林土壤细菌:真菌显著高于其他几种处理(Carney et al ., 2007 ).此外, 土壤MBC:MBN在一定程度上是表征土壤细菌:真菌相对丰度的重要参数, 其中MBC:MBN比值越大, 表明真菌相对比例越高, 土壤细菌:真菌比值则越小(Zhao et al ., 2010 ).然而, 在本研究中, 我们发现基于土壤MBC:MBN比值所反馈的两种林分微生物群落结构变化趋势与通过PLFA方法所获得的微生物群落结构变化趋势并不完全一致.例如, N-only、C:N = 100和C-only几种处理导致天然林土壤MBC:MBN比值显著降低, 从而增加了土壤细菌:真菌比值; 而人工林除C:N = 10之外, 其他几种添加处理对土壤MBC:MBN无明显影响, 具体原因还有待进一步深入探究.虽然两种方法表征的土壤微生物群落结构变化信息并不完全吻合, 但二者从不同角度均强有力地证实了根系分泌物添加处理诱导天然林和人工林微生物群落结构发生了截然不同的改变. ...
The changing global carbon cycle: Linking plant-soil carbon dynamics to global consequences.
1
2009
... 近年来, 随着对森林根系活动和根际过程研究的不断深入, 根系在调控土壤功能和养分代谢过程中的重要作用逐渐被人们所认知和关注(Chapin et al ., 2009 ; Cheng et al ., 2014 ).植物除了通过细根周转向土壤输入碳(C)和养分外, 还可通过根系主动或被动地向周围土壤释放一系列化合物, 即狭义的根系分泌物, 主要包括一些低分子(有机酸、糖类、酚类和各种氨基酸等)和高分子(蛋白质、黏液等)有机化合物(Dijkstra & Cheng, 2007 ).研究表明, 通常森林根系分泌物量占植物光合同化产物的1%-5% (Grayston et al ., 1997 ), 并受森林类型、树种、环境条件以及土壤养分有效性等诸多因素影响Fransson & Johansson, 2010 ; Shi et al ., 2011 ).由于根系分泌物大多为一些易于土壤微生物直接利用的含碳有机物, 可为土壤微生物系统提供重要且丰富的C源和氮(N)源, 有效地改变土壤微生物数量和活性, 而深刻地影响根际土壤有机质分解和养分代谢等过程(Phillips et al ., 2009 ; Yin et al ., 2013 ), 从而导致根系分泌物在调控土壤C养分转化过程中发挥与其含量不成比例的重要作用和功能.植物根系分泌物输入及其介导的根系-土壤-微生物互作效应已成为陆地生态系统碳循环过程研究中一个非常重要但又最缺乏了解的关键环节(Jones et al ., 2004 ; 吴林坤等, 2014 ). ...
Soil C and N availability determine the priming effect: Microbial N mining and stoichiometric decomposition theories.
4
2014
... 尽管当前对根系分泌物及其诱导的生态学效应研究已取得一定的进展, 但是对根系分泌物在森林根际生态过程中的作用及调控机理等方面我们依然缺乏足够的了解(Phillips et al ., 2011 ; Cheng et al ., 2014 ).目前有关森林根系分泌物研究大都仅关注了根系C源输入, 而忽略了根系分泌物中N成分变化及其伴随的C:N化学计量特征对土壤C养分循环过程的激发效应(priming effect), 这种忽略将极大地限制对森林根系-土壤-微生物互作机制的深入认识(Espinosa-Urgel & Ramos, 2001 ; Yin et al ., 2014 ).植物根系分泌物主要为一系列含碳化合物, 其C:N比值通常高于根际微生物的C:N (Cleveland & Liptzin, 2007 ).另外, 由于根系和微生物活动对根际有效N素的获取和激烈竞争, 使得根际区通常成为C过剩而N受限制强烈的区域(Kuzyakov, 2002 ).因此, 根系分泌物N含量或C:N化学计量特征成为驱动根际微生物群落组成和活性的重要调控因子.相应地, 根际微生物利用根系分泌物生长和合成胞外酶的能力严重地受根系分泌物N输入通量的制约, 从而反过来调控土壤生物地球化学过程及其对森林结构和功能的生态反馈效应(Drake et al ., 2013 ).此外, 根系分泌物对森林土壤C养分所诱导的激发效应与供试土壤养分状况(主要指土壤N的有效性)密切相关, 因为后者数量和质量是决定微生物对其生长和胞外酶产生之间能量分配的关键要素(Sullivan & Hart, 2013 ; Chen et al ., 2014 ).然而, 目前该方面研究的直接试验证据几乎还未见报道, 因此开展根系分泌物C:N化学计量特征对土壤C养分转化过程的影响成为一个十分重要但又极度缺乏的研究课题, 尤其在叠加不同土壤养分有效性的条件下. ...
... 土壤是陆地生态系统最大的有机碳库, 外源活性C添加通过土壤激发效应而引起土壤有机质矿化分解过程在短期内发生强烈的改变(Fontaine et al ., 2004 ), 并影响生态系统多个养分循环过程.此外, 由于土壤C-N转化过程密切的偶联效应, 土壤有机质分解和C库动态受根系N源输入以及土壤N养分状况控制.然而, 目前关于N养分有效性对土壤有机质分解的影响效应还未取得一致性的结论, 主要存在截然不同的两种假说: (1)微生物氮矿化(microbial nitrogen mining)假说.该假说认为土壤获N (N-acquiring)微生物通常利用活性外源C作为能源提高其生长和活性, 进而刺激土壤惰性有机质分解和养分转化过程, 以获取微生物生长所需各种养分(Moorhead & Sinsabaugh, 2006 ; 孙悦等, 2014 ); (2)化学计量分解理论(stoichiometric decomposition)假说, 根据该理论假说, 当外源C和N输入与土壤微生物C:N化学计量特征高度匹配时, 土壤微生物才具有最高的活性, 此时土壤有机质分解速率最大(Hessen et al ., 2004 ; Chen et al ., 2014 ).上述两种假说赋予了N养分有效性对土壤有机质稳定性相反的影响效应.第一种假说表明N受限条件(nutrient-poor condition)更有利于土壤有机质分解, 而第二种假说意味着N富集条件(nutrient-rich condition)更有利于土壤有机质分解(Chen et al ., 2014 ). ...
... ).上述两种假说赋予了N养分有效性对土壤有机质稳定性相反的影响效应.第一种假说表明N受限条件(nutrient-poor condition)更有利于土壤有机质分解, 而第二种假说意味着N富集条件(nutrient-rich condition)更有利于土壤有机质分解(Chen et al ., 2014 ). ...
... (1)微生物竞争性假说.正如前面所讨论的, 不同C:N化学计量特征根系分泌物添加所导致的微生物养分需求大小变化是诱导人工林土壤微生物群落结构(细菌/真菌相对优势度)改变的主要原因之一.在本研究中, 除N-only处理之外, 其他几种模拟处理均不同程度地进一步加剧了人工林土壤N限制, 更强的微生物N需求迫使微生物去矿化惰性有机质获取所需要的养分, 并改变微生物活性和微生物群落结构组成, 结果导致那些主要以分解惰性有机质的慢生长k策略功能微生物群(主要为真菌)获得更强的竞争性优势(Fontaine et al ., 2003 ; Ramirez et al ., 2012 ), 从而降低土壤细菌:真菌比值; 相反地, N添加处理则在一定程度上缓和了人工林土壤N限制程度, 从而减轻了微生物通过矿化惰性有机质库以获取N素的需求度(Craine et al ., 2007 ), 并使得那些以分解活性有机质的快生长r策略功能微生物群(主要为细菌)获得更强的竞争性优势(Dorodnikov et al ., 2009 ; Chen et al ., 2014 ), 从而导致N-only处理下人工林土壤细菌:真菌比值显著高于其他几种处理. ...
Synthesis and modeling perspectives of rhizosphere priming.
2
2014
... 近年来, 随着对森林根系活动和根际过程研究的不断深入, 根系在调控土壤功能和养分代谢过程中的重要作用逐渐被人们所认知和关注(Chapin et al ., 2009 ; Cheng et al ., 2014 ).植物除了通过细根周转向土壤输入碳(C)和养分外, 还可通过根系主动或被动地向周围土壤释放一系列化合物, 即狭义的根系分泌物, 主要包括一些低分子(有机酸、糖类、酚类和各种氨基酸等)和高分子(蛋白质、黏液等)有机化合物(Dijkstra & Cheng, 2007 ).研究表明, 通常森林根系分泌物量占植物光合同化产物的1%-5% (Grayston et al ., 1997 ), 并受森林类型、树种、环境条件以及土壤养分有效性等诸多因素影响Fransson & Johansson, 2010 ; Shi et al ., 2011 ).由于根系分泌物大多为一些易于土壤微生物直接利用的含碳有机物, 可为土壤微生物系统提供重要且丰富的C源和氮(N)源, 有效地改变土壤微生物数量和活性, 而深刻地影响根际土壤有机质分解和养分代谢等过程(Phillips et al ., 2009 ; Yin et al ., 2013 ), 从而导致根系分泌物在调控土壤C养分转化过程中发挥与其含量不成比例的重要作用和功能.植物根系分泌物输入及其介导的根系-土壤-微生物互作效应已成为陆地生态系统碳循环过程研究中一个非常重要但又最缺乏了解的关键环节(Jones et al ., 2004 ; 吴林坤等, 2014 ). ...
... 尽管当前对根系分泌物及其诱导的生态学效应研究已取得一定的进展, 但是对根系分泌物在森林根际生态过程中的作用及调控机理等方面我们依然缺乏足够的了解(Phillips et al ., 2011 ; Cheng et al ., 2014 ).目前有关森林根系分泌物研究大都仅关注了根系C源输入, 而忽略了根系分泌物中N成分变化及其伴随的C:N化学计量特征对土壤C养分循环过程的激发效应(priming effect), 这种忽略将极大地限制对森林根系-土壤-微生物互作机制的深入认识(Espinosa-Urgel & Ramos, 2001 ; Yin et al ., 2014 ).植物根系分泌物主要为一系列含碳化合物, 其C:N比值通常高于根际微生物的C:N (Cleveland & Liptzin, 2007 ).另外, 由于根系和微生物活动对根际有效N素的获取和激烈竞争, 使得根际区通常成为C过剩而N受限制强烈的区域(Kuzyakov, 2002 ).因此, 根系分泌物N含量或C:N化学计量特征成为驱动根际微生物群落组成和活性的重要调控因子.相应地, 根际微生物利用根系分泌物生长和合成胞外酶的能力严重地受根系分泌物N输入通量的制约, 从而反过来调控土壤生物地球化学过程及其对森林结构和功能的生态反馈效应(Drake et al ., 2013 ).此外, 根系分泌物对森林土壤C养分所诱导的激发效应与供试土壤养分状况(主要指土壤N的有效性)密切相关, 因为后者数量和质量是决定微生物对其生长和胞外酶产生之间能量分配的关键要素(Sullivan & Hart, 2013 ; Chen et al ., 2014 ).然而, 目前该方面研究的直接试验证据几乎还未见报道, 因此开展根系分泌物C:N化学计量特征对土壤C养分转化过程的影响成为一个十分重要但又极度缺乏的研究课题, 尤其在叠加不同土壤养分有效性的条件下. ...
C:N:P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass?
1
2007
... 尽管当前对根系分泌物及其诱导的生态学效应研究已取得一定的进展, 但是对根系分泌物在森林根际生态过程中的作用及调控机理等方面我们依然缺乏足够的了解(Phillips et al ., 2011 ; Cheng et al ., 2014 ).目前有关森林根系分泌物研究大都仅关注了根系C源输入, 而忽略了根系分泌物中N成分变化及其伴随的C:N化学计量特征对土壤C养分循环过程的激发效应(priming effect), 这种忽略将极大地限制对森林根系-土壤-微生物互作机制的深入认识(Espinosa-Urgel & Ramos, 2001 ; Yin et al ., 2014 ).植物根系分泌物主要为一系列含碳化合物, 其C:N比值通常高于根际微生物的C:N (Cleveland & Liptzin, 2007 ).另外, 由于根系和微生物活动对根际有效N素的获取和激烈竞争, 使得根际区通常成为C过剩而N受限制强烈的区域(Kuzyakov, 2002 ).因此, 根系分泌物N含量或C:N化学计量特征成为驱动根际微生物群落组成和活性的重要调控因子.相应地, 根际微生物利用根系分泌物生长和合成胞外酶的能力严重地受根系分泌物N输入通量的制约, 从而反过来调控土壤生物地球化学过程及其对森林结构和功能的生态反馈效应(Drake et al ., 2013 ).此外, 根系分泌物对森林土壤C养分所诱导的激发效应与供试土壤养分状况(主要指土壤N的有效性)密切相关, 因为后者数量和质量是决定微生物对其生长和胞外酶产生之间能量分配的关键要素(Sullivan & Hart, 2013 ; Chen et al ., 2014 ).然而, 目前该方面研究的直接试验证据几乎还未见报道, 因此开展根系分泌物C:N化学计量特征对土壤C养分转化过程的影响成为一个十分重要但又极度缺乏的研究课题, 尤其在叠加不同土壤养分有效性的条件下. ...
Microbial nitrogen limitation increases decomposition.
1
2007
... (1)微生物竞争性假说.正如前面所讨论的, 不同C:N化学计量特征根系分泌物添加所导致的微生物养分需求大小变化是诱导人工林土壤微生物群落结构(细菌/真菌相对优势度)改变的主要原因之一.在本研究中, 除N-only处理之外, 其他几种模拟处理均不同程度地进一步加剧了人工林土壤N限制, 更强的微生物N需求迫使微生物去矿化惰性有机质获取所需要的养分, 并改变微生物活性和微生物群落结构组成, 结果导致那些主要以分解惰性有机质的慢生长k策略功能微生物群(主要为真菌)获得更强的竞争性优势(Fontaine et al ., 2003 ; Ramirez et al ., 2012 ), 从而降低土壤细菌:真菌比值; 相反地, N添加处理则在一定程度上缓和了人工林土壤N限制程度, 从而减轻了微生物通过矿化惰性有机质库以获取N素的需求度(Craine et al ., 2007 ), 并使得那些以分解活性有机质的快生长r策略功能微生物群(主要为细菌)获得更强的竞争性优势(Dorodnikov et al ., 2009 ; Chen et al ., 2014 ), 从而导致N-only处理下人工林土壤细菌:真菌比值显著高于其他几种处理. ...
Labile soil carbon inputs mediate the soil microbial community composition and plant residue decomposition rates.
0
2010
Interactions between soil and tree roots accelerate long-term soil carbon decomposition.
1
2007
... 近年来, 随着对森林根系活动和根际过程研究的不断深入, 根系在调控土壤功能和养分代谢过程中的重要作用逐渐被人们所认知和关注(Chapin et al ., 2009 ; Cheng et al ., 2014 ).植物除了通过细根周转向土壤输入碳(C)和养分外, 还可通过根系主动或被动地向周围土壤释放一系列化合物, 即狭义的根系分泌物, 主要包括一些低分子(有机酸、糖类、酚类和各种氨基酸等)和高分子(蛋白质、黏液等)有机化合物(Dijkstra & Cheng, 2007 ).研究表明, 通常森林根系分泌物量占植物光合同化产物的1%-5% (Grayston et al ., 1997 ), 并受森林类型、树种、环境条件以及土壤养分有效性等诸多因素影响Fransson & Johansson, 2010 ; Shi et al ., 2011 ).由于根系分泌物大多为一些易于土壤微生物直接利用的含碳有机物, 可为土壤微生物系统提供重要且丰富的C源和氮(N)源, 有效地改变土壤微生物数量和活性, 而深刻地影响根际土壤有机质分解和养分代谢等过程(Phillips et al ., 2009 ; Yin et al ., 2013 ), 从而导致根系分泌物在调控土壤C养分转化过程中发挥与其含量不成比例的重要作用和功能.植物根系分泌物输入及其介导的根系-土壤-微生物互作效应已成为陆地生态系统碳循环过程研究中一个非常重要但又最缺乏了解的关键环节(Jones et al ., 2004 ; 吴林坤等, 2014 ). ...
Stimulation of microbial extracellular enzyme activities by elevated CO2 depends on soil aggregate size.
1
2009
... (1)微生物竞争性假说.正如前面所讨论的, 不同C:N化学计量特征根系分泌物添加所导致的微生物养分需求大小变化是诱导人工林土壤微生物群落结构(细菌/真菌相对优势度)改变的主要原因之一.在本研究中, 除N-only处理之外, 其他几种模拟处理均不同程度地进一步加剧了人工林土壤N限制, 更强的微生物N需求迫使微生物去矿化惰性有机质获取所需要的养分, 并改变微生物活性和微生物群落结构组成, 结果导致那些主要以分解惰性有机质的慢生长k策略功能微生物群(主要为真菌)获得更强的竞争性优势(Fontaine et al ., 2003 ; Ramirez et al ., 2012 ), 从而降低土壤细菌:真菌比值; 相反地, N添加处理则在一定程度上缓和了人工林土壤N限制程度, 从而减轻了微生物通过矿化惰性有机质库以获取N素的需求度(Craine et al ., 2007 ), 并使得那些以分解活性有机质的快生长r策略功能微生物群(主要为细菌)获得更强的竞争性优势(Dorodnikov et al ., 2009 ; Chen et al ., 2014 ), 从而导致N-only处理下人工林土壤细菌:真菌比值显著高于其他几种处理. ...
Stoichiometry constrains microbial response to root exudation-insights from a model and a field experiment in a temperate forest.
1
2013
... 尽管当前对根系分泌物及其诱导的生态学效应研究已取得一定的进展, 但是对根系分泌物在森林根际生态过程中的作用及调控机理等方面我们依然缺乏足够的了解(Phillips et al ., 2011 ; Cheng et al ., 2014 ).目前有关森林根系分泌物研究大都仅关注了根系C源输入, 而忽略了根系分泌物中N成分变化及其伴随的C:N化学计量特征对土壤C养分循环过程的激发效应(priming effect), 这种忽略将极大地限制对森林根系-土壤-微生物互作机制的深入认识(Espinosa-Urgel & Ramos, 2001 ; Yin et al ., 2014 ).植物根系分泌物主要为一系列含碳化合物, 其C:N比值通常高于根际微生物的C:N (Cleveland & Liptzin, 2007 ).另外, 由于根系和微生物活动对根际有效N素的获取和激烈竞争, 使得根际区通常成为C过剩而N受限制强烈的区域(Kuzyakov, 2002 ).因此, 根系分泌物N含量或C:N化学计量特征成为驱动根际微生物群落组成和活性的重要调控因子.相应地, 根际微生物利用根系分泌物生长和合成胞外酶的能力严重地受根系分泌物N输入通量的制约, 从而反过来调控土壤生物地球化学过程及其对森林结构和功能的生态反馈效应(Drake et al ., 2013 ).此外, 根系分泌物对森林土壤C养分所诱导的激发效应与供试土壤养分状况(主要指土壤N的有效性)密切相关, 因为后者数量和质量是决定微生物对其生长和胞外酶产生之间能量分配的关键要素(Sullivan & Hart, 2013 ; Chen et al ., 2014 ).然而, 目前该方面研究的直接试验证据几乎还未见报道, 因此开展根系分泌物C:N化学计量特征对土壤C养分转化过程的影响成为一个十分重要但又极度缺乏的研究课题, 尤其在叠加不同土壤养分有效性的条件下. ...
Expression of a Pseudomonas putida aminotransferase involved in lysine catabolism is induced in the rhizosphere.
1
2001
... 尽管当前对根系分泌物及其诱导的生态学效应研究已取得一定的进展, 但是对根系分泌物在森林根际生态过程中的作用及调控机理等方面我们依然缺乏足够的了解(Phillips et al ., 2011 ; Cheng et al ., 2014 ).目前有关森林根系分泌物研究大都仅关注了根系C源输入, 而忽略了根系分泌物中N成分变化及其伴随的C:N化学计量特征对土壤C养分循环过程的激发效应(priming effect), 这种忽略将极大地限制对森林根系-土壤-微生物互作机制的深入认识(Espinosa-Urgel & Ramos, 2001 ; Yin et al ., 2014 ).植物根系分泌物主要为一系列含碳化合物, 其C:N比值通常高于根际微生物的C:N (Cleveland & Liptzin, 2007 ).另外, 由于根系和微生物活动对根际有效N素的获取和激烈竞争, 使得根际区通常成为C过剩而N受限制强烈的区域(Kuzyakov, 2002 ).因此, 根系分泌物N含量或C:N化学计量特征成为驱动根际微生物群落组成和活性的重要调控因子.相应地, 根际微生物利用根系分泌物生长和合成胞外酶的能力严重地受根系分泌物N输入通量的制约, 从而反过来调控土壤生物地球化学过程及其对森林结构和功能的生态反馈效应(Drake et al ., 2013 ).此外, 根系分泌物对森林土壤C养分所诱导的激发效应与供试土壤养分状况(主要指土壤N的有效性)密切相关, 因为后者数量和质量是决定微生物对其生长和胞外酶产生之间能量分配的关键要素(Sullivan & Hart, 2013 ; Chen et al ., 2014 ).然而, 目前该方面研究的直接试验证据几乎还未见报道, 因此开展根系分泌物C:N化学计量特征对土壤C养分转化过程的影响成为一个十分重要但又极度缺乏的研究课题, 尤其在叠加不同土壤养分有效性的条件下. ...
Carbon input to soil may decrease soil carbon content.
1
2004
... 土壤是陆地生态系统最大的有机碳库, 外源活性C添加通过土壤激发效应而引起土壤有机质矿化分解过程在短期内发生强烈的改变(Fontaine et al ., 2004 ), 并影响生态系统多个养分循环过程.此外, 由于土壤C-N转化过程密切的偶联效应, 土壤有机质分解和C库动态受根系N源输入以及土壤N养分状况控制.然而, 目前关于N养分有效性对土壤有机质分解的影响效应还未取得一致性的结论, 主要存在截然不同的两种假说: (1)微生物氮矿化(microbial nitrogen mining)假说.该假说认为土壤获N (N-acquiring)微生物通常利用活性外源C作为能源提高其生长和活性, 进而刺激土壤惰性有机质分解和养分转化过程, 以获取微生物生长所需各种养分(Moorhead & Sinsabaugh, 2006 ; 孙悦等, 2014 ); (2)化学计量分解理论(stoichiometric decomposition)假说, 根据该理论假说, 当外源C和N输入与土壤微生物C:N化学计量特征高度匹配时, 土壤微生物才具有最高的活性, 此时土壤有机质分解速率最大(Hessen et al ., 2004 ; Chen et al ., 2014 ).上述两种假说赋予了N养分有效性对土壤有机质稳定性相反的影响效应.第一种假说表明N受限条件(nutrient-poor condition)更有利于土壤有机质分解, 而第二种假说意味着N富集条件(nutrient-rich condition)更有利于土壤有机质分解(Chen et al ., 2014 ). ...
Fungi mediate long term sequestration of carbon and nitrogen in soil through their priming effect.
1
2011
... 土壤有机质矿化及其伴随的碳库动态变化实质上是土壤微生物综合作用的结果, 而微生物组成和群落结构变化将深刻影响土壤生物地球化学循环过程(Schmidt et al ., 2011 ).在本研究中, 虽然模拟根系分泌物添加总体降低了天然林和人工林土壤TC含量, 但两种林分土壤对模拟根系分泌物C:N化学计量特征表现出相反的微生物反馈机制(图4 ).除N-only处理之外, 其他几种根系分泌物处理导致人工林土壤微生物PLFA总量和细菌、放线菌、真菌PLFA含量明显增加, 并伴随着更强的土壤有机质(SOM)分解激发效应和TC降低.这与Blagodatskaya 和Kuzyakov (2008)的报道一致, 其研究结果表明土壤微生物生物量大小调控外源活性C添加对土壤激发效应的幅度和方向.与人工林相反, 根系分泌物C源添加处理导致天然林土壤微生物总PLFA含量和细菌、放线菌、真菌PLFA含量明显下降, 这与C源添加后土壤TC含量下降这一结果并不吻合, 我们推测这可能与几种处理下土壤微生物生物量形成和胞外酶分配权衡(Fontaine et al ., 2011 )或特异性微生物代谢酶活性增强有关(de Graaff et al ., 2010).但遗憾的是, 由于试验设计限制, 在本研究中我们并没有同步监测与土壤C变化相关的微生物代谢酶活性变化, 使得无法准确提供不同处理下土壤碳动态变化的机理性证据和解释. ...
The priming effect of organic matter: A question of microbial competition?
1
2003
... (1)微生物竞争性假说.正如前面所讨论的, 不同C:N化学计量特征根系分泌物添加所导致的微生物养分需求大小变化是诱导人工林土壤微生物群落结构(细菌/真菌相对优势度)改变的主要原因之一.在本研究中, 除N-only处理之外, 其他几种模拟处理均不同程度地进一步加剧了人工林土壤N限制, 更强的微生物N需求迫使微生物去矿化惰性有机质获取所需要的养分, 并改变微生物活性和微生物群落结构组成, 结果导致那些主要以分解惰性有机质的慢生长k策略功能微生物群(主要为真菌)获得更强的竞争性优势(Fontaine et al ., 2003 ; Ramirez et al ., 2012 ), 从而降低土壤细菌:真菌比值; 相反地, N添加处理则在一定程度上缓和了人工林土壤N限制程度, 从而减轻了微生物通过矿化惰性有机质库以获取N素的需求度(Craine et al ., 2007 ), 并使得那些以分解活性有机质的快生长r策略功能微生物群(主要为细菌)获得更强的竞争性优势(Dorodnikov et al ., 2009 ; Chen et al ., 2014 ), 从而导致N-only处理下人工林土壤细菌:真菌比值显著高于其他几种处理. ...
Elevated CO2 and nitrogen influence exudation of soluble organic compounds by ectomycorrhizal root systems.
1
2010
... 近年来, 随着对森林根系活动和根际过程研究的不断深入, 根系在调控土壤功能和养分代谢过程中的重要作用逐渐被人们所认知和关注(Chapin et al ., 2009 ; Cheng et al ., 2014 ).植物除了通过细根周转向土壤输入碳(C)和养分外, 还可通过根系主动或被动地向周围土壤释放一系列化合物, 即狭义的根系分泌物, 主要包括一些低分子(有机酸、糖类、酚类和各种氨基酸等)和高分子(蛋白质、黏液等)有机化合物(Dijkstra & Cheng, 2007 ).研究表明, 通常森林根系分泌物量占植物光合同化产物的1%-5% (Grayston et al ., 1997 ), 并受森林类型、树种、环境条件以及土壤养分有效性等诸多因素影响Fransson & Johansson, 2010 ; Shi et al ., 2011 ).由于根系分泌物大多为一些易于土壤微生物直接利用的含碳有机物, 可为土壤微生物系统提供重要且丰富的C源和氮(N)源, 有效地改变土壤微生物数量和活性, 而深刻地影响根际土壤有机质分解和养分代谢等过程(Phillips et al ., 2009 ; Yin et al ., 2013 ), 从而导致根系分泌物在调控土壤C养分转化过程中发挥与其含量不成比例的重要作用和功能.植物根系分泌物输入及其介导的根系-土壤-微生物互作效应已成为陆地生态系统碳循环过程研究中一个非常重要但又最缺乏了解的关键环节(Jones et al ., 2004 ; 吴林坤等, 2014 ). ...
N:P ratios influence litter decomposition and colonization by fungi and bacteria in microcosms.
1
2009
... (2)细菌和真菌化学计量特征差异.导致人工林土壤细菌:真菌比值不同变化的另外一个原因可能与细菌和真菌本身微生物化学计量特征差异有关.研究表明, 真菌微生物量C:N通常高于细菌, 前者C:N一般介于5-15之间, 而后者C:N大多在3-6之间(McGill & Cole, 1981 ), 这表明真菌对养分需求比细菌低(Güsewell & Gessner, 2009 ).在本研究中, 除N-only之外, 其他几种模拟处理其C:N均明显高于细菌C:N比值, 相应地, 使得土壤微生物真菌相对于细菌而言获得更大的竞争性优势, 从而明显降低土壤细菌:真菌比值; 相反, N添加处理则在一定程度上缓和了人工林土壤N限制乃至土壤变为C限制, 土壤C:N降低, 相应地导致土壤细菌优势度更明显, 从而使得N-only处理下人工林土壤细菌:真菌显著高于其他几种处理(Carney et al ., 2007 ).此外, 土壤MBC:MBN在一定程度上是表征土壤细菌:真菌相对丰度的重要参数, 其中MBC:MBN比值越大, 表明真菌相对比例越高, 土壤细菌:真菌比值则越小(Zhao et al ., 2010 ).然而, 在本研究中, 我们发现基于土壤MBC:MBN比值所反馈的两种林分微生物群落结构变化趋势与通过PLFA方法所获得的微生物群落结构变化趋势并不完全一致.例如, N-only、C:N = 100和C-only几种处理导致天然林土壤MBC:MBN比值显著降低, 从而增加了土壤细菌:真菌比值; 而人工林除C:N = 10之外, 其他几种添加处理对土壤MBC:MBN无明显影响, 具体原因还有待进一步深入探究.虽然两种方法表征的土壤微生物群落结构变化信息并不完全吻合, 但二者从不同角度均强有力地证实了根系分泌物添加处理诱导天然林和人工林微生物群落结构发生了截然不同的改变. ...
Rhizosphere carbon flow in trees, in comparison with annual plants: The importance of root exudation and its impact on microbial activity and nutrient availability.
1
1997
... 近年来, 随着对森林根系活动和根际过程研究的不断深入, 根系在调控土壤功能和养分代谢过程中的重要作用逐渐被人们所认知和关注(Chapin et al ., 2009 ; Cheng et al ., 2014 ).植物除了通过细根周转向土壤输入碳(C)和养分外, 还可通过根系主动或被动地向周围土壤释放一系列化合物, 即狭义的根系分泌物, 主要包括一些低分子(有机酸、糖类、酚类和各种氨基酸等)和高分子(蛋白质、黏液等)有机化合物(Dijkstra & Cheng, 2007 ).研究表明, 通常森林根系分泌物量占植物光合同化产物的1%-5% (Grayston et al ., 1997 ), 并受森林类型、树种、环境条件以及土壤养分有效性等诸多因素影响Fransson & Johansson, 2010 ; Shi et al ., 2011 ).由于根系分泌物大多为一些易于土壤微生物直接利用的含碳有机物, 可为土壤微生物系统提供重要且丰富的C源和氮(N)源, 有效地改变土壤微生物数量和活性, 而深刻地影响根际土壤有机质分解和养分代谢等过程(Phillips et al ., 2009 ; Yin et al ., 2013 ), 从而导致根系分泌物在调控土壤C养分转化过程中发挥与其含量不成比例的重要作用和功能.植物根系分泌物输入及其介导的根系-土壤-微生物互作效应已成为陆地生态系统碳循环过程研究中一个非常重要但又最缺乏了解的关键环节(Jones et al ., 2004 ; 吴林坤等, 2014 ). ...
Carbon sequestration in ecosystems: The role of stoichiometry.
1
2004
... 土壤是陆地生态系统最大的有机碳库, 外源活性C添加通过土壤激发效应而引起土壤有机质矿化分解过程在短期内发生强烈的改变(Fontaine et al ., 2004 ), 并影响生态系统多个养分循环过程.此外, 由于土壤C-N转化过程密切的偶联效应, 土壤有机质分解和C库动态受根系N源输入以及土壤N养分状况控制.然而, 目前关于N养分有效性对土壤有机质分解的影响效应还未取得一致性的结论, 主要存在截然不同的两种假说: (1)微生物氮矿化(microbial nitrogen mining)假说.该假说认为土壤获N (N-acquiring)微生物通常利用活性外源C作为能源提高其生长和活性, 进而刺激土壤惰性有机质分解和养分转化过程, 以获取微生物生长所需各种养分(Moorhead & Sinsabaugh, 2006 ; 孙悦等, 2014 ); (2)化学计量分解理论(stoichiometric decomposition)假说, 根据该理论假说, 当外源C和N输入与土壤微生物C:N化学计量特征高度匹配时, 土壤微生物才具有最高的活性, 此时土壤有机质分解速率最大(Hessen et al ., 2004 ; Chen et al ., 2014 ).上述两种假说赋予了N养分有效性对土壤有机质稳定性相反的影响效应.第一种假说表明N受限条件(nutrient-poor condition)更有利于土壤有机质分解, 而第二种假说意味着N富集条件(nutrient-rich condition)更有利于土壤有机质分解(Chen et al ., 2014 ). ...
Plant and mycorrhizal regulation of rhizodeposition.
1
2004
... 近年来, 随着对森林根系活动和根际过程研究的不断深入, 根系在调控土壤功能和养分代谢过程中的重要作用逐渐被人们所认知和关注(Chapin et al ., 2009 ; Cheng et al ., 2014 ).植物除了通过细根周转向土壤输入碳(C)和养分外, 还可通过根系主动或被动地向周围土壤释放一系列化合物, 即狭义的根系分泌物, 主要包括一些低分子(有机酸、糖类、酚类和各种氨基酸等)和高分子(蛋白质、黏液等)有机化合物(Dijkstra & Cheng, 2007 ).研究表明, 通常森林根系分泌物量占植物光合同化产物的1%-5% (Grayston et al ., 1997 ), 并受森林类型、树种、环境条件以及土壤养分有效性等诸多因素影响Fransson & Johansson, 2010 ; Shi et al ., 2011 ).由于根系分泌物大多为一些易于土壤微生物直接利用的含碳有机物, 可为土壤微生物系统提供重要且丰富的C源和氮(N)源, 有效地改变土壤微生物数量和活性, 而深刻地影响根际土壤有机质分解和养分代谢等过程(Phillips et al ., 2009 ; Yin et al ., 2013 ), 从而导致根系分泌物在调控土壤C养分转化过程中发挥与其含量不成比例的重要作用和功能.植物根系分泌物输入及其介导的根系-土壤-微生物互作效应已成为陆地生态系统碳循环过程研究中一个非常重要但又最缺乏了解的关键环节(Jones et al ., 2004 ; 吴林坤等, 2014 ). ...
Review: Factors affecting rhizosphere priming effects.
1
2002
... 尽管当前对根系分泌物及其诱导的生态学效应研究已取得一定的进展, 但是对根系分泌物在森林根际生态过程中的作用及调控机理等方面我们依然缺乏足够的了解(Phillips et al ., 2011 ; Cheng et al ., 2014 ).目前有关森林根系分泌物研究大都仅关注了根系C源输入, 而忽略了根系分泌物中N成分变化及其伴随的C:N化学计量特征对土壤C养分循环过程的激发效应(priming effect), 这种忽略将极大地限制对森林根系-土壤-微生物互作机制的深入认识(Espinosa-Urgel & Ramos, 2001 ; Yin et al ., 2014 ).植物根系分泌物主要为一系列含碳化合物, 其C:N比值通常高于根际微生物的C:N (Cleveland & Liptzin, 2007 ).另外, 由于根系和微生物活动对根际有效N素的获取和激烈竞争, 使得根际区通常成为C过剩而N受限制强烈的区域(Kuzyakov, 2002 ).因此, 根系分泌物N含量或C:N化学计量特征成为驱动根际微生物群落组成和活性的重要调控因子.相应地, 根际微生物利用根系分泌物生长和合成胞外酶的能力严重地受根系分泌物N输入通量的制约, 从而反过来调控土壤生物地球化学过程及其对森林结构和功能的生态反馈效应(Drake et al ., 2013 ).此外, 根系分泌物对森林土壤C养分所诱导的激发效应与供试土壤养分状况(主要指土壤N的有效性)密切相关, 因为后者数量和质量是决定微生物对其生长和胞外酶产生之间能量分配的关键要素(Sullivan & Hart, 2013 ; Chen et al ., 2014 ).然而, 目前该方面研究的直接试验证据几乎还未见报道, 因此开展根系分泌物C:N化学计量特征对土壤C养分转化过程的影响成为一个十分重要但又极度缺乏的研究课题, 尤其在叠加不同土壤养分有效性的条件下. ...
1
2002
... 西南亚高山针叶林(subalpine coniferous forest)是青藏高原东部高寒林区的重要组成部分, 对于维持我国西部地区区域生态安全和促进区域可持续发展具有十分重要的“生态屏障(ecological barrier)”作用(刘庆, 2002 ).随着该区土地利用方式和管理措施的巨大变化, 自20世纪初期该区森林被大规模采伐, 随后在皆伐迹地营造了大面积的亚高山人工针叶林, 成为该区森林的重要组成部分.前期研究发现, 亚高山天然针叶林和云杉人工林具有截然不同的土壤有机质和养分含量(Xu et al ., 2010 ).因此, 本研究以川西亚高山天然林和云杉人工林两种森林林型土壤为试验对象, 通过配制不同C:N化学计量特征的模拟根系分泌物溶液进行人工添加试验, 测定不同处理下两种土壤碳含量动态及其微生物特征变化, 以探究根系分泌物C:N化学计量特征对不同森林类型土壤碳循环过程的影响差异, 为深入揭示森林根系分泌物与土壤C养分循环过程的耦合机制提供理论支持. ...
1
2002
... 西南亚高山针叶林(subalpine coniferous forest)是青藏高原东部高寒林区的重要组成部分, 对于维持我国西部地区区域生态安全和促进区域可持续发展具有十分重要的“生态屏障(ecological barrier)”作用(刘庆, 2002 ).随着该区土地利用方式和管理措施的巨大变化, 自20世纪初期该区森林被大规模采伐, 随后在皆伐迹地营造了大面积的亚高山人工针叶林, 成为该区森林的重要组成部分.前期研究发现, 亚高山天然针叶林和云杉人工林具有截然不同的土壤有机质和养分含量(Xu et al ., 2010 ).因此, 本研究以川西亚高山天然林和云杉人工林两种森林林型土壤为试验对象, 通过配制不同C:N化学计量特征的模拟根系分泌物溶液进行人工添加试验, 测定不同处理下两种土壤碳含量动态及其微生物特征变化, 以探究根系分泌物C:N化学计量特征对不同森林类型土壤碳循环过程的影响差异, 为深入揭示森林根系分泌物与土壤C养分循环过程的耦合机制提供理论支持. ...
Comparative aspects of cycling of organic C, N, S and P through soil organic matter.
1
1981
... (2)细菌和真菌化学计量特征差异.导致人工林土壤细菌:真菌比值不同变化的另外一个原因可能与细菌和真菌本身微生物化学计量特征差异有关.研究表明, 真菌微生物量C:N通常高于细菌, 前者C:N一般介于5-15之间, 而后者C:N大多在3-6之间(McGill & Cole, 1981 ), 这表明真菌对养分需求比细菌低(Güsewell & Gessner, 2009 ).在本研究中, 除N-only之外, 其他几种模拟处理其C:N均明显高于细菌C:N比值, 相应地, 使得土壤微生物真菌相对于细菌而言获得更大的竞争性优势, 从而明显降低土壤细菌:真菌比值; 相反, N添加处理则在一定程度上缓和了人工林土壤N限制乃至土壤变为C限制, 土壤C:N降低, 相应地导致土壤细菌优势度更明显, 从而使得N-only处理下人工林土壤细菌:真菌显著高于其他几种处理(Carney et al ., 2007 ).此外, 土壤MBC:MBN在一定程度上是表征土壤细菌:真菌相对丰度的重要参数, 其中MBC:MBN比值越大, 表明真菌相对比例越高, 土壤细菌:真菌比值则越小(Zhao et al ., 2010 ).然而, 在本研究中, 我们发现基于土壤MBC:MBN比值所反馈的两种林分微生物群落结构变化趋势与通过PLFA方法所获得的微生物群落结构变化趋势并不完全一致.例如, N-only、C:N = 100和C-only几种处理导致天然林土壤MBC:MBN比值显著降低, 从而增加了土壤细菌:真菌比值; 而人工林除C:N = 10之外, 其他几种添加处理对土壤MBC:MBN无明显影响, 具体原因还有待进一步深入探究.虽然两种方法表征的土壤微生物群落结构变化信息并不完全吻合, 但二者从不同角度均强有力地证实了根系分泌物添加处理诱导天然林和人工林微生物群落结构发生了截然不同的改变. ...
A theoretical model of litter decay and microbial interaction.
1
2006
... 土壤是陆地生态系统最大的有机碳库, 外源活性C添加通过土壤激发效应而引起土壤有机质矿化分解过程在短期内发生强烈的改变(Fontaine et al ., 2004 ), 并影响生态系统多个养分循环过程.此外, 由于土壤C-N转化过程密切的偶联效应, 土壤有机质分解和C库动态受根系N源输入以及土壤N养分状况控制.然而, 目前关于N养分有效性对土壤有机质分解的影响效应还未取得一致性的结论, 主要存在截然不同的两种假说: (1)微生物氮矿化(microbial nitrogen mining)假说.该假说认为土壤获N (N-acquiring)微生物通常利用活性外源C作为能源提高其生长和活性, 进而刺激土壤惰性有机质分解和养分转化过程, 以获取微生物生长所需各种养分(Moorhead & Sinsabaugh, 2006 ; 孙悦等, 2014 ); (2)化学计量分解理论(stoichiometric decomposition)假说, 根据该理论假说, 当外源C和N输入与土壤微生物C:N化学计量特征高度匹配时, 土壤微生物才具有最高的活性, 此时土壤有机质分解速率最大(Hessen et al ., 2004 ; Chen et al ., 2014 ).上述两种假说赋予了N养分有效性对土壤有机质稳定性相反的影响效应.第一种假说表明N受限条件(nutrient-poor condition)更有利于土壤有机质分解, 而第二种假说意味着N富集条件(nutrient-rich condition)更有利于土壤有机质分解(Chen et al ., 2014 ). ...
Elevated CO2 increases root exudation from loblolly pine (Pinus taeda) seedlings as an N-mediated response.
1
2009
... 近年来, 随着对森林根系活动和根际过程研究的不断深入, 根系在调控土壤功能和养分代谢过程中的重要作用逐渐被人们所认知和关注(Chapin et al ., 2009 ; Cheng et al ., 2014 ).植物除了通过细根周转向土壤输入碳(C)和养分外, 还可通过根系主动或被动地向周围土壤释放一系列化合物, 即狭义的根系分泌物, 主要包括一些低分子(有机酸、糖类、酚类和各种氨基酸等)和高分子(蛋白质、黏液等)有机化合物(Dijkstra & Cheng, 2007 ).研究表明, 通常森林根系分泌物量占植物光合同化产物的1%-5% (Grayston et al ., 1997 ), 并受森林类型、树种、环境条件以及土壤养分有效性等诸多因素影响Fransson & Johansson, 2010 ; Shi et al ., 2011 ).由于根系分泌物大多为一些易于土壤微生物直接利用的含碳有机物, 可为土壤微生物系统提供重要且丰富的C源和氮(N)源, 有效地改变土壤微生物数量和活性, 而深刻地影响根际土壤有机质分解和养分代谢等过程(Phillips et al ., 2009 ; Yin et al ., 2013 ), 从而导致根系分泌物在调控土壤C养分转化过程中发挥与其含量不成比例的重要作用和功能.植物根系分泌物输入及其介导的根系-土壤-微生物互作效应已成为陆地生态系统碳循环过程研究中一个非常重要但又最缺乏了解的关键环节(Jones et al ., 2004 ; 吴林坤等, 2014 ). ...
Enhanced root exudation induces microbial feedbacks to N cycling in a pine forest under long-term CO2 fumigation.
1
2011
... 尽管当前对根系分泌物及其诱导的生态学效应研究已取得一定的进展, 但是对根系分泌物在森林根际生态过程中的作用及调控机理等方面我们依然缺乏足够的了解(Phillips et al ., 2011 ; Cheng et al ., 2014 ).目前有关森林根系分泌物研究大都仅关注了根系C源输入, 而忽略了根系分泌物中N成分变化及其伴随的C:N化学计量特征对土壤C养分循环过程的激发效应(priming effect), 这种忽略将极大地限制对森林根系-土壤-微生物互作机制的深入认识(Espinosa-Urgel & Ramos, 2001 ; Yin et al ., 2014 ).植物根系分泌物主要为一系列含碳化合物, 其C:N比值通常高于根际微生物的C:N (Cleveland & Liptzin, 2007 ).另外, 由于根系和微生物活动对根际有效N素的获取和激烈竞争, 使得根际区通常成为C过剩而N受限制强烈的区域(Kuzyakov, 2002 ).因此, 根系分泌物N含量或C:N化学计量特征成为驱动根际微生物群落组成和活性的重要调控因子.相应地, 根际微生物利用根系分泌物生长和合成胞外酶的能力严重地受根系分泌物N输入通量的制约, 从而反过来调控土壤生物地球化学过程及其对森林结构和功能的生态反馈效应(Drake et al ., 2013 ).此外, 根系分泌物对森林土壤C养分所诱导的激发效应与供试土壤养分状况(主要指土壤N的有效性)密切相关, 因为后者数量和质量是决定微生物对其生长和胞外酶产生之间能量分配的关键要素(Sullivan & Hart, 2013 ; Chen et al ., 2014 ).然而, 目前该方面研究的直接试验证据几乎还未见报道, 因此开展根系分泌物C:N化学计量特征对土壤C养分转化过程的影响成为一个十分重要但又极度缺乏的研究课题, 尤其在叠加不同土壤养分有效性的条件下. ...
Consistent effects of nitrogen amendments on soil microbial communities and processes across biomes.
1
2012
... (1)微生物竞争性假说.正如前面所讨论的, 不同C:N化学计量特征根系分泌物添加所导致的微生物养分需求大小变化是诱导人工林土壤微生物群落结构(细菌/真菌相对优势度)改变的主要原因之一.在本研究中, 除N-only处理之外, 其他几种模拟处理均不同程度地进一步加剧了人工林土壤N限制, 更强的微生物N需求迫使微生物去矿化惰性有机质获取所需要的养分, 并改变微生物活性和微生物群落结构组成, 结果导致那些主要以分解惰性有机质的慢生长k策略功能微生物群(主要为真菌)获得更强的竞争性优势(Fontaine et al ., 2003 ; Ramirez et al ., 2012 ), 从而降低土壤细菌:真菌比值; 相反地, N添加处理则在一定程度上缓和了人工林土壤N限制程度, 从而减轻了微生物通过矿化惰性有机质库以获取N素的需求度(Craine et al ., 2007 ), 并使得那些以分解活性有机质的快生长r策略功能微生物群(主要为细菌)获得更强的竞争性优势(Dorodnikov et al ., 2009 ; Chen et al ., 2014 ), 从而导致N-only处理下人工林土壤细菌:真菌比值显著高于其他几种处理. ...
Labile and recalcitrant pools of carbon and nitrogen in organic matter decomposing at different depths in soil: An acid hydrolysis approach.
1
2002
... 土壤pH值采用电位法测定; 土壤含水量采用烘干法测定; 土壤总碳(TC)采用元素分析仪(MACRO cube, Elementar, Hanau, Germany)测定; 土壤活性和惰性碳库采用化学酸提法进行测定(Rovira & Vallejo, 2002 ).具体步骤如下: 称量过筛土壤0.5 g,加20 mL 2.5 mol·L-1 H2 SO4 溶液, 然后放在105 ℃下水解30 min, 离心, 把上清液转移到三角瓶中, 并用20 mL去离子水小心、缓慢地冲洗离心管, 上清液也加入三角瓶中, 这部分浸提液体中的土壤C含量被定义为活性碳库I (labile pool I carbon, LPI-C).剩余土壤残渣用2 mL13 mol·L-1 H2 SO4 溶液在室温下(不停摇动)浸提12 h, 然后把浸提液用去离子水稀释到1 mol·L-1 , 在105 ℃下水解3 h, 这部分浸提液中的C含量被定义为活性碳库II (labile pool II carbon, LPII-C).土壤惰性C库(recalcitrant pool carbon, RP-C)为土壤TC与活性C库(LPI-C与LPII-C之和)差值.土壤微生物生物量碳(MBC)和微生物生物量氮(MBN)采用氯仿熏蒸法测定. ...
Persistence of soil organic matter as an ecosystem property.
1
2011
... 土壤有机质矿化及其伴随的碳库动态变化实质上是土壤微生物综合作用的结果, 而微生物组成和群落结构变化将深刻影响土壤生物地球化学循环过程(Schmidt et al ., 2011 ).在本研究中, 虽然模拟根系分泌物添加总体降低了天然林和人工林土壤TC含量, 但两种林分土壤对模拟根系分泌物C:N化学计量特征表现出相反的微生物反馈机制(图4 ).除N-only处理之外, 其他几种根系分泌物处理导致人工林土壤微生物PLFA总量和细菌、放线菌、真菌PLFA含量明显增加, 并伴随着更强的土壤有机质(SOM)分解激发效应和TC降低.这与Blagodatskaya 和Kuzyakov (2008)的报道一致, 其研究结果表明土壤微生物生物量大小调控外源活性C添加对土壤激发效应的幅度和方向.与人工林相反, 根系分泌物C源添加处理导致天然林土壤微生物总PLFA含量和细菌、放线菌、真菌PLFA含量明显下降, 这与C源添加后土壤TC含量下降这一结果并不吻合, 我们推测这可能与几种处理下土壤微生物生物量形成和胞外酶分配权衡(Fontaine et al ., 2011 )或特异性微生物代谢酶活性增强有关(de Graaff et al ., 2010).但遗憾的是, 由于试验设计限制, 在本研究中我们并没有同步监测与土壤C变化相关的微生物代谢酶活性变化, 使得无法准确提供不同处理下土壤碳动态变化的机理性证据和解释. ...
In situ sampling of low molecular weight organic anions from rhizosphere of radiata pine (Pinus radiata) grown in a rhizotron system.
1
2011
... 近年来, 随着对森林根系活动和根际过程研究的不断深入, 根系在调控土壤功能和养分代谢过程中的重要作用逐渐被人们所认知和关注(Chapin et al ., 2009 ; Cheng et al ., 2014 ).植物除了通过细根周转向土壤输入碳(C)和养分外, 还可通过根系主动或被动地向周围土壤释放一系列化合物, 即狭义的根系分泌物, 主要包括一些低分子(有机酸、糖类、酚类和各种氨基酸等)和高分子(蛋白质、黏液等)有机化合物(Dijkstra & Cheng, 2007 ).研究表明, 通常森林根系分泌物量占植物光合同化产物的1%-5% (Grayston et al ., 1997 ), 并受森林类型、树种、环境条件以及土壤养分有效性等诸多因素影响Fransson & Johansson, 2010 ; Shi et al ., 2011 ).由于根系分泌物大多为一些易于土壤微生物直接利用的含碳有机物, 可为土壤微生物系统提供重要且丰富的C源和氮(N)源, 有效地改变土壤微生物数量和活性, 而深刻地影响根际土壤有机质分解和养分代谢等过程(Phillips et al ., 2009 ; Yin et al ., 2013 ), 从而导致根系分泌物在调控土壤C养分转化过程中发挥与其含量不成比例的重要作用和功能.植物根系分泌物输入及其介导的根系-土壤-微生物互作效应已成为陆地生态系统碳循环过程研究中一个非常重要但又最缺乏了解的关键环节(Jones et al ., 2004 ; 吴林坤等, 2014 ). ...
Considering fungal: Bacterial dominance in soils-Methods, controls, and ecosystem implications.
1
2010
... 此外, 根系分泌物处理诱导两种林分土壤微生物群落结构也产生了不同的改变.虽然两种林分土壤微生物群落结构变化在C:N = 10、C:N = 50、C:N = 100和C-only几种处理之间无明显差异, 但通过PLFA分析结果总体表明N添加倾向增加两种林分土壤细菌:真菌比值, 而C添加显著降低人工林土壤细菌:真菌比值, 但对天然林土壤细菌:真菌的影响并不明显.土壤微生物群落结构及其细菌:真菌相对优势度改变深刻地影响和调控土壤C养分循环过程(Strickland & Rousk, 2010 ).目前有关根系分泌物输入对森林土壤微生物群落结构影响的具体机理并不清楚, 基于文献调研和本文的试验结果, 我们提出如下两种假说解释不同化学计量特征根系分泌物处理后人工林土壤微生物群落结构变化的原因. ...
Evaluation of mechanisms controlling the priming of soil carbon along a substrate age gradient.
1
2013
... 尽管当前对根系分泌物及其诱导的生态学效应研究已取得一定的进展, 但是对根系分泌物在森林根际生态过程中的作用及调控机理等方面我们依然缺乏足够的了解(Phillips et al ., 2011 ; Cheng et al ., 2014 ).目前有关森林根系分泌物研究大都仅关注了根系C源输入, 而忽略了根系分泌物中N成分变化及其伴随的C:N化学计量特征对土壤C养分循环过程的激发效应(priming effect), 这种忽略将极大地限制对森林根系-土壤-微生物互作机制的深入认识(Espinosa-Urgel & Ramos, 2001 ; Yin et al ., 2014 ).植物根系分泌物主要为一系列含碳化合物, 其C:N比值通常高于根际微生物的C:N (Cleveland & Liptzin, 2007 ).另外, 由于根系和微生物活动对根际有效N素的获取和激烈竞争, 使得根际区通常成为C过剩而N受限制强烈的区域(Kuzyakov, 2002 ).因此, 根系分泌物N含量或C:N化学计量特征成为驱动根际微生物群落组成和活性的重要调控因子.相应地, 根际微生物利用根系分泌物生长和合成胞外酶的能力严重地受根系分泌物N输入通量的制约, 从而反过来调控土壤生物地球化学过程及其对森林结构和功能的生态反馈效应(Drake et al ., 2013 ).此外, 根系分泌物对森林土壤C养分所诱导的激发效应与供试土壤养分状况(主要指土壤N的有效性)密切相关, 因为后者数量和质量是决定微生物对其生长和胞外酶产生之间能量分配的关键要素(Sullivan & Hart, 2013 ; Chen et al ., 2014 ).然而, 目前该方面研究的直接试验证据几乎还未见报道, 因此开展根系分泌物C:N化学计量特征对土壤C养分转化过程的影响成为一个十分重要但又极度缺乏的研究课题, 尤其在叠加不同土壤养分有效性的条件下. ...
根际激发效应的发生机制及其生态重要性
1
2014
... 土壤是陆地生态系统最大的有机碳库, 外源活性C添加通过土壤激发效应而引起土壤有机质矿化分解过程在短期内发生强烈的改变(Fontaine et al ., 2004 ), 并影响生态系统多个养分循环过程.此外, 由于土壤C-N转化过程密切的偶联效应, 土壤有机质分解和C库动态受根系N源输入以及土壤N养分状况控制.然而, 目前关于N养分有效性对土壤有机质分解的影响效应还未取得一致性的结论, 主要存在截然不同的两种假说: (1)微生物氮矿化(microbial nitrogen mining)假说.该假说认为土壤获N (N-acquiring)微生物通常利用活性外源C作为能源提高其生长和活性, 进而刺激土壤惰性有机质分解和养分转化过程, 以获取微生物生长所需各种养分(Moorhead & Sinsabaugh, 2006 ; 孙悦等, 2014 ); (2)化学计量分解理论(stoichiometric decomposition)假说, 根据该理论假说, 当外源C和N输入与土壤微生物C:N化学计量特征高度匹配时, 土壤微生物才具有最高的活性, 此时土壤有机质分解速率最大(Hessen et al ., 2004 ; Chen et al ., 2014 ).上述两种假说赋予了N养分有效性对土壤有机质稳定性相反的影响效应.第一种假说表明N受限条件(nutrient-poor condition)更有利于土壤有机质分解, 而第二种假说意味着N富集条件(nutrient-rich condition)更有利于土壤有机质分解(Chen et al ., 2014 ). ...
根际激发效应的发生机制及其生态重要性
1
2014
... 土壤是陆地生态系统最大的有机碳库, 外源活性C添加通过土壤激发效应而引起土壤有机质矿化分解过程在短期内发生强烈的改变(Fontaine et al ., 2004 ), 并影响生态系统多个养分循环过程.此外, 由于土壤C-N转化过程密切的偶联效应, 土壤有机质分解和C库动态受根系N源输入以及土壤N养分状况控制.然而, 目前关于N养分有效性对土壤有机质分解的影响效应还未取得一致性的结论, 主要存在截然不同的两种假说: (1)微生物氮矿化(microbial nitrogen mining)假说.该假说认为土壤获N (N-acquiring)微生物通常利用活性外源C作为能源提高其生长和活性, 进而刺激土壤惰性有机质分解和养分转化过程, 以获取微生物生长所需各种养分(Moorhead & Sinsabaugh, 2006 ; 孙悦等, 2014 ); (2)化学计量分解理论(stoichiometric decomposition)假说, 根据该理论假说, 当外源C和N输入与土壤微生物C:N化学计量特征高度匹配时, 土壤微生物才具有最高的活性, 此时土壤有机质分解速率最大(Hessen et al ., 2004 ; Chen et al ., 2014 ).上述两种假说赋予了N养分有效性对土壤有机质稳定性相反的影响效应.第一种假说表明N受限条件(nutrient-poor condition)更有利于土壤有机质分解, 而第二种假说意味着N富集条件(nutrient-rich condition)更有利于土壤有机质分解(Chen et al ., 2014 ). ...
根系分泌物介导下植物-土壤-微生物互作关系研究进展与展望
1
2014
... 近年来, 随着对森林根系活动和根际过程研究的不断深入, 根系在调控土壤功能和养分代谢过程中的重要作用逐渐被人们所认知和关注(Chapin et al ., 2009 ; Cheng et al ., 2014 ).植物除了通过细根周转向土壤输入碳(C)和养分外, 还可通过根系主动或被动地向周围土壤释放一系列化合物, 即狭义的根系分泌物, 主要包括一些低分子(有机酸、糖类、酚类和各种氨基酸等)和高分子(蛋白质、黏液等)有机化合物(Dijkstra & Cheng, 2007 ).研究表明, 通常森林根系分泌物量占植物光合同化产物的1%-5% (Grayston et al ., 1997 ), 并受森林类型、树种、环境条件以及土壤养分有效性等诸多因素影响Fransson & Johansson, 2010 ; Shi et al ., 2011 ).由于根系分泌物大多为一些易于土壤微生物直接利用的含碳有机物, 可为土壤微生物系统提供重要且丰富的C源和氮(N)源, 有效地改变土壤微生物数量和活性, 而深刻地影响根际土壤有机质分解和养分代谢等过程(Phillips et al ., 2009 ; Yin et al ., 2013 ), 从而导致根系分泌物在调控土壤C养分转化过程中发挥与其含量不成比例的重要作用和功能.植物根系分泌物输入及其介导的根系-土壤-微生物互作效应已成为陆地生态系统碳循环过程研究中一个非常重要但又最缺乏了解的关键环节(Jones et al ., 2004 ; 吴林坤等, 2014 ). ...
根系分泌物介导下植物-土壤-微生物互作关系研究进展与展望
1
2014
... 近年来, 随着对森林根系活动和根际过程研究的不断深入, 根系在调控土壤功能和养分代谢过程中的重要作用逐渐被人们所认知和关注(Chapin et al ., 2009 ; Cheng et al ., 2014 ).植物除了通过细根周转向土壤输入碳(C)和养分外, 还可通过根系主动或被动地向周围土壤释放一系列化合物, 即狭义的根系分泌物, 主要包括一些低分子(有机酸、糖类、酚类和各种氨基酸等)和高分子(蛋白质、黏液等)有机化合物(Dijkstra & Cheng, 2007 ).研究表明, 通常森林根系分泌物量占植物光合同化产物的1%-5% (Grayston et al ., 1997 ), 并受森林类型、树种、环境条件以及土壤养分有效性等诸多因素影响Fransson & Johansson, 2010 ; Shi et al ., 2011 ).由于根系分泌物大多为一些易于土壤微生物直接利用的含碳有机物, 可为土壤微生物系统提供重要且丰富的C源和氮(N)源, 有效地改变土壤微生物数量和活性, 而深刻地影响根际土壤有机质分解和养分代谢等过程(Phillips et al ., 2009 ; Yin et al ., 2013 ), 从而导致根系分泌物在调控土壤C养分转化过程中发挥与其含量不成比例的重要作用和功能.植物根系分泌物输入及其介导的根系-土壤-微生物互作效应已成为陆地生态系统碳循环过程研究中一个非常重要但又最缺乏了解的关键环节(Jones et al ., 2004 ; 吴林坤等, 2014 ). ...
Initial responses of soil CO2 efflux and C, N pools to experimental warming in two contrasting forest ecosys- tems, Eastern Tibetan Plateau, China.
2
2010
... 西南亚高山针叶林(subalpine coniferous forest)是青藏高原东部高寒林区的重要组成部分, 对于维持我国西部地区区域生态安全和促进区域可持续发展具有十分重要的“生态屏障(ecological barrier)”作用(刘庆, 2002 ).随着该区土地利用方式和管理措施的巨大变化, 自20世纪初期该区森林被大规模采伐, 随后在皆伐迹地营造了大面积的亚高山人工针叶林, 成为该区森林的重要组成部分.前期研究发现, 亚高山天然针叶林和云杉人工林具有截然不同的土壤有机质和养分含量(Xu et al ., 2010 ).因此, 本研究以川西亚高山天然林和云杉人工林两种森林林型土壤为试验对象, 通过配制不同C:N化学计量特征的模拟根系分泌物溶液进行人工添加试验, 测定不同处理下两种土壤碳含量动态及其微生物特征变化, 以探究根系分泌物C:N化学计量特征对不同森林类型土壤碳循环过程的影响差异, 为深入揭示森林根系分泌物与土壤C养分循环过程的耦合机制提供理论支持. ...
... 在本研究中, C-only和C:N = 100 (后一种处理N浓度仅为0.6 mg·L-1 , 因此其N含量基本可忽略)两种处理显著降低了两种林分土壤TC含量, 而适量的N添加(N-only、C:N = 50和C:N = 10)则在一定程度上缓和了土壤TC含量的降低(图1 ).土壤N限制是西南亚高山针叶林主要特征之一(Xu et al ., 2010 ), 人工林更是如此(表1 ), 根系分泌物外源C添加进一步诱导了该区两种林分土壤N限制, 土壤微生物利用输入C源并刺激微生物生长和活性, 导致土壤有效N被快速消耗(Bengtson et al ., 2012 ; Burns et al ., 2013 ).低的有效N促使微生物分泌更多的胞外酶以加速土壤有机质分解, 释放无机N来满足微生物生长所需的养分, 从而促进土壤有机质分解速率和CO2 释放, 使两种林分土壤TC含量显著降低.然而, 添加一定量的N组分在一定程度上缓和了两种林分土壤N养分限制, 使得微生物不需要通过分解难以利用的土壤有机质获取N, 而是偏好利用易于分解的有机化合物, 相应地减少土壤胞外酶的分泌, 从而降低土壤有机质分解和CO2 释放, 并缓和了两种针叶林土壤TC含量的降低幅度.上述研究结果极好地印证了前人所提出的N矿化假说理论, 即N养分有效性受限会促进土壤有机质分解. ...
Enhanced root exudation stimulates soil nitrogen transformations in a subalpine coniferous forest under experimental warming.
1
2013
... 近年来, 随着对森林根系活动和根际过程研究的不断深入, 根系在调控土壤功能和养分代谢过程中的重要作用逐渐被人们所认知和关注(Chapin et al ., 2009 ; Cheng et al ., 2014 ).植物除了通过细根周转向土壤输入碳(C)和养分外, 还可通过根系主动或被动地向周围土壤释放一系列化合物, 即狭义的根系分泌物, 主要包括一些低分子(有机酸、糖类、酚类和各种氨基酸等)和高分子(蛋白质、黏液等)有机化合物(Dijkstra & Cheng, 2007 ).研究表明, 通常森林根系分泌物量占植物光合同化产物的1%-5% (Grayston et al ., 1997 ), 并受森林类型、树种、环境条件以及土壤养分有效性等诸多因素影响Fransson & Johansson, 2010 ; Shi et al ., 2011 ).由于根系分泌物大多为一些易于土壤微生物直接利用的含碳有机物, 可为土壤微生物系统提供重要且丰富的C源和氮(N)源, 有效地改变土壤微生物数量和活性, 而深刻地影响根际土壤有机质分解和养分代谢等过程(Phillips et al ., 2009 ; Yin et al ., 2013 ), 从而导致根系分泌物在调控土壤C养分转化过程中发挥与其含量不成比例的重要作用和功能.植物根系分泌物输入及其介导的根系-土壤-微生物互作效应已成为陆地生态系统碳循环过程研究中一个非常重要但又最缺乏了解的关键环节(Jones et al ., 2004 ; 吴林坤等, 2014 ). ...
Root-induced changes in nutrient cycling in forests depend on exudation rates.
1
2014
... 尽管当前对根系分泌物及其诱导的生态学效应研究已取得一定的进展, 但是对根系分泌物在森林根际生态过程中的作用及调控机理等方面我们依然缺乏足够的了解(Phillips et al ., 2011 ; Cheng et al ., 2014 ).目前有关森林根系分泌物研究大都仅关注了根系C源输入, 而忽略了根系分泌物中N成分变化及其伴随的C:N化学计量特征对土壤C养分循环过程的激发效应(priming effect), 这种忽略将极大地限制对森林根系-土壤-微生物互作机制的深入认识(Espinosa-Urgel & Ramos, 2001 ; Yin et al ., 2014 ).植物根系分泌物主要为一系列含碳化合物, 其C:N比值通常高于根际微生物的C:N (Cleveland & Liptzin, 2007 ).另外, 由于根系和微生物活动对根际有效N素的获取和激烈竞争, 使得根际区通常成为C过剩而N受限制强烈的区域(Kuzyakov, 2002 ).因此, 根系分泌物N含量或C:N化学计量特征成为驱动根际微生物群落组成和活性的重要调控因子.相应地, 根际微生物利用根系分泌物生长和合成胞外酶的能力严重地受根系分泌物N输入通量的制约, 从而反过来调控土壤生物地球化学过程及其对森林结构和功能的生态反馈效应(Drake et al ., 2013 ).此外, 根系分泌物对森林土壤C养分所诱导的激发效应与供试土壤养分状况(主要指土壤N的有效性)密切相关, 因为后者数量和质量是决定微生物对其生长和胞外酶产生之间能量分配的关键要素(Sullivan & Hart, 2013 ; Chen et al ., 2014 ).然而, 目前该方面研究的直接试验证据几乎还未见报道, 因此开展根系分泌物C:N化学计量特征对土壤C养分转化过程的影响成为一个十分重要但又极度缺乏的研究课题, 尤其在叠加不同土壤养分有效性的条件下. ...
Addition of external organic carbon and native soil organic carbon decomposi- tion: A meta-analysis.
0
2013
Nitrogen and phosphorus transformations in the rhizospheres of three tree species in a nutrient-poor sandy soil.
1
2010
... (2)细菌和真菌化学计量特征差异.导致人工林土壤细菌:真菌比值不同变化的另外一个原因可能与细菌和真菌本身微生物化学计量特征差异有关.研究表明, 真菌微生物量C:N通常高于细菌, 前者C:N一般介于5-15之间, 而后者C:N大多在3-6之间(McGill & Cole, 1981 ), 这表明真菌对养分需求比细菌低(Güsewell & Gessner, 2009 ).在本研究中, 除N-only之外, 其他几种模拟处理其C:N均明显高于细菌C:N比值, 相应地, 使得土壤微生物真菌相对于细菌而言获得更大的竞争性优势, 从而明显降低土壤细菌:真菌比值; 相反, N添加处理则在一定程度上缓和了人工林土壤N限制乃至土壤变为C限制, 土壤C:N降低, 相应地导致土壤细菌优势度更明显, 从而使得N-only处理下人工林土壤细菌:真菌显著高于其他几种处理(Carney et al ., 2007 ).此外, 土壤MBC:MBN在一定程度上是表征土壤细菌:真菌相对丰度的重要参数, 其中MBC:MBN比值越大, 表明真菌相对比例越高, 土壤细菌:真菌比值则越小(Zhao et al ., 2010 ).然而, 在本研究中, 我们发现基于土壤MBC:MBN比值所反馈的两种林分微生物群落结构变化趋势与通过PLFA方法所获得的微生物群落结构变化趋势并不完全一致.例如, N-only、C:N = 100和C-only几种处理导致天然林土壤MBC:MBN比值显著降低, 从而增加了土壤细菌:真菌比值; 而人工林除C:N = 10之外, 其他几种添加处理对土壤MBC:MBN无明显影响, 具体原因还有待进一步深入探究.虽然两种方法表征的土壤微生物群落结构变化信息并不完全吻合, 但二者从不同角度均强有力地证实了根系分泌物添加处理诱导天然林和人工林微生物群落结构发生了截然不同的改变. ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}