外来物种入侵已成为严重危害陆地及海洋环境的生态问题, 被认为是一种生物污染而被广泛关注和研究(Weis & Weis, 2003 )。深入了解外来物种的入侵机制, 并进行有效的预测、预防和管理是减少入侵生物危害的最有效手段(Byers, 2002 ), 而从区域尺度上分析入侵物种的分布特征并探索其形成原因是认识生物入侵问题的重要基础(Erik, 2004 )。
针对入侵植物的分布问题, Losdale (1999)研究了全球入侵植物的分布特征, 并发现温带地区的耕地、城镇和生物多样性热点地区更易遭受外来植物的入侵, 而荒漠、稀树草原等生态系统的可入侵空间较小; Barney等(2008)研究了两种不同入侵历史的外来植物在北美的分布和扩散格局, 发现其入侵范围均随着时间的推移而扩大; 此外, 还有不少学者运用气候模拟技术对入侵植物的潜在分布区进行了预测(Erik, 2004 ; Ibáñez et al ., 2009 ; Cross- man et al ., 2011 ); 而国内的研究多集中在入侵植物多样性的区域分异及其影响因素等方面, 如吴晓雯等(2006)研究了入侵植物在我国的分布格局, 发现其物种数从南到北逐渐减少, 而物种密度由东南海岸向内陆递减的分布特征, 并推测中国的东南部地区将更容易遭受外来植物入侵; 王苏铭等(2012)研究了入侵植物在北京地区的分布特征; 张帅等(2010)研究了入侵植物在我国74个地理单元的分布情况, 发现入侵植物的分布具有明显的区域分化现象, 而降水与温度是影响其空间分布格局的主要 因素。
有关入侵植物影响生态系统功能的报道有很多, 并多关注其负面效应Williamson, 2001 ; Byers et al ., 2002 ; Simberloff et al ., 2003 ), 认为植物入侵导致本地群落中生物多样性的下降李冰和李玉瑛, 2009 ), 例如: 丁晖等(2007)研究发现, 入侵森林林隙中的紫茎泽兰(Ageratina adenophora )使群落中本地植物的物种丰富度显著下降; Fox等在澳洲西部一个草地生态系统中的研究表明群落中的外来物种比例与本地植物种数呈负相关关系(Stohlgren et al ., 1999 ); Tilman (1997)在美国Bethel市一个草地生态系统中的实验也得出了一致的结论。因此一般认为, 外来植物成功入侵后将抑制群落中本地植物的多样性。但也有研究发现: 入侵植物对群落植物多样性的影响与入侵种及本地植物的生活型有关(Hejda et al ., 2009 ); 植物入侵的生态效应与入侵地的演替阶段有关, 例如: 豆科植物入侵生态环境恶劣的尾矿区后, 能有效地改善土壤肥力, 为演替中期过渡种的定居和发育创造良好的条件, 并促进物种多样性的增加Gozlan & Newton, 2009 ); 李安定等(2013)研究发现, 喜旱莲子草(空心莲子草)(Alternanthera philoxeroides )在入侵的早期阶段能改变群落生境条件, 进而促进其他物种的生存与定居, 并增加物种的多样性, 而只有当其种群盖度持续增加后, 才可能会威胁其他物种的生存; Hejda等(2009)研究发现, 入侵植物的盖度是决定群落物种丰富度和均匀度的主要因子, 入侵植物对物种多样性的影响与本地植物优势种的盖度、高度有密切关系。因此, 植物入侵的生态效应受生态系统类型、群落演替阶段的影响; 不同功能群植物对植物入侵的响应也有差异。本文通过研究7种生态系统类型中入侵植物的入侵特征及其对本地植物的影响, 旨在检验以下科学假设: 1)植物入侵的生态效应与入侵地的环境特征和演替阶段有关, 入侵植物可能对入侵地产生正面或负面影响; 2)植物入侵主要影响群落中某一功能群的物种多样性, 而不是影响全部群落个体; 3)多数入侵植物有偏好的入侵生境, 而人为干扰少的生态系统类型对植物入侵有抵制作用。
海南岛虽然气候条件宜人、生物资源丰富, 但生态环境脆弱, 是许多外来入侵物种的“天然大温室”, 目前有关海南岛入侵植物的研究主要集中在以下两方面: 一是区域性入侵植物种类调查或群落组成调查(单家林, 2003 ; 秦新生等, 2008 ); 二是入侵植物生理生态的研究(Huang et al ., 2013 ; 黄乔乔等, 2013 )。这些研究有效地整合了特定区域内外来植物种类组成的信息, 并在入侵生态学方面做了一定的探索, 但还没有在海南全岛范围内开展主要入侵植物的分布特征及其入侵强度的研究, 有关入侵植物在不同生态系统中的入侵特征及对本地植物多样性影响的研究都还缺乏。本文研究了国家农业部指定的20种恶性入侵植物在海南岛的分布特征及其对当地植物的影响, 并比较了6种主要入侵植物在7种生态系统中的入侵强度, 以期进一步了解海南岛入侵植物的分布现状, 并为外来植物的防控、生物多样性保护与生态安全的管理提供理论依据。
1 材料和方法
1.1 研究区域地理、生态环境概况
海南岛地处热带北缘, 面积3.4万km2 , 是我国第二大岛屿。海南岛气候温和, 年平均气温为22.5-26.0 ℃, 年降水量为1500-2000 mm。海南岛的地势中部高、四周低, 这种地理地貌使得其年总积温、日照时数等都具有自中部向沿海递增的特征, 受季风与台风影响, 年降水量分布不均匀, 干湿两季明显, 且不同区域的年降水量存在差异。如东部与中部属于湿润区, 西部与南部属于半干旱区, 水分相对不足, 北部地区则为半湿润区(唐少霞等, 2008 )。海南岛植物资源丰富, 目前记载的野生植物有4000多种, 外来植物165种, 其中入侵植物有58种(彭宗波等, 2013 ; 杨小波等, 2013 )。
1.2 研究方法与对象
1.2.1 入侵植物种的确定
根据国家农业部2012年对海南省恶性入侵植物的调查要求, 确定以下20种为调查的目的种: 飞机草(Chromolaena odorata )、假臭草(Praxelis clematidea )、含羞草(Mimosa pudica )、银胶菊(Parthenium hysterophorus )、微甘菊(Mikania micrantha )、凤眼蓝(Eichhornia crassipes )、假高粱(Pseudosorghum fasciculare )、三裂叶蟛蜞菊(Wedelia trilobata )、苏门白酒草(Conyza sumatrensis )、喜旱莲子草(Altern- anthera philoxeroides )、马缨丹(Lantana camara )、黄顶菊(Flaveria bidentis )、Solanum rostratum 、野莴苣(Lactuca serriola )、加拿大一枝黄花(Solidago canadensis )、Sicyos angulatus 、紫茎泽兰、豚草(Ambrosia artemisiifolia )、Cenchrus pauciflorus 、齿裂大戟(Euphorbia dentata )。
1.2.2 样方设置
调查于2012年4-5月进行, 采用路线法与样方调查相结合的方法, 抽样调查了海南岛18个市县的近郊区和自然保护林区。调查样方随机布设于农田、村落、种植园、林缘、弃耕地、草地、天然次生林等不同生态系统, 共布设了351个样地。根据所处的演替阶段的不同, 调查的生态系统类型可分为处于演替早期的农田、村落、种植园和草地生态系统; 处于演替中后期的林缘、天然次生林生态系统。为避免季节性收割对本地植物变化的影响, 本文数据分析中选取的种植园生态系统以胡椒园、茶树园、桉树林、橡胶园等轮作周期长的种植园为主。天然次生林主要为热带雨林或季雨林, 以下简称次生林, 两种植被类型分布的树种主要有高山榕(Ficus altissima )、蓝树(Wrightia laevis )、郎伞木(Ardisia elegans )、九节(Psychotria rubra )、牛矢果(Osmanthus matsumuranus )、银珠(Peltophorum tonkinense )、山麻秆(Alchornea davidii )、银柴(Aporosa dioica )、榼藤(Entada phaseoloides )、相思子(Abrus precatorius )、海南假砂仁(Amomum chinense )等。发现目的种种群后, 以该种群为中心, 设置一个2 m × 2 m的样方, 然后随机取一方向, 沿该方向每隔10 m设一个样方, 共设5个2 m × 2 m的样方(1套样方), 然后沿垂直于第1套样方的方向设置第2套样方, 两套样方共10个样方, 计40 m2 (1个样地)。记录样地中的土壤类型及植被特征, 统计样方内的植物种类。采用目测估值法计算群落中植物群落的盖度, 计算方法为: 盖度=草本植物的植株投影面积/样方面积。
1.2.3 数据分析
选择农田、村落、种植园、林缘、弃耕地、草地、次生林7种人为干扰程度有差异和不同演替特征的生境类型作为研究对象, 结合调查中各生态系统类型的入侵程度和实地调查的可行性, 分别在农田、村落、种植园、林缘、弃耕地、草地、次生林中选取14、14、20、11、17、11、10个共97个样地的信息用于数据统计分析(图1 ), 分析的样地共含970个样方, 总面积为3904 m2 。以1套样方中入侵样方所占比例作为入侵频率, 用以比较不同生态系统中入侵植物的入侵频率, 这样, 每个样地有2个重复。目前, 有关植物盖度等级的划分多出现在植被覆盖度估算的研究中(盖永芹等, 2009 ; 李锦荣等; 2010 ), 而对样方中种群盖度等级的划分没有统一的标准, 因此, 在划分盖度等级时, 本文根据评估标准及分布范围, 把入侵植物的种群盖度分为一级(1%-20%)、二级(21%-40%)、三级(41%-60%)、四级(≥61%)共4个等级。运用Excel 2003和SPSS 17.0软件进行数据统计分析。用Systat SigmaPlot 10.0软件绘图。由于调查数据中入侵频率和盖度两项指标是服从二项分布的百分数的数据资料, 并且含有小于30%或大于70%的数值, 因此, 数据分析前对其进行反正弦转换Sokal & Rohlf, 1995 )。
图1 调查及分析的样地在海南岛的分布。
Fig. 1 Distribution of surveyed and analyzed sites in Hainan Island.
2 结果
2.1 海南岛入侵植物种类组成
在国家农业部指定的20种恶性入侵植物中, 海南岛分布有11种, 占总数的55.0%; 包括飞机草、假臭草、含羞草、银胶菊、微甘菊、三裂叶蟛蜞菊、假高粱、苏门白酒草、喜旱莲子草、马缨丹、凤眼蓝。
2.2 海南岛入侵植物的水平分布格局
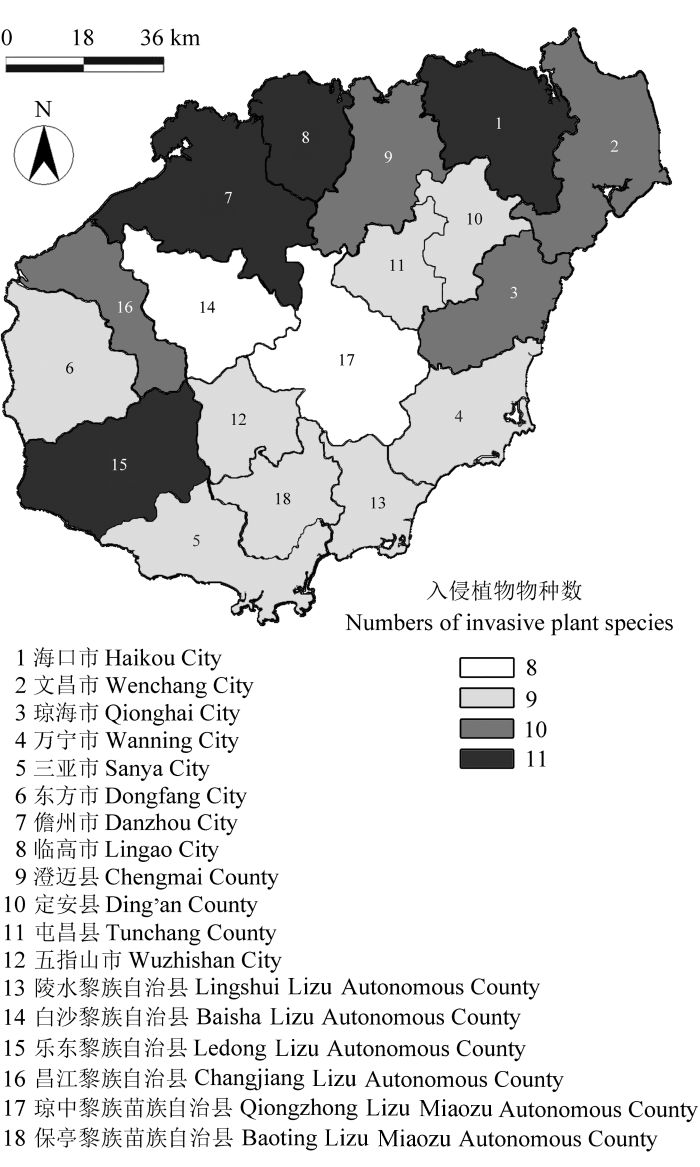
海南岛恶性入侵植物的分布情况如图2 , 其中, 海口市、临高县、儋州市和乐东县是分布入侵植物种数最多的4个市县, 均有11种, 其次是琼海市、文昌市、澄迈县和昌江县, 均有10种, 而琼中市和白沙县分布种数最少, 只有8种, 其余8个市县均有9种(附录1)。海南岛东北部是入侵植物分布比较集中的区域, 单个市县分布有入侵植物9-11种; 其次是西南部的昌江市和乐东县, 分别有10和11种, 而中部地区分布较少, 其余市县均分布有9种。入侵植物在海南岛的分布特征表现为分布广泛, 区域间种数相差不大, 但整体表现为东北部、西南部地区种类最多, 东南部地区次之, 中部地区种类最少。
图2 海南岛入侵植物在各市县分布的数量梯度图。
Fig. 2 The richness of invasive species and its distribution across cities and counties of Hainan Island.
海南岛广布的恶性入侵植物为假臭草、飞机草、含羞草、苏门白酒草、马缨丹、三裂叶蟛蜞菊, 这6种入侵植物在抽查的每个市县均有分布, 入侵频率高; 而其他5种入侵植物在不同区域的分布情况存在差异, 如银胶菊在西南部和北部地区的入侵频率高, 而在中部的五指山市、琼中县等地区入侵频率较低; 凤眼蓝则主要分布在北部和东部市县, 在西南部的昌江、东方和乐东县的入侵频率较低; 微甘菊与假高粱在全岛的分布广泛, 但目前入侵强度小, 应该加强防范。
2.3 不同生态系统中入侵植物的种类组成
从表1 可以看出, 除次生林外, 其余6种生态系统中都有入侵植物分布, 其中弃耕地、村落、农田中分布的入侵植物种数最多, 均有10种, 草地和种植园生态系统中有7种, 而林缘中分布最少, 只有6种。飞机草、假臭草、含羞草、马缨丹、三裂叶蟛蜞菊、苏门白酒草的入侵生境广泛, 在除次生林外的其他6种生境下均有分布; 银胶菊主要分布在村落、弃耕地中; 而假高粱常入侵到弃耕地、农田、村落生态系统中; 凤眼蓝主要分布于村落及农田边的河道中。
2.4 主要入侵植物的入侵频率及其与生境的关系
比较11种入侵植物在6种生境下的入侵频率发现: 单以累计入侵频率分析时, 6种生态系统的整体入侵强度没有显著差异(p > 0.05); 但种植园、农田和弃耕地中累计入侵频率高, 可入侵空间较大, 而在林缘生态系统中, 累计入侵频率最低, 可入侵空间小, 体现了林缘生境对入侵植物的制约性(表2 )。
分析单一入侵植物在不同生态系统中的入侵频率可知: 苏门白酒草, 马缨丹在6种生境中的入侵频率没有显著差异(p > 0.05), 表明这两种入侵植物的生态适应性广泛, 而假臭草、含羞草及三裂叶蟛蜞菊均有最适的入侵生境。例如: 假臭草在种植园和弃耕地中的入侵频率较高, 分别为0.946和0.706, 显著大于其他4种生境下的入侵频率(p < 0.01); 含羞草在农田生境中的入侵频率为0.592, 显著高于其他5种生境中的入侵频率(p < 0.01); 三裂叶蟛蜞菊在种植园下的入侵频率也显著高于其他生境下的入侵频率(p < 0.01); 而微甘菊、假高粱、银胶菊在入侵生境中的入侵频率也没有显著差异性; 与陆生入侵植物相比, 凤眼蓝和喜旱莲子草的入侵生境相对局限, 主要分布于农田等具有湿生环境特征的生态系统中, 体现出入侵生境的局限性。
2.5 不同生态系统中主要入侵植物的入侵强度比较分析
6种恶性入侵植物的种群盖度变化范围大, 但除三裂叶蟛蜞菊外, 其余入侵植物的种群盖度均集中分布在1%-20%之间(表3 ); 进一步分析种群盖度与入侵频率之间的关系可知, 苏门白酒草的入侵频率与种群盖度没有相关性; 而假臭草在弃耕地, 含羞草在农田, 三裂叶蟛蜞菊在种植园生态系统中, 其种群盖度与入侵频率呈正相关关系, 表明这3种入侵植物有偏好的入侵生境; 而飞机草的种群盖度与入侵频率在除种植园外的生态系统中均表现为显著或极显著的负相关关系; 在林缘生态系统中, 除马缨丹和苏门白酒草外, 其余4种入侵植物的入侵频率都随着盖度等级的增加而显著降低, 表明在林缘生态系统中, 多数入侵植物很难成片分布生长, 林缘对植物入侵存在制约作用。
2.6 不同生态系统中入侵植物种数与本地植物种数的关系
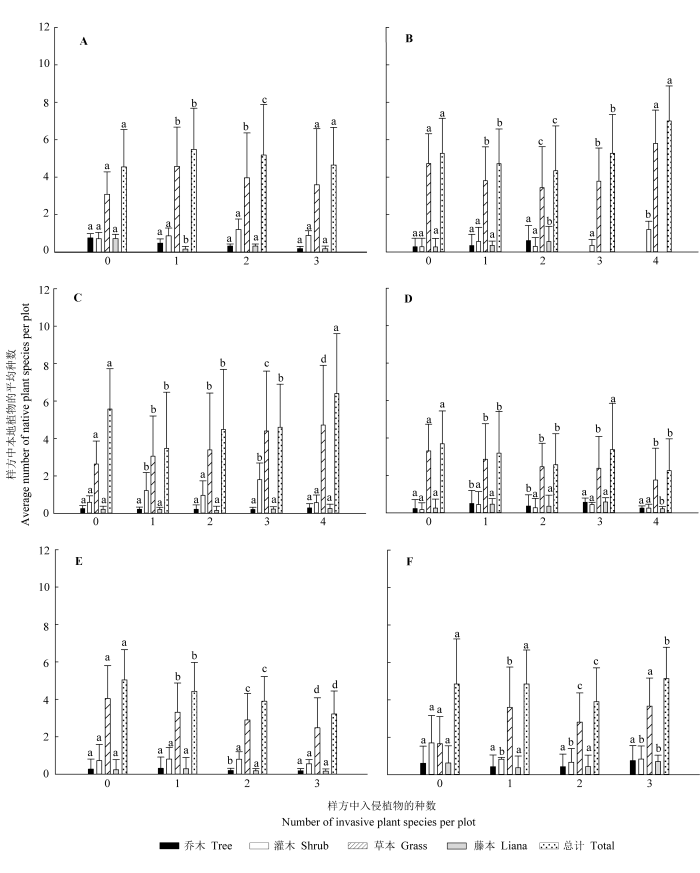
针对6种生态系统类型, 研究单个样方中入侵植物种数变化对本地植物平均种数的影响, 并按生活型把本地植物分成乔木、灌木、草本、藤本4种类型, 分别研究4种类型植物的平均种数对入侵植物种数变化的响应, 结果表明: 在弃耕地、种植园、林缘3种生态系统中, 群落中本地植物的平均种数与入侵植物种数变化无明显相关性(图3C 、3D、3F), 而在农田生态系统中, 两者呈先增加后减少的变化趋势(图3A ), 其中当样方中入侵植物种数为1时, 本地植物平均种数最多; 而在草地生态系统中, 两者表现出先减少后增加的变化趋势, 其中当入侵植物种数为2时, 本地植物平均种数最少(图3B ); 在村落生态系统中, 本地植物平均种数随着入侵植物种数的增加而显著减少, 两者呈负相关关系(图3E )。
样方中乔木、灌木及藤本的平均种数与入侵种数的变化在研究的6类生态系统中均没有明显的相关性, 这3类生活型植物受入侵植物种数变化的影响小; 而草本植物的平均种数在不同的生态系统中对入侵植物种数增加的响应不同, 如在种植园和林缘生态系统中, 两者无相关性(图3D 、3F); 在农田生态系统中, 表现出先增加后减少的趋势(图3A ), 而在草地生态系统中, 表现出先减少后增加的趋势(图3B ); 在弃耕地生态系统中, 虽然当入侵植物种数为1和2时, 群落中草本植物的平均种数无显著差异, 但其整体变化趋势为随着入侵植物种数的增加而增加(图3C ), 而在村落中, 两者呈负相关关系(图3E )。
图3 不同生态系统中样方中入侵植物种数与本地植物平均种数的变化关系(平均值±标准误差)。A, 农田。B, 草地。C, 弃耕地。D, 种植园。E, 村落。F, 林缘。不同小写字母表示差异显著(p < 0.05)。
Fig. 3 The relationship between the number of invasive plant species and the average number of native plant species per plot in different types of ecosystems (mean ± SE ). A, Farmland. B, Grassland. C, Abandoned land. D, Plantation. E, Village. F, Forest edge. Different lowercase letters indicate significant differences (p < 0.05).
2.7 不同生态系统中主要入侵植物盖度总值与本地植物种数的关系
以调查的单个样地为对象, 统计10个样方中所有入侵植物的盖度总值与本地植物种数, 并分析不同生态系统中两者之间的相关性, 结果表明: 在种植园、农田和村落3种生态系统中, 入侵植物盖度总值与本地植物的物种丰富度没有显著相关性(p > 0.05) (图4A 、4B、4C); 而在林缘(n = 12, p = 0.031, R 2 = 0.387)和弃耕地(n = 18, p = 0.045, R 2 = 0.251)生态系统中, 样地中入侵植物的盖度总值与本地植物种数呈显著负相关关系(p < 0.05), 本地植物的物种丰富度将随着入侵植物盖度总值的增大而减少(图4E 、4F), 高盖度的入侵植物居群将抑制本地植物的多样性; 但在草地(n = 11, p = 0.014, R 2 = 0.449)生态系统中, 在单个样方平均盖度低于49.1%时, 入侵植物的盖度总值与本地植物种数却呈正相关关系(p < 0.05), 即本地植物的物种丰富度将随着入侵植物盖度总值的增加而增加, 表现在特定的入侵阶段, 增加入侵植物的居群盖度有助于促进本地植物的多样性(图4D )。
图4 6种生态系统中样地中入侵植物盖度总值与本地植物种数的关系。A, 种植园。B, 村落。C, 农田。D, 草地。E, 林缘。F, 弃耕地。
Fig. 4 The relationship between the total coverage of invasive plants and the number of native plant species sampled from quadrats of six ecosystems. A, Plantation. B, Village. C, Farmland. D, Grassland. E, Forest edge. F, Abandoned land.
3 讨论
3.1 入侵植物在海南岛的分布特征
入侵植物在海南岛分布广泛, 且不同区域间种数相差不大, 但整体呈现为东北部、西南部地区种类较多, 东南部次之, 而中部较少的分布特征。入侵植物的分布主要取决于环境的可入侵性和物种的入侵能力两个方面Newsome & Noble, 1986 ; 刘建等, 2010 )。如“理想”的入侵植物通常具备生活史短, 自交亲和性、结实率高, 可进行无性繁殖等特点, 并能迅速响应环境中的干扰Rejmánek & Richardson, 1996 )。本文研究的11种入侵植物中, 除马缨丹外, 其余10种均为一、二年生的草本植物, 其中有6种为菊科植物, 如飞机草、假臭草、微甘菊等, 这些植物产生的种子数量大而质量轻, 通常还附有冠毛, 便于长距离传播和扩散李振宇和谢焱, 2002 ; Weber et al ., 2008 ), 而喜旱莲子草、微甘菊、凤眼蓝、三裂叶蟛蜞菊具有无性繁殖的特性, 其繁殖体可以借助水流进行传播, 这些生物学特征和传播机制增强了其入侵能力, 有助于入侵植物的扩散和迅速占据入侵地, 并能在较短的时间内扩大种群, 从而导致其在海南岛的分布范围很广。但对比不同区域的分布特点发现, 东北部和西南部的入侵种数相对较多, 为10-11种, 而中部和东南部分布较少, 这种差异可能与不同区域间的气候条件和人为干扰强度有关。由于海南岛的西部地区贯穿有环西粤海铁路, 具有较长的运输史, 而连接该地区的海口市、儋州市、东方市都是水陆交通的重要枢纽, 境内人流密集, 干扰强度大, 这些因素都能有效地促进入侵植物的传播和扩散(吴晓雯等, 2006 ; 张帅等, 2010 )。结合气候条件进一步分析, 海南岛西部地区的年晴天数平均为300天, 明显高于中部和东部地区, 但降水量较少, 而中部地区是全岛降水集中分布的地区, 年降水量达2000 mm, 属于湿润地区。此外, 中部地区的年平均气温为23 ℃左右, 处于全岛平均气温(23-26 ℃)的下限, 低于海南岛其他地区的温度(蔡大鑫等, 2010 )。综合气候条件和入侵种数的分布特征发现, 入侵植物在年平均气温较高、降水量较低、而日照时间较长的东北、西南部地区分布的种类较多, 在年平均气温较低、降水量较高、日照时间较短的中部、东部地区分布的种类较少, 这可能与入侵植物的生活习性有关, 因为本文研究的入侵植物多为草本阳性植物, 它们的种子萌发和种群发展都需要足够的光温条件, 过高湿和低温的环境可能对其生长和扩散具有抑制作用。
3.2 不同生境下入侵植物的发生频率和种群特征
本文研究表明: 弃耕地、村落、农田生态系统中所含的入侵种数最多, 均有10种; 而在林缘生境中, 11种入侵植物的入侵频率和累计入侵频率都较低, 在抽样调查的次生林内, 没有发现入侵植物的分布(表1 ), 说明入侵植物主要分布在人为干扰强度大的生境中, 而在人为干扰强度小、群落结构复杂的生境中分布较少。结合6种生态系统的演替阶段分析, 弃耕地、村落、草地和农田均属于演替早期的生境类型, 受人为干扰的强度大, 而机械和人为活动的介导能有效地减少入侵植物的传播障碍, 或形成干扰间的协同效应(Hobbs, 1991 ); 入侵植物通常能够迅速响应环境中的干扰, 并且具有较强的表型可塑性, 这些特征促成了其在干扰生境中的成功入侵(Hobbs & Huenneke, 1992 )。植物对光能的捕获和利用效率高低是决定其能否适应环境并很好地存活下去的重要因素Baruch & Goldstein, 1999 ; Durand & Goldstein, 2001 ), 因此, 入侵生境的光环境对植物的入侵过程有很大影响, 如弃耕地通常具有高光照、高肥力的环境特征, 这为入侵种群的建立和发展提供了条件, 导致弃耕地常成为入侵植物(如假臭草、三裂叶蟛蜞菊等)集中分布的生境, 这与钟军弟等(2014)在广西的研究结果一致, 表明弃耕地具有较大的可入侵空间。林缘和天然次生林生态系统属于演替中后期的生境类型, 与演替前期的生境类型相比, 受人为干扰强度较低, 而增加的物种密度也将减少群落中的空缺生态位, 从而降低植物成功入侵的几率Stachowicz & Tilman, 2005 )。另一方面, 林缘或林下的光照强度和光合有效辐射都比较低, 这种环境特征也将限制植物入侵林缘的深度Brothers & Spingarn, 1992 ; Shea & Chesson, 2002 ), 因为本文研究的入侵植物多是处于演替初期的草本植物, 这些植物从种子萌发到幼苗生长及种群的发育均需要充足的光照, 它们的入侵可能在演替前期的生境类型, 如农田、草地和村落中具有更好的生态适应性; 但在林缘和林下的入侵却受到光照条件的制约, 且林缘的群落结构对低矮生活型的入侵植物也具有阻碍作用(Honnay et al ., 2002 ), 如喜旱莲子草、含羞草等在林缘中的入侵频率都很低。因此, 入侵植物虽然在林区广泛分布, 但它们仅分布在光照充足的路边或新垦的裸地上, 而无法入侵到森林内部, 国外的不少报道也证实了这种分布特点Parendes & Jones, 2000 )。分析入侵种群的盖度可知, 除三裂叶蟛蜞菊的种群盖度较高外, 其余5种入侵植物的盖度都集中分布在1%-20%之间, 并与入侵地的环境特征存在相关性, 如含羞草在农田、假臭草在弃耕地、三裂叶蟛蜞菊在种植园中通常具有成群分布生长的特点, 体现出这3种入侵植物在对应的生境中有更好的生态适应性; 而在林缘生态系统中, 除马缨丹和苏门白酒草外, 其余4种入侵植物的入侵频率与种群盖度呈负相关关系, 表明在林缘生态系统中, 入侵植物在入侵群落中的密度小, 难以形成单优群落, 这与入侵植物的种子是否能到达林缘, 能否存活及能否萌芽有密切的关系, 这些问题有待进一步实验验证。
3.3 入侵群落中入侵植物对本地植物的影响
入侵植物导致本地植物多样性降低的现象已被广泛认识, 但在不同生态系统中可能存在差异(Daehler, 2003 )。本文的研究结果显示, 样方中入侵植物种数与本地植物平均种数在弃耕地、种植园和林缘3种生态系统中无明显相关性, 当样方中入侵植物种数由0到4种变化时, 群落中本地植物平均种数均没有明显的递变规律; 进一步分析不同生活型植物与入侵植物种数变化的关系发现, 仅弃耕地中草本植物的平均种数略有上升的趋势, 但差异不显著(图3C ); 而乔木、灌木、藤本生活型植物的平均种数与入侵植物的种数变化不相关。在农田生态系统中, 本地植物平均种数随入侵植物种数的增加呈先增加后减少的变化趋势, 其中当入侵植物种数为1时, 本地植物平均种数最多, 随后呈减少的趋势(图3A ), 表明群落中少数入侵种的存在具有促进本地植物多样性的作用, 其原因可能是早期的植物入侵过程改善了群落环境并促进了本地种的生存和定居, 从而增加了本地植物的物种丰富度, 但持续扩大的入侵种群将会抑制本地植物的多样性, 这与李安定等(2013)对莲子草的研究结果是一致的。本文的研究结果也表明: 针对不同的生态系统类型和不同的入侵阶段, 植物入侵对本地植物的影响存在差异; 而一定强度的植物入侵可能具有正面的生态效应。
Stohlgren等(1999)对草地生态系统的研究表明, 在群落水平上, 本地植物种数与入侵植物种数呈正相关关系, 并认为这与两类植物对草地环境的变化有相同的响应有关。而本文发现, 草地群落中本地植物平均种数随入侵种数的增加呈先减少后增加的趋势, 且在入侵种数为2时最少, 草本植物也呈一致的变化趋势, 而乔木、灌木和藤本植物的种数变化不显著, 其原因可能是当入侵种数较少时, 某些入侵植物排挤本地植物后形成了单优群落, 并导致本地植物物种丰富度下降, 而随着入侵种数的增加, 新入侵的植物改变了优势种的组成, 这种变化进一步促进了新的本地植物迁入, 从而增加了本地植物的多样性, 因为群落优势种对新物种的迁入具有决定性作用(Liancourt et al ., 2009 )。Huston (1997)的研究也表明, 群落的可入侵性更多取决于关键种, 而并非整个群落中的物种数, 但在较小尺度上取样时, 物种多样性与入侵关系的模式比较复杂, 群落类型和地域的差异都会导致不同的结果, 还需要考虑其他因素的影响(Fridley et al ., 2007 )。针对本文研究的10种陆生入侵植物, 在入侵植物组合不同的群落中, 具体是哪一类群的入侵植物对本地植物的种数有直接影响, 还需要针对某些单一入侵植物的群落进行比较研究, 并考虑草地演替过程与入侵植物迁入的关系。在村落生态系统中, 本地植物平均种数与入侵植物种数呈负相关关系, 其原因可能是人为地对居住环境进行改造后, 减少了本地植物的多样性, 使入侵植物更容易占据空缺生态位, 并逐渐成为群落优势种, 随着干扰强度的增加, 其优势地位变得更突出, 而群落中的次优种和伴生种的地位逐渐降低甚至消失, 这与王丽丽(2013)对农村生态系统中伴人植物的研究结果是一致的, 体现出人为干扰介导下入侵植物对本地植物多样性的影响。
在本文研究的6种生态系统中, 样方中乔木、灌木和藤本植物的平均种数与入侵植物种数的变化均没有显著相关性, 说明这3种生活型植物的物种丰富度受植物入侵的影响小, 而草本植物在不同生态系统中对入侵植物种数的变化有不同的响应, 如在农田生态系统中, 当样方中入侵植物种数由0到3种变化时, 群落中草本植物平均种数呈先增加后减少的变化趋势, 其中当入侵植物种数为1时达到最多; 而在草地生态系统中, 草本植物平均种数随入侵植物种数的增加呈先减少后增加的变化趋势(图3B ), 其中当入侵植物种数为2时最少, 并且这两类生态系统中草本植物种数的变化都与群落中本地植物平均种数的变化趋势一致。虽然在弃耕地中, 本地植物平均种数与入侵植物种数没有显著相关性, 但群落中草本植物的平均种数却表现出随入侵植物种数的增加而增加的趋势(图3C ), 表明在农田、草地和弃耕地3类生态系统中, 群落中入侵植物种数的变化对乔木、灌木和藤本植物物种丰富度的影响并不显著, 但对草本植物的物种丰富度有显著的影响, 进一步表明: 植物入侵后并非影响群落中的所有个体, 而是对其中某一植物功能群有较直接的作用。
4 结论
(1)在国家农业部确定的20种恶性入侵植物中, 海南岛分布有11种, 占目的种总数的55%; 海南岛的东北部、西南部地区是入侵植物分布种数较多的区域, 东南部次之, 中部最少; 入侵植物在各市县的分布种数相差不大, 表现出在全岛广布的特征; 飞机草、假臭草、含羞草、苏门白酒草、三裂叶蟛蜞菊、马缨丹是海南岛目前分布最广泛、入侵强度较大的6种入侵植物。
(2)结合海南岛的土地利用方式和气候条件发现, 弃耕地、种植园以及农田是入侵植物分布的主要生境, 而林缘和天然次生林生态系统的可入侵空间较小, 并对植物入侵表现出制约作用; 入侵植物主要分布在人为干扰强度大的生境中, 多数入侵植物的种群盖度小, 并没有形成大面积爆发的入侵态势。
(3)植物入侵对本地植物的影响因生态系统类型存在差异, 一种是负面效应, 表现在村落生态系统中; 一种是正面效应(至少是阶段性的), 体现在草地、农田生态系统中, 即一定强度范围内的植物入侵过程能改善入侵群落的生境条件, 促进本地植物的多样性; 而另外一种表现为以人为干扰为介导的综合效应, 如林缘、种植园和弃耕地3种生态系统类型, 植物入侵的效应可能受干扰强度的影响而存在较大的差异。
(4)植物入侵主要对群落中草本功能群的物种多样性有显著影响, 而对乔木、灌木、藤本的影响不显著; 入侵植物的盖度总值对本地植物物种多样性的影响因生态系统类型而存在差异, 具有正面、负面或者不相关3种效应。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Barney JN Whitlow TH Lembo AJ Jr 2008 ). Revealing historic invasion patterns and potential invasion sites for two non-native plant species.
PLoS ONE , 3 (2 ), e1635 , doi: 10.1371/journal.pone.0001635 .
[2]
Baruch Z Goldstein G 1999 ). Leaf construction cost, nutrient concentration, and net CO2 assimilation of native and invasive species in Hawaii.
Oecologia , 121 , 183 -192 .
[本文引用: 1]
[3]
Brothers TS Spingarn A 1992 ). Forest fragmentation and alien plant invasion of central Indiana old-growth forests.
Conservation Biology , 6 , 91 -100 .
[本文引用: 1]
[4]
Byers JE 2002 ). Physical habitat attribute mediates biotic resistant to non-indigenous species invasion.
Oecologia , 130 , 146 -156 .
[本文引用: 2]
[5]
Byers JE Reichard SH Randall JM Parker IM Smith CS Lonsdale WM Atkinson IAE Seastedt TR Williamson M Chornesky E Hayes D 2002 ). Directing research to reduce the impacts of nonindigenous species.
Conservation Biology , 16 , 630 -640 .
[6]
Cai DX Liu SJ Tian GH Xu XC Cui D Zhang JH 2010 ). Analysis on tourism climate resource of Hainan Island.
Modern Agricultural Science and Technology , (16 ), 19 -24 (in Chinese with English abstract).
[本文引用: 1]
[蔡大鑫 , 刘少军 , 田光辉 , 许向春 , 崔丹 , 张京红 (2010 ). 海南岛旅游气候资源分析
. 现代农业科技 , (16 ), 19 -24 .]
[本文引用: 1]
[7]
Crossman ND Bryan BA Cooke DA 2011 ). An invasive plant and climate change threat index for weed risk management: Integrating habitat distribution pattern and dispersal process.
Ecological Indicators , 11 , 183 -198 .
[本文引用: 1]
[8]
Daehler CC 2003 ). Performance’s comparisons of co-occurr- ing native and alien invasive plants: Implications for conservation and restoration.
Annual Review of Ecology and Systematics , 34 , 183 -211 .
[本文引用: 1]
[9]
Ding H Xu HG Liu ZL 2007 ). Impacts of invasion of Eupatorium adenophorum on vegetation diversity.
Journal of Ecology and Rural Environment , 23 (2 ), 29 -32, 75 (in Chinese with English abstract).
[丁晖 , 徐海根 , 刘志磊 (2007 ). 外来入侵植物紫茎泽兰对植物多样性的影响
. 生态与农村环境学报 , 23 (2 ), 29 -32, 75 .]
[10]
Durand LZ Goldstein G 2001 ). Photosynthesis, photo inhibition, and nitrogen use efficiency in native and invasive tree ferns in Hawaii.
Oecologia , 126 , 345 -354 .
[本文引用: 1]
[11]
Erik W 2004 ). Constraints in range predictions of invasive plant species due to non-equilibrium distribution patterns: Purple loosestrife (Lythrum salicaria) in North America.
Ecological Modelling , 179 , 551 -567 .
[本文引用: 2]
[12]
Fridley JD Stachowicz JJ Naeem S Sax DF Seabloom EW Smith MD Stohlgren TJ Tilman D Von Holle B 2007 ). The invasion paradox: reconciling pattern and process in species invasions.
Ecology , 88 , 3 -17 .
[本文引用: 1]
[13]
Gai YQ Li XB Zhang L Li C Liang Y 2009 ). Land use/cover change and vegetation coverage monitoring by remote sensing: A case study of Miyun, Beijing.
Resources Science , 31 , 523 -529 (in Chinese with English abstract).
[本文引用: 1]
[盖永芹 , 李晓兵 , 张立 , 李超 , 梁燕 (2009 ). 土地利用/覆被变化与植被盖度的遥感监测——以北京市密云县为例
. 资源科学 , 31 , 523 -529 .]
[本文引用: 1]
[14]
Gozlan RE Newton AC 2009 ). Biological invasions: Benefits versus risks.
Science , 324 , 1015 -1016 .
[本文引用: 1]
[15]
Hejda M Pyšek P Jarošík V 2009 ). Impact of invasive plants on the species richness, diversity and composition of invaded communities.
Journal of Ecology , 97 , 393 -403 .
[本文引用: 1]
[16]
Hobbs RJ 1991 ). Disturbance―A precursor to weed invasion in native vegetation.
Plant Protection Quarterly , 6 , 99 -104 .
[本文引用: 1]
[17]
Hobbs RJ Huenneke LF 1992 ). Disturbance, diversity, and invasion: Implications for conservation.
Conservation Biology , 6 , 324 -337 .
[本文引用: 1]
[18]
Honnay O Verheyen K Hermy M 2002 ). Permeability of ancient forest edges for weedy plant species invasion.
Forest Ecology and Management , 161 , 109 -122 .
[本文引用: 1]
[19]
Huang QQ Shen YD Li XX Fan ZW Li MG Cheng HT 2013 ). Native expanding Merremia boisiana is not more allelopathic than its non-expanding congener M. vitifolia in the expanded range in Hainan.
American Journal of Plant Sciences , 4 , 774 -779 .
[本文引用: 1]
[20]
Huang QQ Shen YD Fan ZW Li XX Song X Cheng HT Hou YP 2013 ). Effects of soil from different forest types in Wuzhi Mountain on the seedling growth of Merremia boisiana.
Ecology and Environmental Sciences , 22 , 95 -99 (in Chinese with English abstract).
[本文引用: 1]
[黄乔乔 , 沈奕德 , 范志伟 , 李晓霞 , 宋鑫 , 程汉亭 , 侯玉平 (2013 ). 五指山不同林型土壤对金钟藤幼苗生长的影响
. 生态环境学报 , 22 , 95 -99 .]
[本文引用: 1]
[21]
Huston MA 1997 ). Hidden treatments in ecological experiments: Re-evaluating the ecosystem function of biodiversity.
Oecologia , 110 , 449 -460 .
[22]
Ibáñez I Silander Jr JA Wilson AM LaFleur N Tanaka N Tsuyama I 2009 ). Multivariate forecasts of potential distributions of invasive plant species.
Ecological Applications , 19 , 359 -375 .
[本文引用: 1]
[23]
Li AD Xie YG Zhang JL Long XQ 2013 ). Impacts of the invasion of Alternathera philoxeroides on species composition and diversity of heterogeneous habitat in Caohai wetland.
Ecology and Environmental Sciences , 22 , 1322 -1328 (in Chinese with English abstract).
[李安定 , 谢元贵 , 张建利 , 龙秀琴 (2013 ). 异质生境空心莲子草植物群落组成及物种多样性研究
. 生态环境学报 , 22 , 1322 -1328 .]
[24]
Li B Li YY 2009 ). The Interaction between invasive and native species in the biological invasion.
Sichuan Environment , 28 , 64 -67 (in Chinese with English abstract).
[本文引用: 1]
[李冰 , 李玉瑛 (2009 ). 生物入侵中入侵种与土著种的相互作用
. 四川环境 , 28 , 64 -67 .]
[本文引用: 1]
[25]
Li JR Sun BP Ling X Guo JY Wang TS Zhou WY Zhao Y Zhou XS 2010 ). Study on land use and dynamic changes of vegetation cover in and after the project of returning farmland to forests―taking the Ansai County as example.
Journal of Inner Mongolia Agricultural University , 31 , 130 -135 (in Chinese with English abstract) .
[本文引用: 1]
[李锦荣 , 孙保平 , 凌侠 , 郭建英 , 王同顺 , 周文渊 , 赵岩 , 周湘山 (2010 ). 退耕还林工程前后土地利用与植被盖度动态变化研究——以安塞县为例
. 内蒙古农业大学学报 , 31 , 130 -135 .]
[本文引用: 1]
[26]
Li ZY Xie Y 2002). Invasive Alien Species in China . China Forestry Publishing House, Beijing (in Chinese).
[本文引用: 1]
[李振宇 , 解焱 (2002 ). 中国外来入侵种 . 中国林业出版社 , 北京 .]
[本文引用: 1]
[27]
Liancourt P Viard-Crétat F Michalet R 2009 ). Contrasting community responses to fertilization and the role of the competitive ability of dominant species.
Journal of Vegetation Science , 20 , 138 -147 .
[本文引用: 1]
[28]
Liu J Li JM Yu H He WM Yu FH Sang WG Liu GF Dong M 2010 ). The relationship between functional traits and invasiveness of alien plants.
Biodiversity Science , 18 , 569 -576 (in Chinese with English abstract).
[本文引用: 1]
[刘建 , 李钧敏 , 余华 , 何维明 , 于飞海 , 桑卫国 , 刘国方 , 董鸣 (2010 ). 植物功能性状与外来植物入侵
. 生物多样性 , 18 , 569 -576 .]
[本文引用: 1]
[29]
Losdale WM 1999 ). Global patterns of plant invasions and the concept of invasibility.
Ecology , 80 , 1522 -1536 .
[30]
Newsome AE Noble IR 1986 ). Ecological and physiological characters of invading species. In: Groves RH, Burdon JJ eds. Ecology of Biological Invasions . Cambridge University Press , Cambridge, UK . 1 -20 .
[本文引用: 1]
[31]
Parendes LA Jones JA 2000 ). Role of light availability and dispersal in exotic plant invasion along roads and streams in the H. J. Andrews experimental forest, Oregon.
Conservation Biology , 14 , 64 -75 .
[本文引用: 1]
[32]
Peng ZB Jiang Y Jiang JS 2013 ). Risk evaluation indicator system for exotic plant invasion in Hainan Island, South China.
Chinese Journal of Ecology , 32 , 2029 -2034 (in Chinese with English abstract).
[本文引用: 1]
[彭宗波 , 蒋英 , 蒋菊生 (2013 ). 海南岛外来植物入侵风险评价指标体系
. 生态学杂志 , 32 , 2029 -2034 .]
[本文引用: 1]
[33]
Qin XS Zhang RJ Chen HF Yan YH Zheng XL Xing FW 2008 ). Alien plants in limestone regions of Hainan Island, China.
Chinese Journal of Ecology , 27 , 1861 -1868 (in Chinese with English abstract).
[本文引用: 1]
[秦新生 , 张荣京 , 陈红锋 , 严岳鸿 , 郑希龙 , 邢福武 (2008 ). 海南岛石灰岩地区的外来植物
. 生态学杂志 , 27 , 1861 -1868 .]
[本文引用: 1]
[34]
Rejmánek M Richardson DM 1996 ). What attributes make some plant species more invasive?
Ecology , 77 , 1655 -1661 .
[本文引用: 1]
[35]
Shan JL 2003 ). Preliminary studies on exotic plant communities in Hainan.
Chinese Journal of Tropical Agriculture , 23 (3 ), 1 -4 (in Chinese with English abstract).
[本文引用: 1]
[单家林 (2003 ). 海南岛外来植物群落初探
. 热带农业科学 , 23 (3 ), 1 -4 .]
[本文引用: 1]
[36]
Shea K Chesson P 2002 ). Community ecology theory as a framework for biological invasions.
Trends in Ecology & Evolution , 17 , 170 -176 .
[本文引用: 1]
[37]
Simberloff D Relva MA Nuñez MA 2003 ). Introduced species and management of a Nothofagus/Austrocedrus forest.
Environmental Management , 31 , 263 -275 .
[本文引用: 1]
[38]
Sokal RR Rohlf FJ 1995 ). Biometry. 3rd edn
. Freeman, New York. 23 -28 .
[本文引用: 1]
[39]
Stachowicz JJ Tilman D Species invasions and the relationships between species diversity, community saturation, and ecosystem functioning. In: Sax DF, Stachowicz JJ, Gaines SD eds. Species Invasions: Insights into Ecology, Evolution, and Biogeography . Sinauer Associates : , Berlin .
[本文引用: 1]
[40]
Stohlgren TJ Binkley D Chong GW Kalkhan MA Schell LD Bull KA Otsuki Y Newman G Bashkin M Son Y 1999 ). Exotic plant species invade hot spots of native plant diversity.
Ecological Monographs , 69 , 25 -46 .
[本文引用: 1]
[41]
Tang SX Zhao ZZ Bi H Xie GZ 2008 ). Climate resources in Hainan. Journal of Hainan Normal University (
Natural Science ), 21 , 343 -346 (in Chinese with English abstract).
[本文引用: 1]
[唐少霞 , 赵志忠 , 毕华 , 谢跟踪 (2008 ). 海南岛气候资源特征及其开发利用
. 海南师范大学学报 (自然科学版) , 21 , 343 -346 .]
[本文引用: 1]
[42]
Tilman D 1997 ). Community invisibility, recruitment limitation and grassland biodiversity.
Ecology , 78 , 81 -92 .
[43]
Wang LL 2013 ). Effects of the New Countryside Construction on Synarthropic Plant Diversity: A Case Study of Yaodu District of Linfen City
. Master degree dissertation, Shanxi Normal University , Linfen . 1 -6 (in Chinese with English abstract).
[王丽丽 (2013 ). 新农村建设对伴人植物多样性的影响——以临汾市尧都区为例
. 硕士学位论文, 山西师范大学, 临汾 . 1 -6 .]
[44]
Wang SM Zhang N Yu LQ Zhao RH Hao P Li JW Jiang YS Sha HF Liu Y Zhang ZX 2012 ). Distribution pattern and their influencing factors of invasive alien plants in Beijing.
Acta Ecologica Sinica , 32 , 4619 -4628 (in Chinese with English abstract).
[王苏铭 , 张楠 , 于琳倩 , 赵容慧 , 郝鹏 , 李景文 , 姜英淑 , 沙海峰 , 刘义 , 张志翔 (2012 ). 北京地区外来入侵植物分布特征及其影响因素
. 生态学报 , 32 , 4619 -4628 .]
[45]
Weber E Sun SG Li B 2008 ). Invasive alien plants in China: Diversity and ecological insights.
Biological Invasions , 10 , 1411 -1429 .
[本文引用: 1]
[46]
Weis JS Weis P 2003 ). Is the invasion of the common reed, Phragmites australis, into tidal marshes of the Eastern US an ecological disaster?
Marine Pollution Bulletin , 46 , 816 -820 .
[本文引用: 1]
[47]
Williamson M Can the impact of invasive species be predicted? In: Groves RH, Panetta FD, Virtue JG eds . Weed Risk Assessment . CSIRO, Canberra . 20 -33 .
[本文引用: 1]
[48]
Wu XW Luo J Chen JK Li B 2006 ). Spatial patterns of invasive alien plants in China and its relationship with environmental and antropological factors. Journal of Plant Ecology (formerly Acta Phytoecologica Sinica)
, 30 , 576 -584 (in Chinese with English abstract).
[本文引用: 1]
[吴晓雯 , 罗晶 , 陈家宽 , 李博 (2006 ). 中国外来入侵植物的分布格局及其与环境因子和人类活动的关系
. 植物生态学报 , 30 , 576 -584 .]
[本文引用: 1]
[49]
Yang XB Chen YK Li DH Long WX 2013 ). A Checklist of Plants in Hainan . Science Press , Beijing .(in Chinese)
[本文引用: 1]
[杨小波 , 陈玉凯 , 李东海 , 龙文兴 (2013 ). 海南植物名录 . 科学出版社 , 北京 .]
[本文引用: 1]
[50]
Zhang S Guo SL Guan M Yin LP Zhang RX 2010 ). Diversity differentiation of invasive plants at a regional scale in China and its influencing factors: According to analyses on the data from 74 regions.
Acta Ecologica Sinica , 30 , 4241 -4256 .(in Chinese with English abstract)
[本文引用: 1]
[张帅 , 郭水良 , 管铭 , 印丽萍 , 张若轩 (2010 ). 我国入侵植物多样性的区域分异及其影响因素——以74个地区数据为基础
. 生态学报 , 30 , 4241 -4256 .]
[本文引用: 1]
[51]
Zhong JD Xu YM Zeng FH Liu WG Chen Y Liu JX 2014 ). Analysis on the growth characteristics of Eupatorium catarium in different types of habitats.
Guihaia , 34 , 68 -73 .(in Chinese with English abstract)
[钟军弟 , 徐意媚 , 曾富华 , 刘晚苟 , 陈燕 , 刘金祥 (2014 ). 不同生境下假臭草生长特征分析
. 广西植物 , 34 , 68 -73 .]
Revealing historic invasion patterns and potential invasion sites for two non-native plant species.
0
2008
Leaf construction cost, nutrient concentration, and net CO2 assimilation of native and invasive species in Hawaii.
1
1999
... 本文研究表明: 弃耕地、村落、农田生态系统中所含的入侵种数最多, 均有10种; 而在林缘生境中, 11种入侵植物的入侵频率和累计入侵频率都较低, 在抽样调查的次生林内, 没有发现入侵植物的分布(表1 ), 说明入侵植物主要分布在人为干扰强度大的生境中, 而在人为干扰强度小、群落结构复杂的生境中分布较少.结合6种生态系统的演替阶段分析, 弃耕地、村落、草地和农田均属于演替早期的生境类型, 受人为干扰的强度大, 而机械和人为活动的介导能有效地减少入侵植物的传播障碍, 或形成干扰间的协同效应(Hobbs, 1991 ); 入侵植物通常能够迅速响应环境中的干扰, 并且具有较强的表型可塑性, 这些特征促成了其在干扰生境中的成功入侵(Hobbs & Huenneke, 1992 ).植物对光能的捕获和利用效率高低是决定其能否适应环境并很好地存活下去的重要因素Baruch & Goldstein, 1999 ; Durand & Goldstein, 2001 ), 因此, 入侵生境的光环境对植物的入侵过程有很大影响, 如弃耕地通常具有高光照、高肥力的环境特征, 这为入侵种群的建立和发展提供了条件, 导致弃耕地常成为入侵植物(如假臭草、三裂叶蟛蜞菊等)集中分布的生境, 这与钟军弟等(2014)在广西的研究结果一致, 表明弃耕地具有较大的可入侵空间.林缘和天然次生林生态系统属于演替中后期的生境类型, 与演替前期的生境类型相比, 受人为干扰强度较低, 而增加的物种密度也将减少群落中的空缺生态位, 从而降低植物成功入侵的几率Stachowicz & Tilman, 2005 ).另一方面, 林缘或林下的光照强度和光合有效辐射都比较低, 这种环境特征也将限制植物入侵林缘的深度Brothers & Spingarn, 1992 ; Shea & Chesson, 2002 ), 因为本文研究的入侵植物多是处于演替初期的草本植物, 这些植物从种子萌发到幼苗生长及种群的发育均需要充足的光照, 它们的入侵可能在演替前期的生境类型, 如农田、草地和村落中具有更好的生态适应性; 但在林缘和林下的入侵却受到光照条件的制约, 且林缘的群落结构对低矮生活型的入侵植物也具有阻碍作用(Honnay et al ., 2002 ), 如喜旱莲子草、含羞草等在林缘中的入侵频率都很低.因此, 入侵植物虽然在林区广泛分布, 但它们仅分布在光照充足的路边或新垦的裸地上, 而无法入侵到森林内部, 国外的不少报道也证实了这种分布特点Parendes & Jones, 2000 ).分析入侵种群的盖度可知, 除三裂叶蟛蜞菊的种群盖度较高外, 其余5种入侵植物的盖度都集中分布在1%-20%之间, 并与入侵地的环境特征存在相关性, 如含羞草在农田、假臭草在弃耕地、三裂叶蟛蜞菊在种植园中通常具有成群分布生长的特点, 体现出这3种入侵植物在对应的生境中有更好的生态适应性; 而在林缘生态系统中, 除马缨丹和苏门白酒草外, 其余4种入侵植物的入侵频率与种群盖度呈负相关关系, 表明在林缘生态系统中, 入侵植物在入侵群落中的密度小, 难以形成单优群落, 这与入侵植物的种子是否能到达林缘, 能否存活及能否萌芽有密切的关系, 这些问题有待进一步实验验证. ...
Forest fragmentation and alien plant invasion of central Indiana old-growth forests.
1
1992
... 本文研究表明: 弃耕地、村落、农田生态系统中所含的入侵种数最多, 均有10种; 而在林缘生境中, 11种入侵植物的入侵频率和累计入侵频率都较低, 在抽样调查的次生林内, 没有发现入侵植物的分布(表1 ), 说明入侵植物主要分布在人为干扰强度大的生境中, 而在人为干扰强度小、群落结构复杂的生境中分布较少.结合6种生态系统的演替阶段分析, 弃耕地、村落、草地和农田均属于演替早期的生境类型, 受人为干扰的强度大, 而机械和人为活动的介导能有效地减少入侵植物的传播障碍, 或形成干扰间的协同效应(Hobbs, 1991 ); 入侵植物通常能够迅速响应环境中的干扰, 并且具有较强的表型可塑性, 这些特征促成了其在干扰生境中的成功入侵(Hobbs & Huenneke, 1992 ).植物对光能的捕获和利用效率高低是决定其能否适应环境并很好地存活下去的重要因素Baruch & Goldstein, 1999 ; Durand & Goldstein, 2001 ), 因此, 入侵生境的光环境对植物的入侵过程有很大影响, 如弃耕地通常具有高光照、高肥力的环境特征, 这为入侵种群的建立和发展提供了条件, 导致弃耕地常成为入侵植物(如假臭草、三裂叶蟛蜞菊等)集中分布的生境, 这与钟军弟等(2014)在广西的研究结果一致, 表明弃耕地具有较大的可入侵空间.林缘和天然次生林生态系统属于演替中后期的生境类型, 与演替前期的生境类型相比, 受人为干扰强度较低, 而增加的物种密度也将减少群落中的空缺生态位, 从而降低植物成功入侵的几率Stachowicz & Tilman, 2005 ).另一方面, 林缘或林下的光照强度和光合有效辐射都比较低, 这种环境特征也将限制植物入侵林缘的深度Brothers & Spingarn, 1992 ; Shea & Chesson, 2002 ), 因为本文研究的入侵植物多是处于演替初期的草本植物, 这些植物从种子萌发到幼苗生长及种群的发育均需要充足的光照, 它们的入侵可能在演替前期的生境类型, 如农田、草地和村落中具有更好的生态适应性; 但在林缘和林下的入侵却受到光照条件的制约, 且林缘的群落结构对低矮生活型的入侵植物也具有阻碍作用(Honnay et al ., 2002 ), 如喜旱莲子草、含羞草等在林缘中的入侵频率都很低.因此, 入侵植物虽然在林区广泛分布, 但它们仅分布在光照充足的路边或新垦的裸地上, 而无法入侵到森林内部, 国外的不少报道也证实了这种分布特点Parendes & Jones, 2000 ).分析入侵种群的盖度可知, 除三裂叶蟛蜞菊的种群盖度较高外, 其余5种入侵植物的盖度都集中分布在1%-20%之间, 并与入侵地的环境特征存在相关性, 如含羞草在农田、假臭草在弃耕地、三裂叶蟛蜞菊在种植园中通常具有成群分布生长的特点, 体现出这3种入侵植物在对应的生境中有更好的生态适应性; 而在林缘生态系统中, 除马缨丹和苏门白酒草外, 其余4种入侵植物的入侵频率与种群盖度呈负相关关系, 表明在林缘生态系统中, 入侵植物在入侵群落中的密度小, 难以形成单优群落, 这与入侵植物的种子是否能到达林缘, 能否存活及能否萌芽有密切的关系, 这些问题有待进一步实验验证. ...
Physical habitat attribute mediates biotic resistant to non-indigenous species invasion.
2
2002
... 外来物种入侵已成为严重危害陆地及海洋环境的生态问题, 被认为是一种生物污染而被广泛关注和研究(Weis & Weis, 2003 ).深入了解外来物种的入侵机制, 并进行有效的预测、预防和管理是减少入侵生物危害的最有效手段(Byers, 2002 ), 而从区域尺度上分析入侵物种的分布特征并探索其形成原因是认识生物入侵问题的重要基础(Erik, 2004 ). ...
... 有关入侵植物影响生态系统功能的报道有很多, 并多关注其负面效应Williamson, 2001 ; Byers et al ., 2002 ; Simberloff et al ., 2003 ), 认为植物入侵导致本地群落中生物多样性的下降李冰和李玉瑛, 2009 ), 例如: 丁晖等(2007)研究发现, 入侵森林林隙中的紫茎泽兰(Ageratina adenophora )使群落中本地植物的物种丰富度显著下降; Fox等在澳洲西部一个草地生态系统中的研究表明群落中的外来物种比例与本地植物种数呈负相关关系(Stohlgren et al ., 1999 ); Tilman (1997)在美国Bethel市一个草地生态系统中的实验也得出了一致的结论.因此一般认为, 外来植物成功入侵后将抑制群落中本地植物的多样性.但也有研究发现: 入侵植物对群落植物多样性的影响与入侵种及本地植物的生活型有关(Hejda et al ., 2009 ); 植物入侵的生态效应与入侵地的演替阶段有关, 例如: 豆科植物入侵生态环境恶劣的尾矿区后, 能有效地改善土壤肥力, 为演替中期过渡种的定居和发育创造良好的条件, 并促进物种多样性的增加Gozlan & Newton, 2009 ); 李安定等(2013)研究发现, 喜旱莲子草(空心莲子草)(Alternanthera philoxeroides )在入侵的早期阶段能改变群落生境条件, 进而促进其他物种的生存与定居, 并增加物种的多样性, 而只有当其种群盖度持续增加后, 才可能会威胁其他物种的生存; Hejda等(2009)研究发现, 入侵植物的盖度是决定群落物种丰富度和均匀度的主要因子, 入侵植物对物种多样性的影响与本地植物优势种的盖度、高度有密切关系.因此, 植物入侵的生态效应受生态系统类型、群落演替阶段的影响; 不同功能群植物对植物入侵的响应也有差异.本文通过研究7种生态系统类型中入侵植物的入侵特征及其对本地植物的影响, 旨在检验以下科学假设: 1)植物入侵的生态效应与入侵地的环境特征和演替阶段有关, 入侵植物可能对入侵地产生正面或负面影响; 2)植物入侵主要影响群落中某一功能群的物种多样性, 而不是影响全部群落个体; 3)多数入侵植物有偏好的入侵生境, 而人为干扰少的生态系统类型对植物入侵有抵制作用. ...
Directing research to reduce the impacts of nonindigenous species.
0
2002
海南岛旅游气候资源分析
1
2010
... 入侵植物在海南岛分布广泛, 且不同区域间种数相差不大, 但整体呈现为东北部、西南部地区种类较多, 东南部次之, 而中部较少的分布特征.入侵植物的分布主要取决于环境的可入侵性和物种的入侵能力两个方面Newsome & Noble, 1986 ; 刘建等, 2010 ).如“理想”的入侵植物通常具备生活史短, 自交亲和性、结实率高, 可进行无性繁殖等特点, 并能迅速响应环境中的干扰Rejmánek & Richardson, 1996 ).本文研究的11种入侵植物中, 除马缨丹外, 其余10种均为一、二年生的草本植物, 其中有6种为菊科植物, 如飞机草、假臭草、微甘菊等, 这些植物产生的种子数量大而质量轻, 通常还附有冠毛, 便于长距离传播和扩散李振宇和谢焱, 2002 ; Weber et al ., 2008 ), 而喜旱莲子草、微甘菊、凤眼蓝、三裂叶蟛蜞菊具有无性繁殖的特性, 其繁殖体可以借助水流进行传播, 这些生物学特征和传播机制增强了其入侵能力, 有助于入侵植物的扩散和迅速占据入侵地, 并能在较短的时间内扩大种群, 从而导致其在海南岛的分布范围很广.但对比不同区域的分布特点发现, 东北部和西南部的入侵种数相对较多, 为10-11种, 而中部和东南部分布较少, 这种差异可能与不同区域间的气候条件和人为干扰强度有关.由于海南岛的西部地区贯穿有环西粤海铁路, 具有较长的运输史, 而连接该地区的海口市、儋州市、东方市都是水陆交通的重要枢纽, 境内人流密集, 干扰强度大, 这些因素都能有效地促进入侵植物的传播和扩散(吴晓雯等, 2006 ; 张帅等, 2010 ).结合气候条件进一步分析, 海南岛西部地区的年晴天数平均为300天, 明显高于中部和东部地区, 但降水量较少, 而中部地区是全岛降水集中分布的地区, 年降水量达2000 mm, 属于湿润地区.此外, 中部地区的年平均气温为23 ℃左右, 处于全岛平均气温(23-26 ℃)的下限, 低于海南岛其他地区的温度(蔡大鑫等, 2010 ).综合气候条件和入侵种数的分布特征发现, 入侵植物在年平均气温较高、降水量较低、而日照时间较长的东北、西南部地区分布的种类较多, 在年平均气温较低、降水量较高、日照时间较短的中部、东部地区分布的种类较少, 这可能与入侵植物的生活习性有关, 因为本文研究的入侵植物多为草本阳性植物, 它们的种子萌发和种群发展都需要足够的光温条件, 过高湿和低温的环境可能对其生长和扩散具有抑制作用. ...
海南岛旅游气候资源分析
1
2010
... 入侵植物在海南岛分布广泛, 且不同区域间种数相差不大, 但整体呈现为东北部、西南部地区种类较多, 东南部次之, 而中部较少的分布特征.入侵植物的分布主要取决于环境的可入侵性和物种的入侵能力两个方面Newsome & Noble, 1986 ; 刘建等, 2010 ).如“理想”的入侵植物通常具备生活史短, 自交亲和性、结实率高, 可进行无性繁殖等特点, 并能迅速响应环境中的干扰Rejmánek & Richardson, 1996 ).本文研究的11种入侵植物中, 除马缨丹外, 其余10种均为一、二年生的草本植物, 其中有6种为菊科植物, 如飞机草、假臭草、微甘菊等, 这些植物产生的种子数量大而质量轻, 通常还附有冠毛, 便于长距离传播和扩散李振宇和谢焱, 2002 ; Weber et al ., 2008 ), 而喜旱莲子草、微甘菊、凤眼蓝、三裂叶蟛蜞菊具有无性繁殖的特性, 其繁殖体可以借助水流进行传播, 这些生物学特征和传播机制增强了其入侵能力, 有助于入侵植物的扩散和迅速占据入侵地, 并能在较短的时间内扩大种群, 从而导致其在海南岛的分布范围很广.但对比不同区域的分布特点发现, 东北部和西南部的入侵种数相对较多, 为10-11种, 而中部和东南部分布较少, 这种差异可能与不同区域间的气候条件和人为干扰强度有关.由于海南岛的西部地区贯穿有环西粤海铁路, 具有较长的运输史, 而连接该地区的海口市、儋州市、东方市都是水陆交通的重要枢纽, 境内人流密集, 干扰强度大, 这些因素都能有效地促进入侵植物的传播和扩散(吴晓雯等, 2006 ; 张帅等, 2010 ).结合气候条件进一步分析, 海南岛西部地区的年晴天数平均为300天, 明显高于中部和东部地区, 但降水量较少, 而中部地区是全岛降水集中分布的地区, 年降水量达2000 mm, 属于湿润地区.此外, 中部地区的年平均气温为23 ℃左右, 处于全岛平均气温(23-26 ℃)的下限, 低于海南岛其他地区的温度(蔡大鑫等, 2010 ).综合气候条件和入侵种数的分布特征发现, 入侵植物在年平均气温较高、降水量较低、而日照时间较长的东北、西南部地区分布的种类较多, 在年平均气温较低、降水量较高、日照时间较短的中部、东部地区分布的种类较少, 这可能与入侵植物的生活习性有关, 因为本文研究的入侵植物多为草本阳性植物, 它们的种子萌发和种群发展都需要足够的光温条件, 过高湿和低温的环境可能对其生长和扩散具有抑制作用. ...
An invasive plant and climate change threat index for weed risk management: Integrating habitat distribution pattern and dispersal process.
1
2011
... 针对入侵植物的分布问题, Losdale (1999)研究了全球入侵植物的分布特征, 并发现温带地区的耕地、城镇和生物多样性热点地区更易遭受外来植物的入侵, 而荒漠、稀树草原等生态系统的可入侵空间较小; Barney等(2008)研究了两种不同入侵历史的外来植物在北美的分布和扩散格局, 发现其入侵范围均随着时间的推移而扩大; 此外, 还有不少学者运用气候模拟技术对入侵植物的潜在分布区进行了预测(Erik, 2004 ; Ibáñez et al ., 2009 ; Cross- man et al ., 2011 ); 而国内的研究多集中在入侵植物多样性的区域分异及其影响因素等方面, 如吴晓雯等(2006)研究了入侵植物在我国的分布格局, 发现其物种数从南到北逐渐减少, 而物种密度由东南海岸向内陆递减的分布特征, 并推测中国的东南部地区将更容易遭受外来植物入侵; 王苏铭等(2012)研究了入侵植物在北京地区的分布特征; 张帅等(2010)研究了入侵植物在我国74个地理单元的分布情况, 发现入侵植物的分布具有明显的区域分化现象, 而降水与温度是影响其空间分布格局的主要 因素. ...
Performance’s comparisons of co-occurr- ing native and alien invasive plants: Implications for conservation and restoration.
1
2003
... 入侵植物导致本地植物多样性降低的现象已被广泛认识, 但在不同生态系统中可能存在差异(Daehler, 2003 ).本文的研究结果显示, 样方中入侵植物种数与本地植物平均种数在弃耕地、种植园和林缘3种生态系统中无明显相关性, 当样方中入侵植物种数由0到4种变化时, 群落中本地植物平均种数均没有明显的递变规律; 进一步分析不同生活型植物与入侵植物种数变化的关系发现, 仅弃耕地中草本植物的平均种数略有上升的趋势, 但差异不显著(图3C ); 而乔木、灌木、藤本生活型植物的平均种数与入侵植物的种数变化不相关.在农田生态系统中, 本地植物平均种数随入侵植物种数的增加呈先增加后减少的变化趋势, 其中当入侵植物种数为1时, 本地植物平均种数最多, 随后呈减少的趋势(图3A ), 表明群落中少数入侵种的存在具有促进本地植物多样性的作用, 其原因可能是早期的植物入侵过程改善了群落环境并促进了本地种的生存和定居, 从而增加了本地植物的物种丰富度, 但持续扩大的入侵种群将会抑制本地植物的多样性, 这与李安定等(2013)对莲子草的研究结果是一致的.本文的研究结果也表明: 针对不同的生态系统类型和不同的入侵阶段, 植物入侵对本地植物的影响存在差异; 而一定强度的植物入侵可能具有正面的生态效应. ...
外来入侵植物紫茎泽兰对植物多样性的影响
0
2007
外来入侵植物紫茎泽兰对植物多样性的影响
0
2007
Photosynthesis, photo inhibition, and nitrogen use efficiency in native and invasive tree ferns in Hawaii.
1
2001
... 本文研究表明: 弃耕地、村落、农田生态系统中所含的入侵种数最多, 均有10种; 而在林缘生境中, 11种入侵植物的入侵频率和累计入侵频率都较低, 在抽样调查的次生林内, 没有发现入侵植物的分布(表1 ), 说明入侵植物主要分布在人为干扰强度大的生境中, 而在人为干扰强度小、群落结构复杂的生境中分布较少.结合6种生态系统的演替阶段分析, 弃耕地、村落、草地和农田均属于演替早期的生境类型, 受人为干扰的强度大, 而机械和人为活动的介导能有效地减少入侵植物的传播障碍, 或形成干扰间的协同效应(Hobbs, 1991 ); 入侵植物通常能够迅速响应环境中的干扰, 并且具有较强的表型可塑性, 这些特征促成了其在干扰生境中的成功入侵(Hobbs & Huenneke, 1992 ).植物对光能的捕获和利用效率高低是决定其能否适应环境并很好地存活下去的重要因素Baruch & Goldstein, 1999 ; Durand & Goldstein, 2001 ), 因此, 入侵生境的光环境对植物的入侵过程有很大影响, 如弃耕地通常具有高光照、高肥力的环境特征, 这为入侵种群的建立和发展提供了条件, 导致弃耕地常成为入侵植物(如假臭草、三裂叶蟛蜞菊等)集中分布的生境, 这与钟军弟等(2014)在广西的研究结果一致, 表明弃耕地具有较大的可入侵空间.林缘和天然次生林生态系统属于演替中后期的生境类型, 与演替前期的生境类型相比, 受人为干扰强度较低, 而增加的物种密度也将减少群落中的空缺生态位, 从而降低植物成功入侵的几率Stachowicz & Tilman, 2005 ).另一方面, 林缘或林下的光照强度和光合有效辐射都比较低, 这种环境特征也将限制植物入侵林缘的深度Brothers & Spingarn, 1992 ; Shea & Chesson, 2002 ), 因为本文研究的入侵植物多是处于演替初期的草本植物, 这些植物从种子萌发到幼苗生长及种群的发育均需要充足的光照, 它们的入侵可能在演替前期的生境类型, 如农田、草地和村落中具有更好的生态适应性; 但在林缘和林下的入侵却受到光照条件的制约, 且林缘的群落结构对低矮生活型的入侵植物也具有阻碍作用(Honnay et al ., 2002 ), 如喜旱莲子草、含羞草等在林缘中的入侵频率都很低.因此, 入侵植物虽然在林区广泛分布, 但它们仅分布在光照充足的路边或新垦的裸地上, 而无法入侵到森林内部, 国外的不少报道也证实了这种分布特点Parendes & Jones, 2000 ).分析入侵种群的盖度可知, 除三裂叶蟛蜞菊的种群盖度较高外, 其余5种入侵植物的盖度都集中分布在1%-20%之间, 并与入侵地的环境特征存在相关性, 如含羞草在农田、假臭草在弃耕地、三裂叶蟛蜞菊在种植园中通常具有成群分布生长的特点, 体现出这3种入侵植物在对应的生境中有更好的生态适应性; 而在林缘生态系统中, 除马缨丹和苏门白酒草外, 其余4种入侵植物的入侵频率与种群盖度呈负相关关系, 表明在林缘生态系统中, 入侵植物在入侵群落中的密度小, 难以形成单优群落, 这与入侵植物的种子是否能到达林缘, 能否存活及能否萌芽有密切的关系, 这些问题有待进一步实验验证. ...
Constraints in range predictions of invasive plant species due to non-equilibrium distribution patterns: Purple loosestrife (Lythrum salicaria) in North America.
2
2004
... 外来物种入侵已成为严重危害陆地及海洋环境的生态问题, 被认为是一种生物污染而被广泛关注和研究(Weis & Weis, 2003 ).深入了解外来物种的入侵机制, 并进行有效的预测、预防和管理是减少入侵生物危害的最有效手段(Byers, 2002 ), 而从区域尺度上分析入侵物种的分布特征并探索其形成原因是认识生物入侵问题的重要基础(Erik, 2004 ). ...
... 针对入侵植物的分布问题, Losdale (1999)研究了全球入侵植物的分布特征, 并发现温带地区的耕地、城镇和生物多样性热点地区更易遭受外来植物的入侵, 而荒漠、稀树草原等生态系统的可入侵空间较小; Barney等(2008)研究了两种不同入侵历史的外来植物在北美的分布和扩散格局, 发现其入侵范围均随着时间的推移而扩大; 此外, 还有不少学者运用气候模拟技术对入侵植物的潜在分布区进行了预测(Erik, 2004 ; Ibáñez et al ., 2009 ; Cross- man et al ., 2011 ); 而国内的研究多集中在入侵植物多样性的区域分异及其影响因素等方面, 如吴晓雯等(2006)研究了入侵植物在我国的分布格局, 发现其物种数从南到北逐渐减少, 而物种密度由东南海岸向内陆递减的分布特征, 并推测中国的东南部地区将更容易遭受外来植物入侵; 王苏铭等(2012)研究了入侵植物在北京地区的分布特征; 张帅等(2010)研究了入侵植物在我国74个地理单元的分布情况, 发现入侵植物的分布具有明显的区域分化现象, 而降水与温度是影响其空间分布格局的主要 因素. ...
The invasion paradox: reconciling pattern and process in species invasions.
1
2007
... Stohlgren等(1999)对草地生态系统的研究表明, 在群落水平上, 本地植物种数与入侵植物种数呈正相关关系, 并认为这与两类植物对草地环境的变化有相同的响应有关.而本文发现, 草地群落中本地植物平均种数随入侵种数的增加呈先减少后增加的趋势, 且在入侵种数为2时最少, 草本植物也呈一致的变化趋势, 而乔木、灌木和藤本植物的种数变化不显著, 其原因可能是当入侵种数较少时, 某些入侵植物排挤本地植物后形成了单优群落, 并导致本地植物物种丰富度下降, 而随着入侵种数的增加, 新入侵的植物改变了优势种的组成, 这种变化进一步促进了新的本地植物迁入, 从而增加了本地植物的多样性, 因为群落优势种对新物种的迁入具有决定性作用(Liancourt et al ., 2009 ).Huston (1997)的研究也表明, 群落的可入侵性更多取决于关键种, 而并非整个群落中的物种数, 但在较小尺度上取样时, 物种多样性与入侵关系的模式比较复杂, 群落类型和地域的差异都会导致不同的结果, 还需要考虑其他因素的影响(Fridley et al ., 2007 ).针对本文研究的10种陆生入侵植物, 在入侵植物组合不同的群落中, 具体是哪一类群的入侵植物对本地植物的种数有直接影响, 还需要针对某些单一入侵植物的群落进行比较研究, 并考虑草地演替过程与入侵植物迁入的关系.在村落生态系统中, 本地植物平均种数与入侵植物种数呈负相关关系, 其原因可能是人为地对居住环境进行改造后, 减少了本地植物的多样性, 使入侵植物更容易占据空缺生态位, 并逐渐成为群落优势种, 随着干扰强度的增加, 其优势地位变得更突出, 而群落中的次优种和伴生种的地位逐渐降低甚至消失, 这与王丽丽(2013)对农村生态系统中伴人植物的研究结果是一致的, 体现出人为干扰介导下入侵植物对本地植物多样性的影响. ...
土地利用/覆被变化与植被盖度的遥感监测——以北京市密云县为例
1
2009
... 选择农田、村落、种植园、林缘、弃耕地、草地、次生林7种人为干扰程度有差异和不同演替特征的生境类型作为研究对象, 结合调查中各生态系统类型的入侵程度和实地调查的可行性, 分别在农田、村落、种植园、林缘、弃耕地、草地、次生林中选取14、14、20、11、17、11、10个共97个样地的信息用于数据统计分析(图1 ), 分析的样地共含970个样方, 总面积为3904 m2 .以1套样方中入侵样方所占比例作为入侵频率, 用以比较不同生态系统中入侵植物的入侵频率, 这样, 每个样地有2个重复.目前, 有关植物盖度等级的划分多出现在植被覆盖度估算的研究中(盖永芹等, 2009 ; 李锦荣等; 2010 ), 而对样方中种群盖度等级的划分没有统一的标准, 因此, 在划分盖度等级时, 本文根据评估标准及分布范围, 把入侵植物的种群盖度分为一级(1%-20%)、二级(21%-40%)、三级(41%-60%)、四级(≥61%)共4个等级.运用Excel 2003和SPSS 17.0软件进行数据统计分析.用Systat SigmaPlot 10.0软件绘图.由于调查数据中入侵频率和盖度两项指标是服从二项分布的百分数的数据资料, 并且含有小于30%或大于70%的数值, 因此, 数据分析前对其进行反正弦转换Sokal & Rohlf, 1995 ). ...
土地利用/覆被变化与植被盖度的遥感监测——以北京市密云县为例
1
2009
... 选择农田、村落、种植园、林缘、弃耕地、草地、次生林7种人为干扰程度有差异和不同演替特征的生境类型作为研究对象, 结合调查中各生态系统类型的入侵程度和实地调查的可行性, 分别在农田、村落、种植园、林缘、弃耕地、草地、次生林中选取14、14、20、11、17、11、10个共97个样地的信息用于数据统计分析(图1 ), 分析的样地共含970个样方, 总面积为3904 m2 .以1套样方中入侵样方所占比例作为入侵频率, 用以比较不同生态系统中入侵植物的入侵频率, 这样, 每个样地有2个重复.目前, 有关植物盖度等级的划分多出现在植被覆盖度估算的研究中(盖永芹等, 2009 ; 李锦荣等; 2010 ), 而对样方中种群盖度等级的划分没有统一的标准, 因此, 在划分盖度等级时, 本文根据评估标准及分布范围, 把入侵植物的种群盖度分为一级(1%-20%)、二级(21%-40%)、三级(41%-60%)、四级(≥61%)共4个等级.运用Excel 2003和SPSS 17.0软件进行数据统计分析.用Systat SigmaPlot 10.0软件绘图.由于调查数据中入侵频率和盖度两项指标是服从二项分布的百分数的数据资料, 并且含有小于30%或大于70%的数值, 因此, 数据分析前对其进行反正弦转换Sokal & Rohlf, 1995 ). ...
Biological invasions: Benefits versus risks.
1
2009
... 有关入侵植物影响生态系统功能的报道有很多, 并多关注其负面效应Williamson, 2001 ; Byers et al ., 2002 ; Simberloff et al ., 2003 ), 认为植物入侵导致本地群落中生物多样性的下降李冰和李玉瑛, 2009 ), 例如: 丁晖等(2007)研究发现, 入侵森林林隙中的紫茎泽兰(Ageratina adenophora )使群落中本地植物的物种丰富度显著下降; Fox等在澳洲西部一个草地生态系统中的研究表明群落中的外来物种比例与本地植物种数呈负相关关系(Stohlgren et al ., 1999 ); Tilman (1997)在美国Bethel市一个草地生态系统中的实验也得出了一致的结论.因此一般认为, 外来植物成功入侵后将抑制群落中本地植物的多样性.但也有研究发现: 入侵植物对群落植物多样性的影响与入侵种及本地植物的生活型有关(Hejda et al ., 2009 ); 植物入侵的生态效应与入侵地的演替阶段有关, 例如: 豆科植物入侵生态环境恶劣的尾矿区后, 能有效地改善土壤肥力, 为演替中期过渡种的定居和发育创造良好的条件, 并促进物种多样性的增加Gozlan & Newton, 2009 ); 李安定等(2013)研究发现, 喜旱莲子草(空心莲子草)(Alternanthera philoxeroides )在入侵的早期阶段能改变群落生境条件, 进而促进其他物种的生存与定居, 并增加物种的多样性, 而只有当其种群盖度持续增加后, 才可能会威胁其他物种的生存; Hejda等(2009)研究发现, 入侵植物的盖度是决定群落物种丰富度和均匀度的主要因子, 入侵植物对物种多样性的影响与本地植物优势种的盖度、高度有密切关系.因此, 植物入侵的生态效应受生态系统类型、群落演替阶段的影响; 不同功能群植物对植物入侵的响应也有差异.本文通过研究7种生态系统类型中入侵植物的入侵特征及其对本地植物的影响, 旨在检验以下科学假设: 1)植物入侵的生态效应与入侵地的环境特征和演替阶段有关, 入侵植物可能对入侵地产生正面或负面影响; 2)植物入侵主要影响群落中某一功能群的物种多样性, 而不是影响全部群落个体; 3)多数入侵植物有偏好的入侵生境, 而人为干扰少的生态系统类型对植物入侵有抵制作用. ...
Impact of invasive plants on the species richness, diversity and composition of invaded communities.
1
2009
... 有关入侵植物影响生态系统功能的报道有很多, 并多关注其负面效应Williamson, 2001 ; Byers et al ., 2002 ; Simberloff et al ., 2003 ), 认为植物入侵导致本地群落中生物多样性的下降李冰和李玉瑛, 2009 ), 例如: 丁晖等(2007)研究发现, 入侵森林林隙中的紫茎泽兰(Ageratina adenophora )使群落中本地植物的物种丰富度显著下降; Fox等在澳洲西部一个草地生态系统中的研究表明群落中的外来物种比例与本地植物种数呈负相关关系(Stohlgren et al ., 1999 ); Tilman (1997)在美国Bethel市一个草地生态系统中的实验也得出了一致的结论.因此一般认为, 外来植物成功入侵后将抑制群落中本地植物的多样性.但也有研究发现: 入侵植物对群落植物多样性的影响与入侵种及本地植物的生活型有关(Hejda et al ., 2009 ); 植物入侵的生态效应与入侵地的演替阶段有关, 例如: 豆科植物入侵生态环境恶劣的尾矿区后, 能有效地改善土壤肥力, 为演替中期过渡种的定居和发育创造良好的条件, 并促进物种多样性的增加Gozlan & Newton, 2009 ); 李安定等(2013)研究发现, 喜旱莲子草(空心莲子草)(Alternanthera philoxeroides )在入侵的早期阶段能改变群落生境条件, 进而促进其他物种的生存与定居, 并增加物种的多样性, 而只有当其种群盖度持续增加后, 才可能会威胁其他物种的生存; Hejda等(2009)研究发现, 入侵植物的盖度是决定群落物种丰富度和均匀度的主要因子, 入侵植物对物种多样性的影响与本地植物优势种的盖度、高度有密切关系.因此, 植物入侵的生态效应受生态系统类型、群落演替阶段的影响; 不同功能群植物对植物入侵的响应也有差异.本文通过研究7种生态系统类型中入侵植物的入侵特征及其对本地植物的影响, 旨在检验以下科学假设: 1)植物入侵的生态效应与入侵地的环境特征和演替阶段有关, 入侵植物可能对入侵地产生正面或负面影响; 2)植物入侵主要影响群落中某一功能群的物种多样性, 而不是影响全部群落个体; 3)多数入侵植物有偏好的入侵生境, 而人为干扰少的生态系统类型对植物入侵有抵制作用. ...
Disturbance―A precursor to weed invasion in native vegetation.
1
1991
... 本文研究表明: 弃耕地、村落、农田生态系统中所含的入侵种数最多, 均有10种; 而在林缘生境中, 11种入侵植物的入侵频率和累计入侵频率都较低, 在抽样调查的次生林内, 没有发现入侵植物的分布(表1 ), 说明入侵植物主要分布在人为干扰强度大的生境中, 而在人为干扰强度小、群落结构复杂的生境中分布较少.结合6种生态系统的演替阶段分析, 弃耕地、村落、草地和农田均属于演替早期的生境类型, 受人为干扰的强度大, 而机械和人为活动的介导能有效地减少入侵植物的传播障碍, 或形成干扰间的协同效应(Hobbs, 1991 ); 入侵植物通常能够迅速响应环境中的干扰, 并且具有较强的表型可塑性, 这些特征促成了其在干扰生境中的成功入侵(Hobbs & Huenneke, 1992 ).植物对光能的捕获和利用效率高低是决定其能否适应环境并很好地存活下去的重要因素Baruch & Goldstein, 1999 ; Durand & Goldstein, 2001 ), 因此, 入侵生境的光环境对植物的入侵过程有很大影响, 如弃耕地通常具有高光照、高肥力的环境特征, 这为入侵种群的建立和发展提供了条件, 导致弃耕地常成为入侵植物(如假臭草、三裂叶蟛蜞菊等)集中分布的生境, 这与钟军弟等(2014)在广西的研究结果一致, 表明弃耕地具有较大的可入侵空间.林缘和天然次生林生态系统属于演替中后期的生境类型, 与演替前期的生境类型相比, 受人为干扰强度较低, 而增加的物种密度也将减少群落中的空缺生态位, 从而降低植物成功入侵的几率Stachowicz & Tilman, 2005 ).另一方面, 林缘或林下的光照强度和光合有效辐射都比较低, 这种环境特征也将限制植物入侵林缘的深度Brothers & Spingarn, 1992 ; Shea & Chesson, 2002 ), 因为本文研究的入侵植物多是处于演替初期的草本植物, 这些植物从种子萌发到幼苗生长及种群的发育均需要充足的光照, 它们的入侵可能在演替前期的生境类型, 如农田、草地和村落中具有更好的生态适应性; 但在林缘和林下的入侵却受到光照条件的制约, 且林缘的群落结构对低矮生活型的入侵植物也具有阻碍作用(Honnay et al ., 2002 ), 如喜旱莲子草、含羞草等在林缘中的入侵频率都很低.因此, 入侵植物虽然在林区广泛分布, 但它们仅分布在光照充足的路边或新垦的裸地上, 而无法入侵到森林内部, 国外的不少报道也证实了这种分布特点Parendes & Jones, 2000 ).分析入侵种群的盖度可知, 除三裂叶蟛蜞菊的种群盖度较高外, 其余5种入侵植物的盖度都集中分布在1%-20%之间, 并与入侵地的环境特征存在相关性, 如含羞草在农田、假臭草在弃耕地、三裂叶蟛蜞菊在种植园中通常具有成群分布生长的特点, 体现出这3种入侵植物在对应的生境中有更好的生态适应性; 而在林缘生态系统中, 除马缨丹和苏门白酒草外, 其余4种入侵植物的入侵频率与种群盖度呈负相关关系, 表明在林缘生态系统中, 入侵植物在入侵群落中的密度小, 难以形成单优群落, 这与入侵植物的种子是否能到达林缘, 能否存活及能否萌芽有密切的关系, 这些问题有待进一步实验验证. ...
Disturbance, diversity, and invasion: Implications for conservation.
1
1992
... 本文研究表明: 弃耕地、村落、农田生态系统中所含的入侵种数最多, 均有10种; 而在林缘生境中, 11种入侵植物的入侵频率和累计入侵频率都较低, 在抽样调查的次生林内, 没有发现入侵植物的分布(表1 ), 说明入侵植物主要分布在人为干扰强度大的生境中, 而在人为干扰强度小、群落结构复杂的生境中分布较少.结合6种生态系统的演替阶段分析, 弃耕地、村落、草地和农田均属于演替早期的生境类型, 受人为干扰的强度大, 而机械和人为活动的介导能有效地减少入侵植物的传播障碍, 或形成干扰间的协同效应(Hobbs, 1991 ); 入侵植物通常能够迅速响应环境中的干扰, 并且具有较强的表型可塑性, 这些特征促成了其在干扰生境中的成功入侵(Hobbs & Huenneke, 1992 ).植物对光能的捕获和利用效率高低是决定其能否适应环境并很好地存活下去的重要因素Baruch & Goldstein, 1999 ; Durand & Goldstein, 2001 ), 因此, 入侵生境的光环境对植物的入侵过程有很大影响, 如弃耕地通常具有高光照、高肥力的环境特征, 这为入侵种群的建立和发展提供了条件, 导致弃耕地常成为入侵植物(如假臭草、三裂叶蟛蜞菊等)集中分布的生境, 这与钟军弟等(2014)在广西的研究结果一致, 表明弃耕地具有较大的可入侵空间.林缘和天然次生林生态系统属于演替中后期的生境类型, 与演替前期的生境类型相比, 受人为干扰强度较低, 而增加的物种密度也将减少群落中的空缺生态位, 从而降低植物成功入侵的几率Stachowicz & Tilman, 2005 ).另一方面, 林缘或林下的光照强度和光合有效辐射都比较低, 这种环境特征也将限制植物入侵林缘的深度Brothers & Spingarn, 1992 ; Shea & Chesson, 2002 ), 因为本文研究的入侵植物多是处于演替初期的草本植物, 这些植物从种子萌发到幼苗生长及种群的发育均需要充足的光照, 它们的入侵可能在演替前期的生境类型, 如农田、草地和村落中具有更好的生态适应性; 但在林缘和林下的入侵却受到光照条件的制约, 且林缘的群落结构对低矮生活型的入侵植物也具有阻碍作用(Honnay et al ., 2002 ), 如喜旱莲子草、含羞草等在林缘中的入侵频率都很低.因此, 入侵植物虽然在林区广泛分布, 但它们仅分布在光照充足的路边或新垦的裸地上, 而无法入侵到森林内部, 国外的不少报道也证实了这种分布特点Parendes & Jones, 2000 ).分析入侵种群的盖度可知, 除三裂叶蟛蜞菊的种群盖度较高外, 其余5种入侵植物的盖度都集中分布在1%-20%之间, 并与入侵地的环境特征存在相关性, 如含羞草在农田、假臭草在弃耕地、三裂叶蟛蜞菊在种植园中通常具有成群分布生长的特点, 体现出这3种入侵植物在对应的生境中有更好的生态适应性; 而在林缘生态系统中, 除马缨丹和苏门白酒草外, 其余4种入侵植物的入侵频率与种群盖度呈负相关关系, 表明在林缘生态系统中, 入侵植物在入侵群落中的密度小, 难以形成单优群落, 这与入侵植物的种子是否能到达林缘, 能否存活及能否萌芽有密切的关系, 这些问题有待进一步实验验证. ...
Permeability of ancient forest edges for weedy plant species invasion.
1
2002
... 本文研究表明: 弃耕地、村落、农田生态系统中所含的入侵种数最多, 均有10种; 而在林缘生境中, 11种入侵植物的入侵频率和累计入侵频率都较低, 在抽样调查的次生林内, 没有发现入侵植物的分布(表1 ), 说明入侵植物主要分布在人为干扰强度大的生境中, 而在人为干扰强度小、群落结构复杂的生境中分布较少.结合6种生态系统的演替阶段分析, 弃耕地、村落、草地和农田均属于演替早期的生境类型, 受人为干扰的强度大, 而机械和人为活动的介导能有效地减少入侵植物的传播障碍, 或形成干扰间的协同效应(Hobbs, 1991 ); 入侵植物通常能够迅速响应环境中的干扰, 并且具有较强的表型可塑性, 这些特征促成了其在干扰生境中的成功入侵(Hobbs & Huenneke, 1992 ).植物对光能的捕获和利用效率高低是决定其能否适应环境并很好地存活下去的重要因素Baruch & Goldstein, 1999 ; Durand & Goldstein, 2001 ), 因此, 入侵生境的光环境对植物的入侵过程有很大影响, 如弃耕地通常具有高光照、高肥力的环境特征, 这为入侵种群的建立和发展提供了条件, 导致弃耕地常成为入侵植物(如假臭草、三裂叶蟛蜞菊等)集中分布的生境, 这与钟军弟等(2014)在广西的研究结果一致, 表明弃耕地具有较大的可入侵空间.林缘和天然次生林生态系统属于演替中后期的生境类型, 与演替前期的生境类型相比, 受人为干扰强度较低, 而增加的物种密度也将减少群落中的空缺生态位, 从而降低植物成功入侵的几率Stachowicz & Tilman, 2005 ).另一方面, 林缘或林下的光照强度和光合有效辐射都比较低, 这种环境特征也将限制植物入侵林缘的深度Brothers & Spingarn, 1992 ; Shea & Chesson, 2002 ), 因为本文研究的入侵植物多是处于演替初期的草本植物, 这些植物从种子萌发到幼苗生长及种群的发育均需要充足的光照, 它们的入侵可能在演替前期的生境类型, 如农田、草地和村落中具有更好的生态适应性; 但在林缘和林下的入侵却受到光照条件的制约, 且林缘的群落结构对低矮生活型的入侵植物也具有阻碍作用(Honnay et al ., 2002 ), 如喜旱莲子草、含羞草等在林缘中的入侵频率都很低.因此, 入侵植物虽然在林区广泛分布, 但它们仅分布在光照充足的路边或新垦的裸地上, 而无法入侵到森林内部, 国外的不少报道也证实了这种分布特点Parendes & Jones, 2000 ).分析入侵种群的盖度可知, 除三裂叶蟛蜞菊的种群盖度较高外, 其余5种入侵植物的盖度都集中分布在1%-20%之间, 并与入侵地的环境特征存在相关性, 如含羞草在农田、假臭草在弃耕地、三裂叶蟛蜞菊在种植园中通常具有成群分布生长的特点, 体现出这3种入侵植物在对应的生境中有更好的生态适应性; 而在林缘生态系统中, 除马缨丹和苏门白酒草外, 其余4种入侵植物的入侵频率与种群盖度呈负相关关系, 表明在林缘生态系统中, 入侵植物在入侵群落中的密度小, 难以形成单优群落, 这与入侵植物的种子是否能到达林缘, 能否存活及能否萌芽有密切的关系, 这些问题有待进一步实验验证. ...
Native expanding Merremia boisiana is not more allelopathic than its non-expanding congener M. vitifolia in the expanded range in Hainan.
1
2013
... 海南岛虽然气候条件宜人、生物资源丰富, 但生态环境脆弱, 是许多外来入侵物种的“天然大温室”, 目前有关海南岛入侵植物的研究主要集中在以下两方面: 一是区域性入侵植物种类调查或群落组成调查(单家林, 2003 ; 秦新生等, 2008 ); 二是入侵植物生理生态的研究(Huang et al ., 2013 ; 黄乔乔等, 2013 ).这些研究有效地整合了特定区域内外来植物种类组成的信息, 并在入侵生态学方面做了一定的探索, 但还没有在海南全岛范围内开展主要入侵植物的分布特征及其入侵强度的研究, 有关入侵植物在不同生态系统中的入侵特征及对本地植物多样性影响的研究都还缺乏.本文研究了国家农业部指定的20种恶性入侵植物在海南岛的分布特征及其对当地植物的影响, 并比较了6种主要入侵植物在7种生态系统中的入侵强度, 以期进一步了解海南岛入侵植物的分布现状, 并为外来植物的防控、生物多样性保护与生态安全的管理提供理论依据. ...
五指山不同林型土壤对金钟藤幼苗生长的影响
1
2013
... 海南岛虽然气候条件宜人、生物资源丰富, 但生态环境脆弱, 是许多外来入侵物种的“天然大温室”, 目前有关海南岛入侵植物的研究主要集中在以下两方面: 一是区域性入侵植物种类调查或群落组成调查(单家林, 2003 ; 秦新生等, 2008 ); 二是入侵植物生理生态的研究(Huang et al ., 2013 ; 黄乔乔等, 2013 ).这些研究有效地整合了特定区域内外来植物种类组成的信息, 并在入侵生态学方面做了一定的探索, 但还没有在海南全岛范围内开展主要入侵植物的分布特征及其入侵强度的研究, 有关入侵植物在不同生态系统中的入侵特征及对本地植物多样性影响的研究都还缺乏.本文研究了国家农业部指定的20种恶性入侵植物在海南岛的分布特征及其对当地植物的影响, 并比较了6种主要入侵植物在7种生态系统中的入侵强度, 以期进一步了解海南岛入侵植物的分布现状, 并为外来植物的防控、生物多样性保护与生态安全的管理提供理论依据. ...
五指山不同林型土壤对金钟藤幼苗生长的影响
1
2013
... 海南岛虽然气候条件宜人、生物资源丰富, 但生态环境脆弱, 是许多外来入侵物种的“天然大温室”, 目前有关海南岛入侵植物的研究主要集中在以下两方面: 一是区域性入侵植物种类调查或群落组成调查(单家林, 2003 ; 秦新生等, 2008 ); 二是入侵植物生理生态的研究(Huang et al ., 2013 ; 黄乔乔等, 2013 ).这些研究有效地整合了特定区域内外来植物种类组成的信息, 并在入侵生态学方面做了一定的探索, 但还没有在海南全岛范围内开展主要入侵植物的分布特征及其入侵强度的研究, 有关入侵植物在不同生态系统中的入侵特征及对本地植物多样性影响的研究都还缺乏.本文研究了国家农业部指定的20种恶性入侵植物在海南岛的分布特征及其对当地植物的影响, 并比较了6种主要入侵植物在7种生态系统中的入侵强度, 以期进一步了解海南岛入侵植物的分布现状, 并为外来植物的防控、生物多样性保护与生态安全的管理提供理论依据. ...
Hidden treatments in ecological experiments: Re-evaluating the ecosystem function of biodiversity.
0
1997
Multivariate forecasts of potential distributions of invasive plant species.
1
2009
... 针对入侵植物的分布问题, Losdale (1999)研究了全球入侵植物的分布特征, 并发现温带地区的耕地、城镇和生物多样性热点地区更易遭受外来植物的入侵, 而荒漠、稀树草原等生态系统的可入侵空间较小; Barney等(2008)研究了两种不同入侵历史的外来植物在北美的分布和扩散格局, 发现其入侵范围均随着时间的推移而扩大; 此外, 还有不少学者运用气候模拟技术对入侵植物的潜在分布区进行了预测(Erik, 2004 ; Ibáñez et al ., 2009 ; Cross- man et al ., 2011 ); 而国内的研究多集中在入侵植物多样性的区域分异及其影响因素等方面, 如吴晓雯等(2006)研究了入侵植物在我国的分布格局, 发现其物种数从南到北逐渐减少, 而物种密度由东南海岸向内陆递减的分布特征, 并推测中国的东南部地区将更容易遭受外来植物入侵; 王苏铭等(2012)研究了入侵植物在北京地区的分布特征; 张帅等(2010)研究了入侵植物在我国74个地理单元的分布情况, 发现入侵植物的分布具有明显的区域分化现象, 而降水与温度是影响其空间分布格局的主要 因素. ...
异质生境空心莲子草植物群落组成及物种多样性研究
0
2013
异质生境空心莲子草植物群落组成及物种多样性研究
0
2013
生物入侵中入侵种与土著种的相互作用
1
2009
... 有关入侵植物影响生态系统功能的报道有很多, 并多关注其负面效应Williamson, 2001 ; Byers et al ., 2002 ; Simberloff et al ., 2003 ), 认为植物入侵导致本地群落中生物多样性的下降李冰和李玉瑛, 2009 ), 例如: 丁晖等(2007)研究发现, 入侵森林林隙中的紫茎泽兰(Ageratina adenophora )使群落中本地植物的物种丰富度显著下降; Fox等在澳洲西部一个草地生态系统中的研究表明群落中的外来物种比例与本地植物种数呈负相关关系(Stohlgren et al ., 1999 ); Tilman (1997)在美国Bethel市一个草地生态系统中的实验也得出了一致的结论.因此一般认为, 外来植物成功入侵后将抑制群落中本地植物的多样性.但也有研究发现: 入侵植物对群落植物多样性的影响与入侵种及本地植物的生活型有关(Hejda et al ., 2009 ); 植物入侵的生态效应与入侵地的演替阶段有关, 例如: 豆科植物入侵生态环境恶劣的尾矿区后, 能有效地改善土壤肥力, 为演替中期过渡种的定居和发育创造良好的条件, 并促进物种多样性的增加Gozlan & Newton, 2009 ); 李安定等(2013)研究发现, 喜旱莲子草(空心莲子草)(Alternanthera philoxeroides )在入侵的早期阶段能改变群落生境条件, 进而促进其他物种的生存与定居, 并增加物种的多样性, 而只有当其种群盖度持续增加后, 才可能会威胁其他物种的生存; Hejda等(2009)研究发现, 入侵植物的盖度是决定群落物种丰富度和均匀度的主要因子, 入侵植物对物种多样性的影响与本地植物优势种的盖度、高度有密切关系.因此, 植物入侵的生态效应受生态系统类型、群落演替阶段的影响; 不同功能群植物对植物入侵的响应也有差异.本文通过研究7种生态系统类型中入侵植物的入侵特征及其对本地植物的影响, 旨在检验以下科学假设: 1)植物入侵的生态效应与入侵地的环境特征和演替阶段有关, 入侵植物可能对入侵地产生正面或负面影响; 2)植物入侵主要影响群落中某一功能群的物种多样性, 而不是影响全部群落个体; 3)多数入侵植物有偏好的入侵生境, 而人为干扰少的生态系统类型对植物入侵有抵制作用. ...
生物入侵中入侵种与土著种的相互作用
1
2009
... 有关入侵植物影响生态系统功能的报道有很多, 并多关注其负面效应Williamson, 2001 ; Byers et al ., 2002 ; Simberloff et al ., 2003 ), 认为植物入侵导致本地群落中生物多样性的下降李冰和李玉瑛, 2009 ), 例如: 丁晖等(2007)研究发现, 入侵森林林隙中的紫茎泽兰(Ageratina adenophora )使群落中本地植物的物种丰富度显著下降; Fox等在澳洲西部一个草地生态系统中的研究表明群落中的外来物种比例与本地植物种数呈负相关关系(Stohlgren et al ., 1999 ); Tilman (1997)在美国Bethel市一个草地生态系统中的实验也得出了一致的结论.因此一般认为, 外来植物成功入侵后将抑制群落中本地植物的多样性.但也有研究发现: 入侵植物对群落植物多样性的影响与入侵种及本地植物的生活型有关(Hejda et al ., 2009 ); 植物入侵的生态效应与入侵地的演替阶段有关, 例如: 豆科植物入侵生态环境恶劣的尾矿区后, 能有效地改善土壤肥力, 为演替中期过渡种的定居和发育创造良好的条件, 并促进物种多样性的增加Gozlan & Newton, 2009 ); 李安定等(2013)研究发现, 喜旱莲子草(空心莲子草)(Alternanthera philoxeroides )在入侵的早期阶段能改变群落生境条件, 进而促进其他物种的生存与定居, 并增加物种的多样性, 而只有当其种群盖度持续增加后, 才可能会威胁其他物种的生存; Hejda等(2009)研究发现, 入侵植物的盖度是决定群落物种丰富度和均匀度的主要因子, 入侵植物对物种多样性的影响与本地植物优势种的盖度、高度有密切关系.因此, 植物入侵的生态效应受生态系统类型、群落演替阶段的影响; 不同功能群植物对植物入侵的响应也有差异.本文通过研究7种生态系统类型中入侵植物的入侵特征及其对本地植物的影响, 旨在检验以下科学假设: 1)植物入侵的生态效应与入侵地的环境特征和演替阶段有关, 入侵植物可能对入侵地产生正面或负面影响; 2)植物入侵主要影响群落中某一功能群的物种多样性, 而不是影响全部群落个体; 3)多数入侵植物有偏好的入侵生境, 而人为干扰少的生态系统类型对植物入侵有抵制作用. ...
退耕还林工程前后土地利用与植被盖度动态变化研究——以安塞县为例
1
2010
... 选择农田、村落、种植园、林缘、弃耕地、草地、次生林7种人为干扰程度有差异和不同演替特征的生境类型作为研究对象, 结合调查中各生态系统类型的入侵程度和实地调查的可行性, 分别在农田、村落、种植园、林缘、弃耕地、草地、次生林中选取14、14、20、11、17、11、10个共97个样地的信息用于数据统计分析(图1 ), 分析的样地共含970个样方, 总面积为3904 m2 .以1套样方中入侵样方所占比例作为入侵频率, 用以比较不同生态系统中入侵植物的入侵频率, 这样, 每个样地有2个重复.目前, 有关植物盖度等级的划分多出现在植被覆盖度估算的研究中(盖永芹等, 2009 ; 李锦荣等; 2010 ), 而对样方中种群盖度等级的划分没有统一的标准, 因此, 在划分盖度等级时, 本文根据评估标准及分布范围, 把入侵植物的种群盖度分为一级(1%-20%)、二级(21%-40%)、三级(41%-60%)、四级(≥61%)共4个等级.运用Excel 2003和SPSS 17.0软件进行数据统计分析.用Systat SigmaPlot 10.0软件绘图.由于调查数据中入侵频率和盖度两项指标是服从二项分布的百分数的数据资料, 并且含有小于30%或大于70%的数值, 因此, 数据分析前对其进行反正弦转换Sokal & Rohlf, 1995 ). ...
退耕还林工程前后土地利用与植被盖度动态变化研究——以安塞县为例
1
2010
... 选择农田、村落、种植园、林缘、弃耕地、草地、次生林7种人为干扰程度有差异和不同演替特征的生境类型作为研究对象, 结合调查中各生态系统类型的入侵程度和实地调查的可行性, 分别在农田、村落、种植园、林缘、弃耕地、草地、次生林中选取14、14、20、11、17、11、10个共97个样地的信息用于数据统计分析(图1 ), 分析的样地共含970个样方, 总面积为3904 m2 .以1套样方中入侵样方所占比例作为入侵频率, 用以比较不同生态系统中入侵植物的入侵频率, 这样, 每个样地有2个重复.目前, 有关植物盖度等级的划分多出现在植被覆盖度估算的研究中(盖永芹等, 2009 ; 李锦荣等; 2010 ), 而对样方中种群盖度等级的划分没有统一的标准, 因此, 在划分盖度等级时, 本文根据评估标准及分布范围, 把入侵植物的种群盖度分为一级(1%-20%)、二级(21%-40%)、三级(41%-60%)、四级(≥61%)共4个等级.运用Excel 2003和SPSS 17.0软件进行数据统计分析.用Systat SigmaPlot 10.0软件绘图.由于调查数据中入侵频率和盖度两项指标是服从二项分布的百分数的数据资料, 并且含有小于30%或大于70%的数值, 因此, 数据分析前对其进行反正弦转换Sokal & Rohlf, 1995 ). ...
1
2002
... 入侵植物在海南岛分布广泛, 且不同区域间种数相差不大, 但整体呈现为东北部、西南部地区种类较多, 东南部次之, 而中部较少的分布特征.入侵植物的分布主要取决于环境的可入侵性和物种的入侵能力两个方面Newsome & Noble, 1986 ; 刘建等, 2010 ).如“理想”的入侵植物通常具备生活史短, 自交亲和性、结实率高, 可进行无性繁殖等特点, 并能迅速响应环境中的干扰Rejmánek & Richardson, 1996 ).本文研究的11种入侵植物中, 除马缨丹外, 其余10种均为一、二年生的草本植物, 其中有6种为菊科植物, 如飞机草、假臭草、微甘菊等, 这些植物产生的种子数量大而质量轻, 通常还附有冠毛, 便于长距离传播和扩散李振宇和谢焱, 2002 ; Weber et al ., 2008 ), 而喜旱莲子草、微甘菊、凤眼蓝、三裂叶蟛蜞菊具有无性繁殖的特性, 其繁殖体可以借助水流进行传播, 这些生物学特征和传播机制增强了其入侵能力, 有助于入侵植物的扩散和迅速占据入侵地, 并能在较短的时间内扩大种群, 从而导致其在海南岛的分布范围很广.但对比不同区域的分布特点发现, 东北部和西南部的入侵种数相对较多, 为10-11种, 而中部和东南部分布较少, 这种差异可能与不同区域间的气候条件和人为干扰强度有关.由于海南岛的西部地区贯穿有环西粤海铁路, 具有较长的运输史, 而连接该地区的海口市、儋州市、东方市都是水陆交通的重要枢纽, 境内人流密集, 干扰强度大, 这些因素都能有效地促进入侵植物的传播和扩散(吴晓雯等, 2006 ; 张帅等, 2010 ).结合气候条件进一步分析, 海南岛西部地区的年晴天数平均为300天, 明显高于中部和东部地区, 但降水量较少, 而中部地区是全岛降水集中分布的地区, 年降水量达2000 mm, 属于湿润地区.此外, 中部地区的年平均气温为23 ℃左右, 处于全岛平均气温(23-26 ℃)的下限, 低于海南岛其他地区的温度(蔡大鑫等, 2010 ).综合气候条件和入侵种数的分布特征发现, 入侵植物在年平均气温较高、降水量较低、而日照时间较长的东北、西南部地区分布的种类较多, 在年平均气温较低、降水量较高、日照时间较短的中部、东部地区分布的种类较少, 这可能与入侵植物的生活习性有关, 因为本文研究的入侵植物多为草本阳性植物, 它们的种子萌发和种群发展都需要足够的光温条件, 过高湿和低温的环境可能对其生长和扩散具有抑制作用. ...
1
2002
... 入侵植物在海南岛分布广泛, 且不同区域间种数相差不大, 但整体呈现为东北部、西南部地区种类较多, 东南部次之, 而中部较少的分布特征.入侵植物的分布主要取决于环境的可入侵性和物种的入侵能力两个方面Newsome & Noble, 1986 ; 刘建等, 2010 ).如“理想”的入侵植物通常具备生活史短, 自交亲和性、结实率高, 可进行无性繁殖等特点, 并能迅速响应环境中的干扰Rejmánek & Richardson, 1996 ).本文研究的11种入侵植物中, 除马缨丹外, 其余10种均为一、二年生的草本植物, 其中有6种为菊科植物, 如飞机草、假臭草、微甘菊等, 这些植物产生的种子数量大而质量轻, 通常还附有冠毛, 便于长距离传播和扩散李振宇和谢焱, 2002 ; Weber et al ., 2008 ), 而喜旱莲子草、微甘菊、凤眼蓝、三裂叶蟛蜞菊具有无性繁殖的特性, 其繁殖体可以借助水流进行传播, 这些生物学特征和传播机制增强了其入侵能力, 有助于入侵植物的扩散和迅速占据入侵地, 并能在较短的时间内扩大种群, 从而导致其在海南岛的分布范围很广.但对比不同区域的分布特点发现, 东北部和西南部的入侵种数相对较多, 为10-11种, 而中部和东南部分布较少, 这种差异可能与不同区域间的气候条件和人为干扰强度有关.由于海南岛的西部地区贯穿有环西粤海铁路, 具有较长的运输史, 而连接该地区的海口市、儋州市、东方市都是水陆交通的重要枢纽, 境内人流密集, 干扰强度大, 这些因素都能有效地促进入侵植物的传播和扩散(吴晓雯等, 2006 ; 张帅等, 2010 ).结合气候条件进一步分析, 海南岛西部地区的年晴天数平均为300天, 明显高于中部和东部地区, 但降水量较少, 而中部地区是全岛降水集中分布的地区, 年降水量达2000 mm, 属于湿润地区.此外, 中部地区的年平均气温为23 ℃左右, 处于全岛平均气温(23-26 ℃)的下限, 低于海南岛其他地区的温度(蔡大鑫等, 2010 ).综合气候条件和入侵种数的分布特征发现, 入侵植物在年平均气温较高、降水量较低、而日照时间较长的东北、西南部地区分布的种类较多, 在年平均气温较低、降水量较高、日照时间较短的中部、东部地区分布的种类较少, 这可能与入侵植物的生活习性有关, 因为本文研究的入侵植物多为草本阳性植物, 它们的种子萌发和种群发展都需要足够的光温条件, 过高湿和低温的环境可能对其生长和扩散具有抑制作用. ...
Contrasting community responses to fertilization and the role of the competitive ability of dominant species.
1
2009
... Stohlgren等(1999)对草地生态系统的研究表明, 在群落水平上, 本地植物种数与入侵植物种数呈正相关关系, 并认为这与两类植物对草地环境的变化有相同的响应有关.而本文发现, 草地群落中本地植物平均种数随入侵种数的增加呈先减少后增加的趋势, 且在入侵种数为2时最少, 草本植物也呈一致的变化趋势, 而乔木、灌木和藤本植物的种数变化不显著, 其原因可能是当入侵种数较少时, 某些入侵植物排挤本地植物后形成了单优群落, 并导致本地植物物种丰富度下降, 而随着入侵种数的增加, 新入侵的植物改变了优势种的组成, 这种变化进一步促进了新的本地植物迁入, 从而增加了本地植物的多样性, 因为群落优势种对新物种的迁入具有决定性作用(Liancourt et al ., 2009 ).Huston (1997)的研究也表明, 群落的可入侵性更多取决于关键种, 而并非整个群落中的物种数, 但在较小尺度上取样时, 物种多样性与入侵关系的模式比较复杂, 群落类型和地域的差异都会导致不同的结果, 还需要考虑其他因素的影响(Fridley et al ., 2007 ).针对本文研究的10种陆生入侵植物, 在入侵植物组合不同的群落中, 具体是哪一类群的入侵植物对本地植物的种数有直接影响, 还需要针对某些单一入侵植物的群落进行比较研究, 并考虑草地演替过程与入侵植物迁入的关系.在村落生态系统中, 本地植物平均种数与入侵植物种数呈负相关关系, 其原因可能是人为地对居住环境进行改造后, 减少了本地植物的多样性, 使入侵植物更容易占据空缺生态位, 并逐渐成为群落优势种, 随着干扰强度的增加, 其优势地位变得更突出, 而群落中的次优种和伴生种的地位逐渐降低甚至消失, 这与王丽丽(2013)对农村生态系统中伴人植物的研究结果是一致的, 体现出人为干扰介导下入侵植物对本地植物多样性的影响. ...
植物功能性状与外来植物入侵
1
2010
... 入侵植物在海南岛分布广泛, 且不同区域间种数相差不大, 但整体呈现为东北部、西南部地区种类较多, 东南部次之, 而中部较少的分布特征.入侵植物的分布主要取决于环境的可入侵性和物种的入侵能力两个方面Newsome & Noble, 1986 ; 刘建等, 2010 ).如“理想”的入侵植物通常具备生活史短, 自交亲和性、结实率高, 可进行无性繁殖等特点, 并能迅速响应环境中的干扰Rejmánek & Richardson, 1996 ).本文研究的11种入侵植物中, 除马缨丹外, 其余10种均为一、二年生的草本植物, 其中有6种为菊科植物, 如飞机草、假臭草、微甘菊等, 这些植物产生的种子数量大而质量轻, 通常还附有冠毛, 便于长距离传播和扩散李振宇和谢焱, 2002 ; Weber et al ., 2008 ), 而喜旱莲子草、微甘菊、凤眼蓝、三裂叶蟛蜞菊具有无性繁殖的特性, 其繁殖体可以借助水流进行传播, 这些生物学特征和传播机制增强了其入侵能力, 有助于入侵植物的扩散和迅速占据入侵地, 并能在较短的时间内扩大种群, 从而导致其在海南岛的分布范围很广.但对比不同区域的分布特点发现, 东北部和西南部的入侵种数相对较多, 为10-11种, 而中部和东南部分布较少, 这种差异可能与不同区域间的气候条件和人为干扰强度有关.由于海南岛的西部地区贯穿有环西粤海铁路, 具有较长的运输史, 而连接该地区的海口市、儋州市、东方市都是水陆交通的重要枢纽, 境内人流密集, 干扰强度大, 这些因素都能有效地促进入侵植物的传播和扩散(吴晓雯等, 2006 ; 张帅等, 2010 ).结合气候条件进一步分析, 海南岛西部地区的年晴天数平均为300天, 明显高于中部和东部地区, 但降水量较少, 而中部地区是全岛降水集中分布的地区, 年降水量达2000 mm, 属于湿润地区.此外, 中部地区的年平均气温为23 ℃左右, 处于全岛平均气温(23-26 ℃)的下限, 低于海南岛其他地区的温度(蔡大鑫等, 2010 ).综合气候条件和入侵种数的分布特征发现, 入侵植物在年平均气温较高、降水量较低、而日照时间较长的东北、西南部地区分布的种类较多, 在年平均气温较低、降水量较高、日照时间较短的中部、东部地区分布的种类较少, 这可能与入侵植物的生活习性有关, 因为本文研究的入侵植物多为草本阳性植物, 它们的种子萌发和种群发展都需要足够的光温条件, 过高湿和低温的环境可能对其生长和扩散具有抑制作用. ...
植物功能性状与外来植物入侵
1
2010
... 入侵植物在海南岛分布广泛, 且不同区域间种数相差不大, 但整体呈现为东北部、西南部地区种类较多, 东南部次之, 而中部较少的分布特征.入侵植物的分布主要取决于环境的可入侵性和物种的入侵能力两个方面Newsome & Noble, 1986 ; 刘建等, 2010 ).如“理想”的入侵植物通常具备生活史短, 自交亲和性、结实率高, 可进行无性繁殖等特点, 并能迅速响应环境中的干扰Rejmánek & Richardson, 1996 ).本文研究的11种入侵植物中, 除马缨丹外, 其余10种均为一、二年生的草本植物, 其中有6种为菊科植物, 如飞机草、假臭草、微甘菊等, 这些植物产生的种子数量大而质量轻, 通常还附有冠毛, 便于长距离传播和扩散李振宇和谢焱, 2002 ; Weber et al ., 2008 ), 而喜旱莲子草、微甘菊、凤眼蓝、三裂叶蟛蜞菊具有无性繁殖的特性, 其繁殖体可以借助水流进行传播, 这些生物学特征和传播机制增强了其入侵能力, 有助于入侵植物的扩散和迅速占据入侵地, 并能在较短的时间内扩大种群, 从而导致其在海南岛的分布范围很广.但对比不同区域的分布特点发现, 东北部和西南部的入侵种数相对较多, 为10-11种, 而中部和东南部分布较少, 这种差异可能与不同区域间的气候条件和人为干扰强度有关.由于海南岛的西部地区贯穿有环西粤海铁路, 具有较长的运输史, 而连接该地区的海口市、儋州市、东方市都是水陆交通的重要枢纽, 境内人流密集, 干扰强度大, 这些因素都能有效地促进入侵植物的传播和扩散(吴晓雯等, 2006 ; 张帅等, 2010 ).结合气候条件进一步分析, 海南岛西部地区的年晴天数平均为300天, 明显高于中部和东部地区, 但降水量较少, 而中部地区是全岛降水集中分布的地区, 年降水量达2000 mm, 属于湿润地区.此外, 中部地区的年平均气温为23 ℃左右, 处于全岛平均气温(23-26 ℃)的下限, 低于海南岛其他地区的温度(蔡大鑫等, 2010 ).综合气候条件和入侵种数的分布特征发现, 入侵植物在年平均气温较高、降水量较低、而日照时间较长的东北、西南部地区分布的种类较多, 在年平均气温较低、降水量较高、日照时间较短的中部、东部地区分布的种类较少, 这可能与入侵植物的生活习性有关, 因为本文研究的入侵植物多为草本阳性植物, 它们的种子萌发和种群发展都需要足够的光温条件, 过高湿和低温的环境可能对其生长和扩散具有抑制作用. ...
Global patterns of plant invasions and the concept of invasibility.
0
1999
1
1986
... 入侵植物在海南岛分布广泛, 且不同区域间种数相差不大, 但整体呈现为东北部、西南部地区种类较多, 东南部次之, 而中部较少的分布特征.入侵植物的分布主要取决于环境的可入侵性和物种的入侵能力两个方面Newsome & Noble, 1986 ; 刘建等, 2010 ).如“理想”的入侵植物通常具备生活史短, 自交亲和性、结实率高, 可进行无性繁殖等特点, 并能迅速响应环境中的干扰Rejmánek & Richardson, 1996 ).本文研究的11种入侵植物中, 除马缨丹外, 其余10种均为一、二年生的草本植物, 其中有6种为菊科植物, 如飞机草、假臭草、微甘菊等, 这些植物产生的种子数量大而质量轻, 通常还附有冠毛, 便于长距离传播和扩散李振宇和谢焱, 2002 ; Weber et al ., 2008 ), 而喜旱莲子草、微甘菊、凤眼蓝、三裂叶蟛蜞菊具有无性繁殖的特性, 其繁殖体可以借助水流进行传播, 这些生物学特征和传播机制增强了其入侵能力, 有助于入侵植物的扩散和迅速占据入侵地, 并能在较短的时间内扩大种群, 从而导致其在海南岛的分布范围很广.但对比不同区域的分布特点发现, 东北部和西南部的入侵种数相对较多, 为10-11种, 而中部和东南部分布较少, 这种差异可能与不同区域间的气候条件和人为干扰强度有关.由于海南岛的西部地区贯穿有环西粤海铁路, 具有较长的运输史, 而连接该地区的海口市、儋州市、东方市都是水陆交通的重要枢纽, 境内人流密集, 干扰强度大, 这些因素都能有效地促进入侵植物的传播和扩散(吴晓雯等, 2006 ; 张帅等, 2010 ).结合气候条件进一步分析, 海南岛西部地区的年晴天数平均为300天, 明显高于中部和东部地区, 但降水量较少, 而中部地区是全岛降水集中分布的地区, 年降水量达2000 mm, 属于湿润地区.此外, 中部地区的年平均气温为23 ℃左右, 处于全岛平均气温(23-26 ℃)的下限, 低于海南岛其他地区的温度(蔡大鑫等, 2010 ).综合气候条件和入侵种数的分布特征发现, 入侵植物在年平均气温较高、降水量较低、而日照时间较长的东北、西南部地区分布的种类较多, 在年平均气温较低、降水量较高、日照时间较短的中部、东部地区分布的种类较少, 这可能与入侵植物的生活习性有关, 因为本文研究的入侵植物多为草本阳性植物, 它们的种子萌发和种群发展都需要足够的光温条件, 过高湿和低温的环境可能对其生长和扩散具有抑制作用. ...
Role of light availability and dispersal in exotic plant invasion along roads and streams in the H. J. Andrews experimental forest, Oregon.
1
2000
... 本文研究表明: 弃耕地、村落、农田生态系统中所含的入侵种数最多, 均有10种; 而在林缘生境中, 11种入侵植物的入侵频率和累计入侵频率都较低, 在抽样调查的次生林内, 没有发现入侵植物的分布(表1 ), 说明入侵植物主要分布在人为干扰强度大的生境中, 而在人为干扰强度小、群落结构复杂的生境中分布较少.结合6种生态系统的演替阶段分析, 弃耕地、村落、草地和农田均属于演替早期的生境类型, 受人为干扰的强度大, 而机械和人为活动的介导能有效地减少入侵植物的传播障碍, 或形成干扰间的协同效应(Hobbs, 1991 ); 入侵植物通常能够迅速响应环境中的干扰, 并且具有较强的表型可塑性, 这些特征促成了其在干扰生境中的成功入侵(Hobbs & Huenneke, 1992 ).植物对光能的捕获和利用效率高低是决定其能否适应环境并很好地存活下去的重要因素Baruch & Goldstein, 1999 ; Durand & Goldstein, 2001 ), 因此, 入侵生境的光环境对植物的入侵过程有很大影响, 如弃耕地通常具有高光照、高肥力的环境特征, 这为入侵种群的建立和发展提供了条件, 导致弃耕地常成为入侵植物(如假臭草、三裂叶蟛蜞菊等)集中分布的生境, 这与钟军弟等(2014)在广西的研究结果一致, 表明弃耕地具有较大的可入侵空间.林缘和天然次生林生态系统属于演替中后期的生境类型, 与演替前期的生境类型相比, 受人为干扰强度较低, 而增加的物种密度也将减少群落中的空缺生态位, 从而降低植物成功入侵的几率Stachowicz & Tilman, 2005 ).另一方面, 林缘或林下的光照强度和光合有效辐射都比较低, 这种环境特征也将限制植物入侵林缘的深度Brothers & Spingarn, 1992 ; Shea & Chesson, 2002 ), 因为本文研究的入侵植物多是处于演替初期的草本植物, 这些植物从种子萌发到幼苗生长及种群的发育均需要充足的光照, 它们的入侵可能在演替前期的生境类型, 如农田、草地和村落中具有更好的生态适应性; 但在林缘和林下的入侵却受到光照条件的制约, 且林缘的群落结构对低矮生活型的入侵植物也具有阻碍作用(Honnay et al ., 2002 ), 如喜旱莲子草、含羞草等在林缘中的入侵频率都很低.因此, 入侵植物虽然在林区广泛分布, 但它们仅分布在光照充足的路边或新垦的裸地上, 而无法入侵到森林内部, 国外的不少报道也证实了这种分布特点Parendes & Jones, 2000 ).分析入侵种群的盖度可知, 除三裂叶蟛蜞菊的种群盖度较高外, 其余5种入侵植物的盖度都集中分布在1%-20%之间, 并与入侵地的环境特征存在相关性, 如含羞草在农田、假臭草在弃耕地、三裂叶蟛蜞菊在种植园中通常具有成群分布生长的特点, 体现出这3种入侵植物在对应的生境中有更好的生态适应性; 而在林缘生态系统中, 除马缨丹和苏门白酒草外, 其余4种入侵植物的入侵频率与种群盖度呈负相关关系, 表明在林缘生态系统中, 入侵植物在入侵群落中的密度小, 难以形成单优群落, 这与入侵植物的种子是否能到达林缘, 能否存活及能否萌芽有密切的关系, 这些问题有待进一步实验验证. ...
海南岛外来植物入侵风险评价指标体系
1
2013
... 海南岛地处热带北缘, 面积3.4万km2 , 是我国第二大岛屿.海南岛气候温和, 年平均气温为22.5-26.0 ℃, 年降水量为1500-2000 mm.海南岛的地势中部高、四周低, 这种地理地貌使得其年总积温、日照时数等都具有自中部向沿海递增的特征, 受季风与台风影响, 年降水量分布不均匀, 干湿两季明显, 且不同区域的年降水量存在差异.如东部与中部属于湿润区, 西部与南部属于半干旱区, 水分相对不足, 北部地区则为半湿润区(唐少霞等, 2008 ).海南岛植物资源丰富, 目前记载的野生植物有4000多种, 外来植物165种, 其中入侵植物有58种(彭宗波等, 2013 ; 杨小波等, 2013 ). ...
海南岛外来植物入侵风险评价指标体系
1
2013
... 海南岛地处热带北缘, 面积3.4万km2 , 是我国第二大岛屿.海南岛气候温和, 年平均气温为22.5-26.0 ℃, 年降水量为1500-2000 mm.海南岛的地势中部高、四周低, 这种地理地貌使得其年总积温、日照时数等都具有自中部向沿海递增的特征, 受季风与台风影响, 年降水量分布不均匀, 干湿两季明显, 且不同区域的年降水量存在差异.如东部与中部属于湿润区, 西部与南部属于半干旱区, 水分相对不足, 北部地区则为半湿润区(唐少霞等, 2008 ).海南岛植物资源丰富, 目前记载的野生植物有4000多种, 外来植物165种, 其中入侵植物有58种(彭宗波等, 2013 ; 杨小波等, 2013 ). ...
海南岛石灰岩地区的外来植物
1
2008
... 海南岛虽然气候条件宜人、生物资源丰富, 但生态环境脆弱, 是许多外来入侵物种的“天然大温室”, 目前有关海南岛入侵植物的研究主要集中在以下两方面: 一是区域性入侵植物种类调查或群落组成调查(单家林, 2003 ; 秦新生等, 2008 ); 二是入侵植物生理生态的研究(Huang et al ., 2013 ; 黄乔乔等, 2013 ).这些研究有效地整合了特定区域内外来植物种类组成的信息, 并在入侵生态学方面做了一定的探索, 但还没有在海南全岛范围内开展主要入侵植物的分布特征及其入侵强度的研究, 有关入侵植物在不同生态系统中的入侵特征及对本地植物多样性影响的研究都还缺乏.本文研究了国家农业部指定的20种恶性入侵植物在海南岛的分布特征及其对当地植物的影响, 并比较了6种主要入侵植物在7种生态系统中的入侵强度, 以期进一步了解海南岛入侵植物的分布现状, 并为外来植物的防控、生物多样性保护与生态安全的管理提供理论依据. ...
海南岛石灰岩地区的外来植物
1
2008
... 海南岛虽然气候条件宜人、生物资源丰富, 但生态环境脆弱, 是许多外来入侵物种的“天然大温室”, 目前有关海南岛入侵植物的研究主要集中在以下两方面: 一是区域性入侵植物种类调查或群落组成调查(单家林, 2003 ; 秦新生等, 2008 ); 二是入侵植物生理生态的研究(Huang et al ., 2013 ; 黄乔乔等, 2013 ).这些研究有效地整合了特定区域内外来植物种类组成的信息, 并在入侵生态学方面做了一定的探索, 但还没有在海南全岛范围内开展主要入侵植物的分布特征及其入侵强度的研究, 有关入侵植物在不同生态系统中的入侵特征及对本地植物多样性影响的研究都还缺乏.本文研究了国家农业部指定的20种恶性入侵植物在海南岛的分布特征及其对当地植物的影响, 并比较了6种主要入侵植物在7种生态系统中的入侵强度, 以期进一步了解海南岛入侵植物的分布现状, 并为外来植物的防控、生物多样性保护与生态安全的管理提供理论依据. ...
What attributes make some plant species more invasive?
1
1996
... 入侵植物在海南岛分布广泛, 且不同区域间种数相差不大, 但整体呈现为东北部、西南部地区种类较多, 东南部次之, 而中部较少的分布特征.入侵植物的分布主要取决于环境的可入侵性和物种的入侵能力两个方面Newsome & Noble, 1986 ; 刘建等, 2010 ).如“理想”的入侵植物通常具备生活史短, 自交亲和性、结实率高, 可进行无性繁殖等特点, 并能迅速响应环境中的干扰Rejmánek & Richardson, 1996 ).本文研究的11种入侵植物中, 除马缨丹外, 其余10种均为一、二年生的草本植物, 其中有6种为菊科植物, 如飞机草、假臭草、微甘菊等, 这些植物产生的种子数量大而质量轻, 通常还附有冠毛, 便于长距离传播和扩散李振宇和谢焱, 2002 ; Weber et al ., 2008 ), 而喜旱莲子草、微甘菊、凤眼蓝、三裂叶蟛蜞菊具有无性繁殖的特性, 其繁殖体可以借助水流进行传播, 这些生物学特征和传播机制增强了其入侵能力, 有助于入侵植物的扩散和迅速占据入侵地, 并能在较短的时间内扩大种群, 从而导致其在海南岛的分布范围很广.但对比不同区域的分布特点发现, 东北部和西南部的入侵种数相对较多, 为10-11种, 而中部和东南部分布较少, 这种差异可能与不同区域间的气候条件和人为干扰强度有关.由于海南岛的西部地区贯穿有环西粤海铁路, 具有较长的运输史, 而连接该地区的海口市、儋州市、东方市都是水陆交通的重要枢纽, 境内人流密集, 干扰强度大, 这些因素都能有效地促进入侵植物的传播和扩散(吴晓雯等, 2006 ; 张帅等, 2010 ).结合气候条件进一步分析, 海南岛西部地区的年晴天数平均为300天, 明显高于中部和东部地区, 但降水量较少, 而中部地区是全岛降水集中分布的地区, 年降水量达2000 mm, 属于湿润地区.此外, 中部地区的年平均气温为23 ℃左右, 处于全岛平均气温(23-26 ℃)的下限, 低于海南岛其他地区的温度(蔡大鑫等, 2010 ).综合气候条件和入侵种数的分布特征发现, 入侵植物在年平均气温较高、降水量较低、而日照时间较长的东北、西南部地区分布的种类较多, 在年平均气温较低、降水量较高、日照时间较短的中部、东部地区分布的种类较少, 这可能与入侵植物的生活习性有关, 因为本文研究的入侵植物多为草本阳性植物, 它们的种子萌发和种群发展都需要足够的光温条件, 过高湿和低温的环境可能对其生长和扩散具有抑制作用. ...
海南岛外来植物群落初探
1
2003
... 海南岛虽然气候条件宜人、生物资源丰富, 但生态环境脆弱, 是许多外来入侵物种的“天然大温室”, 目前有关海南岛入侵植物的研究主要集中在以下两方面: 一是区域性入侵植物种类调查或群落组成调查(单家林, 2003 ; 秦新生等, 2008 ); 二是入侵植物生理生态的研究(Huang et al ., 2013 ; 黄乔乔等, 2013 ).这些研究有效地整合了特定区域内外来植物种类组成的信息, 并在入侵生态学方面做了一定的探索, 但还没有在海南全岛范围内开展主要入侵植物的分布特征及其入侵强度的研究, 有关入侵植物在不同生态系统中的入侵特征及对本地植物多样性影响的研究都还缺乏.本文研究了国家农业部指定的20种恶性入侵植物在海南岛的分布特征及其对当地植物的影响, 并比较了6种主要入侵植物在7种生态系统中的入侵强度, 以期进一步了解海南岛入侵植物的分布现状, 并为外来植物的防控、生物多样性保护与生态安全的管理提供理论依据. ...
海南岛外来植物群落初探
1
2003
... 海南岛虽然气候条件宜人、生物资源丰富, 但生态环境脆弱, 是许多外来入侵物种的“天然大温室”, 目前有关海南岛入侵植物的研究主要集中在以下两方面: 一是区域性入侵植物种类调查或群落组成调查(单家林, 2003 ; 秦新生等, 2008 ); 二是入侵植物生理生态的研究(Huang et al ., 2013 ; 黄乔乔等, 2013 ).这些研究有效地整合了特定区域内外来植物种类组成的信息, 并在入侵生态学方面做了一定的探索, 但还没有在海南全岛范围内开展主要入侵植物的分布特征及其入侵强度的研究, 有关入侵植物在不同生态系统中的入侵特征及对本地植物多样性影响的研究都还缺乏.本文研究了国家农业部指定的20种恶性入侵植物在海南岛的分布特征及其对当地植物的影响, 并比较了6种主要入侵植物在7种生态系统中的入侵强度, 以期进一步了解海南岛入侵植物的分布现状, 并为外来植物的防控、生物多样性保护与生态安全的管理提供理论依据. ...
Community ecology theory as a framework for biological invasions.
1
2002
... 本文研究表明: 弃耕地、村落、农田生态系统中所含的入侵种数最多, 均有10种; 而在林缘生境中, 11种入侵植物的入侵频率和累计入侵频率都较低, 在抽样调查的次生林内, 没有发现入侵植物的分布(表1 ), 说明入侵植物主要分布在人为干扰强度大的生境中, 而在人为干扰强度小、群落结构复杂的生境中分布较少.结合6种生态系统的演替阶段分析, 弃耕地、村落、草地和农田均属于演替早期的生境类型, 受人为干扰的强度大, 而机械和人为活动的介导能有效地减少入侵植物的传播障碍, 或形成干扰间的协同效应(Hobbs, 1991 ); 入侵植物通常能够迅速响应环境中的干扰, 并且具有较强的表型可塑性, 这些特征促成了其在干扰生境中的成功入侵(Hobbs & Huenneke, 1992 ).植物对光能的捕获和利用效率高低是决定其能否适应环境并很好地存活下去的重要因素Baruch & Goldstein, 1999 ; Durand & Goldstein, 2001 ), 因此, 入侵生境的光环境对植物的入侵过程有很大影响, 如弃耕地通常具有高光照、高肥力的环境特征, 这为入侵种群的建立和发展提供了条件, 导致弃耕地常成为入侵植物(如假臭草、三裂叶蟛蜞菊等)集中分布的生境, 这与钟军弟等(2014)在广西的研究结果一致, 表明弃耕地具有较大的可入侵空间.林缘和天然次生林生态系统属于演替中后期的生境类型, 与演替前期的生境类型相比, 受人为干扰强度较低, 而增加的物种密度也将减少群落中的空缺生态位, 从而降低植物成功入侵的几率Stachowicz & Tilman, 2005 ).另一方面, 林缘或林下的光照强度和光合有效辐射都比较低, 这种环境特征也将限制植物入侵林缘的深度Brothers & Spingarn, 1992 ; Shea & Chesson, 2002 ), 因为本文研究的入侵植物多是处于演替初期的草本植物, 这些植物从种子萌发到幼苗生长及种群的发育均需要充足的光照, 它们的入侵可能在演替前期的生境类型, 如农田、草地和村落中具有更好的生态适应性; 但在林缘和林下的入侵却受到光照条件的制约, 且林缘的群落结构对低矮生活型的入侵植物也具有阻碍作用(Honnay et al ., 2002 ), 如喜旱莲子草、含羞草等在林缘中的入侵频率都很低.因此, 入侵植物虽然在林区广泛分布, 但它们仅分布在光照充足的路边或新垦的裸地上, 而无法入侵到森林内部, 国外的不少报道也证实了这种分布特点Parendes & Jones, 2000 ).分析入侵种群的盖度可知, 除三裂叶蟛蜞菊的种群盖度较高外, 其余5种入侵植物的盖度都集中分布在1%-20%之间, 并与入侵地的环境特征存在相关性, 如含羞草在农田、假臭草在弃耕地、三裂叶蟛蜞菊在种植园中通常具有成群分布生长的特点, 体现出这3种入侵植物在对应的生境中有更好的生态适应性; 而在林缘生态系统中, 除马缨丹和苏门白酒草外, 其余4种入侵植物的入侵频率与种群盖度呈负相关关系, 表明在林缘生态系统中, 入侵植物在入侵群落中的密度小, 难以形成单优群落, 这与入侵植物的种子是否能到达林缘, 能否存活及能否萌芽有密切的关系, 这些问题有待进一步实验验证. ...
Introduced species and management of a Nothofagus/Austrocedrus forest.
1
2003
... 有关入侵植物影响生态系统功能的报道有很多, 并多关注其负面效应Williamson, 2001 ; Byers et al ., 2002 ; Simberloff et al ., 2003 ), 认为植物入侵导致本地群落中生物多样性的下降李冰和李玉瑛, 2009 ), 例如: 丁晖等(2007)研究发现, 入侵森林林隙中的紫茎泽兰(Ageratina adenophora )使群落中本地植物的物种丰富度显著下降; Fox等在澳洲西部一个草地生态系统中的研究表明群落中的外来物种比例与本地植物种数呈负相关关系(Stohlgren et al ., 1999 ); Tilman (1997)在美国Bethel市一个草地生态系统中的实验也得出了一致的结论.因此一般认为, 外来植物成功入侵后将抑制群落中本地植物的多样性.但也有研究发现: 入侵植物对群落植物多样性的影响与入侵种及本地植物的生活型有关(Hejda et al ., 2009 ); 植物入侵的生态效应与入侵地的演替阶段有关, 例如: 豆科植物入侵生态环境恶劣的尾矿区后, 能有效地改善土壤肥力, 为演替中期过渡种的定居和发育创造良好的条件, 并促进物种多样性的增加Gozlan & Newton, 2009 ); 李安定等(2013)研究发现, 喜旱莲子草(空心莲子草)(Alternanthera philoxeroides )在入侵的早期阶段能改变群落生境条件, 进而促进其他物种的生存与定居, 并增加物种的多样性, 而只有当其种群盖度持续增加后, 才可能会威胁其他物种的生存; Hejda等(2009)研究发现, 入侵植物的盖度是决定群落物种丰富度和均匀度的主要因子, 入侵植物对物种多样性的影响与本地植物优势种的盖度、高度有密切关系.因此, 植物入侵的生态效应受生态系统类型、群落演替阶段的影响; 不同功能群植物对植物入侵的响应也有差异.本文通过研究7种生态系统类型中入侵植物的入侵特征及其对本地植物的影响, 旨在检验以下科学假设: 1)植物入侵的生态效应与入侵地的环境特征和演替阶段有关, 入侵植物可能对入侵地产生正面或负面影响; 2)植物入侵主要影响群落中某一功能群的物种多样性, 而不是影响全部群落个体; 3)多数入侵植物有偏好的入侵生境, 而人为干扰少的生态系统类型对植物入侵有抵制作用. ...
Biometry. 3rd edn
1
1995
... 选择农田、村落、种植园、林缘、弃耕地、草地、次生林7种人为干扰程度有差异和不同演替特征的生境类型作为研究对象, 结合调查中各生态系统类型的入侵程度和实地调查的可行性, 分别在农田、村落、种植园、林缘、弃耕地、草地、次生林中选取14、14、20、11、17、11、10个共97个样地的信息用于数据统计分析(图1 ), 分析的样地共含970个样方, 总面积为3904 m2 .以1套样方中入侵样方所占比例作为入侵频率, 用以比较不同生态系统中入侵植物的入侵频率, 这样, 每个样地有2个重复.目前, 有关植物盖度等级的划分多出现在植被覆盖度估算的研究中(盖永芹等, 2009 ; 李锦荣等; 2010 ), 而对样方中种群盖度等级的划分没有统一的标准, 因此, 在划分盖度等级时, 本文根据评估标准及分布范围, 把入侵植物的种群盖度分为一级(1%-20%)、二级(21%-40%)、三级(41%-60%)、四级(≥61%)共4个等级.运用Excel 2003和SPSS 17.0软件进行数据统计分析.用Systat SigmaPlot 10.0软件绘图.由于调查数据中入侵频率和盖度两项指标是服从二项分布的百分数的数据资料, 并且含有小于30%或大于70%的数值, 因此, 数据分析前对其进行反正弦转换Sokal & Rohlf, 1995 ). ...
1
... 本文研究表明: 弃耕地、村落、农田生态系统中所含的入侵种数最多, 均有10种; 而在林缘生境中, 11种入侵植物的入侵频率和累计入侵频率都较低, 在抽样调查的次生林内, 没有发现入侵植物的分布(表1 ), 说明入侵植物主要分布在人为干扰强度大的生境中, 而在人为干扰强度小、群落结构复杂的生境中分布较少.结合6种生态系统的演替阶段分析, 弃耕地、村落、草地和农田均属于演替早期的生境类型, 受人为干扰的强度大, 而机械和人为活动的介导能有效地减少入侵植物的传播障碍, 或形成干扰间的协同效应(Hobbs, 1991 ); 入侵植物通常能够迅速响应环境中的干扰, 并且具有较强的表型可塑性, 这些特征促成了其在干扰生境中的成功入侵(Hobbs & Huenneke, 1992 ).植物对光能的捕获和利用效率高低是决定其能否适应环境并很好地存活下去的重要因素Baruch & Goldstein, 1999 ; Durand & Goldstein, 2001 ), 因此, 入侵生境的光环境对植物的入侵过程有很大影响, 如弃耕地通常具有高光照、高肥力的环境特征, 这为入侵种群的建立和发展提供了条件, 导致弃耕地常成为入侵植物(如假臭草、三裂叶蟛蜞菊等)集中分布的生境, 这与钟军弟等(2014)在广西的研究结果一致, 表明弃耕地具有较大的可入侵空间.林缘和天然次生林生态系统属于演替中后期的生境类型, 与演替前期的生境类型相比, 受人为干扰强度较低, 而增加的物种密度也将减少群落中的空缺生态位, 从而降低植物成功入侵的几率Stachowicz & Tilman, 2005 ).另一方面, 林缘或林下的光照强度和光合有效辐射都比较低, 这种环境特征也将限制植物入侵林缘的深度Brothers & Spingarn, 1992 ; Shea & Chesson, 2002 ), 因为本文研究的入侵植物多是处于演替初期的草本植物, 这些植物从种子萌发到幼苗生长及种群的发育均需要充足的光照, 它们的入侵可能在演替前期的生境类型, 如农田、草地和村落中具有更好的生态适应性; 但在林缘和林下的入侵却受到光照条件的制约, 且林缘的群落结构对低矮生活型的入侵植物也具有阻碍作用(Honnay et al ., 2002 ), 如喜旱莲子草、含羞草等在林缘中的入侵频率都很低.因此, 入侵植物虽然在林区广泛分布, 但它们仅分布在光照充足的路边或新垦的裸地上, 而无法入侵到森林内部, 国外的不少报道也证实了这种分布特点Parendes & Jones, 2000 ).分析入侵种群的盖度可知, 除三裂叶蟛蜞菊的种群盖度较高外, 其余5种入侵植物的盖度都集中分布在1%-20%之间, 并与入侵地的环境特征存在相关性, 如含羞草在农田、假臭草在弃耕地、三裂叶蟛蜞菊在种植园中通常具有成群分布生长的特点, 体现出这3种入侵植物在对应的生境中有更好的生态适应性; 而在林缘生态系统中, 除马缨丹和苏门白酒草外, 其余4种入侵植物的入侵频率与种群盖度呈负相关关系, 表明在林缘生态系统中, 入侵植物在入侵群落中的密度小, 难以形成单优群落, 这与入侵植物的种子是否能到达林缘, 能否存活及能否萌芽有密切的关系, 这些问题有待进一步实验验证. ...
Exotic plant species invade hot spots of native plant diversity.
1
1999
... 有关入侵植物影响生态系统功能的报道有很多, 并多关注其负面效应Williamson, 2001 ; Byers et al ., 2002 ; Simberloff et al ., 2003 ), 认为植物入侵导致本地群落中生物多样性的下降李冰和李玉瑛, 2009 ), 例如: 丁晖等(2007)研究发现, 入侵森林林隙中的紫茎泽兰(Ageratina adenophora )使群落中本地植物的物种丰富度显著下降; Fox等在澳洲西部一个草地生态系统中的研究表明群落中的外来物种比例与本地植物种数呈负相关关系(Stohlgren et al ., 1999 ); Tilman (1997)在美国Bethel市一个草地生态系统中的实验也得出了一致的结论.因此一般认为, 外来植物成功入侵后将抑制群落中本地植物的多样性.但也有研究发现: 入侵植物对群落植物多样性的影响与入侵种及本地植物的生活型有关(Hejda et al ., 2009 ); 植物入侵的生态效应与入侵地的演替阶段有关, 例如: 豆科植物入侵生态环境恶劣的尾矿区后, 能有效地改善土壤肥力, 为演替中期过渡种的定居和发育创造良好的条件, 并促进物种多样性的增加Gozlan & Newton, 2009 ); 李安定等(2013)研究发现, 喜旱莲子草(空心莲子草)(Alternanthera philoxeroides )在入侵的早期阶段能改变群落生境条件, 进而促进其他物种的生存与定居, 并增加物种的多样性, 而只有当其种群盖度持续增加后, 才可能会威胁其他物种的生存; Hejda等(2009)研究发现, 入侵植物的盖度是决定群落物种丰富度和均匀度的主要因子, 入侵植物对物种多样性的影响与本地植物优势种的盖度、高度有密切关系.因此, 植物入侵的生态效应受生态系统类型、群落演替阶段的影响; 不同功能群植物对植物入侵的响应也有差异.本文通过研究7种生态系统类型中入侵植物的入侵特征及其对本地植物的影响, 旨在检验以下科学假设: 1)植物入侵的生态效应与入侵地的环境特征和演替阶段有关, 入侵植物可能对入侵地产生正面或负面影响; 2)植物入侵主要影响群落中某一功能群的物种多样性, 而不是影响全部群落个体; 3)多数入侵植物有偏好的入侵生境, 而人为干扰少的生态系统类型对植物入侵有抵制作用. ...
海南岛气候资源特征及其开发利用
1
2008
... 海南岛地处热带北缘, 面积3.4万km2 , 是我国第二大岛屿.海南岛气候温和, 年平均气温为22.5-26.0 ℃, 年降水量为1500-2000 mm.海南岛的地势中部高、四周低, 这种地理地貌使得其年总积温、日照时数等都具有自中部向沿海递增的特征, 受季风与台风影响, 年降水量分布不均匀, 干湿两季明显, 且不同区域的年降水量存在差异.如东部与中部属于湿润区, 西部与南部属于半干旱区, 水分相对不足, 北部地区则为半湿润区(唐少霞等, 2008 ).海南岛植物资源丰富, 目前记载的野生植物有4000多种, 外来植物165种, 其中入侵植物有58种(彭宗波等, 2013 ; 杨小波等, 2013 ). ...
海南岛气候资源特征及其开发利用
1
2008
... 海南岛地处热带北缘, 面积3.4万km2 , 是我国第二大岛屿.海南岛气候温和, 年平均气温为22.5-26.0 ℃, 年降水量为1500-2000 mm.海南岛的地势中部高、四周低, 这种地理地貌使得其年总积温、日照时数等都具有自中部向沿海递增的特征, 受季风与台风影响, 年降水量分布不均匀, 干湿两季明显, 且不同区域的年降水量存在差异.如东部与中部属于湿润区, 西部与南部属于半干旱区, 水分相对不足, 北部地区则为半湿润区(唐少霞等, 2008 ).海南岛植物资源丰富, 目前记载的野生植物有4000多种, 外来植物165种, 其中入侵植物有58种(彭宗波等, 2013 ; 杨小波等, 2013 ). ...
Community invisibility, recruitment limitation and grassland biodiversity.
0
1997
新农村建设对伴人植物多样性的影响——以临汾市尧都区为例
0
2013
新农村建设对伴人植物多样性的影响——以临汾市尧都区为例
0
2013
北京地区外来入侵植物分布特征及其影响因素
0
2012
北京地区外来入侵植物分布特征及其影响因素
0
2012
Invasive alien plants in China: Diversity and ecological insights.
1
2008
... 入侵植物在海南岛分布广泛, 且不同区域间种数相差不大, 但整体呈现为东北部、西南部地区种类较多, 东南部次之, 而中部较少的分布特征.入侵植物的分布主要取决于环境的可入侵性和物种的入侵能力两个方面Newsome & Noble, 1986 ; 刘建等, 2010 ).如“理想”的入侵植物通常具备生活史短, 自交亲和性、结实率高, 可进行无性繁殖等特点, 并能迅速响应环境中的干扰Rejmánek & Richardson, 1996 ).本文研究的11种入侵植物中, 除马缨丹外, 其余10种均为一、二年生的草本植物, 其中有6种为菊科植物, 如飞机草、假臭草、微甘菊等, 这些植物产生的种子数量大而质量轻, 通常还附有冠毛, 便于长距离传播和扩散李振宇和谢焱, 2002 ; Weber et al ., 2008 ), 而喜旱莲子草、微甘菊、凤眼蓝、三裂叶蟛蜞菊具有无性繁殖的特性, 其繁殖体可以借助水流进行传播, 这些生物学特征和传播机制增强了其入侵能力, 有助于入侵植物的扩散和迅速占据入侵地, 并能在较短的时间内扩大种群, 从而导致其在海南岛的分布范围很广.但对比不同区域的分布特点发现, 东北部和西南部的入侵种数相对较多, 为10-11种, 而中部和东南部分布较少, 这种差异可能与不同区域间的气候条件和人为干扰强度有关.由于海南岛的西部地区贯穿有环西粤海铁路, 具有较长的运输史, 而连接该地区的海口市、儋州市、东方市都是水陆交通的重要枢纽, 境内人流密集, 干扰强度大, 这些因素都能有效地促进入侵植物的传播和扩散(吴晓雯等, 2006 ; 张帅等, 2010 ).结合气候条件进一步分析, 海南岛西部地区的年晴天数平均为300天, 明显高于中部和东部地区, 但降水量较少, 而中部地区是全岛降水集中分布的地区, 年降水量达2000 mm, 属于湿润地区.此外, 中部地区的年平均气温为23 ℃左右, 处于全岛平均气温(23-26 ℃)的下限, 低于海南岛其他地区的温度(蔡大鑫等, 2010 ).综合气候条件和入侵种数的分布特征发现, 入侵植物在年平均气温较高、降水量较低、而日照时间较长的东北、西南部地区分布的种类较多, 在年平均气温较低、降水量较高、日照时间较短的中部、东部地区分布的种类较少, 这可能与入侵植物的生活习性有关, 因为本文研究的入侵植物多为草本阳性植物, 它们的种子萌发和种群发展都需要足够的光温条件, 过高湿和低温的环境可能对其生长和扩散具有抑制作用. ...
Is the invasion of the common reed, Phragmites australis, into tidal marshes of the Eastern US an ecological disaster?
1
2003
... 外来物种入侵已成为严重危害陆地及海洋环境的生态问题, 被认为是一种生物污染而被广泛关注和研究(Weis & Weis, 2003 ).深入了解外来物种的入侵机制, 并进行有效的预测、预防和管理是减少入侵生物危害的最有效手段(Byers, 2002 ), 而从区域尺度上分析入侵物种的分布特征并探索其形成原因是认识生物入侵问题的重要基础(Erik, 2004 ). ...
1
... 有关入侵植物影响生态系统功能的报道有很多, 并多关注其负面效应Williamson, 2001 ; Byers et al ., 2002 ; Simberloff et al ., 2003 ), 认为植物入侵导致本地群落中生物多样性的下降李冰和李玉瑛, 2009 ), 例如: 丁晖等(2007)研究发现, 入侵森林林隙中的紫茎泽兰(Ageratina adenophora )使群落中本地植物的物种丰富度显著下降; Fox等在澳洲西部一个草地生态系统中的研究表明群落中的外来物种比例与本地植物种数呈负相关关系(Stohlgren et al ., 1999 ); Tilman (1997)在美国Bethel市一个草地生态系统中的实验也得出了一致的结论.因此一般认为, 外来植物成功入侵后将抑制群落中本地植物的多样性.但也有研究发现: 入侵植物对群落植物多样性的影响与入侵种及本地植物的生活型有关(Hejda et al ., 2009 ); 植物入侵的生态效应与入侵地的演替阶段有关, 例如: 豆科植物入侵生态环境恶劣的尾矿区后, 能有效地改善土壤肥力, 为演替中期过渡种的定居和发育创造良好的条件, 并促进物种多样性的增加Gozlan & Newton, 2009 ); 李安定等(2013)研究发现, 喜旱莲子草(空心莲子草)(Alternanthera philoxeroides )在入侵的早期阶段能改变群落生境条件, 进而促进其他物种的生存与定居, 并增加物种的多样性, 而只有当其种群盖度持续增加后, 才可能会威胁其他物种的生存; Hejda等(2009)研究发现, 入侵植物的盖度是决定群落物种丰富度和均匀度的主要因子, 入侵植物对物种多样性的影响与本地植物优势种的盖度、高度有密切关系.因此, 植物入侵的生态效应受生态系统类型、群落演替阶段的影响; 不同功能群植物对植物入侵的响应也有差异.本文通过研究7种生态系统类型中入侵植物的入侵特征及其对本地植物的影响, 旨在检验以下科学假设: 1)植物入侵的生态效应与入侵地的环境特征和演替阶段有关, 入侵植物可能对入侵地产生正面或负面影响; 2)植物入侵主要影响群落中某一功能群的物种多样性, 而不是影响全部群落个体; 3)多数入侵植物有偏好的入侵生境, 而人为干扰少的生态系统类型对植物入侵有抵制作用. ...
中国外来入侵植物的分布格局及其与环境因子和人类活动的关系
1
2006
... 入侵植物在海南岛分布广泛, 且不同区域间种数相差不大, 但整体呈现为东北部、西南部地区种类较多, 东南部次之, 而中部较少的分布特征.入侵植物的分布主要取决于环境的可入侵性和物种的入侵能力两个方面Newsome & Noble, 1986 ; 刘建等, 2010 ).如“理想”的入侵植物通常具备生活史短, 自交亲和性、结实率高, 可进行无性繁殖等特点, 并能迅速响应环境中的干扰Rejmánek & Richardson, 1996 ).本文研究的11种入侵植物中, 除马缨丹外, 其余10种均为一、二年生的草本植物, 其中有6种为菊科植物, 如飞机草、假臭草、微甘菊等, 这些植物产生的种子数量大而质量轻, 通常还附有冠毛, 便于长距离传播和扩散李振宇和谢焱, 2002 ; Weber et al ., 2008 ), 而喜旱莲子草、微甘菊、凤眼蓝、三裂叶蟛蜞菊具有无性繁殖的特性, 其繁殖体可以借助水流进行传播, 这些生物学特征和传播机制增强了其入侵能力, 有助于入侵植物的扩散和迅速占据入侵地, 并能在较短的时间内扩大种群, 从而导致其在海南岛的分布范围很广.但对比不同区域的分布特点发现, 东北部和西南部的入侵种数相对较多, 为10-11种, 而中部和东南部分布较少, 这种差异可能与不同区域间的气候条件和人为干扰强度有关.由于海南岛的西部地区贯穿有环西粤海铁路, 具有较长的运输史, 而连接该地区的海口市、儋州市、东方市都是水陆交通的重要枢纽, 境内人流密集, 干扰强度大, 这些因素都能有效地促进入侵植物的传播和扩散(吴晓雯等, 2006 ; 张帅等, 2010 ).结合气候条件进一步分析, 海南岛西部地区的年晴天数平均为300天, 明显高于中部和东部地区, 但降水量较少, 而中部地区是全岛降水集中分布的地区, 年降水量达2000 mm, 属于湿润地区.此外, 中部地区的年平均气温为23 ℃左右, 处于全岛平均气温(23-26 ℃)的下限, 低于海南岛其他地区的温度(蔡大鑫等, 2010 ).综合气候条件和入侵种数的分布特征发现, 入侵植物在年平均气温较高、降水量较低、而日照时间较长的东北、西南部地区分布的种类较多, 在年平均气温较低、降水量较高、日照时间较短的中部、东部地区分布的种类较少, 这可能与入侵植物的生活习性有关, 因为本文研究的入侵植物多为草本阳性植物, 它们的种子萌发和种群发展都需要足够的光温条件, 过高湿和低温的环境可能对其生长和扩散具有抑制作用. ...
中国外来入侵植物的分布格局及其与环境因子和人类活动的关系
1
2006
... 入侵植物在海南岛分布广泛, 且不同区域间种数相差不大, 但整体呈现为东北部、西南部地区种类较多, 东南部次之, 而中部较少的分布特征.入侵植物的分布主要取决于环境的可入侵性和物种的入侵能力两个方面Newsome & Noble, 1986 ; 刘建等, 2010 ).如“理想”的入侵植物通常具备生活史短, 自交亲和性、结实率高, 可进行无性繁殖等特点, 并能迅速响应环境中的干扰Rejmánek & Richardson, 1996 ).本文研究的11种入侵植物中, 除马缨丹外, 其余10种均为一、二年生的草本植物, 其中有6种为菊科植物, 如飞机草、假臭草、微甘菊等, 这些植物产生的种子数量大而质量轻, 通常还附有冠毛, 便于长距离传播和扩散李振宇和谢焱, 2002 ; Weber et al ., 2008 ), 而喜旱莲子草、微甘菊、凤眼蓝、三裂叶蟛蜞菊具有无性繁殖的特性, 其繁殖体可以借助水流进行传播, 这些生物学特征和传播机制增强了其入侵能力, 有助于入侵植物的扩散和迅速占据入侵地, 并能在较短的时间内扩大种群, 从而导致其在海南岛的分布范围很广.但对比不同区域的分布特点发现, 东北部和西南部的入侵种数相对较多, 为10-11种, 而中部和东南部分布较少, 这种差异可能与不同区域间的气候条件和人为干扰强度有关.由于海南岛的西部地区贯穿有环西粤海铁路, 具有较长的运输史, 而连接该地区的海口市、儋州市、东方市都是水陆交通的重要枢纽, 境内人流密集, 干扰强度大, 这些因素都能有效地促进入侵植物的传播和扩散(吴晓雯等, 2006 ; 张帅等, 2010 ).结合气候条件进一步分析, 海南岛西部地区的年晴天数平均为300天, 明显高于中部和东部地区, 但降水量较少, 而中部地区是全岛降水集中分布的地区, 年降水量达2000 mm, 属于湿润地区.此外, 中部地区的年平均气温为23 ℃左右, 处于全岛平均气温(23-26 ℃)的下限, 低于海南岛其他地区的温度(蔡大鑫等, 2010 ).综合气候条件和入侵种数的分布特征发现, 入侵植物在年平均气温较高、降水量较低、而日照时间较长的东北、西南部地区分布的种类较多, 在年平均气温较低、降水量较高、日照时间较短的中部、东部地区分布的种类较少, 这可能与入侵植物的生活习性有关, 因为本文研究的入侵植物多为草本阳性植物, 它们的种子萌发和种群发展都需要足够的光温条件, 过高湿和低温的环境可能对其生长和扩散具有抑制作用. ...
1
2013
... 海南岛地处热带北缘, 面积3.4万km2 , 是我国第二大岛屿.海南岛气候温和, 年平均气温为22.5-26.0 ℃, 年降水量为1500-2000 mm.海南岛的地势中部高、四周低, 这种地理地貌使得其年总积温、日照时数等都具有自中部向沿海递增的特征, 受季风与台风影响, 年降水量分布不均匀, 干湿两季明显, 且不同区域的年降水量存在差异.如东部与中部属于湿润区, 西部与南部属于半干旱区, 水分相对不足, 北部地区则为半湿润区(唐少霞等, 2008 ).海南岛植物资源丰富, 目前记载的野生植物有4000多种, 外来植物165种, 其中入侵植物有58种(彭宗波等, 2013 ; 杨小波等, 2013 ). ...
1
2013
... 海南岛地处热带北缘, 面积3.4万km2 , 是我国第二大岛屿.海南岛气候温和, 年平均气温为22.5-26.0 ℃, 年降水量为1500-2000 mm.海南岛的地势中部高、四周低, 这种地理地貌使得其年总积温、日照时数等都具有自中部向沿海递增的特征, 受季风与台风影响, 年降水量分布不均匀, 干湿两季明显, 且不同区域的年降水量存在差异.如东部与中部属于湿润区, 西部与南部属于半干旱区, 水分相对不足, 北部地区则为半湿润区(唐少霞等, 2008 ).海南岛植物资源丰富, 目前记载的野生植物有4000多种, 外来植物165种, 其中入侵植物有58种(彭宗波等, 2013 ; 杨小波等, 2013 ). ...
我国入侵植物多样性的区域分异及其影响因素——以74个地区数据为基础
1
2010
... 入侵植物在海南岛分布广泛, 且不同区域间种数相差不大, 但整体呈现为东北部、西南部地区种类较多, 东南部次之, 而中部较少的分布特征.入侵植物的分布主要取决于环境的可入侵性和物种的入侵能力两个方面Newsome & Noble, 1986 ; 刘建等, 2010 ).如“理想”的入侵植物通常具备生活史短, 自交亲和性、结实率高, 可进行无性繁殖等特点, 并能迅速响应环境中的干扰Rejmánek & Richardson, 1996 ).本文研究的11种入侵植物中, 除马缨丹外, 其余10种均为一、二年生的草本植物, 其中有6种为菊科植物, 如飞机草、假臭草、微甘菊等, 这些植物产生的种子数量大而质量轻, 通常还附有冠毛, 便于长距离传播和扩散李振宇和谢焱, 2002 ; Weber et al ., 2008 ), 而喜旱莲子草、微甘菊、凤眼蓝、三裂叶蟛蜞菊具有无性繁殖的特性, 其繁殖体可以借助水流进行传播, 这些生物学特征和传播机制增强了其入侵能力, 有助于入侵植物的扩散和迅速占据入侵地, 并能在较短的时间内扩大种群, 从而导致其在海南岛的分布范围很广.但对比不同区域的分布特点发现, 东北部和西南部的入侵种数相对较多, 为10-11种, 而中部和东南部分布较少, 这种差异可能与不同区域间的气候条件和人为干扰强度有关.由于海南岛的西部地区贯穿有环西粤海铁路, 具有较长的运输史, 而连接该地区的海口市、儋州市、东方市都是水陆交通的重要枢纽, 境内人流密集, 干扰强度大, 这些因素都能有效地促进入侵植物的传播和扩散(吴晓雯等, 2006 ; 张帅等, 2010 ).结合气候条件进一步分析, 海南岛西部地区的年晴天数平均为300天, 明显高于中部和东部地区, 但降水量较少, 而中部地区是全岛降水集中分布的地区, 年降水量达2000 mm, 属于湿润地区.此外, 中部地区的年平均气温为23 ℃左右, 处于全岛平均气温(23-26 ℃)的下限, 低于海南岛其他地区的温度(蔡大鑫等, 2010 ).综合气候条件和入侵种数的分布特征发现, 入侵植物在年平均气温较高、降水量较低、而日照时间较长的东北、西南部地区分布的种类较多, 在年平均气温较低、降水量较高、日照时间较短的中部、东部地区分布的种类较少, 这可能与入侵植物的生活习性有关, 因为本文研究的入侵植物多为草本阳性植物, 它们的种子萌发和种群发展都需要足够的光温条件, 过高湿和低温的环境可能对其生长和扩散具有抑制作用. ...
我国入侵植物多样性的区域分异及其影响因素——以74个地区数据为基础
1
2010
... 入侵植物在海南岛分布广泛, 且不同区域间种数相差不大, 但整体呈现为东北部、西南部地区种类较多, 东南部次之, 而中部较少的分布特征.入侵植物的分布主要取决于环境的可入侵性和物种的入侵能力两个方面Newsome & Noble, 1986 ; 刘建等, 2010 ).如“理想”的入侵植物通常具备生活史短, 自交亲和性、结实率高, 可进行无性繁殖等特点, 并能迅速响应环境中的干扰Rejmánek & Richardson, 1996 ).本文研究的11种入侵植物中, 除马缨丹外, 其余10种均为一、二年生的草本植物, 其中有6种为菊科植物, 如飞机草、假臭草、微甘菊等, 这些植物产生的种子数量大而质量轻, 通常还附有冠毛, 便于长距离传播和扩散李振宇和谢焱, 2002 ; Weber et al ., 2008 ), 而喜旱莲子草、微甘菊、凤眼蓝、三裂叶蟛蜞菊具有无性繁殖的特性, 其繁殖体可以借助水流进行传播, 这些生物学特征和传播机制增强了其入侵能力, 有助于入侵植物的扩散和迅速占据入侵地, 并能在较短的时间内扩大种群, 从而导致其在海南岛的分布范围很广.但对比不同区域的分布特点发现, 东北部和西南部的入侵种数相对较多, 为10-11种, 而中部和东南部分布较少, 这种差异可能与不同区域间的气候条件和人为干扰强度有关.由于海南岛的西部地区贯穿有环西粤海铁路, 具有较长的运输史, 而连接该地区的海口市、儋州市、东方市都是水陆交通的重要枢纽, 境内人流密集, 干扰强度大, 这些因素都能有效地促进入侵植物的传播和扩散(吴晓雯等, 2006 ; 张帅等, 2010 ).结合气候条件进一步分析, 海南岛西部地区的年晴天数平均为300天, 明显高于中部和东部地区, 但降水量较少, 而中部地区是全岛降水集中分布的地区, 年降水量达2000 mm, 属于湿润地区.此外, 中部地区的年平均气温为23 ℃左右, 处于全岛平均气温(23-26 ℃)的下限, 低于海南岛其他地区的温度(蔡大鑫等, 2010 ).综合气候条件和入侵种数的分布特征发现, 入侵植物在年平均气温较高、降水量较低、而日照时间较长的东北、西南部地区分布的种类较多, 在年平均气温较低、降水量较高、日照时间较短的中部、东部地区分布的种类较少, 这可能与入侵植物的生活习性有关, 因为本文研究的入侵植物多为草本阳性植物, 它们的种子萌发和种群发展都需要足够的光温条件, 过高湿和低温的环境可能对其生长和扩散具有抑制作用. ...
 , 陈玉凯
, 陈玉凯


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}