苔藓植物是仅次于被子植物的第二大类高等植物类群, 在沼泽、森林、草地和荒漠区均起着重要的生态作用(Mallón et al ., 2007 )。作为环境演替的先锋物种, 耐旱苔藓植物在荒漠生态系统中分布广泛, 它们的存在能够在地表构筑一个粗糙的平面, 对荒漠生态系统的维持和稳定(Kidron et al ., 2009 )、土壤微环境的改善(Harper & Belnap, 2001 )具有重要 作用。
荒漠苔藓作为典型的抗胁迫变水植物已被广泛关注, 诸多研究表明藓类植物能够适应极端高低温、干燥、强光照等恶劣环境(张静和张元明, 2014 )。为适应极端环境, 藓类植物在种群分布、形态特征(郑云普等, 2009 ; 陶冶和张元明, 2012 )、繁殖方式(Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 )、光合活性(张静和张元明, 2011 )、渗透调节物质、抗氧化酶(回嵘等, 2012 ; 陈文佳等, 2013 )等方面进化出多样的抗胁迫特性。研究发现齿肋赤藓(Syntrichia caninervis )叶片表面具有蜡质层, 并在细胞表面富集疣状突起, 在一定程度上能够减弱热气流的影响(许书军, 2007 )。另外, 在高温干燥环境下藓类植物能够通过失水降低代谢活性而进入休眠状态, 并积累大量的脯氨酸和可溶性糖来提高细胞渗透压, 维持渗透压平衡(Oliver et al ., 1997 ; Crowe et al ., 1998 ; 徐杰等, 2005 ; 吴楠等, 2009 )。同时, 通过抗氧化酶的大量合成清除过多的氧自由基, 进而保护植物免受过度伤害(Sgherri et al ., 1994 ; Herrnstadt & Kidron, 2005 ; Pintado et al ., 2005 ; 沈蕾等, 2011 )。但对荒漠藓类植物如何通过生理生化调节并成功度过冬季的极端低温环境, 还有待于深入研究。
在荒漠干燥少雨环境中, 藓类植物在全年的大部分时间均处于失活休眠状态。然而, 有研究发现, 荒漠苔藓的生长和繁殖主要发生在寒冷和湿润的冬季月份Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 )。既使已经冷适应的苔藓, 在温度、光照和湿度达到阈值时仍能够被快速激活(Lange, 2003 ; Kappen & Valladares, 2007 ; Bjerke 2011 ; Su et al ., 2013 )。初冬和早春的积雪融化和低温能够激活藓类植物的光合活性, 延长生物结皮光合活性时间, 增加结皮土壤的碳截获量(Su et al ., 2013 ; 尹本丰和张元明, 2014 )。因此, 苔藓植物更可能在初冬或初春积雪融化时表现出良好的生长特性。但关于寒冷生态系统不同微生境苔藓的生态学特征还知之甚少(Bjerke et al ., 2011 )。
已有的研究和我们前期的实验表明, 微生境的不同也会造成藓类植物生存策略的差异。不少野外调查和研究发现, 遮阴下(灌丛或阴坡)苔藓植物的盖度和生长状况好于暴露环境下的苔藓(Pintado et al ., 2005 ; Zhang et al ., 2007 )。灌丛下的低温、低光照和高含水量与裸露地的高温、高光照和低含水量造成了不同微生境间苔藓形态的变化。为了适应不同微生境, 苔藓植物能够随环境变化而变化, 但是短期微生境变化对苔藓形态的影响并不明显, 而其光合作用与现有生存环境相似(Reynolds & Mc- Letchie, 2011 )。Benassi等(2011)的研究发现: 不同生境下苔藓的繁殖方式也存在显著差异, 遮阴下的苔藓植株更趋于有性繁殖。然而, 不同微生境下苔藓植物对冻融过程中温度和水分变化的适应性是否一致尚不得而知。
因此, 本研究选取古尔班通古特沙漠生物结皮中的优势藓类齿肋赤藓为研究对象, 通过比较3个不同时期(冬季冻结期、早春融雪期和春季中旬干旱无雨期)和3种不同微生境(活灌丛、死灌丛、裸露地) (图1 , 图2 )下齿肋赤藓植株的含水量、渗透调节物质含量、丙二醛含量和抗氧化酶活性的变化特征。试图回答以下科学问题: (1)荒漠藓类植物是怎样通过其生理变化来适应由冬季低温冻结到早春融雪复水再到春季自然干燥过程的? (2)不同微生境下的齿肋赤藓对这种变化的适应性是否存在差异?
图1 古尔班通古特沙漠3种不同微生境下生长的齿肋赤藓。A, 裸露地。B, 死灌丛。C, 活灌丛。
Fig. 1 Syntrichia caninervis grown under three microhabitats in Gurbantünggüt Desert. A, Exposed ground. B, Dead shrub. C, Live shrub.
图2 古尔班通古特沙漠三种不同微生境下齿肋赤藓植株。A, 裸露地。B, 死灌丛。C, 活灌丛。
Fig. 2 Syntrichia caninervis shoots collected from three microhabitats in Gurbantünggüt Desert. A, Exposed ground. B, Dead shrub. C, Live shrub.
1 材料和方法
1.1 研究区概况
实验地点位于新疆准噶尔盆地中部的古尔班通古特沙漠(44.18°-46.33° N, 80.52°-90.00° E, 海拔300-600 m)。该沙漠面积4.88 × 104 km2 , 是中国最大的固定和半固定沙漠。受喜马拉雅山的阻隔作用, 来自印度洋西南季风的水汽不能到达内陆, 致使该沙漠属于典型的温带大陆性气候。沙漠年降水量仅70-150 mm, 年蒸发量达2606.6 mm, 年平均气温7.3 ℃, 冬季平均气温-16.8 ℃。与其他沙漠 不同, 该沙漠在冬季长期积雪覆盖, 稳定积雪日数一般在100-150天, 平均积雪厚度20-30 cm, 占全年降水的25%以上(Zhang et al ., 2007 ; 周宏飞等, 2010 )。
该沙漠春季融雪使得地表土壤含水量显著增加, 荒漠短命植物、类短命植物、一年及多年生草本和隐花植物广泛发育和生长。白梭梭(Haloxylon persicum) 和梭梭(Haloxylon ammodendron )等小半乔木作为古尔班通古特沙漠的建群种, 分别占据了沙垄的上部和中下部。蛇麻黄(Ephedra distachya )、淡枝沙拐枣(Calligonum leucocladum )等灌木也在沙垄间广泛分布。同时, 灌木和生物结皮镶嵌分布是荒漠区的主要地表景观。该沙漠生物结皮主要分为藻结皮、地衣结皮和苔藓结皮3种类型(张元明, 2005 )。其中, 藻结皮多分布在沙垄中上部, 地衣和苔藓结皮多分布在中下部和丘间低地, 尤其在蛇麻黄、沙拐枣(Calligonum spp.)等灌丛下通常丛聚着发育良好的苔藓结皮。
1.2 研究对象的选择
齿肋赤藓属于丛藓科赤藓属, 具有良好的耐旱性和变水特性, 是古尔班通古特沙漠苔藓结皮的优势种。该种较山墙藓(Tortula ruralis )和北方赤藓(Syntrichia norvegica )更能适应极端干旱、低海拔和高温环境, 在世界冷沙漠中具有广泛分布(Reynolds & McLetchie, 2011 )。在干燥条件下, 其个体通常呈现为黑色或淡棕色, 复水后能快速变为绿色, 并在短时间内恢复光合和生理活性张静和张元明, 2014 )。齿肋赤藓种群的大量存在对沙漠的固定和微环境的改善具有重要作用(张元明等, 2004 ; 吴楠等, 2009 )。
1.3 样方设置和样品采集
实验样地设在古尔班通古特沙漠腹地(45.40° N, 88.00° E), 于2013年11月10日, 在研究区选择苔藓结皮发育良好的丘间低地, 设置30 m × 30 m的样方。在样方内分别选择活灌丛(蛇麻黄)、死灌丛(蛇麻黄)和裸露地3种微生境各5块, 大小为1 m × 1 m, 代表5 个重复, 各小样方之间相距2-5 m。并在各小样方上进行标记, 以便在积雪覆盖下能准确找到待测位点。在各生境样方附近地表下5 cm土层埋设ECH2 O监测系统(5TM, Li-COR, Lincoln, USA), 对地表土壤含水量、温度进行连续监测, 每15 min记录一次。同时, 在苔藓表面和距地面50 cm处空中分别安置Microlab lite U盘地温计(liter 5016, Fourier, Israel)和LI-6400光量子探头(LI-COR, Lincoln, USA), 对苔藓表面和大气温度、光照强度进行连续监测, 每30 min记录一次。
实验于2014年2至4月进行。采样时间分别为积雪覆盖下(2月冻结期)、积雪融化期(3月)和春季干燥无雨期(4月)。分别于17:00左右采集不同微生境下的齿肋赤藓样品, 迅速置于装有冰袋的保温箱内。由于齿肋赤藓随环境变化其生理生化特性变化较快(Wu et al ., 2014 ), 为尽可能保持样品原状, 在日落后的低温状态下, 借助辅助光源, 在室外用刀片快速切取苔藓(随用随取)茎叶鲜样。样品过筛3次后(去除夹杂的沙粒), 为每一指标称取150 mg鲜质量(0.001 g精度天平), 用锡箔纸包裹并系上棉线, 置于液氮罐中快速冷冻保存, 随后带回实验室进行后续测定和分析(Robinson et al ., 2005 )。
1.4 生理生化指标的测定
1.4.1 植株含水量
将取回的齿肋赤藓植株样品, 首先称取鲜质量, 然后105 ℃烘干15 min, 调至80 ℃烘干至恒质量。用电子天平称量(0.001 g精度天平), 根据Munné- Bosch等(2003)方法计算其相对含水量。
1.4.2 渗透调节物含量的测定
本研究所测定的渗透调节物主要包括可溶性糖、可溶性蛋白和脯氨酸3种, 分别用蒽酮法、考马斯亮蓝法和酸性茚三酮法进行测定, 其吸光度值由Lamda35紫外分光光度计(PE, Waltham, USA)测定Gonzalez & Pignata; 1994 ; Monreal et al ., 2007 ; Lassouane et al ., 2013 )。
1.4.3 丙二醛含量和抗氧化酶活性的测定
丙二醛(MDA)含量和超氧化物歧化酶(SOD)、过氧化物酶(POD)及过氧化氢酶(CAT) 3种抗氧化酶活性的测定分别采用TBA法、羟胺法、愈创木酚法和可见光法进行测定Choudhury & Panda, 2005 ; Sun et al ., 2009 ; Wu et al ., 2012 )。
1.5 土壤理化性质的测定
在每个样方内, 用高度为5 cm的环刀取0-5 cm土层土壤带回实验室风干磨碎过2 mm筛。用重铬酸钾容量法-外加热法测量土壤有机碳, 高氯酸-硫酸消化法测定全氮、酸溶-钼锑抗比色法测定全磷、酸溶-火焰光度法测定全钾。pH和电导率用pHS-3C精密pH计和DDS-307电导率仪(上海雷磁仪器厂)测定(水土稀释质量比5:1)。采用残渣烘干-质量法测定土壤总盐含量尹本丰和张元明, 2014 )。
1.6 数据处理
使用SPSS 19.0软件, 对不同微生境下齿肋赤藓的各项指标进行描述统计分析。用一般线性模型(GLM)对不同冻融期和微生境是否对苔藓各项指标存在交互作用进行分析。同时对不同处理间的数据进行单因素方差分析(one-way ANOVA)和多重比较(LSD ), 不同时期进行Repeated measure ANO- VA分析。用Pearson相关系数检验不同时期和微生境下齿肋赤藓的含水量、温度与游离脯氨酸、可溶性糖、可溶性蛋白、MDA等含量和POD、SOD及CAT活性之间的相关性, 并用Origin 8.0软件对数据做图。
2 结果和分析
2.1 不同微生境环境因子差异分析
结果表明, 不同微生境下土壤的养分存在显著差异, 活灌丛与死灌丛的土壤有机碳、全氮、全钾含量显著高于裸露地土壤, 死灌丛土壤的全磷含量显著高于裸露地(表1 )。活灌丛下土壤的pH较其他两个生境更趋向于碱性, 但其电导率之间无显著差异。同时, 在积雪冻结期活灌丛下地表温度较死灌丛和裸露地平均高1.7-2.3 ℃。而在积雪融化期和春季中旬干旱无雨期灌丛下平均日均温较其他两生境低1.9-5.0 ℃。
2.2 苔藓植株含水量
由图3 可知, 积雪融水和灌丛的存在均能在一定程度上增加齿肋赤藓的植株含水量。3月积雪的融化使得齿肋赤藓植株含水量显著高于冬季冻结期的2月和春季干旱无雨期的4月。同时, 不同时期活灌丛下齿肋赤藓植株含水量均显著高于死灌丛和裸露地, 死灌丛和裸露地之间植株含水量差异不显著。
图3 冻融期不同微生境下齿肋赤藓植株含水量的比较(平均值±标准偏差)。不同小写字母和大写字母分别表示不同微生境下和不同冻融期齿肋赤藓植株含水量差异显著(p < 0.05)。
Fig. 3 Comparison of water content in Syntrichia caninervis shoots from different microhabitats and freezing-thawing periods (mean ± SD ). Different lowercase and capital letters denote statistically significant differences in water content (p < 0.05) between microhabitats and periods, respectively.
2.3 渗透调节物质
2.3.1 脯氨酸
不同时期和不同微生境对齿肋赤藓脯氨酸含量均有显著影响, 且二者存在显著的交互作用(表2 )。春季中旬干燥无雨期的齿肋赤藓脯氨酸含量显著高于积雪融化期和冬季低温冻结期。在春季中旬干燥环境下, 齿肋赤藓脯氨酸含量表现为裸露地>死灌丛>活灌丛下, 而在积雪期和积雪融化期, 各微生境之间差异不显著(图4A )。
2.3.2 可溶性糖
由表2 和图4B 可以看出, 积雪融化期的可溶性糖含量显著低于其他两个时期, 春季中旬干旱无雨期拥有最高的可溶性糖含量。同时, 不同微生境下齿肋赤藓可溶性糖含量在不同时期表现出不同的差异。在积雪期和积雪融化期, 灌丛下齿肋赤藓可溶性糖含量显著高于裸露地, 而在春季中旬干旱无雨期表现出相反的结果。
2.3.3 可溶性蛋白
积雪融化期的3种不同微生境下齿肋赤藓可溶性蛋白含量均显著高于冬季低温冻结期和春季中旬干旱无雨期(图4C )。在3个不同时期、不同微生境下齿肋赤藓的可溶性蛋白含量表现出相似的变化趋势, 均为活灌丛下>死灌丛>裸露地。
图4 冻融期不同微生境下齿肋赤藓脯氨酸、可溶性糖和可溶性蛋白含量的比较(平均值±标准偏差)。不同小写字母和大写字母分别表示不同微生境下和不同冻融期齿肋赤藓渗透调节物质含量差异显著(p < 0.05)。
Fig. 4 Comparison of the contents of compatible solutes (osmoprotectants) in Syntrichia caninervis shoots from different microhabitats and freezing-thawing periods (mean ± SD ). Different lowercase and capital letters denote statistically significant differences in solutes (p < 0.05) between microhabitats and periods, respectively.
2.4 丙二醛
由表2 和图5 可以看出, 积雪覆盖下的齿肋赤藓植株MDA含量显著高于春季中旬干旱无雨期, 积雪融化期植株MDA含量最低。微生境对齿肋赤藓MDA含量的影响因不同时期而存在差异。在积雪融化期活灌丛下齿肋赤藓植株MDA含量显著高于裸露地, 而在春季中旬干旱无雨期活灌丛下齿肋赤藓植株MDA含量却显著低于裸露地和死灌丛。
图5 冻融期不同微生境下齿肋赤藓丙二醛(MDA)含量的比较(平均值±标准偏差)。不同小写字母和大写字母分别表示不同微生境下和不同冻融期齿肋赤藓MDA含量差异显著(p < 0.05)。
Fig. 5 Comparison of the content of malonyldialdehyde (MDA) in Syntrichia caninervis shoots from different microhabitats and freezing-thawing periods (mean ± SD ). Different lowercase and capital letters denote statistically significant differences in MDA (p < 0.05) between microhabitats and periods, respectively.
2.5 抗氧化酶
不同微生境和积雪期及二者的交互作用均对齿肋赤藓体内3种抗氧化酶活性有显著影响(表2 )。与MDA含量对应, 在积雪融化期3种抗氧化酶活性均显著低于其他两个时期。由图6 可以看出, 不同微生境下齿肋赤藓植株的3种抗氧化酶在不同时期均表现出相似的趋势。除积雪融化期的SOD活性在3种微生境间差异不显著外, 其余各时期3种抗氧化酶活性均表现为灌丛下显著低于裸露地。
图6 冻融期不同微生境下齿肋赤藓过氧化物酶(POD)、超氧化物歧化酶(SOD)和过氧化氢酶(CAT)活性的比较(平均值±标准偏差)。不同小写字母和大写字母分别表示不同微生境下和不同冻融期齿肋赤藓抗氧化酶活性差异显著(p < 0.05)。
Fig. 6 Comparisons of the activities of peroxidase (POD), superoxide dismutase (SOD), catalase (CAT) in Syntrichia caninervis shoots from different microhabitats and freezing-thawing periods (mean ± SD ). Different lowercase and capital letters denote statistically significant differences in enzyme activities (p < 0.05) between microhabitats and periods, respectively.
2.6 生理生化指标与含水量和温度的相关性分析
不同微生境下齿肋赤藓生理生化特征在不同时期与环境因子表现出不同的相关性。在冬季低温冻结期可溶性糖和可溶性蛋白含量与温度和含水量表现出极显著的正相关, 而3种抗氧化酶(SOD、POD、CAT)活性却表现出与温度和含水量极显著的负相关。在积雪融化期, 可溶性糖、可溶性蛋白和MDA含量与温度表现出显著的负相关, POD和CAT与温度表现为正相关。可溶性蛋白含量与含水量呈显著正相关, 而POD与含水量呈现极显著负相关。在春季中旬干旱无雨期, 除可溶性蛋白含量与含水量呈极显著正相关外, 脯氨酸、可溶性蛋白和MDA含量及SOD、POD和CAT 3种抗氧化酶活性均与植株含水量呈极显著负相关; 各生理特征与温度之间的相关性与其与含水量之间的相关性相反。
3 讨论
在温带荒漠冬、春季节, 温度和水分能够间接或直接的影响植物的生理生化过程张静和张元明, 2014 )。研究发现, 与荒漠藓类的形态特征随周围环境的变化相比, 其生理特征变化更为明显, 反应更为迅速Reynolds & McLetchie, 2011 )。本实验发现: 不同时期3种微生境的温度和水分等环境因子均存在显著差异, 随着不同时期水热条件的变化, 齿肋赤藓的含水量、可溶性糖、游离脯氨酸、可溶性蛋白、MDA等含量以及CAT、POD、SOD等抗氧化酶活性均发生显著变化。
3.1 冻融期水热变化对齿肋赤藓渗透调节物含量和抗氧化酶活性的影响
水分是干旱半干旱荒漠生态系统植物生长的主要限制因素, 对于变水植物齿肋赤藓, 水分能够在一定程度上反映植物的活性程度Brown & Ernest, 2002 ; Huxman et al ., 2004 ; Benassi et al ., 2011 )。而在冬春季节的冻融期, 水分的变化常常是受温度变化驱使的。研究发现, 在冻融过程中温度和水分会发生剧烈变化, 在温度和水分胁迫下, 藓类植物能够通过失复水和调节体内渗透调节物质浓度来维持细胞渗透压(Nagao et al ., 2006 ; Lenne et al ., 2010 ), 同时通过抗氧化酶的合成来清除体内多余的活性氧和超氧物阴离子自由基Zhang & Kirkham, 1994 ; Meloni et al ., 2003 ), 进而减少其对植物组织的过度损伤, 稳定和维持细胞内大分子物质的结构与功能。此外, 在极地的研究也发现在极端低温状态下, 苔藓植物能够通过脱水来缓解低温对藓类植物的伤害(Lenne et al ., 2010 ), 这也可能是在冬季低温冻结期齿肋赤藓植株含水量相对较低的重要原因。
冬季的极端低温环境极易使得植物体内的水分凝结形成冰晶, 并对植物造成严重伤害。而可溶性糖的大量积累能够增加细胞液的浓度, 降低细胞内水势, 使细胞液的冰点下降从而增加植物的抗寒能力。不少研究发现随着低温处理时间的延长, 植物体内的可溶性糖含量显著增加(沈蕾等, 2011 ; 朱政等, 2011 )。这在本实验中也得到证实, 在冬季低温冻结期植物体内的可溶性糖含量显著高于融雪期, 这说明齿肋赤藓在极端低温环境下能够通过积累大量可溶性糖来保护自身(Bhyan et al ., 2012 )。同时研究发现, 在积雪融化后的干燥环境下, 藓类植物体内脯氨酸和可溶性糖含量均较其他两个时期显著增加, 这可能是由于地表土壤含水量减少使得藓类植物处于干旱胁迫状态引起的。这也与张静和张元明(2014)在模拟降水对齿肋赤藓生理生化特性时发现的随含水量的减少齿肋赤藓体内可溶性糖和与脯氨酸呈递增趋势的结果相一致。此外, 徐杰等(2005)在不同区域真藓(Bryum argenteum )和土生对齿藓(Didymodon vinealis )的研究也表明, 植物体内脯氨酸与可溶性糖含量在降水量相对较少的草原化荒漠区显著高于降水量较多的典型草原。
可溶性蛋白也是植物重要的调节物质之一, 其含量在一定程度上能够反映植物的新陈代谢水平, 含量越高说明植物代谢越旺盛。不少研究发现极端的低温或干旱环境使得植物脱水而活性降低或失活, 新陈代谢降低。石勇等(2012a)的研究发现随着干旱胁迫的增加植物体内的可溶性蛋白含量逐渐减少, 本实验中也发现在积雪融化期齿肋赤藓可溶性蛋白含量显著高于其他两个时期, 这可能与积雪融化期地表土壤含水量极显著高于冬季冻结期和春季中旬的干旱无雨期有关, 积雪的融化使土壤含水量迅速增加, 植物能够利用短暂的积雪融水快速恢复其生理活性。但沙伟等(2010)在对东亚砂藓(Racomitrium japonicum )的研究中发现, 随基质含水量的减少其体内可溶性蛋白含量呈增加趋势。
水分的严重匮乏与极端的温度均会造成细胞膜脂的过氧化程度加剧, 使得细胞产生大量的活性氧和超氧物阴离子自由基, 进而对细胞质膜的组成和完整性造成严重伤害, 主要表现在植物体内的MDA含量迅速增加(范苏鲁等, 2011 )。研究发现随着积雪融水的逐渐消失, 干旱胁迫加剧使得春季中旬干旱无雨期植物体内MDA含量显著高于融雪期。同时春季中旬干旱无雨期的相关分析也表明MDA含量与植株含水量呈显著负相关关系(表3 )。石勇等(2012b)在研究土生对齿藓(Didymodon vinealis )时也发现随着干旱的逐渐增加, 植物体内的MDA含量显著增加, 但在真藓中表现出相反的结果。也有研究发现水分胁迫对尖叶提灯藓(Mnium cuspidatum )、青藓(Brachythecium albicans )、石地藓(Reboulia hemisphaerica )的MDA含量影响较小(项俊等, 2010 )。这可能说明不同藓类植物在应对水分胁迫时具有不同的响应机制。此外, 在冬季低温冻结期植株的MDA含量显著高于积雪融化期和春季中旬干旱无雨期, 尽管在冬季低温冻结期的植株含水量高于春季中旬干旱无雨期。在水分丰富的积雪融化期, 灌丛下齿肋赤藓的MDA含量却显著高于裸露地, 这也进一步说明冬季的低温对植物造成了严重损伤, 这与沈蕾等(2011)对多蒴灰藓(Hypnum fertile )的低温处理结果相似。
为了缓解干旱和极端温度胁迫对细胞的过度损伤, 植物细胞能够通过合成大量的CAT、POD和SOD等抗氧化酶来清除多余的活性氧和超氧物阴离子自由基(Zhang & Kirkham, 1994 ; Meloni et al ., 2003 )。研究发现: 在水分充足的积雪融化期, 3种抗氧化酶活性均显著低于冬季积雪冻结期和春季中旬干旱无雨期。春季中旬干旱无雨期的相关性分析也表明3种抗氧化酶活性与植株含水量极显著负相关, 在水分相对较好的灌丛下齿肋赤藓3种抗氧化酶活性显著低于其他两个水分相对较差的区域, 这与张静等在研究模拟降水对齿肋赤藓生理生化特性的结果一致, 在干燥状态下齿肋赤藓能够通过增加CAT、POD和SOD 3种抗氧化酶的活性来抵御干旱环境对植物的伤害(张静和张元明, 2014 )。与干旱胁迫一样, 在应对极端温度时植物也会合成大量的抗氧化酶来清除多余的氧自由基。已有的研究发现在高温胁迫下植物体内SOD和POD等抗氧化酶活性显著增加(沈蕾等, 2011 )。本研究发现尽管冬季冻结期的含水量显著高于春季中旬干旱无雨期, 但极端的低温环境使得其表现为更高的CAT、POD和SOD等抗氧化酶活性, 这说明藓类植物在应对极端低温时也会通过合成大量的抗氧化酶来抵御极端低温对植物体的伤害。
3.2 微生境对齿肋赤藓渗透调节物含量和抗氧化酶活性的影响
植物的生存环境可以分为大环境和小环境, 大环境影响生物的生存和分布, 而小环境直接影响到生物的生活, 所以微生境间的差异也势必对荒漠藓类植物的生理生化特征产生一定程度的影响。研究发现在积雪融化期灌丛下的可溶性糖含量显著高于其他两个生境, 这可能是由于积雪融化期灌丛形成的“冷岛效应”使活灌丛温度明显低于裸露地和死灌丛(Nagao et al ., 2006 ; 张亚峰等, 2013 )。同时, 相关分析也表明, 在水分为非胁迫条件下可溶性糖含量与温度呈极显著负相关而与植株含水量无关。随着积雪融水的逐渐消失, 在春季中旬的干旱无雨期, 含水量相对较多的灌丛下齿肋赤藓的可溶性糖含量和脯氨酸含量显著低于死灌丛和裸露地, 且在3种不同微生境下与含水量存在显著负相关, 这与之前在对初冬降雪前后齿肋赤藓生理特性响应的研究结果一致尹本丰和张元明, 2014 )。
本研究发现齿肋赤藓植株体内可溶性蛋白含量与植物含水量呈显著正相关关系(表3 ), 随含水量的增加, 植物可溶性蛋白含量越多, 植物活性越高。灌丛的遮阴减少了地表水分的快速蒸发, 加之灌丛对积雪的阻挡和堆积作用, 使得灌丛下含水量显著高于其他生境(范连连等, 2012 )。所以生活于灌丛下的齿肋赤藓较其他生境的苔藓拥有更多水分和更长时间的水分持续时间。这也可能是灌丛下齿肋赤藓可溶性蛋白含量显著高于其他微生境的重要原因。
尽管冻融期的不同阶段是齿肋赤藓生理特征变化的主要驱动力, 但不同微生境间的差异也会直接影响到齿肋赤藓的生存状况。研究发现在含水量丰富的积雪融化期, 灌丛下齿肋赤藓的MDA含量显著高于裸露地, 而在春季中旬的干旱无雨期表现出相反的结果。这说明灌丛的存在在不同时期对藓类植物存在不同的影响, 尽管在融雪期灌丛的存在对水分起到了一定的富集和延缓蒸发效应, 但与裸露地相比, 其形成的低温区却在一定程度上加重了齿肋赤藓的冻害。相反, 在春季中旬的干旱无雨期灌丛所形成的低温效应引起的水分蒸发减缓, 对齿肋赤藓应对水分的胁迫起到了积极作用。总之, 从整个冻融期来看, 灌丛的存在对齿肋赤藓的生长具有一定的促进作用。
有意义的是, 纵观整个冻融期, 3种微生境下齿肋赤藓的渗透调节物含量和MDA含量, 在冻融期的不同阶段均表现出不同趋势, 但其3种抗氧化酶活性却一致表现为灌丛下显著低于裸露地。这可能是由于裸露地的齿肋赤藓表现出更高的生理耐受性Reynolds & McLetchie, 2011 ), 这也与Stark等(2010)的研究认为的活灌丛下齿肋赤藓较裸露地齿肋赤藓抗旱性稍差相一致。但在不同生境、不同水热组合条件下, 齿肋赤藓与抗逆相关的基因表达是否存在差异还需进一步研究。
感谢中国科学院新疆生态与地理研究所吴楠博士、吴林博士在论文写作修改过程中的指导和帮助。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Benassi M Stark LR Brinda JC McLetchie DN Bonine M Mishler BD 2011 ). Plant size, sex expression and sexual reproduction along an elevation gradient in a desert moss.
The Bryologist , 114 , 277 -288 .
[本文引用: 3]
[2]
Bhyan SB Minami A Kaneko Y Suzuki S Arakawa K Sakata Y Takezawa D 2012 ). Cold acclimation in the moss Physcomitrella patens involves abscisic acid-dependent signaling.
Journal of Plant Physiology , 169 , 137 -145 .
[本文引用: 1]
[3]
Bjerke JW 2011 ). Winter climate change: Ice encapsulation at mild subfreezing temperatures kills freeze-tolerant lichens.
Environmental and Experimental Botany , 72 , 404 -408 .
[本文引用: 2]
[4]
Bjerke JW Bokhorst S Zielke M Callaghan TV Bowles FW Phoenix GK 2011 ). Contrasting sensitivity to extreme winter warming events of dominant sub-Arctic heathland bryophyte and lichen species.
Journal of Ecology , 99 , 1481 -1488 .
[5]
Brown JH Ernest SKM 2002 ). Rain and rodents: Complex dynamics of desert consumers although water is the pri- mary limiting resource in desert ecosystems, the relationship between rodent population dynamics and precipitation is complex and nonlinear.
BioScience , 52 , 979 -987 .
[本文引用: 1]
[6]
Chen WJ Zhang N Hang LL Wang Y Ji MC 2013 ). Influence of shading during the processes of drought stress and re-watering on the physiological and biochemical characteristics of Haplocladium microphyllum.
Chinese Journal of Applied Ecology , 24 , 57 -62 (in Chinese with English abstract).
[本文引用: 1]
[陈文佳 , 张楠 , 杭璐璐 , 王媛 , 季梦成 (2013 ). 干旱胁迫与复水过程中遮光对细叶小羽藓的生理生化影响
. 应用生态学报 , 24 , 57 -62 .]
[本文引用: 1]
[7]
Choudhury S Panda SK 2005 ). Toxic effects, oxidative stress and ultrastructural changes in moss Taxithelium nepalense (Schwaegr.) Broth. under chromium and lead phytotoxicity.
Water, Air, and Soil Pollution , 167 , 73 -90 .
[本文引用: 1]
[8]
Crowe JH Carpenter JF Crowe LM 1998 ). The role of vitrification in anhydrobiosis.
Annual Review of Physiology , 60 , 73 -103 .
[本文引用: 1]
[9]
Fan LL Ma J Wu LF Xu GQ Li Y Tang LS 2012 ). Response of the herbaceous layer to snow variability at the south margin of the Gurbantonggut desert of China.
Chinese Journal of Plant Ecology , 36 , 126 -135 (in Chinese with English abstract).
[本文引用: 1]
[范连连 , 马健 , 吴林峰 , 徐贵青 , 李彦 , 唐立松 (2012 ). 古尔班通古特沙漠南缘草本层对积雪变化的响应
. 植物生态学报 , 36 , 126 -135 .]
[本文引用: 1]
[10]
Fan SL Yuan ZH Feng LJ Wang XH Ding XM Zhen HL 2011 ). Effects of drought stress on physiological and biochemical parameters of Dahlia pinnata.
Chinese Journal of Applied Ecology , 22 , 651 -657 (in Chinese with English abstract).
[本文引用: 1]
[范苏鲁 , 苑兆和 , 冯立娟 , 王晓慧 , 丁雪梅 , 甄红丽 (2011 ). 干旱胁迫对大丽花生理生化指标的影响
. 应用生态学报 , 22 , 651 -657 .]
[本文引用: 1]
[11]
Gonzalez CM Pignata ML 1994 ). The influence of air pollution on soluble proteins, chlorophyll degradation, MDA, sulphur and heavy metals in a transplanted lichen.
Chemistry and Ecology , 9 , 105 -113 .
[本文引用: 1]
[12]
Harper KT Belnap J 2001 ). The influence of biological soil crusts on mineral uptake by associated vascular plants.
Journal of Arid Environments , 47 , 347 -357 .
[本文引用: 1]
[13]
Herrnstadt I Kidron GJ 2005 ). Reproductive strategies of Bryum dunense in three microhabitats in the Negev Desert.
The Bryologist , 108 , 101 -109 .
[本文引用: 3]
[14]
Hui R Li XR Jia RL Zhao X Liu YM Chen CY 2012 ). Effects of enhanced UV-B radiation on physiological characteristics of Bryum argenteum.
Chinese Journal of Ecology , 31 , 38 -43 (in Chinese with English abstract).
[本文引用: 1]
[回嵘 , 李新荣 , 贾荣亮 , 赵昕 , 刘艳梅 , 陈翠云 (2012 ). 增强 UV-B 辐射对真藓结皮生理特性的影响
. 生态学杂志 , 31 , 38 -43 .]
[本文引用: 1]
[15]
Huxman TE Smith MD Fay PA Knapp AK Shaw MR Loik ME Smith SD Tissue DT Zak JC Weltzin JF 2004 ). Convergence across biomes to a common rain-use efficiency.
Nature , 429 , 651 -654 .
[本文引用: 1]
[16]
Kappen L Valladares F 2007 ). Opportunistic growth and des- iccation tolerance: The ecological success of poikilohy- drous autotrophs.
Functional Plant Ecology , 2 , 7 -67 .
[本文引用: 1]
[17]
Kidron GJ Vonshak A Abeliovich A 2009 ). Microbiotic crusts as biomarkers for surface stability and wetness duration in the Negev Desert.
Earth Surface Processes and Landforms , 34 , 1594 -1604 .
[本文引用: 1]
[18]
Lange OL 2003 ). Photosynthetic productivity of the epilithic lichen Lecanora muralis: Long-term field monitoring of CO2 exchange and its physiological interpretation: III. Diel, seasonal, and annual carbon budgets.
Flora-Mor- phology, Distribution, Functional Ecology of Plants , 198 , 277 -292 .
[本文引用: 1]
[19]
Lassouane N Aïd F Lutts S 2013 ). Water stress impact on young seedling growth of Acacia arabica.
Acta Physiologiae Plantarum , 35 , 2157 -2169 .
[本文引用: 1]
[20]
Lenne T Bryant G Hocart CH Huang CX Ball MC 2010 ). Freeze avoidance: A dehydrating moss gathers no ice.
Plant, Cell & Environment , 33 , 1731 -1741 .
[本文引用: 2]
[21]
Mallón R Barros P Luzardo A González ML 2007 ). Encapsulation of moss buds: An efficient method for the in vitro conservation and regeneration of the endangered moss Splachnum ampullaceum.
Plant Cell, Tissue and Organ Culture , 88 , 41 -49 .
[本文引用: 1]
[22]
Meloni DA Oliva MA Martinez CA Cambraia J 2003 ). Pho- tosynthesis and activity of superoxide dismutase, peroxidase and glutathione reductase in cotton under salt stress.
Environmental and Experimental Botany , 49 , 69 -76 .
[本文引用: 2]
[23]
Monreal J Jiménez E Remesal E Morillo-Velarde R García-Mauriño S Echevarría C 2007 ). Proline content of sugar beet storage roots: Response to water deficit and nitrogen fertilization at field conditions.
Environmental and Experimental Botany , 60 , 257 -267 .
[本文引用: 1]
[24]
Munné-Bosch S Jubany-Marí T Alegre L 2003 ). Enhanced photo-and antioxidative protection, and hydrogen peroxide accumulation in drought-stressed Cistus clusii and Cistus albidus plants.
Tree Physiology , 23 , 1 -12 .
[25]
Nagao M Oku K Minami A Mizuno K Sakurai M Arakawa K Fujikawa S Takezawa D 2006 ). Accumulation of theanderose in association with development of freezing tolerance in the moss Physcomitrella patens.
Phytochemistry , 67 , 702 -709 .
[本文引用: 2]
[26]
Oliver MJ Wood AJ O’Mahony P 1997 ). How some plants recover from vegetative desiccation: A repair based strategy.
Acta Physiologiae Plantarum , 19 , 419 -425 .
[本文引用: 1]
[27]
Pintado A Sancho LG Green T Blanquer JM Lázaro R 2005 ). Functional ecology of the biological soil crust in semiarid SE Spain: Sun and shade populations of Diploschistes diacapsis (Ach.) Lumbsch.
The Lichenologist , 37 , 425 -432 .
[本文引用: 2]
[28]
Reynolds LA McLetchie DN 2011 ). Short distances between extreme microhabitats do not result in ecotypes in Syntrichia caninervis.
Journal of Bryology , 33 , 148 -153 .
[本文引用: 4]
[29]
Robinson SA Turnbull JD Lovelock CE 2005 ). Impact of changes in natural ultraviolet radiation on pigment composition, physiological and morphological characteristics of the Antarctic moss, Grimmia antarctici.
Global Change Biology , 11 , 476 -489 .
[本文引用: 1]
[30]
Sgherri CLM Loggini B Puliga S Navari-Izzo F 1994 ). Antioxidant system in Sporobolus stapfianus: Changes in response to desiccation and rehydration.
Phytochemistry , 35 , 561 -565 .
[本文引用: 1]
[31]
Sha W Wang H Shi S 2010 ). Effects of rewatering on the physio-biochemical indexes of Racomitrium japonicum in long-time drought.
Journal of Wuhan Botanical Research , 28 , 246 -249 (in Chinese with English abstract).
[沙伟 , 王欢 , 师帅 (2010 ). 旱后复水对东亚砂藓生理生化指标的影响
. 武汉植物学研究 , 28 , 246 -249 .]
[32]
Shen L Guo SL Yang W Cao T Glime JM 2011 ). Physiological responses of Hypnum fertile Sendtn. (Musci: Hypnaceae) to short-term extreme temperature stress.
Bulletin of Botanical Research , 31 , 40 -48 (in English with Chinese abstract).
[本文引用: 3]
[沈蕾 , 郭水良 , 杨武 , 曹同 , Glime JM 2011 ). 多蒴灰藓(苔藓植物门: 藓纲)对短期极端温度的生理响应
. 植物研究 , 31 , 40 -48 .]
[本文引用: 3]
[33]
Shi Y Yang XJ Zhao X Li XR 2012 a). Responses of signal transduction substances of Didymodon vinealis and Bryum argenteum in biological soil crust to gradual drought stress.
Chinese Journal of Ecology , 31 , 1136 -1142 (in Chinese with English abstract).
[石勇 , 杨小菊 , 赵昕 , 李新荣 (2012a ). 逐渐干旱胁迫下生物土壤结皮中土生对齿藓和真藓信号转导物质的响应
. 生态学杂志 , 31 , 1136 -1142 .]
[34]
Shi Y Zhao X Jia RL Li XR 2012 b). The reactive oxygen species scavenging mechanism of Bryum argenteum and Didymodon vinealis in biological soil crusts under gradual drought stress.
Journal of Desert Research , 32 , 683 -690 (in Chinese with English abstract).
[石勇 , 赵昕 , 贾荣亮 , 李新荣 (2012b ). 逐渐干旱胁迫下生物土壤结皮中真藓和土生对齿藓的活性氧清除机制
. 中国沙漠 , 32 , 683 -690 .]
[35]
Stark LR McLetchie DN Eppley SM 2010 ). Sex ratios and the shy male hypothesis in the moss Bryum argenteum (Bryaceae).
The Bryologist , 113 , 788 -797 .
[36]
Su YG Wu L Zhou ZB Liu YB Zhang YM 2013 ). Carbon flux in deserts depends on soil cover type: A case study in the Gurbantunggut desert, North China.
Soil Biology and Biochemistry , 58 , 332 -340 .
[本文引用: 2]
[37]
Sun SQ He M Cao T Zhang YC Han W 2009 ). Response mechanisms of antioxidants in bryophyte (Hypnum plumaeforme) under the stress of single or combined Pb and/or Ni.
Environmental Monitoring and Assessment , 149 , 291 -302 .
[本文引用: 1]
[38]
Tao Y Zhang YM 2012 ). Effects of leaf hair points on dew deposition and rainfall evaporation rates in moss crusts dominated by Syntrichia caninervis, Gurbantunggut desert, northwestern China.
Acta Ecologica Sinica , 32 , 7 -16 (in Chinese with English abstract).
[本文引用: 1]
[陶冶 , 张元明 (2012 ). 叶片毛尖对齿肋赤藓结皮凝结水形成及蒸发的影响
. 生态学报 , 32 , 7 -16 .]
[本文引用: 1]
[39]
Wu HL Wu XL Li ZH Duan LS Zhang MC 2012 ). Physiological evaluation of drought stress tolerance and recovery in cauliflower (Brassica oleracea L.) seedlings treated with methyl jasmonate and coronatine.
Journal of Plant Growth Regulation , 31 , 113 -123 .
[本文引用: 1]
[40]
Wu N Wei ML Zhang YM 2009 ). Membrane permeability of Syntrichia caninervis in response to dehydration and rehydration in biological soil crust.
Progress in Nature Science , 19 , 942 -951 (in Chinese).
[本文引用: 2]
[吴楠 , 魏美丽 , 张元明 (2009 ). 生物土壤结皮中刺叶赤藓质膜透性对脱水, 复水过程的响应
. 自然科学进展 , 19 , 942 -951 .]
[本文引用: 2]
[41]
Wu N Zhang YM Downing A Aanderud ZT Tao Y Williams S 2014 ). Rapid adjustment of leaf angle explains how the desert moss, Syntrichia caninervis, copes with multiple resource limitations during rehydration.
Functional Plant Biology , 41 , 168 -177 .
[本文引用: 1]
[42]
Xiang J Zhao F Fang YP Chen J 2010 ). Effects of calcium and water stress on physiological and biochemical indexes of bryophytes.
Environmental Science & Technology , 33 , 70 -74 (in Chinese with English abstract).
[本文引用: 1]
[项俊 , 赵芳 , 方元平 , 陈娟 (2010 ). 水分和钙胁迫对苔藓植物生理生化指标的影响
. 环境科学与技术 , 33 , 70 -74 .]
[本文引用: 1]
[43]
Xu J Bai XL Tian GQ Yao YP Gao TY 2005 ). Study on moss, the content of amino acid, the feature of nutritive elements and its resistance to draught in the biotic crusts in arid and semi-arid regions.
Acta Ecologica Sinica , 25 , 1247 -1255 (in Chinese with English abstract).
[本文引用: 1]
[徐杰 , 白学良 , 田桂泉 , 姚一萍 , 高天云 (2005 ). 干旱半干旱地区生物结皮层藓类植物氨基酸和营养物质组成特征及适应性分析
. 生态学报 , 25 , 1247 -1255 .]
[本文引用: 1]
[44]
Xu SJ 2007 ). Reproduction Characteristics of the Typical Desert Moss and Mechanisms of Physiological and Biochemical Tolerance to Environmental Dehydration and Thermistress
. PhD dissertation, Shanghai Jiaotong University, Shanghai (in Chinese with English abstract) .
[本文引用: 1]
[许书军 (2007 ). 典型荒漠苔藓人工繁殖特征与抗御干热环境胁迫的生理生化机制研究
. 博士学位论文, 上海交通大学, 上海 .]
[本文引用: 1]
[45]
Yin BF Zhang YM 2014 ). Physiological and biochemical response of Syntrichia caninervis at a snowfall event in different desert habitats.
Chinese Journal of Plant Ecology , 38 , 978 -989 (in Chinese with English abstract).
[本文引用: 3]
[尹本丰 , 张元明 (2014 ). 荒漠区不同微生境下齿肋赤藓对一次降雪的生理生化响应
. 植物生态学报 , 38 , 978 -989 .]
[本文引用: 3]
[46]
Zhang J Zhang YM 2011 ). Effects of freezing and thawing on chlorophyll fluorescence of Syntrichia caninervis in biological soil crusts.
Journal of Desert Research , 31 , 1479 -1487 .
[本文引用: 1]
(in Chinese with English abstract) [张静 , 张元明 (2011 ). 冻融过程对生物结皮中齿肋赤藓叶绿素荧光特性的影响
. 中国沙漠 , 31 , 1479 -1487 .]
[本文引用: 1]
[47]
Zhang J Zhang YM 2014 ). Influence of simulated rainfall on the physiological characteristics of Syntrichia caninervis.
Journal of Desert Research , 34 , 433 -440 (in Chinese with English abstract).
[本文引用: 4]
[张静 , 张元明 (2014 ). 模拟降雨对齿肋赤藓(Syntrichia caninervis)生理特性的影响
. 中国沙漠 , 34 , 433 -440 .]
[本文引用: 4]
[48]
Zhang JX Kirkham MB 1994 ). Drought-stress-induced changes in activities of superoxide dismutase, catalase, and peroxidase in wheat species.
Plant and Cell Physiology , 35 , 785 -791 .
[本文引用: 2]
[49]
Zhang YF Wang XP Hu R Pan YX 2013 ). Effects of shrubs and precipitation on spatial-temporal variation of soil temperature at the microhabitats induced by desert shrubs.
Journal of Desert Research , 33 , 536 -542 (in Chinese with English abstract).
[本文引用: 1]
[张亚峰 , 王新平 , 虎瑞 , 潘颜霞 (2013 ). 荒漠灌丛微生境土壤温度的时空变异特征——灌丛与降水的影响
. 中国沙漠 , 33 , 536 -542 .]
[本文引用: 1]
[50]
Zhang YM 2005 ). Microstructure and development characters in early stage of biological soil crust of desert.
Chinese Science Bulletin , 50 42 -47 (in Chinese).
[本文引用: 1]
[张元明 (2005 ). 荒漠地表生物土壤结皮的微结构及其早期发育特征
. 科学通报 , 50 , 42 -47 .]
[本文引用: 1]
[51]
Zhang YM Chen J Wang L Wang XQ Gu ZH 2007 ). The spatial distribution patterns of biological soil crusts in the Gurbantunggut Desert, Northern Xinjiang, China.
Journal of Arid Environments , 68 , 599 -610 .
[本文引用: 2]
[52]
Zhang YM Pan HX Pan BR 2004 ). Distribution characteristics of biological soil crust on sand dune surface in Gurbantunggut Desert, Xinjiang.
Journal of Soil and Water Conservation , 18 (4 ), 61 -64 .
[本文引用: 1]
[张元明 , 潘惠霞 , 潘伯荣 (2004 ). 古尔班通古特沙漠不同地貌部位生物结皮的选择性分布
. 水土保持学报 , 18 (4 ), 61 -64 .]
[本文引用: 1]
[53]
Zheng YP Zhao JC Zhang BC Zhang YM 2009 ). Mor- phological and structural adaptation and characteristics of protonemal development of Syntrichia caninervis in the mosses crust layer.
Journal of Desert Research , 29 , 878 -884 (in Chinese with English abstract).
[本文引用: 1]
[郑云普 , 赵建成 , 张丙昌 , 张元明 (2009 ). 荒漠藓类结皮层中齿肋赤藓形态结构适应性及其原丝体发育特征
. 中国沙漠 , 29 , 878 -884 .]
[本文引用: 1]
[54]
Zhou HF Zhou BJ Dai Q 2010 ). Observational analysis of rime condensation on plants over the Gurbantunggut desert in China.
Advances in Water Science , 21 , 56 -62 (in Chinese with English abstract).
[本文引用: 1]
[周宏飞 , 周宝佳 , 代琼 (2010 ). 古尔班通古特沙漠植物雾凇凝结特征
. 水科学进展 , 21 , 56 -62 .]
[本文引用: 1]
[55]
Zhu Z Jiang JY Jiang CJ Li W 2011 ). Effects of low temperature stress on SOD activity, soluble protein content and soluble sugar content in Camellia sinensis leaves.
Journal of Anhui Agricultural University , 38 , 24 -26 (in Chinese with English abstract).
[本文引用: 1]
[朱政 , 蒋家月 , 江昌俊 , 李雯 (2011 ). 低温胁迫对茶树叶片SOD、可溶性蛋白和可溶性糖含量的影响
. 安徽农业大学学报 , 38 , 24 -26 .]
[本文引用: 1]
Plant size, sex expression and sexual reproduction along an elevation gradient in a desert moss.
3
2011
... 荒漠苔藓作为典型的抗胁迫变水植物已被广泛关注, 诸多研究表明藓类植物能够适应极端高低温、干燥、强光照等恶劣环境(张静和张元明, 2014 ).为适应极端环境, 藓类植物在种群分布、形态特征(郑云普等, 2009 ; 陶冶和张元明, 2012 )、繁殖方式(Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 )、光合活性(张静和张元明, 2011 )、渗透调节物质、抗氧化酶(回嵘等, 2012 ; 陈文佳等, 2013 )等方面进化出多样的抗胁迫特性.研究发现齿肋赤藓(Syntrichia caninervis )叶片表面具有蜡质层, 并在细胞表面富集疣状突起, 在一定程度上能够减弱热气流的影响(许书军, 2007 ).另外, 在高温干燥环境下藓类植物能够通过失水降低代谢活性而进入休眠状态, 并积累大量的脯氨酸和可溶性糖来提高细胞渗透压, 维持渗透压平衡(Oliver et al ., 1997 ; Crowe et al ., 1998 ; 徐杰等, 2005 ; 吴楠等, 2009 ).同时, 通过抗氧化酶的大量合成清除过多的氧自由基, 进而保护植物免受过度伤害(Sgherri et al ., 1994 ; Herrnstadt & Kidron, 2005 ; Pintado et al ., 2005 ; 沈蕾等, 2011 ).但对荒漠藓类植物如何通过生理生化调节并成功度过冬季的极端低温环境, 还有待于深入研究. ...
... 在荒漠干燥少雨环境中, 藓类植物在全年的大部分时间均处于失活休眠状态.然而, 有研究发现, 荒漠苔藓的生长和繁殖主要发生在寒冷和湿润的冬季月份Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 ).既使已经冷适应的苔藓, 在温度、光照和湿度达到阈值时仍能够被快速激活(Lange, 2003 ; Kappen & Valladares, 2007 ; Bjerke 2011 ; Su et al ., 2013 ).初冬和早春的积雪融化和低温能够激活藓类植物的光合活性, 延长生物结皮光合活性时间, 增加结皮土壤的碳截获量(Su et al ., 2013 ; 尹本丰和张元明, 2014 ).因此, 苔藓植物更可能在初冬或初春积雪融化时表现出良好的生长特性.但关于寒冷生态系统不同微生境苔藓的生态学特征还知之甚少(Bjerke et al ., 2011 ). ...
... 水分是干旱半干旱荒漠生态系统植物生长的主要限制因素, 对于变水植物齿肋赤藓, 水分能够在一定程度上反映植物的活性程度Brown & Ernest, 2002 ; Huxman et al ., 2004 ; Benassi et al ., 2011 ).而在冬春季节的冻融期, 水分的变化常常是受温度变化驱使的.研究发现, 在冻融过程中温度和水分会发生剧烈变化, 在温度和水分胁迫下, 藓类植物能够通过失复水和调节体内渗透调节物质浓度来维持细胞渗透压(Nagao et al ., 2006 ; Lenne et al ., 2010 ), 同时通过抗氧化酶的合成来清除体内多余的活性氧和超氧物阴离子自由基Zhang & Kirkham, 1994 ; Meloni et al ., 2003 ), 进而减少其对植物组织的过度损伤, 稳定和维持细胞内大分子物质的结构与功能.此外, 在极地的研究也发现在极端低温状态下, 苔藓植物能够通过脱水来缓解低温对藓类植物的伤害(Lenne et al ., 2010 ), 这也可能是在冬季低温冻结期齿肋赤藓植株含水量相对较低的重要原因. ...
Cold acclimation in the moss Physcomitrella patens involves abscisic acid-dependent signaling.
1
2012
... 冬季的极端低温环境极易使得植物体内的水分凝结形成冰晶, 并对植物造成严重伤害.而可溶性糖的大量积累能够增加细胞液的浓度, 降低细胞内水势, 使细胞液的冰点下降从而增加植物的抗寒能力.不少研究发现随着低温处理时间的延长, 植物体内的可溶性糖含量显著增加(沈蕾等, 2011 ; 朱政等, 2011 ).这在本实验中也得到证实, 在冬季低温冻结期植物体内的可溶性糖含量显著高于融雪期, 这说明齿肋赤藓在极端低温环境下能够通过积累大量可溶性糖来保护自身(Bhyan et al ., 2012 ).同时研究发现, 在积雪融化后的干燥环境下, 藓类植物体内脯氨酸和可溶性糖含量均较其他两个时期显著增加, 这可能是由于地表土壤含水量减少使得藓类植物处于干旱胁迫状态引起的.这也与张静和张元明(2014)在模拟降水对齿肋赤藓生理生化特性时发现的随含水量的减少齿肋赤藓体内可溶性糖和与脯氨酸呈递增趋势的结果相一致.此外, 徐杰等(2005)在不同区域真藓(Bryum argenteum )和土生对齿藓(Didymodon vinealis )的研究也表明, 植物体内脯氨酸与可溶性糖含量在降水量相对较少的草原化荒漠区显著高于降水量较多的典型草原. ...
Winter climate change: Ice encapsulation at mild subfreezing temperatures kills freeze-tolerant lichens.
2
2011
... 在荒漠干燥少雨环境中, 藓类植物在全年的大部分时间均处于失活休眠状态.然而, 有研究发现, 荒漠苔藓的生长和繁殖主要发生在寒冷和湿润的冬季月份Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 ).既使已经冷适应的苔藓, 在温度、光照和湿度达到阈值时仍能够被快速激活(Lange, 2003 ; Kappen & Valladares, 2007 ; Bjerke 2011 ; Su et al ., 2013 ).初冬和早春的积雪融化和低温能够激活藓类植物的光合活性, 延长生物结皮光合活性时间, 增加结皮土壤的碳截获量(Su et al ., 2013 ; 尹本丰和张元明, 2014 ).因此, 苔藓植物更可能在初冬或初春积雪融化时表现出良好的生长特性.但关于寒冷生态系统不同微生境苔藓的生态学特征还知之甚少(Bjerke et al ., 2011 ). ...
... ).因此, 苔藓植物更可能在初冬或初春积雪融化时表现出良好的生长特性.但关于寒冷生态系统不同微生境苔藓的生态学特征还知之甚少(Bjerke et al ., 2011 ). ...
Contrasting sensitivity to extreme winter warming events of dominant sub-Arctic heathland bryophyte and lichen species.
0
2011
Rain and rodents: Complex dynamics of desert consumers although water is the pri- mary limiting resource in desert ecosystems, the relationship between rodent population dynamics and precipitation is complex and nonlinear.
1
2002
... 水分是干旱半干旱荒漠生态系统植物生长的主要限制因素, 对于变水植物齿肋赤藓, 水分能够在一定程度上反映植物的活性程度Brown & Ernest, 2002 ; Huxman et al ., 2004 ; Benassi et al ., 2011 ).而在冬春季节的冻融期, 水分的变化常常是受温度变化驱使的.研究发现, 在冻融过程中温度和水分会发生剧烈变化, 在温度和水分胁迫下, 藓类植物能够通过失复水和调节体内渗透调节物质浓度来维持细胞渗透压(Nagao et al ., 2006 ; Lenne et al ., 2010 ), 同时通过抗氧化酶的合成来清除体内多余的活性氧和超氧物阴离子自由基Zhang & Kirkham, 1994 ; Meloni et al ., 2003 ), 进而减少其对植物组织的过度损伤, 稳定和维持细胞内大分子物质的结构与功能.此外, 在极地的研究也发现在极端低温状态下, 苔藓植物能够通过脱水来缓解低温对藓类植物的伤害(Lenne et al ., 2010 ), 这也可能是在冬季低温冻结期齿肋赤藓植株含水量相对较低的重要原因. ...
干旱胁迫与复水过程中遮光对细叶小羽藓的生理生化影响
1
2013
... 荒漠苔藓作为典型的抗胁迫变水植物已被广泛关注, 诸多研究表明藓类植物能够适应极端高低温、干燥、强光照等恶劣环境(张静和张元明, 2014 ).为适应极端环境, 藓类植物在种群分布、形态特征(郑云普等, 2009 ; 陶冶和张元明, 2012 )、繁殖方式(Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 )、光合活性(张静和张元明, 2011 )、渗透调节物质、抗氧化酶(回嵘等, 2012 ; 陈文佳等, 2013 )等方面进化出多样的抗胁迫特性.研究发现齿肋赤藓(Syntrichia caninervis )叶片表面具有蜡质层, 并在细胞表面富集疣状突起, 在一定程度上能够减弱热气流的影响(许书军, 2007 ).另外, 在高温干燥环境下藓类植物能够通过失水降低代谢活性而进入休眠状态, 并积累大量的脯氨酸和可溶性糖来提高细胞渗透压, 维持渗透压平衡(Oliver et al ., 1997 ; Crowe et al ., 1998 ; 徐杰等, 2005 ; 吴楠等, 2009 ).同时, 通过抗氧化酶的大量合成清除过多的氧自由基, 进而保护植物免受过度伤害(Sgherri et al ., 1994 ; Herrnstadt & Kidron, 2005 ; Pintado et al ., 2005 ; 沈蕾等, 2011 ).但对荒漠藓类植物如何通过生理生化调节并成功度过冬季的极端低温环境, 还有待于深入研究. ...
干旱胁迫与复水过程中遮光对细叶小羽藓的生理生化影响
1
2013
... 荒漠苔藓作为典型的抗胁迫变水植物已被广泛关注, 诸多研究表明藓类植物能够适应极端高低温、干燥、强光照等恶劣环境(张静和张元明, 2014 ).为适应极端环境, 藓类植物在种群分布、形态特征(郑云普等, 2009 ; 陶冶和张元明, 2012 )、繁殖方式(Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 )、光合活性(张静和张元明, 2011 )、渗透调节物质、抗氧化酶(回嵘等, 2012 ; 陈文佳等, 2013 )等方面进化出多样的抗胁迫特性.研究发现齿肋赤藓(Syntrichia caninervis )叶片表面具有蜡质层, 并在细胞表面富集疣状突起, 在一定程度上能够减弱热气流的影响(许书军, 2007 ).另外, 在高温干燥环境下藓类植物能够通过失水降低代谢活性而进入休眠状态, 并积累大量的脯氨酸和可溶性糖来提高细胞渗透压, 维持渗透压平衡(Oliver et al ., 1997 ; Crowe et al ., 1998 ; 徐杰等, 2005 ; 吴楠等, 2009 ).同时, 通过抗氧化酶的大量合成清除过多的氧自由基, 进而保护植物免受过度伤害(Sgherri et al ., 1994 ; Herrnstadt & Kidron, 2005 ; Pintado et al ., 2005 ; 沈蕾等, 2011 ).但对荒漠藓类植物如何通过生理生化调节并成功度过冬季的极端低温环境, 还有待于深入研究. ...
Toxic effects, oxidative stress and ultrastructural changes in moss Taxithelium nepalense (Schwaegr.) Broth. under chromium and lead phytotoxicity.
1
2005
... 丙二醛(MDA)含量和超氧化物歧化酶(SOD)、过氧化物酶(POD)及过氧化氢酶(CAT) 3种抗氧化酶活性的测定分别采用TBA法、羟胺法、愈创木酚法和可见光法进行测定Choudhury & Panda, 2005 ; Sun et al ., 2009 ; Wu et al ., 2012 ). ...
The role of vitrification in anhydrobiosis.
1
1998
... 荒漠苔藓作为典型的抗胁迫变水植物已被广泛关注, 诸多研究表明藓类植物能够适应极端高低温、干燥、强光照等恶劣环境(张静和张元明, 2014 ).为适应极端环境, 藓类植物在种群分布、形态特征(郑云普等, 2009 ; 陶冶和张元明, 2012 )、繁殖方式(Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 )、光合活性(张静和张元明, 2011 )、渗透调节物质、抗氧化酶(回嵘等, 2012 ; 陈文佳等, 2013 )等方面进化出多样的抗胁迫特性.研究发现齿肋赤藓(Syntrichia caninervis )叶片表面具有蜡质层, 并在细胞表面富集疣状突起, 在一定程度上能够减弱热气流的影响(许书军, 2007 ).另外, 在高温干燥环境下藓类植物能够通过失水降低代谢活性而进入休眠状态, 并积累大量的脯氨酸和可溶性糖来提高细胞渗透压, 维持渗透压平衡(Oliver et al ., 1997 ; Crowe et al ., 1998 ; 徐杰等, 2005 ; 吴楠等, 2009 ).同时, 通过抗氧化酶的大量合成清除过多的氧自由基, 进而保护植物免受过度伤害(Sgherri et al ., 1994 ; Herrnstadt & Kidron, 2005 ; Pintado et al ., 2005 ; 沈蕾等, 2011 ).但对荒漠藓类植物如何通过生理生化调节并成功度过冬季的极端低温环境, 还有待于深入研究. ...
古尔班通古特沙漠南缘草本层对积雪变化的响应
1
2012
... 本研究发现齿肋赤藓植株体内可溶性蛋白含量与植物含水量呈显著正相关关系(表3 ), 随含水量的增加, 植物可溶性蛋白含量越多, 植物活性越高.灌丛的遮阴减少了地表水分的快速蒸发, 加之灌丛对积雪的阻挡和堆积作用, 使得灌丛下含水量显著高于其他生境(范连连等, 2012 ).所以生活于灌丛下的齿肋赤藓较其他生境的苔藓拥有更多水分和更长时间的水分持续时间.这也可能是灌丛下齿肋赤藓可溶性蛋白含量显著高于其他微生境的重要原因. ...
古尔班通古特沙漠南缘草本层对积雪变化的响应
1
2012
... 本研究发现齿肋赤藓植株体内可溶性蛋白含量与植物含水量呈显著正相关关系(表3 ), 随含水量的增加, 植物可溶性蛋白含量越多, 植物活性越高.灌丛的遮阴减少了地表水分的快速蒸发, 加之灌丛对积雪的阻挡和堆积作用, 使得灌丛下含水量显著高于其他生境(范连连等, 2012 ).所以生活于灌丛下的齿肋赤藓较其他生境的苔藓拥有更多水分和更长时间的水分持续时间.这也可能是灌丛下齿肋赤藓可溶性蛋白含量显著高于其他微生境的重要原因. ...
干旱胁迫对大丽花生理生化指标的影响
1
2011
... 水分的严重匮乏与极端的温度均会造成细胞膜脂的过氧化程度加剧, 使得细胞产生大量的活性氧和超氧物阴离子自由基, 进而对细胞质膜的组成和完整性造成严重伤害, 主要表现在植物体内的MDA含量迅速增加(范苏鲁等, 2011 ).研究发现随着积雪融水的逐渐消失, 干旱胁迫加剧使得春季中旬干旱无雨期植物体内MDA含量显著高于融雪期.同时春季中旬干旱无雨期的相关分析也表明MDA含量与植株含水量呈显著负相关关系(表3 ).石勇等(2012b)在研究土生对齿藓(Didymodon vinealis )时也发现随着干旱的逐渐增加, 植物体内的MDA含量显著增加, 但在真藓中表现出相反的结果.也有研究发现水分胁迫对尖叶提灯藓(Mnium cuspidatum )、青藓(Brachythecium albicans )、石地藓(Reboulia hemisphaerica )的MDA含量影响较小(项俊等, 2010 ).这可能说明不同藓类植物在应对水分胁迫时具有不同的响应机制.此外, 在冬季低温冻结期植株的MDA含量显著高于积雪融化期和春季中旬干旱无雨期, 尽管在冬季低温冻结期的植株含水量高于春季中旬干旱无雨期.在水分丰富的积雪融化期, 灌丛下齿肋赤藓的MDA含量却显著高于裸露地, 这也进一步说明冬季的低温对植物造成了严重损伤, 这与沈蕾等(2011)对多蒴灰藓(Hypnum fertile )的低温处理结果相似. ...
干旱胁迫对大丽花生理生化指标的影响
1
2011
... 水分的严重匮乏与极端的温度均会造成细胞膜脂的过氧化程度加剧, 使得细胞产生大量的活性氧和超氧物阴离子自由基, 进而对细胞质膜的组成和完整性造成严重伤害, 主要表现在植物体内的MDA含量迅速增加(范苏鲁等, 2011 ).研究发现随着积雪融水的逐渐消失, 干旱胁迫加剧使得春季中旬干旱无雨期植物体内MDA含量显著高于融雪期.同时春季中旬干旱无雨期的相关分析也表明MDA含量与植株含水量呈显著负相关关系(表3 ).石勇等(2012b)在研究土生对齿藓(Didymodon vinealis )时也发现随着干旱的逐渐增加, 植物体内的MDA含量显著增加, 但在真藓中表现出相反的结果.也有研究发现水分胁迫对尖叶提灯藓(Mnium cuspidatum )、青藓(Brachythecium albicans )、石地藓(Reboulia hemisphaerica )的MDA含量影响较小(项俊等, 2010 ).这可能说明不同藓类植物在应对水分胁迫时具有不同的响应机制.此外, 在冬季低温冻结期植株的MDA含量显著高于积雪融化期和春季中旬干旱无雨期, 尽管在冬季低温冻结期的植株含水量高于春季中旬干旱无雨期.在水分丰富的积雪融化期, 灌丛下齿肋赤藓的MDA含量却显著高于裸露地, 这也进一步说明冬季的低温对植物造成了严重损伤, 这与沈蕾等(2011)对多蒴灰藓(Hypnum fertile )的低温处理结果相似. ...
The influence of air pollution on soluble proteins, chlorophyll degradation, MDA, sulphur and heavy metals in a transplanted lichen.
1
1994
... 本研究所测定的渗透调节物主要包括可溶性糖、可溶性蛋白和脯氨酸3种, 分别用蒽酮法、考马斯亮蓝法和酸性茚三酮法进行测定, 其吸光度值由Lamda35紫外分光光度计(PE, Waltham, USA)测定Gonzalez & Pignata; 1994 ; Monreal et al ., 2007 ; Lassouane et al ., 2013 ). ...
The influence of biological soil crusts on mineral uptake by associated vascular plants.
1
2001
... 苔藓植物是仅次于被子植物的第二大类高等植物类群, 在沼泽、森林、草地和荒漠区均起着重要的生态作用(Mallón et al ., 2007 ).作为环境演替的先锋物种, 耐旱苔藓植物在荒漠生态系统中分布广泛, 它们的存在能够在地表构筑一个粗糙的平面, 对荒漠生态系统的维持和稳定(Kidron et al ., 2009 )、土壤微环境的改善(Harper & Belnap, 2001 )具有重要 作用. ...
Reproductive strategies of Bryum dunense in three microhabitats in the Negev Desert.
3
2005
... 荒漠苔藓作为典型的抗胁迫变水植物已被广泛关注, 诸多研究表明藓类植物能够适应极端高低温、干燥、强光照等恶劣环境(张静和张元明, 2014 ).为适应极端环境, 藓类植物在种群分布、形态特征(郑云普等, 2009 ; 陶冶和张元明, 2012 )、繁殖方式(Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 )、光合活性(张静和张元明, 2011 )、渗透调节物质、抗氧化酶(回嵘等, 2012 ; 陈文佳等, 2013 )等方面进化出多样的抗胁迫特性.研究发现齿肋赤藓(Syntrichia caninervis )叶片表面具有蜡质层, 并在细胞表面富集疣状突起, 在一定程度上能够减弱热气流的影响(许书军, 2007 ).另外, 在高温干燥环境下藓类植物能够通过失水降低代谢活性而进入休眠状态, 并积累大量的脯氨酸和可溶性糖来提高细胞渗透压, 维持渗透压平衡(Oliver et al ., 1997 ; Crowe et al ., 1998 ; 徐杰等, 2005 ; 吴楠等, 2009 ).同时, 通过抗氧化酶的大量合成清除过多的氧自由基, 进而保护植物免受过度伤害(Sgherri et al ., 1994 ; Herrnstadt & Kidron, 2005 ; Pintado et al ., 2005 ; 沈蕾等, 2011 ).但对荒漠藓类植物如何通过生理生化调节并成功度过冬季的极端低温环境, 还有待于深入研究. ...
... ; Herrnstadt & Kidron, 2005 ; Pintado et al ., 2005 ; 沈蕾等, 2011 ).但对荒漠藓类植物如何通过生理生化调节并成功度过冬季的极端低温环境, 还有待于深入研究. ...
... 在荒漠干燥少雨环境中, 藓类植物在全年的大部分时间均处于失活休眠状态.然而, 有研究发现, 荒漠苔藓的生长和繁殖主要发生在寒冷和湿润的冬季月份Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 ).既使已经冷适应的苔藓, 在温度、光照和湿度达到阈值时仍能够被快速激活(Lange, 2003 ; Kappen & Valladares, 2007 ; Bjerke 2011 ; Su et al ., 2013 ).初冬和早春的积雪融化和低温能够激活藓类植物的光合活性, 延长生物结皮光合活性时间, 增加结皮土壤的碳截获量(Su et al ., 2013 ; 尹本丰和张元明, 2014 ).因此, 苔藓植物更可能在初冬或初春积雪融化时表现出良好的生长特性.但关于寒冷生态系统不同微生境苔藓的生态学特征还知之甚少(Bjerke et al ., 2011 ). ...
增强 UV-B 辐射对真藓结皮生理特性的影响
1
2012
... 荒漠苔藓作为典型的抗胁迫变水植物已被广泛关注, 诸多研究表明藓类植物能够适应极端高低温、干燥、强光照等恶劣环境(张静和张元明, 2014 ).为适应极端环境, 藓类植物在种群分布、形态特征(郑云普等, 2009 ; 陶冶和张元明, 2012 )、繁殖方式(Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 )、光合活性(张静和张元明, 2011 )、渗透调节物质、抗氧化酶(回嵘等, 2012 ; 陈文佳等, 2013 )等方面进化出多样的抗胁迫特性.研究发现齿肋赤藓(Syntrichia caninervis )叶片表面具有蜡质层, 并在细胞表面富集疣状突起, 在一定程度上能够减弱热气流的影响(许书军, 2007 ).另外, 在高温干燥环境下藓类植物能够通过失水降低代谢活性而进入休眠状态, 并积累大量的脯氨酸和可溶性糖来提高细胞渗透压, 维持渗透压平衡(Oliver et al ., 1997 ; Crowe et al ., 1998 ; 徐杰等, 2005 ; 吴楠等, 2009 ).同时, 通过抗氧化酶的大量合成清除过多的氧自由基, 进而保护植物免受过度伤害(Sgherri et al ., 1994 ; Herrnstadt & Kidron, 2005 ; Pintado et al ., 2005 ; 沈蕾等, 2011 ).但对荒漠藓类植物如何通过生理生化调节并成功度过冬季的极端低温环境, 还有待于深入研究. ...
增强 UV-B 辐射对真藓结皮生理特性的影响
1
2012
... 荒漠苔藓作为典型的抗胁迫变水植物已被广泛关注, 诸多研究表明藓类植物能够适应极端高低温、干燥、强光照等恶劣环境(张静和张元明, 2014 ).为适应极端环境, 藓类植物在种群分布、形态特征(郑云普等, 2009 ; 陶冶和张元明, 2012 )、繁殖方式(Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 )、光合活性(张静和张元明, 2011 )、渗透调节物质、抗氧化酶(回嵘等, 2012 ; 陈文佳等, 2013 )等方面进化出多样的抗胁迫特性.研究发现齿肋赤藓(Syntrichia caninervis )叶片表面具有蜡质层, 并在细胞表面富集疣状突起, 在一定程度上能够减弱热气流的影响(许书军, 2007 ).另外, 在高温干燥环境下藓类植物能够通过失水降低代谢活性而进入休眠状态, 并积累大量的脯氨酸和可溶性糖来提高细胞渗透压, 维持渗透压平衡(Oliver et al ., 1997 ; Crowe et al ., 1998 ; 徐杰等, 2005 ; 吴楠等, 2009 ).同时, 通过抗氧化酶的大量合成清除过多的氧自由基, 进而保护植物免受过度伤害(Sgherri et al ., 1994 ; Herrnstadt & Kidron, 2005 ; Pintado et al ., 2005 ; 沈蕾等, 2011 ).但对荒漠藓类植物如何通过生理生化调节并成功度过冬季的极端低温环境, 还有待于深入研究. ...
Convergence across biomes to a common rain-use efficiency.
1
2004
... 水分是干旱半干旱荒漠生态系统植物生长的主要限制因素, 对于变水植物齿肋赤藓, 水分能够在一定程度上反映植物的活性程度Brown & Ernest, 2002 ; Huxman et al ., 2004 ; Benassi et al ., 2011 ).而在冬春季节的冻融期, 水分的变化常常是受温度变化驱使的.研究发现, 在冻融过程中温度和水分会发生剧烈变化, 在温度和水分胁迫下, 藓类植物能够通过失复水和调节体内渗透调节物质浓度来维持细胞渗透压(Nagao et al ., 2006 ; Lenne et al ., 2010 ), 同时通过抗氧化酶的合成来清除体内多余的活性氧和超氧物阴离子自由基Zhang & Kirkham, 1994 ; Meloni et al ., 2003 ), 进而减少其对植物组织的过度损伤, 稳定和维持细胞内大分子物质的结构与功能.此外, 在极地的研究也发现在极端低温状态下, 苔藓植物能够通过脱水来缓解低温对藓类植物的伤害(Lenne et al ., 2010 ), 这也可能是在冬季低温冻结期齿肋赤藓植株含水量相对较低的重要原因. ...
Opportunistic growth and des- iccation tolerance: The ecological success of poikilohy- drous autotrophs.
1
2007
... 在荒漠干燥少雨环境中, 藓类植物在全年的大部分时间均处于失活休眠状态.然而, 有研究发现, 荒漠苔藓的生长和繁殖主要发生在寒冷和湿润的冬季月份Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 ).既使已经冷适应的苔藓, 在温度、光照和湿度达到阈值时仍能够被快速激活(Lange, 2003 ; Kappen & Valladares, 2007 ; Bjerke 2011 ; Su et al ., 2013 ).初冬和早春的积雪融化和低温能够激活藓类植物的光合活性, 延长生物结皮光合活性时间, 增加结皮土壤的碳截获量(Su et al ., 2013 ; 尹本丰和张元明, 2014 ).因此, 苔藓植物更可能在初冬或初春积雪融化时表现出良好的生长特性.但关于寒冷生态系统不同微生境苔藓的生态学特征还知之甚少(Bjerke et al ., 2011 ). ...
Microbiotic crusts as biomarkers for surface stability and wetness duration in the Negev Desert.
1
2009
... 苔藓植物是仅次于被子植物的第二大类高等植物类群, 在沼泽、森林、草地和荒漠区均起着重要的生态作用(Mallón et al ., 2007 ).作为环境演替的先锋物种, 耐旱苔藓植物在荒漠生态系统中分布广泛, 它们的存在能够在地表构筑一个粗糙的平面, 对荒漠生态系统的维持和稳定(Kidron et al ., 2009 )、土壤微环境的改善(Harper & Belnap, 2001 )具有重要 作用. ...
Photosynthetic productivity of the epilithic lichen Lecanora muralis: Long-term field monitoring of CO2 exchange and its physiological interpretation: III. Diel, seasonal, and annual carbon budgets.
1
2003
... 在荒漠干燥少雨环境中, 藓类植物在全年的大部分时间均处于失活休眠状态.然而, 有研究发现, 荒漠苔藓的生长和繁殖主要发生在寒冷和湿润的冬季月份Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 ).既使已经冷适应的苔藓, 在温度、光照和湿度达到阈值时仍能够被快速激活(Lange, 2003 ; Kappen & Valladares, 2007 ; Bjerke 2011 ; Su et al ., 2013 ).初冬和早春的积雪融化和低温能够激活藓类植物的光合活性, 延长生物结皮光合活性时间, 增加结皮土壤的碳截获量(Su et al ., 2013 ; 尹本丰和张元明, 2014 ).因此, 苔藓植物更可能在初冬或初春积雪融化时表现出良好的生长特性.但关于寒冷生态系统不同微生境苔藓的生态学特征还知之甚少(Bjerke et al ., 2011 ). ...
Water stress impact on young seedling growth of Acacia arabica.
1
2013
... 本研究所测定的渗透调节物主要包括可溶性糖、可溶性蛋白和脯氨酸3种, 分别用蒽酮法、考马斯亮蓝法和酸性茚三酮法进行测定, 其吸光度值由Lamda35紫外分光光度计(PE, Waltham, USA)测定Gonzalez & Pignata; 1994 ; Monreal et al ., 2007 ; Lassouane et al ., 2013 ). ...
Freeze avoidance: A dehydrating moss gathers no ice.
2
2010
... 水分是干旱半干旱荒漠生态系统植物生长的主要限制因素, 对于变水植物齿肋赤藓, 水分能够在一定程度上反映植物的活性程度Brown & Ernest, 2002 ; Huxman et al ., 2004 ; Benassi et al ., 2011 ).而在冬春季节的冻融期, 水分的变化常常是受温度变化驱使的.研究发现, 在冻融过程中温度和水分会发生剧烈变化, 在温度和水分胁迫下, 藓类植物能够通过失复水和调节体内渗透调节物质浓度来维持细胞渗透压(Nagao et al ., 2006 ; Lenne et al ., 2010 ), 同时通过抗氧化酶的合成来清除体内多余的活性氧和超氧物阴离子自由基Zhang & Kirkham, 1994 ; Meloni et al ., 2003 ), 进而减少其对植物组织的过度损伤, 稳定和维持细胞内大分子物质的结构与功能.此外, 在极地的研究也发现在极端低温状态下, 苔藓植物能够通过脱水来缓解低温对藓类植物的伤害(Lenne et al ., 2010 ), 这也可能是在冬季低温冻结期齿肋赤藓植株含水量相对较低的重要原因. ...
... ), 进而减少其对植物组织的过度损伤, 稳定和维持细胞内大分子物质的结构与功能.此外, 在极地的研究也发现在极端低温状态下, 苔藓植物能够通过脱水来缓解低温对藓类植物的伤害(Lenne et al ., 2010 ), 这也可能是在冬季低温冻结期齿肋赤藓植株含水量相对较低的重要原因. ...
Encapsulation of moss buds: An efficient method for the in vitro conservation and regeneration of the endangered moss Splachnum ampullaceum.
1
2007
... 苔藓植物是仅次于被子植物的第二大类高等植物类群, 在沼泽、森林、草地和荒漠区均起着重要的生态作用(Mallón et al ., 2007 ).作为环境演替的先锋物种, 耐旱苔藓植物在荒漠生态系统中分布广泛, 它们的存在能够在地表构筑一个粗糙的平面, 对荒漠生态系统的维持和稳定(Kidron et al ., 2009 )、土壤微环境的改善(Harper & Belnap, 2001 )具有重要 作用. ...
Pho- tosynthesis and activity of superoxide dismutase, peroxidase and glutathione reductase in cotton under salt stress.
2
2003
... 水分是干旱半干旱荒漠生态系统植物生长的主要限制因素, 对于变水植物齿肋赤藓, 水分能够在一定程度上反映植物的活性程度Brown & Ernest, 2002 ; Huxman et al ., 2004 ; Benassi et al ., 2011 ).而在冬春季节的冻融期, 水分的变化常常是受温度变化驱使的.研究发现, 在冻融过程中温度和水分会发生剧烈变化, 在温度和水分胁迫下, 藓类植物能够通过失复水和调节体内渗透调节物质浓度来维持细胞渗透压(Nagao et al ., 2006 ; Lenne et al ., 2010 ), 同时通过抗氧化酶的合成来清除体内多余的活性氧和超氧物阴离子自由基Zhang & Kirkham, 1994 ; Meloni et al ., 2003 ), 进而减少其对植物组织的过度损伤, 稳定和维持细胞内大分子物质的结构与功能.此外, 在极地的研究也发现在极端低温状态下, 苔藓植物能够通过脱水来缓解低温对藓类植物的伤害(Lenne et al ., 2010 ), 这也可能是在冬季低温冻结期齿肋赤藓植株含水量相对较低的重要原因. ...
... 为了缓解干旱和极端温度胁迫对细胞的过度损伤, 植物细胞能够通过合成大量的CAT、POD和SOD等抗氧化酶来清除多余的活性氧和超氧物阴离子自由基(Zhang & Kirkham, 1994 ; Meloni et al ., 2003 ).研究发现: 在水分充足的积雪融化期, 3种抗氧化酶活性均显著低于冬季积雪冻结期和春季中旬干旱无雨期.春季中旬干旱无雨期的相关性分析也表明3种抗氧化酶活性与植株含水量极显著负相关, 在水分相对较好的灌丛下齿肋赤藓3种抗氧化酶活性显著低于其他两个水分相对较差的区域, 这与张静等在研究模拟降水对齿肋赤藓生理生化特性的结果一致, 在干燥状态下齿肋赤藓能够通过增加CAT、POD和SOD 3种抗氧化酶的活性来抵御干旱环境对植物的伤害(张静和张元明, 2014 ).与干旱胁迫一样, 在应对极端温度时植物也会合成大量的抗氧化酶来清除多余的氧自由基.已有的研究发现在高温胁迫下植物体内SOD和POD等抗氧化酶活性显著增加(沈蕾等, 2011 ).本研究发现尽管冬季冻结期的含水量显著高于春季中旬干旱无雨期, 但极端的低温环境使得其表现为更高的CAT、POD和SOD等抗氧化酶活性, 这说明藓类植物在应对极端低温时也会通过合成大量的抗氧化酶来抵御极端低温对植物体的伤害. ...
Proline content of sugar beet storage roots: Response to water deficit and nitrogen fertilization at field conditions.
1
2007
... 本研究所测定的渗透调节物主要包括可溶性糖、可溶性蛋白和脯氨酸3种, 分别用蒽酮法、考马斯亮蓝法和酸性茚三酮法进行测定, 其吸光度值由Lamda35紫外分光光度计(PE, Waltham, USA)测定Gonzalez & Pignata; 1994 ; Monreal et al ., 2007 ; Lassouane et al ., 2013 ). ...
Enhanced photo-and antioxidative protection, and hydrogen peroxide accumulation in drought-stressed Cistus clusii and Cistus albidus plants.
0
2003
Accumulation of theanderose in association with development of freezing tolerance in the moss Physcomitrella patens.
2
2006
... 水分是干旱半干旱荒漠生态系统植物生长的主要限制因素, 对于变水植物齿肋赤藓, 水分能够在一定程度上反映植物的活性程度Brown & Ernest, 2002 ; Huxman et al ., 2004 ; Benassi et al ., 2011 ).而在冬春季节的冻融期, 水分的变化常常是受温度变化驱使的.研究发现, 在冻融过程中温度和水分会发生剧烈变化, 在温度和水分胁迫下, 藓类植物能够通过失复水和调节体内渗透调节物质浓度来维持细胞渗透压(Nagao et al ., 2006 ; Lenne et al ., 2010 ), 同时通过抗氧化酶的合成来清除体内多余的活性氧和超氧物阴离子自由基Zhang & Kirkham, 1994 ; Meloni et al ., 2003 ), 进而减少其对植物组织的过度损伤, 稳定和维持细胞内大分子物质的结构与功能.此外, 在极地的研究也发现在极端低温状态下, 苔藓植物能够通过脱水来缓解低温对藓类植物的伤害(Lenne et al ., 2010 ), 这也可能是在冬季低温冻结期齿肋赤藓植株含水量相对较低的重要原因. ...
... 植物的生存环境可以分为大环境和小环境, 大环境影响生物的生存和分布, 而小环境直接影响到生物的生活, 所以微生境间的差异也势必对荒漠藓类植物的生理生化特征产生一定程度的影响.研究发现在积雪融化期灌丛下的可溶性糖含量显著高于其他两个生境, 这可能是由于积雪融化期灌丛形成的“冷岛效应”使活灌丛温度明显低于裸露地和死灌丛(Nagao et al ., 2006 ; 张亚峰等, 2013 ).同时, 相关分析也表明, 在水分为非胁迫条件下可溶性糖含量与温度呈极显著负相关而与植株含水量无关.随着积雪融水的逐渐消失, 在春季中旬的干旱无雨期, 含水量相对较多的灌丛下齿肋赤藓的可溶性糖含量和脯氨酸含量显著低于死灌丛和裸露地, 且在3种不同微生境下与含水量存在显著负相关, 这与之前在对初冬降雪前后齿肋赤藓生理特性响应的研究结果一致尹本丰和张元明, 2014 ). ...
How some plants recover from vegetative desiccation: A repair based strategy.
1
1997
... 荒漠苔藓作为典型的抗胁迫变水植物已被广泛关注, 诸多研究表明藓类植物能够适应极端高低温、干燥、强光照等恶劣环境(张静和张元明, 2014 ).为适应极端环境, 藓类植物在种群分布、形态特征(郑云普等, 2009 ; 陶冶和张元明, 2012 )、繁殖方式(Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 )、光合活性(张静和张元明, 2011 )、渗透调节物质、抗氧化酶(回嵘等, 2012 ; 陈文佳等, 2013 )等方面进化出多样的抗胁迫特性.研究发现齿肋赤藓(Syntrichia caninervis )叶片表面具有蜡质层, 并在细胞表面富集疣状突起, 在一定程度上能够减弱热气流的影响(许书军, 2007 ).另外, 在高温干燥环境下藓类植物能够通过失水降低代谢活性而进入休眠状态, 并积累大量的脯氨酸和可溶性糖来提高细胞渗透压, 维持渗透压平衡(Oliver et al ., 1997 ; Crowe et al ., 1998 ; 徐杰等, 2005 ; 吴楠等, 2009 ).同时, 通过抗氧化酶的大量合成清除过多的氧自由基, 进而保护植物免受过度伤害(Sgherri et al ., 1994 ; Herrnstadt & Kidron, 2005 ; Pintado et al ., 2005 ; 沈蕾等, 2011 ).但对荒漠藓类植物如何通过生理生化调节并成功度过冬季的极端低温环境, 还有待于深入研究. ...
Functional ecology of the biological soil crust in semiarid SE Spain: Sun and shade populations of Diploschistes diacapsis (Ach.) Lumbsch.
2
2005
... 荒漠苔藓作为典型的抗胁迫变水植物已被广泛关注, 诸多研究表明藓类植物能够适应极端高低温、干燥、强光照等恶劣环境(张静和张元明, 2014 ).为适应极端环境, 藓类植物在种群分布、形态特征(郑云普等, 2009 ; 陶冶和张元明, 2012 )、繁殖方式(Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 )、光合活性(张静和张元明, 2011 )、渗透调节物质、抗氧化酶(回嵘等, 2012 ; 陈文佳等, 2013 )等方面进化出多样的抗胁迫特性.研究发现齿肋赤藓(Syntrichia caninervis )叶片表面具有蜡质层, 并在细胞表面富集疣状突起, 在一定程度上能够减弱热气流的影响(许书军, 2007 ).另外, 在高温干燥环境下藓类植物能够通过失水降低代谢活性而进入休眠状态, 并积累大量的脯氨酸和可溶性糖来提高细胞渗透压, 维持渗透压平衡(Oliver et al ., 1997 ; Crowe et al ., 1998 ; 徐杰等, 2005 ; 吴楠等, 2009 ).同时, 通过抗氧化酶的大量合成清除过多的氧自由基, 进而保护植物免受过度伤害(Sgherri et al ., 1994 ; Herrnstadt & Kidron, 2005 ; Pintado et al ., 2005 ; 沈蕾等, 2011 ).但对荒漠藓类植物如何通过生理生化调节并成功度过冬季的极端低温环境, 还有待于深入研究. ...
... 已有的研究和我们前期的实验表明, 微生境的不同也会造成藓类植物生存策略的差异.不少野外调查和研究发现, 遮阴下(灌丛或阴坡)苔藓植物的盖度和生长状况好于暴露环境下的苔藓(Pintado et al ., 2005 ; Zhang et al ., 2007 ).灌丛下的低温、低光照和高含水量与裸露地的高温、高光照和低含水量造成了不同微生境间苔藓形态的变化.为了适应不同微生境, 苔藓植物能够随环境变化而变化, 但是短期微生境变化对苔藓形态的影响并不明显, 而其光合作用与现有生存环境相似(Reynolds & Mc- Letchie, 2011 ).Benassi等(2011)的研究发现: 不同生境下苔藓的繁殖方式也存在显著差异, 遮阴下的苔藓植株更趋于有性繁殖.然而, 不同微生境下苔藓植物对冻融过程中温度和水分变化的适应性是否一致尚不得而知. ...
Short distances between extreme microhabitats do not result in ecotypes in Syntrichia caninervis.
4
2011
... 已有的研究和我们前期的实验表明, 微生境的不同也会造成藓类植物生存策略的差异.不少野外调查和研究发现, 遮阴下(灌丛或阴坡)苔藓植物的盖度和生长状况好于暴露环境下的苔藓(Pintado et al ., 2005 ; Zhang et al ., 2007 ).灌丛下的低温、低光照和高含水量与裸露地的高温、高光照和低含水量造成了不同微生境间苔藓形态的变化.为了适应不同微生境, 苔藓植物能够随环境变化而变化, 但是短期微生境变化对苔藓形态的影响并不明显, 而其光合作用与现有生存环境相似(Reynolds & Mc- Letchie, 2011 ).Benassi等(2011)的研究发现: 不同生境下苔藓的繁殖方式也存在显著差异, 遮阴下的苔藓植株更趋于有性繁殖.然而, 不同微生境下苔藓植物对冻融过程中温度和水分变化的适应性是否一致尚不得而知. ...
... 齿肋赤藓属于丛藓科赤藓属, 具有良好的耐旱性和变水特性, 是古尔班通古特沙漠苔藓结皮的优势种.该种较山墙藓(Tortula ruralis )和北方赤藓(Syntrichia norvegica )更能适应极端干旱、低海拔和高温环境, 在世界冷沙漠中具有广泛分布(Reynolds & McLetchie, 2011 ).在干燥条件下, 其个体通常呈现为黑色或淡棕色, 复水后能快速变为绿色, 并在短时间内恢复光合和生理活性张静和张元明, 2014 ).齿肋赤藓种群的大量存在对沙漠的固定和微环境的改善具有重要作用(张元明等, 2004 ; 吴楠等, 2009 ). ...
... 在温带荒漠冬、春季节, 温度和水分能够间接或直接的影响植物的生理生化过程张静和张元明, 2014 ).研究发现, 与荒漠藓类的形态特征随周围环境的变化相比, 其生理特征变化更为明显, 反应更为迅速Reynolds & McLetchie, 2011 ).本实验发现: 不同时期3种微生境的温度和水分等环境因子均存在显著差异, 随着不同时期水热条件的变化, 齿肋赤藓的含水量、可溶性糖、游离脯氨酸、可溶性蛋白、MDA等含量以及CAT、POD、SOD等抗氧化酶活性均发生显著变化. ...
... 有意义的是, 纵观整个冻融期, 3种微生境下齿肋赤藓的渗透调节物含量和MDA含量, 在冻融期的不同阶段均表现出不同趋势, 但其3种抗氧化酶活性却一致表现为灌丛下显著低于裸露地.这可能是由于裸露地的齿肋赤藓表现出更高的生理耐受性Reynolds & McLetchie, 2011 ), 这也与Stark等(2010)的研究认为的活灌丛下齿肋赤藓较裸露地齿肋赤藓抗旱性稍差相一致.但在不同生境、不同水热组合条件下, 齿肋赤藓与抗逆相关的基因表达是否存在差异还需进一步研究. ...
Impact of changes in natural ultraviolet radiation on pigment composition, physiological and morphological characteristics of the Antarctic moss, Grimmia antarctici.
1
2005
... 实验于2014年2至4月进行.采样时间分别为积雪覆盖下(2月冻结期)、积雪融化期(3月)和春季干燥无雨期(4月).分别于17:00左右采集不同微生境下的齿肋赤藓样品, 迅速置于装有冰袋的保温箱内.由于齿肋赤藓随环境变化其生理生化特性变化较快(Wu et al ., 2014 ), 为尽可能保持样品原状, 在日落后的低温状态下, 借助辅助光源, 在室外用刀片快速切取苔藓(随用随取)茎叶鲜样.样品过筛3次后(去除夹杂的沙粒), 为每一指标称取150 mg鲜质量(0.001 g精度天平), 用锡箔纸包裹并系上棉线, 置于液氮罐中快速冷冻保存, 随后带回实验室进行后续测定和分析(Robinson et al ., 2005 ). ...
Antioxidant system in Sporobolus stapfianus: Changes in response to desiccation and rehydration.
1
1994
... 荒漠苔藓作为典型的抗胁迫变水植物已被广泛关注, 诸多研究表明藓类植物能够适应极端高低温、干燥、强光照等恶劣环境(张静和张元明, 2014 ).为适应极端环境, 藓类植物在种群分布、形态特征(郑云普等, 2009 ; 陶冶和张元明, 2012 )、繁殖方式(Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 )、光合活性(张静和张元明, 2011 )、渗透调节物质、抗氧化酶(回嵘等, 2012 ; 陈文佳等, 2013 )等方面进化出多样的抗胁迫特性.研究发现齿肋赤藓(Syntrichia caninervis )叶片表面具有蜡质层, 并在细胞表面富集疣状突起, 在一定程度上能够减弱热气流的影响(许书军, 2007 ).另外, 在高温干燥环境下藓类植物能够通过失水降低代谢活性而进入休眠状态, 并积累大量的脯氨酸和可溶性糖来提高细胞渗透压, 维持渗透压平衡(Oliver et al ., 1997 ; Crowe et al ., 1998 ; 徐杰等, 2005 ; 吴楠等, 2009 ).同时, 通过抗氧化酶的大量合成清除过多的氧自由基, 进而保护植物免受过度伤害(Sgherri et al ., 1994 ; Herrnstadt & Kidron, 2005 ; Pintado et al ., 2005 ; 沈蕾等, 2011 ).但对荒漠藓类植物如何通过生理生化调节并成功度过冬季的极端低温环境, 还有待于深入研究. ...
旱后复水对东亚砂藓生理生化指标的影响
0
2010
旱后复水对东亚砂藓生理生化指标的影响
0
2010
多蒴灰藓(苔藓植物门: 藓纲)对短期极端温度的生理响应
3
2011
... 荒漠苔藓作为典型的抗胁迫变水植物已被广泛关注, 诸多研究表明藓类植物能够适应极端高低温、干燥、强光照等恶劣环境(张静和张元明, 2014 ).为适应极端环境, 藓类植物在种群分布、形态特征(郑云普等, 2009 ; 陶冶和张元明, 2012 )、繁殖方式(Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 )、光合活性(张静和张元明, 2011 )、渗透调节物质、抗氧化酶(回嵘等, 2012 ; 陈文佳等, 2013 )等方面进化出多样的抗胁迫特性.研究发现齿肋赤藓(Syntrichia caninervis )叶片表面具有蜡质层, 并在细胞表面富集疣状突起, 在一定程度上能够减弱热气流的影响(许书军, 2007 ).另外, 在高温干燥环境下藓类植物能够通过失水降低代谢活性而进入休眠状态, 并积累大量的脯氨酸和可溶性糖来提高细胞渗透压, 维持渗透压平衡(Oliver et al ., 1997 ; Crowe et al ., 1998 ; 徐杰等, 2005 ; 吴楠等, 2009 ).同时, 通过抗氧化酶的大量合成清除过多的氧自由基, 进而保护植物免受过度伤害(Sgherri et al ., 1994 ; Herrnstadt & Kidron, 2005 ; Pintado et al ., 2005 ; 沈蕾等, 2011 ).但对荒漠藓类植物如何通过生理生化调节并成功度过冬季的极端低温环境, 还有待于深入研究. ...
... 冬季的极端低温环境极易使得植物体内的水分凝结形成冰晶, 并对植物造成严重伤害.而可溶性糖的大量积累能够增加细胞液的浓度, 降低细胞内水势, 使细胞液的冰点下降从而增加植物的抗寒能力.不少研究发现随着低温处理时间的延长, 植物体内的可溶性糖含量显著增加(沈蕾等, 2011 ; 朱政等, 2011 ).这在本实验中也得到证实, 在冬季低温冻结期植物体内的可溶性糖含量显著高于融雪期, 这说明齿肋赤藓在极端低温环境下能够通过积累大量可溶性糖来保护自身(Bhyan et al ., 2012 ).同时研究发现, 在积雪融化后的干燥环境下, 藓类植物体内脯氨酸和可溶性糖含量均较其他两个时期显著增加, 这可能是由于地表土壤含水量减少使得藓类植物处于干旱胁迫状态引起的.这也与张静和张元明(2014)在模拟降水对齿肋赤藓生理生化特性时发现的随含水量的减少齿肋赤藓体内可溶性糖和与脯氨酸呈递增趋势的结果相一致.此外, 徐杰等(2005)在不同区域真藓(Bryum argenteum )和土生对齿藓(Didymodon vinealis )的研究也表明, 植物体内脯氨酸与可溶性糖含量在降水量相对较少的草原化荒漠区显著高于降水量较多的典型草原. ...
... 为了缓解干旱和极端温度胁迫对细胞的过度损伤, 植物细胞能够通过合成大量的CAT、POD和SOD等抗氧化酶来清除多余的活性氧和超氧物阴离子自由基(Zhang & Kirkham, 1994 ; Meloni et al ., 2003 ).研究发现: 在水分充足的积雪融化期, 3种抗氧化酶活性均显著低于冬季积雪冻结期和春季中旬干旱无雨期.春季中旬干旱无雨期的相关性分析也表明3种抗氧化酶活性与植株含水量极显著负相关, 在水分相对较好的灌丛下齿肋赤藓3种抗氧化酶活性显著低于其他两个水分相对较差的区域, 这与张静等在研究模拟降水对齿肋赤藓生理生化特性的结果一致, 在干燥状态下齿肋赤藓能够通过增加CAT、POD和SOD 3种抗氧化酶的活性来抵御干旱环境对植物的伤害(张静和张元明, 2014 ).与干旱胁迫一样, 在应对极端温度时植物也会合成大量的抗氧化酶来清除多余的氧自由基.已有的研究发现在高温胁迫下植物体内SOD和POD等抗氧化酶活性显著增加(沈蕾等, 2011 ).本研究发现尽管冬季冻结期的含水量显著高于春季中旬干旱无雨期, 但极端的低温环境使得其表现为更高的CAT、POD和SOD等抗氧化酶活性, 这说明藓类植物在应对极端低温时也会通过合成大量的抗氧化酶来抵御极端低温对植物体的伤害. ...
多蒴灰藓(苔藓植物门: 藓纲)对短期极端温度的生理响应
3
2011
... 荒漠苔藓作为典型的抗胁迫变水植物已被广泛关注, 诸多研究表明藓类植物能够适应极端高低温、干燥、强光照等恶劣环境(张静和张元明, 2014 ).为适应极端环境, 藓类植物在种群分布、形态特征(郑云普等, 2009 ; 陶冶和张元明, 2012 )、繁殖方式(Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 )、光合活性(张静和张元明, 2011 )、渗透调节物质、抗氧化酶(回嵘等, 2012 ; 陈文佳等, 2013 )等方面进化出多样的抗胁迫特性.研究发现齿肋赤藓(Syntrichia caninervis )叶片表面具有蜡质层, 并在细胞表面富集疣状突起, 在一定程度上能够减弱热气流的影响(许书军, 2007 ).另外, 在高温干燥环境下藓类植物能够通过失水降低代谢活性而进入休眠状态, 并积累大量的脯氨酸和可溶性糖来提高细胞渗透压, 维持渗透压平衡(Oliver et al ., 1997 ; Crowe et al ., 1998 ; 徐杰等, 2005 ; 吴楠等, 2009 ).同时, 通过抗氧化酶的大量合成清除过多的氧自由基, 进而保护植物免受过度伤害(Sgherri et al ., 1994 ; Herrnstadt & Kidron, 2005 ; Pintado et al ., 2005 ; 沈蕾等, 2011 ).但对荒漠藓类植物如何通过生理生化调节并成功度过冬季的极端低温环境, 还有待于深入研究. ...
... 冬季的极端低温环境极易使得植物体内的水分凝结形成冰晶, 并对植物造成严重伤害.而可溶性糖的大量积累能够增加细胞液的浓度, 降低细胞内水势, 使细胞液的冰点下降从而增加植物的抗寒能力.不少研究发现随着低温处理时间的延长, 植物体内的可溶性糖含量显著增加(沈蕾等, 2011 ; 朱政等, 2011 ).这在本实验中也得到证实, 在冬季低温冻结期植物体内的可溶性糖含量显著高于融雪期, 这说明齿肋赤藓在极端低温环境下能够通过积累大量可溶性糖来保护自身(Bhyan et al ., 2012 ).同时研究发现, 在积雪融化后的干燥环境下, 藓类植物体内脯氨酸和可溶性糖含量均较其他两个时期显著增加, 这可能是由于地表土壤含水量减少使得藓类植物处于干旱胁迫状态引起的.这也与张静和张元明(2014)在模拟降水对齿肋赤藓生理生化特性时发现的随含水量的减少齿肋赤藓体内可溶性糖和与脯氨酸呈递增趋势的结果相一致.此外, 徐杰等(2005)在不同区域真藓(Bryum argenteum )和土生对齿藓(Didymodon vinealis )的研究也表明, 植物体内脯氨酸与可溶性糖含量在降水量相对较少的草原化荒漠区显著高于降水量较多的典型草原. ...
... 为了缓解干旱和极端温度胁迫对细胞的过度损伤, 植物细胞能够通过合成大量的CAT、POD和SOD等抗氧化酶来清除多余的活性氧和超氧物阴离子自由基(Zhang & Kirkham, 1994 ; Meloni et al ., 2003 ).研究发现: 在水分充足的积雪融化期, 3种抗氧化酶活性均显著低于冬季积雪冻结期和春季中旬干旱无雨期.春季中旬干旱无雨期的相关性分析也表明3种抗氧化酶活性与植株含水量极显著负相关, 在水分相对较好的灌丛下齿肋赤藓3种抗氧化酶活性显著低于其他两个水分相对较差的区域, 这与张静等在研究模拟降水对齿肋赤藓生理生化特性的结果一致, 在干燥状态下齿肋赤藓能够通过增加CAT、POD和SOD 3种抗氧化酶的活性来抵御干旱环境对植物的伤害(张静和张元明, 2014 ).与干旱胁迫一样, 在应对极端温度时植物也会合成大量的抗氧化酶来清除多余的氧自由基.已有的研究发现在高温胁迫下植物体内SOD和POD等抗氧化酶活性显著增加(沈蕾等, 2011 ).本研究发现尽管冬季冻结期的含水量显著高于春季中旬干旱无雨期, 但极端的低温环境使得其表现为更高的CAT、POD和SOD等抗氧化酶活性, 这说明藓类植物在应对极端低温时也会通过合成大量的抗氧化酶来抵御极端低温对植物体的伤害. ...
逐渐干旱胁迫下生物土壤结皮中土生对齿藓和真藓信号转导物质的响应
0
2012a
逐渐干旱胁迫下生物土壤结皮中土生对齿藓和真藓信号转导物质的响应
0
2012a
逐渐干旱胁迫下生物土壤结皮中真藓和土生对齿藓的活性氧清除机制
0
2012b
逐渐干旱胁迫下生物土壤结皮中真藓和土生对齿藓的活性氧清除机制
0
2012b
Sex ratios and the shy male hypothesis in the moss Bryum argenteum (Bryaceae).
0
2010
Carbon flux in deserts depends on soil cover type: A case study in the Gurbantunggut desert, North China.
2
2013
... 在荒漠干燥少雨环境中, 藓类植物在全年的大部分时间均处于失活休眠状态.然而, 有研究发现, 荒漠苔藓的生长和繁殖主要发生在寒冷和湿润的冬季月份Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 ).既使已经冷适应的苔藓, 在温度、光照和湿度达到阈值时仍能够被快速激活(Lange, 2003 ; Kappen & Valladares, 2007 ; Bjerke 2011 ; Su et al ., 2013 ).初冬和早春的积雪融化和低温能够激活藓类植物的光合活性, 延长生物结皮光合活性时间, 增加结皮土壤的碳截获量(Su et al ., 2013 ; 尹本丰和张元明, 2014 ).因此, 苔藓植物更可能在初冬或初春积雪融化时表现出良好的生长特性.但关于寒冷生态系统不同微生境苔藓的生态学特征还知之甚少(Bjerke et al ., 2011 ). ...
... ).初冬和早春的积雪融化和低温能够激活藓类植物的光合活性, 延长生物结皮光合活性时间, 增加结皮土壤的碳截获量(Su et al ., 2013 ; 尹本丰和张元明, 2014 ).因此, 苔藓植物更可能在初冬或初春积雪融化时表现出良好的生长特性.但关于寒冷生态系统不同微生境苔藓的生态学特征还知之甚少(Bjerke et al ., 2011 ). ...
Response mechanisms of antioxidants in bryophyte (Hypnum plumaeforme) under the stress of single or combined Pb and/or Ni.
1
2009
... 丙二醛(MDA)含量和超氧化物歧化酶(SOD)、过氧化物酶(POD)及过氧化氢酶(CAT) 3种抗氧化酶活性的测定分别采用TBA法、羟胺法、愈创木酚法和可见光法进行测定Choudhury & Panda, 2005 ; Sun et al ., 2009 ; Wu et al ., 2012 ). ...
叶片毛尖对齿肋赤藓结皮凝结水形成及蒸发的影响
1
2012
... 荒漠苔藓作为典型的抗胁迫变水植物已被广泛关注, 诸多研究表明藓类植物能够适应极端高低温、干燥、强光照等恶劣环境(张静和张元明, 2014 ).为适应极端环境, 藓类植物在种群分布、形态特征(郑云普等, 2009 ; 陶冶和张元明, 2012 )、繁殖方式(Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 )、光合活性(张静和张元明, 2011 )、渗透调节物质、抗氧化酶(回嵘等, 2012 ; 陈文佳等, 2013 )等方面进化出多样的抗胁迫特性.研究发现齿肋赤藓(Syntrichia caninervis )叶片表面具有蜡质层, 并在细胞表面富集疣状突起, 在一定程度上能够减弱热气流的影响(许书军, 2007 ).另外, 在高温干燥环境下藓类植物能够通过失水降低代谢活性而进入休眠状态, 并积累大量的脯氨酸和可溶性糖来提高细胞渗透压, 维持渗透压平衡(Oliver et al ., 1997 ; Crowe et al ., 1998 ; 徐杰等, 2005 ; 吴楠等, 2009 ).同时, 通过抗氧化酶的大量合成清除过多的氧自由基, 进而保护植物免受过度伤害(Sgherri et al ., 1994 ; Herrnstadt & Kidron, 2005 ; Pintado et al ., 2005 ; 沈蕾等, 2011 ).但对荒漠藓类植物如何通过生理生化调节并成功度过冬季的极端低温环境, 还有待于深入研究. ...
叶片毛尖对齿肋赤藓结皮凝结水形成及蒸发的影响
1
2012
... 荒漠苔藓作为典型的抗胁迫变水植物已被广泛关注, 诸多研究表明藓类植物能够适应极端高低温、干燥、强光照等恶劣环境(张静和张元明, 2014 ).为适应极端环境, 藓类植物在种群分布、形态特征(郑云普等, 2009 ; 陶冶和张元明, 2012 )、繁殖方式(Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 )、光合活性(张静和张元明, 2011 )、渗透调节物质、抗氧化酶(回嵘等, 2012 ; 陈文佳等, 2013 )等方面进化出多样的抗胁迫特性.研究发现齿肋赤藓(Syntrichia caninervis )叶片表面具有蜡质层, 并在细胞表面富集疣状突起, 在一定程度上能够减弱热气流的影响(许书军, 2007 ).另外, 在高温干燥环境下藓类植物能够通过失水降低代谢活性而进入休眠状态, 并积累大量的脯氨酸和可溶性糖来提高细胞渗透压, 维持渗透压平衡(Oliver et al ., 1997 ; Crowe et al ., 1998 ; 徐杰等, 2005 ; 吴楠等, 2009 ).同时, 通过抗氧化酶的大量合成清除过多的氧自由基, 进而保护植物免受过度伤害(Sgherri et al ., 1994 ; Herrnstadt & Kidron, 2005 ; Pintado et al ., 2005 ; 沈蕾等, 2011 ).但对荒漠藓类植物如何通过生理生化调节并成功度过冬季的极端低温环境, 还有待于深入研究. ...
Physiological evaluation of drought stress tolerance and recovery in cauliflower (Brassica oleracea L.) seedlings treated with methyl jasmonate and coronatine.
1
2012
... 丙二醛(MDA)含量和超氧化物歧化酶(SOD)、过氧化物酶(POD)及过氧化氢酶(CAT) 3种抗氧化酶活性的测定分别采用TBA法、羟胺法、愈创木酚法和可见光法进行测定Choudhury & Panda, 2005 ; Sun et al ., 2009 ; Wu et al ., 2012 ). ...
生物土壤结皮中刺叶赤藓质膜透性对脱水, 复水过程的响应
2
2009
... 荒漠苔藓作为典型的抗胁迫变水植物已被广泛关注, 诸多研究表明藓类植物能够适应极端高低温、干燥、强光照等恶劣环境(张静和张元明, 2014 ).为适应极端环境, 藓类植物在种群分布、形态特征(郑云普等, 2009 ; 陶冶和张元明, 2012 )、繁殖方式(Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 )、光合活性(张静和张元明, 2011 )、渗透调节物质、抗氧化酶(回嵘等, 2012 ; 陈文佳等, 2013 )等方面进化出多样的抗胁迫特性.研究发现齿肋赤藓(Syntrichia caninervis )叶片表面具有蜡质层, 并在细胞表面富集疣状突起, 在一定程度上能够减弱热气流的影响(许书军, 2007 ).另外, 在高温干燥环境下藓类植物能够通过失水降低代谢活性而进入休眠状态, 并积累大量的脯氨酸和可溶性糖来提高细胞渗透压, 维持渗透压平衡(Oliver et al ., 1997 ; Crowe et al ., 1998 ; 徐杰等, 2005 ; 吴楠等, 2009 ).同时, 通过抗氧化酶的大量合成清除过多的氧自由基, 进而保护植物免受过度伤害(Sgherri et al ., 1994 ; Herrnstadt & Kidron, 2005 ; Pintado et al ., 2005 ; 沈蕾等, 2011 ).但对荒漠藓类植物如何通过生理生化调节并成功度过冬季的极端低温环境, 还有待于深入研究. ...
... 齿肋赤藓属于丛藓科赤藓属, 具有良好的耐旱性和变水特性, 是古尔班通古特沙漠苔藓结皮的优势种.该种较山墙藓(Tortula ruralis )和北方赤藓(Syntrichia norvegica )更能适应极端干旱、低海拔和高温环境, 在世界冷沙漠中具有广泛分布(Reynolds & McLetchie, 2011 ).在干燥条件下, 其个体通常呈现为黑色或淡棕色, 复水后能快速变为绿色, 并在短时间内恢复光合和生理活性张静和张元明, 2014 ).齿肋赤藓种群的大量存在对沙漠的固定和微环境的改善具有重要作用(张元明等, 2004 ; 吴楠等, 2009 ). ...
生物土壤结皮中刺叶赤藓质膜透性对脱水, 复水过程的响应
2
2009
... 荒漠苔藓作为典型的抗胁迫变水植物已被广泛关注, 诸多研究表明藓类植物能够适应极端高低温、干燥、强光照等恶劣环境(张静和张元明, 2014 ).为适应极端环境, 藓类植物在种群分布、形态特征(郑云普等, 2009 ; 陶冶和张元明, 2012 )、繁殖方式(Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 )、光合活性(张静和张元明, 2011 )、渗透调节物质、抗氧化酶(回嵘等, 2012 ; 陈文佳等, 2013 )等方面进化出多样的抗胁迫特性.研究发现齿肋赤藓(Syntrichia caninervis )叶片表面具有蜡质层, 并在细胞表面富集疣状突起, 在一定程度上能够减弱热气流的影响(许书军, 2007 ).另外, 在高温干燥环境下藓类植物能够通过失水降低代谢活性而进入休眠状态, 并积累大量的脯氨酸和可溶性糖来提高细胞渗透压, 维持渗透压平衡(Oliver et al ., 1997 ; Crowe et al ., 1998 ; 徐杰等, 2005 ; 吴楠等, 2009 ).同时, 通过抗氧化酶的大量合成清除过多的氧自由基, 进而保护植物免受过度伤害(Sgherri et al ., 1994 ; Herrnstadt & Kidron, 2005 ; Pintado et al ., 2005 ; 沈蕾等, 2011 ).但对荒漠藓类植物如何通过生理生化调节并成功度过冬季的极端低温环境, 还有待于深入研究. ...
... 齿肋赤藓属于丛藓科赤藓属, 具有良好的耐旱性和变水特性, 是古尔班通古特沙漠苔藓结皮的优势种.该种较山墙藓(Tortula ruralis )和北方赤藓(Syntrichia norvegica )更能适应极端干旱、低海拔和高温环境, 在世界冷沙漠中具有广泛分布(Reynolds & McLetchie, 2011 ).在干燥条件下, 其个体通常呈现为黑色或淡棕色, 复水后能快速变为绿色, 并在短时间内恢复光合和生理活性张静和张元明, 2014 ).齿肋赤藓种群的大量存在对沙漠的固定和微环境的改善具有重要作用(张元明等, 2004 ; 吴楠等, 2009 ). ...
Rapid adjustment of leaf angle explains how the desert moss, Syntrichia caninervis, copes with multiple resource limitations during rehydration.
1
2014
... 实验于2014年2至4月进行.采样时间分别为积雪覆盖下(2月冻结期)、积雪融化期(3月)和春季干燥无雨期(4月).分别于17:00左右采集不同微生境下的齿肋赤藓样品, 迅速置于装有冰袋的保温箱内.由于齿肋赤藓随环境变化其生理生化特性变化较快(Wu et al ., 2014 ), 为尽可能保持样品原状, 在日落后的低温状态下, 借助辅助光源, 在室外用刀片快速切取苔藓(随用随取)茎叶鲜样.样品过筛3次后(去除夹杂的沙粒), 为每一指标称取150 mg鲜质量(0.001 g精度天平), 用锡箔纸包裹并系上棉线, 置于液氮罐中快速冷冻保存, 随后带回实验室进行后续测定和分析(Robinson et al ., 2005 ). ...
水分和钙胁迫对苔藓植物生理生化指标的影响
1
2010
... 水分的严重匮乏与极端的温度均会造成细胞膜脂的过氧化程度加剧, 使得细胞产生大量的活性氧和超氧物阴离子自由基, 进而对细胞质膜的组成和完整性造成严重伤害, 主要表现在植物体内的MDA含量迅速增加(范苏鲁等, 2011 ).研究发现随着积雪融水的逐渐消失, 干旱胁迫加剧使得春季中旬干旱无雨期植物体内MDA含量显著高于融雪期.同时春季中旬干旱无雨期的相关分析也表明MDA含量与植株含水量呈显著负相关关系(表3 ).石勇等(2012b)在研究土生对齿藓(Didymodon vinealis )时也发现随着干旱的逐渐增加, 植物体内的MDA含量显著增加, 但在真藓中表现出相反的结果.也有研究发现水分胁迫对尖叶提灯藓(Mnium cuspidatum )、青藓(Brachythecium albicans )、石地藓(Reboulia hemisphaerica )的MDA含量影响较小(项俊等, 2010 ).这可能说明不同藓类植物在应对水分胁迫时具有不同的响应机制.此外, 在冬季低温冻结期植株的MDA含量显著高于积雪融化期和春季中旬干旱无雨期, 尽管在冬季低温冻结期的植株含水量高于春季中旬干旱无雨期.在水分丰富的积雪融化期, 灌丛下齿肋赤藓的MDA含量却显著高于裸露地, 这也进一步说明冬季的低温对植物造成了严重损伤, 这与沈蕾等(2011)对多蒴灰藓(Hypnum fertile )的低温处理结果相似. ...
水分和钙胁迫对苔藓植物生理生化指标的影响
1
2010
... 水分的严重匮乏与极端的温度均会造成细胞膜脂的过氧化程度加剧, 使得细胞产生大量的活性氧和超氧物阴离子自由基, 进而对细胞质膜的组成和完整性造成严重伤害, 主要表现在植物体内的MDA含量迅速增加(范苏鲁等, 2011 ).研究发现随着积雪融水的逐渐消失, 干旱胁迫加剧使得春季中旬干旱无雨期植物体内MDA含量显著高于融雪期.同时春季中旬干旱无雨期的相关分析也表明MDA含量与植株含水量呈显著负相关关系(表3 ).石勇等(2012b)在研究土生对齿藓(Didymodon vinealis )时也发现随着干旱的逐渐增加, 植物体内的MDA含量显著增加, 但在真藓中表现出相反的结果.也有研究发现水分胁迫对尖叶提灯藓(Mnium cuspidatum )、青藓(Brachythecium albicans )、石地藓(Reboulia hemisphaerica )的MDA含量影响较小(项俊等, 2010 ).这可能说明不同藓类植物在应对水分胁迫时具有不同的响应机制.此外, 在冬季低温冻结期植株的MDA含量显著高于积雪融化期和春季中旬干旱无雨期, 尽管在冬季低温冻结期的植株含水量高于春季中旬干旱无雨期.在水分丰富的积雪融化期, 灌丛下齿肋赤藓的MDA含量却显著高于裸露地, 这也进一步说明冬季的低温对植物造成了严重损伤, 这与沈蕾等(2011)对多蒴灰藓(Hypnum fertile )的低温处理结果相似. ...
干旱半干旱地区生物结皮层藓类植物氨基酸和营养物质组成特征及适应性分析
1
2005
... 荒漠苔藓作为典型的抗胁迫变水植物已被广泛关注, 诸多研究表明藓类植物能够适应极端高低温、干燥、强光照等恶劣环境(张静和张元明, 2014 ).为适应极端环境, 藓类植物在种群分布、形态特征(郑云普等, 2009 ; 陶冶和张元明, 2012 )、繁殖方式(Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 )、光合活性(张静和张元明, 2011 )、渗透调节物质、抗氧化酶(回嵘等, 2012 ; 陈文佳等, 2013 )等方面进化出多样的抗胁迫特性.研究发现齿肋赤藓(Syntrichia caninervis )叶片表面具有蜡质层, 并在细胞表面富集疣状突起, 在一定程度上能够减弱热气流的影响(许书军, 2007 ).另外, 在高温干燥环境下藓类植物能够通过失水降低代谢活性而进入休眠状态, 并积累大量的脯氨酸和可溶性糖来提高细胞渗透压, 维持渗透压平衡(Oliver et al ., 1997 ; Crowe et al ., 1998 ; 徐杰等, 2005 ; 吴楠等, 2009 ).同时, 通过抗氧化酶的大量合成清除过多的氧自由基, 进而保护植物免受过度伤害(Sgherri et al ., 1994 ; Herrnstadt & Kidron, 2005 ; Pintado et al ., 2005 ; 沈蕾等, 2011 ).但对荒漠藓类植物如何通过生理生化调节并成功度过冬季的极端低温环境, 还有待于深入研究. ...
干旱半干旱地区生物结皮层藓类植物氨基酸和营养物质组成特征及适应性分析
1
2005
... 荒漠苔藓作为典型的抗胁迫变水植物已被广泛关注, 诸多研究表明藓类植物能够适应极端高低温、干燥、强光照等恶劣环境(张静和张元明, 2014 ).为适应极端环境, 藓类植物在种群分布、形态特征(郑云普等, 2009 ; 陶冶和张元明, 2012 )、繁殖方式(Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 )、光合活性(张静和张元明, 2011 )、渗透调节物质、抗氧化酶(回嵘等, 2012 ; 陈文佳等, 2013 )等方面进化出多样的抗胁迫特性.研究发现齿肋赤藓(Syntrichia caninervis )叶片表面具有蜡质层, 并在细胞表面富集疣状突起, 在一定程度上能够减弱热气流的影响(许书军, 2007 ).另外, 在高温干燥环境下藓类植物能够通过失水降低代谢活性而进入休眠状态, 并积累大量的脯氨酸和可溶性糖来提高细胞渗透压, 维持渗透压平衡(Oliver et al ., 1997 ; Crowe et al ., 1998 ; 徐杰等, 2005 ; 吴楠等, 2009 ).同时, 通过抗氧化酶的大量合成清除过多的氧自由基, 进而保护植物免受过度伤害(Sgherri et al ., 1994 ; Herrnstadt & Kidron, 2005 ; Pintado et al ., 2005 ; 沈蕾等, 2011 ).但对荒漠藓类植物如何通过生理生化调节并成功度过冬季的极端低温环境, 还有待于深入研究. ...
典型荒漠苔藓人工繁殖特征与抗御干热环境胁迫的生理生化机制研究
1
2007
... 荒漠苔藓作为典型的抗胁迫变水植物已被广泛关注, 诸多研究表明藓类植物能够适应极端高低温、干燥、强光照等恶劣环境(张静和张元明, 2014 ).为适应极端环境, 藓类植物在种群分布、形态特征(郑云普等, 2009 ; 陶冶和张元明, 2012 )、繁殖方式(Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 )、光合活性(张静和张元明, 2011 )、渗透调节物质、抗氧化酶(回嵘等, 2012 ; 陈文佳等, 2013 )等方面进化出多样的抗胁迫特性.研究发现齿肋赤藓(Syntrichia caninervis )叶片表面具有蜡质层, 并在细胞表面富集疣状突起, 在一定程度上能够减弱热气流的影响(许书军, 2007 ).另外, 在高温干燥环境下藓类植物能够通过失水降低代谢活性而进入休眠状态, 并积累大量的脯氨酸和可溶性糖来提高细胞渗透压, 维持渗透压平衡(Oliver et al ., 1997 ; Crowe et al ., 1998 ; 徐杰等, 2005 ; 吴楠等, 2009 ).同时, 通过抗氧化酶的大量合成清除过多的氧自由基, 进而保护植物免受过度伤害(Sgherri et al ., 1994 ; Herrnstadt & Kidron, 2005 ; Pintado et al ., 2005 ; 沈蕾等, 2011 ).但对荒漠藓类植物如何通过生理生化调节并成功度过冬季的极端低温环境, 还有待于深入研究. ...
典型荒漠苔藓人工繁殖特征与抗御干热环境胁迫的生理生化机制研究
1
2007
... 荒漠苔藓作为典型的抗胁迫变水植物已被广泛关注, 诸多研究表明藓类植物能够适应极端高低温、干燥、强光照等恶劣环境(张静和张元明, 2014 ).为适应极端环境, 藓类植物在种群分布、形态特征(郑云普等, 2009 ; 陶冶和张元明, 2012 )、繁殖方式(Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 )、光合活性(张静和张元明, 2011 )、渗透调节物质、抗氧化酶(回嵘等, 2012 ; 陈文佳等, 2013 )等方面进化出多样的抗胁迫特性.研究发现齿肋赤藓(Syntrichia caninervis )叶片表面具有蜡质层, 并在细胞表面富集疣状突起, 在一定程度上能够减弱热气流的影响(许书军, 2007 ).另外, 在高温干燥环境下藓类植物能够通过失水降低代谢活性而进入休眠状态, 并积累大量的脯氨酸和可溶性糖来提高细胞渗透压, 维持渗透压平衡(Oliver et al ., 1997 ; Crowe et al ., 1998 ; 徐杰等, 2005 ; 吴楠等, 2009 ).同时, 通过抗氧化酶的大量合成清除过多的氧自由基, 进而保护植物免受过度伤害(Sgherri et al ., 1994 ; Herrnstadt & Kidron, 2005 ; Pintado et al ., 2005 ; 沈蕾等, 2011 ).但对荒漠藓类植物如何通过生理生化调节并成功度过冬季的极端低温环境, 还有待于深入研究. ...
荒漠区不同微生境下齿肋赤藓对一次降雪的生理生化响应
3
2014
... 在荒漠干燥少雨环境中, 藓类植物在全年的大部分时间均处于失活休眠状态.然而, 有研究发现, 荒漠苔藓的生长和繁殖主要发生在寒冷和湿润的冬季月份Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 ).既使已经冷适应的苔藓, 在温度、光照和湿度达到阈值时仍能够被快速激活(Lange, 2003 ; Kappen & Valladares, 2007 ; Bjerke 2011 ; Su et al ., 2013 ).初冬和早春的积雪融化和低温能够激活藓类植物的光合活性, 延长生物结皮光合活性时间, 增加结皮土壤的碳截获量(Su et al ., 2013 ; 尹本丰和张元明, 2014 ).因此, 苔藓植物更可能在初冬或初春积雪融化时表现出良好的生长特性.但关于寒冷生态系统不同微生境苔藓的生态学特征还知之甚少(Bjerke et al ., 2011 ). ...
... 在每个样方内, 用高度为5 cm的环刀取0-5 cm土层土壤带回实验室风干磨碎过2 mm筛.用重铬酸钾容量法-外加热法测量土壤有机碳, 高氯酸-硫酸消化法测定全氮、酸溶-钼锑抗比色法测定全磷、酸溶-火焰光度法测定全钾.pH和电导率用pHS-3C精密pH计和DDS-307电导率仪(上海雷磁仪器厂)测定(水土稀释质量比5:1).采用残渣烘干-质量法测定土壤总盐含量尹本丰和张元明, 2014 ). ...
... 植物的生存环境可以分为大环境和小环境, 大环境影响生物的生存和分布, 而小环境直接影响到生物的生活, 所以微生境间的差异也势必对荒漠藓类植物的生理生化特征产生一定程度的影响.研究发现在积雪融化期灌丛下的可溶性糖含量显著高于其他两个生境, 这可能是由于积雪融化期灌丛形成的“冷岛效应”使活灌丛温度明显低于裸露地和死灌丛(Nagao et al ., 2006 ; 张亚峰等, 2013 ).同时, 相关分析也表明, 在水分为非胁迫条件下可溶性糖含量与温度呈极显著负相关而与植株含水量无关.随着积雪融水的逐渐消失, 在春季中旬的干旱无雨期, 含水量相对较多的灌丛下齿肋赤藓的可溶性糖含量和脯氨酸含量显著低于死灌丛和裸露地, 且在3种不同微生境下与含水量存在显著负相关, 这与之前在对初冬降雪前后齿肋赤藓生理特性响应的研究结果一致尹本丰和张元明, 2014 ). ...
荒漠区不同微生境下齿肋赤藓对一次降雪的生理生化响应
3
2014
... 在荒漠干燥少雨环境中, 藓类植物在全年的大部分时间均处于失活休眠状态.然而, 有研究发现, 荒漠苔藓的生长和繁殖主要发生在寒冷和湿润的冬季月份Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 ).既使已经冷适应的苔藓, 在温度、光照和湿度达到阈值时仍能够被快速激活(Lange, 2003 ; Kappen & Valladares, 2007 ; Bjerke 2011 ; Su et al ., 2013 ).初冬和早春的积雪融化和低温能够激活藓类植物的光合活性, 延长生物结皮光合活性时间, 增加结皮土壤的碳截获量(Su et al ., 2013 ; 尹本丰和张元明, 2014 ).因此, 苔藓植物更可能在初冬或初春积雪融化时表现出良好的生长特性.但关于寒冷生态系统不同微生境苔藓的生态学特征还知之甚少(Bjerke et al ., 2011 ). ...
... 在每个样方内, 用高度为5 cm的环刀取0-5 cm土层土壤带回实验室风干磨碎过2 mm筛.用重铬酸钾容量法-外加热法测量土壤有机碳, 高氯酸-硫酸消化法测定全氮、酸溶-钼锑抗比色法测定全磷、酸溶-火焰光度法测定全钾.pH和电导率用pHS-3C精密pH计和DDS-307电导率仪(上海雷磁仪器厂)测定(水土稀释质量比5:1).采用残渣烘干-质量法测定土壤总盐含量尹本丰和张元明, 2014 ). ...
... 植物的生存环境可以分为大环境和小环境, 大环境影响生物的生存和分布, 而小环境直接影响到生物的生活, 所以微生境间的差异也势必对荒漠藓类植物的生理生化特征产生一定程度的影响.研究发现在积雪融化期灌丛下的可溶性糖含量显著高于其他两个生境, 这可能是由于积雪融化期灌丛形成的“冷岛效应”使活灌丛温度明显低于裸露地和死灌丛(Nagao et al ., 2006 ; 张亚峰等, 2013 ).同时, 相关分析也表明, 在水分为非胁迫条件下可溶性糖含量与温度呈极显著负相关而与植株含水量无关.随着积雪融水的逐渐消失, 在春季中旬的干旱无雨期, 含水量相对较多的灌丛下齿肋赤藓的可溶性糖含量和脯氨酸含量显著低于死灌丛和裸露地, 且在3种不同微生境下与含水量存在显著负相关, 这与之前在对初冬降雪前后齿肋赤藓生理特性响应的研究结果一致尹本丰和张元明, 2014 ). ...
冻融过程对生物结皮中齿肋赤藓叶绿素荧光特性的影响
1
2011
... 荒漠苔藓作为典型的抗胁迫变水植物已被广泛关注, 诸多研究表明藓类植物能够适应极端高低温、干燥、强光照等恶劣环境(张静和张元明, 2014 ).为适应极端环境, 藓类植物在种群分布、形态特征(郑云普等, 2009 ; 陶冶和张元明, 2012 )、繁殖方式(Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 )、光合活性(张静和张元明, 2011 )、渗透调节物质、抗氧化酶(回嵘等, 2012 ; 陈文佳等, 2013 )等方面进化出多样的抗胁迫特性.研究发现齿肋赤藓(Syntrichia caninervis )叶片表面具有蜡质层, 并在细胞表面富集疣状突起, 在一定程度上能够减弱热气流的影响(许书军, 2007 ).另外, 在高温干燥环境下藓类植物能够通过失水降低代谢活性而进入休眠状态, 并积累大量的脯氨酸和可溶性糖来提高细胞渗透压, 维持渗透压平衡(Oliver et al ., 1997 ; Crowe et al ., 1998 ; 徐杰等, 2005 ; 吴楠等, 2009 ).同时, 通过抗氧化酶的大量合成清除过多的氧自由基, 进而保护植物免受过度伤害(Sgherri et al ., 1994 ; Herrnstadt & Kidron, 2005 ; Pintado et al ., 2005 ; 沈蕾等, 2011 ).但对荒漠藓类植物如何通过生理生化调节并成功度过冬季的极端低温环境, 还有待于深入研究. ...
冻融过程对生物结皮中齿肋赤藓叶绿素荧光特性的影响
1
2011
... 荒漠苔藓作为典型的抗胁迫变水植物已被广泛关注, 诸多研究表明藓类植物能够适应极端高低温、干燥、强光照等恶劣环境(张静和张元明, 2014 ).为适应极端环境, 藓类植物在种群分布、形态特征(郑云普等, 2009 ; 陶冶和张元明, 2012 )、繁殖方式(Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 )、光合活性(张静和张元明, 2011 )、渗透调节物质、抗氧化酶(回嵘等, 2012 ; 陈文佳等, 2013 )等方面进化出多样的抗胁迫特性.研究发现齿肋赤藓(Syntrichia caninervis )叶片表面具有蜡质层, 并在细胞表面富集疣状突起, 在一定程度上能够减弱热气流的影响(许书军, 2007 ).另外, 在高温干燥环境下藓类植物能够通过失水降低代谢活性而进入休眠状态, 并积累大量的脯氨酸和可溶性糖来提高细胞渗透压, 维持渗透压平衡(Oliver et al ., 1997 ; Crowe et al ., 1998 ; 徐杰等, 2005 ; 吴楠等, 2009 ).同时, 通过抗氧化酶的大量合成清除过多的氧自由基, 进而保护植物免受过度伤害(Sgherri et al ., 1994 ; Herrnstadt & Kidron, 2005 ; Pintado et al ., 2005 ; 沈蕾等, 2011 ).但对荒漠藓类植物如何通过生理生化调节并成功度过冬季的极端低温环境, 还有待于深入研究. ...
模拟降雨对齿肋赤藓(Syntrichia caninervis)生理特性的影响
4
2014
... 荒漠苔藓作为典型的抗胁迫变水植物已被广泛关注, 诸多研究表明藓类植物能够适应极端高低温、干燥、强光照等恶劣环境(张静和张元明, 2014 ).为适应极端环境, 藓类植物在种群分布、形态特征(郑云普等, 2009 ; 陶冶和张元明, 2012 )、繁殖方式(Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 )、光合活性(张静和张元明, 2011 )、渗透调节物质、抗氧化酶(回嵘等, 2012 ; 陈文佳等, 2013 )等方面进化出多样的抗胁迫特性.研究发现齿肋赤藓(Syntrichia caninervis )叶片表面具有蜡质层, 并在细胞表面富集疣状突起, 在一定程度上能够减弱热气流的影响(许书军, 2007 ).另外, 在高温干燥环境下藓类植物能够通过失水降低代谢活性而进入休眠状态, 并积累大量的脯氨酸和可溶性糖来提高细胞渗透压, 维持渗透压平衡(Oliver et al ., 1997 ; Crowe et al ., 1998 ; 徐杰等, 2005 ; 吴楠等, 2009 ).同时, 通过抗氧化酶的大量合成清除过多的氧自由基, 进而保护植物免受过度伤害(Sgherri et al ., 1994 ; Herrnstadt & Kidron, 2005 ; Pintado et al ., 2005 ; 沈蕾等, 2011 ).但对荒漠藓类植物如何通过生理生化调节并成功度过冬季的极端低温环境, 还有待于深入研究. ...
... 齿肋赤藓属于丛藓科赤藓属, 具有良好的耐旱性和变水特性, 是古尔班通古特沙漠苔藓结皮的优势种.该种较山墙藓(Tortula ruralis )和北方赤藓(Syntrichia norvegica )更能适应极端干旱、低海拔和高温环境, 在世界冷沙漠中具有广泛分布(Reynolds & McLetchie, 2011 ).在干燥条件下, 其个体通常呈现为黑色或淡棕色, 复水后能快速变为绿色, 并在短时间内恢复光合和生理活性张静和张元明, 2014 ).齿肋赤藓种群的大量存在对沙漠的固定和微环境的改善具有重要作用(张元明等, 2004 ; 吴楠等, 2009 ). ...
... 在温带荒漠冬、春季节, 温度和水分能够间接或直接的影响植物的生理生化过程张静和张元明, 2014 ).研究发现, 与荒漠藓类的形态特征随周围环境的变化相比, 其生理特征变化更为明显, 反应更为迅速Reynolds & McLetchie, 2011 ).本实验发现: 不同时期3种微生境的温度和水分等环境因子均存在显著差异, 随着不同时期水热条件的变化, 齿肋赤藓的含水量、可溶性糖、游离脯氨酸、可溶性蛋白、MDA等含量以及CAT、POD、SOD等抗氧化酶活性均发生显著变化. ...
... 为了缓解干旱和极端温度胁迫对细胞的过度损伤, 植物细胞能够通过合成大量的CAT、POD和SOD等抗氧化酶来清除多余的活性氧和超氧物阴离子自由基(Zhang & Kirkham, 1994 ; Meloni et al ., 2003 ).研究发现: 在水分充足的积雪融化期, 3种抗氧化酶活性均显著低于冬季积雪冻结期和春季中旬干旱无雨期.春季中旬干旱无雨期的相关性分析也表明3种抗氧化酶活性与植株含水量极显著负相关, 在水分相对较好的灌丛下齿肋赤藓3种抗氧化酶活性显著低于其他两个水分相对较差的区域, 这与张静等在研究模拟降水对齿肋赤藓生理生化特性的结果一致, 在干燥状态下齿肋赤藓能够通过增加CAT、POD和SOD 3种抗氧化酶的活性来抵御干旱环境对植物的伤害(张静和张元明, 2014 ).与干旱胁迫一样, 在应对极端温度时植物也会合成大量的抗氧化酶来清除多余的氧自由基.已有的研究发现在高温胁迫下植物体内SOD和POD等抗氧化酶活性显著增加(沈蕾等, 2011 ).本研究发现尽管冬季冻结期的含水量显著高于春季中旬干旱无雨期, 但极端的低温环境使得其表现为更高的CAT、POD和SOD等抗氧化酶活性, 这说明藓类植物在应对极端低温时也会通过合成大量的抗氧化酶来抵御极端低温对植物体的伤害. ...
模拟降雨对齿肋赤藓(Syntrichia caninervis)生理特性的影响
4
2014
... 荒漠苔藓作为典型的抗胁迫变水植物已被广泛关注, 诸多研究表明藓类植物能够适应极端高低温、干燥、强光照等恶劣环境(张静和张元明, 2014 ).为适应极端环境, 藓类植物在种群分布、形态特征(郑云普等, 2009 ; 陶冶和张元明, 2012 )、繁殖方式(Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 )、光合活性(张静和张元明, 2011 )、渗透调节物质、抗氧化酶(回嵘等, 2012 ; 陈文佳等, 2013 )等方面进化出多样的抗胁迫特性.研究发现齿肋赤藓(Syntrichia caninervis )叶片表面具有蜡质层, 并在细胞表面富集疣状突起, 在一定程度上能够减弱热气流的影响(许书军, 2007 ).另外, 在高温干燥环境下藓类植物能够通过失水降低代谢活性而进入休眠状态, 并积累大量的脯氨酸和可溶性糖来提高细胞渗透压, 维持渗透压平衡(Oliver et al ., 1997 ; Crowe et al ., 1998 ; 徐杰等, 2005 ; 吴楠等, 2009 ).同时, 通过抗氧化酶的大量合成清除过多的氧自由基, 进而保护植物免受过度伤害(Sgherri et al ., 1994 ; Herrnstadt & Kidron, 2005 ; Pintado et al ., 2005 ; 沈蕾等, 2011 ).但对荒漠藓类植物如何通过生理生化调节并成功度过冬季的极端低温环境, 还有待于深入研究. ...
... 齿肋赤藓属于丛藓科赤藓属, 具有良好的耐旱性和变水特性, 是古尔班通古特沙漠苔藓结皮的优势种.该种较山墙藓(Tortula ruralis )和北方赤藓(Syntrichia norvegica )更能适应极端干旱、低海拔和高温环境, 在世界冷沙漠中具有广泛分布(Reynolds & McLetchie, 2011 ).在干燥条件下, 其个体通常呈现为黑色或淡棕色, 复水后能快速变为绿色, 并在短时间内恢复光合和生理活性张静和张元明, 2014 ).齿肋赤藓种群的大量存在对沙漠的固定和微环境的改善具有重要作用(张元明等, 2004 ; 吴楠等, 2009 ). ...
... 在温带荒漠冬、春季节, 温度和水分能够间接或直接的影响植物的生理生化过程张静和张元明, 2014 ).研究发现, 与荒漠藓类的形态特征随周围环境的变化相比, 其生理特征变化更为明显, 反应更为迅速Reynolds & McLetchie, 2011 ).本实验发现: 不同时期3种微生境的温度和水分等环境因子均存在显著差异, 随着不同时期水热条件的变化, 齿肋赤藓的含水量、可溶性糖、游离脯氨酸、可溶性蛋白、MDA等含量以及CAT、POD、SOD等抗氧化酶活性均发生显著变化. ...
... 为了缓解干旱和极端温度胁迫对细胞的过度损伤, 植物细胞能够通过合成大量的CAT、POD和SOD等抗氧化酶来清除多余的活性氧和超氧物阴离子自由基(Zhang & Kirkham, 1994 ; Meloni et al ., 2003 ).研究发现: 在水分充足的积雪融化期, 3种抗氧化酶活性均显著低于冬季积雪冻结期和春季中旬干旱无雨期.春季中旬干旱无雨期的相关性分析也表明3种抗氧化酶活性与植株含水量极显著负相关, 在水分相对较好的灌丛下齿肋赤藓3种抗氧化酶活性显著低于其他两个水分相对较差的区域, 这与张静等在研究模拟降水对齿肋赤藓生理生化特性的结果一致, 在干燥状态下齿肋赤藓能够通过增加CAT、POD和SOD 3种抗氧化酶的活性来抵御干旱环境对植物的伤害(张静和张元明, 2014 ).与干旱胁迫一样, 在应对极端温度时植物也会合成大量的抗氧化酶来清除多余的氧自由基.已有的研究发现在高温胁迫下植物体内SOD和POD等抗氧化酶活性显著增加(沈蕾等, 2011 ).本研究发现尽管冬季冻结期的含水量显著高于春季中旬干旱无雨期, 但极端的低温环境使得其表现为更高的CAT、POD和SOD等抗氧化酶活性, 这说明藓类植物在应对极端低温时也会通过合成大量的抗氧化酶来抵御极端低温对植物体的伤害. ...
Drought-stress-induced changes in activities of superoxide dismutase, catalase, and peroxidase in wheat species.
2
1994
... 水分是干旱半干旱荒漠生态系统植物生长的主要限制因素, 对于变水植物齿肋赤藓, 水分能够在一定程度上反映植物的活性程度Brown & Ernest, 2002 ; Huxman et al ., 2004 ; Benassi et al ., 2011 ).而在冬春季节的冻融期, 水分的变化常常是受温度变化驱使的.研究发现, 在冻融过程中温度和水分会发生剧烈变化, 在温度和水分胁迫下, 藓类植物能够通过失复水和调节体内渗透调节物质浓度来维持细胞渗透压(Nagao et al ., 2006 ; Lenne et al ., 2010 ), 同时通过抗氧化酶的合成来清除体内多余的活性氧和超氧物阴离子自由基Zhang & Kirkham, 1994 ; Meloni et al ., 2003 ), 进而减少其对植物组织的过度损伤, 稳定和维持细胞内大分子物质的结构与功能.此外, 在极地的研究也发现在极端低温状态下, 苔藓植物能够通过脱水来缓解低温对藓类植物的伤害(Lenne et al ., 2010 ), 这也可能是在冬季低温冻结期齿肋赤藓植株含水量相对较低的重要原因. ...
... 为了缓解干旱和极端温度胁迫对细胞的过度损伤, 植物细胞能够通过合成大量的CAT、POD和SOD等抗氧化酶来清除多余的活性氧和超氧物阴离子自由基(Zhang & Kirkham, 1994 ; Meloni et al ., 2003 ).研究发现: 在水分充足的积雪融化期, 3种抗氧化酶活性均显著低于冬季积雪冻结期和春季中旬干旱无雨期.春季中旬干旱无雨期的相关性分析也表明3种抗氧化酶活性与植株含水量极显著负相关, 在水分相对较好的灌丛下齿肋赤藓3种抗氧化酶活性显著低于其他两个水分相对较差的区域, 这与张静等在研究模拟降水对齿肋赤藓生理生化特性的结果一致, 在干燥状态下齿肋赤藓能够通过增加CAT、POD和SOD 3种抗氧化酶的活性来抵御干旱环境对植物的伤害(张静和张元明, 2014 ).与干旱胁迫一样, 在应对极端温度时植物也会合成大量的抗氧化酶来清除多余的氧自由基.已有的研究发现在高温胁迫下植物体内SOD和POD等抗氧化酶活性显著增加(沈蕾等, 2011 ).本研究发现尽管冬季冻结期的含水量显著高于春季中旬干旱无雨期, 但极端的低温环境使得其表现为更高的CAT、POD和SOD等抗氧化酶活性, 这说明藓类植物在应对极端低温时也会通过合成大量的抗氧化酶来抵御极端低温对植物体的伤害. ...
荒漠灌丛微生境土壤温度的时空变异特征——灌丛与降水的影响
1
2013
... 植物的生存环境可以分为大环境和小环境, 大环境影响生物的生存和分布, 而小环境直接影响到生物的生活, 所以微生境间的差异也势必对荒漠藓类植物的生理生化特征产生一定程度的影响.研究发现在积雪融化期灌丛下的可溶性糖含量显著高于其他两个生境, 这可能是由于积雪融化期灌丛形成的“冷岛效应”使活灌丛温度明显低于裸露地和死灌丛(Nagao et al ., 2006 ; 张亚峰等, 2013 ).同时, 相关分析也表明, 在水分为非胁迫条件下可溶性糖含量与温度呈极显著负相关而与植株含水量无关.随着积雪融水的逐渐消失, 在春季中旬的干旱无雨期, 含水量相对较多的灌丛下齿肋赤藓的可溶性糖含量和脯氨酸含量显著低于死灌丛和裸露地, 且在3种不同微生境下与含水量存在显著负相关, 这与之前在对初冬降雪前后齿肋赤藓生理特性响应的研究结果一致尹本丰和张元明, 2014 ). ...
荒漠灌丛微生境土壤温度的时空变异特征——灌丛与降水的影响
1
2013
... 植物的生存环境可以分为大环境和小环境, 大环境影响生物的生存和分布, 而小环境直接影响到生物的生活, 所以微生境间的差异也势必对荒漠藓类植物的生理生化特征产生一定程度的影响.研究发现在积雪融化期灌丛下的可溶性糖含量显著高于其他两个生境, 这可能是由于积雪融化期灌丛形成的“冷岛效应”使活灌丛温度明显低于裸露地和死灌丛(Nagao et al ., 2006 ; 张亚峰等, 2013 ).同时, 相关分析也表明, 在水分为非胁迫条件下可溶性糖含量与温度呈极显著负相关而与植株含水量无关.随着积雪融水的逐渐消失, 在春季中旬的干旱无雨期, 含水量相对较多的灌丛下齿肋赤藓的可溶性糖含量和脯氨酸含量显著低于死灌丛和裸露地, 且在3种不同微生境下与含水量存在显著负相关, 这与之前在对初冬降雪前后齿肋赤藓生理特性响应的研究结果一致尹本丰和张元明, 2014 ). ...
荒漠地表生物土壤结皮的微结构及其早期发育特征
1
2005
... 该沙漠春季融雪使得地表土壤含水量显著增加, 荒漠短命植物、类短命植物、一年及多年生草本和隐花植物广泛发育和生长.白梭梭(Haloxylon persicum) 和梭梭(Haloxylon ammodendron )等小半乔木作为古尔班通古特沙漠的建群种, 分别占据了沙垄的上部和中下部.蛇麻黄(Ephedra distachya )、淡枝沙拐枣(Calligonum leucocladum )等灌木也在沙垄间广泛分布.同时, 灌木和生物结皮镶嵌分布是荒漠区的主要地表景观.该沙漠生物结皮主要分为藻结皮、地衣结皮和苔藓结皮3种类型(张元明, 2005 ).其中, 藻结皮多分布在沙垄中上部, 地衣和苔藓结皮多分布在中下部和丘间低地, 尤其在蛇麻黄、沙拐枣(Calligonum spp.)等灌丛下通常丛聚着发育良好的苔藓结皮. ...
荒漠地表生物土壤结皮的微结构及其早期发育特征
1
2005
... 该沙漠春季融雪使得地表土壤含水量显著增加, 荒漠短命植物、类短命植物、一年及多年生草本和隐花植物广泛发育和生长.白梭梭(Haloxylon persicum) 和梭梭(Haloxylon ammodendron )等小半乔木作为古尔班通古特沙漠的建群种, 分别占据了沙垄的上部和中下部.蛇麻黄(Ephedra distachya )、淡枝沙拐枣(Calligonum leucocladum )等灌木也在沙垄间广泛分布.同时, 灌木和生物结皮镶嵌分布是荒漠区的主要地表景观.该沙漠生物结皮主要分为藻结皮、地衣结皮和苔藓结皮3种类型(张元明, 2005 ).其中, 藻结皮多分布在沙垄中上部, 地衣和苔藓结皮多分布在中下部和丘间低地, 尤其在蛇麻黄、沙拐枣(Calligonum spp.)等灌丛下通常丛聚着发育良好的苔藓结皮. ...
The spatial distribution patterns of biological soil crusts in the Gurbantunggut Desert, Northern Xinjiang, China.
2
2007
... 已有的研究和我们前期的实验表明, 微生境的不同也会造成藓类植物生存策略的差异.不少野外调查和研究发现, 遮阴下(灌丛或阴坡)苔藓植物的盖度和生长状况好于暴露环境下的苔藓(Pintado et al ., 2005 ; Zhang et al ., 2007 ).灌丛下的低温、低光照和高含水量与裸露地的高温、高光照和低含水量造成了不同微生境间苔藓形态的变化.为了适应不同微生境, 苔藓植物能够随环境变化而变化, 但是短期微生境变化对苔藓形态的影响并不明显, 而其光合作用与现有生存环境相似(Reynolds & Mc- Letchie, 2011 ).Benassi等(2011)的研究发现: 不同生境下苔藓的繁殖方式也存在显著差异, 遮阴下的苔藓植株更趋于有性繁殖.然而, 不同微生境下苔藓植物对冻融过程中温度和水分变化的适应性是否一致尚不得而知. ...
... 实验地点位于新疆准噶尔盆地中部的古尔班通古特沙漠(44.18°-46.33° N, 80.52°-90.00° E, 海拔300-600 m).该沙漠面积4.88 × 104 km2 , 是中国最大的固定和半固定沙漠.受喜马拉雅山的阻隔作用, 来自印度洋西南季风的水汽不能到达内陆, 致使该沙漠属于典型的温带大陆性气候.沙漠年降水量仅70-150 mm, 年蒸发量达2606.6 mm, 年平均气温7.3 ℃, 冬季平均气温-16.8 ℃.与其他沙漠 不同, 该沙漠在冬季长期积雪覆盖, 稳定积雪日数一般在100-150天, 平均积雪厚度20-30 cm, 占全年降水的25%以上(Zhang et al ., 2007 ; 周宏飞等, 2010 ). ...
古尔班通古特沙漠不同地貌部位生物结皮的选择性分布
1
2004
... 齿肋赤藓属于丛藓科赤藓属, 具有良好的耐旱性和变水特性, 是古尔班通古特沙漠苔藓结皮的优势种.该种较山墙藓(Tortula ruralis )和北方赤藓(Syntrichia norvegica )更能适应极端干旱、低海拔和高温环境, 在世界冷沙漠中具有广泛分布(Reynolds & McLetchie, 2011 ).在干燥条件下, 其个体通常呈现为黑色或淡棕色, 复水后能快速变为绿色, 并在短时间内恢复光合和生理活性张静和张元明, 2014 ).齿肋赤藓种群的大量存在对沙漠的固定和微环境的改善具有重要作用(张元明等, 2004 ; 吴楠等, 2009 ). ...
古尔班通古特沙漠不同地貌部位生物结皮的选择性分布
1
2004
... 齿肋赤藓属于丛藓科赤藓属, 具有良好的耐旱性和变水特性, 是古尔班通古特沙漠苔藓结皮的优势种.该种较山墙藓(Tortula ruralis )和北方赤藓(Syntrichia norvegica )更能适应极端干旱、低海拔和高温环境, 在世界冷沙漠中具有广泛分布(Reynolds & McLetchie, 2011 ).在干燥条件下, 其个体通常呈现为黑色或淡棕色, 复水后能快速变为绿色, 并在短时间内恢复光合和生理活性张静和张元明, 2014 ).齿肋赤藓种群的大量存在对沙漠的固定和微环境的改善具有重要作用(张元明等, 2004 ; 吴楠等, 2009 ). ...
荒漠藓类结皮层中齿肋赤藓形态结构适应性及其原丝体发育特征
1
2009
... 荒漠苔藓作为典型的抗胁迫变水植物已被广泛关注, 诸多研究表明藓类植物能够适应极端高低温、干燥、强光照等恶劣环境(张静和张元明, 2014 ).为适应极端环境, 藓类植物在种群分布、形态特征(郑云普等, 2009 ; 陶冶和张元明, 2012 )、繁殖方式(Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 )、光合活性(张静和张元明, 2011 )、渗透调节物质、抗氧化酶(回嵘等, 2012 ; 陈文佳等, 2013 )等方面进化出多样的抗胁迫特性.研究发现齿肋赤藓(Syntrichia caninervis )叶片表面具有蜡质层, 并在细胞表面富集疣状突起, 在一定程度上能够减弱热气流的影响(许书军, 2007 ).另外, 在高温干燥环境下藓类植物能够通过失水降低代谢活性而进入休眠状态, 并积累大量的脯氨酸和可溶性糖来提高细胞渗透压, 维持渗透压平衡(Oliver et al ., 1997 ; Crowe et al ., 1998 ; 徐杰等, 2005 ; 吴楠等, 2009 ).同时, 通过抗氧化酶的大量合成清除过多的氧自由基, 进而保护植物免受过度伤害(Sgherri et al ., 1994 ; Herrnstadt & Kidron, 2005 ; Pintado et al ., 2005 ; 沈蕾等, 2011 ).但对荒漠藓类植物如何通过生理生化调节并成功度过冬季的极端低温环境, 还有待于深入研究. ...
荒漠藓类结皮层中齿肋赤藓形态结构适应性及其原丝体发育特征
1
2009
... 荒漠苔藓作为典型的抗胁迫变水植物已被广泛关注, 诸多研究表明藓类植物能够适应极端高低温、干燥、强光照等恶劣环境(张静和张元明, 2014 ).为适应极端环境, 藓类植物在种群分布、形态特征(郑云普等, 2009 ; 陶冶和张元明, 2012 )、繁殖方式(Herrnstadt & Kidron, 2005 ; Benassi et al ., 2011 )、光合活性(张静和张元明, 2011 )、渗透调节物质、抗氧化酶(回嵘等, 2012 ; 陈文佳等, 2013 )等方面进化出多样的抗胁迫特性.研究发现齿肋赤藓(Syntrichia caninervis )叶片表面具有蜡质层, 并在细胞表面富集疣状突起, 在一定程度上能够减弱热气流的影响(许书军, 2007 ).另外, 在高温干燥环境下藓类植物能够通过失水降低代谢活性而进入休眠状态, 并积累大量的脯氨酸和可溶性糖来提高细胞渗透压, 维持渗透压平衡(Oliver et al ., 1997 ; Crowe et al ., 1998 ; 徐杰等, 2005 ; 吴楠等, 2009 ).同时, 通过抗氧化酶的大量合成清除过多的氧自由基, 进而保护植物免受过度伤害(Sgherri et al ., 1994 ; Herrnstadt & Kidron, 2005 ; Pintado et al ., 2005 ; 沈蕾等, 2011 ).但对荒漠藓类植物如何通过生理生化调节并成功度过冬季的极端低温环境, 还有待于深入研究. ...
古尔班通古特沙漠植物雾凇凝结特征
1
2010
... 实验地点位于新疆准噶尔盆地中部的古尔班通古特沙漠(44.18°-46.33° N, 80.52°-90.00° E, 海拔300-600 m).该沙漠面积4.88 × 104 km2 , 是中国最大的固定和半固定沙漠.受喜马拉雅山的阻隔作用, 来自印度洋西南季风的水汽不能到达内陆, 致使该沙漠属于典型的温带大陆性气候.沙漠年降水量仅70-150 mm, 年蒸发量达2606.6 mm, 年平均气温7.3 ℃, 冬季平均气温-16.8 ℃.与其他沙漠 不同, 该沙漠在冬季长期积雪覆盖, 稳定积雪日数一般在100-150天, 平均积雪厚度20-30 cm, 占全年降水的25%以上(Zhang et al ., 2007 ; 周宏飞等, 2010 ). ...
古尔班通古特沙漠植物雾凇凝结特征
1
2010
... 实验地点位于新疆准噶尔盆地中部的古尔班通古特沙漠(44.18°-46.33° N, 80.52°-90.00° E, 海拔300-600 m).该沙漠面积4.88 × 104 km2 , 是中国最大的固定和半固定沙漠.受喜马拉雅山的阻隔作用, 来自印度洋西南季风的水汽不能到达内陆, 致使该沙漠属于典型的温带大陆性气候.沙漠年降水量仅70-150 mm, 年蒸发量达2606.6 mm, 年平均气温7.3 ℃, 冬季平均气温-16.8 ℃.与其他沙漠 不同, 该沙漠在冬季长期积雪覆盖, 稳定积雪日数一般在100-150天, 平均积雪厚度20-30 cm, 占全年降水的25%以上(Zhang et al ., 2007 ; 周宏飞等, 2010 ). ...
低温胁迫对茶树叶片SOD、可溶性蛋白和可溶性糖含量的影响
1
2011
... 冬季的极端低温环境极易使得植物体内的水分凝结形成冰晶, 并对植物造成严重伤害.而可溶性糖的大量积累能够增加细胞液的浓度, 降低细胞内水势, 使细胞液的冰点下降从而增加植物的抗寒能力.不少研究发现随着低温处理时间的延长, 植物体内的可溶性糖含量显著增加(沈蕾等, 2011 ; 朱政等, 2011 ).这在本实验中也得到证实, 在冬季低温冻结期植物体内的可溶性糖含量显著高于融雪期, 这说明齿肋赤藓在极端低温环境下能够通过积累大量可溶性糖来保护自身(Bhyan et al ., 2012 ).同时研究发现, 在积雪融化后的干燥环境下, 藓类植物体内脯氨酸和可溶性糖含量均较其他两个时期显著增加, 这可能是由于地表土壤含水量减少使得藓类植物处于干旱胁迫状态引起的.这也与张静和张元明(2014)在模拟降水对齿肋赤藓生理生化特性时发现的随含水量的减少齿肋赤藓体内可溶性糖和与脯氨酸呈递增趋势的结果相一致.此外, 徐杰等(2005)在不同区域真藓(Bryum argenteum )和土生对齿藓(Didymodon vinealis )的研究也表明, 植物体内脯氨酸与可溶性糖含量在降水量相对较少的草原化荒漠区显著高于降水量较多的典型草原. ...
低温胁迫对茶树叶片SOD、可溶性蛋白和可溶性糖含量的影响
1
2011
... 冬季的极端低温环境极易使得植物体内的水分凝结形成冰晶, 并对植物造成严重伤害.而可溶性糖的大量积累能够增加细胞液的浓度, 降低细胞内水势, 使细胞液的冰点下降从而增加植物的抗寒能力.不少研究发现随着低温处理时间的延长, 植物体内的可溶性糖含量显著增加(沈蕾等, 2011 ; 朱政等, 2011 ).这在本实验中也得到证实, 在冬季低温冻结期植物体内的可溶性糖含量显著高于融雪期, 这说明齿肋赤藓在极端低温环境下能够通过积累大量可溶性糖来保护自身(Bhyan et al ., 2012 ).同时研究发现, 在积雪融化后的干燥环境下, 藓类植物体内脯氨酸和可溶性糖含量均较其他两个时期显著增加, 这可能是由于地表土壤含水量减少使得藓类植物处于干旱胁迫状态引起的.这也与张静和张元明(2014)在模拟降水对齿肋赤藓生理生化特性时发现的随含水量的减少齿肋赤藓体内可溶性糖和与脯氨酸呈递增趋势的结果相一致.此外, 徐杰等(2005)在不同区域真藓(Bryum argenteum )和土生对齿藓(Didymodon vinealis )的研究也表明, 植物体内脯氨酸与可溶性糖含量在降水量相对较少的草原化荒漠区显著高于降水量较多的典型草原. ...




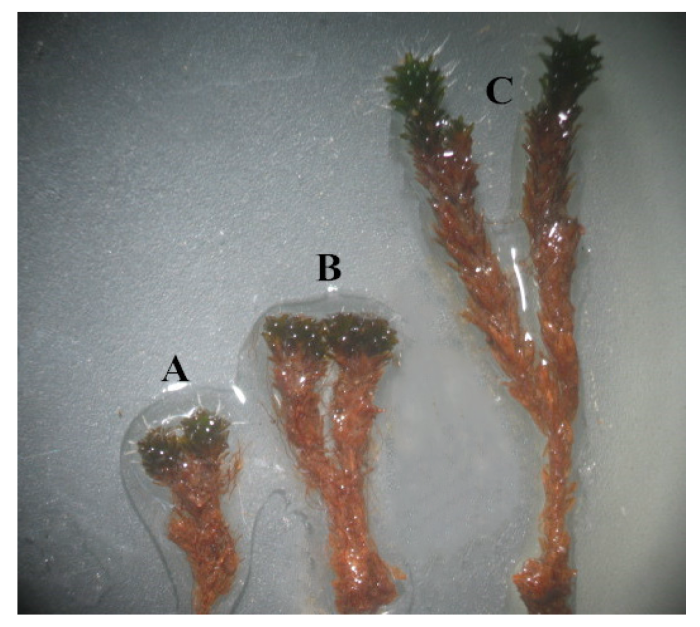
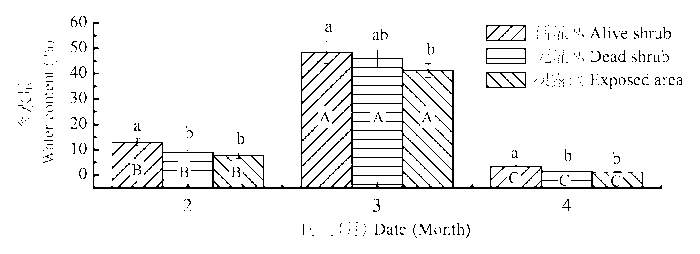
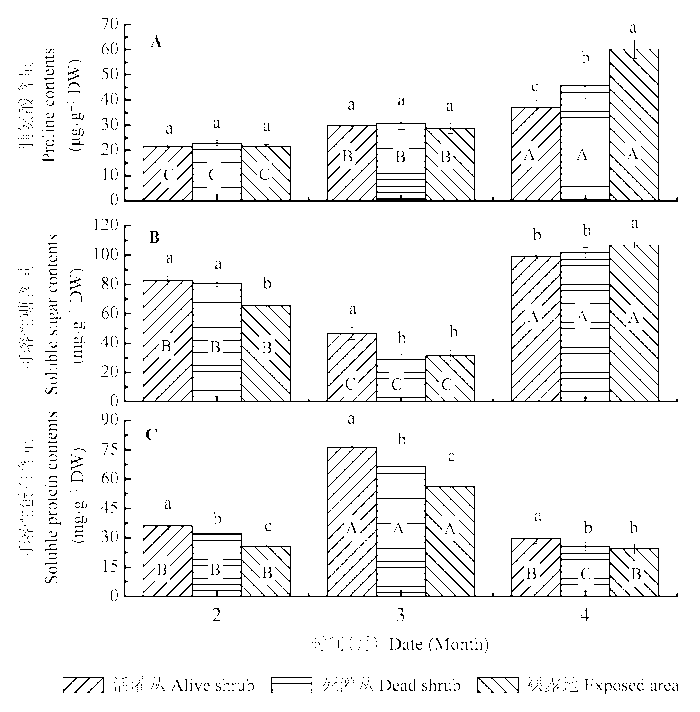
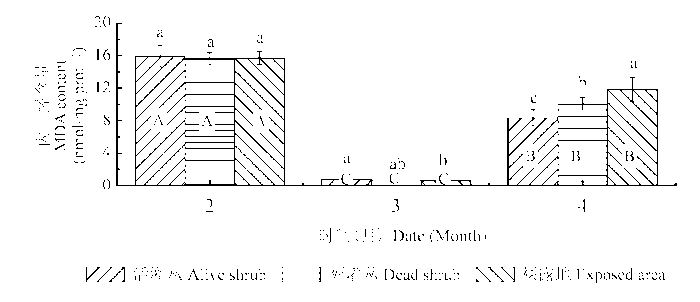
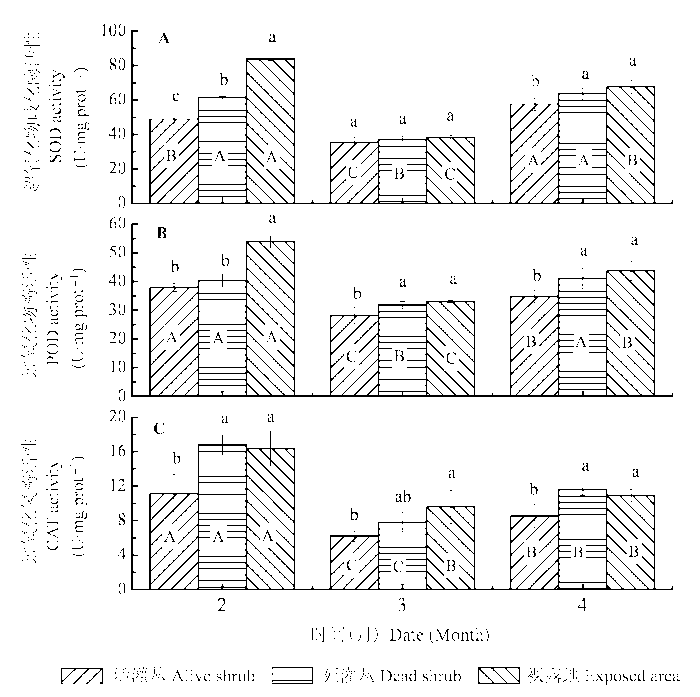
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}