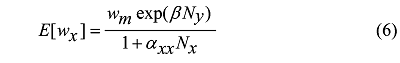竞争是陆生植物生长过程中的重要现象, 竞争作用是群落结构组建的主导因子和决定物种进化模式的重要因素(王刚和张大勇, 1996 )。对物种竞争的研究由来已久, 19世纪50年代末, 达尔文曾指出生活需求相似的近缘物种间经常发生激烈的竞争。随后, 洛特和沃尔特拉用数学公式描述了物种间的竞争关系。高斯假说从生态位角度展示了物种竞争的结果。竞争的研究领域涉及个体、种群、群落、生态系统等不同层次, 如竞争对个体大小不整齐性的影响(Weiner & Thomas, 1986 )、竞争对种群数量的调节(Deng et al ., 2006 )、竞争与群落空间分布格局的关系(Lin et al ., 2012 )、竞争在维持生态系统物种多样性中的作用(Borer et al ., 2014 )等等。随着植物间相互作用研究的不断拓展, 不断有新的概念和理论模型被提出, 甚至有的学者试图打破竞争与正相互作用的清晰界限(Warren et al ., 2014 )。在逐渐深入研究植物间相互作用的过程中, 植物地下根系间的竞争及其生态过程更受关注(Sutherland et al ., 2013 )。本文在前人研究的基础上, 从竞争模式角度着重对植物竞争(包括地上竞争和地下竞争)的对称性和非对称性进行综述和分析, 以期为国内相关研究提供参考和借鉴。
1 对称性竞争与非对称性竞争的概念内涵及其研究意义
迄今为止, 有关植物竞争的概念和类型有很多, 它们之间既有很多共性, 也存在很大差异。根据物种差异、地上地下和竞争模式, 植物竞争类型大致可分为: 种内竞争(intraspecific competition)和种间竞争(interspecific competition)、地上竞争(above- ground competition)和地下竞争(belowground competition) (Deng et al ., 2006 ; 王平等, 2007 )、对称性竞争(symmetric competition)和非对称性竞争(asymm- etric competition) (王刚和张大勇, 1996 )。对称性竞争又可再分为完全对称性竞争和大小依赖的对称性竞争; 非对称性竞争也可再分为完全非对称性竞争和大小依赖的非对称性竞争。种内竞争和种间竞争是基于植物物种的类型进行分类, 强调物种差异对竞争的影响; 而地上竞争和地下竞争的划分主要是基于植物体地上冠层对光资源的竞争和地下根系部分对水和营养元素的竞争, 打破了物种差异的明确界限。从某种意义上讲, 地上、地下竞争将植物竞争的研究范围进一步扩大了, 使得有关植物竞争的研究结论更具普适性。实际上, 对称性竞争与非对称性竞争同样涵盖了植物地上竞争和地下竞争, 但对称性竞争与非对称性竞争从概念上将植物竞争研究的内涵进一步深化, 体现了植物竞争的具体性质; 而地上竞争和地下竞争仅体现为植物竞争的形式。理论上, 地上竞争和地下竞争都可以是对称性或非对称性的; 反之, 植物的对称性竞争与非对称性竞争既可发生在植物的地上部分, 也可发生在植物的地下部分。然而, 对称性竞争与非对称性竞争更加强调了植物最大化获取资源的策略, 从一个新颖的角度阐释环境资源有限的情况下植物个体间分配资源的方式。近年来, 对称性竞争与非对称性竞争的研究逐渐成为热点, 下面将进一步阐释它们的概念及其内涵。
1.1 对称性竞争
大小依赖的对称性竞争是指存在资源竞争的植物间依据植物个体大小来分配资源, 即某株植物的生物量占全部植物总生物量的百分比就是其获得资源的百分比(Schwinning & Weiner, 1998 ; Weiner et al ., 2001 )。例如, 3株植物的生物量分别是B 1 、B 2 、B 3 , 总资源量为A, 则3株植物占有的资源量分别为: AB 1 /( B 1 + B 2 + B 3 )、AB 2 /( B 1 + B 2 + B 3 )、AB 3 /( B 1 + B 2 + B 3 )。植物体由幼苗生长至成熟个体的过程中, 可以表征个体大小的生物量逐渐增大, 如果其竞争模式为大小依赖的对称性竞争, 则其占有的资源量将逐渐增多, 竞争能力逐渐增强。完全对称性竞争是指植物在生长过程中竞争资源时, 根据特定区域内的植株个体数平均分配该区域内的资源Schwin- ning & Weiner, 1998 )。例如, 某一区域内有n 株植物, 则每株植物均分得该区域内资源总量的1/n 。
1.2 非对称性竞争
大小依赖的非对称性竞争是指处于竞争状态的植物间不均衡分配资源, 不按植物个体大小比例分配资源, 而是较大的植物占据大于相应生物量比例的资源, 较小的植物占据小于相应生物量比例的资源Schwinning & Weiner, 1998 )。例如, 两株植物生物量比例为1:3, 但资源分配可能是1:4或1:5。这种不均衡分配, 导致较大植物在下一阶段生长中占有更多资源, 而较小植物占有更少资源, 就像经济学中经常提到的“马太效应”一样, 强者愈强, 弱者愈弱, 最终致使较小植物因得不到足够的资源而死亡。这种具有“马太效应”的非对称性竞争的极端形式便是完全非对称性竞争, 即较大植物占有所有被竞争的资源(contested resource), 而较小植物不占有任何资源, 最终只能因资源枯竭而死亡和被淘汰。
1.3 对称性竞争与非对称性竞争的研究意义
有关植物竞争类型的理论研究一直是生态学领域的研究重点, 因为这是厘清植物邻体间竞争过程和揭示其竞争机制的必备环节。而且植物竞争类型也是在群落水平上研究植物群落结构组成、生态位分化和物种多样性及其稳定性维系机制的基础。竞争的对称性和非对称性问题同样会影响到一系列以此为基础的科学研究。比如, Hautier等(2009)在《Science》上发表文章认为, 立地环境的富营养化促进了植物对光非对称性竞争的发生, 进而导致物种多样性发生了变化。这主要是由于一些对光竞争处于劣势地位的物种得不到足够的光资源最终死亡。在开展植物对光非对称性竞争研究的同时, 植物根系非对称性竞争的研究也进行得如火如荼。近年来, 越来越多的研究证实在环境胁迫条件下植物根系竞争也为非对称性竞争Rajaniem, 2003 )。基于Hautier等(2009)的研究, 我们可以假设: 在以水分和营养元素为关键限制性资源的干旱荒漠生态系统中, 植物群落多样性也将由根系非对称性竞争所调控。而在植物生物量分配方面, 陆生植物体间的相互作用(竞争和促进)也会影响植物体地上和地下的生物量分配(Deng et al ., 2006 ; Chu et al ., 2008 ), 对植物对称性与非对称性竞争理论的深入研究或许能更清晰地揭示这种响应机理。至于植物个体大小不整齐性随种群密度的增加而增加这一现象(Weiner, 1986 ), 究竟是由于空间格局不一致引起植物密度变化而产生的Thomas & Weiner, 1989 ), 还是由于植物体的非对称性竞争产生的, 或者是植物本身具有较强的可塑性而导致的, 需要生态学家对其相关理论进一步深入研究。物种共存、生物入侵以及生物多样性等问题也与植物体间非对称性竞争有着千丝万缕的联系(Osunkoya et al ., 2013 ; Skácelová & Lepš, 2014 ), 研究非对称性竞争的产生机制和生理原理对开展相关工作具有重要意义。
2 对称性竞争与非对称性竞争相关研究进展
2.1 地上、地下竞争模式的提出
早期的竞争理论认为: 植物体地上部分主要竞争光资源, 属于非对称性竞争; 地下部分主要竞争水肥资源, 属于对称性竞争(Weiner, 1986 ; Weiner et al ., 1997 ; Schwinning & Weiner, 1998 ; Zhang et al ., 2013 )。其原理是: 高大植物凭借株高优势遮挡周围的矮小植物, 阻碍其获得充足的光照, 而矮小植物对高大植物的光吸收影响不大, 因此, 植物光竞争为非对称性竞争Weiner & Thomas, 1986 ; Connoily & Wayne, 1996 ); 由于地下水肥资源的流动特性, 使不同个体大小的植物均能得到资源, 因此植物水肥竞争为对称性竞争Bolte & Villanueva, 2006 )。但近年也有研究Rajaniem, 2003 ; Rewald & Leuschner, 2009 ; Lei et al ., 2012 ; Weremijewicz & Janos, 2013 )发现, 地下竞争为非对称性竞争或存在非对称性竞争倾向。比如, Rajaniemi (2003)研究发现, 植物在不同种群密度条件下, 目标植物周围存在不同大小的植物时, 较大的植物对目标植物单位生物量的影响大于较小植物对目标植物的影响, 并证明地下竞争可能为非对称性竞争。而Rewald和Leuschner (2009)以水青冈属(Fagus )和栎属(Quercus )为研究对象, 将野外生长的两树种的部分根系非破坏性挖出, 插进设计好的根生长室中, 使这部分根系发生种内、种间竞争作用, 并给予不同的水分梯度处理, 研究表明植物根系的非对称性竞争还会受到水分梯度的影响, 即植物间的非对称性竞争程度随水分胁迫梯度的增加而增加。植物根系间发生非对称性竞争现象的作用机理通常可以解释为: 生长于同一环境中的两株大小不同的植物, 较大的植物根系伸展范围广, 根系接触土壤中营养元素的表面积大, 获取的水肥资源多, 进而促进植物生长, 而进一步的生长又会使其根系伸展范围更广, 根系接触土壤中营养元素的表面积更大, 吸收消耗土壤中水分营养元素资源的能力更强。在土壤中资源有限的情况下, 大个体植物因其具有更强的吸收消耗能力而占有更多小个体根际周围的水肥资源, 进而导致小个体植物根际周围水肥资源的供应速率越来越低, 加剧了对小个体植物生长的限制。因此, 随着植物生长, 小个体与大个体间对土壤中有限资源的竞争能力和资源的占有量会随着植物的生长而进一步扩大, 最终同样可能导致小个体因资源的枯竭而死亡, 形成了典型的根系非对称性竞争。此外, 也有研究者认为, 在同等环境条件下, 植物个体仅仅通过对有限资源的抢先占有, 便能在竞争中胜出(Craine et al ., 2005 ), 也就是说, 植物个体对有限资源获取的起始时间不一致, 导致了对资源的非对称性竞争。
2.2 相关理论模型的建立及其检验方法
最常用的模型是由Schwinning和Fox (1995)基于植物对有限资源竞争的原理建立的对称性和非对称性竞争模型。Schwinning和Fox (1995)对对称性和非对称性竞争给出了明确的定义, 我们称之为“念模型”:
其中, r 是n 个植物体占有的总资源量, bi 代表种群中某植物个体i 的生物量, ri 代表植物个体i占有的资源量, 参数θ 的取值大小反映了竞争模式。当θ 取值为0或趋近于∞时, 植物间分别以完全对称性竞争和完全非对称性竞争方式分配资源; 当在区间(0, ∞)取值时, 则代表植物间竞争模式从对称性竞争向非对称性竞争的连续性过渡。该模型奠定了研究竞争模式的基础, 此后的许多实验或理论研究均运用此模型或以此模型为基础解决生态学竞争领域内的一些复杂问题(Chu et al ., 2008 , 2009, 2010 ; Lin et al ., 2012 )。但这一模型仅仅是对对称性和非对称性竞争概念的界定, 如用于实际研究, 则显得过于简单, 以至于会忽略掉很多影响植物间竞争模式的重要因子, 譬如物种特异性等。
上述概念模型的不足得到学者进一步的弥补和完善, Freckleton和Watkinson (2001)建立的有关植物竞争模式的模型将植物体间表现的竞争模式与植物体所处空间位置的概率和预期表现整合在一起, 该模型区分了个体间对称性竞争与非对称性竞争和物种间对称性竞争与非对称性竞争。Freckleton和Watkinson (2001)同时从个体水平和种群水平考虑, 根据不同条件下生长的植物建立了不同的生长方程。具体的研究方法是研究种群或群落中植物体密度-生物量关系的变化, 根据此关系的变化判断植物体属于何种竞争类型(对称性竞争和非对称性竞争)。
Freckleton和Watkinson (2001)认为, 在植物群落中既存在种内竞争也存在种间竞争的情况下, 单株目标植物体的预期平均生物量可用以下公式表示:
其中, E [wx ]代表在物种x 与物种y 相互作用的情况下物种x 目标个体的预期平均生物量; Nx 、Ny 分别表示物种x 和y 与目标植物进行相互作用的个体数; p (k )表示从0到Nx + Ny 个位置中, 目标植物占据k 位置的概率; wm 则为在无邻体竞争情况下单株植物体的预期生物量。这里f (Ax Nx ,Ay Ny |k )表示在物种x 和物种y 密度分别为Nx 和Ny 的情况下, 目标植物个体占据k 位置的几率下降的函数, 其中, f 值取决于两个邻体物种消耗资源的程度及邻体植物与目标植物间的竞争关系(种内竞争/种间竞争), Ax 和Ay 则分别表示目标植物与邻体植物物种x 或y 重叠区域占总区域的平均比例。
公式(2)阐释了植物体在个体水平上密度与其生物量的关系。Freckleton和Watkinson (2001)关于植物个体间和物种间对称性与非对称性的研究均以此方程为基础。下面我们将分别介绍Freckleton和Watkinson (2001)建立的有关植物个体和种间水平对称性竞争和非对称性竞争的理论模型。
2.2.1 个体间的对称性竞争
首先, 假设在植物群落中植物物种x 和物种y 的个体分布均为随机分布Firbank & Watkinson, 1985a ), 而且无论植物体处于什么空间位置, 其预期表现(即生态位和对资源获取的能力)都是相同的, 并且每株植物处于任何位置的概率也都是相同的, 物种x 处于k 处的概率px (k )与物种y 处于k 处的概率py (k )相等, 而且:
px (k ) = py (k ) = 1 / (1 + Nx + Ny )
fx (k ) = 1 / (1 + Ax Nx + Ay Ny )
因此, 竞争方程Freckleton & Watkinson, 2001 )是:
其中, E [wx ]代表在物种x 与物种y 相互作用情况下, 物种x 目标个体的预期平均生物量; wm 代表在无邻体竞争情况下单株植物体的预期生物量; Ax 和Ay 则分别表示目标植物与邻体植物物种x 或y 重叠区域占总区域的平均比例; Nx 、Ny 分别表示物种x 和y 与目标植物进行相互作用的个体数。由此推知, 在由物种x 和y 组成的群落中, 植物体的平均生物量与群落密度间的关系呈双曲线型的负相关关系, 即个体平均生物量随该植物邻体数量增加而下降。这与其他学者对平均个体大小与密度关系的研究结果Firbank & Watkinson, 1985b ; Deng et al ., 2006 , 2012a , 2012b )一致, 模型的正确性因此得到检验。这种情况下种群中的个体无论是物种x 还是物种y, 生物量随密度变化的趋势及变化速率均相同; 但是在自然群落中, 很难验证这种对称性竞争, 因为通常在不同的物种间生态位一般都发生了明显分化。上述模型(公式(3)), 同样可应用于植物种内的个体间竞争, 即N x 或N y 为0的情况下, 体现为种内的植物个体间竞争。
2.2.2 个体间非对称性竞争
竞争方程要考虑两点: 1)某一物种占据某一邻体位置的概率; 2)每个位置被邻体植株消耗的资源量。因此, 在位置i 处, 物种x 的资源消耗量就是物种x 占据该位置的概率(Nx px )与单株植物体所耗资源量(1 - Ax dx )的乘积Freckleton & Watkinson, 2001 ):
其中, dx 和dy 分别表示目标植物为物种x 和物种y 时占据的资源被邻体分得的比例。将此方程代入公式(2), 即可预测个体间非对称性竞争情况下, 植物个体平均生物量与群体密度的关系。个体间非对称性竞争方程可用于研究以光资源竞争为主的某一种群或群落个体平均生物量与群体密度间的异速比例关系; 反之, 也可根据该理论模型预测的生物量与密度间的关系检验群落中个体间是否存在非对称性竞争 现象。
2.2.3 物种间对称性竞争
在某一特定的植物群落中, 目标植物体占据任何位置的概率都是相同的, 这一点与个体间的对称性竞争情况相同, 所以在位置k 处有: px (k ) = py (k ) = 1 / (1+ Nx + Ny )。结合公式(2)和公式(4), 在Nx 和Ny 的极限值趋于无穷时, 有公式(5) Freckleton & Watkinson, 2001 ):
其中, αxx = Ax dx , αxy = Ay dy 。E [w ]代表在物种x 与物种y 相互作用情况下, 物种x 或物种y 目标个体的预期平均生物量; wm 为在无邻体竞争情况下单株植物体的预期生物量; Nx 、Ny 分别表示物种x 和y 与目标植物进行相互作用的个体数; Ax 和Ay 分别表示目标植物与邻体植物物种x 或y 重叠区域占总区域的平均比例; dx 和dy 分别表示目标植物为物种x 和物种y 时占据的资源被邻体分得的比例。
对比公式(3)和公式(5)可发现, 个体间和物种间的对称性竞争方程形式相同, 但是具体含义又有区别, 不同物种间存在对称性竞争时, 植物群落中平均个体生物量随密度增加呈指数下降, 其下降速度快于不同个体间发生对称性竞争的情况。
2.2.4 物种间非对称性竞争
物种x 与物种y 共存的群落体系中, 如果物种y 均能较物种x 抢先占领资源, 那么对于体系中处于位置k = 1, …, Ny 的植物来说, py (k ) = Ny -1 , px (k ) = 0, 同时有f (k ) = (1 - Ay dy )k k = 1 + Ny …1 + Nx + Ny , 有py (k ) = 0, px (k ) = 1 / (1 + Nx ), 结合公式(2)有:
其中, β = ln(1- Ay dy ), αxx = Ax dx 。E [wx ]代表在物种x 与物种y 相互作用的情况下物种x 目标个体的预期平均生物量; wm 为在无邻体竞争情况下单株植物体的预期生物量; Nx 、Ny 分别表示物种x 和y 与目标植物进行相互作用的个体数。Ax 和Ay 分别表示目标植物与邻体植物物种x 或y 重叠区域占总区域的平均比例; dx 和dy 分别表示目标植物为物种x 和物种y 时占据的资源被邻体分得的比例。
学者们除了通过目标植物生物量与邻体数量或密度间的变化关系来研究植物间竞争类型外, 还以个体影响域模型为基础, 通过数学建模和对植物个体生长过程的计算机模拟来研究植物间的对称性和非对称性竞争以及正相互作用强度等(Chu et al ., 2008 ), 并逐渐由植物体单层影响域模型向将植物体地上、地下部分区分开的双层影响域模型发展(Zhang et al ., 2013 )。关于运用个体影响域模型来解决对称性与非对称性竞争问题的研究, 未来可能更多关注于模型的进一步完善以及在建模过程中逐步将植物体根、茎、叶等器官区分开来。
检验植物体竞争对称性与非对称性的理论模型最主要和最直接的方法就是野外实验或控制实验。通常实验中具体的操作方法是测量生物量(Weiner, 1986 )、株高(Zhi et al ., 2007 )、冠幅(Iponga et al ., 2008 )、叶面积(Zhi et al ., 2007 )、根长(Reubens et al ., 2011 )、根表面积(Rewald & Leuschner, 2009 )等指标, 根据这些指标可以计算竞争强度(competition intensity)、竞争重要性(competition importance)和竞争反应(competition response)等参数(王平等, 2007 )。而这些参数的具体计算方法又有很大不同, 如根据计算竞争强度是否随生产力变化, 可将竞争强度分为相对竞争强度(relative competition intensity)和绝对竞争强度(absolute competition intensity) (Grace, 1995 ); 此外, 还可以根据植物地上和地下生长环境不同计算地上竞争强度、地下竞争强度以及总竞争强度, 实际运用中应根据实验设计和实验目的选择合适的参数, 各个参数的计算主要通过实施对照实验并分析植物体在有无竞争情况下的差异来进行。
生物量等测量指标能较好地解决植物对称性与非对称性问题, 但这些测量指标大多反映了植物个体大小的基本形态特征。而要揭示对称性竞争与非对称性竞争的生理生态机制, 除了通常以个体大小为衡量指标外, 或许还要将植物竞争对植物生长速率及其物质能量分配规律以及整个生命史和生活周期等方面的影响进行综合分析。比如在有植物竞争的情况下, 目标植物达到某一生长期的时间(例如开花期或成熟期)可能会延长。我们认为: 如果在竞争状态下目标植物的这种延长效应差距随植物生长逐渐增大, 类似这种植物间的竞争现象也应该归为非对称性竞争。
野外实验和实地考察有其客观性和可验证性, 但不确定因素也较多, 准确地研究对称性竞争和非对称性竞争效应较为困难(Firbank & Watkinson, 1987 ), 而模拟实验在研究精度、可操作性和避开不确定因素影响等方面具有明显优势。对相关模型的检验也可通过编程在计算机上进行模拟实验, 尤其是随着植物生长的动态竞争过程更适合通过模拟实验来实现。通常运用Netlogo软件(http://ccl.north- western.edu/netlogo)来实现植物竞争或生长动态等过程的模拟。该软件是由美国西北大学开发的一款能够对自然和社会现象进行仿真模拟的建模软件平台Wilensky, 1999 )。实际操作中, 可通过编程或根据系统动力学原理建立模型中各变量间的关系流程图来完成模拟过程。近年来发展起来的具有强大统计分析功能的R软件也可用于建模仿真和统计模拟, 具体模拟过程主要依据蒙特卡罗方法, 通过计算机产生的随机数(伪随机数)模拟离散随机变量或连续随机变量, 并将预解决的科学问题构建为合适的数学模型, 使模拟的变量正好是数学模型的解。
3 影响植物对称性竞争与非对称性竞争的主要因素
3.1 光照
尽管地下竞争通常大于地上竞争, 尤其是在营养元素贫瘠的土壤中Casper & Jackson, 1997 ; Kiær et al ., 2013 ), 但地上竞争也不可忽略。影响植物地上竞争的主要因素是光照, 绿色植物进行正常的光合作用需要充足的光照条件, 而在较为拥挤的植物种群或群落中, 植物个体间因相互遮阴而发生光竞争, 高大植物会遮挡其冠下植物对光的吸收, 但冠下植物对上层植物光吸收的影响较小。因此, 植物个体间对光的竞争是单方面的, 随着植物的生长, 高大植物的遮阴效果会越来越强, 形成典型的非对称性竞争, 即由于植物体个体大小不同(表现为株高不同)导致上层植物与冠下植物间对光的竞争为非对称性竞争van Leeuwen & Etienne, 2013 )。这种关于地上竞争的非对称性理论还仅局限于将光照作为一个笼统的概念进行整体分析。在未来的研究中可以对光资源进一步细化分析。
事实上, 光照可以通过光质、光强、光周期等对植物生长及竞争产生不同影响。不同的生长环境和植物生理特性可能会对光质、光强、光周期做出不同的响应, 最终改变植物间的竞争类型。例如, 高海拔地区的植物可能进化出适应紫外线的生长机制, 甚至有的物种只有当紫外线达到一定强度才能进行正常的光合作用, 这时植物间的竞争类型就可能随着高海拔地区臭氧空洞是否形成, 植物能否得到一定量的紫外线而发生改变, 即植物的对称性与非对称性竞争模式因光质不同而不同。喜阴植物和喜阳植物对光照强度具有明显不同的反馈作用机制, 其光合作用饱和点也明显不同, 所以可能出现这种情况: 某一光强条件下, 冠下喜阴植物拥有较充足的光资源, 而上层喜阳植物得到的光资源略显匮乏, 这时上层植物与下层植物对光的竞争可能就不表现为非对称性竞争关系, 因此若这些植物生长在不同光强条件下, 就不能单凭植物冠层高低来衡量个体间是否为非对称性竞争; 此外, 光周期的长短能调控植物开花的时间, 植物开花即对生殖器官的投资会影响植物体营养部分的生长速度或动态, 植物体内不同器官间如何分配光合作用获得的能量进而改变个体间的相对大小等性状特征, 也可能会对植物竞争类型产生重要影响。
3.2 土壤异质性
土壤营养异质性会加剧土壤斑块中植物间的竞争程度(Rajaniemi, 2011 )。Rajaniemi通过研究无芒雀麦(Bromus inermis )发现了异质性土壤中生长的植物根系具有非对称性竞争现象(Rajaniemi, 2003 ); Facelli和Facelli (2002)通过研究三叶草属植物Trifolium subterraneum 发现了类似现象。土壤异质性在多数情况下表现为土壤肥力不均匀, 肥力较丰富区域中的植物生长较快, 而周围能与其发生相互作用的生长在肥力较贫瘠土壤中的植物生长较慢, 两者间的植物个体大小差异随生长过程逐渐凸显, 当因密度效应发生地上光竞争时, 土壤肥力丰富地区的植物占据优势地位, 地上和地下同时存在的优势使得肥力丰富地区与贫瘠地区的植物逐渐形成非对称性竞争关系。
如上所述, 研究生物体间的非对称性竞争问题可以与生物的共存、生物多样性、生物入侵等问题相联系。在一定的条件下, 生物多样性降低很可能是由物种间非对称性竞争而造成的(Šmilauer & Šmilauerová, 2013 )。在相同生境中两物种间出现完全的非对称性竞争时, 一物种完全处于竞争优势地位, 而另一物种不占据任何资源优势, 在竞争中逐渐衰败, 最终死亡。因此, 当物种间出现非对称性竞争时, 物种无法共存, 它们所处的生态系统生物多样性会下降, 生物入侵容易发生。而这几个方面恰恰与土壤异质性有关, 或者说土壤异质性可以部分解释物种共存、生物多样性和生物入侵等问题。首先, 土壤异质性与物种入侵间可以发生相互作用和反馈调节。已有证据表明, 外来物种入侵会显著改变生态系统N循环, 入侵物种增加了N含量, 而且加快了N的流动循环Castro-Díez et al ., 2014 )。因此, 随着物种入侵, 土壤中营养元素的成分和含量会发生大的变化, 致使土壤异质性加强。同时, 物种入侵也会受土壤异质性影响。Burns和Brandt (2014)研究发现, 土壤异质性与种群周转率相互作用影响物种入侵, 增加土壤异质性会增加物种入侵。两者间的相互作用关系启示我们在研究物种间非对称性竞争问题时应与生物入侵问题相联系。其次, 土壤异质性会影响生物多样性, 多数情况下土壤异质性会增加生物多样性(Belfrage et al ., 2015 )。从生态位角度考虑, 土壤异质性可以为物种提供更多的生态位, 适应不同物种对不同生境的需求, 而且不同的生态位缓解了物种间对各种资源的强烈竞争, 使生物出现共存局面, 最终可能表现为植物间较弱的非对称性竞争或转变为对称性竞争。但是, 近来研究发现, 土壤异质性高并不意味着生物多样性也升高, 例如当土壤中某种元素含量很高而出现富营养化时, 情况会大不相同。已有研究表明土壤富营养化会削弱生物多样性产生的稳定性效应(Hautier et al ., 2014 )。也就是说, 物种稳定共存的局面不会长期维持下去, 总有一些物种会在竞争中被淘汰掉。因此, 土壤异质性升高、物种多样性降低的环境中, 物种间易于出现非对称性竞争。
3.3 土壤微生物
一些研究发现, 群落中的优势物种往往是那些能够高效利用环境中限制资源的物种(Fargione & Tilman, 2006 )。菌根中的微生物可以使一些植物得到充足的必需元素, 例如豆科植物中的根瘤菌为豆科植物提供充足的氮源, 促使其快速生长。因此, 这类微生物提升了植物根系在土壤中的竞争能力。最终, 豆科植物越长越大, 根系越来越发达, 需要和吸收的营养元素也越来越多(这些元素包括P、K 等必需元素, 而不仅仅是N), 留给供非豆科植物生长的元素就越来越少, 因此抑制非豆科植物生长的程度越来越大, 形成了物种间根系非对称性竞争。土壤中的微生物还有致病菌, 它们会导致相反的结果。一些植物所携带的细菌、真菌等微生物会对其他植物造成伤害, 尤其是一些入侵物种携带的土壤微生物会限制当地物种利用土壤中的资源, 破坏其根系生长, 严重危及当地物种的正常生长; 或者土壤微生物对植物地下生长的调节影响入侵物种间的相互作用关系(Wolfe & Klironomos, 2005 )。土壤中不同的微生物群落以及根系周围不同的菌群成分会对植物间根系竞争产生不同的影响, 不同的微生物通过增加或降低根际周围营养物质的可利用性, 直接影响竞争根系对营养物质的利用效率, 使得仅有一些植物得到较多资源, 最终不同植物因适合度不同而呈现出非对称性竞争趋势(Yang et al ., 2014 )。土壤微生物还可能通过与土壤中营养元素的相互作用来影响物种间的相互作用关系。研究表明, 在土壤中添加大量P元素会降低生物多样性, 即部分物种因不能共存而被淘汰, 而植物根际周围菌群的存在能够增加生物多样性, 因此豆科植物可以依赖于自身拥有的根瘤菌生存于富营养化的土壤中(Yang et al ., 2014 )。调节土壤中的微生物含量和种类, 同时调整土壤中营养元素的含量和种类, 将两种因素进行排列组合, 或许可以找到特定环境, 适合某种植物生存而不利于另一种植物生存, 若使两种植物相邻生长, 物种间非对称性竞争较易体现。
3.4 毒素和信号物质
研究发现一些植物能够通过化感作用产生有毒物质, 限制周围植物生长甚至致其死亡(Schenk, 2006 )。通常不同物种间会出现这种情况, 一种植物根系分泌物会影响另一种植物生长。但也有一些物种成年植物体释放化学物质影响和抑制其幼苗个体生长, 例如银桦(Grevillea robusta )。除此之外, 一些植物根系还会释放一种对其他植物无害的化学物质, 帮助植物感知周围其他植物根系的生长及土壤中营养物质等非生命物质的存在。研究发现, 一些豆科植物根系能通过“化学雷达”而不用物理接触就能感知其周围的物理障碍, 从而改变根的生长形态(Falik et al ., 2005 )。资源获取器官若能够感知非生命物质的存在, 在生长中就占据很大优势。例如, 植物根系若能感知土壤中肥力不足的区域或不利于自身生长的因素, 就不朝肥力不足区域生长而转向肥力丰富区域生长或避开不利于自身生长的因素, 从而保证植物体快速生长, 增强其在逆境中的生存竞争能力。
一系列对草本和灌木物种的研究(Mahall & Callaway, 1996 ; de Kroon et al., 2003 ; Falik et al ., 2003 ; Holzapfel & Alpert, 2003 ; Gruntman & Novoplansky, 2004 )发现, 植物的根系能够通过信号感知并且区分“自己”的和“非己”的根系。这对植物的生长是有利的, 同境中生长的相邻两株植物, 每株植物可以通过化学信号感知周围另一株植物根系的存在, 并且抑制自己根系向对方根系生长所占区域延伸, 尽量避免自己受到伤害。
植物体的这一切通过毒素和信号物质提高自身竞争能力的生存方式和策略, 不仅能有效地保证其在较恶劣环境中存活与生长, 而且会因为某一种群或群落中目标植物与邻体植物竞争能力差异的改变而导致个体间竞争类型发生改变(Mariotte et al., 2012 )。例如, 均产生毒素的目标植物与邻体植物, 如果两者产生的毒素对对方的影响效应基本相同, 那么目标植物与邻体植物间可能因竞争能力相同而形成对称性竞争的竞争模式; 若两者产生的毒素对对方影响效应显著不同, 则可能形成非对称性竞争的竞争模式。
3.5 植物形态可塑性
植物的形态可塑性常常通过改变植物地上和地下的生物量分配、增大某部分器官的表面积等途径来影响植物间的竞争关系(Schwinning & Weiner, 1998 )。当土壤中水肥资源受限时, 植物就竭力分配生物量给地下(通常干旱区植物有发达的根系), 同时增大根系与土壤接触的表面积, 竭力使植物体在水肥资源竞争中占优势。所以当两株相邻植物获取资源的能力相差不大时, 处于劣势的植物很可能通过形态可塑性, 分配更多生物量给地下, 藉此来弥补根系所处的劣势。但当两株植物的相对优势很明显时, 形态可塑性无法弥补劣势植物的劣势, 致使其无法得到足够的资源, 也就无法快速生长, 陷入了恶性循环, 而优势植物由于获得充足的资源, 可以进一步生长, 根系更加强大, 获取资源能力更强, 形成了良性循环, 两株植物间根系的非对称性竞争模式就形成了。形态可塑性在减弱植物个体间非对称性竞争中的作用已经得到一系列转基因实验的证明(Ballaré et al ., 1994 ; Ballaré & Scopel, 1997 )。同样, 形态可塑性也会影响植物体地上部分对光资源的竞争。研究发现, 植物可塑性和时空上的生态位分化能够提高植物获取光的能力并维持生物多样性的积极影响(Sapijanskas et al ., 2014 )。生物多样性局面的维持表明物种间可以共存, 植物可塑性等因素使得同生境中生长的为光资源而发生激烈非对称性竞争的植物能够实现共存。
4 前景展望
植物竞争类型的研究方兴未艾, 并有繁荣发展的势头, 尤其是关于植物地下竞争类型的研究, 更是发展迅速, 有研究认为地下竞争与物种多样性密切相关(Rajaniemi et al ., 2003 )。过去由于探测和测量仪器的局限性, 人工挖掘又会导致部分根系损失, 所以很难准确得到植物地下部分的相关指标和参数, 开展地下竞争的实验和研究也相对较少。但是近年来, 随着科技水平的提升以及一批根系测量与分析系统的完善和发展, 不仅可以准确地测得根系的长度、直径、表面积, 还可以进行根尖计数, 分叉和交叠计数, 使得开展地下竞争研究越来越容易。植物体的地下世界逐渐被发掘, 地下竞争关系越来越清晰。地下竞争或许比地上竞争更加重要, 因为植物体地上部分所需的主要资源是取之不尽用之不竭的光能, 而深入开展植物体地下部分的竞争研究, 尤其是地下部分的非对称性竞争研究, 能帮助人们更加明确对植物体的水肥投入的重要性, 节约营养资源, 增大投入产出效益; 同时也会使人们懂得如何更好地维护整个生态系统, 拯救濒临灭绝的物种, 保证生物圈的物种多样性。
CO2 和光照间的相互作用如何影响植物对称性与非对称性竞争的综合表现尚不明确。分析植物地上竞争时, 一般只考虑植物的光竞争。但是植物进行光合作用时, 不仅需要光照条件, 还需要CO2 作为原料, 植物生长所处环境中CO2 的浓度或许会显著影响植物竞争类型。假设植物地上对光的竞争为非对称性竞争, 即高大植物生物量增量越来越大于矮小植物, 则高大植物同化作用减去异化作用而获得的净生物量会显著大于矮小植物, 而植物生长及进行光合作用的碳源主要来自空气中的CO2 , 所以高大植物消耗的净CO2 量要显著高于矮小植物。然而, 从物理学角度得知, 大气中CO2 的质量和密度相对较大, 往往高处的CO2 浓度要低于低处或地表。因此, 高大植物叶片周围的CO2 浓度可能会显著低于矮小植物周围的CO2 浓度。这就在一定程度上弥补了矮小植物在对光资源的非对称性竞争中所处的劣势地位。因此, 综合考虑这两种影响因素以及它们之间的相互作用, 或许可以发现植物对称性与非对称性竞争的新的响应机制。例如, 通过改变这两种相对竞争优势的强度, 矮小植物获得的足量CO2 可能会弥补其在光资源获取上的劣势地位, 最终减弱植物间非对称性竞争的程度。
植物对称性与非对称性竞争如何影响地上地下生物量分配格局也是值得进一步研究的课题。植物体地上和地下生物量分配模式在一定意义上代表了植物对环境资源的拥有量(包括水、营养元素、光和CO2 等资源)的响应, 而植物对称性与非对称性竞争强调了植物体间对占有的资源如何划分, 它们之间密切的相关关系预示着植物竞争类型可能强烈地影响地上和地下生物量分配格局, 在建立植物地上和地下单独的生长模型时, 可以将对称性与非对称性竞争因素考虑在内, 通过计算机模拟实验分别分析植物地上和地下的生长曲线以及生物量分配模式。
另外, 竞争类型渐变发生机制也是一个重要的研究方向。目前, 关于植物体竞争类型的研究主要集中在大小依赖的对称性和非对称性竞争以及完全对称性和非对称性竞争, 而对这几种竞争类型间相互转化和渐变的机制研究较少, 未来在研究竞争类型时可以将进化因素考虑进来, 将植物间的对称性与非对称性竞争与生物进化机制有机结合, 这样或许可以揭示植物体间的相互作用如何影响植物进化历程以及竞争与进化间是如何相互作用的。例如, 生长在恶劣环境中的植物为了获得生存和发展,逐渐进化出抗寒、抗旱、耐盐等适应机制, 在这一过程中或许植物的竞争类型也会发生改变, 即从对称性竞争向非对称性竞争渐变, 或者从非对称性竞争向对称性竞争渐变, 植物体处于何种环境能够迫使植物竞争类型发生渐变, 值得深入研究。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Ballaré CL Scopel AL 1997 ). Phytochrome signalling in plant canopies: Testing its population-level implications with photoreceptor mutants of Arabidopsis.
Functional Ecology , 11 , 441 -450 .
[本文引用: 1]
[2]
Ballaré CL Scopel AL Jordan ET Vierstra RD 1994 ). Signaling among neighboring plants and the development of size inequalities in plant populations.
Proceedings of the National Academy of Sciences of the United States of America , 91 , 10094 -10098 .
[本文引用: 1]
[3]
Belfrage K Björklund J Salomonsson L 2015 ). Effects of farm size and on-farm landscape heterogeneity on biodiversity-case study of twelve farms in a Swedish landscape.
Agroecology and Sustainable Food Systems , 39 , 170 -188 .
[本文引用: 1]
[4]
Bolte A Villanueva I 2006 ). Interspecific competition impacts on the morphology and distribution of fine roots in European beech (Fagus sylvatica L.) and Norway spruce (Picea abies (L.) Karst.).
European Journal of Forest Research , 125 , 15 -26 .
[本文引用: 1]
[5]
Borer ET Seabloom EW Gruner DS Harpole WS Hillebrand H Lind EM Adler PB Alberti J Anderson TM Bakker JD Biederman L Blumenthal D Brown CS Brudvig LA Buckley YM Cadotte M Chu CJ Cleland EE Crawley MJ Daleo P Damschen EI Davies KF DeCrappeo NM Du GZ Firn J Hautier Y Heckman RW Hector A Iribarne O Klein JA Knops JMH Pierre KJL Leakey ADB Li W MacDougall AS McCulley RL Melbourne BA Mitchell CE Moore JL Mortensen B O’Halloran LR Orrock JL Pascual J Prober SM Pyke DA Risch AC Schuetz M Smith MD Stevens CJ Sullivan LL Williams RJ Wragg PD Wright JP Yang LH 2014 ). Herbivores and nutrients control grassland plant diversity via light limitation.
Nature , 508 , 517 -520 .
[本文引用: 1]
[6]
Burns JH Brandt AJ 2014 ). Heterogeneity in plant-soil feedbacks and resident population dynamics affect mutual invasibility.
Journal of Ecology , 102 , 1048 -1057 .
[7]
Casper BB Jackson RB 1997 ). Plant competition underground.
Annual Review of Ecology and Systematics , 28 , 545 -570 .
[本文引用: 1]
[8]
Castro-Díez P Godoy O Alonso A Gallardo A Saldana A 2014 ). What explains variation in the impacts of exotic plant invasions on the nitrogen cycle? A meta-analysis.
Ecology Letters , 17 , 1 -12 .
[本文引用: 1]
[9]
Chu CJ Maestre FT Xiao S Weiner J Wang YS Zheng HD Wang G 2008 ). Balance between facilitation and resource competition determines biomass-density relationships in plant populations.
Ecology Letters , 11 , 1189 -1197 .
[本文引用: 3]
[10]
Chu CJ Wang YS Li Q Zhao LQ Ren ZW Xiao S Yuan JL Wang G 2009 ). Effects of traits, species identity and local environmental conditions on the assessment of interactions: Insights from an alpine meadow community.
Journal of Plant Ecology , 23 , 135 -141 .
[本文引用: 1]
[11]
Chu CJ Weiner J Maestre FT Wang YS Morris C Xiao S Yuan JL Du GZ Wang G 2010 ). Effects of positive interactions, size symmetry of competition and abiotic stress on self-thinning in simulated plant populations.
Annals of Botany , 106 , 647 -652 .
[12]
Connoily J Wayne P 1996 ). Asymmetric competition between plant species.
Oecologia , 108 , 311 -320 .
[本文引用: 1]
[13]
Craine JM Fargione J Sugita S 2005 ). Supply pre-emption, not concentration reduction, is the mechanism of competition for nutrients.
New Phytologist , 166 , 933 -940 .
[本文引用: 1]
[14]
de Kroon H Mommer L Nishiwaki A 2003 ). Root competition: Towards a mechanistic understanding.
Root Ecology , 168 , 215 -234 .
[本文引用: 1]
[15]
Deng JM Wang GX Morris EC Wei XP Li DX Chen BM Zhao CM Liu J Wang Y 2006 ). Plant mass-density relationship along a moisture gradient in north-west China.
Journal of Ecology , 94 , 953 -958 .
[本文引用: 4]
[16]
Deng JM Ran JZ Wang ZQ Fan ZX Wang GX Ji MF Liu J Wang Y Liu JQ Brown JH 2012a ). Models and tests of optimal density and maximal yield for crop plants.
Proceedings of the National Academy of Sciences of the United States of America ,109 , 15823 -15828 .
[本文引用: 1]
[17]
Deng JM Zuo WY Wang ZQ Fan ZX Ji MF Wang GX Ran JZ Zhao CM Liu JQ Niklas KJ Hammond ST Brown JH 2012b ). Insights into plant size-density relationships from models and agricultural crops.
Proceedings of the National Academy of Sciences of the United States of America ,109 , 8600 -8605 .
[本文引用: 1]
[18]
Facelli E Facelli JM 2002 ). Soil phosphorus heterogeneity and mycorrhizal symbiosis regulate plant intra-specific competition and size distribution.
Oecologia , 133 , 54 -61 .
[19]
Falik O Reides P Gersani M Novoplansky A 2003 ). Self/non-self discrimination in roots.
Journal of Ecology , 91 , 525 -531 .
[本文引用: 1]
[20]
Falik O Reides P Gersani M Novoplansky A 2005 ). Root navigation by self inhibition.
Plant, Cell & Environment , 28 , 562 -569 .
[本文引用: 1]
[21]
Fargione J Tilman D 2006 ). Plant species traits and capacity for resource reduction predict yield and abundance under competition in nitrogen-limited grassland.
Functional Ecology , 20 , 533 -540 .
[本文引用: 1]
[22]
Firbank LG Watkinson AR 1985a ). A model of interference within plant monocultures.
Journal of Theoretical Biology , 116 , 291 -311 .
[本文引用: 1]
[23]
Firbank LG Watkinson AR 1985b ). On the analysis of competition within two-species mixtures of plants.
Journal of Applied Ecology , 22 , 503 -517 .
[本文引用: 1]
[24]
Firbank LG Watkinson AR 1987 ). On the analysis of competition at the level of the individual plant.
Oecologia , 71 , 308 -317 .
[本文引用: 1]
[25]
Freckleton RP Watkinson AR 2001 ). Asymmetric competition between plant species.
Functional Ecology , 15 , 615 -623 .
[本文引用: 3]
[26]
Grace JB 1995 ). On the measurement of plant competition intensity.
Ecology , 76 , 305 -308 .
[本文引用: 1]
[27]
Gruntman M Novoplansky A 2004 ). Physiologically mediated self/non-self discrimination in roots.
Proceedings of the National Academy of Sciences of the United States of America , 101 , 3863 -3867 .
[本文引用: 1]
[28]
Hautier Y Niklaus PA Hector A 2009 ). Competition for light causes plant biodiversity loss after eutrophication.
Science , 324 , 636 -638 .
[29]
Hautier Y Seabloom EW Borer ET Adler PB Harpole WS Hillebrand H Lind EM MacDougall AS Stevens CJ Bakker JD Buckley YM Chu CJ Collins SL Daleo P Damschen EI Davies KF Fay PA Firn J Gruner DS Jin VL Klein JA Knops JMH La Pierre KJ Li W McCulley RL Melbourne BA Moore JL O’Halloran LR Prober SM Risch AC Sankaran M Schuetz M Hector A 2014 ). Eutrophication weakens stabilizing effects of diversity in natural grasslands.
Nature , 508 , 521 -525 .
[本文引用: 1]
[30]
Holzapfel C Alpert P 2003 ). Root cooperation in a clonal plant: Connected strawberries segregate roots.
Oecologia , 134 , 72 -77 .
[本文引用: 1]
[31]
Iponga DM Milton SJ Richardson DM 2008 ). Superiority in competition for light: A crucial attribute defining the impact of the invasive alien tree Schinus molle (Anacardiaceae) in South African Savanna.
Journal of Arid Environments , 72 , 612 -623 .
[本文引用: 1]
[32]
Kiær LP Weisbach AN Weiner J 2013 ). Root and shoot competition: A meta-analysis.
Journal of Ecology , 101 , 1298 -1312 .
[本文引用: 1]
[33]
Lei PF Scherer-Lorenzen M Bauhus J 2012 ). Belowground facilitation and competition in young tree species mixtures.
Forest Ecology and Management , 265 , 191 -200 .
[本文引用: 1]
[34]
Lin Y Berger U Grimm V Ji QR 2012 ). Differences between symmetric and asymmetric facilitation matter: Exploring the interplay between modes of positive and negative plant interactions.
Journal of Ecology , 100 , 1482 -1491 .
[本文引用: 2]
[35]
Mahall BE Callaway RM 1996 ). Effects of regional origin and genotype on intraspecific root communication in the desert shrub Ambrosia dumosa (Asteraceae).
American Journal of Botany , 83 , 93 -98 .
[本文引用: 1]
[36]
Mariotte P Buttler A Johnson D Thébault A Vandenberghe C 2012 ). Exclusion of root competition increases competitive abilities of subordinate plant species through root-shoot interactions.
Journal of Vegetation Science , 23 , 1148 -1158 .
[本文引用: 1]
[37]
Osunkoya OO Perrett C Fernando C Clark C 2013 ). Patterns of seed bank and size asymmetry of plant growth across varying sites in the invasive Lantana camara L. (Verbenaceae).
Plant Ecology , 214 , 725 -736 .
[本文引用: 1]
[38]
Rajaniemi TK 2003 ). Evidence for size asymmetry of belowground competition.
Basic and Applied Ecology , 4 , 239 -247 .
[本文引用: 3]
[39]
Rajaniemi TK 2011 ). Competition for patchy soil resources reduces community evenness.
Oecologia , 165 , 169 -174 .
[本文引用: 1]
[40]
Rajaniemi TK Allison VJ Goldberg DE 2003 ). Root competition can cause a decline in diversity with increased productivity.
Journal of Ecology , 91 , 407 -416 .
[本文引用: 1]
[41]
Reubens B Achten WMJ Maes WH Danjon F Aerts R Poesen J Muys B 2011 ). More than biofuel? Jatropha curcas root system symmetry and potential for soil erosion control.
Journal of Arid Environments , 75 , 201 -205 .
[本文引用: 1]
[42]
Rewald B Leuschner C 2009 ). Does root competition asymmetry increase with water availability?
Plant Ecology & Diversity , 2 , 255 -264 .
[本文引用: 2]
[43]
Sapijanskas J Paquette A Potvin C Kunert N Loreau M 2014 ). Tropical tree diversity enhances light capture through crown plasticity and spatial and temporal niche differences.
Ecology , 95 , 2479 -2492 .
[本文引用: 1]
[44]
Schenk JH 2006 ). Root competition: Beyond resource depletion.
Journal of Ecology , 94 , 725 -739 .
[本文引用: 1]
[45]
Schwinning S Fox GA 1995 ). Population dynamic consequences of competitive symmetry in annual plants.
Oikos , 72 , 422 -432 .
[46]
Schwinning S Weiner J 1998 ). Mechanisms determining the degree of size asymmetry in competition among plants.
Oecologia , 113 , 447 -455 .
[本文引用: 5]
[47]
Skácelová O Lepš J 2014 ). The relationship of diversity and biomass in phytoplankton communities weakens when accounting for species proportions.
Hydrobiologia , 724 , 67 -77 .
[本文引用: 1]
[48]
Šmilauer P Šmilauerová M 2013 ). Asymmetric relationship between grasses and forbs: Results from a field experiment under nutrient limitation.
Grass and Forage Science , 68 , 186 -198 .
[本文引用: 1]
[49]
Sutherland WJ Freckleton RP Godfray HCJ Beissinger SR Benton T Cameron DD Carmel Y Coomes DA Coulson T Emmerson MC Hails RS Hays GC Hodgson DJ Hutchings MJ Johnson D Jones JPG Keeling MJ Kokko H Kunin WE Lambin X Lewis OT Malhi Y Mieszkowska N Milner-Gulland EJ Norris K Phillimore AB Purves DW Reid JM Reuman DC Thompson K Travis JMJ Turnbull LA Wardle DA Wiegand T 2013 ). Identification of 100 fundamental ecological questions.
Journal of Ecology , 101 , 58 -67 .
[本文引用: 1]
[50]
Thomas EM Weiner J 1989 ). Local density variation may mimic effects of asymmetric competition on plant size variability.
Ecology , 70 , 1188 -1191 .
[本文引用: 1]
[51]
van Leeuwen E Etienne RS 2013 ). Caught in the middle: Asymmetric competition causes high variance in intermediate trait abundances.
Theoretical Population Biology , 85 , 26 -37 .
[本文引用: 1]
[52]
Wang G Zhang DY 1996 ). Theory of Biology Competition . Shaanxi Science and Technology Press , Xi’an (in Chinese)
[本文引用: 2]
[王刚 , 张大勇 (1996 ). 生物竞争理论 . 陕西科学技术出版社 , 西安 .]
[本文引用: 2]
[53]
Wang P Wang TH Zhou DW Zhang HX 2007 ). A literature review on the above- and below-ground competition.
Acta Ecologica Sinica , 27 , 3489 -3499 .(in Chinese with English abstract)
[本文引用: 2]
[王平 , 王天慧 , 周道玮 , 张红香 (2007 ). 植物地上竞争与地下竞争研究进展
. 生态学报 , 27 , 3489 -3499 .]
[本文引用: 2]
[54]
Warren RJ Giladi I Bradford MA 2014 ). Competition as a mechanism structuring mutualisms.
Journal of Ecology , 102 , 486 -495 .
[本文引用: 1]
[55]
Weiner J 1986 ). How competition for light and nutrients affects size variability in ipomoea tricolor populations.
Ecology , 67 , 1425 -1427 .
[本文引用: 3]
[56]
Weiner J Wright DB Castro S 1997 ). Symmetry of below- ground competition between Kochia scoparia individuals.
Oikos , 79 , 85 -91 .
[本文引用: 1]
[57]
Weiner J Griepentrog HW Kristensen L 2001 ). Suppression of weeds by spring wheat Triticum aestivum increases with crop density and spatial uniformity.
Journal of Applied Ecology , 38 , 784 -790 .
[本文引用: 1]
[58]
Weiner J Thomas SC 1986 ). Size variability and competition in plant monocultures.
Oikos , 47 , 211 -222 .
[本文引用: 2]
[59]
Weremijewicz J Janos DP 2013 ). Common mycorrhizal networks amplify size inequality in Andropogon gerardii monocultures.
New Phytologist , 198 , 203 -213 .
[本文引用: 1]
[60]
Wilensky U 2014 -11-10.
URL
[本文引用: 1]
[61]
Wolfe BE Klironomos JN 2005 ). Breaking new ground: Soil communities and exotic plant invasion.
BioScience , 55 , 477 -487 .
[本文引用: 3]
[62]
Yang GW Liu N Lu WJ Wang S Kan HM Zhang YJ Xu L Chen YL 2014 ). The interaction between arbuscular mycorrhizal fungi and soil phosphorus availability influences plant community productivity and ecosystem stability.
Journal of Ecology , 102 , 1072 -1082 .
[本文引用: 2]
[63]
Zhang WP Jia X Damgaard C Morris EC Bai YY Pan S Wang GX 2013 ). The interplay between above- and below-ground plant-plant interactions along an environmental gradient: Insights from two-layer zone-of-influence models.
Oikos , 122 , 1147 -1156 .
[本文引用: 2]
[64]
Zhi YB Li HL An SQ Zhao L Zhou CF Deng ZF 2007 ). Inter-specific competition: Spartina alterniflora is replacing Spartina anglica in coastal China.
Estuarine, Coastal and Shelf Science , 74 , 437 -448 .
Phytochrome signalling in plant canopies: Testing its population-level implications with photoreceptor mutants of Arabidopsis.
1
1997
... 植物的形态可塑性常常通过改变植物地上和地下的生物量分配、增大某部分器官的表面积等途径来影响植物间的竞争关系(Schwinning & Weiner, 1998 ).当土壤中水肥资源受限时, 植物就竭力分配生物量给地下(通常干旱区植物有发达的根系), 同时增大根系与土壤接触的表面积, 竭力使植物体在水肥资源竞争中占优势.所以当两株相邻植物获取资源的能力相差不大时, 处于劣势的植物很可能通过形态可塑性, 分配更多生物量给地下, 藉此来弥补根系所处的劣势.但当两株植物的相对优势很明显时, 形态可塑性无法弥补劣势植物的劣势, 致使其无法得到足够的资源, 也就无法快速生长, 陷入了恶性循环, 而优势植物由于获得充足的资源, 可以进一步生长, 根系更加强大, 获取资源能力更强, 形成了良性循环, 两株植物间根系的非对称性竞争模式就形成了.形态可塑性在减弱植物个体间非对称性竞争中的作用已经得到一系列转基因实验的证明(Ballaré et al ., 1994 ; Ballaré & Scopel, 1997 ).同样, 形态可塑性也会影响植物体地上部分对光资源的竞争.研究发现, 植物可塑性和时空上的生态位分化能够提高植物获取光的能力并维持生物多样性的积极影响(Sapijanskas et al ., 2014 ).生物多样性局面的维持表明物种间可以共存, 植物可塑性等因素使得同生境中生长的为光资源而发生激烈非对称性竞争的植物能够实现共存. ...
Signaling among neighboring plants and the development of size inequalities in plant populations.
1
1994
... 植物的形态可塑性常常通过改变植物地上和地下的生物量分配、增大某部分器官的表面积等途径来影响植物间的竞争关系(Schwinning & Weiner, 1998 ).当土壤中水肥资源受限时, 植物就竭力分配生物量给地下(通常干旱区植物有发达的根系), 同时增大根系与土壤接触的表面积, 竭力使植物体在水肥资源竞争中占优势.所以当两株相邻植物获取资源的能力相差不大时, 处于劣势的植物很可能通过形态可塑性, 分配更多生物量给地下, 藉此来弥补根系所处的劣势.但当两株植物的相对优势很明显时, 形态可塑性无法弥补劣势植物的劣势, 致使其无法得到足够的资源, 也就无法快速生长, 陷入了恶性循环, 而优势植物由于获得充足的资源, 可以进一步生长, 根系更加强大, 获取资源能力更强, 形成了良性循环, 两株植物间根系的非对称性竞争模式就形成了.形态可塑性在减弱植物个体间非对称性竞争中的作用已经得到一系列转基因实验的证明(Ballaré et al ., 1994 ; Ballaré & Scopel, 1997 ).同样, 形态可塑性也会影响植物体地上部分对光资源的竞争.研究发现, 植物可塑性和时空上的生态位分化能够提高植物获取光的能力并维持生物多样性的积极影响(Sapijanskas et al ., 2014 ).生物多样性局面的维持表明物种间可以共存, 植物可塑性等因素使得同生境中生长的为光资源而发生激烈非对称性竞争的植物能够实现共存. ...
Effects of farm size and on-farm landscape heterogeneity on biodiversity-case study of twelve farms in a Swedish landscape.
1
2015
... 如上所述, 研究生物体间的非对称性竞争问题可以与生物的共存、生物多样性、生物入侵等问题相联系.在一定的条件下, 生物多样性降低很可能是由物种间非对称性竞争而造成的(Šmilauer & Šmilauerová, 2013 ).在相同生境中两物种间出现完全的非对称性竞争时, 一物种完全处于竞争优势地位, 而另一物种不占据任何资源优势, 在竞争中逐渐衰败, 最终死亡.因此, 当物种间出现非对称性竞争时, 物种无法共存, 它们所处的生态系统生物多样性会下降, 生物入侵容易发生.而这几个方面恰恰与土壤异质性有关, 或者说土壤异质性可以部分解释物种共存、生物多样性和生物入侵等问题.首先, 土壤异质性与物种入侵间可以发生相互作用和反馈调节.已有证据表明, 外来物种入侵会显著改变生态系统N循环, 入侵物种增加了N含量, 而且加快了N的流动循环Castro-Díez et al ., 2014 ).因此, 随着物种入侵, 土壤中营养元素的成分和含量会发生大的变化, 致使土壤异质性加强.同时, 物种入侵也会受土壤异质性影响.Burns和Brandt (2014)研究发现, 土壤异质性与种群周转率相互作用影响物种入侵, 增加土壤异质性会增加物种入侵.两者间的相互作用关系启示我们在研究物种间非对称性竞争问题时应与生物入侵问题相联系.其次, 土壤异质性会影响生物多样性, 多数情况下土壤异质性会增加生物多样性(Belfrage et al ., 2015 ).从生态位角度考虑, 土壤异质性可以为物种提供更多的生态位, 适应不同物种对不同生境的需求, 而且不同的生态位缓解了物种间对各种资源的强烈竞争, 使生物出现共存局面, 最终可能表现为植物间较弱的非对称性竞争或转变为对称性竞争.但是, 近来研究发现, 土壤异质性高并不意味着生物多样性也升高, 例如当土壤中某种元素含量很高而出现富营养化时, 情况会大不相同.已有研究表明土壤富营养化会削弱生物多样性产生的稳定性效应(Hautier et al ., 2014 ).也就是说, 物种稳定共存的局面不会长期维持下去, 总有一些物种会在竞争中被淘汰掉.因此, 土壤异质性升高、物种多样性降低的环境中, 物种间易于出现非对称性竞争. ...
Interspecific competition impacts on the morphology and distribution of fine roots in European beech (Fagus sylvatica L.) and Norway spruce (Picea abies (L.) Karst.).
1
2006
... 早期的竞争理论认为: 植物体地上部分主要竞争光资源, 属于非对称性竞争; 地下部分主要竞争水肥资源, 属于对称性竞争(Weiner, 1986 ; Weiner et al ., 1997 ; Schwinning & Weiner, 1998 ; Zhang et al ., 2013 ).其原理是: 高大植物凭借株高优势遮挡周围的矮小植物, 阻碍其获得充足的光照, 而矮小植物对高大植物的光吸收影响不大, 因此, 植物光竞争为非对称性竞争Weiner & Thomas, 1986 ; Connoily & Wayne, 1996 ); 由于地下水肥资源的流动特性, 使不同个体大小的植物均能得到资源, 因此植物水肥竞争为对称性竞争Bolte & Villanueva, 2006 ).但近年也有研究Rajaniem, 2003 ; Rewald & Leuschner, 2009 ; Lei et al ., 2012 ; Weremijewicz & Janos, 2013 )发现, 地下竞争为非对称性竞争或存在非对称性竞争倾向.比如, Rajaniemi (2003)研究发现, 植物在不同种群密度条件下, 目标植物周围存在不同大小的植物时, 较大的植物对目标植物单位生物量的影响大于较小植物对目标植物的影响, 并证明地下竞争可能为非对称性竞争.而Rewald和Leuschner (2009)以水青冈属(Fagus )和栎属(Quercus )为研究对象, 将野外生长的两树种的部分根系非破坏性挖出, 插进设计好的根生长室中, 使这部分根系发生种内、种间竞争作用, 并给予不同的水分梯度处理, 研究表明植物根系的非对称性竞争还会受到水分梯度的影响, 即植物间的非对称性竞争程度随水分胁迫梯度的增加而增加.植物根系间发生非对称性竞争现象的作用机理通常可以解释为: 生长于同一环境中的两株大小不同的植物, 较大的植物根系伸展范围广, 根系接触土壤中营养元素的表面积大, 获取的水肥资源多, 进而促进植物生长, 而进一步的生长又会使其根系伸展范围更广, 根系接触土壤中营养元素的表面积更大, 吸收消耗土壤中水分营养元素资源的能力更强.在土壤中资源有限的情况下, 大个体植物因其具有更强的吸收消耗能力而占有更多小个体根际周围的水肥资源, 进而导致小个体植物根际周围水肥资源的供应速率越来越低, 加剧了对小个体植物生长的限制.因此, 随着植物生长, 小个体与大个体间对土壤中有限资源的竞争能力和资源的占有量会随着植物的生长而进一步扩大, 最终同样可能导致小个体因资源的枯竭而死亡, 形成了典型的根系非对称性竞争.此外, 也有研究者认为, 在同等环境条件下, 植物个体仅仅通过对有限资源的抢先占有, 便能在竞争中胜出(Craine et al ., 2005 ), 也就是说, 植物个体对有限资源获取的起始时间不一致, 导致了对资源的非对称性竞争. ...
Herbivores and nutrients control grassland plant diversity via light limitation.
1
2014
... 竞争是陆生植物生长过程中的重要现象, 竞争作用是群落结构组建的主导因子和决定物种进化模式的重要因素(王刚和张大勇, 1996 ).对物种竞争的研究由来已久, 19世纪50年代末, 达尔文曾指出生活需求相似的近缘物种间经常发生激烈的竞争.随后, 洛特和沃尔特拉用数学公式描述了物种间的竞争关系.高斯假说从生态位角度展示了物种竞争的结果.竞争的研究领域涉及个体、种群、群落、生态系统等不同层次, 如竞争对个体大小不整齐性的影响(Weiner & Thomas, 1986 )、竞争对种群数量的调节(Deng et al ., 2006 )、竞争与群落空间分布格局的关系(Lin et al ., 2012 )、竞争在维持生态系统物种多样性中的作用(Borer et al ., 2014 )等等.随着植物间相互作用研究的不断拓展, 不断有新的概念和理论模型被提出, 甚至有的学者试图打破竞争与正相互作用的清晰界限(Warren et al ., 2014 ).在逐渐深入研究植物间相互作用的过程中, 植物地下根系间的竞争及其生态过程更受关注(Sutherland et al ., 2013 ).本文在前人研究的基础上, 从竞争模式角度着重对植物竞争(包括地上竞争和地下竞争)的对称性和非对称性进行综述和分析, 以期为国内相关研究提供参考和借鉴. ...
Heterogeneity in plant-soil feedbacks and resident population dynamics affect mutual invasibility.
0
2014
Plant competition underground.
1
1997
... 尽管地下竞争通常大于地上竞争, 尤其是在营养元素贫瘠的土壤中Casper & Jackson, 1997 ; Kiær et al ., 2013 ), 但地上竞争也不可忽略.影响植物地上竞争的主要因素是光照, 绿色植物进行正常的光合作用需要充足的光照条件, 而在较为拥挤的植物种群或群落中, 植物个体间因相互遮阴而发生光竞争, 高大植物会遮挡其冠下植物对光的吸收, 但冠下植物对上层植物光吸收的影响较小.因此, 植物个体间对光的竞争是单方面的, 随着植物的生长, 高大植物的遮阴效果会越来越强, 形成典型的非对称性竞争, 即由于植物体个体大小不同(表现为株高不同)导致上层植物与冠下植物间对光的竞争为非对称性竞争van Leeuwen & Etienne, 2013 ).这种关于地上竞争的非对称性理论还仅局限于将光照作为一个笼统的概念进行整体分析.在未来的研究中可以对光资源进一步细化分析. ...
What explains variation in the impacts of exotic plant invasions on the nitrogen cycle? A meta-analysis.
1
2014
... 如上所述, 研究生物体间的非对称性竞争问题可以与生物的共存、生物多样性、生物入侵等问题相联系.在一定的条件下, 生物多样性降低很可能是由物种间非对称性竞争而造成的(Šmilauer & Šmilauerová, 2013 ).在相同生境中两物种间出现完全的非对称性竞争时, 一物种完全处于竞争优势地位, 而另一物种不占据任何资源优势, 在竞争中逐渐衰败, 最终死亡.因此, 当物种间出现非对称性竞争时, 物种无法共存, 它们所处的生态系统生物多样性会下降, 生物入侵容易发生.而这几个方面恰恰与土壤异质性有关, 或者说土壤异质性可以部分解释物种共存、生物多样性和生物入侵等问题.首先, 土壤异质性与物种入侵间可以发生相互作用和反馈调节.已有证据表明, 外来物种入侵会显著改变生态系统N循环, 入侵物种增加了N含量, 而且加快了N的流动循环Castro-Díez et al ., 2014 ).因此, 随着物种入侵, 土壤中营养元素的成分和含量会发生大的变化, 致使土壤异质性加强.同时, 物种入侵也会受土壤异质性影响.Burns和Brandt (2014)研究发现, 土壤异质性与种群周转率相互作用影响物种入侵, 增加土壤异质性会增加物种入侵.两者间的相互作用关系启示我们在研究物种间非对称性竞争问题时应与生物入侵问题相联系.其次, 土壤异质性会影响生物多样性, 多数情况下土壤异质性会增加生物多样性(Belfrage et al ., 2015 ).从生态位角度考虑, 土壤异质性可以为物种提供更多的生态位, 适应不同物种对不同生境的需求, 而且不同的生态位缓解了物种间对各种资源的强烈竞争, 使生物出现共存局面, 最终可能表现为植物间较弱的非对称性竞争或转变为对称性竞争.但是, 近来研究发现, 土壤异质性高并不意味着生物多样性也升高, 例如当土壤中某种元素含量很高而出现富营养化时, 情况会大不相同.已有研究表明土壤富营养化会削弱生物多样性产生的稳定性效应(Hautier et al ., 2014 ).也就是说, 物种稳定共存的局面不会长期维持下去, 总有一些物种会在竞争中被淘汰掉.因此, 土壤异质性升高、物种多样性降低的环境中, 物种间易于出现非对称性竞争. ...
Balance between facilitation and resource competition determines biomass-density relationships in plant populations.
3
2008
... 有关植物竞争类型的理论研究一直是生态学领域的研究重点, 因为这是厘清植物邻体间竞争过程和揭示其竞争机制的必备环节.而且植物竞争类型也是在群落水平上研究植物群落结构组成、生态位分化和物种多样性及其稳定性维系机制的基础.竞争的对称性和非对称性问题同样会影响到一系列以此为基础的科学研究.比如, Hautier等(2009)在《Science》上发表文章认为, 立地环境的富营养化促进了植物对光非对称性竞争的发生, 进而导致物种多样性发生了变化.这主要是由于一些对光竞争处于劣势地位的物种得不到足够的光资源最终死亡.在开展植物对光非对称性竞争研究的同时, 植物根系非对称性竞争的研究也进行得如火如荼.近年来, 越来越多的研究证实在环境胁迫条件下植物根系竞争也为非对称性竞争Rajaniem, 2003 ).基于Hautier等(2009)的研究, 我们可以假设: 在以水分和营养元素为关键限制性资源的干旱荒漠生态系统中, 植物群落多样性也将由根系非对称性竞争所调控.而在植物生物量分配方面, 陆生植物体间的相互作用(竞争和促进)也会影响植物体地上和地下的生物量分配(Deng et al ., 2006 ; Chu et al ., 2008 ), 对植物对称性与非对称性竞争理论的深入研究或许能更清晰地揭示这种响应机理.至于植物个体大小不整齐性随种群密度的增加而增加这一现象(Weiner, 1986 ), 究竟是由于空间格局不一致引起植物密度变化而产生的Thomas & Weiner, 1989 ), 还是由于植物体的非对称性竞争产生的, 或者是植物本身具有较强的可塑性而导致的, 需要生态学家对其相关理论进一步深入研究.物种共存、生物入侵以及生物多样性等问题也与植物体间非对称性竞争有着千丝万缕的联系(Osunkoya et al ., 2013 ; Skácelová & Lepš, 2014 ), 研究非对称性竞争的产生机制和生理原理对开展相关工作具有重要意义. ...
... 其中, r 是n 个植物体占有的总资源量, bi 代表种群中某植物个体i 的生物量, ri 代表植物个体i占有的资源量, 参数θ 的取值大小反映了竞争模式.当θ 取值为0或趋近于∞时, 植物间分别以完全对称性竞争和完全非对称性竞争方式分配资源; 当在区间(0, ∞)取值时, 则代表植物间竞争模式从对称性竞争向非对称性竞争的连续性过渡.该模型奠定了研究竞争模式的基础, 此后的许多实验或理论研究均运用此模型或以此模型为基础解决生态学竞争领域内的一些复杂问题(Chu et al ., 2008 , 2009, 2010 ; Lin et al ., 2012 ).但这一模型仅仅是对对称性和非对称性竞争概念的界定, 如用于实际研究, 则显得过于简单, 以至于会忽略掉很多影响植物间竞争模式的重要因子, 譬如物种特异性等. ...
... 学者们除了通过目标植物生物量与邻体数量或密度间的变化关系来研究植物间竞争类型外, 还以个体影响域模型为基础, 通过数学建模和对植物个体生长过程的计算机模拟来研究植物间的对称性和非对称性竞争以及正相互作用强度等(Chu et al ., 2008 ), 并逐渐由植物体单层影响域模型向将植物体地上、地下部分区分开的双层影响域模型发展(Zhang et al ., 2013 ).关于运用个体影响域模型来解决对称性与非对称性竞争问题的研究, 未来可能更多关注于模型的进一步完善以及在建模过程中逐步将植物体根、茎、叶等器官区分开来. ...
Effects of traits, species identity and local environmental conditions on the assessment of interactions: Insights from an alpine meadow community.
1
2009
... 其中, r 是n 个植物体占有的总资源量, bi 代表种群中某植物个体i 的生物量, ri 代表植物个体i占有的资源量, 参数θ 的取值大小反映了竞争模式.当θ 取值为0或趋近于∞时, 植物间分别以完全对称性竞争和完全非对称性竞争方式分配资源; 当在区间(0, ∞)取值时, 则代表植物间竞争模式从对称性竞争向非对称性竞争的连续性过渡.该模型奠定了研究竞争模式的基础, 此后的许多实验或理论研究均运用此模型或以此模型为基础解决生态学竞争领域内的一些复杂问题(Chu et al ., 2008 , 2009, 2010 ; Lin et al ., 2012 ).但这一模型仅仅是对对称性和非对称性竞争概念的界定, 如用于实际研究, 则显得过于简单, 以至于会忽略掉很多影响植物间竞争模式的重要因子, 譬如物种特异性等. ...
Effects of positive interactions, size symmetry of competition and abiotic stress on self-thinning in simulated plant populations.
0
2010
Asymmetric competition between plant species.
1
1996
... 早期的竞争理论认为: 植物体地上部分主要竞争光资源, 属于非对称性竞争; 地下部分主要竞争水肥资源, 属于对称性竞争(Weiner, 1986 ; Weiner et al ., 1997 ; Schwinning & Weiner, 1998 ; Zhang et al ., 2013 ).其原理是: 高大植物凭借株高优势遮挡周围的矮小植物, 阻碍其获得充足的光照, 而矮小植物对高大植物的光吸收影响不大, 因此, 植物光竞争为非对称性竞争Weiner & Thomas, 1986 ; Connoily & Wayne, 1996 ); 由于地下水肥资源的流动特性, 使不同个体大小的植物均能得到资源, 因此植物水肥竞争为对称性竞争Bolte & Villanueva, 2006 ).但近年也有研究Rajaniem, 2003 ; Rewald & Leuschner, 2009 ; Lei et al ., 2012 ; Weremijewicz & Janos, 2013 )发现, 地下竞争为非对称性竞争或存在非对称性竞争倾向.比如, Rajaniemi (2003)研究发现, 植物在不同种群密度条件下, 目标植物周围存在不同大小的植物时, 较大的植物对目标植物单位生物量的影响大于较小植物对目标植物的影响, 并证明地下竞争可能为非对称性竞争.而Rewald和Leuschner (2009)以水青冈属(Fagus )和栎属(Quercus )为研究对象, 将野外生长的两树种的部分根系非破坏性挖出, 插进设计好的根生长室中, 使这部分根系发生种内、种间竞争作用, 并给予不同的水分梯度处理, 研究表明植物根系的非对称性竞争还会受到水分梯度的影响, 即植物间的非对称性竞争程度随水分胁迫梯度的增加而增加.植物根系间发生非对称性竞争现象的作用机理通常可以解释为: 生长于同一环境中的两株大小不同的植物, 较大的植物根系伸展范围广, 根系接触土壤中营养元素的表面积大, 获取的水肥资源多, 进而促进植物生长, 而进一步的生长又会使其根系伸展范围更广, 根系接触土壤中营养元素的表面积更大, 吸收消耗土壤中水分营养元素资源的能力更强.在土壤中资源有限的情况下, 大个体植物因其具有更强的吸收消耗能力而占有更多小个体根际周围的水肥资源, 进而导致小个体植物根际周围水肥资源的供应速率越来越低, 加剧了对小个体植物生长的限制.因此, 随着植物生长, 小个体与大个体间对土壤中有限资源的竞争能力和资源的占有量会随着植物的生长而进一步扩大, 最终同样可能导致小个体因资源的枯竭而死亡, 形成了典型的根系非对称性竞争.此外, 也有研究者认为, 在同等环境条件下, 植物个体仅仅通过对有限资源的抢先占有, 便能在竞争中胜出(Craine et al ., 2005 ), 也就是说, 植物个体对有限资源获取的起始时间不一致, 导致了对资源的非对称性竞争. ...
Supply pre-emption, not concentration reduction, is the mechanism of competition for nutrients.
1
2005
... 早期的竞争理论认为: 植物体地上部分主要竞争光资源, 属于非对称性竞争; 地下部分主要竞争水肥资源, 属于对称性竞争(Weiner, 1986 ; Weiner et al ., 1997 ; Schwinning & Weiner, 1998 ; Zhang et al ., 2013 ).其原理是: 高大植物凭借株高优势遮挡周围的矮小植物, 阻碍其获得充足的光照, 而矮小植物对高大植物的光吸收影响不大, 因此, 植物光竞争为非对称性竞争Weiner & Thomas, 1986 ; Connoily & Wayne, 1996 ); 由于地下水肥资源的流动特性, 使不同个体大小的植物均能得到资源, 因此植物水肥竞争为对称性竞争Bolte & Villanueva, 2006 ).但近年也有研究Rajaniem, 2003 ; Rewald & Leuschner, 2009 ; Lei et al ., 2012 ; Weremijewicz & Janos, 2013 )发现, 地下竞争为非对称性竞争或存在非对称性竞争倾向.比如, Rajaniemi (2003)研究发现, 植物在不同种群密度条件下, 目标植物周围存在不同大小的植物时, 较大的植物对目标植物单位生物量的影响大于较小植物对目标植物的影响, 并证明地下竞争可能为非对称性竞争.而Rewald和Leuschner (2009)以水青冈属(Fagus )和栎属(Quercus )为研究对象, 将野外生长的两树种的部分根系非破坏性挖出, 插进设计好的根生长室中, 使这部分根系发生种内、种间竞争作用, 并给予不同的水分梯度处理, 研究表明植物根系的非对称性竞争还会受到水分梯度的影响, 即植物间的非对称性竞争程度随水分胁迫梯度的增加而增加.植物根系间发生非对称性竞争现象的作用机理通常可以解释为: 生长于同一环境中的两株大小不同的植物, 较大的植物根系伸展范围广, 根系接触土壤中营养元素的表面积大, 获取的水肥资源多, 进而促进植物生长, 而进一步的生长又会使其根系伸展范围更广, 根系接触土壤中营养元素的表面积更大, 吸收消耗土壤中水分营养元素资源的能力更强.在土壤中资源有限的情况下, 大个体植物因其具有更强的吸收消耗能力而占有更多小个体根际周围的水肥资源, 进而导致小个体植物根际周围水肥资源的供应速率越来越低, 加剧了对小个体植物生长的限制.因此, 随着植物生长, 小个体与大个体间对土壤中有限资源的竞争能力和资源的占有量会随着植物的生长而进一步扩大, 最终同样可能导致小个体因资源的枯竭而死亡, 形成了典型的根系非对称性竞争.此外, 也有研究者认为, 在同等环境条件下, 植物个体仅仅通过对有限资源的抢先占有, 便能在竞争中胜出(Craine et al ., 2005 ), 也就是说, 植物个体对有限资源获取的起始时间不一致, 导致了对资源的非对称性竞争. ...
Root competition: Towards a mechanistic understanding.
1
2003
... 一系列对草本和灌木物种的研究(Mahall & Callaway, 1996 ; de Kroon et al., 2003 ; Falik et al ., 2003 ; Holzapfel & Alpert, 2003 ; Gruntman & Novoplansky, 2004 )发现, 植物的根系能够通过信号感知并且区分“自己”的和“非己”的根系.这对植物的生长是有利的, 同境中生长的相邻两株植物, 每株植物可以通过化学信号感知周围另一株植物根系的存在, 并且抑制自己根系向对方根系生长所占区域延伸, 尽量避免自己受到伤害. ...
Plant mass-density relationship along a moisture gradient in north-west China.
4
2006
... 竞争是陆生植物生长过程中的重要现象, 竞争作用是群落结构组建的主导因子和决定物种进化模式的重要因素(王刚和张大勇, 1996 ).对物种竞争的研究由来已久, 19世纪50年代末, 达尔文曾指出生活需求相似的近缘物种间经常发生激烈的竞争.随后, 洛特和沃尔特拉用数学公式描述了物种间的竞争关系.高斯假说从生态位角度展示了物种竞争的结果.竞争的研究领域涉及个体、种群、群落、生态系统等不同层次, 如竞争对个体大小不整齐性的影响(Weiner & Thomas, 1986 )、竞争对种群数量的调节(Deng et al ., 2006 )、竞争与群落空间分布格局的关系(Lin et al ., 2012 )、竞争在维持生态系统物种多样性中的作用(Borer et al ., 2014 )等等.随着植物间相互作用研究的不断拓展, 不断有新的概念和理论模型被提出, 甚至有的学者试图打破竞争与正相互作用的清晰界限(Warren et al ., 2014 ).在逐渐深入研究植物间相互作用的过程中, 植物地下根系间的竞争及其生态过程更受关注(Sutherland et al ., 2013 ).本文在前人研究的基础上, 从竞争模式角度着重对植物竞争(包括地上竞争和地下竞争)的对称性和非对称性进行综述和分析, 以期为国内相关研究提供参考和借鉴. ...
... 迄今为止, 有关植物竞争的概念和类型有很多, 它们之间既有很多共性, 也存在很大差异.根据物种差异、地上地下和竞争模式, 植物竞争类型大致可分为: 种内竞争(intraspecific competition)和种间竞争(interspecific competition)、地上竞争(above- ground competition)和地下竞争(belowground competition) (Deng et al ., 2006 ; 王平等, 2007 )、对称性竞争(symmetric competition)和非对称性竞争(asymm- etric competition) (王刚和张大勇, 1996 ).对称性竞争又可再分为完全对称性竞争和大小依赖的对称性竞争; 非对称性竞争也可再分为完全非对称性竞争和大小依赖的非对称性竞争.种内竞争和种间竞争是基于植物物种的类型进行分类, 强调物种差异对竞争的影响; 而地上竞争和地下竞争的划分主要是基于植物体地上冠层对光资源的竞争和地下根系部分对水和营养元素的竞争, 打破了物种差异的明确界限.从某种意义上讲, 地上、地下竞争将植物竞争的研究范围进一步扩大了, 使得有关植物竞争的研究结论更具普适性.实际上, 对称性竞争与非对称性竞争同样涵盖了植物地上竞争和地下竞争, 但对称性竞争与非对称性竞争从概念上将植物竞争研究的内涵进一步深化, 体现了植物竞争的具体性质; 而地上竞争和地下竞争仅体现为植物竞争的形式.理论上, 地上竞争和地下竞争都可以是对称性或非对称性的; 反之, 植物的对称性竞争与非对称性竞争既可发生在植物的地上部分, 也可发生在植物的地下部分.然而, 对称性竞争与非对称性竞争更加强调了植物最大化获取资源的策略, 从一个新颖的角度阐释环境资源有限的情况下植物个体间分配资源的方式.近年来, 对称性竞争与非对称性竞争的研究逐渐成为热点, 下面将进一步阐释它们的概念及其内涵. ...
... 有关植物竞争类型的理论研究一直是生态学领域的研究重点, 因为这是厘清植物邻体间竞争过程和揭示其竞争机制的必备环节.而且植物竞争类型也是在群落水平上研究植物群落结构组成、生态位分化和物种多样性及其稳定性维系机制的基础.竞争的对称性和非对称性问题同样会影响到一系列以此为基础的科学研究.比如, Hautier等(2009)在《Science》上发表文章认为, 立地环境的富营养化促进了植物对光非对称性竞争的发生, 进而导致物种多样性发生了变化.这主要是由于一些对光竞争处于劣势地位的物种得不到足够的光资源最终死亡.在开展植物对光非对称性竞争研究的同时, 植物根系非对称性竞争的研究也进行得如火如荼.近年来, 越来越多的研究证实在环境胁迫条件下植物根系竞争也为非对称性竞争Rajaniem, 2003 ).基于Hautier等(2009)的研究, 我们可以假设: 在以水分和营养元素为关键限制性资源的干旱荒漠生态系统中, 植物群落多样性也将由根系非对称性竞争所调控.而在植物生物量分配方面, 陆生植物体间的相互作用(竞争和促进)也会影响植物体地上和地下的生物量分配(Deng et al ., 2006 ; Chu et al ., 2008 ), 对植物对称性与非对称性竞争理论的深入研究或许能更清晰地揭示这种响应机理.至于植物个体大小不整齐性随种群密度的增加而增加这一现象(Weiner, 1986 ), 究竟是由于空间格局不一致引起植物密度变化而产生的Thomas & Weiner, 1989 ), 还是由于植物体的非对称性竞争产生的, 或者是植物本身具有较强的可塑性而导致的, 需要生态学家对其相关理论进一步深入研究.物种共存、生物入侵以及生物多样性等问题也与植物体间非对称性竞争有着千丝万缕的联系(Osunkoya et al ., 2013 ; Skácelová & Lepš, 2014 ), 研究非对称性竞争的产生机制和生理原理对开展相关工作具有重要意义. ...
... 其中, E [wx ]代表在物种x 与物种y 相互作用情况下, 物种x 目标个体的预期平均生物量; wm 代表在无邻体竞争情况下单株植物体的预期生物量; Ax 和Ay 则分别表示目标植物与邻体植物物种x 或y 重叠区域占总区域的平均比例; Nx 、Ny 分别表示物种x 和y 与目标植物进行相互作用的个体数.由此推知, 在由物种x 和y 组成的群落中, 植物体的平均生物量与群落密度间的关系呈双曲线型的负相关关系, 即个体平均生物量随该植物邻体数量增加而下降.这与其他学者对平均个体大小与密度关系的研究结果Firbank & Watkinson, 1985b ; Deng et al ., 2006 , 2012a , 2012b )一致, 模型的正确性因此得到检验.这种情况下种群中的个体无论是物种x 还是物种y, 生物量随密度变化的趋势及变化速率均相同; 但是在自然群落中, 很难验证这种对称性竞争, 因为通常在不同的物种间生态位一般都发生了明显分化.上述模型(公式(3)), 同样可应用于植物种内的个体间竞争, 即N x 或N y 为0的情况下, 体现为种内的植物个体间竞争. ...
Models and tests of optimal density and maximal yield for crop plants.
1
2012a
... 其中, E [wx ]代表在物种x 与物种y 相互作用情况下, 物种x 目标个体的预期平均生物量; wm 代表在无邻体竞争情况下单株植物体的预期生物量; Ax 和Ay 则分别表示目标植物与邻体植物物种x 或y 重叠区域占总区域的平均比例; Nx 、Ny 分别表示物种x 和y 与目标植物进行相互作用的个体数.由此推知, 在由物种x 和y 组成的群落中, 植物体的平均生物量与群落密度间的关系呈双曲线型的负相关关系, 即个体平均生物量随该植物邻体数量增加而下降.这与其他学者对平均个体大小与密度关系的研究结果Firbank & Watkinson, 1985b ; Deng et al ., 2006 , 2012a , 2012b )一致, 模型的正确性因此得到检验.这种情况下种群中的个体无论是物种x 还是物种y, 生物量随密度变化的趋势及变化速率均相同; 但是在自然群落中, 很难验证这种对称性竞争, 因为通常在不同的物种间生态位一般都发生了明显分化.上述模型(公式(3)), 同样可应用于植物种内的个体间竞争, 即N x 或N y 为0的情况下, 体现为种内的植物个体间竞争. ...
Insights into plant size-density relationships from models and agricultural crops.
1
2012b
... 其中, E [wx ]代表在物种x 与物种y 相互作用情况下, 物种x 目标个体的预期平均生物量; wm 代表在无邻体竞争情况下单株植物体的预期生物量; Ax 和Ay 则分别表示目标植物与邻体植物物种x 或y 重叠区域占总区域的平均比例; Nx 、Ny 分别表示物种x 和y 与目标植物进行相互作用的个体数.由此推知, 在由物种x 和y 组成的群落中, 植物体的平均生物量与群落密度间的关系呈双曲线型的负相关关系, 即个体平均生物量随该植物邻体数量增加而下降.这与其他学者对平均个体大小与密度关系的研究结果Firbank & Watkinson, 1985b ; Deng et al ., 2006 , 2012a , 2012b )一致, 模型的正确性因此得到检验.这种情况下种群中的个体无论是物种x 还是物种y, 生物量随密度变化的趋势及变化速率均相同; 但是在自然群落中, 很难验证这种对称性竞争, 因为通常在不同的物种间生态位一般都发生了明显分化.上述模型(公式(3)), 同样可应用于植物种内的个体间竞争, 即N x 或N y 为0的情况下, 体现为种内的植物个体间竞争. ...
Soil phosphorus heterogeneity and mycorrhizal symbiosis regulate plant intra-specific competition and size distribution.
0
2002
Self/non-self discrimination in roots.
1
2003
... 一系列对草本和灌木物种的研究(Mahall & Callaway, 1996 ; de Kroon et al., 2003 ; Falik et al ., 2003 ; Holzapfel & Alpert, 2003 ; Gruntman & Novoplansky, 2004 )发现, 植物的根系能够通过信号感知并且区分“自己”的和“非己”的根系.这对植物的生长是有利的, 同境中生长的相邻两株植物, 每株植物可以通过化学信号感知周围另一株植物根系的存在, 并且抑制自己根系向对方根系生长所占区域延伸, 尽量避免自己受到伤害. ...
Root navigation by self inhibition.
1
2005
... 研究发现一些植物能够通过化感作用产生有毒物质, 限制周围植物生长甚至致其死亡(Schenk, 2006 ).通常不同物种间会出现这种情况, 一种植物根系分泌物会影响另一种植物生长.但也有一些物种成年植物体释放化学物质影响和抑制其幼苗个体生长, 例如银桦(Grevillea robusta ).除此之外, 一些植物根系还会释放一种对其他植物无害的化学物质, 帮助植物感知周围其他植物根系的生长及土壤中营养物质等非生命物质的存在.研究发现, 一些豆科植物根系能通过“化学雷达”而不用物理接触就能感知其周围的物理障碍, 从而改变根的生长形态(Falik et al ., 2005 ).资源获取器官若能够感知非生命物质的存在, 在生长中就占据很大优势.例如, 植物根系若能感知土壤中肥力不足的区域或不利于自身生长的因素, 就不朝肥力不足区域生长而转向肥力丰富区域生长或避开不利于自身生长的因素, 从而保证植物体快速生长, 增强其在逆境中的生存竞争能力. ...
Plant species traits and capacity for resource reduction predict yield and abundance under competition in nitrogen-limited grassland.
1
2006
... 一些研究发现, 群落中的优势物种往往是那些能够高效利用环境中限制资源的物种(Fargione & Tilman, 2006 ).菌根中的微生物可以使一些植物得到充足的必需元素, 例如豆科植物中的根瘤菌为豆科植物提供充足的氮源, 促使其快速生长.因此, 这类微生物提升了植物根系在土壤中的竞争能力.最终, 豆科植物越长越大, 根系越来越发达, 需要和吸收的营养元素也越来越多(这些元素包括P、K 等必需元素, 而不仅仅是N), 留给供非豆科植物生长的元素就越来越少, 因此抑制非豆科植物生长的程度越来越大, 形成了物种间根系非对称性竞争.土壤中的微生物还有致病菌, 它们会导致相反的结果.一些植物所携带的细菌、真菌等微生物会对其他植物造成伤害, 尤其是一些入侵物种携带的土壤微生物会限制当地物种利用土壤中的资源, 破坏其根系生长, 严重危及当地物种的正常生长; 或者土壤微生物对植物地下生长的调节影响入侵物种间的相互作用关系(Wolfe & Klironomos, 2005 ).土壤中不同的微生物群落以及根系周围不同的菌群成分会对植物间根系竞争产生不同的影响, 不同的微生物通过增加或降低根际周围营养物质的可利用性, 直接影响竞争根系对营养物质的利用效率, 使得仅有一些植物得到较多资源, 最终不同植物因适合度不同而呈现出非对称性竞争趋势(Yang et al ., 2014 ).土壤微生物还可能通过与土壤中营养元素的相互作用来影响物种间的相互作用关系.研究表明, 在土壤中添加大量P元素会降低生物多样性, 即部分物种因不能共存而被淘汰, 而植物根际周围菌群的存在能够增加生物多样性, 因此豆科植物可以依赖于自身拥有的根瘤菌生存于富营养化的土壤中(Yang et al ., 2014 ).调节土壤中的微生物含量和种类, 同时调整土壤中营养元素的含量和种类, 将两种因素进行排列组合, 或许可以找到特定环境, 适合某种植物生存而不利于另一种植物生存, 若使两种植物相邻生长, 物种间非对称性竞争较易体现. ...
A model of interference within plant monocultures.
1
1985a
... 首先, 假设在植物群落中植物物种x 和物种y 的个体分布均为随机分布Firbank & Watkinson, 1985a ), 而且无论植物体处于什么空间位置, 其预期表现(即生态位和对资源获取的能力)都是相同的, 并且每株植物处于任何位置的概率也都是相同的, 物种x 处于k 处的概率px (k )与物种y 处于k 处的概率py (k )相等, 而且: ...
On the analysis of competition within two-species mixtures of plants.
1
1985b
... 其中, E [wx ]代表在物种x 与物种y 相互作用情况下, 物种x 目标个体的预期平均生物量; wm 代表在无邻体竞争情况下单株植物体的预期生物量; Ax 和Ay 则分别表示目标植物与邻体植物物种x 或y 重叠区域占总区域的平均比例; Nx 、Ny 分别表示物种x 和y 与目标植物进行相互作用的个体数.由此推知, 在由物种x 和y 组成的群落中, 植物体的平均生物量与群落密度间的关系呈双曲线型的负相关关系, 即个体平均生物量随该植物邻体数量增加而下降.这与其他学者对平均个体大小与密度关系的研究结果Firbank & Watkinson, 1985b ; Deng et al ., 2006 , 2012a , 2012b )一致, 模型的正确性因此得到检验.这种情况下种群中的个体无论是物种x 还是物种y, 生物量随密度变化的趋势及变化速率均相同; 但是在自然群落中, 很难验证这种对称性竞争, 因为通常在不同的物种间生态位一般都发生了明显分化.上述模型(公式(3)), 同样可应用于植物种内的个体间竞争, 即N x 或N y 为0的情况下, 体现为种内的植物个体间竞争. ...
On the analysis of competition at the level of the individual plant.
1
1987
... 野外实验和实地考察有其客观性和可验证性, 但不确定因素也较多, 准确地研究对称性竞争和非对称性竞争效应较为困难(Firbank & Watkinson, 1987 ), 而模拟实验在研究精度、可操作性和避开不确定因素影响等方面具有明显优势.对相关模型的检验也可通过编程在计算机上进行模拟实验, 尤其是随着植物生长的动态竞争过程更适合通过模拟实验来实现.通常运用Netlogo软件(http://ccl.north- western.edu/netlogo)来实现植物竞争或生长动态等过程的模拟.该软件是由美国西北大学开发的一款能够对自然和社会现象进行仿真模拟的建模软件平台Wilensky, 1999 ).实际操作中, 可通过编程或根据系统动力学原理建立模型中各变量间的关系流程图来完成模拟过程.近年来发展起来的具有强大统计分析功能的R软件也可用于建模仿真和统计模拟, 具体模拟过程主要依据蒙特卡罗方法, 通过计算机产生的随机数(伪随机数)模拟离散随机变量或连续随机变量, 并将预解决的科学问题构建为合适的数学模型, 使模拟的变量正好是数学模型的解. ...
Asymmetric competition between plant species.
3
2001
... 因此, 竞争方程Freckleton & Watkinson, 2001 )是: ...
... 竞争方程要考虑两点: 1)某一物种占据某一邻体位置的概率; 2)每个位置被邻体植株消耗的资源量.因此, 在位置i 处, 物种x 的资源消耗量就是物种x 占据该位置的概率(Nx px )与单株植物体所耗资源量(1 - Ax dx )的乘积Freckleton & Watkinson, 2001 ): ...
... 在某一特定的植物群落中, 目标植物体占据任何位置的概率都是相同的, 这一点与个体间的对称性竞争情况相同, 所以在位置k 处有: px (k ) = py (k ) = 1 / (1+ Nx + Ny ).结合公式(2)和公式(4), 在Nx 和Ny 的极限值趋于无穷时, 有公式(5) Freckleton & Watkinson, 2001 ): ...
On the measurement of plant competition intensity.
1
1995
... 检验植物体竞争对称性与非对称性的理论模型最主要和最直接的方法就是野外实验或控制实验.通常实验中具体的操作方法是测量生物量(Weiner, 1986 )、株高(Zhi et al ., 2007 )、冠幅(Iponga et al ., 2008 )、叶面积(Zhi et al ., 2007 )、根长(Reubens et al ., 2011 )、根表面积(Rewald & Leuschner, 2009 )等指标, 根据这些指标可以计算竞争强度(competition intensity)、竞争重要性(competition importance)和竞争反应(competition response)等参数(王平等, 2007 ).而这些参数的具体计算方法又有很大不同, 如根据计算竞争强度是否随生产力变化, 可将竞争强度分为相对竞争强度(relative competition intensity)和绝对竞争强度(absolute competition intensity) (Grace, 1995 ); 此外, 还可以根据植物地上和地下生长环境不同计算地上竞争强度、地下竞争强度以及总竞争强度, 实际运用中应根据实验设计和实验目的选择合适的参数, 各个参数的计算主要通过实施对照实验并分析植物体在有无竞争情况下的差异来进行. ...
Physiologically mediated self/non-self discrimination in roots.
1
2004
... 一系列对草本和灌木物种的研究(Mahall & Callaway, 1996 ; de Kroon et al., 2003 ; Falik et al ., 2003 ; Holzapfel & Alpert, 2003 ; Gruntman & Novoplansky, 2004 )发现, 植物的根系能够通过信号感知并且区分“自己”的和“非己”的根系.这对植物的生长是有利的, 同境中生长的相邻两株植物, 每株植物可以通过化学信号感知周围另一株植物根系的存在, 并且抑制自己根系向对方根系生长所占区域延伸, 尽量避免自己受到伤害. ...
Competition for light causes plant biodiversity loss after eutrophication.
0
2009
Eutrophication weakens stabilizing effects of diversity in natural grasslands.
1
2014
... 如上所述, 研究生物体间的非对称性竞争问题可以与生物的共存、生物多样性、生物入侵等问题相联系.在一定的条件下, 生物多样性降低很可能是由物种间非对称性竞争而造成的(Šmilauer & Šmilauerová, 2013 ).在相同生境中两物种间出现完全的非对称性竞争时, 一物种完全处于竞争优势地位, 而另一物种不占据任何资源优势, 在竞争中逐渐衰败, 最终死亡.因此, 当物种间出现非对称性竞争时, 物种无法共存, 它们所处的生态系统生物多样性会下降, 生物入侵容易发生.而这几个方面恰恰与土壤异质性有关, 或者说土壤异质性可以部分解释物种共存、生物多样性和生物入侵等问题.首先, 土壤异质性与物种入侵间可以发生相互作用和反馈调节.已有证据表明, 外来物种入侵会显著改变生态系统N循环, 入侵物种增加了N含量, 而且加快了N的流动循环Castro-Díez et al ., 2014 ).因此, 随着物种入侵, 土壤中营养元素的成分和含量会发生大的变化, 致使土壤异质性加强.同时, 物种入侵也会受土壤异质性影响.Burns和Brandt (2014)研究发现, 土壤异质性与种群周转率相互作用影响物种入侵, 增加土壤异质性会增加物种入侵.两者间的相互作用关系启示我们在研究物种间非对称性竞争问题时应与生物入侵问题相联系.其次, 土壤异质性会影响生物多样性, 多数情况下土壤异质性会增加生物多样性(Belfrage et al ., 2015 ).从生态位角度考虑, 土壤异质性可以为物种提供更多的生态位, 适应不同物种对不同生境的需求, 而且不同的生态位缓解了物种间对各种资源的强烈竞争, 使生物出现共存局面, 最终可能表现为植物间较弱的非对称性竞争或转变为对称性竞争.但是, 近来研究发现, 土壤异质性高并不意味着生物多样性也升高, 例如当土壤中某种元素含量很高而出现富营养化时, 情况会大不相同.已有研究表明土壤富营养化会削弱生物多样性产生的稳定性效应(Hautier et al ., 2014 ).也就是说, 物种稳定共存的局面不会长期维持下去, 总有一些物种会在竞争中被淘汰掉.因此, 土壤异质性升高、物种多样性降低的环境中, 物种间易于出现非对称性竞争. ...
Root cooperation in a clonal plant: Connected strawberries segregate roots.
1
2003
... 一系列对草本和灌木物种的研究(Mahall & Callaway, 1996 ; de Kroon et al., 2003 ; Falik et al ., 2003 ; Holzapfel & Alpert, 2003 ; Gruntman & Novoplansky, 2004 )发现, 植物的根系能够通过信号感知并且区分“自己”的和“非己”的根系.这对植物的生长是有利的, 同境中生长的相邻两株植物, 每株植物可以通过化学信号感知周围另一株植物根系的存在, 并且抑制自己根系向对方根系生长所占区域延伸, 尽量避免自己受到伤害. ...
Superiority in competition for light: A crucial attribute defining the impact of the invasive alien tree Schinus molle (Anacardiaceae) in South African Savanna.
1
2008
... 检验植物体竞争对称性与非对称性的理论模型最主要和最直接的方法就是野外实验或控制实验.通常实验中具体的操作方法是测量生物量(Weiner, 1986 )、株高(Zhi et al ., 2007 )、冠幅(Iponga et al ., 2008 )、叶面积(Zhi et al ., 2007 )、根长(Reubens et al ., 2011 )、根表面积(Rewald & Leuschner, 2009 )等指标, 根据这些指标可以计算竞争强度(competition intensity)、竞争重要性(competition importance)和竞争反应(competition response)等参数(王平等, 2007 ).而这些参数的具体计算方法又有很大不同, 如根据计算竞争强度是否随生产力变化, 可将竞争强度分为相对竞争强度(relative competition intensity)和绝对竞争强度(absolute competition intensity) (Grace, 1995 ); 此外, 还可以根据植物地上和地下生长环境不同计算地上竞争强度、地下竞争强度以及总竞争强度, 实际运用中应根据实验设计和实验目的选择合适的参数, 各个参数的计算主要通过实施对照实验并分析植物体在有无竞争情况下的差异来进行. ...
Root and shoot competition: A meta-analysis.
1
2013
... 尽管地下竞争通常大于地上竞争, 尤其是在营养元素贫瘠的土壤中Casper & Jackson, 1997 ; Kiær et al ., 2013 ), 但地上竞争也不可忽略.影响植物地上竞争的主要因素是光照, 绿色植物进行正常的光合作用需要充足的光照条件, 而在较为拥挤的植物种群或群落中, 植物个体间因相互遮阴而发生光竞争, 高大植物会遮挡其冠下植物对光的吸收, 但冠下植物对上层植物光吸收的影响较小.因此, 植物个体间对光的竞争是单方面的, 随着植物的生长, 高大植物的遮阴效果会越来越强, 形成典型的非对称性竞争, 即由于植物体个体大小不同(表现为株高不同)导致上层植物与冠下植物间对光的竞争为非对称性竞争van Leeuwen & Etienne, 2013 ).这种关于地上竞争的非对称性理论还仅局限于将光照作为一个笼统的概念进行整体分析.在未来的研究中可以对光资源进一步细化分析. ...
Belowground facilitation and competition in young tree species mixtures.
1
2012
... 早期的竞争理论认为: 植物体地上部分主要竞争光资源, 属于非对称性竞争; 地下部分主要竞争水肥资源, 属于对称性竞争(Weiner, 1986 ; Weiner et al ., 1997 ; Schwinning & Weiner, 1998 ; Zhang et al ., 2013 ).其原理是: 高大植物凭借株高优势遮挡周围的矮小植物, 阻碍其获得充足的光照, 而矮小植物对高大植物的光吸收影响不大, 因此, 植物光竞争为非对称性竞争Weiner & Thomas, 1986 ; Connoily & Wayne, 1996 ); 由于地下水肥资源的流动特性, 使不同个体大小的植物均能得到资源, 因此植物水肥竞争为对称性竞争Bolte & Villanueva, 2006 ).但近年也有研究Rajaniem, 2003 ; Rewald & Leuschner, 2009 ; Lei et al ., 2012 ; Weremijewicz & Janos, 2013 )发现, 地下竞争为非对称性竞争或存在非对称性竞争倾向.比如, Rajaniemi (2003)研究发现, 植物在不同种群密度条件下, 目标植物周围存在不同大小的植物时, 较大的植物对目标植物单位生物量的影响大于较小植物对目标植物的影响, 并证明地下竞争可能为非对称性竞争.而Rewald和Leuschner (2009)以水青冈属(Fagus )和栎属(Quercus )为研究对象, 将野外生长的两树种的部分根系非破坏性挖出, 插进设计好的根生长室中, 使这部分根系发生种内、种间竞争作用, 并给予不同的水分梯度处理, 研究表明植物根系的非对称性竞争还会受到水分梯度的影响, 即植物间的非对称性竞争程度随水分胁迫梯度的增加而增加.植物根系间发生非对称性竞争现象的作用机理通常可以解释为: 生长于同一环境中的两株大小不同的植物, 较大的植物根系伸展范围广, 根系接触土壤中营养元素的表面积大, 获取的水肥资源多, 进而促进植物生长, 而进一步的生长又会使其根系伸展范围更广, 根系接触土壤中营养元素的表面积更大, 吸收消耗土壤中水分营养元素资源的能力更强.在土壤中资源有限的情况下, 大个体植物因其具有更强的吸收消耗能力而占有更多小个体根际周围的水肥资源, 进而导致小个体植物根际周围水肥资源的供应速率越来越低, 加剧了对小个体植物生长的限制.因此, 随着植物生长, 小个体与大个体间对土壤中有限资源的竞争能力和资源的占有量会随着植物的生长而进一步扩大, 最终同样可能导致小个体因资源的枯竭而死亡, 形成了典型的根系非对称性竞争.此外, 也有研究者认为, 在同等环境条件下, 植物个体仅仅通过对有限资源的抢先占有, 便能在竞争中胜出(Craine et al ., 2005 ), 也就是说, 植物个体对有限资源获取的起始时间不一致, 导致了对资源的非对称性竞争. ...
Differences between symmetric and asymmetric facilitation matter: Exploring the interplay between modes of positive and negative plant interactions.
2
2012
... 竞争是陆生植物生长过程中的重要现象, 竞争作用是群落结构组建的主导因子和决定物种进化模式的重要因素(王刚和张大勇, 1996 ).对物种竞争的研究由来已久, 19世纪50年代末, 达尔文曾指出生活需求相似的近缘物种间经常发生激烈的竞争.随后, 洛特和沃尔特拉用数学公式描述了物种间的竞争关系.高斯假说从生态位角度展示了物种竞争的结果.竞争的研究领域涉及个体、种群、群落、生态系统等不同层次, 如竞争对个体大小不整齐性的影响(Weiner & Thomas, 1986 )、竞争对种群数量的调节(Deng et al ., 2006 )、竞争与群落空间分布格局的关系(Lin et al ., 2012 )、竞争在维持生态系统物种多样性中的作用(Borer et al ., 2014 )等等.随着植物间相互作用研究的不断拓展, 不断有新的概念和理论模型被提出, 甚至有的学者试图打破竞争与正相互作用的清晰界限(Warren et al ., 2014 ).在逐渐深入研究植物间相互作用的过程中, 植物地下根系间的竞争及其生态过程更受关注(Sutherland et al ., 2013 ).本文在前人研究的基础上, 从竞争模式角度着重对植物竞争(包括地上竞争和地下竞争)的对称性和非对称性进行综述和分析, 以期为国内相关研究提供参考和借鉴. ...
... 其中, r 是n 个植物体占有的总资源量, bi 代表种群中某植物个体i 的生物量, ri 代表植物个体i占有的资源量, 参数θ 的取值大小反映了竞争模式.当θ 取值为0或趋近于∞时, 植物间分别以完全对称性竞争和完全非对称性竞争方式分配资源; 当在区间(0, ∞)取值时, 则代表植物间竞争模式从对称性竞争向非对称性竞争的连续性过渡.该模型奠定了研究竞争模式的基础, 此后的许多实验或理论研究均运用此模型或以此模型为基础解决生态学竞争领域内的一些复杂问题(Chu et al ., 2008 , 2009, 2010 ; Lin et al ., 2012 ).但这一模型仅仅是对对称性和非对称性竞争概念的界定, 如用于实际研究, 则显得过于简单, 以至于会忽略掉很多影响植物间竞争模式的重要因子, 譬如物种特异性等. ...
Effects of regional origin and genotype on intraspecific root communication in the desert shrub Ambrosia dumosa (Asteraceae).
1
1996
... 一系列对草本和灌木物种的研究(Mahall & Callaway, 1996 ; de Kroon et al., 2003 ; Falik et al ., 2003 ; Holzapfel & Alpert, 2003 ; Gruntman & Novoplansky, 2004 )发现, 植物的根系能够通过信号感知并且区分“自己”的和“非己”的根系.这对植物的生长是有利的, 同境中生长的相邻两株植物, 每株植物可以通过化学信号感知周围另一株植物根系的存在, 并且抑制自己根系向对方根系生长所占区域延伸, 尽量避免自己受到伤害. ...
Exclusion of root competition increases competitive abilities of subordinate plant species through root-shoot interactions.
1
2012
... 植物体的这一切通过毒素和信号物质提高自身竞争能力的生存方式和策略, 不仅能有效地保证其在较恶劣环境中存活与生长, 而且会因为某一种群或群落中目标植物与邻体植物竞争能力差异的改变而导致个体间竞争类型发生改变(Mariotte et al., 2012 ).例如, 均产生毒素的目标植物与邻体植物, 如果两者产生的毒素对对方的影响效应基本相同, 那么目标植物与邻体植物间可能因竞争能力相同而形成对称性竞争的竞争模式; 若两者产生的毒素对对方影响效应显著不同, 则可能形成非对称性竞争的竞争模式. ...
Patterns of seed bank and size asymmetry of plant growth across varying sites in the invasive Lantana camara L. (Verbenaceae).
1
2013
... 有关植物竞争类型的理论研究一直是生态学领域的研究重点, 因为这是厘清植物邻体间竞争过程和揭示其竞争机制的必备环节.而且植物竞争类型也是在群落水平上研究植物群落结构组成、生态位分化和物种多样性及其稳定性维系机制的基础.竞争的对称性和非对称性问题同样会影响到一系列以此为基础的科学研究.比如, Hautier等(2009)在《Science》上发表文章认为, 立地环境的富营养化促进了植物对光非对称性竞争的发生, 进而导致物种多样性发生了变化.这主要是由于一些对光竞争处于劣势地位的物种得不到足够的光资源最终死亡.在开展植物对光非对称性竞争研究的同时, 植物根系非对称性竞争的研究也进行得如火如荼.近年来, 越来越多的研究证实在环境胁迫条件下植物根系竞争也为非对称性竞争Rajaniem, 2003 ).基于Hautier等(2009)的研究, 我们可以假设: 在以水分和营养元素为关键限制性资源的干旱荒漠生态系统中, 植物群落多样性也将由根系非对称性竞争所调控.而在植物生物量分配方面, 陆生植物体间的相互作用(竞争和促进)也会影响植物体地上和地下的生物量分配(Deng et al ., 2006 ; Chu et al ., 2008 ), 对植物对称性与非对称性竞争理论的深入研究或许能更清晰地揭示这种响应机理.至于植物个体大小不整齐性随种群密度的增加而增加这一现象(Weiner, 1986 ), 究竟是由于空间格局不一致引起植物密度变化而产生的Thomas & Weiner, 1989 ), 还是由于植物体的非对称性竞争产生的, 或者是植物本身具有较强的可塑性而导致的, 需要生态学家对其相关理论进一步深入研究.物种共存、生物入侵以及生物多样性等问题也与植物体间非对称性竞争有着千丝万缕的联系(Osunkoya et al ., 2013 ; Skácelová & Lepš, 2014 ), 研究非对称性竞争的产生机制和生理原理对开展相关工作具有重要意义. ...
Evidence for size asymmetry of belowground competition.
3
2003
... 有关植物竞争类型的理论研究一直是生态学领域的研究重点, 因为这是厘清植物邻体间竞争过程和揭示其竞争机制的必备环节.而且植物竞争类型也是在群落水平上研究植物群落结构组成、生态位分化和物种多样性及其稳定性维系机制的基础.竞争的对称性和非对称性问题同样会影响到一系列以此为基础的科学研究.比如, Hautier等(2009)在《Science》上发表文章认为, 立地环境的富营养化促进了植物对光非对称性竞争的发生, 进而导致物种多样性发生了变化.这主要是由于一些对光竞争处于劣势地位的物种得不到足够的光资源最终死亡.在开展植物对光非对称性竞争研究的同时, 植物根系非对称性竞争的研究也进行得如火如荼.近年来, 越来越多的研究证实在环境胁迫条件下植物根系竞争也为非对称性竞争Rajaniem, 2003 ).基于Hautier等(2009)的研究, 我们可以假设: 在以水分和营养元素为关键限制性资源的干旱荒漠生态系统中, 植物群落多样性也将由根系非对称性竞争所调控.而在植物生物量分配方面, 陆生植物体间的相互作用(竞争和促进)也会影响植物体地上和地下的生物量分配(Deng et al ., 2006 ; Chu et al ., 2008 ), 对植物对称性与非对称性竞争理论的深入研究或许能更清晰地揭示这种响应机理.至于植物个体大小不整齐性随种群密度的增加而增加这一现象(Weiner, 1986 ), 究竟是由于空间格局不一致引起植物密度变化而产生的Thomas & Weiner, 1989 ), 还是由于植物体的非对称性竞争产生的, 或者是植物本身具有较强的可塑性而导致的, 需要生态学家对其相关理论进一步深入研究.物种共存、生物入侵以及生物多样性等问题也与植物体间非对称性竞争有着千丝万缕的联系(Osunkoya et al ., 2013 ; Skácelová & Lepš, 2014 ), 研究非对称性竞争的产生机制和生理原理对开展相关工作具有重要意义. ...
... 早期的竞争理论认为: 植物体地上部分主要竞争光资源, 属于非对称性竞争; 地下部分主要竞争水肥资源, 属于对称性竞争(Weiner, 1986 ; Weiner et al ., 1997 ; Schwinning & Weiner, 1998 ; Zhang et al ., 2013 ).其原理是: 高大植物凭借株高优势遮挡周围的矮小植物, 阻碍其获得充足的光照, 而矮小植物对高大植物的光吸收影响不大, 因此, 植物光竞争为非对称性竞争Weiner & Thomas, 1986 ; Connoily & Wayne, 1996 ); 由于地下水肥资源的流动特性, 使不同个体大小的植物均能得到资源, 因此植物水肥竞争为对称性竞争Bolte & Villanueva, 2006 ).但近年也有研究Rajaniem, 2003 ; Rewald & Leuschner, 2009 ; Lei et al ., 2012 ; Weremijewicz & Janos, 2013 )发现, 地下竞争为非对称性竞争或存在非对称性竞争倾向.比如, Rajaniemi (2003)研究发现, 植物在不同种群密度条件下, 目标植物周围存在不同大小的植物时, 较大的植物对目标植物单位生物量的影响大于较小植物对目标植物的影响, 并证明地下竞争可能为非对称性竞争.而Rewald和Leuschner (2009)以水青冈属(Fagus )和栎属(Quercus )为研究对象, 将野外生长的两树种的部分根系非破坏性挖出, 插进设计好的根生长室中, 使这部分根系发生种内、种间竞争作用, 并给予不同的水分梯度处理, 研究表明植物根系的非对称性竞争还会受到水分梯度的影响, 即植物间的非对称性竞争程度随水分胁迫梯度的增加而增加.植物根系间发生非对称性竞争现象的作用机理通常可以解释为: 生长于同一环境中的两株大小不同的植物, 较大的植物根系伸展范围广, 根系接触土壤中营养元素的表面积大, 获取的水肥资源多, 进而促进植物生长, 而进一步的生长又会使其根系伸展范围更广, 根系接触土壤中营养元素的表面积更大, 吸收消耗土壤中水分营养元素资源的能力更强.在土壤中资源有限的情况下, 大个体植物因其具有更强的吸收消耗能力而占有更多小个体根际周围的水肥资源, 进而导致小个体植物根际周围水肥资源的供应速率越来越低, 加剧了对小个体植物生长的限制.因此, 随着植物生长, 小个体与大个体间对土壤中有限资源的竞争能力和资源的占有量会随着植物的生长而进一步扩大, 最终同样可能导致小个体因资源的枯竭而死亡, 形成了典型的根系非对称性竞争.此外, 也有研究者认为, 在同等环境条件下, 植物个体仅仅通过对有限资源的抢先占有, 便能在竞争中胜出(Craine et al ., 2005 ), 也就是说, 植物个体对有限资源获取的起始时间不一致, 导致了对资源的非对称性竞争. ...
... 土壤营养异质性会加剧土壤斑块中植物间的竞争程度(Rajaniemi, 2011 ).Rajaniemi通过研究无芒雀麦(Bromus inermis )发现了异质性土壤中生长的植物根系具有非对称性竞争现象(Rajaniemi, 2003 ); Facelli和Facelli (2002)通过研究三叶草属植物Trifolium subterraneum 发现了类似现象.土壤异质性在多数情况下表现为土壤肥力不均匀, 肥力较丰富区域中的植物生长较快, 而周围能与其发生相互作用的生长在肥力较贫瘠土壤中的植物生长较慢, 两者间的植物个体大小差异随生长过程逐渐凸显, 当因密度效应发生地上光竞争时, 土壤肥力丰富地区的植物占据优势地位, 地上和地下同时存在的优势使得肥力丰富地区与贫瘠地区的植物逐渐形成非对称性竞争关系. ...
Competition for patchy soil resources reduces community evenness.
1
2011
... 土壤营养异质性会加剧土壤斑块中植物间的竞争程度(Rajaniemi, 2011 ).Rajaniemi通过研究无芒雀麦(Bromus inermis )发现了异质性土壤中生长的植物根系具有非对称性竞争现象(Rajaniemi, 2003 ); Facelli和Facelli (2002)通过研究三叶草属植物Trifolium subterraneum 发现了类似现象.土壤异质性在多数情况下表现为土壤肥力不均匀, 肥力较丰富区域中的植物生长较快, 而周围能与其发生相互作用的生长在肥力较贫瘠土壤中的植物生长较慢, 两者间的植物个体大小差异随生长过程逐渐凸显, 当因密度效应发生地上光竞争时, 土壤肥力丰富地区的植物占据优势地位, 地上和地下同时存在的优势使得肥力丰富地区与贫瘠地区的植物逐渐形成非对称性竞争关系. ...
Root competition can cause a decline in diversity with increased productivity.
1
2003
... 植物竞争类型的研究方兴未艾, 并有繁荣发展的势头, 尤其是关于植物地下竞争类型的研究, 更是发展迅速, 有研究认为地下竞争与物种多样性密切相关(Rajaniemi et al ., 2003 ).过去由于探测和测量仪器的局限性, 人工挖掘又会导致部分根系损失, 所以很难准确得到植物地下部分的相关指标和参数, 开展地下竞争的实验和研究也相对较少.但是近年来, 随着科技水平的提升以及一批根系测量与分析系统的完善和发展, 不仅可以准确地测得根系的长度、直径、表面积, 还可以进行根尖计数, 分叉和交叠计数, 使得开展地下竞争研究越来越容易.植物体的地下世界逐渐被发掘, 地下竞争关系越来越清晰.地下竞争或许比地上竞争更加重要, 因为植物体地上部分所需的主要资源是取之不尽用之不竭的光能, 而深入开展植物体地下部分的竞争研究, 尤其是地下部分的非对称性竞争研究, 能帮助人们更加明确对植物体的水肥投入的重要性, 节约营养资源, 增大投入产出效益; 同时也会使人们懂得如何更好地维护整个生态系统, 拯救濒临灭绝的物种, 保证生物圈的物种多样性. ...
More than biofuel? Jatropha curcas root system symmetry and potential for soil erosion control.
1
2011
... 检验植物体竞争对称性与非对称性的理论模型最主要和最直接的方法就是野外实验或控制实验.通常实验中具体的操作方法是测量生物量(Weiner, 1986 )、株高(Zhi et al ., 2007 )、冠幅(Iponga et al ., 2008 )、叶面积(Zhi et al ., 2007 )、根长(Reubens et al ., 2011 )、根表面积(Rewald & Leuschner, 2009 )等指标, 根据这些指标可以计算竞争强度(competition intensity)、竞争重要性(competition importance)和竞争反应(competition response)等参数(王平等, 2007 ).而这些参数的具体计算方法又有很大不同, 如根据计算竞争强度是否随生产力变化, 可将竞争强度分为相对竞争强度(relative competition intensity)和绝对竞争强度(absolute competition intensity) (Grace, 1995 ); 此外, 还可以根据植物地上和地下生长环境不同计算地上竞争强度、地下竞争强度以及总竞争强度, 实际运用中应根据实验设计和实验目的选择合适的参数, 各个参数的计算主要通过实施对照实验并分析植物体在有无竞争情况下的差异来进行. ...
Does root competition asymmetry increase with water availability?
2
2009
... 早期的竞争理论认为: 植物体地上部分主要竞争光资源, 属于非对称性竞争; 地下部分主要竞争水肥资源, 属于对称性竞争(Weiner, 1986 ; Weiner et al ., 1997 ; Schwinning & Weiner, 1998 ; Zhang et al ., 2013 ).其原理是: 高大植物凭借株高优势遮挡周围的矮小植物, 阻碍其获得充足的光照, 而矮小植物对高大植物的光吸收影响不大, 因此, 植物光竞争为非对称性竞争Weiner & Thomas, 1986 ; Connoily & Wayne, 1996 ); 由于地下水肥资源的流动特性, 使不同个体大小的植物均能得到资源, 因此植物水肥竞争为对称性竞争Bolte & Villanueva, 2006 ).但近年也有研究Rajaniem, 2003 ; Rewald & Leuschner, 2009 ; Lei et al ., 2012 ; Weremijewicz & Janos, 2013 )发现, 地下竞争为非对称性竞争或存在非对称性竞争倾向.比如, Rajaniemi (2003)研究发现, 植物在不同种群密度条件下, 目标植物周围存在不同大小的植物时, 较大的植物对目标植物单位生物量的影响大于较小植物对目标植物的影响, 并证明地下竞争可能为非对称性竞争.而Rewald和Leuschner (2009)以水青冈属(Fagus )和栎属(Quercus )为研究对象, 将野外生长的两树种的部分根系非破坏性挖出, 插进设计好的根生长室中, 使这部分根系发生种内、种间竞争作用, 并给予不同的水分梯度处理, 研究表明植物根系的非对称性竞争还会受到水分梯度的影响, 即植物间的非对称性竞争程度随水分胁迫梯度的增加而增加.植物根系间发生非对称性竞争现象的作用机理通常可以解释为: 生长于同一环境中的两株大小不同的植物, 较大的植物根系伸展范围广, 根系接触土壤中营养元素的表面积大, 获取的水肥资源多, 进而促进植物生长, 而进一步的生长又会使其根系伸展范围更广, 根系接触土壤中营养元素的表面积更大, 吸收消耗土壤中水分营养元素资源的能力更强.在土壤中资源有限的情况下, 大个体植物因其具有更强的吸收消耗能力而占有更多小个体根际周围的水肥资源, 进而导致小个体植物根际周围水肥资源的供应速率越来越低, 加剧了对小个体植物生长的限制.因此, 随着植物生长, 小个体与大个体间对土壤中有限资源的竞争能力和资源的占有量会随着植物的生长而进一步扩大, 最终同样可能导致小个体因资源的枯竭而死亡, 形成了典型的根系非对称性竞争.此外, 也有研究者认为, 在同等环境条件下, 植物个体仅仅通过对有限资源的抢先占有, 便能在竞争中胜出(Craine et al ., 2005 ), 也就是说, 植物个体对有限资源获取的起始时间不一致, 导致了对资源的非对称性竞争. ...
... 检验植物体竞争对称性与非对称性的理论模型最主要和最直接的方法就是野外实验或控制实验.通常实验中具体的操作方法是测量生物量(Weiner, 1986 )、株高(Zhi et al ., 2007 )、冠幅(Iponga et al ., 2008 )、叶面积(Zhi et al ., 2007 )、根长(Reubens et al ., 2011 )、根表面积(Rewald & Leuschner, 2009 )等指标, 根据这些指标可以计算竞争强度(competition intensity)、竞争重要性(competition importance)和竞争反应(competition response)等参数(王平等, 2007 ).而这些参数的具体计算方法又有很大不同, 如根据计算竞争强度是否随生产力变化, 可将竞争强度分为相对竞争强度(relative competition intensity)和绝对竞争强度(absolute competition intensity) (Grace, 1995 ); 此外, 还可以根据植物地上和地下生长环境不同计算地上竞争强度、地下竞争强度以及总竞争强度, 实际运用中应根据实验设计和实验目的选择合适的参数, 各个参数的计算主要通过实施对照实验并分析植物体在有无竞争情况下的差异来进行. ...
Tropical tree diversity enhances light capture through crown plasticity and spatial and temporal niche differences.
1
2014
... 植物的形态可塑性常常通过改变植物地上和地下的生物量分配、增大某部分器官的表面积等途径来影响植物间的竞争关系(Schwinning & Weiner, 1998 ).当土壤中水肥资源受限时, 植物就竭力分配生物量给地下(通常干旱区植物有发达的根系), 同时增大根系与土壤接触的表面积, 竭力使植物体在水肥资源竞争中占优势.所以当两株相邻植物获取资源的能力相差不大时, 处于劣势的植物很可能通过形态可塑性, 分配更多生物量给地下, 藉此来弥补根系所处的劣势.但当两株植物的相对优势很明显时, 形态可塑性无法弥补劣势植物的劣势, 致使其无法得到足够的资源, 也就无法快速生长, 陷入了恶性循环, 而优势植物由于获得充足的资源, 可以进一步生长, 根系更加强大, 获取资源能力更强, 形成了良性循环, 两株植物间根系的非对称性竞争模式就形成了.形态可塑性在减弱植物个体间非对称性竞争中的作用已经得到一系列转基因实验的证明(Ballaré et al ., 1994 ; Ballaré & Scopel, 1997 ).同样, 形态可塑性也会影响植物体地上部分对光资源的竞争.研究发现, 植物可塑性和时空上的生态位分化能够提高植物获取光的能力并维持生物多样性的积极影响(Sapijanskas et al ., 2014 ).生物多样性局面的维持表明物种间可以共存, 植物可塑性等因素使得同生境中生长的为光资源而发生激烈非对称性竞争的植物能够实现共存. ...
Root competition: Beyond resource depletion.
1
2006
... 研究发现一些植物能够通过化感作用产生有毒物质, 限制周围植物生长甚至致其死亡(Schenk, 2006 ).通常不同物种间会出现这种情况, 一种植物根系分泌物会影响另一种植物生长.但也有一些物种成年植物体释放化学物质影响和抑制其幼苗个体生长, 例如银桦(Grevillea robusta ).除此之外, 一些植物根系还会释放一种对其他植物无害的化学物质, 帮助植物感知周围其他植物根系的生长及土壤中营养物质等非生命物质的存在.研究发现, 一些豆科植物根系能通过“化学雷达”而不用物理接触就能感知其周围的物理障碍, 从而改变根的生长形态(Falik et al ., 2005 ).资源获取器官若能够感知非生命物质的存在, 在生长中就占据很大优势.例如, 植物根系若能感知土壤中肥力不足的区域或不利于自身生长的因素, 就不朝肥力不足区域生长而转向肥力丰富区域生长或避开不利于自身生长的因素, 从而保证植物体快速生长, 增强其在逆境中的生存竞争能力. ...
Population dynamic consequences of competitive symmetry in annual plants.
0
1995
Mechanisms determining the degree of size asymmetry in competition among plants.
5
1998
... 大小依赖的对称性竞争是指存在资源竞争的植物间依据植物个体大小来分配资源, 即某株植物的生物量占全部植物总生物量的百分比就是其获得资源的百分比(Schwinning & Weiner, 1998 ; Weiner et al ., 2001 ).例如, 3株植物的生物量分别是B 1 、B 2 、B 3 , 总资源量为A, 则3株植物占有的资源量分别为: AB 1 /( B 1 + B 2 + B 3 )、AB 2 /( B 1 + B 2 + B 3 )、AB 3 /( B 1 + B 2 + B 3 ).植物体由幼苗生长至成熟个体的过程中, 可以表征个体大小的生物量逐渐增大, 如果其竞争模式为大小依赖的对称性竞争, 则其占有的资源量将逐渐增多, 竞争能力逐渐增强.完全对称性竞争是指植物在生长过程中竞争资源时, 根据特定区域内的植株个体数平均分配该区域内的资源Schwin- ning & Weiner, 1998 ).例如, 某一区域内有n 株植物, 则每株植物均分得该区域内资源总量的1/n . ...
... ).植物体由幼苗生长至成熟个体的过程中, 可以表征个体大小的生物量逐渐增大, 如果其竞争模式为大小依赖的对称性竞争, 则其占有的资源量将逐渐增多, 竞争能力逐渐增强.完全对称性竞争是指植物在生长过程中竞争资源时, 根据特定区域内的植株个体数平均分配该区域内的资源Schwin- ning & Weiner, 1998 ).例如, 某一区域内有n 株植物, 则每株植物均分得该区域内资源总量的1/n . ...
... 大小依赖的非对称性竞争是指处于竞争状态的植物间不均衡分配资源, 不按植物个体大小比例分配资源, 而是较大的植物占据大于相应生物量比例的资源, 较小的植物占据小于相应生物量比例的资源Schwinning & Weiner, 1998 ).例如, 两株植物生物量比例为1:3, 但资源分配可能是1:4或1:5.这种不均衡分配, 导致较大植物在下一阶段生长中占有更多资源, 而较小植物占有更少资源, 就像经济学中经常提到的“马太效应”一样, 强者愈强, 弱者愈弱, 最终致使较小植物因得不到足够的资源而死亡.这种具有“马太效应”的非对称性竞争的极端形式便是完全非对称性竞争, 即较大植物占有所有被竞争的资源(contested resource), 而较小植物不占有任何资源, 最终只能因资源枯竭而死亡和被淘汰. ...
... 早期的竞争理论认为: 植物体地上部分主要竞争光资源, 属于非对称性竞争; 地下部分主要竞争水肥资源, 属于对称性竞争(Weiner, 1986 ; Weiner et al ., 1997 ; Schwinning & Weiner, 1998 ; Zhang et al ., 2013 ).其原理是: 高大植物凭借株高优势遮挡周围的矮小植物, 阻碍其获得充足的光照, 而矮小植物对高大植物的光吸收影响不大, 因此, 植物光竞争为非对称性竞争Weiner & Thomas, 1986 ; Connoily & Wayne, 1996 ); 由于地下水肥资源的流动特性, 使不同个体大小的植物均能得到资源, 因此植物水肥竞争为对称性竞争Bolte & Villanueva, 2006 ).但近年也有研究Rajaniem, 2003 ; Rewald & Leuschner, 2009 ; Lei et al ., 2012 ; Weremijewicz & Janos, 2013 )发现, 地下竞争为非对称性竞争或存在非对称性竞争倾向.比如, Rajaniemi (2003)研究发现, 植物在不同种群密度条件下, 目标植物周围存在不同大小的植物时, 较大的植物对目标植物单位生物量的影响大于较小植物对目标植物的影响, 并证明地下竞争可能为非对称性竞争.而Rewald和Leuschner (2009)以水青冈属(Fagus )和栎属(Quercus )为研究对象, 将野外生长的两树种的部分根系非破坏性挖出, 插进设计好的根生长室中, 使这部分根系发生种内、种间竞争作用, 并给予不同的水分梯度处理, 研究表明植物根系的非对称性竞争还会受到水分梯度的影响, 即植物间的非对称性竞争程度随水分胁迫梯度的增加而增加.植物根系间发生非对称性竞争现象的作用机理通常可以解释为: 生长于同一环境中的两株大小不同的植物, 较大的植物根系伸展范围广, 根系接触土壤中营养元素的表面积大, 获取的水肥资源多, 进而促进植物生长, 而进一步的生长又会使其根系伸展范围更广, 根系接触土壤中营养元素的表面积更大, 吸收消耗土壤中水分营养元素资源的能力更强.在土壤中资源有限的情况下, 大个体植物因其具有更强的吸收消耗能力而占有更多小个体根际周围的水肥资源, 进而导致小个体植物根际周围水肥资源的供应速率越来越低, 加剧了对小个体植物生长的限制.因此, 随着植物生长, 小个体与大个体间对土壤中有限资源的竞争能力和资源的占有量会随着植物的生长而进一步扩大, 最终同样可能导致小个体因资源的枯竭而死亡, 形成了典型的根系非对称性竞争.此外, 也有研究者认为, 在同等环境条件下, 植物个体仅仅通过对有限资源的抢先占有, 便能在竞争中胜出(Craine et al ., 2005 ), 也就是说, 植物个体对有限资源获取的起始时间不一致, 导致了对资源的非对称性竞争. ...
... 植物的形态可塑性常常通过改变植物地上和地下的生物量分配、增大某部分器官的表面积等途径来影响植物间的竞争关系(Schwinning & Weiner, 1998 ).当土壤中水肥资源受限时, 植物就竭力分配生物量给地下(通常干旱区植物有发达的根系), 同时增大根系与土壤接触的表面积, 竭力使植物体在水肥资源竞争中占优势.所以当两株相邻植物获取资源的能力相差不大时, 处于劣势的植物很可能通过形态可塑性, 分配更多生物量给地下, 藉此来弥补根系所处的劣势.但当两株植物的相对优势很明显时, 形态可塑性无法弥补劣势植物的劣势, 致使其无法得到足够的资源, 也就无法快速生长, 陷入了恶性循环, 而优势植物由于获得充足的资源, 可以进一步生长, 根系更加强大, 获取资源能力更强, 形成了良性循环, 两株植物间根系的非对称性竞争模式就形成了.形态可塑性在减弱植物个体间非对称性竞争中的作用已经得到一系列转基因实验的证明(Ballaré et al ., 1994 ; Ballaré & Scopel, 1997 ).同样, 形态可塑性也会影响植物体地上部分对光资源的竞争.研究发现, 植物可塑性和时空上的生态位分化能够提高植物获取光的能力并维持生物多样性的积极影响(Sapijanskas et al ., 2014 ).生物多样性局面的维持表明物种间可以共存, 植物可塑性等因素使得同生境中生长的为光资源而发生激烈非对称性竞争的植物能够实现共存. ...
The relationship of diversity and biomass in phytoplankton communities weakens when accounting for species proportions.
1
2014
... 有关植物竞争类型的理论研究一直是生态学领域的研究重点, 因为这是厘清植物邻体间竞争过程和揭示其竞争机制的必备环节.而且植物竞争类型也是在群落水平上研究植物群落结构组成、生态位分化和物种多样性及其稳定性维系机制的基础.竞争的对称性和非对称性问题同样会影响到一系列以此为基础的科学研究.比如, Hautier等(2009)在《Science》上发表文章认为, 立地环境的富营养化促进了植物对光非对称性竞争的发生, 进而导致物种多样性发生了变化.这主要是由于一些对光竞争处于劣势地位的物种得不到足够的光资源最终死亡.在开展植物对光非对称性竞争研究的同时, 植物根系非对称性竞争的研究也进行得如火如荼.近年来, 越来越多的研究证实在环境胁迫条件下植物根系竞争也为非对称性竞争Rajaniem, 2003 ).基于Hautier等(2009)的研究, 我们可以假设: 在以水分和营养元素为关键限制性资源的干旱荒漠生态系统中, 植物群落多样性也将由根系非对称性竞争所调控.而在植物生物量分配方面, 陆生植物体间的相互作用(竞争和促进)也会影响植物体地上和地下的生物量分配(Deng et al ., 2006 ; Chu et al ., 2008 ), 对植物对称性与非对称性竞争理论的深入研究或许能更清晰地揭示这种响应机理.至于植物个体大小不整齐性随种群密度的增加而增加这一现象(Weiner, 1986 ), 究竟是由于空间格局不一致引起植物密度变化而产生的Thomas & Weiner, 1989 ), 还是由于植物体的非对称性竞争产生的, 或者是植物本身具有较强的可塑性而导致的, 需要生态学家对其相关理论进一步深入研究.物种共存、生物入侵以及生物多样性等问题也与植物体间非对称性竞争有着千丝万缕的联系(Osunkoya et al ., 2013 ; Skácelová & Lepš, 2014 ), 研究非对称性竞争的产生机制和生理原理对开展相关工作具有重要意义. ...
Asymmetric relationship between grasses and forbs: Results from a field experiment under nutrient limitation.
1
2013
... 如上所述, 研究生物体间的非对称性竞争问题可以与生物的共存、生物多样性、生物入侵等问题相联系.在一定的条件下, 生物多样性降低很可能是由物种间非对称性竞争而造成的(Šmilauer & Šmilauerová, 2013 ).在相同生境中两物种间出现完全的非对称性竞争时, 一物种完全处于竞争优势地位, 而另一物种不占据任何资源优势, 在竞争中逐渐衰败, 最终死亡.因此, 当物种间出现非对称性竞争时, 物种无法共存, 它们所处的生态系统生物多样性会下降, 生物入侵容易发生.而这几个方面恰恰与土壤异质性有关, 或者说土壤异质性可以部分解释物种共存、生物多样性和生物入侵等问题.首先, 土壤异质性与物种入侵间可以发生相互作用和反馈调节.已有证据表明, 外来物种入侵会显著改变生态系统N循环, 入侵物种增加了N含量, 而且加快了N的流动循环Castro-Díez et al ., 2014 ).因此, 随着物种入侵, 土壤中营养元素的成分和含量会发生大的变化, 致使土壤异质性加强.同时, 物种入侵也会受土壤异质性影响.Burns和Brandt (2014)研究发现, 土壤异质性与种群周转率相互作用影响物种入侵, 增加土壤异质性会增加物种入侵.两者间的相互作用关系启示我们在研究物种间非对称性竞争问题时应与生物入侵问题相联系.其次, 土壤异质性会影响生物多样性, 多数情况下土壤异质性会增加生物多样性(Belfrage et al ., 2015 ).从生态位角度考虑, 土壤异质性可以为物种提供更多的生态位, 适应不同物种对不同生境的需求, 而且不同的生态位缓解了物种间对各种资源的强烈竞争, 使生物出现共存局面, 最终可能表现为植物间较弱的非对称性竞争或转变为对称性竞争.但是, 近来研究发现, 土壤异质性高并不意味着生物多样性也升高, 例如当土壤中某种元素含量很高而出现富营养化时, 情况会大不相同.已有研究表明土壤富营养化会削弱生物多样性产生的稳定性效应(Hautier et al ., 2014 ).也就是说, 物种稳定共存的局面不会长期维持下去, 总有一些物种会在竞争中被淘汰掉.因此, 土壤异质性升高、物种多样性降低的环境中, 物种间易于出现非对称性竞争. ...
Identification of 100 fundamental ecological questions.
1
2013
... 竞争是陆生植物生长过程中的重要现象, 竞争作用是群落结构组建的主导因子和决定物种进化模式的重要因素(王刚和张大勇, 1996 ).对物种竞争的研究由来已久, 19世纪50年代末, 达尔文曾指出生活需求相似的近缘物种间经常发生激烈的竞争.随后, 洛特和沃尔特拉用数学公式描述了物种间的竞争关系.高斯假说从生态位角度展示了物种竞争的结果.竞争的研究领域涉及个体、种群、群落、生态系统等不同层次, 如竞争对个体大小不整齐性的影响(Weiner & Thomas, 1986 )、竞争对种群数量的调节(Deng et al ., 2006 )、竞争与群落空间分布格局的关系(Lin et al ., 2012 )、竞争在维持生态系统物种多样性中的作用(Borer et al ., 2014 )等等.随着植物间相互作用研究的不断拓展, 不断有新的概念和理论模型被提出, 甚至有的学者试图打破竞争与正相互作用的清晰界限(Warren et al ., 2014 ).在逐渐深入研究植物间相互作用的过程中, 植物地下根系间的竞争及其生态过程更受关注(Sutherland et al ., 2013 ).本文在前人研究的基础上, 从竞争模式角度着重对植物竞争(包括地上竞争和地下竞争)的对称性和非对称性进行综述和分析, 以期为国内相关研究提供参考和借鉴. ...
Local density variation may mimic effects of asymmetric competition on plant size variability.
1
1989
... 有关植物竞争类型的理论研究一直是生态学领域的研究重点, 因为这是厘清植物邻体间竞争过程和揭示其竞争机制的必备环节.而且植物竞争类型也是在群落水平上研究植物群落结构组成、生态位分化和物种多样性及其稳定性维系机制的基础.竞争的对称性和非对称性问题同样会影响到一系列以此为基础的科学研究.比如, Hautier等(2009)在《Science》上发表文章认为, 立地环境的富营养化促进了植物对光非对称性竞争的发生, 进而导致物种多样性发生了变化.这主要是由于一些对光竞争处于劣势地位的物种得不到足够的光资源最终死亡.在开展植物对光非对称性竞争研究的同时, 植物根系非对称性竞争的研究也进行得如火如荼.近年来, 越来越多的研究证实在环境胁迫条件下植物根系竞争也为非对称性竞争Rajaniem, 2003 ).基于Hautier等(2009)的研究, 我们可以假设: 在以水分和营养元素为关键限制性资源的干旱荒漠生态系统中, 植物群落多样性也将由根系非对称性竞争所调控.而在植物生物量分配方面, 陆生植物体间的相互作用(竞争和促进)也会影响植物体地上和地下的生物量分配(Deng et al ., 2006 ; Chu et al ., 2008 ), 对植物对称性与非对称性竞争理论的深入研究或许能更清晰地揭示这种响应机理.至于植物个体大小不整齐性随种群密度的增加而增加这一现象(Weiner, 1986 ), 究竟是由于空间格局不一致引起植物密度变化而产生的Thomas & Weiner, 1989 ), 还是由于植物体的非对称性竞争产生的, 或者是植物本身具有较强的可塑性而导致的, 需要生态学家对其相关理论进一步深入研究.物种共存、生物入侵以及生物多样性等问题也与植物体间非对称性竞争有着千丝万缕的联系(Osunkoya et al ., 2013 ; Skácelová & Lepš, 2014 ), 研究非对称性竞争的产生机制和生理原理对开展相关工作具有重要意义. ...
Caught in the middle: Asymmetric competition causes high variance in intermediate trait abundances.
1
2013
... 尽管地下竞争通常大于地上竞争, 尤其是在营养元素贫瘠的土壤中Casper & Jackson, 1997 ; Kiær et al ., 2013 ), 但地上竞争也不可忽略.影响植物地上竞争的主要因素是光照, 绿色植物进行正常的光合作用需要充足的光照条件, 而在较为拥挤的植物种群或群落中, 植物个体间因相互遮阴而发生光竞争, 高大植物会遮挡其冠下植物对光的吸收, 但冠下植物对上层植物光吸收的影响较小.因此, 植物个体间对光的竞争是单方面的, 随着植物的生长, 高大植物的遮阴效果会越来越强, 形成典型的非对称性竞争, 即由于植物体个体大小不同(表现为株高不同)导致上层植物与冠下植物间对光的竞争为非对称性竞争van Leeuwen & Etienne, 2013 ).这种关于地上竞争的非对称性理论还仅局限于将光照作为一个笼统的概念进行整体分析.在未来的研究中可以对光资源进一步细化分析. ...
2
1996
... 竞争是陆生植物生长过程中的重要现象, 竞争作用是群落结构组建的主导因子和决定物种进化模式的重要因素(王刚和张大勇, 1996 ).对物种竞争的研究由来已久, 19世纪50年代末, 达尔文曾指出生活需求相似的近缘物种间经常发生激烈的竞争.随后, 洛特和沃尔特拉用数学公式描述了物种间的竞争关系.高斯假说从生态位角度展示了物种竞争的结果.竞争的研究领域涉及个体、种群、群落、生态系统等不同层次, 如竞争对个体大小不整齐性的影响(Weiner & Thomas, 1986 )、竞争对种群数量的调节(Deng et al ., 2006 )、竞争与群落空间分布格局的关系(Lin et al ., 2012 )、竞争在维持生态系统物种多样性中的作用(Borer et al ., 2014 )等等.随着植物间相互作用研究的不断拓展, 不断有新的概念和理论模型被提出, 甚至有的学者试图打破竞争与正相互作用的清晰界限(Warren et al ., 2014 ).在逐渐深入研究植物间相互作用的过程中, 植物地下根系间的竞争及其生态过程更受关注(Sutherland et al ., 2013 ).本文在前人研究的基础上, 从竞争模式角度着重对植物竞争(包括地上竞争和地下竞争)的对称性和非对称性进行综述和分析, 以期为国内相关研究提供参考和借鉴. ...
... 迄今为止, 有关植物竞争的概念和类型有很多, 它们之间既有很多共性, 也存在很大差异.根据物种差异、地上地下和竞争模式, 植物竞争类型大致可分为: 种内竞争(intraspecific competition)和种间竞争(interspecific competition)、地上竞争(above- ground competition)和地下竞争(belowground competition) (Deng et al ., 2006 ; 王平等, 2007 )、对称性竞争(symmetric competition)和非对称性竞争(asymm- etric competition) (王刚和张大勇, 1996 ).对称性竞争又可再分为完全对称性竞争和大小依赖的对称性竞争; 非对称性竞争也可再分为完全非对称性竞争和大小依赖的非对称性竞争.种内竞争和种间竞争是基于植物物种的类型进行分类, 强调物种差异对竞争的影响; 而地上竞争和地下竞争的划分主要是基于植物体地上冠层对光资源的竞争和地下根系部分对水和营养元素的竞争, 打破了物种差异的明确界限.从某种意义上讲, 地上、地下竞争将植物竞争的研究范围进一步扩大了, 使得有关植物竞争的研究结论更具普适性.实际上, 对称性竞争与非对称性竞争同样涵盖了植物地上竞争和地下竞争, 但对称性竞争与非对称性竞争从概念上将植物竞争研究的内涵进一步深化, 体现了植物竞争的具体性质; 而地上竞争和地下竞争仅体现为植物竞争的形式.理论上, 地上竞争和地下竞争都可以是对称性或非对称性的; 反之, 植物的对称性竞争与非对称性竞争既可发生在植物的地上部分, 也可发生在植物的地下部分.然而, 对称性竞争与非对称性竞争更加强调了植物最大化获取资源的策略, 从一个新颖的角度阐释环境资源有限的情况下植物个体间分配资源的方式.近年来, 对称性竞争与非对称性竞争的研究逐渐成为热点, 下面将进一步阐释它们的概念及其内涵. ...
2
1996
... 竞争是陆生植物生长过程中的重要现象, 竞争作用是群落结构组建的主导因子和决定物种进化模式的重要因素(王刚和张大勇, 1996 ).对物种竞争的研究由来已久, 19世纪50年代末, 达尔文曾指出生活需求相似的近缘物种间经常发生激烈的竞争.随后, 洛特和沃尔特拉用数学公式描述了物种间的竞争关系.高斯假说从生态位角度展示了物种竞争的结果.竞争的研究领域涉及个体、种群、群落、生态系统等不同层次, 如竞争对个体大小不整齐性的影响(Weiner & Thomas, 1986 )、竞争对种群数量的调节(Deng et al ., 2006 )、竞争与群落空间分布格局的关系(Lin et al ., 2012 )、竞争在维持生态系统物种多样性中的作用(Borer et al ., 2014 )等等.随着植物间相互作用研究的不断拓展, 不断有新的概念和理论模型被提出, 甚至有的学者试图打破竞争与正相互作用的清晰界限(Warren et al ., 2014 ).在逐渐深入研究植物间相互作用的过程中, 植物地下根系间的竞争及其生态过程更受关注(Sutherland et al ., 2013 ).本文在前人研究的基础上, 从竞争模式角度着重对植物竞争(包括地上竞争和地下竞争)的对称性和非对称性进行综述和分析, 以期为国内相关研究提供参考和借鉴. ...
... 迄今为止, 有关植物竞争的概念和类型有很多, 它们之间既有很多共性, 也存在很大差异.根据物种差异、地上地下和竞争模式, 植物竞争类型大致可分为: 种内竞争(intraspecific competition)和种间竞争(interspecific competition)、地上竞争(above- ground competition)和地下竞争(belowground competition) (Deng et al ., 2006 ; 王平等, 2007 )、对称性竞争(symmetric competition)和非对称性竞争(asymm- etric competition) (王刚和张大勇, 1996 ).对称性竞争又可再分为完全对称性竞争和大小依赖的对称性竞争; 非对称性竞争也可再分为完全非对称性竞争和大小依赖的非对称性竞争.种内竞争和种间竞争是基于植物物种的类型进行分类, 强调物种差异对竞争的影响; 而地上竞争和地下竞争的划分主要是基于植物体地上冠层对光资源的竞争和地下根系部分对水和营养元素的竞争, 打破了物种差异的明确界限.从某种意义上讲, 地上、地下竞争将植物竞争的研究范围进一步扩大了, 使得有关植物竞争的研究结论更具普适性.实际上, 对称性竞争与非对称性竞争同样涵盖了植物地上竞争和地下竞争, 但对称性竞争与非对称性竞争从概念上将植物竞争研究的内涵进一步深化, 体现了植物竞争的具体性质; 而地上竞争和地下竞争仅体现为植物竞争的形式.理论上, 地上竞争和地下竞争都可以是对称性或非对称性的; 反之, 植物的对称性竞争与非对称性竞争既可发生在植物的地上部分, 也可发生在植物的地下部分.然而, 对称性竞争与非对称性竞争更加强调了植物最大化获取资源的策略, 从一个新颖的角度阐释环境资源有限的情况下植物个体间分配资源的方式.近年来, 对称性竞争与非对称性竞争的研究逐渐成为热点, 下面将进一步阐释它们的概念及其内涵. ...
植物地上竞争与地下竞争研究进展
2
2007
... 迄今为止, 有关植物竞争的概念和类型有很多, 它们之间既有很多共性, 也存在很大差异.根据物种差异、地上地下和竞争模式, 植物竞争类型大致可分为: 种内竞争(intraspecific competition)和种间竞争(interspecific competition)、地上竞争(above- ground competition)和地下竞争(belowground competition) (Deng et al ., 2006 ; 王平等, 2007 )、对称性竞争(symmetric competition)和非对称性竞争(asymm- etric competition) (王刚和张大勇, 1996 ).对称性竞争又可再分为完全对称性竞争和大小依赖的对称性竞争; 非对称性竞争也可再分为完全非对称性竞争和大小依赖的非对称性竞争.种内竞争和种间竞争是基于植物物种的类型进行分类, 强调物种差异对竞争的影响; 而地上竞争和地下竞争的划分主要是基于植物体地上冠层对光资源的竞争和地下根系部分对水和营养元素的竞争, 打破了物种差异的明确界限.从某种意义上讲, 地上、地下竞争将植物竞争的研究范围进一步扩大了, 使得有关植物竞争的研究结论更具普适性.实际上, 对称性竞争与非对称性竞争同样涵盖了植物地上竞争和地下竞争, 但对称性竞争与非对称性竞争从概念上将植物竞争研究的内涵进一步深化, 体现了植物竞争的具体性质; 而地上竞争和地下竞争仅体现为植物竞争的形式.理论上, 地上竞争和地下竞争都可以是对称性或非对称性的; 反之, 植物的对称性竞争与非对称性竞争既可发生在植物的地上部分, 也可发生在植物的地下部分.然而, 对称性竞争与非对称性竞争更加强调了植物最大化获取资源的策略, 从一个新颖的角度阐释环境资源有限的情况下植物个体间分配资源的方式.近年来, 对称性竞争与非对称性竞争的研究逐渐成为热点, 下面将进一步阐释它们的概念及其内涵. ...
... 检验植物体竞争对称性与非对称性的理论模型最主要和最直接的方法就是野外实验或控制实验.通常实验中具体的操作方法是测量生物量(Weiner, 1986 )、株高(Zhi et al ., 2007 )、冠幅(Iponga et al ., 2008 )、叶面积(Zhi et al ., 2007 )、根长(Reubens et al ., 2011 )、根表面积(Rewald & Leuschner, 2009 )等指标, 根据这些指标可以计算竞争强度(competition intensity)、竞争重要性(competition importance)和竞争反应(competition response)等参数(王平等, 2007 ).而这些参数的具体计算方法又有很大不同, 如根据计算竞争强度是否随生产力变化, 可将竞争强度分为相对竞争强度(relative competition intensity)和绝对竞争强度(absolute competition intensity) (Grace, 1995 ); 此外, 还可以根据植物地上和地下生长环境不同计算地上竞争强度、地下竞争强度以及总竞争强度, 实际运用中应根据实验设计和实验目的选择合适的参数, 各个参数的计算主要通过实施对照实验并分析植物体在有无竞争情况下的差异来进行. ...
植物地上竞争与地下竞争研究进展
2
2007
... 迄今为止, 有关植物竞争的概念和类型有很多, 它们之间既有很多共性, 也存在很大差异.根据物种差异、地上地下和竞争模式, 植物竞争类型大致可分为: 种内竞争(intraspecific competition)和种间竞争(interspecific competition)、地上竞争(above- ground competition)和地下竞争(belowground competition) (Deng et al ., 2006 ; 王平等, 2007 )、对称性竞争(symmetric competition)和非对称性竞争(asymm- etric competition) (王刚和张大勇, 1996 ).对称性竞争又可再分为完全对称性竞争和大小依赖的对称性竞争; 非对称性竞争也可再分为完全非对称性竞争和大小依赖的非对称性竞争.种内竞争和种间竞争是基于植物物种的类型进行分类, 强调物种差异对竞争的影响; 而地上竞争和地下竞争的划分主要是基于植物体地上冠层对光资源的竞争和地下根系部分对水和营养元素的竞争, 打破了物种差异的明确界限.从某种意义上讲, 地上、地下竞争将植物竞争的研究范围进一步扩大了, 使得有关植物竞争的研究结论更具普适性.实际上, 对称性竞争与非对称性竞争同样涵盖了植物地上竞争和地下竞争, 但对称性竞争与非对称性竞争从概念上将植物竞争研究的内涵进一步深化, 体现了植物竞争的具体性质; 而地上竞争和地下竞争仅体现为植物竞争的形式.理论上, 地上竞争和地下竞争都可以是对称性或非对称性的; 反之, 植物的对称性竞争与非对称性竞争既可发生在植物的地上部分, 也可发生在植物的地下部分.然而, 对称性竞争与非对称性竞争更加强调了植物最大化获取资源的策略, 从一个新颖的角度阐释环境资源有限的情况下植物个体间分配资源的方式.近年来, 对称性竞争与非对称性竞争的研究逐渐成为热点, 下面将进一步阐释它们的概念及其内涵. ...
... 检验植物体竞争对称性与非对称性的理论模型最主要和最直接的方法就是野外实验或控制实验.通常实验中具体的操作方法是测量生物量(Weiner, 1986 )、株高(Zhi et al ., 2007 )、冠幅(Iponga et al ., 2008 )、叶面积(Zhi et al ., 2007 )、根长(Reubens et al ., 2011 )、根表面积(Rewald & Leuschner, 2009 )等指标, 根据这些指标可以计算竞争强度(competition intensity)、竞争重要性(competition importance)和竞争反应(competition response)等参数(王平等, 2007 ).而这些参数的具体计算方法又有很大不同, 如根据计算竞争强度是否随生产力变化, 可将竞争强度分为相对竞争强度(relative competition intensity)和绝对竞争强度(absolute competition intensity) (Grace, 1995 ); 此外, 还可以根据植物地上和地下生长环境不同计算地上竞争强度、地下竞争强度以及总竞争强度, 实际运用中应根据实验设计和实验目的选择合适的参数, 各个参数的计算主要通过实施对照实验并分析植物体在有无竞争情况下的差异来进行. ...
Competition as a mechanism structuring mutualisms.
1
2014
... 竞争是陆生植物生长过程中的重要现象, 竞争作用是群落结构组建的主导因子和决定物种进化模式的重要因素(王刚和张大勇, 1996 ).对物种竞争的研究由来已久, 19世纪50年代末, 达尔文曾指出生活需求相似的近缘物种间经常发生激烈的竞争.随后, 洛特和沃尔特拉用数学公式描述了物种间的竞争关系.高斯假说从生态位角度展示了物种竞争的结果.竞争的研究领域涉及个体、种群、群落、生态系统等不同层次, 如竞争对个体大小不整齐性的影响(Weiner & Thomas, 1986 )、竞争对种群数量的调节(Deng et al ., 2006 )、竞争与群落空间分布格局的关系(Lin et al ., 2012 )、竞争在维持生态系统物种多样性中的作用(Borer et al ., 2014 )等等.随着植物间相互作用研究的不断拓展, 不断有新的概念和理论模型被提出, 甚至有的学者试图打破竞争与正相互作用的清晰界限(Warren et al ., 2014 ).在逐渐深入研究植物间相互作用的过程中, 植物地下根系间的竞争及其生态过程更受关注(Sutherland et al ., 2013 ).本文在前人研究的基础上, 从竞争模式角度着重对植物竞争(包括地上竞争和地下竞争)的对称性和非对称性进行综述和分析, 以期为国内相关研究提供参考和借鉴. ...
How competition for light and nutrients affects size variability in ipomoea tricolor populations.
3
1986
... 有关植物竞争类型的理论研究一直是生态学领域的研究重点, 因为这是厘清植物邻体间竞争过程和揭示其竞争机制的必备环节.而且植物竞争类型也是在群落水平上研究植物群落结构组成、生态位分化和物种多样性及其稳定性维系机制的基础.竞争的对称性和非对称性问题同样会影响到一系列以此为基础的科学研究.比如, Hautier等(2009)在《Science》上发表文章认为, 立地环境的富营养化促进了植物对光非对称性竞争的发生, 进而导致物种多样性发生了变化.这主要是由于一些对光竞争处于劣势地位的物种得不到足够的光资源最终死亡.在开展植物对光非对称性竞争研究的同时, 植物根系非对称性竞争的研究也进行得如火如荼.近年来, 越来越多的研究证实在环境胁迫条件下植物根系竞争也为非对称性竞争Rajaniem, 2003 ).基于Hautier等(2009)的研究, 我们可以假设: 在以水分和营养元素为关键限制性资源的干旱荒漠生态系统中, 植物群落多样性也将由根系非对称性竞争所调控.而在植物生物量分配方面, 陆生植物体间的相互作用(竞争和促进)也会影响植物体地上和地下的生物量分配(Deng et al ., 2006 ; Chu et al ., 2008 ), 对植物对称性与非对称性竞争理论的深入研究或许能更清晰地揭示这种响应机理.至于植物个体大小不整齐性随种群密度的增加而增加这一现象(Weiner, 1986 ), 究竟是由于空间格局不一致引起植物密度变化而产生的Thomas & Weiner, 1989 ), 还是由于植物体的非对称性竞争产生的, 或者是植物本身具有较强的可塑性而导致的, 需要生态学家对其相关理论进一步深入研究.物种共存、生物入侵以及生物多样性等问题也与植物体间非对称性竞争有着千丝万缕的联系(Osunkoya et al ., 2013 ; Skácelová & Lepš, 2014 ), 研究非对称性竞争的产生机制和生理原理对开展相关工作具有重要意义. ...
... 早期的竞争理论认为: 植物体地上部分主要竞争光资源, 属于非对称性竞争; 地下部分主要竞争水肥资源, 属于对称性竞争(Weiner, 1986 ; Weiner et al ., 1997 ; Schwinning & Weiner, 1998 ; Zhang et al ., 2013 ).其原理是: 高大植物凭借株高优势遮挡周围的矮小植物, 阻碍其获得充足的光照, 而矮小植物对高大植物的光吸收影响不大, 因此, 植物光竞争为非对称性竞争Weiner & Thomas, 1986 ; Connoily & Wayne, 1996 ); 由于地下水肥资源的流动特性, 使不同个体大小的植物均能得到资源, 因此植物水肥竞争为对称性竞争Bolte & Villanueva, 2006 ).但近年也有研究Rajaniem, 2003 ; Rewald & Leuschner, 2009 ; Lei et al ., 2012 ; Weremijewicz & Janos, 2013 )发现, 地下竞争为非对称性竞争或存在非对称性竞争倾向.比如, Rajaniemi (2003)研究发现, 植物在不同种群密度条件下, 目标植物周围存在不同大小的植物时, 较大的植物对目标植物单位生物量的影响大于较小植物对目标植物的影响, 并证明地下竞争可能为非对称性竞争.而Rewald和Leuschner (2009)以水青冈属(Fagus )和栎属(Quercus )为研究对象, 将野外生长的两树种的部分根系非破坏性挖出, 插进设计好的根生长室中, 使这部分根系发生种内、种间竞争作用, 并给予不同的水分梯度处理, 研究表明植物根系的非对称性竞争还会受到水分梯度的影响, 即植物间的非对称性竞争程度随水分胁迫梯度的增加而增加.植物根系间发生非对称性竞争现象的作用机理通常可以解释为: 生长于同一环境中的两株大小不同的植物, 较大的植物根系伸展范围广, 根系接触土壤中营养元素的表面积大, 获取的水肥资源多, 进而促进植物生长, 而进一步的生长又会使其根系伸展范围更广, 根系接触土壤中营养元素的表面积更大, 吸收消耗土壤中水分营养元素资源的能力更强.在土壤中资源有限的情况下, 大个体植物因其具有更强的吸收消耗能力而占有更多小个体根际周围的水肥资源, 进而导致小个体植物根际周围水肥资源的供应速率越来越低, 加剧了对小个体植物生长的限制.因此, 随着植物生长, 小个体与大个体间对土壤中有限资源的竞争能力和资源的占有量会随着植物的生长而进一步扩大, 最终同样可能导致小个体因资源的枯竭而死亡, 形成了典型的根系非对称性竞争.此外, 也有研究者认为, 在同等环境条件下, 植物个体仅仅通过对有限资源的抢先占有, 便能在竞争中胜出(Craine et al ., 2005 ), 也就是说, 植物个体对有限资源获取的起始时间不一致, 导致了对资源的非对称性竞争. ...
... 检验植物体竞争对称性与非对称性的理论模型最主要和最直接的方法就是野外实验或控制实验.通常实验中具体的操作方法是测量生物量(Weiner, 1986 )、株高(Zhi et al ., 2007 )、冠幅(Iponga et al ., 2008 )、叶面积(Zhi et al ., 2007 )、根长(Reubens et al ., 2011 )、根表面积(Rewald & Leuschner, 2009 )等指标, 根据这些指标可以计算竞争强度(competition intensity)、竞争重要性(competition importance)和竞争反应(competition response)等参数(王平等, 2007 ).而这些参数的具体计算方法又有很大不同, 如根据计算竞争强度是否随生产力变化, 可将竞争强度分为相对竞争强度(relative competition intensity)和绝对竞争强度(absolute competition intensity) (Grace, 1995 ); 此外, 还可以根据植物地上和地下生长环境不同计算地上竞争强度、地下竞争强度以及总竞争强度, 实际运用中应根据实验设计和实验目的选择合适的参数, 各个参数的计算主要通过实施对照实验并分析植物体在有无竞争情况下的差异来进行. ...
Symmetry of below- ground competition between Kochia scoparia individuals.
1
1997
... 早期的竞争理论认为: 植物体地上部分主要竞争光资源, 属于非对称性竞争; 地下部分主要竞争水肥资源, 属于对称性竞争(Weiner, 1986 ; Weiner et al ., 1997 ; Schwinning & Weiner, 1998 ; Zhang et al ., 2013 ).其原理是: 高大植物凭借株高优势遮挡周围的矮小植物, 阻碍其获得充足的光照, 而矮小植物对高大植物的光吸收影响不大, 因此, 植物光竞争为非对称性竞争Weiner & Thomas, 1986 ; Connoily & Wayne, 1996 ); 由于地下水肥资源的流动特性, 使不同个体大小的植物均能得到资源, 因此植物水肥竞争为对称性竞争Bolte & Villanueva, 2006 ).但近年也有研究Rajaniem, 2003 ; Rewald & Leuschner, 2009 ; Lei et al ., 2012 ; Weremijewicz & Janos, 2013 )发现, 地下竞争为非对称性竞争或存在非对称性竞争倾向.比如, Rajaniemi (2003)研究发现, 植物在不同种群密度条件下, 目标植物周围存在不同大小的植物时, 较大的植物对目标植物单位生物量的影响大于较小植物对目标植物的影响, 并证明地下竞争可能为非对称性竞争.而Rewald和Leuschner (2009)以水青冈属(Fagus )和栎属(Quercus )为研究对象, 将野外生长的两树种的部分根系非破坏性挖出, 插进设计好的根生长室中, 使这部分根系发生种内、种间竞争作用, 并给予不同的水分梯度处理, 研究表明植物根系的非对称性竞争还会受到水分梯度的影响, 即植物间的非对称性竞争程度随水分胁迫梯度的增加而增加.植物根系间发生非对称性竞争现象的作用机理通常可以解释为: 生长于同一环境中的两株大小不同的植物, 较大的植物根系伸展范围广, 根系接触土壤中营养元素的表面积大, 获取的水肥资源多, 进而促进植物生长, 而进一步的生长又会使其根系伸展范围更广, 根系接触土壤中营养元素的表面积更大, 吸收消耗土壤中水分营养元素资源的能力更强.在土壤中资源有限的情况下, 大个体植物因其具有更强的吸收消耗能力而占有更多小个体根际周围的水肥资源, 进而导致小个体植物根际周围水肥资源的供应速率越来越低, 加剧了对小个体植物生长的限制.因此, 随着植物生长, 小个体与大个体间对土壤中有限资源的竞争能力和资源的占有量会随着植物的生长而进一步扩大, 最终同样可能导致小个体因资源的枯竭而死亡, 形成了典型的根系非对称性竞争.此外, 也有研究者认为, 在同等环境条件下, 植物个体仅仅通过对有限资源的抢先占有, 便能在竞争中胜出(Craine et al ., 2005 ), 也就是说, 植物个体对有限资源获取的起始时间不一致, 导致了对资源的非对称性竞争. ...
Suppression of weeds by spring wheat Triticum aestivum increases with crop density and spatial uniformity.
1
2001
... 大小依赖的对称性竞争是指存在资源竞争的植物间依据植物个体大小来分配资源, 即某株植物的生物量占全部植物总生物量的百分比就是其获得资源的百分比(Schwinning & Weiner, 1998 ; Weiner et al ., 2001 ).例如, 3株植物的生物量分别是B 1 、B 2 、B 3 , 总资源量为A, 则3株植物占有的资源量分别为: AB 1 /( B 1 + B 2 + B 3 )、AB 2 /( B 1 + B 2 + B 3 )、AB 3 /( B 1 + B 2 + B 3 ).植物体由幼苗生长至成熟个体的过程中, 可以表征个体大小的生物量逐渐增大, 如果其竞争模式为大小依赖的对称性竞争, 则其占有的资源量将逐渐增多, 竞争能力逐渐增强.完全对称性竞争是指植物在生长过程中竞争资源时, 根据特定区域内的植株个体数平均分配该区域内的资源Schwin- ning & Weiner, 1998 ).例如, 某一区域内有n 株植物, 则每株植物均分得该区域内资源总量的1/n . ...
Size variability and competition in plant monocultures.
2
1986
... 竞争是陆生植物生长过程中的重要现象, 竞争作用是群落结构组建的主导因子和决定物种进化模式的重要因素(王刚和张大勇, 1996 ).对物种竞争的研究由来已久, 19世纪50年代末, 达尔文曾指出生活需求相似的近缘物种间经常发生激烈的竞争.随后, 洛特和沃尔特拉用数学公式描述了物种间的竞争关系.高斯假说从生态位角度展示了物种竞争的结果.竞争的研究领域涉及个体、种群、群落、生态系统等不同层次, 如竞争对个体大小不整齐性的影响(Weiner & Thomas, 1986 )、竞争对种群数量的调节(Deng et al ., 2006 )、竞争与群落空间分布格局的关系(Lin et al ., 2012 )、竞争在维持生态系统物种多样性中的作用(Borer et al ., 2014 )等等.随着植物间相互作用研究的不断拓展, 不断有新的概念和理论模型被提出, 甚至有的学者试图打破竞争与正相互作用的清晰界限(Warren et al ., 2014 ).在逐渐深入研究植物间相互作用的过程中, 植物地下根系间的竞争及其生态过程更受关注(Sutherland et al ., 2013 ).本文在前人研究的基础上, 从竞争模式角度着重对植物竞争(包括地上竞争和地下竞争)的对称性和非对称性进行综述和分析, 以期为国内相关研究提供参考和借鉴. ...
... 早期的竞争理论认为: 植物体地上部分主要竞争光资源, 属于非对称性竞争; 地下部分主要竞争水肥资源, 属于对称性竞争(Weiner, 1986 ; Weiner et al ., 1997 ; Schwinning & Weiner, 1998 ; Zhang et al ., 2013 ).其原理是: 高大植物凭借株高优势遮挡周围的矮小植物, 阻碍其获得充足的光照, 而矮小植物对高大植物的光吸收影响不大, 因此, 植物光竞争为非对称性竞争Weiner & Thomas, 1986 ; Connoily & Wayne, 1996 ); 由于地下水肥资源的流动特性, 使不同个体大小的植物均能得到资源, 因此植物水肥竞争为对称性竞争Bolte & Villanueva, 2006 ).但近年也有研究Rajaniem, 2003 ; Rewald & Leuschner, 2009 ; Lei et al ., 2012 ; Weremijewicz & Janos, 2013 )发现, 地下竞争为非对称性竞争或存在非对称性竞争倾向.比如, Rajaniemi (2003)研究发现, 植物在不同种群密度条件下, 目标植物周围存在不同大小的植物时, 较大的植物对目标植物单位生物量的影响大于较小植物对目标植物的影响, 并证明地下竞争可能为非对称性竞争.而Rewald和Leuschner (2009)以水青冈属(Fagus )和栎属(Quercus )为研究对象, 将野外生长的两树种的部分根系非破坏性挖出, 插进设计好的根生长室中, 使这部分根系发生种内、种间竞争作用, 并给予不同的水分梯度处理, 研究表明植物根系的非对称性竞争还会受到水分梯度的影响, 即植物间的非对称性竞争程度随水分胁迫梯度的增加而增加.植物根系间发生非对称性竞争现象的作用机理通常可以解释为: 生长于同一环境中的两株大小不同的植物, 较大的植物根系伸展范围广, 根系接触土壤中营养元素的表面积大, 获取的水肥资源多, 进而促进植物生长, 而进一步的生长又会使其根系伸展范围更广, 根系接触土壤中营养元素的表面积更大, 吸收消耗土壤中水分营养元素资源的能力更强.在土壤中资源有限的情况下, 大个体植物因其具有更强的吸收消耗能力而占有更多小个体根际周围的水肥资源, 进而导致小个体植物根际周围水肥资源的供应速率越来越低, 加剧了对小个体植物生长的限制.因此, 随着植物生长, 小个体与大个体间对土壤中有限资源的竞争能力和资源的占有量会随着植物的生长而进一步扩大, 最终同样可能导致小个体因资源的枯竭而死亡, 形成了典型的根系非对称性竞争.此外, 也有研究者认为, 在同等环境条件下, 植物个体仅仅通过对有限资源的抢先占有, 便能在竞争中胜出(Craine et al ., 2005 ), 也就是说, 植物个体对有限资源获取的起始时间不一致, 导致了对资源的非对称性竞争. ...
Common mycorrhizal networks amplify size inequality in Andropogon gerardii monocultures.
1
2013
... 早期的竞争理论认为: 植物体地上部分主要竞争光资源, 属于非对称性竞争; 地下部分主要竞争水肥资源, 属于对称性竞争(Weiner, 1986 ; Weiner et al ., 1997 ; Schwinning & Weiner, 1998 ; Zhang et al ., 2013 ).其原理是: 高大植物凭借株高优势遮挡周围的矮小植物, 阻碍其获得充足的光照, 而矮小植物对高大植物的光吸收影响不大, 因此, 植物光竞争为非对称性竞争Weiner & Thomas, 1986 ; Connoily & Wayne, 1996 ); 由于地下水肥资源的流动特性, 使不同个体大小的植物均能得到资源, 因此植物水肥竞争为对称性竞争Bolte & Villanueva, 2006 ).但近年也有研究Rajaniem, 2003 ; Rewald & Leuschner, 2009 ; Lei et al ., 2012 ; Weremijewicz & Janos, 2013 )发现, 地下竞争为非对称性竞争或存在非对称性竞争倾向.比如, Rajaniemi (2003)研究发现, 植物在不同种群密度条件下, 目标植物周围存在不同大小的植物时, 较大的植物对目标植物单位生物量的影响大于较小植物对目标植物的影响, 并证明地下竞争可能为非对称性竞争.而Rewald和Leuschner (2009)以水青冈属(Fagus )和栎属(Quercus )为研究对象, 将野外生长的两树种的部分根系非破坏性挖出, 插进设计好的根生长室中, 使这部分根系发生种内、种间竞争作用, 并给予不同的水分梯度处理, 研究表明植物根系的非对称性竞争还会受到水分梯度的影响, 即植物间的非对称性竞争程度随水分胁迫梯度的增加而增加.植物根系间发生非对称性竞争现象的作用机理通常可以解释为: 生长于同一环境中的两株大小不同的植物, 较大的植物根系伸展范围广, 根系接触土壤中营养元素的表面积大, 获取的水肥资源多, 进而促进植物生长, 而进一步的生长又会使其根系伸展范围更广, 根系接触土壤中营养元素的表面积更大, 吸收消耗土壤中水分营养元素资源的能力更强.在土壤中资源有限的情况下, 大个体植物因其具有更强的吸收消耗能力而占有更多小个体根际周围的水肥资源, 进而导致小个体植物根际周围水肥资源的供应速率越来越低, 加剧了对小个体植物生长的限制.因此, 随着植物生长, 小个体与大个体间对土壤中有限资源的竞争能力和资源的占有量会随着植物的生长而进一步扩大, 最终同样可能导致小个体因资源的枯竭而死亡, 形成了典型的根系非对称性竞争.此外, 也有研究者认为, 在同等环境条件下, 植物个体仅仅通过对有限资源的抢先占有, 便能在竞争中胜出(Craine et al ., 2005 ), 也就是说, 植物个体对有限资源获取的起始时间不一致, 导致了对资源的非对称性竞争. ...
1
2014
... 野外实验和实地考察有其客观性和可验证性, 但不确定因素也较多, 准确地研究对称性竞争和非对称性竞争效应较为困难(Firbank & Watkinson, 1987 ), 而模拟实验在研究精度、可操作性和避开不确定因素影响等方面具有明显优势.对相关模型的检验也可通过编程在计算机上进行模拟实验, 尤其是随着植物生长的动态竞争过程更适合通过模拟实验来实现.通常运用Netlogo软件(http://ccl.north- western.edu/netlogo)来实现植物竞争或生长动态等过程的模拟.该软件是由美国西北大学开发的一款能够对自然和社会现象进行仿真模拟的建模软件平台Wilensky, 1999 ).实际操作中, 可通过编程或根据系统动力学原理建立模型中各变量间的关系流程图来完成模拟过程.近年来发展起来的具有强大统计分析功能的R软件也可用于建模仿真和统计模拟, 具体模拟过程主要依据蒙特卡罗方法, 通过计算机产生的随机数(伪随机数)模拟离散随机变量或连续随机变量, 并将预解决的科学问题构建为合适的数学模型, 使模拟的变量正好是数学模型的解. ...
Breaking new ground: Soil communities and exotic plant invasion.
3
2005
... 一些研究发现, 群落中的优势物种往往是那些能够高效利用环境中限制资源的物种(Fargione & Tilman, 2006 ).菌根中的微生物可以使一些植物得到充足的必需元素, 例如豆科植物中的根瘤菌为豆科植物提供充足的氮源, 促使其快速生长.因此, 这类微生物提升了植物根系在土壤中的竞争能力.最终, 豆科植物越长越大, 根系越来越发达, 需要和吸收的营养元素也越来越多(这些元素包括P、K 等必需元素, 而不仅仅是N), 留给供非豆科植物生长的元素就越来越少, 因此抑制非豆科植物生长的程度越来越大, 形成了物种间根系非对称性竞争.土壤中的微生物还有致病菌, 它们会导致相反的结果.一些植物所携带的细菌、真菌等微生物会对其他植物造成伤害, 尤其是一些入侵物种携带的土壤微生物会限制当地物种利用土壤中的资源, 破坏其根系生长, 严重危及当地物种的正常生长; 或者土壤微生物对植物地下生长的调节影响入侵物种间的相互作用关系(Wolfe & Klironomos, 2005 ).土壤中不同的微生物群落以及根系周围不同的菌群成分会对植物间根系竞争产生不同的影响, 不同的微生物通过增加或降低根际周围营养物质的可利用性, 直接影响竞争根系对营养物质的利用效率, 使得仅有一些植物得到较多资源, 最终不同植物因适合度不同而呈现出非对称性竞争趋势(Yang et al ., 2014 ).土壤微生物还可能通过与土壤中营养元素的相互作用来影响物种间的相互作用关系.研究表明, 在土壤中添加大量P元素会降低生物多样性, 即部分物种因不能共存而被淘汰, 而植物根际周围菌群的存在能够增加生物多样性, 因此豆科植物可以依赖于自身拥有的根瘤菌生存于富营养化的土壤中(Yang et al ., 2014 ).调节土壤中的微生物含量和种类, 同时调整土壤中营养元素的含量和种类, 将两种因素进行排列组合, 或许可以找到特定环境, 适合某种植物生存而不利于另一种植物生存, 若使两种植物相邻生长, 物种间非对称性竞争较易体现. ...
... ).土壤中不同的微生物群落以及根系周围不同的菌群成分会对植物间根系竞争产生不同的影响, 不同的微生物通过增加或降低根际周围营养物质的可利用性, 直接影响竞争根系对营养物质的利用效率, 使得仅有一些植物得到较多资源, 最终不同植物因适合度不同而呈现出非对称性竞争趋势(Yang et al ., 2014 ).土壤微生物还可能通过与土壤中营养元素的相互作用来影响物种间的相互作用关系.研究表明, 在土壤中添加大量P元素会降低生物多样性, 即部分物种因不能共存而被淘汰, 而植物根际周围菌群的存在能够增加生物多样性, 因此豆科植物可以依赖于自身拥有的根瘤菌生存于富营养化的土壤中(Yang et al ., 2014 ).调节土壤中的微生物含量和种类, 同时调整土壤中营养元素的含量和种类, 将两种因素进行排列组合, 或许可以找到特定环境, 适合某种植物生存而不利于另一种植物生存, 若使两种植物相邻生长, 物种间非对称性竞争较易体现. ...
... ).土壤微生物还可能通过与土壤中营养元素的相互作用来影响物种间的相互作用关系.研究表明, 在土壤中添加大量P元素会降低生物多样性, 即部分物种因不能共存而被淘汰, 而植物根际周围菌群的存在能够增加生物多样性, 因此豆科植物可以依赖于自身拥有的根瘤菌生存于富营养化的土壤中(Yang et al ., 2014 ).调节土壤中的微生物含量和种类, 同时调整土壤中营养元素的含量和种类, 将两种因素进行排列组合, 或许可以找到特定环境, 适合某种植物生存而不利于另一种植物生存, 若使两种植物相邻生长, 物种间非对称性竞争较易体现. ...
The interaction between arbuscular mycorrhizal fungi and soil phosphorus availability influences plant community productivity and ecosystem stability.
2
2014
... 早期的竞争理论认为: 植物体地上部分主要竞争光资源, 属于非对称性竞争; 地下部分主要竞争水肥资源, 属于对称性竞争(Weiner, 1986 ; Weiner et al ., 1997 ; Schwinning & Weiner, 1998 ; Zhang et al ., 2013 ).其原理是: 高大植物凭借株高优势遮挡周围的矮小植物, 阻碍其获得充足的光照, 而矮小植物对高大植物的光吸收影响不大, 因此, 植物光竞争为非对称性竞争Weiner & Thomas, 1986 ; Connoily & Wayne, 1996 ); 由于地下水肥资源的流动特性, 使不同个体大小的植物均能得到资源, 因此植物水肥竞争为对称性竞争Bolte & Villanueva, 2006 ).但近年也有研究Rajaniem, 2003 ; Rewald & Leuschner, 2009 ; Lei et al ., 2012 ; Weremijewicz & Janos, 2013 )发现, 地下竞争为非对称性竞争或存在非对称性竞争倾向.比如, Rajaniemi (2003)研究发现, 植物在不同种群密度条件下, 目标植物周围存在不同大小的植物时, 较大的植物对目标植物单位生物量的影响大于较小植物对目标植物的影响, 并证明地下竞争可能为非对称性竞争.而Rewald和Leuschner (2009)以水青冈属(Fagus )和栎属(Quercus )为研究对象, 将野外生长的两树种的部分根系非破坏性挖出, 插进设计好的根生长室中, 使这部分根系发生种内、种间竞争作用, 并给予不同的水分梯度处理, 研究表明植物根系的非对称性竞争还会受到水分梯度的影响, 即植物间的非对称性竞争程度随水分胁迫梯度的增加而增加.植物根系间发生非对称性竞争现象的作用机理通常可以解释为: 生长于同一环境中的两株大小不同的植物, 较大的植物根系伸展范围广, 根系接触土壤中营养元素的表面积大, 获取的水肥资源多, 进而促进植物生长, 而进一步的生长又会使其根系伸展范围更广, 根系接触土壤中营养元素的表面积更大, 吸收消耗土壤中水分营养元素资源的能力更强.在土壤中资源有限的情况下, 大个体植物因其具有更强的吸收消耗能力而占有更多小个体根际周围的水肥资源, 进而导致小个体植物根际周围水肥资源的供应速率越来越低, 加剧了对小个体植物生长的限制.因此, 随着植物生长, 小个体与大个体间对土壤中有限资源的竞争能力和资源的占有量会随着植物的生长而进一步扩大, 最终同样可能导致小个体因资源的枯竭而死亡, 形成了典型的根系非对称性竞争.此外, 也有研究者认为, 在同等环境条件下, 植物个体仅仅通过对有限资源的抢先占有, 便能在竞争中胜出(Craine et al ., 2005 ), 也就是说, 植物个体对有限资源获取的起始时间不一致, 导致了对资源的非对称性竞争. ...
... 学者们除了通过目标植物生物量与邻体数量或密度间的变化关系来研究植物间竞争类型外, 还以个体影响域模型为基础, 通过数学建模和对植物个体生长过程的计算机模拟来研究植物间的对称性和非对称性竞争以及正相互作用强度等(Chu et al ., 2008 ), 并逐渐由植物体单层影响域模型向将植物体地上、地下部分区分开的双层影响域模型发展(Zhang et al ., 2013 ).关于运用个体影响域模型来解决对称性与非对称性竞争问题的研究, 未来可能更多关注于模型的进一步完善以及在建模过程中逐步将植物体根、茎、叶等器官区分开来. ...
The interplay between above- and below-ground plant-plant interactions along an environmental gradient: Insights from two-layer zone-of-influence models.
2
2013
... 检验植物体竞争对称性与非对称性的理论模型最主要和最直接的方法就是野外实验或控制实验.通常实验中具体的操作方法是测量生物量(Weiner, 1986 )、株高(Zhi et al ., 2007 )、冠幅(Iponga et al ., 2008 )、叶面积(Zhi et al ., 2007 )、根长(Reubens et al ., 2011 )、根表面积(Rewald & Leuschner, 2009 )等指标, 根据这些指标可以计算竞争强度(competition intensity)、竞争重要性(competition importance)和竞争反应(competition response)等参数(王平等, 2007 ).而这些参数的具体计算方法又有很大不同, 如根据计算竞争强度是否随生产力变化, 可将竞争强度分为相对竞争强度(relative competition intensity)和绝对竞争强度(absolute competition intensity) (Grace, 1995 ); 此外, 还可以根据植物地上和地下生长环境不同计算地上竞争强度、地下竞争强度以及总竞争强度, 实际运用中应根据实验设计和实验目的选择合适的参数, 各个参数的计算主要通过实施对照实验并分析植物体在有无竞争情况下的差异来进行. ...
... )、叶面积(Zhi et al ., 2007 )、根长(Reubens et al ., 2011 )、根表面积(Rewald & Leuschner, 2009 )等指标, 根据这些指标可以计算竞争强度(competition intensity)、竞争重要性(competition importance)和竞争反应(competition response)等参数(王平等, 2007 ).而这些参数的具体计算方法又有很大不同, 如根据计算竞争强度是否随生产力变化, 可将竞争强度分为相对竞争强度(relative competition intensity)和绝对竞争强度(absolute competition intensity) (Grace, 1995 ); 此外, 还可以根据植物地上和地下生长环境不同计算地上竞争强度、地下竞争强度以及总竞争强度, 实际运用中应根据实验设计和实验目的选择合适的参数, 各个参数的计算主要通过实施对照实验并分析植物体在有无竞争情况下的差异来进行. ...
Inter-specific competition: Spartina alterniflora is replacing Spartina anglica in coastal China.
0
2007

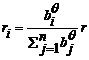 (1)
(1) 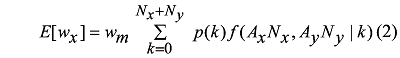
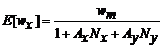 (3)
(3) 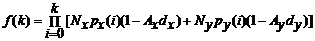 (4)
(4) 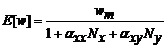 (5)
(5)