Net ecosystem exchange of CO2 on sunny and cloudy days over a reed wetland in the Yellow River Delta, China
CHU Xiao-Jing1, 2, HAN Guang-Xuan1, *, , XING Qing-Hui1, 2, YU Jun-Bao1, WU Li-Xin3, LIU Hai-Fang3, WANG Guang-Mei1, MAO Pei-Li1
1Key Laboratory of Coastal Environmental Processes and Ecological Remediation, Chinese Academy of Sciences, Yantai Institute of Coastal Zone Research, Chinese Academy of Sciences, Yantai, Shandong 264003, China2University of Chinese Academy of Sciences, Beijing 100049, China3Administration Bureau of the Yellow River Delta National Nature Reserve, Dongying, Shandong 257091, China
通讯作者: * 通讯作者 Author for correspondence (E-mail: gxhan@yic.ac.cn)
Aims Clouds and aerosols change the radiation level on the land surface and indirectly alter the microclimate. Shifts in sunny and cloudy days, for example, would affect the net ecosystem exchange of CO2 (NEE) between land surface and the atmosphere. Our objective was to analyze the influence of shifts in sunny and cloudy days on NEE, its responses to light and temperature in a reed (Phragmites australis) wetland in the Yellow River Delta, China. Methods Using the eddy covariance technique, we measured the temporal changes in NEE during the growing season over the reed wetland. We selected 12 paired-days during the measurement period following two criteria: (1) the two paired days are adjacent, with one sunny day and another cloudy day; (2) no rain event during the two days. We assumed that: (1) live biomass and leaf area index (LAI) are the same during any paired-days; (2) soil moisture has no significant difference between the two adjacent days. With these criteria, we expected that radiation condition exerted the major control on NEE. Important findings Diurnal change of NEE showed a distinct U-shaped pattern on both sunny and cloudy days, but with substantial variation in its amplitude. During the daytime, NEE on sunny days was significantly higher (p < 0.01) than that on the cloudy days (n = 12). The daytime NEE response to photosynthetically active radiation (PAR) was modeled with the rectangular hyperbolic function (Eq. (1)) for both sunny and cloudy days. There appeared a significant reduction (p < 0.01) in light-saturated NEE (Amax) on cloudy days compared to the sunny days. Similarly, there was a significant decrease (p < 0.01) in daytime ecosystem respiration (Reco,daytime) on cloudy days as compared to that of the sunny day although there existed significant exponential relationships between Reco,daytime and air temperature on both sunny and cloudy days. In addition, the temperature sensitivity of ecosystem respiration (Q10) on cloudy days (1.9) was significantly lower than that of sunny days (5.5). Stepwise multiple regression analyses suggested that PAR and T explained 63% of the changes in NEE between sunny and cloudy days. By taking advantage of the natural shift of sunny and cloudy days without disturbance to the plant-soil system, our results indicated that cloud cover significantly reduced the absorption capacity of CO2 in the wetland. Thus, it is necessary to take into account the shits between sunny and cloudy days on NEE when predicting the ecosystem responses to future climate in the wetland.
Keywords:sunny day
;
cloudy day
;
net ecosystem CO2 exchange (NEE)
;
daytime ecosystem respiration (Reco
;
daytime)
;
light response
;
temperature response
CHUXiao-Jing, HANGuang-Xuan, XINGQing-Hui, YUJun-Bao, WULi-Xin, LIUHai-Fang, WANGGuang-Mei, MAOPei-Li. Net ecosystem exchange of CO2 on sunny and cloudy days over a reed wetland in the Yellow River Delta, China[J]. Chinese Journal of Plant Ecology, 2015, 39(7): 661-673 https://doi.org/10.17521/cjpe.2015.0063
Fig. 1 Diurnal changes in net ecosystem exchange of CO2 (NEE) and photosynthetically active radiation (PAR) on the 12 paired-days (i.e., a sunny day and an adjacent cloudy day) during the 2013 growing season in the Yellow River Delta wetland. Black and grey solid lines represent PAR on sunny and cloudy days, respectively.
图2 2013年黄河三角洲湿地生长季阴天和晴天净生态系统CO2交换(NEE)、光合有效辐射(PAR)与气温(T)的平均日动态(平均值±标准误差)。***, p < 0.001。
Fig. 2 Average diurnal variations of net ecosystem exchange of CO2 (NEE), photosynthetically active radiation (PAR) and air temperature (T) on sunny days and cloudy days during the 2013 growing season in the Yellow River Delta. Bars represent standard errors of the means of 12 sunny days and 12 adjacent cloudy days (mean ± SE). ***, p < 0.001.
Fig. 3 Relationships between daytime net ecosystem exchange of CO2 (NEE) and photosynthetically active radiation (PAR) between sunny days and cloudy days during the 2013 growing season in the Yellow River Delta wetland. Black solid line represents fitting curve of sunny days, and grey line represents fitting curve of cloudy days.
Fig. 4 Relationships between daytime ecosystem respiration (Reco,daytime) and air temperature (T) on sunny days and cloudy days during the 2013 growing season in the Yellow River Delta wetland. Black solid line represents fitting curve of sunny days, and grey line represents fitting curve of cloudy days.
2.4 阴天和晴天PAR与T对湿地生态系统NEE的协同影响
多元回归分析发现, T和PAR对晴天NEE的协同影响达到40% (p < 0.01), 对阴天的协同影响为41%, 偏相关分析发现T是晴天NEE变异的主控因子(T: R2 = 0.37, p < 0.001; PAR: R2 = 0.20, p < 0.001), PAR是阴天NEE变异的主控因子(T: R2 = 0.10, p < 0.001; PAR: R2 = 0.38, p < 0.001)。多元回归分析表明, ΔPAR与ΔT对ΔNEE的协同影响达到63% (R2 = 0.63, p < 0.001; 表2), 说明PAR与T是引起阴天和晴天NEE差异的主要环境因子。
Table 1 Comparison of the analog parameters from daytime net ecosystem exchange of CO2(NEE) and photosynthetically active radiation (PAR) using a Michaelis-Menten model (Eq.(3)) between sunny days and cloudy days in the wetland
Table 2 Estimated empirical coefficient of multiple liner regression models for changes in net ecosystem exchange (NEE) with photosynthetically active radiation (PAR) and air temperature (T) on sunny and cloudy days during the growing season in the Yellow River Delta
方程 Equation
R2
p
n
晴天 Sunny day
NEE = -0.005PAR - 0.28T + 4.06
0.41
<0.001
288
阴天 Cloudy day
NEE = -0.006PAR - 0.107T + 2.67
0.42
<0.001
288
阴天和晴天差量 Difference between sunny and cloudy days
ΔNEE = -0.004ΔPAR - 0.123ΔT - 2.54
0.63
<0.001
288
ΔNEE, NEE difference between sunny and cloudy days; ΔT, air temperature difference between sunny and cloudy days.ΔNEE, 阴天和晴天NEE差量; ΔT, 气温差量。
Respiration of the external mycelium in the arbuscular mycorrhizal symbiosis shows strong dependence on recent photosynthates and acclimation to temperature.
Annual variation of temperature sensitivity of soil organic carbon decomposition in North peatlands: Implications for thermal responses of carbon cycling to global warming.
Carbon dioxide exchange above a Mediterranean C3/C4 grassland during two climatologically contrasting years.
1
2008
... 另外, 太阳辐射变化引起的温度变化也影响NEE对光的响应特征.首先, 温度通过影响酶系列化学反应对光合作用产生影响(Syed et al., 2006), 温度升高有利于光合速率的提高(Pingintha et al., 2010).相关分析发现, 阴天和晴天Amax与T均为显著的线性正相关(晴天: R2 = 0.80, p < 0.05; 阴天: R2 = 0.63, p < 0.05), 其次, 生态系统呼吸与气温通常显著正相关(Aires et al., 2008; Pingintha et al., 2010).本研究中虽然晴天条件下Reco,daytime较阴天提高了20%, 但NEE却显著高于阴天的67%, 说明晴天温度对湿地光合固碳能力的提高远超过生态系统呼吸的消耗, 表现为晴天生态系统的碳汇功能更强, 这与已有的研究结果一致(Han et al., 2014a). ...
Reduced carbon sequestration in terrestrial ecosystems under overcast skies compared to clear skies.
2008
The impact of diffuse sunlight on canopy light-use efficiency, gross photosynthetic product and net ecosystem exchange in three forest biomes.
1
2007
... 已有研究表明, 阴天和晴天所引起的辐射差异影响着陆地生态系统的交换过程和碳吸收能力(Roderick et al., 2001; Still et al., 2009).但由于研究方法、地域环境条件以及不同植被类型的差异, 研究结论存在较大差异(Gu et al., 1999, 2002; Alton et al., 2007; Urban et al., 2007).例如, 中国渭北刺槐(Robinia pseudoacacia)生长初期和旺盛期晴天条件下森林日均固碳量比阴天高8%-16% (郑元等, 2011).中国西北玉米(Zea mays)农田生态系统阴天条件下净生态系统CO2交换(NEE)的增长速率却高于晴天, 且生态系统总初级生产力(GPP)最大值出现于阴天(Zhang et al., 2011).对中国西北草地(Bai et al., 2012)和不同温带森林类型(Zhang et al., 2010)的研究也得出类似结论.另外, 也有研究发现加拿大常绿灌木(Letts et al., 2005)和西藏高寒草甸(范玉枝等, 2009)生态系统CO2交换不受阴天和晴天天气条件影响. ...
Comparing the impact of cloudiness on carbon dioxide exchange in a grassland and a maize cropland in northwestern China.
1
2012
... 已有研究表明, 阴天和晴天所引起的辐射差异影响着陆地生态系统的交换过程和碳吸收能力(Roderick et al., 2001; Still et al., 2009).但由于研究方法、地域环境条件以及不同植被类型的差异, 研究结论存在较大差异(Gu et al., 1999, 2002; Alton et al., 2007; Urban et al., 2007).例如, 中国渭北刺槐(Robinia pseudoacacia)生长初期和旺盛期晴天条件下森林日均固碳量比阴天高8%-16% (郑元等, 2011).中国西北玉米(Zea mays)农田生态系统阴天条件下净生态系统CO2交换(NEE)的增长速率却高于晴天, 且生态系统总初级生产力(GPP)最大值出现于阴天(Zhang et al., 2011).对中国西北草地(Bai et al., 2012)和不同温带森林类型(Zhang et al., 2010)的研究也得出类似结论.另外, 也有研究发现加拿大常绿灌木(Letts et al., 2005)和西藏高寒草甸(范玉枝等, 2009)生态系统CO2交换不受阴天和晴天天气条件影响. ...
Measuring and modelling carbon dioxide and water vapour exchange over a temperate broad-leaved forest during the 1995 summer drought.
1
1997
... 本文研究发现晴天和阴天芦苇湿地NEE对PAR的响应趋势一致, 均呈直角双曲线关系, 与先前的很多研究结果一致(Falge et al., 2001; Monson et al., 2002; Zhou et al., 2009; 张弥等, 2009; Han et al., 2014a, 2014b; 邢庆会等, 2014).PAR是控制阴天和晴天生态系统生长季昼间NEE变化的主要因素(Flanagan & Johnson, 2005; Liu et al., 2006; 薛红喜等, 2012), Michaelis-Menten方程拟合发现, PAR可以解释晴天单天NEE变化的57%-82%, 解释阴天单天NEE变化的53%-81%.云层变化会改变地表接收太阳辐射的强度, 影响PAR、T等环境因子, 进而影响阴天和晴天条件下NEE对光的响应特征(Baldocchi, 1997; Freedman et al., 2001). ...
How switches and lags in biophysical regulators affect spatial-temporal variation of soil respiration in an oak-grass savanna.
1
2006
... 本研究以黄河三角洲芦苇湿地为研究对象, 在自然条件下选择相邻的阴天和晴天, 在生物要素(生物量、叶面积指数)、土壤水分以及养分特征保持不变的前提下, 分析阴天和晴天引起的辐射变化对生态系统NEE的影响机制.目前, 有关太阳辐射对生态系统NEE的影响机制大都以晴空指数为指标(Gu et al., 2003; Liu et al., 2006; 范玉枝等, 2009; Zhang et al., 2010; 周丽艳等, 2010; 蒋琰等, 2011; Zhao et al., 2011; 刘佳等, 2014).这种研究方法的最大不足就是研究阶段中, 太阳辐射、生物要素(生物量、叶面积指数)、土壤湿度等环境因子都在发生变化, 这对阐明太阳辐射这一单因素对NEE的控制机制研究带来一定困难.而本研究巧妙地在自然条件下选择相邻的阴天和晴天, 在日尺度上实现了对单一变量PAR的控制.利用这种研究方法, Han等(2014a)通过对比分析阴天和晴天植物冠层光合作用的差异性, 定量分析了光合作用在日尺度上对土壤呼吸的调节作用.此外, 我们已有研究发现PAR改变所引起的植物光合作用变化对生态系统呼吸会产生一定的影响(Tang et al., 2005; Baldocchi et al., 2006; Moyano et al., 2008; Wingate et al., 2010; Han et al., 2012).但是, 以往的研究多通过夜间生态系统呼吸与气温函数关系外延的方法来估算白天生态系统呼吸(Doughty et al., 2010; Zhang et al., 2011), 忽略了PAR对白天生态系统呼吸的影响, 因而具有一定的局限性.考虑到白天PAR的变化, 本研究采用Michaelis-Menten模型对白天生态系统呼吸进行推算, 使得生态系统呼吸的温度响应数据更具有说服力. ...
Estimating cloud field coverage using morphological analysis.
1
2010
... 近年来, 大气污染、气溶胶粒子增加等环境问题日益严重, 云层和大气气溶胶含量的变化给地面接收的直接辐射和散射辐射带来很大影响(Niyogi et al., 2004; Bar-Or et al., 2010; 马金玉等, 2011).我国华北地区大气环境由于受到西北地区沙尘的影响, 大量滞留在大气中的沙尘成为对流层气溶胶的主要成分, 影响地-气系统辐射能收支, 从而影响区域的气候及生态环境(Min, 2005; 贾漩等, 2010).研究表明: 近50年来, 地面太阳总辐射从减少到增加(齐月等, 2014).而云作为一种天气现象会对局地的微气候环境产生综合效应, 当天空有云层出现时, 地面接收的太阳辐射强度及散射辐射与直射辐射的比例会发生变化, 引起生态系统光能与热能的变化, 植被的光合与呼吸作用发生变化, 最终生态系统与大气间的净CO2交换也会受到影响(Letts et al., 2005; Urban et al., 2007). ...
Cloud pattern and water relations in Picea rubens and Abies fraseri, southern Appalachian Mountains, USA.
1
2012
... 阴天和晴天生态系统Reco,daytime与T均呈显著的正相关关系, 两者呈指数关系, 与以往很多研究的结果一致(石培礼等, 2006; Zhou et al., 2009; Zhao et al., 2010; Han et al., 2013, 2014a; 邢庆会等, 2014).T是影响生态系统呼吸的主要因素(Law et al., 2002; Reichstein et al., 2005; Urban et al., 2007), 本文研究发现: 黄河三角洲芦苇湿地晴天气温相对阴天提高了10%, 其阴天和晴天的气温差高于美国阿尔卑斯山脉南部森林的阴天和晴天的气温差(Berry & Smith, 2012), 黄河三角洲芦苇湿地晴天对应的生态系统呼吸相对阴天提高了20% (p < 0.05), 说明阴天条件下, 植被地上部分尤其是叶面温度较晴朗天气强太阳辐射条件下的低, 叶片以及植被茎干呼吸受到一定的抑制(Urban et al., 2007). ...
Net ecosystem CO2 exchange in a temperate cattail marsh in relation to biophysical properties.
1
2008
... Q10值被广泛应用于评价生态系统呼吸对温度变化的敏感程度(杨庆朋等, 2011; 杨利琼等, 2013; Han et al., 2014a, 2014b), Q10值越大, 说明生态系统呼吸对温度变化的响应越敏感.黄河三角洲芦苇湿地生态系统阴天和晴天Q10值均在已有的湿地Q10范围(1.0-7.7)内(Chapman & Thurlow, 1996; Silvola et al., 1996; Lafleur et al., 2001; Bonneville et al., 2008; Zhou et al., 2009; 周丽艳等, 2010), 其中晴天白天Q10为5.5, 远高于阴天白天的Q10 (1.9), 说明晴天白天生态系统呼吸对温度变化的敏感性相对较高.分析其原因, 一方面生态系统温度敏感性与气温有关(Kirschbaum, 1995; Xu & Qi, 2001; Janssens & Pilegaard, 2003; Davidson & Janssens., 2006; 张雷明等, 2006; Jassal et al., 2008), 气温对Q10产生影响主要通过影响光合与呼吸作用酶的活性实现(Davidson & Janssens, 2006).晴天气温T ((23.56 ± 2.62) ℃)显著大于阴天((21.26 ± 2.34) ℃), 阴天由于酶促反应需要一定的活化酶, 光合与呼吸作用酶的活性一般受到限制; 而晴天随着辐射增强, 气温升高, 越来越多的分子达到或超过了自身的活化能, 反应加快(Xiang & Freeman, 2009; Karhu et al., 2010).另一方面, 晴天相对阴天较高的PAR使得晴天光合作用更强, 产生输送的有机质增多(Curiel Yuste et al., 2007; Jassal et al., 2008), 自养呼吸增强, 同时地上植被以及土壤异养微生物呼吸对气温的响应能力增强(Edwards et al., 2004; Heinemeyer et al., 2006; Moyano et al., 2008), 而阴天由于底物基质供应不足使得自养、异养呼吸受到抑制(Gershenson et al., 2009). ...
黄河三角洲景观组分判定与景观破碎化分析
1
1999
... 黄河口近海与海岸湿地是暖温带增长速度最快和最具代表性的新生近海与海岸湿地, 具有海陆过渡性、原生性和脆弱性的特点(布仁仓等, 1999; 崔树强, 2002).受水沙量变化、陆地和河流淡水径流、咸水海流多重影响以及人类活动的干扰, 黄河三角洲湿地生态系统CO2交换存在着极大的复杂性和不确定性(Sasaki et al., 2009;Han et al., 2014a, 2014b; 邢庆会等, 2014).研究表明黄河三角洲芦苇湿地生长季NEE占全年的80%以上, 且碳吸收集中在植物生长旺盛期(7-9月)(杨利琼等, 2013; Han et al., 2014a, 2014b).而这一时期正值黄河三角洲雨季, 其云雨状况影响太阳辐射, 进而影响湿地生态系统的碳吸收功能.在以往的研究中, 温度、水分、太阳辐射等环境因子, 生物量、叶面积等生物因子以及地下水位、地表水深等水文要素对湿地生态系统NEE的影响机制受到研究者的极大关注(牟长城等, 2009; Zhao et al., 2010; Han et al., 2013; 邢庆会等, 2014).目前, 有关阴天和晴天条件对生态系统碳交换影响的研究主要集中在森林生态系统(Liu et al., 2006; Zhang et al., 2010; 周丽艳等, 2010; Jiang et al., 2011; Zhao et al., 2011; 刘佳等, 2014)和草原生态系统(Gu et al., 2003; 范玉枝等, 2009).阴天和晴天状况引起的太阳辐射变化对湿地生态系统NEE和碳汇功能究竟产生怎样的影响目前尚不清楚. ...
黄河三角洲景观组分判定与景观破碎化分析
1
1999
... 黄河口近海与海岸湿地是暖温带增长速度最快和最具代表性的新生近海与海岸湿地, 具有海陆过渡性、原生性和脆弱性的特点(布仁仓等, 1999; 崔树强, 2002).受水沙量变化、陆地和河流淡水径流、咸水海流多重影响以及人类活动的干扰, 黄河三角洲湿地生态系统CO2交换存在着极大的复杂性和不确定性(Sasaki et al., 2009;Han et al., 2014a, 2014b; 邢庆会等, 2014).研究表明黄河三角洲芦苇湿地生长季NEE占全年的80%以上, 且碳吸收集中在植物生长旺盛期(7-9月)(杨利琼等, 2013; Han et al., 2014a, 2014b).而这一时期正值黄河三角洲雨季, 其云雨状况影响太阳辐射, 进而影响湿地生态系统的碳吸收功能.在以往的研究中, 温度、水分、太阳辐射等环境因子, 生物量、叶面积等生物因子以及地下水位、地表水深等水文要素对湿地生态系统NEE的影响机制受到研究者的极大关注(牟长城等, 2009; Zhao et al., 2010; Han et al., 2013; 邢庆会等, 2014).目前, 有关阴天和晴天条件对生态系统碳交换影响的研究主要集中在森林生态系统(Liu et al., 2006; Zhang et al., 2010; 周丽艳等, 2010; Jiang et al., 2011; Zhao et al., 2011; 刘佳等, 2014)和草原生态系统(Gu et al., 2003; 范玉枝等, 2009).阴天和晴天状况引起的太阳辐射变化对湿地生态系统NEE和碳汇功能究竟产生怎样的影响目前尚不清楚. ...
The influence of climate on CO2 and CH4 emissions from organic soils.
1
1996
... Q10值被广泛应用于评价生态系统呼吸对温度变化的敏感程度(杨庆朋等, 2011; 杨利琼等, 2013; Han et al., 2014a, 2014b), Q10值越大, 说明生态系统呼吸对温度变化的响应越敏感.黄河三角洲芦苇湿地生态系统阴天和晴天Q10值均在已有的湿地Q10范围(1.0-7.7)内(Chapman & Thurlow, 1996; Silvola et al., 1996; Lafleur et al., 2001; Bonneville et al., 2008; Zhou et al., 2009; 周丽艳等, 2010), 其中晴天白天Q10为5.5, 远高于阴天白天的Q10 (1.9), 说明晴天白天生态系统呼吸对温度变化的敏感性相对较高.分析其原因, 一方面生态系统温度敏感性与气温有关(Kirschbaum, 1995; Xu & Qi, 2001; Janssens & Pilegaard, 2003; Davidson & Janssens., 2006; 张雷明等, 2006; Jassal et al., 2008), 气温对Q10产生影响主要通过影响光合与呼吸作用酶的活性实现(Davidson & Janssens, 2006).晴天气温T ((23.56 ± 2.62) ℃)显著大于阴天((21.26 ± 2.34) ℃), 阴天由于酶促反应需要一定的活化酶, 光合与呼吸作用酶的活性一般受到限制; 而晴天随着辐射增强, 气温升高, 越来越多的分子达到或超过了自身的活化能, 反应加快(Xiang & Freeman, 2009; Karhu et al., 2010).另一方面, 晴天相对阴天较高的PAR使得晴天光合作用更强, 产生输送的有机质增多(Curiel Yuste et al., 2007; Jassal et al., 2008), 自养呼吸增强, 同时地上植被以及土壤异养微生物呼吸对气温的响应能力增强(Edwards et al., 2004; Heinemeyer et al., 2006; Moyano et al., 2008), 而阴天由于底物基质供应不足使得自养、异养呼吸受到抑制(Gershenson et al., 2009). ...
黄河断流对黄河三角洲生态环境的影响
1
2002
... 黄河口近海与海岸湿地是暖温带增长速度最快和最具代表性的新生近海与海岸湿地, 具有海陆过渡性、原生性和脆弱性的特点(布仁仓等, 1999; 崔树强, 2002).受水沙量变化、陆地和河流淡水径流、咸水海流多重影响以及人类活动的干扰, 黄河三角洲湿地生态系统CO2交换存在着极大的复杂性和不确定性(Sasaki et al., 2009;Han et al., 2014a, 2014b; 邢庆会等, 2014).研究表明黄河三角洲芦苇湿地生长季NEE占全年的80%以上, 且碳吸收集中在植物生长旺盛期(7-9月)(杨利琼等, 2013; Han et al., 2014a, 2014b).而这一时期正值黄河三角洲雨季, 其云雨状况影响太阳辐射, 进而影响湿地生态系统的碳吸收功能.在以往的研究中, 温度、水分、太阳辐射等环境因子, 生物量、叶面积等生物因子以及地下水位、地表水深等水文要素对湿地生态系统NEE的影响机制受到研究者的极大关注(牟长城等, 2009; Zhao et al., 2010; Han et al., 2013; 邢庆会等, 2014).目前, 有关阴天和晴天条件对生态系统碳交换影响的研究主要集中在森林生态系统(Liu et al., 2006; Zhang et al., 2010; 周丽艳等, 2010; Jiang et al., 2011; Zhao et al., 2011; 刘佳等, 2014)和草原生态系统(Gu et al., 2003; 范玉枝等, 2009).阴天和晴天状况引起的太阳辐射变化对湿地生态系统NEE和碳汇功能究竟产生怎样的影响目前尚不清楚. ...
黄河断流对黄河三角洲生态环境的影响
1
2002
... 黄河口近海与海岸湿地是暖温带增长速度最快和最具代表性的新生近海与海岸湿地, 具有海陆过渡性、原生性和脆弱性的特点(布仁仓等, 1999; 崔树强, 2002).受水沙量变化、陆地和河流淡水径流、咸水海流多重影响以及人类活动的干扰, 黄河三角洲湿地生态系统CO2交换存在着极大的复杂性和不确定性(Sasaki et al., 2009;Han et al., 2014a, 2014b; 邢庆会等, 2014).研究表明黄河三角洲芦苇湿地生长季NEE占全年的80%以上, 且碳吸收集中在植物生长旺盛期(7-9月)(杨利琼等, 2013; Han et al., 2014a, 2014b).而这一时期正值黄河三角洲雨季, 其云雨状况影响太阳辐射, 进而影响湿地生态系统的碳吸收功能.在以往的研究中, 温度、水分、太阳辐射等环境因子, 生物量、叶面积等生物因子以及地下水位、地表水深等水文要素对湿地生态系统NEE的影响机制受到研究者的极大关注(牟长城等, 2009; Zhao et al., 2010; Han et al., 2013; 邢庆会等, 2014).目前, 有关阴天和晴天条件对生态系统碳交换影响的研究主要集中在森林生态系统(Liu et al., 2006; Zhang et al., 2010; 周丽艳等, 2010; Jiang et al., 2011; Zhao et al., 2011; 刘佳等, 2014)和草原生态系统(Gu et al., 2003; 范玉枝等, 2009).阴天和晴天状况引起的太阳辐射变化对湿地生态系统NEE和碳汇功能究竟产生怎样的影响目前尚不清楚. ...
Microbial soil respiration and its dependency on carbon inputs, soil temperature and moisture.
1
2007
... Q10值被广泛应用于评价生态系统呼吸对温度变化的敏感程度(杨庆朋等, 2011; 杨利琼等, 2013; Han et al., 2014a, 2014b), Q10值越大, 说明生态系统呼吸对温度变化的响应越敏感.黄河三角洲芦苇湿地生态系统阴天和晴天Q10值均在已有的湿地Q10范围(1.0-7.7)内(Chapman & Thurlow, 1996; Silvola et al., 1996; Lafleur et al., 2001; Bonneville et al., 2008; Zhou et al., 2009; 周丽艳等, 2010), 其中晴天白天Q10为5.5, 远高于阴天白天的Q10 (1.9), 说明晴天白天生态系统呼吸对温度变化的敏感性相对较高.分析其原因, 一方面生态系统温度敏感性与气温有关(Kirschbaum, 1995; Xu & Qi, 2001; Janssens & Pilegaard, 2003; Davidson & Janssens., 2006; 张雷明等, 2006; Jassal et al., 2008), 气温对Q10产生影响主要通过影响光合与呼吸作用酶的活性实现(Davidson & Janssens, 2006).晴天气温T ((23.56 ± 2.62) ℃)显著大于阴天((21.26 ± 2.34) ℃), 阴天由于酶促反应需要一定的活化酶, 光合与呼吸作用酶的活性一般受到限制; 而晴天随着辐射增强, 气温升高, 越来越多的分子达到或超过了自身的活化能, 反应加快(Xiang & Freeman, 2009; Karhu et al., 2010).另一方面, 晴天相对阴天较高的PAR使得晴天光合作用更强, 产生输送的有机质增多(Curiel Yuste et al., 2007; Jassal et al., 2008), 自养呼吸增强, 同时地上植被以及土壤异养微生物呼吸对气温的响应能力增强(Edwards et al., 2004; Heinemeyer et al., 2006; Moyano et al., 2008), 而阴天由于底物基质供应不足使得自养、异养呼吸受到抑制(Gershenson et al., 2009). ...
Temperature sensitivity of soil carbon decomposition and feedbacks to climate change.
1
2006
... Q10值被广泛应用于评价生态系统呼吸对温度变化的敏感程度(杨庆朋等, 2011; 杨利琼等, 2013; Han et al., 2014a, 2014b), Q10值越大, 说明生态系统呼吸对温度变化的响应越敏感.黄河三角洲芦苇湿地生态系统阴天和晴天Q10值均在已有的湿地Q10范围(1.0-7.7)内(Chapman & Thurlow, 1996; Silvola et al., 1996; Lafleur et al., 2001; Bonneville et al., 2008; Zhou et al., 2009; 周丽艳等, 2010), 其中晴天白天Q10为5.5, 远高于阴天白天的Q10 (1.9), 说明晴天白天生态系统呼吸对温度变化的敏感性相对较高.分析其原因, 一方面生态系统温度敏感性与气温有关(Kirschbaum, 1995; Xu & Qi, 2001; Janssens & Pilegaard, 2003; Davidson & Janssens., 2006; 张雷明等, 2006; Jassal et al., 2008), 气温对Q10产生影响主要通过影响光合与呼吸作用酶的活性实现(Davidson & Janssens, 2006).晴天气温T ((23.56 ± 2.62) ℃)显著大于阴天((21.26 ± 2.34) ℃), 阴天由于酶促反应需要一定的活化酶, 光合与呼吸作用酶的活性一般受到限制; 而晴天随着辐射增强, 气温升高, 越来越多的分子达到或超过了自身的活化能, 反应加快(Xiang & Freeman, 2009; Karhu et al., 2010).另一方面, 晴天相对阴天较高的PAR使得晴天光合作用更强, 产生输送的有机质增多(Curiel Yuste et al., 2007; Jassal et al., 2008), 自养呼吸增强, 同时地上植被以及土壤异养微生物呼吸对气温的响应能力增强(Edwards et al., 2004; Heinemeyer et al., 2006; Moyano et al., 2008), 而阴天由于底物基质供应不足使得自养、异养呼吸受到抑制(Gershenson et al., 2009). ...
Carbon dioxide exchange and canopy conductance of two coniferous forests under various sky conditions.
1
2010
... 在一定光照强度范围内, 光合作用随PAR的增强而增加(Glenn et al., 2006; Syed et al., 2006; Hao et al., 2011; 同小娟等, 2011).光能过剩会对光合作用产生不良影响, 出现光抑制, 致使叶绿体光合效率下降(Jiang et al., 2011; Tholen et al., 2012).晴天PAR显著大于阴天(图2B), 因此, 晴天条件下植被叶片光合作用可能大于阴天, 表现为晴天Amax显著大于阴天(表1).Zhou等(2009)研究认为Amax与植被的环境条件有关, 其中晴天Amax较阴天条件下提高了30%, 说明在植被生长旺季, 天空有云层覆盖时芦苇的光合能力比晴朗天气无云时有所降低, Law等(2002)总结全球森林、草地、农田以及苔原生态系统碳交换相关研究也得出类似结论.阴天条件下, 伴随天空云量的增多, PAR显著降低(图2B), 尽管阴天和晴天条件下, NEE均随PAR的增大而增加(图3), 但由于阴天PAR较低, 且晴天未出现光抑制现象, 因此其NEE显著低于晴天(图2A).Alton (2008)在分析北美洲和欧洲6种森林类型2-3年的碳通量观测数据后发现, 这些森林阴天的日固碳量比晴天少60%-80%.相反, 在研究太阳辐射对黄河小浪底人工混交林生态系统碳交换的影响中发现, 多云天气下净碳交换会增加(刘佳等, 2014).造成研究结论不一致的原因是多方面的, 不同的阴天和晴天划分指标(Graham et al., 2003; Dengel & Grace, 2010)和拟合模型(Zhang et al., 2011)来估计天气对碳通量的影响, 散射光和直射光下的碳交换没有一直处在相同的天气辐射条件下进行对比.此外, 研究地域的差异和不同植被类型的特性也在很大程度上影响了碳交换过程对天气的响应过程. ...
Effect of smoke on subcanopy shaded light, canopy temperature, and carbon dioxide uptake in an Amazon rainforest.
1
2010
... 本研究以黄河三角洲芦苇湿地为研究对象, 在自然条件下选择相邻的阴天和晴天, 在生物要素(生物量、叶面积指数)、土壤水分以及养分特征保持不变的前提下, 分析阴天和晴天引起的辐射变化对生态系统NEE的影响机制.目前, 有关太阳辐射对生态系统NEE的影响机制大都以晴空指数为指标(Gu et al., 2003; Liu et al., 2006; 范玉枝等, 2009; Zhang et al., 2010; 周丽艳等, 2010; 蒋琰等, 2011; Zhao et al., 2011; 刘佳等, 2014).这种研究方法的最大不足就是研究阶段中, 太阳辐射、生物要素(生物量、叶面积指数)、土壤湿度等环境因子都在发生变化, 这对阐明太阳辐射这一单因素对NEE的控制机制研究带来一定困难.而本研究巧妙地在自然条件下选择相邻的阴天和晴天, 在日尺度上实现了对单一变量PAR的控制.利用这种研究方法, Han等(2014a)通过对比分析阴天和晴天植物冠层光合作用的差异性, 定量分析了光合作用在日尺度上对土壤呼吸的调节作用.此外, 我们已有研究发现PAR改变所引起的植物光合作用变化对生态系统呼吸会产生一定的影响(Tang et al., 2005; Baldocchi et al., 2006; Moyano et al., 2008; Wingate et al., 2010; Han et al., 2012).但是, 以往的研究多通过夜间生态系统呼吸与气温函数关系外延的方法来估算白天生态系统呼吸(Doughty et al., 2010; Zhang et al., 2011), 忽略了PAR对白天生态系统呼吸的影响, 因而具有一定的局限性.考虑到白天PAR的变化, 本研究采用Michaelis-Menten模型对白天生态系统呼吸进行推算, 使得生态系统呼吸的温度响应数据更具有说服力. ...
Root production is determined by radiation flux in a temperate grassland community.
1
2004
... Q10值被广泛应用于评价生态系统呼吸对温度变化的敏感程度(杨庆朋等, 2011; 杨利琼等, 2013; Han et al., 2014a, 2014b), Q10值越大, 说明生态系统呼吸对温度变化的响应越敏感.黄河三角洲芦苇湿地生态系统阴天和晴天Q10值均在已有的湿地Q10范围(1.0-7.7)内(Chapman & Thurlow, 1996; Silvola et al., 1996; Lafleur et al., 2001; Bonneville et al., 2008; Zhou et al., 2009; 周丽艳等, 2010), 其中晴天白天Q10为5.5, 远高于阴天白天的Q10 (1.9), 说明晴天白天生态系统呼吸对温度变化的敏感性相对较高.分析其原因, 一方面生态系统温度敏感性与气温有关(Kirschbaum, 1995; Xu & Qi, 2001; Janssens & Pilegaard, 2003; Davidson & Janssens., 2006; 张雷明等, 2006; Jassal et al., 2008), 气温对Q10产生影响主要通过影响光合与呼吸作用酶的活性实现(Davidson & Janssens, 2006).晴天气温T ((23.56 ± 2.62) ℃)显著大于阴天((21.26 ± 2.34) ℃), 阴天由于酶促反应需要一定的活化酶, 光合与呼吸作用酶的活性一般受到限制; 而晴天随着辐射增强, 气温升高, 越来越多的分子达到或超过了自身的活化能, 反应加快(Xiang & Freeman, 2009; Karhu et al., 2010).另一方面, 晴天相对阴天较高的PAR使得晴天光合作用更强, 产生输送的有机质增多(Curiel Yuste et al., 2007; Jassal et al., 2008), 自养呼吸增强, 同时地上植被以及土壤异养微生物呼吸对气温的响应能力增强(Edwards et al., 2004; Heinemeyer et al., 2006; Moyano et al., 2008), 而阴天由于底物基质供应不足使得自养、异养呼吸受到抑制(Gershenson et al., 2009). ...
Gap filling strategies for defensible annual sums of net ecosystem exchange.
... 本文研究发现晴天和阴天芦苇湿地NEE对PAR的响应趋势一致, 均呈直角双曲线关系, 与先前的很多研究结果一致(Falge et al., 2001; Monson et al., 2002; Zhou et al., 2009; 张弥等, 2009; Han et al., 2014a, 2014b; 邢庆会等, 2014).PAR是控制阴天和晴天生态系统生长季昼间NEE变化的主要因素(Flanagan & Johnson, 2005; Liu et al., 2006; 薛红喜等, 2012), Michaelis-Menten方程拟合发现, PAR可以解释晴天单天NEE变化的57%-82%, 解释阴天单天NEE变化的53%-81%.云层变化会改变地表接收太阳辐射的强度, 影响PAR、T等环境因子, 进而影响阴天和晴天条件下NEE对光的响应特征(Baldocchi, 1997; Freedman et al., 2001). ...
散射辐射对西藏高原高寒草甸净生态系统CO2交换的影响
3
2009
... 已有研究表明, 阴天和晴天所引起的辐射差异影响着陆地生态系统的交换过程和碳吸收能力(Roderick et al., 2001; Still et al., 2009).但由于研究方法、地域环境条件以及不同植被类型的差异, 研究结论存在较大差异(Gu et al., 1999, 2002; Alton et al., 2007; Urban et al., 2007).例如, 中国渭北刺槐(Robinia pseudoacacia)生长初期和旺盛期晴天条件下森林日均固碳量比阴天高8%-16% (郑元等, 2011).中国西北玉米(Zea mays)农田生态系统阴天条件下净生态系统CO2交换(NEE)的增长速率却高于晴天, 且生态系统总初级生产力(GPP)最大值出现于阴天(Zhang et al., 2011).对中国西北草地(Bai et al., 2012)和不同温带森林类型(Zhang et al., 2010)的研究也得出类似结论.另外, 也有研究发现加拿大常绿灌木(Letts et al., 2005)和西藏高寒草甸(范玉枝等, 2009)生态系统CO2交换不受阴天和晴天天气条件影响. ...
... 黄河口近海与海岸湿地是暖温带增长速度最快和最具代表性的新生近海与海岸湿地, 具有海陆过渡性、原生性和脆弱性的特点(布仁仓等, 1999; 崔树强, 2002).受水沙量变化、陆地和河流淡水径流、咸水海流多重影响以及人类活动的干扰, 黄河三角洲湿地生态系统CO2交换存在着极大的复杂性和不确定性(Sasaki et al., 2009;Han et al., 2014a, 2014b; 邢庆会等, 2014).研究表明黄河三角洲芦苇湿地生长季NEE占全年的80%以上, 且碳吸收集中在植物生长旺盛期(7-9月)(杨利琼等, 2013; Han et al., 2014a, 2014b).而这一时期正值黄河三角洲雨季, 其云雨状况影响太阳辐射, 进而影响湿地生态系统的碳吸收功能.在以往的研究中, 温度、水分、太阳辐射等环境因子, 生物量、叶面积等生物因子以及地下水位、地表水深等水文要素对湿地生态系统NEE的影响机制受到研究者的极大关注(牟长城等, 2009; Zhao et al., 2010; Han et al., 2013; 邢庆会等, 2014).目前, 有关阴天和晴天条件对生态系统碳交换影响的研究主要集中在森林生态系统(Liu et al., 2006; Zhang et al., 2010; 周丽艳等, 2010; Jiang et al., 2011; Zhao et al., 2011; 刘佳等, 2014)和草原生态系统(Gu et al., 2003; 范玉枝等, 2009).阴天和晴天状况引起的太阳辐射变化对湿地生态系统NEE和碳汇功能究竟产生怎样的影响目前尚不清楚. ...
... 本研究以黄河三角洲芦苇湿地为研究对象, 在自然条件下选择相邻的阴天和晴天, 在生物要素(生物量、叶面积指数)、土壤水分以及养分特征保持不变的前提下, 分析阴天和晴天引起的辐射变化对生态系统NEE的影响机制.目前, 有关太阳辐射对生态系统NEE的影响机制大都以晴空指数为指标(Gu et al., 2003; Liu et al., 2006; 范玉枝等, 2009; Zhang et al., 2010; 周丽艳等, 2010; 蒋琰等, 2011; Zhao et al., 2011; 刘佳等, 2014).这种研究方法的最大不足就是研究阶段中, 太阳辐射、生物要素(生物量、叶面积指数)、土壤湿度等环境因子都在发生变化, 这对阐明太阳辐射这一单因素对NEE的控制机制研究带来一定困难.而本研究巧妙地在自然条件下选择相邻的阴天和晴天, 在日尺度上实现了对单一变量PAR的控制.利用这种研究方法, Han等(2014a)通过对比分析阴天和晴天植物冠层光合作用的差异性, 定量分析了光合作用在日尺度上对土壤呼吸的调节作用.此外, 我们已有研究发现PAR改变所引起的植物光合作用变化对生态系统呼吸会产生一定的影响(Tang et al., 2005; Baldocchi et al., 2006; Moyano et al., 2008; Wingate et al., 2010; Han et al., 2012).但是, 以往的研究多通过夜间生态系统呼吸与气温函数关系外延的方法来估算白天生态系统呼吸(Doughty et al., 2010; Zhang et al., 2011), 忽略了PAR对白天生态系统呼吸的影响, 因而具有一定的局限性.考虑到白天PAR的变化, 本研究采用Michaelis-Menten模型对白天生态系统呼吸进行推算, 使得生态系统呼吸的温度响应数据更具有说服力. ...
散射辐射对西藏高原高寒草甸净生态系统CO2交换的影响
3
2009
... 已有研究表明, 阴天和晴天所引起的辐射差异影响着陆地生态系统的交换过程和碳吸收能力(Roderick et al., 2001; Still et al., 2009).但由于研究方法、地域环境条件以及不同植被类型的差异, 研究结论存在较大差异(Gu et al., 1999, 2002; Alton et al., 2007; Urban et al., 2007).例如, 中国渭北刺槐(Robinia pseudoacacia)生长初期和旺盛期晴天条件下森林日均固碳量比阴天高8%-16% (郑元等, 2011).中国西北玉米(Zea mays)农田生态系统阴天条件下净生态系统CO2交换(NEE)的增长速率却高于晴天, 且生态系统总初级生产力(GPP)最大值出现于阴天(Zhang et al., 2011).对中国西北草地(Bai et al., 2012)和不同温带森林类型(Zhang et al., 2010)的研究也得出类似结论.另外, 也有研究发现加拿大常绿灌木(Letts et al., 2005)和西藏高寒草甸(范玉枝等, 2009)生态系统CO2交换不受阴天和晴天天气条件影响. ...
... 黄河口近海与海岸湿地是暖温带增长速度最快和最具代表性的新生近海与海岸湿地, 具有海陆过渡性、原生性和脆弱性的特点(布仁仓等, 1999; 崔树强, 2002).受水沙量变化、陆地和河流淡水径流、咸水海流多重影响以及人类活动的干扰, 黄河三角洲湿地生态系统CO2交换存在着极大的复杂性和不确定性(Sasaki et al., 2009;Han et al., 2014a, 2014b; 邢庆会等, 2014).研究表明黄河三角洲芦苇湿地生长季NEE占全年的80%以上, 且碳吸收集中在植物生长旺盛期(7-9月)(杨利琼等, 2013; Han et al., 2014a, 2014b).而这一时期正值黄河三角洲雨季, 其云雨状况影响太阳辐射, 进而影响湿地生态系统的碳吸收功能.在以往的研究中, 温度、水分、太阳辐射等环境因子, 生物量、叶面积等生物因子以及地下水位、地表水深等水文要素对湿地生态系统NEE的影响机制受到研究者的极大关注(牟长城等, 2009; Zhao et al., 2010; Han et al., 2013; 邢庆会等, 2014).目前, 有关阴天和晴天条件对生态系统碳交换影响的研究主要集中在森林生态系统(Liu et al., 2006; Zhang et al., 2010; 周丽艳等, 2010; Jiang et al., 2011; Zhao et al., 2011; 刘佳等, 2014)和草原生态系统(Gu et al., 2003; 范玉枝等, 2009).阴天和晴天状况引起的太阳辐射变化对湿地生态系统NEE和碳汇功能究竟产生怎样的影响目前尚不清楚. ...
... 本研究以黄河三角洲芦苇湿地为研究对象, 在自然条件下选择相邻的阴天和晴天, 在生物要素(生物量、叶面积指数)、土壤水分以及养分特征保持不变的前提下, 分析阴天和晴天引起的辐射变化对生态系统NEE的影响机制.目前, 有关太阳辐射对生态系统NEE的影响机制大都以晴空指数为指标(Gu et al., 2003; Liu et al., 2006; 范玉枝等, 2009; Zhang et al., 2010; 周丽艳等, 2010; 蒋琰等, 2011; Zhao et al., 2011; 刘佳等, 2014).这种研究方法的最大不足就是研究阶段中, 太阳辐射、生物要素(生物量、叶面积指数)、土壤湿度等环境因子都在发生变化, 这对阐明太阳辐射这一单因素对NEE的控制机制研究带来一定困难.而本研究巧妙地在自然条件下选择相邻的阴天和晴天, 在日尺度上实现了对单一变量PAR的控制.利用这种研究方法, Han等(2014a)通过对比分析阴天和晴天植物冠层光合作用的差异性, 定量分析了光合作用在日尺度上对土壤呼吸的调节作用.此外, 我们已有研究发现PAR改变所引起的植物光合作用变化对生态系统呼吸会产生一定的影响(Tang et al., 2005; Baldocchi et al., 2006; Moyano et al., 2008; Wingate et al., 2010; Han et al., 2012).但是, 以往的研究多通过夜间生态系统呼吸与气温函数关系外延的方法来估算白天生态系统呼吸(Doughty et al., 2010; Zhang et al., 2011), 忽略了PAR对白天生态系统呼吸的影响, 因而具有一定的局限性.考虑到白天PAR的变化, 本研究采用Michaelis-Menten模型对白天生态系统呼吸进行推算, 使得生态系统呼吸的温度响应数据更具有说服力. ...
Interacting effects of temperature, soil moisture and plant biomass production on ecosystem respiration in a northern temperate grassland.
1
2005
... 本文研究发现晴天和阴天芦苇湿地NEE对PAR的响应趋势一致, 均呈直角双曲线关系, 与先前的很多研究结果一致(Falge et al., 2001; Monson et al., 2002; Zhou et al., 2009; 张弥等, 2009; Han et al., 2014a, 2014b; 邢庆会等, 2014).PAR是控制阴天和晴天生态系统生长季昼间NEE变化的主要因素(Flanagan & Johnson, 2005; Liu et al., 2006; 薛红喜等, 2012), Michaelis-Menten方程拟合发现, PAR可以解释晴天单天NEE变化的57%-82%, 解释阴天单天NEE变化的53%-81%.云层变化会改变地表接收太阳辐射的强度, 影响PAR、T等环境因子, 进而影响阴天和晴天条件下NEE对光的响应特征(Baldocchi, 1997; Freedman et al., 2001). ...
Boundary layer clouds and vegetation-atmosphere feedbacks.
1
2001
... 本文研究发现晴天和阴天芦苇湿地NEE对PAR的响应趋势一致, 均呈直角双曲线关系, 与先前的很多研究结果一致(Falge et al., 2001; Monson et al., 2002; Zhou et al., 2009; 张弥等, 2009; Han et al., 2014a, 2014b; 邢庆会等, 2014).PAR是控制阴天和晴天生态系统生长季昼间NEE变化的主要因素(Flanagan & Johnson, 2005; Liu et al., 2006; 薛红喜等, 2012), Michaelis-Menten方程拟合发现, PAR可以解释晴天单天NEE变化的57%-82%, 解释阴天单天NEE变化的53%-81%.云层变化会改变地表接收太阳辐射的强度, 影响PAR、T等环境因子, 进而影响阴天和晴天条件下NEE对光的响应特征(Baldocchi, 1997; Freedman et al., 2001). ...
Effects of substrate availability on the temperature sensitivity of soil organic matter decomposition.
1
2009
... Q10值被广泛应用于评价生态系统呼吸对温度变化的敏感程度(杨庆朋等, 2011; 杨利琼等, 2013; Han et al., 2014a, 2014b), Q10值越大, 说明生态系统呼吸对温度变化的响应越敏感.黄河三角洲芦苇湿地生态系统阴天和晴天Q10值均在已有的湿地Q10范围(1.0-7.7)内(Chapman & Thurlow, 1996; Silvola et al., 1996; Lafleur et al., 2001; Bonneville et al., 2008; Zhou et al., 2009; 周丽艳等, 2010), 其中晴天白天Q10为5.5, 远高于阴天白天的Q10 (1.9), 说明晴天白天生态系统呼吸对温度变化的敏感性相对较高.分析其原因, 一方面生态系统温度敏感性与气温有关(Kirschbaum, 1995; Xu & Qi, 2001; Janssens & Pilegaard, 2003; Davidson & Janssens., 2006; 张雷明等, 2006; Jassal et al., 2008), 气温对Q10产生影响主要通过影响光合与呼吸作用酶的活性实现(Davidson & Janssens, 2006).晴天气温T ((23.56 ± 2.62) ℃)显著大于阴天((21.26 ± 2.34) ℃), 阴天由于酶促反应需要一定的活化酶, 光合与呼吸作用酶的活性一般受到限制; 而晴天随着辐射增强, 气温升高, 越来越多的分子达到或超过了自身的活化能, 反应加快(Xiang & Freeman, 2009; Karhu et al., 2010).另一方面, 晴天相对阴天较高的PAR使得晴天光合作用更强, 产生输送的有机质增多(Curiel Yuste et al., 2007; Jassal et al., 2008), 自养呼吸增强, 同时地上植被以及土壤异养微生物呼吸对气温的响应能力增强(Edwards et al., 2004; Heinemeyer et al., 2006; Moyano et al., 2008), 而阴天由于底物基质供应不足使得自养、异养呼吸受到抑制(Gershenson et al., 2009). ...
Comparison of net ecosystem CO2 exchange in two peatlands in western Canada with contrasting dominant vegetation, Sphagnum and Carex.
1
2006
... 在一定光照强度范围内, 光合作用随PAR的增强而增加(Glenn et al., 2006; Syed et al., 2006; Hao et al., 2011; 同小娟等, 2011).光能过剩会对光合作用产生不良影响, 出现光抑制, 致使叶绿体光合效率下降(Jiang et al., 2011; Tholen et al., 2012).晴天PAR显著大于阴天(图2B), 因此, 晴天条件下植被叶片光合作用可能大于阴天, 表现为晴天Amax显著大于阴天(表1).Zhou等(2009)研究认为Amax与植被的环境条件有关, 其中晴天Amax较阴天条件下提高了30%, 说明在植被生长旺季, 天空有云层覆盖时芦苇的光合能力比晴朗天气无云时有所降低, Law等(2002)总结全球森林、草地、农田以及苔原生态系统碳交换相关研究也得出类似结论.阴天条件下, 伴随天空云量的增多, PAR显著降低(图2B), 尽管阴天和晴天条件下, NEE均随PAR的增大而增加(图3), 但由于阴天PAR较低, 且晴天未出现光抑制现象, 因此其NEE显著低于晴天(图2A).Alton (2008)在分析北美洲和欧洲6种森林类型2-3年的碳通量观测数据后发现, 这些森林阴天的日固碳量比晴天少60%-80%.相反, 在研究太阳辐射对黄河小浪底人工混交林生态系统碳交换的影响中发现, 多云天气下净碳交换会增加(刘佳等, 2014).造成研究结论不一致的原因是多方面的, 不同的阴天和晴天划分指标(Graham et al., 2003; Dengel & Grace, 2010)和拟合模型(Zhang et al., 2011)来估计天气对碳通量的影响, 散射光和直射光下的碳交换没有一直处在相同的天气辐射条件下进行对比.此外, 研究地域的差异和不同植被类型的特性也在很大程度上影响了碳交换过程对天气的响应过程. ...
Cloud cover limits net CO2 uptake and growth of a rainforest tree during tropical rainy seasons.
1
2003
... 在一定光照强度范围内, 光合作用随PAR的增强而增加(Glenn et al., 2006; Syed et al., 2006; Hao et al., 2011; 同小娟等, 2011).光能过剩会对光合作用产生不良影响, 出现光抑制, 致使叶绿体光合效率下降(Jiang et al., 2011; Tholen et al., 2012).晴天PAR显著大于阴天(图2B), 因此, 晴天条件下植被叶片光合作用可能大于阴天, 表现为晴天Amax显著大于阴天(表1).Zhou等(2009)研究认为Amax与植被的环境条件有关, 其中晴天Amax较阴天条件下提高了30%, 说明在植被生长旺季, 天空有云层覆盖时芦苇的光合能力比晴朗天气无云时有所降低, Law等(2002)总结全球森林、草地、农田以及苔原生态系统碳交换相关研究也得出类似结论.阴天条件下, 伴随天空云量的增多, PAR显著降低(图2B), 尽管阴天和晴天条件下, NEE均随PAR的增大而增加(图3), 但由于阴天PAR较低, 且晴天未出现光抑制现象, 因此其NEE显著低于晴天(图2A).Alton (2008)在分析北美洲和欧洲6种森林类型2-3年的碳通量观测数据后发现, 这些森林阴天的日固碳量比晴天少60%-80%.相反, 在研究太阳辐射对黄河小浪底人工混交林生态系统碳交换的影响中发现, 多云天气下净碳交换会增加(刘佳等, 2014).造成研究结论不一致的原因是多方面的, 不同的阴天和晴天划分指标(Graham et al., 2003; Dengel & Grace, 2010)和拟合模型(Zhang et al., 2011)来估计天气对碳通量的影响, 散射光和直射光下的碳交换没有一直处在相同的天气辐射条件下进行对比.此外, 研究地域的差异和不同植被类型的特性也在很大程度上影响了碳交换过程对天气的响应过程. ...
Advantages of diffuse radiation for terrestrial ecosystem productivity.
1
2002
... 已有研究表明, 阴天和晴天所引起的辐射差异影响着陆地生态系统的交换过程和碳吸收能力(Roderick et al., 2001; Still et al., 2009).但由于研究方法、地域环境条件以及不同植被类型的差异, 研究结论存在较大差异(Gu et al., 1999, 2002; Alton et al., 2007; Urban et al., 2007).例如, 中国渭北刺槐(Robinia pseudoacacia)生长初期和旺盛期晴天条件下森林日均固碳量比阴天高8%-16% (郑元等, 2011).中国西北玉米(Zea mays)农田生态系统阴天条件下净生态系统CO2交换(NEE)的增长速率却高于晴天, 且生态系统总初级生产力(GPP)最大值出现于阴天(Zhang et al., 2011).对中国西北草地(Bai et al., 2012)和不同温带森林类型(Zhang et al., 2010)的研究也得出类似结论.另外, 也有研究发现加拿大常绿灌木(Letts et al., 2005)和西藏高寒草甸(范玉枝等, 2009)生态系统CO2交换不受阴天和晴天天气条件影响. ...
Responses of net ecosystem exchanges of carbon dioxide to changes in cloudiness: Results from two North American deciduous forests.
1
1999
... 已有研究表明, 阴天和晴天所引起的辐射差异影响着陆地生态系统的交换过程和碳吸收能力(Roderick et al., 2001; Still et al., 2009).但由于研究方法、地域环境条件以及不同植被类型的差异, 研究结论存在较大差异(Gu et al., 1999, 2002; Alton et al., 2007; Urban et al., 2007).例如, 中国渭北刺槐(Robinia pseudoacacia)生长初期和旺盛期晴天条件下森林日均固碳量比阴天高8%-16% (郑元等, 2011).中国西北玉米(Zea mays)农田生态系统阴天条件下净生态系统CO2交换(NEE)的增长速率却高于晴天, 且生态系统总初级生产力(GPP)最大值出现于阴天(Zhang et al., 2011).对中国西北草地(Bai et al., 2012)和不同温带森林类型(Zhang et al., 2010)的研究也得出类似结论.另外, 也有研究发现加拿大常绿灌木(Letts et al., 2005)和西藏高寒草甸(范玉枝等, 2009)生态系统CO2交换不受阴天和晴天天气条件影响. ...
Short-term variation of CO2 flux in relation to environmental controls in an alpine meadow on the Qinghai- Tibetan Plateau.
2
2003
... 黄河口近海与海岸湿地是暖温带增长速度最快和最具代表性的新生近海与海岸湿地, 具有海陆过渡性、原生性和脆弱性的特点(布仁仓等, 1999; 崔树强, 2002).受水沙量变化、陆地和河流淡水径流、咸水海流多重影响以及人类活动的干扰, 黄河三角洲湿地生态系统CO2交换存在着极大的复杂性和不确定性(Sasaki et al., 2009;Han et al., 2014a, 2014b; 邢庆会等, 2014).研究表明黄河三角洲芦苇湿地生长季NEE占全年的80%以上, 且碳吸收集中在植物生长旺盛期(7-9月)(杨利琼等, 2013; Han et al., 2014a, 2014b).而这一时期正值黄河三角洲雨季, 其云雨状况影响太阳辐射, 进而影响湿地生态系统的碳吸收功能.在以往的研究中, 温度、水分、太阳辐射等环境因子, 生物量、叶面积等生物因子以及地下水位、地表水深等水文要素对湿地生态系统NEE的影响机制受到研究者的极大关注(牟长城等, 2009; Zhao et al., 2010; Han et al., 2013; 邢庆会等, 2014).目前, 有关阴天和晴天条件对生态系统碳交换影响的研究主要集中在森林生态系统(Liu et al., 2006; Zhang et al., 2010; 周丽艳等, 2010; Jiang et al., 2011; Zhao et al., 2011; 刘佳等, 2014)和草原生态系统(Gu et al., 2003; 范玉枝等, 2009).阴天和晴天状况引起的太阳辐射变化对湿地生态系统NEE和碳汇功能究竟产生怎样的影响目前尚不清楚. ...
... 本研究以黄河三角洲芦苇湿地为研究对象, 在自然条件下选择相邻的阴天和晴天, 在生物要素(生物量、叶面积指数)、土壤水分以及养分特征保持不变的前提下, 分析阴天和晴天引起的辐射变化对生态系统NEE的影响机制.目前, 有关太阳辐射对生态系统NEE的影响机制大都以晴空指数为指标(Gu et al., 2003; Liu et al., 2006; 范玉枝等, 2009; Zhang et al., 2010; 周丽艳等, 2010; 蒋琰等, 2011; Zhao et al., 2011; 刘佳等, 2014).这种研究方法的最大不足就是研究阶段中, 太阳辐射、生物要素(生物量、叶面积指数)、土壤湿度等环境因子都在发生变化, 这对阐明太阳辐射这一单因素对NEE的控制机制研究带来一定困难.而本研究巧妙地在自然条件下选择相邻的阴天和晴天, 在日尺度上实现了对单一变量PAR的控制.利用这种研究方法, Han等(2014a)通过对比分析阴天和晴天植物冠层光合作用的差异性, 定量分析了光合作用在日尺度上对土壤呼吸的调节作用.此外, 我们已有研究发现PAR改变所引起的植物光合作用变化对生态系统呼吸会产生一定的影响(Tang et al., 2005; Baldocchi et al., 2006; Moyano et al., 2008; Wingate et al., 2010; Han et al., 2012).但是, 以往的研究多通过夜间生态系统呼吸与气温函数关系外延的方法来估算白天生态系统呼吸(Doughty et al., 2010; Zhang et al., 2011), 忽略了PAR对白天生态系统呼吸的影响, 因而具有一定的局限性.考虑到白天PAR的变化, 本研究采用Michaelis-Menten模型对白天生态系统呼吸进行推算, 使得生态系统呼吸的温度响应数据更具有说服力. ...
Ecosystem photosynthesis regulates soil respiration on a diurnal scale with a short-term time lag in a coastal wetland.
6
2014a
... 黄河口近海与海岸湿地是暖温带增长速度最快和最具代表性的新生近海与海岸湿地, 具有海陆过渡性、原生性和脆弱性的特点(布仁仓等, 1999; 崔树强, 2002).受水沙量变化、陆地和河流淡水径流、咸水海流多重影响以及人类活动的干扰, 黄河三角洲湿地生态系统CO2交换存在着极大的复杂性和不确定性(Sasaki et al., 2009;Han et al., 2014a, 2014b; 邢庆会等, 2014).研究表明黄河三角洲芦苇湿地生长季NEE占全年的80%以上, 且碳吸收集中在植物生长旺盛期(7-9月)(杨利琼等, 2013; Han et al., 2014a, 2014b).而这一时期正值黄河三角洲雨季, 其云雨状况影响太阳辐射, 进而影响湿地生态系统的碳吸收功能.在以往的研究中, 温度、水分、太阳辐射等环境因子, 生物量、叶面积等生物因子以及地下水位、地表水深等水文要素对湿地生态系统NEE的影响机制受到研究者的极大关注(牟长城等, 2009; Zhao et al., 2010; Han et al., 2013; 邢庆会等, 2014).目前, 有关阴天和晴天条件对生态系统碳交换影响的研究主要集中在森林生态系统(Liu et al., 2006; Zhang et al., 2010; 周丽艳等, 2010; Jiang et al., 2011; Zhao et al., 2011; 刘佳等, 2014)和草原生态系统(Gu et al., 2003; 范玉枝等, 2009).阴天和晴天状况引起的太阳辐射变化对湿地生态系统NEE和碳汇功能究竟产生怎样的影响目前尚不清楚. ...
... ; Han et al., 2014a, 2014b).而这一时期正值黄河三角洲雨季, 其云雨状况影响太阳辐射, 进而影响湿地生态系统的碳吸收功能.在以往的研究中, 温度、水分、太阳辐射等环境因子, 生物量、叶面积等生物因子以及地下水位、地表水深等水文要素对湿地生态系统NEE的影响机制受到研究者的极大关注(牟长城等, 2009; Zhao et al., 2010; Han et al., 2013; 邢庆会等, 2014).目前, 有关阴天和晴天条件对生态系统碳交换影响的研究主要集中在森林生态系统(Liu et al., 2006; Zhang et al., 2010; 周丽艳等, 2010; Jiang et al., 2011; Zhao et al., 2011; 刘佳等, 2014)和草原生态系统(Gu et al., 2003; 范玉枝等, 2009).阴天和晴天状况引起的太阳辐射变化对湿地生态系统NEE和碳汇功能究竟产生怎样的影响目前尚不清楚. ...
... 本文研究发现晴天和阴天芦苇湿地NEE对PAR的响应趋势一致, 均呈直角双曲线关系, 与先前的很多研究结果一致(Falge et al., 2001; Monson et al., 2002; Zhou et al., 2009; 张弥等, 2009; Han et al., 2014a, 2014b; 邢庆会等, 2014).PAR是控制阴天和晴天生态系统生长季昼间NEE变化的主要因素(Flanagan & Johnson, 2005; Liu et al., 2006; 薛红喜等, 2012), Michaelis-Menten方程拟合发现, PAR可以解释晴天单天NEE变化的57%-82%, 解释阴天单天NEE变化的53%-81%.云层变化会改变地表接收太阳辐射的强度, 影响PAR、T等环境因子, 进而影响阴天和晴天条件下NEE对光的响应特征(Baldocchi, 1997; Freedman et al., 2001). ...
... 另外, 太阳辐射变化引起的温度变化也影响NEE对光的响应特征.首先, 温度通过影响酶系列化学反应对光合作用产生影响(Syed et al., 2006), 温度升高有利于光合速率的提高(Pingintha et al., 2010).相关分析发现, 阴天和晴天Amax与T均为显著的线性正相关(晴天: R2 = 0.80, p < 0.05; 阴天: R2 = 0.63, p < 0.05), 其次, 生态系统呼吸与气温通常显著正相关(Aires et al., 2008; Pingintha et al., 2010).本研究中虽然晴天条件下Reco,daytime较阴天提高了20%, 但NEE却显著高于阴天的67%, 说明晴天温度对湿地光合固碳能力的提高远超过生态系统呼吸的消耗, 表现为晴天生态系统的碳汇功能更强, 这与已有的研究结果一致(Han et al., 2014a). ...
... 阴天和晴天生态系统Reco,daytime与T均呈显著的正相关关系, 两者呈指数关系, 与以往很多研究的结果一致(石培礼等, 2006; Zhou et al., 2009; Zhao et al., 2010; Han et al., 2013, 2014a; 邢庆会等, 2014).T是影响生态系统呼吸的主要因素(Law et al., 2002; Reichstein et al., 2005; Urban et al., 2007), 本文研究发现: 黄河三角洲芦苇湿地晴天气温相对阴天提高了10%, 其阴天和晴天的气温差高于美国阿尔卑斯山脉南部森林的阴天和晴天的气温差(Berry & Smith, 2012), 黄河三角洲芦苇湿地晴天对应的生态系统呼吸相对阴天提高了20% (p < 0.05), 说明阴天条件下, 植被地上部分尤其是叶面温度较晴朗天气强太阳辐射条件下的低, 叶片以及植被茎干呼吸受到一定的抑制(Urban et al., 2007). ...
... Q10值被广泛应用于评价生态系统呼吸对温度变化的敏感程度(杨庆朋等, 2011; 杨利琼等, 2013; Han et al., 2014a, 2014b), Q10值越大, 说明生态系统呼吸对温度变化的响应越敏感.黄河三角洲芦苇湿地生态系统阴天和晴天Q10值均在已有的湿地Q10范围(1.0-7.7)内(Chapman & Thurlow, 1996; Silvola et al., 1996; Lafleur et al., 2001; Bonneville et al., 2008; Zhou et al., 2009; 周丽艳等, 2010), 其中晴天白天Q10为5.5, 远高于阴天白天的Q10 (1.9), 说明晴天白天生态系统呼吸对温度变化的敏感性相对较高.分析其原因, 一方面生态系统温度敏感性与气温有关(Kirschbaum, 1995; Xu & Qi, 2001; Janssens & Pilegaard, 2003; Davidson & Janssens., 2006; 张雷明等, 2006; Jassal et al., 2008), 气温对Q10产生影响主要通过影响光合与呼吸作用酶的活性实现(Davidson & Janssens, 2006).晴天气温T ((23.56 ± 2.62) ℃)显著大于阴天((21.26 ± 2.34) ℃), 阴天由于酶促反应需要一定的活化酶, 光合与呼吸作用酶的活性一般受到限制; 而晴天随着辐射增强, 气温升高, 越来越多的分子达到或超过了自身的活化能, 反应加快(Xiang & Freeman, 2009; Karhu et al., 2010).另一方面, 晴天相对阴天较高的PAR使得晴天光合作用更强, 产生输送的有机质增多(Curiel Yuste et al., 2007; Jassal et al., 2008), 自养呼吸增强, 同时地上植被以及土壤异养微生物呼吸对气温的响应能力增强(Edwards et al., 2004; Heinemeyer et al., 2006; Moyano et al., 2008), 而阴天由于底物基质供应不足使得自养、异养呼吸受到抑制(Gershenson et al., 2009). ...
Agricultural reclamation effects on ecosystem CO2 exchange of a coastal wetland in the Yellow River Delta.
4
2014b
... 黄河口近海与海岸湿地是暖温带增长速度最快和最具代表性的新生近海与海岸湿地, 具有海陆过渡性、原生性和脆弱性的特点(布仁仓等, 1999; 崔树强, 2002).受水沙量变化、陆地和河流淡水径流、咸水海流多重影响以及人类活动的干扰, 黄河三角洲湿地生态系统CO2交换存在着极大的复杂性和不确定性(Sasaki et al., 2009;Han et al., 2014a, 2014b; 邢庆会等, 2014).研究表明黄河三角洲芦苇湿地生长季NEE占全年的80%以上, 且碳吸收集中在植物生长旺盛期(7-9月)(杨利琼等, 2013; Han et al., 2014a, 2014b).而这一时期正值黄河三角洲雨季, 其云雨状况影响太阳辐射, 进而影响湿地生态系统的碳吸收功能.在以往的研究中, 温度、水分、太阳辐射等环境因子, 生物量、叶面积等生物因子以及地下水位、地表水深等水文要素对湿地生态系统NEE的影响机制受到研究者的极大关注(牟长城等, 2009; Zhao et al., 2010; Han et al., 2013; 邢庆会等, 2014).目前, 有关阴天和晴天条件对生态系统碳交换影响的研究主要集中在森林生态系统(Liu et al., 2006; Zhang et al., 2010; 周丽艳等, 2010; Jiang et al., 2011; Zhao et al., 2011; 刘佳等, 2014)和草原生态系统(Gu et al., 2003; 范玉枝等, 2009).阴天和晴天状况引起的太阳辐射变化对湿地生态系统NEE和碳汇功能究竟产生怎样的影响目前尚不清楚. ...
... , 2014b).而这一时期正值黄河三角洲雨季, 其云雨状况影响太阳辐射, 进而影响湿地生态系统的碳吸收功能.在以往的研究中, 温度、水分、太阳辐射等环境因子, 生物量、叶面积等生物因子以及地下水位、地表水深等水文要素对湿地生态系统NEE的影响机制受到研究者的极大关注(牟长城等, 2009; Zhao et al., 2010; Han et al., 2013; 邢庆会等, 2014).目前, 有关阴天和晴天条件对生态系统碳交换影响的研究主要集中在森林生态系统(Liu et al., 2006; Zhang et al., 2010; 周丽艳等, 2010; Jiang et al., 2011; Zhao et al., 2011; 刘佳等, 2014)和草原生态系统(Gu et al., 2003; 范玉枝等, 2009).阴天和晴天状况引起的太阳辐射变化对湿地生态系统NEE和碳汇功能究竟产生怎样的影响目前尚不清楚. ...
... 本文研究发现晴天和阴天芦苇湿地NEE对PAR的响应趋势一致, 均呈直角双曲线关系, 与先前的很多研究结果一致(Falge et al., 2001; Monson et al., 2002; Zhou et al., 2009; 张弥等, 2009; Han et al., 2014a, 2014b; 邢庆会等, 2014).PAR是控制阴天和晴天生态系统生长季昼间NEE变化的主要因素(Flanagan & Johnson, 2005; Liu et al., 2006; 薛红喜等, 2012), Michaelis-Menten方程拟合发现, PAR可以解释晴天单天NEE变化的57%-82%, 解释阴天单天NEE变化的53%-81%.云层变化会改变地表接收太阳辐射的强度, 影响PAR、T等环境因子, 进而影响阴天和晴天条件下NEE对光的响应特征(Baldocchi, 1997; Freedman et al., 2001). ...
... Q10值被广泛应用于评价生态系统呼吸对温度变化的敏感程度(杨庆朋等, 2011; 杨利琼等, 2013; Han et al., 2014a, 2014b), Q10值越大, 说明生态系统呼吸对温度变化的响应越敏感.黄河三角洲芦苇湿地生态系统阴天和晴天Q10值均在已有的湿地Q10范围(1.0-7.7)内(Chapman & Thurlow, 1996; Silvola et al., 1996; Lafleur et al., 2001; Bonneville et al., 2008; Zhou et al., 2009; 周丽艳等, 2010), 其中晴天白天Q10为5.5, 远高于阴天白天的Q10 (1.9), 说明晴天白天生态系统呼吸对温度变化的敏感性相对较高.分析其原因, 一方面生态系统温度敏感性与气温有关(Kirschbaum, 1995; Xu & Qi, 2001; Janssens & Pilegaard, 2003; Davidson & Janssens., 2006; 张雷明等, 2006; Jassal et al., 2008), 气温对Q10产生影响主要通过影响光合与呼吸作用酶的活性实现(Davidson & Janssens, 2006).晴天气温T ((23.56 ± 2.62) ℃)显著大于阴天((21.26 ± 2.34) ℃), 阴天由于酶促反应需要一定的活化酶, 光合与呼吸作用酶的活性一般受到限制; 而晴天随着辐射增强, 气温升高, 越来越多的分子达到或超过了自身的活化能, 反应加快(Xiang & Freeman, 2009; Karhu et al., 2010).另一方面, 晴天相对阴天较高的PAR使得晴天光合作用更强, 产生输送的有机质增多(Curiel Yuste et al., 2007; Jassal et al., 2008), 自养呼吸增强, 同时地上植被以及土壤异养微生物呼吸对气温的响应能力增强(Edwards et al., 2004; Heinemeyer et al., 2006; Moyano et al., 2008), 而阴天由于底物基质供应不足使得自养、异养呼吸受到抑制(Gershenson et al., 2009). ...
Environmental controls on net ecosystem CO2 exchange over a reed (Phragmites australis) wetland in the Yellow River Delta, China.
2
2013
... 黄河口近海与海岸湿地是暖温带增长速度最快和最具代表性的新生近海与海岸湿地, 具有海陆过渡性、原生性和脆弱性的特点(布仁仓等, 1999; 崔树强, 2002).受水沙量变化、陆地和河流淡水径流、咸水海流多重影响以及人类活动的干扰, 黄河三角洲湿地生态系统CO2交换存在着极大的复杂性和不确定性(Sasaki et al., 2009;Han et al., 2014a, 2014b; 邢庆会等, 2014).研究表明黄河三角洲芦苇湿地生长季NEE占全年的80%以上, 且碳吸收集中在植物生长旺盛期(7-9月)(杨利琼等, 2013; Han et al., 2014a, 2014b).而这一时期正值黄河三角洲雨季, 其云雨状况影响太阳辐射, 进而影响湿地生态系统的碳吸收功能.在以往的研究中, 温度、水分、太阳辐射等环境因子, 生物量、叶面积等生物因子以及地下水位、地表水深等水文要素对湿地生态系统NEE的影响机制受到研究者的极大关注(牟长城等, 2009; Zhao et al., 2010; Han et al., 2013; 邢庆会等, 2014).目前, 有关阴天和晴天条件对生态系统碳交换影响的研究主要集中在森林生态系统(Liu et al., 2006; Zhang et al., 2010; 周丽艳等, 2010; Jiang et al., 2011; Zhao et al., 2011; 刘佳等, 2014)和草原生态系统(Gu et al., 2003; 范玉枝等, 2009).阴天和晴天状况引起的太阳辐射变化对湿地生态系统NEE和碳汇功能究竟产生怎样的影响目前尚不清楚. ...
... 阴天和晴天生态系统Reco,daytime与T均呈显著的正相关关系, 两者呈指数关系, 与以往很多研究的结果一致(石培礼等, 2006; Zhou et al., 2009; Zhao et al., 2010; Han et al., 2013, 2014a; 邢庆会等, 2014).T是影响生态系统呼吸的主要因素(Law et al., 2002; Reichstein et al., 2005; Urban et al., 2007), 本文研究发现: 黄河三角洲芦苇湿地晴天气温相对阴天提高了10%, 其阴天和晴天的气温差高于美国阿尔卑斯山脉南部森林的阴天和晴天的气温差(Berry & Smith, 2012), 黄河三角洲芦苇湿地晴天对应的生态系统呼吸相对阴天提高了20% (p < 0.05), 说明阴天条件下, 植被地上部分尤其是叶面温度较晴朗天气强太阳辐射条件下的低, 叶片以及植被茎干呼吸受到一定的抑制(Urban et al., 2007). ...
Winter soil respiration from different vegetation patches in the Yellow River Delta, China.
1
2012
... 本研究以黄河三角洲芦苇湿地为研究对象, 在自然条件下选择相邻的阴天和晴天, 在生物要素(生物量、叶面积指数)、土壤水分以及养分特征保持不变的前提下, 分析阴天和晴天引起的辐射变化对生态系统NEE的影响机制.目前, 有关太阳辐射对生态系统NEE的影响机制大都以晴空指数为指标(Gu et al., 2003; Liu et al., 2006; 范玉枝等, 2009; Zhang et al., 2010; 周丽艳等, 2010; 蒋琰等, 2011; Zhao et al., 2011; 刘佳等, 2014).这种研究方法的最大不足就是研究阶段中, 太阳辐射、生物要素(生物量、叶面积指数)、土壤湿度等环境因子都在发生变化, 这对阐明太阳辐射这一单因素对NEE的控制机制研究带来一定困难.而本研究巧妙地在自然条件下选择相邻的阴天和晴天, 在日尺度上实现了对单一变量PAR的控制.利用这种研究方法, Han等(2014a)通过对比分析阴天和晴天植物冠层光合作用的差异性, 定量分析了光合作用在日尺度上对土壤呼吸的调节作用.此外, 我们已有研究发现PAR改变所引起的植物光合作用变化对生态系统呼吸会产生一定的影响(Tang et al., 2005; Baldocchi et al., 2006; Moyano et al., 2008; Wingate et al., 2010; Han et al., 2012).但是, 以往的研究多通过夜间生态系统呼吸与气温函数关系外延的方法来估算白天生态系统呼吸(Doughty et al., 2010; Zhang et al., 2011), 忽略了PAR对白天生态系统呼吸的影响, 因而具有一定的局限性.考虑到白天PAR的变化, 本研究采用Michaelis-Menten模型对白天生态系统呼吸进行推算, 使得生态系统呼吸的温度响应数据更具有说服力. ...
Predominance of precipitation and temperature controls on ecosystem CO2 exchange in Zoige alpine wetlands of southwest China.
1
2011
... 在一定光照强度范围内, 光合作用随PAR的增强而增加(Glenn et al., 2006; Syed et al., 2006; Hao et al., 2011; 同小娟等, 2011).光能过剩会对光合作用产生不良影响, 出现光抑制, 致使叶绿体光合效率下降(Jiang et al., 2011; Tholen et al., 2012).晴天PAR显著大于阴天(图2B), 因此, 晴天条件下植被叶片光合作用可能大于阴天, 表现为晴天Amax显著大于阴天(表1).Zhou等(2009)研究认为Amax与植被的环境条件有关, 其中晴天Amax较阴天条件下提高了30%, 说明在植被生长旺季, 天空有云层覆盖时芦苇的光合能力比晴朗天气无云时有所降低, Law等(2002)总结全球森林、草地、农田以及苔原生态系统碳交换相关研究也得出类似结论.阴天条件下, 伴随天空云量的增多, PAR显著降低(图2B), 尽管阴天和晴天条件下, NEE均随PAR的增大而增加(图3), 但由于阴天PAR较低, 且晴天未出现光抑制现象, 因此其NEE显著低于晴天(图2A).Alton (2008)在分析北美洲和欧洲6种森林类型2-3年的碳通量观测数据后发现, 这些森林阴天的日固碳量比晴天少60%-80%.相反, 在研究太阳辐射对黄河小浪底人工混交林生态系统碳交换的影响中发现, 多云天气下净碳交换会增加(刘佳等, 2014).造成研究结论不一致的原因是多方面的, 不同的阴天和晴天划分指标(Graham et al., 2003; Dengel & Grace, 2010)和拟合模型(Zhang et al., 2011)来估计天气对碳通量的影响, 散射光和直射光下的碳交换没有一直处在相同的天气辐射条件下进行对比.此外, 研究地域的差异和不同植被类型的特性也在很大程度上影响了碳交换过程对天气的响应过程. ...
Respiration of the external mycelium in the arbuscular mycorrhizal symbiosis shows strong dependence on recent photosynthates and acclimation to temperature.
1
2006
... Q10值被广泛应用于评价生态系统呼吸对温度变化的敏感程度(杨庆朋等, 2011; 杨利琼等, 2013; Han et al., 2014a, 2014b), Q10值越大, 说明生态系统呼吸对温度变化的响应越敏感.黄河三角洲芦苇湿地生态系统阴天和晴天Q10值均在已有的湿地Q10范围(1.0-7.7)内(Chapman & Thurlow, 1996; Silvola et al., 1996; Lafleur et al., 2001; Bonneville et al., 2008; Zhou et al., 2009; 周丽艳等, 2010), 其中晴天白天Q10为5.5, 远高于阴天白天的Q10 (1.9), 说明晴天白天生态系统呼吸对温度变化的敏感性相对较高.分析其原因, 一方面生态系统温度敏感性与气温有关(Kirschbaum, 1995; Xu & Qi, 2001; Janssens & Pilegaard, 2003; Davidson & Janssens., 2006; 张雷明等, 2006; Jassal et al., 2008), 气温对Q10产生影响主要通过影响光合与呼吸作用酶的活性实现(Davidson & Janssens, 2006).晴天气温T ((23.56 ± 2.62) ℃)显著大于阴天((21.26 ± 2.34) ℃), 阴天由于酶促反应需要一定的活化酶, 光合与呼吸作用酶的活性一般受到限制; 而晴天随着辐射增强, 气温升高, 越来越多的分子达到或超过了自身的活化能, 反应加快(Xiang & Freeman, 2009; Karhu et al., 2010).另一方面, 晴天相对阴天较高的PAR使得晴天光合作用更强, 产生输送的有机质增多(Curiel Yuste et al., 2007; Jassal et al., 2008), 自养呼吸增强, 同时地上植被以及土壤异养微生物呼吸对气温的响应能力增强(Edwards et al., 2004; Heinemeyer et al., 2006; Moyano et al., 2008), 而阴天由于底物基质供应不足使得自养、异养呼吸受到抑制(Gershenson et al., 2009). ...
Large seasonal changes in Q10 of soil respiration in a beech forest.
1
2003
... Q10值被广泛应用于评价生态系统呼吸对温度变化的敏感程度(杨庆朋等, 2011; 杨利琼等, 2013; Han et al., 2014a, 2014b), Q10值越大, 说明生态系统呼吸对温度变化的响应越敏感.黄河三角洲芦苇湿地生态系统阴天和晴天Q10值均在已有的湿地Q10范围(1.0-7.7)内(Chapman & Thurlow, 1996; Silvola et al., 1996; Lafleur et al., 2001; Bonneville et al., 2008; Zhou et al., 2009; 周丽艳等, 2010), 其中晴天白天Q10为5.5, 远高于阴天白天的Q10 (1.9), 说明晴天白天生态系统呼吸对温度变化的敏感性相对较高.分析其原因, 一方面生态系统温度敏感性与气温有关(Kirschbaum, 1995; Xu & Qi, 2001; Janssens & Pilegaard, 2003; Davidson & Janssens., 2006; 张雷明等, 2006; Jassal et al., 2008), 气温对Q10产生影响主要通过影响光合与呼吸作用酶的活性实现(Davidson & Janssens, 2006).晴天气温T ((23.56 ± 2.62) ℃)显著大于阴天((21.26 ± 2.34) ℃), 阴天由于酶促反应需要一定的活化酶, 光合与呼吸作用酶的活性一般受到限制; 而晴天随着辐射增强, 气温升高, 越来越多的分子达到或超过了自身的活化能, 反应加快(Xiang & Freeman, 2009; Karhu et al., 2010).另一方面, 晴天相对阴天较高的PAR使得晴天光合作用更强, 产生输送的有机质增多(Curiel Yuste et al., 2007; Jassal et al., 2008), 自养呼吸增强, 同时地上植被以及土壤异养微生物呼吸对气温的响应能力增强(Edwards et al., 2004; Heinemeyer et al., 2006; Moyano et al., 2008), 而阴天由于底物基质供应不足使得自养、异养呼吸受到抑制(Gershenson et al., 2009). ...
Effect of soil water stress on soil respiration and its temperature sensitivity in an 18-year-old temperate Douglas- fir stand.
2
2008
... Q10值被广泛应用于评价生态系统呼吸对温度变化的敏感程度(杨庆朋等, 2011; 杨利琼等, 2013; Han et al., 2014a, 2014b), Q10值越大, 说明生态系统呼吸对温度变化的响应越敏感.黄河三角洲芦苇湿地生态系统阴天和晴天Q10值均在已有的湿地Q10范围(1.0-7.7)内(Chapman & Thurlow, 1996; Silvola et al., 1996; Lafleur et al., 2001; Bonneville et al., 2008; Zhou et al., 2009; 周丽艳等, 2010), 其中晴天白天Q10为5.5, 远高于阴天白天的Q10 (1.9), 说明晴天白天生态系统呼吸对温度变化的敏感性相对较高.分析其原因, 一方面生态系统温度敏感性与气温有关(Kirschbaum, 1995; Xu & Qi, 2001; Janssens & Pilegaard, 2003; Davidson & Janssens., 2006; 张雷明等, 2006; Jassal et al., 2008), 气温对Q10产生影响主要通过影响光合与呼吸作用酶的活性实现(Davidson & Janssens, 2006).晴天气温T ((23.56 ± 2.62) ℃)显著大于阴天((21.26 ± 2.34) ℃), 阴天由于酶促反应需要一定的活化酶, 光合与呼吸作用酶的活性一般受到限制; 而晴天随着辐射增强, 气温升高, 越来越多的分子达到或超过了自身的活化能, 反应加快(Xiang & Freeman, 2009; Karhu et al., 2010).另一方面, 晴天相对阴天较高的PAR使得晴天光合作用更强, 产生输送的有机质增多(Curiel Yuste et al., 2007; Jassal et al., 2008), 自养呼吸增强, 同时地上植被以及土壤异养微生物呼吸对气温的响应能力增强(Edwards et al., 2004; Heinemeyer et al., 2006; Moyano et al., 2008), 而阴天由于底物基质供应不足使得自养、异养呼吸受到抑制(Gershenson et al., 2009). ...
... ; Jassal et al., 2008), 自养呼吸增强, 同时地上植被以及土壤异养微生物呼吸对气温的响应能力增强(Edwards et al., 2004; Heinemeyer et al., 2006; Moyano et al., 2008), 而阴天由于底物基质供应不足使得自养、异养呼吸受到抑制(Gershenson et al., 2009). ...
华北地区沙尘气溶胶对云辐射强迫的影响
1
2010
... 近年来, 大气污染、气溶胶粒子增加等环境问题日益严重, 云层和大气气溶胶含量的变化给地面接收的直接辐射和散射辐射带来很大影响(Niyogi et al., 2004; Bar-Or et al., 2010; 马金玉等, 2011).我国华北地区大气环境由于受到西北地区沙尘的影响, 大量滞留在大气中的沙尘成为对流层气溶胶的主要成分, 影响地-气系统辐射能收支, 从而影响区域的气候及生态环境(Min, 2005; 贾漩等, 2010).研究表明: 近50年来, 地面太阳总辐射从减少到增加(齐月等, 2014).而云作为一种天气现象会对局地的微气候环境产生综合效应, 当天空有云层出现时, 地面接收的太阳辐射强度及散射辐射与直射辐射的比例会发生变化, 引起生态系统光能与热能的变化, 植被的光合与呼吸作用发生变化, 最终生态系统与大气间的净CO2交换也会受到影响(Letts et al., 2005; Urban et al., 2007). ...
华北地区沙尘气溶胶对云辐射强迫的影响
1
2010
... 近年来, 大气污染、气溶胶粒子增加等环境问题日益严重, 云层和大气气溶胶含量的变化给地面接收的直接辐射和散射辐射带来很大影响(Niyogi et al., 2004; Bar-Or et al., 2010; 马金玉等, 2011).我国华北地区大气环境由于受到西北地区沙尘的影响, 大量滞留在大气中的沙尘成为对流层气溶胶的主要成分, 影响地-气系统辐射能收支, 从而影响区域的气候及生态环境(Min, 2005; 贾漩等, 2010).研究表明: 近50年来, 地面太阳总辐射从减少到增加(齐月等, 2014).而云作为一种天气现象会对局地的微气候环境产生综合效应, 当天空有云层出现时, 地面接收的太阳辐射强度及散射辐射与直射辐射的比例会发生变化, 引起生态系统光能与热能的变化, 植被的光合与呼吸作用发生变化, 最终生态系统与大气间的净CO2交换也会受到影响(Letts et al., 2005; Urban et al., 2007). ...
Systemic regulation of leaf anatomical structure, photosynthetic performance, and high-light tolerance in Sorghum.
2
2011
... 黄河口近海与海岸湿地是暖温带增长速度最快和最具代表性的新生近海与海岸湿地, 具有海陆过渡性、原生性和脆弱性的特点(布仁仓等, 1999; 崔树强, 2002).受水沙量变化、陆地和河流淡水径流、咸水海流多重影响以及人类活动的干扰, 黄河三角洲湿地生态系统CO2交换存在着极大的复杂性和不确定性(Sasaki et al., 2009;Han et al., 2014a, 2014b; 邢庆会等, 2014).研究表明黄河三角洲芦苇湿地生长季NEE占全年的80%以上, 且碳吸收集中在植物生长旺盛期(7-9月)(杨利琼等, 2013; Han et al., 2014a, 2014b).而这一时期正值黄河三角洲雨季, 其云雨状况影响太阳辐射, 进而影响湿地生态系统的碳吸收功能.在以往的研究中, 温度、水分、太阳辐射等环境因子, 生物量、叶面积等生物因子以及地下水位、地表水深等水文要素对湿地生态系统NEE的影响机制受到研究者的极大关注(牟长城等, 2009; Zhao et al., 2010; Han et al., 2013; 邢庆会等, 2014).目前, 有关阴天和晴天条件对生态系统碳交换影响的研究主要集中在森林生态系统(Liu et al., 2006; Zhang et al., 2010; 周丽艳等, 2010; Jiang et al., 2011; Zhao et al., 2011; 刘佳等, 2014)和草原生态系统(Gu et al., 2003; 范玉枝等, 2009).阴天和晴天状况引起的太阳辐射变化对湿地生态系统NEE和碳汇功能究竟产生怎样的影响目前尚不清楚. ...
... 在一定光照强度范围内, 光合作用随PAR的增强而增加(Glenn et al., 2006; Syed et al., 2006; Hao et al., 2011; 同小娟等, 2011).光能过剩会对光合作用产生不良影响, 出现光抑制, 致使叶绿体光合效率下降(Jiang et al., 2011; Tholen et al., 2012).晴天PAR显著大于阴天(图2B), 因此, 晴天条件下植被叶片光合作用可能大于阴天, 表现为晴天Amax显著大于阴天(表1).Zhou等(2009)研究认为Amax与植被的环境条件有关, 其中晴天Amax较阴天条件下提高了30%, 说明在植被生长旺季, 天空有云层覆盖时芦苇的光合能力比晴朗天气无云时有所降低, Law等(2002)总结全球森林、草地、农田以及苔原生态系统碳交换相关研究也得出类似结论.阴天条件下, 伴随天空云量的增多, PAR显著降低(图2B), 尽管阴天和晴天条件下, NEE均随PAR的增大而增加(图3), 但由于阴天PAR较低, 且晴天未出现光抑制现象, 因此其NEE显著低于晴天(图2A).Alton (2008)在分析北美洲和欧洲6种森林类型2-3年的碳通量观测数据后发现, 这些森林阴天的日固碳量比晴天少60%-80%.相反, 在研究太阳辐射对黄河小浪底人工混交林生态系统碳交换的影响中发现, 多云天气下净碳交换会增加(刘佳等, 2014).造成研究结论不一致的原因是多方面的, 不同的阴天和晴天划分指标(Graham et al., 2003; Dengel & Grace, 2010)和拟合模型(Zhang et al., 2011)来估计天气对碳通量的影响, 散射光和直射光下的碳交换没有一直处在相同的天气辐射条件下进行对比.此外, 研究地域的差异和不同植被类型的特性也在很大程度上影响了碳交换过程对天气的响应过程. ...
北亚热带次生栎林碳通量及其影响因子研究
1
2011
... 本研究以黄河三角洲芦苇湿地为研究对象, 在自然条件下选择相邻的阴天和晴天, 在生物要素(生物量、叶面积指数)、土壤水分以及养分特征保持不变的前提下, 分析阴天和晴天引起的辐射变化对生态系统NEE的影响机制.目前, 有关太阳辐射对生态系统NEE的影响机制大都以晴空指数为指标(Gu et al., 2003; Liu et al., 2006; 范玉枝等, 2009; Zhang et al., 2010; 周丽艳等, 2010; 蒋琰等, 2011; Zhao et al., 2011; 刘佳等, 2014).这种研究方法的最大不足就是研究阶段中, 太阳辐射、生物要素(生物量、叶面积指数)、土壤湿度等环境因子都在发生变化, 这对阐明太阳辐射这一单因素对NEE的控制机制研究带来一定困难.而本研究巧妙地在自然条件下选择相邻的阴天和晴天, 在日尺度上实现了对单一变量PAR的控制.利用这种研究方法, Han等(2014a)通过对比分析阴天和晴天植物冠层光合作用的差异性, 定量分析了光合作用在日尺度上对土壤呼吸的调节作用.此外, 我们已有研究发现PAR改变所引起的植物光合作用变化对生态系统呼吸会产生一定的影响(Tang et al., 2005; Baldocchi et al., 2006; Moyano et al., 2008; Wingate et al., 2010; Han et al., 2012).但是, 以往的研究多通过夜间生态系统呼吸与气温函数关系外延的方法来估算白天生态系统呼吸(Doughty et al., 2010; Zhang et al., 2011), 忽略了PAR对白天生态系统呼吸的影响, 因而具有一定的局限性.考虑到白天PAR的变化, 本研究采用Michaelis-Menten模型对白天生态系统呼吸进行推算, 使得生态系统呼吸的温度响应数据更具有说服力. ...
北亚热带次生栎林碳通量及其影响因子研究
1
2011
... 本研究以黄河三角洲芦苇湿地为研究对象, 在自然条件下选择相邻的阴天和晴天, 在生物要素(生物量、叶面积指数)、土壤水分以及养分特征保持不变的前提下, 分析阴天和晴天引起的辐射变化对生态系统NEE的影响机制.目前, 有关太阳辐射对生态系统NEE的影响机制大都以晴空指数为指标(Gu et al., 2003; Liu et al., 2006; 范玉枝等, 2009; Zhang et al., 2010; 周丽艳等, 2010; 蒋琰等, 2011; Zhao et al., 2011; 刘佳等, 2014).这种研究方法的最大不足就是研究阶段中, 太阳辐射、生物要素(生物量、叶面积指数)、土壤湿度等环境因子都在发生变化, 这对阐明太阳辐射这一单因素对NEE的控制机制研究带来一定困难.而本研究巧妙地在自然条件下选择相邻的阴天和晴天, 在日尺度上实现了对单一变量PAR的控制.利用这种研究方法, Han等(2014a)通过对比分析阴天和晴天植物冠层光合作用的差异性, 定量分析了光合作用在日尺度上对土壤呼吸的调节作用.此外, 我们已有研究发现PAR改变所引起的植物光合作用变化对生态系统呼吸会产生一定的影响(Tang et al., 2005; Baldocchi et al., 2006; Moyano et al., 2008; Wingate et al., 2010; Han et al., 2012).但是, 以往的研究多通过夜间生态系统呼吸与气温函数关系外延的方法来估算白天生态系统呼吸(Doughty et al., 2010; Zhang et al., 2011), 忽略了PAR对白天生态系统呼吸的影响, 因而具有一定的局限性.考虑到白天PAR的变化, 本研究采用Michaelis-Menten模型对白天生态系统呼吸进行推算, 使得生态系统呼吸的温度响应数据更具有说服力. ...
Temperature sensitivity of organic matter decomposition in two boreal forest soil profiles.
1
2010
... Q10值被广泛应用于评价生态系统呼吸对温度变化的敏感程度(杨庆朋等, 2011; 杨利琼等, 2013; Han et al., 2014a, 2014b), Q10值越大, 说明生态系统呼吸对温度变化的响应越敏感.黄河三角洲芦苇湿地生态系统阴天和晴天Q10值均在已有的湿地Q10范围(1.0-7.7)内(Chapman & Thurlow, 1996; Silvola et al., 1996; Lafleur et al., 2001; Bonneville et al., 2008; Zhou et al., 2009; 周丽艳等, 2010), 其中晴天白天Q10为5.5, 远高于阴天白天的Q10 (1.9), 说明晴天白天生态系统呼吸对温度变化的敏感性相对较高.分析其原因, 一方面生态系统温度敏感性与气温有关(Kirschbaum, 1995; Xu & Qi, 2001; Janssens & Pilegaard, 2003; Davidson & Janssens., 2006; 张雷明等, 2006; Jassal et al., 2008), 气温对Q10产生影响主要通过影响光合与呼吸作用酶的活性实现(Davidson & Janssens, 2006).晴天气温T ((23.56 ± 2.62) ℃)显著大于阴天((21.26 ± 2.34) ℃), 阴天由于酶促反应需要一定的活化酶, 光合与呼吸作用酶的活性一般受到限制; 而晴天随着辐射增强, 气温升高, 越来越多的分子达到或超过了自身的活化能, 反应加快(Xiang & Freeman, 2009; Karhu et al., 2010).另一方面, 晴天相对阴天较高的PAR使得晴天光合作用更强, 产生输送的有机质增多(Curiel Yuste et al., 2007; Jassal et al., 2008), 自养呼吸增强, 同时地上植被以及土壤异养微生物呼吸对气温的响应能力增强(Edwards et al., 2004; Heinemeyer et al., 2006; Moyano et al., 2008), 而阴天由于底物基质供应不足使得自养、异养呼吸受到抑制(Gershenson et al., 2009). ...
The temperature dependence of soil organic-matter decomposition, and the effect of global warming on soil organic C storage.
1
1995
... Q10值被广泛应用于评价生态系统呼吸对温度变化的敏感程度(杨庆朋等, 2011; 杨利琼等, 2013; Han et al., 2014a, 2014b), Q10值越大, 说明生态系统呼吸对温度变化的响应越敏感.黄河三角洲芦苇湿地生态系统阴天和晴天Q10值均在已有的湿地Q10范围(1.0-7.7)内(Chapman & Thurlow, 1996; Silvola et al., 1996; Lafleur et al., 2001; Bonneville et al., 2008; Zhou et al., 2009; 周丽艳等, 2010), 其中晴天白天Q10为5.5, 远高于阴天白天的Q10 (1.9), 说明晴天白天生态系统呼吸对温度变化的敏感性相对较高.分析其原因, 一方面生态系统温度敏感性与气温有关(Kirschbaum, 1995; Xu & Qi, 2001; Janssens & Pilegaard, 2003; Davidson & Janssens., 2006; 张雷明等, 2006; Jassal et al., 2008), 气温对Q10产生影响主要通过影响光合与呼吸作用酶的活性实现(Davidson & Janssens, 2006).晴天气温T ((23.56 ± 2.62) ℃)显著大于阴天((21.26 ± 2.34) ℃), 阴天由于酶促反应需要一定的活化酶, 光合与呼吸作用酶的活性一般受到限制; 而晴天随着辐射增强, 气温升高, 越来越多的分子达到或超过了自身的活化能, 反应加快(Xiang & Freeman, 2009; Karhu et al., 2010).另一方面, 晴天相对阴天较高的PAR使得晴天光合作用更强, 产生输送的有机质增多(Curiel Yuste et al., 2007; Jassal et al., 2008), 自养呼吸增强, 同时地上植被以及土壤异养微生物呼吸对气温的响应能力增强(Edwards et al., 2004; Heinemeyer et al., 2006; Moyano et al., 2008), 而阴天由于底物基质供应不足使得自养、异养呼吸受到抑制(Gershenson et al., 2009). ...
Annual cycle of CO2 exchange at a bog peatland.
1
2001
... Q10值被广泛应用于评价生态系统呼吸对温度变化的敏感程度(杨庆朋等, 2011; 杨利琼等, 2013; Han et al., 2014a, 2014b), Q10值越大, 说明生态系统呼吸对温度变化的响应越敏感.黄河三角洲芦苇湿地生态系统阴天和晴天Q10值均在已有的湿地Q10范围(1.0-7.7)内(Chapman & Thurlow, 1996; Silvola et al., 1996; Lafleur et al., 2001; Bonneville et al., 2008; Zhou et al., 2009; 周丽艳等, 2010), 其中晴天白天Q10为5.5, 远高于阴天白天的Q10 (1.9), 说明晴天白天生态系统呼吸对温度变化的敏感性相对较高.分析其原因, 一方面生态系统温度敏感性与气温有关(Kirschbaum, 1995; Xu & Qi, 2001; Janssens & Pilegaard, 2003; Davidson & Janssens., 2006; 张雷明等, 2006; Jassal et al., 2008), 气温对Q10产生影响主要通过影响光合与呼吸作用酶的活性实现(Davidson & Janssens, 2006).晴天气温T ((23.56 ± 2.62) ℃)显著大于阴天((21.26 ± 2.34) ℃), 阴天由于酶促反应需要一定的活化酶, 光合与呼吸作用酶的活性一般受到限制; 而晴天随着辐射增强, 气温升高, 越来越多的分子达到或超过了自身的活化能, 反应加快(Xiang & Freeman, 2009; Karhu et al., 2010).另一方面, 晴天相对阴天较高的PAR使得晴天光合作用更强, 产生输送的有机质增多(Curiel Yuste et al., 2007; Jassal et al., 2008), 自养呼吸增强, 同时地上植被以及土壤异养微生物呼吸对气温的响应能力增强(Edwards et al., 2004; Heinemeyer et al., 2006; Moyano et al., 2008), 而阴天由于底物基质供应不足使得自养、异养呼吸受到抑制(Gershenson et al., 2009). ...
Environmental controls over carbon dioxide and water vapor exchange of terrestrial vegetation.
1
2002
... 阴天和晴天生态系统Reco,daytime与T均呈显著的正相关关系, 两者呈指数关系, 与以往很多研究的结果一致(石培礼等, 2006; Zhou et al., 2009; Zhao et al., 2010; Han et al., 2013, 2014a; 邢庆会等, 2014).T是影响生态系统呼吸的主要因素(Law et al., 2002; Reichstein et al., 2005; Urban et al., 2007), 本文研究发现: 黄河三角洲芦苇湿地晴天气温相对阴天提高了10%, 其阴天和晴天的气温差高于美国阿尔卑斯山脉南部森林的阴天和晴天的气温差(Berry & Smith, 2012), 黄河三角洲芦苇湿地晴天对应的生态系统呼吸相对阴天提高了20% (p < 0.05), 说明阴天条件下, 植被地上部分尤其是叶面温度较晴朗天气强太阳辐射条件下的低, 叶片以及植被茎干呼吸受到一定的抑制(Urban et al., 2007). ...
On the relationship between cloudiness and net ecosystem carbon dioxide exchange in a peatland ecosystem.
2
2005
... 近年来, 大气污染、气溶胶粒子增加等环境问题日益严重, 云层和大气气溶胶含量的变化给地面接收的直接辐射和散射辐射带来很大影响(Niyogi et al., 2004; Bar-Or et al., 2010; 马金玉等, 2011).我国华北地区大气环境由于受到西北地区沙尘的影响, 大量滞留在大气中的沙尘成为对流层气溶胶的主要成分, 影响地-气系统辐射能收支, 从而影响区域的气候及生态环境(Min, 2005; 贾漩等, 2010).研究表明: 近50年来, 地面太阳总辐射从减少到增加(齐月等, 2014).而云作为一种天气现象会对局地的微气候环境产生综合效应, 当天空有云层出现时, 地面接收的太阳辐射强度及散射辐射与直射辐射的比例会发生变化, 引起生态系统光能与热能的变化, 植被的光合与呼吸作用发生变化, 最终生态系统与大气间的净CO2交换也会受到影响(Letts et al., 2005; Urban et al., 2007). ...
... 已有研究表明, 阴天和晴天所引起的辐射差异影响着陆地生态系统的交换过程和碳吸收能力(Roderick et al., 2001; Still et al., 2009).但由于研究方法、地域环境条件以及不同植被类型的差异, 研究结论存在较大差异(Gu et al., 1999, 2002; Alton et al., 2007; Urban et al., 2007).例如, 中国渭北刺槐(Robinia pseudoacacia)生长初期和旺盛期晴天条件下森林日均固碳量比阴天高8%-16% (郑元等, 2011).中国西北玉米(Zea mays)农田生态系统阴天条件下净生态系统CO2交换(NEE)的增长速率却高于晴天, 且生态系统总初级生产力(GPP)最大值出现于阴天(Zhang et al., 2011).对中国西北草地(Bai et al., 2012)和不同温带森林类型(Zhang et al., 2010)的研究也得出类似结论.另外, 也有研究发现加拿大常绿灌木(Letts et al., 2005)和西藏高寒草甸(范玉枝等, 2009)生态系统CO2交换不受阴天和晴天天气条件影响. ...
太阳辐射对黄河小浪底人工混交林净生态系统碳交换的影响
3
2014
... 黄河口近海与海岸湿地是暖温带增长速度最快和最具代表性的新生近海与海岸湿地, 具有海陆过渡性、原生性和脆弱性的特点(布仁仓等, 1999; 崔树强, 2002).受水沙量变化、陆地和河流淡水径流、咸水海流多重影响以及人类活动的干扰, 黄河三角洲湿地生态系统CO2交换存在着极大的复杂性和不确定性(Sasaki et al., 2009;Han et al., 2014a, 2014b; 邢庆会等, 2014).研究表明黄河三角洲芦苇湿地生长季NEE占全年的80%以上, 且碳吸收集中在植物生长旺盛期(7-9月)(杨利琼等, 2013; Han et al., 2014a, 2014b).而这一时期正值黄河三角洲雨季, 其云雨状况影响太阳辐射, 进而影响湿地生态系统的碳吸收功能.在以往的研究中, 温度、水分、太阳辐射等环境因子, 生物量、叶面积等生物因子以及地下水位、地表水深等水文要素对湿地生态系统NEE的影响机制受到研究者的极大关注(牟长城等, 2009; Zhao et al., 2010; Han et al., 2013; 邢庆会等, 2014).目前, 有关阴天和晴天条件对生态系统碳交换影响的研究主要集中在森林生态系统(Liu et al., 2006; Zhang et al., 2010; 周丽艳等, 2010; Jiang et al., 2011; Zhao et al., 2011; 刘佳等, 2014)和草原生态系统(Gu et al., 2003; 范玉枝等, 2009).阴天和晴天状况引起的太阳辐射变化对湿地生态系统NEE和碳汇功能究竟产生怎样的影响目前尚不清楚. ...
... 在一定光照强度范围内, 光合作用随PAR的增强而增加(Glenn et al., 2006; Syed et al., 2006; Hao et al., 2011; 同小娟等, 2011).光能过剩会对光合作用产生不良影响, 出现光抑制, 致使叶绿体光合效率下降(Jiang et al., 2011; Tholen et al., 2012).晴天PAR显著大于阴天(图2B), 因此, 晴天条件下植被叶片光合作用可能大于阴天, 表现为晴天Amax显著大于阴天(表1).Zhou等(2009)研究认为Amax与植被的环境条件有关, 其中晴天Amax较阴天条件下提高了30%, 说明在植被生长旺季, 天空有云层覆盖时芦苇的光合能力比晴朗天气无云时有所降低, Law等(2002)总结全球森林、草地、农田以及苔原生态系统碳交换相关研究也得出类似结论.阴天条件下, 伴随天空云量的增多, PAR显著降低(图2B), 尽管阴天和晴天条件下, NEE均随PAR的增大而增加(图3), 但由于阴天PAR较低, 且晴天未出现光抑制现象, 因此其NEE显著低于晴天(图2A).Alton (2008)在分析北美洲和欧洲6种森林类型2-3年的碳通量观测数据后发现, 这些森林阴天的日固碳量比晴天少60%-80%.相反, 在研究太阳辐射对黄河小浪底人工混交林生态系统碳交换的影响中发现, 多云天气下净碳交换会增加(刘佳等, 2014).造成研究结论不一致的原因是多方面的, 不同的阴天和晴天划分指标(Graham et al., 2003; Dengel & Grace, 2010)和拟合模型(Zhang et al., 2011)来估计天气对碳通量的影响, 散射光和直射光下的碳交换没有一直处在相同的天气辐射条件下进行对比.此外, 研究地域的差异和不同植被类型的特性也在很大程度上影响了碳交换过程对天气的响应过程. ...
... 本研究以黄河三角洲芦苇湿地为研究对象, 在自然条件下选择相邻的阴天和晴天, 在生物要素(生物量、叶面积指数)、土壤水分以及养分特征保持不变的前提下, 分析阴天和晴天引起的辐射变化对生态系统NEE的影响机制.目前, 有关太阳辐射对生态系统NEE的影响机制大都以晴空指数为指标(Gu et al., 2003; Liu et al., 2006; 范玉枝等, 2009; Zhang et al., 2010; 周丽艳等, 2010; 蒋琰等, 2011; Zhao et al., 2011; 刘佳等, 2014).这种研究方法的最大不足就是研究阶段中, 太阳辐射、生物要素(生物量、叶面积指数)、土壤湿度等环境因子都在发生变化, 这对阐明太阳辐射这一单因素对NEE的控制机制研究带来一定困难.而本研究巧妙地在自然条件下选择相邻的阴天和晴天, 在日尺度上实现了对单一变量PAR的控制.利用这种研究方法, Han等(2014a)通过对比分析阴天和晴天植物冠层光合作用的差异性, 定量分析了光合作用在日尺度上对土壤呼吸的调节作用.此外, 我们已有研究发现PAR改变所引起的植物光合作用变化对生态系统呼吸会产生一定的影响(Tang et al., 2005; Baldocchi et al., 2006; Moyano et al., 2008; Wingate et al., 2010; Han et al., 2012).但是, 以往的研究多通过夜间生态系统呼吸与气温函数关系外延的方法来估算白天生态系统呼吸(Doughty et al., 2010; Zhang et al., 2011), 忽略了PAR对白天生态系统呼吸的影响, 因而具有一定的局限性.考虑到白天PAR的变化, 本研究采用Michaelis-Menten模型对白天生态系统呼吸进行推算, 使得生态系统呼吸的温度响应数据更具有说服力. ...
太阳辐射对黄河小浪底人工混交林净生态系统碳交换的影响
3
2014
... 黄河口近海与海岸湿地是暖温带增长速度最快和最具代表性的新生近海与海岸湿地, 具有海陆过渡性、原生性和脆弱性的特点(布仁仓等, 1999; 崔树强, 2002).受水沙量变化、陆地和河流淡水径流、咸水海流多重影响以及人类活动的干扰, 黄河三角洲湿地生态系统CO2交换存在着极大的复杂性和不确定性(Sasaki et al., 2009;Han et al., 2014a, 2014b; 邢庆会等, 2014).研究表明黄河三角洲芦苇湿地生长季NEE占全年的80%以上, 且碳吸收集中在植物生长旺盛期(7-9月)(杨利琼等, 2013; Han et al., 2014a, 2014b).而这一时期正值黄河三角洲雨季, 其云雨状况影响太阳辐射, 进而影响湿地生态系统的碳吸收功能.在以往的研究中, 温度、水分、太阳辐射等环境因子, 生物量、叶面积等生物因子以及地下水位、地表水深等水文要素对湿地生态系统NEE的影响机制受到研究者的极大关注(牟长城等, 2009; Zhao et al., 2010; Han et al., 2013; 邢庆会等, 2014).目前, 有关阴天和晴天条件对生态系统碳交换影响的研究主要集中在森林生态系统(Liu et al., 2006; Zhang et al., 2010; 周丽艳等, 2010; Jiang et al., 2011; Zhao et al., 2011; 刘佳等, 2014)和草原生态系统(Gu et al., 2003; 范玉枝等, 2009).阴天和晴天状况引起的太阳辐射变化对湿地生态系统NEE和碳汇功能究竟产生怎样的影响目前尚不清楚. ...
... 在一定光照强度范围内, 光合作用随PAR的增强而增加(Glenn et al., 2006; Syed et al., 2006; Hao et al., 2011; 同小娟等, 2011).光能过剩会对光合作用产生不良影响, 出现光抑制, 致使叶绿体光合效率下降(Jiang et al., 2011; Tholen et al., 2012).晴天PAR显著大于阴天(图2B), 因此, 晴天条件下植被叶片光合作用可能大于阴天, 表现为晴天Amax显著大于阴天(表1).Zhou等(2009)研究认为Amax与植被的环境条件有关, 其中晴天Amax较阴天条件下提高了30%, 说明在植被生长旺季, 天空有云层覆盖时芦苇的光合能力比晴朗天气无云时有所降低, Law等(2002)总结全球森林、草地、农田以及苔原生态系统碳交换相关研究也得出类似结论.阴天条件下, 伴随天空云量的增多, PAR显著降低(图2B), 尽管阴天和晴天条件下, NEE均随PAR的增大而增加(图3), 但由于阴天PAR较低, 且晴天未出现光抑制现象, 因此其NEE显著低于晴天(图2A).Alton (2008)在分析北美洲和欧洲6种森林类型2-3年的碳通量观测数据后发现, 这些森林阴天的日固碳量比晴天少60%-80%.相反, 在研究太阳辐射对黄河小浪底人工混交林生态系统碳交换的影响中发现, 多云天气下净碳交换会增加(刘佳等, 2014).造成研究结论不一致的原因是多方面的, 不同的阴天和晴天划分指标(Graham et al., 2003; Dengel & Grace, 2010)和拟合模型(Zhang et al., 2011)来估计天气对碳通量的影响, 散射光和直射光下的碳交换没有一直处在相同的天气辐射条件下进行对比.此外, 研究地域的差异和不同植被类型的特性也在很大程度上影响了碳交换过程对天气的响应过程. ...
... 本研究以黄河三角洲芦苇湿地为研究对象, 在自然条件下选择相邻的阴天和晴天, 在生物要素(生物量、叶面积指数)、土壤水分以及养分特征保持不变的前提下, 分析阴天和晴天引起的辐射变化对生态系统NEE的影响机制.目前, 有关太阳辐射对生态系统NEE的影响机制大都以晴空指数为指标(Gu et al., 2003; Liu et al., 2006; 范玉枝等, 2009; Zhang et al., 2010; 周丽艳等, 2010; 蒋琰等, 2011; Zhao et al., 2011; 刘佳等, 2014).这种研究方法的最大不足就是研究阶段中, 太阳辐射、生物要素(生物量、叶面积指数)、土壤湿度等环境因子都在发生变化, 这对阐明太阳辐射这一单因素对NEE的控制机制研究带来一定困难.而本研究巧妙地在自然条件下选择相邻的阴天和晴天, 在日尺度上实现了对单一变量PAR的控制.利用这种研究方法, Han等(2014a)通过对比分析阴天和晴天植物冠层光合作用的差异性, 定量分析了光合作用在日尺度上对土壤呼吸的调节作用.此外, 我们已有研究发现PAR改变所引起的植物光合作用变化对生态系统呼吸会产生一定的影响(Tang et al., 2005; Baldocchi et al., 2006; Moyano et al., 2008; Wingate et al., 2010; Han et al., 2012).但是, 以往的研究多通过夜间生态系统呼吸与气温函数关系外延的方法来估算白天生态系统呼吸(Doughty et al., 2010; Zhang et al., 2011), 忽略了PAR对白天生态系统呼吸的影响, 因而具有一定的局限性.考虑到白天PAR的变化, 本研究采用Michaelis-Menten模型对白天生态系统呼吸进行推算, 使得生态系统呼吸的温度响应数据更具有说服力. ...
Seasonal dynamics of CO2 fluxes from subtropical plantation coniferous ecosystem.
3
2006
... 黄河口近海与海岸湿地是暖温带增长速度最快和最具代表性的新生近海与海岸湿地, 具有海陆过渡性、原生性和脆弱性的特点(布仁仓等, 1999; 崔树强, 2002).受水沙量变化、陆地和河流淡水径流、咸水海流多重影响以及人类活动的干扰, 黄河三角洲湿地生态系统CO2交换存在着极大的复杂性和不确定性(Sasaki et al., 2009;Han et al., 2014a, 2014b; 邢庆会等, 2014).研究表明黄河三角洲芦苇湿地生长季NEE占全年的80%以上, 且碳吸收集中在植物生长旺盛期(7-9月)(杨利琼等, 2013; Han et al., 2014a, 2014b).而这一时期正值黄河三角洲雨季, 其云雨状况影响太阳辐射, 进而影响湿地生态系统的碳吸收功能.在以往的研究中, 温度、水分、太阳辐射等环境因子, 生物量、叶面积等生物因子以及地下水位、地表水深等水文要素对湿地生态系统NEE的影响机制受到研究者的极大关注(牟长城等, 2009; Zhao et al., 2010; Han et al., 2013; 邢庆会等, 2014).目前, 有关阴天和晴天条件对生态系统碳交换影响的研究主要集中在森林生态系统(Liu et al., 2006; Zhang et al., 2010; 周丽艳等, 2010; Jiang et al., 2011; Zhao et al., 2011; 刘佳等, 2014)和草原生态系统(Gu et al., 2003; 范玉枝等, 2009).阴天和晴天状况引起的太阳辐射变化对湿地生态系统NEE和碳汇功能究竟产生怎样的影响目前尚不清楚. ...
... 本文研究发现晴天和阴天芦苇湿地NEE对PAR的响应趋势一致, 均呈直角双曲线关系, 与先前的很多研究结果一致(Falge et al., 2001; Monson et al., 2002; Zhou et al., 2009; 张弥等, 2009; Han et al., 2014a, 2014b; 邢庆会等, 2014).PAR是控制阴天和晴天生态系统生长季昼间NEE变化的主要因素(Flanagan & Johnson, 2005; Liu et al., 2006; 薛红喜等, 2012), Michaelis-Menten方程拟合发现, PAR可以解释晴天单天NEE变化的57%-82%, 解释阴天单天NEE变化的53%-81%.云层变化会改变地表接收太阳辐射的强度, 影响PAR、T等环境因子, 进而影响阴天和晴天条件下NEE对光的响应特征(Baldocchi, 1997; Freedman et al., 2001). ...
... 本研究以黄河三角洲芦苇湿地为研究对象, 在自然条件下选择相邻的阴天和晴天, 在生物要素(生物量、叶面积指数)、土壤水分以及养分特征保持不变的前提下, 分析阴天和晴天引起的辐射变化对生态系统NEE的影响机制.目前, 有关太阳辐射对生态系统NEE的影响机制大都以晴空指数为指标(Gu et al., 2003; Liu et al., 2006; 范玉枝等, 2009; Zhang et al., 2010; 周丽艳等, 2010; 蒋琰等, 2011; Zhao et al., 2011; 刘佳等, 2014).这种研究方法的最大不足就是研究阶段中, 太阳辐射、生物要素(生物量、叶面积指数)、土壤湿度等环境因子都在发生变化, 这对阐明太阳辐射这一单因素对NEE的控制机制研究带来一定困难.而本研究巧妙地在自然条件下选择相邻的阴天和晴天, 在日尺度上实现了对单一变量PAR的控制.利用这种研究方法, Han等(2014a)通过对比分析阴天和晴天植物冠层光合作用的差异性, 定量分析了光合作用在日尺度上对土壤呼吸的调节作用.此外, 我们已有研究发现PAR改变所引起的植物光合作用变化对生态系统呼吸会产生一定的影响(Tang et al., 2005; Baldocchi et al., 2006; Moyano et al., 2008; Wingate et al., 2010; Han et al., 2012).但是, 以往的研究多通过夜间生态系统呼吸与气温函数关系外延的方法来估算白天生态系统呼吸(Doughty et al., 2010; Zhang et al., 2011), 忽略了PAR对白天生态系统呼吸的影响, 因而具有一定的局限性.考虑到白天PAR的变化, 本研究采用Michaelis-Menten模型对白天生态系统呼吸进行推算, 使得生态系统呼吸的温度响应数据更具有说服力. ...
On the temperature dependence of soil respiration.
... 近年来, 大气污染、气溶胶粒子增加等环境问题日益严重, 云层和大气气溶胶含量的变化给地面接收的直接辐射和散射辐射带来很大影响(Niyogi et al., 2004; Bar-Or et al., 2010; 马金玉等, 2011).我国华北地区大气环境由于受到西北地区沙尘的影响, 大量滞留在大气中的沙尘成为对流层气溶胶的主要成分, 影响地-气系统辐射能收支, 从而影响区域的气候及生态环境(Min, 2005; 贾漩等, 2010).研究表明: 近50年来, 地面太阳总辐射从减少到增加(齐月等, 2014).而云作为一种天气现象会对局地的微气候环境产生综合效应, 当天空有云层出现时, 地面接收的太阳辐射强度及散射辐射与直射辐射的比例会发生变化, 引起生态系统光能与热能的变化, 植被的光合与呼吸作用发生变化, 最终生态系统与大气间的净CO2交换也会受到影响(Letts et al., 2005; Urban et al., 2007). ...
中国近50年太阳直接辐射和散射辐射变化趋势特征
1
2011
... 近年来, 大气污染、气溶胶粒子增加等环境问题日益严重, 云层和大气气溶胶含量的变化给地面接收的直接辐射和散射辐射带来很大影响(Niyogi et al., 2004; Bar-Or et al., 2010; 马金玉等, 2011).我国华北地区大气环境由于受到西北地区沙尘的影响, 大量滞留在大气中的沙尘成为对流层气溶胶的主要成分, 影响地-气系统辐射能收支, 从而影响区域的气候及生态环境(Min, 2005; 贾漩等, 2010).研究表明: 近50年来, 地面太阳总辐射从减少到增加(齐月等, 2014).而云作为一种天气现象会对局地的微气候环境产生综合效应, 当天空有云层出现时, 地面接收的太阳辐射强度及散射辐射与直射辐射的比例会发生变化, 引起生态系统光能与热能的变化, 植被的光合与呼吸作用发生变化, 最终生态系统与大气间的净CO2交换也会受到影响(Letts et al., 2005; Urban et al., 2007). ...
Impacts of aerosols and clouds on forest- atmosphere carbon exchange.
1
2005
... 近年来, 大气污染、气溶胶粒子增加等环境问题日益严重, 云层和大气气溶胶含量的变化给地面接收的直接辐射和散射辐射带来很大影响(Niyogi et al., 2004; Bar-Or et al., 2010; 马金玉等, 2011).我国华北地区大气环境由于受到西北地区沙尘的影响, 大量滞留在大气中的沙尘成为对流层气溶胶的主要成分, 影响地-气系统辐射能收支, 从而影响区域的气候及生态环境(Min, 2005; 贾漩等, 2010).研究表明: 近50年来, 地面太阳总辐射从减少到增加(齐月等, 2014).而云作为一种天气现象会对局地的微气候环境产生综合效应, 当天空有云层出现时, 地面接收的太阳辐射强度及散射辐射与直射辐射的比例会发生变化, 引起生态系统光能与热能的变化, 植被的光合与呼吸作用发生变化, 最终生态系统与大气间的净CO2交换也会受到影响(Letts et al., 2005; Urban et al., 2007). ...
Carbon sequestration in a high-elevation, subalpine forest.
1
2002
... 本文研究发现晴天和阴天芦苇湿地NEE对PAR的响应趋势一致, 均呈直角双曲线关系, 与先前的很多研究结果一致(Falge et al., 2001; Monson et al., 2002; Zhou et al., 2009; 张弥等, 2009; Han et al., 2014a, 2014b; 邢庆会等, 2014).PAR是控制阴天和晴天生态系统生长季昼间NEE变化的主要因素(Flanagan & Johnson, 2005; Liu et al., 2006; 薛红喜等, 2012), Michaelis-Menten方程拟合发现, PAR可以解释晴天单天NEE变化的57%-82%, 解释阴天单天NEE变化的53%-81%.云层变化会改变地表接收太阳辐射的强度, 影响PAR、T等环境因子, 进而影响阴天和晴天条件下NEE对光的响应特征(Baldocchi, 1997; Freedman et al., 2001). ...
Soil respiration fluxes in relation to photosynthetic activity in broad-leaf and needle-leaf forest stands.
2
2008
... Q10值被广泛应用于评价生态系统呼吸对温度变化的敏感程度(杨庆朋等, 2011; 杨利琼等, 2013; Han et al., 2014a, 2014b), Q10值越大, 说明生态系统呼吸对温度变化的响应越敏感.黄河三角洲芦苇湿地生态系统阴天和晴天Q10值均在已有的湿地Q10范围(1.0-7.7)内(Chapman & Thurlow, 1996; Silvola et al., 1996; Lafleur et al., 2001; Bonneville et al., 2008; Zhou et al., 2009; 周丽艳等, 2010), 其中晴天白天Q10为5.5, 远高于阴天白天的Q10 (1.9), 说明晴天白天生态系统呼吸对温度变化的敏感性相对较高.分析其原因, 一方面生态系统温度敏感性与气温有关(Kirschbaum, 1995; Xu & Qi, 2001; Janssens & Pilegaard, 2003; Davidson & Janssens., 2006; 张雷明等, 2006; Jassal et al., 2008), 气温对Q10产生影响主要通过影响光合与呼吸作用酶的活性实现(Davidson & Janssens, 2006).晴天气温T ((23.56 ± 2.62) ℃)显著大于阴天((21.26 ± 2.34) ℃), 阴天由于酶促反应需要一定的活化酶, 光合与呼吸作用酶的活性一般受到限制; 而晴天随着辐射增强, 气温升高, 越来越多的分子达到或超过了自身的活化能, 反应加快(Xiang & Freeman, 2009; Karhu et al., 2010).另一方面, 晴天相对阴天较高的PAR使得晴天光合作用更强, 产生输送的有机质增多(Curiel Yuste et al., 2007; Jassal et al., 2008), 自养呼吸增强, 同时地上植被以及土壤异养微生物呼吸对气温的响应能力增强(Edwards et al., 2004; Heinemeyer et al., 2006; Moyano et al., 2008), 而阴天由于底物基质供应不足使得自养、异养呼吸受到抑制(Gershenson et al., 2009). ...
... 本研究以黄河三角洲芦苇湿地为研究对象, 在自然条件下选择相邻的阴天和晴天, 在生物要素(生物量、叶面积指数)、土壤水分以及养分特征保持不变的前提下, 分析阴天和晴天引起的辐射变化对生态系统NEE的影响机制.目前, 有关太阳辐射对生态系统NEE的影响机制大都以晴空指数为指标(Gu et al., 2003; Liu et al., 2006; 范玉枝等, 2009; Zhang et al., 2010; 周丽艳等, 2010; 蒋琰等, 2011; Zhao et al., 2011; 刘佳等, 2014).这种研究方法的最大不足就是研究阶段中, 太阳辐射、生物要素(生物量、叶面积指数)、土壤湿度等环境因子都在发生变化, 这对阐明太阳辐射这一单因素对NEE的控制机制研究带来一定困难.而本研究巧妙地在自然条件下选择相邻的阴天和晴天, 在日尺度上实现了对单一变量PAR的控制.利用这种研究方法, Han等(2014a)通过对比分析阴天和晴天植物冠层光合作用的差异性, 定量分析了光合作用在日尺度上对土壤呼吸的调节作用.此外, 我们已有研究发现PAR改变所引起的植物光合作用变化对生态系统呼吸会产生一定的影响(Tang et al., 2005; Baldocchi et al., 2006; Moyano et al., 2008; Wingate et al., 2010; Han et al., 2012).但是, 以往的研究多通过夜间生态系统呼吸与气温函数关系外延的方法来估算白天生态系统呼吸(Doughty et al., 2010; Zhang et al., 2011), 忽略了PAR对白天生态系统呼吸的影响, 因而具有一定的局限性.考虑到白天PAR的变化, 本研究采用Michaelis-Menten模型对白天生态系统呼吸进行推算, 使得生态系统呼吸的温度响应数据更具有说服力. ...
小兴安岭典型草丛沼泽湿地CO2、CH4和N2O的排放动态及其影响因素
1
2009
... 黄河口近海与海岸湿地是暖温带增长速度最快和最具代表性的新生近海与海岸湿地, 具有海陆过渡性、原生性和脆弱性的特点(布仁仓等, 1999; 崔树强, 2002).受水沙量变化、陆地和河流淡水径流、咸水海流多重影响以及人类活动的干扰, 黄河三角洲湿地生态系统CO2交换存在着极大的复杂性和不确定性(Sasaki et al., 2009;Han et al., 2014a, 2014b; 邢庆会等, 2014).研究表明黄河三角洲芦苇湿地生长季NEE占全年的80%以上, 且碳吸收集中在植物生长旺盛期(7-9月)(杨利琼等, 2013; Han et al., 2014a, 2014b).而这一时期正值黄河三角洲雨季, 其云雨状况影响太阳辐射, 进而影响湿地生态系统的碳吸收功能.在以往的研究中, 温度、水分、太阳辐射等环境因子, 生物量、叶面积等生物因子以及地下水位、地表水深等水文要素对湿地生态系统NEE的影响机制受到研究者的极大关注(牟长城等, 2009; Zhao et al., 2010; Han et al., 2013; 邢庆会等, 2014).目前, 有关阴天和晴天条件对生态系统碳交换影响的研究主要集中在森林生态系统(Liu et al., 2006; Zhang et al., 2010; 周丽艳等, 2010; Jiang et al., 2011; Zhao et al., 2011; 刘佳等, 2014)和草原生态系统(Gu et al., 2003; 范玉枝等, 2009).阴天和晴天状况引起的太阳辐射变化对湿地生态系统NEE和碳汇功能究竟产生怎样的影响目前尚不清楚. ...
小兴安岭典型草丛沼泽湿地CO2、CH4和N2O的排放动态及其影响因素
1
2009
... 黄河口近海与海岸湿地是暖温带增长速度最快和最具代表性的新生近海与海岸湿地, 具有海陆过渡性、原生性和脆弱性的特点(布仁仓等, 1999; 崔树强, 2002).受水沙量变化、陆地和河流淡水径流、咸水海流多重影响以及人类活动的干扰, 黄河三角洲湿地生态系统CO2交换存在着极大的复杂性和不确定性(Sasaki et al., 2009;Han et al., 2014a, 2014b; 邢庆会等, 2014).研究表明黄河三角洲芦苇湿地生长季NEE占全年的80%以上, 且碳吸收集中在植物生长旺盛期(7-9月)(杨利琼等, 2013; Han et al., 2014a, 2014b).而这一时期正值黄河三角洲雨季, 其云雨状况影响太阳辐射, 进而影响湿地生态系统的碳吸收功能.在以往的研究中, 温度、水分、太阳辐射等环境因子, 生物量、叶面积等生物因子以及地下水位、地表水深等水文要素对湿地生态系统NEE的影响机制受到研究者的极大关注(牟长城等, 2009; Zhao et al., 2010; Han et al., 2013; 邢庆会等, 2014).目前, 有关阴天和晴天条件对生态系统碳交换影响的研究主要集中在森林生态系统(Liu et al., 2006; Zhang et al., 2010; 周丽艳等, 2010; Jiang et al., 2011; Zhao et al., 2011; 刘佳等, 2014)和草原生态系统(Gu et al., 2003; 范玉枝等, 2009).阴天和晴天状况引起的太阳辐射变化对湿地生态系统NEE和碳汇功能究竟产生怎样的影响目前尚不清楚. ...
Direct observations of the effects of aerosol loading on net ecosystem CO2 exchanges over different landscapes.
1
2004
... 近年来, 大气污染、气溶胶粒子增加等环境问题日益严重, 云层和大气气溶胶含量的变化给地面接收的直接辐射和散射辐射带来很大影响(Niyogi et al., 2004; Bar-Or et al., 2010; 马金玉等, 2011).我国华北地区大气环境由于受到西北地区沙尘的影响, 大量滞留在大气中的沙尘成为对流层气溶胶的主要成分, 影响地-气系统辐射能收支, 从而影响区域的气候及生态环境(Min, 2005; 贾漩等, 2010).研究表明: 近50年来, 地面太阳总辐射从减少到增加(齐月等, 2014).而云作为一种天气现象会对局地的微气候环境产生综合效应, 当天空有云层出现时, 地面接收的太阳辐射强度及散射辐射与直射辐射的比例会发生变化, 引起生态系统光能与热能的变化, 植被的光合与呼吸作用发生变化, 最终生态系统与大气间的净CO2交换也会受到影响(Letts et al., 2005; Urban et al., 2007). ...
Hysteresis response of daytime net ecosystem exchange during drought.
2
2010
... 另外, 太阳辐射变化引起的温度变化也影响NEE对光的响应特征.首先, 温度通过影响酶系列化学反应对光合作用产生影响(Syed et al., 2006), 温度升高有利于光合速率的提高(Pingintha et al., 2010).相关分析发现, 阴天和晴天Amax与T均为显著的线性正相关(晴天: R2 = 0.80, p < 0.05; 阴天: R2 = 0.63, p < 0.05), 其次, 生态系统呼吸与气温通常显著正相关(Aires et al., 2008; Pingintha et al., 2010).本研究中虽然晴天条件下Reco,daytime较阴天提高了20%, 但NEE却显著高于阴天的67%, 说明晴天温度对湿地光合固碳能力的提高远超过生态系统呼吸的消耗, 表现为晴天生态系统的碳汇功能更强, 这与已有的研究结果一致(Han et al., 2014a). ...
... ; Pingintha et al., 2010).本研究中虽然晴天条件下Reco,daytime较阴天提高了20%, 但NEE却显著高于阴天的67%, 说明晴天温度对湿地光合固碳能力的提高远超过生态系统呼吸的消耗, 表现为晴天生态系统的碳汇功能更强, 这与已有的研究结果一致(Han et al., 2014a). ...
近50年来中国地面太阳辐射变化及其空间分布
1
2014
... 近年来, 大气污染、气溶胶粒子增加等环境问题日益严重, 云层和大气气溶胶含量的变化给地面接收的直接辐射和散射辐射带来很大影响(Niyogi et al., 2004; Bar-Or et al., 2010; 马金玉等, 2011).我国华北地区大气环境由于受到西北地区沙尘的影响, 大量滞留在大气中的沙尘成为对流层气溶胶的主要成分, 影响地-气系统辐射能收支, 从而影响区域的气候及生态环境(Min, 2005; 贾漩等, 2010).研究表明: 近50年来, 地面太阳总辐射从减少到增加(齐月等, 2014).而云作为一种天气现象会对局地的微气候环境产生综合效应, 当天空有云层出现时, 地面接收的太阳辐射强度及散射辐射与直射辐射的比例会发生变化, 引起生态系统光能与热能的变化, 植被的光合与呼吸作用发生变化, 最终生态系统与大气间的净CO2交换也会受到影响(Letts et al., 2005; Urban et al., 2007). ...
近50年来中国地面太阳辐射变化及其空间分布
1
2014
... 近年来, 大气污染、气溶胶粒子增加等环境问题日益严重, 云层和大气气溶胶含量的变化给地面接收的直接辐射和散射辐射带来很大影响(Niyogi et al., 2004; Bar-Or et al., 2010; 马金玉等, 2011).我国华北地区大气环境由于受到西北地区沙尘的影响, 大量滞留在大气中的沙尘成为对流层气溶胶的主要成分, 影响地-气系统辐射能收支, 从而影响区域的气候及生态环境(Min, 2005; 贾漩等, 2010).研究表明: 近50年来, 地面太阳总辐射从减少到增加(齐月等, 2014).而云作为一种天气现象会对局地的微气候环境产生综合效应, 当天空有云层出现时, 地面接收的太阳辐射强度及散射辐射与直射辐射的比例会发生变化, 引起生态系统光能与热能的变化, 植被的光合与呼吸作用发生变化, 最终生态系统与大气间的净CO2交换也会受到影响(Letts et al., 2005; Urban et al., 2007). ...
On the separation of net ecosystem exchange into assimilation and ecosystem respiration: Review and improved algorithm.
1
2005
... 阴天和晴天生态系统Reco,daytime与T均呈显著的正相关关系, 两者呈指数关系, 与以往很多研究的结果一致(石培礼等, 2006; Zhou et al., 2009; Zhao et al., 2010; Han et al., 2013, 2014a; 邢庆会等, 2014).T是影响生态系统呼吸的主要因素(Law et al., 2002; Reichstein et al., 2005; Urban et al., 2007), 本文研究发现: 黄河三角洲芦苇湿地晴天气温相对阴天提高了10%, 其阴天和晴天的气温差高于美国阿尔卑斯山脉南部森林的阴天和晴天的气温差(Berry & Smith, 2012), 黄河三角洲芦苇湿地晴天对应的生态系统呼吸相对阴天提高了20% (p < 0.05), 说明阴天条件下, 植被地上部分尤其是叶面温度较晴朗天气强太阳辐射条件下的低, 叶片以及植被茎干呼吸受到一定的抑制(Urban et al., 2007). ...
On the direct effect of clouds and atmospheric particles on the productivity and structure of vegetation.
1
2001
... 已有研究表明, 阴天和晴天所引起的辐射差异影响着陆地生态系统的交换过程和碳吸收能力(Roderick et al., 2001; Still et al., 2009).但由于研究方法、地域环境条件以及不同植被类型的差异, 研究结论存在较大差异(Gu et al., 1999, 2002; Alton et al., 2007; Urban et al., 2007).例如, 中国渭北刺槐(Robinia pseudoacacia)生长初期和旺盛期晴天条件下森林日均固碳量比阴天高8%-16% (郑元等, 2011).中国西北玉米(Zea mays)农田生态系统阴天条件下净生态系统CO2交换(NEE)的增长速率却高于晴天, 且生态系统总初级生产力(GPP)最大值出现于阴天(Zhang et al., 2011).对中国西北草地(Bai et al., 2012)和不同温带森林类型(Zhang et al., 2010)的研究也得出类似结论.另外, 也有研究发现加拿大常绿灌木(Letts et al., 2005)和西藏高寒草甸(范玉枝等, 2009)生态系统CO2交换不受阴天和晴天天气条件影响. ...
CO2 fluxes over plant canopies and solar radiation: A review.
1
1995
... 白天缺失的NEE数据(净辐射> 20 W·m-2), 采用Michaelis-Menten模型(Ruimy et al., 1995)进行插补. ...
Tidal effects on the organic carbon mineralization rate under aerobic conditions in sediments of an intertidal estuary.
1
2009
... 黄河口近海与海岸湿地是暖温带增长速度最快和最具代表性的新生近海与海岸湿地, 具有海陆过渡性、原生性和脆弱性的特点(布仁仓等, 1999; 崔树强, 2002).受水沙量变化、陆地和河流淡水径流、咸水海流多重影响以及人类活动的干扰, 黄河三角洲湿地生态系统CO2交换存在着极大的复杂性和不确定性(Sasaki et al., 2009;Han et al., 2014a, 2014b; 邢庆会等, 2014).研究表明黄河三角洲芦苇湿地生长季NEE占全年的80%以上, 且碳吸收集中在植物生长旺盛期(7-9月)(杨利琼等, 2013; Han et al., 2014a, 2014b).而这一时期正值黄河三角洲雨季, 其云雨状况影响太阳辐射, 进而影响湿地生态系统的碳吸收功能.在以往的研究中, 温度、水分、太阳辐射等环境因子, 生物量、叶面积等生物因子以及地下水位、地表水深等水文要素对湿地生态系统NEE的影响机制受到研究者的极大关注(牟长城等, 2009; Zhao et al., 2010; Han et al., 2013; 邢庆会等, 2014).目前, 有关阴天和晴天条件对生态系统碳交换影响的研究主要集中在森林生态系统(Liu et al., 2006; Zhang et al., 2010; 周丽艳等, 2010; Jiang et al., 2011; Zhao et al., 2011; 刘佳等, 2014)和草原生态系统(Gu et al., 2003; 范玉枝等, 2009).阴天和晴天状况引起的太阳辐射变化对湿地生态系统NEE和碳汇功能究竟产生怎样的影响目前尚不清楚. ...
西藏高原草原化嵩草草甸生态系统CO2净交换及其影响因子
1
2006
... 阴天和晴天生态系统Reco,daytime与T均呈显著的正相关关系, 两者呈指数关系, 与以往很多研究的结果一致(石培礼等, 2006; Zhou et al., 2009; Zhao et al., 2010; Han et al., 2013, 2014a; 邢庆会等, 2014).T是影响生态系统呼吸的主要因素(Law et al., 2002; Reichstein et al., 2005; Urban et al., 2007), 本文研究发现: 黄河三角洲芦苇湿地晴天气温相对阴天提高了10%, 其阴天和晴天的气温差高于美国阿尔卑斯山脉南部森林的阴天和晴天的气温差(Berry & Smith, 2012), 黄河三角洲芦苇湿地晴天对应的生态系统呼吸相对阴天提高了20% (p < 0.05), 说明阴天条件下, 植被地上部分尤其是叶面温度较晴朗天气强太阳辐射条件下的低, 叶片以及植被茎干呼吸受到一定的抑制(Urban et al., 2007). ...
西藏高原草原化嵩草草甸生态系统CO2净交换及其影响因子
1
2006
... 阴天和晴天生态系统Reco,daytime与T均呈显著的正相关关系, 两者呈指数关系, 与以往很多研究的结果一致(石培礼等, 2006; Zhou et al., 2009; Zhao et al., 2010; Han et al., 2013, 2014a; 邢庆会等, 2014).T是影响生态系统呼吸的主要因素(Law et al., 2002; Reichstein et al., 2005; Urban et al., 2007), 本文研究发现: 黄河三角洲芦苇湿地晴天气温相对阴天提高了10%, 其阴天和晴天的气温差高于美国阿尔卑斯山脉南部森林的阴天和晴天的气温差(Berry & Smith, 2012), 黄河三角洲芦苇湿地晴天对应的生态系统呼吸相对阴天提高了20% (p < 0.05), 说明阴天条件下, 植被地上部分尤其是叶面温度较晴朗天气强太阳辐射条件下的低, 叶片以及植被茎干呼吸受到一定的抑制(Urban et al., 2007). ...
CO2 fluxes from peat in boreal mires under varying temperature and moisture conditions.
1
1996
... Q10值被广泛应用于评价生态系统呼吸对温度变化的敏感程度(杨庆朋等, 2011; 杨利琼等, 2013; Han et al., 2014a, 2014b), Q10值越大, 说明生态系统呼吸对温度变化的响应越敏感.黄河三角洲芦苇湿地生态系统阴天和晴天Q10值均在已有的湿地Q10范围(1.0-7.7)内(Chapman & Thurlow, 1996; Silvola et al., 1996; Lafleur et al., 2001; Bonneville et al., 2008; Zhou et al., 2009; 周丽艳等, 2010), 其中晴天白天Q10为5.5, 远高于阴天白天的Q10 (1.9), 说明晴天白天生态系统呼吸对温度变化的敏感性相对较高.分析其原因, 一方面生态系统温度敏感性与气温有关(Kirschbaum, 1995; Xu & Qi, 2001; Janssens & Pilegaard, 2003; Davidson & Janssens., 2006; 张雷明等, 2006; Jassal et al., 2008), 气温对Q10产生影响主要通过影响光合与呼吸作用酶的活性实现(Davidson & Janssens, 2006).晴天气温T ((23.56 ± 2.62) ℃)显著大于阴天((21.26 ± 2.34) ℃), 阴天由于酶促反应需要一定的活化酶, 光合与呼吸作用酶的活性一般受到限制; 而晴天随着辐射增强, 气温升高, 越来越多的分子达到或超过了自身的活化能, 反应加快(Xiang & Freeman, 2009; Karhu et al., 2010).另一方面, 晴天相对阴天较高的PAR使得晴天光合作用更强, 产生输送的有机质增多(Curiel Yuste et al., 2007; Jassal et al., 2008), 自养呼吸增强, 同时地上植被以及土壤异养微生物呼吸对气温的响应能力增强(Edwards et al., 2004; Heinemeyer et al., 2006; Moyano et al., 2008), 而阴天由于底物基质供应不足使得自养、异养呼吸受到抑制(Gershenson et al., 2009). ...
Influence of clouds and diffuse radiation on ecosystem-atmosphere CO2 and CO18O exchanges.
1
2009
... 已有研究表明, 阴天和晴天所引起的辐射差异影响着陆地生态系统的交换过程和碳吸收能力(Roderick et al., 2001; Still et al., 2009).但由于研究方法、地域环境条件以及不同植被类型的差异, 研究结论存在较大差异(Gu et al., 1999, 2002; Alton et al., 2007; Urban et al., 2007).例如, 中国渭北刺槐(Robinia pseudoacacia)生长初期和旺盛期晴天条件下森林日均固碳量比阴天高8%-16% (郑元等, 2011).中国西北玉米(Zea mays)农田生态系统阴天条件下净生态系统CO2交换(NEE)的增长速率却高于晴天, 且生态系统总初级生产力(GPP)最大值出现于阴天(Zhang et al., 2011).对中国西北草地(Bai et al., 2012)和不同温带森林类型(Zhang et al., 2010)的研究也得出类似结论.另外, 也有研究发现加拿大常绿灌木(Letts et al., 2005)和西藏高寒草甸(范玉枝等, 2009)生态系统CO2交换不受阴天和晴天天气条件影响. ...
Environmental control of net ecosystem CO2 exchange in a treed, moderately rich fen in northern Alberta.
2
2006
... 在一定光照强度范围内, 光合作用随PAR的增强而增加(Glenn et al., 2006; Syed et al., 2006; Hao et al., 2011; 同小娟等, 2011).光能过剩会对光合作用产生不良影响, 出现光抑制, 致使叶绿体光合效率下降(Jiang et al., 2011; Tholen et al., 2012).晴天PAR显著大于阴天(图2B), 因此, 晴天条件下植被叶片光合作用可能大于阴天, 表现为晴天Amax显著大于阴天(表1).Zhou等(2009)研究认为Amax与植被的环境条件有关, 其中晴天Amax较阴天条件下提高了30%, 说明在植被生长旺季, 天空有云层覆盖时芦苇的光合能力比晴朗天气无云时有所降低, Law等(2002)总结全球森林、草地、农田以及苔原生态系统碳交换相关研究也得出类似结论.阴天条件下, 伴随天空云量的增多, PAR显著降低(图2B), 尽管阴天和晴天条件下, NEE均随PAR的增大而增加(图3), 但由于阴天PAR较低, 且晴天未出现光抑制现象, 因此其NEE显著低于晴天(图2A).Alton (2008)在分析北美洲和欧洲6种森林类型2-3年的碳通量观测数据后发现, 这些森林阴天的日固碳量比晴天少60%-80%.相反, 在研究太阳辐射对黄河小浪底人工混交林生态系统碳交换的影响中发现, 多云天气下净碳交换会增加(刘佳等, 2014).造成研究结论不一致的原因是多方面的, 不同的阴天和晴天划分指标(Graham et al., 2003; Dengel & Grace, 2010)和拟合模型(Zhang et al., 2011)来估计天气对碳通量的影响, 散射光和直射光下的碳交换没有一直处在相同的天气辐射条件下进行对比.此外, 研究地域的差异和不同植被类型的特性也在很大程度上影响了碳交换过程对天气的响应过程. ...
... 另外, 太阳辐射变化引起的温度变化也影响NEE对光的响应特征.首先, 温度通过影响酶系列化学反应对光合作用产生影响(Syed et al., 2006), 温度升高有利于光合速率的提高(Pingintha et al., 2010).相关分析发现, 阴天和晴天Amax与T均为显著的线性正相关(晴天: R2 = 0.80, p < 0.05; 阴天: R2 = 0.63, p < 0.05), 其次, 生态系统呼吸与气温通常显著正相关(Aires et al., 2008; Pingintha et al., 2010).本研究中虽然晴天条件下Reco,daytime较阴天提高了20%, 但NEE却显著高于阴天的67%, 说明晴天温度对湿地光合固碳能力的提高远超过生态系统呼吸的消耗, 表现为晴天生态系统的碳汇功能更强, 这与已有的研究结果一致(Han et al., 2014a). ...
Tree photosynthesis modulates soil respiration on a diurnal time scale.
1
2005
... 本研究以黄河三角洲芦苇湿地为研究对象, 在自然条件下选择相邻的阴天和晴天, 在生物要素(生物量、叶面积指数)、土壤水分以及养分特征保持不变的前提下, 分析阴天和晴天引起的辐射变化对生态系统NEE的影响机制.目前, 有关太阳辐射对生态系统NEE的影响机制大都以晴空指数为指标(Gu et al., 2003; Liu et al., 2006; 范玉枝等, 2009; Zhang et al., 2010; 周丽艳等, 2010; 蒋琰等, 2011; Zhao et al., 2011; 刘佳等, 2014).这种研究方法的最大不足就是研究阶段中, 太阳辐射、生物要素(生物量、叶面积指数)、土壤湿度等环境因子都在发生变化, 这对阐明太阳辐射这一单因素对NEE的控制机制研究带来一定困难.而本研究巧妙地在自然条件下选择相邻的阴天和晴天, 在日尺度上实现了对单一变量PAR的控制.利用这种研究方法, Han等(2014a)通过对比分析阴天和晴天植物冠层光合作用的差异性, 定量分析了光合作用在日尺度上对土壤呼吸的调节作用.此外, 我们已有研究发现PAR改变所引起的植物光合作用变化对生态系统呼吸会产生一定的影响(Tang et al., 2005; Baldocchi et al., 2006; Moyano et al., 2008; Wingate et al., 2010; Han et al., 2012).但是, 以往的研究多通过夜间生态系统呼吸与气温函数关系外延的方法来估算白天生态系统呼吸(Doughty et al., 2010; Zhang et al., 2011), 忽略了PAR对白天生态系统呼吸的影响, 因而具有一定的局限性.考虑到白天PAR的变化, 本研究采用Michaelis-Menten模型对白天生态系统呼吸进行推算, 使得生态系统呼吸的温度响应数据更具有说服力. ...
Opinion: Prospects for improving photosynthesis by altering leaf anatomy.
1
2012
... 在一定光照强度范围内, 光合作用随PAR的增强而增加(Glenn et al., 2006; Syed et al., 2006; Hao et al., 2011; 同小娟等, 2011).光能过剩会对光合作用产生不良影响, 出现光抑制, 致使叶绿体光合效率下降(Jiang et al., 2011; Tholen et al., 2012).晴天PAR显著大于阴天(图2B), 因此, 晴天条件下植被叶片光合作用可能大于阴天, 表现为晴天Amax显著大于阴天(表1).Zhou等(2009)研究认为Amax与植被的环境条件有关, 其中晴天Amax较阴天条件下提高了30%, 说明在植被生长旺季, 天空有云层覆盖时芦苇的光合能力比晴朗天气无云时有所降低, Law等(2002)总结全球森林、草地、农田以及苔原生态系统碳交换相关研究也得出类似结论.阴天条件下, 伴随天空云量的增多, PAR显著降低(图2B), 尽管阴天和晴天条件下, NEE均随PAR的增大而增加(图3), 但由于阴天PAR较低, 且晴天未出现光抑制现象, 因此其NEE显著低于晴天(图2A).Alton (2008)在分析北美洲和欧洲6种森林类型2-3年的碳通量观测数据后发现, 这些森林阴天的日固碳量比晴天少60%-80%.相反, 在研究太阳辐射对黄河小浪底人工混交林生态系统碳交换的影响中发现, 多云天气下净碳交换会增加(刘佳等, 2014).造成研究结论不一致的原因是多方面的, 不同的阴天和晴天划分指标(Graham et al., 2003; Dengel & Grace, 2010)和拟合模型(Zhang et al., 2011)来估计天气对碳通量的影响, 散射光和直射光下的碳交换没有一直处在相同的天气辐射条件下进行对比.此外, 研究地域的差异和不同植被类型的特性也在很大程度上影响了碳交换过程对天气的响应过程. ...
华北平原玉米田生态系统光合作用特征及影响因素
1
2011
... 在一定光照强度范围内, 光合作用随PAR的增强而增加(Glenn et al., 2006; Syed et al., 2006; Hao et al., 2011; 同小娟等, 2011).光能过剩会对光合作用产生不良影响, 出现光抑制, 致使叶绿体光合效率下降(Jiang et al., 2011; Tholen et al., 2012).晴天PAR显著大于阴天(图2B), 因此, 晴天条件下植被叶片光合作用可能大于阴天, 表现为晴天Amax显著大于阴天(表1).Zhou等(2009)研究认为Amax与植被的环境条件有关, 其中晴天Amax较阴天条件下提高了30%, 说明在植被生长旺季, 天空有云层覆盖时芦苇的光合能力比晴朗天气无云时有所降低, Law等(2002)总结全球森林、草地、农田以及苔原生态系统碳交换相关研究也得出类似结论.阴天条件下, 伴随天空云量的增多, PAR显著降低(图2B), 尽管阴天和晴天条件下, NEE均随PAR的增大而增加(图3), 但由于阴天PAR较低, 且晴天未出现光抑制现象, 因此其NEE显著低于晴天(图2A).Alton (2008)在分析北美洲和欧洲6种森林类型2-3年的碳通量观测数据后发现, 这些森林阴天的日固碳量比晴天少60%-80%.相反, 在研究太阳辐射对黄河小浪底人工混交林生态系统碳交换的影响中发现, 多云天气下净碳交换会增加(刘佳等, 2014).造成研究结论不一致的原因是多方面的, 不同的阴天和晴天划分指标(Graham et al., 2003; Dengel & Grace, 2010)和拟合模型(Zhang et al., 2011)来估计天气对碳通量的影响, 散射光和直射光下的碳交换没有一直处在相同的天气辐射条件下进行对比.此外, 研究地域的差异和不同植被类型的特性也在很大程度上影响了碳交换过程对天气的响应过程. ...
华北平原玉米田生态系统光合作用特征及影响因素
1
2011
... 在一定光照强度范围内, 光合作用随PAR的增强而增加(Glenn et al., 2006; Syed et al., 2006; Hao et al., 2011; 同小娟等, 2011).光能过剩会对光合作用产生不良影响, 出现光抑制, 致使叶绿体光合效率下降(Jiang et al., 2011; Tholen et al., 2012).晴天PAR显著大于阴天(图2B), 因此, 晴天条件下植被叶片光合作用可能大于阴天, 表现为晴天Amax显著大于阴天(表1).Zhou等(2009)研究认为Amax与植被的环境条件有关, 其中晴天Amax较阴天条件下提高了30%, 说明在植被生长旺季, 天空有云层覆盖时芦苇的光合能力比晴朗天气无云时有所降低, Law等(2002)总结全球森林、草地、农田以及苔原生态系统碳交换相关研究也得出类似结论.阴天条件下, 伴随天空云量的增多, PAR显著降低(图2B), 尽管阴天和晴天条件下, NEE均随PAR的增大而增加(图3), 但由于阴天PAR较低, 且晴天未出现光抑制现象, 因此其NEE显著低于晴天(图2A).Alton (2008)在分析北美洲和欧洲6种森林类型2-3年的碳通量观测数据后发现, 这些森林阴天的日固碳量比晴天少60%-80%.相反, 在研究太阳辐射对黄河小浪底人工混交林生态系统碳交换的影响中发现, 多云天气下净碳交换会增加(刘佳等, 2014).造成研究结论不一致的原因是多方面的, 不同的阴天和晴天划分指标(Graham et al., 2003; Dengel & Grace, 2010)和拟合模型(Zhang et al., 2011)来估计天气对碳通量的影响, 散射光和直射光下的碳交换没有一直处在相同的天气辐射条件下进行对比.此外, 研究地域的差异和不同植被类型的特性也在很大程度上影响了碳交换过程对天气的响应过程. ...
Ecophysiological controls over the net ecosystem exchange of mountain spruce stand. Comparison of the response in direct vs. diffuse solar radiation.
3
2007
... 近年来, 大气污染、气溶胶粒子增加等环境问题日益严重, 云层和大气气溶胶含量的变化给地面接收的直接辐射和散射辐射带来很大影响(Niyogi et al., 2004; Bar-Or et al., 2010; 马金玉等, 2011).我国华北地区大气环境由于受到西北地区沙尘的影响, 大量滞留在大气中的沙尘成为对流层气溶胶的主要成分, 影响地-气系统辐射能收支, 从而影响区域的气候及生态环境(Min, 2005; 贾漩等, 2010).研究表明: 近50年来, 地面太阳总辐射从减少到增加(齐月等, 2014).而云作为一种天气现象会对局地的微气候环境产生综合效应, 当天空有云层出现时, 地面接收的太阳辐射强度及散射辐射与直射辐射的比例会发生变化, 引起生态系统光能与热能的变化, 植被的光合与呼吸作用发生变化, 最终生态系统与大气间的净CO2交换也会受到影响(Letts et al., 2005; Urban et al., 2007). ...
... 已有研究表明, 阴天和晴天所引起的辐射差异影响着陆地生态系统的交换过程和碳吸收能力(Roderick et al., 2001; Still et al., 2009).但由于研究方法、地域环境条件以及不同植被类型的差异, 研究结论存在较大差异(Gu et al., 1999, 2002; Alton et al., 2007; Urban et al., 2007).例如, 中国渭北刺槐(Robinia pseudoacacia)生长初期和旺盛期晴天条件下森林日均固碳量比阴天高8%-16% (郑元等, 2011).中国西北玉米(Zea mays)农田生态系统阴天条件下净生态系统CO2交换(NEE)的增长速率却高于晴天, 且生态系统总初级生产力(GPP)最大值出现于阴天(Zhang et al., 2011).对中国西北草地(Bai et al., 2012)和不同温带森林类型(Zhang et al., 2010)的研究也得出类似结论.另外, 也有研究发现加拿大常绿灌木(Letts et al., 2005)和西藏高寒草甸(范玉枝等, 2009)生态系统CO2交换不受阴天和晴天天气条件影响. ...
... 阴天和晴天生态系统Reco,daytime与T均呈显著的正相关关系, 两者呈指数关系, 与以往很多研究的结果一致(石培礼等, 2006; Zhou et al., 2009; Zhao et al., 2010; Han et al., 2013, 2014a; 邢庆会等, 2014).T是影响生态系统呼吸的主要因素(Law et al., 2002; Reichstein et al., 2005; Urban et al., 2007), 本文研究发现: 黄河三角洲芦苇湿地晴天气温相对阴天提高了10%, 其阴天和晴天的气温差高于美国阿尔卑斯山脉南部森林的阴天和晴天的气温差(Berry & Smith, 2012), 黄河三角洲芦苇湿地晴天对应的生态系统呼吸相对阴天提高了20% (p < 0.05), 说明阴天条件下, 植被地上部分尤其是叶面温度较晴朗天气强太阳辐射条件下的低, 叶片以及植被茎干呼吸受到一定的抑制(Urban et al., 2007). ...
Photosynthetic carbon isotope discrimination and its relationship to the carbon isotope signals of stem, soil and ecosystem respiration.
1
2010
... 本研究以黄河三角洲芦苇湿地为研究对象, 在自然条件下选择相邻的阴天和晴天, 在生物要素(生物量、叶面积指数)、土壤水分以及养分特征保持不变的前提下, 分析阴天和晴天引起的辐射变化对生态系统NEE的影响机制.目前, 有关太阳辐射对生态系统NEE的影响机制大都以晴空指数为指标(Gu et al., 2003; Liu et al., 2006; 范玉枝等, 2009; Zhang et al., 2010; 周丽艳等, 2010; 蒋琰等, 2011; Zhao et al., 2011; 刘佳等, 2014).这种研究方法的最大不足就是研究阶段中, 太阳辐射、生物要素(生物量、叶面积指数)、土壤湿度等环境因子都在发生变化, 这对阐明太阳辐射这一单因素对NEE的控制机制研究带来一定困难.而本研究巧妙地在自然条件下选择相邻的阴天和晴天, 在日尺度上实现了对单一变量PAR的控制.利用这种研究方法, Han等(2014a)通过对比分析阴天和晴天植物冠层光合作用的差异性, 定量分析了光合作用在日尺度上对土壤呼吸的调节作用.此外, 我们已有研究发现PAR改变所引起的植物光合作用变化对生态系统呼吸会产生一定的影响(Tang et al., 2005; Baldocchi et al., 2006; Moyano et al., 2008; Wingate et al., 2010; Han et al., 2012).但是, 以往的研究多通过夜间生态系统呼吸与气温函数关系外延的方法来估算白天生态系统呼吸(Doughty et al., 2010; Zhang et al., 2011), 忽略了PAR对白天生态系统呼吸的影响, 因而具有一定的局限性.考虑到白天PAR的变化, 本研究采用Michaelis-Menten模型对白天生态系统呼吸进行推算, 使得生态系统呼吸的温度响应数据更具有说服力. ...
Annual variation of temperature sensitivity of soil organic carbon decomposition in North peatlands: Implications for thermal responses of carbon cycling to global warming.
1
2009
... Q10值被广泛应用于评价生态系统呼吸对温度变化的敏感程度(杨庆朋等, 2011; 杨利琼等, 2013; Han et al., 2014a, 2014b), Q10值越大, 说明生态系统呼吸对温度变化的响应越敏感.黄河三角洲芦苇湿地生态系统阴天和晴天Q10值均在已有的湿地Q10范围(1.0-7.7)内(Chapman & Thurlow, 1996; Silvola et al., 1996; Lafleur et al., 2001; Bonneville et al., 2008; Zhou et al., 2009; 周丽艳等, 2010), 其中晴天白天Q10为5.5, 远高于阴天白天的Q10 (1.9), 说明晴天白天生态系统呼吸对温度变化的敏感性相对较高.分析其原因, 一方面生态系统温度敏感性与气温有关(Kirschbaum, 1995; Xu & Qi, 2001; Janssens & Pilegaard, 2003; Davidson & Janssens., 2006; 张雷明等, 2006; Jassal et al., 2008), 气温对Q10产生影响主要通过影响光合与呼吸作用酶的活性实现(Davidson & Janssens, 2006).晴天气温T ((23.56 ± 2.62) ℃)显著大于阴天((21.26 ± 2.34) ℃), 阴天由于酶促反应需要一定的活化酶, 光合与呼吸作用酶的活性一般受到限制; 而晴天随着辐射增强, 气温升高, 越来越多的分子达到或超过了自身的活化能, 反应加快(Xiang & Freeman, 2009; Karhu et al., 2010).另一方面, 晴天相对阴天较高的PAR使得晴天光合作用更强, 产生输送的有机质增多(Curiel Yuste et al., 2007; Jassal et al., 2008), 自养呼吸增强, 同时地上植被以及土壤异养微生物呼吸对气温的响应能力增强(Edwards et al., 2004; Heinemeyer et al., 2006; Moyano et al., 2008), 而阴天由于底物基质供应不足使得自养、异养呼吸受到抑制(Gershenson et al., 2009). ...
黄河口潮间盐沼湿地生长季净生态系统CO2交换特征及其影响因素
4
2014
... 黄河口近海与海岸湿地是暖温带增长速度最快和最具代表性的新生近海与海岸湿地, 具有海陆过渡性、原生性和脆弱性的特点(布仁仓等, 1999; 崔树强, 2002).受水沙量变化、陆地和河流淡水径流、咸水海流多重影响以及人类活动的干扰, 黄河三角洲湿地生态系统CO2交换存在着极大的复杂性和不确定性(Sasaki et al., 2009;Han et al., 2014a, 2014b; 邢庆会等, 2014).研究表明黄河三角洲芦苇湿地生长季NEE占全年的80%以上, 且碳吸收集中在植物生长旺盛期(7-9月)(杨利琼等, 2013; Han et al., 2014a, 2014b).而这一时期正值黄河三角洲雨季, 其云雨状况影响太阳辐射, 进而影响湿地生态系统的碳吸收功能.在以往的研究中, 温度、水分、太阳辐射等环境因子, 生物量、叶面积等生物因子以及地下水位、地表水深等水文要素对湿地生态系统NEE的影响机制受到研究者的极大关注(牟长城等, 2009; Zhao et al., 2010; Han et al., 2013; 邢庆会等, 2014).目前, 有关阴天和晴天条件对生态系统碳交换影响的研究主要集中在森林生态系统(Liu et al., 2006; Zhang et al., 2010; 周丽艳等, 2010; Jiang et al., 2011; Zhao et al., 2011; 刘佳等, 2014)和草原生态系统(Gu et al., 2003; 范玉枝等, 2009).阴天和晴天状况引起的太阳辐射变化对湿地生态系统NEE和碳汇功能究竟产生怎样的影响目前尚不清楚. ...
... ; 邢庆会等, 2014).目前, 有关阴天和晴天条件对生态系统碳交换影响的研究主要集中在森林生态系统(Liu et al., 2006; Zhang et al., 2010; 周丽艳等, 2010; Jiang et al., 2011; Zhao et al., 2011; 刘佳等, 2014)和草原生态系统(Gu et al., 2003; 范玉枝等, 2009).阴天和晴天状况引起的太阳辐射变化对湿地生态系统NEE和碳汇功能究竟产生怎样的影响目前尚不清楚. ...
... 本文研究发现晴天和阴天芦苇湿地NEE对PAR的响应趋势一致, 均呈直角双曲线关系, 与先前的很多研究结果一致(Falge et al., 2001; Monson et al., 2002; Zhou et al., 2009; 张弥等, 2009; Han et al., 2014a, 2014b; 邢庆会等, 2014).PAR是控制阴天和晴天生态系统生长季昼间NEE变化的主要因素(Flanagan & Johnson, 2005; Liu et al., 2006; 薛红喜等, 2012), Michaelis-Menten方程拟合发现, PAR可以解释晴天单天NEE变化的57%-82%, 解释阴天单天NEE变化的53%-81%.云层变化会改变地表接收太阳辐射的强度, 影响PAR、T等环境因子, 进而影响阴天和晴天条件下NEE对光的响应特征(Baldocchi, 1997; Freedman et al., 2001). ...
... 阴天和晴天生态系统Reco,daytime与T均呈显著的正相关关系, 两者呈指数关系, 与以往很多研究的结果一致(石培礼等, 2006; Zhou et al., 2009; Zhao et al., 2010; Han et al., 2013, 2014a; 邢庆会等, 2014).T是影响生态系统呼吸的主要因素(Law et al., 2002; Reichstein et al., 2005; Urban et al., 2007), 本文研究发现: 黄河三角洲芦苇湿地晴天气温相对阴天提高了10%, 其阴天和晴天的气温差高于美国阿尔卑斯山脉南部森林的阴天和晴天的气温差(Berry & Smith, 2012), 黄河三角洲芦苇湿地晴天对应的生态系统呼吸相对阴天提高了20% (p < 0.05), 说明阴天条件下, 植被地上部分尤其是叶面温度较晴朗天气强太阳辐射条件下的低, 叶片以及植被茎干呼吸受到一定的抑制(Urban et al., 2007). ...
黄河口潮间盐沼湿地生长季净生态系统CO2交换特征及其影响因素
4
2014
... 黄河口近海与海岸湿地是暖温带增长速度最快和最具代表性的新生近海与海岸湿地, 具有海陆过渡性、原生性和脆弱性的特点(布仁仓等, 1999; 崔树强, 2002).受水沙量变化、陆地和河流淡水径流、咸水海流多重影响以及人类活动的干扰, 黄河三角洲湿地生态系统CO2交换存在着极大的复杂性和不确定性(Sasaki et al., 2009;Han et al., 2014a, 2014b; 邢庆会等, 2014).研究表明黄河三角洲芦苇湿地生长季NEE占全年的80%以上, 且碳吸收集中在植物生长旺盛期(7-9月)(杨利琼等, 2013; Han et al., 2014a, 2014b).而这一时期正值黄河三角洲雨季, 其云雨状况影响太阳辐射, 进而影响湿地生态系统的碳吸收功能.在以往的研究中, 温度、水分、太阳辐射等环境因子, 生物量、叶面积等生物因子以及地下水位、地表水深等水文要素对湿地生态系统NEE的影响机制受到研究者的极大关注(牟长城等, 2009; Zhao et al., 2010; Han et al., 2013; 邢庆会等, 2014).目前, 有关阴天和晴天条件对生态系统碳交换影响的研究主要集中在森林生态系统(Liu et al., 2006; Zhang et al., 2010; 周丽艳等, 2010; Jiang et al., 2011; Zhao et al., 2011; 刘佳等, 2014)和草原生态系统(Gu et al., 2003; 范玉枝等, 2009).阴天和晴天状况引起的太阳辐射变化对湿地生态系统NEE和碳汇功能究竟产生怎样的影响目前尚不清楚. ...
... ; 邢庆会等, 2014).目前, 有关阴天和晴天条件对生态系统碳交换影响的研究主要集中在森林生态系统(Liu et al., 2006; Zhang et al., 2010; 周丽艳等, 2010; Jiang et al., 2011; Zhao et al., 2011; 刘佳等, 2014)和草原生态系统(Gu et al., 2003; 范玉枝等, 2009).阴天和晴天状况引起的太阳辐射变化对湿地生态系统NEE和碳汇功能究竟产生怎样的影响目前尚不清楚. ...
... 本文研究发现晴天和阴天芦苇湿地NEE对PAR的响应趋势一致, 均呈直角双曲线关系, 与先前的很多研究结果一致(Falge et al., 2001; Monson et al., 2002; Zhou et al., 2009; 张弥等, 2009; Han et al., 2014a, 2014b; 邢庆会等, 2014).PAR是控制阴天和晴天生态系统生长季昼间NEE变化的主要因素(Flanagan & Johnson, 2005; Liu et al., 2006; 薛红喜等, 2012), Michaelis-Menten方程拟合发现, PAR可以解释晴天单天NEE变化的57%-82%, 解释阴天单天NEE变化的53%-81%.云层变化会改变地表接收太阳辐射的强度, 影响PAR、T等环境因子, 进而影响阴天和晴天条件下NEE对光的响应特征(Baldocchi, 1997; Freedman et al., 2001). ...
... 阴天和晴天生态系统Reco,daytime与T均呈显著的正相关关系, 两者呈指数关系, 与以往很多研究的结果一致(石培礼等, 2006; Zhou et al., 2009; Zhao et al., 2010; Han et al., 2013, 2014a; 邢庆会等, 2014).T是影响生态系统呼吸的主要因素(Law et al., 2002; Reichstein et al., 2005; Urban et al., 2007), 本文研究发现: 黄河三角洲芦苇湿地晴天气温相对阴天提高了10%, 其阴天和晴天的气温差高于美国阿尔卑斯山脉南部森林的阴天和晴天的气温差(Berry & Smith, 2012), 黄河三角洲芦苇湿地晴天对应的生态系统呼吸相对阴天提高了20% (p < 0.05), 说明阴天条件下, 植被地上部分尤其是叶面温度较晴朗天气强太阳辐射条件下的低, 叶片以及植被茎干呼吸受到一定的抑制(Urban et al., 2007). ...
Spatial and seasonal variations of Q10 determined by soil respiration measurements at a Sierra Nevadan forest.
1
2001
... Q10值被广泛应用于评价生态系统呼吸对温度变化的敏感程度(杨庆朋等, 2011; 杨利琼等, 2013; Han et al., 2014a, 2014b), Q10值越大, 说明生态系统呼吸对温度变化的响应越敏感.黄河三角洲芦苇湿地生态系统阴天和晴天Q10值均在已有的湿地Q10范围(1.0-7.7)内(Chapman & Thurlow, 1996; Silvola et al., 1996; Lafleur et al., 2001; Bonneville et al., 2008; Zhou et al., 2009; 周丽艳等, 2010), 其中晴天白天Q10为5.5, 远高于阴天白天的Q10 (1.9), 说明晴天白天生态系统呼吸对温度变化的敏感性相对较高.分析其原因, 一方面生态系统温度敏感性与气温有关(Kirschbaum, 1995; Xu & Qi, 2001; Janssens & Pilegaard, 2003; Davidson & Janssens., 2006; 张雷明等, 2006; Jassal et al., 2008), 气温对Q10产生影响主要通过影响光合与呼吸作用酶的活性实现(Davidson & Janssens, 2006).晴天气温T ((23.56 ± 2.62) ℃)显著大于阴天((21.26 ± 2.34) ℃), 阴天由于酶促反应需要一定的活化酶, 光合与呼吸作用酶的活性一般受到限制; 而晴天随着辐射增强, 气温升高, 越来越多的分子达到或超过了自身的活化能, 反应加快(Xiang & Freeman, 2009; Karhu et al., 2010).另一方面, 晴天相对阴天较高的PAR使得晴天光合作用更强, 产生输送的有机质增多(Curiel Yuste et al., 2007; Jassal et al., 2008), 自养呼吸增强, 同时地上植被以及土壤异养微生物呼吸对气温的响应能力增强(Edwards et al., 2004; Heinemeyer et al., 2006; Moyano et al., 2008), 而阴天由于底物基质供应不足使得自养、异养呼吸受到抑制(Gershenson et al., 2009). ...
基于涡度相关法的中国农田生态系统碳通量研究进展
1
2012
... 本文研究发现晴天和阴天芦苇湿地NEE对PAR的响应趋势一致, 均呈直角双曲线关系, 与先前的很多研究结果一致(Falge et al., 2001; Monson et al., 2002; Zhou et al., 2009; 张弥等, 2009; Han et al., 2014a, 2014b; 邢庆会等, 2014).PAR是控制阴天和晴天生态系统生长季昼间NEE变化的主要因素(Flanagan & Johnson, 2005; Liu et al., 2006; 薛红喜等, 2012), Michaelis-Menten方程拟合发现, PAR可以解释晴天单天NEE变化的57%-82%, 解释阴天单天NEE变化的53%-81%.云层变化会改变地表接收太阳辐射的强度, 影响PAR、T等环境因子, 进而影响阴天和晴天条件下NEE对光的响应特征(Baldocchi, 1997; Freedman et al., 2001). ...
基于涡度相关法的中国农田生态系统碳通量研究进展
1
2012
... 本文研究发现晴天和阴天芦苇湿地NEE对PAR的响应趋势一致, 均呈直角双曲线关系, 与先前的很多研究结果一致(Falge et al., 2001; Monson et al., 2002; Zhou et al., 2009; 张弥等, 2009; Han et al., 2014a, 2014b; 邢庆会等, 2014).PAR是控制阴天和晴天生态系统生长季昼间NEE变化的主要因素(Flanagan & Johnson, 2005; Liu et al., 2006; 薛红喜等, 2012), Michaelis-Menten方程拟合发现, PAR可以解释晴天单天NEE变化的57%-82%, 解释阴天单天NEE变化的53%-81%.云层变化会改变地表接收太阳辐射的强度, 影响PAR、T等环境因子, 进而影响阴天和晴天条件下NEE对光的响应特征(Baldocchi, 1997; Freedman et al., 2001). ...
开垦对黄河三角洲湿地净生态系统CO2交换的影响
2
2013
... 黄河口近海与海岸湿地是暖温带增长速度最快和最具代表性的新生近海与海岸湿地, 具有海陆过渡性、原生性和脆弱性的特点(布仁仓等, 1999; 崔树强, 2002).受水沙量变化、陆地和河流淡水径流、咸水海流多重影响以及人类活动的干扰, 黄河三角洲湿地生态系统CO2交换存在着极大的复杂性和不确定性(Sasaki et al., 2009;Han et al., 2014a, 2014b; 邢庆会等, 2014).研究表明黄河三角洲芦苇湿地生长季NEE占全年的80%以上, 且碳吸收集中在植物生长旺盛期(7-9月)(杨利琼等, 2013; Han et al., 2014a, 2014b).而这一时期正值黄河三角洲雨季, 其云雨状况影响太阳辐射, 进而影响湿地生态系统的碳吸收功能.在以往的研究中, 温度、水分、太阳辐射等环境因子, 生物量、叶面积等生物因子以及地下水位、地表水深等水文要素对湿地生态系统NEE的影响机制受到研究者的极大关注(牟长城等, 2009; Zhao et al., 2010; Han et al., 2013; 邢庆会等, 2014).目前, 有关阴天和晴天条件对生态系统碳交换影响的研究主要集中在森林生态系统(Liu et al., 2006; Zhang et al., 2010; 周丽艳等, 2010; Jiang et al., 2011; Zhao et al., 2011; 刘佳等, 2014)和草原生态系统(Gu et al., 2003; 范玉枝等, 2009).阴天和晴天状况引起的太阳辐射变化对湿地生态系统NEE和碳汇功能究竟产生怎样的影响目前尚不清楚. ...
... Q10值被广泛应用于评价生态系统呼吸对温度变化的敏感程度(杨庆朋等, 2011; 杨利琼等, 2013; Han et al., 2014a, 2014b), Q10值越大, 说明生态系统呼吸对温度变化的响应越敏感.黄河三角洲芦苇湿地生态系统阴天和晴天Q10值均在已有的湿地Q10范围(1.0-7.7)内(Chapman & Thurlow, 1996; Silvola et al., 1996; Lafleur et al., 2001; Bonneville et al., 2008; Zhou et al., 2009; 周丽艳等, 2010), 其中晴天白天Q10为5.5, 远高于阴天白天的Q10 (1.9), 说明晴天白天生态系统呼吸对温度变化的敏感性相对较高.分析其原因, 一方面生态系统温度敏感性与气温有关(Kirschbaum, 1995; Xu & Qi, 2001; Janssens & Pilegaard, 2003; Davidson & Janssens., 2006; 张雷明等, 2006; Jassal et al., 2008), 气温对Q10产生影响主要通过影响光合与呼吸作用酶的活性实现(Davidson & Janssens, 2006).晴天气温T ((23.56 ± 2.62) ℃)显著大于阴天((21.26 ± 2.34) ℃), 阴天由于酶促反应需要一定的活化酶, 光合与呼吸作用酶的活性一般受到限制; 而晴天随着辐射增强, 气温升高, 越来越多的分子达到或超过了自身的活化能, 反应加快(Xiang & Freeman, 2009; Karhu et al., 2010).另一方面, 晴天相对阴天较高的PAR使得晴天光合作用更强, 产生输送的有机质增多(Curiel Yuste et al., 2007; Jassal et al., 2008), 自养呼吸增强, 同时地上植被以及土壤异养微生物呼吸对气温的响应能力增强(Edwards et al., 2004; Heinemeyer et al., 2006; Moyano et al., 2008), 而阴天由于底物基质供应不足使得自养、异养呼吸受到抑制(Gershenson et al., 2009). ...
开垦对黄河三角洲湿地净生态系统CO2交换的影响
2
2013
... 黄河口近海与海岸湿地是暖温带增长速度最快和最具代表性的新生近海与海岸湿地, 具有海陆过渡性、原生性和脆弱性的特点(布仁仓等, 1999; 崔树强, 2002).受水沙量变化、陆地和河流淡水径流、咸水海流多重影响以及人类活动的干扰, 黄河三角洲湿地生态系统CO2交换存在着极大的复杂性和不确定性(Sasaki et al., 2009;Han et al., 2014a, 2014b; 邢庆会等, 2014).研究表明黄河三角洲芦苇湿地生长季NEE占全年的80%以上, 且碳吸收集中在植物生长旺盛期(7-9月)(杨利琼等, 2013; Han et al., 2014a, 2014b).而这一时期正值黄河三角洲雨季, 其云雨状况影响太阳辐射, 进而影响湿地生态系统的碳吸收功能.在以往的研究中, 温度、水分、太阳辐射等环境因子, 生物量、叶面积等生物因子以及地下水位、地表水深等水文要素对湿地生态系统NEE的影响机制受到研究者的极大关注(牟长城等, 2009; Zhao et al., 2010; Han et al., 2013; 邢庆会等, 2014).目前, 有关阴天和晴天条件对生态系统碳交换影响的研究主要集中在森林生态系统(Liu et al., 2006; Zhang et al., 2010; 周丽艳等, 2010; Jiang et al., 2011; Zhao et al., 2011; 刘佳等, 2014)和草原生态系统(Gu et al., 2003; 范玉枝等, 2009).阴天和晴天状况引起的太阳辐射变化对湿地生态系统NEE和碳汇功能究竟产生怎样的影响目前尚不清楚. ...
... Q10值被广泛应用于评价生态系统呼吸对温度变化的敏感程度(杨庆朋等, 2011; 杨利琼等, 2013; Han et al., 2014a, 2014b), Q10值越大, 说明生态系统呼吸对温度变化的响应越敏感.黄河三角洲芦苇湿地生态系统阴天和晴天Q10值均在已有的湿地Q10范围(1.0-7.7)内(Chapman & Thurlow, 1996; Silvola et al., 1996; Lafleur et al., 2001; Bonneville et al., 2008; Zhou et al., 2009; 周丽艳等, 2010), 其中晴天白天Q10为5.5, 远高于阴天白天的Q10 (1.9), 说明晴天白天生态系统呼吸对温度变化的敏感性相对较高.分析其原因, 一方面生态系统温度敏感性与气温有关(Kirschbaum, 1995; Xu & Qi, 2001; Janssens & Pilegaard, 2003; Davidson & Janssens., 2006; 张雷明等, 2006; Jassal et al., 2008), 气温对Q10产生影响主要通过影响光合与呼吸作用酶的活性实现(Davidson & Janssens, 2006).晴天气温T ((23.56 ± 2.62) ℃)显著大于阴天((21.26 ± 2.34) ℃), 阴天由于酶促反应需要一定的活化酶, 光合与呼吸作用酶的活性一般受到限制; 而晴天随着辐射增强, 气温升高, 越来越多的分子达到或超过了自身的活化能, 反应加快(Xiang & Freeman, 2009; Karhu et al., 2010).另一方面, 晴天相对阴天较高的PAR使得晴天光合作用更强, 产生输送的有机质增多(Curiel Yuste et al., 2007; Jassal et al., 2008), 自养呼吸增强, 同时地上植被以及土壤异养微生物呼吸对气温的响应能力增强(Edwards et al., 2004; Heinemeyer et al., 2006; Moyano et al., 2008), 而阴天由于底物基质供应不足使得自养、异养呼吸受到抑制(Gershenson et al., 2009). ...
土壤呼吸温度敏感性的影响因素和不确定性
1
2011
... Q10值被广泛应用于评价生态系统呼吸对温度变化的敏感程度(杨庆朋等, 2011; 杨利琼等, 2013; Han et al., 2014a, 2014b), Q10值越大, 说明生态系统呼吸对温度变化的响应越敏感.黄河三角洲芦苇湿地生态系统阴天和晴天Q10值均在已有的湿地Q10范围(1.0-7.7)内(Chapman & Thurlow, 1996; Silvola et al., 1996; Lafleur et al., 2001; Bonneville et al., 2008; Zhou et al., 2009; 周丽艳等, 2010), 其中晴天白天Q10为5.5, 远高于阴天白天的Q10 (1.9), 说明晴天白天生态系统呼吸对温度变化的敏感性相对较高.分析其原因, 一方面生态系统温度敏感性与气温有关(Kirschbaum, 1995; Xu & Qi, 2001; Janssens & Pilegaard, 2003; Davidson & Janssens., 2006; 张雷明等, 2006; Jassal et al., 2008), 气温对Q10产生影响主要通过影响光合与呼吸作用酶的活性实现(Davidson & Janssens, 2006).晴天气温T ((23.56 ± 2.62) ℃)显著大于阴天((21.26 ± 2.34) ℃), 阴天由于酶促反应需要一定的活化酶, 光合与呼吸作用酶的活性一般受到限制; 而晴天随着辐射增强, 气温升高, 越来越多的分子达到或超过了自身的活化能, 反应加快(Xiang & Freeman, 2009; Karhu et al., 2010).另一方面, 晴天相对阴天较高的PAR使得晴天光合作用更强, 产生输送的有机质增多(Curiel Yuste et al., 2007; Jassal et al., 2008), 自养呼吸增强, 同时地上植被以及土壤异养微生物呼吸对气温的响应能力增强(Edwards et al., 2004; Heinemeyer et al., 2006; Moyano et al., 2008), 而阴天由于底物基质供应不足使得自养、异养呼吸受到抑制(Gershenson et al., 2009). ...
土壤呼吸温度敏感性的影响因素和不确定性
1
2011
... Q10值被广泛应用于评价生态系统呼吸对温度变化的敏感程度(杨庆朋等, 2011; 杨利琼等, 2013; Han et al., 2014a, 2014b), Q10值越大, 说明生态系统呼吸对温度变化的响应越敏感.黄河三角洲芦苇湿地生态系统阴天和晴天Q10值均在已有的湿地Q10范围(1.0-7.7)内(Chapman & Thurlow, 1996; Silvola et al., 1996; Lafleur et al., 2001; Bonneville et al., 2008; Zhou et al., 2009; 周丽艳等, 2010), 其中晴天白天Q10为5.5, 远高于阴天白天的Q10 (1.9), 说明晴天白天生态系统呼吸对温度变化的敏感性相对较高.分析其原因, 一方面生态系统温度敏感性与气温有关(Kirschbaum, 1995; Xu & Qi, 2001; Janssens & Pilegaard, 2003; Davidson & Janssens., 2006; 张雷明等, 2006; Jassal et al., 2008), 气温对Q10产生影响主要通过影响光合与呼吸作用酶的活性实现(Davidson & Janssens, 2006).晴天气温T ((23.56 ± 2.62) ℃)显著大于阴天((21.26 ± 2.34) ℃), 阴天由于酶促反应需要一定的活化酶, 光合与呼吸作用酶的活性一般受到限制; 而晴天随着辐射增强, 气温升高, 越来越多的分子达到或超过了自身的活化能, 反应加快(Xiang & Freeman, 2009; Karhu et al., 2010).另一方面, 晴天相对阴天较高的PAR使得晴天光合作用更强, 产生输送的有机质增多(Curiel Yuste et al., 2007; Jassal et al., 2008), 自养呼吸增强, 同时地上植被以及土壤异养微生物呼吸对气温的响应能力增强(Edwards et al., 2004; Heinemeyer et al., 2006; Moyano et al., 2008), 而阴天由于底物基质供应不足使得自养、异养呼吸受到抑制(Gershenson et al., 2009). ...
Effects of cloudiness on carbon dioxide exchange over an irrigated maize cropland in northwestern China.
3
2011
... 已有研究表明, 阴天和晴天所引起的辐射差异影响着陆地生态系统的交换过程和碳吸收能力(Roderick et al., 2001; Still et al., 2009).但由于研究方法、地域环境条件以及不同植被类型的差异, 研究结论存在较大差异(Gu et al., 1999, 2002; Alton et al., 2007; Urban et al., 2007).例如, 中国渭北刺槐(Robinia pseudoacacia)生长初期和旺盛期晴天条件下森林日均固碳量比阴天高8%-16% (郑元等, 2011).中国西北玉米(Zea mays)农田生态系统阴天条件下净生态系统CO2交换(NEE)的增长速率却高于晴天, 且生态系统总初级生产力(GPP)最大值出现于阴天(Zhang et al., 2011).对中国西北草地(Bai et al., 2012)和不同温带森林类型(Zhang et al., 2010)的研究也得出类似结论.另外, 也有研究发现加拿大常绿灌木(Letts et al., 2005)和西藏高寒草甸(范玉枝等, 2009)生态系统CO2交换不受阴天和晴天天气条件影响. ...
... 在一定光照强度范围内, 光合作用随PAR的增强而增加(Glenn et al., 2006; Syed et al., 2006; Hao et al., 2011; 同小娟等, 2011).光能过剩会对光合作用产生不良影响, 出现光抑制, 致使叶绿体光合效率下降(Jiang et al., 2011; Tholen et al., 2012).晴天PAR显著大于阴天(图2B), 因此, 晴天条件下植被叶片光合作用可能大于阴天, 表现为晴天Amax显著大于阴天(表1).Zhou等(2009)研究认为Amax与植被的环境条件有关, 其中晴天Amax较阴天条件下提高了30%, 说明在植被生长旺季, 天空有云层覆盖时芦苇的光合能力比晴朗天气无云时有所降低, Law等(2002)总结全球森林、草地、农田以及苔原生态系统碳交换相关研究也得出类似结论.阴天条件下, 伴随天空云量的增多, PAR显著降低(图2B), 尽管阴天和晴天条件下, NEE均随PAR的增大而增加(图3), 但由于阴天PAR较低, 且晴天未出现光抑制现象, 因此其NEE显著低于晴天(图2A).Alton (2008)在分析北美洲和欧洲6种森林类型2-3年的碳通量观测数据后发现, 这些森林阴天的日固碳量比晴天少60%-80%.相反, 在研究太阳辐射对黄河小浪底人工混交林生态系统碳交换的影响中发现, 多云天气下净碳交换会增加(刘佳等, 2014).造成研究结论不一致的原因是多方面的, 不同的阴天和晴天划分指标(Graham et al., 2003; Dengel & Grace, 2010)和拟合模型(Zhang et al., 2011)来估计天气对碳通量的影响, 散射光和直射光下的碳交换没有一直处在相同的天气辐射条件下进行对比.此外, 研究地域的差异和不同植被类型的特性也在很大程度上影响了碳交换过程对天气的响应过程. ...
... 本研究以黄河三角洲芦苇湿地为研究对象, 在自然条件下选择相邻的阴天和晴天, 在生物要素(生物量、叶面积指数)、土壤水分以及养分特征保持不变的前提下, 分析阴天和晴天引起的辐射变化对生态系统NEE的影响机制.目前, 有关太阳辐射对生态系统NEE的影响机制大都以晴空指数为指标(Gu et al., 2003; Liu et al., 2006; 范玉枝等, 2009; Zhang et al., 2010; 周丽艳等, 2010; 蒋琰等, 2011; Zhao et al., 2011; 刘佳等, 2014).这种研究方法的最大不足就是研究阶段中, 太阳辐射、生物要素(生物量、叶面积指数)、土壤湿度等环境因子都在发生变化, 这对阐明太阳辐射这一单因素对NEE的控制机制研究带来一定困难.而本研究巧妙地在自然条件下选择相邻的阴天和晴天, 在日尺度上实现了对单一变量PAR的控制.利用这种研究方法, Han等(2014a)通过对比分析阴天和晴天植物冠层光合作用的差异性, 定量分析了光合作用在日尺度上对土壤呼吸的调节作用.此外, 我们已有研究发现PAR改变所引起的植物光合作用变化对生态系统呼吸会产生一定的影响(Tang et al., 2005; Baldocchi et al., 2006; Moyano et al., 2008; Wingate et al., 2010; Han et al., 2012).但是, 以往的研究多通过夜间生态系统呼吸与气温函数关系外延的方法来估算白天生态系统呼吸(Doughty et al., 2010; Zhang et al., 2011), 忽略了PAR对白天生态系统呼吸的影响, 因而具有一定的局限性.考虑到白天PAR的变化, 本研究采用Michaelis-Menten模型对白天生态系统呼吸进行推算, 使得生态系统呼吸的温度响应数据更具有说服力. ...
中国东部森林样带典型生态系统碳收支的季节变化
1
2006
... Q10值被广泛应用于评价生态系统呼吸对温度变化的敏感程度(杨庆朋等, 2011; 杨利琼等, 2013; Han et al., 2014a, 2014b), Q10值越大, 说明生态系统呼吸对温度变化的响应越敏感.黄河三角洲芦苇湿地生态系统阴天和晴天Q10值均在已有的湿地Q10范围(1.0-7.7)内(Chapman & Thurlow, 1996; Silvola et al., 1996; Lafleur et al., 2001; Bonneville et al., 2008; Zhou et al., 2009; 周丽艳等, 2010), 其中晴天白天Q10为5.5, 远高于阴天白天的Q10 (1.9), 说明晴天白天生态系统呼吸对温度变化的敏感性相对较高.分析其原因, 一方面生态系统温度敏感性与气温有关(Kirschbaum, 1995; Xu & Qi, 2001; Janssens & Pilegaard, 2003; Davidson & Janssens., 2006; 张雷明等, 2006; Jassal et al., 2008), 气温对Q10产生影响主要通过影响光合与呼吸作用酶的活性实现(Davidson & Janssens, 2006).晴天气温T ((23.56 ± 2.62) ℃)显著大于阴天((21.26 ± 2.34) ℃), 阴天由于酶促反应需要一定的活化酶, 光合与呼吸作用酶的活性一般受到限制; 而晴天随着辐射增强, 气温升高, 越来越多的分子达到或超过了自身的活化能, 反应加快(Xiang & Freeman, 2009; Karhu et al., 2010).另一方面, 晴天相对阴天较高的PAR使得晴天光合作用更强, 产生输送的有机质增多(Curiel Yuste et al., 2007; Jassal et al., 2008), 自养呼吸增强, 同时地上植被以及土壤异养微生物呼吸对气温的响应能力增强(Edwards et al., 2004; Heinemeyer et al., 2006; Moyano et al., 2008), 而阴天由于底物基质供应不足使得自养、异养呼吸受到抑制(Gershenson et al., 2009). ...
中国东部森林样带典型生态系统碳收支的季节变化
1
2006
... Q10值被广泛应用于评价生态系统呼吸对温度变化的敏感程度(杨庆朋等, 2011; 杨利琼等, 2013; Han et al., 2014a, 2014b), Q10值越大, 说明生态系统呼吸对温度变化的响应越敏感.黄河三角洲芦苇湿地生态系统阴天和晴天Q10值均在已有的湿地Q10范围(1.0-7.7)内(Chapman & Thurlow, 1996; Silvola et al., 1996; Lafleur et al., 2001; Bonneville et al., 2008; Zhou et al., 2009; 周丽艳等, 2010), 其中晴天白天Q10为5.5, 远高于阴天白天的Q10 (1.9), 说明晴天白天生态系统呼吸对温度变化的敏感性相对较高.分析其原因, 一方面生态系统温度敏感性与气温有关(Kirschbaum, 1995; Xu & Qi, 2001; Janssens & Pilegaard, 2003; Davidson & Janssens., 2006; 张雷明等, 2006; Jassal et al., 2008), 气温对Q10产生影响主要通过影响光合与呼吸作用酶的活性实现(Davidson & Janssens, 2006).晴天气温T ((23.56 ± 2.62) ℃)显著大于阴天((21.26 ± 2.34) ℃), 阴天由于酶促反应需要一定的活化酶, 光合与呼吸作用酶的活性一般受到限制; 而晴天随着辐射增强, 气温升高, 越来越多的分子达到或超过了自身的活化能, 反应加快(Xiang & Freeman, 2009; Karhu et al., 2010).另一方面, 晴天相对阴天较高的PAR使得晴天光合作用更强, 产生输送的有机质增多(Curiel Yuste et al., 2007; Jassal et al., 2008), 自养呼吸增强, 同时地上植被以及土壤异养微生物呼吸对气温的响应能力增强(Edwards et al., 2004; Heinemeyer et al., 2006; Moyano et al., 2008), 而阴天由于底物基质供应不足使得自养、异养呼吸受到抑制(Gershenson et al., 2009). ...
太阳辐射对长白山阔叶红松林净生态系统碳交换的影响
1
2009
... 本文研究发现晴天和阴天芦苇湿地NEE对PAR的响应趋势一致, 均呈直角双曲线关系, 与先前的很多研究结果一致(Falge et al., 2001; Monson et al., 2002; Zhou et al., 2009; 张弥等, 2009; Han et al., 2014a, 2014b; 邢庆会等, 2014).PAR是控制阴天和晴天生态系统生长季昼间NEE变化的主要因素(Flanagan & Johnson, 2005; Liu et al., 2006; 薛红喜等, 2012), Michaelis-Menten方程拟合发现, PAR可以解释晴天单天NEE变化的57%-82%, 解释阴天单天NEE变化的53%-81%.云层变化会改变地表接收太阳辐射的强度, 影响PAR、T等环境因子, 进而影响阴天和晴天条件下NEE对光的响应特征(Baldocchi, 1997; Freedman et al., 2001). ...
太阳辐射对长白山阔叶红松林净生态系统碳交换的影响
1
2009
... 本文研究发现晴天和阴天芦苇湿地NEE对PAR的响应趋势一致, 均呈直角双曲线关系, 与先前的很多研究结果一致(Falge et al., 2001; Monson et al., 2002; Zhou et al., 2009; 张弥等, 2009; Han et al., 2014a, 2014b; 邢庆会等, 2014).PAR是控制阴天和晴天生态系统生长季昼间NEE变化的主要因素(Flanagan & Johnson, 2005; Liu et al., 2006; 薛红喜等, 2012), Michaelis-Menten方程拟合发现, PAR可以解释晴天单天NEE变化的57%-82%, 解释阴天单天NEE变化的53%-81%.云层变化会改变地表接收太阳辐射的强度, 影响PAR、T等环境因子, 进而影响阴天和晴天条件下NEE对光的响应特征(Baldocchi, 1997; Freedman et al., 2001). ...
Impact of cloudiness on net ecosystem exchange of carbon dioxide in different types of forest ecosystems in China.
3
2010
... 已有研究表明, 阴天和晴天所引起的辐射差异影响着陆地生态系统的交换过程和碳吸收能力(Roderick et al., 2001; Still et al., 2009).但由于研究方法、地域环境条件以及不同植被类型的差异, 研究结论存在较大差异(Gu et al., 1999, 2002; Alton et al., 2007; Urban et al., 2007).例如, 中国渭北刺槐(Robinia pseudoacacia)生长初期和旺盛期晴天条件下森林日均固碳量比阴天高8%-16% (郑元等, 2011).中国西北玉米(Zea mays)农田生态系统阴天条件下净生态系统CO2交换(NEE)的增长速率却高于晴天, 且生态系统总初级生产力(GPP)最大值出现于阴天(Zhang et al., 2011).对中国西北草地(Bai et al., 2012)和不同温带森林类型(Zhang et al., 2010)的研究也得出类似结论.另外, 也有研究发现加拿大常绿灌木(Letts et al., 2005)和西藏高寒草甸(范玉枝等, 2009)生态系统CO2交换不受阴天和晴天天气条件影响. ...
... 黄河口近海与海岸湿地是暖温带增长速度最快和最具代表性的新生近海与海岸湿地, 具有海陆过渡性、原生性和脆弱性的特点(布仁仓等, 1999; 崔树强, 2002).受水沙量变化、陆地和河流淡水径流、咸水海流多重影响以及人类活动的干扰, 黄河三角洲湿地生态系统CO2交换存在着极大的复杂性和不确定性(Sasaki et al., 2009;Han et al., 2014a, 2014b; 邢庆会等, 2014).研究表明黄河三角洲芦苇湿地生长季NEE占全年的80%以上, 且碳吸收集中在植物生长旺盛期(7-9月)(杨利琼等, 2013; Han et al., 2014a, 2014b).而这一时期正值黄河三角洲雨季, 其云雨状况影响太阳辐射, 进而影响湿地生态系统的碳吸收功能.在以往的研究中, 温度、水分、太阳辐射等环境因子, 生物量、叶面积等生物因子以及地下水位、地表水深等水文要素对湿地生态系统NEE的影响机制受到研究者的极大关注(牟长城等, 2009; Zhao et al., 2010; Han et al., 2013; 邢庆会等, 2014).目前, 有关阴天和晴天条件对生态系统碳交换影响的研究主要集中在森林生态系统(Liu et al., 2006; Zhang et al., 2010; 周丽艳等, 2010; Jiang et al., 2011; Zhao et al., 2011; 刘佳等, 2014)和草原生态系统(Gu et al., 2003; 范玉枝等, 2009).阴天和晴天状况引起的太阳辐射变化对湿地生态系统NEE和碳汇功能究竟产生怎样的影响目前尚不清楚. ...
... 本研究以黄河三角洲芦苇湿地为研究对象, 在自然条件下选择相邻的阴天和晴天, 在生物要素(生物量、叶面积指数)、土壤水分以及养分特征保持不变的前提下, 分析阴天和晴天引起的辐射变化对生态系统NEE的影响机制.目前, 有关太阳辐射对生态系统NEE的影响机制大都以晴空指数为指标(Gu et al., 2003; Liu et al., 2006; 范玉枝等, 2009; Zhang et al., 2010; 周丽艳等, 2010; 蒋琰等, 2011; Zhao et al., 2011; 刘佳等, 2014).这种研究方法的最大不足就是研究阶段中, 太阳辐射、生物要素(生物量、叶面积指数)、土壤湿度等环境因子都在发生变化, 这对阐明太阳辐射这一单因素对NEE的控制机制研究带来一定困难.而本研究巧妙地在自然条件下选择相邻的阴天和晴天, 在日尺度上实现了对单一变量PAR的控制.利用这种研究方法, Han等(2014a)通过对比分析阴天和晴天植物冠层光合作用的差异性, 定量分析了光合作用在日尺度上对土壤呼吸的调节作用.此外, 我们已有研究发现PAR改变所引起的植物光合作用变化对生态系统呼吸会产生一定的影响(Tang et al., 2005; Baldocchi et al., 2006; Moyano et al., 2008; Wingate et al., 2010; Han et al., 2012).但是, 以往的研究多通过夜间生态系统呼吸与气温函数关系外延的方法来估算白天生态系统呼吸(Doughty et al., 2010; Zhang et al., 2011), 忽略了PAR对白天生态系统呼吸的影响, 因而具有一定的局限性.考虑到白天PAR的变化, 本研究采用Michaelis-Menten模型对白天生态系统呼吸进行推算, 使得生态系统呼吸的温度响应数据更具有说服力. ...
Seasonal variations in carbon dioxide exchange in an alpine wetland meadow on the Qinghai-Tibetan Plateau.
2
2010
... 黄河口近海与海岸湿地是暖温带增长速度最快和最具代表性的新生近海与海岸湿地, 具有海陆过渡性、原生性和脆弱性的特点(布仁仓等, 1999; 崔树强, 2002).受水沙量变化、陆地和河流淡水径流、咸水海流多重影响以及人类活动的干扰, 黄河三角洲湿地生态系统CO2交换存在着极大的复杂性和不确定性(Sasaki et al., 2009;Han et al., 2014a, 2014b; 邢庆会等, 2014).研究表明黄河三角洲芦苇湿地生长季NEE占全年的80%以上, 且碳吸收集中在植物生长旺盛期(7-9月)(杨利琼等, 2013; Han et al., 2014a, 2014b).而这一时期正值黄河三角洲雨季, 其云雨状况影响太阳辐射, 进而影响湿地生态系统的碳吸收功能.在以往的研究中, 温度、水分、太阳辐射等环境因子, 生物量、叶面积等生物因子以及地下水位、地表水深等水文要素对湿地生态系统NEE的影响机制受到研究者的极大关注(牟长城等, 2009; Zhao et al., 2010; Han et al., 2013; 邢庆会等, 2014).目前, 有关阴天和晴天条件对生态系统碳交换影响的研究主要集中在森林生态系统(Liu et al., 2006; Zhang et al., 2010; 周丽艳等, 2010; Jiang et al., 2011; Zhao et al., 2011; 刘佳等, 2014)和草原生态系统(Gu et al., 2003; 范玉枝等, 2009).阴天和晴天状况引起的太阳辐射变化对湿地生态系统NEE和碳汇功能究竟产生怎样的影响目前尚不清楚. ...
... 阴天和晴天生态系统Reco,daytime与T均呈显著的正相关关系, 两者呈指数关系, 与以往很多研究的结果一致(石培礼等, 2006; Zhou et al., 2009; Zhao et al., 2010; Han et al., 2013, 2014a; 邢庆会等, 2014).T是影响生态系统呼吸的主要因素(Law et al., 2002; Reichstein et al., 2005; Urban et al., 2007), 本文研究发现: 黄河三角洲芦苇湿地晴天气温相对阴天提高了10%, 其阴天和晴天的气温差高于美国阿尔卑斯山脉南部森林的阴天和晴天的气温差(Berry & Smith, 2012), 黄河三角洲芦苇湿地晴天对应的生态系统呼吸相对阴天提高了20% (p < 0.05), 说明阴天条件下, 植被地上部分尤其是叶面温度较晴朗天气强太阳辐射条件下的低, 叶片以及植被茎干呼吸受到一定的抑制(Urban et al., 2007). ...
Characteristics of CO2 flux in a Chinese fir plantation ecosystem in Huitong County, Hunan Province.
2
2011
... 黄河口近海与海岸湿地是暖温带增长速度最快和最具代表性的新生近海与海岸湿地, 具有海陆过渡性、原生性和脆弱性的特点(布仁仓等, 1999; 崔树强, 2002).受水沙量变化、陆地和河流淡水径流、咸水海流多重影响以及人类活动的干扰, 黄河三角洲湿地生态系统CO2交换存在着极大的复杂性和不确定性(Sasaki et al., 2009;Han et al., 2014a, 2014b; 邢庆会等, 2014).研究表明黄河三角洲芦苇湿地生长季NEE占全年的80%以上, 且碳吸收集中在植物生长旺盛期(7-9月)(杨利琼等, 2013; Han et al., 2014a, 2014b).而这一时期正值黄河三角洲雨季, 其云雨状况影响太阳辐射, 进而影响湿地生态系统的碳吸收功能.在以往的研究中, 温度、水分、太阳辐射等环境因子, 生物量、叶面积等生物因子以及地下水位、地表水深等水文要素对湿地生态系统NEE的影响机制受到研究者的极大关注(牟长城等, 2009; Zhao et al., 2010; Han et al., 2013; 邢庆会等, 2014).目前, 有关阴天和晴天条件对生态系统碳交换影响的研究主要集中在森林生态系统(Liu et al., 2006; Zhang et al., 2010; 周丽艳等, 2010; Jiang et al., 2011; Zhao et al., 2011; 刘佳等, 2014)和草原生态系统(Gu et al., 2003; 范玉枝等, 2009).阴天和晴天状况引起的太阳辐射变化对湿地生态系统NEE和碳汇功能究竟产生怎样的影响目前尚不清楚. ...
... 本研究以黄河三角洲芦苇湿地为研究对象, 在自然条件下选择相邻的阴天和晴天, 在生物要素(生物量、叶面积指数)、土壤水分以及养分特征保持不变的前提下, 分析阴天和晴天引起的辐射变化对生态系统NEE的影响机制.目前, 有关太阳辐射对生态系统NEE的影响机制大都以晴空指数为指标(Gu et al., 2003; Liu et al., 2006; 范玉枝等, 2009; Zhang et al., 2010; 周丽艳等, 2010; 蒋琰等, 2011; Zhao et al., 2011; 刘佳等, 2014).这种研究方法的最大不足就是研究阶段中, 太阳辐射、生物要素(生物量、叶面积指数)、土壤湿度等环境因子都在发生变化, 这对阐明太阳辐射这一单因素对NEE的控制机制研究带来一定困难.而本研究巧妙地在自然条件下选择相邻的阴天和晴天, 在日尺度上实现了对单一变量PAR的控制.利用这种研究方法, Han等(2014a)通过对比分析阴天和晴天植物冠层光合作用的差异性, 定量分析了光合作用在日尺度上对土壤呼吸的调节作用.此外, 我们已有研究发现PAR改变所引起的植物光合作用变化对生态系统呼吸会产生一定的影响(Tang et al., 2005; Baldocchi et al., 2006; Moyano et al., 2008; Wingate et al., 2010; Han et al., 2012).但是, 以往的研究多通过夜间生态系统呼吸与气温函数关系外延的方法来估算白天生态系统呼吸(Doughty et al., 2010; Zhang et al., 2011), 忽略了PAR对白天生态系统呼吸的影响, 因而具有一定的局限性.考虑到白天PAR的变化, 本研究采用Michaelis-Menten模型对白天生态系统呼吸进行推算, 使得生态系统呼吸的温度响应数据更具有说服力. ...
晴天和阴天对刺槐光合生理特性的影响
1
2011
... 已有研究表明, 阴天和晴天所引起的辐射差异影响着陆地生态系统的交换过程和碳吸收能力(Roderick et al., 2001; Still et al., 2009).但由于研究方法、地域环境条件以及不同植被类型的差异, 研究结论存在较大差异(Gu et al., 1999, 2002; Alton et al., 2007; Urban et al., 2007).例如, 中国渭北刺槐(Robinia pseudoacacia)生长初期和旺盛期晴天条件下森林日均固碳量比阴天高8%-16% (郑元等, 2011).中国西北玉米(Zea mays)农田生态系统阴天条件下净生态系统CO2交换(NEE)的增长速率却高于晴天, 且生态系统总初级生产力(GPP)最大值出现于阴天(Zhang et al., 2011).对中国西北草地(Bai et al., 2012)和不同温带森林类型(Zhang et al., 2010)的研究也得出类似结论.另外, 也有研究发现加拿大常绿灌木(Letts et al., 2005)和西藏高寒草甸(范玉枝等, 2009)生态系统CO2交换不受阴天和晴天天气条件影响. ...
晴天和阴天对刺槐光合生理特性的影响
1
2011
... 已有研究表明, 阴天和晴天所引起的辐射差异影响着陆地生态系统的交换过程和碳吸收能力(Roderick et al., 2001; Still et al., 2009).但由于研究方法、地域环境条件以及不同植被类型的差异, 研究结论存在较大差异(Gu et al., 1999, 2002; Alton et al., 2007; Urban et al., 2007).例如, 中国渭北刺槐(Robinia pseudoacacia)生长初期和旺盛期晴天条件下森林日均固碳量比阴天高8%-16% (郑元等, 2011).中国西北玉米(Zea mays)农田生态系统阴天条件下净生态系统CO2交换(NEE)的增长速率却高于晴天, 且生态系统总初级生产力(GPP)最大值出现于阴天(Zhang et al., 2011).对中国西北草地(Bai et al., 2012)和不同温带森林类型(Zhang et al., 2010)的研究也得出类似结论.另外, 也有研究发现加拿大常绿灌木(Letts et al., 2005)和西藏高寒草甸(范玉枝等, 2009)生态系统CO2交换不受阴天和晴天天气条件影响. ...
Annual cycle of CO2 exchange over a reed (Phragmites australis) wetland in Northeast China.
3
2009
... 本文研究发现晴天和阴天芦苇湿地NEE对PAR的响应趋势一致, 均呈直角双曲线关系, 与先前的很多研究结果一致(Falge et al., 2001; Monson et al., 2002; Zhou et al., 2009; 张弥等, 2009; Han et al., 2014a, 2014b; 邢庆会等, 2014).PAR是控制阴天和晴天生态系统生长季昼间NEE变化的主要因素(Flanagan & Johnson, 2005; Liu et al., 2006; 薛红喜等, 2012), Michaelis-Menten方程拟合发现, PAR可以解释晴天单天NEE变化的57%-82%, 解释阴天单天NEE变化的53%-81%.云层变化会改变地表接收太阳辐射的强度, 影响PAR、T等环境因子, 进而影响阴天和晴天条件下NEE对光的响应特征(Baldocchi, 1997; Freedman et al., 2001). ...
... 阴天和晴天生态系统Reco,daytime与T均呈显著的正相关关系, 两者呈指数关系, 与以往很多研究的结果一致(石培礼等, 2006; Zhou et al., 2009; Zhao et al., 2010; Han et al., 2013, 2014a; 邢庆会等, 2014).T是影响生态系统呼吸的主要因素(Law et al., 2002; Reichstein et al., 2005; Urban et al., 2007), 本文研究发现: 黄河三角洲芦苇湿地晴天气温相对阴天提高了10%, 其阴天和晴天的气温差高于美国阿尔卑斯山脉南部森林的阴天和晴天的气温差(Berry & Smith, 2012), 黄河三角洲芦苇湿地晴天对应的生态系统呼吸相对阴天提高了20% (p < 0.05), 说明阴天条件下, 植被地上部分尤其是叶面温度较晴朗天气强太阳辐射条件下的低, 叶片以及植被茎干呼吸受到一定的抑制(Urban et al., 2007). ...
... Q10值被广泛应用于评价生态系统呼吸对温度变化的敏感程度(杨庆朋等, 2011; 杨利琼等, 2013; Han et al., 2014a, 2014b), Q10值越大, 说明生态系统呼吸对温度变化的响应越敏感.黄河三角洲芦苇湿地生态系统阴天和晴天Q10值均在已有的湿地Q10范围(1.0-7.7)内(Chapman & Thurlow, 1996; Silvola et al., 1996; Lafleur et al., 2001; Bonneville et al., 2008; Zhou et al., 2009; 周丽艳等, 2010), 其中晴天白天Q10为5.5, 远高于阴天白天的Q10 (1.9), 说明晴天白天生态系统呼吸对温度变化的敏感性相对较高.分析其原因, 一方面生态系统温度敏感性与气温有关(Kirschbaum, 1995; Xu & Qi, 2001; Janssens & Pilegaard, 2003; Davidson & Janssens., 2006; 张雷明等, 2006; Jassal et al., 2008), 气温对Q10产生影响主要通过影响光合与呼吸作用酶的活性实现(Davidson & Janssens, 2006).晴天气温T ((23.56 ± 2.62) ℃)显著大于阴天((21.26 ± 2.34) ℃), 阴天由于酶促反应需要一定的活化酶, 光合与呼吸作用酶的活性一般受到限制; 而晴天随着辐射增强, 气温升高, 越来越多的分子达到或超过了自身的活化能, 反应加快(Xiang & Freeman, 2009; Karhu et al., 2010).另一方面, 晴天相对阴天较高的PAR使得晴天光合作用更强, 产生输送的有机质增多(Curiel Yuste et al., 2007; Jassal et al., 2008), 自养呼吸增强, 同时地上植被以及土壤异养微生物呼吸对气温的响应能力增强(Edwards et al., 2004; Heinemeyer et al., 2006; Moyano et al., 2008), 而阴天由于底物基质供应不足使得自养、异养呼吸受到抑制(Gershenson et al., 2009). ...
原始兴安落叶松林生长季净生态系统CO2交换及其光响应特征
3
2010
... 黄河口近海与海岸湿地是暖温带增长速度最快和最具代表性的新生近海与海岸湿地, 具有海陆过渡性、原生性和脆弱性的特点(布仁仓等, 1999; 崔树强, 2002).受水沙量变化、陆地和河流淡水径流、咸水海流多重影响以及人类活动的干扰, 黄河三角洲湿地生态系统CO2交换存在着极大的复杂性和不确定性(Sasaki et al., 2009;Han et al., 2014a, 2014b; 邢庆会等, 2014).研究表明黄河三角洲芦苇湿地生长季NEE占全年的80%以上, 且碳吸收集中在植物生长旺盛期(7-9月)(杨利琼等, 2013; Han et al., 2014a, 2014b).而这一时期正值黄河三角洲雨季, 其云雨状况影响太阳辐射, 进而影响湿地生态系统的碳吸收功能.在以往的研究中, 温度、水分、太阳辐射等环境因子, 生物量、叶面积等生物因子以及地下水位、地表水深等水文要素对湿地生态系统NEE的影响机制受到研究者的极大关注(牟长城等, 2009; Zhao et al., 2010; Han et al., 2013; 邢庆会等, 2014).目前, 有关阴天和晴天条件对生态系统碳交换影响的研究主要集中在森林生态系统(Liu et al., 2006; Zhang et al., 2010; 周丽艳等, 2010; Jiang et al., 2011; Zhao et al., 2011; 刘佳等, 2014)和草原生态系统(Gu et al., 2003; 范玉枝等, 2009).阴天和晴天状况引起的太阳辐射变化对湿地生态系统NEE和碳汇功能究竟产生怎样的影响目前尚不清楚. ...
... Q10值被广泛应用于评价生态系统呼吸对温度变化的敏感程度(杨庆朋等, 2011; 杨利琼等, 2013; Han et al., 2014a, 2014b), Q10值越大, 说明生态系统呼吸对温度变化的响应越敏感.黄河三角洲芦苇湿地生态系统阴天和晴天Q10值均在已有的湿地Q10范围(1.0-7.7)内(Chapman & Thurlow, 1996; Silvola et al., 1996; Lafleur et al., 2001; Bonneville et al., 2008; Zhou et al., 2009; 周丽艳等, 2010), 其中晴天白天Q10为5.5, 远高于阴天白天的Q10 (1.9), 说明晴天白天生态系统呼吸对温度变化的敏感性相对较高.分析其原因, 一方面生态系统温度敏感性与气温有关(Kirschbaum, 1995; Xu & Qi, 2001; Janssens & Pilegaard, 2003; Davidson & Janssens., 2006; 张雷明等, 2006; Jassal et al., 2008), 气温对Q10产生影响主要通过影响光合与呼吸作用酶的活性实现(Davidson & Janssens, 2006).晴天气温T ((23.56 ± 2.62) ℃)显著大于阴天((21.26 ± 2.34) ℃), 阴天由于酶促反应需要一定的活化酶, 光合与呼吸作用酶的活性一般受到限制; 而晴天随着辐射增强, 气温升高, 越来越多的分子达到或超过了自身的活化能, 反应加快(Xiang & Freeman, 2009; Karhu et al., 2010).另一方面, 晴天相对阴天较高的PAR使得晴天光合作用更强, 产生输送的有机质增多(Curiel Yuste et al., 2007; Jassal et al., 2008), 自养呼吸增强, 同时地上植被以及土壤异养微生物呼吸对气温的响应能力增强(Edwards et al., 2004; Heinemeyer et al., 2006; Moyano et al., 2008), 而阴天由于底物基质供应不足使得自养、异养呼吸受到抑制(Gershenson et al., 2009). ...
... 本研究以黄河三角洲芦苇湿地为研究对象, 在自然条件下选择相邻的阴天和晴天, 在生物要素(生物量、叶面积指数)、土壤水分以及养分特征保持不变的前提下, 分析阴天和晴天引起的辐射变化对生态系统NEE的影响机制.目前, 有关太阳辐射对生态系统NEE的影响机制大都以晴空指数为指标(Gu et al., 2003; Liu et al., 2006; 范玉枝等, 2009; Zhang et al., 2010; 周丽艳等, 2010; 蒋琰等, 2011; Zhao et al., 2011; 刘佳等, 2014).这种研究方法的最大不足就是研究阶段中, 太阳辐射、生物要素(生物量、叶面积指数)、土壤湿度等环境因子都在发生变化, 这对阐明太阳辐射这一单因素对NEE的控制机制研究带来一定困难.而本研究巧妙地在自然条件下选择相邻的阴天和晴天, 在日尺度上实现了对单一变量PAR的控制.利用这种研究方法, Han等(2014a)通过对比分析阴天和晴天植物冠层光合作用的差异性, 定量分析了光合作用在日尺度上对土壤呼吸的调节作用.此外, 我们已有研究发现PAR改变所引起的植物光合作用变化对生态系统呼吸会产生一定的影响(Tang et al., 2005; Baldocchi et al., 2006; Moyano et al., 2008; Wingate et al., 2010; Han et al., 2012).但是, 以往的研究多通过夜间生态系统呼吸与气温函数关系外延的方法来估算白天生态系统呼吸(Doughty et al., 2010; Zhang et al., 2011), 忽略了PAR对白天生态系统呼吸的影响, 因而具有一定的局限性.考虑到白天PAR的变化, 本研究采用Michaelis-Menten模型对白天生态系统呼吸进行推算, 使得生态系统呼吸的温度响应数据更具有说服力. ...
原始兴安落叶松林生长季净生态系统CO2交换及其光响应特征
3
2010
... 黄河口近海与海岸湿地是暖温带增长速度最快和最具代表性的新生近海与海岸湿地, 具有海陆过渡性、原生性和脆弱性的特点(布仁仓等, 1999; 崔树强, 2002).受水沙量变化、陆地和河流淡水径流、咸水海流多重影响以及人类活动的干扰, 黄河三角洲湿地生态系统CO2交换存在着极大的复杂性和不确定性(Sasaki et al., 2009;Han et al., 2014a, 2014b; 邢庆会等, 2014).研究表明黄河三角洲芦苇湿地生长季NEE占全年的80%以上, 且碳吸收集中在植物生长旺盛期(7-9月)(杨利琼等, 2013; Han et al., 2014a, 2014b).而这一时期正值黄河三角洲雨季, 其云雨状况影响太阳辐射, 进而影响湿地生态系统的碳吸收功能.在以往的研究中, 温度、水分、太阳辐射等环境因子, 生物量、叶面积等生物因子以及地下水位、地表水深等水文要素对湿地生态系统NEE的影响机制受到研究者的极大关注(牟长城等, 2009; Zhao et al., 2010; Han et al., 2013; 邢庆会等, 2014).目前, 有关阴天和晴天条件对生态系统碳交换影响的研究主要集中在森林生态系统(Liu et al., 2006; Zhang et al., 2010; 周丽艳等, 2010; Jiang et al., 2011; Zhao et al., 2011; 刘佳等, 2014)和草原生态系统(Gu et al., 2003; 范玉枝等, 2009).阴天和晴天状况引起的太阳辐射变化对湿地生态系统NEE和碳汇功能究竟产生怎样的影响目前尚不清楚. ...
... Q10值被广泛应用于评价生态系统呼吸对温度变化的敏感程度(杨庆朋等, 2011; 杨利琼等, 2013; Han et al., 2014a, 2014b), Q10值越大, 说明生态系统呼吸对温度变化的响应越敏感.黄河三角洲芦苇湿地生态系统阴天和晴天Q10值均在已有的湿地Q10范围(1.0-7.7)内(Chapman & Thurlow, 1996; Silvola et al., 1996; Lafleur et al., 2001; Bonneville et al., 2008; Zhou et al., 2009; 周丽艳等, 2010), 其中晴天白天Q10为5.5, 远高于阴天白天的Q10 (1.9), 说明晴天白天生态系统呼吸对温度变化的敏感性相对较高.分析其原因, 一方面生态系统温度敏感性与气温有关(Kirschbaum, 1995; Xu & Qi, 2001; Janssens & Pilegaard, 2003; Davidson & Janssens., 2006; 张雷明等, 2006; Jassal et al., 2008), 气温对Q10产生影响主要通过影响光合与呼吸作用酶的活性实现(Davidson & Janssens, 2006).晴天气温T ((23.56 ± 2.62) ℃)显著大于阴天((21.26 ± 2.34) ℃), 阴天由于酶促反应需要一定的活化酶, 光合与呼吸作用酶的活性一般受到限制; 而晴天随着辐射增强, 气温升高, 越来越多的分子达到或超过了自身的活化能, 反应加快(Xiang & Freeman, 2009; Karhu et al., 2010).另一方面, 晴天相对阴天较高的PAR使得晴天光合作用更强, 产生输送的有机质增多(Curiel Yuste et al., 2007; Jassal et al., 2008), 自养呼吸增强, 同时地上植被以及土壤异养微生物呼吸对气温的响应能力增强(Edwards et al., 2004; Heinemeyer et al., 2006; Moyano et al., 2008), 而阴天由于底物基质供应不足使得自养、异养呼吸受到抑制(Gershenson et al., 2009). ...
... 本研究以黄河三角洲芦苇湿地为研究对象, 在自然条件下选择相邻的阴天和晴天, 在生物要素(生物量、叶面积指数)、土壤水分以及养分特征保持不变的前提下, 分析阴天和晴天引起的辐射变化对生态系统NEE的影响机制.目前, 有关太阳辐射对生态系统NEE的影响机制大都以晴空指数为指标(Gu et al., 2003; Liu et al., 2006; 范玉枝等, 2009; Zhang et al., 2010; 周丽艳等, 2010; 蒋琰等, 2011; Zhao et al., 2011; 刘佳等, 2014).这种研究方法的最大不足就是研究阶段中, 太阳辐射、生物要素(生物量、叶面积指数)、土壤湿度等环境因子都在发生变化, 这对阐明太阳辐射这一单因素对NEE的控制机制研究带来一定困难.而本研究巧妙地在自然条件下选择相邻的阴天和晴天, 在日尺度上实现了对单一变量PAR的控制.利用这种研究方法, Han等(2014a)通过对比分析阴天和晴天植物冠层光合作用的差异性, 定量分析了光合作用在日尺度上对土壤呼吸的调节作用.此外, 我们已有研究发现PAR改变所引起的植物光合作用变化对生态系统呼吸会产生一定的影响(Tang et al., 2005; Baldocchi et al., 2006; Moyano et al., 2008; Wingate et al., 2010; Han et al., 2012).但是, 以往的研究多通过夜间生态系统呼吸与气温函数关系外延的方法来估算白天生态系统呼吸(Doughty et al., 2010; Zhang et al., 2011), 忽略了PAR对白天生态系统呼吸的影响, 因而具有一定的局限性.考虑到白天PAR的变化, 本研究采用Michaelis-Menten模型对白天生态系统呼吸进行推算, 使得生态系统呼吸的温度响应数据更具有说服力. ...
 , 邢庆会
, 邢庆会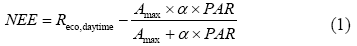



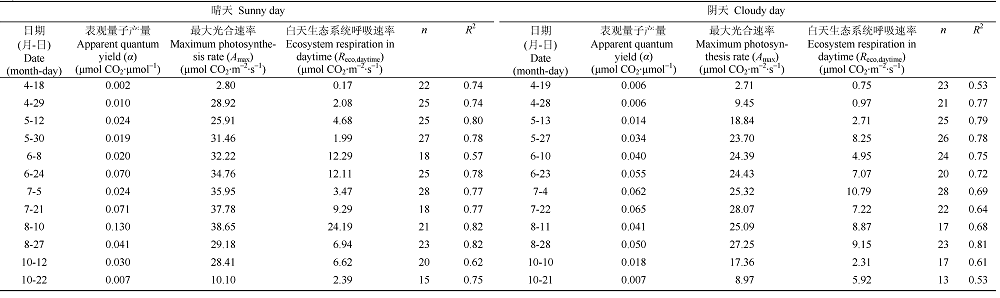


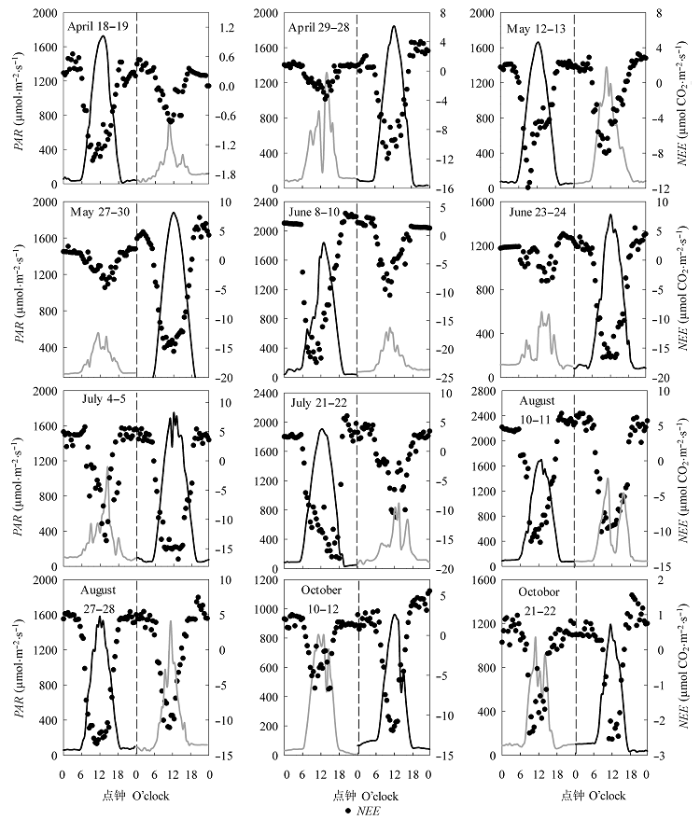
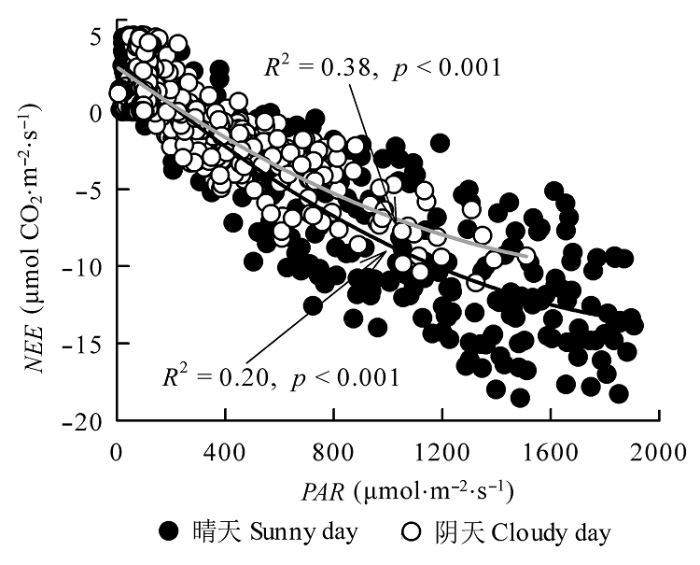
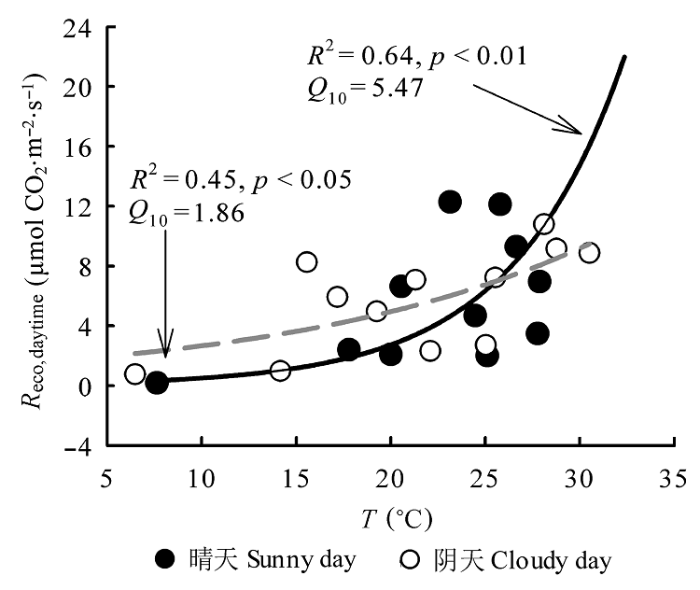
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}