Effects of different sources of dissolved organic matter on soil CO2 emission in subtropical forests
WAN Jing-Juan, GUO Jian-Fen*, , JI Shu-Rong, REN Wei-Ling, SI You-Tao, YANG Yu-Sheng
State Key Laboratory Breeding Base of Humid Subtropical Mountain Ecology, College of Geographical Sciences, Fujian Normal University, Fuzhou, 350007, China
Aims Dissolved organic matter (DOM) is an important carbon and nutrient pool, but the effects of different sources of DOM on soil carbon cycling are less well understood. Our objective in this study was to investigate how differences in the quantity and quality of DOM from fresh leaves and leaf litter of Cunninghamia lanceolata and Castanopsis carlesii affected soil CO2 fluxes in a laboratory incubation experiment. Methods Mineral soils (0-10 cm) from an 11-year-old Cunninghamia lanceolata plantation in Sanming of Fujian Province, China, were incubated for 59 days after adding the DOM from fresh leaves and leaf litter of Cunninghamia lanceolata and Castanopsis carlesii. Carbon (C) mineralization during incubation was determined using CO2 respiration method. Important findings Compared to the controls, the rates of C mineralization significantly increased by 91.5%, 12.8%, 61.0% and 113.3% on day 1, following additions of DOM from fresh leaves and leaf litter of Cunninghamia lanceolata and Castanopsis carlesii, respectively; the magnitudes of the increases declined to 24.1%, 8.3%, 14.6% and 13.2% by day 5, indicating that addition of DOM had significant but short-term influences on soil CO2 emission. DOM from different sources had significant effects on the cumulative CO2 production following addition of DOM by day 31 (p < 0.05). After 59 days of incubation, the cumulative quantity of mineralized C following addition of DOM from fresh leaves and leaf litter of Cunninghamia lanceolata was significantly greater than that from those of Castanopsis carlesii, while there was no significant difference in the cumulative CO2 production between DOM from fresh leaves and leaf litter of the same tree species, suggesting that difference in tree species had a greater influence on C mineralization than difference in the degree of leaf decay. Addition of DOM originated from fresh leaves and leaf litter of Castanopsis carlesii resulted in increased C mineralization by 22.5% and 50.0% of C added over the course of 59 day incubation, whereas increases by additions of DOM from fresh leaves and leaf litter of Cunninghamia lanceolata were 1.76 times and 2.56 times, respectively. Thus, a single addition of different sources of DOM may lead to diverse effects on total soil carbon stocks.
WANJing-Juan, GUOJian-Fen, JIShu-Rong, RENWei-Ling, SIYou-Tao, YANGYu-Sheng. Effects of different sources of dissolved organic matter on soil CO2 emission in subtropical forests[J]. Chinese Journal of Plant Ecology, 2015, 39(7): 674-681 https://doi.org/10.17521/cjpe.2015.0064
Biodegradation and chemical characteristics of hot-water extractable organic matter from soils under four different vegetation types in the Wuyi Mountains, southeastern China.
Dynamics of the water extractable organic carbon pool during mineralisation in soils from a Douglas fir plantation and an oak-beech forest―An incubation experiment.
Increased litter build up and soil organic matter stabilization in a poplar plantation after 6 years of atmospheric CO2 enrichment (FACE): Final results of POP-Euro FACE compared to other forest FACE experiments.
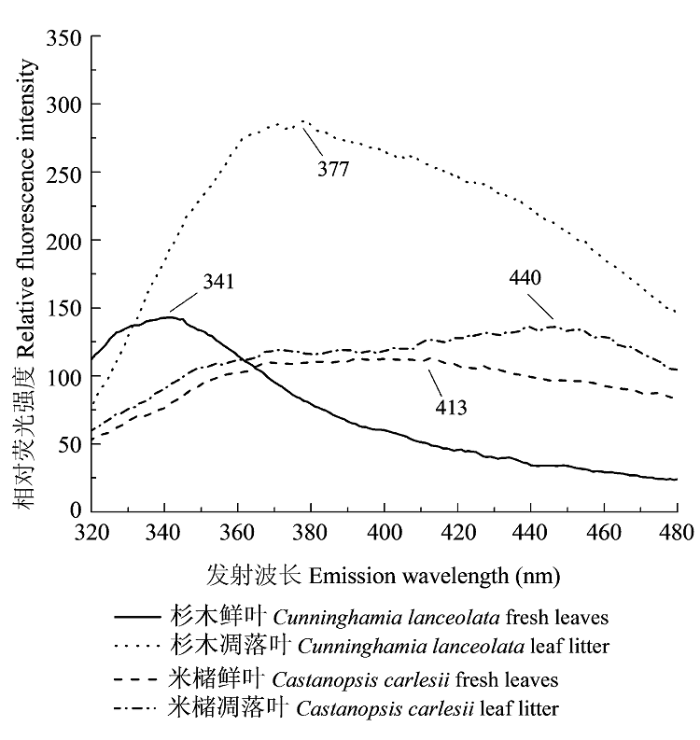
Quantities and spectral characteristics of DOM released from leaf and litterfall in Castanopsis carlesii forest and Cunninghamia lanceolata plantation.
Lack of correlation between turnover of low-molecular-weight dissolved organic carbon and differences in microbial community composition or growth across a soil pH gradient.
Applied and Environmental Microbiology, 77, 2791-2795.
Stellera chamaejasme L. increases soil N availability, turnover rates and microbial biomass in an alpine meadow ecosystem on the eastern Tibetan Plateau of China.
Carbon and nitrogen mineralization dynamics in different soils of the tropics amended with legume residues and contrasting soil moisture contents.
1
2012
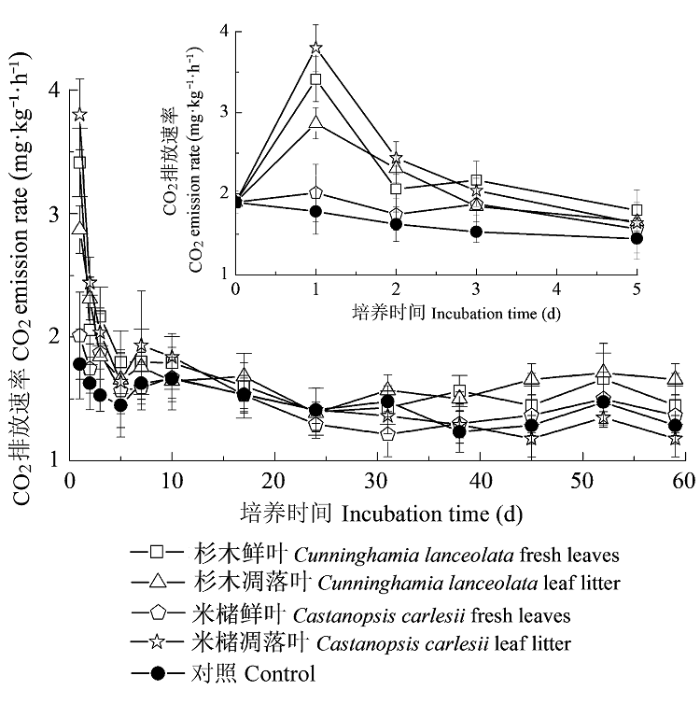
... 研究表明, 外源添加有机物的数量与质量均会影响土壤CO2排放.如Cleveland等(2010)的研究表明土壤CO2排放量随外源添加DOC的增加而增加.Wang等(2013, 2014)认为外源添加有机物的化学性质差异也会影响土壤CO2排放.因异养微生物能快速利用土壤中活性的(低分子量)、易分解的有机物从而促进土壤CO2排放(Abera et al., 2012; He & Ruan, 2014; Yang & Zhu, 2015).本研究结果发现, 土壤CO2排放受到外源添加DOM数量与质量的共同影响.如在培养第1天时, 添加不同来源的DOM的土壤CO2瞬时排放速率随外源添加DOC的增加而增加.而在培养59天时, 添加杉木凋落叶DOM的土壤CO2累积排放量显著高于添加米槠凋落叶DOM的土壤CO2累积排放量, 但外源添加杉木凋落叶DOC (35.7 mg·kg-1)显著低于外源添加米槠凋落叶DOC (55.9 mg·kg-1), 这可能与外源添加DOM的化学性质有关, 因为杉木凋落叶DOM的E2:E3值显著高于米槠凋落叶DOM的, 即外源添加的杉木凋落叶DOM中含有更多的小分子有机物, 而周转快的小分子物质能增加土壤CO2的排放(van Hees et al., 2005; Fujii et al., 2010; Rousk et al., 2011).因此, 为了更好地区分外源添加DOM的数量与质量对土壤CO2排放的影响, 后期研究需要通过控制外源DOM的数量或者质量, 从而更加深入地分析添加不同来源的DOM对土壤CO2排放的影响机制. ...
Mechanisms of real and apparent priming effects and their dependence on soil microbial biomass and community structure: Critical review.
1
2008
... 土壤有机碳库是陆地生态系统最大碳储存库, 储量高达1500 Pg (Eswaran et al., 1993), 是大气碳库的2倍、植物碳库的3倍多, 因此土壤碳库的动态变化对全球碳循环起着重要作用.土壤有机碳库变化受到凋落物输入与土壤有机碳(SOC)矿化的共同影响(Vesterdal et al., 2012).而土壤有机碳矿化不仅受到温度、水等环境因素的调控, 也受外源有机物(包括有机物的质量和数量)的影响(Kirschbaum, 2004; Fierer et al., 2005; Hartley & Ineson, 2008).如输入活性碳到土壤中后, 会增加土壤有机碳矿化(Kuzyakov et al., 2000; Blagodatskaya & Kuzyakov, 2008; Kuzyakov, 2010; Cheng et al., 2014).在森林生态系统中, 降雨淋溶、凋落物淋溶的可溶性有机物(DOM)是土壤有机物(SOM)中活性碳库的重要来源(Gauthier et al., 2010), 亦是土壤微生物分解作用的主要底物来源(Marschner & Kalbit, 2003; Li et al., 2010), 因此, 研究不同来源的DOM对土壤碳库的影响具有重要意义. ...
Biodegradation and chemical characteristics of hot-water extractable organic matter from soils under four different vegetation types in the Wuyi Mountains, southeastern China.
Synthesis and modeling perspectives of rhizosphere priming.
1
2014
... 土壤有机碳库是陆地生态系统最大碳储存库, 储量高达1500 Pg (Eswaran et al., 1993), 是大气碳库的2倍、植物碳库的3倍多, 因此土壤碳库的动态变化对全球碳循环起着重要作用.土壤有机碳库变化受到凋落物输入与土壤有机碳(SOC)矿化的共同影响(Vesterdal et al., 2012).而土壤有机碳矿化不仅受到温度、水等环境因素的调控, 也受外源有机物(包括有机物的质量和数量)的影响(Kirschbaum, 2004; Fierer et al., 2005; Hartley & Ineson, 2008).如输入活性碳到土壤中后, 会增加土壤有机碳矿化(Kuzyakov et al., 2000; Blagodatskaya & Kuzyakov, 2008; Kuzyakov, 2010; Cheng et al., 2014).在森林生态系统中, 降雨淋溶、凋落物淋溶的可溶性有机物(DOM)是土壤有机物(SOM)中活性碳库的重要来源(Gauthier et al., 2010), 亦是土壤微生物分解作用的主要底物来源(Marschner & Kalbit, 2003; Li et al., 2010), 因此, 研究不同来源的DOM对土壤碳库的影响具有重要意义. ...
Composition, dynamics, and fate of leached dissolved organic matter in terrestrial ecosystems: Results from a decomposition experiment.
1
2004
... 已有研究发现不同来源的DOM化学组成和性质差异较大(Inamdar et al., 2012; Kothawala et al., 2012).如阔叶树凋落叶可溶性有机碳(DOC)含量高于针叶树(Wieder et al., 2008), 相同树种的凋落叶DOC含量高于鲜叶(Cleveland et al., 2004), 鲜叶淋溶的DOM中含有更多低分子量、易分解的有机物, 而凋落叶DOM中含有更多的芳香性化合物和腐殖化程度更高的有机物(Kalbitz et al., 2007; Inamdar et al., 2012).因此, 分析不同树种鲜叶与凋落叶DOM的差异对土壤CO2排放的影响是生态系统碳循环的重要环节.已有的野外研究表明不同树种DOM对土壤CO2累积排放量具有显著影响(Kiikkilä et al., 2012).室内试验也证明添加不同树种凋落叶DOM后, 会影响土壤CO2排放而影响土壤碳库(Wieder et al., 2008). ...
Experimental drought in a tropical rain forest increases soil carbon dioxide losses to the atmosphere.
2010
Organic carbon in soils of the world.
1
1993
... 土壤有机碳库是陆地生态系统最大碳储存库, 储量高达1500 Pg (Eswaran et al., 1993), 是大气碳库的2倍、植物碳库的3倍多, 因此土壤碳库的动态变化对全球碳循环起着重要作用.土壤有机碳库变化受到凋落物输入与土壤有机碳(SOC)矿化的共同影响(Vesterdal et al., 2012).而土壤有机碳矿化不仅受到温度、水等环境因素的调控, 也受外源有机物(包括有机物的质量和数量)的影响(Kirschbaum, 2004; Fierer et al., 2005; Hartley & Ineson, 2008).如输入活性碳到土壤中后, 会增加土壤有机碳矿化(Kuzyakov et al., 2000; Blagodatskaya & Kuzyakov, 2008; Kuzyakov, 2010; Cheng et al., 2014).在森林生态系统中, 降雨淋溶、凋落物淋溶的可溶性有机物(DOM)是土壤有机物(SOM)中活性碳库的重要来源(Gauthier et al., 2010), 亦是土壤微生物分解作用的主要底物来源(Marschner & Kalbit, 2003; Li et al., 2010), 因此, 研究不同来源的DOM对土壤碳库的影响具有重要意义. ...
Litter quality and the temperature sensitivity of decomposition.
1
2005
... 土壤有机碳库是陆地生态系统最大碳储存库, 储量高达1500 Pg (Eswaran et al., 1993), 是大气碳库的2倍、植物碳库的3倍多, 因此土壤碳库的动态变化对全球碳循环起着重要作用.土壤有机碳库变化受到凋落物输入与土壤有机碳(SOC)矿化的共同影响(Vesterdal et al., 2012).而土壤有机碳矿化不仅受到温度、水等环境因素的调控, 也受外源有机物(包括有机物的质量和数量)的影响(Kirschbaum, 2004; Fierer et al., 2005; Hartley & Ineson, 2008).如输入活性碳到土壤中后, 会增加土壤有机碳矿化(Kuzyakov et al., 2000; Blagodatskaya & Kuzyakov, 2008; Kuzyakov, 2010; Cheng et al., 2014).在森林生态系统中, 降雨淋溶、凋落物淋溶的可溶性有机物(DOM)是土壤有机物(SOM)中活性碳库的重要来源(Gauthier et al., 2010), 亦是土壤微生物分解作用的主要底物来源(Marschner & Kalbit, 2003; Li et al., 2010), 因此, 研究不同来源的DOM对土壤碳库的影响具有重要意义. ...
Carbon input to soil may decrease soil carbon content.
2004
Biodegradation of low molecular weight organic compounds and their contribution to heterotrophic soil respiration in three Japanese forest soils.
1
2010
... 研究表明, 外源添加有机物的数量与质量均会影响土壤CO2排放.如Cleveland等(2010)的研究表明土壤CO2排放量随外源添加DOC的增加而增加.Wang等(2013, 2014)认为外源添加有机物的化学性质差异也会影响土壤CO2排放.因异养微生物能快速利用土壤中活性的(低分子量)、易分解的有机物从而促进土壤CO2排放(Abera et al., 2012; He & Ruan, 2014; Yang & Zhu, 2015).本研究结果发现, 土壤CO2排放受到外源添加DOM数量与质量的共同影响.如在培养第1天时, 添加不同来源的DOM的土壤CO2瞬时排放速率随外源添加DOC的增加而增加.而在培养59天时, 添加杉木凋落叶DOM的土壤CO2累积排放量显著高于添加米槠凋落叶DOM的土壤CO2累积排放量, 但外源添加杉木凋落叶DOC (35.7 mg·kg-1)显著低于外源添加米槠凋落叶DOC (55.9 mg·kg-1), 这可能与外源添加DOM的化学性质有关, 因为杉木凋落叶DOM的E2:E3值显著高于米槠凋落叶DOM的, 即外源添加的杉木凋落叶DOM中含有更多的小分子有机物, 而周转快的小分子物质能增加土壤CO2的排放(van Hees et al., 2005; Fujii et al., 2010; Rousk et al., 2011).因此, 为了更好地区分外源添加DOM的数量与质量对土壤CO2排放的影响, 后期研究需要通过控制外源DOM的数量或者质量, 从而更加深入地分析添加不同来源的DOM对土壤CO2排放的影响机制. ...
Dynamics of the water extractable organic carbon pool during mineralisation in soils from a Douglas fir plantation and an oak-beech forest―An incubation experiment.
2
2010
... 土壤有机碳库是陆地生态系统最大碳储存库, 储量高达1500 Pg (Eswaran et al., 1993), 是大气碳库的2倍、植物碳库的3倍多, 因此土壤碳库的动态变化对全球碳循环起着重要作用.土壤有机碳库变化受到凋落物输入与土壤有机碳(SOC)矿化的共同影响(Vesterdal et al., 2012).而土壤有机碳矿化不仅受到温度、水等环境因素的调控, 也受外源有机物(包括有机物的质量和数量)的影响(Kirschbaum, 2004; Fierer et al., 2005; Hartley & Ineson, 2008).如输入活性碳到土壤中后, 会增加土壤有机碳矿化(Kuzyakov et al., 2000; Blagodatskaya & Kuzyakov, 2008; Kuzyakov, 2010; Cheng et al., 2014).在森林生态系统中, 降雨淋溶、凋落物淋溶的可溶性有机物(DOM)是土壤有机物(SOM)中活性碳库的重要来源(Gauthier et al., 2010), 亦是土壤微生物分解作用的主要底物来源(Marschner & Kalbit, 2003; Li et al., 2010), 因此, 研究不同来源的DOM对土壤碳库的影响具有重要意义. ...
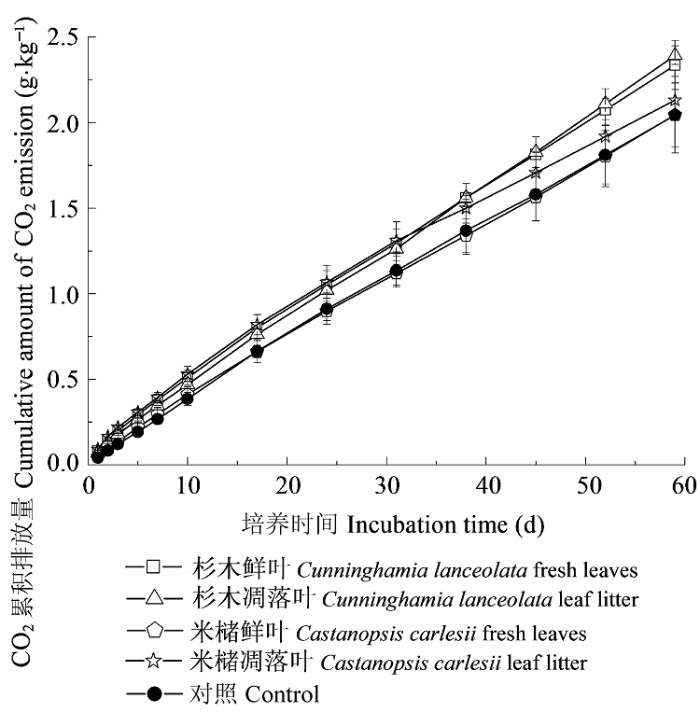
... 培养59天时, 添加米槠鲜叶和凋落叶DOM后增加了土壤总有机碳库, 这与许多外源添加有机物对土壤碳库的影响是一致的(Qiao et al., 2014; Xiao et al., 2014).但添加杉木鲜叶和凋落叶DOM后降低了土壤总有机碳库, Fontaine等(2004)、Hoosbeek和Scarascia-Mugnozza (2009)也得到类似的研究结果, 这与外源添加DOM的差异有关.如Wieder等(2008)的研究表明, 添加不同树种凋落叶DOM到土壤中后, 对土壤总有机碳库的影响是不一致的, 这主要是由外源添加的DOM的化学性质差异引起.在培养前31天时, 添加米槠凋落叶DOM的土壤CO2累积排放量比对照高15.4%, 但培养59天时下降到4.3%, 说明在培养后期添加米槠凋落叶DOM的土壤CO2累积排放量与对照逐渐趋于一致, 这与其他3种处理的结果相反, 其原因除了与培养时间有关外, 还可能由于添加外源DOM的质量对土壤微生物活动的影响(Gauthier et al., 2010), 如本研究发现外源添加的米槠凋落叶DOM中含有更多大分子量、难分解的化合物. ...
Substrate quality and the temperature sensitivity of soil organic matter decomposition.
1
2008
... 土壤有机碳库是陆地生态系统最大碳储存库, 储量高达1500 Pg (Eswaran et al., 1993), 是大气碳库的2倍、植物碳库的3倍多, 因此土壤碳库的动态变化对全球碳循环起着重要作用.土壤有机碳库变化受到凋落物输入与土壤有机碳(SOC)矿化的共同影响(Vesterdal et al., 2012).而土壤有机碳矿化不仅受到温度、水等环境因素的调控, 也受外源有机物(包括有机物的质量和数量)的影响(Kirschbaum, 2004; Fierer et al., 2005; Hartley & Ineson, 2008).如输入活性碳到土壤中后, 会增加土壤有机碳矿化(Kuzyakov et al., 2000; Blagodatskaya & Kuzyakov, 2008; Kuzyakov, 2010; Cheng et al., 2014).在森林生态系统中, 降雨淋溶、凋落物淋溶的可溶性有机物(DOM)是土壤有机物(SOM)中活性碳库的重要来源(Gauthier et al., 2010), 亦是土壤微生物分解作用的主要底物来源(Marschner & Kalbit, 2003; Li et al., 2010), 因此, 研究不同来源的DOM对土壤碳库的影响具有重要意义. ...
Long term effect of land reclamation from Lake on chemical composition of soil organic matter and its mineralization.
1
2014
... 研究表明, 外源添加有机物的数量与质量均会影响土壤CO2排放.如Cleveland等(2010)的研究表明土壤CO2排放量随外源添加DOC的增加而增加.Wang等(2013, 2014)认为外源添加有机物的化学性质差异也会影响土壤CO2排放.因异养微生物能快速利用土壤中活性的(低分子量)、易分解的有机物从而促进土壤CO2排放(Abera et al., 2012; He & Ruan, 2014; Yang & Zhu, 2015).本研究结果发现, 土壤CO2排放受到外源添加DOM数量与质量的共同影响.如在培养第1天时, 添加不同来源的DOM的土壤CO2瞬时排放速率随外源添加DOC的增加而增加.而在培养59天时, 添加杉木凋落叶DOM的土壤CO2累积排放量显著高于添加米槠凋落叶DOM的土壤CO2累积排放量, 但外源添加杉木凋落叶DOC (35.7 mg·kg-1)显著低于外源添加米槠凋落叶DOC (55.9 mg·kg-1), 这可能与外源添加DOM的化学性质有关, 因为杉木凋落叶DOM的E2:E3值显著高于米槠凋落叶DOM的, 即外源添加的杉木凋落叶DOM中含有更多的小分子有机物, 而周转快的小分子物质能增加土壤CO2的排放(van Hees et al., 2005; Fujii et al., 2010; Rousk et al., 2011).因此, 为了更好地区分外源添加DOM的数量与质量对土壤CO2排放的影响, 后期研究需要通过控制外源DOM的数量或者质量, 从而更加深入地分析添加不同来源的DOM对土壤CO2排放的影响机制. ...
Increased litter build up and soil organic matter stabilization in a poplar plantation after 6 years of atmospheric CO2 enrichment (FACE): Final results of POP-Euro FACE compared to other forest FACE experiments.
2009
Dissolved organic matter (DOM) concentration and quality in a forested mid- Atlantic watershed, USA.
3
2012
... 已有研究发现不同来源的DOM化学组成和性质差异较大(Inamdar et al., 2012; Kothawala et al., 2012).如阔叶树凋落叶可溶性有机碳(DOC)含量高于针叶树(Wieder et al., 2008), 相同树种的凋落叶DOC含量高于鲜叶(Cleveland et al., 2004), 鲜叶淋溶的DOM中含有更多低分子量、易分解的有机物, 而凋落叶DOM中含有更多的芳香性化合物和腐殖化程度更高的有机物(Kalbitz et al., 2007; Inamdar et al., 2012).因此, 分析不同树种鲜叶与凋落叶DOM的差异对土壤CO2排放的影响是生态系统碳循环的重要环节.已有的野外研究表明不同树种DOM对土壤CO2累积排放量具有显著影响(Kiikkilä et al., 2012).室内试验也证明添加不同树种凋落叶DOM后, 会影响土壤CO2排放而影响土壤碳库(Wieder et al., 2008). ...
... ; Inamdar et al., 2012).因此, 分析不同树种鲜叶与凋落叶DOM的差异对土壤CO2排放的影响是生态系统碳循环的重要环节.已有的野外研究表明不同树种DOM对土壤CO2累积排放量具有显著影响(Kiikkilä et al., 2012).室内试验也证明添加不同树种凋落叶DOM后, 会影响土壤CO2排放而影响土壤碳库(Wieder et al., 2008). ...
Response of dissolved organic matter in the forest floor to long-term manipulation of litter and throughfall inputs.
1
2007
... 已有研究发现不同来源的DOM化学组成和性质差异较大(Inamdar et al., 2012; Kothawala et al., 2012).如阔叶树凋落叶可溶性有机碳(DOC)含量高于针叶树(Wieder et al., 2008), 相同树种的凋落叶DOC含量高于鲜叶(Cleveland et al., 2004), 鲜叶淋溶的DOM中含有更多低分子量、易分解的有机物, 而凋落叶DOM中含有更多的芳香性化合物和腐殖化程度更高的有机物(Kalbitz et al., 2007; Inamdar et al., 2012).因此, 分析不同树种鲜叶与凋落叶DOM的差异对土壤CO2排放的影响是生态系统碳循环的重要环节.已有的野外研究表明不同树种DOM对土壤CO2累积排放量具有显著影响(Kiikkilä et al., 2012).室内试验也证明添加不同树种凋落叶DOM后, 会影响土壤CO2排放而影响土壤碳库(Wieder et al., 2008). ...
Properties of dissolved organic matter derived from silver birch and Norway spruce stands: Degradability combined with chemical characteristics.
2011
Characterization of dissolved organic matter in decomposing Norway spruce and silver birch litter.
1
2012
... 已有研究发现不同来源的DOM化学组成和性质差异较大(Inamdar et al., 2012; Kothawala et al., 2012).如阔叶树凋落叶可溶性有机碳(DOC)含量高于针叶树(Wieder et al., 2008), 相同树种的凋落叶DOC含量高于鲜叶(Cleveland et al., 2004), 鲜叶淋溶的DOM中含有更多低分子量、易分解的有机物, 而凋落叶DOM中含有更多的芳香性化合物和腐殖化程度更高的有机物(Kalbitz et al., 2007; Inamdar et al., 2012).因此, 分析不同树种鲜叶与凋落叶DOM的差异对土壤CO2排放的影响是生态系统碳循环的重要环节.已有的野外研究表明不同树种DOM对土壤CO2累积排放量具有显著影响(Kiikkilä et al., 2012).室内试验也证明添加不同树种凋落叶DOM后, 会影响土壤CO2排放而影响土壤碳库(Wieder et al., 2008). ...
Soil microbial activity in relation to dissolved organic matter properties under different tree species.
1
2014
... 已有研究表明, 添加外源DOM后会显著影响土壤CO2排放(Wieder et al., 2008; Leff et al., 2012), 本研究也发现, 添加不同来源的DOM后土壤CO2累积排放量均高于对照.在培养31天时, 添加米槠凋落叶DOM的土壤CO2累积排放量高于添加杉木凋落叶DOM的(p = 0.351), 但培养结束时, 添加米槠凋落叶DOM的土壤CO2累积排放量比添加杉木凋落叶DOM的低11%, 这除了与培养时间有关, 可能也与不同树种DOM的差异有关(Kiikkilä et al., 2014).培养31天时, 添加米槠鲜叶DOM的土壤CO2累积排放量显著低于添加米槠凋落叶DOM的, 这可能是因为外源添加DOC:DON大于25, 使外源N成了土壤微生物生长和繁殖的限制性因素, 因而, 含氮量高的DOM进入土壤后微生物迅速繁殖促进了土壤CO2排放(Nourbakhsh & Dick, 2005; Sun et al., 2009).而在培养59天时, 添加米槠鲜叶DOM的土壤CO2累积排放量与添加米槠凋落叶DOM的差异不显著, 表明随着培养时间延长, 添加米槠鲜叶DOM与米槠凋落叶DOM的土壤CO2累积排放量之间的差异在逐渐减小.同时, 添加杉木鲜叶和凋落叶DOM的土壤CO2累积量均显著高于添加米槠鲜叶和凋落叶DOM的, 表明单次添加外源可溶性有机物到土壤中, 添加杉木树种DOM对土壤CO2排放的影响更大. ...
Soil respiration under prolonged soil warming: Are rate reductions caused by acclimation or substrate loss?
1
2004
... 土壤有机碳库是陆地生态系统最大碳储存库, 储量高达1500 Pg (Eswaran et al., 1993), 是大气碳库的2倍、植物碳库的3倍多, 因此土壤碳库的动态变化对全球碳循环起着重要作用.土壤有机碳库变化受到凋落物输入与土壤有机碳(SOC)矿化的共同影响(Vesterdal et al., 2012).而土壤有机碳矿化不仅受到温度、水等环境因素的调控, 也受外源有机物(包括有机物的质量和数量)的影响(Kirschbaum, 2004; Fierer et al., 2005; Hartley & Ineson, 2008).如输入活性碳到土壤中后, 会增加土壤有机碳矿化(Kuzyakov et al., 2000; Blagodatskaya & Kuzyakov, 2008; Kuzyakov, 2010; Cheng et al., 2014).在森林生态系统中, 降雨淋溶、凋落物淋溶的可溶性有机物(DOM)是土壤有机物(SOM)中活性碳库的重要来源(Gauthier et al., 2010), 亦是土壤微生物分解作用的主要底物来源(Marschner & Kalbit, 2003; Li et al., 2010), 因此, 研究不同来源的DOM对土壤碳库的影响具有重要意义. ...
Selective adsorption of dissolved organic matter to mineral soils.
1
2012
... 已有研究发现不同来源的DOM化学组成和性质差异较大(Inamdar et al., 2012; Kothawala et al., 2012).如阔叶树凋落叶可溶性有机碳(DOC)含量高于针叶树(Wieder et al., 2008), 相同树种的凋落叶DOC含量高于鲜叶(Cleveland et al., 2004), 鲜叶淋溶的DOM中含有更多低分子量、易分解的有机物, 而凋落叶DOM中含有更多的芳香性化合物和腐殖化程度更高的有机物(Kalbitz et al., 2007; Inamdar et al., 2012).因此, 分析不同树种鲜叶与凋落叶DOM的差异对土壤CO2排放的影响是生态系统碳循环的重要环节.已有的野外研究表明不同树种DOM对土壤CO2累积排放量具有显著影响(Kiikkilä et al., 2012).室内试验也证明添加不同树种凋落叶DOM后, 会影响土壤CO2排放而影响土壤碳库(Wieder et al., 2008). ...
Priming effects: Interactions between living and dead organic matter.
1
2010
... 土壤有机碳库是陆地生态系统最大碳储存库, 储量高达1500 Pg (Eswaran et al., 1993), 是大气碳库的2倍、植物碳库的3倍多, 因此土壤碳库的动态变化对全球碳循环起着重要作用.土壤有机碳库变化受到凋落物输入与土壤有机碳(SOC)矿化的共同影响(Vesterdal et al., 2012).而土壤有机碳矿化不仅受到温度、水等环境因素的调控, 也受外源有机物(包括有机物的质量和数量)的影响(Kirschbaum, 2004; Fierer et al., 2005; Hartley & Ineson, 2008).如输入活性碳到土壤中后, 会增加土壤有机碳矿化(Kuzyakov et al., 2000; Blagodatskaya & Kuzyakov, 2008; Kuzyakov, 2010; Cheng et al., 2014).在森林生态系统中, 降雨淋溶、凋落物淋溶的可溶性有机物(DOM)是土壤有机物(SOM)中活性碳库的重要来源(Gauthier et al., 2010), 亦是土壤微生物分解作用的主要底物来源(Marschner & Kalbit, 2003; Li et al., 2010), 因此, 研究不同来源的DOM对土壤碳库的影响具有重要意义. ...
Review of mechanisms and quantification of priming effects.
1
2000
... 土壤有机碳库是陆地生态系统最大碳储存库, 储量高达1500 Pg (Eswaran et al., 1993), 是大气碳库的2倍、植物碳库的3倍多, 因此土壤碳库的动态变化对全球碳循环起着重要作用.土壤有机碳库变化受到凋落物输入与土壤有机碳(SOC)矿化的共同影响(Vesterdal et al., 2012).而土壤有机碳矿化不仅受到温度、水等环境因素的调控, 也受外源有机物(包括有机物的质量和数量)的影响(Kirschbaum, 2004; Fierer et al., 2005; Hartley & Ineson, 2008).如输入活性碳到土壤中后, 会增加土壤有机碳矿化(Kuzyakov et al., 2000; Blagodatskaya & Kuzyakov, 2008; Kuzyakov, 2010; Cheng et al., 2014).在森林生态系统中, 降雨淋溶、凋落物淋溶的可溶性有机物(DOM)是土壤有机物(SOM)中活性碳库的重要来源(Gauthier et al., 2010), 亦是土壤微生物分解作用的主要底物来源(Marschner & Kalbit, 2003; Li et al., 2010), 因此, 研究不同来源的DOM对土壤碳库的影响具有重要意义. ...
The effects of soil bacterial community structure on decomposition in a tropical rain forest.
1
2012
... 已有研究表明, 添加外源DOM后会显著影响土壤CO2排放(Wieder et al., 2008; Leff et al., 2012), 本研究也发现, 添加不同来源的DOM后土壤CO2累积排放量均高于对照.在培养31天时, 添加米槠凋落叶DOM的土壤CO2累积排放量高于添加杉木凋落叶DOM的(p = 0.351), 但培养结束时, 添加米槠凋落叶DOM的土壤CO2累积排放量比添加杉木凋落叶DOM的低11%, 这除了与培养时间有关, 可能也与不同树种DOM的差异有关(Kiikkilä et al., 2014).培养31天时, 添加米槠鲜叶DOM的土壤CO2累积排放量显著低于添加米槠凋落叶DOM的, 这可能是因为外源添加DOC:DON大于25, 使外源N成了土壤微生物生长和繁殖的限制性因素, 因而, 含氮量高的DOM进入土壤后微生物迅速繁殖促进了土壤CO2排放(Nourbakhsh & Dick, 2005; Sun et al., 2009).而在培养59天时, 添加米槠鲜叶DOM的土壤CO2累积排放量与添加米槠凋落叶DOM的差异不显著, 表明随着培养时间延长, 添加米槠鲜叶DOM与米槠凋落叶DOM的土壤CO2累积排放量之间的差异在逐渐减小.同时, 添加杉木鲜叶和凋落叶DOM的土壤CO2累积量均显著高于添加米槠鲜叶和凋落叶DOM的, 表明单次添加外源可溶性有机物到土壤中, 添加杉木树种DOM对土壤CO2排放的影响更大. ...
Organic C and N mineralization as affected by dissolved organic matter in paddy soils of subtropical China.
1
2010
... 土壤有机碳库是陆地生态系统最大碳储存库, 储量高达1500 Pg (Eswaran et al., 1993), 是大气碳库的2倍、植物碳库的3倍多, 因此土壤碳库的动态变化对全球碳循环起着重要作用.土壤有机碳库变化受到凋落物输入与土壤有机碳(SOC)矿化的共同影响(Vesterdal et al., 2012).而土壤有机碳矿化不仅受到温度、水等环境因素的调控, 也受外源有机物(包括有机物的质量和数量)的影响(Kirschbaum, 2004; Fierer et al., 2005; Hartley & Ineson, 2008).如输入活性碳到土壤中后, 会增加土壤有机碳矿化(Kuzyakov et al., 2000; Blagodatskaya & Kuzyakov, 2008; Kuzyakov, 2010; Cheng et al., 2014).在森林生态系统中, 降雨淋溶、凋落物淋溶的可溶性有机物(DOM)是土壤有机物(SOM)中活性碳库的重要来源(Gauthier et al., 2010), 亦是土壤微生物分解作用的主要底物来源(Marschner & Kalbit, 2003; Li et al., 2010), 因此, 研究不同来源的DOM对土壤碳库的影响具有重要意义. ...
Controls of bioavailability and biodegradability of dissolved organic matter in soils.
1
2003
... 土壤有机碳库是陆地生态系统最大碳储存库, 储量高达1500 Pg (Eswaran et al., 1993), 是大气碳库的2倍、植物碳库的3倍多, 因此土壤碳库的动态变化对全球碳循环起着重要作用.土壤有机碳库变化受到凋落物输入与土壤有机碳(SOC)矿化的共同影响(Vesterdal et al., 2012).而土壤有机碳矿化不仅受到温度、水等环境因素的调控, 也受外源有机物(包括有机物的质量和数量)的影响(Kirschbaum, 2004; Fierer et al., 2005; Hartley & Ineson, 2008).如输入活性碳到土壤中后, 会增加土壤有机碳矿化(Kuzyakov et al., 2000; Blagodatskaya & Kuzyakov, 2008; Kuzyakov, 2010; Cheng et al., 2014).在森林生态系统中, 降雨淋溶、凋落物淋溶的可溶性有机物(DOM)是土壤有机物(SOM)中活性碳库的重要来源(Gauthier et al., 2010), 亦是土壤微生物分解作用的主要底物来源(Marschner & Kalbit, 2003; Li et al., 2010), 因此, 研究不同来源的DOM对土壤碳库的影响具有重要意义. ...
Net nitrogen mineralization or immobilization potential in a residue―Amended calcareous soil.
1
2005
... 已有研究表明, 添加外源DOM后会显著影响土壤CO2排放(Wieder et al., 2008; Leff et al., 2012), 本研究也发现, 添加不同来源的DOM后土壤CO2累积排放量均高于对照.在培养31天时, 添加米槠凋落叶DOM的土壤CO2累积排放量高于添加杉木凋落叶DOM的(p = 0.351), 但培养结束时, 添加米槠凋落叶DOM的土壤CO2累积排放量比添加杉木凋落叶DOM的低11%, 这除了与培养时间有关, 可能也与不同树种DOM的差异有关(Kiikkilä et al., 2014).培养31天时, 添加米槠鲜叶DOM的土壤CO2累积排放量显著低于添加米槠凋落叶DOM的, 这可能是因为外源添加DOC:DON大于25, 使外源N成了土壤微生物生长和繁殖的限制性因素, 因而, 含氮量高的DOM进入土壤后微生物迅速繁殖促进了土壤CO2排放(Nourbakhsh & Dick, 2005; Sun et al., 2009).而在培养59天时, 添加米槠鲜叶DOM的土壤CO2累积排放量与添加米槠凋落叶DOM的差异不显著, 表明随着培养时间延长, 添加米槠鲜叶DOM与米槠凋落叶DOM的土壤CO2累积排放量之间的差异在逐渐减小.同时, 添加杉木鲜叶和凋落叶DOM的土壤CO2累积量均显著高于添加米槠鲜叶和凋落叶DOM的, 表明单次添加外源可溶性有机物到土壤中, 添加杉木树种DOM对土壤CO2排放的影响更大. ...
Molecular size distribution and spectroscopic properties of aquatic humic substances.
Labile carbon retention compensates for CO2 released by priming in forest soils.
1
2014
... 培养59天时, 添加米槠鲜叶和凋落叶DOM后增加了土壤总有机碳库, 这与许多外源添加有机物对土壤碳库的影响是一致的(Qiao et al., 2014; Xiao et al., 2014).但添加杉木鲜叶和凋落叶DOM后降低了土壤总有机碳库, Fontaine等(2004)、Hoosbeek和Scarascia-Mugnozza (2009)也得到类似的研究结果, 这与外源添加DOM的差异有关.如Wieder等(2008)的研究表明, 添加不同树种凋落叶DOM到土壤中后, 对土壤总有机碳库的影响是不一致的, 这主要是由外源添加的DOM的化学性质差异引起.在培养前31天时, 添加米槠凋落叶DOM的土壤CO2累积排放量比对照高15.4%, 但培养59天时下降到4.3%, 说明在培养后期添加米槠凋落叶DOM的土壤CO2累积排放量与对照逐渐趋于一致, 这与其他3种处理的结果相反, 其原因除了与培养时间有关外, 还可能由于添加外源DOM的质量对土壤微生物活动的影响(Gauthier et al., 2010), 如本研究发现外源添加的米槠凋落叶DOM中含有更多大分子量、难分解的化合物. ...
Lack of correlation between turnover of low-molecular-weight dissolved organic carbon and differences in microbial community composition or growth across a soil pH gradient.
1
2011
... 研究表明, 外源添加有机物的数量与质量均会影响土壤CO2排放.如Cleveland等(2010)的研究表明土壤CO2排放量随外源添加DOC的增加而增加.Wang等(2013, 2014)认为外源添加有机物的化学性质差异也会影响土壤CO2排放.因异养微生物能快速利用土壤中活性的(低分子量)、易分解的有机物从而促进土壤CO2排放(Abera et al., 2012; He & Ruan, 2014; Yang & Zhu, 2015).本研究结果发现, 土壤CO2排放受到外源添加DOM数量与质量的共同影响.如在培养第1天时, 添加不同来源的DOM的土壤CO2瞬时排放速率随外源添加DOC的增加而增加.而在培养59天时, 添加杉木凋落叶DOM的土壤CO2累积排放量显著高于添加米槠凋落叶DOM的土壤CO2累积排放量, 但外源添加杉木凋落叶DOC (35.7 mg·kg-1)显著低于外源添加米槠凋落叶DOC (55.9 mg·kg-1), 这可能与外源添加DOM的化学性质有关, 因为杉木凋落叶DOM的E2:E3值显著高于米槠凋落叶DOM的, 即外源添加的杉木凋落叶DOM中含有更多的小分子有机物, 而周转快的小分子物质能增加土壤CO2的排放(van Hees et al., 2005; Fujii et al., 2010; Rousk et al., 2011).因此, 为了更好地区分外源添加DOM的数量与质量对土壤CO2排放的影响, 后期研究需要通过控制外源DOM的数量或者质量, 从而更加深入地分析添加不同来源的DOM对土壤CO2排放的影响机制. ...
Stellera chamaejasme L. increases soil N availability, turnover rates and microbial biomass in an alpine meadow ecosystem on the eastern Tibetan Plateau of China.
1
2009
... 已有研究表明, 添加外源DOM后会显著影响土壤CO2排放(Wieder et al., 2008; Leff et al., 2012), 本研究也发现, 添加不同来源的DOM后土壤CO2累积排放量均高于对照.在培养31天时, 添加米槠凋落叶DOM的土壤CO2累积排放量高于添加杉木凋落叶DOM的(p = 0.351), 但培养结束时, 添加米槠凋落叶DOM的土壤CO2累积排放量比添加杉木凋落叶DOM的低11%, 这除了与培养时间有关, 可能也与不同树种DOM的差异有关(Kiikkilä et al., 2014).培养31天时, 添加米槠鲜叶DOM的土壤CO2累积排放量显著低于添加米槠凋落叶DOM的, 这可能是因为外源添加DOC:DON大于25, 使外源N成了土壤微生物生长和繁殖的限制性因素, 因而, 含氮量高的DOM进入土壤后微生物迅速繁殖促进了土壤CO2排放(Nourbakhsh & Dick, 2005; Sun et al., 2009).而在培养59天时, 添加米槠鲜叶DOM的土壤CO2累积排放量与添加米槠凋落叶DOM的差异不显著, 表明随着培养时间延长, 添加米槠鲜叶DOM与米槠凋落叶DOM的土壤CO2累积排放量之间的差异在逐渐减小.同时, 添加杉木鲜叶和凋落叶DOM的土壤CO2累积量均显著高于添加米槠鲜叶和凋落叶DOM的, 表明单次添加外源可溶性有机物到土壤中, 添加杉木树种DOM对土壤CO2排放的影响更大. ...
The carbon we do not see-the impact of low molecular weight compounds on carbon dynamics and respiration in forest soils: A review.
1
2005
... 研究表明, 外源添加有机物的数量与质量均会影响土壤CO2排放.如Cleveland等(2010)的研究表明土壤CO2排放量随外源添加DOC的增加而增加.Wang等(2013, 2014)认为外源添加有机物的化学性质差异也会影响土壤CO2排放.因异养微生物能快速利用土壤中活性的(低分子量)、易分解的有机物从而促进土壤CO2排放(Abera et al., 2012; He & Ruan, 2014; Yang & Zhu, 2015).本研究结果发现, 土壤CO2排放受到外源添加DOM数量与质量的共同影响.如在培养第1天时, 添加不同来源的DOM的土壤CO2瞬时排放速率随外源添加DOC的增加而增加.而在培养59天时, 添加杉木凋落叶DOM的土壤CO2累积排放量显著高于添加米槠凋落叶DOM的土壤CO2累积排放量, 但外源添加杉木凋落叶DOC (35.7 mg·kg-1)显著低于外源添加米槠凋落叶DOC (55.9 mg·kg-1), 这可能与外源添加DOM的化学性质有关, 因为杉木凋落叶DOM的E2:E3值显著高于米槠凋落叶DOM的, 即外源添加的杉木凋落叶DOM中含有更多的小分子有机物, 而周转快的小分子物质能增加土壤CO2的排放(van Hees et al., 2005; Fujii et al., 2010; Rousk et al., 2011).因此, 为了更好地区分外源添加DOM的数量与质量对土壤CO2排放的影响, 后期研究需要通过控制外源DOM的数量或者质量, 从而更加深入地分析添加不同来源的DOM对土壤CO2排放的影响机制. ...
Soil respiration and rates of soil carbon turnover differ among six common European tree species.
1
2012
... 土壤有机碳库是陆地生态系统最大碳储存库, 储量高达1500 Pg (Eswaran et al., 1993), 是大气碳库的2倍、植物碳库的3倍多, 因此土壤碳库的动态变化对全球碳循环起着重要作用.土壤有机碳库变化受到凋落物输入与土壤有机碳(SOC)矿化的共同影响(Vesterdal et al., 2012).而土壤有机碳矿化不仅受到温度、水等环境因素的调控, 也受外源有机物(包括有机物的质量和数量)的影响(Kirschbaum, 2004; Fierer et al., 2005; Hartley & Ineson, 2008).如输入活性碳到土壤中后, 会增加土壤有机碳矿化(Kuzyakov et al., 2000; Blagodatskaya & Kuzyakov, 2008; Kuzyakov, 2010; Cheng et al., 2014).在森林生态系统中, 降雨淋溶、凋落物淋溶的可溶性有机物(DOM)是土壤有机物(SOM)中活性碳库的重要来源(Gauthier et al., 2010), 亦是土壤微生物分解作用的主要底物来源(Marschner & Kalbit, 2003; Li et al., 2010), 因此, 研究不同来源的DOM对土壤碳库的影响具有重要意义. ...
Debris manipulation alters soil CO2 efflux in a subtropical plantation forest.
2013
Response of organic carbon mineralization and microbial community to leaf litter and nutrient additions in subtropical forest soils.
2014
Tropical tree species composition affects the oxidation of dissolved organic matter from litter.
3
2008
... 已有研究发现不同来源的DOM化学组成和性质差异较大(Inamdar et al., 2012; Kothawala et al., 2012).如阔叶树凋落叶可溶性有机碳(DOC)含量高于针叶树(Wieder et al., 2008), 相同树种的凋落叶DOC含量高于鲜叶(Cleveland et al., 2004), 鲜叶淋溶的DOM中含有更多低分子量、易分解的有机物, 而凋落叶DOM中含有更多的芳香性化合物和腐殖化程度更高的有机物(Kalbitz et al., 2007; Inamdar et al., 2012).因此, 分析不同树种鲜叶与凋落叶DOM的差异对土壤CO2排放的影响是生态系统碳循环的重要环节.已有的野外研究表明不同树种DOM对土壤CO2累积排放量具有显著影响(Kiikkilä et al., 2012).室内试验也证明添加不同树种凋落叶DOM后, 会影响土壤CO2排放而影响土壤碳库(Wieder et al., 2008). ...
... 排放而影响土壤碳库(Wieder et al., 2008). ...
... 已有研究表明, 添加外源DOM后会显著影响土壤CO2排放(Wieder et al., 2008; Leff et al., 2012), 本研究也发现, 添加不同来源的DOM后土壤CO2累积排放量均高于对照.在培养31天时, 添加米槠凋落叶DOM的土壤CO2累积排放量高于添加杉木凋落叶DOM的(p = 0.351), 但培养结束时, 添加米槠凋落叶DOM的土壤CO2累积排放量比添加杉木凋落叶DOM的低11%, 这除了与培养时间有关, 可能也与不同树种DOM的差异有关(Kiikkilä et al., 2014).培养31天时, 添加米槠鲜叶DOM的土壤CO2累积排放量显著低于添加米槠凋落叶DOM的, 这可能是因为外源添加DOC:DON大于25, 使外源N成了土壤微生物生长和繁殖的限制性因素, 因而, 含氮量高的DOM进入土壤后微生物迅速繁殖促进了土壤CO2排放(Nourbakhsh & Dick, 2005; Sun et al., 2009).而在培养59天时, 添加米槠鲜叶DOM的土壤CO2累积排放量与添加米槠凋落叶DOM的差异不显著, 表明随着培养时间延长, 添加米槠鲜叶DOM与米槠凋落叶DOM的土壤CO2累积排放量之间的差异在逐渐减小.同时, 添加杉木鲜叶和凋落叶DOM的土壤CO2累积量均显著高于添加米槠鲜叶和凋落叶DOM的, 表明单次添加外源可溶性有机物到土壤中, 添加杉木树种DOM对土壤CO2排放的影响更大. ...
米槠和杉木人工林土壤呼吸及其组分分析
1
2014
... 试验地在福建省三明市格氏栲自然保护区(26.17º N, 117.47º E).该保护区气候属于中亚热带季风气候, 试验地附近的三明市年平均气温20.1 ℃, 年降水量1670 mm, 降水多集中于3-8月份(吴君君等, 2014).杉木人工林为2003年米槠次生林皆伐后 营造人工纯林形成, 树龄11年. ...
米槠和杉木人工林土壤呼吸及其组分分析
1
2014
... 试验地在福建省三明市格氏栲自然保护区(26.17º N, 117.47º E).该保护区气候属于中亚热带季风气候, 试验地附近的三明市年平均气温20.1 ℃, 年降水量1670 mm, 降水多集中于3-8月份(吴君君等, 2014).杉木人工林为2003年米槠次生林皆伐后 营造人工纯林形成, 树龄11年. ...
Increasing active biomass carbon may lead to a breakdown of mature forest equilibrium.
1
2014
... 培养59天时, 添加米槠鲜叶和凋落叶DOM后增加了土壤总有机碳库, 这与许多外源添加有机物对土壤碳库的影响是一致的(Qiao et al., 2014; Xiao et al., 2014).但添加杉木鲜叶和凋落叶DOM后降低了土壤总有机碳库, Fontaine等(2004)、Hoosbeek和Scarascia-Mugnozza (2009)也得到类似的研究结果, 这与外源添加DOM的差异有关.如Wieder等(2008)的研究表明, 添加不同树种凋落叶DOM到土壤中后, 对土壤总有机碳库的影响是不一致的, 这主要是由外源添加的DOM的化学性质差异引起.在培养前31天时, 添加米槠凋落叶DOM的土壤CO2累积排放量比对照高15.4%, 但培养59天时下降到4.3%, 说明在培养后期添加米槠凋落叶DOM的土壤CO2累积排放量与对照逐渐趋于一致, 这与其他3种处理的结果相反, 其原因除了与培养时间有关外, 还可能由于添加外源DOM的质量对土壤微生物活动的影响(Gauthier et al., 2010), 如本研究发现外源添加的米槠凋落叶DOM中含有更多大分子量、难分解的化合物. ...
 , 纪淑蓉, 任卫岭, 司友涛, 杨玉盛
, 纪淑蓉, 任卫岭, 司友涛, 杨玉盛


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}