盐沼植物作为全球范围内生产力较高的生物群落之一, 是推动湿地生态系统物质循环和能量流动的基础要素, 其在维持个体生存、种群稳定与群落发展的同时, 也为湿地生态系统中的众多成员提供了丰富的食物, 并创造了良好的栖息环境。然而, 近年来由于全球气候变化和频繁的人类活动, 植物群落组成结构以及空间分布格局发生了巨大的变化, 这不仅影响了湿地中鸟类、底栖生物及鱼类等生物的种群数量, 而且也不断改变着植物类群的种群结构, 甚至可能会导致一些敏感物种的灭绝(童春富, 2012 ; Isacch et al ., 2014 ; Li et al ., 2014b )。因此, 明确湿地中主要植物种群及其分布状况, 掌握其随环境因子的变化规律, 对湿地生态系统的保护、恢复、重建都具有重要的参考价值。虽然前人已就此进行了较多的研究(黄华梅等, 2007 ; Duarte et al ., 2013 ; 曹浩冰等, 2014 ; Hughes, 2014 ; Li et al ., 2014a , 2014b ), 但河口潮滩区域海陆交互作用强烈、人类活动频繁, 使得滩面、潮沟及植被都处于不断变化中, 崇明东滩正是这样一个处在发育过程中的典型盐沼湿地生态系统。2014年崇明东滩互花米草(Spartina alterniflora )生态治理工程的实施, 可能对该区域盐沼植物的空间分布及其生境产生了重大影响(高宇和赵斌, 2006 ; Zhu et al ., 2014 )。因此, 在施工前期对围垦外围区进行全面的植被调查, 不仅能准确地掌握崇明东滩南部的植被类型和现有分布状况, 而且能为今后研究其植被空间动态变化提供基础数据。
近十几年来, 大量研究人员通过遥感影像解译, 对崇明东滩主要植被进行了分区(Gao & Zhang, 2006 ; Tian et al ., 2008 ; 阿也提古丽·斯迪克等, 2010 ; 翟新源和杨苏新, 2013 ; 郑宗生等, 2010 ; 宗玮等, 2011 ), 普遍认为芦苇(Phragmites australis )、互花米草(Spartina alterniflora )及海三棱藨草(Scirpus mariqueter )是崇明东滩的“三大”主要植物类群。然而, 崇明东滩南部长期受长江入海淡水影响, 水体盐度相对较低, 在团结沙至大潮沟(图1 )存在较大面积的藨草(Scirpus triqueter )种群区(闫芊等, 2007 ; 章振亚等, 2012 )。同时发现, 糙叶薹草(Carex scabrifolia )在研究区内频繁出现, 白茅(Imperata cylindrica )在部分空间也已发展成优势种。
图1 研究区域。
Fig. 1 The study area.
上述植物中, 海三棱藨草是中国特有的盐沼植物, 也是典型的潮滩先锋物种, 目前主要分布于长江口沿岸的滩涂湿地(Sun et al ., 2002 , 2003 ; 梅雪英和张修峰, 2007 ; 陈秀芝和孙瑛, 2011 )。崇明东滩作为长江口最大的天然湿地, 有大量的海三棱藨草分布(虞快, 1994 , 1996 ), 但近年来崇明东滩北部互花米草生长泛滥, 海三棱藨草原有生态位受到挤压, 已基本被互花米草取代(陈秀芝和孙瑛, 2011 ), 东旺沙中间(2014年新围垦大堤东南部外围)是海三棱藨草现存种群主要分布区, 同时也是莎草科类群的主要分布区1) (1) 吴纪华 (2014). 上海崇明东滩鸟类国家级自然保护区2012年度资源监测公报, 14-15.) 。海三棱藨草在崇明东滩形成湿地草场, 能为鸻鹬类、雁鸭类、鹤类、鸥类等众多越冬候鸟提供天然的觅食、活动、休憩场所, 同时海三棱藨草的根茎和球茎, 还是国家一级保护鸟类——白头鹤的重要饵料(Ma et al ., 2003 ; Zhu et al ., 2007 ), 因此, 海三棱藨草对崇明东滩所有滨海湿地鸟类都显得至关重要。然而近年来滩涂围垦、堤外放牧及外来物种竞争等导致植物种群分布状况发生变化, 植物群落斑块破碎化严重, 整个生态系统服务功能也随之降低(张利权和雍学葵, 1992; Chen et al ., 2004; 陈中义等, 2005b; Wang et al ., 2010)。因此, 确定包括海三棱藨草在内的关键植物种群在东滩的空间分布, 明确其随环境因子的变化规律, 对该湿地生态系统的修复和保护具有重要的指导意义。
在前人相关研究(Ma et al ., 2003 ; 陈中义等, 2005a ; 贺强等, 2009 ; 詹跃武等, 2011 )的基础上, 本研究选择潮滩高程和表层30 cm土壤盐度作为主要参数, 探讨盐沼植物在东滩南部的空间分布及其与两大环境因子的相互关系。
1 研究区域和方法
1.1 自然概况
崇明东滩是长江口典型淤涨型潮滩湿地, 位于崇明岛最东端(121.83°-122.08° E, 31.42°-31.63° N), 属北亚热带季风气候, 四季分明, 日照充分, 雨量充沛, 气候温暖湿润, 年平均气温15.3 ℃, 年降水量1022 mm。这里是东亚-澳大利西亚迁徙路线上鸟类重要的停歇地, 也是小天鹅、白头鹤和黑脸琵鹭等珍稀濒危水禽重要的越冬栖息地(黄正一等, 1993 )。1997年崇明东滩被正式列入东亚-澳大利西亚涉禽保护区网络, 2002年又被《湿地公约》组织列入国际重要湿地名录(徐宏发和赵云龙, 2005 ), 2005年晋升为国家级自然保护区。本文研究区位于崇明东滩鸟类国家级自然保护区南部, 团结沙和东旺沙之间(121.90°-121.99° E, 31.45°-31.52° N), 北以互花米草生态治理工程2014年新筑大堤为界, 西至98大堤, 东、南两侧到达长江口外水域, 位于保护区的核心部分, 主要由团结沙外滩、大潮沟以及东旺沙外滩等植被覆盖的地区组成, 总面积约1800 hm2 (图1 )。
长江入海径流以崇明岛为界, 形成南、北两个一级支流, 两侧入海淡水不均衡, 南支淡水充裕, 北支偏少, 加之涨潮时北支经常出现咸水倒灌(贺松林等, 2006 ), 因此, 崇明东滩南部水体盐度明显低于北部。受潮汐影响, 海水与淡水在混合过程中又形成了新的盐度梯度, 即低潮带盐度高于中、高潮滩(陈中义等, 2005a ; 贺松林等, 2006 ; 曹利利等, 2012 ; 王卿等, 2012 )。与此同时, 径流中所携带的大量泥沙在岛堤之外沉积并发育形成新的具有不同潮滩高程的滩地, 受潮汐影响, 滩面水流在涨、落潮过程中不断冲刷滩面, 而崇明东滩南部植被相对稀疏, 因此在南部潮滩上形成多条宽窄不一、错综复杂的潮沟(张修桂, 2005 ; 姜亦飞等, 2012 ; 刘建华等, 2012 ; 陈勇等, 2013 ), 这在一定程度上限制了人类活动对保护区的影响。
1.2 研究方法
2013年, 基于GoogleTM Earth地图全景模式, 对研究区进行网格化处理——以90 m × 90 m为单元对研究区划分, 并把网格结点作为采样观测点, 导入全球定位系统GPS用以指导野外调查工作。
1.2.1 盐度数据的测定
2013年1、2月, 在研究区东西方向设立6条平行采样带, 样带间距800-1000 m, 每条采样带设置4-7个采样点, 样点距离400-600 m。利用直径5 cm、长30 cm的柱状采泥器垂直地面进行采样, 每个采样点采集3组样品作为平行样, 将样品用自封袋封装好, 做好相应记录。土壤样置于实验室内40 ℃烘箱内恒温烘干至恒重, 挑出样品中的植物根茎等杂质再对土样进行研磨, 过100目的不锈钢网筛, 采用四分法将土壤样品混合均匀, 取适量土样和超纯水按照1:5质量比进行溶解, 经搅拌充分混合均匀, 再用DDS-11A电导仪(DDS-11A, 上海雷磁创益仪器仪表有限公司, 上海)测量上层析出液的电导率, 并在处理过的样品中随机选择20个, 测定50 g样品析出液烘干后所得晶体的质量。
1.2.2 高程数据的测定
2013年8-10月, 选择WGS-84坐标作为参考坐标, 在崇明东滩基准点处架设Trimble RTK (Trimble-R8, Trimble Navigation Limited, Sunnyvale, USA)的基准站, 校准后, 携Trimble RTK移动端在采样点附近利用断面法测量高程, 待数据稳定后记录并保存。同时架设激光反射柱, 利用Trimble RTK对其一一进行位置和高程测量, 再利用Riegl VZ-4000 (Riegl VZ-4000, RIEGL Laser Measurement Systems GmbH, Salzburg, Austria)进行地形扫描。
1.2.3 植被数据的测定
2013年8-11月, 进行植被调查以及植物样本的采集, 在预先设置的2084个采样点附近随机放置100 cm × 100 cm的样方框。记录框内植物种类, 估测其投影面积与样方框面积的比值, 并将该值记为该植物地表盖度。植物地表部分利用剪刀采集, 每种植物随机选取10株, 测量其高度, 取平均值记为植物高度, 研究区内每间隔10-15个样点选取典型植物样本装入自封袋, 并记录其详细信息。
1.3 数据分析
将采集的数据统计、归类后, 导入ESRI ArcGISTM 10.0软件。依据植物空间分布状况, 在研究区划分出各种群分布范围, 并计算各自的分布面积。然后, 利用克里金(Kriging)空间插值法, 对分布面积最大的6个物种进行盖度插值, 经掩膜提取(extract by mask)后, 得出各物种在植物分布区内盖度变化的趋势图。同时, 通过比较同一栅格中不同物种的盖度, 划出优势植被分布区。划分过程中, 根据每种植物不同生长特征给出相应的划分标准(表1 ), 满足条件的部分即划入优势植被区。在同一栅格中植物总盖度值低于5%的记为光滩, 若在同一栅格中同时出现多种植物, 则选择盖度值最大的代表该栅格物种属性。
土壤盐度可通过电导率换算成土壤全盐含量: 首先对电导率数据和全盐含量进行线性回归分析, 得出回归方程S = 0.05s + 0.011 (R 2 = 0.733, p < 0.01, S 表示全盐含量, 单位: g·kg-1 , s 表示电导率, 单位: ms·cm-1 ), 再根据DDS-11A电导仪所测得的电导率计算各样品的土壤全盐含量。
有植被覆盖的部分, Riegl VZ4000激光扫描仪会同时接收到植物的反射, 无法直接生成数字高程图。因此, 利用克里金(Kriging)空间插值对植物高度进行插值处理, 再将扫描数据处理后导入ESRI ArcGISTM 10.0进行植被剔除, 得到潮滩高程数值, 再利用RTK数据对栅格数据进行检验。由于扫描数据是以点云模式采集, 扫描得到的数据能够更好地反映地形的变化趋势。植被分类后分别提取高程值和盐度数据, 并利用SPSS 18.0进行数据统计, 土壤盐度和潮滩高程分别符合正偏态和负偏态分布, 同时利用单因素方差分析(one-way ANOVA)对高程和土壤全盐含量进行环境因子差异显著性多重比较(Duncan法), 并利用OriginPro8.0作图, 探讨植物类型与其分布区潮滩高程、土壤盐度的关系。
2 结果
2.1 崇明东滩南部主要植物种群的空间分布
本次调查共发现植物17种, 隶属于7科16属, 其中禾本科7种, 莎草科5种, 香蒲科、菊科、藜科、雨久花科、苋科各1种, 植物在研究区内平均盖度高达85%。其中, 除互花米草、喜旱莲子草(Alternanthera philoxeroides )是外来物种外, 其余都是土著物种。所调查的植物中, 禾本科的芦苇、互花米草、白茅和莎草科的糙叶薹草、藨草、海三棱藨草等6种植物, 分布面积和盖度都占较大优势, 因此将它们作为本文研究的主要对象, 各物种在研究区的详细分布情况见图2 。
图2 主要盐沼植物种群分布。A, 芦苇。B, 白茅。C, 互花米草。D, 糙叶薹草。E, 藨草。F, 海三棱藨草。
Fig. 2 The distribution of major salt marsh species. A, Phragmites australis . B, Imperata cylindrica . C, Spartina alterniflora . D, Carex scabrifolia . E, Scirpus triqueter . F, Scirpus mariqueter.
为便于对植物分布进行研究, 在由陆地向近海方向上, 将研究区域划分为两个植被带, 距离大堤1.3 km范围内记为“内带”, 1.3 km到光滩之间记为“外带”。禾本科植物主要分布在研究区北部和“内带”, 莎草科主要分布在南部和“外带”, 禾本科平均盖度(50%)高于莎草科(36%), 因此研究区北部植物盖度显著高于南部, “内带”大于“外带”。禾本科植物(图2A , 2B, 2C)除互花米草(图2C )外, 主要集中分布在离大堤较近的“内带”, 莎草科植物除糙叶薹草(图2D )分布广泛外, 其余两种则分布在“外带” (图2E , 2F)。芦苇(图2A )和糙叶薹草(图2D )两大优势物种几乎遍及研究区, 两者是禾本科、莎草科植物中分布面积最大的物种。芦苇在大堤附近盖度最大, 可达96%, 往光滩方向逐渐减小, 南部盖度最低仅为3.5%。糙叶薹草在“外带”区盖度达到最大值65%, 往“内带”方向植被盖度值逐渐降低, 在98大堤附近盖度值最低(6%)。芦苇在研究区内平均盖度值为48%, 远高于糙叶薹草的32%。
禾本科中另一种植物——互花米草, 则集中分布在“外带”, 主要分布在研究区东北部, 种群外延毗邻光滩及海三棱藨草, 互花米草占禾本科植物总分布面积的20%, 远远低于芦苇, 但其种群平均盖度却高达62%, 种群中心区盖度极高, 一般为100%, 在3种禾本科植物中最高(图2C )。白茅(图2B )种群斑块多、面积小, 平均盖度偏低, 仅为40%, 而分布于大堤附近的白茅盖度却高达70%。芦苇、白茅、互花米草虽然都属于禾本科, 但在空间分布格局上存在很大差异。芦苇种群均匀分布; 互花米草植株密集, 并且个体均匀, 属于聚集型分布; 而白茅种群植株是典型的随机分布。
藨草(图2E )和海三棱藨草(图2F )盖度最大值都出现在各自分布区中部, 分别为73%和90%, 而最小盖度值出现在种群边缘, 分别为7%和8%, 海三棱藨草在莎草科中平均盖度最大达46%。作为先锋植物, 莎草科植物在演替初期成片状占据光滩水域, 植被盖度值较低, 为20%-35%, 但群落斑块之间连接度高、渗透性好, 相对于“内带”, 光滩区域人类活动少、影响小、干扰少, 仅受潮汐等自然因素影响, 因此莎草科植物能够均匀分布, 并且斑块之间逐渐连接, 成条带状。在群落相对稳定时, 群落中心植被总盖度值最大可达90%。而演替后期大部分空间被禾本科逐步占领, 即“内带”莎草科植物多以斑块形式随机分布, 盖度维持在24%-41%。整体而言, 莎草科中藨草和海三棱藨草集中在“外带”, 植株均匀分布, “内带”数量稀少, 呈随机分布, 并且两种群中间盖度高, 边缘盖度低; 糙叶薹草无论是“内带”还是“外带”都呈现典型的均匀分布, 但“外带”植被盖度显著高于“内带”, 莎草科植物的空间分布类型与群落发育时间关系密切(王卿等, 2012 )。
值得关注的是, 海三棱藨草(图2F )作为中国特有盐沼植物, 其种群面积与前人研究结果(黄华梅等, 2007 ; Yuan & Zhang, 2008 )存在较大差异。此外, 作为特有先锋物种, 海三棱藨草分布在“外带”, 但优势带也与互花米草存在较大的重叠空间, 两者存在显著的竞争关系, 然而互花米草在高度、盖度、地上生物量、地下生物量方面都比海三棱藨草具有明显优势, 为其获取更多光照、营养盐, 快速取代海三棱藨草创造了条件(Chen et al ., 2004 ; 陈中义等, 2005b , He et al ., 2011 ), 因此, 两种植物的分布存在交错带对海三棱藨草种群的发展极其不利。
2.2 优势植物种群分布
崇明东滩南部植物优势种群大多呈条带状分布, 部分较小的种群斑块会随机分布在光滩乃至其他植物群落中。例如, 芦苇(图3 )作为一个主要优势种, 集中分布于2014年新筑大堤以南1 km空间范围内, 其在大潮沟至东旺沙形成了一个长约2.4 km、宽约1.5 km的矩形分布区, 芦苇因盖度优势成为优势种。沿98大堤东向0.4-0.7 km范围内, 芦苇也呈显著的条带状分布。此外, 散布在研究区内的其他芦苇斑块, 有的虽然通过狭窄的廊道相互连接, 但盖度值相对偏低, 因此不能成为优势种。
图3 崇明东滩南部优势盐沼植物分布图。
Fig. 3 The distribution of dominant salt marsh species in southern Chongming Dongtan.
研究区东北部的互花米草也因较高的盖度值成为优势种, 同时, 无性繁殖方式, 使得种群扩散速度快、发育周期短, 并能够以母体为中心快速向周围空间扩散, 加之互花米草萌发时间早, 植物生长迅速, 抑制相邻其他植物的正常生长, 在其分布区内占有绝对优势(Li et al ., 2009 ; Lu & Zhang, 2013 ; Liu et al ., 2014 ; 徐伟伟等, 2014 )。研究区内互花米草与糙叶薹草、海三棱藨草、芦苇的邻接带分别长达1.2 km、2.0 km、3.0 km (图3 ), 这为互花米草向周围快速扩散提供了可能。与芦苇和互花米草不同, 海三棱藨草种群破碎化明显, 虽然外侧与光滩的边界长达2.4 km, 为种群向光滩扩张创造了良好的演替条件, 但其群落中部被芦苇和互花米草等植物所隔离, 外部受潮汐以及波浪作用明显, 因此生长条件比其他物种更为恶劣, 种群优势因此降低。其余3种植物空间分布形式各不相同, 白茅空间分布不均, 种群连接度不高, 斑块面积小, 但是数量较多, 因此种群总面积能占一定优势。糙叶薹草种群连接度高, 种群斑块面积大, 分布广, 能够承受较高的环境风险。藨草虽然在“外带”附近种群分布连续, 且面积较大, 但夹在糙叶薹草内部的藨草种群面积小, 较为分散。所有植物优势种群在东北至西南方向上都呈明显的条带状分布。
进一步分析6种主要植物在研究区内的分布面积与优势面积关系, 发现植物优势面积与其分布面积之比约为1:2 (图4 ), 禾本科植物的分布面积以及优势面积显著大于莎草科, 面积由大到小依次为: 芦苇>糙叶薹草>互花米草>藨草>白茅>海三棱藨草。研究中划定的崇明东滩传统三大优势种群芦苇、互花米草、海三棱藨草, 在本研究区目前的分布面积分别为1298 hm2 、422 hm2 、294 hm2 , 优势面积仅为700 hm2 、200 hm2 、120 hm2 , 而以前未受重视, 常被划入上述优势种群的糙叶薹草、藨草和白茅, 优势面积分别达到454 hm2 、189 hm2 、132 hm2 。海三棱藨草在6种植物中, 分布面积最小, 仅占研究区域总面积的6.9%, 同时也仅占莎草科植物的15.7%, 远低于前人报道的优势面积2091 hm2 (陈晖等, 2009; 黄华梅, 2009)。
图4 6种主要植物种群分布与优势面积对比。
Fig. 4 Comparison of total area and dominant area for six species.
2.3 环境因子与植被分布
研究区内, 潮滩高程介于-1.22-7.15 m之间(图5A ), 总体呈西高东低, 东旺沙附近地势较高, 在3.50-7.15 m之间, 在东旺沙中心部分出现高程最大值, 大潮沟南部至团结沙区, 介于2.30-5.10 m之间。长江口咸、淡水交互作用, 在崇明东滩外部区域形成了天然盐度梯度变化(陈中义等, 2005a ; 詹跃武等, 2011 ), 研究区内土壤盐度范围介于1.60-4.42 g·kg-1 , 并且空间上具有明显的变化规律——东北高、西南低, 因此土壤盐度最大值出现在东北部, 达到4.42 g·kg-1 。
图5 潮滩高程(A)和土壤盐度(B)分布。
Fig. 5 Distribution of tidal flat elevation (A) and soil salinity (B).
为明确植被与潮滩高程因子之间的关系, 我们对各植物类群分别进行显著性差异分析, 发现除了白茅和芦苇, 藨草和海三棱藨草两两之间无显著性差异外, 其余各植物群落之间差异非常显著(p < 0.01)。各物种相应高程分别为白茅(3.85 ± 0.32 m)≈芦苇(3.75 ± 0.57 m) >糙叶薹草(3.64 ± 0.48 m) >互花米草(3.55 ± 0.73 m) >藨草(3.26 ± 0.57 m)≈海三棱藨草(3.25 ± 0.72 m)(图6 )。自然状态下, 植被分布区潮滩高程与群落演替发育的时间存在密切联系(王卿等, 2012 ), 潮滩原生演替初期, 莎草科植物作为先锋植物首先出现在高程较低的光滩, 各植物种群发育成熟后, 变成相对稳定的莎草科植物群落。成熟的草滩具有明显的促淤沉降功能, 加速潮滩高程演变, 伴随着草滩“内带”泥沙淤积程度不断加深, 潮滩高程不断增高, 由此禾本科逐步占领莎草科生存空间(Wang et al ., 2010 ; 曹浩冰等, 2014 )。总之, 植被的出现加快高程的抬升, 高程的增加又促进植物群落的演替, 但禾本科分布区的潮滩高程始终普遍高于莎草科。
图6 不同植物分布区潮滩高程和土壤盐度差异性比较(平均值±标准偏差)。不同字母表示显著差异(p < 0.01)。大写字母代表高程, 小写字母代表盐度。
Fig. 6 Changes in elevation and soil salinity with vegetation types (mean ± SD ). Different letters represent significant differences (p < 0.01). Capital letters represent elevation, while lowercase letters represent salinity.
潮滩高程固然是划分植被分布范围的重要参考依据, 但植被空间分布同时受到其他环境因子的影响, 盐度就是其中一个重要的限定因子。植物对盐度的变化非常敏感, 盐沼植物对于盐度虽已形成一系列的响应和适应机制, 但盐度依然是盐沼植物生长过程中重要的影响因素(陈中义等, 2005a ; Zhang et al ., 2011 )。因为潮汐的不连续性, 加之长江口水体盐度较土壤盐度波动频繁, 因此选择土壤盐度为研究对象。从图6 可知, 互花米草分布区的土壤盐度最高, 为(3.21 ± 0.51 g·kg-1 ), 白茅区盐度值最低, 为(1.72 ± 0.15 g·kg-1 ), 另外4种主要植物的土壤盐度分别为: 海三棱藨草(2.86 ± 0.64 g·kg-1 ) >芦苇(2.12 ± 0.57 g·kg-1 )≈藨草(2.10 ± 0.37 g·kg-1 ) >糙叶薹草(1.90 ± 0.26 g·kg-1 ), 除了芦苇和藨草, 其余各类群植物土壤盐度差异性非常显著(p < 0.01), 因此土壤盐度在空间的变化必然影响甚至决定了各植物类群间的分布状况。
综合考虑高程和土壤盐度两大环境变量, 我们发现白茅群落在本研究区内滩面高程显著高于其他植被区, 而土壤盐度又显著低于其他植物群落, 白茅群落对两种环境因子的选择分别处于极大值和极小值, 能够同时满足其生长所需高程和盐度的范围有限, 因此, 白茅群落分布面积较小, 在6种植物中位于第5位。芦苇群落的滩面高程虽然仅次于白茅, 并且两者滩面高程无显著性差异, 但芦苇、白茅两者的土壤盐度却存在非常显著的差异(p < 0.01), 芦苇区的盐度适应范围在所有植物中处于中间值, 并且变幅较大, 因此能够适应更大范围生境, 分布面积最大。由此可以推断, 土壤盐度在芦苇和白茅两种植物分布上起着关键作用。糙叶薹草群落的滩面高程仅次于芦苇, 盐度也仅次于芦苇, 与芦苇比较, 其两大环境因子都存在非常显著的差异(p < 0.01), 糙叶薹草对两种环境变量的适应范围都处于中间水平, 因此, 糙叶薹草也可以广泛分布于研究区内。
海三棱藨草盐度耐受性次于互花米草, 同时两者潮滩高程存在显著差异(p < 0.01), 但两种植物对两大环境因子的适应范围相近(图6 ), 因此, 两种植物分布区的环境十分近似, 导致两者存在较大面积的交错分布。而在植被交错区内, 互花米草和海三棱藨草将形成激烈的竞争关系, 争夺种群发展的自然资源。
另一方面, 芦苇与互花米草的交错边界最长, 但是两者在高程以及盐度上都存在非常显著的差异(p < 0.01), 并且区间偏离较大, 因此互花米草不具备向芦苇种群扩张的优势。与海三棱藨草相比, 互花米草能够适应更高的高程和土壤盐度, 海三棱藨草-互花米草植被交错区往大堤方向高程变高, 反而适合互花米草生长; 交错带偏海三棱藨草一侧土壤盐度逐渐升高, 限制了海三棱藨草进一步扩张, 而在交错区内, 互花米草通过根茎、分蘖等繁殖方式, 争夺空间资源, 在植株高度、盖度等方面的优势限制了海三棱藨草生长(Chen et al ., 2004 ; Wang et al ., 2010 )。无论在交错区内部, 还是在与交错带毗邻的缓冲空间, 互花米草在高程和土壤盐度的综合作用下, 生长状况会更加良好, 而海三棱藨草很可能会被互花米草逐步取代。
3 讨论
崇明东滩属于典型的淤涨型潮滩, 目前滩涂植被覆盖区总面积4265 hm2 , 2012年调查数据显示, 崇明东滩中部、北部植物以芦苇和互花米草为主, 而莎草科植物主要集中分布在崇明东滩南部1) ( 1) 吴纪华 (2014). 上海崇明东滩鸟类国家级自然保护区2012年度资源监测公报, 14-15.) 。在崇明东滩的相关研究中, 众多学者根据研究需要将崇明东滩的海三棱藨草、藨草、糙叶薹草3种植物统一归入海三棱藨草类群(黄华梅等, 2007 ; Yuan & Zhang, 2008 ; 严格等, 2014 )。部分研究虽然将海三棱藨草、藨草、糙叶薹草统一划分为莎草科植物群落或者藨草群落(曹浩冰等, 2014 ; 任璘婧等, 2014 ), 但没有对这3种植物进行细致的划分, 各种群面积大小及其具体分布目前尚无相关报道。上述3种植物虽然同属于莎草科, 但占据不同的生态位, 承担着不同的生态功能, 例如在崇明东滩三者虽然都可以作为湿地鸟类的饵料, 但不同的鸟类会取食不同植物的不同部分(Ma et al ., 2003 ; Zhu et al ., 2007 ), 且3种植物组织、器官发育和成熟时间也各不相同。本文基于崇明东滩南部各植物种群的细致调查, 明确了各主要植物种群的空间分布以及各自的面积, 为进一步研究植被分布格局变化以及重要鸟类觅食生境提供基础。
潮滩植物群落在空间上呈现不同的分布状况, 潮滩高程作为影响植物分布的重要环境因子, 与植物的淹水时间和频度, 水体中的溶解氧, 土壤粒径、养分、氧化还原电位、pH值等生境因子存在密切联系(Silvestri et al ., 2005 ; 何彦龙, 2014 ), 与此同时, 植物的密度、高度等因素也会影响高程的变化(Duarte et al ., 2013 )。本研究表明崇明东滩南部植被覆盖空间6种主要植物分布在4个差异显著的高程区间内(p < 0.01), 黄华梅等(2007)和严格等(2014)认为崇明东滩芦苇分布在高潮带(高程>2.9 m), 海三棱藨草分布在中潮带(高程2.0-2.9 m), 互花米草在中、高潮带都有分布。同时对比何彦龙等(2010)在2008年对崇明东滩互花米草(3.21 ± 0.01 m)和海三棱藨草(3.10 ± 0.02 m)的研究结果, 发现上述植物在高程数值上有所差异, 但各植物种群生长空间的高程相对大小保持稳定, 表明在崇明东滩无论是大尺度区域范围, 还是小尺度空间, 各主要植物类群的潮滩高程相对高度保持恒定, 因此潮滩高程是决定植被空间分布的重要因素, 而具体数值差异可能是由于野外调查的时间、地点以及高程仪器精度差异所致。盐度是植物生长过程中的又一重要条件性因子, 不同的植物在不同的生长阶段适应不同范围的盐度(Zhang et al ., 2011 )。陈中义等(2005b)研究了水体中盐度的变化对互花米草和海三棱藨草的影响, 认为互花米草更适应高盐度海水。本研究中互花米草生长空间的土壤盐度确实显著高于海三棱藨草区(p < 0.01), 除了与互花米草本身的高耐盐特性有关, 与植株地上部分的泌盐作用也有一定关系(He et al ., 2011 )。但是无论崇明东滩土壤盐度还是潮滩高程都直接受到潮汐影响, 在空间上都呈现梯度变化(王卿等, 2012 ), 因此, 崇明东滩南部6种主要植物受到两种限定因子的共同制约, 在空间上形成现有的分布格局, 而对潮滩高程和土壤盐度在不同空间尺度下作用的权重仍有待进一步研究。
黄华梅等(2007) 认为, 影响崇明东滩植被分布格局的主要因素是互花米草入侵和围堤工程。但2014年互花米草治理工程实施后, 围堤外仍存在大面积的互花米草, 根据土壤盐度和潮滩高程的空间分布趋势, 互花米草具有向土壤盐度较高的东北方向扩张的可能, 广泛的高程适应范围又增加了其向南部扩张的几率, 在盐度和高程变化的影响下, 互花米草种群在发展过程中与海三棱藨草之间必然存在激烈的竞争关系, 这极有可能破坏该区植物群落的稳定现状。目前进行的互花米草生态治理工程虽未直接影响东滩南部的植被分布状况, 但是围堤之后, 大坝外围的水动力条件必然发生改变, 影响堤外泥沙淤积速率、土壤粒径等环境因子, 进而影响南部潮滩高程和土壤盐度, 改变植被空间格局。但具体的变化过程和趋势以及由此造成的影响还需要更长时间尺度的监测和分析, 其中本地特有物种海三棱藨草的面积和分布变化尤其值得关注。
致谢 感谢河口海岸大楼414办公室全体成员在样品采集以及文章撰写过程中给予的帮助。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Ayetiguli , Zhao SH Zuo P Wang CH 2010 ). The classification of salt marsh vegetation for Chongming Dongtan Nature Reserve based on knowledge engineer.
Remote Sensing for Land & Resources , (1 ), 123 -126 .
[本文引用: 1]
(in Chinese) [阿也提古丽·斯迪克 , 赵书河 , 左平 , 王春红 (2010 ). 基于知识工程师的崇明岛东滩自然保护区盐沼植被分类研究
. 国土资源遥感 , (1 ), 123 -126 .]
[本文引用: 1]
[2]
Cao HB Ge ZM Zhu ZC Zhang LQ 2014 ). The expansion pattern of saltmarshes at Chongming Dongtan and its underlying mechanism.
Acta Ecologica Sinica , 34 , 3944 -3952 .
[本文引用: 3]
(in Chinese with English abstract) [曹浩冰 , 葛振鸣 , 祝振昌 , 张利权 (2014 ). 崇明东滩盐沼植被扩散格局及其形成机制
. 生态学报 , 34 , 3944 -3952 .]
[本文引用: 3]
[3]
Cao LL Tao JF Zhang CK Liu GP 2012 ). Characteristics of intrusion in dry season of the North branch of the Yangtze River estuary.
Yangtze River , 43 (S1 ), 90 -92 .
[本文引用: 1]
(in Chinese) [曹利利 , 陶建峰 , 张长宽 , 刘桂平 (2012 ). 长江河口枯季北支盐水倒灌特征分析
. 人民长江 , 43 (S1 ), 90 -92 .]
[本文引用: 1]
[4]
Chen H Liu M Hou LJ Xu SY Yan HM Lin X 2009 ). Distribution of biogenic silica in Scirpus mariqueter of Chongming eastern tidal flat and its seasonal changes.
China Environmental Science , 29 , 73 -77 .
(in Chinese with English abstract) [陈晖 , 刘敏 , 侯立军 , 许世远 , 闫惠敏 , 林啸 (2009 ). 崇明东滩海三棱藨草生物硅分布及季节变化
. 中国环境科学 , 29 , 73 -77 .]
[5]
Chen XZ Sun Y 2011 ). Chinese endemic salt marsh plant——bulrush (Scirpus mariqueter), current status of its conservation and utilization.
Wetland Science & Management , 7 (1 ), 60 -63 .
[本文引用: 2]
(in Chinese with English abstract) [陈秀芝 , 孙瑛 (2011 ). 中国特有盐沼植物——海三棱藨草的现状及保护利用
. 湿地科学与管理 , 7 (1 ), 60 -63 .]
[本文引用: 2]
[6]
Chen Y He ZF Li B Zhao BC 2013 ). Spatial distribution of tidal creeks and quantitative analysis of its driving factors in Chongming Dongtan, Shanghai. Journal of Jilin University (Earth Science Edition)
, 43 , 212 -219 .
[本文引用: 1]
(in Chinese with English abstract) [陈勇 , 何中发 , 黎兵 , 赵宝成 (2013 ). 崇明东滩潮沟发育特征及其影响因素定量分析
. 吉林大学学报 (地球科学版) , 43 , 212 -219 .]
[本文引用: 1]
[7]
Chen ZY Li B Chen JK 2005a ). Effects of salt stress and elevation of tideland on the growth of introduced Spartina alerniflora at Dongtan of Chongming, the Yangtze River Estuary. Journal of Yangtze University (
Nature Science Edition ), 2 (2 ), 6 -9 .
[本文引用: 4]
(in Chinese with English abstract) [陈中义 , 李博 , 陈家宽 (2005a ). 长江口崇明东滩土壤盐度和潮间带高程对外来种互花米草生长的影响
. 长江大学学报(自然科学版) , 2 (2 ), 6 -9 .]
[本文引用: 4]
[8]
Chen ZY Li B Chen JK 2005b ). Some growth characteristics and relative competitive ability of invasive Spartina alterniflora and native Scirpus mariqueter.
Biodiversity Science , 13 , 130 -136 .
[本文引用: 1]
(in Chinese with English abstract) [陈中义 , 李博 , 陈家宽 (2005b ). 互花米草与海三棱藨草的生长特征和相对竞争能力
. 生物多样性 , 13 , 130 -136 .]
[本文引用: 1]
[9]
Chen ZY Li B Yang Z Chen JK 2004 ). Local competitive effects of introduced Spartina alterniflora on Scirpus mariqueter at Dongtan of Chongming Island, the Yangtze River estuary and their potential ecological consequences.
Hydrobiologia , 528 , 99 -106 .
[本文引用: 2]
[10]
Duarte CM Losada IJ Hendriks IE Mazarrasa I Marbà N 2013 ). The role of coastal plant communities for climate change mitigation and adaptation.
Nature Climate Change , 3 , 961 -968 .
[本文引用: 2]
[11]
Gao Y Zhao B 2006 ). The effect of reclamation on mud flat development in Chongming Island, Shanghai.
Chinese Agricultural Science Bulletin , 22 , 475 -479 .
[本文引用: 2]
(in Chinese with English abstract) [高宇 , 赵斌 (2006 ). 人类围垦活动对上海崇明东滩滩涂发育的影响
. 中国农学通报 , 22 , 475 -479 .]
[本文引用: 2]
[12]
Gao ZG Zhang LQ 2006 ). Multi-seasonal spectral characteristics analysis of coastal salt marsh vegetation in Shanghai, China.
Estuarine, Coastal and Shelf Science , 69 , 217 -224 .
[13]
He Q Cui BS Zhao XS Fu HL Liao XL 2009 ). Relationships between salt marsh vegetation distribution/diversity and soil chemical factors in the Yellow River Estuary, China.
Acta Ecologica Sinica , 29 , 676 -687 .
[本文引用: 1]
(in Chinese with English abstract) [贺强 , 崔保山 , 赵欣胜 , 付华龄 , 廖晓琳 (2009 ). 黄河河口盐沼植被分布、多样性与土壤化学因子的相关关系
. 生态学报 , 29 , 676 -687 .]
[本文引用: 1]
[14]
He SL Ding PX Kong YZ 2006 ). Saltwater intrusion of the North branch and salinity variability of the South branch river of the Yangtze River in dry season.
Progress in Natural Science , 16 , 584 -589 .
[本文引用: 2]
(in Chinese) [贺松林 , 丁平兴 , 孔亚珍 (2006 ). 长江口南支河段枯季盐度变异与北支咸水倒灌
. 自然科学进展 , 16 , 584 -589 .]
[本文引用: 2]
[15]
He YL 2014 ). The Mechanism of Vegetation Differentiation in the Lower Salt Marsh of Yangtze River Estuary.
PhD dissertation, East China Normal University, Shanghai . 6-7, 33 , 96 -97 .
[本文引用: 1]
(in Chinese with English abstract) [何彦龙 (2014 ). 中低潮滩盐沼植被分异的形成机制研究
. 博士学位论文, 华东师范大学, 上海 . 6-7, 33 , 96 -97 .]
[本文引用: 1]
[16]
He YL Li XZ Craft C Ma ZG Sun YG 2011 ). Relationships between vegetation zonation and environmental factors in newly formed tidal marshes of the Yangtze River estuary.
Wetlands Ecology and Management , 19 , 341 -349 .
[本文引用: 2]
[17]
He YL Li XZ Ma ZG Sun YG Jia Y 2010 ). Vegetation zonation related to the edaphic factors in the East headland of Chongming Island.
Acta Ecologica Sinica , 30 , 4919 -4927 .
(in Chinese with English abstract) [何彦龙 , 李秀珍 , 马志刚 , 孙永光 , 贾悦 (2010 ). 崇明东滩盐沼植被成带性对土壤因子的响应
. 生态学报 , 30 , 4919 -4927 .]
[18]
Huang HM 2009 ). A Research on Spatial-Temporal Dynamics of Salt Marsh Vegetation at the Intertidal Zone in Shanghai.
PhD dissertation, East China Normal University, Shanghai . I-III , 85 -91 .
(in Chinese with English abstract) [黄华梅 (2009 ). 上海滩涂盐沼植被的分布格局和时空动态研究
. 博士学位论文, 华东师范大学, 上海 . I-III , 85 -91 .]
[19]
Huang HM Zhang LQ Yuan L 2007 ). The spatio-temporal dynamics of salt marsh vegetation for Chongming Dongtan National Nature Reserve, Shanghai.
Acta Ecologica Sinica , 27 , 4166 -4172 .
[本文引用: 4]
(in Chinese with English abstract) [黄华梅 , 张利权 , 袁琳 (2007 ). 崇明东滩自然保护区盐沼植被的时空动态
. 生态学报 , 27 , 4166 -4172 .]
[本文引用: 4]
[20]
Huang ZY Sun ZH Yu K 1993 ). Bird Resources and Habitats in Shanghai . Fudan University Press , Shanghai .
[本文引用: 1]
(in Chinese) [黄正一 , 孙振华 , 虞快 (1993 ). 上海鸟类资源及其生境 . 复旦大学出版社 , 上海 .]
[本文引用: 1]
[21]
Hughes AR 2014 ). Genotypic diversity and trait variance interact to affect marsh plant performance.
Journal of Ecology , 102 , 651 -658 .
[本文引用: 1]
[22]
Isacch JP Cardoni DA Ieibaren OO 2014 ). Diversity and habitat distribution of birds in coastal marshes and comparisons with surrounding upland habitats in Southeastern South America.
Estuaries and Coasts , 37 , 229 -239 .
[本文引用: 1]
[23]
Jiang YF Du JZ Zhang J Zhang WX Zhang J 2012 ). The determination of sedimentation rates in various vegetational zones of Chongming tidal flat of the Changjiang Estuary.
Acta Oceanologica Sinica , 34 , 114 -121 .
[本文引用: 1]
(in Chinese with English abstract) [姜亦飞 , 杜金洲 , 张敬 , 张文祥 , 张经 (2012 ). 长江口崇明东滩不同植被带沉积速率研究
. 海洋学报 , 34 , 114 -121 .]
[本文引用: 1]
[24]
Li B Liao CH Zhang XD Chen HL Wang Q Chen ZY Gan XJ Wu JH Zhao B Ma ZJ Cheng XL Jiang LF Chen JK 2009 ). Spartina alterniflora invasions in the Yangtze River estuary, China: An overview of current status and ecosystem effects.
Ecological Engineering , 35 , 511 -520 .
[本文引用: 1]
[25]
Li H Zhang XM Zheng RH Li X Elmer WH Wolfe LM Li B 2014a) . Indirect effects of non-native Spartina alterniflora and its fungal pathogen (Fusarium palustre) on native saltmarsh plants in China.
Journal of Ecology , 102 , 1112 -1119 .
[本文引用: 1]
[26]
Li XZ Ren LJ Liu Y Christopher C Mander Ü Yang SL 2014b ). The impact of the change in vegetation structure on the ecological functions of salt marshes: The example of the Yangtze estuary.
Regional Environmental Change , 14 , 623 -632 .
[本文引用: 2]
[27]
Liu HY Lin ZS Qi XZ Zhang MY Yang H 2014 ). The relative importance of sexual and asexual reproduction in the spread of Spartina alterniflora using a spatially explicit individual-based model.
Ecological Research , 29 , 905 -915 .
[本文引用: 1]
[28]
Liu JH Yang SL Shi BW Luo XX Fu XK 2012 ). Topography and evolution of the tidal trench in the eastern Chongming tidal flat, Changjiang River Estuary.
Journal of Marine Sciences , 30 (2 ), 43 -50 .
[本文引用: 1]
(in Chinese with English abstract) [刘建华 , 杨世伦 , 史本伟 , 罗向欣 , 付信坤 (2012 ). 长江口崇明东滩潮沟地貌形态和演变
. 海洋学研究 , 30 (2 ), 43 -50 .]
[本文引用: 1]
[29]
Lu JB Zhang Y 2013 ). Spatial distribution of an invasive plant Spartina alterniflora and its potential as biofuels in China
. Ecological Engineering , 52 , 175 -181 .
[本文引用: 1]
[30]
Ma ZJ Li B Jing K Zhao B Tang SM Chen JK 2003 ). Effects of tidewater on the feeding ecology of hooded crane (Grus monacha) and conservation of their wintering habitats at Chongming Dongtan, China.
Ecological Research , 18 , 321 -329 .
[本文引用: 3]
[31]
Mei XY Zhang XF 2007 ). Carbon storage and fixation function of Scirpus mariqueter in Changjiang River Estuary: A case study of Chongming Dongtan wetland.
Journal of Agro-Environment Science , 26 , 360 -363 .
[本文引用: 1]
(in Chinese with English abstract) [梅雪英 , 张修峰 (2007 ). 长江口湿地海三棱藨草(Scirpus mariqueter)的储碳、固碳功能研究——以崇明东滩为例
. 农业环境科学学报 , 26 , 360 -363 .]
[本文引用: 1]
[32]
Ren LJ Li XZ Yang SL Yan ZZ Huang X 2014 ). The impact of salt marsh change on sediment accumulation and wave attenuation at the East Chongming Island.
Acta Ecologica Sinica , 34 , 3350 -3358 .
[本文引用: 1]
(in Chinese with English abstract) [任璘婧 , 李秀珍 , 杨世伦 , 闫中正 , 黄星 (2014 ). 崇明东滩盐沼植被变化对滩涂湿地促淤消浪功能的影响
. 生态学报 , 34 , 3350 -3358 .
[本文引用: 1]
[33]
Silvestri S Defina A Marani M 2005 ). Tidal regime, salinity and salt marsh plant zonation.
Estuarine, Coastal and Shelf Science , 62 , 119 -130 .
[本文引用: 1]
[34]
Sun SC Cai YL An SQ 2002 ). Differences in morphology and biomass allocation of Scirpus mariqueter between creekside and inland communities in the Changjiang estuary, China.
Wetlands , 22 , 786 -793 .
[本文引用: 1]
[35]
Sun SC Cai YL Tian XJ 2003 ). Salt marsh vegetation change after a short-term tidal restriction in the Changjiang estuary.
Wetlands , 23 , 257 -266 .
[本文引用: 1]
[36]
Tian B Zhou YX Zhang LQ Yuan L 2008 ). Analyzing the habitat suitability for migratory birds at the Chongming Dongtan Nature Reserve in Shanghai, China.
Estuarine Coastal and Shelf Science , 80 , 296 -302 .
[本文引用: 1]
[37]
Tong CF 2012 ). Characteristics of the fish assemblages in the intertidal salt marsh zone and adjacent mudflat in the Yangtze Estuary.
Acta Ecologica Sinica , 32 , 6501 -6510 .
[本文引用: 1]
(in Chinese with English abstract) [童春富 (2012 ). 长江河口潮间带盐沼植被分布区及邻近光滩鱼类组成特征
. 生态学报 , 32 , 6501 -6510 .]
[本文引用: 1]
[38]
Wang CH Lu M Yang B Yang Q Zhang XD Hara T Li B 2010 ). Effects of environmental gradients on the performances of four dominant plants in a Chinese saltmarsh: Implications for plant zonation.
Ecological Research , 25 , 347 -358 .
[本文引用: 2]
[39]
Wang Q Wang CH Huang SF Sha CY Ruan JJ Wang M 2012 ). Review on salt marsh plant communities: Distribution, succession and impact factors.
Ecology and Environmental Sciences , 21 , 375 -388 .
[本文引用: 4]
(in Chinese with English abstract) [王卿 , 汪承焕 , 黄沈发 , 沙晨燕 , 阮俊杰 , 王敏 (2012 ). 盐沼植物群落研究进展: 分布、演替及影响因子
. 生态环境学报 , 21 , 375 -388 .]
[本文引用: 4]
[40]
Xu HF Zhao YL 2005 ). Scientific Survey on Chongming Dongtan Migratory Birds Nature Reserve of Shanghai . China Forestry Publishing House , Beijing .
[本文引用: 1]
(in Chinese) [徐宏发 , 赵云龙 (2005 ). 上海市崇明东滩鸟类自然保护区科学考察集 . 中国林业出版社 , 北京 .]
[本文引用: 1]
[41]
Xu WW Wang GX Liu JE Chen ZY Hang ZQ Wang H 2014 ). Two reproductive mode of Spartina alterniflora on coastal wetland of North Jiangsu.
Acta Ecologica Sinica , 34 , 3839 -3847 .
[本文引用: 1]
(in Chinese with English abstract) [徐伟伟 , 王国祥 , 刘金娥 , 陈正勇 , 杭子清 , 王会 (2014 ). 苏北海滨湿地互花米草种群繁殖方式
. 生态学报 , 34 , 3839 -3847 .]
[本文引用: 1]
[42]
Yan G Ge ZM Zhang LQ 2014 ). Distribution of soil carbon storage in different saltmarsh plant communities in Chongming Dongtan wetland.
Chinese Journal of Applied Ecology , 25 , 85 -91 .
[本文引用: 1]
(in Chinese with English abstract) [严格 , 葛振鸣 , 张利权 (2014 ). 崇明东滩湿地不同盐沼植物群落土壤碳储量分布
. 应用生态学报 , 25 , 85 -91 .]
[本文引用: 1]
[43]
Yan Q Lu JJ He WS 2007 ). Succession character of salt marsh vegetations in Chongming Dongtan wetland.
Chinese Journal of Applied Ecology , 18 , 1097 -1101 .
[本文引用: 1]
(in Chinese with English abstract) [闫芊 , 陆健健 , 何文珊 (2007 ). 崇明东滩湿地高等植被演替特征
. 应用生态学报 , 18 , 1097 -1101 .]
[本文引用: 1]
[44]
Yu K 1994 ). Anseries and Scirpus mariqueter in Chongming Dongtan, Shanghai.
China Nature , (3 ), 7 -8 .
[本文引用: 1]
(in Chinese) [虞快 (1994 ). 崇明东滩的雁鸭与海三棱藨草
. 大自然 , (3 ), 7 -8 .]
[本文引用: 1]
[45]
Yu K 1996 ). An endangered species, Scirpus mariqueter.
China Nature , (3 ), 35 -36 .
[本文引用: 1]
(in Chinese) [虞快 (1996 ). 亟待保护的海三棱藨草
. 大自然 , (3 ), 35 -36 .]
[本文引用: 1]
[46]
Yuan L Zhang LQ 2008 ). Mapping large-scale distribution of submerged aquatic vegetation coverage using remote sensing.
Ecological Informatics , 3 , 245 -251 .
[本文引用: 2]
[47]
Zhai XY Yang SX 2013 ). The remote sensing monitoring study of saltmarsh vegetation in Chongming Dongtan Nature Reserve.
China Resources Comprehensive Utilization , 31 (7 ), 46 -48 .
[本文引用: 1]
(in Chinese) [翟新源 , 杨苏新 (2013 ). 崇明东滩湿地植被遥感监测研究
. 中国资源综合利用 , 31 (7 ), 46 -48 .]
[本文引用: 1]
[48]
Zhan YW He CQ Meng ZS Lei YR Zheng H 2011 ). Spatial and temporal distribution characteristics of Scripus mariqueter in salt soil environmental factors.
Marine Environmental Science , 30 , 765 -770 .
[本文引用: 2]
(in Chinese with English abstract) [詹跃武 , 何池全 , 孟昭寿 , 雷雁茹 , 郑虹 (2011 ). 海三棱藨草盐沼土壤环境因子的时空分布特征
. 海洋环境科学 , 30 , 765 -770 .]
[本文引用: 2]
[49]
Zhang LQ Yong XK 1992 ). Studies on phenology and spatial distribution pattern of Scirpus mariqueter population.
Acta Phytoecologica et Geobotanica Sinica , 16 , 43 -51 .
(in Chinese with English abstract) [张利权 , 雍学葵 (1992 ). 海三棱藨草种群的物候与分布格局研究
. 植物生态学与地植物学学报 , 16 , 43 -51 .]
[50]
Zhang TT Zeng SL Gao Y Ouyang ZT Li B Fang CM 2011 ). Using hyperspectral vegetation indices as a proxy to monitor soil salinity.
Ecological Indicators , 11 , 1552 -1562 .
[本文引用: 2]
[51]
Zhang XG 2005 ). The historical formation of Chongming Island. Fudan Journal (
Social Sciences ), (3 ), 57 -66 .
[本文引用: 1]
(in Chinese with English abstract) [张修桂 (2005 ). 崇明岛形成的历史过程
. 复旦学报(社会科学版) , (3 ), 57 -66 .]
[本文引用: 1]
[52]
Zhang ZY Ding CL Xiao M 2012 ). The diversity of invasive plant Spartina alterniflora rhizosphere bacteria in tidal salt marshes at Chongming Dongtan in the Yangtze River estuary.
Acta Ecologica Sinica , 32 , 6636 -6646 .
[本文引用: 1]
(in Chinese with English abstract) [章振亚 , 丁陈利 , 肖明 (2012 ). 崇明东滩湿地不同潮汐带入侵植物互花米草根际细菌的多样性
. 生态学报 , 32 , 6636 -6646 .]
[本文引用: 1]
[53]
Zheng ZS Zhou YX Li X Kuang RY 2010 ). Relationship between mudflat evolution dynamic and vegetation at Chongming Dongtan integrating with remote sensing and hydrodynamic model.
Resources and Environment in the Yangtze Basin , 19 , 1368 -1373 .
[本文引用: 1]
(in Chinese with English abstract) [郑宗生 , 周云轩 , 李行 , 况润元 (2010 ). 基于遥感及数值模拟的崇明东滩冲淤与植被关系探讨
. 长江流域资源与环境 , 19 , 1368 -1373 .]
[本文引用: 1]
[54]
Zhu J Jing K Gan XJ Ma ZJ 2007 ). Food supply in intertidal area for shorebirds during stopover at Chongming Dongtan, China
. Acta Ecologica Sinica , 27 , 2149 -2159 .
[本文引用: 2]
[55]
Zhu MS Sun T Shao DD 2014 ). Impact of land reclamation on the evolution of shoreline change and nearshore vegetation distribution in Yangtze River Estuary.
Wetlands, doi: 10. 1007/s13157-014-0610-6 .
[本文引用: 1]
[56]
Zong W Lin WP Zhou YX Rui JX 2011 ). Estimation of typical wetland vegetation NPP in Shanghai Chongming Dongtan based on remote sensing.
Resources and Environment in the Yangtze Basin , 20 , 1355 -1360 .
[本文引用: 1]
(in Chinese with English abstract) [宗玮 , 林文鹏 , 周云轩 , 芮建勋 (2011 ). 基于遥感的上海崇明东滩湿地典型植被净初级生产力估算
. 长江流域资源与环境 , 20 , 1355 -1360 .]
[本文引用: 1]
基于知识工程师的崇明岛东滩自然保护区盐沼植被分类研究
1
2010
... 近十几年来, 大量研究人员通过遥感影像解译, 对崇明东滩主要植被进行了分区(Gao & Zhang, 2006 ; Tian et al ., 2008 ; 阿也提古丽·斯迪克等, 2010 ; 翟新源和杨苏新, 2013 ; 郑宗生等, 2010 ; 宗玮等, 2011 ), 普遍认为芦苇(Phragmites australis )、互花米草(Spartina alterniflora )及海三棱藨草(Scirpus mariqueter )是崇明东滩的“三大”主要植物类群.然而, 崇明东滩南部长期受长江入海淡水影响, 水体盐度相对较低, 在团结沙至大潮沟(图1 )存在较大面积的藨草(Scirpus triqueter )种群区(闫芊等, 2007 ; 章振亚等, 2012 ).同时发现, 糙叶薹草(Carex scabrifolia )在研究区内频繁出现, 白茅(Imperata cylindrica )在部分空间也已发展成优势种. ...
基于知识工程师的崇明岛东滩自然保护区盐沼植被分类研究
1
2010
... 近十几年来, 大量研究人员通过遥感影像解译, 对崇明东滩主要植被进行了分区(Gao & Zhang, 2006 ; Tian et al ., 2008 ; 阿也提古丽·斯迪克等, 2010 ; 翟新源和杨苏新, 2013 ; 郑宗生等, 2010 ; 宗玮等, 2011 ), 普遍认为芦苇(Phragmites australis )、互花米草(Spartina alterniflora )及海三棱藨草(Scirpus mariqueter )是崇明东滩的“三大”主要植物类群.然而, 崇明东滩南部长期受长江入海淡水影响, 水体盐度相对较低, 在团结沙至大潮沟(图1 )存在较大面积的藨草(Scirpus triqueter )种群区(闫芊等, 2007 ; 章振亚等, 2012 ).同时发现, 糙叶薹草(Carex scabrifolia )在研究区内频繁出现, 白茅(Imperata cylindrica )在部分空间也已发展成优势种. ...
崇明东滩盐沼植被扩散格局及其形成机制
3
2014
... 盐沼植物作为全球范围内生产力较高的生物群落之一, 是推动湿地生态系统物质循环和能量流动的基础要素, 其在维持个体生存、种群稳定与群落发展的同时, 也为湿地生态系统中的众多成员提供了丰富的食物, 并创造了良好的栖息环境.然而, 近年来由于全球气候变化和频繁的人类活动, 植物群落组成结构以及空间分布格局发生了巨大的变化, 这不仅影响了湿地中鸟类、底栖生物及鱼类等生物的种群数量, 而且也不断改变着植物类群的种群结构, 甚至可能会导致一些敏感物种的灭绝(童春富, 2012 ; Isacch et al ., 2014 ; Li et al ., 2014b ).因此, 明确湿地中主要植物种群及其分布状况, 掌握其随环境因子的变化规律, 对湿地生态系统的保护、恢复、重建都具有重要的参考价值.虽然前人已就此进行了较多的研究(黄华梅等, 2007 ; Duarte et al ., 2013 ; 曹浩冰等, 2014 ; Hughes, 2014 ; Li et al ., 2014a , 2014b ), 但河口潮滩区域海陆交互作用强烈、人类活动频繁, 使得滩面、潮沟及植被都处于不断变化中, 崇明东滩正是这样一个处在发育过程中的典型盐沼湿地生态系统.2014年崇明东滩互花米草(Spartina alterniflora )生态治理工程的实施, 可能对该区域盐沼植物的空间分布及其生境产生了重大影响(高宇和赵斌, 2006 ; Zhu et al ., 2014 ).因此, 在施工前期对围垦外围区进行全面的植被调查, 不仅能准确地掌握崇明东滩南部的植被类型和现有分布状况, 而且能为今后研究其植被空间动态变化提供基础数据. ...
... 为明确植被与潮滩高程因子之间的关系, 我们对各植物类群分别进行显著性差异分析, 发现除了白茅和芦苇, 藨草和海三棱藨草两两之间无显著性差异外, 其余各植物群落之间差异非常显著(p < 0.01).各物种相应高程分别为白茅(3.85 ± 0.32 m)≈芦苇(3.75 ± 0.57 m) >糙叶薹草(3.64 ± 0.48 m) >互花米草(3.55 ± 0.73 m) >藨草(3.26 ± 0.57 m)≈海三棱藨草(3.25 ± 0.72 m)(图6 ).自然状态下, 植被分布区潮滩高程与群落演替发育的时间存在密切联系(王卿等, 2012 ), 潮滩原生演替初期, 莎草科植物作为先锋植物首先出现在高程较低的光滩, 各植物种群发育成熟后, 变成相对稳定的莎草科植物群落.成熟的草滩具有明显的促淤沉降功能, 加速潮滩高程演变, 伴随着草滩“内带”泥沙淤积程度不断加深, 潮滩高程不断增高, 由此禾本科逐步占领莎草科生存空间(Wang et al ., 2010 ; 曹浩冰等, 2014 ).总之, 植被的出现加快高程的抬升, 高程的增加又促进植物群落的演替, 但禾本科分布区的潮滩高程始终普遍高于莎草科. ...
... 崇明东滩属于典型的淤涨型潮滩, 目前滩涂植被覆盖区总面积4265 hm2 , 2012年调查数据显示, 崇明东滩中部、北部植物以芦苇和互花米草为主, 而莎草科植物主要集中分布在崇明东滩南部1) ( 1) 吴纪华 (2014). 上海崇明东滩鸟类国家级自然保护区2012年度资源监测公报, 14-15.) .在崇明东滩的相关研究中, 众多学者根据研究需要将崇明东滩的海三棱藨草、藨草、糙叶薹草3种植物统一归入海三棱藨草类群(黄华梅等, 2007 ; Yuan & Zhang, 2008 ; 严格等, 2014 ).部分研究虽然将海三棱藨草、藨草、糙叶薹草统一划分为莎草科植物群落或者藨草群落(曹浩冰等, 2014 ; 任璘婧等, 2014 ), 但没有对这3种植物进行细致的划分, 各种群面积大小及其具体分布目前尚无相关报道.上述3种植物虽然同属于莎草科, 但占据不同的生态位, 承担着不同的生态功能, 例如在崇明东滩三者虽然都可以作为湿地鸟类的饵料, 但不同的鸟类会取食不同植物的不同部分(Ma et al ., 2003 ; Zhu et al ., 2007 ), 且3种植物组织、器官发育和成熟时间也各不相同.本文基于崇明东滩南部各植物种群的细致调查, 明确了各主要植物种群的空间分布以及各自的面积, 为进一步研究植被分布格局变化以及重要鸟类觅食生境提供基础. ...
崇明东滩盐沼植被扩散格局及其形成机制
3
2014
... 盐沼植物作为全球范围内生产力较高的生物群落之一, 是推动湿地生态系统物质循环和能量流动的基础要素, 其在维持个体生存、种群稳定与群落发展的同时, 也为湿地生态系统中的众多成员提供了丰富的食物, 并创造了良好的栖息环境.然而, 近年来由于全球气候变化和频繁的人类活动, 植物群落组成结构以及空间分布格局发生了巨大的变化, 这不仅影响了湿地中鸟类、底栖生物及鱼类等生物的种群数量, 而且也不断改变着植物类群的种群结构, 甚至可能会导致一些敏感物种的灭绝(童春富, 2012 ; Isacch et al ., 2014 ; Li et al ., 2014b ).因此, 明确湿地中主要植物种群及其分布状况, 掌握其随环境因子的变化规律, 对湿地生态系统的保护、恢复、重建都具有重要的参考价值.虽然前人已就此进行了较多的研究(黄华梅等, 2007 ; Duarte et al ., 2013 ; 曹浩冰等, 2014 ; Hughes, 2014 ; Li et al ., 2014a , 2014b ), 但河口潮滩区域海陆交互作用强烈、人类活动频繁, 使得滩面、潮沟及植被都处于不断变化中, 崇明东滩正是这样一个处在发育过程中的典型盐沼湿地生态系统.2014年崇明东滩互花米草(Spartina alterniflora )生态治理工程的实施, 可能对该区域盐沼植物的空间分布及其生境产生了重大影响(高宇和赵斌, 2006 ; Zhu et al ., 2014 ).因此, 在施工前期对围垦外围区进行全面的植被调查, 不仅能准确地掌握崇明东滩南部的植被类型和现有分布状况, 而且能为今后研究其植被空间动态变化提供基础数据. ...
... 为明确植被与潮滩高程因子之间的关系, 我们对各植物类群分别进行显著性差异分析, 发现除了白茅和芦苇, 藨草和海三棱藨草两两之间无显著性差异外, 其余各植物群落之间差异非常显著(p < 0.01).各物种相应高程分别为白茅(3.85 ± 0.32 m)≈芦苇(3.75 ± 0.57 m) >糙叶薹草(3.64 ± 0.48 m) >互花米草(3.55 ± 0.73 m) >藨草(3.26 ± 0.57 m)≈海三棱藨草(3.25 ± 0.72 m)(图6 ).自然状态下, 植被分布区潮滩高程与群落演替发育的时间存在密切联系(王卿等, 2012 ), 潮滩原生演替初期, 莎草科植物作为先锋植物首先出现在高程较低的光滩, 各植物种群发育成熟后, 变成相对稳定的莎草科植物群落.成熟的草滩具有明显的促淤沉降功能, 加速潮滩高程演变, 伴随着草滩“内带”泥沙淤积程度不断加深, 潮滩高程不断增高, 由此禾本科逐步占领莎草科生存空间(Wang et al ., 2010 ; 曹浩冰等, 2014 ).总之, 植被的出现加快高程的抬升, 高程的增加又促进植物群落的演替, 但禾本科分布区的潮滩高程始终普遍高于莎草科. ...
... 崇明东滩属于典型的淤涨型潮滩, 目前滩涂植被覆盖区总面积4265 hm2 , 2012年调查数据显示, 崇明东滩中部、北部植物以芦苇和互花米草为主, 而莎草科植物主要集中分布在崇明东滩南部1) ( 1) 吴纪华 (2014). 上海崇明东滩鸟类国家级自然保护区2012年度资源监测公报, 14-15.) .在崇明东滩的相关研究中, 众多学者根据研究需要将崇明东滩的海三棱藨草、藨草、糙叶薹草3种植物统一归入海三棱藨草类群(黄华梅等, 2007 ; Yuan & Zhang, 2008 ; 严格等, 2014 ).部分研究虽然将海三棱藨草、藨草、糙叶薹草统一划分为莎草科植物群落或者藨草群落(曹浩冰等, 2014 ; 任璘婧等, 2014 ), 但没有对这3种植物进行细致的划分, 各种群面积大小及其具体分布目前尚无相关报道.上述3种植物虽然同属于莎草科, 但占据不同的生态位, 承担着不同的生态功能, 例如在崇明东滩三者虽然都可以作为湿地鸟类的饵料, 但不同的鸟类会取食不同植物的不同部分(Ma et al ., 2003 ; Zhu et al ., 2007 ), 且3种植物组织、器官发育和成熟时间也各不相同.本文基于崇明东滩南部各植物种群的细致调查, 明确了各主要植物种群的空间分布以及各自的面积, 为进一步研究植被分布格局变化以及重要鸟类觅食生境提供基础. ...
长江河口枯季北支盐水倒灌特征分析
1
2012
... 长江入海径流以崇明岛为界, 形成南、北两个一级支流, 两侧入海淡水不均衡, 南支淡水充裕, 北支偏少, 加之涨潮时北支经常出现咸水倒灌(贺松林等, 2006 ), 因此, 崇明东滩南部水体盐度明显低于北部.受潮汐影响, 海水与淡水在混合过程中又形成了新的盐度梯度, 即低潮带盐度高于中、高潮滩(陈中义等, 2005a ; 贺松林等, 2006 ; 曹利利等, 2012 ; 王卿等, 2012 ).与此同时, 径流中所携带的大量泥沙在岛堤之外沉积并发育形成新的具有不同潮滩高程的滩地, 受潮汐影响, 滩面水流在涨、落潮过程中不断冲刷滩面, 而崇明东滩南部植被相对稀疏, 因此在南部潮滩上形成多条宽窄不一、错综复杂的潮沟(张修桂, 2005 ; 姜亦飞等, 2012 ; 刘建华等, 2012 ; 陈勇等, 2013 ), 这在一定程度上限制了人类活动对保护区的影响. ...
长江河口枯季北支盐水倒灌特征分析
1
2012
... 长江入海径流以崇明岛为界, 形成南、北两个一级支流, 两侧入海淡水不均衡, 南支淡水充裕, 北支偏少, 加之涨潮时北支经常出现咸水倒灌(贺松林等, 2006 ), 因此, 崇明东滩南部水体盐度明显低于北部.受潮汐影响, 海水与淡水在混合过程中又形成了新的盐度梯度, 即低潮带盐度高于中、高潮滩(陈中义等, 2005a ; 贺松林等, 2006 ; 曹利利等, 2012 ; 王卿等, 2012 ).与此同时, 径流中所携带的大量泥沙在岛堤之外沉积并发育形成新的具有不同潮滩高程的滩地, 受潮汐影响, 滩面水流在涨、落潮过程中不断冲刷滩面, 而崇明东滩南部植被相对稀疏, 因此在南部潮滩上形成多条宽窄不一、错综复杂的潮沟(张修桂, 2005 ; 姜亦飞等, 2012 ; 刘建华等, 2012 ; 陈勇等, 2013 ), 这在一定程度上限制了人类活动对保护区的影响. ...
中国特有盐沼植物——海三棱藨草的现状及保护利用
2
2011
... 上述植物中, 海三棱藨草是中国特有的盐沼植物, 也是典型的潮滩先锋物种, 目前主要分布于长江口沿岸的滩涂湿地(Sun et al ., 2002 , 2003 ; 梅雪英和张修峰, 2007 ; 陈秀芝和孙瑛, 2011 ).崇明东滩作为长江口最大的天然湿地, 有大量的海三棱藨草分布(虞快, 1994 , 1996 ), 但近年来崇明东滩北部互花米草生长泛滥, 海三棱藨草原有生态位受到挤压, 已基本被互花米草取代(陈秀芝和孙瑛, 2011 ), 东旺沙中间(2014年新围垦大堤东南部外围)是海三棱藨草现存种群主要分布区, 同时也是莎草科类群的主要分布区1) (1) 吴纪华 (2014). 上海崇明东滩鸟类国家级自然保护区2012年度资源监测公报, 14-15.) .海三棱藨草在崇明东滩形成湿地草场, 能为鸻鹬类、雁鸭类、鹤类、鸥类等众多越冬候鸟提供天然的觅食、活动、休憩场所, 同时海三棱藨草的根茎和球茎, 还是国家一级保护鸟类——白头鹤的重要饵料(Ma et al ., 2003 ; Zhu et al ., 2007 ), 因此, 海三棱藨草对崇明东滩所有滨海湿地鸟类都显得至关重要.然而近年来滩涂围垦、堤外放牧及外来物种竞争等导致植物种群分布状况发生变化, 植物群落斑块破碎化严重, 整个生态系统服务功能也随之降低(张利权和雍学葵, 1992; Chen et al ., 2004; 陈中义等, 2005b; Wang et al ., 2010).因此, 确定包括海三棱藨草在内的关键植物种群在东滩的空间分布, 明确其随环境因子的变化规律, 对该湿地生态系统的修复和保护具有重要的指导意义. ...
... ), 但近年来崇明东滩北部互花米草生长泛滥, 海三棱藨草原有生态位受到挤压, 已基本被互花米草取代(陈秀芝和孙瑛, 2011 ), 东旺沙中间(2014年新围垦大堤东南部外围)是海三棱藨草现存种群主要分布区, 同时也是莎草科类群的主要分布区1) (1) 吴纪华 (2014). 上海崇明东滩鸟类国家级自然保护区2012年度资源监测公报, 14-15.) .海三棱藨草在崇明东滩形成湿地草场, 能为鸻鹬类、雁鸭类、鹤类、鸥类等众多越冬候鸟提供天然的觅食、活动、休憩场所, 同时海三棱藨草的根茎和球茎, 还是国家一级保护鸟类——白头鹤的重要饵料(Ma et al ., 2003 ; Zhu et al ., 2007 ), 因此, 海三棱藨草对崇明东滩所有滨海湿地鸟类都显得至关重要.然而近年来滩涂围垦、堤外放牧及外来物种竞争等导致植物种群分布状况发生变化, 植物群落斑块破碎化严重, 整个生态系统服务功能也随之降低(张利权和雍学葵, 1992; Chen et al ., 2004; 陈中义等, 2005b; Wang et al ., 2010).因此, 确定包括海三棱藨草在内的关键植物种群在东滩的空间分布, 明确其随环境因子的变化规律, 对该湿地生态系统的修复和保护具有重要的指导意义. ...
中国特有盐沼植物——海三棱藨草的现状及保护利用
2
2011
... 上述植物中, 海三棱藨草是中国特有的盐沼植物, 也是典型的潮滩先锋物种, 目前主要分布于长江口沿岸的滩涂湿地(Sun et al ., 2002 , 2003 ; 梅雪英和张修峰, 2007 ; 陈秀芝和孙瑛, 2011 ).崇明东滩作为长江口最大的天然湿地, 有大量的海三棱藨草分布(虞快, 1994 , 1996 ), 但近年来崇明东滩北部互花米草生长泛滥, 海三棱藨草原有生态位受到挤压, 已基本被互花米草取代(陈秀芝和孙瑛, 2011 ), 东旺沙中间(2014年新围垦大堤东南部外围)是海三棱藨草现存种群主要分布区, 同时也是莎草科类群的主要分布区1) (1) 吴纪华 (2014). 上海崇明东滩鸟类国家级自然保护区2012年度资源监测公报, 14-15.) .海三棱藨草在崇明东滩形成湿地草场, 能为鸻鹬类、雁鸭类、鹤类、鸥类等众多越冬候鸟提供天然的觅食、活动、休憩场所, 同时海三棱藨草的根茎和球茎, 还是国家一级保护鸟类——白头鹤的重要饵料(Ma et al ., 2003 ; Zhu et al ., 2007 ), 因此, 海三棱藨草对崇明东滩所有滨海湿地鸟类都显得至关重要.然而近年来滩涂围垦、堤外放牧及外来物种竞争等导致植物种群分布状况发生变化, 植物群落斑块破碎化严重, 整个生态系统服务功能也随之降低(张利权和雍学葵, 1992; Chen et al ., 2004; 陈中义等, 2005b; Wang et al ., 2010).因此, 确定包括海三棱藨草在内的关键植物种群在东滩的空间分布, 明确其随环境因子的变化规律, 对该湿地生态系统的修复和保护具有重要的指导意义. ...
... ), 但近年来崇明东滩北部互花米草生长泛滥, 海三棱藨草原有生态位受到挤压, 已基本被互花米草取代(陈秀芝和孙瑛, 2011 ), 东旺沙中间(2014年新围垦大堤东南部外围)是海三棱藨草现存种群主要分布区, 同时也是莎草科类群的主要分布区1) (1) 吴纪华 (2014). 上海崇明东滩鸟类国家级自然保护区2012年度资源监测公报, 14-15.) .海三棱藨草在崇明东滩形成湿地草场, 能为鸻鹬类、雁鸭类、鹤类、鸥类等众多越冬候鸟提供天然的觅食、活动、休憩场所, 同时海三棱藨草的根茎和球茎, 还是国家一级保护鸟类——白头鹤的重要饵料(Ma et al ., 2003 ; Zhu et al ., 2007 ), 因此, 海三棱藨草对崇明东滩所有滨海湿地鸟类都显得至关重要.然而近年来滩涂围垦、堤外放牧及外来物种竞争等导致植物种群分布状况发生变化, 植物群落斑块破碎化严重, 整个生态系统服务功能也随之降低(张利权和雍学葵, 1992; Chen et al ., 2004; 陈中义等, 2005b; Wang et al ., 2010).因此, 确定包括海三棱藨草在内的关键植物种群在东滩的空间分布, 明确其随环境因子的变化规律, 对该湿地生态系统的修复和保护具有重要的指导意义. ...
崇明东滩潮沟发育特征及其影响因素定量分析
1
2013
... 长江入海径流以崇明岛为界, 形成南、北两个一级支流, 两侧入海淡水不均衡, 南支淡水充裕, 北支偏少, 加之涨潮时北支经常出现咸水倒灌(贺松林等, 2006 ), 因此, 崇明东滩南部水体盐度明显低于北部.受潮汐影响, 海水与淡水在混合过程中又形成了新的盐度梯度, 即低潮带盐度高于中、高潮滩(陈中义等, 2005a ; 贺松林等, 2006 ; 曹利利等, 2012 ; 王卿等, 2012 ).与此同时, 径流中所携带的大量泥沙在岛堤之外沉积并发育形成新的具有不同潮滩高程的滩地, 受潮汐影响, 滩面水流在涨、落潮过程中不断冲刷滩面, 而崇明东滩南部植被相对稀疏, 因此在南部潮滩上形成多条宽窄不一、错综复杂的潮沟(张修桂, 2005 ; 姜亦飞等, 2012 ; 刘建华等, 2012 ; 陈勇等, 2013 ), 这在一定程度上限制了人类活动对保护区的影响. ...
崇明东滩潮沟发育特征及其影响因素定量分析
1
2013
... 长江入海径流以崇明岛为界, 形成南、北两个一级支流, 两侧入海淡水不均衡, 南支淡水充裕, 北支偏少, 加之涨潮时北支经常出现咸水倒灌(贺松林等, 2006 ), 因此, 崇明东滩南部水体盐度明显低于北部.受潮汐影响, 海水与淡水在混合过程中又形成了新的盐度梯度, 即低潮带盐度高于中、高潮滩(陈中义等, 2005a ; 贺松林等, 2006 ; 曹利利等, 2012 ; 王卿等, 2012 ).与此同时, 径流中所携带的大量泥沙在岛堤之外沉积并发育形成新的具有不同潮滩高程的滩地, 受潮汐影响, 滩面水流在涨、落潮过程中不断冲刷滩面, 而崇明东滩南部植被相对稀疏, 因此在南部潮滩上形成多条宽窄不一、错综复杂的潮沟(张修桂, 2005 ; 姜亦飞等, 2012 ; 刘建华等, 2012 ; 陈勇等, 2013 ), 这在一定程度上限制了人类活动对保护区的影响. ...
长江口崇明东滩土壤盐度和潮间带高程对外来种互花米草生长的影响
4
2005a
... 在前人相关研究(Ma et al ., 2003 ; 陈中义等, 2005a ; 贺强等, 2009 ; 詹跃武等, 2011 )的基础上, 本研究选择潮滩高程和表层30 cm土壤盐度作为主要参数, 探讨盐沼植物在东滩南部的空间分布及其与两大环境因子的相互关系. ...
... 长江入海径流以崇明岛为界, 形成南、北两个一级支流, 两侧入海淡水不均衡, 南支淡水充裕, 北支偏少, 加之涨潮时北支经常出现咸水倒灌(贺松林等, 2006 ), 因此, 崇明东滩南部水体盐度明显低于北部.受潮汐影响, 海水与淡水在混合过程中又形成了新的盐度梯度, 即低潮带盐度高于中、高潮滩(陈中义等, 2005a ; 贺松林等, 2006 ; 曹利利等, 2012 ; 王卿等, 2012 ).与此同时, 径流中所携带的大量泥沙在岛堤之外沉积并发育形成新的具有不同潮滩高程的滩地, 受潮汐影响, 滩面水流在涨、落潮过程中不断冲刷滩面, 而崇明东滩南部植被相对稀疏, 因此在南部潮滩上形成多条宽窄不一、错综复杂的潮沟(张修桂, 2005 ; 姜亦飞等, 2012 ; 刘建华等, 2012 ; 陈勇等, 2013 ), 这在一定程度上限制了人类活动对保护区的影响. ...
... 研究区内, 潮滩高程介于-1.22-7.15 m之间(图5A ), 总体呈西高东低, 东旺沙附近地势较高, 在3.50-7.15 m之间, 在东旺沙中心部分出现高程最大值, 大潮沟南部至团结沙区, 介于2.30-5.10 m之间.长江口咸、淡水交互作用, 在崇明东滩外部区域形成了天然盐度梯度变化(陈中义等, 2005a ; 詹跃武等, 2011 ), 研究区内土壤盐度范围介于1.60-4.42 g·kg-1 , 并且空间上具有明显的变化规律——东北高、西南低, 因此土壤盐度最大值出现在东北部, 达到4.42 g·kg-1 . ...
... 潮滩高程固然是划分植被分布范围的重要参考依据, 但植被空间分布同时受到其他环境因子的影响, 盐度就是其中一个重要的限定因子.植物对盐度的变化非常敏感, 盐沼植物对于盐度虽已形成一系列的响应和适应机制, 但盐度依然是盐沼植物生长过程中重要的影响因素(陈中义等, 2005a ; Zhang et al ., 2011 ).因为潮汐的不连续性, 加之长江口水体盐度较土壤盐度波动频繁, 因此选择土壤盐度为研究对象.从图6 可知, 互花米草分布区的土壤盐度最高, 为(3.21 ± 0.51 g·kg-1 ), 白茅区盐度值最低, 为(1.72 ± 0.15 g·kg-1 ), 另外4种主要植物的土壤盐度分别为: 海三棱藨草(2.86 ± 0.64 g·kg-1 ) >芦苇(2.12 ± 0.57 g·kg-1 )≈藨草(2.10 ± 0.37 g·kg-1 ) >糙叶薹草(1.90 ± 0.26 g·kg-1 ), 除了芦苇和藨草, 其余各类群植物土壤盐度差异性非常显著(p < 0.01), 因此土壤盐度在空间的变化必然影响甚至决定了各植物类群间的分布状况. ...
长江口崇明东滩土壤盐度和潮间带高程对外来种互花米草生长的影响
4
2005a
... 在前人相关研究(Ma et al ., 2003 ; 陈中义等, 2005a ; 贺强等, 2009 ; 詹跃武等, 2011 )的基础上, 本研究选择潮滩高程和表层30 cm土壤盐度作为主要参数, 探讨盐沼植物在东滩南部的空间分布及其与两大环境因子的相互关系. ...
... 长江入海径流以崇明岛为界, 形成南、北两个一级支流, 两侧入海淡水不均衡, 南支淡水充裕, 北支偏少, 加之涨潮时北支经常出现咸水倒灌(贺松林等, 2006 ), 因此, 崇明东滩南部水体盐度明显低于北部.受潮汐影响, 海水与淡水在混合过程中又形成了新的盐度梯度, 即低潮带盐度高于中、高潮滩(陈中义等, 2005a ; 贺松林等, 2006 ; 曹利利等, 2012 ; 王卿等, 2012 ).与此同时, 径流中所携带的大量泥沙在岛堤之外沉积并发育形成新的具有不同潮滩高程的滩地, 受潮汐影响, 滩面水流在涨、落潮过程中不断冲刷滩面, 而崇明东滩南部植被相对稀疏, 因此在南部潮滩上形成多条宽窄不一、错综复杂的潮沟(张修桂, 2005 ; 姜亦飞等, 2012 ; 刘建华等, 2012 ; 陈勇等, 2013 ), 这在一定程度上限制了人类活动对保护区的影响. ...
... 研究区内, 潮滩高程介于-1.22-7.15 m之间(图5A ), 总体呈西高东低, 东旺沙附近地势较高, 在3.50-7.15 m之间, 在东旺沙中心部分出现高程最大值, 大潮沟南部至团结沙区, 介于2.30-5.10 m之间.长江口咸、淡水交互作用, 在崇明东滩外部区域形成了天然盐度梯度变化(陈中义等, 2005a ; 詹跃武等, 2011 ), 研究区内土壤盐度范围介于1.60-4.42 g·kg-1 , 并且空间上具有明显的变化规律——东北高、西南低, 因此土壤盐度最大值出现在东北部, 达到4.42 g·kg-1 . ...
... 潮滩高程固然是划分植被分布范围的重要参考依据, 但植被空间分布同时受到其他环境因子的影响, 盐度就是其中一个重要的限定因子.植物对盐度的变化非常敏感, 盐沼植物对于盐度虽已形成一系列的响应和适应机制, 但盐度依然是盐沼植物生长过程中重要的影响因素(陈中义等, 2005a ; Zhang et al ., 2011 ).因为潮汐的不连续性, 加之长江口水体盐度较土壤盐度波动频繁, 因此选择土壤盐度为研究对象.从图6 可知, 互花米草分布区的土壤盐度最高, 为(3.21 ± 0.51 g·kg-1 ), 白茅区盐度值最低, 为(1.72 ± 0.15 g·kg-1 ), 另外4种主要植物的土壤盐度分别为: 海三棱藨草(2.86 ± 0.64 g·kg-1 ) >芦苇(2.12 ± 0.57 g·kg-1 )≈藨草(2.10 ± 0.37 g·kg-1 ) >糙叶薹草(1.90 ± 0.26 g·kg-1 ), 除了芦苇和藨草, 其余各类群植物土壤盐度差异性非常显著(p < 0.01), 因此土壤盐度在空间的变化必然影响甚至决定了各植物类群间的分布状况. ...
互花米草与海三棱藨草的生长特征和相对竞争能力
1
2005b
... 值得关注的是, 海三棱藨草(图2F )作为中国特有盐沼植物, 其种群面积与前人研究结果(黄华梅等, 2007 ; Yuan & Zhang, 2008 )存在较大差异.此外, 作为特有先锋物种, 海三棱藨草分布在“外带”, 但优势带也与互花米草存在较大的重叠空间, 两者存在显著的竞争关系, 然而互花米草在高度、盖度、地上生物量、地下生物量方面都比海三棱藨草具有明显优势, 为其获取更多光照、营养盐, 快速取代海三棱藨草创造了条件(Chen et al ., 2004 ; 陈中义等, 2005b , He et al ., 2011 ), 因此, 两种植物的分布存在交错带对海三棱藨草种群的发展极其不利. ...
互花米草与海三棱藨草的生长特征和相对竞争能力
1
2005b
... 值得关注的是, 海三棱藨草(图2F )作为中国特有盐沼植物, 其种群面积与前人研究结果(黄华梅等, 2007 ; Yuan & Zhang, 2008 )存在较大差异.此外, 作为特有先锋物种, 海三棱藨草分布在“外带”, 但优势带也与互花米草存在较大的重叠空间, 两者存在显著的竞争关系, 然而互花米草在高度、盖度、地上生物量、地下生物量方面都比海三棱藨草具有明显优势, 为其获取更多光照、营养盐, 快速取代海三棱藨草创造了条件(Chen et al ., 2004 ; 陈中义等, 2005b , He et al ., 2011 ), 因此, 两种植物的分布存在交错带对海三棱藨草种群的发展极其不利. ...
Local competitive effects of introduced Spartina alterniflora on Scirpus mariqueter at Dongtan of Chongming Island, the Yangtze River estuary and their potential ecological consequences.
2
2004
... 值得关注的是, 海三棱藨草(图2F )作为中国特有盐沼植物, 其种群面积与前人研究结果(黄华梅等, 2007 ; Yuan & Zhang, 2008 )存在较大差异.此外, 作为特有先锋物种, 海三棱藨草分布在“外带”, 但优势带也与互花米草存在较大的重叠空间, 两者存在显著的竞争关系, 然而互花米草在高度、盖度、地上生物量、地下生物量方面都比海三棱藨草具有明显优势, 为其获取更多光照、营养盐, 快速取代海三棱藨草创造了条件(Chen et al ., 2004 ; 陈中义等, 2005b , He et al ., 2011 ), 因此, 两种植物的分布存在交错带对海三棱藨草种群的发展极其不利. ...
... 另一方面, 芦苇与互花米草的交错边界最长, 但是两者在高程以及盐度上都存在非常显著的差异(p < 0.01), 并且区间偏离较大, 因此互花米草不具备向芦苇种群扩张的优势.与海三棱藨草相比, 互花米草能够适应更高的高程和土壤盐度, 海三棱藨草-互花米草植被交错区往大堤方向高程变高, 反而适合互花米草生长; 交错带偏海三棱藨草一侧土壤盐度逐渐升高, 限制了海三棱藨草进一步扩张, 而在交错区内, 互花米草通过根茎、分蘖等繁殖方式, 争夺空间资源, 在植株高度、盖度等方面的优势限制了海三棱藨草生长(Chen et al ., 2004 ; Wang et al ., 2010 ).无论在交错区内部, 还是在与交错带毗邻的缓冲空间, 互花米草在高程和土壤盐度的综合作用下, 生长状况会更加良好, 而海三棱藨草很可能会被互花米草逐步取代. ...
The role of coastal plant communities for climate change mitigation and adaptation.
2
2013
... 盐沼植物作为全球范围内生产力较高的生物群落之一, 是推动湿地生态系统物质循环和能量流动的基础要素, 其在维持个体生存、种群稳定与群落发展的同时, 也为湿地生态系统中的众多成员提供了丰富的食物, 并创造了良好的栖息环境.然而, 近年来由于全球气候变化和频繁的人类活动, 植物群落组成结构以及空间分布格局发生了巨大的变化, 这不仅影响了湿地中鸟类、底栖生物及鱼类等生物的种群数量, 而且也不断改变着植物类群的种群结构, 甚至可能会导致一些敏感物种的灭绝(童春富, 2012 ; Isacch et al ., 2014 ; Li et al ., 2014b ).因此, 明确湿地中主要植物种群及其分布状况, 掌握其随环境因子的变化规律, 对湿地生态系统的保护、恢复、重建都具有重要的参考价值.虽然前人已就此进行了较多的研究(黄华梅等, 2007 ; Duarte et al ., 2013 ; 曹浩冰等, 2014 ; Hughes, 2014 ; Li et al ., 2014a , 2014b ), 但河口潮滩区域海陆交互作用强烈、人类活动频繁, 使得滩面、潮沟及植被都处于不断变化中, 崇明东滩正是这样一个处在发育过程中的典型盐沼湿地生态系统.2014年崇明东滩互花米草(Spartina alterniflora )生态治理工程的实施, 可能对该区域盐沼植物的空间分布及其生境产生了重大影响(高宇和赵斌, 2006 ; Zhu et al ., 2014 ).因此, 在施工前期对围垦外围区进行全面的植被调查, 不仅能准确地掌握崇明东滩南部的植被类型和现有分布状况, 而且能为今后研究其植被空间动态变化提供基础数据. ...
... 潮滩植物群落在空间上呈现不同的分布状况, 潮滩高程作为影响植物分布的重要环境因子, 与植物的淹水时间和频度, 水体中的溶解氧, 土壤粒径、养分、氧化还原电位、pH值等生境因子存在密切联系(Silvestri et al ., 2005 ; 何彦龙, 2014 ), 与此同时, 植物的密度、高度等因素也会影响高程的变化(Duarte et al ., 2013 ).本研究表明崇明东滩南部植被覆盖空间6种主要植物分布在4个差异显著的高程区间内(p < 0.01), 黄华梅等(2007)和严格等(2014)认为崇明东滩芦苇分布在高潮带(高程>2.9 m), 海三棱藨草分布在中潮带(高程2.0-2.9 m), 互花米草在中、高潮带都有分布.同时对比何彦龙等(2010)在2008年对崇明东滩互花米草(3.21 ± 0.01 m)和海三棱藨草(3.10 ± 0.02 m)的研究结果, 发现上述植物在高程数值上有所差异, 但各植物种群生长空间的高程相对大小保持稳定, 表明在崇明东滩无论是大尺度区域范围, 还是小尺度空间, 各主要植物类群的潮滩高程相对高度保持恒定, 因此潮滩高程是决定植被空间分布的重要因素, 而具体数值差异可能是由于野外调查的时间、地点以及高程仪器精度差异所致.盐度是植物生长过程中的又一重要条件性因子, 不同的植物在不同的生长阶段适应不同范围的盐度(Zhang et al ., 2011 ).陈中义等(2005b)研究了水体中盐度的变化对互花米草和海三棱藨草的影响, 认为互花米草更适应高盐度海水.本研究中互花米草生长空间的土壤盐度确实显著高于海三棱藨草区(p < 0.01), 除了与互花米草本身的高耐盐特性有关, 与植株地上部分的泌盐作用也有一定关系(He et al ., 2011 ).但是无论崇明东滩土壤盐度还是潮滩高程都直接受到潮汐影响, 在空间上都呈现梯度变化(王卿等, 2012 ), 因此, 崇明东滩南部6种主要植物受到两种限定因子的共同制约, 在空间上形成现有的分布格局, 而对潮滩高程和土壤盐度在不同空间尺度下作用的权重仍有待进一步研究. ...
人类围垦活动对上海崇明东滩滩涂发育的影响
2
2006
... 盐沼植物作为全球范围内生产力较高的生物群落之一, 是推动湿地生态系统物质循环和能量流动的基础要素, 其在维持个体生存、种群稳定与群落发展的同时, 也为湿地生态系统中的众多成员提供了丰富的食物, 并创造了良好的栖息环境.然而, 近年来由于全球气候变化和频繁的人类活动, 植物群落组成结构以及空间分布格局发生了巨大的变化, 这不仅影响了湿地中鸟类、底栖生物及鱼类等生物的种群数量, 而且也不断改变着植物类群的种群结构, 甚至可能会导致一些敏感物种的灭绝(童春富, 2012 ; Isacch et al ., 2014 ; Li et al ., 2014b ).因此, 明确湿地中主要植物种群及其分布状况, 掌握其随环境因子的变化规律, 对湿地生态系统的保护、恢复、重建都具有重要的参考价值.虽然前人已就此进行了较多的研究(黄华梅等, 2007 ; Duarte et al ., 2013 ; 曹浩冰等, 2014 ; Hughes, 2014 ; Li et al ., 2014a , 2014b ), 但河口潮滩区域海陆交互作用强烈、人类活动频繁, 使得滩面、潮沟及植被都处于不断变化中, 崇明东滩正是这样一个处在发育过程中的典型盐沼湿地生态系统.2014年崇明东滩互花米草(Spartina alterniflora )生态治理工程的实施, 可能对该区域盐沼植物的空间分布及其生境产生了重大影响(高宇和赵斌, 2006 ; Zhu et al ., 2014 ).因此, 在施工前期对围垦外围区进行全面的植被调查, 不仅能准确地掌握崇明东滩南部的植被类型和现有分布状况, 而且能为今后研究其植被空间动态变化提供基础数据. ...
... 近十几年来, 大量研究人员通过遥感影像解译, 对崇明东滩主要植被进行了分区(Gao & Zhang, 2006 ; Tian et al ., 2008 ; 阿也提古丽·斯迪克等, 2010 ; 翟新源和杨苏新, 2013 ; 郑宗生等, 2010 ; 宗玮等, 2011 ), 普遍认为芦苇(Phragmites australis )、互花米草(Spartina alterniflora )及海三棱藨草(Scirpus mariqueter )是崇明东滩的“三大”主要植物类群.然而, 崇明东滩南部长期受长江入海淡水影响, 水体盐度相对较低, 在团结沙至大潮沟(图1 )存在较大面积的藨草(Scirpus triqueter )种群区(闫芊等, 2007 ; 章振亚等, 2012 ).同时发现, 糙叶薹草(Carex scabrifolia )在研究区内频繁出现, 白茅(Imperata cylindrica )在部分空间也已发展成优势种. ...
人类围垦活动对上海崇明东滩滩涂发育的影响
2
2006
... 盐沼植物作为全球范围内生产力较高的生物群落之一, 是推动湿地生态系统物质循环和能量流动的基础要素, 其在维持个体生存、种群稳定与群落发展的同时, 也为湿地生态系统中的众多成员提供了丰富的食物, 并创造了良好的栖息环境.然而, 近年来由于全球气候变化和频繁的人类活动, 植物群落组成结构以及空间分布格局发生了巨大的变化, 这不仅影响了湿地中鸟类、底栖生物及鱼类等生物的种群数量, 而且也不断改变着植物类群的种群结构, 甚至可能会导致一些敏感物种的灭绝(童春富, 2012 ; Isacch et al ., 2014 ; Li et al ., 2014b ).因此, 明确湿地中主要植物种群及其分布状况, 掌握其随环境因子的变化规律, 对湿地生态系统的保护、恢复、重建都具有重要的参考价值.虽然前人已就此进行了较多的研究(黄华梅等, 2007 ; Duarte et al ., 2013 ; 曹浩冰等, 2014 ; Hughes, 2014 ; Li et al ., 2014a , 2014b ), 但河口潮滩区域海陆交互作用强烈、人类活动频繁, 使得滩面、潮沟及植被都处于不断变化中, 崇明东滩正是这样一个处在发育过程中的典型盐沼湿地生态系统.2014年崇明东滩互花米草(Spartina alterniflora )生态治理工程的实施, 可能对该区域盐沼植物的空间分布及其生境产生了重大影响(高宇和赵斌, 2006 ; Zhu et al ., 2014 ).因此, 在施工前期对围垦外围区进行全面的植被调查, 不仅能准确地掌握崇明东滩南部的植被类型和现有分布状况, 而且能为今后研究其植被空间动态变化提供基础数据. ...
... 近十几年来, 大量研究人员通过遥感影像解译, 对崇明东滩主要植被进行了分区(Gao & Zhang, 2006 ; Tian et al ., 2008 ; 阿也提古丽·斯迪克等, 2010 ; 翟新源和杨苏新, 2013 ; 郑宗生等, 2010 ; 宗玮等, 2011 ), 普遍认为芦苇(Phragmites australis )、互花米草(Spartina alterniflora )及海三棱藨草(Scirpus mariqueter )是崇明东滩的“三大”主要植物类群.然而, 崇明东滩南部长期受长江入海淡水影响, 水体盐度相对较低, 在团结沙至大潮沟(图1 )存在较大面积的藨草(Scirpus triqueter )种群区(闫芊等, 2007 ; 章振亚等, 2012 ).同时发现, 糙叶薹草(Carex scabrifolia )在研究区内频繁出现, 白茅(Imperata cylindrica )在部分空间也已发展成优势种. ...
Multi-seasonal spectral characteristics analysis of coastal salt marsh vegetation in Shanghai, China.
2006
黄河河口盐沼植被分布、多样性与土壤化学因子的相关关系
1
2009
... 在前人相关研究(Ma et al ., 2003 ; 陈中义等, 2005a ; 贺强等, 2009 ; 詹跃武等, 2011 )的基础上, 本研究选择潮滩高程和表层30 cm土壤盐度作为主要参数, 探讨盐沼植物在东滩南部的空间分布及其与两大环境因子的相互关系. ...
黄河河口盐沼植被分布、多样性与土壤化学因子的相关关系
1
2009
... 在前人相关研究(Ma et al ., 2003 ; 陈中义等, 2005a ; 贺强等, 2009 ; 詹跃武等, 2011 )的基础上, 本研究选择潮滩高程和表层30 cm土壤盐度作为主要参数, 探讨盐沼植物在东滩南部的空间分布及其与两大环境因子的相互关系. ...
长江口南支河段枯季盐度变异与北支咸水倒灌
2
2006
... 长江入海径流以崇明岛为界, 形成南、北两个一级支流, 两侧入海淡水不均衡, 南支淡水充裕, 北支偏少, 加之涨潮时北支经常出现咸水倒灌(贺松林等, 2006 ), 因此, 崇明东滩南部水体盐度明显低于北部.受潮汐影响, 海水与淡水在混合过程中又形成了新的盐度梯度, 即低潮带盐度高于中、高潮滩(陈中义等, 2005a ; 贺松林等, 2006 ; 曹利利等, 2012 ; 王卿等, 2012 ).与此同时, 径流中所携带的大量泥沙在岛堤之外沉积并发育形成新的具有不同潮滩高程的滩地, 受潮汐影响, 滩面水流在涨、落潮过程中不断冲刷滩面, 而崇明东滩南部植被相对稀疏, 因此在南部潮滩上形成多条宽窄不一、错综复杂的潮沟(张修桂, 2005 ; 姜亦飞等, 2012 ; 刘建华等, 2012 ; 陈勇等, 2013 ), 这在一定程度上限制了人类活动对保护区的影响. ...
... ; 贺松林等, 2006 ; 曹利利等, 2012 ; 王卿等, 2012 ).与此同时, 径流中所携带的大量泥沙在岛堤之外沉积并发育形成新的具有不同潮滩高程的滩地, 受潮汐影响, 滩面水流在涨、落潮过程中不断冲刷滩面, 而崇明东滩南部植被相对稀疏, 因此在南部潮滩上形成多条宽窄不一、错综复杂的潮沟(张修桂, 2005 ; 姜亦飞等, 2012 ; 刘建华等, 2012 ; 陈勇等, 2013 ), 这在一定程度上限制了人类活动对保护区的影响. ...
长江口南支河段枯季盐度变异与北支咸水倒灌
2
2006
... 长江入海径流以崇明岛为界, 形成南、北两个一级支流, 两侧入海淡水不均衡, 南支淡水充裕, 北支偏少, 加之涨潮时北支经常出现咸水倒灌(贺松林等, 2006 ), 因此, 崇明东滩南部水体盐度明显低于北部.受潮汐影响, 海水与淡水在混合过程中又形成了新的盐度梯度, 即低潮带盐度高于中、高潮滩(陈中义等, 2005a ; 贺松林等, 2006 ; 曹利利等, 2012 ; 王卿等, 2012 ).与此同时, 径流中所携带的大量泥沙在岛堤之外沉积并发育形成新的具有不同潮滩高程的滩地, 受潮汐影响, 滩面水流在涨、落潮过程中不断冲刷滩面, 而崇明东滩南部植被相对稀疏, 因此在南部潮滩上形成多条宽窄不一、错综复杂的潮沟(张修桂, 2005 ; 姜亦飞等, 2012 ; 刘建华等, 2012 ; 陈勇等, 2013 ), 这在一定程度上限制了人类活动对保护区的影响. ...
... ; 贺松林等, 2006 ; 曹利利等, 2012 ; 王卿等, 2012 ).与此同时, 径流中所携带的大量泥沙在岛堤之外沉积并发育形成新的具有不同潮滩高程的滩地, 受潮汐影响, 滩面水流在涨、落潮过程中不断冲刷滩面, 而崇明东滩南部植被相对稀疏, 因此在南部潮滩上形成多条宽窄不一、错综复杂的潮沟(张修桂, 2005 ; 姜亦飞等, 2012 ; 刘建华等, 2012 ; 陈勇等, 2013 ), 这在一定程度上限制了人类活动对保护区的影响. ...
中低潮滩盐沼植被分异的形成机制研究
1
2014
... 潮滩植物群落在空间上呈现不同的分布状况, 潮滩高程作为影响植物分布的重要环境因子, 与植物的淹水时间和频度, 水体中的溶解氧, 土壤粒径、养分、氧化还原电位、pH值等生境因子存在密切联系(Silvestri et al ., 2005 ; 何彦龙, 2014 ), 与此同时, 植物的密度、高度等因素也会影响高程的变化(Duarte et al ., 2013 ).本研究表明崇明东滩南部植被覆盖空间6种主要植物分布在4个差异显著的高程区间内(p < 0.01), 黄华梅等(2007)和严格等(2014)认为崇明东滩芦苇分布在高潮带(高程>2.9 m), 海三棱藨草分布在中潮带(高程2.0-2.9 m), 互花米草在中、高潮带都有分布.同时对比何彦龙等(2010)在2008年对崇明东滩互花米草(3.21 ± 0.01 m)和海三棱藨草(3.10 ± 0.02 m)的研究结果, 发现上述植物在高程数值上有所差异, 但各植物种群生长空间的高程相对大小保持稳定, 表明在崇明东滩无论是大尺度区域范围, 还是小尺度空间, 各主要植物类群的潮滩高程相对高度保持恒定, 因此潮滩高程是决定植被空间分布的重要因素, 而具体数值差异可能是由于野外调查的时间、地点以及高程仪器精度差异所致.盐度是植物生长过程中的又一重要条件性因子, 不同的植物在不同的生长阶段适应不同范围的盐度(Zhang et al ., 2011 ).陈中义等(2005b)研究了水体中盐度的变化对互花米草和海三棱藨草的影响, 认为互花米草更适应高盐度海水.本研究中互花米草生长空间的土壤盐度确实显著高于海三棱藨草区(p < 0.01), 除了与互花米草本身的高耐盐特性有关, 与植株地上部分的泌盐作用也有一定关系(He et al ., 2011 ).但是无论崇明东滩土壤盐度还是潮滩高程都直接受到潮汐影响, 在空间上都呈现梯度变化(王卿等, 2012 ), 因此, 崇明东滩南部6种主要植物受到两种限定因子的共同制约, 在空间上形成现有的分布格局, 而对潮滩高程和土壤盐度在不同空间尺度下作用的权重仍有待进一步研究. ...
中低潮滩盐沼植被分异的形成机制研究
1
2014
... 潮滩植物群落在空间上呈现不同的分布状况, 潮滩高程作为影响植物分布的重要环境因子, 与植物的淹水时间和频度, 水体中的溶解氧, 土壤粒径、养分、氧化还原电位、pH值等生境因子存在密切联系(Silvestri et al ., 2005 ; 何彦龙, 2014 ), 与此同时, 植物的密度、高度等因素也会影响高程的变化(Duarte et al ., 2013 ).本研究表明崇明东滩南部植被覆盖空间6种主要植物分布在4个差异显著的高程区间内(p < 0.01), 黄华梅等(2007)和严格等(2014)认为崇明东滩芦苇分布在高潮带(高程>2.9 m), 海三棱藨草分布在中潮带(高程2.0-2.9 m), 互花米草在中、高潮带都有分布.同时对比何彦龙等(2010)在2008年对崇明东滩互花米草(3.21 ± 0.01 m)和海三棱藨草(3.10 ± 0.02 m)的研究结果, 发现上述植物在高程数值上有所差异, 但各植物种群生长空间的高程相对大小保持稳定, 表明在崇明东滩无论是大尺度区域范围, 还是小尺度空间, 各主要植物类群的潮滩高程相对高度保持恒定, 因此潮滩高程是决定植被空间分布的重要因素, 而具体数值差异可能是由于野外调查的时间、地点以及高程仪器精度差异所致.盐度是植物生长过程中的又一重要条件性因子, 不同的植物在不同的生长阶段适应不同范围的盐度(Zhang et al ., 2011 ).陈中义等(2005b)研究了水体中盐度的变化对互花米草和海三棱藨草的影响, 认为互花米草更适应高盐度海水.本研究中互花米草生长空间的土壤盐度确实显著高于海三棱藨草区(p < 0.01), 除了与互花米草本身的高耐盐特性有关, 与植株地上部分的泌盐作用也有一定关系(He et al ., 2011 ).但是无论崇明东滩土壤盐度还是潮滩高程都直接受到潮汐影响, 在空间上都呈现梯度变化(王卿等, 2012 ), 因此, 崇明东滩南部6种主要植物受到两种限定因子的共同制约, 在空间上形成现有的分布格局, 而对潮滩高程和土壤盐度在不同空间尺度下作用的权重仍有待进一步研究. ...
Relationships between vegetation zonation and environmental factors in newly formed tidal marshes of the Yangtze River estuary.
2
2011
... 值得关注的是, 海三棱藨草(图2F )作为中国特有盐沼植物, 其种群面积与前人研究结果(黄华梅等, 2007 ; Yuan & Zhang, 2008 )存在较大差异.此外, 作为特有先锋物种, 海三棱藨草分布在“外带”, 但优势带也与互花米草存在较大的重叠空间, 两者存在显著的竞争关系, 然而互花米草在高度、盖度、地上生物量、地下生物量方面都比海三棱藨草具有明显优势, 为其获取更多光照、营养盐, 快速取代海三棱藨草创造了条件(Chen et al ., 2004 ; 陈中义等, 2005b , He et al ., 2011 ), 因此, 两种植物的分布存在交错带对海三棱藨草种群的发展极其不利. ...
... 潮滩植物群落在空间上呈现不同的分布状况, 潮滩高程作为影响植物分布的重要环境因子, 与植物的淹水时间和频度, 水体中的溶解氧, 土壤粒径、养分、氧化还原电位、pH值等生境因子存在密切联系(Silvestri et al ., 2005 ; 何彦龙, 2014 ), 与此同时, 植物的密度、高度等因素也会影响高程的变化(Duarte et al ., 2013 ).本研究表明崇明东滩南部植被覆盖空间6种主要植物分布在4个差异显著的高程区间内(p < 0.01), 黄华梅等(2007)和严格等(2014)认为崇明东滩芦苇分布在高潮带(高程>2.9 m), 海三棱藨草分布在中潮带(高程2.0-2.9 m), 互花米草在中、高潮带都有分布.同时对比何彦龙等(2010)在2008年对崇明东滩互花米草(3.21 ± 0.01 m)和海三棱藨草(3.10 ± 0.02 m)的研究结果, 发现上述植物在高程数值上有所差异, 但各植物种群生长空间的高程相对大小保持稳定, 表明在崇明东滩无论是大尺度区域范围, 还是小尺度空间, 各主要植物类群的潮滩高程相对高度保持恒定, 因此潮滩高程是决定植被空间分布的重要因素, 而具体数值差异可能是由于野外调查的时间、地点以及高程仪器精度差异所致.盐度是植物生长过程中的又一重要条件性因子, 不同的植物在不同的生长阶段适应不同范围的盐度(Zhang et al ., 2011 ).陈中义等(2005b)研究了水体中盐度的变化对互花米草和海三棱藨草的影响, 认为互花米草更适应高盐度海水.本研究中互花米草生长空间的土壤盐度确实显著高于海三棱藨草区(p < 0.01), 除了与互花米草本身的高耐盐特性有关, 与植株地上部分的泌盐作用也有一定关系(He et al ., 2011 ).但是无论崇明东滩土壤盐度还是潮滩高程都直接受到潮汐影响, 在空间上都呈现梯度变化(王卿等, 2012 ), 因此, 崇明东滩南部6种主要植物受到两种限定因子的共同制约, 在空间上形成现有的分布格局, 而对潮滩高程和土壤盐度在不同空间尺度下作用的权重仍有待进一步研究. ...
上海滩涂盐沼植被的分布格局和时空动态研究
2009
上海滩涂盐沼植被的分布格局和时空动态研究
2009
崇明东滩自然保护区盐沼植被的时空动态
4
2007
... 盐沼植物作为全球范围内生产力较高的生物群落之一, 是推动湿地生态系统物质循环和能量流动的基础要素, 其在维持个体生存、种群稳定与群落发展的同时, 也为湿地生态系统中的众多成员提供了丰富的食物, 并创造了良好的栖息环境.然而, 近年来由于全球气候变化和频繁的人类活动, 植物群落组成结构以及空间分布格局发生了巨大的变化, 这不仅影响了湿地中鸟类、底栖生物及鱼类等生物的种群数量, 而且也不断改变着植物类群的种群结构, 甚至可能会导致一些敏感物种的灭绝(童春富, 2012 ; Isacch et al ., 2014 ; Li et al ., 2014b ).因此, 明确湿地中主要植物种群及其分布状况, 掌握其随环境因子的变化规律, 对湿地生态系统的保护、恢复、重建都具有重要的参考价值.虽然前人已就此进行了较多的研究(黄华梅等, 2007 ; Duarte et al ., 2013 ; 曹浩冰等, 2014 ; Hughes, 2014 ; Li et al ., 2014a , 2014b ), 但河口潮滩区域海陆交互作用强烈、人类活动频繁, 使得滩面、潮沟及植被都处于不断变化中, 崇明东滩正是这样一个处在发育过程中的典型盐沼湿地生态系统.2014年崇明东滩互花米草(Spartina alterniflora )生态治理工程的实施, 可能对该区域盐沼植物的空间分布及其生境产生了重大影响(高宇和赵斌, 2006 ; Zhu et al ., 2014 ).因此, 在施工前期对围垦外围区进行全面的植被调查, 不仅能准确地掌握崇明东滩南部的植被类型和现有分布状况, 而且能为今后研究其植被空间动态变化提供基础数据. ...
... 值得关注的是, 海三棱藨草(图2F )作为中国特有盐沼植物, 其种群面积与前人研究结果(黄华梅等, 2007 ; Yuan & Zhang, 2008 )存在较大差异.此外, 作为特有先锋物种, 海三棱藨草分布在“外带”, 但优势带也与互花米草存在较大的重叠空间, 两者存在显著的竞争关系, 然而互花米草在高度、盖度、地上生物量、地下生物量方面都比海三棱藨草具有明显优势, 为其获取更多光照、营养盐, 快速取代海三棱藨草创造了条件(Chen et al ., 2004 ; 陈中义等, 2005b , He et al ., 2011 ), 因此, 两种植物的分布存在交错带对海三棱藨草种群的发展极其不利. ...
... 崇明东滩属于典型的淤涨型潮滩, 目前滩涂植被覆盖区总面积4265 hm2 , 2012年调查数据显示, 崇明东滩中部、北部植物以芦苇和互花米草为主, 而莎草科植物主要集中分布在崇明东滩南部1) ( 1) 吴纪华 (2014). 上海崇明东滩鸟类国家级自然保护区2012年度资源监测公报, 14-15.) .在崇明东滩的相关研究中, 众多学者根据研究需要将崇明东滩的海三棱藨草、藨草、糙叶薹草3种植物统一归入海三棱藨草类群(黄华梅等, 2007 ; Yuan & Zhang, 2008 ; 严格等, 2014 ).部分研究虽然将海三棱藨草、藨草、糙叶薹草统一划分为莎草科植物群落或者藨草群落(曹浩冰等, 2014 ; 任璘婧等, 2014 ), 但没有对这3种植物进行细致的划分, 各种群面积大小及其具体分布目前尚无相关报道.上述3种植物虽然同属于莎草科, 但占据不同的生态位, 承担着不同的生态功能, 例如在崇明东滩三者虽然都可以作为湿地鸟类的饵料, 但不同的鸟类会取食不同植物的不同部分(Ma et al ., 2003 ; Zhu et al ., 2007 ), 且3种植物组织、器官发育和成熟时间也各不相同.本文基于崇明东滩南部各植物种群的细致调查, 明确了各主要植物种群的空间分布以及各自的面积, 为进一步研究植被分布格局变化以及重要鸟类觅食生境提供基础. ...
... 黄华梅等(2007) 认为, 影响崇明东滩植被分布格局的主要因素是互花米草入侵和围堤工程.但2014年互花米草治理工程实施后, 围堤外仍存在大面积的互花米草, 根据土壤盐度和潮滩高程的空间分布趋势, 互花米草具有向土壤盐度较高的东北方向扩张的可能, 广泛的高程适应范围又增加了其向南部扩张的几率, 在盐度和高程变化的影响下, 互花米草种群在发展过程中与海三棱藨草之间必然存在激烈的竞争关系, 这极有可能破坏该区植物群落的稳定现状.目前进行的互花米草生态治理工程虽未直接影响东滩南部的植被分布状况, 但是围堤之后, 大坝外围的水动力条件必然发生改变, 影响堤外泥沙淤积速率、土壤粒径等环境因子, 进而影响南部潮滩高程和土壤盐度, 改变植被空间格局.但具体的变化过程和趋势以及由此造成的影响还需要更长时间尺度的监测和分析, 其中本地特有物种海三棱藨草的面积和分布变化尤其值得关注. ...
崇明东滩自然保护区盐沼植被的时空动态
4
2007
... 盐沼植物作为全球范围内生产力较高的生物群落之一, 是推动湿地生态系统物质循环和能量流动的基础要素, 其在维持个体生存、种群稳定与群落发展的同时, 也为湿地生态系统中的众多成员提供了丰富的食物, 并创造了良好的栖息环境.然而, 近年来由于全球气候变化和频繁的人类活动, 植物群落组成结构以及空间分布格局发生了巨大的变化, 这不仅影响了湿地中鸟类、底栖生物及鱼类等生物的种群数量, 而且也不断改变着植物类群的种群结构, 甚至可能会导致一些敏感物种的灭绝(童春富, 2012 ; Isacch et al ., 2014 ; Li et al ., 2014b ).因此, 明确湿地中主要植物种群及其分布状况, 掌握其随环境因子的变化规律, 对湿地生态系统的保护、恢复、重建都具有重要的参考价值.虽然前人已就此进行了较多的研究(黄华梅等, 2007 ; Duarte et al ., 2013 ; 曹浩冰等, 2014 ; Hughes, 2014 ; Li et al ., 2014a , 2014b ), 但河口潮滩区域海陆交互作用强烈、人类活动频繁, 使得滩面、潮沟及植被都处于不断变化中, 崇明东滩正是这样一个处在发育过程中的典型盐沼湿地生态系统.2014年崇明东滩互花米草(Spartina alterniflora )生态治理工程的实施, 可能对该区域盐沼植物的空间分布及其生境产生了重大影响(高宇和赵斌, 2006 ; Zhu et al ., 2014 ).因此, 在施工前期对围垦外围区进行全面的植被调查, 不仅能准确地掌握崇明东滩南部的植被类型和现有分布状况, 而且能为今后研究其植被空间动态变化提供基础数据. ...
... 值得关注的是, 海三棱藨草(图2F )作为中国特有盐沼植物, 其种群面积与前人研究结果(黄华梅等, 2007 ; Yuan & Zhang, 2008 )存在较大差异.此外, 作为特有先锋物种, 海三棱藨草分布在“外带”, 但优势带也与互花米草存在较大的重叠空间, 两者存在显著的竞争关系, 然而互花米草在高度、盖度、地上生物量、地下生物量方面都比海三棱藨草具有明显优势, 为其获取更多光照、营养盐, 快速取代海三棱藨草创造了条件(Chen et al ., 2004 ; 陈中义等, 2005b , He et al ., 2011 ), 因此, 两种植物的分布存在交错带对海三棱藨草种群的发展极其不利. ...
... 崇明东滩属于典型的淤涨型潮滩, 目前滩涂植被覆盖区总面积4265 hm2 , 2012年调查数据显示, 崇明东滩中部、北部植物以芦苇和互花米草为主, 而莎草科植物主要集中分布在崇明东滩南部1) ( 1) 吴纪华 (2014). 上海崇明东滩鸟类国家级自然保护区2012年度资源监测公报, 14-15.) .在崇明东滩的相关研究中, 众多学者根据研究需要将崇明东滩的海三棱藨草、藨草、糙叶薹草3种植物统一归入海三棱藨草类群(黄华梅等, 2007 ; Yuan & Zhang, 2008 ; 严格等, 2014 ).部分研究虽然将海三棱藨草、藨草、糙叶薹草统一划分为莎草科植物群落或者藨草群落(曹浩冰等, 2014 ; 任璘婧等, 2014 ), 但没有对这3种植物进行细致的划分, 各种群面积大小及其具体分布目前尚无相关报道.上述3种植物虽然同属于莎草科, 但占据不同的生态位, 承担着不同的生态功能, 例如在崇明东滩三者虽然都可以作为湿地鸟类的饵料, 但不同的鸟类会取食不同植物的不同部分(Ma et al ., 2003 ; Zhu et al ., 2007 ), 且3种植物组织、器官发育和成熟时间也各不相同.本文基于崇明东滩南部各植物种群的细致调查, 明确了各主要植物种群的空间分布以及各自的面积, 为进一步研究植被分布格局变化以及重要鸟类觅食生境提供基础. ...
... 黄华梅等(2007) 认为, 影响崇明东滩植被分布格局的主要因素是互花米草入侵和围堤工程.但2014年互花米草治理工程实施后, 围堤外仍存在大面积的互花米草, 根据土壤盐度和潮滩高程的空间分布趋势, 互花米草具有向土壤盐度较高的东北方向扩张的可能, 广泛的高程适应范围又增加了其向南部扩张的几率, 在盐度和高程变化的影响下, 互花米草种群在发展过程中与海三棱藨草之间必然存在激烈的竞争关系, 这极有可能破坏该区植物群落的稳定现状.目前进行的互花米草生态治理工程虽未直接影响东滩南部的植被分布状况, 但是围堤之后, 大坝外围的水动力条件必然发生改变, 影响堤外泥沙淤积速率、土壤粒径等环境因子, 进而影响南部潮滩高程和土壤盐度, 改变植被空间格局.但具体的变化过程和趋势以及由此造成的影响还需要更长时间尺度的监测和分析, 其中本地特有物种海三棱藨草的面积和分布变化尤其值得关注. ...
1
1993
... 崇明东滩是长江口典型淤涨型潮滩湿地, 位于崇明岛最东端(121.83°-122.08° E, 31.42°-31.63° N), 属北亚热带季风气候, 四季分明, 日照充分, 雨量充沛, 气候温暖湿润, 年平均气温15.3 ℃, 年降水量1022 mm.这里是东亚-澳大利西亚迁徙路线上鸟类重要的停歇地, 也是小天鹅、白头鹤和黑脸琵鹭等珍稀濒危水禽重要的越冬栖息地(黄正一等, 1993 ).1997年崇明东滩被正式列入东亚-澳大利西亚涉禽保护区网络, 2002年又被《湿地公约》组织列入国际重要湿地名录(徐宏发和赵云龙, 2005 ), 2005年晋升为国家级自然保护区.本文研究区位于崇明东滩鸟类国家级自然保护区南部, 团结沙和东旺沙之间(121.90°-121.99° E, 31.45°-31.52° N), 北以互花米草生态治理工程2014年新筑大堤为界, 西至98大堤, 东、南两侧到达长江口外水域, 位于保护区的核心部分, 主要由团结沙外滩、大潮沟以及东旺沙外滩等植被覆盖的地区组成, 总面积约1800 hm2 (图1 ). ...
1
1993
... 崇明东滩是长江口典型淤涨型潮滩湿地, 位于崇明岛最东端(121.83°-122.08° E, 31.42°-31.63° N), 属北亚热带季风气候, 四季分明, 日照充分, 雨量充沛, 气候温暖湿润, 年平均气温15.3 ℃, 年降水量1022 mm.这里是东亚-澳大利西亚迁徙路线上鸟类重要的停歇地, 也是小天鹅、白头鹤和黑脸琵鹭等珍稀濒危水禽重要的越冬栖息地(黄正一等, 1993 ).1997年崇明东滩被正式列入东亚-澳大利西亚涉禽保护区网络, 2002年又被《湿地公约》组织列入国际重要湿地名录(徐宏发和赵云龙, 2005 ), 2005年晋升为国家级自然保护区.本文研究区位于崇明东滩鸟类国家级自然保护区南部, 团结沙和东旺沙之间(121.90°-121.99° E, 31.45°-31.52° N), 北以互花米草生态治理工程2014年新筑大堤为界, 西至98大堤, 东、南两侧到达长江口外水域, 位于保护区的核心部分, 主要由团结沙外滩、大潮沟以及东旺沙外滩等植被覆盖的地区组成, 总面积约1800 hm2 (图1 ). ...
Genotypic diversity and trait variance interact to affect marsh plant performance.
1
2014
... 盐沼植物作为全球范围内生产力较高的生物群落之一, 是推动湿地生态系统物质循环和能量流动的基础要素, 其在维持个体生存、种群稳定与群落发展的同时, 也为湿地生态系统中的众多成员提供了丰富的食物, 并创造了良好的栖息环境.然而, 近年来由于全球气候变化和频繁的人类活动, 植物群落组成结构以及空间分布格局发生了巨大的变化, 这不仅影响了湿地中鸟类、底栖生物及鱼类等生物的种群数量, 而且也不断改变着植物类群的种群结构, 甚至可能会导致一些敏感物种的灭绝(童春富, 2012 ; Isacch et al ., 2014 ; Li et al ., 2014b ).因此, 明确湿地中主要植物种群及其分布状况, 掌握其随环境因子的变化规律, 对湿地生态系统的保护、恢复、重建都具有重要的参考价值.虽然前人已就此进行了较多的研究(黄华梅等, 2007 ; Duarte et al ., 2013 ; 曹浩冰等, 2014 ; Hughes, 2014 ; Li et al ., 2014a , 2014b ), 但河口潮滩区域海陆交互作用强烈、人类活动频繁, 使得滩面、潮沟及植被都处于不断变化中, 崇明东滩正是这样一个处在发育过程中的典型盐沼湿地生态系统.2014年崇明东滩互花米草(Spartina alterniflora )生态治理工程的实施, 可能对该区域盐沼植物的空间分布及其生境产生了重大影响(高宇和赵斌, 2006 ; Zhu et al ., 2014 ).因此, 在施工前期对围垦外围区进行全面的植被调查, 不仅能准确地掌握崇明东滩南部的植被类型和现有分布状况, 而且能为今后研究其植被空间动态变化提供基础数据. ...
Diversity and habitat distribution of birds in coastal marshes and comparisons with surrounding upland habitats in Southeastern South America.
1
2014
... 盐沼植物作为全球范围内生产力较高的生物群落之一, 是推动湿地生态系统物质循环和能量流动的基础要素, 其在维持个体生存、种群稳定与群落发展的同时, 也为湿地生态系统中的众多成员提供了丰富的食物, 并创造了良好的栖息环境.然而, 近年来由于全球气候变化和频繁的人类活动, 植物群落组成结构以及空间分布格局发生了巨大的变化, 这不仅影响了湿地中鸟类、底栖生物及鱼类等生物的种群数量, 而且也不断改变着植物类群的种群结构, 甚至可能会导致一些敏感物种的灭绝(童春富, 2012 ; Isacch et al ., 2014 ; Li et al ., 2014b ).因此, 明确湿地中主要植物种群及其分布状况, 掌握其随环境因子的变化规律, 对湿地生态系统的保护、恢复、重建都具有重要的参考价值.虽然前人已就此进行了较多的研究(黄华梅等, 2007 ; Duarte et al ., 2013 ; 曹浩冰等, 2014 ; Hughes, 2014 ; Li et al ., 2014a , 2014b ), 但河口潮滩区域海陆交互作用强烈、人类活动频繁, 使得滩面、潮沟及植被都处于不断变化中, 崇明东滩正是这样一个处在发育过程中的典型盐沼湿地生态系统.2014年崇明东滩互花米草(Spartina alterniflora )生态治理工程的实施, 可能对该区域盐沼植物的空间分布及其生境产生了重大影响(高宇和赵斌, 2006 ; Zhu et al ., 2014 ).因此, 在施工前期对围垦外围区进行全面的植被调查, 不仅能准确地掌握崇明东滩南部的植被类型和现有分布状况, 而且能为今后研究其植被空间动态变化提供基础数据. ...
长江口崇明东滩不同植被带沉积速率研究
1
2012
... 长江入海径流以崇明岛为界, 形成南、北两个一级支流, 两侧入海淡水不均衡, 南支淡水充裕, 北支偏少, 加之涨潮时北支经常出现咸水倒灌(贺松林等, 2006 ), 因此, 崇明东滩南部水体盐度明显低于北部.受潮汐影响, 海水与淡水在混合过程中又形成了新的盐度梯度, 即低潮带盐度高于中、高潮滩(陈中义等, 2005a ; 贺松林等, 2006 ; 曹利利等, 2012 ; 王卿等, 2012 ).与此同时, 径流中所携带的大量泥沙在岛堤之外沉积并发育形成新的具有不同潮滩高程的滩地, 受潮汐影响, 滩面水流在涨、落潮过程中不断冲刷滩面, 而崇明东滩南部植被相对稀疏, 因此在南部潮滩上形成多条宽窄不一、错综复杂的潮沟(张修桂, 2005 ; 姜亦飞等, 2012 ; 刘建华等, 2012 ; 陈勇等, 2013 ), 这在一定程度上限制了人类活动对保护区的影响. ...
长江口崇明东滩不同植被带沉积速率研究
1
2012
... 长江入海径流以崇明岛为界, 形成南、北两个一级支流, 两侧入海淡水不均衡, 南支淡水充裕, 北支偏少, 加之涨潮时北支经常出现咸水倒灌(贺松林等, 2006 ), 因此, 崇明东滩南部水体盐度明显低于北部.受潮汐影响, 海水与淡水在混合过程中又形成了新的盐度梯度, 即低潮带盐度高于中、高潮滩(陈中义等, 2005a ; 贺松林等, 2006 ; 曹利利等, 2012 ; 王卿等, 2012 ).与此同时, 径流中所携带的大量泥沙在岛堤之外沉积并发育形成新的具有不同潮滩高程的滩地, 受潮汐影响, 滩面水流在涨、落潮过程中不断冲刷滩面, 而崇明东滩南部植被相对稀疏, 因此在南部潮滩上形成多条宽窄不一、错综复杂的潮沟(张修桂, 2005 ; 姜亦飞等, 2012 ; 刘建华等, 2012 ; 陈勇等, 2013 ), 这在一定程度上限制了人类活动对保护区的影响. ...
Spartina alterniflora invasions in the Yangtze River estuary, China: An overview of current status and ecosystem effects.
1
2009
... 研究区东北部的互花米草也因较高的盖度值成为优势种, 同时, 无性繁殖方式, 使得种群扩散速度快、发育周期短, 并能够以母体为中心快速向周围空间扩散, 加之互花米草萌发时间早, 植物生长迅速, 抑制相邻其他植物的正常生长, 在其分布区内占有绝对优势(Li et al ., 2009 ; Lu & Zhang, 2013 ; Liu et al ., 2014 ; 徐伟伟等, 2014 ).研究区内互花米草与糙叶薹草、海三棱藨草、芦苇的邻接带分别长达1.2 km、2.0 km、3.0 km (图3 ), 这为互花米草向周围快速扩散提供了可能.与芦苇和互花米草不同, 海三棱藨草种群破碎化明显, 虽然外侧与光滩的边界长达2.4 km, 为种群向光滩扩张创造了良好的演替条件, 但其群落中部被芦苇和互花米草等植物所隔离, 外部受潮汐以及波浪作用明显, 因此生长条件比其他物种更为恶劣, 种群优势因此降低.其余3种植物空间分布形式各不相同, 白茅空间分布不均, 种群连接度不高, 斑块面积小, 但是数量较多, 因此种群总面积能占一定优势.糙叶薹草种群连接度高, 种群斑块面积大, 分布广, 能够承受较高的环境风险.藨草虽然在“外带”附近种群分布连续, 且面积较大, 但夹在糙叶薹草内部的藨草种群面积小, 较为分散.所有植物优势种群在东北至西南方向上都呈明显的条带状分布. ...
Indirect effects of non-native Spartina alterniflora and its fungal pathogen (Fusarium palustre) on native saltmarsh plants in China.
1
2014a)
... 盐沼植物作为全球范围内生产力较高的生物群落之一, 是推动湿地生态系统物质循环和能量流动的基础要素, 其在维持个体生存、种群稳定与群落发展的同时, 也为湿地生态系统中的众多成员提供了丰富的食物, 并创造了良好的栖息环境.然而, 近年来由于全球气候变化和频繁的人类活动, 植物群落组成结构以及空间分布格局发生了巨大的变化, 这不仅影响了湿地中鸟类、底栖生物及鱼类等生物的种群数量, 而且也不断改变着植物类群的种群结构, 甚至可能会导致一些敏感物种的灭绝(童春富, 2012 ; Isacch et al ., 2014 ; Li et al ., 2014b ).因此, 明确湿地中主要植物种群及其分布状况, 掌握其随环境因子的变化规律, 对湿地生态系统的保护、恢复、重建都具有重要的参考价值.虽然前人已就此进行了较多的研究(黄华梅等, 2007 ; Duarte et al ., 2013 ; 曹浩冰等, 2014 ; Hughes, 2014 ; Li et al ., 2014a , 2014b ), 但河口潮滩区域海陆交互作用强烈、人类活动频繁, 使得滩面、潮沟及植被都处于不断变化中, 崇明东滩正是这样一个处在发育过程中的典型盐沼湿地生态系统.2014年崇明东滩互花米草(Spartina alterniflora )生态治理工程的实施, 可能对该区域盐沼植物的空间分布及其生境产生了重大影响(高宇和赵斌, 2006 ; Zhu et al ., 2014 ).因此, 在施工前期对围垦外围区进行全面的植被调查, 不仅能准确地掌握崇明东滩南部的植被类型和现有分布状况, 而且能为今后研究其植被空间动态变化提供基础数据. ...
The impact of the change in vegetation structure on the ecological functions of salt marshes: The example of the Yangtze estuary.
2
2014b
... 盐沼植物作为全球范围内生产力较高的生物群落之一, 是推动湿地生态系统物质循环和能量流动的基础要素, 其在维持个体生存、种群稳定与群落发展的同时, 也为湿地生态系统中的众多成员提供了丰富的食物, 并创造了良好的栖息环境.然而, 近年来由于全球气候变化和频繁的人类活动, 植物群落组成结构以及空间分布格局发生了巨大的变化, 这不仅影响了湿地中鸟类、底栖生物及鱼类等生物的种群数量, 而且也不断改变着植物类群的种群结构, 甚至可能会导致一些敏感物种的灭绝(童春富, 2012 ; Isacch et al ., 2014 ; Li et al ., 2014b ).因此, 明确湿地中主要植物种群及其分布状况, 掌握其随环境因子的变化规律, 对湿地生态系统的保护、恢复、重建都具有重要的参考价值.虽然前人已就此进行了较多的研究(黄华梅等, 2007 ; Duarte et al ., 2013 ; 曹浩冰等, 2014 ; Hughes, 2014 ; Li et al ., 2014a , 2014b ), 但河口潮滩区域海陆交互作用强烈、人类活动频繁, 使得滩面、潮沟及植被都处于不断变化中, 崇明东滩正是这样一个处在发育过程中的典型盐沼湿地生态系统.2014年崇明东滩互花米草(Spartina alterniflora )生态治理工程的实施, 可能对该区域盐沼植物的空间分布及其生境产生了重大影响(高宇和赵斌, 2006 ; Zhu et al ., 2014 ).因此, 在施工前期对围垦外围区进行全面的植被调查, 不仅能准确地掌握崇明东滩南部的植被类型和现有分布状况, 而且能为今后研究其植被空间动态变化提供基础数据. ...
... , 2014b ), 但河口潮滩区域海陆交互作用强烈、人类活动频繁, 使得滩面、潮沟及植被都处于不断变化中, 崇明东滩正是这样一个处在发育过程中的典型盐沼湿地生态系统.2014年崇明东滩互花米草(Spartina alterniflora )生态治理工程的实施, 可能对该区域盐沼植物的空间分布及其生境产生了重大影响(高宇和赵斌, 2006 ; Zhu et al ., 2014 ).因此, 在施工前期对围垦外围区进行全面的植被调查, 不仅能准确地掌握崇明东滩南部的植被类型和现有分布状况, 而且能为今后研究其植被空间动态变化提供基础数据. ...
The relative importance of sexual and asexual reproduction in the spread of Spartina alterniflora using a spatially explicit individual-based model.
1
2014
... 研究区东北部的互花米草也因较高的盖度值成为优势种, 同时, 无性繁殖方式, 使得种群扩散速度快、发育周期短, 并能够以母体为中心快速向周围空间扩散, 加之互花米草萌发时间早, 植物生长迅速, 抑制相邻其他植物的正常生长, 在其分布区内占有绝对优势(Li et al ., 2009 ; Lu & Zhang, 2013 ; Liu et al ., 2014 ; 徐伟伟等, 2014 ).研究区内互花米草与糙叶薹草、海三棱藨草、芦苇的邻接带分别长达1.2 km、2.0 km、3.0 km (图3 ), 这为互花米草向周围快速扩散提供了可能.与芦苇和互花米草不同, 海三棱藨草种群破碎化明显, 虽然外侧与光滩的边界长达2.4 km, 为种群向光滩扩张创造了良好的演替条件, 但其群落中部被芦苇和互花米草等植物所隔离, 外部受潮汐以及波浪作用明显, 因此生长条件比其他物种更为恶劣, 种群优势因此降低.其余3种植物空间分布形式各不相同, 白茅空间分布不均, 种群连接度不高, 斑块面积小, 但是数量较多, 因此种群总面积能占一定优势.糙叶薹草种群连接度高, 种群斑块面积大, 分布广, 能够承受较高的环境风险.藨草虽然在“外带”附近种群分布连续, 且面积较大, 但夹在糙叶薹草内部的藨草种群面积小, 较为分散.所有植物优势种群在东北至西南方向上都呈明显的条带状分布. ...
长江口崇明东滩潮沟地貌形态和演变
1
2012
... 长江入海径流以崇明岛为界, 形成南、北两个一级支流, 两侧入海淡水不均衡, 南支淡水充裕, 北支偏少, 加之涨潮时北支经常出现咸水倒灌(贺松林等, 2006 ), 因此, 崇明东滩南部水体盐度明显低于北部.受潮汐影响, 海水与淡水在混合过程中又形成了新的盐度梯度, 即低潮带盐度高于中、高潮滩(陈中义等, 2005a ; 贺松林等, 2006 ; 曹利利等, 2012 ; 王卿等, 2012 ).与此同时, 径流中所携带的大量泥沙在岛堤之外沉积并发育形成新的具有不同潮滩高程的滩地, 受潮汐影响, 滩面水流在涨、落潮过程中不断冲刷滩面, 而崇明东滩南部植被相对稀疏, 因此在南部潮滩上形成多条宽窄不一、错综复杂的潮沟(张修桂, 2005 ; 姜亦飞等, 2012 ; 刘建华等, 2012 ; 陈勇等, 2013 ), 这在一定程度上限制了人类活动对保护区的影响. ...
长江口崇明东滩潮沟地貌形态和演变
1
2012
... 长江入海径流以崇明岛为界, 形成南、北两个一级支流, 两侧入海淡水不均衡, 南支淡水充裕, 北支偏少, 加之涨潮时北支经常出现咸水倒灌(贺松林等, 2006 ), 因此, 崇明东滩南部水体盐度明显低于北部.受潮汐影响, 海水与淡水在混合过程中又形成了新的盐度梯度, 即低潮带盐度高于中、高潮滩(陈中义等, 2005a ; 贺松林等, 2006 ; 曹利利等, 2012 ; 王卿等, 2012 ).与此同时, 径流中所携带的大量泥沙在岛堤之外沉积并发育形成新的具有不同潮滩高程的滩地, 受潮汐影响, 滩面水流在涨、落潮过程中不断冲刷滩面, 而崇明东滩南部植被相对稀疏, 因此在南部潮滩上形成多条宽窄不一、错综复杂的潮沟(张修桂, 2005 ; 姜亦飞等, 2012 ; 刘建华等, 2012 ; 陈勇等, 2013 ), 这在一定程度上限制了人类活动对保护区的影响. ...
Spatial distribution of an invasive plant Spartina alterniflora and its potential as biofuels in China
1
2013
... 研究区东北部的互花米草也因较高的盖度值成为优势种, 同时, 无性繁殖方式, 使得种群扩散速度快、发育周期短, 并能够以母体为中心快速向周围空间扩散, 加之互花米草萌发时间早, 植物生长迅速, 抑制相邻其他植物的正常生长, 在其分布区内占有绝对优势(Li et al ., 2009 ; Lu & Zhang, 2013 ; Liu et al ., 2014 ; 徐伟伟等, 2014 ).研究区内互花米草与糙叶薹草、海三棱藨草、芦苇的邻接带分别长达1.2 km、2.0 km、3.0 km (图3 ), 这为互花米草向周围快速扩散提供了可能.与芦苇和互花米草不同, 海三棱藨草种群破碎化明显, 虽然外侧与光滩的边界长达2.4 km, 为种群向光滩扩张创造了良好的演替条件, 但其群落中部被芦苇和互花米草等植物所隔离, 外部受潮汐以及波浪作用明显, 因此生长条件比其他物种更为恶劣, 种群优势因此降低.其余3种植物空间分布形式各不相同, 白茅空间分布不均, 种群连接度不高, 斑块面积小, 但是数量较多, 因此种群总面积能占一定优势.糙叶薹草种群连接度高, 种群斑块面积大, 分布广, 能够承受较高的环境风险.藨草虽然在“外带”附近种群分布连续, 且面积较大, 但夹在糙叶薹草内部的藨草种群面积小, 较为分散.所有植物优势种群在东北至西南方向上都呈明显的条带状分布. ...
Effects of tidewater on the feeding ecology of hooded crane (Grus monacha) and conservation of their wintering habitats at Chongming Dongtan, China.
3
2003
... 上述植物中, 海三棱藨草是中国特有的盐沼植物, 也是典型的潮滩先锋物种, 目前主要分布于长江口沿岸的滩涂湿地(Sun et al ., 2002 , 2003 ; 梅雪英和张修峰, 2007 ; 陈秀芝和孙瑛, 2011 ).崇明东滩作为长江口最大的天然湿地, 有大量的海三棱藨草分布(虞快, 1994 , 1996 ), 但近年来崇明东滩北部互花米草生长泛滥, 海三棱藨草原有生态位受到挤压, 已基本被互花米草取代(陈秀芝和孙瑛, 2011 ), 东旺沙中间(2014年新围垦大堤东南部外围)是海三棱藨草现存种群主要分布区, 同时也是莎草科类群的主要分布区1) (1) 吴纪华 (2014). 上海崇明东滩鸟类国家级自然保护区2012年度资源监测公报, 14-15.) .海三棱藨草在崇明东滩形成湿地草场, 能为鸻鹬类、雁鸭类、鹤类、鸥类等众多越冬候鸟提供天然的觅食、活动、休憩场所, 同时海三棱藨草的根茎和球茎, 还是国家一级保护鸟类——白头鹤的重要饵料(Ma et al ., 2003 ; Zhu et al ., 2007 ), 因此, 海三棱藨草对崇明东滩所有滨海湿地鸟类都显得至关重要.然而近年来滩涂围垦、堤外放牧及外来物种竞争等导致植物种群分布状况发生变化, 植物群落斑块破碎化严重, 整个生态系统服务功能也随之降低(张利权和雍学葵, 1992; Chen et al ., 2004; 陈中义等, 2005b; Wang et al ., 2010).因此, 确定包括海三棱藨草在内的关键植物种群在东滩的空间分布, 明确其随环境因子的变化规律, 对该湿地生态系统的修复和保护具有重要的指导意义. ...
... 在前人相关研究(Ma et al ., 2003 ; 陈中义等, 2005a ; 贺强等, 2009 ; 詹跃武等, 2011 )的基础上, 本研究选择潮滩高程和表层30 cm土壤盐度作为主要参数, 探讨盐沼植物在东滩南部的空间分布及其与两大环境因子的相互关系. ...
... 崇明东滩属于典型的淤涨型潮滩, 目前滩涂植被覆盖区总面积4265 hm2 , 2012年调查数据显示, 崇明东滩中部、北部植物以芦苇和互花米草为主, 而莎草科植物主要集中分布在崇明东滩南部1) ( 1) 吴纪华 (2014). 上海崇明东滩鸟类国家级自然保护区2012年度资源监测公报, 14-15.) .在崇明东滩的相关研究中, 众多学者根据研究需要将崇明东滩的海三棱藨草、藨草、糙叶薹草3种植物统一归入海三棱藨草类群(黄华梅等, 2007 ; Yuan & Zhang, 2008 ; 严格等, 2014 ).部分研究虽然将海三棱藨草、藨草、糙叶薹草统一划分为莎草科植物群落或者藨草群落(曹浩冰等, 2014 ; 任璘婧等, 2014 ), 但没有对这3种植物进行细致的划分, 各种群面积大小及其具体分布目前尚无相关报道.上述3种植物虽然同属于莎草科, 但占据不同的生态位, 承担着不同的生态功能, 例如在崇明东滩三者虽然都可以作为湿地鸟类的饵料, 但不同的鸟类会取食不同植物的不同部分(Ma et al ., 2003 ; Zhu et al ., 2007 ), 且3种植物组织、器官发育和成熟时间也各不相同.本文基于崇明东滩南部各植物种群的细致调查, 明确了各主要植物种群的空间分布以及各自的面积, 为进一步研究植被分布格局变化以及重要鸟类觅食生境提供基础. ...
长江口湿地海三棱藨草(Scirpus mariqueter)的储碳、固碳功能研究——以崇明东滩为例
1
2007
... 上述植物中, 海三棱藨草是中国特有的盐沼植物, 也是典型的潮滩先锋物种, 目前主要分布于长江口沿岸的滩涂湿地(Sun et al ., 2002 , 2003 ; 梅雪英和张修峰, 2007 ; 陈秀芝和孙瑛, 2011 ).崇明东滩作为长江口最大的天然湿地, 有大量的海三棱藨草分布(虞快, 1994 , 1996 ), 但近年来崇明东滩北部互花米草生长泛滥, 海三棱藨草原有生态位受到挤压, 已基本被互花米草取代(陈秀芝和孙瑛, 2011 ), 东旺沙中间(2014年新围垦大堤东南部外围)是海三棱藨草现存种群主要分布区, 同时也是莎草科类群的主要分布区1) (1) 吴纪华 (2014). 上海崇明东滩鸟类国家级自然保护区2012年度资源监测公报, 14-15.) .海三棱藨草在崇明东滩形成湿地草场, 能为鸻鹬类、雁鸭类、鹤类、鸥类等众多越冬候鸟提供天然的觅食、活动、休憩场所, 同时海三棱藨草的根茎和球茎, 还是国家一级保护鸟类——白头鹤的重要饵料(Ma et al ., 2003 ; Zhu et al ., 2007 ), 因此, 海三棱藨草对崇明东滩所有滨海湿地鸟类都显得至关重要.然而近年来滩涂围垦、堤外放牧及外来物种竞争等导致植物种群分布状况发生变化, 植物群落斑块破碎化严重, 整个生态系统服务功能也随之降低(张利权和雍学葵, 1992; Chen et al ., 2004; 陈中义等, 2005b; Wang et al ., 2010).因此, 确定包括海三棱藨草在内的关键植物种群在东滩的空间分布, 明确其随环境因子的变化规律, 对该湿地生态系统的修复和保护具有重要的指导意义. ...
长江口湿地海三棱藨草(Scirpus mariqueter)的储碳、固碳功能研究——以崇明东滩为例
1
2007
... 上述植物中, 海三棱藨草是中国特有的盐沼植物, 也是典型的潮滩先锋物种, 目前主要分布于长江口沿岸的滩涂湿地(Sun et al ., 2002 , 2003 ; 梅雪英和张修峰, 2007 ; 陈秀芝和孙瑛, 2011 ).崇明东滩作为长江口最大的天然湿地, 有大量的海三棱藨草分布(虞快, 1994 , 1996 ), 但近年来崇明东滩北部互花米草生长泛滥, 海三棱藨草原有生态位受到挤压, 已基本被互花米草取代(陈秀芝和孙瑛, 2011 ), 东旺沙中间(2014年新围垦大堤东南部外围)是海三棱藨草现存种群主要分布区, 同时也是莎草科类群的主要分布区1) (1) 吴纪华 (2014). 上海崇明东滩鸟类国家级自然保护区2012年度资源监测公报, 14-15.) .海三棱藨草在崇明东滩形成湿地草场, 能为鸻鹬类、雁鸭类、鹤类、鸥类等众多越冬候鸟提供天然的觅食、活动、休憩场所, 同时海三棱藨草的根茎和球茎, 还是国家一级保护鸟类——白头鹤的重要饵料(Ma et al ., 2003 ; Zhu et al ., 2007 ), 因此, 海三棱藨草对崇明东滩所有滨海湿地鸟类都显得至关重要.然而近年来滩涂围垦、堤外放牧及外来物种竞争等导致植物种群分布状况发生变化, 植物群落斑块破碎化严重, 整个生态系统服务功能也随之降低(张利权和雍学葵, 1992; Chen et al ., 2004; 陈中义等, 2005b; Wang et al ., 2010).因此, 确定包括海三棱藨草在内的关键植物种群在东滩的空间分布, 明确其随环境因子的变化规律, 对该湿地生态系统的修复和保护具有重要的指导意义. ...
崇明东滩盐沼植被变化对滩涂湿地促淤消浪功能的影响
1
2014
... 崇明东滩属于典型的淤涨型潮滩, 目前滩涂植被覆盖区总面积4265 hm2 , 2012年调查数据显示, 崇明东滩中部、北部植物以芦苇和互花米草为主, 而莎草科植物主要集中分布在崇明东滩南部1) ( 1) 吴纪华 (2014). 上海崇明东滩鸟类国家级自然保护区2012年度资源监测公报, 14-15.) .在崇明东滩的相关研究中, 众多学者根据研究需要将崇明东滩的海三棱藨草、藨草、糙叶薹草3种植物统一归入海三棱藨草类群(黄华梅等, 2007 ; Yuan & Zhang, 2008 ; 严格等, 2014 ).部分研究虽然将海三棱藨草、藨草、糙叶薹草统一划分为莎草科植物群落或者藨草群落(曹浩冰等, 2014 ; 任璘婧等, 2014 ), 但没有对这3种植物进行细致的划分, 各种群面积大小及其具体分布目前尚无相关报道.上述3种植物虽然同属于莎草科, 但占据不同的生态位, 承担着不同的生态功能, 例如在崇明东滩三者虽然都可以作为湿地鸟类的饵料, 但不同的鸟类会取食不同植物的不同部分(Ma et al ., 2003 ; Zhu et al ., 2007 ), 且3种植物组织、器官发育和成熟时间也各不相同.本文基于崇明东滩南部各植物种群的细致调查, 明确了各主要植物种群的空间分布以及各自的面积, 为进一步研究植被分布格局变化以及重要鸟类觅食生境提供基础. ...
崇明东滩盐沼植被变化对滩涂湿地促淤消浪功能的影响
1
2014
... 崇明东滩属于典型的淤涨型潮滩, 目前滩涂植被覆盖区总面积4265 hm2 , 2012年调查数据显示, 崇明东滩中部、北部植物以芦苇和互花米草为主, 而莎草科植物主要集中分布在崇明东滩南部1) ( 1) 吴纪华 (2014). 上海崇明东滩鸟类国家级自然保护区2012年度资源监测公报, 14-15.) .在崇明东滩的相关研究中, 众多学者根据研究需要将崇明东滩的海三棱藨草、藨草、糙叶薹草3种植物统一归入海三棱藨草类群(黄华梅等, 2007 ; Yuan & Zhang, 2008 ; 严格等, 2014 ).部分研究虽然将海三棱藨草、藨草、糙叶薹草统一划分为莎草科植物群落或者藨草群落(曹浩冰等, 2014 ; 任璘婧等, 2014 ), 但没有对这3种植物进行细致的划分, 各种群面积大小及其具体分布目前尚无相关报道.上述3种植物虽然同属于莎草科, 但占据不同的生态位, 承担着不同的生态功能, 例如在崇明东滩三者虽然都可以作为湿地鸟类的饵料, 但不同的鸟类会取食不同植物的不同部分(Ma et al ., 2003 ; Zhu et al ., 2007 ), 且3种植物组织、器官发育和成熟时间也各不相同.本文基于崇明东滩南部各植物种群的细致调查, 明确了各主要植物种群的空间分布以及各自的面积, 为进一步研究植被分布格局变化以及重要鸟类觅食生境提供基础. ...
Tidal regime, salinity and salt marsh plant zonation.
1
2005
... 潮滩植物群落在空间上呈现不同的分布状况, 潮滩高程作为影响植物分布的重要环境因子, 与植物的淹水时间和频度, 水体中的溶解氧, 土壤粒径、养分、氧化还原电位、pH值等生境因子存在密切联系(Silvestri et al ., 2005 ; 何彦龙, 2014 ), 与此同时, 植物的密度、高度等因素也会影响高程的变化(Duarte et al ., 2013 ).本研究表明崇明东滩南部植被覆盖空间6种主要植物分布在4个差异显著的高程区间内(p < 0.01), 黄华梅等(2007)和严格等(2014)认为崇明东滩芦苇分布在高潮带(高程>2.9 m), 海三棱藨草分布在中潮带(高程2.0-2.9 m), 互花米草在中、高潮带都有分布.同时对比何彦龙等(2010)在2008年对崇明东滩互花米草(3.21 ± 0.01 m)和海三棱藨草(3.10 ± 0.02 m)的研究结果, 发现上述植物在高程数值上有所差异, 但各植物种群生长空间的高程相对大小保持稳定, 表明在崇明东滩无论是大尺度区域范围, 还是小尺度空间, 各主要植物类群的潮滩高程相对高度保持恒定, 因此潮滩高程是决定植被空间分布的重要因素, 而具体数值差异可能是由于野外调查的时间、地点以及高程仪器精度差异所致.盐度是植物生长过程中的又一重要条件性因子, 不同的植物在不同的生长阶段适应不同范围的盐度(Zhang et al ., 2011 ).陈中义等(2005b)研究了水体中盐度的变化对互花米草和海三棱藨草的影响, 认为互花米草更适应高盐度海水.本研究中互花米草生长空间的土壤盐度确实显著高于海三棱藨草区(p < 0.01), 除了与互花米草本身的高耐盐特性有关, 与植株地上部分的泌盐作用也有一定关系(He et al ., 2011 ).但是无论崇明东滩土壤盐度还是潮滩高程都直接受到潮汐影响, 在空间上都呈现梯度变化(王卿等, 2012 ), 因此, 崇明东滩南部6种主要植物受到两种限定因子的共同制约, 在空间上形成现有的分布格局, 而对潮滩高程和土壤盐度在不同空间尺度下作用的权重仍有待进一步研究. ...
Differences in morphology and biomass allocation of Scirpus mariqueter between creekside and inland communities in the Changjiang estuary, China.
1
2002
... 上述植物中, 海三棱藨草是中国特有的盐沼植物, 也是典型的潮滩先锋物种, 目前主要分布于长江口沿岸的滩涂湿地(Sun et al ., 2002 , 2003 ; 梅雪英和张修峰, 2007 ; 陈秀芝和孙瑛, 2011 ).崇明东滩作为长江口最大的天然湿地, 有大量的海三棱藨草分布(虞快, 1994 , 1996 ), 但近年来崇明东滩北部互花米草生长泛滥, 海三棱藨草原有生态位受到挤压, 已基本被互花米草取代(陈秀芝和孙瑛, 2011 ), 东旺沙中间(2014年新围垦大堤东南部外围)是海三棱藨草现存种群主要分布区, 同时也是莎草科类群的主要分布区1) (1) 吴纪华 (2014). 上海崇明东滩鸟类国家级自然保护区2012年度资源监测公报, 14-15.) .海三棱藨草在崇明东滩形成湿地草场, 能为鸻鹬类、雁鸭类、鹤类、鸥类等众多越冬候鸟提供天然的觅食、活动、休憩场所, 同时海三棱藨草的根茎和球茎, 还是国家一级保护鸟类——白头鹤的重要饵料(Ma et al ., 2003 ; Zhu et al ., 2007 ), 因此, 海三棱藨草对崇明东滩所有滨海湿地鸟类都显得至关重要.然而近年来滩涂围垦、堤外放牧及外来物种竞争等导致植物种群分布状况发生变化, 植物群落斑块破碎化严重, 整个生态系统服务功能也随之降低(张利权和雍学葵, 1992; Chen et al ., 2004; 陈中义等, 2005b; Wang et al ., 2010).因此, 确定包括海三棱藨草在内的关键植物种群在东滩的空间分布, 明确其随环境因子的变化规律, 对该湿地生态系统的修复和保护具有重要的指导意义. ...
Salt marsh vegetation change after a short-term tidal restriction in the Changjiang estuary.
1
2003
... 上述植物中, 海三棱藨草是中国特有的盐沼植物, 也是典型的潮滩先锋物种, 目前主要分布于长江口沿岸的滩涂湿地(Sun et al ., 2002 , 2003 ; 梅雪英和张修峰, 2007 ; 陈秀芝和孙瑛, 2011 ).崇明东滩作为长江口最大的天然湿地, 有大量的海三棱藨草分布(虞快, 1994 , 1996 ), 但近年来崇明东滩北部互花米草生长泛滥, 海三棱藨草原有生态位受到挤压, 已基本被互花米草取代(陈秀芝和孙瑛, 2011 ), 东旺沙中间(2014年新围垦大堤东南部外围)是海三棱藨草现存种群主要分布区, 同时也是莎草科类群的主要分布区1) (1) 吴纪华 (2014). 上海崇明东滩鸟类国家级自然保护区2012年度资源监测公报, 14-15.) .海三棱藨草在崇明东滩形成湿地草场, 能为鸻鹬类、雁鸭类、鹤类、鸥类等众多越冬候鸟提供天然的觅食、活动、休憩场所, 同时海三棱藨草的根茎和球茎, 还是国家一级保护鸟类——白头鹤的重要饵料(Ma et al ., 2003 ; Zhu et al ., 2007 ), 因此, 海三棱藨草对崇明东滩所有滨海湿地鸟类都显得至关重要.然而近年来滩涂围垦、堤外放牧及外来物种竞争等导致植物种群分布状况发生变化, 植物群落斑块破碎化严重, 整个生态系统服务功能也随之降低(张利权和雍学葵, 1992; Chen et al ., 2004; 陈中义等, 2005b; Wang et al ., 2010).因此, 确定包括海三棱藨草在内的关键植物种群在东滩的空间分布, 明确其随环境因子的变化规律, 对该湿地生态系统的修复和保护具有重要的指导意义. ...
Analyzing the habitat suitability for migratory birds at the Chongming Dongtan Nature Reserve in Shanghai, China.
1
2008
... 近十几年来, 大量研究人员通过遥感影像解译, 对崇明东滩主要植被进行了分区(Gao & Zhang, 2006 ; Tian et al ., 2008 ; 阿也提古丽·斯迪克等, 2010 ; 翟新源和杨苏新, 2013 ; 郑宗生等, 2010 ; 宗玮等, 2011 ), 普遍认为芦苇(Phragmites australis )、互花米草(Spartina alterniflora )及海三棱藨草(Scirpus mariqueter )是崇明东滩的“三大”主要植物类群.然而, 崇明东滩南部长期受长江入海淡水影响, 水体盐度相对较低, 在团结沙至大潮沟(图1 )存在较大面积的藨草(Scirpus triqueter )种群区(闫芊等, 2007 ; 章振亚等, 2012 ).同时发现, 糙叶薹草(Carex scabrifolia )在研究区内频繁出现, 白茅(Imperata cylindrica )在部分空间也已发展成优势种. ...
长江河口潮间带盐沼植被分布区及邻近光滩鱼类组成特征
1
2012
... 盐沼植物作为全球范围内生产力较高的生物群落之一, 是推动湿地生态系统物质循环和能量流动的基础要素, 其在维持个体生存、种群稳定与群落发展的同时, 也为湿地生态系统中的众多成员提供了丰富的食物, 并创造了良好的栖息环境.然而, 近年来由于全球气候变化和频繁的人类活动, 植物群落组成结构以及空间分布格局发生了巨大的变化, 这不仅影响了湿地中鸟类、底栖生物及鱼类等生物的种群数量, 而且也不断改变着植物类群的种群结构, 甚至可能会导致一些敏感物种的灭绝(童春富, 2012 ; Isacch et al ., 2014 ; Li et al ., 2014b ).因此, 明确湿地中主要植物种群及其分布状况, 掌握其随环境因子的变化规律, 对湿地生态系统的保护、恢复、重建都具有重要的参考价值.虽然前人已就此进行了较多的研究(黄华梅等, 2007 ; Duarte et al ., 2013 ; 曹浩冰等, 2014 ; Hughes, 2014 ; Li et al ., 2014a , 2014b ), 但河口潮滩区域海陆交互作用强烈、人类活动频繁, 使得滩面、潮沟及植被都处于不断变化中, 崇明东滩正是这样一个处在发育过程中的典型盐沼湿地生态系统.2014年崇明东滩互花米草(Spartina alterniflora )生态治理工程的实施, 可能对该区域盐沼植物的空间分布及其生境产生了重大影响(高宇和赵斌, 2006 ; Zhu et al ., 2014 ).因此, 在施工前期对围垦外围区进行全面的植被调查, 不仅能准确地掌握崇明东滩南部的植被类型和现有分布状况, 而且能为今后研究其植被空间动态变化提供基础数据. ...
长江河口潮间带盐沼植被分布区及邻近光滩鱼类组成特征
1
2012
... 盐沼植物作为全球范围内生产力较高的生物群落之一, 是推动湿地生态系统物质循环和能量流动的基础要素, 其在维持个体生存、种群稳定与群落发展的同时, 也为湿地生态系统中的众多成员提供了丰富的食物, 并创造了良好的栖息环境.然而, 近年来由于全球气候变化和频繁的人类活动, 植物群落组成结构以及空间分布格局发生了巨大的变化, 这不仅影响了湿地中鸟类、底栖生物及鱼类等生物的种群数量, 而且也不断改变着植物类群的种群结构, 甚至可能会导致一些敏感物种的灭绝(童春富, 2012 ; Isacch et al ., 2014 ; Li et al ., 2014b ).因此, 明确湿地中主要植物种群及其分布状况, 掌握其随环境因子的变化规律, 对湿地生态系统的保护、恢复、重建都具有重要的参考价值.虽然前人已就此进行了较多的研究(黄华梅等, 2007 ; Duarte et al ., 2013 ; 曹浩冰等, 2014 ; Hughes, 2014 ; Li et al ., 2014a , 2014b ), 但河口潮滩区域海陆交互作用强烈、人类活动频繁, 使得滩面、潮沟及植被都处于不断变化中, 崇明东滩正是这样一个处在发育过程中的典型盐沼湿地生态系统.2014年崇明东滩互花米草(Spartina alterniflora )生态治理工程的实施, 可能对该区域盐沼植物的空间分布及其生境产生了重大影响(高宇和赵斌, 2006 ; Zhu et al ., 2014 ).因此, 在施工前期对围垦外围区进行全面的植被调查, 不仅能准确地掌握崇明东滩南部的植被类型和现有分布状况, 而且能为今后研究其植被空间动态变化提供基础数据. ...
Effects of environmental gradients on the performances of four dominant plants in a Chinese saltmarsh: Implications for plant zonation.
2
2010
... 为明确植被与潮滩高程因子之间的关系, 我们对各植物类群分别进行显著性差异分析, 发现除了白茅和芦苇, 藨草和海三棱藨草两两之间无显著性差异外, 其余各植物群落之间差异非常显著(p < 0.01).各物种相应高程分别为白茅(3.85 ± 0.32 m)≈芦苇(3.75 ± 0.57 m) >糙叶薹草(3.64 ± 0.48 m) >互花米草(3.55 ± 0.73 m) >藨草(3.26 ± 0.57 m)≈海三棱藨草(3.25 ± 0.72 m)(图6 ).自然状态下, 植被分布区潮滩高程与群落演替发育的时间存在密切联系(王卿等, 2012 ), 潮滩原生演替初期, 莎草科植物作为先锋植物首先出现在高程较低的光滩, 各植物种群发育成熟后, 变成相对稳定的莎草科植物群落.成熟的草滩具有明显的促淤沉降功能, 加速潮滩高程演变, 伴随着草滩“内带”泥沙淤积程度不断加深, 潮滩高程不断增高, 由此禾本科逐步占领莎草科生存空间(Wang et al ., 2010 ; 曹浩冰等, 2014 ).总之, 植被的出现加快高程的抬升, 高程的增加又促进植物群落的演替, 但禾本科分布区的潮滩高程始终普遍高于莎草科. ...
... 另一方面, 芦苇与互花米草的交错边界最长, 但是两者在高程以及盐度上都存在非常显著的差异(p < 0.01), 并且区间偏离较大, 因此互花米草不具备向芦苇种群扩张的优势.与海三棱藨草相比, 互花米草能够适应更高的高程和土壤盐度, 海三棱藨草-互花米草植被交错区往大堤方向高程变高, 反而适合互花米草生长; 交错带偏海三棱藨草一侧土壤盐度逐渐升高, 限制了海三棱藨草进一步扩张, 而在交错区内, 互花米草通过根茎、分蘖等繁殖方式, 争夺空间资源, 在植株高度、盖度等方面的优势限制了海三棱藨草生长(Chen et al ., 2004 ; Wang et al ., 2010 ).无论在交错区内部, 还是在与交错带毗邻的缓冲空间, 互花米草在高程和土壤盐度的综合作用下, 生长状况会更加良好, 而海三棱藨草很可能会被互花米草逐步取代. ...
盐沼植物群落研究进展: 分布、演替及影响因子
4
2012
... 长江入海径流以崇明岛为界, 形成南、北两个一级支流, 两侧入海淡水不均衡, 南支淡水充裕, 北支偏少, 加之涨潮时北支经常出现咸水倒灌(贺松林等, 2006 ), 因此, 崇明东滩南部水体盐度明显低于北部.受潮汐影响, 海水与淡水在混合过程中又形成了新的盐度梯度, 即低潮带盐度高于中、高潮滩(陈中义等, 2005a ; 贺松林等, 2006 ; 曹利利等, 2012 ; 王卿等, 2012 ).与此同时, 径流中所携带的大量泥沙在岛堤之外沉积并发育形成新的具有不同潮滩高程的滩地, 受潮汐影响, 滩面水流在涨、落潮过程中不断冲刷滩面, 而崇明东滩南部植被相对稀疏, 因此在南部潮滩上形成多条宽窄不一、错综复杂的潮沟(张修桂, 2005 ; 姜亦飞等, 2012 ; 刘建华等, 2012 ; 陈勇等, 2013 ), 这在一定程度上限制了人类活动对保护区的影响. ...
... 藨草(图2E )和海三棱藨草(图2F )盖度最大值都出现在各自分布区中部, 分别为73%和90%, 而最小盖度值出现在种群边缘, 分别为7%和8%, 海三棱藨草在莎草科中平均盖度最大达46%.作为先锋植物, 莎草科植物在演替初期成片状占据光滩水域, 植被盖度值较低, 为20%-35%, 但群落斑块之间连接度高、渗透性好, 相对于“内带”, 光滩区域人类活动少、影响小、干扰少, 仅受潮汐等自然因素影响, 因此莎草科植物能够均匀分布, 并且斑块之间逐渐连接, 成条带状.在群落相对稳定时, 群落中心植被总盖度值最大可达90%.而演替后期大部分空间被禾本科逐步占领, 即“内带”莎草科植物多以斑块形式随机分布, 盖度维持在24%-41%.整体而言, 莎草科中藨草和海三棱藨草集中在“外带”, 植株均匀分布, “内带”数量稀少, 呈随机分布, 并且两种群中间盖度高, 边缘盖度低; 糙叶薹草无论是“内带”还是“外带”都呈现典型的均匀分布, 但“外带”植被盖度显著高于“内带”, 莎草科植物的空间分布类型与群落发育时间关系密切(王卿等, 2012 ). ...
... 为明确植被与潮滩高程因子之间的关系, 我们对各植物类群分别进行显著性差异分析, 发现除了白茅和芦苇, 藨草和海三棱藨草两两之间无显著性差异外, 其余各植物群落之间差异非常显著(p < 0.01).各物种相应高程分别为白茅(3.85 ± 0.32 m)≈芦苇(3.75 ± 0.57 m) >糙叶薹草(3.64 ± 0.48 m) >互花米草(3.55 ± 0.73 m) >藨草(3.26 ± 0.57 m)≈海三棱藨草(3.25 ± 0.72 m)(图6 ).自然状态下, 植被分布区潮滩高程与群落演替发育的时间存在密切联系(王卿等, 2012 ), 潮滩原生演替初期, 莎草科植物作为先锋植物首先出现在高程较低的光滩, 各植物种群发育成熟后, 变成相对稳定的莎草科植物群落.成熟的草滩具有明显的促淤沉降功能, 加速潮滩高程演变, 伴随着草滩“内带”泥沙淤积程度不断加深, 潮滩高程不断增高, 由此禾本科逐步占领莎草科生存空间(Wang et al ., 2010 ; 曹浩冰等, 2014 ).总之, 植被的出现加快高程的抬升, 高程的增加又促进植物群落的演替, 但禾本科分布区的潮滩高程始终普遍高于莎草科. ...
... 潮滩植物群落在空间上呈现不同的分布状况, 潮滩高程作为影响植物分布的重要环境因子, 与植物的淹水时间和频度, 水体中的溶解氧, 土壤粒径、养分、氧化还原电位、pH值等生境因子存在密切联系(Silvestri et al ., 2005 ; 何彦龙, 2014 ), 与此同时, 植物的密度、高度等因素也会影响高程的变化(Duarte et al ., 2013 ).本研究表明崇明东滩南部植被覆盖空间6种主要植物分布在4个差异显著的高程区间内(p < 0.01), 黄华梅等(2007)和严格等(2014)认为崇明东滩芦苇分布在高潮带(高程>2.9 m), 海三棱藨草分布在中潮带(高程2.0-2.9 m), 互花米草在中、高潮带都有分布.同时对比何彦龙等(2010)在2008年对崇明东滩互花米草(3.21 ± 0.01 m)和海三棱藨草(3.10 ± 0.02 m)的研究结果, 发现上述植物在高程数值上有所差异, 但各植物种群生长空间的高程相对大小保持稳定, 表明在崇明东滩无论是大尺度区域范围, 还是小尺度空间, 各主要植物类群的潮滩高程相对高度保持恒定, 因此潮滩高程是决定植被空间分布的重要因素, 而具体数值差异可能是由于野外调查的时间、地点以及高程仪器精度差异所致.盐度是植物生长过程中的又一重要条件性因子, 不同的植物在不同的生长阶段适应不同范围的盐度(Zhang et al ., 2011 ).陈中义等(2005b)研究了水体中盐度的变化对互花米草和海三棱藨草的影响, 认为互花米草更适应高盐度海水.本研究中互花米草生长空间的土壤盐度确实显著高于海三棱藨草区(p < 0.01), 除了与互花米草本身的高耐盐特性有关, 与植株地上部分的泌盐作用也有一定关系(He et al ., 2011 ).但是无论崇明东滩土壤盐度还是潮滩高程都直接受到潮汐影响, 在空间上都呈现梯度变化(王卿等, 2012 ), 因此, 崇明东滩南部6种主要植物受到两种限定因子的共同制约, 在空间上形成现有的分布格局, 而对潮滩高程和土壤盐度在不同空间尺度下作用的权重仍有待进一步研究. ...
盐沼植物群落研究进展: 分布、演替及影响因子
4
2012
... 长江入海径流以崇明岛为界, 形成南、北两个一级支流, 两侧入海淡水不均衡, 南支淡水充裕, 北支偏少, 加之涨潮时北支经常出现咸水倒灌(贺松林等, 2006 ), 因此, 崇明东滩南部水体盐度明显低于北部.受潮汐影响, 海水与淡水在混合过程中又形成了新的盐度梯度, 即低潮带盐度高于中、高潮滩(陈中义等, 2005a ; 贺松林等, 2006 ; 曹利利等, 2012 ; 王卿等, 2012 ).与此同时, 径流中所携带的大量泥沙在岛堤之外沉积并发育形成新的具有不同潮滩高程的滩地, 受潮汐影响, 滩面水流在涨、落潮过程中不断冲刷滩面, 而崇明东滩南部植被相对稀疏, 因此在南部潮滩上形成多条宽窄不一、错综复杂的潮沟(张修桂, 2005 ; 姜亦飞等, 2012 ; 刘建华等, 2012 ; 陈勇等, 2013 ), 这在一定程度上限制了人类活动对保护区的影响. ...
... 藨草(图2E )和海三棱藨草(图2F )盖度最大值都出现在各自分布区中部, 分别为73%和90%, 而最小盖度值出现在种群边缘, 分别为7%和8%, 海三棱藨草在莎草科中平均盖度最大达46%.作为先锋植物, 莎草科植物在演替初期成片状占据光滩水域, 植被盖度值较低, 为20%-35%, 但群落斑块之间连接度高、渗透性好, 相对于“内带”, 光滩区域人类活动少、影响小、干扰少, 仅受潮汐等自然因素影响, 因此莎草科植物能够均匀分布, 并且斑块之间逐渐连接, 成条带状.在群落相对稳定时, 群落中心植被总盖度值最大可达90%.而演替后期大部分空间被禾本科逐步占领, 即“内带”莎草科植物多以斑块形式随机分布, 盖度维持在24%-41%.整体而言, 莎草科中藨草和海三棱藨草集中在“外带”, 植株均匀分布, “内带”数量稀少, 呈随机分布, 并且两种群中间盖度高, 边缘盖度低; 糙叶薹草无论是“内带”还是“外带”都呈现典型的均匀分布, 但“外带”植被盖度显著高于“内带”, 莎草科植物的空间分布类型与群落发育时间关系密切(王卿等, 2012 ). ...
... 为明确植被与潮滩高程因子之间的关系, 我们对各植物类群分别进行显著性差异分析, 发现除了白茅和芦苇, 藨草和海三棱藨草两两之间无显著性差异外, 其余各植物群落之间差异非常显著(p < 0.01).各物种相应高程分别为白茅(3.85 ± 0.32 m)≈芦苇(3.75 ± 0.57 m) >糙叶薹草(3.64 ± 0.48 m) >互花米草(3.55 ± 0.73 m) >藨草(3.26 ± 0.57 m)≈海三棱藨草(3.25 ± 0.72 m)(图6 ).自然状态下, 植被分布区潮滩高程与群落演替发育的时间存在密切联系(王卿等, 2012 ), 潮滩原生演替初期, 莎草科植物作为先锋植物首先出现在高程较低的光滩, 各植物种群发育成熟后, 变成相对稳定的莎草科植物群落.成熟的草滩具有明显的促淤沉降功能, 加速潮滩高程演变, 伴随着草滩“内带”泥沙淤积程度不断加深, 潮滩高程不断增高, 由此禾本科逐步占领莎草科生存空间(Wang et al ., 2010 ; 曹浩冰等, 2014 ).总之, 植被的出现加快高程的抬升, 高程的增加又促进植物群落的演替, 但禾本科分布区的潮滩高程始终普遍高于莎草科. ...
... 潮滩植物群落在空间上呈现不同的分布状况, 潮滩高程作为影响植物分布的重要环境因子, 与植物的淹水时间和频度, 水体中的溶解氧, 土壤粒径、养分、氧化还原电位、pH值等生境因子存在密切联系(Silvestri et al ., 2005 ; 何彦龙, 2014 ), 与此同时, 植物的密度、高度等因素也会影响高程的变化(Duarte et al ., 2013 ).本研究表明崇明东滩南部植被覆盖空间6种主要植物分布在4个差异显著的高程区间内(p < 0.01), 黄华梅等(2007)和严格等(2014)认为崇明东滩芦苇分布在高潮带(高程>2.9 m), 海三棱藨草分布在中潮带(高程2.0-2.9 m), 互花米草在中、高潮带都有分布.同时对比何彦龙等(2010)在2008年对崇明东滩互花米草(3.21 ± 0.01 m)和海三棱藨草(3.10 ± 0.02 m)的研究结果, 发现上述植物在高程数值上有所差异, 但各植物种群生长空间的高程相对大小保持稳定, 表明在崇明东滩无论是大尺度区域范围, 还是小尺度空间, 各主要植物类群的潮滩高程相对高度保持恒定, 因此潮滩高程是决定植被空间分布的重要因素, 而具体数值差异可能是由于野外调查的时间、地点以及高程仪器精度差异所致.盐度是植物生长过程中的又一重要条件性因子, 不同的植物在不同的生长阶段适应不同范围的盐度(Zhang et al ., 2011 ).陈中义等(2005b)研究了水体中盐度的变化对互花米草和海三棱藨草的影响, 认为互花米草更适应高盐度海水.本研究中互花米草生长空间的土壤盐度确实显著高于海三棱藨草区(p < 0.01), 除了与互花米草本身的高耐盐特性有关, 与植株地上部分的泌盐作用也有一定关系(He et al ., 2011 ).但是无论崇明东滩土壤盐度还是潮滩高程都直接受到潮汐影响, 在空间上都呈现梯度变化(王卿等, 2012 ), 因此, 崇明东滩南部6种主要植物受到两种限定因子的共同制约, 在空间上形成现有的分布格局, 而对潮滩高程和土壤盐度在不同空间尺度下作用的权重仍有待进一步研究. ...
1
2005
... 崇明东滩是长江口典型淤涨型潮滩湿地, 位于崇明岛最东端(121.83°-122.08° E, 31.42°-31.63° N), 属北亚热带季风气候, 四季分明, 日照充分, 雨量充沛, 气候温暖湿润, 年平均气温15.3 ℃, 年降水量1022 mm.这里是东亚-澳大利西亚迁徙路线上鸟类重要的停歇地, 也是小天鹅、白头鹤和黑脸琵鹭等珍稀濒危水禽重要的越冬栖息地(黄正一等, 1993 ).1997年崇明东滩被正式列入东亚-澳大利西亚涉禽保护区网络, 2002年又被《湿地公约》组织列入国际重要湿地名录(徐宏发和赵云龙, 2005 ), 2005年晋升为国家级自然保护区.本文研究区位于崇明东滩鸟类国家级自然保护区南部, 团结沙和东旺沙之间(121.90°-121.99° E, 31.45°-31.52° N), 北以互花米草生态治理工程2014年新筑大堤为界, 西至98大堤, 东、南两侧到达长江口外水域, 位于保护区的核心部分, 主要由团结沙外滩、大潮沟以及东旺沙外滩等植被覆盖的地区组成, 总面积约1800 hm2 (图1 ). ...
1
2005
... 崇明东滩是长江口典型淤涨型潮滩湿地, 位于崇明岛最东端(121.83°-122.08° E, 31.42°-31.63° N), 属北亚热带季风气候, 四季分明, 日照充分, 雨量充沛, 气候温暖湿润, 年平均气温15.3 ℃, 年降水量1022 mm.这里是东亚-澳大利西亚迁徙路线上鸟类重要的停歇地, 也是小天鹅、白头鹤和黑脸琵鹭等珍稀濒危水禽重要的越冬栖息地(黄正一等, 1993 ).1997年崇明东滩被正式列入东亚-澳大利西亚涉禽保护区网络, 2002年又被《湿地公约》组织列入国际重要湿地名录(徐宏发和赵云龙, 2005 ), 2005年晋升为国家级自然保护区.本文研究区位于崇明东滩鸟类国家级自然保护区南部, 团结沙和东旺沙之间(121.90°-121.99° E, 31.45°-31.52° N), 北以互花米草生态治理工程2014年新筑大堤为界, 西至98大堤, 东、南两侧到达长江口外水域, 位于保护区的核心部分, 主要由团结沙外滩、大潮沟以及东旺沙外滩等植被覆盖的地区组成, 总面积约1800 hm2 (图1 ). ...
苏北海滨湿地互花米草种群繁殖方式
1
2014
... 研究区东北部的互花米草也因较高的盖度值成为优势种, 同时, 无性繁殖方式, 使得种群扩散速度快、发育周期短, 并能够以母体为中心快速向周围空间扩散, 加之互花米草萌发时间早, 植物生长迅速, 抑制相邻其他植物的正常生长, 在其分布区内占有绝对优势(Li et al ., 2009 ; Lu & Zhang, 2013 ; Liu et al ., 2014 ; 徐伟伟等, 2014 ).研究区内互花米草与糙叶薹草、海三棱藨草、芦苇的邻接带分别长达1.2 km、2.0 km、3.0 km (图3 ), 这为互花米草向周围快速扩散提供了可能.与芦苇和互花米草不同, 海三棱藨草种群破碎化明显, 虽然外侧与光滩的边界长达2.4 km, 为种群向光滩扩张创造了良好的演替条件, 但其群落中部被芦苇和互花米草等植物所隔离, 外部受潮汐以及波浪作用明显, 因此生长条件比其他物种更为恶劣, 种群优势因此降低.其余3种植物空间分布形式各不相同, 白茅空间分布不均, 种群连接度不高, 斑块面积小, 但是数量较多, 因此种群总面积能占一定优势.糙叶薹草种群连接度高, 种群斑块面积大, 分布广, 能够承受较高的环境风险.藨草虽然在“外带”附近种群分布连续, 且面积较大, 但夹在糙叶薹草内部的藨草种群面积小, 较为分散.所有植物优势种群在东北至西南方向上都呈明显的条带状分布. ...
苏北海滨湿地互花米草种群繁殖方式
1
2014
... 研究区东北部的互花米草也因较高的盖度值成为优势种, 同时, 无性繁殖方式, 使得种群扩散速度快、发育周期短, 并能够以母体为中心快速向周围空间扩散, 加之互花米草萌发时间早, 植物生长迅速, 抑制相邻其他植物的正常生长, 在其分布区内占有绝对优势(Li et al ., 2009 ; Lu & Zhang, 2013 ; Liu et al ., 2014 ; 徐伟伟等, 2014 ).研究区内互花米草与糙叶薹草、海三棱藨草、芦苇的邻接带分别长达1.2 km、2.0 km、3.0 km (图3 ), 这为互花米草向周围快速扩散提供了可能.与芦苇和互花米草不同, 海三棱藨草种群破碎化明显, 虽然外侧与光滩的边界长达2.4 km, 为种群向光滩扩张创造了良好的演替条件, 但其群落中部被芦苇和互花米草等植物所隔离, 外部受潮汐以及波浪作用明显, 因此生长条件比其他物种更为恶劣, 种群优势因此降低.其余3种植物空间分布形式各不相同, 白茅空间分布不均, 种群连接度不高, 斑块面积小, 但是数量较多, 因此种群总面积能占一定优势.糙叶薹草种群连接度高, 种群斑块面积大, 分布广, 能够承受较高的环境风险.藨草虽然在“外带”附近种群分布连续, 且面积较大, 但夹在糙叶薹草内部的藨草种群面积小, 较为分散.所有植物优势种群在东北至西南方向上都呈明显的条带状分布. ...
崇明东滩湿地不同盐沼植物群落土壤碳储量分布
1
2014
... 崇明东滩属于典型的淤涨型潮滩, 目前滩涂植被覆盖区总面积4265 hm2 , 2012年调查数据显示, 崇明东滩中部、北部植物以芦苇和互花米草为主, 而莎草科植物主要集中分布在崇明东滩南部1) ( 1) 吴纪华 (2014). 上海崇明东滩鸟类国家级自然保护区2012年度资源监测公报, 14-15.) .在崇明东滩的相关研究中, 众多学者根据研究需要将崇明东滩的海三棱藨草、藨草、糙叶薹草3种植物统一归入海三棱藨草类群(黄华梅等, 2007 ; Yuan & Zhang, 2008 ; 严格等, 2014 ).部分研究虽然将海三棱藨草、藨草、糙叶薹草统一划分为莎草科植物群落或者藨草群落(曹浩冰等, 2014 ; 任璘婧等, 2014 ), 但没有对这3种植物进行细致的划分, 各种群面积大小及其具体分布目前尚无相关报道.上述3种植物虽然同属于莎草科, 但占据不同的生态位, 承担着不同的生态功能, 例如在崇明东滩三者虽然都可以作为湿地鸟类的饵料, 但不同的鸟类会取食不同植物的不同部分(Ma et al ., 2003 ; Zhu et al ., 2007 ), 且3种植物组织、器官发育和成熟时间也各不相同.本文基于崇明东滩南部各植物种群的细致调查, 明确了各主要植物种群的空间分布以及各自的面积, 为进一步研究植被分布格局变化以及重要鸟类觅食生境提供基础. ...
崇明东滩湿地不同盐沼植物群落土壤碳储量分布
1
2014
... 崇明东滩属于典型的淤涨型潮滩, 目前滩涂植被覆盖区总面积4265 hm2 , 2012年调查数据显示, 崇明东滩中部、北部植物以芦苇和互花米草为主, 而莎草科植物主要集中分布在崇明东滩南部1) ( 1) 吴纪华 (2014). 上海崇明东滩鸟类国家级自然保护区2012年度资源监测公报, 14-15.) .在崇明东滩的相关研究中, 众多学者根据研究需要将崇明东滩的海三棱藨草、藨草、糙叶薹草3种植物统一归入海三棱藨草类群(黄华梅等, 2007 ; Yuan & Zhang, 2008 ; 严格等, 2014 ).部分研究虽然将海三棱藨草、藨草、糙叶薹草统一划分为莎草科植物群落或者藨草群落(曹浩冰等, 2014 ; 任璘婧等, 2014 ), 但没有对这3种植物进行细致的划分, 各种群面积大小及其具体分布目前尚无相关报道.上述3种植物虽然同属于莎草科, 但占据不同的生态位, 承担着不同的生态功能, 例如在崇明东滩三者虽然都可以作为湿地鸟类的饵料, 但不同的鸟类会取食不同植物的不同部分(Ma et al ., 2003 ; Zhu et al ., 2007 ), 且3种植物组织、器官发育和成熟时间也各不相同.本文基于崇明东滩南部各植物种群的细致调查, 明确了各主要植物种群的空间分布以及各自的面积, 为进一步研究植被分布格局变化以及重要鸟类觅食生境提供基础. ...
崇明东滩湿地高等植被演替特征
1
2007
... 近十几年来, 大量研究人员通过遥感影像解译, 对崇明东滩主要植被进行了分区(Gao & Zhang, 2006 ; Tian et al ., 2008 ; 阿也提古丽·斯迪克等, 2010 ; 翟新源和杨苏新, 2013 ; 郑宗生等, 2010 ; 宗玮等, 2011 ), 普遍认为芦苇(Phragmites australis )、互花米草(Spartina alterniflora )及海三棱藨草(Scirpus mariqueter )是崇明东滩的“三大”主要植物类群.然而, 崇明东滩南部长期受长江入海淡水影响, 水体盐度相对较低, 在团结沙至大潮沟(图1 )存在较大面积的藨草(Scirpus triqueter )种群区(闫芊等, 2007 ; 章振亚等, 2012 ).同时发现, 糙叶薹草(Carex scabrifolia )在研究区内频繁出现, 白茅(Imperata cylindrica )在部分空间也已发展成优势种. ...
崇明东滩湿地高等植被演替特征
1
2007
... 近十几年来, 大量研究人员通过遥感影像解译, 对崇明东滩主要植被进行了分区(Gao & Zhang, 2006 ; Tian et al ., 2008 ; 阿也提古丽·斯迪克等, 2010 ; 翟新源和杨苏新, 2013 ; 郑宗生等, 2010 ; 宗玮等, 2011 ), 普遍认为芦苇(Phragmites australis )、互花米草(Spartina alterniflora )及海三棱藨草(Scirpus mariqueter )是崇明东滩的“三大”主要植物类群.然而, 崇明东滩南部长期受长江入海淡水影响, 水体盐度相对较低, 在团结沙至大潮沟(图1 )存在较大面积的藨草(Scirpus triqueter )种群区(闫芊等, 2007 ; 章振亚等, 2012 ).同时发现, 糙叶薹草(Carex scabrifolia )在研究区内频繁出现, 白茅(Imperata cylindrica )在部分空间也已发展成优势种. ...
崇明东滩的雁鸭与海三棱藨草
1
1994
... 上述植物中, 海三棱藨草是中国特有的盐沼植物, 也是典型的潮滩先锋物种, 目前主要分布于长江口沿岸的滩涂湿地(Sun et al ., 2002 , 2003 ; 梅雪英和张修峰, 2007 ; 陈秀芝和孙瑛, 2011 ).崇明东滩作为长江口最大的天然湿地, 有大量的海三棱藨草分布(虞快, 1994 , 1996 ), 但近年来崇明东滩北部互花米草生长泛滥, 海三棱藨草原有生态位受到挤压, 已基本被互花米草取代(陈秀芝和孙瑛, 2011 ), 东旺沙中间(2014年新围垦大堤东南部外围)是海三棱藨草现存种群主要分布区, 同时也是莎草科类群的主要分布区1) (1) 吴纪华 (2014). 上海崇明东滩鸟类国家级自然保护区2012年度资源监测公报, 14-15.) .海三棱藨草在崇明东滩形成湿地草场, 能为鸻鹬类、雁鸭类、鹤类、鸥类等众多越冬候鸟提供天然的觅食、活动、休憩场所, 同时海三棱藨草的根茎和球茎, 还是国家一级保护鸟类——白头鹤的重要饵料(Ma et al ., 2003 ; Zhu et al ., 2007 ), 因此, 海三棱藨草对崇明东滩所有滨海湿地鸟类都显得至关重要.然而近年来滩涂围垦、堤外放牧及外来物种竞争等导致植物种群分布状况发生变化, 植物群落斑块破碎化严重, 整个生态系统服务功能也随之降低(张利权和雍学葵, 1992; Chen et al ., 2004; 陈中义等, 2005b; Wang et al ., 2010).因此, 确定包括海三棱藨草在内的关键植物种群在东滩的空间分布, 明确其随环境因子的变化规律, 对该湿地生态系统的修复和保护具有重要的指导意义. ...
崇明东滩的雁鸭与海三棱藨草
1
1994
... 上述植物中, 海三棱藨草是中国特有的盐沼植物, 也是典型的潮滩先锋物种, 目前主要分布于长江口沿岸的滩涂湿地(Sun et al ., 2002 , 2003 ; 梅雪英和张修峰, 2007 ; 陈秀芝和孙瑛, 2011 ).崇明东滩作为长江口最大的天然湿地, 有大量的海三棱藨草分布(虞快, 1994 , 1996 ), 但近年来崇明东滩北部互花米草生长泛滥, 海三棱藨草原有生态位受到挤压, 已基本被互花米草取代(陈秀芝和孙瑛, 2011 ), 东旺沙中间(2014年新围垦大堤东南部外围)是海三棱藨草现存种群主要分布区, 同时也是莎草科类群的主要分布区1) (1) 吴纪华 (2014). 上海崇明东滩鸟类国家级自然保护区2012年度资源监测公报, 14-15.) .海三棱藨草在崇明东滩形成湿地草场, 能为鸻鹬类、雁鸭类、鹤类、鸥类等众多越冬候鸟提供天然的觅食、活动、休憩场所, 同时海三棱藨草的根茎和球茎, 还是国家一级保护鸟类——白头鹤的重要饵料(Ma et al ., 2003 ; Zhu et al ., 2007 ), 因此, 海三棱藨草对崇明东滩所有滨海湿地鸟类都显得至关重要.然而近年来滩涂围垦、堤外放牧及外来物种竞争等导致植物种群分布状况发生变化, 植物群落斑块破碎化严重, 整个生态系统服务功能也随之降低(张利权和雍学葵, 1992; Chen et al ., 2004; 陈中义等, 2005b; Wang et al ., 2010).因此, 确定包括海三棱藨草在内的关键植物种群在东滩的空间分布, 明确其随环境因子的变化规律, 对该湿地生态系统的修复和保护具有重要的指导意义. ...
亟待保护的海三棱藨草
1
1996
... 上述植物中, 海三棱藨草是中国特有的盐沼植物, 也是典型的潮滩先锋物种, 目前主要分布于长江口沿岸的滩涂湿地(Sun et al ., 2002 , 2003 ; 梅雪英和张修峰, 2007 ; 陈秀芝和孙瑛, 2011 ).崇明东滩作为长江口最大的天然湿地, 有大量的海三棱藨草分布(虞快, 1994 , 1996 ), 但近年来崇明东滩北部互花米草生长泛滥, 海三棱藨草原有生态位受到挤压, 已基本被互花米草取代(陈秀芝和孙瑛, 2011 ), 东旺沙中间(2014年新围垦大堤东南部外围)是海三棱藨草现存种群主要分布区, 同时也是莎草科类群的主要分布区1) (1) 吴纪华 (2014). 上海崇明东滩鸟类国家级自然保护区2012年度资源监测公报, 14-15.) .海三棱藨草在崇明东滩形成湿地草场, 能为鸻鹬类、雁鸭类、鹤类、鸥类等众多越冬候鸟提供天然的觅食、活动、休憩场所, 同时海三棱藨草的根茎和球茎, 还是国家一级保护鸟类——白头鹤的重要饵料(Ma et al ., 2003 ; Zhu et al ., 2007 ), 因此, 海三棱藨草对崇明东滩所有滨海湿地鸟类都显得至关重要.然而近年来滩涂围垦、堤外放牧及外来物种竞争等导致植物种群分布状况发生变化, 植物群落斑块破碎化严重, 整个生态系统服务功能也随之降低(张利权和雍学葵, 1992; Chen et al ., 2004; 陈中义等, 2005b; Wang et al ., 2010).因此, 确定包括海三棱藨草在内的关键植物种群在东滩的空间分布, 明确其随环境因子的变化规律, 对该湿地生态系统的修复和保护具有重要的指导意义. ...
亟待保护的海三棱藨草
1
1996
... 上述植物中, 海三棱藨草是中国特有的盐沼植物, 也是典型的潮滩先锋物种, 目前主要分布于长江口沿岸的滩涂湿地(Sun et al ., 2002 , 2003 ; 梅雪英和张修峰, 2007 ; 陈秀芝和孙瑛, 2011 ).崇明东滩作为长江口最大的天然湿地, 有大量的海三棱藨草分布(虞快, 1994 , 1996 ), 但近年来崇明东滩北部互花米草生长泛滥, 海三棱藨草原有生态位受到挤压, 已基本被互花米草取代(陈秀芝和孙瑛, 2011 ), 东旺沙中间(2014年新围垦大堤东南部外围)是海三棱藨草现存种群主要分布区, 同时也是莎草科类群的主要分布区1) (1) 吴纪华 (2014). 上海崇明东滩鸟类国家级自然保护区2012年度资源监测公报, 14-15.) .海三棱藨草在崇明东滩形成湿地草场, 能为鸻鹬类、雁鸭类、鹤类、鸥类等众多越冬候鸟提供天然的觅食、活动、休憩场所, 同时海三棱藨草的根茎和球茎, 还是国家一级保护鸟类——白头鹤的重要饵料(Ma et al ., 2003 ; Zhu et al ., 2007 ), 因此, 海三棱藨草对崇明东滩所有滨海湿地鸟类都显得至关重要.然而近年来滩涂围垦、堤外放牧及外来物种竞争等导致植物种群分布状况发生变化, 植物群落斑块破碎化严重, 整个生态系统服务功能也随之降低(张利权和雍学葵, 1992; Chen et al ., 2004; 陈中义等, 2005b; Wang et al ., 2010).因此, 确定包括海三棱藨草在内的关键植物种群在东滩的空间分布, 明确其随环境因子的变化规律, 对该湿地生态系统的修复和保护具有重要的指导意义. ...
Mapping large-scale distribution of submerged aquatic vegetation coverage using remote sensing.
2
2008
... 值得关注的是, 海三棱藨草(图2F )作为中国特有盐沼植物, 其种群面积与前人研究结果(黄华梅等, 2007 ; Yuan & Zhang, 2008 )存在较大差异.此外, 作为特有先锋物种, 海三棱藨草分布在“外带”, 但优势带也与互花米草存在较大的重叠空间, 两者存在显著的竞争关系, 然而互花米草在高度、盖度、地上生物量、地下生物量方面都比海三棱藨草具有明显优势, 为其获取更多光照、营养盐, 快速取代海三棱藨草创造了条件(Chen et al ., 2004 ; 陈中义等, 2005b , He et al ., 2011 ), 因此, 两种植物的分布存在交错带对海三棱藨草种群的发展极其不利. ...
... 崇明东滩属于典型的淤涨型潮滩, 目前滩涂植被覆盖区总面积4265 hm2 , 2012年调查数据显示, 崇明东滩中部、北部植物以芦苇和互花米草为主, 而莎草科植物主要集中分布在崇明东滩南部1) ( 1) 吴纪华 (2014). 上海崇明东滩鸟类国家级自然保护区2012年度资源监测公报, 14-15.) .在崇明东滩的相关研究中, 众多学者根据研究需要将崇明东滩的海三棱藨草、藨草、糙叶薹草3种植物统一归入海三棱藨草类群(黄华梅等, 2007 ; Yuan & Zhang, 2008 ; 严格等, 2014 ).部分研究虽然将海三棱藨草、藨草、糙叶薹草统一划分为莎草科植物群落或者藨草群落(曹浩冰等, 2014 ; 任璘婧等, 2014 ), 但没有对这3种植物进行细致的划分, 各种群面积大小及其具体分布目前尚无相关报道.上述3种植物虽然同属于莎草科, 但占据不同的生态位, 承担着不同的生态功能, 例如在崇明东滩三者虽然都可以作为湿地鸟类的饵料, 但不同的鸟类会取食不同植物的不同部分(Ma et al ., 2003 ; Zhu et al ., 2007 ), 且3种植物组织、器官发育和成熟时间也各不相同.本文基于崇明东滩南部各植物种群的细致调查, 明确了各主要植物种群的空间分布以及各自的面积, 为进一步研究植被分布格局变化以及重要鸟类觅食生境提供基础. ...
崇明东滩湿地植被遥感监测研究
1
2013
... 近十几年来, 大量研究人员通过遥感影像解译, 对崇明东滩主要植被进行了分区(Gao & Zhang, 2006 ; Tian et al ., 2008 ; 阿也提古丽·斯迪克等, 2010 ; 翟新源和杨苏新, 2013 ; 郑宗生等, 2010 ; 宗玮等, 2011 ), 普遍认为芦苇(Phragmites australis )、互花米草(Spartina alterniflora )及海三棱藨草(Scirpus mariqueter )是崇明东滩的“三大”主要植物类群.然而, 崇明东滩南部长期受长江入海淡水影响, 水体盐度相对较低, 在团结沙至大潮沟(图1 )存在较大面积的藨草(Scirpus triqueter )种群区(闫芊等, 2007 ; 章振亚等, 2012 ).同时发现, 糙叶薹草(Carex scabrifolia )在研究区内频繁出现, 白茅(Imperata cylindrica )在部分空间也已发展成优势种. ...
崇明东滩湿地植被遥感监测研究
1
2013
... 近十几年来, 大量研究人员通过遥感影像解译, 对崇明东滩主要植被进行了分区(Gao & Zhang, 2006 ; Tian et al ., 2008 ; 阿也提古丽·斯迪克等, 2010 ; 翟新源和杨苏新, 2013 ; 郑宗生等, 2010 ; 宗玮等, 2011 ), 普遍认为芦苇(Phragmites australis )、互花米草(Spartina alterniflora )及海三棱藨草(Scirpus mariqueter )是崇明东滩的“三大”主要植物类群.然而, 崇明东滩南部长期受长江入海淡水影响, 水体盐度相对较低, 在团结沙至大潮沟(图1 )存在较大面积的藨草(Scirpus triqueter )种群区(闫芊等, 2007 ; 章振亚等, 2012 ).同时发现, 糙叶薹草(Carex scabrifolia )在研究区内频繁出现, 白茅(Imperata cylindrica )在部分空间也已发展成优势种. ...
海三棱藨草盐沼土壤环境因子的时空分布特征
2
2011
... 在前人相关研究(Ma et al ., 2003 ; 陈中义等, 2005a ; 贺强等, 2009 ; 詹跃武等, 2011 )的基础上, 本研究选择潮滩高程和表层30 cm土壤盐度作为主要参数, 探讨盐沼植物在东滩南部的空间分布及其与两大环境因子的相互关系. ...
... 研究区内, 潮滩高程介于-1.22-7.15 m之间(图5A ), 总体呈西高东低, 东旺沙附近地势较高, 在3.50-7.15 m之间, 在东旺沙中心部分出现高程最大值, 大潮沟南部至团结沙区, 介于2.30-5.10 m之间.长江口咸、淡水交互作用, 在崇明东滩外部区域形成了天然盐度梯度变化(陈中义等, 2005a ; 詹跃武等, 2011 ), 研究区内土壤盐度范围介于1.60-4.42 g·kg-1 , 并且空间上具有明显的变化规律——东北高、西南低, 因此土壤盐度最大值出现在东北部, 达到4.42 g·kg-1 . ...
海三棱藨草盐沼土壤环境因子的时空分布特征
2
2011
... 在前人相关研究(Ma et al ., 2003 ; 陈中义等, 2005a ; 贺强等, 2009 ; 詹跃武等, 2011 )的基础上, 本研究选择潮滩高程和表层30 cm土壤盐度作为主要参数, 探讨盐沼植物在东滩南部的空间分布及其与两大环境因子的相互关系. ...
... 研究区内, 潮滩高程介于-1.22-7.15 m之间(图5A ), 总体呈西高东低, 东旺沙附近地势较高, 在3.50-7.15 m之间, 在东旺沙中心部分出现高程最大值, 大潮沟南部至团结沙区, 介于2.30-5.10 m之间.长江口咸、淡水交互作用, 在崇明东滩外部区域形成了天然盐度梯度变化(陈中义等, 2005a ; 詹跃武等, 2011 ), 研究区内土壤盐度范围介于1.60-4.42 g·kg-1 , 并且空间上具有明显的变化规律——东北高、西南低, 因此土壤盐度最大值出现在东北部, 达到4.42 g·kg-1 . ...
Using hyperspectral vegetation indices as a proxy to monitor soil salinity.
2
2011
... 潮滩高程固然是划分植被分布范围的重要参考依据, 但植被空间分布同时受到其他环境因子的影响, 盐度就是其中一个重要的限定因子.植物对盐度的变化非常敏感, 盐沼植物对于盐度虽已形成一系列的响应和适应机制, 但盐度依然是盐沼植物生长过程中重要的影响因素(陈中义等, 2005a ; Zhang et al ., 2011 ).因为潮汐的不连续性, 加之长江口水体盐度较土壤盐度波动频繁, 因此选择土壤盐度为研究对象.从图6 可知, 互花米草分布区的土壤盐度最高, 为(3.21 ± 0.51 g·kg-1 ), 白茅区盐度值最低, 为(1.72 ± 0.15 g·kg-1 ), 另外4种主要植物的土壤盐度分别为: 海三棱藨草(2.86 ± 0.64 g·kg-1 ) >芦苇(2.12 ± 0.57 g·kg-1 )≈藨草(2.10 ± 0.37 g·kg-1 ) >糙叶薹草(1.90 ± 0.26 g·kg-1 ), 除了芦苇和藨草, 其余各类群植物土壤盐度差异性非常显著(p < 0.01), 因此土壤盐度在空间的变化必然影响甚至决定了各植物类群间的分布状况. ...
... 潮滩植物群落在空间上呈现不同的分布状况, 潮滩高程作为影响植物分布的重要环境因子, 与植物的淹水时间和频度, 水体中的溶解氧, 土壤粒径、养分、氧化还原电位、pH值等生境因子存在密切联系(Silvestri et al ., 2005 ; 何彦龙, 2014 ), 与此同时, 植物的密度、高度等因素也会影响高程的变化(Duarte et al ., 2013 ).本研究表明崇明东滩南部植被覆盖空间6种主要植物分布在4个差异显著的高程区间内(p < 0.01), 黄华梅等(2007)和严格等(2014)认为崇明东滩芦苇分布在高潮带(高程>2.9 m), 海三棱藨草分布在中潮带(高程2.0-2.9 m), 互花米草在中、高潮带都有分布.同时对比何彦龙等(2010)在2008年对崇明东滩互花米草(3.21 ± 0.01 m)和海三棱藨草(3.10 ± 0.02 m)的研究结果, 发现上述植物在高程数值上有所差异, 但各植物种群生长空间的高程相对大小保持稳定, 表明在崇明东滩无论是大尺度区域范围, 还是小尺度空间, 各主要植物类群的潮滩高程相对高度保持恒定, 因此潮滩高程是决定植被空间分布的重要因素, 而具体数值差异可能是由于野外调查的时间、地点以及高程仪器精度差异所致.盐度是植物生长过程中的又一重要条件性因子, 不同的植物在不同的生长阶段适应不同范围的盐度(Zhang et al ., 2011 ).陈中义等(2005b)研究了水体中盐度的变化对互花米草和海三棱藨草的影响, 认为互花米草更适应高盐度海水.本研究中互花米草生长空间的土壤盐度确实显著高于海三棱藨草区(p < 0.01), 除了与互花米草本身的高耐盐特性有关, 与植株地上部分的泌盐作用也有一定关系(He et al ., 2011 ).但是无论崇明东滩土壤盐度还是潮滩高程都直接受到潮汐影响, 在空间上都呈现梯度变化(王卿等, 2012 ), 因此, 崇明东滩南部6种主要植物受到两种限定因子的共同制约, 在空间上形成现有的分布格局, 而对潮滩高程和土壤盐度在不同空间尺度下作用的权重仍有待进一步研究. ...
崇明岛形成的历史过程
1
2005
... 长江入海径流以崇明岛为界, 形成南、北两个一级支流, 两侧入海淡水不均衡, 南支淡水充裕, 北支偏少, 加之涨潮时北支经常出现咸水倒灌(贺松林等, 2006 ), 因此, 崇明东滩南部水体盐度明显低于北部.受潮汐影响, 海水与淡水在混合过程中又形成了新的盐度梯度, 即低潮带盐度高于中、高潮滩(陈中义等, 2005a ; 贺松林等, 2006 ; 曹利利等, 2012 ; 王卿等, 2012 ).与此同时, 径流中所携带的大量泥沙在岛堤之外沉积并发育形成新的具有不同潮滩高程的滩地, 受潮汐影响, 滩面水流在涨、落潮过程中不断冲刷滩面, 而崇明东滩南部植被相对稀疏, 因此在南部潮滩上形成多条宽窄不一、错综复杂的潮沟(张修桂, 2005 ; 姜亦飞等, 2012 ; 刘建华等, 2012 ; 陈勇等, 2013 ), 这在一定程度上限制了人类活动对保护区的影响. ...
崇明岛形成的历史过程
1
2005
... 长江入海径流以崇明岛为界, 形成南、北两个一级支流, 两侧入海淡水不均衡, 南支淡水充裕, 北支偏少, 加之涨潮时北支经常出现咸水倒灌(贺松林等, 2006 ), 因此, 崇明东滩南部水体盐度明显低于北部.受潮汐影响, 海水与淡水在混合过程中又形成了新的盐度梯度, 即低潮带盐度高于中、高潮滩(陈中义等, 2005a ; 贺松林等, 2006 ; 曹利利等, 2012 ; 王卿等, 2012 ).与此同时, 径流中所携带的大量泥沙在岛堤之外沉积并发育形成新的具有不同潮滩高程的滩地, 受潮汐影响, 滩面水流在涨、落潮过程中不断冲刷滩面, 而崇明东滩南部植被相对稀疏, 因此在南部潮滩上形成多条宽窄不一、错综复杂的潮沟(张修桂, 2005 ; 姜亦飞等, 2012 ; 刘建华等, 2012 ; 陈勇等, 2013 ), 这在一定程度上限制了人类活动对保护区的影响. ...
崇明东滩湿地不同潮汐带入侵植物互花米草根际细菌的多样性
1
2012
... 近十几年来, 大量研究人员通过遥感影像解译, 对崇明东滩主要植被进行了分区(Gao & Zhang, 2006 ; Tian et al ., 2008 ; 阿也提古丽·斯迪克等, 2010 ; 翟新源和杨苏新, 2013 ; 郑宗生等, 2010 ; 宗玮等, 2011 ), 普遍认为芦苇(Phragmites australis )、互花米草(Spartina alterniflora )及海三棱藨草(Scirpus mariqueter )是崇明东滩的“三大”主要植物类群.然而, 崇明东滩南部长期受长江入海淡水影响, 水体盐度相对较低, 在团结沙至大潮沟(图1 )存在较大面积的藨草(Scirpus triqueter )种群区(闫芊等, 2007 ; 章振亚等, 2012 ).同时发现, 糙叶薹草(Carex scabrifolia )在研究区内频繁出现, 白茅(Imperata cylindrica )在部分空间也已发展成优势种. ...
崇明东滩湿地不同潮汐带入侵植物互花米草根际细菌的多样性
1
2012
... 近十几年来, 大量研究人员通过遥感影像解译, 对崇明东滩主要植被进行了分区(Gao & Zhang, 2006 ; Tian et al ., 2008 ; 阿也提古丽·斯迪克等, 2010 ; 翟新源和杨苏新, 2013 ; 郑宗生等, 2010 ; 宗玮等, 2011 ), 普遍认为芦苇(Phragmites australis )、互花米草(Spartina alterniflora )及海三棱藨草(Scirpus mariqueter )是崇明东滩的“三大”主要植物类群.然而, 崇明东滩南部长期受长江入海淡水影响, 水体盐度相对较低, 在团结沙至大潮沟(图1 )存在较大面积的藨草(Scirpus triqueter )种群区(闫芊等, 2007 ; 章振亚等, 2012 ).同时发现, 糙叶薹草(Carex scabrifolia )在研究区内频繁出现, 白茅(Imperata cylindrica )在部分空间也已发展成优势种. ...
基于遥感及数值模拟的崇明东滩冲淤与植被关系探讨
1
2010
... 近十几年来, 大量研究人员通过遥感影像解译, 对崇明东滩主要植被进行了分区(Gao & Zhang, 2006 ; Tian et al ., 2008 ; 阿也提古丽·斯迪克等, 2010 ; 翟新源和杨苏新, 2013 ; 郑宗生等, 2010 ; 宗玮等, 2011 ), 普遍认为芦苇(Phragmites australis )、互花米草(Spartina alterniflora )及海三棱藨草(Scirpus mariqueter )是崇明东滩的“三大”主要植物类群.然而, 崇明东滩南部长期受长江入海淡水影响, 水体盐度相对较低, 在团结沙至大潮沟(图1 )存在较大面积的藨草(Scirpus triqueter )种群区(闫芊等, 2007 ; 章振亚等, 2012 ).同时发现, 糙叶薹草(Carex scabrifolia )在研究区内频繁出现, 白茅(Imperata cylindrica )在部分空间也已发展成优势种. ...
基于遥感及数值模拟的崇明东滩冲淤与植被关系探讨
1
2010
... 近十几年来, 大量研究人员通过遥感影像解译, 对崇明东滩主要植被进行了分区(Gao & Zhang, 2006 ; Tian et al ., 2008 ; 阿也提古丽·斯迪克等, 2010 ; 翟新源和杨苏新, 2013 ; 郑宗生等, 2010 ; 宗玮等, 2011 ), 普遍认为芦苇(Phragmites australis )、互花米草(Spartina alterniflora )及海三棱藨草(Scirpus mariqueter )是崇明东滩的“三大”主要植物类群.然而, 崇明东滩南部长期受长江入海淡水影响, 水体盐度相对较低, 在团结沙至大潮沟(图1 )存在较大面积的藨草(Scirpus triqueter )种群区(闫芊等, 2007 ; 章振亚等, 2012 ).同时发现, 糙叶薹草(Carex scabrifolia )在研究区内频繁出现, 白茅(Imperata cylindrica )在部分空间也已发展成优势种. ...
Food supply in intertidal area for shorebirds during stopover at Chongming Dongtan, China
2
2007
... 上述植物中, 海三棱藨草是中国特有的盐沼植物, 也是典型的潮滩先锋物种, 目前主要分布于长江口沿岸的滩涂湿地(Sun et al ., 2002 , 2003 ; 梅雪英和张修峰, 2007 ; 陈秀芝和孙瑛, 2011 ).崇明东滩作为长江口最大的天然湿地, 有大量的海三棱藨草分布(虞快, 1994 , 1996 ), 但近年来崇明东滩北部互花米草生长泛滥, 海三棱藨草原有生态位受到挤压, 已基本被互花米草取代(陈秀芝和孙瑛, 2011 ), 东旺沙中间(2014年新围垦大堤东南部外围)是海三棱藨草现存种群主要分布区, 同时也是莎草科类群的主要分布区1) (1) 吴纪华 (2014). 上海崇明东滩鸟类国家级自然保护区2012年度资源监测公报, 14-15.) .海三棱藨草在崇明东滩形成湿地草场, 能为鸻鹬类、雁鸭类、鹤类、鸥类等众多越冬候鸟提供天然的觅食、活动、休憩场所, 同时海三棱藨草的根茎和球茎, 还是国家一级保护鸟类——白头鹤的重要饵料(Ma et al ., 2003 ; Zhu et al ., 2007 ), 因此, 海三棱藨草对崇明东滩所有滨海湿地鸟类都显得至关重要.然而近年来滩涂围垦、堤外放牧及外来物种竞争等导致植物种群分布状况发生变化, 植物群落斑块破碎化严重, 整个生态系统服务功能也随之降低(张利权和雍学葵, 1992; Chen et al ., 2004; 陈中义等, 2005b; Wang et al ., 2010).因此, 确定包括海三棱藨草在内的关键植物种群在东滩的空间分布, 明确其随环境因子的变化规律, 对该湿地生态系统的修复和保护具有重要的指导意义. ...
... 崇明东滩属于典型的淤涨型潮滩, 目前滩涂植被覆盖区总面积4265 hm2 , 2012年调查数据显示, 崇明东滩中部、北部植物以芦苇和互花米草为主, 而莎草科植物主要集中分布在崇明东滩南部1) ( 1) 吴纪华 (2014). 上海崇明东滩鸟类国家级自然保护区2012年度资源监测公报, 14-15.) .在崇明东滩的相关研究中, 众多学者根据研究需要将崇明东滩的海三棱藨草、藨草、糙叶薹草3种植物统一归入海三棱藨草类群(黄华梅等, 2007 ; Yuan & Zhang, 2008 ; 严格等, 2014 ).部分研究虽然将海三棱藨草、藨草、糙叶薹草统一划分为莎草科植物群落或者藨草群落(曹浩冰等, 2014 ; 任璘婧等, 2014 ), 但没有对这3种植物进行细致的划分, 各种群面积大小及其具体分布目前尚无相关报道.上述3种植物虽然同属于莎草科, 但占据不同的生态位, 承担着不同的生态功能, 例如在崇明东滩三者虽然都可以作为湿地鸟类的饵料, 但不同的鸟类会取食不同植物的不同部分(Ma et al ., 2003 ; Zhu et al ., 2007 ), 且3种植物组织、器官发育和成熟时间也各不相同.本文基于崇明东滩南部各植物种群的细致调查, 明确了各主要植物种群的空间分布以及各自的面积, 为进一步研究植被分布格局变化以及重要鸟类觅食生境提供基础. ...
Impact of land reclamation on the evolution of shoreline change and nearshore vegetation distribution in Yangtze River Estuary.
1
2014
... 盐沼植物作为全球范围内生产力较高的生物群落之一, 是推动湿地生态系统物质循环和能量流动的基础要素, 其在维持个体生存、种群稳定与群落发展的同时, 也为湿地生态系统中的众多成员提供了丰富的食物, 并创造了良好的栖息环境.然而, 近年来由于全球气候变化和频繁的人类活动, 植物群落组成结构以及空间分布格局发生了巨大的变化, 这不仅影响了湿地中鸟类、底栖生物及鱼类等生物的种群数量, 而且也不断改变着植物类群的种群结构, 甚至可能会导致一些敏感物种的灭绝(童春富, 2012 ; Isacch et al ., 2014 ; Li et al ., 2014b ).因此, 明确湿地中主要植物种群及其分布状况, 掌握其随环境因子的变化规律, 对湿地生态系统的保护、恢复、重建都具有重要的参考价值.虽然前人已就此进行了较多的研究(黄华梅等, 2007 ; Duarte et al ., 2013 ; 曹浩冰等, 2014 ; Hughes, 2014 ; Li et al ., 2014a , 2014b ), 但河口潮滩区域海陆交互作用强烈、人类活动频繁, 使得滩面、潮沟及植被都处于不断变化中, 崇明东滩正是这样一个处在发育过程中的典型盐沼湿地生态系统.2014年崇明东滩互花米草(Spartina alterniflora )生态治理工程的实施, 可能对该区域盐沼植物的空间分布及其生境产生了重大影响(高宇和赵斌, 2006 ; Zhu et al ., 2014 ).因此, 在施工前期对围垦外围区进行全面的植被调查, 不仅能准确地掌握崇明东滩南部的植被类型和现有分布状况, 而且能为今后研究其植被空间动态变化提供基础数据. ...
基于遥感的上海崇明东滩湿地典型植被净初级生产力估算
1
2011
... 近十几年来, 大量研究人员通过遥感影像解译, 对崇明东滩主要植被进行了分区(Gao & Zhang, 2006 ; Tian et al ., 2008 ; 阿也提古丽·斯迪克等, 2010 ; 翟新源和杨苏新, 2013 ; 郑宗生等, 2010 ; 宗玮等, 2011 ), 普遍认为芦苇(Phragmites australis )、互花米草(Spartina alterniflora )及海三棱藨草(Scirpus mariqueter )是崇明东滩的“三大”主要植物类群.然而, 崇明东滩南部长期受长江入海淡水影响, 水体盐度相对较低, 在团结沙至大潮沟(图1 )存在较大面积的藨草(Scirpus triqueter )种群区(闫芊等, 2007 ; 章振亚等, 2012 ).同时发现, 糙叶薹草(Carex scabrifolia )在研究区内频繁出现, 白茅(Imperata cylindrica )在部分空间也已发展成优势种. ...
基于遥感的上海崇明东滩湿地典型植被净初级生产力估算
1
2011
... 近十几年来, 大量研究人员通过遥感影像解译, 对崇明东滩主要植被进行了分区(Gao & Zhang, 2006 ; Tian et al ., 2008 ; 阿也提古丽·斯迪克等, 2010 ; 翟新源和杨苏新, 2013 ; 郑宗生等, 2010 ; 宗玮等, 2011 ), 普遍认为芦苇(Phragmites australis )、互花米草(Spartina alterniflora )及海三棱藨草(Scirpus mariqueter )是崇明东滩的“三大”主要植物类群.然而, 崇明东滩南部长期受长江入海淡水影响, 水体盐度相对较低, 在团结沙至大潮沟(图1 )存在较大面积的藨草(Scirpus triqueter )种群区(闫芊等, 2007 ; 章振亚等, 2012 ).同时发现, 糙叶薹草(Carex scabrifolia )在研究区内频繁出现, 白茅(Imperata cylindrica )在部分空间也已发展成优势种. ...
 , 黄星
, 黄星

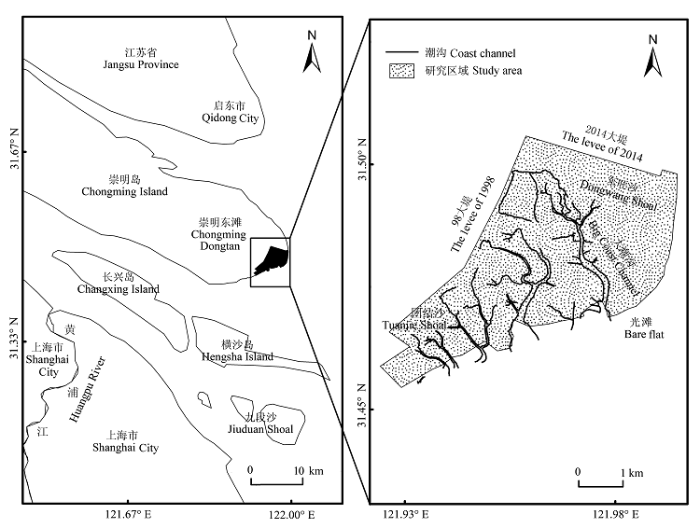
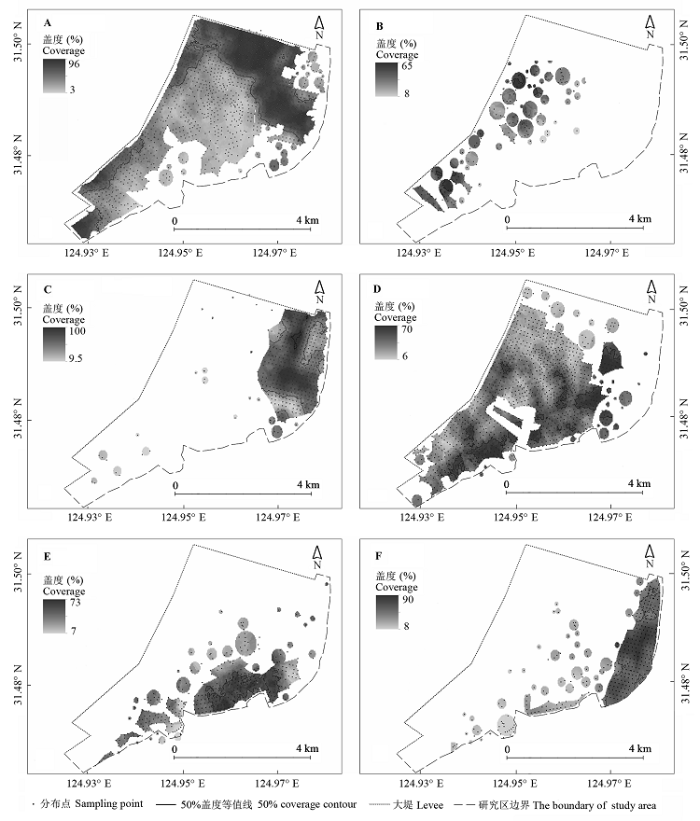
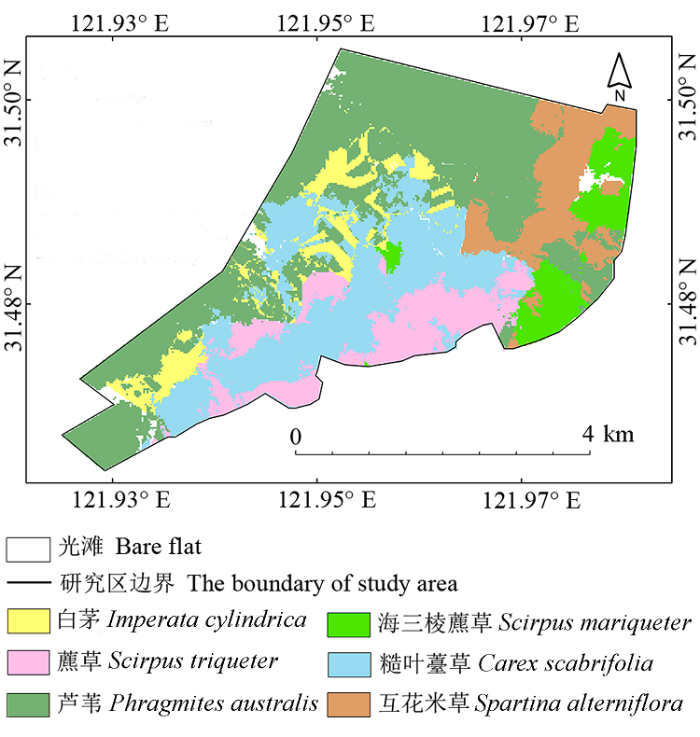
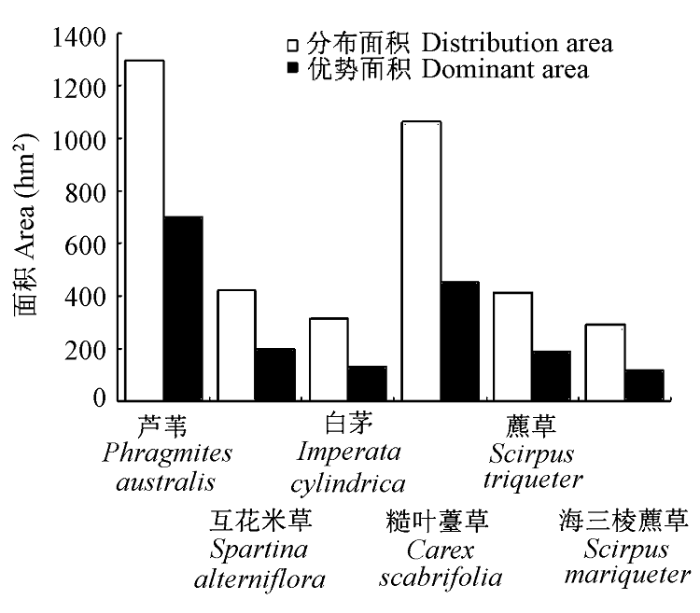

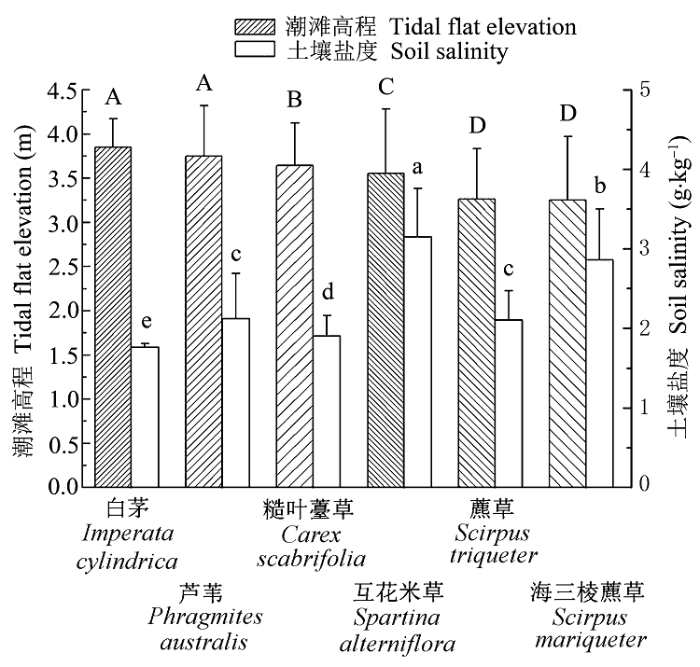
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}