在植物与昆虫的相互作用中, 有关花形态塑造和昆虫的适应进化是生态学和植物进化研究中最吸引人的一个领域(黄双全和郭友好, 2000 ; 黄双全, 2007 ; 方强和黄双全, 2014 )。鼠尾草属(Salvia )植物具有花器官两侧对称、二强雄蕊、虫媒异交和杠杆传粉机制等特征, 与传粉者的协同进化程度较高(张勃等, 2010 ), 典型的杠杆传粉过程基于雄蕊药隔的显著伸长、联合具阻挡作用的药隔下壁和明显可运动的“关节”等结构, 在传粉昆虫进入花冠取食花蜜时, 迫使其推动下臂做杠杆状运动, 上臂可育花药将花粉涂抹在昆虫背部从而携带出花粉。鼠尾草属雄蕊结构与功能的分化可能促进了该属物种多样性的形成(Claßen-Bockhoff et al ., 2004 )。根据传粉者类型, 鼠尾草属可分为蜂媒传粉(蜂媒花, melittophilae)和鸟媒传粉(鸟媒花, ornithophily), 蜂媒传粉物种主要分布于欧亚大陆、地中海沿岸、非洲东部及南非、大洋洲以及北美部分地区; 鸟媒传粉物种分布于北美西部至墨西哥、中美洲和南美洲安第斯山脉及东部(魏宇昆等, 2015 )。就雄蕊结构和传粉特征而言, 鼠尾草属中还具有非典型的杠杆状雄蕊结构和传粉模式。
黄艳波等(2014)根据花器官结构和花粉接触传粉昆虫的部位, 将中国的鼠尾草属物种划分为3种传粉模式类型: 短药隔杠杆、长药隔杠杆和退化杠杆。3个类群大致形成以短药隔杠杆物种集中分布于中国西南, 长药隔杠杆物种集中于华中, 退化杠杆类型以华东为核心的分布格局。对鼠尾草属部分物种的杠杆传粉机制了解较多, 如短药隔杠杆类型的圆苞鼠尾草(S. cyclostegia )和长药隔杠杆类型丹参(S. miltiorrhiza )均借助传粉昆虫推动雄蕊做杠杆状运动, 完成花粉分发和授粉作用(张勃等, 2012 ; 黄艳波等, 2014 )。在中国, 鼠尾草亚属(subgenus Allagospadonopsis )的16个物种具有退化的杠杆结构, 推测它们具有非典型的杠杆传粉特征(黄艳波等, 2014 )。鼠尾草属雄蕊的分化是物种形成和多样性产生的关键因素(Claßen-Bockhoff et al ., 2003 ), 这种适应辐射的物种形成事件也是各种环境选择压力综合作用的结果。尽管对西南分布的短药隔类型鼠尾草的传粉结构、进化响应和表型选择等已有所了解(Zhang et al ., 2011 ; 张勃等, 2012 ; Zhang & Li, 2014 ), 但相比鼠尾草属多样性的传粉结构和传粉过程, 对其中一种类型的认识显然不足以揭示鼠尾草属面对的传粉者选择压力和进化方向。更何况, 至今对该属植物的传粉生态学和繁育系统仍然了解不多(Claßen-Bockhoff et al ., 2003 )。
中国分布的鼠尾草属物种虽不算丰富, 但雄蕊类型比较全面。对鼠尾草属雄蕊结构类型和传粉昆虫相互作用的系统研究, 将阐明该类群物种多样性的起源和不同选择压力下鼠尾草属物种的进化方向。舌瓣鼠尾草(S. liguliloba )是鼠尾草亚属退化杠杆类型的代表性物种, 是分布于华东天目山-黄山区系的特有种。本文将对其花器官结构及传粉过程进行分析, 了解其传粉昆虫组成和访花特征, 同时与短药隔类型的毛地黄鼠尾草(S. digitaloides )进行比较, 以揭示退化杠杆可能的进化选择压力及其生态学意义。
1 材料和方法
1.1 样地自然概况
舌瓣鼠尾草研究地点位于天目山国家级自然保护区(30.39° N, 119.48° E), 海拔677 m。天目山保护区属北亚热带气候, 季风强盛、四季分明、气候温和、雨水丰沛、光照适宜。年平均气温8.9-15.8 ℃, 最冷月气温为2.6-3.4 ℃, 最热月气温为19.9-28.1 ℃, ≥ 10 ℃积温2500-5100 ℃, 年降水量1390-1870 mm, 全年无霜期209-235天, 年太阳总辐射3770- 4460 MJ·m-2 (杨逢春, 1992 )。天目山植物区系的温带、亚热带东亚区系成分特征显著, 在地理上和发生上与东亚日本、北美、东南亚等有着不同程度的联系(杨逢春, 1992 )。研究样地植被属于常绿阔叶林(杨逢春, 1992 ), 其群落组成主要为菊科一年蓬(Erigeron annuus )、蒲儿根(Sinosenecio oldhamianus ), 报春花科过路黄(Lysimachia christinae ), 荨麻科青叶苎麻(Boehmeria nivea var. tenacissima ), 山茶科茶(Camellia sinensis ), 禾本科阔叶箬竹(Indocalamus latifolius )、早竹(Phyllostachys violascens )、求米草(Oplismenus undulatifolius ), 唇形科紫花香薷(Elsholtzia argyi ), 菝葜科菝葜(Smilax china ), 杉科杉木(Cunninghamia lanceolata ), 胡桃科山核桃(Carya cathayensis )。其中优势种主要是青叶苎麻、早竹和山核桃等。
毛地黄鼠尾草研究样地位于云南省丽江市玉龙雪山自然保护区甘海子(27.06° N, 100.26° E), 海拔2990 m, 属青藏高原东南缘, 横断山系南端。玉龙雪山海拔落差及气候垂直差异极大, 最高峰海拔5596 m, 相对高差3846 m (冯建孟等, 2006 )。区内植被类型多样, 植物区系起源古老, 地理成分复杂, 特有种丰富, 分化显著。气候干湿季分明, 5-10月为雨季, 降水量为全年降水量的85%-99% (辛惠娟等, 2013 ), 其中2009年海拔3046 m记录的5-10月降水量为1884.3 mm (辛惠娟等, 2012 )。年平均气温12.79 ℃, 最热月7月平均气温为17.9 ℃, 最冷月1月平均气温为5.9 ℃。植被属高山云南松(Pinus yunnanensis )、丽江云杉(Picea likiangensis )和长苞冷杉(Abies georgei )针叶林。研究样地生境是林缘间开阔草地群落, 毛地黄鼠尾草是其中常见的优势物种。
1.2 研究材料
舌瓣鼠尾草为鼠尾草亚属一至二年生直立草本, 天目山-黄山地区特有物种, 株高30-70 cm, 植株单一或偶有分枝, 单叶披针形, 以基生叶为主, 2-4对茎生叶, 叶长3-11 cm, 宽1.2-3.0 cm。唇形花冠淡蓝紫色, 雄蕊及雌蕊上部颜色较深, 花药乳白色。花期5月中旬至6月中旬, 果期从5月下旬开始, 6月10日至30日为成熟期, 每朵花可产生4枚棕褐色小坚果。在本研究区域, 舌瓣鼠尾草为常见草本植物, 多分布于林下、路边及溪边, 同域分布的鼠尾草属植物有南丹参(Salvia bowleyana )。研究样地属舌瓣鼠尾草自然分布种群, 样地面积约50 m2 , 种群密度为8.6株·m-2 (图1 )。
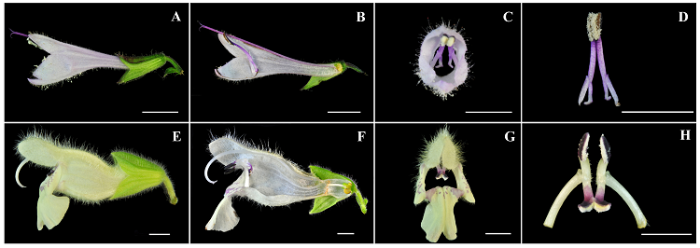
图1 舌瓣鼠尾草种群、植株、花器官形态及访花昆虫。A, 生境。B, 植株。C, 花侧面。D, 花纵剖。E, 雌蕊。F, 花正面。G, 雄蕊。H, 果实。I, 三条熊蜂。J, 隧蜂科一种(示倒向取食)。K, 隧蜂科一种(示正向取食)。L, 天蛾科一种。图中标尺均为2 mm。
Fig. 1 Photographic images of Salvia liguliloba and its visiting insects. A, Habitat. B, Plant. C, Lateral view of flower. D, Longitudinal section of flower. E, Pistil. F, Front view of flower. G, Stamens. H, Fruits. I, Bombus trifasciatus (pollinator). J, Ventral visiting by a species of Halictidae. K, Dorsal visiting by a species of Halictidae. L, A visitor of Sphingidae species. Bar = 2 mm.
毛地黄鼠尾草是弧隔鼠尾草亚属(subgenus Salvia )的多年生草本植物, 产于云南西北部和四川西南部, 株高30-60 cm, 叶通常基出, 间有茎生叶, 叶片较宽大, 长圆状椭圆形, 长3.5-11.0 cm, 宽2-5 cm, 叶片两面尤其叶背面密被白色短柔毛, 花冠黄色具淡紫色斑点, 冠筒基部较窄, 向上逐渐增大并在冠筒中部有一明显上弯, 至喉部宽达1 cm, 花期4-5月和8-9月。4枚小坚果灰黑色, 倒卵圆形光滑。
1.3 研究方法
1.3.1 花器官和昆虫的测量
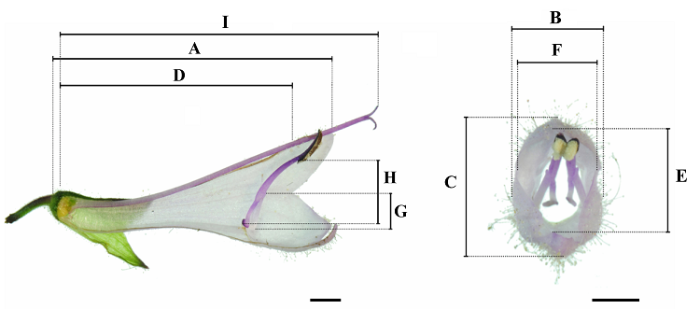
在植物盛花期, 随机选取样地内不同植株上完全开放的花, 用电子游标卡尺对解剖后的花各部结构进行测量。测量性状包括花冠长(A)、花冠宽(B)、花冠高(C)、冠筒长(D)、冠口高(E)、冠口宽(F)、花丝长(G)、药隔长(H)、雌蕊长(I), 方法及位置如图2 所示, 2种鼠尾草各测量10-20个重复。
图2 舌瓣鼠尾草花器官测量示意图。A, 花冠长。B, 花冠宽。C, 花冠高。D, 冠筒长。E, 冠口高。F, 冠口宽。G, 花丝长。H, 药隔长。I, 雌蕊长。图中标尺均为2 mm。
Fig. 2 Diagram of Salvia liguliloba flower measurement. A, Corolla length. B, Corolla width. C, Corolla height. D, Tube length. E, Entrance height. F, Entrance width. G, Filament length. H, Connective length. I, Pistil length. Bar = 2 mm.
采集植物种群中出现的访花昆虫, 首先用乙酸乙酯将昆虫毒死, 用镊子辅助保持自然伸展状态, 用电子游标卡尺进行形态特征测量, 包括昆虫体长、胸宽、胸厚和喙长4个指标。喙长测量时用镊子将其拉出至自然伸展状态, 测量完成后用昆虫针将其固定, 制成标本带回实验室进行拍照鉴定。对舌瓣鼠尾草采集样地内出现的所有访花昆虫进行鉴定测量, 毛地黄鼠尾草的传粉昆虫参考张勃等(2012) 在甘海子的测量结果。
1.3.2 开花与传粉观察
在盛花期对样地内舌瓣鼠尾草的开花时间、开放式样、单花持续时间、雌雄蕊的形态变化、花药散粉时间、柱头可授期进行观察和拍照。记录访花昆虫的种类、访花姿态、花间飞行方式、是否取食花蜜或花粉、访花时接触的花器官部位等。在天气晴朗条件下, 统计访花昆虫的单花访问时间, 单位时间内某一只昆虫的访问花朵数量。利用数码摄录机(SONY, HDR-CX510E)对全天(7:00-18:00)固定样方内的植物访花情况进行不间断拍摄(样方内共计75朵开放花), 记录所有进入样方的访花昆虫, 统计样方内的昆虫种类、出现和访花次数。
1.3.3 传粉生态学特征
参考Talavera等(2001) 以及龚燕兵和黄双全(2007)的方法, 确定并计算: (1)有效传粉昆虫(effective pollinator), 可携带花粉并完成授粉的昆虫; (2)相对频次(relative frequency, F ), 在固定样方内, 某一种访花昆虫的出现次数占所有访花昆虫出现总次数的相对比例; (3)昆虫传粉的日活动变化规律, 用固定样方内每小时的昆虫访花总次数表示; (4)昆虫单花访问时间(T ), 即访花者从登陆花冠口到访花结束后飞离花冠的时间; (5)活跃度(AR ), 以单只昆虫每分钟的访问花朵数量表示, 代表了此种昆虫对某一种植物花的偏好程度和传粉潜力; (6)访花频率, 用指数IVR = F × AR 表示, 访花频率同时考虑了昆虫的活跃度及其出现的相对频次; (7)传粉回报, 传粉者为植物传粉获得的物质补偿, 一般是花粉或花蜜。毛地黄鼠尾草访花昆虫的访花特征参考张勃等(2012)在甘海子样地的观测结果。
2 结果和分析
2.1 花器官特征
舌瓣鼠尾草花序的基本单位是由6朵小花构成的2个二歧聚伞花序, 再组成1个轮伞花序。每一花序分枝通常由8-19层轮伞花序组成, 整个植株花序又由3或5或7个花序分枝构成假圆锥花序, 植株的小花总数量为144-798朵, 整株花序的小花偏向一侧生长(图1 )。舌瓣鼠尾草种群始花期为5月中旬, 至6月中旬花期基本结束。整株花序为自下而上的开花顺序, 在轮伞花序中, 2个二歧聚伞花序均为中间2朵小花最先同时开放, 每个分枝花序中同一时间至少保证有相邻的2层轮伞花序均有在盛花期的小花, 同时完全开放的小花数量为3-10朵。舌瓣鼠尾草的单花平均开放时间(单花花期)为24.3 h (n = 23), 花粉囊在花朵刚刚开放时即已成熟开裂, 而雌蕊成熟较晚, 当柱头顶端明显开裂就预示其具有了可 授性。
舌瓣鼠尾草花器官的形态特征如图3 A-3D所示, 花萼窄钟形, 花冠呈现两侧对称的细筒状结构, 花冠口较宽, 至喉部以下明显变窄。雄蕊内藏于花冠内, 浅紫色, 但较其他器官颜色深。花粉囊中的药隔显著伸长将2个药室撑开, 在花丝与药隔的连接处形成“关节”, 将药隔分为上臂和下臂。上臂可育花粉囊成熟后向下开裂, 2药室靠合, 紧贴冠筒内壁, 不伸出花冠上唇, 但在冠口正面可见浅黄色花粉团。下臂花粉囊退化不育, 短小呈弯曲状, 顶端不联合, 花丝(2.39 ± 0.14) mm生于冠筒底部两侧, 略向冠筒中央倾斜, 显著短于药隔(5.19 ± 0.33) mm。雌蕊略伸出花冠, 成熟后明显二裂。花冠外被较密的短柔毛及腺毛。
图3 舌瓣鼠尾草(A-D)与毛地黄鼠尾草(E-H)花器官形态。A, E, 花侧面。B, F, 花器官侧剖面。C, G, 花正面。D, H, 雄蕊。图中标尺为5 mm
Fig. 3 Comparisons of morphometric data between Salvia liguliloba (A to D) and S . digitaloides (E to H). A, E, Lateral view of a flower. B, F, Longitudinal section of a flower. C, G, Front view of a flower. D, H, Stamens. Bar = 5 mm.
毛地黄鼠尾草花器官特征如图3E -3H所示, 花序生长方式与舌瓣鼠尾草相同, 每个轮伞花序4-6朵花, 3-8层组成总状花序, 再由下部分枝组成假圆锥花序。盛花期单株开放的小花总数量平均为(32 ± 35)(n = 16)朵。花萼宽钟形, 花冠呈现两侧对称的宽筒状结构, 花冠浅黄色, 冠口较宽, 至花萼处变窄且有一定弯曲。雄蕊内藏于花冠内, 粉紫色, 冠筒内部靠近冠口有淡紫色斑点, 花粉囊中的药隔伸长将2个药室撑开, 形成雄蕊药隔的上下臂。上、下臂花粉囊均可育产生花粉, 上臂2个花粉囊分离, 散粉时向外侧开裂, 下臂花粉囊略小联合, 向下开裂散粉。从冠筒正面只可见下臂淡黄色花药, 上臂花药完全隐藏于上唇内。花丝(7.33 ± 0.46) mm生于冠筒底部两侧, 向冠筒中央倾斜, 花丝长于药隔(5.10 ± 0.43) mm。雌蕊伸出冠筒之外, 向下内卷。花冠外密被长柔毛。
2.2 昆虫的访花行为
在舌瓣鼠尾草的样方内记录观察到3种访花昆虫, 分别是蜜蜂科熊蜂属(Bombus )的三条熊蜂(B. trifasciatus ), 隧蜂科(Halictidae)和天蛾科(Sphing- idae)昆虫(图1 )。因天蛾科昆虫数量少、飞行速度快, 未能捕捉到活体, 故只测量了三条熊蜂和隧蜂科昆虫。从体型大小看, 3种访花昆虫形态特征差异明显(表2 ), 并且访花行为也完全不同。如图4 所示, 三条熊蜂体型较大, 只有一种访花姿态, 即直接飞至冠口, 依靠其前、中足抓住花冠前部保持平衡, 头部插入冠筒, 利用其长喙吸食花蜜, 此时前额紧贴雄蕊上臂的花粉囊, 将花粉带出, 完整的访花时间为(1.54 ± 0.60) s。当它再次进入另一朵具成熟雌蕊的花冠时, 便可将其头部的花粉授予柱头, 完成授粉。隧蜂科昆虫体型小, 通常飞落至花冠口再缓慢爬进冠筒, 自由地取食花粉, 并未取食花蜜, 而此时雌蕊尚未成熟。其单花访花时间很长、变幅较大, 平均为(73.9 ± 55.6) s。另一种天蛾科昆虫的飞行速率极快, 单花访花时间短, 平均只有(0.67 ± 0.23) s, 因其喙很长, 取食花蜜时没有触碰到花器官任何部位(图1 L)。在甘海子样地, 毛地黄鼠尾草的访花昆虫有寄生性拟熊蜂(Psithyrus sp.)、小雅熊蜂(Bombus lepidus )和Bombus infrequens 。三种访花昆虫主要进行背部传粉, 小雅熊蜂兼进行腹部传粉(表3 )(张勃等, 2012 )。它们的访花姿态相似, 先落至花冠下唇, 再爬进冠筒取食花蜜, 在此过程中推动雄蕊下臂做杠杆状运动, 上臂花粉囊向下打在昆虫背部或腹部, 待其离开花器官时携带出花粉, 访花时间较长(本文作者观察)。
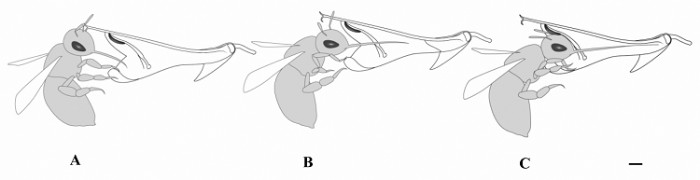
图4 舌瓣鼠尾草传粉昆虫三条熊蜂的传粉过程示意图。A, 接近花冠。B, 准备进入冠筒, 喙逐渐伸出。C, 喙已完全深入冠筒取食花蜜, 头部顶住花药, 上臂花粉落至其前额部。图中标尺为2 mm。
Fig. 4 The pollination process of Bombus trifasciatus, an effective pollinator of Salvia liguliloba . A, Approaching to the corolla. B, Accessing to flower tube and the pollinator’s tongue is extending. C, B . trifasciatus is sucking nectar and its forehead touching the fertile anther. Bar = 2 mm.
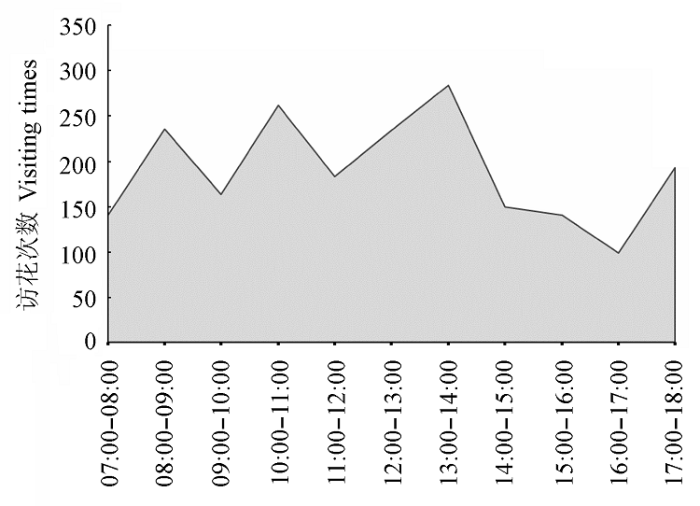
三条熊蜂是舌瓣鼠尾草的唯一传粉昆虫, 对一天中样方内访花昆虫访问次数的统计表明: 三条熊蜂出现的相对频次为0.959, 占绝对优势, 隧蜂科和天蛾科昆虫分别仅占0.016和0.024。我们对三条熊蜂在一天中的访花进程进行进一步观察, 统计每小时样方内三条熊蜂的总访花次数, 以此了解舌瓣鼠尾草传粉昆虫的活动规律。从图5 反映出三条熊蜂从7:00至18:00均有访花行为, 从早至晚的活动频率呈现波浪状起伏, 但整体看上午的访花相对频繁, 且逐渐升高, 至14:00达到最高峰, 随后明显下降, 至18:00左右又略有上升。
图5 三条熊蜂对舌瓣鼠尾草的访花日动态。
Fig. 5 The daily dynamic of flower-visiting times for Salvia liguliloba by Bombus trifasciatus .
3 讨论
本文的研究表明, 退化杠杆类型的舌瓣鼠尾草无论从花器官结构还是昆虫访花特征, 都与短药隔杠杆类型的毛地黄鼠尾草明显不同。这体现了鼠尾草属花器官结构与生态功能的高度适应, 同时暗示该属不同类型的传粉模式源于各异的选择压力, 经历了不同的进化途径。但究竟鼠尾草属不同类型的传粉结构和机制是如何形成的?传粉昆虫在其中起到了何种作用?东亚鼠尾草属内是单系起源还是平行进化? 对这些问题都还不清楚, 未来还需对以上几个问题做深入研究和明确阐述。
相比毛地黄鼠尾草, 舌瓣鼠尾草传粉昆虫的单花访问时间、活跃度和访花频率均较高(表3 ), 说明小型花冠和退化的杠杆结构缩短了访花时间, 提高了活跃度, 可能促进了昆虫的访花效率。在传粉稳定性方面, 舌瓣鼠尾草花结构能够固定访花时昆虫取食花蜜的姿态, 保证传粉的可靠性和稳定性, 同时固定的花药和昆虫接触部位, 进一步提高了传粉的精确性。传粉结构和本地传粉者的特征共同决定了舌瓣鼠尾草的传粉方式。短药隔杠杆的毛地黄鼠尾草, 传粉昆虫进入冠筒有2种方式, 并且在冠筒中有自由活动的空间, 身体各部位都有接触花药的可能, 沾在腹部的花粉在飞行途中也有可能掉落, 增大了花粉损失的可能(张勃等, 2012 )。本研究仅从花器官结构和传粉昆虫的相互作用角度进行了深入研究, 具退化杠杆的物种是否具有选择优势?哪一种传粉模式更有效?还需从鼠尾草属植物的雄性适合度和繁育系统方面进行比较分析, 从而更深入地了解它们的生态学意义和进化选择压力。
舌瓣鼠尾草有专一性的传粉昆虫三条熊蜂, 毛地黄鼠尾草则有3种。专一性的提高是否暗示植物与昆虫的相互关系更紧密, 协同进化程度更高?舌瓣鼠尾草传粉者的高度专一性可能还与所在地的昆虫种类组成相关, 究竟是退化杠杆结构过滤了其他传粉者, 还是传粉者的改变导致杠杆功能丧失? 都需要进一步深入研究。另一方面, 专一性的传粉昆虫面临的风险较大, 一旦失去传粉者, 就有无法延续生存的可能。相反, 具有多个传粉者的物种, 这样的风险则大大降低。目前对鼠尾草属短药隔杠杆机制的作用以及传粉者响应的生态学意义等方面研究较充分(Zhang et al ., 2011 ; 张勃等, 2012 ; Zhang & Li, 2014 ), 而其他类型杠杆的研究还很少, 对该属多种类型杠杆机制的阐明, 无疑会对揭示其起源和进化方向以及该物种多样性的产生、共存和维持机制提供更多证据。
在宏观形态学方面, 鼠尾草属杠杆呈现出雄蕊药隔逐渐伸长、药隔下臂逐渐退化形成附属结构, 直至最后完全丧失功能这样的进化历程(Claßen- Bockhoff et al ., 2003 ; 黄艳波等, 2014 )。毛地黄鼠尾草与舌瓣鼠尾草恰似占据此进化历程的两极。Li等(2013)将中国产43种鼠尾草属植物构建了系统进化树, 尽管涵盖了舌瓣鼠尾草所在的鼠尾草亚属和毛地黄鼠尾草所在的弧隔鼠尾草亚属, 但并没有采用毛地黄鼠尾草这个材料。仅从该文中两个种所在的亚属来看, 舌瓣鼠尾草所在的鼠尾草亚属首先与丹参所在的荔枝草亚属(subgenus Sclarea )聚为一支, 再与毛地黄鼠尾草所在的弧隔鼠尾草亚属聚为一支, 可以间接判断毛地黄鼠尾草所在的弧隔鼠尾草亚属可能更古老原始, 而舌瓣鼠尾草的进化时间更短, 成种时间较晚。由于该属可能处在迅速的辐射成种阶段, 仅使用几个叶绿体和核基因片段来研究鼠尾草属的种间系统关系, 信息量似乎远远不足, 未来需借助更多的标记或二代测序技术, 结合物种分布格局和谱系地理研究(王琦等, 2015 ), 从而更深入地理解不同传粉模式的系统发育关系。
致谢 中国科学院动物研究所朱朝东研究员和姚建高级工程师在鉴定昆虫标本中提供帮助, 中国科学院计算生物学研究所刘欣雨协助绘图, 一并致谢!
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Claßen-Bockhoff R Speck T Tweraser E Wester P Thimm S Reith M 2004 ). The staminal lever mechanism in Salvia L. (Lamiaceae): A key innovation for adaptive radiation?
Organisms Diversity & Evolution , 4 , 189 -205 .
[本文引用: 1]
[2]
Claßen-Bockhoff R Wester P Tweraser E 2003 ). The staminal lever mechanism in Salvia L. (Lamiaceae)—A review.
Plant Biology , 5 , 33 -41 .
[本文引用: 3]
[3]
Fang Q Huang SQ 2014 ). Progress in pollination ecology at the community level.
Chinese Science Bulletin , 59 , 449 -458 .
[本文引用: 1]
(in Chinese with English abstract) [方强 , 黄双全 (2014 ). 群落水平上传粉生态学的研究进展
. 科学通报 , 59 , 449 -458 .]
[本文引用: 1]
[4]
Feng JM Wang XP Xu CD Yang YH Fang JY 2006 ). Altitudinal patterns of plant species diversity and community structure on Yulong Mountains, Yunnan, China.
Journal of Mountain Science , 24 , 110 -116 .
[本文引用: 1]
(in Chinese with English abstract) [冯建孟 , 王襄平 , 徐成东 , 杨元合 , 方精云 (2006 ). 玉龙雪山植物物种多样性和群落结构沿海拔梯度的分布格局
. 山地学报 , 24 , 110 -116 .]
[本文引用: 1]
[5]
Gong YB Huang SQ 2007 ). On methodology of foraging behavior of pollinating insects.
Biodiversity Science , 15 , 576 -583 .
(in Chinese with English abstract) [龚燕兵 , 黄双全 (2007 ). 传粉昆虫行为的研究方法探讨
. 生物多样性 , 15 , 576 -583 .]
[6]
Huang SQ 2007 ). Studies on plant-pollinator interaction and its significances.
Biodiversity Science , 15 , 569 -575 .
[本文引用: 1]
(in Chinese with English abstract) [黄双全 (2007 ). 植物与传粉者相互作用的研究及其意义
. 生物多样性 , 15 , 569 -575 .]
[本文引用: 1]
[7]
Huang SQ Guo YH 2000 ). New advances in pollination biology.
Chinese Science Bulletin , 45 , 225 -237 .
[本文引用: 1]
(in Chinese) [黄双全 , 郭友好 (2000 ). 传粉生物学的研究进展
. 科学通报 , 45 , 225 -237 .]
[本文引用: 1]
[8]
Huang YB Wei YK Ge BJ Wang Q 2014 ). Pollination Mechanisms of genus Salvia (Lamiaceae) in East Asia (China).
Acta Ecologica Sinica , 34 , 2282 -2289 .
[本文引用: 3]
(in Chinese with English abstract) [黄艳波 , 魏宇昆 , 葛斌杰 , 王琦 (2014 ). 鼠尾草属东亚分支的传粉模式
. 生态学报 , 34 , 2282 -2289 .]
[本文引用: 3]
[9]
Li QQ Li MH Yuan QJ Cui ZH Huang LQ Xiao PG 2013 ) Phylogenetic relationships of Salvia (Lamiaceae) in China: Evidence from DNA sequence datasets.
Journal of Systematics and Evolution , 51 , 184 -195 .
[10]
Talavera S Bastida F Ortiz PL Arista M 2001 ). Pollinator attendance and reproductive success in Cistus libanotis L. (Cistaceae).
International Journal of Plant Sciences , 162 , 343 -352 .
[本文引用: 1]
[11]
Wang Q Wei YK Huang YB 2015 ). Research on distribution pattern of Subg. Salvia Benth. (Lamiaceae), an important group of medicinal plants in East Asia.
Acta Ecologica Sinica , 35 , 1470 -1479 .
[本文引用: 1]
(in Chinese with English abstract) [王琦 , 魏宇昆 , 黄艳波 (2015 ). 中国弧隔鼠尾草亚属(唇形科)的分布格局
. 生态学报 , 35 , 1470 -1479 .]
[本文引用: 1]
[12]
Wei YK Wang Q Huang YB 2015 ). Species diversity and distribution of Salvia (Lamiaceae).
Biodiversity Science , 23 , 3 -10 .
[本文引用: 1]
(in Chinese with English abstract) [魏宇昆 , 王琦 , 黄艳波 (2015 ). 唇形科鼠尾草属的物种多样性与分布
. 生物多样性 , 23 , 3 -10 .]
[本文引用: 1]
[13]
Xin HJ He YQ Li ZX Wang SJ Du JK Wang CF Pu T Zhang W 2012 ). Inter-annual variation of temperature and precipitation gradient at the Eastern slope of Yulong Snow Mountain. Earth Science-
Journal of China University of Geosciences , 37 (Suppl. ), 188 -194 .
[本文引用: 1]
(in Chinese with English abstract) [辛惠娟 , 何元庆 , 李宗省 , 王世金 , 杜建括 , 王春凤 , 蒲焘 , 张蔚 (2012 ). 玉龙雪山东坡气温和降水梯度年内变化特征
. 地球科学——中国地质大学学报 , 37 (Suppl. ), 188 -194 .]
[本文引用: 1]
[14]
Xin HJ He YQ Zhang T Niu HW Du JK 2013 ). The features of climate variation and glacier response in Mt. Yulong, southeastern Tibetan Plateau.
Advances in Earth Science , 28 , 1257 -1268 .
[本文引用: 1]
(in Chinese with English abstract) [辛惠娟 , 何元庆 , 张涛 , 牛贺文 , 杜建括 (2013 ). 青藏高原东南缘丽江玉龙雪山气候变化特征及其对冰川变化的影响
. 地球科学进展 , 28 , 1257 -1268 .]
[本文引用: 1]
[15]
Yang FC 1992 ). Comprehensive Investigation Report on Natural Resource of Tianmu Mountain Nature Reserve . Zhejiang Science and Technology Press , Hangzhou .
[本文引用: 3]
(in Chinese) [杨逢春 (1992 ). 天目山自然保护区自然资源综合考察报告 . 浙江科学技术出版社 , 杭州 .]
[本文引用: 3]
[16]
Zhang B Claßen-Bockhoff R Zhang ZQ Sun S Luo YJ Li QJ 2011 ). Functional implications of the staminal lever mechanism in Salvia cyclostegia (Lamiaceae).
Annals of Botany , 107 , 621 -628 .
[本文引用: 2]
[17]
Zhang B Li QJ 2014 ). Phenotypic selection on the staminal lever mechanism in Salvia digitaloides (Labiaceae).
Evolutionary Ecology , 28 , 373 -386 .
[本文引用: 2]
[18]
Zhang B Sun S Fang QE Bai XM 2012 ). Evolutionary response of staminal lever mechanism of different species in Salvia to spatial variation in pollinators.
Chinese Journal of Plant Ecology , 36 , 681 -689 .
[本文引用: 8]
(in Chinese with English abstract) [张勃 , 孙杉 , 方强恩 , 白小明 (2012 ). 鼠尾草属不同物种的雄蕊杠杆机制对传粉者空间变异的进化响应
. 植物生态学报 , 36 , 681 -689 .]
[本文引用: 8]
[19]
Zhang B Sun S Zhang ZQ Li QJ 2010 ). A review of the evolutionary and ecological significance of lever-like stamens.
Chinese Journal of Plant Ecology , 34 , 89 -99 .
[本文引用: 1]
(in Chinese with English abstract) [张勃 , 孙杉 , 张志强 , 李庆军 (2010 ). 杠杆状雄蕊及其进化生态学意义
. 植物生态学报 , 34 , 89 -99 .]
[本文引用: 1]
The staminal lever mechanism in Salvia L. (Lamiaceae): A key innovation for adaptive radiation?
1
2004
... 在植物与昆虫的相互作用中, 有关花形态塑造和昆虫的适应进化是生态学和植物进化研究中最吸引人的一个领域(黄双全和郭友好, 2000 ; 黄双全, 2007 ; 方强和黄双全, 2014 ).鼠尾草属(Salvia )植物具有花器官两侧对称、二强雄蕊、虫媒异交和杠杆传粉机制等特征, 与传粉者的协同进化程度较高(张勃等, 2010 ), 典型的杠杆传粉过程基于雄蕊药隔的显著伸长、联合具阻挡作用的药隔下壁和明显可运动的“关节”等结构, 在传粉昆虫进入花冠取食花蜜时, 迫使其推动下臂做杠杆状运动, 上臂可育花药将花粉涂抹在昆虫背部从而携带出花粉.鼠尾草属雄蕊结构与功能的分化可能促进了该属物种多样性的形成(Claßen-Bockhoff et al ., 2004 ).根据传粉者类型, 鼠尾草属可分为蜂媒传粉(蜂媒花, melittophilae)和鸟媒传粉(鸟媒花, ornithophily), 蜂媒传粉物种主要分布于欧亚大陆、地中海沿岸、非洲东部及南非、大洋洲以及北美部分地区; 鸟媒传粉物种分布于北美西部至墨西哥、中美洲和南美洲安第斯山脉及东部(魏宇昆等, 2015 ).就雄蕊结构和传粉特征而言, 鼠尾草属中还具有非典型的杠杆状雄蕊结构和传粉模式. ...
The staminal lever mechanism in Salvia L. (Lamiaceae)—A review.
3
2003
... 黄艳波等(2014)根据花器官结构和花粉接触传粉昆虫的部位, 将中国的鼠尾草属物种划分为3种传粉模式类型: 短药隔杠杆、长药隔杠杆和退化杠杆.3个类群大致形成以短药隔杠杆物种集中分布于中国西南, 长药隔杠杆物种集中于华中, 退化杠杆类型以华东为核心的分布格局.对鼠尾草属部分物种的杠杆传粉机制了解较多, 如短药隔杠杆类型的圆苞鼠尾草(S. cyclostegia )和长药隔杠杆类型丹参(S. miltiorrhiza )均借助传粉昆虫推动雄蕊做杠杆状运动, 完成花粉分发和授粉作用(张勃等, 2012 ; 黄艳波等, 2014 ).在中国, 鼠尾草亚属(subgenus Allagospadonopsis )的16个物种具有退化的杠杆结构, 推测它们具有非典型的杠杆传粉特征(黄艳波等, 2014 ).鼠尾草属雄蕊的分化是物种形成和多样性产生的关键因素(Claßen-Bockhoff et al ., 2003 ), 这种适应辐射的物种形成事件也是各种环境选择压力综合作用的结果.尽管对西南分布的短药隔类型鼠尾草的传粉结构、进化响应和表型选择等已有所了解(Zhang et al ., 2011 ; 张勃等, 2012 ; Zhang & Li, 2014 ), 但相比鼠尾草属多样性的传粉结构和传粉过程, 对其中一种类型的认识显然不足以揭示鼠尾草属面对的传粉者选择压力和进化方向.更何况, 至今对该属植物的传粉生态学和繁育系统仍然了解不多(Claßen-Bockhoff et al ., 2003 ). ...
... ), 但相比鼠尾草属多样性的传粉结构和传粉过程, 对其中一种类型的认识显然不足以揭示鼠尾草属面对的传粉者选择压力和进化方向.更何况, 至今对该属植物的传粉生态学和繁育系统仍然了解不多(Claßen-Bockhoff et al ., 2003 ). ...
... 在宏观形态学方面, 鼠尾草属杠杆呈现出雄蕊药隔逐渐伸长、药隔下臂逐渐退化形成附属结构, 直至最后完全丧失功能这样的进化历程(Claßen- Bockhoff et al ., 2003 ; 黄艳波等, 2014 ).毛地黄鼠尾草与舌瓣鼠尾草恰似占据此进化历程的两极.Li等(2013)将中国产43种鼠尾草属植物构建了系统进化树, 尽管涵盖了舌瓣鼠尾草所在的鼠尾草亚属和毛地黄鼠尾草所在的弧隔鼠尾草亚属, 但并没有采用毛地黄鼠尾草这个材料.仅从该文中两个种所在的亚属来看, 舌瓣鼠尾草所在的鼠尾草亚属首先与丹参所在的荔枝草亚属(subgenus Sclarea )聚为一支, 再与毛地黄鼠尾草所在的弧隔鼠尾草亚属聚为一支, 可以间接判断毛地黄鼠尾草所在的弧隔鼠尾草亚属可能更古老原始, 而舌瓣鼠尾草的进化时间更短, 成种时间较晚.由于该属可能处在迅速的辐射成种阶段, 仅使用几个叶绿体和核基因片段来研究鼠尾草属的种间系统关系, 信息量似乎远远不足, 未来需借助更多的标记或二代测序技术, 结合物种分布格局和谱系地理研究(王琦等, 2015 ), 从而更深入地理解不同传粉模式的系统发育关系. ...
群落水平上传粉生态学的研究进展
1
2014
... 在植物与昆虫的相互作用中, 有关花形态塑造和昆虫的适应进化是生态学和植物进化研究中最吸引人的一个领域(黄双全和郭友好, 2000 ; 黄双全, 2007 ; 方强和黄双全, 2014 ).鼠尾草属(Salvia )植物具有花器官两侧对称、二强雄蕊、虫媒异交和杠杆传粉机制等特征, 与传粉者的协同进化程度较高(张勃等, 2010 ), 典型的杠杆传粉过程基于雄蕊药隔的显著伸长、联合具阻挡作用的药隔下壁和明显可运动的“关节”等结构, 在传粉昆虫进入花冠取食花蜜时, 迫使其推动下臂做杠杆状运动, 上臂可育花药将花粉涂抹在昆虫背部从而携带出花粉.鼠尾草属雄蕊结构与功能的分化可能促进了该属物种多样性的形成(Claßen-Bockhoff et al ., 2004 ).根据传粉者类型, 鼠尾草属可分为蜂媒传粉(蜂媒花, melittophilae)和鸟媒传粉(鸟媒花, ornithophily), 蜂媒传粉物种主要分布于欧亚大陆、地中海沿岸、非洲东部及南非、大洋洲以及北美部分地区; 鸟媒传粉物种分布于北美西部至墨西哥、中美洲和南美洲安第斯山脉及东部(魏宇昆等, 2015 ).就雄蕊结构和传粉特征而言, 鼠尾草属中还具有非典型的杠杆状雄蕊结构和传粉模式. ...
群落水平上传粉生态学的研究进展
1
2014
... 在植物与昆虫的相互作用中, 有关花形态塑造和昆虫的适应进化是生态学和植物进化研究中最吸引人的一个领域(黄双全和郭友好, 2000 ; 黄双全, 2007 ; 方强和黄双全, 2014 ).鼠尾草属(Salvia )植物具有花器官两侧对称、二强雄蕊、虫媒异交和杠杆传粉机制等特征, 与传粉者的协同进化程度较高(张勃等, 2010 ), 典型的杠杆传粉过程基于雄蕊药隔的显著伸长、联合具阻挡作用的药隔下壁和明显可运动的“关节”等结构, 在传粉昆虫进入花冠取食花蜜时, 迫使其推动下臂做杠杆状运动, 上臂可育花药将花粉涂抹在昆虫背部从而携带出花粉.鼠尾草属雄蕊结构与功能的分化可能促进了该属物种多样性的形成(Claßen-Bockhoff et al ., 2004 ).根据传粉者类型, 鼠尾草属可分为蜂媒传粉(蜂媒花, melittophilae)和鸟媒传粉(鸟媒花, ornithophily), 蜂媒传粉物种主要分布于欧亚大陆、地中海沿岸、非洲东部及南非、大洋洲以及北美部分地区; 鸟媒传粉物种分布于北美西部至墨西哥、中美洲和南美洲安第斯山脉及东部(魏宇昆等, 2015 ).就雄蕊结构和传粉特征而言, 鼠尾草属中还具有非典型的杠杆状雄蕊结构和传粉模式. ...
玉龙雪山植物物种多样性和群落结构沿海拔梯度的分布格局
1
2006
... 毛地黄鼠尾草研究样地位于云南省丽江市玉龙雪山自然保护区甘海子(27.06° N, 100.26° E), 海拔2990 m, 属青藏高原东南缘, 横断山系南端.玉龙雪山海拔落差及气候垂直差异极大, 最高峰海拔5596 m, 相对高差3846 m (冯建孟等, 2006 ).区内植被类型多样, 植物区系起源古老, 地理成分复杂, 特有种丰富, 分化显著.气候干湿季分明, 5-10月为雨季, 降水量为全年降水量的85%-99% (辛惠娟等, 2013 ), 其中2009年海拔3046 m记录的5-10月降水量为1884.3 mm (辛惠娟等, 2012 ).年平均气温12.79 ℃, 最热月7月平均气温为17.9 ℃, 最冷月1月平均气温为5.9 ℃.植被属高山云南松(Pinus yunnanensis )、丽江云杉(Picea likiangensis )和长苞冷杉(Abies georgei )针叶林.研究样地生境是林缘间开阔草地群落, 毛地黄鼠尾草是其中常见的优势物种. ...
玉龙雪山植物物种多样性和群落结构沿海拔梯度的分布格局
1
2006
... 毛地黄鼠尾草研究样地位于云南省丽江市玉龙雪山自然保护区甘海子(27.06° N, 100.26° E), 海拔2990 m, 属青藏高原东南缘, 横断山系南端.玉龙雪山海拔落差及气候垂直差异极大, 最高峰海拔5596 m, 相对高差3846 m (冯建孟等, 2006 ).区内植被类型多样, 植物区系起源古老, 地理成分复杂, 特有种丰富, 分化显著.气候干湿季分明, 5-10月为雨季, 降水量为全年降水量的85%-99% (辛惠娟等, 2013 ), 其中2009年海拔3046 m记录的5-10月降水量为1884.3 mm (辛惠娟等, 2012 ).年平均气温12.79 ℃, 最热月7月平均气温为17.9 ℃, 最冷月1月平均气温为5.9 ℃.植被属高山云南松(Pinus yunnanensis )、丽江云杉(Picea likiangensis )和长苞冷杉(Abies georgei )针叶林.研究样地生境是林缘间开阔草地群落, 毛地黄鼠尾草是其中常见的优势物种. ...
植物与传粉者相互作用的研究及其意义
1
2007
... 在植物与昆虫的相互作用中, 有关花形态塑造和昆虫的适应进化是生态学和植物进化研究中最吸引人的一个领域(黄双全和郭友好, 2000 ; 黄双全, 2007 ; 方强和黄双全, 2014 ).鼠尾草属(Salvia )植物具有花器官两侧对称、二强雄蕊、虫媒异交和杠杆传粉机制等特征, 与传粉者的协同进化程度较高(张勃等, 2010 ), 典型的杠杆传粉过程基于雄蕊药隔的显著伸长、联合具阻挡作用的药隔下壁和明显可运动的“关节”等结构, 在传粉昆虫进入花冠取食花蜜时, 迫使其推动下臂做杠杆状运动, 上臂可育花药将花粉涂抹在昆虫背部从而携带出花粉.鼠尾草属雄蕊结构与功能的分化可能促进了该属物种多样性的形成(Claßen-Bockhoff et al ., 2004 ).根据传粉者类型, 鼠尾草属可分为蜂媒传粉(蜂媒花, melittophilae)和鸟媒传粉(鸟媒花, ornithophily), 蜂媒传粉物种主要分布于欧亚大陆、地中海沿岸、非洲东部及南非、大洋洲以及北美部分地区; 鸟媒传粉物种分布于北美西部至墨西哥、中美洲和南美洲安第斯山脉及东部(魏宇昆等, 2015 ).就雄蕊结构和传粉特征而言, 鼠尾草属中还具有非典型的杠杆状雄蕊结构和传粉模式. ...
植物与传粉者相互作用的研究及其意义
1
2007
... 在植物与昆虫的相互作用中, 有关花形态塑造和昆虫的适应进化是生态学和植物进化研究中最吸引人的一个领域(黄双全和郭友好, 2000 ; 黄双全, 2007 ; 方强和黄双全, 2014 ).鼠尾草属(Salvia )植物具有花器官两侧对称、二强雄蕊、虫媒异交和杠杆传粉机制等特征, 与传粉者的协同进化程度较高(张勃等, 2010 ), 典型的杠杆传粉过程基于雄蕊药隔的显著伸长、联合具阻挡作用的药隔下壁和明显可运动的“关节”等结构, 在传粉昆虫进入花冠取食花蜜时, 迫使其推动下臂做杠杆状运动, 上臂可育花药将花粉涂抹在昆虫背部从而携带出花粉.鼠尾草属雄蕊结构与功能的分化可能促进了该属物种多样性的形成(Claßen-Bockhoff et al ., 2004 ).根据传粉者类型, 鼠尾草属可分为蜂媒传粉(蜂媒花, melittophilae)和鸟媒传粉(鸟媒花, ornithophily), 蜂媒传粉物种主要分布于欧亚大陆、地中海沿岸、非洲东部及南非、大洋洲以及北美部分地区; 鸟媒传粉物种分布于北美西部至墨西哥、中美洲和南美洲安第斯山脉及东部(魏宇昆等, 2015 ).就雄蕊结构和传粉特征而言, 鼠尾草属中还具有非典型的杠杆状雄蕊结构和传粉模式. ...
传粉生物学的研究进展
1
2000
... 在植物与昆虫的相互作用中, 有关花形态塑造和昆虫的适应进化是生态学和植物进化研究中最吸引人的一个领域(黄双全和郭友好, 2000 ; 黄双全, 2007 ; 方强和黄双全, 2014 ).鼠尾草属(Salvia )植物具有花器官两侧对称、二强雄蕊、虫媒异交和杠杆传粉机制等特征, 与传粉者的协同进化程度较高(张勃等, 2010 ), 典型的杠杆传粉过程基于雄蕊药隔的显著伸长、联合具阻挡作用的药隔下壁和明显可运动的“关节”等结构, 在传粉昆虫进入花冠取食花蜜时, 迫使其推动下臂做杠杆状运动, 上臂可育花药将花粉涂抹在昆虫背部从而携带出花粉.鼠尾草属雄蕊结构与功能的分化可能促进了该属物种多样性的形成(Claßen-Bockhoff et al ., 2004 ).根据传粉者类型, 鼠尾草属可分为蜂媒传粉(蜂媒花, melittophilae)和鸟媒传粉(鸟媒花, ornithophily), 蜂媒传粉物种主要分布于欧亚大陆、地中海沿岸、非洲东部及南非、大洋洲以及北美部分地区; 鸟媒传粉物种分布于北美西部至墨西哥、中美洲和南美洲安第斯山脉及东部(魏宇昆等, 2015 ).就雄蕊结构和传粉特征而言, 鼠尾草属中还具有非典型的杠杆状雄蕊结构和传粉模式. ...
传粉生物学的研究进展
1
2000
... 在植物与昆虫的相互作用中, 有关花形态塑造和昆虫的适应进化是生态学和植物进化研究中最吸引人的一个领域(黄双全和郭友好, 2000 ; 黄双全, 2007 ; 方强和黄双全, 2014 ).鼠尾草属(Salvia )植物具有花器官两侧对称、二强雄蕊、虫媒异交和杠杆传粉机制等特征, 与传粉者的协同进化程度较高(张勃等, 2010 ), 典型的杠杆传粉过程基于雄蕊药隔的显著伸长、联合具阻挡作用的药隔下壁和明显可运动的“关节”等结构, 在传粉昆虫进入花冠取食花蜜时, 迫使其推动下臂做杠杆状运动, 上臂可育花药将花粉涂抹在昆虫背部从而携带出花粉.鼠尾草属雄蕊结构与功能的分化可能促进了该属物种多样性的形成(Claßen-Bockhoff et al ., 2004 ).根据传粉者类型, 鼠尾草属可分为蜂媒传粉(蜂媒花, melittophilae)和鸟媒传粉(鸟媒花, ornithophily), 蜂媒传粉物种主要分布于欧亚大陆、地中海沿岸、非洲东部及南非、大洋洲以及北美部分地区; 鸟媒传粉物种分布于北美西部至墨西哥、中美洲和南美洲安第斯山脉及东部(魏宇昆等, 2015 ).就雄蕊结构和传粉特征而言, 鼠尾草属中还具有非典型的杠杆状雄蕊结构和传粉模式. ...
鼠尾草属东亚分支的传粉模式
3
2014
... 黄艳波等(2014)根据花器官结构和花粉接触传粉昆虫的部位, 将中国的鼠尾草属物种划分为3种传粉模式类型: 短药隔杠杆、长药隔杠杆和退化杠杆.3个类群大致形成以短药隔杠杆物种集中分布于中国西南, 长药隔杠杆物种集中于华中, 退化杠杆类型以华东为核心的分布格局.对鼠尾草属部分物种的杠杆传粉机制了解较多, 如短药隔杠杆类型的圆苞鼠尾草(S. cyclostegia )和长药隔杠杆类型丹参(S. miltiorrhiza )均借助传粉昆虫推动雄蕊做杠杆状运动, 完成花粉分发和授粉作用(张勃等, 2012 ; 黄艳波等, 2014 ).在中国, 鼠尾草亚属(subgenus Allagospadonopsis )的16个物种具有退化的杠杆结构, 推测它们具有非典型的杠杆传粉特征(黄艳波等, 2014 ).鼠尾草属雄蕊的分化是物种形成和多样性产生的关键因素(Claßen-Bockhoff et al ., 2003 ), 这种适应辐射的物种形成事件也是各种环境选择压力综合作用的结果.尽管对西南分布的短药隔类型鼠尾草的传粉结构、进化响应和表型选择等已有所了解(Zhang et al ., 2011 ; 张勃等, 2012 ; Zhang & Li, 2014 ), 但相比鼠尾草属多样性的传粉结构和传粉过程, 对其中一种类型的认识显然不足以揭示鼠尾草属面对的传粉者选择压力和进化方向.更何况, 至今对该属植物的传粉生态学和繁育系统仍然了解不多(Claßen-Bockhoff et al ., 2003 ). ...
... )的16个物种具有退化的杠杆结构, 推测它们具有非典型的杠杆传粉特征(黄艳波等, 2014 ).鼠尾草属雄蕊的分化是物种形成和多样性产生的关键因素(Claßen-Bockhoff et al ., 2003 ), 这种适应辐射的物种形成事件也是各种环境选择压力综合作用的结果.尽管对西南分布的短药隔类型鼠尾草的传粉结构、进化响应和表型选择等已有所了解(Zhang et al ., 2011 ; 张勃等, 2012 ; Zhang & Li, 2014 ), 但相比鼠尾草属多样性的传粉结构和传粉过程, 对其中一种类型的认识显然不足以揭示鼠尾草属面对的传粉者选择压力和进化方向.更何况, 至今对该属植物的传粉生态学和繁育系统仍然了解不多(Claßen-Bockhoff et al ., 2003 ). ...
... 在宏观形态学方面, 鼠尾草属杠杆呈现出雄蕊药隔逐渐伸长、药隔下臂逐渐退化形成附属结构, 直至最后完全丧失功能这样的进化历程(Claßen- Bockhoff et al ., 2003 ; 黄艳波等, 2014 ).毛地黄鼠尾草与舌瓣鼠尾草恰似占据此进化历程的两极.Li等(2013)将中国产43种鼠尾草属植物构建了系统进化树, 尽管涵盖了舌瓣鼠尾草所在的鼠尾草亚属和毛地黄鼠尾草所在的弧隔鼠尾草亚属, 但并没有采用毛地黄鼠尾草这个材料.仅从该文中两个种所在的亚属来看, 舌瓣鼠尾草所在的鼠尾草亚属首先与丹参所在的荔枝草亚属(subgenus Sclarea )聚为一支, 再与毛地黄鼠尾草所在的弧隔鼠尾草亚属聚为一支, 可以间接判断毛地黄鼠尾草所在的弧隔鼠尾草亚属可能更古老原始, 而舌瓣鼠尾草的进化时间更短, 成种时间较晚.由于该属可能处在迅速的辐射成种阶段, 仅使用几个叶绿体和核基因片段来研究鼠尾草属的种间系统关系, 信息量似乎远远不足, 未来需借助更多的标记或二代测序技术, 结合物种分布格局和谱系地理研究(王琦等, 2015 ), 从而更深入地理解不同传粉模式的系统发育关系. ...
鼠尾草属东亚分支的传粉模式
3
2014
... 黄艳波等(2014)根据花器官结构和花粉接触传粉昆虫的部位, 将中国的鼠尾草属物种划分为3种传粉模式类型: 短药隔杠杆、长药隔杠杆和退化杠杆.3个类群大致形成以短药隔杠杆物种集中分布于中国西南, 长药隔杠杆物种集中于华中, 退化杠杆类型以华东为核心的分布格局.对鼠尾草属部分物种的杠杆传粉机制了解较多, 如短药隔杠杆类型的圆苞鼠尾草(S. cyclostegia )和长药隔杠杆类型丹参(S. miltiorrhiza )均借助传粉昆虫推动雄蕊做杠杆状运动, 完成花粉分发和授粉作用(张勃等, 2012 ; 黄艳波等, 2014 ).在中国, 鼠尾草亚属(subgenus Allagospadonopsis )的16个物种具有退化的杠杆结构, 推测它们具有非典型的杠杆传粉特征(黄艳波等, 2014 ).鼠尾草属雄蕊的分化是物种形成和多样性产生的关键因素(Claßen-Bockhoff et al ., 2003 ), 这种适应辐射的物种形成事件也是各种环境选择压力综合作用的结果.尽管对西南分布的短药隔类型鼠尾草的传粉结构、进化响应和表型选择等已有所了解(Zhang et al ., 2011 ; 张勃等, 2012 ; Zhang & Li, 2014 ), 但相比鼠尾草属多样性的传粉结构和传粉过程, 对其中一种类型的认识显然不足以揭示鼠尾草属面对的传粉者选择压力和进化方向.更何况, 至今对该属植物的传粉生态学和繁育系统仍然了解不多(Claßen-Bockhoff et al ., 2003 ). ...
... )的16个物种具有退化的杠杆结构, 推测它们具有非典型的杠杆传粉特征(黄艳波等, 2014 ).鼠尾草属雄蕊的分化是物种形成和多样性产生的关键因素(Claßen-Bockhoff et al ., 2003 ), 这种适应辐射的物种形成事件也是各种环境选择压力综合作用的结果.尽管对西南分布的短药隔类型鼠尾草的传粉结构、进化响应和表型选择等已有所了解(Zhang et al ., 2011 ; 张勃等, 2012 ; Zhang & Li, 2014 ), 但相比鼠尾草属多样性的传粉结构和传粉过程, 对其中一种类型的认识显然不足以揭示鼠尾草属面对的传粉者选择压力和进化方向.更何况, 至今对该属植物的传粉生态学和繁育系统仍然了解不多(Claßen-Bockhoff et al ., 2003 ). ...
... 在宏观形态学方面, 鼠尾草属杠杆呈现出雄蕊药隔逐渐伸长、药隔下臂逐渐退化形成附属结构, 直至最后完全丧失功能这样的进化历程(Claßen- Bockhoff et al ., 2003 ; 黄艳波等, 2014 ).毛地黄鼠尾草与舌瓣鼠尾草恰似占据此进化历程的两极.Li等(2013)将中国产43种鼠尾草属植物构建了系统进化树, 尽管涵盖了舌瓣鼠尾草所在的鼠尾草亚属和毛地黄鼠尾草所在的弧隔鼠尾草亚属, 但并没有采用毛地黄鼠尾草这个材料.仅从该文中两个种所在的亚属来看, 舌瓣鼠尾草所在的鼠尾草亚属首先与丹参所在的荔枝草亚属(subgenus Sclarea )聚为一支, 再与毛地黄鼠尾草所在的弧隔鼠尾草亚属聚为一支, 可以间接判断毛地黄鼠尾草所在的弧隔鼠尾草亚属可能更古老原始, 而舌瓣鼠尾草的进化时间更短, 成种时间较晚.由于该属可能处在迅速的辐射成种阶段, 仅使用几个叶绿体和核基因片段来研究鼠尾草属的种间系统关系, 信息量似乎远远不足, 未来需借助更多的标记或二代测序技术, 结合物种分布格局和谱系地理研究(王琦等, 2015 ), 从而更深入地理解不同传粉模式的系统发育关系. ...
Phylogenetic relationships of Salvia (Lamiaceae) in China: Evidence from DNA sequence datasets.
2013
Pollinator attendance and reproductive success in Cistus libanotis L. (Cistaceae).
1
2001
... 参考Talavera等(2001) 以及龚燕兵和黄双全(2007)的方法, 确定并计算: (1)有效传粉昆虫(effective pollinator), 可携带花粉并完成授粉的昆虫; (2)相对频次(relative frequency, F ), 在固定样方内, 某一种访花昆虫的出现次数占所有访花昆虫出现总次数的相对比例; (3)昆虫传粉的日活动变化规律, 用固定样方内每小时的昆虫访花总次数表示; (4)昆虫单花访问时间(T ), 即访花者从登陆花冠口到访花结束后飞离花冠的时间; (5)活跃度(AR ), 以单只昆虫每分钟的访问花朵数量表示, 代表了此种昆虫对某一种植物花的偏好程度和传粉潜力; (6)访花频率, 用指数IVR = F × AR 表示, 访花频率同时考虑了昆虫的活跃度及其出现的相对频次; (7)传粉回报, 传粉者为植物传粉获得的物质补偿, 一般是花粉或花蜜.毛地黄鼠尾草访花昆虫的访花特征参考张勃等(2012)在甘海子样地的观测结果. ...
中国弧隔鼠尾草亚属(唇形科)的分布格局
1
2015
... 在宏观形态学方面, 鼠尾草属杠杆呈现出雄蕊药隔逐渐伸长、药隔下臂逐渐退化形成附属结构, 直至最后完全丧失功能这样的进化历程(Claßen- Bockhoff et al ., 2003 ; 黄艳波等, 2014 ).毛地黄鼠尾草与舌瓣鼠尾草恰似占据此进化历程的两极.Li等(2013)将中国产43种鼠尾草属植物构建了系统进化树, 尽管涵盖了舌瓣鼠尾草所在的鼠尾草亚属和毛地黄鼠尾草所在的弧隔鼠尾草亚属, 但并没有采用毛地黄鼠尾草这个材料.仅从该文中两个种所在的亚属来看, 舌瓣鼠尾草所在的鼠尾草亚属首先与丹参所在的荔枝草亚属(subgenus Sclarea )聚为一支, 再与毛地黄鼠尾草所在的弧隔鼠尾草亚属聚为一支, 可以间接判断毛地黄鼠尾草所在的弧隔鼠尾草亚属可能更古老原始, 而舌瓣鼠尾草的进化时间更短, 成种时间较晚.由于该属可能处在迅速的辐射成种阶段, 仅使用几个叶绿体和核基因片段来研究鼠尾草属的种间系统关系, 信息量似乎远远不足, 未来需借助更多的标记或二代测序技术, 结合物种分布格局和谱系地理研究(王琦等, 2015 ), 从而更深入地理解不同传粉模式的系统发育关系. ...
中国弧隔鼠尾草亚属(唇形科)的分布格局
1
2015
... 在宏观形态学方面, 鼠尾草属杠杆呈现出雄蕊药隔逐渐伸长、药隔下臂逐渐退化形成附属结构, 直至最后完全丧失功能这样的进化历程(Claßen- Bockhoff et al ., 2003 ; 黄艳波等, 2014 ).毛地黄鼠尾草与舌瓣鼠尾草恰似占据此进化历程的两极.Li等(2013)将中国产43种鼠尾草属植物构建了系统进化树, 尽管涵盖了舌瓣鼠尾草所在的鼠尾草亚属和毛地黄鼠尾草所在的弧隔鼠尾草亚属, 但并没有采用毛地黄鼠尾草这个材料.仅从该文中两个种所在的亚属来看, 舌瓣鼠尾草所在的鼠尾草亚属首先与丹参所在的荔枝草亚属(subgenus Sclarea )聚为一支, 再与毛地黄鼠尾草所在的弧隔鼠尾草亚属聚为一支, 可以间接判断毛地黄鼠尾草所在的弧隔鼠尾草亚属可能更古老原始, 而舌瓣鼠尾草的进化时间更短, 成种时间较晚.由于该属可能处在迅速的辐射成种阶段, 仅使用几个叶绿体和核基因片段来研究鼠尾草属的种间系统关系, 信息量似乎远远不足, 未来需借助更多的标记或二代测序技术, 结合物种分布格局和谱系地理研究(王琦等, 2015 ), 从而更深入地理解不同传粉模式的系统发育关系. ...
唇形科鼠尾草属的物种多样性与分布
1
2015
... 在植物与昆虫的相互作用中, 有关花形态塑造和昆虫的适应进化是生态学和植物进化研究中最吸引人的一个领域(黄双全和郭友好, 2000 ; 黄双全, 2007 ; 方强和黄双全, 2014 ).鼠尾草属(Salvia )植物具有花器官两侧对称、二强雄蕊、虫媒异交和杠杆传粉机制等特征, 与传粉者的协同进化程度较高(张勃等, 2010 ), 典型的杠杆传粉过程基于雄蕊药隔的显著伸长、联合具阻挡作用的药隔下壁和明显可运动的“关节”等结构, 在传粉昆虫进入花冠取食花蜜时, 迫使其推动下臂做杠杆状运动, 上臂可育花药将花粉涂抹在昆虫背部从而携带出花粉.鼠尾草属雄蕊结构与功能的分化可能促进了该属物种多样性的形成(Claßen-Bockhoff et al ., 2004 ).根据传粉者类型, 鼠尾草属可分为蜂媒传粉(蜂媒花, melittophilae)和鸟媒传粉(鸟媒花, ornithophily), 蜂媒传粉物种主要分布于欧亚大陆、地中海沿岸、非洲东部及南非、大洋洲以及北美部分地区; 鸟媒传粉物种分布于北美西部至墨西哥、中美洲和南美洲安第斯山脉及东部(魏宇昆等, 2015 ).就雄蕊结构和传粉特征而言, 鼠尾草属中还具有非典型的杠杆状雄蕊结构和传粉模式. ...
唇形科鼠尾草属的物种多样性与分布
1
2015
... 在植物与昆虫的相互作用中, 有关花形态塑造和昆虫的适应进化是生态学和植物进化研究中最吸引人的一个领域(黄双全和郭友好, 2000 ; 黄双全, 2007 ; 方强和黄双全, 2014 ).鼠尾草属(Salvia )植物具有花器官两侧对称、二强雄蕊、虫媒异交和杠杆传粉机制等特征, 与传粉者的协同进化程度较高(张勃等, 2010 ), 典型的杠杆传粉过程基于雄蕊药隔的显著伸长、联合具阻挡作用的药隔下壁和明显可运动的“关节”等结构, 在传粉昆虫进入花冠取食花蜜时, 迫使其推动下臂做杠杆状运动, 上臂可育花药将花粉涂抹在昆虫背部从而携带出花粉.鼠尾草属雄蕊结构与功能的分化可能促进了该属物种多样性的形成(Claßen-Bockhoff et al ., 2004 ).根据传粉者类型, 鼠尾草属可分为蜂媒传粉(蜂媒花, melittophilae)和鸟媒传粉(鸟媒花, ornithophily), 蜂媒传粉物种主要分布于欧亚大陆、地中海沿岸、非洲东部及南非、大洋洲以及北美部分地区; 鸟媒传粉物种分布于北美西部至墨西哥、中美洲和南美洲安第斯山脉及东部(魏宇昆等, 2015 ).就雄蕊结构和传粉特征而言, 鼠尾草属中还具有非典型的杠杆状雄蕊结构和传粉模式. ...
玉龙雪山东坡气温和降水梯度年内变化特征
1
2012
... 毛地黄鼠尾草研究样地位于云南省丽江市玉龙雪山自然保护区甘海子(27.06° N, 100.26° E), 海拔2990 m, 属青藏高原东南缘, 横断山系南端.玉龙雪山海拔落差及气候垂直差异极大, 最高峰海拔5596 m, 相对高差3846 m (冯建孟等, 2006 ).区内植被类型多样, 植物区系起源古老, 地理成分复杂, 特有种丰富, 分化显著.气候干湿季分明, 5-10月为雨季, 降水量为全年降水量的85%-99% (辛惠娟等, 2013 ), 其中2009年海拔3046 m记录的5-10月降水量为1884.3 mm (辛惠娟等, 2012 ).年平均气温12.79 ℃, 最热月7月平均气温为17.9 ℃, 最冷月1月平均气温为5.9 ℃.植被属高山云南松(Pinus yunnanensis )、丽江云杉(Picea likiangensis )和长苞冷杉(Abies georgei )针叶林.研究样地生境是林缘间开阔草地群落, 毛地黄鼠尾草是其中常见的优势物种. ...
玉龙雪山东坡气温和降水梯度年内变化特征
1
2012
... 毛地黄鼠尾草研究样地位于云南省丽江市玉龙雪山自然保护区甘海子(27.06° N, 100.26° E), 海拔2990 m, 属青藏高原东南缘, 横断山系南端.玉龙雪山海拔落差及气候垂直差异极大, 最高峰海拔5596 m, 相对高差3846 m (冯建孟等, 2006 ).区内植被类型多样, 植物区系起源古老, 地理成分复杂, 特有种丰富, 分化显著.气候干湿季分明, 5-10月为雨季, 降水量为全年降水量的85%-99% (辛惠娟等, 2013 ), 其中2009年海拔3046 m记录的5-10月降水量为1884.3 mm (辛惠娟等, 2012 ).年平均气温12.79 ℃, 最热月7月平均气温为17.9 ℃, 最冷月1月平均气温为5.9 ℃.植被属高山云南松(Pinus yunnanensis )、丽江云杉(Picea likiangensis )和长苞冷杉(Abies georgei )针叶林.研究样地生境是林缘间开阔草地群落, 毛地黄鼠尾草是其中常见的优势物种. ...
青藏高原东南缘丽江玉龙雪山气候变化特征及其对冰川变化的影响
1
2013
... 毛地黄鼠尾草研究样地位于云南省丽江市玉龙雪山自然保护区甘海子(27.06° N, 100.26° E), 海拔2990 m, 属青藏高原东南缘, 横断山系南端.玉龙雪山海拔落差及气候垂直差异极大, 最高峰海拔5596 m, 相对高差3846 m (冯建孟等, 2006 ).区内植被类型多样, 植物区系起源古老, 地理成分复杂, 特有种丰富, 分化显著.气候干湿季分明, 5-10月为雨季, 降水量为全年降水量的85%-99% (辛惠娟等, 2013 ), 其中2009年海拔3046 m记录的5-10月降水量为1884.3 mm (辛惠娟等, 2012 ).年平均气温12.79 ℃, 最热月7月平均气温为17.9 ℃, 最冷月1月平均气温为5.9 ℃.植被属高山云南松(Pinus yunnanensis )、丽江云杉(Picea likiangensis )和长苞冷杉(Abies georgei )针叶林.研究样地生境是林缘间开阔草地群落, 毛地黄鼠尾草是其中常见的优势物种. ...
青藏高原东南缘丽江玉龙雪山气候变化特征及其对冰川变化的影响
1
2013
... 毛地黄鼠尾草研究样地位于云南省丽江市玉龙雪山自然保护区甘海子(27.06° N, 100.26° E), 海拔2990 m, 属青藏高原东南缘, 横断山系南端.玉龙雪山海拔落差及气候垂直差异极大, 最高峰海拔5596 m, 相对高差3846 m (冯建孟等, 2006 ).区内植被类型多样, 植物区系起源古老, 地理成分复杂, 特有种丰富, 分化显著.气候干湿季分明, 5-10月为雨季, 降水量为全年降水量的85%-99% (辛惠娟等, 2013 ), 其中2009年海拔3046 m记录的5-10月降水量为1884.3 mm (辛惠娟等, 2012 ).年平均气温12.79 ℃, 最热月7月平均气温为17.9 ℃, 最冷月1月平均气温为5.9 ℃.植被属高山云南松(Pinus yunnanensis )、丽江云杉(Picea likiangensis )和长苞冷杉(Abies georgei )针叶林.研究样地生境是林缘间开阔草地群落, 毛地黄鼠尾草是其中常见的优势物种. ...
3
1992
... 舌瓣鼠尾草研究地点位于天目山国家级自然保护区(30.39° N, 119.48° E), 海拔677 m.天目山保护区属北亚热带气候, 季风强盛、四季分明、气候温和、雨水丰沛、光照适宜.年平均气温8.9-15.8 ℃, 最冷月气温为2.6-3.4 ℃, 最热月气温为19.9-28.1 ℃, ≥ 10 ℃积温2500-5100 ℃, 年降水量1390-1870 mm, 全年无霜期209-235天, 年太阳总辐射3770- 4460 MJ·m-2 (杨逢春, 1992 ).天目山植物区系的温带、亚热带东亚区系成分特征显著, 在地理上和发生上与东亚日本、北美、东南亚等有着不同程度的联系(杨逢春, 1992 ).研究样地植被属于常绿阔叶林(杨逢春, 1992 ), 其群落组成主要为菊科一年蓬(Erigeron annuus )、蒲儿根(Sinosenecio oldhamianus ), 报春花科过路黄(Lysimachia christinae ), 荨麻科青叶苎麻(Boehmeria nivea var. tenacissima ), 山茶科茶(Camellia sinensis ), 禾本科阔叶箬竹(Indocalamus latifolius )、早竹(Phyllostachys violascens )、求米草(Oplismenus undulatifolius ), 唇形科紫花香薷(Elsholtzia argyi ), 菝葜科菝葜(Smilax china ), 杉科杉木(Cunninghamia lanceolata ), 胡桃科山核桃(Carya cathayensis ).其中优势种主要是青叶苎麻、早竹和山核桃等. ...
... ).天目山植物区系的温带、亚热带东亚区系成分特征显著, 在地理上和发生上与东亚日本、北美、东南亚等有着不同程度的联系(杨逢春, 1992 ).研究样地植被属于常绿阔叶林(杨逢春, 1992 ), 其群落组成主要为菊科一年蓬(Erigeron annuus )、蒲儿根(Sinosenecio oldhamianus ), 报春花科过路黄(Lysimachia christinae ), 荨麻科青叶苎麻(Boehmeria nivea var. tenacissima ), 山茶科茶(Camellia sinensis ), 禾本科阔叶箬竹(Indocalamus latifolius )、早竹(Phyllostachys violascens )、求米草(Oplismenus undulatifolius ), 唇形科紫花香薷(Elsholtzia argyi ), 菝葜科菝葜(Smilax china ), 杉科杉木(Cunninghamia lanceolata ), 胡桃科山核桃(Carya cathayensis ).其中优势种主要是青叶苎麻、早竹和山核桃等. ...
... ).研究样地植被属于常绿阔叶林(杨逢春, 1992 ), 其群落组成主要为菊科一年蓬(Erigeron annuus )、蒲儿根(Sinosenecio oldhamianus ), 报春花科过路黄(Lysimachia christinae ), 荨麻科青叶苎麻(Boehmeria nivea var. tenacissima ), 山茶科茶(Camellia sinensis ), 禾本科阔叶箬竹(Indocalamus latifolius )、早竹(Phyllostachys violascens )、求米草(Oplismenus undulatifolius ), 唇形科紫花香薷(Elsholtzia argyi ), 菝葜科菝葜(Smilax china ), 杉科杉木(Cunninghamia lanceolata ), 胡桃科山核桃(Carya cathayensis ).其中优势种主要是青叶苎麻、早竹和山核桃等. ...
3
1992
... 舌瓣鼠尾草研究地点位于天目山国家级自然保护区(30.39° N, 119.48° E), 海拔677 m.天目山保护区属北亚热带气候, 季风强盛、四季分明、气候温和、雨水丰沛、光照适宜.年平均气温8.9-15.8 ℃, 最冷月气温为2.6-3.4 ℃, 最热月气温为19.9-28.1 ℃, ≥ 10 ℃积温2500-5100 ℃, 年降水量1390-1870 mm, 全年无霜期209-235天, 年太阳总辐射3770- 4460 MJ·m-2 (杨逢春, 1992 ).天目山植物区系的温带、亚热带东亚区系成分特征显著, 在地理上和发生上与东亚日本、北美、东南亚等有着不同程度的联系(杨逢春, 1992 ).研究样地植被属于常绿阔叶林(杨逢春, 1992 ), 其群落组成主要为菊科一年蓬(Erigeron annuus )、蒲儿根(Sinosenecio oldhamianus ), 报春花科过路黄(Lysimachia christinae ), 荨麻科青叶苎麻(Boehmeria nivea var. tenacissima ), 山茶科茶(Camellia sinensis ), 禾本科阔叶箬竹(Indocalamus latifolius )、早竹(Phyllostachys violascens )、求米草(Oplismenus undulatifolius ), 唇形科紫花香薷(Elsholtzia argyi ), 菝葜科菝葜(Smilax china ), 杉科杉木(Cunninghamia lanceolata ), 胡桃科山核桃(Carya cathayensis ).其中优势种主要是青叶苎麻、早竹和山核桃等. ...
... ).天目山植物区系的温带、亚热带东亚区系成分特征显著, 在地理上和发生上与东亚日本、北美、东南亚等有着不同程度的联系(杨逢春, 1992 ).研究样地植被属于常绿阔叶林(杨逢春, 1992 ), 其群落组成主要为菊科一年蓬(Erigeron annuus )、蒲儿根(Sinosenecio oldhamianus ), 报春花科过路黄(Lysimachia christinae ), 荨麻科青叶苎麻(Boehmeria nivea var. tenacissima ), 山茶科茶(Camellia sinensis ), 禾本科阔叶箬竹(Indocalamus latifolius )、早竹(Phyllostachys violascens )、求米草(Oplismenus undulatifolius ), 唇形科紫花香薷(Elsholtzia argyi ), 菝葜科菝葜(Smilax china ), 杉科杉木(Cunninghamia lanceolata ), 胡桃科山核桃(Carya cathayensis ).其中优势种主要是青叶苎麻、早竹和山核桃等. ...
... ).研究样地植被属于常绿阔叶林(杨逢春, 1992 ), 其群落组成主要为菊科一年蓬(Erigeron annuus )、蒲儿根(Sinosenecio oldhamianus ), 报春花科过路黄(Lysimachia christinae ), 荨麻科青叶苎麻(Boehmeria nivea var. tenacissima ), 山茶科茶(Camellia sinensis ), 禾本科阔叶箬竹(Indocalamus latifolius )、早竹(Phyllostachys violascens )、求米草(Oplismenus undulatifolius ), 唇形科紫花香薷(Elsholtzia argyi ), 菝葜科菝葜(Smilax china ), 杉科杉木(Cunninghamia lanceolata ), 胡桃科山核桃(Carya cathayensis ).其中优势种主要是青叶苎麻、早竹和山核桃等. ...
Functional implications of the staminal lever mechanism in Salvia cyclostegia (Lamiaceae).
2
2011
... 黄艳波等(2014)根据花器官结构和花粉接触传粉昆虫的部位, 将中国的鼠尾草属物种划分为3种传粉模式类型: 短药隔杠杆、长药隔杠杆和退化杠杆.3个类群大致形成以短药隔杠杆物种集中分布于中国西南, 长药隔杠杆物种集中于华中, 退化杠杆类型以华东为核心的分布格局.对鼠尾草属部分物种的杠杆传粉机制了解较多, 如短药隔杠杆类型的圆苞鼠尾草(S. cyclostegia )和长药隔杠杆类型丹参(S. miltiorrhiza )均借助传粉昆虫推动雄蕊做杠杆状运动, 完成花粉分发和授粉作用(张勃等, 2012 ; 黄艳波等, 2014 ).在中国, 鼠尾草亚属(subgenus Allagospadonopsis )的16个物种具有退化的杠杆结构, 推测它们具有非典型的杠杆传粉特征(黄艳波等, 2014 ).鼠尾草属雄蕊的分化是物种形成和多样性产生的关键因素(Claßen-Bockhoff et al ., 2003 ), 这种适应辐射的物种形成事件也是各种环境选择压力综合作用的结果.尽管对西南分布的短药隔类型鼠尾草的传粉结构、进化响应和表型选择等已有所了解(Zhang et al ., 2011 ; 张勃等, 2012 ; Zhang & Li, 2014 ), 但相比鼠尾草属多样性的传粉结构和传粉过程, 对其中一种类型的认识显然不足以揭示鼠尾草属面对的传粉者选择压力和进化方向.更何况, 至今对该属植物的传粉生态学和繁育系统仍然了解不多(Claßen-Bockhoff et al ., 2003 ). ...
... 舌瓣鼠尾草有专一性的传粉昆虫三条熊蜂, 毛地黄鼠尾草则有3种.专一性的提高是否暗示植物与昆虫的相互关系更紧密, 协同进化程度更高?舌瓣鼠尾草传粉者的高度专一性可能还与所在地的昆虫种类组成相关, 究竟是退化杠杆结构过滤了其他传粉者, 还是传粉者的改变导致杠杆功能丧失? 都需要进一步深入研究.另一方面, 专一性的传粉昆虫面临的风险较大, 一旦失去传粉者, 就有无法延续生存的可能.相反, 具有多个传粉者的物种, 这样的风险则大大降低.目前对鼠尾草属短药隔杠杆机制的作用以及传粉者响应的生态学意义等方面研究较充分(Zhang et al ., 2011 ; 张勃等, 2012 ; Zhang & Li, 2014 ), 而其他类型杠杆的研究还很少, 对该属多种类型杠杆机制的阐明, 无疑会对揭示其起源和进化方向以及该物种多样性的产生、共存和维持机制提供更多证据. ...
Phenotypic selection on the staminal lever mechanism in Salvia digitaloides (Labiaceae).
2
2014
... 黄艳波等(2014)根据花器官结构和花粉接触传粉昆虫的部位, 将中国的鼠尾草属物种划分为3种传粉模式类型: 短药隔杠杆、长药隔杠杆和退化杠杆.3个类群大致形成以短药隔杠杆物种集中分布于中国西南, 长药隔杠杆物种集中于华中, 退化杠杆类型以华东为核心的分布格局.对鼠尾草属部分物种的杠杆传粉机制了解较多, 如短药隔杠杆类型的圆苞鼠尾草(S. cyclostegia )和长药隔杠杆类型丹参(S. miltiorrhiza )均借助传粉昆虫推动雄蕊做杠杆状运动, 完成花粉分发和授粉作用(张勃等, 2012 ; 黄艳波等, 2014 ).在中国, 鼠尾草亚属(subgenus Allagospadonopsis )的16个物种具有退化的杠杆结构, 推测它们具有非典型的杠杆传粉特征(黄艳波等, 2014 ).鼠尾草属雄蕊的分化是物种形成和多样性产生的关键因素(Claßen-Bockhoff et al ., 2003 ), 这种适应辐射的物种形成事件也是各种环境选择压力综合作用的结果.尽管对西南分布的短药隔类型鼠尾草的传粉结构、进化响应和表型选择等已有所了解(Zhang et al ., 2011 ; 张勃等, 2012 ; Zhang & Li, 2014 ), 但相比鼠尾草属多样性的传粉结构和传粉过程, 对其中一种类型的认识显然不足以揭示鼠尾草属面对的传粉者选择压力和进化方向.更何况, 至今对该属植物的传粉生态学和繁育系统仍然了解不多(Claßen-Bockhoff et al ., 2003 ). ...
... 舌瓣鼠尾草有专一性的传粉昆虫三条熊蜂, 毛地黄鼠尾草则有3种.专一性的提高是否暗示植物与昆虫的相互关系更紧密, 协同进化程度更高?舌瓣鼠尾草传粉者的高度专一性可能还与所在地的昆虫种类组成相关, 究竟是退化杠杆结构过滤了其他传粉者, 还是传粉者的改变导致杠杆功能丧失? 都需要进一步深入研究.另一方面, 专一性的传粉昆虫面临的风险较大, 一旦失去传粉者, 就有无法延续生存的可能.相反, 具有多个传粉者的物种, 这样的风险则大大降低.目前对鼠尾草属短药隔杠杆机制的作用以及传粉者响应的生态学意义等方面研究较充分(Zhang et al ., 2011 ; 张勃等, 2012 ; Zhang & Li, 2014 ), 而其他类型杠杆的研究还很少, 对该属多种类型杠杆机制的阐明, 无疑会对揭示其起源和进化方向以及该物种多样性的产生、共存和维持机制提供更多证据. ...
鼠尾草属不同物种的雄蕊杠杆机制对传粉者空间变异的进化响应
8
2012
... 黄艳波等(2014)根据花器官结构和花粉接触传粉昆虫的部位, 将中国的鼠尾草属物种划分为3种传粉模式类型: 短药隔杠杆、长药隔杠杆和退化杠杆.3个类群大致形成以短药隔杠杆物种集中分布于中国西南, 长药隔杠杆物种集中于华中, 退化杠杆类型以华东为核心的分布格局.对鼠尾草属部分物种的杠杆传粉机制了解较多, 如短药隔杠杆类型的圆苞鼠尾草(S. cyclostegia )和长药隔杠杆类型丹参(S. miltiorrhiza )均借助传粉昆虫推动雄蕊做杠杆状运动, 完成花粉分发和授粉作用(张勃等, 2012 ; 黄艳波等, 2014 ).在中国, 鼠尾草亚属(subgenus Allagospadonopsis )的16个物种具有退化的杠杆结构, 推测它们具有非典型的杠杆传粉特征(黄艳波等, 2014 ).鼠尾草属雄蕊的分化是物种形成和多样性产生的关键因素(Claßen-Bockhoff et al ., 2003 ), 这种适应辐射的物种形成事件也是各种环境选择压力综合作用的结果.尽管对西南分布的短药隔类型鼠尾草的传粉结构、进化响应和表型选择等已有所了解(Zhang et al ., 2011 ; 张勃等, 2012 ; Zhang & Li, 2014 ), 但相比鼠尾草属多样性的传粉结构和传粉过程, 对其中一种类型的认识显然不足以揭示鼠尾草属面对的传粉者选择压力和进化方向.更何况, 至今对该属植物的传粉生态学和繁育系统仍然了解不多(Claßen-Bockhoff et al ., 2003 ). ...
... ; 张勃等, 2012 ; Zhang & Li, 2014 ), 但相比鼠尾草属多样性的传粉结构和传粉过程, 对其中一种类型的认识显然不足以揭示鼠尾草属面对的传粉者选择压力和进化方向.更何况, 至今对该属植物的传粉生态学和繁育系统仍然了解不多(Claßen-Bockhoff et al ., 2003 ). ...
... 采集植物种群中出现的访花昆虫, 首先用乙酸乙酯将昆虫毒死, 用镊子辅助保持自然伸展状态, 用电子游标卡尺进行形态特征测量, 包括昆虫体长、胸宽、胸厚和喙长4个指标.喙长测量时用镊子将其拉出至自然伸展状态, 测量完成后用昆虫针将其固定, 制成标本带回实验室进行拍照鉴定.对舌瓣鼠尾草采集样地内出现的所有访花昆虫进行鉴定测量, 毛地黄鼠尾草的传粉昆虫参考张勃等(2012) 在甘海子的测量结果. ...
... 在舌瓣鼠尾草的样方内记录观察到3种访花昆虫, 分别是蜜蜂科熊蜂属(Bombus )的三条熊蜂(B. trifasciatus ), 隧蜂科(Halictidae)和天蛾科(Sphing- idae)昆虫(图1 ).因天蛾科昆虫数量少、飞行速度快, 未能捕捉到活体, 故只测量了三条熊蜂和隧蜂科昆虫.从体型大小看, 3种访花昆虫形态特征差异明显(表2 ), 并且访花行为也完全不同.如图4 所示, 三条熊蜂体型较大, 只有一种访花姿态, 即直接飞至冠口, 依靠其前、中足抓住花冠前部保持平衡, 头部插入冠筒, 利用其长喙吸食花蜜, 此时前额紧贴雄蕊上臂的花粉囊, 将花粉带出, 完整的访花时间为(1.54 ± 0.60) s.当它再次进入另一朵具成熟雌蕊的花冠时, 便可将其头部的花粉授予柱头, 完成授粉.隧蜂科昆虫体型小, 通常飞落至花冠口再缓慢爬进冠筒, 自由地取食花粉, 并未取食花蜜, 而此时雌蕊尚未成熟.其单花访花时间很长、变幅较大, 平均为(73.9 ± 55.6) s.另一种天蛾科昆虫的飞行速率极快, 单花访花时间短, 平均只有(0.67 ± 0.23) s, 因其喙很长, 取食花蜜时没有触碰到花器官任何部位(图1 L).在甘海子样地, 毛地黄鼠尾草的访花昆虫有寄生性拟熊蜂(Psithyrus sp.)、小雅熊蜂(Bombus lepidus )和Bombus infrequens .三种访花昆虫主要进行背部传粉, 小雅熊蜂兼进行腹部传粉(表3 )(张勃等, 2012 ).它们的访花姿态相似, 先落至花冠下唇, 再爬进冠筒取食花蜜, 在此过程中推动雄蕊下臂做杠杆状运动, 上臂花粉囊向下打在昆虫背部或腹部, 待其离开花器官时携带出花粉, 访花时间较长(本文作者观察). ...
... 相比毛地黄鼠尾草, 舌瓣鼠尾草传粉昆虫的单花访问时间、活跃度和访花频率均较高(表3 ), 说明小型花冠和退化的杠杆结构缩短了访花时间, 提高了活跃度, 可能促进了昆虫的访花效率.在传粉稳定性方面, 舌瓣鼠尾草花结构能够固定访花时昆虫取食花蜜的姿态, 保证传粉的可靠性和稳定性, 同时固定的花药和昆虫接触部位, 进一步提高了传粉的精确性.传粉结构和本地传粉者的特征共同决定了舌瓣鼠尾草的传粉方式.短药隔杠杆的毛地黄鼠尾草, 传粉昆虫进入冠筒有2种方式, 并且在冠筒中有自由活动的空间, 身体各部位都有接触花药的可能, 沾在腹部的花粉在飞行途中也有可能掉落, 增大了花粉损失的可能(张勃等, 2012 ).本研究仅从花器官结构和传粉昆虫的相互作用角度进行了深入研究, 具退化杠杆的物种是否具有选择优势?哪一种传粉模式更有效?还需从鼠尾草属植物的雄性适合度和繁育系统方面进行比较分析, 从而更深入地了解它们的生态学意义和进化选择压力. ...
... The data of S . digitaloides refer to Zhang et al . (2012) . F, forehead pollination; D, dorsal pollination; D/V, dorsal pollination prevails over ventral pollination. -, no data measured. ...
... 毛地黄鼠尾草传粉昆虫数据来源于张勃等(2012) .F, 头部传粉; D, 背部传粉; D/V, 背部兼腹部传粉.-, 无相关数据. ...
... 舌瓣鼠尾草有专一性的传粉昆虫三条熊蜂, 毛地黄鼠尾草则有3种.专一性的提高是否暗示植物与昆虫的相互关系更紧密, 协同进化程度更高?舌瓣鼠尾草传粉者的高度专一性可能还与所在地的昆虫种类组成相关, 究竟是退化杠杆结构过滤了其他传粉者, 还是传粉者的改变导致杠杆功能丧失? 都需要进一步深入研究.另一方面, 专一性的传粉昆虫面临的风险较大, 一旦失去传粉者, 就有无法延续生存的可能.相反, 具有多个传粉者的物种, 这样的风险则大大降低.目前对鼠尾草属短药隔杠杆机制的作用以及传粉者响应的生态学意义等方面研究较充分(Zhang et al ., 2011 ; 张勃等, 2012 ; Zhang & Li, 2014 ), 而其他类型杠杆的研究还很少, 对该属多种类型杠杆机制的阐明, 无疑会对揭示其起源和进化方向以及该物种多样性的产生、共存和维持机制提供更多证据. ...
鼠尾草属不同物种的雄蕊杠杆机制对传粉者空间变异的进化响应
8
2012
... 黄艳波等(2014)根据花器官结构和花粉接触传粉昆虫的部位, 将中国的鼠尾草属物种划分为3种传粉模式类型: 短药隔杠杆、长药隔杠杆和退化杠杆.3个类群大致形成以短药隔杠杆物种集中分布于中国西南, 长药隔杠杆物种集中于华中, 退化杠杆类型以华东为核心的分布格局.对鼠尾草属部分物种的杠杆传粉机制了解较多, 如短药隔杠杆类型的圆苞鼠尾草(S. cyclostegia )和长药隔杠杆类型丹参(S. miltiorrhiza )均借助传粉昆虫推动雄蕊做杠杆状运动, 完成花粉分发和授粉作用(张勃等, 2012 ; 黄艳波等, 2014 ).在中国, 鼠尾草亚属(subgenus Allagospadonopsis )的16个物种具有退化的杠杆结构, 推测它们具有非典型的杠杆传粉特征(黄艳波等, 2014 ).鼠尾草属雄蕊的分化是物种形成和多样性产生的关键因素(Claßen-Bockhoff et al ., 2003 ), 这种适应辐射的物种形成事件也是各种环境选择压力综合作用的结果.尽管对西南分布的短药隔类型鼠尾草的传粉结构、进化响应和表型选择等已有所了解(Zhang et al ., 2011 ; 张勃等, 2012 ; Zhang & Li, 2014 ), 但相比鼠尾草属多样性的传粉结构和传粉过程, 对其中一种类型的认识显然不足以揭示鼠尾草属面对的传粉者选择压力和进化方向.更何况, 至今对该属植物的传粉生态学和繁育系统仍然了解不多(Claßen-Bockhoff et al ., 2003 ). ...
... ; 张勃等, 2012 ; Zhang & Li, 2014 ), 但相比鼠尾草属多样性的传粉结构和传粉过程, 对其中一种类型的认识显然不足以揭示鼠尾草属面对的传粉者选择压力和进化方向.更何况, 至今对该属植物的传粉生态学和繁育系统仍然了解不多(Claßen-Bockhoff et al ., 2003 ). ...
... 采集植物种群中出现的访花昆虫, 首先用乙酸乙酯将昆虫毒死, 用镊子辅助保持自然伸展状态, 用电子游标卡尺进行形态特征测量, 包括昆虫体长、胸宽、胸厚和喙长4个指标.喙长测量时用镊子将其拉出至自然伸展状态, 测量完成后用昆虫针将其固定, 制成标本带回实验室进行拍照鉴定.对舌瓣鼠尾草采集样地内出现的所有访花昆虫进行鉴定测量, 毛地黄鼠尾草的传粉昆虫参考张勃等(2012) 在甘海子的测量结果. ...
... 在舌瓣鼠尾草的样方内记录观察到3种访花昆虫, 分别是蜜蜂科熊蜂属(Bombus )的三条熊蜂(B. trifasciatus ), 隧蜂科(Halictidae)和天蛾科(Sphing- idae)昆虫(图1 ).因天蛾科昆虫数量少、飞行速度快, 未能捕捉到活体, 故只测量了三条熊蜂和隧蜂科昆虫.从体型大小看, 3种访花昆虫形态特征差异明显(表2 ), 并且访花行为也完全不同.如图4 所示, 三条熊蜂体型较大, 只有一种访花姿态, 即直接飞至冠口, 依靠其前、中足抓住花冠前部保持平衡, 头部插入冠筒, 利用其长喙吸食花蜜, 此时前额紧贴雄蕊上臂的花粉囊, 将花粉带出, 完整的访花时间为(1.54 ± 0.60) s.当它再次进入另一朵具成熟雌蕊的花冠时, 便可将其头部的花粉授予柱头, 完成授粉.隧蜂科昆虫体型小, 通常飞落至花冠口再缓慢爬进冠筒, 自由地取食花粉, 并未取食花蜜, 而此时雌蕊尚未成熟.其单花访花时间很长、变幅较大, 平均为(73.9 ± 55.6) s.另一种天蛾科昆虫的飞行速率极快, 单花访花时间短, 平均只有(0.67 ± 0.23) s, 因其喙很长, 取食花蜜时没有触碰到花器官任何部位(图1 L).在甘海子样地, 毛地黄鼠尾草的访花昆虫有寄生性拟熊蜂(Psithyrus sp.)、小雅熊蜂(Bombus lepidus )和Bombus infrequens .三种访花昆虫主要进行背部传粉, 小雅熊蜂兼进行腹部传粉(表3 )(张勃等, 2012 ).它们的访花姿态相似, 先落至花冠下唇, 再爬进冠筒取食花蜜, 在此过程中推动雄蕊下臂做杠杆状运动, 上臂花粉囊向下打在昆虫背部或腹部, 待其离开花器官时携带出花粉, 访花时间较长(本文作者观察). ...
... 相比毛地黄鼠尾草, 舌瓣鼠尾草传粉昆虫的单花访问时间、活跃度和访花频率均较高(表3 ), 说明小型花冠和退化的杠杆结构缩短了访花时间, 提高了活跃度, 可能促进了昆虫的访花效率.在传粉稳定性方面, 舌瓣鼠尾草花结构能够固定访花时昆虫取食花蜜的姿态, 保证传粉的可靠性和稳定性, 同时固定的花药和昆虫接触部位, 进一步提高了传粉的精确性.传粉结构和本地传粉者的特征共同决定了舌瓣鼠尾草的传粉方式.短药隔杠杆的毛地黄鼠尾草, 传粉昆虫进入冠筒有2种方式, 并且在冠筒中有自由活动的空间, 身体各部位都有接触花药的可能, 沾在腹部的花粉在飞行途中也有可能掉落, 增大了花粉损失的可能(张勃等, 2012 ).本研究仅从花器官结构和传粉昆虫的相互作用角度进行了深入研究, 具退化杠杆的物种是否具有选择优势?哪一种传粉模式更有效?还需从鼠尾草属植物的雄性适合度和繁育系统方面进行比较分析, 从而更深入地了解它们的生态学意义和进化选择压力. ...
... The data of S . digitaloides refer to Zhang et al . (2012) . F, forehead pollination; D, dorsal pollination; D/V, dorsal pollination prevails over ventral pollination. -, no data measured. ...
... 毛地黄鼠尾草传粉昆虫数据来源于张勃等(2012) .F, 头部传粉; D, 背部传粉; D/V, 背部兼腹部传粉.-, 无相关数据. ...
... 舌瓣鼠尾草有专一性的传粉昆虫三条熊蜂, 毛地黄鼠尾草则有3种.专一性的提高是否暗示植物与昆虫的相互关系更紧密, 协同进化程度更高?舌瓣鼠尾草传粉者的高度专一性可能还与所在地的昆虫种类组成相关, 究竟是退化杠杆结构过滤了其他传粉者, 还是传粉者的改变导致杠杆功能丧失? 都需要进一步深入研究.另一方面, 专一性的传粉昆虫面临的风险较大, 一旦失去传粉者, 就有无法延续生存的可能.相反, 具有多个传粉者的物种, 这样的风险则大大降低.目前对鼠尾草属短药隔杠杆机制的作用以及传粉者响应的生态学意义等方面研究较充分(Zhang et al ., 2011 ; 张勃等, 2012 ; Zhang & Li, 2014 ), 而其他类型杠杆的研究还很少, 对该属多种类型杠杆机制的阐明, 无疑会对揭示其起源和进化方向以及该物种多样性的产生、共存和维持机制提供更多证据. ...
杠杆状雄蕊及其进化生态学意义
1
2010
... 在植物与昆虫的相互作用中, 有关花形态塑造和昆虫的适应进化是生态学和植物进化研究中最吸引人的一个领域(黄双全和郭友好, 2000 ; 黄双全, 2007 ; 方强和黄双全, 2014 ).鼠尾草属(Salvia )植物具有花器官两侧对称、二强雄蕊、虫媒异交和杠杆传粉机制等特征, 与传粉者的协同进化程度较高(张勃等, 2010 ), 典型的杠杆传粉过程基于雄蕊药隔的显著伸长、联合具阻挡作用的药隔下壁和明显可运动的“关节”等结构, 在传粉昆虫进入花冠取食花蜜时, 迫使其推动下臂做杠杆状运动, 上臂可育花药将花粉涂抹在昆虫背部从而携带出花粉.鼠尾草属雄蕊结构与功能的分化可能促进了该属物种多样性的形成(Claßen-Bockhoff et al ., 2004 ).根据传粉者类型, 鼠尾草属可分为蜂媒传粉(蜂媒花, melittophilae)和鸟媒传粉(鸟媒花, ornithophily), 蜂媒传粉物种主要分布于欧亚大陆、地中海沿岸、非洲东部及南非、大洋洲以及北美部分地区; 鸟媒传粉物种分布于北美西部至墨西哥、中美洲和南美洲安第斯山脉及东部(魏宇昆等, 2015 ).就雄蕊结构和传粉特征而言, 鼠尾草属中还具有非典型的杠杆状雄蕊结构和传粉模式. ...
杠杆状雄蕊及其进化生态学意义
1
2010
... 在植物与昆虫的相互作用中, 有关花形态塑造和昆虫的适应进化是生态学和植物进化研究中最吸引人的一个领域(黄双全和郭友好, 2000 ; 黄双全, 2007 ; 方强和黄双全, 2014 ).鼠尾草属(Salvia )植物具有花器官两侧对称、二强雄蕊、虫媒异交和杠杆传粉机制等特征, 与传粉者的协同进化程度较高(张勃等, 2010 ), 典型的杠杆传粉过程基于雄蕊药隔的显著伸长、联合具阻挡作用的药隔下壁和明显可运动的“关节”等结构, 在传粉昆虫进入花冠取食花蜜时, 迫使其推动下臂做杠杆状运动, 上臂可育花药将花粉涂抹在昆虫背部从而携带出花粉.鼠尾草属雄蕊结构与功能的分化可能促进了该属物种多样性的形成(Claßen-Bockhoff et al ., 2004 ).根据传粉者类型, 鼠尾草属可分为蜂媒传粉(蜂媒花, melittophilae)和鸟媒传粉(鸟媒花, ornithophily), 蜂媒传粉物种主要分布于欧亚大陆、地中海沿岸、非洲东部及南非、大洋洲以及北美部分地区; 鸟媒传粉物种分布于北美西部至墨西哥、中美洲和南美洲安第斯山脉及东部(魏宇昆等, 2015 ).就雄蕊结构和传粉特征而言, 鼠尾草属中还具有非典型的杠杆状雄蕊结构和传粉模式. ...
 , 王琦
, 王琦


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}