植物化学计量学特征在植物生活史(Wright et al ., 2004 )、功能性状(He et al ., 2009 )以及植物死后的生态系统过程(Güsewell & Gessner, 2009 )中扮演极为重要的角色。N、P、K是各类生态系统中限制植物生长的关键元素(Koerselman & Meuleman, 1996 ; Olde Venterink et al ., 2003 ; Harpole et al ., 2011 ), 其在植物体内的含量与植物生长速率具有直接的关联性(Ågren, 2008 )。这些元素的计量学特征对植物生长(阎恩荣等, 2008 ; 刘兴诏等, 2010 ; 宾振钧等, 2014 )、生态系统功能的发挥(Yu et al ., 2010 )具有极为重要的作用。然而, 在生态系统功能中具有重要作用的元素并不仅限于N、P、K, 其他元素如Ca、Mg、Mn以及其他微量元素在植物生长和凋落物分解过程中具有举足轻重的地位(Makkonen et al ., 2012 ; 闫帮国等, 2013 ), 进而可以对土壤或者地表碳库产生重要的影响(Reich et al ., 2005 )。然而迄今为止, 对这些元素的稳定性和变异性的了解却很少(贺金生和韩兴国, 2010 ), 对这些元素计量关系的驱动因素也不清楚。
在全球变化的背景下, 植物的化学计量学与生产力对环境变化的响应无疑会对生物群落乃至生态系统功能变迁(Sistla & Schimel, 2012 )起到重要的级联效应。各种植物的化学计量学特征是植物在各自生境下适应环境的结果, 也与植物生长速率等重要特性紧密关联(Elser et al ., 2000 , 2003 ; Sterner & Elser, 2002 )。然而, 植物的化学计量学特征具有很强的可塑性(Güsewell, 2004 ), 这种可塑性对环境因素的响应可能直接影响植物的生长能力和对环境的适应能力(Yu et al ., 2010 ; Sistla & Schimel, 2012 )。因此, 阐明植物化学计量学变化的驱动机制有利于理解植物在全球变化背景下的适应机制和植物的生态系统功能(Elser et al ., 2010 )。
众所周知, 植物生产力是生态系统功能的主体因子之一, 植物元素计量关系变化与植物生产力变化的耦合必然对生态系统过程和功能产生重要影响。其中养分和水分是影响植物生长生产力的最重要因素。然而养分和水分对植物生长的促进作用极为复杂, 可能存在拮抗效应、协同效应、加和效应等(Harpole et al ., 2007 ; Niu et al ., 2009 ; Lü et al ., 2014 )。有关养分和水分的复杂作用对植物元素平衡影响的研究很少(Lü et al ., 2012 ), 而大量元素与微量元素的计量关系研究更是缺乏, 这限制了我们对全球变化背景下生态系统功能的预测。生物体生长稀释理论(somatic growth dilution)认为重金属在生物体内的富集与生物生长速率有关(Karimi et al ., 2007 )。在生物体快速生长时, 其吸收的重金属可以被稀释(Sunda & Huntsman, 1998 )。然而植物生长快慢对微量元素的影响, 特别是大量元素与微量元素计量比的影响还不清楚。本研究以金沙江干热河谷地区的土壤和植物为实验材料, 研究水分和养分处理对植物生物量的作用, 并探索生物量变化与植物化学计量学特征变化的关联。研究结果将有助于揭示该地区植物化学计量学特征对水分和养分变化的响应机制。
1 材料和方法
1.1 研究区概况
干热河谷地区以其特殊的气候而成为一个独特的地理单元。这其中尤以金沙江干热河谷元谋段最为典型。区域内年平均气温约为21.5 ℃, 降水量小于700 mm, 降水主要集中在雨季(6-10月间的降水量占90%以上) (张建平等, 2000 )。
1.2 实验设计与数据收集
采集干热河谷6种草本植物种子各200 g, 这些植物基本涵盖了当地群落的典型植物, 包括4种优势植物: 黄茅(Heteropogon contortus )、孔颖草(Bothriochloa pertusa )、橘草(Cymbopogon goeringii )、双花草(Dichanthium annulatum ); 一种稀有植物——裂稃草(Schizachyrium brevifolium )和1种引进植物——红毛草(Melinis repens )。
2012年6月, 在温室中设置盆栽实验。每个物种装12盆, 在盆中装填当地典型的燥红土10 kg, 播入5 g种子, 待植物发芽后一周, 除去多余的个体直至每盆4株植物, 再分别作水分处理、养分处理。实验中第一组为对照处理(C), 每个花盆中每星期浇两次水, 每次300 mL; 第二组为养分处理(N), 每隔两个星期添加300 mL 1/6 Hogland溶液代替当次浇水, 其余处理与对照相同; 第三组为高频次水分添加处理(W), 每星期浇3次水, 每次300 mL; 第四组为水分和养分的复合处理(NW), 每星期添加3次水, 每次300 mL, 此外每隔两星期添加300 mL 1/6 Hogland溶液。实验共计2个水分处理× 2个养分处理× 3个重复× 6个物种= 72盆。所有花盆, 包括对照处理每个月灌注1次2000 mL水以淋洗土壤的盐离子, 避免养分处理造成的盐毒害。由于干热河谷地区蒸发量很高, 因此灌注的水很快就会被蒸发, 对随后的处理不会造成太大影响。
实验开始后100天, 收取植物, 测量生物量数据。其中地上部分刈割后带回实验室70 ℃烘干, 分离成叶片、茎干(含叶鞘部分)和花穗部分后各自称质量。根部收集时, 将花盆中所有根系连同土壤一同倒进1 mm的网袋中, 放在水龙头下冲洗掉所有的土壤后, 拣去其他杂物和较大的砂粒, 根系装入信封后70 ℃烘干称质量。
由于花穗样品重量有限, 每种植物的花穗混合后作为一个样品, 其他器官则是每种植物相同处理下的样品混合成一个混样, 粉碎后测定N、P、K、Ca、Mg、Zn、Mn含量。N含量采用凯氏定氮法测量(GB 5009.5-2010), 其他元素采用电感耦合等离子发射光谱仪测定(NY/T 1653-2008)。
1.3 数据分析
植物总体各种元素的含量计算公式为
E = ∑Ei × Oi (1)
E 为植物体内元素的含量, Ei 为该元素在器官中的含量, Oi 为器官在植株质量中的比值。
植物生物量和化学计量学特征对水分和养分处理的响应指数通过以下公式计算
R = (Bt - Bc )/Bc ×100% (2)
R 为响应指数(本文以R 加下标形式指示各种参数的响应, 如R stoichiometry 指示植物各种化学计量特征的响应指数, R biomass 指示生物量对处理的响应指数), Bc 为对照处理的植物生物量或化学计量学特征, Bt 为植物在水分、养分以及水分-养分复合处理下的植物生物量或化学计量学特征。
我们用多因素方差分析法, 分析物种、水分、养分处理及其交互作用对植物生物量的影响, 并对各处理间进行post-hoc检验。采用一元线性回归分析法分析R biomass 与R stoichiometry 的相关性, 采用多元回归模型分析R biomass 、处理类型(或物种)对R stoichiometry 的影响。
2 结果
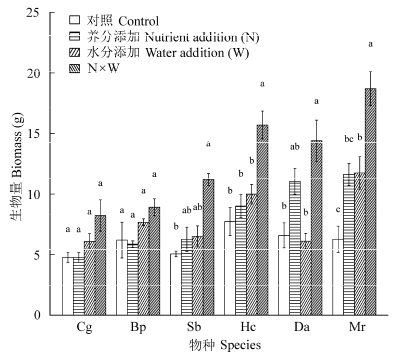
水分、养分、物种以及各因素的二元交互作用对植物生物量具有显著的效应, 只有三元交互作用不显著(表1 )。总体上, 养分添加处理的生物量比对照增加了32.55%, 高频次水分处理增加了31.35%, 水分与养分复合处理下生物量增加了110.60 %, 体现出水分与养分的协同效应(图1 )。
物种生物量均值的排序为: 红毛草> 黄茅> 双花草> 孔颖草> 裂稃草> 橘草。不同的物种对处理的响应特征各不相同。橘草和孔颖草对水分的响应较为明显, 而对养分响应不明显。裂稃草和黄茅对水分和养分的复合处理响应较大, 而对养分和水分的单独处理响应较小, 呈现出极强的协同效应; 双花草只对养分与水分的复合处理展现出显著的响应, 而对高频次水分处理的响应不显著; 红毛草则对水分和养分处理的响应程度相似, 但是对水分和养分的复合处理响应大约是二者的加和, 体现了水分和养分的加性效应(图1 )。
图1 养分、水分处理下的植物生物量(平均值±标准误差)。Hc, 黄茅; Bp, 孔颖草; Cg, 橘草; Da, 双花草; Sb, 裂稃草; Mr, 红毛草。物种内不同字母表示处理间存在显著性差异(p < 0.05)。
Fig. 1 Response of plant biomass to nutrient and water treatments (mean ± SE ). Hc, Heteropogon contortus; Bp, Bothriochloa pertusa; Cg, Cymbopogon goeringii; Da, Dichanthium annulatum; Sb, Schizachyrium brevifolium; Mr, Melinis repens . For each species different letters denote significant difference in biomass between treatments (p < 0.05).
植物体元素含量总体上为K > N > Ca > Mg > P > Mn > Zn (表2 )。植物总体的N:P:K:Ca:Mg:Mn:Zn为141.07:10.96:164.64:70.52:26.06:2.20:1。N的平均值为9.243 mg·g-1 (变化幅度5.872-15.229 mg·g-1 ), P平均值为0.718 mg·g-1 (变化幅度0.535-1.019 mg·g-1 ), K的平均值为10.788 mg·g-1 (变化幅度7.506-15.317 mg·g-1 ), Ca的平均值为4.621 mg·g-1 (变化幅度2.664-7.220 mg·g-1 ), Mg的平均值为1.707 mg·g-1 (变化幅度介于1.183-2.809 mg·g-1 ), Mn的平均值为0.144 mg·g-1 (变化幅度0.094-0.207 mg·g-1 ), Zn的平均值为0.066 mg·g-1 (变化幅度0.041-0.131 mg·g-1 )。
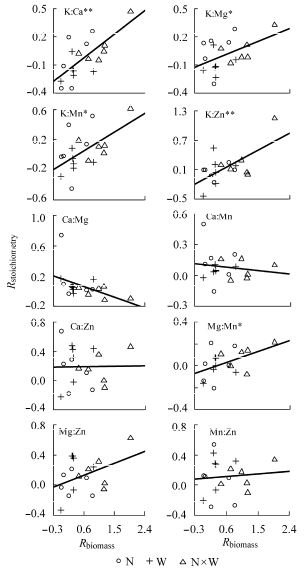
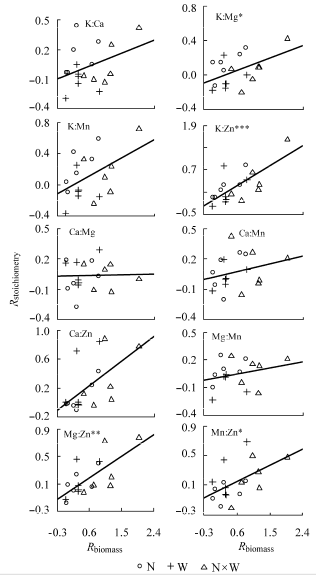
植物总体N、P、K含量与植物生物量响应指数相关性不显著, 而Ca、Mg、Mn、Zn含量的变化与植物生物量变化呈显著负相关(p < 0.05) (表3 )。植物总体N:P以及N、P与其他元素的计量比变化与生物量变化相关性不显著, 植物叶片、茎干和根系中这些化学计量特征与生物量变化相关性也不显著。此后不再讨论N、P与其他元素的计量关系。植物总体K:Ca、K:Mg、K:Mn、K:Zn、Mg:Mn与植物生物量响应指数显著正相关(p < 0.05) (图2 )。
图2 植物总体元素计量学特征响应指数(R stoichiometry )与植物生物量响应指数(R biomass )的关系。**, p < 0.01; *, p < 0.05。N, 养分添加; W, 水分添加。
Fig. 2 Relationships between response index of whole-plant stoichiometry (R stoichiometry ) and response index of plant biomass (R biomass ). N, nutrient addition; W, water addition. **, p < 0.01; *, p < 0.05.
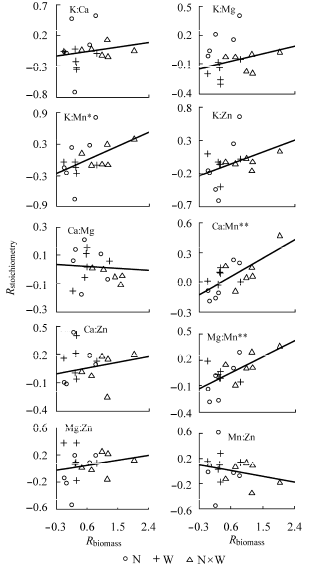
植物叶片Mn含量的变化与植物生物量变化呈显著负相关, 其他元素含量与生物量变化不相关(p < 0.05)。植物叶片K:Mn、Ca:Mn、Mg:Mn与植物生物量响应指数显著正相关(p < 0.05) (图3 )。
图3 植物叶片元素计量学特征响应指数(R stoichiometry )与植物生物量响应指数(R biomass )的关系。N, 养分添加; W, 水分添加。
Fig. 3 Relationship between response index of leaf stoich- iometry (R stoichiometry ) and response index of plant biomass (R stoichiometry ). N, nutrient addition; W, water addition.
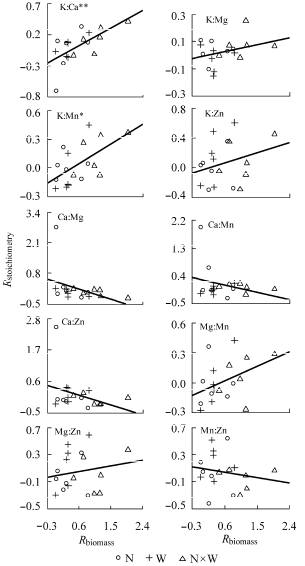
植物根系元素含量的变化与植物生物量变化相关性均不显著(p > 0.05)。植物根系K:Ca、K:Mn与植物生物量响应指数显著正相关(p < 0.05)(图4 )。
图4 植物根系元素计量学特征响应指数(R stoichiometry )与植物生物量响应指数(R biomass )的关系。N, 养分添加; W, 水分添加。
Fig. 4 Relationship between response index of root stoich- iometry (R stoichiometry ) and response index of plant biomass (R biomass ). N, nutrient addition; W, water addition.
植物茎干元素含量的变化与植物生物量变化相关性均不显著(p > 0.05)。植物茎干K:Mg、K:Zn、Mg:Zn、Mn:Zn与植物生物量响应指数显著正相关(p < 0.05) (图5 )。
图5 植物茎干元素计量学特征响应指数(R stoichiometry )与植物生物量响应指数(R biomass )的关系。N, 养分添加; W, 水分添加。
Fig. 5 Relationship between response index of stem stoich- iometry (R stoichiometry ) and response index of plant biomass (R biomass ). N, nutrient addition; W, water addition.
将处理因素和生物量同时作为影响因素, 进行多元回归分析时, 生物量变化仍然显著影响了植物总体K:Ca、K:Mg、K:Mn、K:Zn, 茎干的K:Mg、K:Zn、K:Mn、Ca:Zn、Mg:Zn、Mn:Zn, 叶片的K:Mn、Ca:Mn、Mg:Mn和根系的K:Ca、K:Mn (p < 0.05), 处理类型显著影响了茎干K:Ca、K:Mn (p < 0.05), 对其他元素计量关系影响不显著。将物种因素和生物量同时作为影响因素时, 生物量变化仍然显著影响植物总体的K:Ca、K:Zn、Mg:Zn、茎干中K:Zn、Mg:Zn、叶片中Ca:Mn和根系中K:Ca、K:Mn (p < 0.05)。物种只显著影响了植物总体Mg:Zn (p < 0.05)。
3 讨论
本研究结果显示植物根茎叶以及总体K、Ca、Mg、Mn、Zn含量及其化学计量特征对水分、养分处理下生物量变化具有明显的响应。这些元素基本上可分为三大类: 第一类是K, 第二类是Ca、Mg, 第三类是Mn、Zn。从元素含量的高低顺序来看, 植物含量较高的元素在生物量变化中较为稳定, 而微量元素则随着生物量增加有降低趋势。这使得含量较高的元素与含量较低的元素计量关系发生了极大变化, 导致第一类元素与第二、三类元素的计量比、第二类元素与第三类元素的计量比随着生物量增加而不断增加。
植物在养分和水分处理时的生物量变化可能影响了植物对元素的吸收平衡。本研究发现元素计量比的变化趋势似乎与元素含量具有一定的联系, 如本研究中元素含量的排序为K > Ca > Mg > Mn > Zn, 而大部分情况下这些元素计量比的变化与生物量变化成正比。如植物总体K:Mg、K:Mn、K:Zn比值变化与生物量变化成正比, 这与K含量大于Mn、Zn含量一致, 表明元素含量越高的元素与含量较低的元素比值随着生物量的增大而不断增大。即便是大量元素内部(如植物总体K:Ca、K:Mg), 或者微量元素内部(如茎干Mn:Zn)元素计量关系的变化也与生物量变化呈正相关关系。
水分、养分可以影响植物化学计量学特征(Ågren & Weih, 2012 ; Lü et al ., 2012 ), 但这种影响背后的生物量因素却很少受到重视。本研究还发现, 生物量是这些元素的计量比变化的主要影响因子, 而环境条件(这里为水分处理、养分处理以及二者的复合处理)和物种因素的影响很小, 表明水分和养分对植物化学计量学特征的影响作用具有共同的驱动因素, 即生物量变化。本研究还发现元素化学计量学特征的变化与生物量变化的关系在根茎叶中都存在, 说明这可能是个普遍现象。
植物化学计量学特征与生物量都是生态系统功能的主要载体, 认识这二者之间的变化关系对于理解全球变化背景下生态系统功能具有重要意义。因此, 植物元素含量以及化学计量关系变化的驱动机制一直是生态化学计量学领域重点关注的问题(Güsewell, 2004 ; Ågren, 2008 ; He et al ., 2008 )。Elser等(2000)认为元素的计量学变异范围与元素含量有关, 越是大量元素, 其含量也越为稳定。而Han等(2011)研究认为元素含量的变异范围与该元素的限制情况有关, 越是限制性元素, 其含量变化越小。Karimi和Folt (2006)研究认为相对于大量元素, 微量元素很不稳定。事实上, 植物叶片中这些元素变化之间具有一定的耦合性, 且部分元素的变化速率高于其他元素(闫帮国等, 2012 )。然而, 植物元素的变化受到何种因素驱动却一直没有定论, 植物元素含量及其计量比与生物量变化的关系也很少受到关注。本研究的结果并不否定前人的研究结果, 而是从生物量变化的层面反映和探讨了植物化学计量学特征变化的驱动机制, 表明植物体元素的变异与生物量变化有关。
虽然微量元素是生物体必需的元素, 然而相对于大量元素, 其在生长中的限制作用可能较小(Karimi & Folt, 2006 )。可能如同重金属一样, 在植物快速生长中被大量稀释, 形成稀释效应(Karimi et al ., 2007 )。本研究中微量元素含量变化与生物量变化成反比的关系与上述研究结果相一致(表3 )。相反, 大量元素在生物体中的生物化学功能极为重要, 其具有相对较高的内稳性(Sterner & Elser, 2002 ), 因此, 在生物量变化中必须被不断补充, 从而保持稳定性。此外, 水分和养分可能影响了植物对大量元素的吸收, 促进了植物生长, 在保持大量元素较为稳定的同时稀释了微量元素。这种变化格局可能最终导致大量元素与微量元素的计量关系随生物量增长而增长。N、P含量及其与其他元素的计量比与生物量变化的关系并不显著, 具体的原因还不清楚。N、P在土壤中的相当一部分形态是有机物, 因此其吸收取决于植物与微生物的关系(van der Heijden et al ., 2008 ), 而其他元素主要以无机物形态存在, 其吸收主要取决于植物根系。这种差异可能造成了N、P与其他元素在生物生长中受到了不同因素的影响, 其计量关系因而没有呈现出显著的变化格局。
植物生长能力与植物体元素计量比的变化可以对生态系统的生物地球化学循环造成深远的影响(Jobbágy & Jackson, 2004 ; Mooshammer et al ., 2012 )。微量元素和大量元素一样, 是生态系统中物质循环的重要因素, 一些元素是凋落物分解(Berg et al ., 2007 )、动物生存(Karimi & Folt, 2006 )的重要限制因素, 其作用甚至会超过大量元素。如Mn是一些生态系统凋落物分解的限制因子(Berg & Mcclaugherty, 2003 ), 而其他的微量元素也可能在其中起重要作用(Kaspari et al ., 2008 ), 从而影响碎屑食物链的物质和能量的传输效率。大量元素与微量元素计量比的增大极有可能促进微量元素的限制作用。
水分和养分对植物化学计量学特征的影响必将作用于植物的生态系统功能。但水分和养分对植物化学计量学特征的影响与生物量变化有关, 生物量对水分或养分响应强烈的物种, 其化学计量学特征响应也强烈。因此, 这一切主要取决于物种、水分和养分的互作效应对生物量的作用。植物物种对水分、养分变化的差异性响应对干热地区生态系统的功能变化可能尤为重要。干热河谷地区的水分和养分都较为稀缺。一些植物的生物量对降水增加较为敏感, 另外一部分物种生物量对养分增加较为敏感。这些差异对物种在水分和养分条件不同的斑块中的分布具有重要作用(Condit et al ., 2013 ), 但是也会对植物化学计量学产生影响。水分和养分在提高生物量的同时, 也增加了大量元素与微量元素的计量比。
目前还不了解元素计量特征与生物量变化的关系是否在人工作物中也普遍存在。高强度的水肥投入使得作物产量大幅度提升, 这是否会影响农产品的元素计量特征以及人类的健康(Dellapenna, 1999 )还缺乏研究。
致谢 感谢杨继云同事在实验过程中给予的大力帮助!
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Ågren GI 2008 ). Stoichiometry and nutrition of plant growth in natural communities.
Annual Review of Ecology, Evolution, and Systematics , 39 , 153 -170 .
[本文引用: 2]
[2]
Ågren GI Weih M 2012 ). Plant stoichiometry at different scales: Element concentration patterns reflect environment more than genotype.
New Phytologist , 194 , 944 -952 .
[本文引用: 1]
[3]
Berg B McClaugherty C 2003 ). Plant Litter: Decomposition, Humus Formation, Carbon Sequestration. 2nd edn. Springer-Verlag, Berlin
, Heidelberg.
[本文引用: 1]
[4]
Berg B Steffen KT McClaugherty C 2007 ). Litter decom- position rate is dependent on litter Mn concentrations.
Biogeochemistry , 82 , 29 -39 .
[本文引用: 1]
[5]
Bin ZJ Wang JJ Zhang WP Xu DH Cheng XH Li KJ Cao DH 2014 ). Effects of N addition on ecological stoichiometric characteristics in six dominant plant species of alpine meadow on the Qinghai-Xizang Plateau, China.
Chinese Journal of Plant Ecology , 38 , 231 -237 .(in Chinese with English abstract)
[本文引用: 1]
[宾振钧 , 王静静 , 张文鹏 , 徐当会 , 程雪寒 , 李柯杰 , 曹德昊 (2014 ). 氮肥添加对青藏高原高寒草甸6个群落优势种生态化学计量学特征的影响
. 植物生态学报 , 38 , 231 -237 .]
[本文引用: 1]
[6]
Condit R Engelbrecht BMJ Pino D Pérez R Turner BL 2013 ). Species distributions in response to individual soil nutrients and seasonal drought across a community of tropical trees.
Proceedings of the National Academy of Sciences of the United States of America , 110 , 5064 -5068 .
[本文引用: 1]
[7]
DellaPenna D 1999 ). Nutritional genomics: Manipulating plant micronutrients to improve human health.
Science , 285 , 375 -379 .
[本文引用: 1]
[8]
Elser JJ Acharya K Kyle M Cotner J Makino W Markow T Watts T Hobbie S Fagan W Schade J Hood J Sterner RW 2003 ). Growth rate-stoichiometry couplings in diverse biota.
Ecology Letters , 6 , 936 -943 .
[本文引用: 1]
[9]
Elser JJ Fagan WF Kerkhoff AJ Swenson NG Enquist BJ 2010 ). Biological stoichiometry of plant production: Metabolism, scaling and ecological response to global change.
New Phytologist , 186 , 593 -608 .
[本文引用: 1]
[10]
Elser JJ Sterner RW Gorokhova E Fagan WF Markow TA Cotner JB Harrison JF Hobbie SE Odell GM Weider LW 2000 ). Biological stoichiometry from genes to ecosystems.
Ecology Letters , 3 , 540 -550 .
[本文引用: 1]
[11]
Güsewell S 2004 ). N:P ratios in terrestrial plants: Variation and functional significance.
New Phytologist , 164 , 243 -266 .
[本文引用: 2]
[12]
Güsewell S Gessner MO 2009 ). N:P ratios influence litter decomposition and colonization by fungi and bacteria in microcosms.
Functional Ecology , 23 , 211 -219 .
[本文引用: 1]
[13]
Han WX Fang JY Reich PB Ian Woodward F Wang ZH 2011 ). Biogeography and variability of eleven mineral elements in plant leaves across gradients of climate, soil and plant functional type in China.
Ecology Letters , 14 , 788 -796 .
[14]
Harpole WS Ngai JT Cleland EE Seabloom EW Borer ET Bracken MES Elser JJ Gruner DS Hillebrand H Shurin JB Smith JE 2011 ). Nutrient co-limitation of primary producer communities.
Ecology Letters , 14 , 852 -862 .
[本文引用: 1]
[15]
Harpole WS Potts DL Suding KN 2007 ). Ecosystem responses to water and nitrogen amendment in a California grassland.
Global Change Biology , 13 , 2341 -2348 .
[本文引用: 1]
[16]
He JS Han XG 2010 ). Ecological stoichiometry: Searching for unifying principles from individuals to ecosystems.
Chinese Journal of Plant Ecology , 34 , 2 -6 .(in Chinese)
[本文引用: 1]
[贺金生 , 韩兴国 (2010 ). 生态化学计量学: 探索从个体到生态系统的统一化理论
. 植物生态学报 , 34 , 2 -6 .]
[本文引用: 1]
[17]
He JS Wang L Flynn DFB Wang XP Ma WH Fang JY 2008 ). Leaf nitrogen: Phosphorus stoichiometry across Chinese grassland biomes.
Oecologia , 155 , 301 -310 .
[本文引用: 1]
[18]
He JS Wang XP Flynn DFB Wang L Schmid B Fang JY 2009 ). Taxonomic, phylogenetic, and environmental trade-offs between leaf productivity and persistence.
Ecology , 90 , 2779 -2791 .
[本文引用: 1]
[19]
Jobbágy EG Jackson RB 2004 ). The uplift of soil nutrients by plants: Biogeochemical consequences across scales.
Ecology , 85 , 2380 -2389 .
[本文引用: 1]
[20]
Karimi R Chen CY Pickhardt PC Fisher NS Folt CL 2007 ). Stoichiometric controls of mercury dilution by growth.
Proceedings of the National Academy of Sciences of the United States of America , 104 , 7477 -82 .
[本文引用: 2]
[21]
Karimi R Folt CL 2006 ). Beyond macronutrients: Element variability and multielement stoichiometry in freshwater invertebrates.
Ecology Letters , 9 , 1273 -1283 .
[本文引用: 2]
[22]
Kaspari M Garcia MN Harms KE Santana M Wright SJ Yavitt JB 2008 ). Multiple nutrients limit litterfall and decomposition in a tropical forest.
Ecology Letters , 11 , 35 -43 .
[本文引用: 1]
[23]
Koerselman W Meuleman AFM 1996 ). The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation.
Journal of Applied Ecology , 33 , 1441 -1450 .
[本文引用: 1]
[24]
Liu XZ Zhou GY Zhang DQ Liu SZ Chu GW Yan JH 2010 ). N and P stoichiometry of plant and soil in lower subtropical forest successional series in southern China.
Chinese Journal of Plant Ecology , 34 , 64 -71 .(in Chinese with English abstract)
[本文引用: 1]
[刘兴诏 , 周国逸 , 张德强 , 刘世忠 , 褚国伟 , 闫俊华 (2010 ). 南亚热带森林不同演替阶段植物与土壤中N、P的化学计量特征
. 植物生态学报 , 34 , 64 -71 .]
[本文引用: 1]
[25]
Lü XT Dijkstra FA Kong DL Wang ZW Han XG 2014 ). Plant nitrogen uptake drives responses of productivity to nitrogen and water addition in a grassland.
Scientific Reports , 4 , 4817 .
[本文引用: 1]
[26]
Lü XT Kong DL Pan QM Simmons M Han XG 2012 ). Nitrogen and water availability interact to affect leaf stoichiometry in a semi-arid grassland.
Oecologia , 168 , 301 -310 .
[本文引用: 2]
[27]
Makkonen M Berg MP Handa IT Hättenschwiler S van Ruijven J van Bodegom PM Aerts R 2012 ). Highly consistent effects of plant litter identity and functional traits on decomposition across a latitudinal gradient.
Ecology Letters , 15 , 1033 -1041 .
[本文引用: 1]
[28]
Mooshammer M Wanek W Schnecker J Wild B Leitner S Hofhansl F Blöchl A Hämmerle I Frank AH Fuchslueger L Keiblinger KM Zechmeister-Boltenstern S Richter A 2012 ). Stoichiometric controls of nitrogen and phosphorus cycling in decomposing beech leaf litter.
Ecology , 93 , 770 -782 .
[本文引用: 1]
[29]
Niu SL Yang HJ Zhang Z Wu MY Lu Q Li LH Han XG Wan SQ 2009 ). Non-additive effects of water and nitrogen addition on ecosystem carbon exchange in a temperate steppe.
Ecosystems , 12 , 915 -926 .
[本文引用: 1]
[30]
Olde Venterink H Wassen MJ Verkroost AWM de Ruiter PC 2003 ). Species richness-productivity patterns differ between N-, P-, and K-limited wetlands.
Ecology , 84 , 2191 -2199 .
[本文引用: 1]
[31]
Reich PB Oleksyn J Modrzynski J Mrozinski P Hobbie SE Eissenstat DM Chorover J Chadwick OA Hale CM Tjoelker MG 2005 ). Linking litter calcium, earthworms and soil properties: A common garden test with 14 tree species.
Ecology Letters , 8 , 811 -818 .
[本文引用: 1]
[32]
Sistla SA Schimel JP 2012 ). Stoichiometric flexibility as a regulator of carbon and nutrient cycling in terrestrial ecosystems under change.
New Phytologist , 196 , 68 -78 .
[本文引用: 2]
[33]
Sterner RW Elser JJ 2002 ). Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere . Princeton University Press , Princeton .
[本文引用: 2]
[34]
Sunda WG Huntsman SA 1998 ). Processes regulating cellular metal accumulation and physiological effects: Phytoplankton as model systems.
Science of The Total Environment , 219 , 165 -181 .
[本文引用: 1]
[35]
van der Heijden MGA Bardgett RD Van Straalen NM 2008 ). The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems.
Ecology Letters , 11 , 296 -310 .
[本文引用: 1]
[36]
Wright IJ Reich PB Westoby M Ackerly DD Baruch Z Bongers F Cavender-Bares J Chapin T Cornelissen JHC Diemer M Flexas J Garnier E Groom PK Gulias J Hikosaka K Lamont BB Lee T Lee W Lusk C Midgley JJ Navas M-L Niinemets Ü Oleksyn J Osada N Poorter H Poot P Prior L Pyankov VI Roumet C Thomas SC Tjoelker MG Veneklaas EJ Villar R 2004 ). The worldwide leaf economics spectrum.
Nature , 428 , 821 -827 .
[本文引用: 1]
[37]
Yan BG He GX Li JC Ji ZH 2012 ). Scaling relationships and stoichiometry of plant leaf biogenic elements from the arid-hot valley of Jinsha River, China.
Chinese Journal of Plant Ecology , 36 , 1136 -1144 .(in Chinese with English abstract)
[本文引用: 1]
[闫帮国 , 何光熊 , 李纪潮 , 纪中华 (2012 ). 金沙江干热河谷地区植物叶片中各生源要素的化学计量特征以及异速增长关系
. 植物生态学报 , 36 , 1136 -1144 .]
[本文引用: 1]
[38]
Yan BG Ji ZH He GX Shi LT Pan ZX Li JC Yue XW Wang CX Peng H 2013 ). The effects of leaf stoichiochemistric characters on litter turnover in an arid-hot valley of Jinsha River, China.
Acta Ecologica Sinica , 33 , 5668 -5674 .(in Chinese with English abstract)
[本文引用: 1]
[闫帮国 , 纪中华 , 何光熊 , 史亮涛 , 潘志贤 , 李纪潮 , 岳学文 , 王春雪 , 彭辉 (2013 ). 金沙江干热河谷植物叶片元素含量在地表凋落物周转中的作用
. 生态学报 , 33 , 5668 -5674 .]
[本文引用: 1]
[39]
Yan ER Wang XH Zhou W 2008 ). N:P stoichiometry in secondary succession in evergreen broad-leaved forest, Tiantong, East China. Journal of Plant Ecology (
Chinese Version ), 32 , 13 -22 .(in Chinese with English abstract)
[本文引用: 1]
[阎恩荣 , 王希华 , 周武 (2008 ). 天童常绿阔叶林演替系列植物群落的N:P化学计量特征
. 植物生态学报 , 32 , 13 -22 .]
[本文引用: 1]
[40]
Yu Q Chen QS Elser JJ He NP Wu HH Zhang GM Wu JG Bai YF Han XG 2010 ). Linking stoichiometric homoeostasis with ecosystem structure, functioning and stability.
Ecology Letters , 13 , 1390 -1399 .
[本文引用: 2]
[41]
Zhang JP Wang DJ Wang YK Wen AB 2000 ). Discusses on eco-environment changes in dry-lot valley of Yuanmou.
Scinentia Geographica Sinica , 20 , 148 -152 .(in Chinese with English abstract)
[本文引用: 1]
[张建平 , 王道杰 , 王玉宽 , 文安邦 (2000 ). 元谋干热河谷区生态环境变迁探讨
. 地理科学 , 20 , 148 -152 .]
[本文引用: 1]
Stoichiometry and nutrition of plant growth in natural communities.
2
2008
... 植物化学计量学特征在植物生活史(Wright et al ., 2004 )、功能性状(He et al ., 2009 )以及植物死后的生态系统过程(Güsewell & Gessner, 2009 )中扮演极为重要的角色.N、P、K是各类生态系统中限制植物生长的关键元素(Koerselman & Meuleman, 1996 ; Olde Venterink et al ., 2003 ; Harpole et al ., 2011 ), 其在植物体内的含量与植物生长速率具有直接的关联性(Ågren, 2008 ).这些元素的计量学特征对植物生长(阎恩荣等, 2008 ; 刘兴诏等, 2010 ; 宾振钧等, 2014 )、生态系统功能的发挥(Yu et al ., 2010 )具有极为重要的作用.然而, 在生态系统功能中具有重要作用的元素并不仅限于N、P、K, 其他元素如Ca、Mg、Mn以及其他微量元素在植物生长和凋落物分解过程中具有举足轻重的地位(Makkonen et al ., 2012 ; 闫帮国等, 2013 ), 进而可以对土壤或者地表碳库产生重要的影响(Reich et al ., 2005 ).然而迄今为止, 对这些元素的稳定性和变异性的了解却很少(贺金生和韩兴国, 2010 ), 对这些元素计量关系的驱动因素也不清楚. ...
... 植物化学计量学特征与生物量都是生态系统功能的主要载体, 认识这二者之间的变化关系对于理解全球变化背景下生态系统功能具有重要意义.因此, 植物元素含量以及化学计量关系变化的驱动机制一直是生态化学计量学领域重点关注的问题(Güsewell, 2004 ; Ågren, 2008 ; He et al ., 2008 ).Elser等(2000)认为元素的计量学变异范围与元素含量有关, 越是大量元素, 其含量也越为稳定.而Han等(2011)研究认为元素含量的变异范围与该元素的限制情况有关, 越是限制性元素, 其含量变化越小.Karimi和Folt (2006)研究认为相对于大量元素, 微量元素很不稳定.事实上, 植物叶片中这些元素变化之间具有一定的耦合性, 且部分元素的变化速率高于其他元素(闫帮国等, 2012 ).然而, 植物元素的变化受到何种因素驱动却一直没有定论, 植物元素含量及其计量比与生物量变化的关系也很少受到关注.本研究的结果并不否定前人的研究结果, 而是从生物量变化的层面反映和探讨了植物化学计量学特征变化的驱动机制, 表明植物体元素的变异与生物量变化有关. ...
Plant stoichiometry at different scales: Element concentration patterns reflect environment more than genotype.
1
2012
... 水分、养分可以影响植物化学计量学特征(Ågren & Weih, 2012 ; Lü et al ., 2012 ), 但这种影响背后的生物量因素却很少受到重视.本研究还发现, 生物量是这些元素的计量比变化的主要影响因子, 而环境条件(这里为水分处理、养分处理以及二者的复合处理)和物种因素的影响很小, 表明水分和养分对植物化学计量学特征的影响作用具有共同的驱动因素, 即生物量变化.本研究还发现元素化学计量学特征的变化与生物量变化的关系在根茎叶中都存在, 说明这可能是个普遍现象. ...
Plant Litter: Decomposition, Humus Formation, Carbon Sequestration. 2nd edn. Springer-Verlag, Berlin
1
2003
... 植物生长能力与植物体元素计量比的变化可以对生态系统的生物地球化学循环造成深远的影响(Jobbágy & Jackson, 2004 ; Mooshammer et al ., 2012 ).微量元素和大量元素一样, 是生态系统中物质循环的重要因素, 一些元素是凋落物分解(Berg et al ., 2007 )、动物生存(Karimi & Folt, 2006 )的重要限制因素, 其作用甚至会超过大量元素.如Mn是一些生态系统凋落物分解的限制因子(Berg & Mcclaugherty, 2003 ), 而其他的微量元素也可能在其中起重要作用(Kaspari et al ., 2008 ), 从而影响碎屑食物链的物质和能量的传输效率.大量元素与微量元素计量比的增大极有可能促进微量元素的限制作用. ...
Litter decom- position rate is dependent on litter Mn concentrations.
1
2007
... 植物生长能力与植物体元素计量比的变化可以对生态系统的生物地球化学循环造成深远的影响(Jobbágy & Jackson, 2004 ; Mooshammer et al ., 2012 ).微量元素和大量元素一样, 是生态系统中物质循环的重要因素, 一些元素是凋落物分解(Berg et al ., 2007 )、动物生存(Karimi & Folt, 2006 )的重要限制因素, 其作用甚至会超过大量元素.如Mn是一些生态系统凋落物分解的限制因子(Berg & Mcclaugherty, 2003 ), 而其他的微量元素也可能在其中起重要作用(Kaspari et al ., 2008 ), 从而影响碎屑食物链的物质和能量的传输效率.大量元素与微量元素计量比的增大极有可能促进微量元素的限制作用. ...
氮肥添加对青藏高原高寒草甸6个群落优势种生态化学计量学特征的影响
1
2014
... 植物化学计量学特征在植物生活史(Wright et al ., 2004 )、功能性状(He et al ., 2009 )以及植物死后的生态系统过程(Güsewell & Gessner, 2009 )中扮演极为重要的角色.N、P、K是各类生态系统中限制植物生长的关键元素(Koerselman & Meuleman, 1996 ; Olde Venterink et al ., 2003 ; Harpole et al ., 2011 ), 其在植物体内的含量与植物生长速率具有直接的关联性(Ågren, 2008 ).这些元素的计量学特征对植物生长(阎恩荣等, 2008 ; 刘兴诏等, 2010 ; 宾振钧等, 2014 )、生态系统功能的发挥(Yu et al ., 2010 )具有极为重要的作用.然而, 在生态系统功能中具有重要作用的元素并不仅限于N、P、K, 其他元素如Ca、Mg、Mn以及其他微量元素在植物生长和凋落物分解过程中具有举足轻重的地位(Makkonen et al ., 2012 ; 闫帮国等, 2013 ), 进而可以对土壤或者地表碳库产生重要的影响(Reich et al ., 2005 ).然而迄今为止, 对这些元素的稳定性和变异性的了解却很少(贺金生和韩兴国, 2010 ), 对这些元素计量关系的驱动因素也不清楚. ...
氮肥添加对青藏高原高寒草甸6个群落优势种生态化学计量学特征的影响
1
2014
... 植物化学计量学特征在植物生活史(Wright et al ., 2004 )、功能性状(He et al ., 2009 )以及植物死后的生态系统过程(Güsewell & Gessner, 2009 )中扮演极为重要的角色.N、P、K是各类生态系统中限制植物生长的关键元素(Koerselman & Meuleman, 1996 ; Olde Venterink et al ., 2003 ; Harpole et al ., 2011 ), 其在植物体内的含量与植物生长速率具有直接的关联性(Ågren, 2008 ).这些元素的计量学特征对植物生长(阎恩荣等, 2008 ; 刘兴诏等, 2010 ; 宾振钧等, 2014 )、生态系统功能的发挥(Yu et al ., 2010 )具有极为重要的作用.然而, 在生态系统功能中具有重要作用的元素并不仅限于N、P、K, 其他元素如Ca、Mg、Mn以及其他微量元素在植物生长和凋落物分解过程中具有举足轻重的地位(Makkonen et al ., 2012 ; 闫帮国等, 2013 ), 进而可以对土壤或者地表碳库产生重要的影响(Reich et al ., 2005 ).然而迄今为止, 对这些元素的稳定性和变异性的了解却很少(贺金生和韩兴国, 2010 ), 对这些元素计量关系的驱动因素也不清楚. ...
Species distributions in response to individual soil nutrients and seasonal drought across a community of tropical trees.
1
2013
... 水分和养分对植物化学计量学特征的影响必将作用于植物的生态系统功能.但水分和养分对植物化学计量学特征的影响与生物量变化有关, 生物量对水分或养分响应强烈的物种, 其化学计量学特征响应也强烈.因此, 这一切主要取决于物种、水分和养分的互作效应对生物量的作用.植物物种对水分、养分变化的差异性响应对干热地区生态系统的功能变化可能尤为重要.干热河谷地区的水分和养分都较为稀缺.一些植物的生物量对降水增加较为敏感, 另外一部分物种生物量对养分增加较为敏感.这些差异对物种在水分和养分条件不同的斑块中的分布具有重要作用(Condit et al ., 2013 ), 但是也会对植物化学计量学产生影响.水分和养分在提高生物量的同时, 也增加了大量元素与微量元素的计量比. ...
Nutritional genomics: Manipulating plant micronutrients to improve human health.
1
1999
... 目前还不了解元素计量特征与生物量变化的关系是否在人工作物中也普遍存在.高强度的水肥投入使得作物产量大幅度提升, 这是否会影响农产品的元素计量特征以及人类的健康(Dellapenna, 1999 )还缺乏研究. ...
Growth rate-stoichiometry couplings in diverse biota.
1
2003
... 在全球变化的背景下, 植物的化学计量学与生产力对环境变化的响应无疑会对生物群落乃至生态系统功能变迁(Sistla & Schimel, 2012 )起到重要的级联效应.各种植物的化学计量学特征是植物在各自生境下适应环境的结果, 也与植物生长速率等重要特性紧密关联(Elser et al ., 2000 , 2003 ; Sterner & Elser, 2002 ).然而, 植物的化学计量学特征具有很强的可塑性(Güsewell, 2004 ), 这种可塑性对环境因素的响应可能直接影响植物的生长能力和对环境的适应能力(Yu et al ., 2010 ; Sistla & Schimel, 2012 ).因此, 阐明植物化学计量学变化的驱动机制有利于理解植物在全球变化背景下的适应机制和植物的生态系统功能(Elser et al ., 2010 ). ...
Biological stoichiometry of plant production: Metabolism, scaling and ecological response to global change.
1
2010
... 在全球变化的背景下, 植物的化学计量学与生产力对环境变化的响应无疑会对生物群落乃至生态系统功能变迁(Sistla & Schimel, 2012 )起到重要的级联效应.各种植物的化学计量学特征是植物在各自生境下适应环境的结果, 也与植物生长速率等重要特性紧密关联(Elser et al ., 2000 , 2003 ; Sterner & Elser, 2002 ).然而, 植物的化学计量学特征具有很强的可塑性(Güsewell, 2004 ), 这种可塑性对环境因素的响应可能直接影响植物的生长能力和对环境的适应能力(Yu et al ., 2010 ; Sistla & Schimel, 2012 ).因此, 阐明植物化学计量学变化的驱动机制有利于理解植物在全球变化背景下的适应机制和植物的生态系统功能(Elser et al ., 2010 ). ...
Biological stoichiometry from genes to ecosystems.
1
2000
... 在全球变化的背景下, 植物的化学计量学与生产力对环境变化的响应无疑会对生物群落乃至生态系统功能变迁(Sistla & Schimel, 2012 )起到重要的级联效应.各种植物的化学计量学特征是植物在各自生境下适应环境的结果, 也与植物生长速率等重要特性紧密关联(Elser et al ., 2000 , 2003 ; Sterner & Elser, 2002 ).然而, 植物的化学计量学特征具有很强的可塑性(Güsewell, 2004 ), 这种可塑性对环境因素的响应可能直接影响植物的生长能力和对环境的适应能力(Yu et al ., 2010 ; Sistla & Schimel, 2012 ).因此, 阐明植物化学计量学变化的驱动机制有利于理解植物在全球变化背景下的适应机制和植物的生态系统功能(Elser et al ., 2010 ). ...
N:P ratios in terrestrial plants: Variation and functional significance.
2
2004
... 在全球变化的背景下, 植物的化学计量学与生产力对环境变化的响应无疑会对生物群落乃至生态系统功能变迁(Sistla & Schimel, 2012 )起到重要的级联效应.各种植物的化学计量学特征是植物在各自生境下适应环境的结果, 也与植物生长速率等重要特性紧密关联(Elser et al ., 2000 , 2003 ; Sterner & Elser, 2002 ).然而, 植物的化学计量学特征具有很强的可塑性(Güsewell, 2004 ), 这种可塑性对环境因素的响应可能直接影响植物的生长能力和对环境的适应能力(Yu et al ., 2010 ; Sistla & Schimel, 2012 ).因此, 阐明植物化学计量学变化的驱动机制有利于理解植物在全球变化背景下的适应机制和植物的生态系统功能(Elser et al ., 2010 ). ...
... 植物化学计量学特征与生物量都是生态系统功能的主要载体, 认识这二者之间的变化关系对于理解全球变化背景下生态系统功能具有重要意义.因此, 植物元素含量以及化学计量关系变化的驱动机制一直是生态化学计量学领域重点关注的问题(Güsewell, 2004 ; Ågren, 2008 ; He et al ., 2008 ).Elser等(2000)认为元素的计量学变异范围与元素含量有关, 越是大量元素, 其含量也越为稳定.而Han等(2011)研究认为元素含量的变异范围与该元素的限制情况有关, 越是限制性元素, 其含量变化越小.Karimi和Folt (2006)研究认为相对于大量元素, 微量元素很不稳定.事实上, 植物叶片中这些元素变化之间具有一定的耦合性, 且部分元素的变化速率高于其他元素(闫帮国等, 2012 ).然而, 植物元素的变化受到何种因素驱动却一直没有定论, 植物元素含量及其计量比与生物量变化的关系也很少受到关注.本研究的结果并不否定前人的研究结果, 而是从生物量变化的层面反映和探讨了植物化学计量学特征变化的驱动机制, 表明植物体元素的变异与生物量变化有关. ...
N:P ratios influence litter decomposition and colonization by fungi and bacteria in microcosms.
1
2009
... 植物化学计量学特征在植物生活史(Wright et al ., 2004 )、功能性状(He et al ., 2009 )以及植物死后的生态系统过程(Güsewell & Gessner, 2009 )中扮演极为重要的角色.N、P、K是各类生态系统中限制植物生长的关键元素(Koerselman & Meuleman, 1996 ; Olde Venterink et al ., 2003 ; Harpole et al ., 2011 ), 其在植物体内的含量与植物生长速率具有直接的关联性(Ågren, 2008 ).这些元素的计量学特征对植物生长(阎恩荣等, 2008 ; 刘兴诏等, 2010 ; 宾振钧等, 2014 )、生态系统功能的发挥(Yu et al ., 2010 )具有极为重要的作用.然而, 在生态系统功能中具有重要作用的元素并不仅限于N、P、K, 其他元素如Ca、Mg、Mn以及其他微量元素在植物生长和凋落物分解过程中具有举足轻重的地位(Makkonen et al ., 2012 ; 闫帮国等, 2013 ), 进而可以对土壤或者地表碳库产生重要的影响(Reich et al ., 2005 ).然而迄今为止, 对这些元素的稳定性和变异性的了解却很少(贺金生和韩兴国, 2010 ), 对这些元素计量关系的驱动因素也不清楚. ...
Biogeography and variability of eleven mineral elements in plant leaves across gradients of climate, soil and plant functional type in China.
0
2011
Nutrient co-limitation of primary producer communities.
1
2011
... 植物化学计量学特征在植物生活史(Wright et al ., 2004 )、功能性状(He et al ., 2009 )以及植物死后的生态系统过程(Güsewell & Gessner, 2009 )中扮演极为重要的角色.N、P、K是各类生态系统中限制植物生长的关键元素(Koerselman & Meuleman, 1996 ; Olde Venterink et al ., 2003 ; Harpole et al ., 2011 ), 其在植物体内的含量与植物生长速率具有直接的关联性(Ågren, 2008 ).这些元素的计量学特征对植物生长(阎恩荣等, 2008 ; 刘兴诏等, 2010 ; 宾振钧等, 2014 )、生态系统功能的发挥(Yu et al ., 2010 )具有极为重要的作用.然而, 在生态系统功能中具有重要作用的元素并不仅限于N、P、K, 其他元素如Ca、Mg、Mn以及其他微量元素在植物生长和凋落物分解过程中具有举足轻重的地位(Makkonen et al ., 2012 ; 闫帮国等, 2013 ), 进而可以对土壤或者地表碳库产生重要的影响(Reich et al ., 2005 ).然而迄今为止, 对这些元素的稳定性和变异性的了解却很少(贺金生和韩兴国, 2010 ), 对这些元素计量关系的驱动因素也不清楚. ...
Ecosystem responses to water and nitrogen amendment in a California grassland.
1
2007
... 众所周知, 植物生产力是生态系统功能的主体因子之一, 植物元素计量关系变化与植物生产力变化的耦合必然对生态系统过程和功能产生重要影响.其中养分和水分是影响植物生长生产力的最重要因素.然而养分和水分对植物生长的促进作用极为复杂, 可能存在拮抗效应、协同效应、加和效应等(Harpole et al ., 2007 ; Niu et al ., 2009 ; Lü et al ., 2014 ).有关养分和水分的复杂作用对植物元素平衡影响的研究很少(Lü et al ., 2012 ), 而大量元素与微量元素的计量关系研究更是缺乏, 这限制了我们对全球变化背景下生态系统功能的预测.生物体生长稀释理论(somatic growth dilution)认为重金属在生物体内的富集与生物生长速率有关(Karimi et al ., 2007 ).在生物体快速生长时, 其吸收的重金属可以被稀释(Sunda & Huntsman, 1998 ).然而植物生长快慢对微量元素的影响, 特别是大量元素与微量元素计量比的影响还不清楚.本研究以金沙江干热河谷地区的土壤和植物为实验材料, 研究水分和养分处理对植物生物量的作用, 并探索生物量变化与植物化学计量学特征变化的关联.研究结果将有助于揭示该地区植物化学计量学特征对水分和养分变化的响应机制. ...
生态化学计量学: 探索从个体到生态系统的统一化理论
1
2010
... 植物化学计量学特征在植物生活史(Wright et al ., 2004 )、功能性状(He et al ., 2009 )以及植物死后的生态系统过程(Güsewell & Gessner, 2009 )中扮演极为重要的角色.N、P、K是各类生态系统中限制植物生长的关键元素(Koerselman & Meuleman, 1996 ; Olde Venterink et al ., 2003 ; Harpole et al ., 2011 ), 其在植物体内的含量与植物生长速率具有直接的关联性(Ågren, 2008 ).这些元素的计量学特征对植物生长(阎恩荣等, 2008 ; 刘兴诏等, 2010 ; 宾振钧等, 2014 )、生态系统功能的发挥(Yu et al ., 2010 )具有极为重要的作用.然而, 在生态系统功能中具有重要作用的元素并不仅限于N、P、K, 其他元素如Ca、Mg、Mn以及其他微量元素在植物生长和凋落物分解过程中具有举足轻重的地位(Makkonen et al ., 2012 ; 闫帮国等, 2013 ), 进而可以对土壤或者地表碳库产生重要的影响(Reich et al ., 2005 ).然而迄今为止, 对这些元素的稳定性和变异性的了解却很少(贺金生和韩兴国, 2010 ), 对这些元素计量关系的驱动因素也不清楚. ...
生态化学计量学: 探索从个体到生态系统的统一化理论
1
2010
... 植物化学计量学特征在植物生活史(Wright et al ., 2004 )、功能性状(He et al ., 2009 )以及植物死后的生态系统过程(Güsewell & Gessner, 2009 )中扮演极为重要的角色.N、P、K是各类生态系统中限制植物生长的关键元素(Koerselman & Meuleman, 1996 ; Olde Venterink et al ., 2003 ; Harpole et al ., 2011 ), 其在植物体内的含量与植物生长速率具有直接的关联性(Ågren, 2008 ).这些元素的计量学特征对植物生长(阎恩荣等, 2008 ; 刘兴诏等, 2010 ; 宾振钧等, 2014 )、生态系统功能的发挥(Yu et al ., 2010 )具有极为重要的作用.然而, 在生态系统功能中具有重要作用的元素并不仅限于N、P、K, 其他元素如Ca、Mg、Mn以及其他微量元素在植物生长和凋落物分解过程中具有举足轻重的地位(Makkonen et al ., 2012 ; 闫帮国等, 2013 ), 进而可以对土壤或者地表碳库产生重要的影响(Reich et al ., 2005 ).然而迄今为止, 对这些元素的稳定性和变异性的了解却很少(贺金生和韩兴国, 2010 ), 对这些元素计量关系的驱动因素也不清楚. ...
Leaf nitrogen: Phosphorus stoichiometry across Chinese grassland biomes.
1
2008
... 植物化学计量学特征与生物量都是生态系统功能的主要载体, 认识这二者之间的变化关系对于理解全球变化背景下生态系统功能具有重要意义.因此, 植物元素含量以及化学计量关系变化的驱动机制一直是生态化学计量学领域重点关注的问题(Güsewell, 2004 ; Ågren, 2008 ; He et al ., 2008 ).Elser等(2000)认为元素的计量学变异范围与元素含量有关, 越是大量元素, 其含量也越为稳定.而Han等(2011)研究认为元素含量的变异范围与该元素的限制情况有关, 越是限制性元素, 其含量变化越小.Karimi和Folt (2006)研究认为相对于大量元素, 微量元素很不稳定.事实上, 植物叶片中这些元素变化之间具有一定的耦合性, 且部分元素的变化速率高于其他元素(闫帮国等, 2012 ).然而, 植物元素的变化受到何种因素驱动却一直没有定论, 植物元素含量及其计量比与生物量变化的关系也很少受到关注.本研究的结果并不否定前人的研究结果, 而是从生物量变化的层面反映和探讨了植物化学计量学特征变化的驱动机制, 表明植物体元素的变异与生物量变化有关. ...
Taxonomic, phylogenetic, and environmental trade-offs between leaf productivity and persistence.
1
2009
... 植物化学计量学特征在植物生活史(Wright et al ., 2004 )、功能性状(He et al ., 2009 )以及植物死后的生态系统过程(Güsewell & Gessner, 2009 )中扮演极为重要的角色.N、P、K是各类生态系统中限制植物生长的关键元素(Koerselman & Meuleman, 1996 ; Olde Venterink et al ., 2003 ; Harpole et al ., 2011 ), 其在植物体内的含量与植物生长速率具有直接的关联性(Ågren, 2008 ).这些元素的计量学特征对植物生长(阎恩荣等, 2008 ; 刘兴诏等, 2010 ; 宾振钧等, 2014 )、生态系统功能的发挥(Yu et al ., 2010 )具有极为重要的作用.然而, 在生态系统功能中具有重要作用的元素并不仅限于N、P、K, 其他元素如Ca、Mg、Mn以及其他微量元素在植物生长和凋落物分解过程中具有举足轻重的地位(Makkonen et al ., 2012 ; 闫帮国等, 2013 ), 进而可以对土壤或者地表碳库产生重要的影响(Reich et al ., 2005 ).然而迄今为止, 对这些元素的稳定性和变异性的了解却很少(贺金生和韩兴国, 2010 ), 对这些元素计量关系的驱动因素也不清楚. ...
The uplift of soil nutrients by plants: Biogeochemical consequences across scales.
1
2004
... 植物生长能力与植物体元素计量比的变化可以对生态系统的生物地球化学循环造成深远的影响(Jobbágy & Jackson, 2004 ; Mooshammer et al ., 2012 ).微量元素和大量元素一样, 是生态系统中物质循环的重要因素, 一些元素是凋落物分解(Berg et al ., 2007 )、动物生存(Karimi & Folt, 2006 )的重要限制因素, 其作用甚至会超过大量元素.如Mn是一些生态系统凋落物分解的限制因子(Berg & Mcclaugherty, 2003 ), 而其他的微量元素也可能在其中起重要作用(Kaspari et al ., 2008 ), 从而影响碎屑食物链的物质和能量的传输效率.大量元素与微量元素计量比的增大极有可能促进微量元素的限制作用. ...
Stoichiometric controls of mercury dilution by growth.
2
2007
... 众所周知, 植物生产力是生态系统功能的主体因子之一, 植物元素计量关系变化与植物生产力变化的耦合必然对生态系统过程和功能产生重要影响.其中养分和水分是影响植物生长生产力的最重要因素.然而养分和水分对植物生长的促进作用极为复杂, 可能存在拮抗效应、协同效应、加和效应等(Harpole et al ., 2007 ; Niu et al ., 2009 ; Lü et al ., 2014 ).有关养分和水分的复杂作用对植物元素平衡影响的研究很少(Lü et al ., 2012 ), 而大量元素与微量元素的计量关系研究更是缺乏, 这限制了我们对全球变化背景下生态系统功能的预测.生物体生长稀释理论(somatic growth dilution)认为重金属在生物体内的富集与生物生长速率有关(Karimi et al ., 2007 ).在生物体快速生长时, 其吸收的重金属可以被稀释(Sunda & Huntsman, 1998 ).然而植物生长快慢对微量元素的影响, 特别是大量元素与微量元素计量比的影响还不清楚.本研究以金沙江干热河谷地区的土壤和植物为实验材料, 研究水分和养分处理对植物生物量的作用, 并探索生物量变化与植物化学计量学特征变化的关联.研究结果将有助于揭示该地区植物化学计量学特征对水分和养分变化的响应机制. ...
... 虽然微量元素是生物体必需的元素, 然而相对于大量元素, 其在生长中的限制作用可能较小(Karimi & Folt, 2006 ).可能如同重金属一样, 在植物快速生长中被大量稀释, 形成稀释效应(Karimi et al ., 2007 ).本研究中微量元素含量变化与生物量变化成反比的关系与上述研究结果相一致(表3 ).相反, 大量元素在生物体中的生物化学功能极为重要, 其具有相对较高的内稳性(Sterner & Elser, 2002 ), 因此, 在生物量变化中必须被不断补充, 从而保持稳定性.此外, 水分和养分可能影响了植物对大量元素的吸收, 促进了植物生长, 在保持大量元素较为稳定的同时稀释了微量元素.这种变化格局可能最终导致大量元素与微量元素的计量关系随生物量增长而增长.N、P含量及其与其他元素的计量比与生物量变化的关系并不显著, 具体的原因还不清楚.N、P在土壤中的相当一部分形态是有机物, 因此其吸收取决于植物与微生物的关系(van der Heijden et al ., 2008 ), 而其他元素主要以无机物形态存在, 其吸收主要取决于植物根系.这种差异可能造成了N、P与其他元素在生物生长中受到了不同因素的影响, 其计量关系因而没有呈现出显著的变化格局. ...
Beyond macronutrients: Element variability and multielement stoichiometry in freshwater invertebrates.
2
2006
... 虽然微量元素是生物体必需的元素, 然而相对于大量元素, 其在生长中的限制作用可能较小(Karimi & Folt, 2006 ).可能如同重金属一样, 在植物快速生长中被大量稀释, 形成稀释效应(Karimi et al ., 2007 ).本研究中微量元素含量变化与生物量变化成反比的关系与上述研究结果相一致(表3 ).相反, 大量元素在生物体中的生物化学功能极为重要, 其具有相对较高的内稳性(Sterner & Elser, 2002 ), 因此, 在生物量变化中必须被不断补充, 从而保持稳定性.此外, 水分和养分可能影响了植物对大量元素的吸收, 促进了植物生长, 在保持大量元素较为稳定的同时稀释了微量元素.这种变化格局可能最终导致大量元素与微量元素的计量关系随生物量增长而增长.N、P含量及其与其他元素的计量比与生物量变化的关系并不显著, 具体的原因还不清楚.N、P在土壤中的相当一部分形态是有机物, 因此其吸收取决于植物与微生物的关系(van der Heijden et al ., 2008 ), 而其他元素主要以无机物形态存在, 其吸收主要取决于植物根系.这种差异可能造成了N、P与其他元素在生物生长中受到了不同因素的影响, 其计量关系因而没有呈现出显著的变化格局. ...
... 植物生长能力与植物体元素计量比的变化可以对生态系统的生物地球化学循环造成深远的影响(Jobbágy & Jackson, 2004 ; Mooshammer et al ., 2012 ).微量元素和大量元素一样, 是生态系统中物质循环的重要因素, 一些元素是凋落物分解(Berg et al ., 2007 )、动物生存(Karimi & Folt, 2006 )的重要限制因素, 其作用甚至会超过大量元素.如Mn是一些生态系统凋落物分解的限制因子(Berg & Mcclaugherty, 2003 ), 而其他的微量元素也可能在其中起重要作用(Kaspari et al ., 2008 ), 从而影响碎屑食物链的物质和能量的传输效率.大量元素与微量元素计量比的增大极有可能促进微量元素的限制作用. ...
Multiple nutrients limit litterfall and decomposition in a tropical forest.
1
2008
... 植物生长能力与植物体元素计量比的变化可以对生态系统的生物地球化学循环造成深远的影响(Jobbágy & Jackson, 2004 ; Mooshammer et al ., 2012 ).微量元素和大量元素一样, 是生态系统中物质循环的重要因素, 一些元素是凋落物分解(Berg et al ., 2007 )、动物生存(Karimi & Folt, 2006 )的重要限制因素, 其作用甚至会超过大量元素.如Mn是一些生态系统凋落物分解的限制因子(Berg & Mcclaugherty, 2003 ), 而其他的微量元素也可能在其中起重要作用(Kaspari et al ., 2008 ), 从而影响碎屑食物链的物质和能量的传输效率.大量元素与微量元素计量比的增大极有可能促进微量元素的限制作用. ...
The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation.
1
1996
... 植物化学计量学特征在植物生活史(Wright et al ., 2004 )、功能性状(He et al ., 2009 )以及植物死后的生态系统过程(Güsewell & Gessner, 2009 )中扮演极为重要的角色.N、P、K是各类生态系统中限制植物生长的关键元素(Koerselman & Meuleman, 1996 ; Olde Venterink et al ., 2003 ; Harpole et al ., 2011 ), 其在植物体内的含量与植物生长速率具有直接的关联性(Ågren, 2008 ).这些元素的计量学特征对植物生长(阎恩荣等, 2008 ; 刘兴诏等, 2010 ; 宾振钧等, 2014 )、生态系统功能的发挥(Yu et al ., 2010 )具有极为重要的作用.然而, 在生态系统功能中具有重要作用的元素并不仅限于N、P、K, 其他元素如Ca、Mg、Mn以及其他微量元素在植物生长和凋落物分解过程中具有举足轻重的地位(Makkonen et al ., 2012 ; 闫帮国等, 2013 ), 进而可以对土壤或者地表碳库产生重要的影响(Reich et al ., 2005 ).然而迄今为止, 对这些元素的稳定性和变异性的了解却很少(贺金生和韩兴国, 2010 ), 对这些元素计量关系的驱动因素也不清楚. ...
南亚热带森林不同演替阶段植物与土壤中N、P的化学计量特征
1
2010
... 植物化学计量学特征在植物生活史(Wright et al ., 2004 )、功能性状(He et al ., 2009 )以及植物死后的生态系统过程(Güsewell & Gessner, 2009 )中扮演极为重要的角色.N、P、K是各类生态系统中限制植物生长的关键元素(Koerselman & Meuleman, 1996 ; Olde Venterink et al ., 2003 ; Harpole et al ., 2011 ), 其在植物体内的含量与植物生长速率具有直接的关联性(Ågren, 2008 ).这些元素的计量学特征对植物生长(阎恩荣等, 2008 ; 刘兴诏等, 2010 ; 宾振钧等, 2014 )、生态系统功能的发挥(Yu et al ., 2010 )具有极为重要的作用.然而, 在生态系统功能中具有重要作用的元素并不仅限于N、P、K, 其他元素如Ca、Mg、Mn以及其他微量元素在植物生长和凋落物分解过程中具有举足轻重的地位(Makkonen et al ., 2012 ; 闫帮国等, 2013 ), 进而可以对土壤或者地表碳库产生重要的影响(Reich et al ., 2005 ).然而迄今为止, 对这些元素的稳定性和变异性的了解却很少(贺金生和韩兴国, 2010 ), 对这些元素计量关系的驱动因素也不清楚. ...
南亚热带森林不同演替阶段植物与土壤中N、P的化学计量特征
1
2010
... 植物化学计量学特征在植物生活史(Wright et al ., 2004 )、功能性状(He et al ., 2009 )以及植物死后的生态系统过程(Güsewell & Gessner, 2009 )中扮演极为重要的角色.N、P、K是各类生态系统中限制植物生长的关键元素(Koerselman & Meuleman, 1996 ; Olde Venterink et al ., 2003 ; Harpole et al ., 2011 ), 其在植物体内的含量与植物生长速率具有直接的关联性(Ågren, 2008 ).这些元素的计量学特征对植物生长(阎恩荣等, 2008 ; 刘兴诏等, 2010 ; 宾振钧等, 2014 )、生态系统功能的发挥(Yu et al ., 2010 )具有极为重要的作用.然而, 在生态系统功能中具有重要作用的元素并不仅限于N、P、K, 其他元素如Ca、Mg、Mn以及其他微量元素在植物生长和凋落物分解过程中具有举足轻重的地位(Makkonen et al ., 2012 ; 闫帮国等, 2013 ), 进而可以对土壤或者地表碳库产生重要的影响(Reich et al ., 2005 ).然而迄今为止, 对这些元素的稳定性和变异性的了解却很少(贺金生和韩兴国, 2010 ), 对这些元素计量关系的驱动因素也不清楚. ...
Plant nitrogen uptake drives responses of productivity to nitrogen and water addition in a grassland.
1
2014
... 众所周知, 植物生产力是生态系统功能的主体因子之一, 植物元素计量关系变化与植物生产力变化的耦合必然对生态系统过程和功能产生重要影响.其中养分和水分是影响植物生长生产力的最重要因素.然而养分和水分对植物生长的促进作用极为复杂, 可能存在拮抗效应、协同效应、加和效应等(Harpole et al ., 2007 ; Niu et al ., 2009 ; Lü et al ., 2014 ).有关养分和水分的复杂作用对植物元素平衡影响的研究很少(Lü et al ., 2012 ), 而大量元素与微量元素的计量关系研究更是缺乏, 这限制了我们对全球变化背景下生态系统功能的预测.生物体生长稀释理论(somatic growth dilution)认为重金属在生物体内的富集与生物生长速率有关(Karimi et al ., 2007 ).在生物体快速生长时, 其吸收的重金属可以被稀释(Sunda & Huntsman, 1998 ).然而植物生长快慢对微量元素的影响, 特别是大量元素与微量元素计量比的影响还不清楚.本研究以金沙江干热河谷地区的土壤和植物为实验材料, 研究水分和养分处理对植物生物量的作用, 并探索生物量变化与植物化学计量学特征变化的关联.研究结果将有助于揭示该地区植物化学计量学特征对水分和养分变化的响应机制. ...
Nitrogen and water availability interact to affect leaf stoichiometry in a semi-arid grassland.
2
2012
... 众所周知, 植物生产力是生态系统功能的主体因子之一, 植物元素计量关系变化与植物生产力变化的耦合必然对生态系统过程和功能产生重要影响.其中养分和水分是影响植物生长生产力的最重要因素.然而养分和水分对植物生长的促进作用极为复杂, 可能存在拮抗效应、协同效应、加和效应等(Harpole et al ., 2007 ; Niu et al ., 2009 ; Lü et al ., 2014 ).有关养分和水分的复杂作用对植物元素平衡影响的研究很少(Lü et al ., 2012 ), 而大量元素与微量元素的计量关系研究更是缺乏, 这限制了我们对全球变化背景下生态系统功能的预测.生物体生长稀释理论(somatic growth dilution)认为重金属在生物体内的富集与生物生长速率有关(Karimi et al ., 2007 ).在生物体快速生长时, 其吸收的重金属可以被稀释(Sunda & Huntsman, 1998 ).然而植物生长快慢对微量元素的影响, 特别是大量元素与微量元素计量比的影响还不清楚.本研究以金沙江干热河谷地区的土壤和植物为实验材料, 研究水分和养分处理对植物生物量的作用, 并探索生物量变化与植物化学计量学特征变化的关联.研究结果将有助于揭示该地区植物化学计量学特征对水分和养分变化的响应机制. ...
... 水分、养分可以影响植物化学计量学特征(Ågren & Weih, 2012 ; Lü et al ., 2012 ), 但这种影响背后的生物量因素却很少受到重视.本研究还发现, 生物量是这些元素的计量比变化的主要影响因子, 而环境条件(这里为水分处理、养分处理以及二者的复合处理)和物种因素的影响很小, 表明水分和养分对植物化学计量学特征的影响作用具有共同的驱动因素, 即生物量变化.本研究还发现元素化学计量学特征的变化与生物量变化的关系在根茎叶中都存在, 说明这可能是个普遍现象. ...
Highly consistent effects of plant litter identity and functional traits on decomposition across a latitudinal gradient.
1
2012
... 植物化学计量学特征在植物生活史(Wright et al ., 2004 )、功能性状(He et al ., 2009 )以及植物死后的生态系统过程(Güsewell & Gessner, 2009 )中扮演极为重要的角色.N、P、K是各类生态系统中限制植物生长的关键元素(Koerselman & Meuleman, 1996 ; Olde Venterink et al ., 2003 ; Harpole et al ., 2011 ), 其在植物体内的含量与植物生长速率具有直接的关联性(Ågren, 2008 ).这些元素的计量学特征对植物生长(阎恩荣等, 2008 ; 刘兴诏等, 2010 ; 宾振钧等, 2014 )、生态系统功能的发挥(Yu et al ., 2010 )具有极为重要的作用.然而, 在生态系统功能中具有重要作用的元素并不仅限于N、P、K, 其他元素如Ca、Mg、Mn以及其他微量元素在植物生长和凋落物分解过程中具有举足轻重的地位(Makkonen et al ., 2012 ; 闫帮国等, 2013 ), 进而可以对土壤或者地表碳库产生重要的影响(Reich et al ., 2005 ).然而迄今为止, 对这些元素的稳定性和变异性的了解却很少(贺金生和韩兴国, 2010 ), 对这些元素计量关系的驱动因素也不清楚. ...
Stoichiometric controls of nitrogen and phosphorus cycling in decomposing beech leaf litter.
1
2012
... 植物生长能力与植物体元素计量比的变化可以对生态系统的生物地球化学循环造成深远的影响(Jobbágy & Jackson, 2004 ; Mooshammer et al ., 2012 ).微量元素和大量元素一样, 是生态系统中物质循环的重要因素, 一些元素是凋落物分解(Berg et al ., 2007 )、动物生存(Karimi & Folt, 2006 )的重要限制因素, 其作用甚至会超过大量元素.如Mn是一些生态系统凋落物分解的限制因子(Berg & Mcclaugherty, 2003 ), 而其他的微量元素也可能在其中起重要作用(Kaspari et al ., 2008 ), 从而影响碎屑食物链的物质和能量的传输效率.大量元素与微量元素计量比的增大极有可能促进微量元素的限制作用. ...
Non-additive effects of water and nitrogen addition on ecosystem carbon exchange in a temperate steppe.
1
2009
... 众所周知, 植物生产力是生态系统功能的主体因子之一, 植物元素计量关系变化与植物生产力变化的耦合必然对生态系统过程和功能产生重要影响.其中养分和水分是影响植物生长生产力的最重要因素.然而养分和水分对植物生长的促进作用极为复杂, 可能存在拮抗效应、协同效应、加和效应等(Harpole et al ., 2007 ; Niu et al ., 2009 ; Lü et al ., 2014 ).有关养分和水分的复杂作用对植物元素平衡影响的研究很少(Lü et al ., 2012 ), 而大量元素与微量元素的计量关系研究更是缺乏, 这限制了我们对全球变化背景下生态系统功能的预测.生物体生长稀释理论(somatic growth dilution)认为重金属在生物体内的富集与生物生长速率有关(Karimi et al ., 2007 ).在生物体快速生长时, 其吸收的重金属可以被稀释(Sunda & Huntsman, 1998 ).然而植物生长快慢对微量元素的影响, 特别是大量元素与微量元素计量比的影响还不清楚.本研究以金沙江干热河谷地区的土壤和植物为实验材料, 研究水分和养分处理对植物生物量的作用, 并探索生物量变化与植物化学计量学特征变化的关联.研究结果将有助于揭示该地区植物化学计量学特征对水分和养分变化的响应机制. ...
Species richness-productivity patterns differ between N-, P-, and K-limited wetlands.
1
2003
... 植物化学计量学特征在植物生活史(Wright et al ., 2004 )、功能性状(He et al ., 2009 )以及植物死后的生态系统过程(Güsewell & Gessner, 2009 )中扮演极为重要的角色.N、P、K是各类生态系统中限制植物生长的关键元素(Koerselman & Meuleman, 1996 ; Olde Venterink et al ., 2003 ; Harpole et al ., 2011 ), 其在植物体内的含量与植物生长速率具有直接的关联性(Ågren, 2008 ).这些元素的计量学特征对植物生长(阎恩荣等, 2008 ; 刘兴诏等, 2010 ; 宾振钧等, 2014 )、生态系统功能的发挥(Yu et al ., 2010 )具有极为重要的作用.然而, 在生态系统功能中具有重要作用的元素并不仅限于N、P、K, 其他元素如Ca、Mg、Mn以及其他微量元素在植物生长和凋落物分解过程中具有举足轻重的地位(Makkonen et al ., 2012 ; 闫帮国等, 2013 ), 进而可以对土壤或者地表碳库产生重要的影响(Reich et al ., 2005 ).然而迄今为止, 对这些元素的稳定性和变异性的了解却很少(贺金生和韩兴国, 2010 ), 对这些元素计量关系的驱动因素也不清楚. ...
Linking litter calcium, earthworms and soil properties: A common garden test with 14 tree species.
1
2005
... 植物化学计量学特征在植物生活史(Wright et al ., 2004 )、功能性状(He et al ., 2009 )以及植物死后的生态系统过程(Güsewell & Gessner, 2009 )中扮演极为重要的角色.N、P、K是各类生态系统中限制植物生长的关键元素(Koerselman & Meuleman, 1996 ; Olde Venterink et al ., 2003 ; Harpole et al ., 2011 ), 其在植物体内的含量与植物生长速率具有直接的关联性(Ågren, 2008 ).这些元素的计量学特征对植物生长(阎恩荣等, 2008 ; 刘兴诏等, 2010 ; 宾振钧等, 2014 )、生态系统功能的发挥(Yu et al ., 2010 )具有极为重要的作用.然而, 在生态系统功能中具有重要作用的元素并不仅限于N、P、K, 其他元素如Ca、Mg、Mn以及其他微量元素在植物生长和凋落物分解过程中具有举足轻重的地位(Makkonen et al ., 2012 ; 闫帮国等, 2013 ), 进而可以对土壤或者地表碳库产生重要的影响(Reich et al ., 2005 ).然而迄今为止, 对这些元素的稳定性和变异性的了解却很少(贺金生和韩兴国, 2010 ), 对这些元素计量关系的驱动因素也不清楚. ...
Stoichiometric flexibility as a regulator of carbon and nutrient cycling in terrestrial ecosystems under change.
2
2012
... 在全球变化的背景下, 植物的化学计量学与生产力对环境变化的响应无疑会对生物群落乃至生态系统功能变迁(Sistla & Schimel, 2012 )起到重要的级联效应.各种植物的化学计量学特征是植物在各自生境下适应环境的结果, 也与植物生长速率等重要特性紧密关联(Elser et al ., 2000 , 2003 ; Sterner & Elser, 2002 ).然而, 植物的化学计量学特征具有很强的可塑性(Güsewell, 2004 ), 这种可塑性对环境因素的响应可能直接影响植物的生长能力和对环境的适应能力(Yu et al ., 2010 ; Sistla & Schimel, 2012 ).因此, 阐明植物化学计量学变化的驱动机制有利于理解植物在全球变化背景下的适应机制和植物的生态系统功能(Elser et al ., 2010 ). ...
... ; Sistla & Schimel, 2012 ).因此, 阐明植物化学计量学变化的驱动机制有利于理解植物在全球变化背景下的适应机制和植物的生态系统功能(Elser et al ., 2010 ). ...
2
2002
... 在全球变化的背景下, 植物的化学计量学与生产力对环境变化的响应无疑会对生物群落乃至生态系统功能变迁(Sistla & Schimel, 2012 )起到重要的级联效应.各种植物的化学计量学特征是植物在各自生境下适应环境的结果, 也与植物生长速率等重要特性紧密关联(Elser et al ., 2000 , 2003 ; Sterner & Elser, 2002 ).然而, 植物的化学计量学特征具有很强的可塑性(Güsewell, 2004 ), 这种可塑性对环境因素的响应可能直接影响植物的生长能力和对环境的适应能力(Yu et al ., 2010 ; Sistla & Schimel, 2012 ).因此, 阐明植物化学计量学变化的驱动机制有利于理解植物在全球变化背景下的适应机制和植物的生态系统功能(Elser et al ., 2010 ). ...
... 虽然微量元素是生物体必需的元素, 然而相对于大量元素, 其在生长中的限制作用可能较小(Karimi & Folt, 2006 ).可能如同重金属一样, 在植物快速生长中被大量稀释, 形成稀释效应(Karimi et al ., 2007 ).本研究中微量元素含量变化与生物量变化成反比的关系与上述研究结果相一致(表3 ).相反, 大量元素在生物体中的生物化学功能极为重要, 其具有相对较高的内稳性(Sterner & Elser, 2002 ), 因此, 在生物量变化中必须被不断补充, 从而保持稳定性.此外, 水分和养分可能影响了植物对大量元素的吸收, 促进了植物生长, 在保持大量元素较为稳定的同时稀释了微量元素.这种变化格局可能最终导致大量元素与微量元素的计量关系随生物量增长而增长.N、P含量及其与其他元素的计量比与生物量变化的关系并不显著, 具体的原因还不清楚.N、P在土壤中的相当一部分形态是有机物, 因此其吸收取决于植物与微生物的关系(van der Heijden et al ., 2008 ), 而其他元素主要以无机物形态存在, 其吸收主要取决于植物根系.这种差异可能造成了N、P与其他元素在生物生长中受到了不同因素的影响, 其计量关系因而没有呈现出显著的变化格局. ...
Processes regulating cellular metal accumulation and physiological effects: Phytoplankton as model systems.
1
1998
... 众所周知, 植物生产力是生态系统功能的主体因子之一, 植物元素计量关系变化与植物生产力变化的耦合必然对生态系统过程和功能产生重要影响.其中养分和水分是影响植物生长生产力的最重要因素.然而养分和水分对植物生长的促进作用极为复杂, 可能存在拮抗效应、协同效应、加和效应等(Harpole et al ., 2007 ; Niu et al ., 2009 ; Lü et al ., 2014 ).有关养分和水分的复杂作用对植物元素平衡影响的研究很少(Lü et al ., 2012 ), 而大量元素与微量元素的计量关系研究更是缺乏, 这限制了我们对全球变化背景下生态系统功能的预测.生物体生长稀释理论(somatic growth dilution)认为重金属在生物体内的富集与生物生长速率有关(Karimi et al ., 2007 ).在生物体快速生长时, 其吸收的重金属可以被稀释(Sunda & Huntsman, 1998 ).然而植物生长快慢对微量元素的影响, 特别是大量元素与微量元素计量比的影响还不清楚.本研究以金沙江干热河谷地区的土壤和植物为实验材料, 研究水分和养分处理对植物生物量的作用, 并探索生物量变化与植物化学计量学特征变化的关联.研究结果将有助于揭示该地区植物化学计量学特征对水分和养分变化的响应机制. ...
The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems.
1
2008
... 虽然微量元素是生物体必需的元素, 然而相对于大量元素, 其在生长中的限制作用可能较小(Karimi & Folt, 2006 ).可能如同重金属一样, 在植物快速生长中被大量稀释, 形成稀释效应(Karimi et al ., 2007 ).本研究中微量元素含量变化与生物量变化成反比的关系与上述研究结果相一致(表3 ).相反, 大量元素在生物体中的生物化学功能极为重要, 其具有相对较高的内稳性(Sterner & Elser, 2002 ), 因此, 在生物量变化中必须被不断补充, 从而保持稳定性.此外, 水分和养分可能影响了植物对大量元素的吸收, 促进了植物生长, 在保持大量元素较为稳定的同时稀释了微量元素.这种变化格局可能最终导致大量元素与微量元素的计量关系随生物量增长而增长.N、P含量及其与其他元素的计量比与生物量变化的关系并不显著, 具体的原因还不清楚.N、P在土壤中的相当一部分形态是有机物, 因此其吸收取决于植物与微生物的关系(van der Heijden et al ., 2008 ), 而其他元素主要以无机物形态存在, 其吸收主要取决于植物根系.这种差异可能造成了N、P与其他元素在生物生长中受到了不同因素的影响, 其计量关系因而没有呈现出显著的变化格局. ...
The worldwide leaf economics spectrum.
1
2004
... 植物化学计量学特征在植物生活史(Wright et al ., 2004 )、功能性状(He et al ., 2009 )以及植物死后的生态系统过程(Güsewell & Gessner, 2009 )中扮演极为重要的角色.N、P、K是各类生态系统中限制植物生长的关键元素(Koerselman & Meuleman, 1996 ; Olde Venterink et al ., 2003 ; Harpole et al ., 2011 ), 其在植物体内的含量与植物生长速率具有直接的关联性(Ågren, 2008 ).这些元素的计量学特征对植物生长(阎恩荣等, 2008 ; 刘兴诏等, 2010 ; 宾振钧等, 2014 )、生态系统功能的发挥(Yu et al ., 2010 )具有极为重要的作用.然而, 在生态系统功能中具有重要作用的元素并不仅限于N、P、K, 其他元素如Ca、Mg、Mn以及其他微量元素在植物生长和凋落物分解过程中具有举足轻重的地位(Makkonen et al ., 2012 ; 闫帮国等, 2013 ), 进而可以对土壤或者地表碳库产生重要的影响(Reich et al ., 2005 ).然而迄今为止, 对这些元素的稳定性和变异性的了解却很少(贺金生和韩兴国, 2010 ), 对这些元素计量关系的驱动因素也不清楚. ...
金沙江干热河谷地区植物叶片中各生源要素的化学计量特征以及异速增长关系
1
2012
... 植物化学计量学特征与生物量都是生态系统功能的主要载体, 认识这二者之间的变化关系对于理解全球变化背景下生态系统功能具有重要意义.因此, 植物元素含量以及化学计量关系变化的驱动机制一直是生态化学计量学领域重点关注的问题(Güsewell, 2004 ; Ågren, 2008 ; He et al ., 2008 ).Elser等(2000)认为元素的计量学变异范围与元素含量有关, 越是大量元素, 其含量也越为稳定.而Han等(2011)研究认为元素含量的变异范围与该元素的限制情况有关, 越是限制性元素, 其含量变化越小.Karimi和Folt (2006)研究认为相对于大量元素, 微量元素很不稳定.事实上, 植物叶片中这些元素变化之间具有一定的耦合性, 且部分元素的变化速率高于其他元素(闫帮国等, 2012 ).然而, 植物元素的变化受到何种因素驱动却一直没有定论, 植物元素含量及其计量比与生物量变化的关系也很少受到关注.本研究的结果并不否定前人的研究结果, 而是从生物量变化的层面反映和探讨了植物化学计量学特征变化的驱动机制, 表明植物体元素的变异与生物量变化有关. ...
金沙江干热河谷地区植物叶片中各生源要素的化学计量特征以及异速增长关系
1
2012
... 植物化学计量学特征与生物量都是生态系统功能的主要载体, 认识这二者之间的变化关系对于理解全球变化背景下生态系统功能具有重要意义.因此, 植物元素含量以及化学计量关系变化的驱动机制一直是生态化学计量学领域重点关注的问题(Güsewell, 2004 ; Ågren, 2008 ; He et al ., 2008 ).Elser等(2000)认为元素的计量学变异范围与元素含量有关, 越是大量元素, 其含量也越为稳定.而Han等(2011)研究认为元素含量的变异范围与该元素的限制情况有关, 越是限制性元素, 其含量变化越小.Karimi和Folt (2006)研究认为相对于大量元素, 微量元素很不稳定.事实上, 植物叶片中这些元素变化之间具有一定的耦合性, 且部分元素的变化速率高于其他元素(闫帮国等, 2012 ).然而, 植物元素的变化受到何种因素驱动却一直没有定论, 植物元素含量及其计量比与生物量变化的关系也很少受到关注.本研究的结果并不否定前人的研究结果, 而是从生物量变化的层面反映和探讨了植物化学计量学特征变化的驱动机制, 表明植物体元素的变异与生物量变化有关. ...
金沙江干热河谷植物叶片元素含量在地表凋落物周转中的作用
1
2013
... 植物化学计量学特征在植物生活史(Wright et al ., 2004 )、功能性状(He et al ., 2009 )以及植物死后的生态系统过程(Güsewell & Gessner, 2009 )中扮演极为重要的角色.N、P、K是各类生态系统中限制植物生长的关键元素(Koerselman & Meuleman, 1996 ; Olde Venterink et al ., 2003 ; Harpole et al ., 2011 ), 其在植物体内的含量与植物生长速率具有直接的关联性(Ågren, 2008 ).这些元素的计量学特征对植物生长(阎恩荣等, 2008 ; 刘兴诏等, 2010 ; 宾振钧等, 2014 )、生态系统功能的发挥(Yu et al ., 2010 )具有极为重要的作用.然而, 在生态系统功能中具有重要作用的元素并不仅限于N、P、K, 其他元素如Ca、Mg、Mn以及其他微量元素在植物生长和凋落物分解过程中具有举足轻重的地位(Makkonen et al ., 2012 ; 闫帮国等, 2013 ), 进而可以对土壤或者地表碳库产生重要的影响(Reich et al ., 2005 ).然而迄今为止, 对这些元素的稳定性和变异性的了解却很少(贺金生和韩兴国, 2010 ), 对这些元素计量关系的驱动因素也不清楚. ...
金沙江干热河谷植物叶片元素含量在地表凋落物周转中的作用
1
2013
... 植物化学计量学特征在植物生活史(Wright et al ., 2004 )、功能性状(He et al ., 2009 )以及植物死后的生态系统过程(Güsewell & Gessner, 2009 )中扮演极为重要的角色.N、P、K是各类生态系统中限制植物生长的关键元素(Koerselman & Meuleman, 1996 ; Olde Venterink et al ., 2003 ; Harpole et al ., 2011 ), 其在植物体内的含量与植物生长速率具有直接的关联性(Ågren, 2008 ).这些元素的计量学特征对植物生长(阎恩荣等, 2008 ; 刘兴诏等, 2010 ; 宾振钧等, 2014 )、生态系统功能的发挥(Yu et al ., 2010 )具有极为重要的作用.然而, 在生态系统功能中具有重要作用的元素并不仅限于N、P、K, 其他元素如Ca、Mg、Mn以及其他微量元素在植物生长和凋落物分解过程中具有举足轻重的地位(Makkonen et al ., 2012 ; 闫帮国等, 2013 ), 进而可以对土壤或者地表碳库产生重要的影响(Reich et al ., 2005 ).然而迄今为止, 对这些元素的稳定性和变异性的了解却很少(贺金生和韩兴国, 2010 ), 对这些元素计量关系的驱动因素也不清楚. ...
天童常绿阔叶林演替系列植物群落的N:P化学计量特征
1
2008
... 植物化学计量学特征在植物生活史(Wright et al ., 2004 )、功能性状(He et al ., 2009 )以及植物死后的生态系统过程(Güsewell & Gessner, 2009 )中扮演极为重要的角色.N、P、K是各类生态系统中限制植物生长的关键元素(Koerselman & Meuleman, 1996 ; Olde Venterink et al ., 2003 ; Harpole et al ., 2011 ), 其在植物体内的含量与植物生长速率具有直接的关联性(Ågren, 2008 ).这些元素的计量学特征对植物生长(阎恩荣等, 2008 ; 刘兴诏等, 2010 ; 宾振钧等, 2014 )、生态系统功能的发挥(Yu et al ., 2010 )具有极为重要的作用.然而, 在生态系统功能中具有重要作用的元素并不仅限于N、P、K, 其他元素如Ca、Mg、Mn以及其他微量元素在植物生长和凋落物分解过程中具有举足轻重的地位(Makkonen et al ., 2012 ; 闫帮国等, 2013 ), 进而可以对土壤或者地表碳库产生重要的影响(Reich et al ., 2005 ).然而迄今为止, 对这些元素的稳定性和变异性的了解却很少(贺金生和韩兴国, 2010 ), 对这些元素计量关系的驱动因素也不清楚. ...
天童常绿阔叶林演替系列植物群落的N:P化学计量特征
1
2008
... 植物化学计量学特征在植物生活史(Wright et al ., 2004 )、功能性状(He et al ., 2009 )以及植物死后的生态系统过程(Güsewell & Gessner, 2009 )中扮演极为重要的角色.N、P、K是各类生态系统中限制植物生长的关键元素(Koerselman & Meuleman, 1996 ; Olde Venterink et al ., 2003 ; Harpole et al ., 2011 ), 其在植物体内的含量与植物生长速率具有直接的关联性(Ågren, 2008 ).这些元素的计量学特征对植物生长(阎恩荣等, 2008 ; 刘兴诏等, 2010 ; 宾振钧等, 2014 )、生态系统功能的发挥(Yu et al ., 2010 )具有极为重要的作用.然而, 在生态系统功能中具有重要作用的元素并不仅限于N、P、K, 其他元素如Ca、Mg、Mn以及其他微量元素在植物生长和凋落物分解过程中具有举足轻重的地位(Makkonen et al ., 2012 ; 闫帮国等, 2013 ), 进而可以对土壤或者地表碳库产生重要的影响(Reich et al ., 2005 ).然而迄今为止, 对这些元素的稳定性和变异性的了解却很少(贺金生和韩兴国, 2010 ), 对这些元素计量关系的驱动因素也不清楚. ...
Linking stoichiometric homoeostasis with ecosystem structure, functioning and stability.
2
2010
... 植物化学计量学特征在植物生活史(Wright et al ., 2004 )、功能性状(He et al ., 2009 )以及植物死后的生态系统过程(Güsewell & Gessner, 2009 )中扮演极为重要的角色.N、P、K是各类生态系统中限制植物生长的关键元素(Koerselman & Meuleman, 1996 ; Olde Venterink et al ., 2003 ; Harpole et al ., 2011 ), 其在植物体内的含量与植物生长速率具有直接的关联性(Ågren, 2008 ).这些元素的计量学特征对植物生长(阎恩荣等, 2008 ; 刘兴诏等, 2010 ; 宾振钧等, 2014 )、生态系统功能的发挥(Yu et al ., 2010 )具有极为重要的作用.然而, 在生态系统功能中具有重要作用的元素并不仅限于N、P、K, 其他元素如Ca、Mg、Mn以及其他微量元素在植物生长和凋落物分解过程中具有举足轻重的地位(Makkonen et al ., 2012 ; 闫帮国等, 2013 ), 进而可以对土壤或者地表碳库产生重要的影响(Reich et al ., 2005 ).然而迄今为止, 对这些元素的稳定性和变异性的了解却很少(贺金生和韩兴国, 2010 ), 对这些元素计量关系的驱动因素也不清楚. ...
... 在全球变化的背景下, 植物的化学计量学与生产力对环境变化的响应无疑会对生物群落乃至生态系统功能变迁(Sistla & Schimel, 2012 )起到重要的级联效应.各种植物的化学计量学特征是植物在各自生境下适应环境的结果, 也与植物生长速率等重要特性紧密关联(Elser et al ., 2000 , 2003 ; Sterner & Elser, 2002 ).然而, 植物的化学计量学特征具有很强的可塑性(Güsewell, 2004 ), 这种可塑性对环境因素的响应可能直接影响植物的生长能力和对环境的适应能力(Yu et al ., 2010 ; Sistla & Schimel, 2012 ).因此, 阐明植物化学计量学变化的驱动机制有利于理解植物在全球变化背景下的适应机制和植物的生态系统功能(Elser et al ., 2010 ). ...
元谋干热河谷区生态环境变迁探讨
1
2000
... 干热河谷地区以其特殊的气候而成为一个独特的地理单元.这其中尤以金沙江干热河谷元谋段最为典型.区域内年平均气温约为21.5 ℃, 降水量小于700 mm, 降水主要集中在雨季(6-10月间的降水量占90%以上) (张建平等, 2000 ). ...
元谋干热河谷区生态环境变迁探讨
1
2000
... 干热河谷地区以其特殊的气候而成为一个独特的地理单元.这其中尤以金沙江干热河谷元谋段最为典型.区域内年平均气温约为21.5 ℃, 降水量小于700 mm, 降水主要集中在雨季(6-10月间的降水量占90%以上) (张建平等, 2000 ). ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}