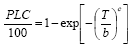A review of new research progress on the vulnerability of xylem embolism of woody plants
LI Rong1, *, JIANG Zai-Min2, *, ZHANG Shuo-Xin1, 3, CAI Jing1, 3, **,
1College of Forestry, Northwest A&F University, Yangling, Shaanxi 712100, China 2College of Life Sciences, Northwest A&F University, Yangling, Shaanxi 712100, China3Qinling National Forest Ecosystem Research Station, Yangling, Shaanxi 712100, China
Xylem cavitation/embolism is the blockage of xylem conduits when woody plants suffer from water stress under drought and other environmental conditions, the study of embolism has become a hot and key topic under global climate change. Recent researches on the relationship between the vulnerability of xylem embolism and hydraulic architecture/drought tolerance have made some progress, however, scholars reached different conclusions based on results from different regions or different materials. This paper reviews the current achievements and controversial viewpoints, which includes indicator of xylem embolism vulnerability (P50), method of vulnerability curve establishment, the relationship between embolism vulnerability and hydraulic architecture (vessel diameter, vessel length, pit area, wood density, fiber and fiber tracheid) and the relationship between embolism vulnerability and drought tolerance of woody plants. Future studies should use Cochard Cavitron centrifuge and Sperry centrifuge coupled with traditional methods to establish vulnerability curves, calculate P50, analyze the difference among different organisms (root, stem, leaf), and measure physiological and ecological indexes. Future studies should be aimed to explore the relationship between the vulnerability of xylem embolism and hydraulic architecture/drought tolerance and to assess drought tolerance ability of different species under future climate change.
LIRong, JIANGZai-Min, ZHANGShuo-Xin, CAIJing. A review of new research progress on the vulnerability of xylem embolism of woody plants. Chinese Journal of Plant Ecology, 2015, 39(8): 838-848 https://doi.org/10.17521/cjpe.2015.0080
FichotR, BarigahTS, ChamaillardS, Le ThiecD, LauransF, CochardH, BrignolasF (2010).
Common trade-offs between xylem resistance to cavitation and other physiological traits do not hold among unrelated Populus deltoides × Populus nigra hybrids.
Drought induced xylem embolism in four riparian trees from the Western Cape Province: Insights and implications for planning and evaluation of restoration.
Vulnerability to cavitation in Olea europaea current-year shoots: Further evidence of an open-vessel artifact associated with centrifuge and air-injection techniques.
A computational algorithm addressing how vessel length might depend on vessel diameter.
0
2010b
Structure and function of bordered pits: New discoveries and impacts on whole- plant hydraulic function.
1
2008
... 纹孔是贯穿于导管分子间次生细胞壁的窄通道.纹孔分为单纹孔和具缘纹孔.单纹孔是次生壁上普遍存在的纹孔形式, 由纹孔腔和纹孔膜组成.具缘纹孔的构造较单纹孔复杂, 主要存在于木质部的组成分子中, 是导管或管胞的重要特征.每一个纹孔的中央是相对较薄的纹孔膜, 纹孔膜是由初生壁形成, 且由亲水性纤维素聚合物构成.纹孔膜在木质部水分运输传导上具有很重要的作用, 其上有大量的微孔, 是水流横向运输从一个导管(管胞)流向另一个导管(管胞)的入口(Choat et al., 2008; Loepfe et al., 2007). ...
Measurement of vulnerability to water stress-induced cavitation in grapevine: A comparison of four techniques applied to a long-vesseled species.
1
2010
... 栓塞脆弱性可通过栓塞脆弱曲线来反应, 该曲线的建立有许多方法, 早期常用的是自然干燥法(bench dehydration)、空气注入法(air-injection)和压力室法(pressure chamber)(Cochard et al., 1992), 目前国际上常用的快速测定栓塞脆弱曲线的方法是Cochard Cavitron离心机法(Cochard, 2002; Cochard et al., 2005)和Sperry离心机技术(Sperry et al., 1988).自然干燥法是最直接、最原始的诱导植物产生空穴化和栓塞的方法, 将植物(通常为带有叶片且长于1 m的枝条)不做任何处理直接放在空气中进行脱水干燥, 木质部压力可用压力势法测量未蒸腾的叶片而得(Tyree et al., 1992).空气注入法是将压缩的空气通过压力套注入木质部导管中, 木质部的导水率随压力的变化而变化, 从而建立枝条的栓塞脆弱曲线(Cochard et al., 1992).离心机法是将枝条放在离心机中旋转, 其原理是利用不断增大的离心力模拟活体植株中的木质部负压来诱导枝条产生栓塞, 从而测定枝条导水率损失值(PLC)(Pockman et al., 1995).不同之处是: 1)自然干燥法建立脆弱曲线需要数周时间和多个枝条才能得到, 因此比较费时, 而且原位诱导产生的栓塞可能影响导水率与木质部负压的关系, 并且最近研究发现, 在被切开的木质部导管压力下该方法易于建立错误曲线(Li et al., 2009; Wheeler et al., 2013; Torres-Ruiz et al., 2015).2)空气注入法的优势在于水分可以在几分钟内精确地调整到人为想要控制的程度, 相比于自然干燥法, 该方法可以在数小时内建立一条完整的脆弱曲线, 但实际操作中压力套在高压下会发生漏气现象, 同时注入导管内的空气很可能影响导水率, 所以需要等压力恢复到大气压时才能进行测定(Cochard et al., 2010).空气注入法和自然干燥法更适合于建立长导管植物脆弱曲线(Choat et al., 2010).3) Cochard Cavitron离心机是在Pockman离心机(Pockman et al., 1995)的基础上改装而来的, 它具有一个大转子(所容纳枝条长度为27.4 cm, 转子长27.5 cm), Sperry离心机具有一个小转子(所容纳枝条长度为14.5 cm), 不同之处是Cochard Cavitron离心机法仅需要一根枝条且通常在0.5 h内就可建立一条完整的脆弱曲线, 而Sperry离心机技术则需要3-5根枝条、一周时间才能建立一条完整的脆弱曲线, 离心机法具有空气注入法的优点, 且不用考虑由于已经存在的空气带来的不便, 此外, 离心机法还可以用于测定许多草本植物脆弱曲线, 这一点空气注入法是无法完成的(Tixier et al., 2013).离心机法所得脆弱曲线可将植物生理生态特性和木质部结构结合起来(Hacke et al., 2001, 2006; Jacobsen et al., 2005; Cai & Tyree, 2010; Lens et al., 2011), 可筛选出杨属(Populus)和柳属(Salix)栓塞抗性强的树种 (Cochard et al., 2007; Arango-Velez et al., 2011; Schreiber et al., 2011), 可将抗旱性和生活史结合起来评估植物抵御干旱的能力(Pratt et al., 2007), 也可用来探究干旱环境下植物的死亡率(Choat et al., 2012; Plaut et al., 2012; Anderegg et al., 2013; Paddock et al., 2013). ...
Global convergence in the vulnerability of forests to drought.
3
2012
... 依据Domec理论(Domec & Gartner, 2001), 测定和分析导水率损失最大值时的枝条临界水势和导水率50%时对应的枝条水势(P50)值(P50为负值), 可以很好地描述不同树种木质部的栓塞脆弱性、比较不同树种木质部抵抗栓塞的能力(Choat et al., 2012).但也有研究表明, 植物栓塞后的剩余导水率可能比P50值更适合描述植物在水分亏缺条件下抵抗栓塞的能力(Pammenter & van der Willigen, 1998; Swift et al., 2008).目前, 国际上大多数学者用P50值来表示栓塞脆弱性. ...
... 栓塞脆弱性可通过栓塞脆弱曲线来反应, 该曲线的建立有许多方法, 早期常用的是自然干燥法(bench dehydration)、空气注入法(air-injection)和压力室法(pressure chamber)(Cochard et al., 1992), 目前国际上常用的快速测定栓塞脆弱曲线的方法是Cochard Cavitron离心机法(Cochard, 2002; Cochard et al., 2005)和Sperry离心机技术(Sperry et al., 1988).自然干燥法是最直接、最原始的诱导植物产生空穴化和栓塞的方法, 将植物(通常为带有叶片且长于1 m的枝条)不做任何处理直接放在空气中进行脱水干燥, 木质部压力可用压力势法测量未蒸腾的叶片而得(Tyree et al., 1992).空气注入法是将压缩的空气通过压力套注入木质部导管中, 木质部的导水率随压力的变化而变化, 从而建立枝条的栓塞脆弱曲线(Cochard et al., 1992).离心机法是将枝条放在离心机中旋转, 其原理是利用不断增大的离心力模拟活体植株中的木质部负压来诱导枝条产生栓塞, 从而测定枝条导水率损失值(PLC)(Pockman et al., 1995).不同之处是: 1)自然干燥法建立脆弱曲线需要数周时间和多个枝条才能得到, 因此比较费时, 而且原位诱导产生的栓塞可能影响导水率与木质部负压的关系, 并且最近研究发现, 在被切开的木质部导管压力下该方法易于建立错误曲线(Li et al., 2009; Wheeler et al., 2013; Torres-Ruiz et al., 2015).2)空气注入法的优势在于水分可以在几分钟内精确地调整到人为想要控制的程度, 相比于自然干燥法, 该方法可以在数小时内建立一条完整的脆弱曲线, 但实际操作中压力套在高压下会发生漏气现象, 同时注入导管内的空气很可能影响导水率, 所以需要等压力恢复到大气压时才能进行测定(Cochard et al., 2010).空气注入法和自然干燥法更适合于建立长导管植物脆弱曲线(Choat et al., 2010).3) Cochard Cavitron离心机是在Pockman离心机(Pockman et al., 1995)的基础上改装而来的, 它具有一个大转子(所容纳枝条长度为27.4 cm, 转子长27.5 cm), Sperry离心机具有一个小转子(所容纳枝条长度为14.5 cm), 不同之处是Cochard Cavitron离心机法仅需要一根枝条且通常在0.5 h内就可建立一条完整的脆弱曲线, 而Sperry离心机技术则需要3-5根枝条、一周时间才能建立一条完整的脆弱曲线, 离心机法具有空气注入法的优点, 且不用考虑由于已经存在的空气带来的不便, 此外, 离心机法还可以用于测定许多草本植物脆弱曲线, 这一点空气注入法是无法完成的(Tixier et al., 2013).离心机法所得脆弱曲线可将植物生理生态特性和木质部结构结合起来(Hacke et al., 2001, 2006; Jacobsen et al., 2005; Cai & Tyree, 2010; Lens et al., 2011), 可筛选出杨属(Populus)和柳属(Salix)栓塞抗性强的树种 (Cochard et al., 2007; Arango-Velez et al., 2011; Schreiber et al., 2011), 可将抗旱性和生活史结合起来评估植物抵御干旱的能力(Pratt et al., 2007), 也可用来探究干旱环境下植物的死亡率(Choat et al., 2012; Plaut et al., 2012; Anderegg et al., 2013; Paddock et al., 2013). ...
... 离心机法建立栓塞脆弱曲线前是否需要对枝条进行冲洗还没有统一的认识.Wang等(2014)发现冲洗的枝条和未被冲洗的枝条在离心时没有区别, 原因是未被冲洗的枝条树液在旋转时很快被流动的液体所代替.相反, Choat等(2010)用离心方法对葡萄(Vitis vinifera)枝条研究时发现未被冲洗的枝条更不易栓塞.但与自然干燥法相比, 未被冲洗的枝条更易栓塞, 是因为气泡在浮力的作用下进入旋转的枝条, 导致导管发生栓塞(Hacke et al., 2015).目前, 用离心机法已经获得数百个树种的栓塞脆弱曲线(Choat et al., 2012), 其中, 绝大多数曲线可以用单威布尔累积分布函数拟合: ...
Frost fatigue and spring recovery of xylem vessels in three diffuse-porous trees in situ.
Cavitation and water storage capacity in bole xylem segments of mature and young Douglas-fir trees.
1
2001
... 依据Domec理论(Domec & Gartner, 2001), 测定和分析导水率损失最大值时的枝条临界水势和导水率50%时对应的枝条水势(P50)值(P50为负值), 可以很好地描述不同树种木质部的栓塞脆弱性、比较不同树种木质部抵抗栓塞的能力(Choat et al., 2012).但也有研究表明, 植物栓塞后的剩余导水率可能比P50值更适合描述植物在水分亏缺条件下抵抗栓塞的能力(Pammenter & van der Willigen, 1998; Swift et al., 2008).目前, 国际上大多数学者用P50值来表示栓塞脆弱性. ...
Fluid flow in the outermost xylem increment of a ring-porous tree, Ulmus americana.
Common trade-offs between xylem resistance to cavitation and other physiological traits do not hold among unrelated Populus deltoides × Populus nigra hybrids.
1
2010
... 木质部导管的导水率与导管直径的4次方成正比(D4, 源于Hagen-Poiseuille方程), 若增加一倍的导管直径将导致单个导管上16倍的导水率和4倍的横截面积的增大(Tyree & Zimmermann, 2002), 即导管直径越大, 导管的输水效率越高.因此, 多数研究认为木质部导管平均直径与栓塞脆弱性呈正相关关系, 即同一树种内大径导管比小径导管更易发生栓塞(Hargrave et al., 1994; Gullo et al., 1995; Wheeler et al., 2005; Hacke et al., 2006; Cai & Tyree, 2010; 张海昕等, 2013), 如: 阔叶树种是环孔材或散孔材, 环孔材导管直径较大、导管长度较长, 运输水分速度快, 从而容易发生栓塞; 针叶树是无孔材, 其管胞直径小、水分运输速度慢, 因而不易发生栓塞; 散孔材树种则导管直径较小、导管长度较短, 运输水分阻力较大, 栓塞发生的概率在环孔材树种和无孔材树种之间(李吉跃和翟洪波, 2000; 刘晓燕等, 2003).也有研究认为栓塞脆弱性与木质部导管直径成反比(Lemoine et al., 2002; Hacke et al., 2009), 或栓塞脆弱性与导管直径不相关(Rosner et al., 2007; Fichot et al., 2010).Cai和Tyree (2010)以杨属(Populus)植物为例, 按照植物导管直径径级构建的栓塞脆弱曲线表明: 随着导管直径增大, P50值增大(负值), 即导管直径与P50呈正相关关系(Markesteijn et al., 2011).除导管直径外, 木质部栓塞脆弱性与导管内径跨度(b)和导管壁的厚度(t)有关, 但(t/b)2比t或者b与P50相关性更好, 研究表明导管壁(t/b)2与木质部抗栓塞能力有很紧密的比例关系, 如: 12种针叶树种(t/b)2与栓塞脆弱性的R2 = 0.73, 36种被子植物二者R2 = 0.81, 因此(t/b)2大的植物可避免植物在负压下木质部导管壁的破坏, 从而防止栓塞的形成(Hacke et al., 2001).Cochard等(2008)对蔷薇科李属(Prunus)植物研究得出相同结论, 并指出导管壁为37 μm宽时, P50与(t2/b2)37相关性最好(R2 = 0.76, p = 0.001).但树木在长期的对外界环境条件适应的进化过程中, 可以通过形态和结构等自我调节来达到水分运输途中有效性与安全性的协调统一.在一些干旱半干旱地区, 植物木质部导管直径变小, 在一定程度上水分运输的有效性降低, 但小径导管的导管壁加厚, 在木质部压力很低时, 导管坍塌和被破坏的几率降低, 且导管密度增加, 水分运输的有效性即得到了一定的有效补偿. ...
Relations between vulnerability to xylem embolism and xylem conduit dimensions in young trees of Quercus corris.
1
1995
... 木质部导管的导水率与导管直径的4次方成正比(D4, 源于Hagen-Poiseuille方程), 若增加一倍的导管直径将导致单个导管上16倍的导水率和4倍的横截面积的增大(Tyree & Zimmermann, 2002), 即导管直径越大, 导管的输水效率越高.因此, 多数研究认为木质部导管平均直径与栓塞脆弱性呈正相关关系, 即同一树种内大径导管比小径导管更易发生栓塞(Hargrave et al., 1994; Gullo et al., 1995; Wheeler et al., 2005; Hacke et al., 2006; Cai & Tyree, 2010; 张海昕等, 2013), 如: 阔叶树种是环孔材或散孔材, 环孔材导管直径较大、导管长度较长, 运输水分速度快, 从而容易发生栓塞; 针叶树是无孔材, 其管胞直径小、水分运输速度慢, 因而不易发生栓塞; 散孔材树种则导管直径较小、导管长度较短, 运输水分阻力较大, 栓塞发生的概率在环孔材树种和无孔材树种之间(李吉跃和翟洪波, 2000; 刘晓燕等, 2003).也有研究认为栓塞脆弱性与木质部导管直径成反比(Lemoine et al., 2002; Hacke et al., 2009), 或栓塞脆弱性与导管直径不相关(Rosner et al., 2007; Fichot et al., 2010).Cai和Tyree (2010)以杨属(Populus)植物为例, 按照植物导管直径径级构建的栓塞脆弱曲线表明: 随着导管直径增大, P50值增大(负值), 即导管直径与P50呈正相关关系(Markesteijn et al., 2011).除导管直径外, 木质部栓塞脆弱性与导管内径跨度(b)和导管壁的厚度(t)有关, 但(t/b)2比t或者b与P50相关性更好, 研究表明导管壁(t/b)2与木质部抗栓塞能力有很紧密的比例关系, 如: 12种针叶树种(t/b)2与栓塞脆弱性的R2 = 0.73, 36种被子植物二者R2 = 0.81, 因此(t/b)2大的植物可避免植物在负压下木质部导管壁的破坏, 从而防止栓塞的形成(Hacke et al., 2001).Cochard等(2008)对蔷薇科李属(Prunus)植物研究得出相同结论, 并指出导管壁为37 μm宽时, P50与(t2/b2)37相关性最好(R2 = 0.76, p = 0.001).但树木在长期的对外界环境条件适应的进化过程中, 可以通过形态和结构等自我调节来达到水分运输途中有效性与安全性的协调统一.在一些干旱半干旱地区, 植物木质部导管直径变小, 在一定程度上水分运输的有效性降低, 但小径导管的导管壁加厚, 在木质部压力很低时, 导管坍塌和被破坏的几率降低, 且导管密度增加, 水分运输的有效性即得到了一定的有效补偿. ...
Xylem function of arid-land shrubs from California, USA: An ecological and evolutionary analysis.
1
2009
... 木质部导管的导水率与导管直径的4次方成正比(D4, 源于Hagen-Poiseuille方程), 若增加一倍的导管直径将导致单个导管上16倍的导水率和4倍的横截面积的增大(Tyree & Zimmermann, 2002), 即导管直径越大, 导管的输水效率越高.因此, 多数研究认为木质部导管平均直径与栓塞脆弱性呈正相关关系, 即同一树种内大径导管比小径导管更易发生栓塞(Hargrave et al., 1994; Gullo et al., 1995; Wheeler et al., 2005; Hacke et al., 2006; Cai & Tyree, 2010; 张海昕等, 2013), 如: 阔叶树种是环孔材或散孔材, 环孔材导管直径较大、导管长度较长, 运输水分速度快, 从而容易发生栓塞; 针叶树是无孔材, 其管胞直径小、水分运输速度慢, 因而不易发生栓塞; 散孔材树种则导管直径较小、导管长度较短, 运输水分阻力较大, 栓塞发生的概率在环孔材树种和无孔材树种之间(李吉跃和翟洪波, 2000; 刘晓燕等, 2003).也有研究认为栓塞脆弱性与木质部导管直径成反比(Lemoine et al., 2002; Hacke et al., 2009), 或栓塞脆弱性与导管直径不相关(Rosner et al., 2007; Fichot et al., 2010).Cai和Tyree (2010)以杨属(Populus)植物为例, 按照植物导管直径径级构建的栓塞脆弱曲线表明: 随着导管直径增大, P50值增大(负值), 即导管直径与P50呈正相关关系(Markesteijn et al., 2011).除导管直径外, 木质部栓塞脆弱性与导管内径跨度(b)和导管壁的厚度(t)有关, 但(t/b)2比t或者b与P50相关性更好, 研究表明导管壁(t/b)2与木质部抗栓塞能力有很紧密的比例关系, 如: 12种针叶树种(t/b)2与栓塞脆弱性的R2 = 0.73, 36种被子植物二者R2 = 0.81, 因此(t/b)2大的植物可避免植物在负压下木质部导管壁的破坏, 从而防止栓塞的形成(Hacke et al., 2001).Cochard等(2008)对蔷薇科李属(Prunus)植物研究得出相同结论, 并指出导管壁为37 μm宽时, P50与(t2/b2)37相关性最好(R2 = 0.76, p = 0.001).但树木在长期的对外界环境条件适应的进化过程中, 可以通过形态和结构等自我调节来达到水分运输途中有效性与安全性的协调统一.在一些干旱半干旱地区, 植物木质部导管直径变小, 在一定程度上水分运输的有效性降低, 但小径导管的导管壁加厚, 在木质部压力很低时, 导管坍塌和被破坏的几率降低, 且导管密度增加, 水分运输的有效性即得到了一定的有效补偿. ...
Embolism resistance of three boreal conifer species varies with pit structure.
4
2009
... 木质部空穴化和栓塞的发现为植物输水结构体系和植物耐旱机理研究提供了新的思路和方向, 植物对木质部栓塞的易受性可用“脆弱曲线(VC)”来描述(Tyree & Sperry, 1989), 该曲线是根据木质部导水率损失的百分数(PLC)与相对应水势绘制而成的曲线, 且已被广泛引用(Maherali et al., 2004).栓塞脆弱曲线的建立可以使学者们对植物抵御栓塞抗性的大小进行评估, 在研究了木质部栓塞脆弱性与输水结构间的关系(Wheeler et al., 2005; Sperry et al., 2006; Hacke & Jansen, 2009; Cai & Tyree, 2010; 张海昕等, 2013)及植物木质部栓塞脆弱性与耐旱性的关系(Tyree et al., 2003; Maherali et al., 2004; Awad et al., 2010)后, 学者们认为由植物木质部空穴化和栓塞导致的木本植物输水结构功能障碍影响其自身耐旱能力.脆弱曲线与植物的部位、木质部结构、分布、抗旱性等存在一定关系, 因此能够提供许多抗旱信息(安锋和张硕新, 2005).木质部栓塞脆弱性与输水结构间的关系可使人们评估植物对栓塞的抵抗力大小, 从而进一步探究木质部栓塞脆弱性和耐旱性的关系, 但各学者对不同树种研究所得结论不尽相同. ...
... 输水结构被用来描述整个树体上茎段导水率与其结构间的关系(Zimmermann, 1983).栓塞使木质部管道中充有气泡而阻断了水分的运输, 因此, 输水结构的输水效率会受到木质部栓塞脆弱性的影响.木质部栓塞脆弱性与导管或管胞的解剖特征和纹孔超微结构密切相关(Wheeler et al., 2005; Sperry et al., 2006; Hacke & Jansen, 2009). ...
... ; Milburn & McLaughlin, 1974).空穴发生后, 空气会填充进真空的导管内(栓塞化), 使原来连续的水柱断裂, 从而阻碍水分的运输.目前认为“空穴形成”有4种模式: 1)气泡由纹孔进入导管形成空穴; 2)液体流动过程中自然发生的微小气核膨胀形成空穴; 3)驻留在导管内疏水区域的小气泡膨胀形成空穴; 4)导管壁上微小气核与疏水区脱离形成空穴(Tyree & Zimmermann, 2002).气泡使得空穴化在两个相邻导管间扩散, 从而阻滞水分的运输, 最终导致栓塞(Tyree & Sperry, 1989).这一理论得到学者的广泛支持(Tyree et al., 2003; Angeles et al., 2004). ...
Xylem cavitation in the leaf of Prunus laurocerasus and its impact on leaf hydraulics.
0
2001
Factors determining mortality of adult chaparral shrubs in an extreme drought year in California.
1
2013
... 栓塞脆弱性可通过栓塞脆弱曲线来反应, 该曲线的建立有许多方法, 早期常用的是自然干燥法(bench dehydration)、空气注入法(air-injection)和压力室法(pressure chamber)(Cochard et al., 1992), 目前国际上常用的快速测定栓塞脆弱曲线的方法是Cochard Cavitron离心机法(Cochard, 2002; Cochard et al., 2005)和Sperry离心机技术(Sperry et al., 1988).自然干燥法是最直接、最原始的诱导植物产生空穴化和栓塞的方法, 将植物(通常为带有叶片且长于1 m的枝条)不做任何处理直接放在空气中进行脱水干燥, 木质部压力可用压力势法测量未蒸腾的叶片而得(Tyree et al., 1992).空气注入法是将压缩的空气通过压力套注入木质部导管中, 木质部的导水率随压力的变化而变化, 从而建立枝条的栓塞脆弱曲线(Cochard et al., 1992).离心机法是将枝条放在离心机中旋转, 其原理是利用不断增大的离心力模拟活体植株中的木质部负压来诱导枝条产生栓塞, 从而测定枝条导水率损失值(PLC)(Pockman et al., 1995).不同之处是: 1)自然干燥法建立脆弱曲线需要数周时间和多个枝条才能得到, 因此比较费时, 而且原位诱导产生的栓塞可能影响导水率与木质部负压的关系, 并且最近研究发现, 在被切开的木质部导管压力下该方法易于建立错误曲线(Li et al., 2009; Wheeler et al., 2013; Torres-Ruiz et al., 2015).2)空气注入法的优势在于水分可以在几分钟内精确地调整到人为想要控制的程度, 相比于自然干燥法, 该方法可以在数小时内建立一条完整的脆弱曲线, 但实际操作中压力套在高压下会发生漏气现象, 同时注入导管内的空气很可能影响导水率, 所以需要等压力恢复到大气压时才能进行测定(Cochard et al., 2010).空气注入法和自然干燥法更适合于建立长导管植物脆弱曲线(Choat et al., 2010).3) Cochard Cavitron离心机是在Pockman离心机(Pockman et al., 1995)的基础上改装而来的, 它具有一个大转子(所容纳枝条长度为27.4 cm, 转子长27.5 cm), Sperry离心机具有一个小转子(所容纳枝条长度为14.5 cm), 不同之处是Cochard Cavitron离心机法仅需要一根枝条且通常在0.5 h内就可建立一条完整的脆弱曲线, 而Sperry离心机技术则需要3-5根枝条、一周时间才能建立一条完整的脆弱曲线, 离心机法具有空气注入法的优点, 且不用考虑由于已经存在的空气带来的不便, 此外, 离心机法还可以用于测定许多草本植物脆弱曲线, 这一点空气注入法是无法完成的(Tixier et al., 2013).离心机法所得脆弱曲线可将植物生理生态特性和木质部结构结合起来(Hacke et al., 2001, 2006; Jacobsen et al., 2005; Cai & Tyree, 2010; Lens et al., 2011), 可筛选出杨属(Populus)和柳属(Salix)栓塞抗性强的树种 (Cochard et al., 2007; Arango-Velez et al., 2011; Schreiber et al., 2011), 可将抗旱性和生活史结合起来评估植物抵御干旱的能力(Pratt et al., 2007), 也可用来探究干旱环境下植物的死亡率(Choat et al., 2012; Plaut et al., 2012; Anderegg et al., 2013; Paddock et al., 2013). ...
A mathematical and statistical analysis of the curves illustrating vulnerability of xylem to cavitation.
1
1998
... 依据Domec理论(Domec & Gartner, 2001), 测定和分析导水率损失最大值时的枝条临界水势和导水率50%时对应的枝条水势(P50)值(P50为负值), 可以很好地描述不同树种木质部的栓塞脆弱性、比较不同树种木质部抵抗栓塞的能力(Choat et al., 2012).但也有研究表明, 植物栓塞后的剩余导水率可能比P50值更适合描述植物在水分亏缺条件下抵抗栓塞的能力(Pammenter & van der Willigen, 1998; Swift et al., 2008).目前, 国际上大多数学者用P50值来表示栓塞脆弱性. ...
Hydraulic limits preceding mortality in a piñon-juniper woodland under experimental drought.
1
2012
... 栓塞脆弱性可通过栓塞脆弱曲线来反应, 该曲线的建立有许多方法, 早期常用的是自然干燥法(bench dehydration)、空气注入法(air-injection)和压力室法(pressure chamber)(Cochard et al., 1992), 目前国际上常用的快速测定栓塞脆弱曲线的方法是Cochard Cavitron离心机法(Cochard, 2002; Cochard et al., 2005)和Sperry离心机技术(Sperry et al., 1988).自然干燥法是最直接、最原始的诱导植物产生空穴化和栓塞的方法, 将植物(通常为带有叶片且长于1 m的枝条)不做任何处理直接放在空气中进行脱水干燥, 木质部压力可用压力势法测量未蒸腾的叶片而得(Tyree et al., 1992).空气注入法是将压缩的空气通过压力套注入木质部导管中, 木质部的导水率随压力的变化而变化, 从而建立枝条的栓塞脆弱曲线(Cochard et al., 1992).离心机法是将枝条放在离心机中旋转, 其原理是利用不断增大的离心力模拟活体植株中的木质部负压来诱导枝条产生栓塞, 从而测定枝条导水率损失值(PLC)(Pockman et al., 1995).不同之处是: 1)自然干燥法建立脆弱曲线需要数周时间和多个枝条才能得到, 因此比较费时, 而且原位诱导产生的栓塞可能影响导水率与木质部负压的关系, 并且最近研究发现, 在被切开的木质部导管压力下该方法易于建立错误曲线(Li et al., 2009; Wheeler et al., 2013; Torres-Ruiz et al., 2015).2)空气注入法的优势在于水分可以在几分钟内精确地调整到人为想要控制的程度, 相比于自然干燥法, 该方法可以在数小时内建立一条完整的脆弱曲线, 但实际操作中压力套在高压下会发生漏气现象, 同时注入导管内的空气很可能影响导水率, 所以需要等压力恢复到大气压时才能进行测定(Cochard et al., 2010).空气注入法和自然干燥法更适合于建立长导管植物脆弱曲线(Choat et al., 2010).3) Cochard Cavitron离心机是在Pockman离心机(Pockman et al., 1995)的基础上改装而来的, 它具有一个大转子(所容纳枝条长度为27.4 cm, 转子长27.5 cm), Sperry离心机具有一个小转子(所容纳枝条长度为14.5 cm), 不同之处是Cochard Cavitron离心机法仅需要一根枝条且通常在0.5 h内就可建立一条完整的脆弱曲线, 而Sperry离心机技术则需要3-5根枝条、一周时间才能建立一条完整的脆弱曲线, 离心机法具有空气注入法的优点, 且不用考虑由于已经存在的空气带来的不便, 此外, 离心机法还可以用于测定许多草本植物脆弱曲线, 这一点空气注入法是无法完成的(Tixier et al., 2013).离心机法所得脆弱曲线可将植物生理生态特性和木质部结构结合起来(Hacke et al., 2001, 2006; Jacobsen et al., 2005; Cai & Tyree, 2010; Lens et al., 2011), 可筛选出杨属(Populus)和柳属(Salix)栓塞抗性强的树种 (Cochard et al., 2007; Arango-Velez et al., 2011; Schreiber et al., 2011), 可将抗旱性和生活史结合起来评估植物抵御干旱的能力(Pratt et al., 2007), 也可用来探究干旱环境下植物的死亡率(Choat et al., 2012; Plaut et al., 2012; Anderegg et al., 2013; Paddock et al., 2013). ...
Linking irradiance-induced changes in pit membrane ultrastructure with xylem vulnerability to cavitation.
1
2011
... 有研究表明纹孔膜的厚度和整个细胞壁厚度的协同性可能对木质部密度有一定影响(Jansen et al., 2009; Plavcová et al., 2011).Hacke等(2001)对来自不同科的48种植物进行研究, 发现12种针叶树种木质部密度和栓塞脆弱性的R2 = 0.73, 36种被子植物二者R2 = 0.62, 因此得出木质部密度与栓塞脆弱性有很强的相关性, 木质部密度越大, P50值越小(越负), 对栓塞抵抗力越强, 植物越不易栓塞.该结论随后又得到了进一步的验证(Jacobsen et al., 2005).但Cochard等(2008)对蔷薇科李属的10个树种研究发现木质部密度与P50的R2 < 0.4, 其认为利用木质部密度作为评价李属不同种栓塞脆弱性的指标是不现实的. ...
Vulnerability to xylem cavitation and the distribution of Sonoran desert vegetation.
1
2000
... 栓塞脆弱曲线的建立使得人们可以评估植物对栓塞的抵抗力, 有些学者分析了植物木质部栓塞脆弱性与耐旱性的关系(Sperry & Tyree, 1990; Cochard et al., 1992, 2008, 2013; Tyree et al., 2003; Maherali et al., 2004), 认为由植物木质部空穴化和栓塞导致的植物输水结构功能障碍影响了植物的气孔运动及耐旱能力(Sperry et al., 1988; Salleo et al., 2000).栓塞脆弱性决定了干旱地区生态系统中不同物种的生存模式(Davis et al., 1998; Pockman & Sperry, 2000), 但栓塞抗性在不同物种和不同森林生态系统的表现不同(Maherali et al., 2004; Delzon et al., 2010), 因此, 在严重干旱情况下裸子植物和被子植物存活率也不同(Brodribb & Cochard, 2009; Brodribb et al., 2010). ...
Sustained and significant negative water pressure in xylem.
2
1995
... 栓塞脆弱性可通过栓塞脆弱曲线来反应, 该曲线的建立有许多方法, 早期常用的是自然干燥法(bench dehydration)、空气注入法(air-injection)和压力室法(pressure chamber)(Cochard et al., 1992), 目前国际上常用的快速测定栓塞脆弱曲线的方法是Cochard Cavitron离心机法(Cochard, 2002; Cochard et al., 2005)和Sperry离心机技术(Sperry et al., 1988).自然干燥法是最直接、最原始的诱导植物产生空穴化和栓塞的方法, 将植物(通常为带有叶片且长于1 m的枝条)不做任何处理直接放在空气中进行脱水干燥, 木质部压力可用压力势法测量未蒸腾的叶片而得(Tyree et al., 1992).空气注入法是将压缩的空气通过压力套注入木质部导管中, 木质部的导水率随压力的变化而变化, 从而建立枝条的栓塞脆弱曲线(Cochard et al., 1992).离心机法是将枝条放在离心机中旋转, 其原理是利用不断增大的离心力模拟活体植株中的木质部负压来诱导枝条产生栓塞, 从而测定枝条导水率损失值(PLC)(Pockman et al., 1995).不同之处是: 1)自然干燥法建立脆弱曲线需要数周时间和多个枝条才能得到, 因此比较费时, 而且原位诱导产生的栓塞可能影响导水率与木质部负压的关系, 并且最近研究发现, 在被切开的木质部导管压力下该方法易于建立错误曲线(Li et al., 2009; Wheeler et al., 2013; Torres-Ruiz et al., 2015).2)空气注入法的优势在于水分可以在几分钟内精确地调整到人为想要控制的程度, 相比于自然干燥法, 该方法可以在数小时内建立一条完整的脆弱曲线, 但实际操作中压力套在高压下会发生漏气现象, 同时注入导管内的空气很可能影响导水率, 所以需要等压力恢复到大气压时才能进行测定(Cochard et al., 2010).空气注入法和自然干燥法更适合于建立长导管植物脆弱曲线(Choat et al., 2010).3) Cochard Cavitron离心机是在Pockman离心机(Pockman et al., 1995)的基础上改装而来的, 它具有一个大转子(所容纳枝条长度为27.4 cm, 转子长27.5 cm), Sperry离心机具有一个小转子(所容纳枝条长度为14.5 cm), 不同之处是Cochard Cavitron离心机法仅需要一根枝条且通常在0.5 h内就可建立一条完整的脆弱曲线, 而Sperry离心机技术则需要3-5根枝条、一周时间才能建立一条完整的脆弱曲线, 离心机法具有空气注入法的优点, 且不用考虑由于已经存在的空气带来的不便, 此外, 离心机法还可以用于测定许多草本植物脆弱曲线, 这一点空气注入法是无法完成的(Tixier et al., 2013).离心机法所得脆弱曲线可将植物生理生态特性和木质部结构结合起来(Hacke et al., 2001, 2006; Jacobsen et al., 2005; Cai & Tyree, 2010; Lens et al., 2011), 可筛选出杨属(Populus)和柳属(Salix)栓塞抗性强的树种 (Cochard et al., 2007; Arango-Velez et al., 2011; Schreiber et al., 2011), 可将抗旱性和生活史结合起来评估植物抵御干旱的能力(Pratt et al., 2007), 也可用来探究干旱环境下植物的死亡率(Choat et al., 2012; Plaut et al., 2012; Anderegg et al., 2013; Paddock et al., 2013). ...
... ).3) Cochard Cavitron离心机是在Pockman离心机(Pockman et al., 1995)的基础上改装而来的, 它具有一个大转子(所容纳枝条长度为27.4 cm, 转子长27.5 cm), Sperry离心机具有一个小转子(所容纳枝条长度为14.5 cm), 不同之处是Cochard Cavitron离心机法仅需要一根枝条且通常在0.5 h内就可建立一条完整的脆弱曲线, 而Sperry离心机技术则需要3-5根枝条、一周时间才能建立一条完整的脆弱曲线, 离心机法具有空气注入法的优点, 且不用考虑由于已经存在的空气带来的不便, 此外, 离心机法还可以用于测定许多草本植物脆弱曲线, 这一点空气注入法是无法完成的(Tixier et al., 2013).离心机法所得脆弱曲线可将植物生理生态特性和木质部结构结合起来(Hacke et al., 2001, 2006; Jacobsen et al., 2005; Cai & Tyree, 2010; Lens et al., 2011), 可筛选出杨属(Populus)和柳属(Salix)栓塞抗性强的树种 (Cochard et al., 2007; Arango-Velez et al., 2011; Schreiber et al., 2011), 可将抗旱性和生活史结合起来评估植物抵御干旱的能力(Pratt et al., 2007), 也可用来探究干旱环境下植物的死亡率(Choat et al., 2012; Plaut et al., 2012; Anderegg et al., 2013; Paddock et al., 2013). ...
Life history type and water stress tolerance in nine California chaparral species (Rhamnac- eae).
1
2007
... 栓塞脆弱性可通过栓塞脆弱曲线来反应, 该曲线的建立有许多方法, 早期常用的是自然干燥法(bench dehydration)、空气注入法(air-injection)和压力室法(pressure chamber)(Cochard et al., 1992), 目前国际上常用的快速测定栓塞脆弱曲线的方法是Cochard Cavitron离心机法(Cochard, 2002; Cochard et al., 2005)和Sperry离心机技术(Sperry et al., 1988).自然干燥法是最直接、最原始的诱导植物产生空穴化和栓塞的方法, 将植物(通常为带有叶片且长于1 m的枝条)不做任何处理直接放在空气中进行脱水干燥, 木质部压力可用压力势法测量未蒸腾的叶片而得(Tyree et al., 1992).空气注入法是将压缩的空气通过压力套注入木质部导管中, 木质部的导水率随压力的变化而变化, 从而建立枝条的栓塞脆弱曲线(Cochard et al., 1992).离心机法是将枝条放在离心机中旋转, 其原理是利用不断增大的离心力模拟活体植株中的木质部负压来诱导枝条产生栓塞, 从而测定枝条导水率损失值(PLC)(Pockman et al., 1995).不同之处是: 1)自然干燥法建立脆弱曲线需要数周时间和多个枝条才能得到, 因此比较费时, 而且原位诱导产生的栓塞可能影响导水率与木质部负压的关系, 并且最近研究发现, 在被切开的木质部导管压力下该方法易于建立错误曲线(Li et al., 2009; Wheeler et al., 2013; Torres-Ruiz et al., 2015).2)空气注入法的优势在于水分可以在几分钟内精确地调整到人为想要控制的程度, 相比于自然干燥法, 该方法可以在数小时内建立一条完整的脆弱曲线, 但实际操作中压力套在高压下会发生漏气现象, 同时注入导管内的空气很可能影响导水率, 所以需要等压力恢复到大气压时才能进行测定(Cochard et al., 2010).空气注入法和自然干燥法更适合于建立长导管植物脆弱曲线(Choat et al., 2010).3) Cochard Cavitron离心机是在Pockman离心机(Pockman et al., 1995)的基础上改装而来的, 它具有一个大转子(所容纳枝条长度为27.4 cm, 转子长27.5 cm), Sperry离心机具有一个小转子(所容纳枝条长度为14.5 cm), 不同之处是Cochard Cavitron离心机法仅需要一根枝条且通常在0.5 h内就可建立一条完整的脆弱曲线, 而Sperry离心机技术则需要3-5根枝条、一周时间才能建立一条完整的脆弱曲线, 离心机法具有空气注入法的优点, 且不用考虑由于已经存在的空气带来的不便, 此外, 离心机法还可以用于测定许多草本植物脆弱曲线, 这一点空气注入法是无法完成的(Tixier et al., 2013).离心机法所得脆弱曲线可将植物生理生态特性和木质部结构结合起来(Hacke et al., 2001, 2006; Jacobsen et al., 2005; Cai & Tyree, 2010; Lens et al., 2011), 可筛选出杨属(Populus)和柳属(Salix)栓塞抗性强的树种 (Cochard et al., 2007; Arango-Velez et al., 2011; Schreiber et al., 2011), 可将抗旱性和生活史结合起来评估植物抵御干旱的能力(Pratt et al., 2007), 也可用来探究干旱环境下植物的死亡率(Choat et al., 2012; Plaut et al., 2012; Anderegg et al., 2013; Paddock et al., 2013). ...
From genome to function: The Arabidopsis aquaporins.
1
2002
... 在植物生存和发挥功能所需的各种资源中, 水是最重要的, 同时也是对植物生长限制性最强的(Johansson et al., 2000; Kaldenhoff & Fisher, 2006).植物只有在含有一定量水分的条件下才能进行正常的生命活动, 否则生命活动就会受到阻碍, 甚至出现死亡(Martre et al., 2002; Quigley et al., 2002; Cai & Tyree, 2010).而因干旱引起的木质部空穴化和栓塞则是影响树木正常生长发育的一个重要制约因子. ...
Cavitation and its discontents: Opportunities for resolving current controversies.
0
2014
Hydraulic and mechanical properties of young Norway spruce clones related to growth and wood structure.
1
2007
... 木质部导管的导水率与导管直径的4次方成正比(D4, 源于Hagen-Poiseuille方程), 若增加一倍的导管直径将导致单个导管上16倍的导水率和4倍的横截面积的增大(Tyree & Zimmermann, 2002), 即导管直径越大, 导管的输水效率越高.因此, 多数研究认为木质部导管平均直径与栓塞脆弱性呈正相关关系, 即同一树种内大径导管比小径导管更易发生栓塞(Hargrave et al., 1994; Gullo et al., 1995; Wheeler et al., 2005; Hacke et al., 2006; Cai & Tyree, 2010; 张海昕等, 2013), 如: 阔叶树种是环孔材或散孔材, 环孔材导管直径较大、导管长度较长, 运输水分速度快, 从而容易发生栓塞; 针叶树是无孔材, 其管胞直径小、水分运输速度慢, 因而不易发生栓塞; 散孔材树种则导管直径较小、导管长度较短, 运输水分阻力较大, 栓塞发生的概率在环孔材树种和无孔材树种之间(李吉跃和翟洪波, 2000; 刘晓燕等, 2003).也有研究认为栓塞脆弱性与木质部导管直径成反比(Lemoine et al., 2002; Hacke et al., 2009), 或栓塞脆弱性与导管直径不相关(Rosner et al., 2007; Fichot et al., 2010).Cai和Tyree (2010)以杨属(Populus)植物为例, 按照植物导管直径径级构建的栓塞脆弱曲线表明: 随着导管直径增大, P50值增大(负值), 即导管直径与P50呈正相关关系(Markesteijn et al., 2011).除导管直径外, 木质部栓塞脆弱性与导管内径跨度(b)和导管壁的厚度(t)有关, 但(t/b)2比t或者b与P50相关性更好, 研究表明导管壁(t/b)2与木质部抗栓塞能力有很紧密的比例关系, 如: 12种针叶树种(t/b)2与栓塞脆弱性的R2 = 0.73, 36种被子植物二者R2 = 0.81, 因此(t/b)2大的植物可避免植物在负压下木质部导管壁的破坏, 从而防止栓塞的形成(Hacke et al., 2001).Cochard等(2008)对蔷薇科李属(Prunus)植物研究得出相同结论, 并指出导管壁为37 μm宽时, P50与(t2/b2)37相关性最好(R2 = 0.76, p = 0.001).但树木在长期的对外界环境条件适应的进化过程中, 可以通过形态和结构等自我调节来达到水分运输途中有效性与安全性的协调统一.在一些干旱半干旱地区, 植物木质部导管直径变小, 在一定程度上水分运输的有效性降低, 但小径导管的导管壁加厚, 在木质部压力很低时, 导管坍塌和被破坏的几率降低, 且导管密度增加, 水分运输的有效性即得到了一定的有效补偿. ...
Xylem cavitation and hydraulic control of stomatal conductance in Laurel (Laurus nobilis L.).
1
2000
... 栓塞脆弱曲线的建立使得人们可以评估植物对栓塞的抵抗力, 有些学者分析了植物木质部栓塞脆弱性与耐旱性的关系(Sperry & Tyree, 1990; Cochard et al., 1992, 2008, 2013; Tyree et al., 2003; Maherali et al., 2004), 认为由植物木质部空穴化和栓塞导致的植物输水结构功能障碍影响了植物的气孔运动及耐旱能力(Sperry et al., 1988; Salleo et al., 2000).栓塞脆弱性决定了干旱地区生态系统中不同物种的生存模式(Davis et al., 1998; Pockman & Sperry, 2000), 但栓塞抗性在不同物种和不同森林生态系统的表现不同(Maherali et al., 2004; Delzon et al., 2010), 因此, 在严重干旱情况下裸子植物和被子植物存活率也不同(Brodribb & Cochard, 2009; Brodribb et al., 2010). ...
Genetic variation of hydraulic and wood anatomical traits in hybrid poplar and trembling aspen.
1
2011
... 栓塞脆弱性可通过栓塞脆弱曲线来反应, 该曲线的建立有许多方法, 早期常用的是自然干燥法(bench dehydration)、空气注入法(air-injection)和压力室法(pressure chamber)(Cochard et al., 1992), 目前国际上常用的快速测定栓塞脆弱曲线的方法是Cochard Cavitron离心机法(Cochard, 2002; Cochard et al., 2005)和Sperry离心机技术(Sperry et al., 1988).自然干燥法是最直接、最原始的诱导植物产生空穴化和栓塞的方法, 将植物(通常为带有叶片且长于1 m的枝条)不做任何处理直接放在空气中进行脱水干燥, 木质部压力可用压力势法测量未蒸腾的叶片而得(Tyree et al., 1992).空气注入法是将压缩的空气通过压力套注入木质部导管中, 木质部的导水率随压力的变化而变化, 从而建立枝条的栓塞脆弱曲线(Cochard et al., 1992).离心机法是将枝条放在离心机中旋转, 其原理是利用不断增大的离心力模拟活体植株中的木质部负压来诱导枝条产生栓塞, 从而测定枝条导水率损失值(PLC)(Pockman et al., 1995).不同之处是: 1)自然干燥法建立脆弱曲线需要数周时间和多个枝条才能得到, 因此比较费时, 而且原位诱导产生的栓塞可能影响导水率与木质部负压的关系, 并且最近研究发现, 在被切开的木质部导管压力下该方法易于建立错误曲线(Li et al., 2009; Wheeler et al., 2013; Torres-Ruiz et al., 2015).2)空气注入法的优势在于水分可以在几分钟内精确地调整到人为想要控制的程度, 相比于自然干燥法, 该方法可以在数小时内建立一条完整的脆弱曲线, 但实际操作中压力套在高压下会发生漏气现象, 同时注入导管内的空气很可能影响导水率, 所以需要等压力恢复到大气压时才能进行测定(Cochard et al., 2010).空气注入法和自然干燥法更适合于建立长导管植物脆弱曲线(Choat et al., 2010).3) Cochard Cavitron离心机是在Pockman离心机(Pockman et al., 1995)的基础上改装而来的, 它具有一个大转子(所容纳枝条长度为27.4 cm, 转子长27.5 cm), Sperry离心机具有一个小转子(所容纳枝条长度为14.5 cm), 不同之处是Cochard Cavitron离心机法仅需要一根枝条且通常在0.5 h内就可建立一条完整的脆弱曲线, 而Sperry离心机技术则需要3-5根枝条、一周时间才能建立一条完整的脆弱曲线, 离心机法具有空气注入法的优点, 且不用考虑由于已经存在的空气带来的不便, 此外, 离心机法还可以用于测定许多草本植物脆弱曲线, 这一点空气注入法是无法完成的(Tixier et al., 2013).离心机法所得脆弱曲线可将植物生理生态特性和木质部结构结合起来(Hacke et al., 2001, 2006; Jacobsen et al., 2005; Cai & Tyree, 2010; Lens et al., 2011), 可筛选出杨属(Populus)和柳属(Salix)栓塞抗性强的树种 (Cochard et al., 2007; Arango-Velez et al., 2011; Schreiber et al., 2011), 可将抗旱性和生活史结合起来评估植物抵御干旱的能力(Pratt et al., 2007), 也可用来探究干旱环境下植物的死亡率(Choat et al., 2012; Plaut et al., 2012; Anderegg et al., 2013; Paddock et al., 2013). ...
Vulnerability curves by centrifugation: Is there an open vessel artefact, and are ‘r’ shaped curves necessarily invalid?
3
2012
... 式中, PLC是导水率损失值, T是压力, b和c是威布尔常量, 其中参数b是当最大导水率(Kmax)下降63.2%时对应的木质部的负压值, 参数c是曲线的斜率.单威布尔累积分布函数能够拟合很多树种的脆弱曲线, 包括“s”形和“r”形(Sperry et al., 2012).但有些树种脆弱曲线不能用单威布尔累积分布函数很好地拟合, 研究发现沙棘(Hippophae rhamnoides)的脆弱曲线用双威布尔函数比单威布尔函数拟合度更高(Cai et al., 2014). ...
... 目前, 对于一些树种的相同部位材料采用不同方法所得脆弱曲线不同, 特别是对离心机法建立的长导管植物的栓塞脆弱曲线正确与否存在很大争议(Cochard et al., 2005, 2010, 2013; Sperry et al., 2012; Torres-Ruiz et al., 2015).离心机法建立的脆弱曲线有两种形态, 一种为“r”形, 一种为“s”形, “r”形脆弱曲线意味着在水势较高(稍负)时或稍降时, 植物即产生很大的栓塞或导水率损失, “s”形脆弱曲线则表示只有当水分亏缺达到一定程度才会发生栓塞.因此, 脆弱曲线的形式直接影响导水率损失值(P50).有关脆弱曲线的争议主要表现在以下几方面: ...
... 但还有一种研究认为离心机法建立的“r”形曲线是正确的.Sperry等(2012)研究表明采用脱水法、空气注入法与离心机法得到了相同的曲线, 他们认为并不是所有的环孔材导管都被切开(Li et al., 2008; Jacobsen & Pratt, 2012; Sperry et al., 2012; Christman et al., 2012).Jacobsen等(2014)对10种植物(含长导管植物, 最大导管长度>1 m)建立脆弱曲线, 发现利用不同方法所得植物导水率损失P50值无明显差异.Hacke等(2015)对具长导管的橄榄进行研究, 发现利用离心机法和自然干燥法所得曲线均为“r”形, 他认为离心机法是有效的. ...
A method for measuring hydraulic conductivity and embolism in xylem.
2
1988
... 栓塞脆弱性可通过栓塞脆弱曲线来反应, 该曲线的建立有许多方法, 早期常用的是自然干燥法(bench dehydration)、空气注入法(air-injection)和压力室法(pressure chamber)(Cochard et al., 1992), 目前国际上常用的快速测定栓塞脆弱曲线的方法是Cochard Cavitron离心机法(Cochard, 2002; Cochard et al., 2005)和Sperry离心机技术(Sperry et al., 1988).自然干燥法是最直接、最原始的诱导植物产生空穴化和栓塞的方法, 将植物(通常为带有叶片且长于1 m的枝条)不做任何处理直接放在空气中进行脱水干燥, 木质部压力可用压力势法测量未蒸腾的叶片而得(Tyree et al., 1992).空气注入法是将压缩的空气通过压力套注入木质部导管中, 木质部的导水率随压力的变化而变化, 从而建立枝条的栓塞脆弱曲线(Cochard et al., 1992).离心机法是将枝条放在离心机中旋转, 其原理是利用不断增大的离心力模拟活体植株中的木质部负压来诱导枝条产生栓塞, 从而测定枝条导水率损失值(PLC)(Pockman et al., 1995).不同之处是: 1)自然干燥法建立脆弱曲线需要数周时间和多个枝条才能得到, 因此比较费时, 而且原位诱导产生的栓塞可能影响导水率与木质部负压的关系, 并且最近研究发现, 在被切开的木质部导管压力下该方法易于建立错误曲线(Li et al., 2009; Wheeler et al., 2013; Torres-Ruiz et al., 2015).2)空气注入法的优势在于水分可以在几分钟内精确地调整到人为想要控制的程度, 相比于自然干燥法, 该方法可以在数小时内建立一条完整的脆弱曲线, 但实际操作中压力套在高压下会发生漏气现象, 同时注入导管内的空气很可能影响导水率, 所以需要等压力恢复到大气压时才能进行测定(Cochard et al., 2010).空气注入法和自然干燥法更适合于建立长导管植物脆弱曲线(Choat et al., 2010).3) Cochard Cavitron离心机是在Pockman离心机(Pockman et al., 1995)的基础上改装而来的, 它具有一个大转子(所容纳枝条长度为27.4 cm, 转子长27.5 cm), Sperry离心机具有一个小转子(所容纳枝条长度为14.5 cm), 不同之处是Cochard Cavitron离心机法仅需要一根枝条且通常在0.5 h内就可建立一条完整的脆弱曲线, 而Sperry离心机技术则需要3-5根枝条、一周时间才能建立一条完整的脆弱曲线, 离心机法具有空气注入法的优点, 且不用考虑由于已经存在的空气带来的不便, 此外, 离心机法还可以用于测定许多草本植物脆弱曲线, 这一点空气注入法是无法完成的(Tixier et al., 2013).离心机法所得脆弱曲线可将植物生理生态特性和木质部结构结合起来(Hacke et al., 2001, 2006; Jacobsen et al., 2005; Cai & Tyree, 2010; Lens et al., 2011), 可筛选出杨属(Populus)和柳属(Salix)栓塞抗性强的树种 (Cochard et al., 2007; Arango-Velez et al., 2011; Schreiber et al., 2011), 可将抗旱性和生活史结合起来评估植物抵御干旱的能力(Pratt et al., 2007), 也可用来探究干旱环境下植物的死亡率(Choat et al., 2012; Plaut et al., 2012; Anderegg et al., 2013; Paddock et al., 2013). ...
... 栓塞脆弱曲线的建立使得人们可以评估植物对栓塞的抵抗力, 有些学者分析了植物木质部栓塞脆弱性与耐旱性的关系(Sperry & Tyree, 1990; Cochard et al., 1992, 2008, 2013; Tyree et al., 2003; Maherali et al., 2004), 认为由植物木质部空穴化和栓塞导致的植物输水结构功能障碍影响了植物的气孔运动及耐旱能力(Sperry et al., 1988; Salleo et al., 2000).栓塞脆弱性决定了干旱地区生态系统中不同物种的生存模式(Davis et al., 1998; Pockman & Sperry, 2000), 但栓塞抗性在不同物种和不同森林生态系统的表现不同(Maherali et al., 2004; Delzon et al., 2010), 因此, 在严重干旱情况下裸子植物和被子植物存活率也不同(Brodribb & Cochard, 2009; Brodribb et al., 2010). ...
Size and function in conifer tracheids and angiosperm vessels.
2
2006
... 木质部空穴化和栓塞的发现为植物输水结构体系和植物耐旱机理研究提供了新的思路和方向, 植物对木质部栓塞的易受性可用“脆弱曲线(VC)”来描述(Tyree & Sperry, 1989), 该曲线是根据木质部导水率损失的百分数(PLC)与相对应水势绘制而成的曲线, 且已被广泛引用(Maherali et al., 2004).栓塞脆弱曲线的建立可以使学者们对植物抵御栓塞抗性的大小进行评估, 在研究了木质部栓塞脆弱性与输水结构间的关系(Wheeler et al., 2005; Sperry et al., 2006; Hacke & Jansen, 2009; Cai & Tyree, 2010; 张海昕等, 2013)及植物木质部栓塞脆弱性与耐旱性的关系(Tyree et al., 2003; Maherali et al., 2004; Awad et al., 2010)后, 学者们认为由植物木质部空穴化和栓塞导致的木本植物输水结构功能障碍影响其自身耐旱能力.脆弱曲线与植物的部位、木质部结构、分布、抗旱性等存在一定关系, 因此能够提供许多抗旱信息(安锋和张硕新, 2005).木质部栓塞脆弱性与输水结构间的关系可使人们评估植物对栓塞的抵抗力大小, 从而进一步探究木质部栓塞脆弱性和耐旱性的关系, 但各学者对不同树种研究所得结论不尽相同. ...
... 输水结构被用来描述整个树体上茎段导水率与其结构间的关系(Zimmermann, 1983).栓塞使木质部管道中充有气泡而阻断了水分的运输, 因此, 输水结构的输水效率会受到木质部栓塞脆弱性的影响.木质部栓塞脆弱性与导管或管胞的解剖特征和纹孔超微结构密切相关(Wheeler et al., 2005; Sperry et al., 2006; Hacke & Jansen, 2009). ...
Water-stress-induced xylem embolism in three species of conifers.
1
1990
... 栓塞脆弱曲线的建立使得人们可以评估植物对栓塞的抵抗力, 有些学者分析了植物木质部栓塞脆弱性与耐旱性的关系(Sperry & Tyree, 1990; Cochard et al., 1992, 2008, 2013; Tyree et al., 2003; Maherali et al., 2004), 认为由植物木质部空穴化和栓塞导致的植物输水结构功能障碍影响了植物的气孔运动及耐旱能力(Sperry et al., 1988; Salleo et al., 2000).栓塞脆弱性决定了干旱地区生态系统中不同物种的生存模式(Davis et al., 1998; Pockman & Sperry, 2000), 但栓塞抗性在不同物种和不同森林生态系统的表现不同(Maherali et al., 2004; Delzon et al., 2010), 因此, 在严重干旱情况下裸子植物和被子植物存活率也不同(Brodribb & Cochard, 2009; Brodribb et al., 2010). ...
Drought induced xylem embolism in four riparian trees from the Western Cape Province: Insights and implications for planning and evaluation of restoration.
1
2008
... 依据Domec理论(Domec & Gartner, 2001), 测定和分析导水率损失最大值时的枝条临界水势和导水率50%时对应的枝条水势(P50)值(P50为负值), 可以很好地描述不同树种木质部的栓塞脆弱性、比较不同树种木质部抵抗栓塞的能力(Choat et al., 2012).但也有研究表明, 植物栓塞后的剩余导水率可能比P50值更适合描述植物在水分亏缺条件下抵抗栓塞的能力(Pammenter & van der Willigen, 1998; Swift et al., 2008).目前, 国际上大多数学者用P50值来表示栓塞脆弱性. ...
Arabidopsis thaliana as a model species for xylem hydraulics: Does size matter?
1
2013
... 栓塞脆弱性可通过栓塞脆弱曲线来反应, 该曲线的建立有许多方法, 早期常用的是自然干燥法(bench dehydration)、空气注入法(air-injection)和压力室法(pressure chamber)(Cochard et al., 1992), 目前国际上常用的快速测定栓塞脆弱曲线的方法是Cochard Cavitron离心机法(Cochard, 2002; Cochard et al., 2005)和Sperry离心机技术(Sperry et al., 1988).自然干燥法是最直接、最原始的诱导植物产生空穴化和栓塞的方法, 将植物(通常为带有叶片且长于1 m的枝条)不做任何处理直接放在空气中进行脱水干燥, 木质部压力可用压力势法测量未蒸腾的叶片而得(Tyree et al., 1992).空气注入法是将压缩的空气通过压力套注入木质部导管中, 木质部的导水率随压力的变化而变化, 从而建立枝条的栓塞脆弱曲线(Cochard et al., 1992).离心机法是将枝条放在离心机中旋转, 其原理是利用不断增大的离心力模拟活体植株中的木质部负压来诱导枝条产生栓塞, 从而测定枝条导水率损失值(PLC)(Pockman et al., 1995).不同之处是: 1)自然干燥法建立脆弱曲线需要数周时间和多个枝条才能得到, 因此比较费时, 而且原位诱导产生的栓塞可能影响导水率与木质部负压的关系, 并且最近研究发现, 在被切开的木质部导管压力下该方法易于建立错误曲线(Li et al., 2009; Wheeler et al., 2013; Torres-Ruiz et al., 2015).2)空气注入法的优势在于水分可以在几分钟内精确地调整到人为想要控制的程度, 相比于自然干燥法, 该方法可以在数小时内建立一条完整的脆弱曲线, 但实际操作中压力套在高压下会发生漏气现象, 同时注入导管内的空气很可能影响导水率, 所以需要等压力恢复到大气压时才能进行测定(Cochard et al., 2010).空气注入法和自然干燥法更适合于建立长导管植物脆弱曲线(Choat et al., 2010).3) Cochard Cavitron离心机是在Pockman离心机(Pockman et al., 1995)的基础上改装而来的, 它具有一个大转子(所容纳枝条长度为27.4 cm, 转子长27.5 cm), Sperry离心机具有一个小转子(所容纳枝条长度为14.5 cm), 不同之处是Cochard Cavitron离心机法仅需要一根枝条且通常在0.5 h内就可建立一条完整的脆弱曲线, 而Sperry离心机技术则需要3-5根枝条、一周时间才能建立一条完整的脆弱曲线, 离心机法具有空气注入法的优点, 且不用考虑由于已经存在的空气带来的不便, 此外, 离心机法还可以用于测定许多草本植物脆弱曲线, 这一点空气注入法是无法完成的(Tixier et al., 2013).离心机法所得脆弱曲线可将植物生理生态特性和木质部结构结合起来(Hacke et al., 2001, 2006; Jacobsen et al., 2005; Cai & Tyree, 2010; Lens et al., 2011), 可筛选出杨属(Populus)和柳属(Salix)栓塞抗性强的树种 (Cochard et al., 2007; Arango-Velez et al., 2011; Schreiber et al., 2011), 可将抗旱性和生活史结合起来评估植物抵御干旱的能力(Pratt et al., 2007), 也可用来探究干旱环境下植物的死亡率(Choat et al., 2012; Plaut et al., 2012; Anderegg et al., 2013; Paddock et al., 2013). ...
Vulnerability to cavitation in Olea europaea current-year shoots: Further evidence of an open-vessel artifact associated with centrifuge and air-injection techniques.
0
2014
Direct micro-CT observation confirms the induction of embolism upon xylem cutting under tension.
2
2015
... 栓塞脆弱性可通过栓塞脆弱曲线来反应, 该曲线的建立有许多方法, 早期常用的是自然干燥法(bench dehydration)、空气注入法(air-injection)和压力室法(pressure chamber)(Cochard et al., 1992), 目前国际上常用的快速测定栓塞脆弱曲线的方法是Cochard Cavitron离心机法(Cochard, 2002; Cochard et al., 2005)和Sperry离心机技术(Sperry et al., 1988).自然干燥法是最直接、最原始的诱导植物产生空穴化和栓塞的方法, 将植物(通常为带有叶片且长于1 m的枝条)不做任何处理直接放在空气中进行脱水干燥, 木质部压力可用压力势法测量未蒸腾的叶片而得(Tyree et al., 1992).空气注入法是将压缩的空气通过压力套注入木质部导管中, 木质部的导水率随压力的变化而变化, 从而建立枝条的栓塞脆弱曲线(Cochard et al., 1992).离心机法是将枝条放在离心机中旋转, 其原理是利用不断增大的离心力模拟活体植株中的木质部负压来诱导枝条产生栓塞, 从而测定枝条导水率损失值(PLC)(Pockman et al., 1995).不同之处是: 1)自然干燥法建立脆弱曲线需要数周时间和多个枝条才能得到, 因此比较费时, 而且原位诱导产生的栓塞可能影响导水率与木质部负压的关系, 并且最近研究发现, 在被切开的木质部导管压力下该方法易于建立错误曲线(Li et al., 2009; Wheeler et al., 2013; Torres-Ruiz et al., 2015).2)空气注入法的优势在于水分可以在几分钟内精确地调整到人为想要控制的程度, 相比于自然干燥法, 该方法可以在数小时内建立一条完整的脆弱曲线, 但实际操作中压力套在高压下会发生漏气现象, 同时注入导管内的空气很可能影响导水率, 所以需要等压力恢复到大气压时才能进行测定(Cochard et al., 2010).空气注入法和自然干燥法更适合于建立长导管植物脆弱曲线(Choat et al., 2010).3) Cochard Cavitron离心机是在Pockman离心机(Pockman et al., 1995)的基础上改装而来的, 它具有一个大转子(所容纳枝条长度为27.4 cm, 转子长27.5 cm), Sperry离心机具有一个小转子(所容纳枝条长度为14.5 cm), 不同之处是Cochard Cavitron离心机法仅需要一根枝条且通常在0.5 h内就可建立一条完整的脆弱曲线, 而Sperry离心机技术则需要3-5根枝条、一周时间才能建立一条完整的脆弱曲线, 离心机法具有空气注入法的优点, 且不用考虑由于已经存在的空气带来的不便, 此外, 离心机法还可以用于测定许多草本植物脆弱曲线, 这一点空气注入法是无法完成的(Tixier et al., 2013).离心机法所得脆弱曲线可将植物生理生态特性和木质部结构结合起来(Hacke et al., 2001, 2006; Jacobsen et al., 2005; Cai & Tyree, 2010; Lens et al., 2011), 可筛选出杨属(Populus)和柳属(Salix)栓塞抗性强的树种 (Cochard et al., 2007; Arango-Velez et al., 2011; Schreiber et al., 2011), 可将抗旱性和生活史结合起来评估植物抵御干旱的能力(Pratt et al., 2007), 也可用来探究干旱环境下植物的死亡率(Choat et al., 2012; Plaut et al., 2012; Anderegg et al., 2013; Paddock et al., 2013). ...
... 目前, 对于一些树种的相同部位材料采用不同方法所得脆弱曲线不同, 特别是对离心机法建立的长导管植物的栓塞脆弱曲线正确与否存在很大争议(Cochard et al., 2005, 2010, 2013; Sperry et al., 2012; Torres-Ruiz et al., 2015).离心机法建立的脆弱曲线有两种形态, 一种为“r”形, 一种为“s”形, “r”形脆弱曲线意味着在水势较高(稍负)时或稍降时, 植物即产生很大的栓塞或导水率损失, “s”形脆弱曲线则表示只有当水分亏缺达到一定程度才会发生栓塞.因此, 脆弱曲线的形式直接影响导水率损失值(P50).有关脆弱曲线的争议主要表现在以下几方面: ...
Loss of hydraulic conductivity due to water stress in intact juveniles of Quercus rubra and Populus deltoides.
1
1992
... 栓塞脆弱性可通过栓塞脆弱曲线来反应, 该曲线的建立有许多方法, 早期常用的是自然干燥法(bench dehydration)、空气注入法(air-injection)和压力室法(pressure chamber)(Cochard et al., 1992), 目前国际上常用的快速测定栓塞脆弱曲线的方法是Cochard Cavitron离心机法(Cochard, 2002; Cochard et al., 2005)和Sperry离心机技术(Sperry et al., 1988).自然干燥法是最直接、最原始的诱导植物产生空穴化和栓塞的方法, 将植物(通常为带有叶片且长于1 m的枝条)不做任何处理直接放在空气中进行脱水干燥, 木质部压力可用压力势法测量未蒸腾的叶片而得(Tyree et al., 1992).空气注入法是将压缩的空气通过压力套注入木质部导管中, 木质部的导水率随压力的变化而变化, 从而建立枝条的栓塞脆弱曲线(Cochard et al., 1992).离心机法是将枝条放在离心机中旋转, 其原理是利用不断增大的离心力模拟活体植株中的木质部负压来诱导枝条产生栓塞, 从而测定枝条导水率损失值(PLC)(Pockman et al., 1995).不同之处是: 1)自然干燥法建立脆弱曲线需要数周时间和多个枝条才能得到, 因此比较费时, 而且原位诱导产生的栓塞可能影响导水率与木质部负压的关系, 并且最近研究发现, 在被切开的木质部导管压力下该方法易于建立错误曲线(Li et al., 2009; Wheeler et al., 2013; Torres-Ruiz et al., 2015).2)空气注入法的优势在于水分可以在几分钟内精确地调整到人为想要控制的程度, 相比于自然干燥法, 该方法可以在数小时内建立一条完整的脆弱曲线, 但实际操作中压力套在高压下会发生漏气现象, 同时注入导管内的空气很可能影响导水率, 所以需要等压力恢复到大气压时才能进行测定(Cochard et al., 2010).空气注入法和自然干燥法更适合于建立长导管植物脆弱曲线(Choat et al., 2010).3) Cochard Cavitron离心机是在Pockman离心机(Pockman et al., 1995)的基础上改装而来的, 它具有一个大转子(所容纳枝条长度为27.4 cm, 转子长27.5 cm), Sperry离心机具有一个小转子(所容纳枝条长度为14.5 cm), 不同之处是Cochard Cavitron离心机法仅需要一根枝条且通常在0.5 h内就可建立一条完整的脆弱曲线, 而Sperry离心机技术则需要3-5根枝条、一周时间才能建立一条完整的脆弱曲线, 离心机法具有空气注入法的优点, 且不用考虑由于已经存在的空气带来的不便, 此外, 离心机法还可以用于测定许多草本植物脆弱曲线, 这一点空气注入法是无法完成的(Tixier et al., 2013).离心机法所得脆弱曲线可将植物生理生态特性和木质部结构结合起来(Hacke et al., 2001, 2006; Jacobsen et al., 2005; Cai & Tyree, 2010; Lens et al., 2011), 可筛选出杨属(Populus)和柳属(Salix)栓塞抗性强的树种 (Cochard et al., 2007; Arango-Velez et al., 2011; Schreiber et al., 2011), 可将抗旱性和生活史结合起来评估植物抵御干旱的能力(Pratt et al., 2007), 也可用来探究干旱环境下植物的死亡率(Choat et al., 2012; Plaut et al., 2012; Anderegg et al., 2013; Paddock et al., 2013). ...
Desiccation tolerance of five tropical seedlings in Panama. Relationship to a field assessment of drought performance.
3
2003
... 木质部空穴化和栓塞的发现为植物输水结构体系和植物耐旱机理研究提供了新的思路和方向, 植物对木质部栓塞的易受性可用“脆弱曲线(VC)”来描述(Tyree & Sperry, 1989), 该曲线是根据木质部导水率损失的百分数(PLC)与相对应水势绘制而成的曲线, 且已被广泛引用(Maherali et al., 2004).栓塞脆弱曲线的建立可以使学者们对植物抵御栓塞抗性的大小进行评估, 在研究了木质部栓塞脆弱性与输水结构间的关系(Wheeler et al., 2005; Sperry et al., 2006; Hacke & Jansen, 2009; Cai & Tyree, 2010; 张海昕等, 2013)及植物木质部栓塞脆弱性与耐旱性的关系(Tyree et al., 2003; Maherali et al., 2004; Awad et al., 2010)后, 学者们认为由植物木质部空穴化和栓塞导致的木本植物输水结构功能障碍影响其自身耐旱能力.脆弱曲线与植物的部位、木质部结构、分布、抗旱性等存在一定关系, 因此能够提供许多抗旱信息(安锋和张硕新, 2005).木质部栓塞脆弱性与输水结构间的关系可使人们评估植物对栓塞的抵抗力大小, 从而进一步探究木质部栓塞脆弱性和耐旱性的关系, 但各学者对不同树种研究所得结论不尽相同. ...