Effects of external nitrogen additions on soil organic carbon dynamics and the mechanism
LI Rong1, 2, CHANG Rui-Ying1, *,
1Key Laboratory of Mountain Surface Processes and Ecological Regulation, Institute of Mountain Hazards and Environment, Chinese Academy of Sciences, Chengdu 610041, China2University of Chinese Academy of Sciences, Beijing 100049, China
What would be the impact of external nitrogen additions on soil carbon, an issue still under debating, as reported experimental results were either positive, negtive or neutral. Several factors may be related to these seemingly controversial results: differences in ecosystem types and soil properties, soil carbon detection methods, soil depths, and contents of soil labile and recalcitrant carbon that affect the responses to nitrogen additions, all could cause discrepancies and variations in carbon sequestration. The several processes that contribute to enhance soil organic carbon storage include increasing litter input, decresing soil carbon output, particularly, by supressed decomposition of recalcitrant carbon, promoting soil humifiction and formation of recalcitrant carbon storage. However, there are still many uncertainties associated with these issues. To improve our understanding, the research about carbon in deep soil layers, dissolved organic carbon leaching and accumulation, and the effect of labile and recalcitrant soil C ratios on N addition responses, should be further investigated in the future studies.
LIRong, CHANGRui-Ying. Effects of external nitrogen additions on soil organic carbon dynamics and the mechanism. Chinese Journal of Plant Ecology, 2015, 39(10): 1012-1020 https://doi.org/10.17521/cjpe.2015.0098
Fig. 1 Effects of soil nitrogen additions on soil organic carbon storage and cycle. Solid arrows indicate interaction of the input and output of soil organic carbon, and the soil organic carbon storage. Hollow arrows indicate the effect of nitrogen additions. Upward, downward, and parallel arrows show positive, negative, and insignificant effects. The question mark indicates little research was done, or an unanswered question.
Nitrogen deposition impacts on the amount and stability of soil organic matter in an alpine meadow ecosystem depend on the form and rate of applied nitrogen.
Early effect of elevated nitrogen input on above-ground net primary production of a lower montane rain forest, Panama.
1
2009
... 一些研究显示施氮可显著增加植被凋落物量(Bradley et al., 2006; Adamek et al., 2009), 且凋落物量随氮添加水平增加而增加.例如, 樊后保等(2007)研究指出: 高氮添加(240 kg N∙hm-2∙a-1)显著增加森林凋落物量(2599.5 kg∙hm-2∙a-1), 而中氮(120 kg N∙hm-2∙a-1)和低氮(60 kg N∙hm-2∙a-1)对凋落物量增加的促进作用不明显.但也有研究发现: 尽管氮添加可以促进植被生产力增加, 但地上叶片与地下根系凋落物输入对氮添加并无明显的正反馈作用(Zak et al., 2008; Janssens et al., 2010).此外, 氮添加可以增加叶片和细根氮含量(Giardina et al., 2003; Xia & Wan, 2008; Zak et al., 2008; Yuan & Chen, 2012), 可能促进叶片和细根凋落物的分解(Waldrop et al., 2004; Knorr et al., 2005; Parton et al., 2007; Hobbie et al., 2012), 而不利于土壤碳的积累.然而, 有研究显示氮添加只在初期促进凋落物分解, 主要包括水溶性物质和非木质性纤维素的分解, 随着中后期难分解的木质素含量的增多, 氮添加抑制了凋落物的分解(Carreiro et al., 2000). ...
Combining theory and experiment to understand effects of inorganic nitrogen on litter decomposition.
1
2001
... 此外, 有研究认为土壤微生物的碳利用效率(CUE)与土壤碳积累有关(Six et al., 2006).微生物的碳利用效率是指用于微生物生物量积累的碳与微生物所吸收的碳之比(del Giorgio & Cole, 1998).CUE越高表明微生物用于自身有效生长的碳越多, 反之则表明有更多的碳用于微生物呼吸和分泌物的产生, 从而导致碳损耗, 不利于土壤碳积累.土壤微生物的碳利用效率受多种因素影响, 如高温和水分胁迫可通过影响微生物活动和呼吸作用而降低CUE (Conant et al., 2011; Manzoni et al., 2012).氮添加则可通过影响微生物的合成代谢与分解代谢影响CUE.研究认为, 氮添加可提高微生物CUE, 减弱微生物呼吸作用(Blagodatskaya et al., 2014), 从而促进土壤碳积累(Thiet et al., 2006; Manzoni et al., 2012), 也可通过影响土壤胞外酶活性降低稳定性碳分解(Ågren et al., 2001). ...
Litter decomposition and organic matter turnover in northern forest soils.
1
2000
... 氮添加可促进腐殖质的形成, 促进分解残物向稳定性碳的转变, 增加稳定性碳积累(Moran et al., 2005; Whittinghill et al., 2012).氮元素与较难分解的凋落物残体(如木质素等)结合形成更难分解的杂环类物质(如吲哚等)和酚类是氮添加促进难分解碳形成的一个机制(Berg, 2000; Janssens et al., 2010).而氮添加对团聚体(aggregate, 即土壤碳的物理保护)的促进作用可能不明显(Janssens et al., 2010).尽管氮添加对稳定性碳库的促进作用在短期内可能对土壤总有机碳库影响不显著, 但其长期作用明显(Reid et al., 2012).土壤稳定性碳是影响土壤有机碳库稳定与积累的重要因素, 加强对土壤稳定性碳的研究对于从机理上深入认识氮添加对土壤有机碳库的影响及其响应机制具有重要作用. ...
The use of stable carbon isotopes for estimating soil organic matter turnover rates.
1
1998
... 第四, 土壤碳储量巨大, 但其在氮添加下的变化量相对微小.如何减小其变化量的检测误差, 是当前相关研究需重点关注的一个方面.采取连续观测法并通过严格选择采样点可减小测量误差(Ellert et al., 2002; VandenBygaart & Kay, 2004).相比于传统的样地对比法, 碳同位素(13C和14C)示踪方法可以更加敏感地揭示土壤碳的动态变化(de Camargo et al., 1999).碳的稳定性同位素(13C)示踪技术可以阐明土壤碳储量的迁移和转化, 揭示不同形成时期新老土壤有机碳对碳储量的贡献(Bernoux et al., 1998).放射性同位素(14C)可以反映土壤固定和释放碳的时间, 可用于研究不同时间尺度土壤有机碳的动力学机制(Trumbore, 2000).常用的估算土壤有机碳库更新速率的方法包括以下两个方法: 1)将14C作为土壤有机碳更新速率的指示剂, 通过检测1950年以来土壤有机碳中的14C量, 推算土壤碳动力学过程(Hsieh, 1993; Trumbore et al., 1996; Wang et al., 1999); 2)根据大气层残留核爆14C来识别土壤有机碳库的驻留时间(MRT)(Hsieh, 1993).如利用加速质谱仪检出土壤中14C含量在短期内的变化, 可以间接反映土壤碳的分解与更新速率.同时, 15N标记在氮添加实验中的应用不仅可以有效地检验氮素的循环过程, 而且可以在较短时期内(可以假定C:N不变)通过检验不同碳库中的15N含量及碳氮比, 并结合氮平衡法(N-balance method, 测定15N在土壤中的积累量并结合土壤C:N来估算土壤碳的变化量)来检测氮添加下土壤碳变化(de Vries et al., 2006; Mol et al., 2009).此外, 碳氮同位素的应用还可以有效地示踪土壤有机碳的来源和周转过程(Ehleringer et al., 2000), 揭示土壤碳氮耦合变化关系, 这也必将成为今后相关研究的重要方面. ...
Microbial growth and carbon use efficiency in the rhizosphere and root-free soil.
1
2014
... 此外, 有研究认为土壤微生物的碳利用效率(CUE)与土壤碳积累有关(Six et al., 2006).微生物的碳利用效率是指用于微生物生物量积累的碳与微生物所吸收的碳之比(del Giorgio & Cole, 1998).CUE越高表明微生物用于自身有效生长的碳越多, 反之则表明有更多的碳用于微生物呼吸和分泌物的产生, 从而导致碳损耗, 不利于土壤碳积累.土壤微生物的碳利用效率受多种因素影响, 如高温和水分胁迫可通过影响微生物活动和呼吸作用而降低CUE (Conant et al., 2011; Manzoni et al., 2012).氮添加则可通过影响微生物的合成代谢与分解代谢影响CUE.研究认为, 氮添加可提高微生物CUE, 减弱微生物呼吸作用(Blagodatskaya et al., 2014), 从而促进土壤碳积累(Thiet et al., 2006; Manzoni et al., 2012), 也可通过影响土壤胞外酶活性降低稳定性碳分解(Ågren et al., 2001). ...
Negative impact of nitrogen deposition on soil buffering capacity.
1
2008
... 氮素是生物必需的大量元素, 对调控生态系统结构和功能, 维持生态系统的健康和稳定具有重要意义.自工业革命以来, 由于人类活动, 如化肥、化石燃料使用等, 全球范围内的氮生产及大气活性氮沉降均大幅增加(Galloway et al., 2008; Cui et al., 2013).据测算, 全球表面平均大气氮沉降从工业革命前的0.5 kg N·hm-2·a-1增加到当前的10 kg N·hm-2·a-1 (Erisman et al., 2008), 而我国当前的大气氮沉降远高于全球平均水平, 约为21.1 kg N·hm-2·a-1 (Liu et al., 2013), 并且仍将持续增加(Galloway et al., 2008; Liu et al., 2013).大气氮沉降增加作为当前全球变化的一个重要方面, 伴随着大气CO2浓度增加和气温升高, 对生态系统结构、功能和服务产生了深刻影响(Mack et al., 2004; Suding et al., 2005; Magnani et al., 2007; Bowman et al., 2008; Janssens et al., 2010). ...
Increased N availability in grassland soils modifies their microbial communities and decreases the abundance of arbuscular mycorrhizal fungi.
1
2006
... 一些研究显示施氮可显著增加植被凋落物量(Bradley et al., 2006; Adamek et al., 2009), 且凋落物量随氮添加水平增加而增加.例如, 樊后保等(2007)研究指出: 高氮添加(240 kg N∙hm-2∙a-1)显著增加森林凋落物量(2599.5 kg∙hm-2∙a-1), 而中氮(120 kg N∙hm-2∙a-1)和低氮(60 kg N∙hm-2∙a-1)对凋落物量增加的促进作用不明显.但也有研究发现: 尽管氮添加可以促进植被生产力增加, 但地上叶片与地下根系凋落物输入对氮添加并无明显的正反馈作用(Zak et al., 2008; Janssens et al., 2010).此外, 氮添加可以增加叶片和细根氮含量(Giardina et al., 2003; Xia & Wan, 2008; Zak et al., 2008; Yuan & Chen, 2012), 可能促进叶片和细根凋落物的分解(Waldrop et al., 2004; Knorr et al., 2005; Parton et al., 2007; Hobbie et al., 2012), 而不利于土壤碳的积累.然而, 有研究显示氮添加只在初期促进凋落物分解, 主要包括水溶性物质和非木质性纤维素的分解, 随着中后期难分解的木质素含量的增多, 氮添加抑制了凋落物的分解(Carreiro et al., 2000). ...
... 一些研究显示施氮可显著增加植被凋落物量(Bradley et al., 2006; Adamek et al., 2009), 且凋落物量随氮添加水平增加而增加.例如, 樊后保等(2007)研究指出: 高氮添加(240 kg N∙hm-2∙a-1)显著增加森林凋落物量(2599.5 kg∙hm-2∙a-1), 而中氮(120 kg N∙hm-2∙a-1)和低氮(60 kg N∙hm-2∙a-1)对凋落物量增加的促进作用不明显.但也有研究发现: 尽管氮添加可以促进植被生产力增加, 但地上叶片与地下根系凋落物输入对氮添加并无明显的正反馈作用(Zak et al., 2008; Janssens et al., 2010).此外, 氮添加可以增加叶片和细根氮含量(Giardina et al., 2003; Xia & Wan, 2008; Zak et al., 2008; Yuan & Chen, 2012), 可能促进叶片和细根凋落物的分解(Waldrop et al., 2004; Knorr et al., 2005; Parton et al., 2007; Hobbie et al., 2012), 而不利于土壤碳的积累.然而, 有研究显示氮添加只在初期促进凋落物分解, 主要包括水溶性物质和非木质性纤维素的分解, 随着中后期难分解的木质素含量的增多, 氮添加抑制了凋落物的分解(Carreiro et al., 2000). ...
... 氮添加不仅对土壤微生物呼吸产生影响, 而且可以促使土壤微生物群落结构发生改变(Waldrop et al., 2004; Carreiro et al., 2000).土壤微生物对陆地生态系统土壤碳矿化过程及营养元素循环具有重要影响, 由土壤微生物活动、植物根系分泌物和动植物残体腐解过程所产生的土壤酶, 包括游离酶、胞内酶和胞外酶等的活性与土壤有机质的分解合成以及土壤供应植物养分的能力密切相关(关松荫, 1986).研究认为氮添加可改变土壤细菌与真菌组成比例(Demoling et al., 2008; Ramirez et al., 2012), 并通过影响水解酶和氧化还原酶的活性而影响稳定性碳的分解, 如促进纤维素和多糖类等有机质的分解, 抑制木质素等其他难分解有机质的分解(Carreiro et al., 2000; Hobbie et al., 2012; Ramirez et al., 2012).此外, 分子生物学的发展为微生物生态学的研究提供了新的手段.Cusack等(2010)利用磷脂脂肪酸分析和13C核磁共振技术对热带雨林的研究发现, 氮添加通过影响水解酶及氧化酶活性促进土壤中活性碳的分解.此外, 高通量测序技术、RT-PCR技术、PCR-DGGE技术、基于rDNA指纹图谱等的应用, 也将有助于深入认识氮添加下土壤有机碳变化的微生物学机制. ...
... ), 并通过影响水解酶和氧化还原酶的活性而影响稳定性碳的分解, 如促进纤维素和多糖类等有机质的分解, 抑制木质素等其他难分解有机质的分解(Carreiro et al., 2000; Hobbie et al., 2012; Ramirez et al., 2012).此外, 分子生物学的发展为微生物生态学的研究提供了新的手段.Cusack等(2010)利用磷脂脂肪酸分析和13C核磁共振技术对热带雨林的研究发现, 氮添加通过影响水解酶及氧化酶活性促进土壤中活性碳的分解.此外, 高通量测序技术、RT-PCR技术、PCR-DGGE技术、基于rDNA指纹图谱等的应用, 也将有助于深入认识氮添加下土壤有机碳变化的微生物学机制. ...
... 研究表明, 下层碳可能受团聚体的物理保护(隔离了微生物的侵袭), 活性碳输入不会造成下层有机碳由于激发效应产生的损失.激发效应对土壤碳的影响程度和作用时间与生态系统类型、活性碳类型、观测时间以及土壤氮供给水平等密切相关(Cheng et al., 2003; Dijkstra & Cheng, 2007; Drake et al., 2013).目前对淋溶碳输入是否会引发土壤激发效应仍缺乏实证, 需要结合对土壤剖面可溶性有机碳的淋溶过程的观测深入开展研究.因此, 土壤剖面淋溶碳的相关研究对于认识土壤有机碳, 尤其是下层土壤碳在氮素添加下的变化机制具有重要作用. ...
Nutrient additions to a tropical rain forest drive substantial soil carbon dioxide losses to the atmosphere.
2
2006
... 相对于植被碳汇, 目前关于土壤碳汇对氮添加的响应争议较大.Mack等(2004)研究发现氮添加改变了苔原地区植物群落组成并促进了凋落物分解, 从而使土壤有机碳储量减少, 抵消了氮添加下的植被碳汇增量, 导致生态系统碳储量减少.同样, Cleveland和Townsend (2006)研究发现, 氮添加促进土壤呼吸而减小土壤有机碳库储量.Nadelhoffer等(1999)则认为氮添加对欧洲森林生态系统碳库影响较小.此外, 较多研究显示氮添加增加了土壤碳储量(de Vries et al., 2006; Reay et al., 2008).例如, Zak等(2008)指出氮添加(30 kg N∙hm-2∙a-1)处理的森林土壤有机碳含量显著高于对照土壤, 其增幅约12%.氮添加同样可促进北欧云杉林和松树林土壤碳吸收, 其碳汇增量为11 g C (Hyvönen et al., 2008).Reay等(2008)对以往的研究进行了综述, 认为氮添加下的土壤碳库增量为7-23 g C.Meta分析的结果表明, 森林土壤在氮添加下的碳汇增量为19 g C (Janssens et al., 2010); 而Lu等(2011)的结果显示森林和草地的土壤碳库变化很小.经过文献梳理和总结, 氮添加下土壤碳汇增量范围为0-30 g C, 低于植被碳汇增量(de Vries et al., 2006, 2008; Reay et al., 2008; Janssens et al., 2010).整体而言, 目前关于氮添加条件下土壤碳汇的变化方向与强度争议较大, 亟需对其影响因子及机理进行深入研究分析. ...
Temperature and soil organic matter decomposition rates—synthesis of current knowledge and a way forward.
1
2011
... 此外, 有研究认为土壤微生物的碳利用效率(CUE)与土壤碳积累有关(Six et al., 2006).微生物的碳利用效率是指用于微生物生物量积累的碳与微生物所吸收的碳之比(del Giorgio & Cole, 1998).CUE越高表明微生物用于自身有效生长的碳越多, 反之则表明有更多的碳用于微生物呼吸和分泌物的产生, 从而导致碳损耗, 不利于土壤碳积累.土壤微生物的碳利用效率受多种因素影响, 如高温和水分胁迫可通过影响微生物活动和呼吸作用而降低CUE (Conant et al., 2011; Manzoni et al., 2012).氮添加则可通过影响微生物的合成代谢与分解代谢影响CUE.研究认为, 氮添加可提高微生物CUE, 减弱微生物呼吸作用(Blagodatskaya et al., 2014), 从而促进土壤碳积累(Thiet et al., 2006; Manzoni et al., 2012), 也可通过影响土壤胞外酶活性降低稳定性碳分解(Ågren et al., 2001). ...
Centennial-scale analysis of the creation and fate of reactive nitrogen in China (1910-2010).
1
2013
... 氮素是生物必需的大量元素, 对调控生态系统结构和功能, 维持生态系统的健康和稳定具有重要意义.自工业革命以来, 由于人类活动, 如化肥、化石燃料使用等, 全球范围内的氮生产及大气活性氮沉降均大幅增加(Galloway et al., 2008; Cui et al., 2013).据测算, 全球表面平均大气氮沉降从工业革命前的0.5 kg N·hm-2·a-1增加到当前的10 kg N·hm-2·a-1 (Erisman et al., 2008), 而我国当前的大气氮沉降远高于全球平均水平, 约为21.1 kg N·hm-2·a-1 (Liu et al., 2013), 并且仍将持续增加(Galloway et al., 2008; Liu et al., 2013).大气氮沉降增加作为当前全球变化的一个重要方面, 伴随着大气CO2浓度增加和气温升高, 对生态系统结构、功能和服务产生了深刻影响(Mack et al., 2004; Suding et al., 2005; Magnani et al., 2007; Bowman et al., 2008; Janssens et al., 2010). ...
The response of heterotrophic activity and carbon cycling to nitrogen additions and warming in two tropical soils.
4
2010
... 第一, 与生态系统类型及其性质、土壤类型及氮素添加形式、时间长短及添加水平有关(Hyvönen et al., 2008; de Vries et al., 2008; Cusack et al., 2010).氮添加能够显著地促进农田生态系统土壤有机碳积累, 而对林地、草地、湿地及荒漠等非农业生态系统土壤碳积累的作用不明显(Lu et al., 2011).同样, 相比于温带针叶林(temperate conifer forest)和温带混合林(temperate mixed forest)及热带雨林(tropical rain forest), 氮添加对北方泰加林(boreal forest)矿质层土壤有机碳影响更为显著(Liu & Greaver, 2010).不同生态系统土壤有机碳对氮添加的响应差异可能与其本身的性质有关, 如土壤氮含量及碳氮元素计量关系、凋落物质量等(Waldrop et al., 2004; Kirkby et al., 2013).Waldrop等(2004)发现: 在3年的氮添加(80 kg N∙hm-2∙a-1)实验中, 凋落物质量较高(C:N较低)的Acer saccharum林0-20 cm土壤碳下降了20%, 而凋落物质量较低(C:N较高)的Quercus velutina林在同一深度的土壤碳则显著提高了10%.此外, 不同氮添加形式对土壤有机碳具有不同影响, 氨态氮对土壤有机碳累积的促进作用明显高于硝态氮(Lu et al., 2011), 其原因可能是氨态氮促进土壤pH值和微生物酶活性的降低, 不利于土壤有机碳的矿化(Min et al., 2011).微生物群落与土壤有机碳对氮添加的响应存在临界阈值, 长期施氮会使受氮限制的生态系统发生氮饱和, NO3-大量淋溶, 土壤酸化、大量土壤阳离子流失并产生铝毒(Tietema, 1998), 对微生物活性产生不利影响(Lovell & Hatch, 1997; Wallenstein et al., 2006); 而短期施氮可促进土壤碳积累.但也有研究表明: 短期氮添加可能会增加或降低土壤微生物生物量, 或者对其无影响(Dijkstra et al., 2005; Liu et al., 2007), 从而对土壤碳库产生不同影响.适量施氮可促进土壤碳积累, 但过量施氮可能会使有机质分解作用强于其累积, 从而降低土壤碳储量(Soussana et al., 2004).例如, 低氮添加(10 kg N∙hm-2∙a-1)促进土壤碳积累, 而中氮(20 kg N∙hm-2∙a-1)和高氮添加(40 kg N∙hm-2∙a-1)反而会降低土壤碳储量(Fang et al., 2014). ...
... 第三, 氮添加对土壤有机碳影响的差异还可能与不同土壤碳组分, 即易变碳与稳定性碳的差异性响应有关.Neff等(2002)发现, 氮添加(10 g N∙m-2∙a-1)促进了较易分解的轻组土壤碳的分解, 而增加了较难分解的重组土壤碳的稳定性.并且, 不同土壤碳组分对氮添加的响应时间也存在差异.例如, 易变碳可能在短期内就对氮添加做出响应, 促使土壤易变碳库减少(Dijkstra et al., 2005), 而稳定性碳对氮添加响应较为缓慢(Cusack et al., 2010).土壤易变碳与稳定性碳的分布和格局与植被和土壤类型以及土壤深度密切相关, 二者对氮添加的差异性响应可能也是造成不同生态系统和土壤深度碳库对氮添加响应差异的一个重要原因. ...
... 氮添加通过抑制土壤呼吸作用与土壤稳定性碳的分解而促进土壤碳的积累, 这也是目前被广泛接受的解释(Zak et al., 2008; Reay et al., 2008; Janssens & Luyssaert, 2009; Cusack et al., 2010; Ramirez et al., 2012). ...
... 氮添加不仅对土壤微生物呼吸产生影响, 而且可以促使土壤微生物群落结构发生改变(Waldrop et al., 2004; Carreiro et al., 2000).土壤微生物对陆地生态系统土壤碳矿化过程及营养元素循环具有重要影响, 由土壤微生物活动、植物根系分泌物和动植物残体腐解过程所产生的土壤酶, 包括游离酶、胞内酶和胞外酶等的活性与土壤有机质的分解合成以及土壤供应植物养分的能力密切相关(关松荫, 1986).研究认为氮添加可改变土壤细菌与真菌组成比例(Demoling et al., 2008; Ramirez et al., 2012), 并通过影响水解酶和氧化还原酶的活性而影响稳定性碳的分解, 如促进纤维素和多糖类等有机质的分解, 抑制木质素等其他难分解有机质的分解(Carreiro et al., 2000; Hobbie et al., 2012; Ramirez et al., 2012).此外, 分子生物学的发展为微生物生态学的研究提供了新的手段.Cusack等(2010)利用磷脂脂肪酸分析和13C核磁共振技术对热带雨林的研究发现, 氮添加通过影响水解酶及氧化酶活性促进土壤中活性碳的分解.此外, 高通量测序技术、RT-PCR技术、PCR-DGGE技术、基于rDNA指纹图谱等的应用, 也将有助于深入认识氮添加下土壤有机碳变化的微生物学机制. ...
Soil carbon dynamics in regrowing forest of eastern Amazonia.
1
1999
... 第四, 土壤碳储量巨大, 但其在氮添加下的变化量相对微小.如何减小其变化量的检测误差, 是当前相关研究需重点关注的一个方面.采取连续观测法并通过严格选择采样点可减小测量误差(Ellert et al., 2002; VandenBygaart & Kay, 2004).相比于传统的样地对比法, 碳同位素(13C和14C)示踪方法可以更加敏感地揭示土壤碳的动态变化(de Camargo et al., 1999).碳的稳定性同位素(13C)示踪技术可以阐明土壤碳储量的迁移和转化, 揭示不同形成时期新老土壤有机碳对碳储量的贡献(Bernoux et al., 1998).放射性同位素(14C)可以反映土壤固定和释放碳的时间, 可用于研究不同时间尺度土壤有机碳的动力学机制(Trumbore, 2000).常用的估算土壤有机碳库更新速率的方法包括以下两个方法: 1)将14C作为土壤有机碳更新速率的指示剂, 通过检测1950年以来土壤有机碳中的14C量, 推算土壤碳动力学过程(Hsieh, 1993; Trumbore et al., 1996; Wang et al., 1999); 2)根据大气层残留核爆14C来识别土壤有机碳库的驻留时间(MRT)(Hsieh, 1993).如利用加速质谱仪检出土壤中14C含量在短期内的变化, 可以间接反映土壤碳的分解与更新速率.同时, 15N标记在氮添加实验中的应用不仅可以有效地检验氮素的循环过程, 而且可以在较短时期内(可以假定C:N不变)通过检验不同碳库中的15N含量及碳氮比, 并结合氮平衡法(N-balance method, 测定15N在土壤中的积累量并结合土壤C:N来估算土壤碳的变化量)来检测氮添加下土壤碳变化(de Vries et al., 2006; Mol et al., 2009).此外, 碳氮同位素的应用还可以有效地示踪土壤有机碳的来源和周转过程(Ehleringer et al., 2000), 揭示土壤碳氮耦合变化关系, 这也必将成为今后相关研究的重要方面. ...
Bacterial growth efficiency in natural aquatic systems.
1
1998
... 此外, 有研究认为土壤微生物的碳利用效率(CUE)与土壤碳积累有关(Six et al., 2006).微生物的碳利用效率是指用于微生物生物量积累的碳与微生物所吸收的碳之比(del Giorgio & Cole, 1998).CUE越高表明微生物用于自身有效生长的碳越多, 反之则表明有更多的碳用于微生物呼吸和分泌物的产生, 从而导致碳损耗, 不利于土壤碳积累.土壤微生物的碳利用效率受多种因素影响, 如高温和水分胁迫可通过影响微生物活动和呼吸作用而降低CUE (Conant et al., 2011; Manzoni et al., 2012).氮添加则可通过影响微生物的合成代谢与分解代谢影响CUE.研究认为, 氮添加可提高微生物CUE, 减弱微生物呼吸作用(Blagodatskaya et al., 2014), 从而促进土壤碳积累(Thiet et al., 2006; Manzoni et al., 2012), 也可通过影响土壤胞外酶活性降低稳定性碳分解(Ågren et al., 2001). ...
The impact of nitrogen deposition on carbon sequestration in European forests and forest soils.
3
2006
... 相对于植被碳汇, 目前关于土壤碳汇对氮添加的响应争议较大.Mack等(2004)研究发现氮添加改变了苔原地区植物群落组成并促进了凋落物分解, 从而使土壤有机碳储量减少, 抵消了氮添加下的植被碳汇增量, 导致生态系统碳储量减少.同样, Cleveland和Townsend (2006)研究发现, 氮添加促进土壤呼吸而减小土壤有机碳库储量.Nadelhoffer等(1999)则认为氮添加对欧洲森林生态系统碳库影响较小.此外, 较多研究显示氮添加增加了土壤碳储量(de Vries et al., 2006; Reay et al., 2008).例如, Zak等(2008)指出氮添加(30 kg N∙hm-2∙a-1)处理的森林土壤有机碳含量显著高于对照土壤, 其增幅约12%.氮添加同样可促进北欧云杉林和松树林土壤碳吸收, 其碳汇增量为11 g C (Hyvönen et al., 2008).Reay等(2008)对以往的研究进行了综述, 认为氮添加下的土壤碳库增量为7-23 g C.Meta分析的结果表明, 森林土壤在氮添加下的碳汇增量为19 g C (Janssens et al., 2010); 而Lu等(2011)的结果显示森林和草地的土壤碳库变化很小.经过文献梳理和总结, 氮添加下土壤碳汇增量范围为0-30 g C, 低于植被碳汇增量(de Vries et al., 2006, 2008; Reay et al., 2008; Janssens et al., 2010).整体而言, 目前关于氮添加条件下土壤碳汇的变化方向与强度争议较大, 亟需对其影响因子及机理进行深入研究分析. ...
... 的结果显示森林和草地的土壤碳库变化很小.经过文献梳理和总结, 氮添加下土壤碳汇增量范围为0-30 g C, 低于植被碳汇增量(de Vries et al., 2006, 2008; Reay et al., 2008; Janssens et al., 2010).整体而言, 目前关于氮添加条件下土壤碳汇的变化方向与强度争议较大, 亟需对其影响因子及机理进行深入研究分析. ...
... 第四, 土壤碳储量巨大, 但其在氮添加下的变化量相对微小.如何减小其变化量的检测误差, 是当前相关研究需重点关注的一个方面.采取连续观测法并通过严格选择采样点可减小测量误差(Ellert et al., 2002; VandenBygaart & Kay, 2004).相比于传统的样地对比法, 碳同位素(13C和14C)示踪方法可以更加敏感地揭示土壤碳的动态变化(de Camargo et al., 1999).碳的稳定性同位素(13C)示踪技术可以阐明土壤碳储量的迁移和转化, 揭示不同形成时期新老土壤有机碳对碳储量的贡献(Bernoux et al., 1998).放射性同位素(14C)可以反映土壤固定和释放碳的时间, 可用于研究不同时间尺度土壤有机碳的动力学机制(Trumbore, 2000).常用的估算土壤有机碳库更新速率的方法包括以下两个方法: 1)将14C作为土壤有机碳更新速率的指示剂, 通过检测1950年以来土壤有机碳中的14C量, 推算土壤碳动力学过程(Hsieh, 1993; Trumbore et al., 1996; Wang et al., 1999); 2)根据大气层残留核爆14C来识别土壤有机碳库的驻留时间(MRT)(Hsieh, 1993).如利用加速质谱仪检出土壤中14C含量在短期内的变化, 可以间接反映土壤碳的分解与更新速率.同时, 15N标记在氮添加实验中的应用不仅可以有效地检验氮素的循环过程, 而且可以在较短时期内(可以假定C:N不变)通过检验不同碳库中的15N含量及碳氮比, 并结合氮平衡法(N-balance method, 测定15N在土壤中的积累量并结合土壤C:N来估算土壤碳的变化量)来检测氮添加下土壤碳变化(de Vries et al., 2006; Mol et al., 2009).此外, 碳氮同位素的应用还可以有效地示踪土壤有机碳的来源和周转过程(Ehleringer et al., 2000), 揭示土壤碳氮耦合变化关系, 这也必将成为今后相关研究的重要方面. ...
Ecologically implausible carbon response?
3
2008
... 生态系统碳循环与氮素存在耦合关系, 碳积累与氮素供给密切相关(Luo et al., 2006; Kirkby et al., 2013).碳氮耦合作用在植被的光合生产、生长与分配过程以及土壤养分的循环过程中均发挥着重要作用.目前普遍认为氮添加可促进植被净初级生产力增加及植被生长, 提高植被碳储量(Magnani et al., 2007; Reay et al., 2008; Xia & Wan, 2008; LeBauer & Treseder, 2008; Janssens & Luyssaert, 2009; Thomas et al., 2010).不同植被类型及气候条件下的植被碳汇增量(此处将每克氮素输入所增加的碳量暂称为碳汇增量)为30-200 g C (de Vries et al., 2008; Sutton et al., 2008; Reay et al., 2008; Thomas et al., 2010; Templer et al., 2012).Magnani等(2007)研究认为每增加1 g氮输入, 可促使森林生态系统增加约470 g C.de Vries等(2008)则认为Magnani等(2007)的研究高估了该值, 重新估算的碳汇增量为30-70 g C, 其中植被碳库增量为20-40 g C, 土壤碳库增量为10-30 g C. ...
... 相对于植被碳汇, 目前关于土壤碳汇对氮添加的响应争议较大.Mack等(2004)研究发现氮添加改变了苔原地区植物群落组成并促进了凋落物分解, 从而使土壤有机碳储量减少, 抵消了氮添加下的植被碳汇增量, 导致生态系统碳储量减少.同样, Cleveland和Townsend (2006)研究发现, 氮添加促进土壤呼吸而减小土壤有机碳库储量.Nadelhoffer等(1999)则认为氮添加对欧洲森林生态系统碳库影响较小.此外, 较多研究显示氮添加增加了土壤碳储量(de Vries et al., 2006; Reay et al., 2008).例如, Zak等(2008)指出氮添加(30 kg N∙hm-2∙a-1)处理的森林土壤有机碳含量显著高于对照土壤, 其增幅约12%.氮添加同样可促进北欧云杉林和松树林土壤碳吸收, 其碳汇增量为11 g C (Hyvönen et al., 2008).Reay等(2008)对以往的研究进行了综述, 认为氮添加下的土壤碳库增量为7-23 g C.Meta分析的结果表明, 森林土壤在氮添加下的碳汇增量为19 g C (Janssens et al., 2010); 而Lu等(2011)的结果显示森林和草地的土壤碳库变化很小.经过文献梳理和总结, 氮添加下土壤碳汇增量范围为0-30 g C, 低于植被碳汇增量(de Vries et al., 2006, 2008; Reay et al., 2008; Janssens et al., 2010).整体而言, 目前关于氮添加条件下土壤碳汇的变化方向与强度争议较大, 亟需对其影响因子及机理进行深入研究分析. ...
... 第一, 与生态系统类型及其性质、土壤类型及氮素添加形式、时间长短及添加水平有关(Hyvönen et al., 2008; de Vries et al., 2008; Cusack et al., 2010).氮添加能够显著地促进农田生态系统土壤有机碳积累, 而对林地、草地、湿地及荒漠等非农业生态系统土壤碳积累的作用不明显(Lu et al., 2011).同样, 相比于温带针叶林(temperate conifer forest)和温带混合林(temperate mixed forest)及热带雨林(tropical rain forest), 氮添加对北方泰加林(boreal forest)矿质层土壤有机碳影响更为显著(Liu & Greaver, 2010).不同生态系统土壤有机碳对氮添加的响应差异可能与其本身的性质有关, 如土壤氮含量及碳氮元素计量关系、凋落物质量等(Waldrop et al., 2004; Kirkby et al., 2013).Waldrop等(2004)发现: 在3年的氮添加(80 kg N∙hm-2∙a-1)实验中, 凋落物质量较高(C:N较低)的Acer saccharum林0-20 cm土壤碳下降了20%, 而凋落物质量较低(C:N较高)的Quercus velutina林在同一深度的土壤碳则显著提高了10%.此外, 不同氮添加形式对土壤有机碳具有不同影响, 氨态氮对土壤有机碳累积的促进作用明显高于硝态氮(Lu et al., 2011), 其原因可能是氨态氮促进土壤pH值和微生物酶活性的降低, 不利于土壤有机碳的矿化(Min et al., 2011).微生物群落与土壤有机碳对氮添加的响应存在临界阈值, 长期施氮会使受氮限制的生态系统发生氮饱和, NO3-大量淋溶, 土壤酸化、大量土壤阳离子流失并产生铝毒(Tietema, 1998), 对微生物活性产生不利影响(Lovell & Hatch, 1997; Wallenstein et al., 2006); 而短期施氮可促进土壤碳积累.但也有研究表明: 短期氮添加可能会增加或降低土壤微生物生物量, 或者对其无影响(Dijkstra et al., 2005; Liu et al., 2007), 从而对土壤碳库产生不同影响.适量施氮可促进土壤碳积累, 但过量施氮可能会使有机质分解作用强于其累积, 从而降低土壤碳储量(Soussana et al., 2004).例如, 低氮添加(10 kg N∙hm-2∙a-1)促进土壤碳积累, 而中氮(20 kg N∙hm-2∙a-1)和高氮添加(40 kg N∙hm-2∙a-1)反而会降低土壤碳储量(Fang et al., 2014). ...
Bacterial and fungal response to nitrogen fertilization in three coniferous forest soils.
1
2008
... 氮添加不仅对土壤微生物呼吸产生影响, 而且可以促使土壤微生物群落结构发生改变(Waldrop et al., 2004; Carreiro et al., 2000).土壤微生物对陆地生态系统土壤碳矿化过程及营养元素循环具有重要影响, 由土壤微生物活动、植物根系分泌物和动植物残体腐解过程所产生的土壤酶, 包括游离酶、胞内酶和胞外酶等的活性与土壤有机质的分解合成以及土壤供应植物养分的能力密切相关(关松荫, 1986).研究认为氮添加可改变土壤细菌与真菌组成比例(Demoling et al., 2008; Ramirez et al., 2012), 并通过影响水解酶和氧化还原酶的活性而影响稳定性碳的分解, 如促进纤维素和多糖类等有机质的分解, 抑制木质素等其他难分解有机质的分解(Carreiro et al., 2000; Hobbie et al., 2012; Ramirez et al., 2012).此外, 分子生物学的发展为微生物生态学的研究提供了新的手段.Cusack等(2010)利用磷脂脂肪酸分析和13C核磁共振技术对热带雨林的研究发现, 氮添加通过影响水解酶及氧化酶活性促进土壤中活性碳的分解.此外, 高通量测序技术、RT-PCR技术、PCR-DGGE技术、基于rDNA指纹图谱等的应用, 也将有助于深入认识氮添加下土壤有机碳变化的微生物学机制. ...
Divergent effects of elevated CO2, N fertilization, and plant diversity on soil C and N dynamics in a grassland field experiment.
2
2005
... 第一, 与生态系统类型及其性质、土壤类型及氮素添加形式、时间长短及添加水平有关(Hyvönen et al., 2008; de Vries et al., 2008; Cusack et al., 2010).氮添加能够显著地促进农田生态系统土壤有机碳积累, 而对林地、草地、湿地及荒漠等非农业生态系统土壤碳积累的作用不明显(Lu et al., 2011).同样, 相比于温带针叶林(temperate conifer forest)和温带混合林(temperate mixed forest)及热带雨林(tropical rain forest), 氮添加对北方泰加林(boreal forest)矿质层土壤有机碳影响更为显著(Liu & Greaver, 2010).不同生态系统土壤有机碳对氮添加的响应差异可能与其本身的性质有关, 如土壤氮含量及碳氮元素计量关系、凋落物质量等(Waldrop et al., 2004; Kirkby et al., 2013).Waldrop等(2004)发现: 在3年的氮添加(80 kg N∙hm-2∙a-1)实验中, 凋落物质量较高(C:N较低)的Acer saccharum林0-20 cm土壤碳下降了20%, 而凋落物质量较低(C:N较高)的Quercus velutina林在同一深度的土壤碳则显著提高了10%.此外, 不同氮添加形式对土壤有机碳具有不同影响, 氨态氮对土壤有机碳累积的促进作用明显高于硝态氮(Lu et al., 2011), 其原因可能是氨态氮促进土壤pH值和微生物酶活性的降低, 不利于土壤有机碳的矿化(Min et al., 2011).微生物群落与土壤有机碳对氮添加的响应存在临界阈值, 长期施氮会使受氮限制的生态系统发生氮饱和, NO3-大量淋溶, 土壤酸化、大量土壤阳离子流失并产生铝毒(Tietema, 1998), 对微生物活性产生不利影响(Lovell & Hatch, 1997; Wallenstein et al., 2006); 而短期施氮可促进土壤碳积累.但也有研究表明: 短期氮添加可能会增加或降低土壤微生物生物量, 或者对其无影响(Dijkstra et al., 2005; Liu et al., 2007), 从而对土壤碳库产生不同影响.适量施氮可促进土壤碳积累, 但过量施氮可能会使有机质分解作用强于其累积, 从而降低土壤碳储量(Soussana et al., 2004).例如, 低氮添加(10 kg N∙hm-2∙a-1)促进土壤碳积累, 而中氮(20 kg N∙hm-2∙a-1)和高氮添加(40 kg N∙hm-2∙a-1)反而会降低土壤碳储量(Fang et al., 2014). ...
... 第三, 氮添加对土壤有机碳影响的差异还可能与不同土壤碳组分, 即易变碳与稳定性碳的差异性响应有关.Neff等(2002)发现, 氮添加(10 g N∙m-2∙a-1)促进了较易分解的轻组土壤碳的分解, 而增加了较难分解的重组土壤碳的稳定性.并且, 不同土壤碳组分对氮添加的响应时间也存在差异.例如, 易变碳可能在短期内就对氮添加做出响应, 促使土壤易变碳库减少(Dijkstra et al., 2005), 而稳定性碳对氮添加响应较为缓慢(Cusack et al., 2010).土壤易变碳与稳定性碳的分布和格局与植被和土壤类型以及土壤深度密切相关, 二者对氮添加的差异性响应可能也是造成不同生态系统和土壤深度碳库对氮添加响应差异的一个重要原因. ...
Moisture modulates rhizos- phere effects on C decomposition in two different soil types.
Carbon isotope ratios in belowground carbon cycle processes.
1
2000
... 第四, 土壤碳储量巨大, 但其在氮添加下的变化量相对微小.如何减小其变化量的检测误差, 是当前相关研究需重点关注的一个方面.采取连续观测法并通过严格选择采样点可减小测量误差(Ellert et al., 2002; VandenBygaart & Kay, 2004).相比于传统的样地对比法, 碳同位素(13C和14C)示踪方法可以更加敏感地揭示土壤碳的动态变化(de Camargo et al., 1999).碳的稳定性同位素(13C)示踪技术可以阐明土壤碳储量的迁移和转化, 揭示不同形成时期新老土壤有机碳对碳储量的贡献(Bernoux et al., 1998).放射性同位素(14C)可以反映土壤固定和释放碳的时间, 可用于研究不同时间尺度土壤有机碳的动力学机制(Trumbore, 2000).常用的估算土壤有机碳库更新速率的方法包括以下两个方法: 1)将14C作为土壤有机碳更新速率的指示剂, 通过检测1950年以来土壤有机碳中的14C量, 推算土壤碳动力学过程(Hsieh, 1993; Trumbore et al., 1996; Wang et al., 1999); 2)根据大气层残留核爆14C来识别土壤有机碳库的驻留时间(MRT)(Hsieh, 1993).如利用加速质谱仪检出土壤中14C含量在短期内的变化, 可以间接反映土壤碳的分解与更新速率.同时, 15N标记在氮添加实验中的应用不仅可以有效地检验氮素的循环过程, 而且可以在较短时期内(可以假定C:N不变)通过检验不同碳库中的15N含量及碳氮比, 并结合氮平衡法(N-balance method, 测定15N在土壤中的积累量并结合土壤C:N来估算土壤碳的变化量)来检测氮添加下土壤碳变化(de Vries et al., 2006; Mol et al., 2009).此外, 碳氮同位素的应用还可以有效地示踪土壤有机碳的来源和周转过程(Ehleringer et al., 2000), 揭示土壤碳氮耦合变化关系, 这也必将成为今后相关研究的重要方面. ...
Assessment of a method to measure temporal change in soil carbon storage.
1
2002
... 第四, 土壤碳储量巨大, 但其在氮添加下的变化量相对微小.如何减小其变化量的检测误差, 是当前相关研究需重点关注的一个方面.采取连续观测法并通过严格选择采样点可减小测量误差(Ellert et al., 2002; VandenBygaart & Kay, 2004).相比于传统的样地对比法, 碳同位素(13C和14C)示踪方法可以更加敏感地揭示土壤碳的动态变化(de Camargo et al., 1999).碳的稳定性同位素(13C)示踪技术可以阐明土壤碳储量的迁移和转化, 揭示不同形成时期新老土壤有机碳对碳储量的贡献(Bernoux et al., 1998).放射性同位素(14C)可以反映土壤固定和释放碳的时间, 可用于研究不同时间尺度土壤有机碳的动力学机制(Trumbore, 2000).常用的估算土壤有机碳库更新速率的方法包括以下两个方法: 1)将14C作为土壤有机碳更新速率的指示剂, 通过检测1950年以来土壤有机碳中的14C量, 推算土壤碳动力学过程(Hsieh, 1993; Trumbore et al., 1996; Wang et al., 1999); 2)根据大气层残留核爆14C来识别土壤有机碳库的驻留时间(MRT)(Hsieh, 1993).如利用加速质谱仪检出土壤中14C含量在短期内的变化, 可以间接反映土壤碳的分解与更新速率.同时, 15N标记在氮添加实验中的应用不仅可以有效地检验氮素的循环过程, 而且可以在较短时期内(可以假定C:N不变)通过检验不同碳库中的15N含量及碳氮比, 并结合氮平衡法(N-balance method, 测定15N在土壤中的积累量并结合土壤C:N来估算土壤碳的变化量)来检测氮添加下土壤碳变化(de Vries et al., 2006; Mol et al., 2009).此外, 碳氮同位素的应用还可以有效地示踪土壤有机碳的来源和周转过程(Ehleringer et al., 2000), 揭示土壤碳氮耦合变化关系, 这也必将成为今后相关研究的重要方面. ...
How a century of ammonia synthesis changed the world.
1
2008
... 氮素是生物必需的大量元素, 对调控生态系统结构和功能, 维持生态系统的健康和稳定具有重要意义.自工业革命以来, 由于人类活动, 如化肥、化石燃料使用等, 全球范围内的氮生产及大气活性氮沉降均大幅增加(Galloway et al., 2008; Cui et al., 2013).据测算, 全球表面平均大气氮沉降从工业革命前的0.5 kg N·hm-2·a-1增加到当前的10 kg N·hm-2·a-1 (Erisman et al., 2008), 而我国当前的大气氮沉降远高于全球平均水平, 约为21.1 kg N·hm-2·a-1 (Liu et al., 2013), 并且仍将持续增加(Galloway et al., 2008; Liu et al., 2013).大气氮沉降增加作为当前全球变化的一个重要方面, 伴随着大气CO2浓度增加和气温升高, 对生态系统结构、功能和服务产生了深刻影响(Mack et al., 2004; Suding et al., 2005; Magnani et al., 2007; Bowman et al., 2008; Janssens et al., 2010). ...
杉木人工林凋落物量对氮沉降增加的初期响应
1
2007
... 一些研究显示施氮可显著增加植被凋落物量(Bradley et al., 2006; Adamek et al., 2009), 且凋落物量随氮添加水平增加而增加.例如, 樊后保等(2007)研究指出: 高氮添加(240 kg N∙hm-2∙a-1)显著增加森林凋落物量(2599.5 kg∙hm-2∙a-1), 而中氮(120 kg N∙hm-2∙a-1)和低氮(60 kg N∙hm-2∙a-1)对凋落物量增加的促进作用不明显.但也有研究发现: 尽管氮添加可以促进植被生产力增加, 但地上叶片与地下根系凋落物输入对氮添加并无明显的正反馈作用(Zak et al., 2008; Janssens et al., 2010).此外, 氮添加可以增加叶片和细根氮含量(Giardina et al., 2003; Xia & Wan, 2008; Zak et al., 2008; Yuan & Chen, 2012), 可能促进叶片和细根凋落物的分解(Waldrop et al., 2004; Knorr et al., 2005; Parton et al., 2007; Hobbie et al., 2012), 而不利于土壤碳的积累.然而, 有研究显示氮添加只在初期促进凋落物分解, 主要包括水溶性物质和非木质性纤维素的分解, 随着中后期难分解的木质素含量的增多, 氮添加抑制了凋落物的分解(Carreiro et al., 2000). ...
杉木人工林凋落物量对氮沉降增加的初期响应
1
2007
... 一些研究显示施氮可显著增加植被凋落物量(Bradley et al., 2006; Adamek et al., 2009), 且凋落物量随氮添加水平增加而增加.例如, 樊后保等(2007)研究指出: 高氮添加(240 kg N∙hm-2∙a-1)显著增加森林凋落物量(2599.5 kg∙hm-2∙a-1), 而中氮(120 kg N∙hm-2∙a-1)和低氮(60 kg N∙hm-2∙a-1)对凋落物量增加的促进作用不明显.但也有研究发现: 尽管氮添加可以促进植被生产力增加, 但地上叶片与地下根系凋落物输入对氮添加并无明显的正反馈作用(Zak et al., 2008; Janssens et al., 2010).此外, 氮添加可以增加叶片和细根氮含量(Giardina et al., 2003; Xia & Wan, 2008; Zak et al., 2008; Yuan & Chen, 2012), 可能促进叶片和细根凋落物的分解(Waldrop et al., 2004; Knorr et al., 2005; Parton et al., 2007; Hobbie et al., 2012), 而不利于土壤碳的积累.然而, 有研究显示氮添加只在初期促进凋落物分解, 主要包括水溶性物质和非木质性纤维素的分解, 随着中后期难分解的木质素含量的增多, 氮添加抑制了凋落物的分解(Carreiro et al., 2000). ...
Nitrogen deposition impacts on the amount and stability of soil organic matter in an alpine meadow ecosystem depend on the form and rate of applied nitrogen.
1
2014
... 第一, 与生态系统类型及其性质、土壤类型及氮素添加形式、时间长短及添加水平有关(Hyvönen et al., 2008; de Vries et al., 2008; Cusack et al., 2010).氮添加能够显著地促进农田生态系统土壤有机碳积累, 而对林地、草地、湿地及荒漠等非农业生态系统土壤碳积累的作用不明显(Lu et al., 2011).同样, 相比于温带针叶林(temperate conifer forest)和温带混合林(temperate mixed forest)及热带雨林(tropical rain forest), 氮添加对北方泰加林(boreal forest)矿质层土壤有机碳影响更为显著(Liu & Greaver, 2010).不同生态系统土壤有机碳对氮添加的响应差异可能与其本身的性质有关, 如土壤氮含量及碳氮元素计量关系、凋落物质量等(Waldrop et al., 2004; Kirkby et al., 2013).Waldrop等(2004)发现: 在3年的氮添加(80 kg N∙hm-2∙a-1)实验中, 凋落物质量较高(C:N较低)的Acer saccharum林0-20 cm土壤碳下降了20%, 而凋落物质量较低(C:N较高)的Quercus velutina林在同一深度的土壤碳则显著提高了10%.此外, 不同氮添加形式对土壤有机碳具有不同影响, 氨态氮对土壤有机碳累积的促进作用明显高于硝态氮(Lu et al., 2011), 其原因可能是氨态氮促进土壤pH值和微生物酶活性的降低, 不利于土壤有机碳的矿化(Min et al., 2011).微生物群落与土壤有机碳对氮添加的响应存在临界阈值, 长期施氮会使受氮限制的生态系统发生氮饱和, NO3-大量淋溶, 土壤酸化、大量土壤阳离子流失并产生铝毒(Tietema, 1998), 对微生物活性产生不利影响(Lovell & Hatch, 1997; Wallenstein et al., 2006); 而短期施氮可促进土壤碳积累.但也有研究表明: 短期氮添加可能会增加或降低土壤微生物生物量, 或者对其无影响(Dijkstra et al., 2005; Liu et al., 2007), 从而对土壤碳库产生不同影响.适量施氮可促进土壤碳积累, 但过量施氮可能会使有机质分解作用强于其累积, 从而降低土壤碳储量(Soussana et al., 2004).例如, 低氮添加(10 kg N∙hm-2∙a-1)促进土壤碳积累, 而中氮(20 kg N∙hm-2∙a-1)和高氮添加(40 kg N∙hm-2∙a-1)反而会降低土壤碳储量(Fang et al., 2014). ...
Stability of organic carbon in deep soil layers controlled by fresh carbon supply.
Transformation of the nitrogen cycle: Recent trends, questions, and potential solutions.
2
2008
... 氮素是生物必需的大量元素, 对调控生态系统结构和功能, 维持生态系统的健康和稳定具有重要意义.自工业革命以来, 由于人类活动, 如化肥、化石燃料使用等, 全球范围内的氮生产及大气活性氮沉降均大幅增加(Galloway et al., 2008; Cui et al., 2013).据测算, 全球表面平均大气氮沉降从工业革命前的0.5 kg N·hm-2·a-1增加到当前的10 kg N·hm-2·a-1 (Erisman et al., 2008), 而我国当前的大气氮沉降远高于全球平均水平, 约为21.1 kg N·hm-2·a-1 (Liu et al., 2013), 并且仍将持续增加(Galloway et al., 2008; Liu et al., 2013).大气氮沉降增加作为当前全球变化的一个重要方面, 伴随着大气CO2浓度增加和气温升高, 对生态系统结构、功能和服务产生了深刻影响(Mack et al., 2004; Suding et al., 2005; Magnani et al., 2007; Bowman et al., 2008; Janssens et al., 2010). ...
... ), 并且仍将持续增加(Galloway et al., 2008; Liu et al., 2013).大气氮沉降增加作为当前全球变化的一个重要方面, 伴随着大气CO2浓度增加和气温升高, 对生态系统结构、功能和服务产生了深刻影响(Mack et al., 2004; Suding et al., 2005; Magnani et al., 2007; Bowman et al., 2008; Janssens et al., 2010). ...
Primary production and carbon allocation in relation to nutrient supply in a tropical experimental forest.
1
2003
... 一些研究显示施氮可显著增加植被凋落物量(Bradley et al., 2006; Adamek et al., 2009), 且凋落物量随氮添加水平增加而增加.例如, 樊后保等(2007)研究指出: 高氮添加(240 kg N∙hm-2∙a-1)显著增加森林凋落物量(2599.5 kg∙hm-2∙a-1), 而中氮(120 kg N∙hm-2∙a-1)和低氮(60 kg N∙hm-2∙a-1)对凋落物量增加的促进作用不明显.但也有研究发现: 尽管氮添加可以促进植被生产力增加, 但地上叶片与地下根系凋落物输入对氮添加并无明显的正反馈作用(Zak et al., 2008; Janssens et al., 2010).此外, 氮添加可以增加叶片和细根氮含量(Giardina et al., 2003; Xia & Wan, 2008; Zak et al., 2008; Yuan & Chen, 2012), 可能促进叶片和细根凋落物的分解(Waldrop et al., 2004; Knorr et al., 2005; Parton et al., 2007; Hobbie et al., 2012), 而不利于土壤碳的积累.然而, 有研究显示氮添加只在初期促进凋落物分解, 主要包括水溶性物质和非木质性纤维素的分解, 随着中后期难分解的木质素含量的增多, 氮添加抑制了凋落物的分解(Carreiro et al., 2000). ...
1
1986
... 氮添加不仅对土壤微生物呼吸产生影响, 而且可以促使土壤微生物群落结构发生改变(Waldrop et al., 2004; Carreiro et al., 2000).土壤微生物对陆地生态系统土壤碳矿化过程及营养元素循环具有重要影响, 由土壤微生物活动、植物根系分泌物和动植物残体腐解过程所产生的土壤酶, 包括游离酶、胞内酶和胞外酶等的活性与土壤有机质的分解合成以及土壤供应植物养分的能力密切相关(关松荫, 1986).研究认为氮添加可改变土壤细菌与真菌组成比例(Demoling et al., 2008; Ramirez et al., 2012), 并通过影响水解酶和氧化还原酶的活性而影响稳定性碳的分解, 如促进纤维素和多糖类等有机质的分解, 抑制木质素等其他难分解有机质的分解(Carreiro et al., 2000; Hobbie et al., 2012; Ramirez et al., 2012).此外, 分子生物学的发展为微生物生态学的研究提供了新的手段.Cusack等(2010)利用磷脂脂肪酸分析和13C核磁共振技术对热带雨林的研究发现, 氮添加通过影响水解酶及氧化酶活性促进土壤中活性碳的分解.此外, 高通量测序技术、RT-PCR技术、PCR-DGGE技术、基于rDNA指纹图谱等的应用, 也将有助于深入认识氮添加下土壤有机碳变化的微生物学机制. ...
1
1986
... 氮添加不仅对土壤微生物呼吸产生影响, 而且可以促使土壤微生物群落结构发生改变(Waldrop et al., 2004; Carreiro et al., 2000).土壤微生物对陆地生态系统土壤碳矿化过程及营养元素循环具有重要影响, 由土壤微生物活动、植物根系分泌物和动植物残体腐解过程所产生的土壤酶, 包括游离酶、胞内酶和胞外酶等的活性与土壤有机质的分解合成以及土壤供应植物养分的能力密切相关(关松荫, 1986).研究认为氮添加可改变土壤细菌与真菌组成比例(Demoling et al., 2008; Ramirez et al., 2012), 并通过影响水解酶和氧化还原酶的活性而影响稳定性碳的分解, 如促进纤维素和多糖类等有机质的分解, 抑制木质素等其他难分解有机质的分解(Carreiro et al., 2000; Hobbie et al., 2012; Ramirez et al., 2012).此外, 分子生物学的发展为微生物生态学的研究提供了新的手段.Cusack等(2010)利用磷脂脂肪酸分析和13C核磁共振技术对热带雨林的研究发现, 氮添加通过影响水解酶及氧化酶活性促进土壤中活性碳的分解.此外, 高通量测序技术、RT-PCR技术、PCR-DGGE技术、基于rDNA指纹图谱等的应用, 也将有助于深入认识氮添加下土壤有机碳变化的微生物学机制. ...
Simple rules for measuring changes in ecosystem carbon in forestry-offset projects.
1
2000
... 第二, 土壤有机碳对氮添加的响应受不同环境因子与土壤深度的影响.森林土壤有机碳库在氮添加下随纬度的增高而增加, 随年平均温度的增加而降低, 而草地土壤有机碳库在不同环境因子(如温度、纬度及降水)条件下则无显著变化(Lu et al., 2011).不同深度土壤在理化性质及稳定机制等方面均存在明显差异(Salom et al., 2010), 从而影响碳库对氮添加的响应程度与响应速率.研究显示: 氮添加可以显著增加土壤表层0-5 cm有机碳储量, 但对下层土壤有机碳含量影响不明显(Morell et al., 2011).但也有研究认为下层土壤碳同样对氮素添加响应敏感(Mack et al., 2004; Rumpel & Kögel-土壤Knabner, 2011).因此, 仅考虑表层土壤而忽略下层有机碳变化, 可能会对生态系统土壤有机碳积累速度的估算产生误差.VandenBygaart和Angers (2006)认为对农田生态系统土壤固碳研究至少需要考虑整个耕作层土壤碳变化, 甚至还有学者提出需要考虑土壤母质层, 即C层以上或1 m深度土壤碳的变化(Hamburg, 2000). ...
Deep soil horizons: Contribution and importance to soil carbon pools and in assessing whole-ecosystem response to management and global change.
Response of decomposing litter and its microbial community to multiple forms of nitrogen enrichment.
2
2012
... 一些研究显示施氮可显著增加植被凋落物量(Bradley et al., 2006; Adamek et al., 2009), 且凋落物量随氮添加水平增加而增加.例如, 樊后保等(2007)研究指出: 高氮添加(240 kg N∙hm-2∙a-1)显著增加森林凋落物量(2599.5 kg∙hm-2∙a-1), 而中氮(120 kg N∙hm-2∙a-1)和低氮(60 kg N∙hm-2∙a-1)对凋落物量增加的促进作用不明显.但也有研究发现: 尽管氮添加可以促进植被生产力增加, 但地上叶片与地下根系凋落物输入对氮添加并无明显的正反馈作用(Zak et al., 2008; Janssens et al., 2010).此外, 氮添加可以增加叶片和细根氮含量(Giardina et al., 2003; Xia & Wan, 2008; Zak et al., 2008; Yuan & Chen, 2012), 可能促进叶片和细根凋落物的分解(Waldrop et al., 2004; Knorr et al., 2005; Parton et al., 2007; Hobbie et al., 2012), 而不利于土壤碳的积累.然而, 有研究显示氮添加只在初期促进凋落物分解, 主要包括水溶性物质和非木质性纤维素的分解, 随着中后期难分解的木质素含量的增多, 氮添加抑制了凋落物的分解(Carreiro et al., 2000). ...
... 氮添加不仅对土壤微生物呼吸产生影响, 而且可以促使土壤微生物群落结构发生改变(Waldrop et al., 2004; Carreiro et al., 2000).土壤微生物对陆地生态系统土壤碳矿化过程及营养元素循环具有重要影响, 由土壤微生物活动、植物根系分泌物和动植物残体腐解过程所产生的土壤酶, 包括游离酶、胞内酶和胞外酶等的活性与土壤有机质的分解合成以及土壤供应植物养分的能力密切相关(关松荫, 1986).研究认为氮添加可改变土壤细菌与真菌组成比例(Demoling et al., 2008; Ramirez et al., 2012), 并通过影响水解酶和氧化还原酶的活性而影响稳定性碳的分解, 如促进纤维素和多糖类等有机质的分解, 抑制木质素等其他难分解有机质的分解(Carreiro et al., 2000; Hobbie et al., 2012; Ramirez et al., 2012).此外, 分子生物学的发展为微生物生态学的研究提供了新的手段.Cusack等(2010)利用磷脂脂肪酸分析和13C核磁共振技术对热带雨林的研究发现, 氮添加通过影响水解酶及氧化酶活性促进土壤中活性碳的分解.此外, 高通量测序技术、RT-PCR技术、PCR-DGGE技术、基于rDNA指纹图谱等的应用, 也将有助于深入认识氮添加下土壤有机碳变化的微生物学机制. ...
Radiocarbon signatures of turnover rates in active soil organic carbon pools.
2
1993
... 第四, 土壤碳储量巨大, 但其在氮添加下的变化量相对微小.如何减小其变化量的检测误差, 是当前相关研究需重点关注的一个方面.采取连续观测法并通过严格选择采样点可减小测量误差(Ellert et al., 2002; VandenBygaart & Kay, 2004).相比于传统的样地对比法, 碳同位素(13C和14C)示踪方法可以更加敏感地揭示土壤碳的动态变化(de Camargo et al., 1999).碳的稳定性同位素(13C)示踪技术可以阐明土壤碳储量的迁移和转化, 揭示不同形成时期新老土壤有机碳对碳储量的贡献(Bernoux et al., 1998).放射性同位素(14C)可以反映土壤固定和释放碳的时间, 可用于研究不同时间尺度土壤有机碳的动力学机制(Trumbore, 2000).常用的估算土壤有机碳库更新速率的方法包括以下两个方法: 1)将14C作为土壤有机碳更新速率的指示剂, 通过检测1950年以来土壤有机碳中的14C量, 推算土壤碳动力学过程(Hsieh, 1993; Trumbore et al., 1996; Wang et al., 1999); 2)根据大气层残留核爆14C来识别土壤有机碳库的驻留时间(MRT)(Hsieh, 1993).如利用加速质谱仪检出土壤中14C含量在短期内的变化, 可以间接反映土壤碳的分解与更新速率.同时, 15N标记在氮添加实验中的应用不仅可以有效地检验氮素的循环过程, 而且可以在较短时期内(可以假定C:N不变)通过检验不同碳库中的15N含量及碳氮比, 并结合氮平衡法(N-balance method, 测定15N在土壤中的积累量并结合土壤C:N来估算土壤碳的变化量)来检测氮添加下土壤碳变化(de Vries et al., 2006; Mol et al., 2009).此外, 碳氮同位素的应用还可以有效地示踪土壤有机碳的来源和周转过程(Ehleringer et al., 2000), 揭示土壤碳氮耦合变化关系, 这也必将成为今后相关研究的重要方面. ...
... )(Hsieh, 1993).如利用加速质谱仪检出土壤中14C含量在短期内的变化, 可以间接反映土壤碳的分解与更新速率.同时, 15N标记在氮添加实验中的应用不仅可以有效地检验氮素的循环过程, 而且可以在较短时期内(可以假定C:N不变)通过检验不同碳库中的15N含量及碳氮比, 并结合氮平衡法(N-balance method, 测定15N在土壤中的积累量并结合土壤C:N来估算土壤碳的变化量)来检测氮添加下土壤碳变化(de Vries et al., 2006; Mol et al., 2009).此外, 碳氮同位素的应用还可以有效地示踪土壤有机碳的来源和周转过程(Ehleringer et al., 2000), 揭示土壤碳氮耦合变化关系, 这也必将成为今后相关研究的重要方面. ...
Impact of long-term nitrogen addition on carbon stocks in trees and soils in northern Europe.
2
2008
... 相对于植被碳汇, 目前关于土壤碳汇对氮添加的响应争议较大.Mack等(2004)研究发现氮添加改变了苔原地区植物群落组成并促进了凋落物分解, 从而使土壤有机碳储量减少, 抵消了氮添加下的植被碳汇增量, 导致生态系统碳储量减少.同样, Cleveland和Townsend (2006)研究发现, 氮添加促进土壤呼吸而减小土壤有机碳库储量.Nadelhoffer等(1999)则认为氮添加对欧洲森林生态系统碳库影响较小.此外, 较多研究显示氮添加增加了土壤碳储量(de Vries et al., 2006; Reay et al., 2008).例如, Zak等(2008)指出氮添加(30 kg N∙hm-2∙a-1)处理的森林土壤有机碳含量显著高于对照土壤, 其增幅约12%.氮添加同样可促进北欧云杉林和松树林土壤碳吸收, 其碳汇增量为11 g C (Hyvönen et al., 2008).Reay等(2008)对以往的研究进行了综述, 认为氮添加下的土壤碳库增量为7-23 g C.Meta分析的结果表明, 森林土壤在氮添加下的碳汇增量为19 g C (Janssens et al., 2010); 而Lu等(2011)的结果显示森林和草地的土壤碳库变化很小.经过文献梳理和总结, 氮添加下土壤碳汇增量范围为0-30 g C, 低于植被碳汇增量(de Vries et al., 2006, 2008; Reay et al., 2008; Janssens et al., 2010).整体而言, 目前关于氮添加条件下土壤碳汇的变化方向与强度争议较大, 亟需对其影响因子及机理进行深入研究分析. ...
... 第一, 与生态系统类型及其性质、土壤类型及氮素添加形式、时间长短及添加水平有关(Hyvönen et al., 2008; de Vries et al., 2008; Cusack et al., 2010).氮添加能够显著地促进农田生态系统土壤有机碳积累, 而对林地、草地、湿地及荒漠等非农业生态系统土壤碳积累的作用不明显(Lu et al., 2011).同样, 相比于温带针叶林(temperate conifer forest)和温带混合林(temperate mixed forest)及热带雨林(tropical rain forest), 氮添加对北方泰加林(boreal forest)矿质层土壤有机碳影响更为显著(Liu & Greaver, 2010).不同生态系统土壤有机碳对氮添加的响应差异可能与其本身的性质有关, 如土壤氮含量及碳氮元素计量关系、凋落物质量等(Waldrop et al., 2004; Kirkby et al., 2013).Waldrop等(2004)发现: 在3年的氮添加(80 kg N∙hm-2∙a-1)实验中, 凋落物质量较高(C:N较低)的Acer saccharum林0-20 cm土壤碳下降了20%, 而凋落物质量较低(C:N较高)的Quercus velutina林在同一深度的土壤碳则显著提高了10%.此外, 不同氮添加形式对土壤有机碳具有不同影响, 氨态氮对土壤有机碳累积的促进作用明显高于硝态氮(Lu et al., 2011), 其原因可能是氨态氮促进土壤pH值和微生物酶活性的降低, 不利于土壤有机碳的矿化(Min et al., 2011).微生物群落与土壤有机碳对氮添加的响应存在临界阈值, 长期施氮会使受氮限制的生态系统发生氮饱和, NO3-大量淋溶, 土壤酸化、大量土壤阳离子流失并产生铝毒(Tietema, 1998), 对微生物活性产生不利影响(Lovell & Hatch, 1997; Wallenstein et al., 2006); 而短期施氮可促进土壤碳积累.但也有研究表明: 短期氮添加可能会增加或降低土壤微生物生物量, 或者对其无影响(Dijkstra et al., 2005; Liu et al., 2007), 从而对土壤碳库产生不同影响.适量施氮可促进土壤碳积累, 但过量施氮可能会使有机质分解作用强于其累积, 从而降低土壤碳储量(Soussana et al., 2004).例如, 低氮添加(10 kg N∙hm-2∙a-1)促进土壤碳积累, 而中氮(20 kg N∙hm-2∙a-1)和高氮添加(40 kg N∙hm-2∙a-1)反而会降低土壤碳储量(Fang et al., 2014). ...
Reduction of forest soil respiration in response to nitrogen deposition.
7
2010
... 氮素是生物必需的大量元素, 对调控生态系统结构和功能, 维持生态系统的健康和稳定具有重要意义.自工业革命以来, 由于人类活动, 如化肥、化石燃料使用等, 全球范围内的氮生产及大气活性氮沉降均大幅增加(Galloway et al., 2008; Cui et al., 2013).据测算, 全球表面平均大气氮沉降从工业革命前的0.5 kg N·hm-2·a-1增加到当前的10 kg N·hm-2·a-1 (Erisman et al., 2008), 而我国当前的大气氮沉降远高于全球平均水平, 约为21.1 kg N·hm-2·a-1 (Liu et al., 2013), 并且仍将持续增加(Galloway et al., 2008; Liu et al., 2013).大气氮沉降增加作为当前全球变化的一个重要方面, 伴随着大气CO2浓度增加和气温升高, 对生态系统结构、功能和服务产生了深刻影响(Mack et al., 2004; Suding et al., 2005; Magnani et al., 2007; Bowman et al., 2008; Janssens et al., 2010). ...
... 相对于植被碳汇, 目前关于土壤碳汇对氮添加的响应争议较大.Mack等(2004)研究发现氮添加改变了苔原地区植物群落组成并促进了凋落物分解, 从而使土壤有机碳储量减少, 抵消了氮添加下的植被碳汇增量, 导致生态系统碳储量减少.同样, Cleveland和Townsend (2006)研究发现, 氮添加促进土壤呼吸而减小土壤有机碳库储量.Nadelhoffer等(1999)则认为氮添加对欧洲森林生态系统碳库影响较小.此外, 较多研究显示氮添加增加了土壤碳储量(de Vries et al., 2006; Reay et al., 2008).例如, Zak等(2008)指出氮添加(30 kg N∙hm-2∙a-1)处理的森林土壤有机碳含量显著高于对照土壤, 其增幅约12%.氮添加同样可促进北欧云杉林和松树林土壤碳吸收, 其碳汇增量为11 g C (Hyvönen et al., 2008).Reay等(2008)对以往的研究进行了综述, 认为氮添加下的土壤碳库增量为7-23 g C.Meta分析的结果表明, 森林土壤在氮添加下的碳汇增量为19 g C (Janssens et al., 2010); 而Lu等(2011)的结果显示森林和草地的土壤碳库变化很小.经过文献梳理和总结, 氮添加下土壤碳汇增量范围为0-30 g C, 低于植被碳汇增量(de Vries et al., 2006, 2008; Reay et al., 2008; Janssens et al., 2010).整体而言, 目前关于氮添加条件下土壤碳汇的变化方向与强度争议较大, 亟需对其影响因子及机理进行深入研究分析. ...
... ; Janssens et al., 2010).整体而言, 目前关于氮添加条件下土壤碳汇的变化方向与强度争议较大, 亟需对其影响因子及机理进行深入研究分析. ...
... 一些研究显示施氮可显著增加植被凋落物量(Bradley et al., 2006; Adamek et al., 2009), 且凋落物量随氮添加水平增加而增加.例如, 樊后保等(2007)研究指出: 高氮添加(240 kg N∙hm-2∙a-1)显著增加森林凋落物量(2599.5 kg∙hm-2∙a-1), 而中氮(120 kg N∙hm-2∙a-1)和低氮(60 kg N∙hm-2∙a-1)对凋落物量增加的促进作用不明显.但也有研究发现: 尽管氮添加可以促进植被生产力增加, 但地上叶片与地下根系凋落物输入对氮添加并无明显的正反馈作用(Zak et al., 2008; Janssens et al., 2010).此外, 氮添加可以增加叶片和细根氮含量(Giardina et al., 2003; Xia & Wan, 2008; Zak et al., 2008; Yuan & Chen, 2012), 可能促进叶片和细根凋落物的分解(Waldrop et al., 2004; Knorr et al., 2005; Parton et al., 2007; Hobbie et al., 2012), 而不利于土壤碳的积累.然而, 有研究显示氮添加只在初期促进凋落物分解, 主要包括水溶性物质和非木质性纤维素的分解, 随着中后期难分解的木质素含量的增多, 氮添加抑制了凋落物的分解(Carreiro et al., 2000). ...
... 氮添加可促进腐殖质的形成, 促进分解残物向稳定性碳的转变, 增加稳定性碳积累(Moran et al., 2005; Whittinghill et al., 2012).氮元素与较难分解的凋落物残体(如木质素等)结合形成更难分解的杂环类物质(如吲哚等)和酚类是氮添加促进难分解碳形成的一个机制(Berg, 2000; Janssens et al., 2010).而氮添加对团聚体(aggregate, 即土壤碳的物理保护)的促进作用可能不明显(Janssens et al., 2010).尽管氮添加对稳定性碳库的促进作用在短期内可能对土壤总有机碳库影响不显著, 但其长期作用明显(Reid et al., 2012).土壤稳定性碳是影响土壤有机碳库稳定与积累的重要因素, 加强对土壤稳定性碳的研究对于从机理上深入认识氮添加对土壤有机碳库的影响及其响应机制具有重要作用. ...
... ).而氮添加对团聚体(aggregate, 即土壤碳的物理保护)的促进作用可能不明显(Janssens et al., 2010).尽管氮添加对稳定性碳库的促进作用在短期内可能对土壤总有机碳库影响不显著, 但其长期作用明显(Reid et al., 2012).土壤稳定性碳是影响土壤有机碳库稳定与积累的重要因素, 加强对土壤稳定性碳的研究对于从机理上深入认识氮添加对土壤有机碳库的影响及其响应机制具有重要作用. ...
Carbon cycle: Nitrogen’s carbon bonus.
3
2009
... 生态系统碳循环与氮素存在耦合关系, 碳积累与氮素供给密切相关(Luo et al., 2006; Kirkby et al., 2013).碳氮耦合作用在植被的光合生产、生长与分配过程以及土壤养分的循环过程中均发挥着重要作用.目前普遍认为氮添加可促进植被净初级生产力增加及植被生长, 提高植被碳储量(Magnani et al., 2007; Reay et al., 2008; Xia & Wan, 2008; LeBauer & Treseder, 2008; Janssens & Luyssaert, 2009; Thomas et al., 2010).不同植被类型及气候条件下的植被碳汇增量(此处将每克氮素输入所增加的碳量暂称为碳汇增量)为30-200 g C (de Vries et al., 2008; Sutton et al., 2008; Reay et al., 2008; Thomas et al., 2010; Templer et al., 2012).Magnani等(2007)研究认为每增加1 g氮输入, 可促使森林生态系统增加约470 g C.de Vries等(2008)则认为Magnani等(2007)的研究高估了该值, 重新估算的碳汇增量为30-70 g C, 其中植被碳库增量为20-40 g C, 土壤碳库增量为10-30 g C. ...
... 氮添加通过抑制土壤呼吸作用与土壤稳定性碳的分解而促进土壤碳的积累, 这也是目前被广泛接受的解释(Zak et al., 2008; Reay et al., 2008; Janssens & Luyssaert, 2009; Cusack et al., 2010; Ramirez et al., 2012). ...
Carbon-nutrient stoichiometry to increase soil carbon sequestration.
2
2013
... 生态系统碳循环与氮素存在耦合关系, 碳积累与氮素供给密切相关(Luo et al., 2006; Kirkby et al., 2013).碳氮耦合作用在植被的光合生产、生长与分配过程以及土壤养分的循环过程中均发挥着重要作用.目前普遍认为氮添加可促进植被净初级生产力增加及植被生长, 提高植被碳储量(Magnani et al., 2007; Reay et al., 2008; Xia & Wan, 2008; LeBauer & Treseder, 2008; Janssens & Luyssaert, 2009; Thomas et al., 2010).不同植被类型及气候条件下的植被碳汇增量(此处将每克氮素输入所增加的碳量暂称为碳汇增量)为30-200 g C (de Vries et al., 2008; Sutton et al., 2008; Reay et al., 2008; Thomas et al., 2010; Templer et al., 2012).Magnani等(2007)研究认为每增加1 g氮输入, 可促使森林生态系统增加约470 g C.de Vries等(2008)则认为Magnani等(2007)的研究高估了该值, 重新估算的碳汇增量为30-70 g C, 其中植被碳库增量为20-40 g C, 土壤碳库增量为10-30 g C. ...
... 第一, 与生态系统类型及其性质、土壤类型及氮素添加形式、时间长短及添加水平有关(Hyvönen et al., 2008; de Vries et al., 2008; Cusack et al., 2010).氮添加能够显著地促进农田生态系统土壤有机碳积累, 而对林地、草地、湿地及荒漠等非农业生态系统土壤碳积累的作用不明显(Lu et al., 2011).同样, 相比于温带针叶林(temperate conifer forest)和温带混合林(temperate mixed forest)及热带雨林(tropical rain forest), 氮添加对北方泰加林(boreal forest)矿质层土壤有机碳影响更为显著(Liu & Greaver, 2010).不同生态系统土壤有机碳对氮添加的响应差异可能与其本身的性质有关, 如土壤氮含量及碳氮元素计量关系、凋落物质量等(Waldrop et al., 2004; Kirkby et al., 2013).Waldrop等(2004)发现: 在3年的氮添加(80 kg N∙hm-2∙a-1)实验中, 凋落物质量较高(C:N较低)的Acer saccharum林0-20 cm土壤碳下降了20%, 而凋落物质量较低(C:N较高)的Quercus velutina林在同一深度的土壤碳则显著提高了10%.此外, 不同氮添加形式对土壤有机碳具有不同影响, 氨态氮对土壤有机碳累积的促进作用明显高于硝态氮(Lu et al., 2011), 其原因可能是氨态氮促进土壤pH值和微生物酶活性的降低, 不利于土壤有机碳的矿化(Min et al., 2011).微生物群落与土壤有机碳对氮添加的响应存在临界阈值, 长期施氮会使受氮限制的生态系统发生氮饱和, NO3-大量淋溶, 土壤酸化、大量土壤阳离子流失并产生铝毒(Tietema, 1998), 对微生物活性产生不利影响(Lovell & Hatch, 1997; Wallenstein et al., 2006); 而短期施氮可促进土壤碳积累.但也有研究表明: 短期氮添加可能会增加或降低土壤微生物生物量, 或者对其无影响(Dijkstra et al., 2005; Liu et al., 2007), 从而对土壤碳库产生不同影响.适量施氮可促进土壤碳积累, 但过量施氮可能会使有机质分解作用强于其累积, 从而降低土壤碳储量(Soussana et al., 2004).例如, 低氮添加(10 kg N∙hm-2∙a-1)促进土壤碳积累, 而中氮(20 kg N∙hm-2∙a-1)和高氮添加(40 kg N∙hm-2∙a-1)反而会降低土壤碳储量(Fang et al., 2014). ...
Nitrogen additions and litter decomposition: A meta-analysis.
1
2005
... 一些研究显示施氮可显著增加植被凋落物量(Bradley et al., 2006; Adamek et al., 2009), 且凋落物量随氮添加水平增加而增加.例如, 樊后保等(2007)研究指出: 高氮添加(240 kg N∙hm-2∙a-1)显著增加森林凋落物量(2599.5 kg∙hm-2∙a-1), 而中氮(120 kg N∙hm-2∙a-1)和低氮(60 kg N∙hm-2∙a-1)对凋落物量增加的促进作用不明显.但也有研究发现: 尽管氮添加可以促进植被生产力增加, 但地上叶片与地下根系凋落物输入对氮添加并无明显的正反馈作用(Zak et al., 2008; Janssens et al., 2010).此外, 氮添加可以增加叶片和细根氮含量(Giardina et al., 2003; Xia & Wan, 2008; Zak et al., 2008; Yuan & Chen, 2012), 可能促进叶片和细根凋落物的分解(Waldrop et al., 2004; Knorr et al., 2005; Parton et al., 2007; Hobbie et al., 2012), 而不利于土壤碳的积累.然而, 有研究显示氮添加只在初期促进凋落物分解, 主要包括水溶性物质和非木质性纤维素的分解, 随着中后期难分解的木质素含量的增多, 氮添加抑制了凋落物的分解(Carreiro et al., 2000). ...
Review of mechanisms and quantification of priming effects.
Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed.
1
2008
... 生态系统碳循环与氮素存在耦合关系, 碳积累与氮素供给密切相关(Luo et al., 2006; Kirkby et al., 2013).碳氮耦合作用在植被的光合生产、生长与分配过程以及土壤养分的循环过程中均发挥着重要作用.目前普遍认为氮添加可促进植被净初级生产力增加及植被生长, 提高植被碳储量(Magnani et al., 2007; Reay et al., 2008; Xia & Wan, 2008; LeBauer & Treseder, 2008; Janssens & Luyssaert, 2009; Thomas et al., 2010).不同植被类型及气候条件下的植被碳汇增量(此处将每克氮素输入所增加的碳量暂称为碳汇增量)为30-200 g C (de Vries et al., 2008; Sutton et al., 2008; Reay et al., 2008; Thomas et al., 2010; Templer et al., 2012).Magnani等(2007)研究认为每增加1 g氮输入, 可促使森林生态系统增加约470 g C.de Vries等(2008)则认为Magnani等(2007)的研究高估了该值, 重新估算的碳汇增量为30-70 g C, 其中植被碳库增量为20-40 g C, 土壤碳库增量为10-30 g C. ...
A global perspective on belowground carbon dynamics under nitrogen enrichment.
1
2010
... 第一, 与生态系统类型及其性质、土壤类型及氮素添加形式、时间长短及添加水平有关(Hyvönen et al., 2008; de Vries et al., 2008; Cusack et al., 2010).氮添加能够显著地促进农田生态系统土壤有机碳积累, 而对林地、草地、湿地及荒漠等非农业生态系统土壤碳积累的作用不明显(Lu et al., 2011).同样, 相比于温带针叶林(temperate conifer forest)和温带混合林(temperate mixed forest)及热带雨林(tropical rain forest), 氮添加对北方泰加林(boreal forest)矿质层土壤有机碳影响更为显著(Liu & Greaver, 2010).不同生态系统土壤有机碳对氮添加的响应差异可能与其本身的性质有关, 如土壤氮含量及碳氮元素计量关系、凋落物质量等(Waldrop et al., 2004; Kirkby et al., 2013).Waldrop等(2004)发现: 在3年的氮添加(80 kg N∙hm-2∙a-1)实验中, 凋落物质量较高(C:N较低)的Acer saccharum林0-20 cm土壤碳下降了20%, 而凋落物质量较低(C:N较高)的Quercus velutina林在同一深度的土壤碳则显著提高了10%.此外, 不同氮添加形式对土壤有机碳具有不同影响, 氨态氮对土壤有机碳累积的促进作用明显高于硝态氮(Lu et al., 2011), 其原因可能是氨态氮促进土壤pH值和微生物酶活性的降低, 不利于土壤有机碳的矿化(Min et al., 2011).微生物群落与土壤有机碳对氮添加的响应存在临界阈值, 长期施氮会使受氮限制的生态系统发生氮饱和, NO3-大量淋溶, 土壤酸化、大量土壤阳离子流失并产生铝毒(Tietema, 1998), 对微生物活性产生不利影响(Lovell & Hatch, 1997; Wallenstein et al., 2006); 而短期施氮可促进土壤碳积累.但也有研究表明: 短期氮添加可能会增加或降低土壤微生物生物量, 或者对其无影响(Dijkstra et al., 2005; Liu et al., 2007), 从而对土壤碳库产生不同影响.适量施氮可促进土壤碳积累, 但过量施氮可能会使有机质分解作用强于其累积, 从而降低土壤碳储量(Soussana et al., 2004).例如, 低氮添加(10 kg N∙hm-2∙a-1)促进土壤碳积累, 而中氮(20 kg N∙hm-2∙a-1)和高氮添加(40 kg N∙hm-2∙a-1)反而会降低土壤碳储量(Fang et al., 2014). ...
Responses of microbial biomass and respiration of soil to topography, burning, and nitrogen fertilization in a temperate steppe.
1
2007
... 第一, 与生态系统类型及其性质、土壤类型及氮素添加形式、时间长短及添加水平有关(Hyvönen et al., 2008; de Vries et al., 2008; Cusack et al., 2010).氮添加能够显著地促进农田生态系统土壤有机碳积累, 而对林地、草地、湿地及荒漠等非农业生态系统土壤碳积累的作用不明显(Lu et al., 2011).同样, 相比于温带针叶林(temperate conifer forest)和温带混合林(temperate mixed forest)及热带雨林(tropical rain forest), 氮添加对北方泰加林(boreal forest)矿质层土壤有机碳影响更为显著(Liu & Greaver, 2010).不同生态系统土壤有机碳对氮添加的响应差异可能与其本身的性质有关, 如土壤氮含量及碳氮元素计量关系、凋落物质量等(Waldrop et al., 2004; Kirkby et al., 2013).Waldrop等(2004)发现: 在3年的氮添加(80 kg N∙hm-2∙a-1)实验中, 凋落物质量较高(C:N较低)的Acer saccharum林0-20 cm土壤碳下降了20%, 而凋落物质量较低(C:N较高)的Quercus velutina林在同一深度的土壤碳则显著提高了10%.此外, 不同氮添加形式对土壤有机碳具有不同影响, 氨态氮对土壤有机碳累积的促进作用明显高于硝态氮(Lu et al., 2011), 其原因可能是氨态氮促进土壤pH值和微生物酶活性的降低, 不利于土壤有机碳的矿化(Min et al., 2011).微生物群落与土壤有机碳对氮添加的响应存在临界阈值, 长期施氮会使受氮限制的生态系统发生氮饱和, NO3-大量淋溶, 土壤酸化、大量土壤阳离子流失并产生铝毒(Tietema, 1998), 对微生物活性产生不利影响(Lovell & Hatch, 1997; Wallenstein et al., 2006); 而短期施氮可促进土壤碳积累.但也有研究表明: 短期氮添加可能会增加或降低土壤微生物生物量, 或者对其无影响(Dijkstra et al., 2005; Liu et al., 2007), 从而对土壤碳库产生不同影响.适量施氮可促进土壤碳积累, 但过量施氮可能会使有机质分解作用强于其累积, 从而降低土壤碳储量(Soussana et al., 2004).例如, 低氮添加(10 kg N∙hm-2∙a-1)促进土壤碳积累, 而中氮(20 kg N∙hm-2∙a-1)和高氮添加(40 kg N∙hm-2∙a-1)反而会降低土壤碳储量(Fang et al., 2014). ...
Enhanced nitrogen deposition over China.
2
2013
... 氮素是生物必需的大量元素, 对调控生态系统结构和功能, 维持生态系统的健康和稳定具有重要意义.自工业革命以来, 由于人类活动, 如化肥、化石燃料使用等, 全球范围内的氮生产及大气活性氮沉降均大幅增加(Galloway et al., 2008; Cui et al., 2013).据测算, 全球表面平均大气氮沉降从工业革命前的0.5 kg N·hm-2·a-1增加到当前的10 kg N·hm-2·a-1 (Erisman et al., 2008), 而我国当前的大气氮沉降远高于全球平均水平, 约为21.1 kg N·hm-2·a-1 (Liu et al., 2013), 并且仍将持续增加(Galloway et al., 2008; Liu et al., 2013).大气氮沉降增加作为当前全球变化的一个重要方面, 伴随着大气CO2浓度增加和气温升高, 对生态系统结构、功能和服务产生了深刻影响(Mack et al., 2004; Suding et al., 2005; Magnani et al., 2007; Bowman et al., 2008; Janssens et al., 2010). ...
... ; Liu et al., 2013).大气氮沉降增加作为当前全球变化的一个重要方面, 伴随着大气CO2浓度增加和气温升高, 对生态系统结构、功能和服务产生了深刻影响(Mack et al., 2004; Suding et al., 2005; Magnani et al., 2007; Bowman et al., 2008; Janssens et al., 2010). ...
The depth distribution of soil organic carbon in relation to land use and management and the potential of carbon sequestration in subsoil horizons.
Stimulation of microbial activity following spring applications of nitrogen.
1
1997
... 第一, 与生态系统类型及其性质、土壤类型及氮素添加形式、时间长短及添加水平有关(Hyvönen et al., 2008; de Vries et al., 2008; Cusack et al., 2010).氮添加能够显著地促进农田生态系统土壤有机碳积累, 而对林地、草地、湿地及荒漠等非农业生态系统土壤碳积累的作用不明显(Lu et al., 2011).同样, 相比于温带针叶林(temperate conifer forest)和温带混合林(temperate mixed forest)及热带雨林(tropical rain forest), 氮添加对北方泰加林(boreal forest)矿质层土壤有机碳影响更为显著(Liu & Greaver, 2010).不同生态系统土壤有机碳对氮添加的响应差异可能与其本身的性质有关, 如土壤氮含量及碳氮元素计量关系、凋落物质量等(Waldrop et al., 2004; Kirkby et al., 2013).Waldrop等(2004)发现: 在3年的氮添加(80 kg N∙hm-2∙a-1)实验中, 凋落物质量较高(C:N较低)的Acer saccharum林0-20 cm土壤碳下降了20%, 而凋落物质量较低(C:N较高)的Quercus velutina林在同一深度的土壤碳则显著提高了10%.此外, 不同氮添加形式对土壤有机碳具有不同影响, 氨态氮对土壤有机碳累积的促进作用明显高于硝态氮(Lu et al., 2011), 其原因可能是氨态氮促进土壤pH值和微生物酶活性的降低, 不利于土壤有机碳的矿化(Min et al., 2011).微生物群落与土壤有机碳对氮添加的响应存在临界阈值, 长期施氮会使受氮限制的生态系统发生氮饱和, NO3-大量淋溶, 土壤酸化、大量土壤阳离子流失并产生铝毒(Tietema, 1998), 对微生物活性产生不利影响(Lovell & Hatch, 1997; Wallenstein et al., 2006); 而短期施氮可促进土壤碳积累.但也有研究表明: 短期氮添加可能会增加或降低土壤微生物生物量, 或者对其无影响(Dijkstra et al., 2005; Liu et al., 2007), 从而对土壤碳库产生不同影响.适量施氮可促进土壤碳积累, 但过量施氮可能会使有机质分解作用强于其累积, 从而降低土壤碳储量(Soussana et al., 2004).例如, 低氮添加(10 kg N∙hm-2∙a-1)促进土壤碳积累, 而中氮(20 kg N∙hm-2∙a-1)和高氮添加(40 kg N∙hm-2∙a-1)反而会降低土壤碳储量(Fang et al., 2014). ...
Minor stimulation of soil carbon storage by nitrogen addition: A meta-analysis. Agriculture,
4
2011
... 相对于植被碳汇, 目前关于土壤碳汇对氮添加的响应争议较大.Mack等(2004)研究发现氮添加改变了苔原地区植物群落组成并促进了凋落物分解, 从而使土壤有机碳储量减少, 抵消了氮添加下的植被碳汇增量, 导致生态系统碳储量减少.同样, Cleveland和Townsend (2006)研究发现, 氮添加促进土壤呼吸而减小土壤有机碳库储量.Nadelhoffer等(1999)则认为氮添加对欧洲森林生态系统碳库影响较小.此外, 较多研究显示氮添加增加了土壤碳储量(de Vries et al., 2006; Reay et al., 2008).例如, Zak等(2008)指出氮添加(30 kg N∙hm-2∙a-1)处理的森林土壤有机碳含量显著高于对照土壤, 其增幅约12%.氮添加同样可促进北欧云杉林和松树林土壤碳吸收, 其碳汇增量为11 g C (Hyvönen et al., 2008).Reay等(2008)对以往的研究进行了综述, 认为氮添加下的土壤碳库增量为7-23 g C.Meta分析的结果表明, 森林土壤在氮添加下的碳汇增量为19 g C (Janssens et al., 2010); 而Lu等(2011)的结果显示森林和草地的土壤碳库变化很小.经过文献梳理和总结, 氮添加下土壤碳汇增量范围为0-30 g C, 低于植被碳汇增量(de Vries et al., 2006, 2008; Reay et al., 2008; Janssens et al., 2010).整体而言, 目前关于氮添加条件下土壤碳汇的变化方向与强度争议较大, 亟需对其影响因子及机理进行深入研究分析. ...
... 第一, 与生态系统类型及其性质、土壤类型及氮素添加形式、时间长短及添加水平有关(Hyvönen et al., 2008; de Vries et al., 2008; Cusack et al., 2010).氮添加能够显著地促进农田生态系统土壤有机碳积累, 而对林地、草地、湿地及荒漠等非农业生态系统土壤碳积累的作用不明显(Lu et al., 2011).同样, 相比于温带针叶林(temperate conifer forest)和温带混合林(temperate mixed forest)及热带雨林(tropical rain forest), 氮添加对北方泰加林(boreal forest)矿质层土壤有机碳影响更为显著(Liu & Greaver, 2010).不同生态系统土壤有机碳对氮添加的响应差异可能与其本身的性质有关, 如土壤氮含量及碳氮元素计量关系、凋落物质量等(Waldrop et al., 2004; Kirkby et al., 2013).Waldrop等(2004)发现: 在3年的氮添加(80 kg N∙hm-2∙a-1)实验中, 凋落物质量较高(C:N较低)的Acer saccharum林0-20 cm土壤碳下降了20%, 而凋落物质量较低(C:N较高)的Quercus velutina林在同一深度的土壤碳则显著提高了10%.此外, 不同氮添加形式对土壤有机碳具有不同影响, 氨态氮对土壤有机碳累积的促进作用明显高于硝态氮(Lu et al., 2011), 其原因可能是氨态氮促进土壤pH值和微生物酶活性的降低, 不利于土壤有机碳的矿化(Min et al., 2011).微生物群落与土壤有机碳对氮添加的响应存在临界阈值, 长期施氮会使受氮限制的生态系统发生氮饱和, NO3-大量淋溶, 土壤酸化、大量土壤阳离子流失并产生铝毒(Tietema, 1998), 对微生物活性产生不利影响(Lovell & Hatch, 1997; Wallenstein et al., 2006); 而短期施氮可促进土壤碳积累.但也有研究表明: 短期氮添加可能会增加或降低土壤微生物生物量, 或者对其无影响(Dijkstra et al., 2005; Liu et al., 2007), 从而对土壤碳库产生不同影响.适量施氮可促进土壤碳积累, 但过量施氮可能会使有机质分解作用强于其累积, 从而降低土壤碳储量(Soussana et al., 2004).例如, 低氮添加(10 kg N∙hm-2∙a-1)促进土壤碳积累, 而中氮(20 kg N∙hm-2∙a-1)和高氮添加(40 kg N∙hm-2∙a-1)反而会降低土壤碳储量(Fang et al., 2014). ...
... 林在同一深度的土壤碳则显著提高了10%.此外, 不同氮添加形式对土壤有机碳具有不同影响, 氨态氮对土壤有机碳累积的促进作用明显高于硝态氮(Lu et al., 2011), 其原因可能是氨态氮促进土壤pH值和微生物酶活性的降低, 不利于土壤有机碳的矿化(Min et al., 2011).微生物群落与土壤有机碳对氮添加的响应存在临界阈值, 长期施氮会使受氮限制的生态系统发生氮饱和, NO3-大量淋溶, 土壤酸化、大量土壤阳离子流失并产生铝毒(Tietema, 1998), 对微生物活性产生不利影响(Lovell & Hatch, 1997; Wallenstein et al., 2006); 而短期施氮可促进土壤碳积累.但也有研究表明: 短期氮添加可能会增加或降低土壤微生物生物量, 或者对其无影响(Dijkstra et al., 2005; Liu et al., 2007), 从而对土壤碳库产生不同影响.适量施氮可促进土壤碳积累, 但过量施氮可能会使有机质分解作用强于其累积, 从而降低土壤碳储量(Soussana et al., 2004).例如, 低氮添加(10 kg N∙hm-2∙a-1)促进土壤碳积累, 而中氮(20 kg N∙hm-2∙a-1)和高氮添加(40 kg N∙hm-2∙a-1)反而会降低土壤碳储量(Fang et al., 2014). ...
... 第二, 土壤有机碳对氮添加的响应受不同环境因子与土壤深度的影响.森林土壤有机碳库在氮添加下随纬度的增高而增加, 随年平均温度的增加而降低, 而草地土壤有机碳库在不同环境因子(如温度、纬度及降水)条件下则无显著变化(Lu et al., 2011).不同深度土壤在理化性质及稳定机制等方面均存在明显差异(Salom et al., 2010), 从而影响碳库对氮添加的响应程度与响应速率.研究显示: 氮添加可以显著增加土壤表层0-5 cm有机碳储量, 但对下层土壤有机碳含量影响不明显(Morell et al., 2011).但也有研究认为下层土壤碳同样对氮素添加响应敏感(Mack et al., 2004; Rumpel & Kögel-土壤Knabner, 2011).因此, 仅考虑表层土壤而忽略下层有机碳变化, 可能会对生态系统土壤有机碳积累速度的估算产生误差.VandenBygaart和Angers (2006)认为对农田生态系统土壤固碳研究至少需要考虑整个耕作层土壤碳变化, 甚至还有学者提出需要考虑土壤母质层, 即C层以上或1 m深度土壤碳的变化(Hamburg, 2000). ...
Does nitrogen constrain carbon cycling, or does carbon input stimulate nitrogen cycling?
1
2006
... 生态系统碳循环与氮素存在耦合关系, 碳积累与氮素供给密切相关(Luo et al., 2006; Kirkby et al., 2013).碳氮耦合作用在植被的光合生产、生长与分配过程以及土壤养分的循环过程中均发挥着重要作用.目前普遍认为氮添加可促进植被净初级生产力增加及植被生长, 提高植被碳储量(Magnani et al., 2007; Reay et al., 2008; Xia & Wan, 2008; LeBauer & Treseder, 2008; Janssens & Luyssaert, 2009; Thomas et al., 2010).不同植被类型及气候条件下的植被碳汇增量(此处将每克氮素输入所增加的碳量暂称为碳汇增量)为30-200 g C (de Vries et al., 2008; Sutton et al., 2008; Reay et al., 2008; Thomas et al., 2010; Templer et al., 2012).Magnani等(2007)研究认为每增加1 g氮输入, 可促使森林生态系统增加约470 g C.de Vries等(2008)则认为Magnani等(2007)的研究高估了该值, 重新估算的碳汇增量为30-70 g C, 其中植被碳库增量为20-40 g C, 土壤碳库增量为10-30 g C. ...
Ecosystem carbon storage in arctic tundra reduced by long-term nutrient fertilization.
5
2004
... 氮素是生物必需的大量元素, 对调控生态系统结构和功能, 维持生态系统的健康和稳定具有重要意义.自工业革命以来, 由于人类活动, 如化肥、化石燃料使用等, 全球范围内的氮生产及大气活性氮沉降均大幅增加(Galloway et al., 2008; Cui et al., 2013).据测算, 全球表面平均大气氮沉降从工业革命前的0.5 kg N·hm-2·a-1增加到当前的10 kg N·hm-2·a-1 (Erisman et al., 2008), 而我国当前的大气氮沉降远高于全球平均水平, 约为21.1 kg N·hm-2·a-1 (Liu et al., 2013), 并且仍将持续增加(Galloway et al., 2008; Liu et al., 2013).大气氮沉降增加作为当前全球变化的一个重要方面, 伴随着大气CO2浓度增加和气温升高, 对生态系统结构、功能和服务产生了深刻影响(Mack et al., 2004; Suding et al., 2005; Magnani et al., 2007; Bowman et al., 2008; Janssens et al., 2010). ...
... 相对于植被碳汇, 目前关于土壤碳汇对氮添加的响应争议较大.Mack等(2004)研究发现氮添加改变了苔原地区植物群落组成并促进了凋落物分解, 从而使土壤有机碳储量减少, 抵消了氮添加下的植被碳汇增量, 导致生态系统碳储量减少.同样, Cleveland和Townsend (2006)研究发现, 氮添加促进土壤呼吸而减小土壤有机碳库储量.Nadelhoffer等(1999)则认为氮添加对欧洲森林生态系统碳库影响较小.此外, 较多研究显示氮添加增加了土壤碳储量(de Vries et al., 2006; Reay et al., 2008).例如, Zak等(2008)指出氮添加(30 kg N∙hm-2∙a-1)处理的森林土壤有机碳含量显著高于对照土壤, 其增幅约12%.氮添加同样可促进北欧云杉林和松树林土壤碳吸收, 其碳汇增量为11 g C (Hyvönen et al., 2008).Reay等(2008)对以往的研究进行了综述, 认为氮添加下的土壤碳库增量为7-23 g C.Meta分析的结果表明, 森林土壤在氮添加下的碳汇增量为19 g C (Janssens et al., 2010); 而Lu等(2011)的结果显示森林和草地的土壤碳库变化很小.经过文献梳理和总结, 氮添加下土壤碳汇增量范围为0-30 g C, 低于植被碳汇增量(de Vries et al., 2006, 2008; Reay et al., 2008; Janssens et al., 2010).整体而言, 目前关于氮添加条件下土壤碳汇的变化方向与强度争议较大, 亟需对其影响因子及机理进行深入研究分析. ...
... 第二, 土壤有机碳对氮添加的响应受不同环境因子与土壤深度的影响.森林土壤有机碳库在氮添加下随纬度的增高而增加, 随年平均温度的增加而降低, 而草地土壤有机碳库在不同环境因子(如温度、纬度及降水)条件下则无显著变化(Lu et al., 2011).不同深度土壤在理化性质及稳定机制等方面均存在明显差异(Salom et al., 2010), 从而影响碳库对氮添加的响应程度与响应速率.研究显示: 氮添加可以显著增加土壤表层0-5 cm有机碳储量, 但对下层土壤有机碳含量影响不明显(Morell et al., 2011).但也有研究认为下层土壤碳同样对氮素添加响应敏感(Mack et al., 2004; Rumpel & Kögel-土壤Knabner, 2011).因此, 仅考虑表层土壤而忽略下层有机碳变化, 可能会对生态系统土壤有机碳积累速度的估算产生误差.VandenBygaart和Angers (2006)认为对农田生态系统土壤固碳研究至少需要考虑整个耕作层土壤碳变化, 甚至还有学者提出需要考虑土壤母质层, 即C层以上或1 m深度土壤碳的变化(Hamburg, 2000). ...
... 氮添加对土壤有机碳库的影响依赖于碳输入与分解矿化过程的动态平衡(Mack et al., 2004; Trumbore, 2006), 包括植物生长、碳吸收及光合产物的分配、凋落物分解、土壤有机质周转和土壤呼吸等过程.当前氮添加对土壤有机碳库影响的研究也主要围绕以上相关过程展开.本文对氮添加下的土壤有机碳库、碳循环过程及其相关研究进行了梳理(图1), 将氮添加对土壤碳积累与稳定性的响应机理总结为以下3个主要方面. ...
The human footprint in the carbon cycle of temperate and boreal forests.
5
2007
... 氮素是生物必需的大量元素, 对调控生态系统结构和功能, 维持生态系统的健康和稳定具有重要意义.自工业革命以来, 由于人类活动, 如化肥、化石燃料使用等, 全球范围内的氮生产及大气活性氮沉降均大幅增加(Galloway et al., 2008; Cui et al., 2013).据测算, 全球表面平均大气氮沉降从工业革命前的0.5 kg N·hm-2·a-1增加到当前的10 kg N·hm-2·a-1 (Erisman et al., 2008), 而我国当前的大气氮沉降远高于全球平均水平, 约为21.1 kg N·hm-2·a-1 (Liu et al., 2013), 并且仍将持续增加(Galloway et al., 2008; Liu et al., 2013).大气氮沉降增加作为当前全球变化的一个重要方面, 伴随着大气CO2浓度增加和气温升高, 对生态系统结构、功能和服务产生了深刻影响(Mack et al., 2004; Suding et al., 2005; Magnani et al., 2007; Bowman et al., 2008; Janssens et al., 2010). ...
... 生态系统碳循环与氮素存在耦合关系, 碳积累与氮素供给密切相关(Luo et al., 2006; Kirkby et al., 2013).碳氮耦合作用在植被的光合生产、生长与分配过程以及土壤养分的循环过程中均发挥着重要作用.目前普遍认为氮添加可促进植被净初级生产力增加及植被生长, 提高植被碳储量(Magnani et al., 2007; Reay et al., 2008; Xia & Wan, 2008; LeBauer & Treseder, 2008; Janssens & Luyssaert, 2009; Thomas et al., 2010).不同植被类型及气候条件下的植被碳汇增量(此处将每克氮素输入所增加的碳量暂称为碳汇增量)为30-200 g C (de Vries et al., 2008; Sutton et al., 2008; Reay et al., 2008; Thomas et al., 2010; Templer et al., 2012).Magnani等(2007)研究认为每增加1 g氮输入, 可促使森林生态系统增加约470 g C.de Vries等(2008)则认为Magnani等(2007)的研究高估了该值, 重新估算的碳汇增量为30-70 g C, 其中植被碳库增量为20-40 g C, 土壤碳库增量为10-30 g C. ...
... ).Magnani等(2007)研究认为每增加1 g氮输入, 可促使森林生态系统增加约470 g C.de Vries等(2008)则认为Magnani等(2007)的研究高估了该值, 重新估算的碳汇增量为30-70 g C, 其中植被碳库增量为20-40 g C, 土壤碳库增量为10-30 g C. ...
... 研究认为每增加1 g氮输入, 可促使森林生态系统增加约470 g C.de Vries等(2008)则认为Magnani等(2007)的研究高估了该值, 重新估算的碳汇增量为30-70 g C, 其中植被碳库增量为20-40 g C, 土壤碳库增量为10-30 g C. ...
Environmental and stoichiometric controls on microbial carbon-use efficiency in soils.
2
2012
... 此外, 有研究认为土壤微生物的碳利用效率(CUE)与土壤碳积累有关(Six et al., 2006).微生物的碳利用效率是指用于微生物生物量积累的碳与微生物所吸收的碳之比(del Giorgio & Cole, 1998).CUE越高表明微生物用于自身有效生长的碳越多, 反之则表明有更多的碳用于微生物呼吸和分泌物的产生, 从而导致碳损耗, 不利于土壤碳积累.土壤微生物的碳利用效率受多种因素影响, 如高温和水分胁迫可通过影响微生物活动和呼吸作用而降低CUE (Conant et al., 2011; Manzoni et al., 2012).氮添加则可通过影响微生物的合成代谢与分解代谢影响CUE.研究认为, 氮添加可提高微生物CUE, 减弱微生物呼吸作用(Blagodatskaya et al., 2014), 从而促进土壤碳积累(Thiet et al., 2006; Manzoni et al., 2012), 也可通过影响土壤胞外酶活性降低稳定性碳分解(Ågren et al., 2001). ...
... ; Manzoni et al., 2012), 也可通过影响土壤胞外酶活性降低稳定性碳分解(Ågren et al., 2001). ...
Effects of ammonium and nitrate additions on carbon mineralization in wetland soils.
1
2011
... 第一, 与生态系统类型及其性质、土壤类型及氮素添加形式、时间长短及添加水平有关(Hyvönen et al., 2008; de Vries et al., 2008; Cusack et al., 2010).氮添加能够显著地促进农田生态系统土壤有机碳积累, 而对林地、草地、湿地及荒漠等非农业生态系统土壤碳积累的作用不明显(Lu et al., 2011).同样, 相比于温带针叶林(temperate conifer forest)和温带混合林(temperate mixed forest)及热带雨林(tropical rain forest), 氮添加对北方泰加林(boreal forest)矿质层土壤有机碳影响更为显著(Liu & Greaver, 2010).不同生态系统土壤有机碳对氮添加的响应差异可能与其本身的性质有关, 如土壤氮含量及碳氮元素计量关系、凋落物质量等(Waldrop et al., 2004; Kirkby et al., 2013).Waldrop等(2004)发现: 在3年的氮添加(80 kg N∙hm-2∙a-1)实验中, 凋落物质量较高(C:N较低)的Acer saccharum林0-20 cm土壤碳下降了20%, 而凋落物质量较低(C:N较高)的Quercus velutina林在同一深度的土壤碳则显著提高了10%.此外, 不同氮添加形式对土壤有机碳具有不同影响, 氨态氮对土壤有机碳累积的促进作用明显高于硝态氮(Lu et al., 2011), 其原因可能是氨态氮促进土壤pH值和微生物酶活性的降低, 不利于土壤有机碳的矿化(Min et al., 2011).微生物群落与土壤有机碳对氮添加的响应存在临界阈值, 长期施氮会使受氮限制的生态系统发生氮饱和, NO3-大量淋溶, 土壤酸化、大量土壤阳离子流失并产生铝毒(Tietema, 1998), 对微生物活性产生不利影响(Lovell & Hatch, 1997; Wallenstein et al., 2006); 而短期施氮可促进土壤碳积累.但也有研究表明: 短期氮添加可能会增加或降低土壤微生物生物量, 或者对其无影响(Dijkstra et al., 2005; Liu et al., 2007), 从而对土壤碳库产生不同影响.适量施氮可促进土壤碳积累, 但过量施氮可能会使有机质分解作用强于其累积, 从而降低土壤碳储量(Soussana et al., 2004).例如, 低氮添加(10 kg N∙hm-2∙a-1)促进土壤碳积累, 而中氮(20 kg N∙hm-2∙a-1)和高氮添加(40 kg N∙hm-2∙a-1)反而会降低土壤碳储量(Fang et al., 2014). ...
Modelling soil carbon sequestration of intensively monitored forest plots in Europe by three different approaches.
1
2009
... 第四, 土壤碳储量巨大, 但其在氮添加下的变化量相对微小.如何减小其变化量的检测误差, 是当前相关研究需重点关注的一个方面.采取连续观测法并通过严格选择采样点可减小测量误差(Ellert et al., 2002; VandenBygaart & Kay, 2004).相比于传统的样地对比法, 碳同位素(13C和14C)示踪方法可以更加敏感地揭示土壤碳的动态变化(de Camargo et al., 1999).碳的稳定性同位素(13C)示踪技术可以阐明土壤碳储量的迁移和转化, 揭示不同形成时期新老土壤有机碳对碳储量的贡献(Bernoux et al., 1998).放射性同位素(14C)可以反映土壤固定和释放碳的时间, 可用于研究不同时间尺度土壤有机碳的动力学机制(Trumbore, 2000).常用的估算土壤有机碳库更新速率的方法包括以下两个方法: 1)将14C作为土壤有机碳更新速率的指示剂, 通过检测1950年以来土壤有机碳中的14C量, 推算土壤碳动力学过程(Hsieh, 1993; Trumbore et al., 1996; Wang et al., 1999); 2)根据大气层残留核爆14C来识别土壤有机碳库的驻留时间(MRT)(Hsieh, 1993).如利用加速质谱仪检出土壤中14C含量在短期内的变化, 可以间接反映土壤碳的分解与更新速率.同时, 15N标记在氮添加实验中的应用不仅可以有效地检验氮素的循环过程, 而且可以在较短时期内(可以假定C:N不变)通过检验不同碳库中的15N含量及碳氮比, 并结合氮平衡法(N-balance method, 测定15N在土壤中的积累量并结合土壤C:N来估算土壤碳的变化量)来检测氮添加下土壤碳变化(de Vries et al., 2006; Mol et al., 2009).此外, 碳氮同位素的应用还可以有效地示踪土壤有机碳的来源和周转过程(Ehleringer et al., 2000), 揭示土壤碳氮耦合变化关系, 这也必将成为今后相关研究的重要方面. ...
Role of mineral-nitrogen in residue decomposition and stable soil organic matter formation.
1
2005
... 氮添加可促进腐殖质的形成, 促进分解残物向稳定性碳的转变, 增加稳定性碳积累(Moran et al., 2005; Whittinghill et al., 2012).氮元素与较难分解的凋落物残体(如木质素等)结合形成更难分解的杂环类物质(如吲哚等)和酚类是氮添加促进难分解碳形成的一个机制(Berg, 2000; Janssens et al., 2010).而氮添加对团聚体(aggregate, 即土壤碳的物理保护)的促进作用可能不明显(Janssens et al., 2010).尽管氮添加对稳定性碳库的促进作用在短期内可能对土壤总有机碳库影响不显著, 但其长期作用明显(Reid et al., 2012).土壤稳定性碳是影响土壤有机碳库稳定与积累的重要因素, 加强对土壤稳定性碳的研究对于从机理上深入认识氮添加对土壤有机碳库的影响及其响应机制具有重要作用. ...
Soil carbon dioxide flux and organic carbon content: Effects of tillage and nitrogen fertilization.
1
2011
... 第二, 土壤有机碳对氮添加的响应受不同环境因子与土壤深度的影响.森林土壤有机碳库在氮添加下随纬度的增高而增加, 随年平均温度的增加而降低, 而草地土壤有机碳库在不同环境因子(如温度、纬度及降水)条件下则无显著变化(Lu et al., 2011).不同深度土壤在理化性质及稳定机制等方面均存在明显差异(Salom et al., 2010), 从而影响碳库对氮添加的响应程度与响应速率.研究显示: 氮添加可以显著增加土壤表层0-5 cm有机碳储量, 但对下层土壤有机碳含量影响不明显(Morell et al., 2011).但也有研究认为下层土壤碳同样对氮素添加响应敏感(Mack et al., 2004; Rumpel & Kögel-土壤Knabner, 2011).因此, 仅考虑表层土壤而忽略下层有机碳变化, 可能会对生态系统土壤有机碳积累速度的估算产生误差.VandenBygaart和Angers (2006)认为对农田生态系统土壤固碳研究至少需要考虑整个耕作层土壤碳变化, 甚至还有学者提出需要考虑土壤母质层, 即C层以上或1 m深度土壤碳的变化(Hamburg, 2000). ...
Nitrogen deposition makes a minor contribution to carbon sequestration in temperate forests.
1
1999
... 相对于植被碳汇, 目前关于土壤碳汇对氮添加的响应争议较大.Mack等(2004)研究发现氮添加改变了苔原地区植物群落组成并促进了凋落物分解, 从而使土壤有机碳储量减少, 抵消了氮添加下的植被碳汇增量, 导致生态系统碳储量减少.同样, Cleveland和Townsend (2006)研究发现, 氮添加促进土壤呼吸而减小土壤有机碳库储量.Nadelhoffer等(1999)则认为氮添加对欧洲森林生态系统碳库影响较小.此外, 较多研究显示氮添加增加了土壤碳储量(de Vries et al., 2006; Reay et al., 2008).例如, Zak等(2008)指出氮添加(30 kg N∙hm-2∙a-1)处理的森林土壤有机碳含量显著高于对照土壤, 其增幅约12%.氮添加同样可促进北欧云杉林和松树林土壤碳吸收, 其碳汇增量为11 g C (Hyvönen et al., 2008).Reay等(2008)对以往的研究进行了综述, 认为氮添加下的土壤碳库增量为7-23 g C.Meta分析的结果表明, 森林土壤在氮添加下的碳汇增量为19 g C (Janssens et al., 2010); 而Lu等(2011)的结果显示森林和草地的土壤碳库变化很小.经过文献梳理和总结, 氮添加下土壤碳汇增量范围为0-30 g C, 低于植被碳汇增量(de Vries et al., 2006, 2008; Reay et al., 2008; Janssens et al., 2010).整体而言, 目前关于氮添加条件下土壤碳汇的变化方向与强度争议较大, 亟需对其影响因子及机理进行深入研究分析. ...
Variable effects of nitrogen additions on the stability and turnover of soil carbon.
1
2002
... 第三, 氮添加对土壤有机碳影响的差异还可能与不同土壤碳组分, 即易变碳与稳定性碳的差异性响应有关.Neff等(2002)发现, 氮添加(10 g N∙m-2∙a-1)促进了较易分解的轻组土壤碳的分解, 而增加了较难分解的重组土壤碳的稳定性.并且, 不同土壤碳组分对氮添加的响应时间也存在差异.例如, 易变碳可能在短期内就对氮添加做出响应, 促使土壤易变碳库减少(Dijkstra et al., 2005), 而稳定性碳对氮添加响应较为缓慢(Cusack et al., 2010).土壤易变碳与稳定性碳的分布和格局与植被和土壤类型以及土壤深度密切相关, 二者对氮添加的差异性响应可能也是造成不同生态系统和土壤深度碳库对氮添加响应差异的一个重要原因. ...
Global-scale similarities in nitrogen release patterns during long-term decomposition.
1
2007
... 一些研究显示施氮可显著增加植被凋落物量(Bradley et al., 2006; Adamek et al., 2009), 且凋落物量随氮添加水平增加而增加.例如, 樊后保等(2007)研究指出: 高氮添加(240 kg N∙hm-2∙a-1)显著增加森林凋落物量(2599.5 kg∙hm-2∙a-1), 而中氮(120 kg N∙hm-2∙a-1)和低氮(60 kg N∙hm-2∙a-1)对凋落物量增加的促进作用不明显.但也有研究发现: 尽管氮添加可以促进植被生产力增加, 但地上叶片与地下根系凋落物输入对氮添加并无明显的正反馈作用(Zak et al., 2008; Janssens et al., 2010).此外, 氮添加可以增加叶片和细根氮含量(Giardina et al., 2003; Xia & Wan, 2008; Zak et al., 2008; Yuan & Chen, 2012), 可能促进叶片和细根凋落物的分解(Waldrop et al., 2004; Knorr et al., 2005; Parton et al., 2007; Hobbie et al., 2012), 而不利于土壤碳的积累.然而, 有研究显示氮添加只在初期促进凋落物分解, 主要包括水溶性物质和非木质性纤维素的分解, 随着中后期难分解的木质素含量的增多, 氮添加抑制了凋落物的分解(Carreiro et al., 2000). ...
Fertilization effects on fineroot biomass, rhizosphere microbes and respiratory fluxes in hardwood forest soils.
... 生态系统碳循环与氮素存在耦合关系, 碳积累与氮素供给密切相关(Luo et al., 2006; Kirkby et al., 2013).碳氮耦合作用在植被的光合生产、生长与分配过程以及土壤养分的循环过程中均发挥着重要作用.目前普遍认为氮添加可促进植被净初级生产力增加及植被生长, 提高植被碳储量(Magnani et al., 2007; Reay et al., 2008; Xia & Wan, 2008; LeBauer & Treseder, 2008; Janssens & Luyssaert, 2009; Thomas et al., 2010).不同植被类型及气候条件下的植被碳汇增量(此处将每克氮素输入所增加的碳量暂称为碳汇增量)为30-200 g C (de Vries et al., 2008; Sutton et al., 2008; Reay et al., 2008; Thomas et al., 2010; Templer et al., 2012).Magnani等(2007)研究认为每增加1 g氮输入, 可促使森林生态系统增加约470 g C.de Vries等(2008)则认为Magnani等(2007)的研究高估了该值, 重新估算的碳汇增量为30-70 g C, 其中植被碳库增量为20-40 g C, 土壤碳库增量为10-30 g C. ...
... ; Reay et al., 2008; Thomas et al., 2010; Templer et al., 2012).Magnani等(2007)研究认为每增加1 g氮输入, 可促使森林生态系统增加约470 g C.de Vries等(2008)则认为Magnani等(2007)的研究高估了该值, 重新估算的碳汇增量为30-70 g C, 其中植被碳库增量为20-40 g C, 土壤碳库增量为10-30 g C. ...
... 相对于植被碳汇, 目前关于土壤碳汇对氮添加的响应争议较大.Mack等(2004)研究发现氮添加改变了苔原地区植物群落组成并促进了凋落物分解, 从而使土壤有机碳储量减少, 抵消了氮添加下的植被碳汇增量, 导致生态系统碳储量减少.同样, Cleveland和Townsend (2006)研究发现, 氮添加促进土壤呼吸而减小土壤有机碳库储量.Nadelhoffer等(1999)则认为氮添加对欧洲森林生态系统碳库影响较小.此外, 较多研究显示氮添加增加了土壤碳储量(de Vries et al., 2006; Reay et al., 2008).例如, Zak等(2008)指出氮添加(30 kg N∙hm-2∙a-1)处理的森林土壤有机碳含量显著高于对照土壤, 其增幅约12%.氮添加同样可促进北欧云杉林和松树林土壤碳吸收, 其碳汇增量为11 g C (Hyvönen et al., 2008).Reay等(2008)对以往的研究进行了综述, 认为氮添加下的土壤碳库增量为7-23 g C.Meta分析的结果表明, 森林土壤在氮添加下的碳汇增量为19 g C (Janssens et al., 2010); 而Lu等(2011)的结果显示森林和草地的土壤碳库变化很小.经过文献梳理和总结, 氮添加下土壤碳汇增量范围为0-30 g C, 低于植被碳汇增量(de Vries et al., 2006, 2008; Reay et al., 2008; Janssens et al., 2010).整体而言, 目前关于氮添加条件下土壤碳汇的变化方向与强度争议较大, 亟需对其影响因子及机理进行深入研究分析. ...
... ; Reay et al., 2008; Janssens et al., 2010).整体而言, 目前关于氮添加条件下土壤碳汇的变化方向与强度争议较大, 亟需对其影响因子及机理进行深入研究分析. ...
... 氮添加通过抑制土壤呼吸作用与土壤稳定性碳的分解而促进土壤碳的积累, 这也是目前被广泛接受的解释(Zak et al., 2008; Reay et al., 2008; Janssens & Luyssaert, 2009; Cusack et al., 2010; Ramirez et al., 2012). ...
Biodiversity, nitrogen deposition, and CO2 affect grassland soil carbon cycling but not storage.
1
2012
... 氮添加可促进腐殖质的形成, 促进分解残物向稳定性碳的转变, 增加稳定性碳积累(Moran et al., 2005; Whittinghill et al., 2012).氮元素与较难分解的凋落物残体(如木质素等)结合形成更难分解的杂环类物质(如吲哚等)和酚类是氮添加促进难分解碳形成的一个机制(Berg, 2000; Janssens et al., 2010).而氮添加对团聚体(aggregate, 即土壤碳的物理保护)的促进作用可能不明显(Janssens et al., 2010).尽管氮添加对稳定性碳库的促进作用在短期内可能对土壤总有机碳库影响不显著, 但其长期作用明显(Reid et al., 2012).土壤稳定性碳是影响土壤有机碳库稳定与积累的重要因素, 加强对土壤稳定性碳的研究对于从机理上深入认识氮添加对土壤有机碳库的影响及其响应机制具有重要作用. ...
Deep soil organic matter—A key but poorly understood component of terrestrial C cycle.
4
2011
... 第二, 土壤有机碳对氮添加的响应受不同环境因子与土壤深度的影响.森林土壤有机碳库在氮添加下随纬度的增高而增加, 随年平均温度的增加而降低, 而草地土壤有机碳库在不同环境因子(如温度、纬度及降水)条件下则无显著变化(Lu et al., 2011).不同深度土壤在理化性质及稳定机制等方面均存在明显差异(Salom et al., 2010), 从而影响碳库对氮添加的响应程度与响应速率.研究显示: 氮添加可以显著增加土壤表层0-5 cm有机碳储量, 但对下层土壤有机碳含量影响不明显(Morell et al., 2011).但也有研究认为下层土壤碳同样对氮素添加响应敏感(Mack et al., 2004; Rumpel & Kögel-土壤Knabner, 2011).因此, 仅考虑表层土壤而忽略下层有机碳变化, 可能会对生态系统土壤有机碳积累速度的估算产生误差.VandenBygaart和Angers (2006)认为对农田生态系统土壤固碳研究至少需要考虑整个耕作层土壤碳变化, 甚至还有学者提出需要考虑土壤母质层, 即C层以上或1 m深度土壤碳的变化(Hamburg, 2000). ...
Bacterial and fungal contributions to carbon sequestration in agroecosystems.
1
2006
... 此外, 有研究认为土壤微生物的碳利用效率(CUE)与土壤碳积累有关(Six et al., 2006).微生物的碳利用效率是指用于微生物生物量积累的碳与微生物所吸收的碳之比(del Giorgio & Cole, 1998).CUE越高表明微生物用于自身有效生长的碳越多, 反之则表明有更多的碳用于微生物呼吸和分泌物的产生, 从而导致碳损耗, 不利于土壤碳积累.土壤微生物的碳利用效率受多种因素影响, 如高温和水分胁迫可通过影响微生物活动和呼吸作用而降低CUE (Conant et al., 2011; Manzoni et al., 2012).氮添加则可通过影响微生物的合成代谢与分解代谢影响CUE.研究认为, 氮添加可提高微生物CUE, 减弱微生物呼吸作用(Blagodatskaya et al., 2014), 从而促进土壤碳积累(Thiet et al., 2006; Manzoni et al., 2012), 也可通过影响土壤胞外酶活性降低稳定性碳分解(Ågren et al., 2001). ...
Carbon cycling and sequestration opportunities in temperate grasslands.
1
2004
... 第一, 与生态系统类型及其性质、土壤类型及氮素添加形式、时间长短及添加水平有关(Hyvönen et al., 2008; de Vries et al., 2008; Cusack et al., 2010).氮添加能够显著地促进农田生态系统土壤有机碳积累, 而对林地、草地、湿地及荒漠等非农业生态系统土壤碳积累的作用不明显(Lu et al., 2011).同样, 相比于温带针叶林(temperate conifer forest)和温带混合林(temperate mixed forest)及热带雨林(tropical rain forest), 氮添加对北方泰加林(boreal forest)矿质层土壤有机碳影响更为显著(Liu & Greaver, 2010).不同生态系统土壤有机碳对氮添加的响应差异可能与其本身的性质有关, 如土壤氮含量及碳氮元素计量关系、凋落物质量等(Waldrop et al., 2004; Kirkby et al., 2013).Waldrop等(2004)发现: 在3年的氮添加(80 kg N∙hm-2∙a-1)实验中, 凋落物质量较高(C:N较低)的Acer saccharum林0-20 cm土壤碳下降了20%, 而凋落物质量较低(C:N较高)的Quercus velutina林在同一深度的土壤碳则显著提高了10%.此外, 不同氮添加形式对土壤有机碳具有不同影响, 氨态氮对土壤有机碳累积的促进作用明显高于硝态氮(Lu et al., 2011), 其原因可能是氨态氮促进土壤pH值和微生物酶活性的降低, 不利于土壤有机碳的矿化(Min et al., 2011).微生物群落与土壤有机碳对氮添加的响应存在临界阈值, 长期施氮会使受氮限制的生态系统发生氮饱和, NO3-大量淋溶, 土壤酸化、大量土壤阳离子流失并产生铝毒(Tietema, 1998), 对微生物活性产生不利影响(Lovell & Hatch, 1997; Wallenstein et al., 2006); 而短期施氮可促进土壤碳积累.但也有研究表明: 短期氮添加可能会增加或降低土壤微生物生物量, 或者对其无影响(Dijkstra et al., 2005; Liu et al., 2007), 从而对土壤碳库产生不同影响.适量施氮可促进土壤碳积累, 但过量施氮可能会使有机质分解作用强于其累积, 从而降低土壤碳储量(Soussana et al., 2004).例如, 低氮添加(10 kg N∙hm-2∙a-1)促进土壤碳积累, 而中氮(20 kg N∙hm-2∙a-1)和高氮添加(40 kg N∙hm-2∙a-1)反而会降低土壤碳储量(Fang et al., 2014). ...
Functional- and abundance-based mechanisms explain diversity loss due to N fertilization.
1
2005
... 氮素是生物必需的大量元素, 对调控生态系统结构和功能, 维持生态系统的健康和稳定具有重要意义.自工业革命以来, 由于人类活动, 如化肥、化石燃料使用等, 全球范围内的氮生产及大气活性氮沉降均大幅增加(Galloway et al., 2008; Cui et al., 2013).据测算, 全球表面平均大气氮沉降从工业革命前的0.5 kg N·hm-2·a-1增加到当前的10 kg N·hm-2·a-1 (Erisman et al., 2008), 而我国当前的大气氮沉降远高于全球平均水平, 约为21.1 kg N·hm-2·a-1 (Liu et al., 2013), 并且仍将持续增加(Galloway et al., 2008; Liu et al., 2013).大气氮沉降增加作为当前全球变化的一个重要方面, 伴随着大气CO2浓度增加和气温升高, 对生态系统结构、功能和服务产生了深刻影响(Mack et al., 2004; Suding et al., 2005; Magnani et al., 2007; Bowman et al., 2008; Janssens et al., 2010). ...
Uncertainties in the relationship between atmospheric nitrogen deposition and forest carbon sequestration.
1
2008
... 生态系统碳循环与氮素存在耦合关系, 碳积累与氮素供给密切相关(Luo et al., 2006; Kirkby et al., 2013).碳氮耦合作用在植被的光合生产、生长与分配过程以及土壤养分的循环过程中均发挥着重要作用.目前普遍认为氮添加可促进植被净初级生产力增加及植被生长, 提高植被碳储量(Magnani et al., 2007; Reay et al., 2008; Xia & Wan, 2008; LeBauer & Treseder, 2008; Janssens & Luyssaert, 2009; Thomas et al., 2010).不同植被类型及气候条件下的植被碳汇增量(此处将每克氮素输入所增加的碳量暂称为碳汇增量)为30-200 g C (de Vries et al., 2008; Sutton et al., 2008; Reay et al., 2008; Thomas et al., 2010; Templer et al., 2012).Magnani等(2007)研究认为每增加1 g氮输入, 可促使森林生态系统增加约470 g C.de Vries等(2008)则认为Magnani等(2007)的研究高估了该值, 重新估算的碳汇增量为30-70 g C, 其中植被碳库增量为20-40 g C, 土壤碳库增量为10-30 g C. ...
Soil organic carbon pools in the northern circumpolar permafrost region.
Sinks for nitrogen inputs in terrestrial ecosystems: A meta-analysis of 15N tracer field studies.
1
2012
... 生态系统碳循环与氮素存在耦合关系, 碳积累与氮素供给密切相关(Luo et al., 2006; Kirkby et al., 2013).碳氮耦合作用在植被的光合生产、生长与分配过程以及土壤养分的循环过程中均发挥着重要作用.目前普遍认为氮添加可促进植被净初级生产力增加及植被生长, 提高植被碳储量(Magnani et al., 2007; Reay et al., 2008; Xia & Wan, 2008; LeBauer & Treseder, 2008; Janssens & Luyssaert, 2009; Thomas et al., 2010).不同植被类型及气候条件下的植被碳汇增量(此处将每克氮素输入所增加的碳量暂称为碳汇增量)为30-200 g C (de Vries et al., 2008; Sutton et al., 2008; Reay et al., 2008; Thomas et al., 2010; Templer et al., 2012).Magnani等(2007)研究认为每增加1 g氮输入, 可促使森林生态系统增加约470 g C.de Vries等(2008)则认为Magnani等(2007)的研究高估了该值, 重新估算的碳汇增量为30-70 g C, 其中植被碳库增量为20-40 g C, 土壤碳库增量为10-30 g C. ...
Do growth yield efficiencies differ between soil microbial communities differing in fungal: Bacterial ratios? Reality check and methodological issues.
1
2006
... 此外, 有研究认为土壤微生物的碳利用效率(CUE)与土壤碳积累有关(Six et al., 2006).微生物的碳利用效率是指用于微生物生物量积累的碳与微生物所吸收的碳之比(del Giorgio & Cole, 1998).CUE越高表明微生物用于自身有效生长的碳越多, 反之则表明有更多的碳用于微生物呼吸和分泌物的产生, 从而导致碳损耗, 不利于土壤碳积累.土壤微生物的碳利用效率受多种因素影响, 如高温和水分胁迫可通过影响微生物活动和呼吸作用而降低CUE (Conant et al., 2011; Manzoni et al., 2012).氮添加则可通过影响微生物的合成代谢与分解代谢影响CUE.研究认为, 氮添加可提高微生物CUE, 减弱微生物呼吸作用(Blagodatskaya et al., 2014), 从而促进土壤碳积累(Thiet et al., 2006; Manzoni et al., 2012), 也可通过影响土壤胞外酶活性降低稳定性碳分解(Ågren et al., 2001). ...
Increased tree carbon storage in response to nitrogen deposition in the US.
2
2010
... 生态系统碳循环与氮素存在耦合关系, 碳积累与氮素供给密切相关(Luo et al., 2006; Kirkby et al., 2013).碳氮耦合作用在植被的光合生产、生长与分配过程以及土壤养分的循环过程中均发挥着重要作用.目前普遍认为氮添加可促进植被净初级生产力增加及植被生长, 提高植被碳储量(Magnani et al., 2007; Reay et al., 2008; Xia & Wan, 2008; LeBauer & Treseder, 2008; Janssens & Luyssaert, 2009; Thomas et al., 2010).不同植被类型及气候条件下的植被碳汇增量(此处将每克氮素输入所增加的碳量暂称为碳汇增量)为30-200 g C (de Vries et al., 2008; Sutton et al., 2008; Reay et al., 2008; Thomas et al., 2010; Templer et al., 2012).Magnani等(2007)研究认为每增加1 g氮输入, 可促使森林生态系统增加约470 g C.de Vries等(2008)则认为Magnani等(2007)的研究高估了该值, 重新估算的碳汇增量为30-70 g C, 其中植被碳库增量为20-40 g C, 土壤碳库增量为10-30 g C. ...
... ; Thomas et al., 2010; Templer et al., 2012).Magnani等(2007)研究认为每增加1 g氮输入, 可促使森林生态系统增加约470 g C.de Vries等(2008)则认为Magnani等(2007)的研究高估了该值, 重新估算的碳汇增量为30-70 g C, 其中植被碳库增量为20-40 g C, 土壤碳库增量为10-30 g C. ...
Microbial carbon and nitrogen dynamics in coniferous forest floor material collected along a European nitrogen deposition gradient.
1
1998
... 第一, 与生态系统类型及其性质、土壤类型及氮素添加形式、时间长短及添加水平有关(Hyvönen et al., 2008; de Vries et al., 2008; Cusack et al., 2010).氮添加能够显著地促进农田生态系统土壤有机碳积累, 而对林地、草地、湿地及荒漠等非农业生态系统土壤碳积累的作用不明显(Lu et al., 2011).同样, 相比于温带针叶林(temperate conifer forest)和温带混合林(temperate mixed forest)及热带雨林(tropical rain forest), 氮添加对北方泰加林(boreal forest)矿质层土壤有机碳影响更为显著(Liu & Greaver, 2010).不同生态系统土壤有机碳对氮添加的响应差异可能与其本身的性质有关, 如土壤氮含量及碳氮元素计量关系、凋落物质量等(Waldrop et al., 2004; Kirkby et al., 2013).Waldrop等(2004)发现: 在3年的氮添加(80 kg N∙hm-2∙a-1)实验中, 凋落物质量较高(C:N较低)的Acer saccharum林0-20 cm土壤碳下降了20%, 而凋落物质量较低(C:N较高)的Quercus velutina林在同一深度的土壤碳则显著提高了10%.此外, 不同氮添加形式对土壤有机碳具有不同影响, 氨态氮对土壤有机碳累积的促进作用明显高于硝态氮(Lu et al., 2011), 其原因可能是氨态氮促进土壤pH值和微生物酶活性的降低, 不利于土壤有机碳的矿化(Min et al., 2011).微生物群落与土壤有机碳对氮添加的响应存在临界阈值, 长期施氮会使受氮限制的生态系统发生氮饱和, NO3-大量淋溶, 土壤酸化、大量土壤阳离子流失并产生铝毒(Tietema, 1998), 对微生物活性产生不利影响(Lovell & Hatch, 1997; Wallenstein et al., 2006); 而短期施氮可促进土壤碳积累.但也有研究表明: 短期氮添加可能会增加或降低土壤微生物生物量, 或者对其无影响(Dijkstra et al., 2005; Liu et al., 2007), 从而对土壤碳库产生不同影响.适量施氮可促进土壤碳积累, 但过量施氮可能会使有机质分解作用强于其累积, 从而降低土壤碳储量(Soussana et al., 2004).例如, 低氮添加(10 kg N∙hm-2∙a-1)促进土壤碳积累, 而中氮(20 kg N∙hm-2∙a-1)和高氮添加(40 kg N∙hm-2∙a-1)反而会降低土壤碳储量(Fang et al., 2014). ...
Age of soil organic matter and soil respiration: Radiocarbon constraints on belowground C dynamics.
1
2000
... 第四, 土壤碳储量巨大, 但其在氮添加下的变化量相对微小.如何减小其变化量的检测误差, 是当前相关研究需重点关注的一个方面.采取连续观测法并通过严格选择采样点可减小测量误差(Ellert et al., 2002; VandenBygaart & Kay, 2004).相比于传统的样地对比法, 碳同位素(13C和14C)示踪方法可以更加敏感地揭示土壤碳的动态变化(de Camargo et al., 1999).碳的稳定性同位素(13C)示踪技术可以阐明土壤碳储量的迁移和转化, 揭示不同形成时期新老土壤有机碳对碳储量的贡献(Bernoux et al., 1998).放射性同位素(14C)可以反映土壤固定和释放碳的时间, 可用于研究不同时间尺度土壤有机碳的动力学机制(Trumbore, 2000).常用的估算土壤有机碳库更新速率的方法包括以下两个方法: 1)将14C作为土壤有机碳更新速率的指示剂, 通过检测1950年以来土壤有机碳中的14C量, 推算土壤碳动力学过程(Hsieh, 1993; Trumbore et al., 1996; Wang et al., 1999); 2)根据大气层残留核爆14C来识别土壤有机碳库的驻留时间(MRT)(Hsieh, 1993).如利用加速质谱仪检出土壤中14C含量在短期内的变化, 可以间接反映土壤碳的分解与更新速率.同时, 15N标记在氮添加实验中的应用不仅可以有效地检验氮素的循环过程, 而且可以在较短时期内(可以假定C:N不变)通过检验不同碳库中的15N含量及碳氮比, 并结合氮平衡法(N-balance method, 测定15N在土壤中的积累量并结合土壤C:N来估算土壤碳的变化量)来检测氮添加下土壤碳变化(de Vries et al., 2006; Mol et al., 2009).此外, 碳氮同位素的应用还可以有效地示踪土壤有机碳的来源和周转过程(Ehleringer et al., 2000), 揭示土壤碳氮耦合变化关系, 这也必将成为今后相关研究的重要方面. ...
Carbon respired by terrestrial ecosystems—Recent progress and challenges.
1
2006
... 氮添加对土壤有机碳库的影响依赖于碳输入与分解矿化过程的动态平衡(Mack et al., 2004; Trumbore, 2006), 包括植物生长、碳吸收及光合产物的分配、凋落物分解、土壤有机质周转和土壤呼吸等过程.当前氮添加对土壤有机碳库影响的研究也主要围绕以上相关过程展开.本文对氮添加下的土壤有机碳库、碳循环过程及其相关研究进行了梳理(图1), 将氮添加对土壤碳积累与稳定性的响应机理总结为以下3个主要方面. ...
Rapid exchange between soil carbon and atmospheric carbon dioxide driven by temperature change.
1
1996
... 第四, 土壤碳储量巨大, 但其在氮添加下的变化量相对微小.如何减小其变化量的检测误差, 是当前相关研究需重点关注的一个方面.采取连续观测法并通过严格选择采样点可减小测量误差(Ellert et al., 2002; VandenBygaart & Kay, 2004).相比于传统的样地对比法, 碳同位素(13C和14C)示踪方法可以更加敏感地揭示土壤碳的动态变化(de Camargo et al., 1999).碳的稳定性同位素(13C)示踪技术可以阐明土壤碳储量的迁移和转化, 揭示不同形成时期新老土壤有机碳对碳储量的贡献(Bernoux et al., 1998).放射性同位素(14C)可以反映土壤固定和释放碳的时间, 可用于研究不同时间尺度土壤有机碳的动力学机制(Trumbore, 2000).常用的估算土壤有机碳库更新速率的方法包括以下两个方法: 1)将14C作为土壤有机碳更新速率的指示剂, 通过检测1950年以来土壤有机碳中的14C量, 推算土壤碳动力学过程(Hsieh, 1993; Trumbore et al., 1996; Wang et al., 1999); 2)根据大气层残留核爆14C来识别土壤有机碳库的驻留时间(MRT)(Hsieh, 1993).如利用加速质谱仪检出土壤中14C含量在短期内的变化, 可以间接反映土壤碳的分解与更新速率.同时, 15N标记在氮添加实验中的应用不仅可以有效地检验氮素的循环过程, 而且可以在较短时期内(可以假定C:N不变)通过检验不同碳库中的15N含量及碳氮比, 并结合氮平衡法(N-balance method, 测定15N在土壤中的积累量并结合土壤C:N来估算土壤碳的变化量)来检测氮添加下土壤碳变化(de Vries et al., 2006; Mol et al., 2009).此外, 碳氮同位素的应用还可以有效地示踪土壤有机碳的来源和周转过程(Ehleringer et al., 2000), 揭示土壤碳氮耦合变化关系, 这也必将成为今后相关研究的重要方面. ...
Persistence of soil organic carbon after plowing a long-term no-till field in southern Ontario, Canada.
1
2004
... 第四, 土壤碳储量巨大, 但其在氮添加下的变化量相对微小.如何减小其变化量的检测误差, 是当前相关研究需重点关注的一个方面.采取连续观测法并通过严格选择采样点可减小测量误差(Ellert et al., 2002; VandenBygaart & Kay, 2004).相比于传统的样地对比法, 碳同位素(13C和14C)示踪方法可以更加敏感地揭示土壤碳的动态变化(de Camargo et al., 1999).碳的稳定性同位素(13C)示踪技术可以阐明土壤碳储量的迁移和转化, 揭示不同形成时期新老土壤有机碳对碳储量的贡献(Bernoux et al., 1998).放射性同位素(14C)可以反映土壤固定和释放碳的时间, 可用于研究不同时间尺度土壤有机碳的动力学机制(Trumbore, 2000).常用的估算土壤有机碳库更新速率的方法包括以下两个方法: 1)将14C作为土壤有机碳更新速率的指示剂, 通过检测1950年以来土壤有机碳中的14C量, 推算土壤碳动力学过程(Hsieh, 1993; Trumbore et al., 1996; Wang et al., 1999); 2)根据大气层残留核爆14C来识别土壤有机碳库的驻留时间(MRT)(Hsieh, 1993).如利用加速质谱仪检出土壤中14C含量在短期内的变化, 可以间接反映土壤碳的分解与更新速率.同时, 15N标记在氮添加实验中的应用不仅可以有效地检验氮素的循环过程, 而且可以在较短时期内(可以假定C:N不变)通过检验不同碳库中的15N含量及碳氮比, 并结合氮平衡法(N-balance method, 测定15N在土壤中的积累量并结合土壤C:N来估算土壤碳的变化量)来检测氮添加下土壤碳变化(de Vries et al., 2006; Mol et al., 2009).此外, 碳氮同位素的应用还可以有效地示踪土壤有机碳的来源和周转过程(Ehleringer et al., 2000), 揭示土壤碳氮耦合变化关系, 这也必将成为今后相关研究的重要方面. ...
Towards accurate measurements of soil organic carbon stock change in agroecosystems.
1
2006
... 第二, 土壤有机碳对氮添加的响应受不同环境因子与土壤深度的影响.森林土壤有机碳库在氮添加下随纬度的增高而增加, 随年平均温度的增加而降低, 而草地土壤有机碳库在不同环境因子(如温度、纬度及降水)条件下则无显著变化(Lu et al., 2011).不同深度土壤在理化性质及稳定机制等方面均存在明显差异(Salom et al., 2010), 从而影响碳库对氮添加的响应程度与响应速率.研究显示: 氮添加可以显著增加土壤表层0-5 cm有机碳储量, 但对下层土壤有机碳含量影响不明显(Morell et al., 2011).但也有研究认为下层土壤碳同样对氮素添加响应敏感(Mack et al., 2004; Rumpel & Kögel-土壤Knabner, 2011).因此, 仅考虑表层土壤而忽略下层有机碳变化, 可能会对生态系统土壤有机碳积累速度的估算产生误差.VandenBygaart和Angers (2006)认为对农田生态系统土壤固碳研究至少需要考虑整个耕作层土壤碳变化, 甚至还有学者提出需要考虑土壤母质层, 即C层以上或1 m深度土壤碳的变化(Hamburg, 2000). ...
Nitrogen deposition modifies soil carbon storage through changes in microbial enzymatic activity.
4
2004
... 第一, 与生态系统类型及其性质、土壤类型及氮素添加形式、时间长短及添加水平有关(Hyvönen et al., 2008; de Vries et al., 2008; Cusack et al., 2010).氮添加能够显著地促进农田生态系统土壤有机碳积累, 而对林地、草地、湿地及荒漠等非农业生态系统土壤碳积累的作用不明显(Lu et al., 2011).同样, 相比于温带针叶林(temperate conifer forest)和温带混合林(temperate mixed forest)及热带雨林(tropical rain forest), 氮添加对北方泰加林(boreal forest)矿质层土壤有机碳影响更为显著(Liu & Greaver, 2010).不同生态系统土壤有机碳对氮添加的响应差异可能与其本身的性质有关, 如土壤氮含量及碳氮元素计量关系、凋落物质量等(Waldrop et al., 2004; Kirkby et al., 2013).Waldrop等(2004)发现: 在3年的氮添加(80 kg N∙hm-2∙a-1)实验中, 凋落物质量较高(C:N较低)的Acer saccharum林0-20 cm土壤碳下降了20%, 而凋落物质量较低(C:N较高)的Quercus velutina林在同一深度的土壤碳则显著提高了10%.此外, 不同氮添加形式对土壤有机碳具有不同影响, 氨态氮对土壤有机碳累积的促进作用明显高于硝态氮(Lu et al., 2011), 其原因可能是氨态氮促进土壤pH值和微生物酶活性的降低, 不利于土壤有机碳的矿化(Min et al., 2011).微生物群落与土壤有机碳对氮添加的响应存在临界阈值, 长期施氮会使受氮限制的生态系统发生氮饱和, NO3-大量淋溶, 土壤酸化、大量土壤阳离子流失并产生铝毒(Tietema, 1998), 对微生物活性产生不利影响(Lovell & Hatch, 1997; Wallenstein et al., 2006); 而短期施氮可促进土壤碳积累.但也有研究表明: 短期氮添加可能会增加或降低土壤微生物生物量, 或者对其无影响(Dijkstra et al., 2005; Liu et al., 2007), 从而对土壤碳库产生不同影响.适量施氮可促进土壤碳积累, 但过量施氮可能会使有机质分解作用强于其累积, 从而降低土壤碳储量(Soussana et al., 2004).例如, 低氮添加(10 kg N∙hm-2∙a-1)促进土壤碳积累, 而中氮(20 kg N∙hm-2∙a-1)和高氮添加(40 kg N∙hm-2∙a-1)反而会降低土壤碳储量(Fang et al., 2014). ...
... ).Waldrop等(2004)发现: 在3年的氮添加(80 kg N∙hm-2∙a-1)实验中, 凋落物质量较高(C:N较低)的Acer saccharum林0-20 cm土壤碳下降了20%, 而凋落物质量较低(C:N较高)的Quercus velutina林在同一深度的土壤碳则显著提高了10%.此外, 不同氮添加形式对土壤有机碳具有不同影响, 氨态氮对土壤有机碳累积的促进作用明显高于硝态氮(Lu et al., 2011), 其原因可能是氨态氮促进土壤pH值和微生物酶活性的降低, 不利于土壤有机碳的矿化(Min et al., 2011).微生物群落与土壤有机碳对氮添加的响应存在临界阈值, 长期施氮会使受氮限制的生态系统发生氮饱和, NO3-大量淋溶, 土壤酸化、大量土壤阳离子流失并产生铝毒(Tietema, 1998), 对微生物活性产生不利影响(Lovell & Hatch, 1997; Wallenstein et al., 2006); 而短期施氮可促进土壤碳积累.但也有研究表明: 短期氮添加可能会增加或降低土壤微生物生物量, 或者对其无影响(Dijkstra et al., 2005; Liu et al., 2007), 从而对土壤碳库产生不同影响.适量施氮可促进土壤碳积累, 但过量施氮可能会使有机质分解作用强于其累积, 从而降低土壤碳储量(Soussana et al., 2004).例如, 低氮添加(10 kg N∙hm-2∙a-1)促进土壤碳积累, 而中氮(20 kg N∙hm-2∙a-1)和高氮添加(40 kg N∙hm-2∙a-1)反而会降低土壤碳储量(Fang et al., 2014). ...
... 一些研究显示施氮可显著增加植被凋落物量(Bradley et al., 2006; Adamek et al., 2009), 且凋落物量随氮添加水平增加而增加.例如, 樊后保等(2007)研究指出: 高氮添加(240 kg N∙hm-2∙a-1)显著增加森林凋落物量(2599.5 kg∙hm-2∙a-1), 而中氮(120 kg N∙hm-2∙a-1)和低氮(60 kg N∙hm-2∙a-1)对凋落物量增加的促进作用不明显.但也有研究发现: 尽管氮添加可以促进植被生产力增加, 但地上叶片与地下根系凋落物输入对氮添加并无明显的正反馈作用(Zak et al., 2008; Janssens et al., 2010).此外, 氮添加可以增加叶片和细根氮含量(Giardina et al., 2003; Xia & Wan, 2008; Zak et al., 2008; Yuan & Chen, 2012), 可能促进叶片和细根凋落物的分解(Waldrop et al., 2004; Knorr et al., 2005; Parton et al., 2007; Hobbie et al., 2012), 而不利于土壤碳的积累.然而, 有研究显示氮添加只在初期促进凋落物分解, 主要包括水溶性物质和非木质性纤维素的分解, 随着中后期难分解的木质素含量的增多, 氮添加抑制了凋落物的分解(Carreiro et al., 2000). ...
... 氮添加不仅对土壤微生物呼吸产生影响, 而且可以促使土壤微生物群落结构发生改变(Waldrop et al., 2004; Carreiro et al., 2000).土壤微生物对陆地生态系统土壤碳矿化过程及营养元素循环具有重要影响, 由土壤微生物活动、植物根系分泌物和动植物残体腐解过程所产生的土壤酶, 包括游离酶、胞内酶和胞外酶等的活性与土壤有机质的分解合成以及土壤供应植物养分的能力密切相关(关松荫, 1986).研究认为氮添加可改变土壤细菌与真菌组成比例(Demoling et al., 2008; Ramirez et al., 2012), 并通过影响水解酶和氧化还原酶的活性而影响稳定性碳的分解, 如促进纤维素和多糖类等有机质的分解, 抑制木质素等其他难分解有机质的分解(Carreiro et al., 2000; Hobbie et al., 2012; Ramirez et al., 2012).此外, 分子生物学的发展为微生物生态学的研究提供了新的手段.Cusack等(2010)利用磷脂脂肪酸分析和13C核磁共振技术对热带雨林的研究发现, 氮添加通过影响水解酶及氧化酶活性促进土壤中活性碳的分解.此外, 高通量测序技术、RT-PCR技术、PCR-DGGE技术、基于rDNA指纹图谱等的应用, 也将有助于深入认识氮添加下土壤有机碳变化的微生物学机制. ...
Nitrogen fertilization decreases forest soil fungal and bacterial biomass in three long-term experiments.
1
2006
... 第一, 与生态系统类型及其性质、土壤类型及氮素添加形式、时间长短及添加水平有关(Hyvönen et al., 2008; de Vries et al., 2008; Cusack et al., 2010).氮添加能够显著地促进农田生态系统土壤有机碳积累, 而对林地、草地、湿地及荒漠等非农业生态系统土壤碳积累的作用不明显(Lu et al., 2011).同样, 相比于温带针叶林(temperate conifer forest)和温带混合林(temperate mixed forest)及热带雨林(tropical rain forest), 氮添加对北方泰加林(boreal forest)矿质层土壤有机碳影响更为显著(Liu & Greaver, 2010).不同生态系统土壤有机碳对氮添加的响应差异可能与其本身的性质有关, 如土壤氮含量及碳氮元素计量关系、凋落物质量等(Waldrop et al., 2004; Kirkby et al., 2013).Waldrop等(2004)发现: 在3年的氮添加(80 kg N∙hm-2∙a-1)实验中, 凋落物质量较高(C:N较低)的Acer saccharum林0-20 cm土壤碳下降了20%, 而凋落物质量较低(C:N较高)的Quercus velutina林在同一深度的土壤碳则显著提高了10%.此外, 不同氮添加形式对土壤有机碳具有不同影响, 氨态氮对土壤有机碳累积的促进作用明显高于硝态氮(Lu et al., 2011), 其原因可能是氨态氮促进土壤pH值和微生物酶活性的降低, 不利于土壤有机碳的矿化(Min et al., 2011).微生物群落与土壤有机碳对氮添加的响应存在临界阈值, 长期施氮会使受氮限制的生态系统发生氮饱和, NO3-大量淋溶, 土壤酸化、大量土壤阳离子流失并产生铝毒(Tietema, 1998), 对微生物活性产生不利影响(Lovell & Hatch, 1997; Wallenstein et al., 2006); 而短期施氮可促进土壤碳积累.但也有研究表明: 短期氮添加可能会增加或降低土壤微生物生物量, 或者对其无影响(Dijkstra et al., 2005; Liu et al., 2007), 从而对土壤碳库产生不同影响.适量施氮可促进土壤碳积累, 但过量施氮可能会使有机质分解作用强于其累积, 从而降低土壤碳储量(Soussana et al., 2004).例如, 低氮添加(10 kg N∙hm-2∙a-1)促进土壤碳积累, 而中氮(20 kg N∙hm-2∙a-1)和高氮添加(40 kg N∙hm-2∙a-1)反而会降低土壤碳储量(Fang et al., 2014). ...
The impact of land use change on C turnover in soils.
1
1999
... 第四, 土壤碳储量巨大, 但其在氮添加下的变化量相对微小.如何减小其变化量的检测误差, 是当前相关研究需重点关注的一个方面.采取连续观测法并通过严格选择采样点可减小测量误差(Ellert et al., 2002; VandenBygaart & Kay, 2004).相比于传统的样地对比法, 碳同位素(13C和14C)示踪方法可以更加敏感地揭示土壤碳的动态变化(de Camargo et al., 1999).碳的稳定性同位素(13C)示踪技术可以阐明土壤碳储量的迁移和转化, 揭示不同形成时期新老土壤有机碳对碳储量的贡献(Bernoux et al., 1998).放射性同位素(14C)可以反映土壤固定和释放碳的时间, 可用于研究不同时间尺度土壤有机碳的动力学机制(Trumbore, 2000).常用的估算土壤有机碳库更新速率的方法包括以下两个方法: 1)将14C作为土壤有机碳更新速率的指示剂, 通过检测1950年以来土壤有机碳中的14C量, 推算土壤碳动力学过程(Hsieh, 1993; Trumbore et al., 1996; Wang et al., 1999); 2)根据大气层残留核爆14C来识别土壤有机碳库的驻留时间(MRT)(Hsieh, 1993).如利用加速质谱仪检出土壤中14C含量在短期内的变化, 可以间接反映土壤碳的分解与更新速率.同时, 15N标记在氮添加实验中的应用不仅可以有效地检验氮素的循环过程, 而且可以在较短时期内(可以假定C:N不变)通过检验不同碳库中的15N含量及碳氮比, 并结合氮平衡法(N-balance method, 测定15N在土壤中的积累量并结合土壤C:N来估算土壤碳的变化量)来检测氮添加下土壤碳变化(de Vries et al., 2006; Mol et al., 2009).此外, 碳氮同位素的应用还可以有效地示踪土壤有机碳的来源和周转过程(Ehleringer et al., 2000), 揭示土壤碳氮耦合变化关系, 这也必将成为今后相关研究的重要方面. ...
Anthropogenic N deposition increases soil C storage by decreasing the extent of litter decay: Analysis of field observations with an ecosystem model.
1
2012
... 氮添加可促进腐殖质的形成, 促进分解残物向稳定性碳的转变, 增加稳定性碳积累(Moran et al., 2005; Whittinghill et al., 2012).氮元素与较难分解的凋落物残体(如木质素等)结合形成更难分解的杂环类物质(如吲哚等)和酚类是氮添加促进难分解碳形成的一个机制(Berg, 2000; Janssens et al., 2010).而氮添加对团聚体(aggregate, 即土壤碳的物理保护)的促进作用可能不明显(Janssens et al., 2010).尽管氮添加对稳定性碳库的促进作用在短期内可能对土壤总有机碳库影响不显著, 但其长期作用明显(Reid et al., 2012).土壤稳定性碳是影响土壤有机碳库稳定与积累的重要因素, 加强对土壤稳定性碳的研究对于从机理上深入认识氮添加对土壤有机碳库的影响及其响应机制具有重要作用. ...
Global response patterns of terrestrial plant species to nitrogen addition.
2
2008
... 生态系统碳循环与氮素存在耦合关系, 碳积累与氮素供给密切相关(Luo et al., 2006; Kirkby et al., 2013).碳氮耦合作用在植被的光合生产、生长与分配过程以及土壤养分的循环过程中均发挥着重要作用.目前普遍认为氮添加可促进植被净初级生产力增加及植被生长, 提高植被碳储量(Magnani et al., 2007; Reay et al., 2008; Xia & Wan, 2008; LeBauer & Treseder, 2008; Janssens & Luyssaert, 2009; Thomas et al., 2010).不同植被类型及气候条件下的植被碳汇增量(此处将每克氮素输入所增加的碳量暂称为碳汇增量)为30-200 g C (de Vries et al., 2008; Sutton et al., 2008; Reay et al., 2008; Thomas et al., 2010; Templer et al., 2012).Magnani等(2007)研究认为每增加1 g氮输入, 可促使森林生态系统增加约470 g C.de Vries等(2008)则认为Magnani等(2007)的研究高估了该值, 重新估算的碳汇增量为30-70 g C, 其中植被碳库增量为20-40 g C, 土壤碳库增量为10-30 g C. ...
... 一些研究显示施氮可显著增加植被凋落物量(Bradley et al., 2006; Adamek et al., 2009), 且凋落物量随氮添加水平增加而增加.例如, 樊后保等(2007)研究指出: 高氮添加(240 kg N∙hm-2∙a-1)显著增加森林凋落物量(2599.5 kg∙hm-2∙a-1), 而中氮(120 kg N∙hm-2∙a-1)和低氮(60 kg N∙hm-2∙a-1)对凋落物量增加的促进作用不明显.但也有研究发现: 尽管氮添加可以促进植被生产力增加, 但地上叶片与地下根系凋落物输入对氮添加并无明显的正反馈作用(Zak et al., 2008; Janssens et al., 2010).此外, 氮添加可以增加叶片和细根氮含量(Giardina et al., 2003; Xia & Wan, 2008; Zak et al., 2008; Yuan & Chen, 2012), 可能促进叶片和细根凋落物的分解(Waldrop et al., 2004; Knorr et al., 2005; Parton et al., 2007; Hobbie et al., 2012), 而不利于土壤碳的积累.然而, 有研究显示氮添加只在初期促进凋落物分解, 主要包括水溶性物质和非木质性纤维素的分解, 随着中后期难分解的木质素含量的增多, 氮添加抑制了凋落物的分解(Carreiro et al., 2000). ...
A global analysis of fine root production as affected by soil nitrogen and phosphorus.
1
2012
... 一些研究显示施氮可显著增加植被凋落物量(Bradley et al., 2006; Adamek et al., 2009), 且凋落物量随氮添加水平增加而增加.例如, 樊后保等(2007)研究指出: 高氮添加(240 kg N∙hm-2∙a-1)显著增加森林凋落物量(2599.5 kg∙hm-2∙a-1), 而中氮(120 kg N∙hm-2∙a-1)和低氮(60 kg N∙hm-2∙a-1)对凋落物量增加的促进作用不明显.但也有研究发现: 尽管氮添加可以促进植被生产力增加, 但地上叶片与地下根系凋落物输入对氮添加并无明显的正反馈作用(Zak et al., 2008; Janssens et al., 2010).此外, 氮添加可以增加叶片和细根氮含量(Giardina et al., 2003; Xia & Wan, 2008; Zak et al., 2008; Yuan & Chen, 2012), 可能促进叶片和细根凋落物的分解(Waldrop et al., 2004; Knorr et al., 2005; Parton et al., 2007; Hobbie et al., 2012), 而不利于土壤碳的积累.然而, 有研究显示氮添加只在初期促进凋落物分解, 主要包括水溶性物质和非木质性纤维素的分解, 随着中后期难分解的木质素含量的增多, 氮添加抑制了凋落物的分解(Carreiro et al., 2000). ...
... 相对于植被碳汇, 目前关于土壤碳汇对氮添加的响应争议较大.Mack等(2004)研究发现氮添加改变了苔原地区植物群落组成并促进了凋落物分解, 从而使土壤有机碳储量减少, 抵消了氮添加下的植被碳汇增量, 导致生态系统碳储量减少.同样, Cleveland和Townsend (2006)研究发现, 氮添加促进土壤呼吸而减小土壤有机碳库储量.Nadelhoffer等(1999)则认为氮添加对欧洲森林生态系统碳库影响较小.此外, 较多研究显示氮添加增加了土壤碳储量(de Vries et al., 2006; Reay et al., 2008).例如, Zak等(2008)指出氮添加(30 kg N∙hm-2∙a-1)处理的森林土壤有机碳含量显著高于对照土壤, 其增幅约12%.氮添加同样可促进北欧云杉林和松树林土壤碳吸收, 其碳汇增量为11 g C (Hyvönen et al., 2008).Reay等(2008)对以往的研究进行了综述, 认为氮添加下的土壤碳库增量为7-23 g C.Meta分析的结果表明, 森林土壤在氮添加下的碳汇增量为19 g C (Janssens et al., 2010); 而Lu等(2011)的结果显示森林和草地的土壤碳库变化很小.经过文献梳理和总结, 氮添加下土壤碳汇增量范围为0-30 g C, 低于植被碳汇增量(de Vries et al., 2006, 2008; Reay et al., 2008; Janssens et al., 2010).整体而言, 目前关于氮添加条件下土壤碳汇的变化方向与强度争议较大, 亟需对其影响因子及机理进行深入研究分析. ...
... 一些研究显示施氮可显著增加植被凋落物量(Bradley et al., 2006; Adamek et al., 2009), 且凋落物量随氮添加水平增加而增加.例如, 樊后保等(2007)研究指出: 高氮添加(240 kg N∙hm-2∙a-1)显著增加森林凋落物量(2599.5 kg∙hm-2∙a-1), 而中氮(120 kg N∙hm-2∙a-1)和低氮(60 kg N∙hm-2∙a-1)对凋落物量增加的促进作用不明显.但也有研究发现: 尽管氮添加可以促进植被生产力增加, 但地上叶片与地下根系凋落物输入对氮添加并无明显的正反馈作用(Zak et al., 2008; Janssens et al., 2010).此外, 氮添加可以增加叶片和细根氮含量(Giardina et al., 2003; Xia & Wan, 2008; Zak et al., 2008; Yuan & Chen, 2012), 可能促进叶片和细根凋落物的分解(Waldrop et al., 2004; Knorr et al., 2005; Parton et al., 2007; Hobbie et al., 2012), 而不利于土壤碳的积累.然而, 有研究显示氮添加只在初期促进凋落物分解, 主要包括水溶性物质和非木质性纤维素的分解, 随着中后期难分解的木质素含量的增多, 氮添加抑制了凋落物的分解(Carreiro et al., 2000). ...
... ; Zak et al., 2008; Yuan & Chen, 2012), 可能促进叶片和细根凋落物的分解(Waldrop et al., 2004; Knorr et al., 2005; Parton et al., 2007; Hobbie et al., 2012), 而不利于土壤碳的积累.然而, 有研究显示氮添加只在初期促进凋落物分解, 主要包括水溶性物质和非木质性纤维素的分解, 随着中后期难分解的木质素含量的增多, 氮添加抑制了凋落物的分解(Carreiro et al., 2000). ...
... 氮添加通过抑制土壤呼吸作用与土壤稳定性碳的分解而促进土壤碳的积累, 这也是目前被广泛接受的解释(Zak et al., 2008; Reay et al., 2008; Janssens & Luyssaert, 2009; Cusack et al., 2010; Ramirez et al., 2012). ...



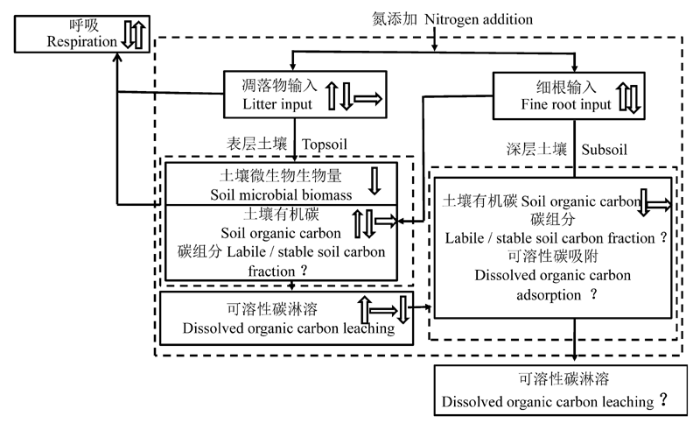
{kind=link}
{kind=link}