Leaf is the most important organ for carbon-water coupling of a plant because it is the primary medium for photosynthesis. It also acts as the hydraulic bottleneck and safety valve against hydraulic catastrophic dysfunctions. The leaf economics spectrum, which reflects the balance between investments and returns of leaf economic traits, provides a useful framework for examining species strategies as shaped by their evolutionary history. Changes in leaf hydraulic traits will influence leaf economic traits as well as plant survival and growth. Exploring trade-offs between leaf hydraulic and economic traits is thus of significance for modeling carbon-water relations, understanding the mechanisms of water/carbon investments, and extending the leaf economic spectrum. In this review, we first examined the trade-offs between leaf hydraulic and economic traits. Specially, we analyzed the relationships between leaf hydraulic conductivity and hydraulic vulnerability, water potential at the turgor loss point, water capacitance, safety margin, and leaf morphological, structural and functional traits. We then discussed potential mechanisms regulating leaf hydraulic and economic traits from leaf morphology, anatomy, venation, and stomatal functions. Finally, we proposed future research to: (1) develop an integrated whole-plant economics spectrum, including carbon-nitrogen-water resources and root-stem-leaf hydraulic transport system that will help revealing ecophysiological mechanisms of plant structure-functional coupling, carbon sequestration and water use; (2) explore a generalized trade-offs among leaf hydraulic safety, hydraulic efficiency and carbon fixation efficiency to advance our understanding of the relationships between biophysical structure and physiological metabolism in plant leaf construction under drought stress; and (3) explore the carbon-water metabolic relationship and coupling of water transport and growth rate for the metabolic theory and predictions at community scale.
JINYing, WANGChuan-Kuan. Trade-offs between plant leaf hydraulic and economic traits. Chinese Journal of Plant Ecology, 2015, 39(10): 1021-1032 https://doi.org/10.17521/cjpe.2015.0099
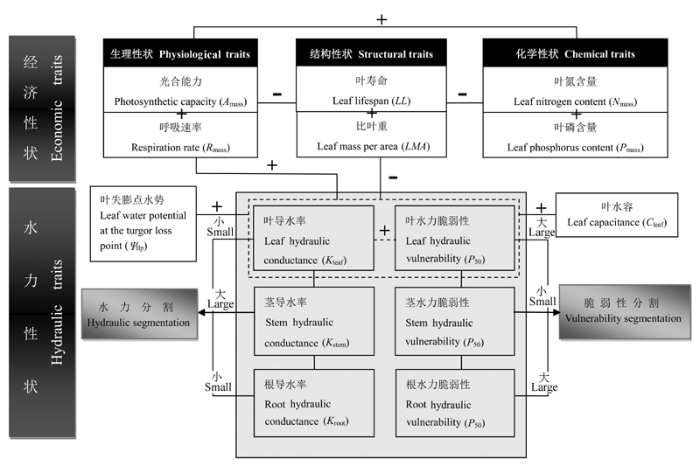
Fig. 1 The trade-offs among leaf hydraulic and economic traits. “+” represents positive relationships, “-” represents negative relationships; the dashed line region represents that the relationships are uncertain, where “+” represents a potential trade-off relationship; the gay region represents individual-level hydraulic traits.
Leaf anatomical characteristics associated with shoot hydraulic conductance, stomatal conductance and stomatal sensitivity to changes of leaf water status in temperate deciduous trees.
Australian Journal of Plant Physiology, 28, 765-774.
Dynamic changes in hydraulic conductivity in petioles of two savanna tree species: Factors and mechanisms contributing to the refilling of embolized vessels.
The stem xylem of Patagonian shrubs operates far from the point of catastrophic dysfunction and is additionally protected from drought-induced embolism by leaves and roots.
Xylem density, biomechanics and anatomical traits correlate with water stress in 17 evergreen shrub species of the Mediterranean-type climate region of South Africa.
Leaf hydraulic conductance, measured in situ, declines and recovers daily: Leaf hydraulics, water potential and stomatal conductance in four temperate and three tropical tree species.
Photosynthetically relevant foliar traits correlating better on a mass vs an area basis: Of ecophysiological relevance or just a case of mathematical imperatives and statistical quicksand?
New Phytologist, 199, 311-321.
[48]
Lo GulloMA, NardiniA, TrifilòP, SalleoS (2003).
Changes in leaf hydraulics and stomatal conductance following drought stress and irrigation in Ceratonia siliqua (Carob tree).
Growth dynamics of root and shoot hydraulic conductance in seedlings of five neotropical tree species: Scaling to show possible adaptation to differing light regimes.
Are symplast tolerance to intense drought conditions and xylem vulnerability to cavitation coordinated? An integrated analysis of photosynthetic, hydraulic and leaf level processes in two Mediterranean drought-resistant species.
Leaf hydraulic conductance in relation to anatomical and functional traits during Populus tremula leaf ontogeny.
1
2005
... 叶片通过一系列形态-解剖结构调整, 提高植物的抗旱能力, 这是植物生存和竞争的关键(Simonin et al., 2012).在水分胁迫条件下, Kleaf的变化与木质部导管和细胞性状直接相关(Cochard et al., 2004; Nardini et al., 2005).Kleaf与中脉木质部导管的宽度(Aasamaa et al., 2001; Sack et al., 2004)和细胞间隙的比例正相关, 而与叶肉细胞和表皮的体积负相关(Aasamaa et al., 2005; 张志亮等, 2014).导管直径降低会导致叶片水分运输效率下降, 但可通过增加导管数量或叶脉密度来平衡(Givnish, 1987).叶片水力脆弱性(P50)与导管直径有关, 与导管壁厚无关(Nardini et al., 2012a); 而导管直径与壁厚、中脉木质部导管密度呈负相关; 表明直径小且数量多的导管P50较低.因此, 通过调整导管直径和数量可以提高叶片的水力安全(低P50).抗旱植物在提高自身水力安全的同时, 也增加了叶片组织构建上的碳投资, 提高了叶片水分利用的成本(Simonin et al., 2012), 因而, 可能会降低在资源丰富地区的竞争能力(Nardini et al., 2012a).由于叶脉密度、导管大小和数量等随着发育程度的变化, Kleaf在不同生长条件下具有高度的可塑性(Sack & Holbrook, 2006).热带雨林树种的Kleaf与栅栏组织的厚度、栅栏组织/海绵组织的比例有关(Sack & Frole, 2006), 而温带树种的Kleaf则与总的叶片厚度和单位叶片面积的储水能力有关(Sack et al., 2003; Cochard et al., 2004).Kröber等(2015)研究了亚热带地区21个落叶树种和19个常绿树种的叶片形态和生理特性, 结果表明 常绿树种通常有较厚的海绵和栅栏组织.这种水力特性与叶片结构上的相关性可能与结构的协调性(解剖和发育结构)、或与功能协调性有关(为适应某一特定环境而进行的特性的选择) (Sack & Holbrook, 2006).综上所述, 木质部结构的变化为Kleaf与P50之间的关系提供了一种解释. ...
Hydraulic conductance and stomatal sensitivity to changes of leaf water status in six deciduous tree species.
1
2001
... 叶片通过一系列形态-解剖结构调整, 提高植物的抗旱能力, 这是植物生存和竞争的关键(Simonin et al., 2012).在水分胁迫条件下, Kleaf的变化与木质部导管和细胞性状直接相关(Cochard et al., 2004; Nardini et al., 2005).Kleaf与中脉木质部导管的宽度(Aasamaa et al., 2001; Sack et al., 2004)和细胞间隙的比例正相关, 而与叶肉细胞和表皮的体积负相关(Aasamaa et al., 2005; 张志亮等, 2014).导管直径降低会导致叶片水分运输效率下降, 但可通过增加导管数量或叶脉密度来平衡(Givnish, 1987).叶片水力脆弱性(P50)与导管直径有关, 与导管壁厚无关(Nardini et al., 2012a); 而导管直径与壁厚、中脉木质部导管密度呈负相关; 表明直径小且数量多的导管P50较低.因此, 通过调整导管直径和数量可以提高叶片的水力安全(低P50).抗旱植物在提高自身水力安全的同时, 也增加了叶片组织构建上的碳投资, 提高了叶片水分利用的成本(Simonin et al., 2012), 因而, 可能会降低在资源丰富地区的竞争能力(Nardini et al., 2012a).由于叶脉密度、导管大小和数量等随着发育程度的变化, Kleaf在不同生长条件下具有高度的可塑性(Sack & Holbrook, 2006).热带雨林树种的Kleaf与栅栏组织的厚度、栅栏组织/海绵组织的比例有关(Sack & Frole, 2006), 而温带树种的Kleaf则与总的叶片厚度和单位叶片面积的储水能力有关(Sack et al., 2003; Cochard et al., 2004).Kröber等(2015)研究了亚热带地区21个落叶树种和19个常绿树种的叶片形态和生理特性, 结果表明 常绿树种通常有较厚的海绵和栅栏组织.这种水力特性与叶片结构上的相关性可能与结构的协调性(解剖和发育结构)、或与功能协调性有关(为适应某一特定环境而进行的特性的选择) (Sack & Holbrook, 2006).综上所述, 木质部结构的变化为Kleaf与P50之间的关系提供了一种解释. ...
Leaf anatomical characteristics associated with shoot hydraulic conductance, stomatal conductance and stomatal sensitivity to changes of leaf water status in temperate deciduous trees.
1
2001
... 叶片水容(Cleaf)表征植物调节根系供水和叶片蒸腾需水之间的不平衡、缓冲叶片水势波动的能力, 是植物避免干旱的水力对策(Lamont & Lamont, 2000; Sack et al., 2003, 2005).Blackman等(2010)测定了20种温带木本植物的叶片体积水容和叶片组织水容, 结果表明Kleaf与叶片组织水容呈正相关, 但与体积水容无关.这说明并不是所有的叶片储存水都可以补充蒸腾的水分损失, 只有叶片组织中的储存水可以与蒸腾流进行自由交换, 支持了叶片水力分区的观点(Zwieniecki et al., 2007).Kleaf与Cleaf功能上的协调性解释了蒸腾速率一定的情况下Kleaf较高的叶片关闭气孔较慢的原因(Tyree et al., 1998; Aasamaa & Sõber, 2001; Hao et al., 2008). ...
Contrasting drought-response strategies in California redwoods.
1
2015
... 叶片水力安全阈值(safety margin)是指叶片气孔关闭所对应的叶片水势(Ψmin)与P50的差值(Brodribb & Holbrook, 2004b; Sperry, 2004), 用于表征植物的水力对策(Apgaua et al., 2015).安全阈值较大的物种, 当Kleaf显著下降时会关闭气孔, 保护木质部安全, 从而最大限度地降低栓塞风险(Cochard, 2002; Cochard et al., 2002; Nardini et al., 2003; Brodribb & Holbrook, 2004b); 相反, 安全阈值较小的物种, 当Kleaf下降时, 气孔并不完全关闭, 从而使植物能够长时间地维持气体交换(Salleo et al., 2000, 2001; Brodribb & Holbrook, 2003, 2004a, 2004b; Bucci et al., 2003; Lo Gullo et al., 2003), 直至Kleaf下降到空穴化不可逆转的水平(Sperry et al., 2002; Brodribb & Holbrook, 2004b).通常, 对空穴化更脆弱的物种, 其水力安全阈值较小, 在非干旱条件下也有可能经历栓塞(Meinzer et al., 2009), 但其修复栓塞的能力较大(Johnson et al., 2009, 2011; Choat et al., 2012; Ambrose et al., 2015), 在修复栓塞的同时也会引起巨大的代谢消耗(Nardini et al., 2011).纵观森林群落中的各种物种, 虽然对干旱发生的响应差异较大, 但总体上被子植物的安全阈值范围很小, 常不能随着水分可利用性的巨大差异而发生变化, 因而在旱季有木质部栓塞的风险(Choat et al., 2012). ...
Functional traits and water transport strategies in lowland tropical rainforest trees.
1
2015
... 叶片水力安全阈值(safety margin)是指叶片气孔关闭所对应的叶片水势(Ψmin)与P50的差值(Brodribb & Holbrook, 2004b; Sperry, 2004), 用于表征植物的水力对策(Apgaua et al., 2015).安全阈值较大的物种, 当Kleaf显著下降时会关闭气孔, 保护木质部安全, 从而最大限度地降低栓塞风险(Cochard, 2002; Cochard et al., 2002; Nardini et al., 2003; Brodribb & Holbrook, 2004b); 相反, 安全阈值较小的物种, 当Kleaf下降时, 气孔并不完全关闭, 从而使植物能够长时间地维持气体交换(Salleo et al., 2000, 2001; Brodribb & Holbrook, 2003, 2004a, 2004b; Bucci et al., 2003; Lo Gullo et al., 2003), 直至Kleaf下降到空穴化不可逆转的水平(Sperry et al., 2002; Brodribb & Holbrook, 2004b).通常, 对空穴化更脆弱的物种, 其水力安全阈值较小, 在非干旱条件下也有可能经历栓塞(Meinzer et al., 2009), 但其修复栓塞的能力较大(Johnson et al., 2009, 2011; Choat et al., 2012; Ambrose et al., 2015), 在修复栓塞的同时也会引起巨大的代谢消耗(Nardini et al., 2011).纵观森林群落中的各种物种, 虽然对干旱发生的响应差异较大, 但总体上被子植物的安全阈值范围很小, 常不能随着水分可利用性的巨大差异而发生变化, 因而在旱季有木质部栓塞的风险(Choat et al., 2012). ...
Plant science: The hidden cost of transpiration.
1
2010
... 植物为了容忍叶片机械破坏和导管栓塞, 需要构建稠密的叶脉冗余网络, 以提供额外的水流路径, 但同时也增加了巨大的碳投资(Corson, 2010; Scoffoni et al., 2011; Nardini et al., 2012a; Pantin et al., 2012).近年来, 叶脉功能性状与叶片水力、光合能力、叶片碳构建等关系受到广泛的关注(Lamont & Lamont, 2000; Beerling & Franks, 2010; Blonder et al., 2011; Sack et al., 2012, 2013; Zhang et al., 2012).Brodribb等(2007)提出较高的叶片导水率和光合能力的耦合与叶脉密度有关.Nardini等(2012a)指出, Kleaf_mass和P50与叶脉密度和LMA均存在负相关关系, 表明植物通过提高叶脉密度, 增加其抗旱阻力, 但同时也增加了单位叶面积的碳构建成本(高LMA) (Blonder et al., 2011).Prado和Maurel (2013)的叶片水力模型表明叶片导水率和不同等级的叶脉密度共同促成最优化的木质部网络水力效率.叶片主脉密度对Kleaf的影响微小(Sack & Frole, 2006), 而细脉密度对Kleaf的影响较大.这是因为细脉密度的增加, 不仅增加了叶片木质部的导水率(Cochard et al., 2004), 同时也增加了木质部与周围叶肉组织接触的表面积, 降低了木质部外水流路径的长度(Roth-Nebelsick et al., 2001; Sack & Frole, 2006).由于低等级叶脉具有较高的导水率, 因此, 与碳资源投资相比, 较高的叶脉密度在经济上是可行的. ...
Leaf hydraulic vulnerability is related to conduit dimensions and drought resistance across a diverse range of woody angiosperms.
5
2010
... P50值(叶片导水率损失50%对应的叶片水势)是常用于量化叶片水力脆弱性的指标.Kleaf和P50是植物适应不同环境条件的主要生理性状(Sack et al., 2005; Nardini et al., 2012a), 决定植物的竞争能力和生产力(Nardini & Luglio, 2014).一般而言, 植物茎干栓塞阻力(用P50表示)的增加常会引起水力效率(Kstem)的降低(Bucci et al., 2006)或碳消耗的增加(Jacobsen et al., 2007).然而, 由于叶片水平的P50与基于叶面积的叶片导水率(Kleaf_area)缺少相关性, 人们常认为不存在水力效率与安全的权衡(Brodribb & Holbrook, 2003, 2004b; Lo Gullo et al., 2003; Nardini et al., 2003, 2012a; Blackman et al., 2010; Scoffoni et al., 2011).但近期Nardini和Luglio (2014)综合分析了全球130个木本被子植物叶片水力数据发现: P50与Kleaf_area确实没有相关性, 但与基于叶干质量的叶片导水率(Kleaf_mass)存在弱正相关关系, 表明叶片水平可能存在水力安全和水力效率的权衡.Kleaf_mass不仅可以解释叶片水力系统的水分运输效率, 还可估算构建叶片水力系统的碳消耗(Nardini et al., 2012b; Simonin et al., 2012); 抗旱阻力强的叶片(P50低), 其碳消耗较大(Nardini et al., 2012a).至于这种权衡关系是否有普适性, 还需验证. ...
... 叶片失膨点水势或萎蔫水势(Ψtlp)是叶片对干旱胁迫耐性的决定因子, 对植株的生理干旱耐性有重要贡献(Maréchaux et al., 2015).有研究报道Ψtlp与P50、Kleaf呈正相关(Nardini et al., 2012a; Villagra et al., 2013), 表明Ψtlp低(即叶片耐旱性高)的物种, 其叶片组织的供水能力低(Kleaf低), 但抗栓塞阻力强(P50低); 相反, 水分运输效率高(Kleaf高)的物种, 能够保持叶片良好的水分状况, 阻止叶片水势的下降, 从而保持气孔开放和碳同化时间长.P50与Ψtlp的协调关系不仅解释了叶片木质部外导水率下降的原因(Brodribb & Holbrook, 2004b; Knipfer & Steudle, 2008), 还表明了叶片内部非原生质体和共质体之间抗旱阻力的协调关系(Blackman et al., 2010; Vilagrosa et al., 2010; Nardini & Luglio, 2014).此外, Brodribb和Holbrook (2003)的研究表明P50和Ψtlp影响水分胁迫期间的气孔关闭时间.在空穴化发生早期, 首先引起叶片气孔关闭(Choat et al., 2012), 在叶片膨胀体积逐渐损失直至完全丧失之前, 逐渐降低叶片蒸腾速率(Blackman et al., 2010; Scoffoni et al., 2011; Nardini et al., 2012a).因此, 叶片是保护活细胞远离临界水分状况的安全阀门. ...
... ), 在叶片膨胀体积逐渐损失直至完全丧失之前, 逐渐降低叶片蒸腾速率(Blackman et al., 2010; Scoffoni et al., 2011; Nardini et al., 2012a).因此, 叶片是保护活细胞远离临界水分状况的安全阀门. ...
... 叶片水容(Cleaf)表征植物调节根系供水和叶片蒸腾需水之间的不平衡、缓冲叶片水势波动的能力, 是植物避免干旱的水力对策(Lamont & Lamont, 2000; Sack et al., 2003, 2005).Blackman等(2010)测定了20种温带木本植物的叶片体积水容和叶片组织水容, 结果表明Kleaf与叶片组织水容呈正相关, 但与体积水容无关.这说明并不是所有的叶片储存水都可以补充蒸腾的水分损失, 只有叶片组织中的储存水可以与蒸腾流进行自由交换, 支持了叶片水力分区的观点(Zwieniecki et al., 2007).Kleaf与Cleaf功能上的协调性解释了蒸腾速率一定的情况下Kleaf较高的叶片关闭气孔较慢的原因(Tyree et al., 1998; Aasamaa & Sõber, 2001; Hao et al., 2008). ...
... 许多研究发现, LL和LMA与Kleaf_mass负相关, 而与Kleaf_area无关(Nardini, 2001; Wright et al., 2001; Shipley, 2002; Sack et al., 2003, 2005; Brodribb et al., 2005; Blackman et al., 2010; Nardini et al., 2012a; Simonin et al., 2012; Bucci et al., 2013).原因可能是Kleaf_area更偏向于发生在叶片-大气界面之间的叶片表面的过程, 而Kleaf_mass是基于整个叶片组织的过程(Nardini et al., 2012a).Nardini等(2012b)研究发现 Quercus ilex阳生叶的Kleaf_mass比阴生叶高40%, 而Kleaf_area是阴生叶的2倍多; 这表明同一冠层叶片对不同光强度的水力驯化增加了单位叶面积的水分运输效率, 但叶片水分运输系统的碳消耗保持相对稳定.近期, Maréchaux等(2015)研究了法属圭亚那地区(French Guiana) 71个热带树种165株树木的Ψtlp与叶片经济性状的关系, 发现Ψtlp与叶片硬度和厚度呈弱正相关关系, 但与LMA、叶面积、叶片氮浓度、叶片碳浓度等性状没有相关性.可见, 叶片水力性状与经济性状之间是否有普适性关系还需要更多的研究加以验证. ...
Venation networks and the origin of the leaf economics spectrum.
2
2011
... 植物为了容忍叶片机械破坏和导管栓塞, 需要构建稠密的叶脉冗余网络, 以提供额外的水流路径, 但同时也增加了巨大的碳投资(Corson, 2010; Scoffoni et al., 2011; Nardini et al., 2012a; Pantin et al., 2012).近年来, 叶脉功能性状与叶片水力、光合能力、叶片碳构建等关系受到广泛的关注(Lamont & Lamont, 2000; Beerling & Franks, 2010; Blonder et al., 2011; Sack et al., 2012, 2013; Zhang et al., 2012).Brodribb等(2007)提出较高的叶片导水率和光合能力的耦合与叶脉密度有关.Nardini等(2012a)指出, Kleaf_mass和P50与叶脉密度和LMA均存在负相关关系, 表明植物通过提高叶脉密度, 增加其抗旱阻力, 但同时也增加了单位叶面积的碳构建成本(高LMA) (Blonder et al., 2011).Prado和Maurel (2013)的叶片水力模型表明叶片导水率和不同等级的叶脉密度共同促成最优化的木质部网络水力效率.叶片主脉密度对Kleaf的影响微小(Sack & Frole, 2006), 而细脉密度对Kleaf的影响较大.这是因为细脉密度的增加, 不仅增加了叶片木质部的导水率(Cochard et al., 2004), 同时也增加了木质部与周围叶肉组织接触的表面积, 降低了木质部外水流路径的长度(Roth-Nebelsick et al., 2001; Sack & Frole, 2006).由于低等级叶脉具有较高的导水率, 因此, 与碳资源投资相比, 较高的叶脉密度在经济上是可行的. ...
... ) (Blonder et al., 2011).Prado和Maurel (2013)的叶片水力模型表明叶片导水率和不同等级的叶脉密度共同促成最优化的木质部网络水力效率.叶片主脉密度对Kleaf的影响微小(Sack & Frole, 2006), 而细脉密度对Kleaf的影响较大.这是因为细脉密度的增加, 不仅增加了叶片木质部的导水率(Cochard et al., 2004), 同时也增加了木质部与周围叶肉组织接触的表面积, 降低了木质部外水流路径的长度(Roth-Nebelsick et al., 2001; Sack & Frole, 2006).由于低等级叶脉具有较高的导水率, 因此, 与碳资源投资相比, 较高的叶脉密度在经济上是可行的. ...
Angiosperm leaf vein evolution was physiologically and environmentally transformative.
2
2009
... Sack等(2013)基于大范围物种研究提出了“flux trait network hypothesis”假说, 阐明了叶脉性状对水力性状和气体交换性状的影响机制: 即叶脉密度通过Kleaf对气孔导度和基于叶面积表达的光合速率(Aarea)产生影响, 进而影响基于叶干质量表达的光合速率(Amass)和相对生长速率(Lamont & Lamont, 2000; Brodribb et al., 2007; Boyce et al., 2009; Brodribb et al., 2010; McKown et al., 2010; Walls, 2011).尽管该假说不一定适用于所有物种, 但它阐明了叶脉、水力、气孔和气体交换性状之间的关系.从某种意义上讲, 叶脉密度反映了资源流通能力(Sack et al., 2013).因此, 叶脉密度对叶片碳、水通量的影响可促成“快-慢”经济型物种.叶脉系统的进化, 也支持了高水分流通速率和其他快速叶片经济型谱(Lamont & Lamont, 2000; McDowell et al., 2008; Boyce et al., 2009; Sack & Scoffoni, 2013).因此, 该假说在预测植物地理分布和对气候变化的响应上有潜在的应用前景. ...
Stem hydraulic supply is linked to leaf photosynthetic capacity: Evidence from New Caledonian and Tasmanian rainforests.
2
2000
... 植物一方面要最大限度地吸收CO2进行光合作用, 另一方面要最小限度地控制水分散失, 这两个生理过程共享同一个气孔路径(Xiong et al., 2015).因此, 植物通过气孔吸收CO2的气孔导度(Gs, 气相水分散失能力指标)与释放水蒸气的水力导度(Kleaf, 液相水分传输能力指标)之间存在权衡机制(Sack & Holbrook, 2006).由于C3植物的Gs与光合速率的耦合关系, 因此Kleaf与光合速率也存在正相关关系(Brodribb et al., 2005, 2007).对系统发生、叶片结构、叶片寿命、地理分布等特性差异较大的大量物种的综合研究发现, Kleaf与Gs和光合速率呈正相关关系(Brodribb et al., 2005, 2007), 表明水力供需能力及碳获取与水分运输能力之间存在平衡关系(Brodribb & Feild, 2000; Meinzer, 2002; Sack et al., 2003, 2005; Brodribb & Holbrook, 2004b; Brodribb et al., 2005; Campanello et al., 2008; Prentice et al., 2014), 也反映叶片水平的气液两相导度的协调性; 有效的叶片水分供应能够最大限度地维持气孔开放(Brodribb et al., 2005, 2007; Lo Gullo et al., 2005; Nardini et al., 2005; Manzoni et al., 2013).Katul等(2003)提出了一个把Gs、光合速率和水分运输效率联系在一起的理论框架模型, 可以直接预测最大羧化能力与土壤-植物水力性状之间的关系. ...
... 不同环境条件下同一物种或不同物种的叶片导水率与最大光合速率密切相关(Zhang & Cao, 2009), 表明水力特性对气孔导度和光合气体交换起限制作用.Scoffoni等(2015)研究不同光强下叶片水力和气体交换、解剖、结构特性等的关系, 发现这些特性均有可塑性, 随着光照环境的不同而协同变化以适应不同光照环境, 达到最优生长(Scoffoni et al., 2015).在水分状况不同的地区, 植物叶片水分运输效率与光合能力之间也存在协调作用(Brodribb & Feild, 2000; Brodribb et al., 2002; Santiago et al., 2004).湿润地区的植物为了快速进行光合作用而有较高的Kleaf, 从而使植物在有较大气体交换速率的同时, 无需增加叶片致命脱水的风险.因此, 高Kleaf植物在湿润地区具有较大的竞争优势(Meinzer, 2002; Brodribb et al., 2007).然而, 干旱地区植物的首要任务是维持叶片水力系统的功能, 其次是具有较高的气体交换速率. ...
Leaf maximum photosynthetic rate and venation are linked by hydraulics.
7
2007
... LES (Wright et al., 2004)认为, 植物的化学、结构和生理功能性状呈一条连续变化的带谱, 在带谱的两端分别是“快速投资-收益”型和“缓慢投资-收益”型物种, 其间是两种类型的过渡梯度类型.叶片水力性状影响叶片碳同化能力、水分利用效率和植物生长(Sack & Holbrook, 2006), 对植物生产力和进化起着重要的作用(Brodribb et al., 2010).叶片导水率(Kleaf)和叶片水力脆弱性(P50)常用于反映叶片水力效率和水力安全; 两者的权衡关系可限制物种地理分布, 对不同空间尺度的植被组成和分布非常重要(Nardini et al., 2012a).因此, 近期越来越多的研究关注叶片水力性状对叶片经济性状的影响(Sack & Frole, 2006; Sack & Scoffoni, 2013; Sack et al., 2013; 龚容和高琼, 2015).例如, Villagra等(2013)提出LES反映的碳和养分“投资-收益”的平衡与叶片水力性状有关.Kleaf高的物种, 叶片组织密度低、寿命短, 资源获取速率快(Reich, 2014).此外, 由于叶片水平水分流动阻力占整个植株阻力的比例较大(≥30%), 且与叶片气体交换有关, 因此, Kleaf对植物碳经济、水经济及两者之间的耦合关系均至关重要(Brodribb et al., 2005, 2007; Reich, 2014).为此, 本文首先综述植物叶片水力性状、经济性状及两者之间的权衡关系; 然后从结构和功能角度探讨叶片水力性状与经济性状的调节机制; 最后针对植物的水力和经济性状耦合关系提出今后的研究方向. ...
... 植物一方面要最大限度地吸收CO2进行光合作用, 另一方面要最小限度地控制水分散失, 这两个生理过程共享同一个气孔路径(Xiong et al., 2015).因此, 植物通过气孔吸收CO2的气孔导度(Gs, 气相水分散失能力指标)与释放水蒸气的水力导度(Kleaf, 液相水分传输能力指标)之间存在权衡机制(Sack & Holbrook, 2006).由于C3植物的Gs与光合速率的耦合关系, 因此Kleaf与光合速率也存在正相关关系(Brodribb et al., 2005, 2007).对系统发生、叶片结构、叶片寿命、地理分布等特性差异较大的大量物种的综合研究发现, Kleaf与Gs和光合速率呈正相关关系(Brodribb et al., 2005, 2007), 表明水力供需能力及碳获取与水分运输能力之间存在平衡关系(Brodribb & Feild, 2000; Meinzer, 2002; Sack et al., 2003, 2005; Brodribb & Holbrook, 2004b; Brodribb et al., 2005; Campanello et al., 2008; Prentice et al., 2014), 也反映叶片水平的气液两相导度的协调性; 有效的叶片水分供应能够最大限度地维持气孔开放(Brodribb et al., 2005, 2007; Lo Gullo et al., 2005; Nardini et al., 2005; Manzoni et al., 2013).Katul等(2003)提出了一个把Gs、光合速率和水分运输效率联系在一起的理论框架模型, 可以直接预测最大羧化能力与土壤-植物水力性状之间的关系. ...
... , 2007), 表明水力供需能力及碳获取与水分运输能力之间存在平衡关系(Brodribb & Feild, 2000; Meinzer, 2002; Sack et al., 2003, 2005; Brodribb & Holbrook, 2004b; Brodribb et al., 2005; Campanello et al., 2008; Prentice et al., 2014), 也反映叶片水平的气液两相导度的协调性; 有效的叶片水分供应能够最大限度地维持气孔开放(Brodribb et al., 2005, 2007; Lo Gullo et al., 2005; Nardini et al., 2005; Manzoni et al., 2013).Katul等(2003)提出了一个把Gs、光合速率和水分运输效率联系在一起的理论框架模型, 可以直接预测最大羧化能力与土壤-植物水力性状之间的关系. ...
... , 2007; Lo Gullo et al., 2005; Nardini et al., 2005; Manzoni et al., 2013).Katul等(2003)提出了一个把Gs、光合速率和水分运输效率联系在一起的理论框架模型, 可以直接预测最大羧化能力与土壤-植物水力性状之间的关系. ...
... 不同环境条件下同一物种或不同物种的叶片导水率与最大光合速率密切相关(Zhang & Cao, 2009), 表明水力特性对气孔导度和光合气体交换起限制作用.Scoffoni等(2015)研究不同光强下叶片水力和气体交换、解剖、结构特性等的关系, 发现这些特性均有可塑性, 随着光照环境的不同而协同变化以适应不同光照环境, 达到最优生长(Scoffoni et al., 2015).在水分状况不同的地区, 植物叶片水分运输效率与光合能力之间也存在协调作用(Brodribb & Feild, 2000; Brodribb et al., 2002; Santiago et al., 2004).湿润地区的植物为了快速进行光合作用而有较高的Kleaf, 从而使植物在有较大气体交换速率的同时, 无需增加叶片致命脱水的风险.因此, 高Kleaf植物在湿润地区具有较大的竞争优势(Meinzer, 2002; Brodribb et al., 2007).然而, 干旱地区植物的首要任务是维持叶片水力系统的功能, 其次是具有较高的气体交换速率. ...
... 植物为了容忍叶片机械破坏和导管栓塞, 需要构建稠密的叶脉冗余网络, 以提供额外的水流路径, 但同时也增加了巨大的碳投资(Corson, 2010; Scoffoni et al., 2011; Nardini et al., 2012a; Pantin et al., 2012).近年来, 叶脉功能性状与叶片水力、光合能力、叶片碳构建等关系受到广泛的关注(Lamont & Lamont, 2000; Beerling & Franks, 2010; Blonder et al., 2011; Sack et al., 2012, 2013; Zhang et al., 2012).Brodribb等(2007)提出较高的叶片导水率和光合能力的耦合与叶脉密度有关.Nardini等(2012a)指出, Kleaf_mass和P50与叶脉密度和LMA均存在负相关关系, 表明植物通过提高叶脉密度, 增加其抗旱阻力, 但同时也增加了单位叶面积的碳构建成本(高LMA) (Blonder et al., 2011).Prado和Maurel (2013)的叶片水力模型表明叶片导水率和不同等级的叶脉密度共同促成最优化的木质部网络水力效率.叶片主脉密度对Kleaf的影响微小(Sack & Frole, 2006), 而细脉密度对Kleaf的影响较大.这是因为细脉密度的增加, 不仅增加了叶片木质部的导水率(Cochard et al., 2004), 同时也增加了木质部与周围叶肉组织接触的表面积, 降低了木质部外水流路径的长度(Roth-Nebelsick et al., 2001; Sack & Frole, 2006).由于低等级叶脉具有较高的导水率, 因此, 与碳资源投资相比, 较高的叶脉密度在经济上是可行的. ...
... Sack等(2013)基于大范围物种研究提出了“flux trait network hypothesis”假说, 阐明了叶脉性状对水力性状和气体交换性状的影响机制: 即叶脉密度通过Kleaf对气孔导度和基于叶面积表达的光合速率(Aarea)产生影响, 进而影响基于叶干质量表达的光合速率(Amass)和相对生长速率(Lamont & Lamont, 2000; Brodribb et al., 2007; Boyce et al., 2009; Brodribb et al., 2010; McKown et al., 2010; Walls, 2011).尽管该假说不一定适用于所有物种, 但它阐明了叶脉、水力、气孔和气体交换性状之间的关系.从某种意义上讲, 叶脉密度反映了资源流通能力(Sack et al., 2013).因此, 叶脉密度对叶片碳、水通量的影响可促成“快-慢”经济型物种.叶脉系统的进化, 也支持了高水分流通速率和其他快速叶片经济型谱(Lamont & Lamont, 2000; McDowell et al., 2008; Boyce et al., 2009; Sack & Scoffoni, 2013).因此, 该假说在预测植物地理分布和对气候变化的响应上有潜在的应用前景. ...
Viewing leaf structure and evolution from a hydraulic perspective.
2
2010
... LES (Wright et al., 2004)认为, 植物的化学、结构和生理功能性状呈一条连续变化的带谱, 在带谱的两端分别是“快速投资-收益”型和“缓慢投资-收益”型物种, 其间是两种类型的过渡梯度类型.叶片水力性状影响叶片碳同化能力、水分利用效率和植物生长(Sack & Holbrook, 2006), 对植物生产力和进化起着重要的作用(Brodribb et al., 2010).叶片导水率(Kleaf)和叶片水力脆弱性(P50)常用于反映叶片水力效率和水力安全; 两者的权衡关系可限制物种地理分布, 对不同空间尺度的植被组成和分布非常重要(Nardini et al., 2012a).因此, 近期越来越多的研究关注叶片水力性状对叶片经济性状的影响(Sack & Frole, 2006; Sack & Scoffoni, 2013; Sack et al., 2013; 龚容和高琼, 2015).例如, Villagra等(2013)提出LES反映的碳和养分“投资-收益”的平衡与叶片水力性状有关.Kleaf高的物种, 叶片组织密度低、寿命短, 资源获取速率快(Reich, 2014).此外, 由于叶片水平水分流动阻力占整个植株阻力的比例较大(≥30%), 且与叶片气体交换有关, 因此, Kleaf对植物碳经济、水经济及两者之间的耦合关系均至关重要(Brodribb et al., 2005, 2007; Reich, 2014).为此, 本文首先综述植物叶片水力性状、经济性状及两者之间的权衡关系; 然后从结构和功能角度探讨叶片水力性状与经济性状的调节机制; 最后针对植物的水力和经济性状耦合关系提出今后的研究方向. ...
... Sack等(2013)基于大范围物种研究提出了“flux trait network hypothesis”假说, 阐明了叶脉性状对水力性状和气体交换性状的影响机制: 即叶脉密度通过Kleaf对气孔导度和基于叶面积表达的光合速率(Aarea)产生影响, 进而影响基于叶干质量表达的光合速率(Amass)和相对生长速率(Lamont & Lamont, 2000; Brodribb et al., 2007; Boyce et al., 2009; Brodribb et al., 2010; McKown et al., 2010; Walls, 2011).尽管该假说不一定适用于所有物种, 但它阐明了叶脉、水力、气孔和气体交换性状之间的关系.从某种意义上讲, 叶脉密度反映了资源流通能力(Sack et al., 2013).因此, 叶脉密度对叶片碳、水通量的影响可促成“快-慢”经济型物种.叶脉系统的进化, 也支持了高水分流通速率和其他快速叶片经济型谱(Lamont & Lamont, 2000; McDowell et al., 2008; Boyce et al., 2009; Sack & Scoffoni, 2013).因此, 该假说在预测植物地理分布和对气候变化的响应上有潜在的应用前景. ...
Stomatal closure during leaf dehydration, correlation with other leaf physiological traits.
4
2003
... P50值(叶片导水率损失50%对应的叶片水势)是常用于量化叶片水力脆弱性的指标.Kleaf和P50是植物适应不同环境条件的主要生理性状(Sack et al., 2005; Nardini et al., 2012a), 决定植物的竞争能力和生产力(Nardini & Luglio, 2014).一般而言, 植物茎干栓塞阻力(用P50表示)的增加常会引起水力效率(Kstem)的降低(Bucci et al., 2006)或碳消耗的增加(Jacobsen et al., 2007).然而, 由于叶片水平的P50与基于叶面积的叶片导水率(Kleaf_area)缺少相关性, 人们常认为不存在水力效率与安全的权衡(Brodribb & Holbrook, 2003, 2004b; Lo Gullo et al., 2003; Nardini et al., 2003, 2012a; Blackman et al., 2010; Scoffoni et al., 2011).但近期Nardini和Luglio (2014)综合分析了全球130个木本被子植物叶片水力数据发现: P50与Kleaf_area确实没有相关性, 但与基于叶干质量的叶片导水率(Kleaf_mass)存在弱正相关关系, 表明叶片水平可能存在水力安全和水力效率的权衡.Kleaf_mass不仅可以解释叶片水力系统的水分运输效率, 还可估算构建叶片水力系统的碳消耗(Nardini et al., 2012b; Simonin et al., 2012); 抗旱阻力强的叶片(P50低), 其碳消耗较大(Nardini et al., 2012a).至于这种权衡关系是否有普适性, 还需验证. ...
... 叶片失膨点水势或萎蔫水势(Ψtlp)是叶片对干旱胁迫耐性的决定因子, 对植株的生理干旱耐性有重要贡献(Maréchaux et al., 2015).有研究报道Ψtlp与P50、Kleaf呈正相关(Nardini et al., 2012a; Villagra et al., 2013), 表明Ψtlp低(即叶片耐旱性高)的物种, 其叶片组织的供水能力低(Kleaf低), 但抗栓塞阻力强(P50低); 相反, 水分运输效率高(Kleaf高)的物种, 能够保持叶片良好的水分状况, 阻止叶片水势的下降, 从而保持气孔开放和碳同化时间长.P50与Ψtlp的协调关系不仅解释了叶片木质部外导水率下降的原因(Brodribb & Holbrook, 2004b; Knipfer & Steudle, 2008), 还表明了叶片内部非原生质体和共质体之间抗旱阻力的协调关系(Blackman et al., 2010; Vilagrosa et al., 2010; Nardini & Luglio, 2014).此外, Brodribb和Holbrook (2003)的研究表明P50和Ψtlp影响水分胁迫期间的气孔关闭时间.在空穴化发生早期, 首先引起叶片气孔关闭(Choat et al., 2012), 在叶片膨胀体积逐渐损失直至完全丧失之前, 逐渐降低叶片蒸腾速率(Blackman et al., 2010; Scoffoni et al., 2011; Nardini et al., 2012a).因此, 叶片是保护活细胞远离临界水分状况的安全阀门. ...
... 叶片水力安全阈值(safety margin)是指叶片气孔关闭所对应的叶片水势(Ψmin)与P50的差值(Brodribb & Holbrook, 2004b; Sperry, 2004), 用于表征植物的水力对策(Apgaua et al., 2015).安全阈值较大的物种, 当Kleaf显著下降时会关闭气孔, 保护木质部安全, 从而最大限度地降低栓塞风险(Cochard, 2002; Cochard et al., 2002; Nardini et al., 2003; Brodribb & Holbrook, 2004b); 相反, 安全阈值较小的物种, 当Kleaf下降时, 气孔并不完全关闭, 从而使植物能够长时间地维持气体交换(Salleo et al., 2000, 2001; Brodribb & Holbrook, 2003, 2004a, 2004b; Bucci et al., 2003; Lo Gullo et al., 2003), 直至Kleaf下降到空穴化不可逆转的水平(Sperry et al., 2002; Brodribb & Holbrook, 2004b).通常, 对空穴化更脆弱的物种, 其水力安全阈值较小, 在非干旱条件下也有可能经历栓塞(Meinzer et al., 2009), 但其修复栓塞的能力较大(Johnson et al., 2009, 2011; Choat et al., 2012; Ambrose et al., 2015), 在修复栓塞的同时也会引起巨大的代谢消耗(Nardini et al., 2011).纵观森林群落中的各种物种, 虽然对干旱发生的响应差异较大, 但总体上被子植物的安全阈值范围很小, 常不能随着水分可利用性的巨大差异而发生变化, 因而在旱季有木质部栓塞的风险(Choat et al., 2012). ...
... 植物个体水平的气体交换速率和生产力依赖于叶片水平气孔对碳获取和水分损失的调节(Tombesi et al., 2014).在干旱胁迫条件下, 不同物种维持正碳平衡的能力在很大程度上取决于叶片水势下降时的气孔响应(Chaves et al., 2010).植物通过是否需要牺牲光合碳获取与防止叶片功能丧失的权衡, 决定是否需要关闭气孔(Brodribb & Holbrook, 2003).为此, 研究者常基于水分胁迫条件下的气孔动力学机制, 把植物分为等水势调节(isohydric)和非等水势调节(anisohydric)两大类(McDowell et al., 2008; Sade et al., 2012).等水势植物通过严格的气孔控制, 以保持相对恒定的中午叶片水势, 免受外界环境条件的限制.等水势调节植物因气孔关闭而有利于减少水分的损失, 但同时也降低了光合速率: 一是植物的光合作用受到限制, 植物对碳水化合物的需求得不到满足; 二是气孔的关闭导致光合速率的下降, 从而植物吸收的光能将大幅度地超过光合作用的需要, 过剩的光能将诱导大量活性氧化物的产生, 结果引起叶片衰老脱落.相反, 非等水势植物采用冒险的水力对策, 在水分亏缺的情况下以牺牲稳定的叶片水势为代价, 保持气孔开放, 维持光合同化和生长.与等水势调节植物相比, 非等水势调节植物木质部对较负的水势有更强的抵抗力, 适宜占据更为干旱的栖息地(McDowell et al., 2008). ...
a). Diurnal depression of leaf hydraulic conductance in a tropical tree species.
1
2004
... 叶片水力安全阈值(safety margin)是指叶片气孔关闭所对应的叶片水势(Ψmin)与P50的差值(Brodribb & Holbrook, 2004b; Sperry, 2004), 用于表征植物的水力对策(Apgaua et al., 2015).安全阈值较大的物种, 当Kleaf显著下降时会关闭气孔, 保护木质部安全, 从而最大限度地降低栓塞风险(Cochard, 2002; Cochard et al., 2002; Nardini et al., 2003; Brodribb & Holbrook, 2004b); 相反, 安全阈值较小的物种, 当Kleaf下降时, 气孔并不完全关闭, 从而使植物能够长时间地维持气体交换(Salleo et al., 2000, 2001; Brodribb & Holbrook, 2003, 2004a, 2004b; Bucci et al., 2003; Lo Gullo et al., 2003), 直至Kleaf下降到空穴化不可逆转的水平(Sperry et al., 2002; Brodribb & Holbrook, 2004b).通常, 对空穴化更脆弱的物种, 其水力安全阈值较小, 在非干旱条件下也有可能经历栓塞(Meinzer et al., 2009), 但其修复栓塞的能力较大(Johnson et al., 2009, 2011; Choat et al., 2012; Ambrose et al., 2015), 在修复栓塞的同时也会引起巨大的代谢消耗(Nardini et al., 2011).纵观森林群落中的各种物种, 虽然对干旱发生的响应差异较大, 但总体上被子植物的安全阈值范围很小, 常不能随着水分可利用性的巨大差异而发生变化, 因而在旱季有木质部栓塞的风险(Choat et al., 2012). ...
b). Stomatal protection against hydraulic failure: A comparison of coexisting ferns and angiosperms.
7
2004
... P50值(叶片导水率损失50%对应的叶片水势)是常用于量化叶片水力脆弱性的指标.Kleaf和P50是植物适应不同环境条件的主要生理性状(Sack et al., 2005; Nardini et al., 2012a), 决定植物的竞争能力和生产力(Nardini & Luglio, 2014).一般而言, 植物茎干栓塞阻力(用P50表示)的增加常会引起水力效率(Kstem)的降低(Bucci et al., 2006)或碳消耗的增加(Jacobsen et al., 2007).然而, 由于叶片水平的P50与基于叶面积的叶片导水率(Kleaf_area)缺少相关性, 人们常认为不存在水力效率与安全的权衡(Brodribb & Holbrook, 2003, 2004b; Lo Gullo et al., 2003; Nardini et al., 2003, 2012a; Blackman et al., 2010; Scoffoni et al., 2011).但近期Nardini和Luglio (2014)综合分析了全球130个木本被子植物叶片水力数据发现: P50与Kleaf_area确实没有相关性, 但与基于叶干质量的叶片导水率(Kleaf_mass)存在弱正相关关系, 表明叶片水平可能存在水力安全和水力效率的权衡.Kleaf_mass不仅可以解释叶片水力系统的水分运输效率, 还可估算构建叶片水力系统的碳消耗(Nardini et al., 2012b; Simonin et al., 2012); 抗旱阻力强的叶片(P50低), 其碳消耗较大(Nardini et al., 2012a).至于这种权衡关系是否有普适性, 还需验证. ...
... 叶片失膨点水势或萎蔫水势(Ψtlp)是叶片对干旱胁迫耐性的决定因子, 对植株的生理干旱耐性有重要贡献(Maréchaux et al., 2015).有研究报道Ψtlp与P50、Kleaf呈正相关(Nardini et al., 2012a; Villagra et al., 2013), 表明Ψtlp低(即叶片耐旱性高)的物种, 其叶片组织的供水能力低(Kleaf低), 但抗栓塞阻力强(P50低); 相反, 水分运输效率高(Kleaf高)的物种, 能够保持叶片良好的水分状况, 阻止叶片水势的下降, 从而保持气孔开放和碳同化时间长.P50与Ψtlp的协调关系不仅解释了叶片木质部外导水率下降的原因(Brodribb & Holbrook, 2004b; Knipfer & Steudle, 2008), 还表明了叶片内部非原生质体和共质体之间抗旱阻力的协调关系(Blackman et al., 2010; Vilagrosa et al., 2010; Nardini & Luglio, 2014).此外, Brodribb和Holbrook (2003)的研究表明P50和Ψtlp影响水分胁迫期间的气孔关闭时间.在空穴化发生早期, 首先引起叶片气孔关闭(Choat et al., 2012), 在叶片膨胀体积逐渐损失直至完全丧失之前, 逐渐降低叶片蒸腾速率(Blackman et al., 2010; Scoffoni et al., 2011; Nardini et al., 2012a).因此, 叶片是保护活细胞远离临界水分状况的安全阀门. ...
... 叶片水力安全阈值(safety margin)是指叶片气孔关闭所对应的叶片水势(Ψmin)与P50的差值(Brodribb & Holbrook, 2004b; Sperry, 2004), 用于表征植物的水力对策(Apgaua et al., 2015).安全阈值较大的物种, 当Kleaf显著下降时会关闭气孔, 保护木质部安全, 从而最大限度地降低栓塞风险(Cochard, 2002; Cochard et al., 2002; Nardini et al., 2003; Brodribb & Holbrook, 2004b); 相反, 安全阈值较小的物种, 当Kleaf下降时, 气孔并不完全关闭, 从而使植物能够长时间地维持气体交换(Salleo et al., 2000, 2001; Brodribb & Holbrook, 2003, 2004a, 2004b; Bucci et al., 2003; Lo Gullo et al., 2003), 直至Kleaf下降到空穴化不可逆转的水平(Sperry et al., 2002; Brodribb & Holbrook, 2004b).通常, 对空穴化更脆弱的物种, 其水力安全阈值较小, 在非干旱条件下也有可能经历栓塞(Meinzer et al., 2009), 但其修复栓塞的能力较大(Johnson et al., 2009, 2011; Choat et al., 2012; Ambrose et al., 2015), 在修复栓塞的同时也会引起巨大的代谢消耗(Nardini et al., 2011).纵观森林群落中的各种物种, 虽然对干旱发生的响应差异较大, 但总体上被子植物的安全阈值范围很小, 常不能随着水分可利用性的巨大差异而发生变化, 因而在旱季有木质部栓塞的风险(Choat et al., 2012). ...
... ; Brodribb & Holbrook, 2004b); 相反, 安全阈值较小的物种, 当Kleaf下降时, 气孔并不完全关闭, 从而使植物能够长时间地维持气体交换(Salleo et al., 2000, 2001; Brodribb & Holbrook, 2003, 2004a, 2004b; Bucci et al., 2003; Lo Gullo et al., 2003), 直至Kleaf下降到空穴化不可逆转的水平(Sperry et al., 2002; Brodribb & Holbrook, 2004b).通常, 对空穴化更脆弱的物种, 其水力安全阈值较小, 在非干旱条件下也有可能经历栓塞(Meinzer et al., 2009), 但其修复栓塞的能力较大(Johnson et al., 2009, 2011; Choat et al., 2012; Ambrose et al., 2015), 在修复栓塞的同时也会引起巨大的代谢消耗(Nardini et al., 2011).纵观森林群落中的各种物种, 虽然对干旱发生的响应差异较大, 但总体上被子植物的安全阈值范围很小, 常不能随着水分可利用性的巨大差异而发生变化, 因而在旱季有木质部栓塞的风险(Choat et al., 2012). ...
... , 2004b; Bucci et al., 2003; Lo Gullo et al., 2003), 直至Kleaf下降到空穴化不可逆转的水平(Sperry et al., 2002; Brodribb & Holbrook, 2004b).通常, 对空穴化更脆弱的物种, 其水力安全阈值较小, 在非干旱条件下也有可能经历栓塞(Meinzer et al., 2009), 但其修复栓塞的能力较大(Johnson et al., 2009, 2011; Choat et al., 2012; Ambrose et al., 2015), 在修复栓塞的同时也会引起巨大的代谢消耗(Nardini et al., 2011).纵观森林群落中的各种物种, 虽然对干旱发生的响应差异较大, 但总体上被子植物的安全阈值范围很小, 常不能随着水分可利用性的巨大差异而发生变化, 因而在旱季有木质部栓塞的风险(Choat et al., 2012). ...
... ; Brodribb & Holbrook, 2004b).通常, 对空穴化更脆弱的物种, 其水力安全阈值较小, 在非干旱条件下也有可能经历栓塞(Meinzer et al., 2009), 但其修复栓塞的能力较大(Johnson et al., 2009, 2011; Choat et al., 2012; Ambrose et al., 2015), 在修复栓塞的同时也会引起巨大的代谢消耗(Nardini et al., 2011).纵观森林群落中的各种物种, 虽然对干旱发生的响应差异较大, 但总体上被子植物的安全阈值范围很小, 常不能随着水分可利用性的巨大差异而发生变化, 因而在旱季有木质部栓塞的风险(Choat et al., 2012). ...
... 植物一方面要最大限度地吸收CO2进行光合作用, 另一方面要最小限度地控制水分散失, 这两个生理过程共享同一个气孔路径(Xiong et al., 2015).因此, 植物通过气孔吸收CO2的气孔导度(Gs, 气相水分散失能力指标)与释放水蒸气的水力导度(Kleaf, 液相水分传输能力指标)之间存在权衡机制(Sack & Holbrook, 2006).由于C3植物的Gs与光合速率的耦合关系, 因此Kleaf与光合速率也存在正相关关系(Brodribb et al., 2005, 2007).对系统发生、叶片结构、叶片寿命、地理分布等特性差异较大的大量物种的综合研究发现, Kleaf与Gs和光合速率呈正相关关系(Brodribb et al., 2005, 2007), 表明水力供需能力及碳获取与水分运输能力之间存在平衡关系(Brodribb & Feild, 2000; Meinzer, 2002; Sack et al., 2003, 2005; Brodribb & Holbrook, 2004b; Brodribb et al., 2005; Campanello et al., 2008; Prentice et al., 2014), 也反映叶片水平的气液两相导度的协调性; 有效的叶片水分供应能够最大限度地维持气孔开放(Brodribb et al., 2005, 2007; Lo Gullo et al., 2005; Nardini et al., 2005; Manzoni et al., 2013).Katul等(2003)提出了一个把Gs、光合速率和水分运输效率联系在一起的理论框架模型, 可以直接预测最大羧化能力与土壤-植物水力性状之间的关系. ...
Hydraulic and photosynthetic co-ordination in seasonally dry tropical forest trees.
1
2002
... 不同环境条件下同一物种或不同物种的叶片导水率与最大光合速率密切相关(Zhang & Cao, 2009), 表明水力特性对气孔导度和光合气体交换起限制作用.Scoffoni等(2015)研究不同光强下叶片水力和气体交换、解剖、结构特性等的关系, 发现这些特性均有可塑性, 随着光照环境的不同而协同变化以适应不同光照环境, 达到最优生长(Scoffoni et al., 2015).在水分状况不同的地区, 植物叶片水分运输效率与光合能力之间也存在协调作用(Brodribb & Feild, 2000; Brodribb et al., 2002; Santiago et al., 2004).湿润地区的植物为了快速进行光合作用而有较高的Kleaf, 从而使植物在有较大气体交换速率的同时, 无需增加叶片致命脱水的风险.因此, 高Kleaf植物在湿润地区具有较大的竞争优势(Meinzer, 2002; Brodribb et al., 2007).然而, 干旱地区植物的首要任务是维持叶片水力系统的功能, 其次是具有较高的气体交换速率. ...
Leaf hydraulic capacity in ferns, conifers and angiosperms: Impacts on photosynthetic maxima.
6
2005
... LES (Wright et al., 2004)认为, 植物的化学、结构和生理功能性状呈一条连续变化的带谱, 在带谱的两端分别是“快速投资-收益”型和“缓慢投资-收益”型物种, 其间是两种类型的过渡梯度类型.叶片水力性状影响叶片碳同化能力、水分利用效率和植物生长(Sack & Holbrook, 2006), 对植物生产力和进化起着重要的作用(Brodribb et al., 2010).叶片导水率(Kleaf)和叶片水力脆弱性(P50)常用于反映叶片水力效率和水力安全; 两者的权衡关系可限制物种地理分布, 对不同空间尺度的植被组成和分布非常重要(Nardini et al., 2012a).因此, 近期越来越多的研究关注叶片水力性状对叶片经济性状的影响(Sack & Frole, 2006; Sack & Scoffoni, 2013; Sack et al., 2013; 龚容和高琼, 2015).例如, Villagra等(2013)提出LES反映的碳和养分“投资-收益”的平衡与叶片水力性状有关.Kleaf高的物种, 叶片组织密度低、寿命短, 资源获取速率快(Reich, 2014).此外, 由于叶片水平水分流动阻力占整个植株阻力的比例较大(≥30%), 且与叶片气体交换有关, 因此, Kleaf对植物碳经济、水经济及两者之间的耦合关系均至关重要(Brodribb et al., 2005, 2007; Reich, 2014).为此, 本文首先综述植物叶片水力性状、经济性状及两者之间的权衡关系; 然后从结构和功能角度探讨叶片水力性状与经济性状的调节机制; 最后针对植物的水力和经济性状耦合关系提出今后的研究方向. ...
... 许多研究发现, LL和LMA与Kleaf_mass负相关, 而与Kleaf_area无关(Nardini, 2001; Wright et al., 2001; Shipley, 2002; Sack et al., 2003, 2005; Brodribb et al., 2005; Blackman et al., 2010; Nardini et al., 2012a; Simonin et al., 2012; Bucci et al., 2013).原因可能是Kleaf_area更偏向于发生在叶片-大气界面之间的叶片表面的过程, 而Kleaf_mass是基于整个叶片组织的过程(Nardini et al., 2012a).Nardini等(2012b)研究发现 Quercus ilex阳生叶的Kleaf_mass比阴生叶高40%, 而Kleaf_area是阴生叶的2倍多; 这表明同一冠层叶片对不同光强度的水力驯化增加了单位叶面积的水分运输效率, 但叶片水分运输系统的碳消耗保持相对稳定.近期, Maréchaux等(2015)研究了法属圭亚那地区(French Guiana) 71个热带树种165株树木的Ψtlp与叶片经济性状的关系, 发现Ψtlp与叶片硬度和厚度呈弱正相关关系, 但与LMA、叶面积、叶片氮浓度、叶片碳浓度等性状没有相关性.可见, 叶片水力性状与经济性状之间是否有普适性关系还需要更多的研究加以验证. ...
... 植物一方面要最大限度地吸收CO2进行光合作用, 另一方面要最小限度地控制水分散失, 这两个生理过程共享同一个气孔路径(Xiong et al., 2015).因此, 植物通过气孔吸收CO2的气孔导度(Gs, 气相水分散失能力指标)与释放水蒸气的水力导度(Kleaf, 液相水分传输能力指标)之间存在权衡机制(Sack & Holbrook, 2006).由于C3植物的Gs与光合速率的耦合关系, 因此Kleaf与光合速率也存在正相关关系(Brodribb et al., 2005, 2007).对系统发生、叶片结构、叶片寿命、地理分布等特性差异较大的大量物种的综合研究发现, Kleaf与Gs和光合速率呈正相关关系(Brodribb et al., 2005, 2007), 表明水力供需能力及碳获取与水分运输能力之间存在平衡关系(Brodribb & Feild, 2000; Meinzer, 2002; Sack et al., 2003, 2005; Brodribb & Holbrook, 2004b; Brodribb et al., 2005; Campanello et al., 2008; Prentice et al., 2014), 也反映叶片水平的气液两相导度的协调性; 有效的叶片水分供应能够最大限度地维持气孔开放(Brodribb et al., 2005, 2007; Lo Gullo et al., 2005; Nardini et al., 2005; Manzoni et al., 2013).Katul等(2003)提出了一个把Gs、光合速率和水分运输效率联系在一起的理论框架模型, 可以直接预测最大羧化能力与土壤-植物水力性状之间的关系. ...
... 和光合速率呈正相关关系(Brodribb et al., 2005, 2007), 表明水力供需能力及碳获取与水分运输能力之间存在平衡关系(Brodribb & Feild, 2000; Meinzer, 2002; Sack et al., 2003, 2005; Brodribb & Holbrook, 2004b; Brodribb et al., 2005; Campanello et al., 2008; Prentice et al., 2014), 也反映叶片水平的气液两相导度的协调性; 有效的叶片水分供应能够最大限度地维持气孔开放(Brodribb et al., 2005, 2007; Lo Gullo et al., 2005; Nardini et al., 2005; Manzoni et al., 2013).Katul等(2003)提出了一个把Gs、光合速率和水分运输效率联系在一起的理论框架模型, 可以直接预测最大羧化能力与土壤-植物水力性状之间的关系. ...
... ; Brodribb et al., 2005; Campanello et al., 2008; Prentice et al., 2014), 也反映叶片水平的气液两相导度的协调性; 有效的叶片水分供应能够最大限度地维持气孔开放(Brodribb et al., 2005, 2007; Lo Gullo et al., 2005; Nardini et al., 2005; Manzoni et al., 2013).Katul等(2003)提出了一个把Gs、光合速率和水分运输效率联系在一起的理论框架模型, 可以直接预测最大羧化能力与土壤-植物水力性状之间的关系. ...
... ), 也反映叶片水平的气液两相导度的协调性; 有效的叶片水分供应能够最大限度地维持气孔开放(Brodribb et al., 2005, 2007; Lo Gullo et al., 2005; Nardini et al., 2005; Manzoni et al., 2013).Katul等(2003)提出了一个把Gs、光合速率和水分运输效率联系在一起的理论框架模型, 可以直接预测最大羧化能力与土壤-植物水力性状之间的关系. ...
Water supply and demand remain balanced during leaf acclimation of Nothofagus cunninghamii trees.
Hydraulic differences along the water transport system of South American Nothofagus species: Do leaves protect the stem functionality?
1
2012
... Zimmermann (1983)首次提出了水力分割假说(HSH), 并得到许多研究的证实(Hao et al., 2008; Chen et al., 2009; Johnson et al., 2011; Bucci et al., 2012; Pivovaroff et al., 2014).HSH假说认为, 末端器官(根和叶)比主干或茎的导水率低或对空穴化更脆弱(P50高), 这种解耦现象是以牺牲寿命短、相对投资少且容易被替代的末端器官为代价, 以维持寿命长、碳投资大的器官(主干或茎)的水分状况.由于末端器官水力阻力(水力效率的倒数)较大, 在整株水分流动上起到瓶颈作用(Sack & Holbrook, 2006).因此, HSH假说可以解释木本植物体内水力阻力的分布.然而, Villagra等(2013)在亚热带森林的研究表明, 该地区的物种叶片和茎对空穴化的脆弱性相似, 并不存在分割作用; 这可能是由于该地区雨水的季节性变化不大, 没有产生增加叶片和茎之间水力分割的选择性压力.迄今为止, 尽管根、茎、叶水力性状的单独研究不少, 但相互之间水分传输交互作用的研究还刚起步; 对根、茎、叶木质部的栓塞关系以及植株各部分间水分传输的协同关系还需要深入研究. ...
Nutrient availability constrains the hydraulic architecture and water relations of savannah trees.
1
2006
... P50值(叶片导水率损失50%对应的叶片水势)是常用于量化叶片水力脆弱性的指标.Kleaf和P50是植物适应不同环境条件的主要生理性状(Sack et al., 2005; Nardini et al., 2012a), 决定植物的竞争能力和生产力(Nardini & Luglio, 2014).一般而言, 植物茎干栓塞阻力(用P50表示)的增加常会引起水力效率(Kstem)的降低(Bucci et al., 2006)或碳消耗的增加(Jacobsen et al., 2007).然而, 由于叶片水平的P50与基于叶面积的叶片导水率(Kleaf_area)缺少相关性, 人们常认为不存在水力效率与安全的权衡(Brodribb & Holbrook, 2003, 2004b; Lo Gullo et al., 2003; Nardini et al., 2003, 2012a; Blackman et al., 2010; Scoffoni et al., 2011).但近期Nardini和Luglio (2014)综合分析了全球130个木本被子植物叶片水力数据发现: P50与Kleaf_area确实没有相关性, 但与基于叶干质量的叶片导水率(Kleaf_mass)存在弱正相关关系, 表明叶片水平可能存在水力安全和水力效率的权衡.Kleaf_mass不仅可以解释叶片水力系统的水分运输效率, 还可估算构建叶片水力系统的碳消耗(Nardini et al., 2012b; Simonin et al., 2012); 抗旱阻力强的叶片(P50低), 其碳消耗较大(Nardini et al., 2012a).至于这种权衡关系是否有普适性, 还需验证. ...
Dynamic changes in hydraulic conductivity in petioles of two savanna tree species: Factors and mechanisms contributing to the refilling of embolized vessels.
1
2003
... 叶片水力安全阈值(safety margin)是指叶片气孔关闭所对应的叶片水势(Ψmin)与P50的差值(Brodribb & Holbrook, 2004b; Sperry, 2004), 用于表征植物的水力对策(Apgaua et al., 2015).安全阈值较大的物种, 当Kleaf显著下降时会关闭气孔, 保护木质部安全, 从而最大限度地降低栓塞风险(Cochard, 2002; Cochard et al., 2002; Nardini et al., 2003; Brodribb & Holbrook, 2004b); 相反, 安全阈值较小的物种, 当Kleaf下降时, 气孔并不完全关闭, 从而使植物能够长时间地维持气体交换(Salleo et al., 2000, 2001; Brodribb & Holbrook, 2003, 2004a, 2004b; Bucci et al., 2003; Lo Gullo et al., 2003), 直至Kleaf下降到空穴化不可逆转的水平(Sperry et al., 2002; Brodribb & Holbrook, 2004b).通常, 对空穴化更脆弱的物种, 其水力安全阈值较小, 在非干旱条件下也有可能经历栓塞(Meinzer et al., 2009), 但其修复栓塞的能力较大(Johnson et al., 2009, 2011; Choat et al., 2012; Ambrose et al., 2015), 在修复栓塞的同时也会引起巨大的代谢消耗(Nardini et al., 2011).纵观森林群落中的各种物种, 虽然对干旱发生的响应差异较大, 但总体上被子植物的安全阈值范围很小, 常不能随着水分可利用性的巨大差异而发生变化, 因而在旱季有木质部栓塞的风险(Choat et al., 2012). ...
The stem xylem of Patagonian shrubs operates far from the point of catastrophic dysfunction and is additionally protected from drought-induced embolism by leaves and roots.
1
2013
... 许多研究发现, LL和LMA与Kleaf_mass负相关, 而与Kleaf_area无关(Nardini, 2001; Wright et al., 2001; Shipley, 2002; Sack et al., 2003, 2005; Brodribb et al., 2005; Blackman et al., 2010; Nardini et al., 2012a; Simonin et al., 2012; Bucci et al., 2013).原因可能是Kleaf_area更偏向于发生在叶片-大气界面之间的叶片表面的过程, 而Kleaf_mass是基于整个叶片组织的过程(Nardini et al., 2012a).Nardini等(2012b)研究发现 Quercus ilex阳生叶的Kleaf_mass比阴生叶高40%, 而Kleaf_area是阴生叶的2倍多; 这表明同一冠层叶片对不同光强度的水力驯化增加了单位叶面积的水分运输效率, 但叶片水分运输系统的碳消耗保持相对稳定.近期, Maréchaux等(2015)研究了法属圭亚那地区(French Guiana) 71个热带树种165株树木的Ψtlp与叶片经济性状的关系, 发现Ψtlp与叶片硬度和厚度呈弱正相关关系, 但与LMA、叶面积、叶片氮浓度、叶片碳浓度等性状没有相关性.可见, 叶片水力性状与经济性状之间是否有普适性关系还需要更多的研究加以验证. ...
Coordination between water-transport efficiency and photosynthetic capacity in canopy tree species at different growth irradiances.
1
2008
... 植物一方面要最大限度地吸收CO2进行光合作用, 另一方面要最小限度地控制水分散失, 这两个生理过程共享同一个气孔路径(Xiong et al., 2015).因此, 植物通过气孔吸收CO2的气孔导度(Gs, 气相水分散失能力指标)与释放水蒸气的水力导度(Kleaf, 液相水分传输能力指标)之间存在权衡机制(Sack & Holbrook, 2006).由于C3植物的Gs与光合速率的耦合关系, 因此Kleaf与光合速率也存在正相关关系(Brodribb et al., 2005, 2007).对系统发生、叶片结构、叶片寿命、地理分布等特性差异较大的大量物种的综合研究发现, Kleaf与Gs和光合速率呈正相关关系(Brodribb et al., 2005, 2007), 表明水力供需能力及碳获取与水分运输能力之间存在平衡关系(Brodribb & Feild, 2000; Meinzer, 2002; Sack et al., 2003, 2005; Brodribb & Holbrook, 2004b; Brodribb et al., 2005; Campanello et al., 2008; Prentice et al., 2014), 也反映叶片水平的气液两相导度的协调性; 有效的叶片水分供应能够最大限度地维持气孔开放(Brodribb et al., 2005, 2007; Lo Gullo et al., 2005; Nardini et al., 2005; Manzoni et al., 2013).Katul等(2003)提出了一个把Gs、光合速率和水分运输效率联系在一起的理论框架模型, 可以直接预测最大羧化能力与土壤-植物水力性状之间的关系. ...
Grapevine under deficit irrigation: Hints from physiological and molecular data.
1
2010
... 植物个体水平的气体交换速率和生产力依赖于叶片水平气孔对碳获取和水分损失的调节(Tombesi et al., 2014).在干旱胁迫条件下, 不同物种维持正碳平衡的能力在很大程度上取决于叶片水势下降时的气孔响应(Chaves et al., 2010).植物通过是否需要牺牲光合碳获取与防止叶片功能丧失的权衡, 决定是否需要关闭气孔(Brodribb & Holbrook, 2003).为此, 研究者常基于水分胁迫条件下的气孔动力学机制, 把植物分为等水势调节(isohydric)和非等水势调节(anisohydric)两大类(McDowell et al., 2008; Sade et al., 2012).等水势植物通过严格的气孔控制, 以保持相对恒定的中午叶片水势, 免受外界环境条件的限制.等水势调节植物因气孔关闭而有利于减少水分的损失, 但同时也降低了光合速率: 一是植物的光合作用受到限制, 植物对碳水化合物的需求得不到满足; 二是气孔的关闭导致光合速率的下降, 从而植物吸收的光能将大幅度地超过光合作用的需要, 过剩的光能将诱导大量活性氧化物的产生, 结果引起叶片衰老脱落.相反, 非等水势植物采用冒险的水力对策, 在水分亏缺的情况下以牺牲稳定的叶片水势为代价, 保持气孔开放, 维持光合同化和生长.与等水势调节植物相比, 非等水势调节植物木质部对较负的水势有更强的抵抗力, 适宜占据更为干旱的栖息地(McDowell et al., 2008). ...
Independence of stem and leaf hydraulic traits in six Euphorbiaceae tree species with contrasting leaf phenology.
1
2009
... Zimmermann (1983)首次提出了水力分割假说(HSH), 并得到许多研究的证实(Hao et al., 2008; Chen et al., 2009; Johnson et al., 2011; Bucci et al., 2012; Pivovaroff et al., 2014).HSH假说认为, 末端器官(根和叶)比主干或茎的导水率低或对空穴化更脆弱(P50高), 这种解耦现象是以牺牲寿命短、相对投资少且容易被替代的末端器官为代价, 以维持寿命长、碳投资大的器官(主干或茎)的水分状况.由于末端器官水力阻力(水力效率的倒数)较大, 在整株水分流动上起到瓶颈作用(Sack & Holbrook, 2006).因此, HSH假说可以解释木本植物体内水力阻力的分布.然而, Villagra等(2013)在亚热带森林的研究表明, 该地区的物种叶片和茎对空穴化的脆弱性相似, 并不存在分割作用; 这可能是由于该地区雨水的季节性变化不大, 没有产生增加叶片和茎之间水力分割的选择性压力.迄今为止, 尽管根、茎、叶水力性状的单独研究不少, 但相互之间水分传输交互作用的研究还刚起步; 对根、茎、叶木质部的栓塞关系以及植株各部分间水分传输的协同关系还需要深入研究. ...
... 由此可见, 植物叶片经济性状呈现出一种普适的快-慢型谱(Wright et al., 2004; Reich, 2014).在谱的一端为“快速投资-收益”型物种, 它们具有光合速率大、呼吸速率快、养分含量高、但寿命短、比叶重小、碳和养分归还快等特点(Wright et al., 2004; Gagliardi et al., 2015); 这种类型的植物因其养分投资高且回收快而支持了高的生长速率和叶片周转速率(Mason & Donovan, 2015), 从而适宜生长在资源丰富的地区.相反, 在谱的另一端为“缓慢投资-收益”型物种, 它们具有叶寿命长、叶片坚硬的特点, 用于叶结构建成的资源较多, 因而减少对光合和呼吸功能的资源投入, 从而使光合速率和呼吸速率都偏低(Wright et al., 2005; Ordoñez et al., 2009; Freschet et al., 2010; Osnas et al., 2013); 这种类型的植物由于养分投资低且回收慢而使其生长速率和叶片周转速率均较慢(Kleyer & Minden, 2015; Mason & Donovan, 2015), 从而适宜生长在低资源环境的地区.自然群落中的植物种均处于“快速投资-收益”型和“缓慢投资-收益”型之间的连续体中. ...
Comparative studies of leaf form: Assessing the relative roles of selective pressures and phylogenetic constraints.
2
1987
... Kleaf与叶片大小(周长或面积)的关系受叶片结构和功能协调性的影响(Sack et al., 2003).从湿润到干旱地区, 叶片逐渐变大(Givnish, 1987), 这可能受叶片主脉密度和抗旱阻力大小的影响(Sack & Scoffoni, 2013).Kleaf与叶片形状均与叶脉性状相关(Dengler & Kang, 2001; Nardini et al., 2001; Sack et al., 2002; Zwieniecki et al., 2002).叶脉像灌溉系统一样为整个叶片提供水分.低等级的主脉(一、二级叶脉)水流阻力较低, 主要起提供水分的功能; 而高等级的细脉水流阻力较大, 主要负责给整个叶片分配水分(Zwieniecki et al., 2002; Sack & Frole, 2006).因此, 叶片越大, 使得部分叶肉组织与细脉接触较少, 导致供水相对较差, 从而降低了Kleaf.由于较大叶片的主脉密度和木质部导管直径较大, 因此, 大叶片在应对干旱胁迫时更为脆弱(即P50大) (Sack & Scoffoni, 2013).也有研究表明, Kleaf与叶片大小无关(Sack et al., 2003, 2012).主要理由是: 较大的叶片有较大的主脉, 包括数量较多且直径较大的导管, 使得单位叶片面积的运输能力独立于叶片大小(McKown et al., 2010), 且细脉的直径和细胞大小也与叶片大小无关(Sack et al., 2012).因此, 普适性的水力性状与叶片大小的关系还需验证. ...
... 叶片通过一系列形态-解剖结构调整, 提高植物的抗旱能力, 这是植物生存和竞争的关键(Simonin et al., 2012).在水分胁迫条件下, Kleaf的变化与木质部导管和细胞性状直接相关(Cochard et al., 2004; Nardini et al., 2005).Kleaf与中脉木质部导管的宽度(Aasamaa et al., 2001; Sack et al., 2004)和细胞间隙的比例正相关, 而与叶肉细胞和表皮的体积负相关(Aasamaa et al., 2005; 张志亮等, 2014).导管直径降低会导致叶片水分运输效率下降, 但可通过增加导管数量或叶脉密度来平衡(Givnish, 1987).叶片水力脆弱性(P50)与导管直径有关, 与导管壁厚无关(Nardini et al., 2012a); 而导管直径与壁厚、中脉木质部导管密度呈负相关; 表明直径小且数量多的导管P50较低.因此, 通过调整导管直径和数量可以提高叶片的水力安全(低P50).抗旱植物在提高自身水力安全的同时, 也增加了叶片组织构建上的碳投资, 提高了叶片水分利用的成本(Simonin et al., 2012), 因而, 可能会降低在资源丰富地区的竞争能力(Nardini et al., 2012a).由于叶脉密度、导管大小和数量等随着发育程度的变化, Kleaf在不同生长条件下具有高度的可塑性(Sack & Holbrook, 2006).热带雨林树种的Kleaf与栅栏组织的厚度、栅栏组织/海绵组织的比例有关(Sack & Frole, 2006), 而温带树种的Kleaf则与总的叶片厚度和单位叶片面积的储水能力有关(Sack et al., 2003; Cochard et al., 2004).Kröber等(2015)研究了亚热带地区21个落叶树种和19个常绿树种的叶片形态和生理特性, 结果表明 常绿树种通常有较厚的海绵和栅栏组织.这种水力特性与叶片结构上的相关性可能与结构的协调性(解剖和发育结构)、或与功能协调性有关(为适应某一特定环境而进行的特性的选择) (Sack & Holbrook, 2006).综上所述, 木质部结构的变化为Kleaf与P50之间的关系提供了一种解释. ...
叶片结构的水力学特性对植物生理功能影响的研究进展
1
2015
... LES (Wright et al., 2004)认为, 植物的化学、结构和生理功能性状呈一条连续变化的带谱, 在带谱的两端分别是“快速投资-收益”型和“缓慢投资-收益”型物种, 其间是两种类型的过渡梯度类型.叶片水力性状影响叶片碳同化能力、水分利用效率和植物生长(Sack & Holbrook, 2006), 对植物生产力和进化起着重要的作用(Brodribb et al., 2010).叶片导水率(Kleaf)和叶片水力脆弱性(P50)常用于反映叶片水力效率和水力安全; 两者的权衡关系可限制物种地理分布, 对不同空间尺度的植被组成和分布非常重要(Nardini et al., 2012a).因此, 近期越来越多的研究关注叶片水力性状对叶片经济性状的影响(Sack & Frole, 2006; Sack & Scoffoni, 2013; Sack et al., 2013; 龚容和高琼, 2015).例如, Villagra等(2013)提出LES反映的碳和养分“投资-收益”的平衡与叶片水力性状有关.Kleaf高的物种, 叶片组织密度低、寿命短, 资源获取速率快(Reich, 2014).此外, 由于叶片水平水分流动阻力占整个植株阻力的比例较大(≥30%), 且与叶片气体交换有关, 因此, Kleaf对植物碳经济、水经济及两者之间的耦合关系均至关重要(Brodribb et al., 2005, 2007; Reich, 2014).为此, 本文首先综述植物叶片水力性状、经济性状及两者之间的权衡关系; 然后从结构和功能角度探讨叶片水力性状与经济性状的调节机制; 最后针对植物的水力和经济性状耦合关系提出今后的研究方向. ...
叶片结构的水力学特性对植物生理功能影响的研究进展
1
2015
... LES (Wright et al., 2004)认为, 植物的化学、结构和生理功能性状呈一条连续变化的带谱, 在带谱的两端分别是“快速投资-收益”型和“缓慢投资-收益”型物种, 其间是两种类型的过渡梯度类型.叶片水力性状影响叶片碳同化能力、水分利用效率和植物生长(Sack & Holbrook, 2006), 对植物生产力和进化起着重要的作用(Brodribb et al., 2010).叶片导水率(Kleaf)和叶片水力脆弱性(P50)常用于反映叶片水力效率和水力安全; 两者的权衡关系可限制物种地理分布, 对不同空间尺度的植被组成和分布非常重要(Nardini et al., 2012a).因此, 近期越来越多的研究关注叶片水力性状对叶片经济性状的影响(Sack & Frole, 2006; Sack & Scoffoni, 2013; Sack et al., 2013; 龚容和高琼, 2015).例如, Villagra等(2013)提出LES反映的碳和养分“投资-收益”的平衡与叶片水力性状有关.Kleaf高的物种, 叶片组织密度低、寿命短, 资源获取速率快(Reich, 2014).此外, 由于叶片水平水分流动阻力占整个植株阻力的比例较大(≥30%), 且与叶片气体交换有关, 因此, Kleaf对植物碳经济、水经济及两者之间的耦合关系均至关重要(Brodribb et al., 2005, 2007; Reich, 2014).为此, 本文首先综述植物叶片水力性状、经济性状及两者之间的权衡关系; 然后从结构和功能角度探讨叶片水力性状与经济性状的调节机制; 最后针对植物的水力和经济性状耦合关系提出今后的研究方向. ...
Stem and leaf hydraulics of congeneric tree species from adjacent tropical savanna and forest ecosystems.
2
2008
... 叶片水容(Cleaf)表征植物调节根系供水和叶片蒸腾需水之间的不平衡、缓冲叶片水势波动的能力, 是植物避免干旱的水力对策(Lamont & Lamont, 2000; Sack et al., 2003, 2005).Blackman等(2010)测定了20种温带木本植物的叶片体积水容和叶片组织水容, 结果表明Kleaf与叶片组织水容呈正相关, 但与体积水容无关.这说明并不是所有的叶片储存水都可以补充蒸腾的水分损失, 只有叶片组织中的储存水可以与蒸腾流进行自由交换, 支持了叶片水力分区的观点(Zwieniecki et al., 2007).Kleaf与Cleaf功能上的协调性解释了蒸腾速率一定的情况下Kleaf较高的叶片关闭气孔较慢的原因(Tyree et al., 1998; Aasamaa & Sõber, 2001; Hao et al., 2008). ...
... Zimmermann (1983)首次提出了水力分割假说(HSH), 并得到许多研究的证实(Hao et al., 2008; Chen et al., 2009; Johnson et al., 2011; Bucci et al., 2012; Pivovaroff et al., 2014).HSH假说认为, 末端器官(根和叶)比主干或茎的导水率低或对空穴化更脆弱(P50高), 这种解耦现象是以牺牲寿命短、相对投资少且容易被替代的末端器官为代价, 以维持寿命长、碳投资大的器官(主干或茎)的水分状况.由于末端器官水力阻力(水力效率的倒数)较大, 在整株水分流动上起到瓶颈作用(Sack & Holbrook, 2006).因此, HSH假说可以解释木本植物体内水力阻力的分布.然而, Villagra等(2013)在亚热带森林的研究表明, 该地区的物种叶片和茎对空穴化的脆弱性相似, 并不存在分割作用; 这可能是由于该地区雨水的季节性变化不大, 没有产生增加叶片和茎之间水力分割的选择性压力.迄今为止, 尽管根、茎、叶水力性状的单独研究不少, 但相互之间水分传输交互作用的研究还刚起步; 对根、茎、叶木质部的栓塞关系以及植株各部分间水分传输的协同关系还需要深入研究. ...
Xylem density, biomechanics and anatomical traits correlate with water stress in 17 evergreen shrub species of the Mediterranean-type climate region of South Africa.
1
2007
... P50值(叶片导水率损失50%对应的叶片水势)是常用于量化叶片水力脆弱性的指标.Kleaf和P50是植物适应不同环境条件的主要生理性状(Sack et al., 2005; Nardini et al., 2012a), 决定植物的竞争能力和生产力(Nardini & Luglio, 2014).一般而言, 植物茎干栓塞阻力(用P50表示)的增加常会引起水力效率(Kstem)的降低(Bucci et al., 2006)或碳消耗的增加(Jacobsen et al., 2007).然而, 由于叶片水平的P50与基于叶面积的叶片导水率(Kleaf_area)缺少相关性, 人们常认为不存在水力效率与安全的权衡(Brodribb & Holbrook, 2003, 2004b; Lo Gullo et al., 2003; Nardini et al., 2003, 2012a; Blackman et al., 2010; Scoffoni et al., 2011).但近期Nardini和Luglio (2014)综合分析了全球130个木本被子植物叶片水力数据发现: P50与Kleaf_area确实没有相关性, 但与基于叶干质量的叶片导水率(Kleaf_mass)存在弱正相关关系, 表明叶片水平可能存在水力安全和水力效率的权衡.Kleaf_mass不仅可以解释叶片水力系统的水分运输效率, 还可估算构建叶片水力系统的碳消耗(Nardini et al., 2012b; Simonin et al., 2012); 抗旱阻力强的叶片(P50低), 其碳消耗较大(Nardini et al., 2012a).至于这种权衡关系是否有普适性, 还需验证. ...
Hydraulic patterns and safety margins, from stem to stomata, in three eastern US tree species.
2
2011
... 叶片水力安全阈值(safety margin)是指叶片气孔关闭所对应的叶片水势(Ψmin)与P50的差值(Brodribb & Holbrook, 2004b; Sperry, 2004), 用于表征植物的水力对策(Apgaua et al., 2015).安全阈值较大的物种, 当Kleaf显著下降时会关闭气孔, 保护木质部安全, 从而最大限度地降低栓塞风险(Cochard, 2002; Cochard et al., 2002; Nardini et al., 2003; Brodribb & Holbrook, 2004b); 相反, 安全阈值较小的物种, 当Kleaf下降时, 气孔并不完全关闭, 从而使植物能够长时间地维持气体交换(Salleo et al., 2000, 2001; Brodribb & Holbrook, 2003, 2004a, 2004b; Bucci et al., 2003; Lo Gullo et al., 2003), 直至Kleaf下降到空穴化不可逆转的水平(Sperry et al., 2002; Brodribb & Holbrook, 2004b).通常, 对空穴化更脆弱的物种, 其水力安全阈值较小, 在非干旱条件下也有可能经历栓塞(Meinzer et al., 2009), 但其修复栓塞的能力较大(Johnson et al., 2009, 2011; Choat et al., 2012; Ambrose et al., 2015), 在修复栓塞的同时也会引起巨大的代谢消耗(Nardini et al., 2011).纵观森林群落中的各种物种, 虽然对干旱发生的响应差异较大, 但总体上被子植物的安全阈值范围很小, 常不能随着水分可利用性的巨大差异而发生变化, 因而在旱季有木质部栓塞的风险(Choat et al., 2012). ...
... Zimmermann (1983)首次提出了水力分割假说(HSH), 并得到许多研究的证实(Hao et al., 2008; Chen et al., 2009; Johnson et al., 2011; Bucci et al., 2012; Pivovaroff et al., 2014).HSH假说认为, 末端器官(根和叶)比主干或茎的导水率低或对空穴化更脆弱(P50高), 这种解耦现象是以牺牲寿命短、相对投资少且容易被替代的末端器官为代价, 以维持寿命长、碳投资大的器官(主干或茎)的水分状况.由于末端器官水力阻力(水力效率的倒数)较大, 在整株水分流动上起到瓶颈作用(Sack & Holbrook, 2006).因此, HSH假说可以解释木本植物体内水力阻力的分布.然而, Villagra等(2013)在亚热带森林的研究表明, 该地区的物种叶片和茎对空穴化的脆弱性相似, 并不存在分割作用; 这可能是由于该地区雨水的季节性变化不大, 没有产生增加叶片和茎之间水力分割的选择性压力.迄今为止, 尽管根、茎、叶水力性状的单独研究不少, 但相互之间水分传输交互作用的研究还刚起步; 对根、茎、叶木质部的栓塞关系以及植株各部分间水分传输的协同关系还需要深入研究. ...
Leaf hydraulic conductance, measured in situ, declines and recovers daily: Leaf hydraulics, water potential and stomatal conductance in four temperate and three tropical tree species.
1
2009
... 叶片水力安全阈值(safety margin)是指叶片气孔关闭所对应的叶片水势(Ψmin)与P50的差值(Brodribb & Holbrook, 2004b; Sperry, 2004), 用于表征植物的水力对策(Apgaua et al., 2015).安全阈值较大的物种, 当Kleaf显著下降时会关闭气孔, 保护木质部安全, 从而最大限度地降低栓塞风险(Cochard, 2002; Cochard et al., 2002; Nardini et al., 2003; Brodribb & Holbrook, 2004b); 相反, 安全阈值较小的物种, 当Kleaf下降时, 气孔并不完全关闭, 从而使植物能够长时间地维持气体交换(Salleo et al., 2000, 2001; Brodribb & Holbrook, 2003, 2004a, 2004b; Bucci et al., 2003; Lo Gullo et al., 2003), 直至Kleaf下降到空穴化不可逆转的水平(Sperry et al., 2002; Brodribb & Holbrook, 2004b).通常, 对空穴化更脆弱的物种, 其水力安全阈值较小, 在非干旱条件下也有可能经历栓塞(Meinzer et al., 2009), 但其修复栓塞的能力较大(Johnson et al., 2009, 2011; Choat et al., 2012; Ambrose et al., 2015), 在修复栓塞的同时也会引起巨大的代谢消耗(Nardini et al., 2011).纵观森林群落中的各种物种, 虽然对干旱发生的响应差异较大, 但总体上被子植物的安全阈值范围很小, 常不能随着水分可利用性的巨大差异而发生变化, 因而在旱季有木质部栓塞的风险(Choat et al., 2012). ...
Relationship between plant hydraulic and biochemical properties derived from a steady-state coupled water and carbon transport model.
0
2003
Why functional ecology should consider all plant organs: An allocation-based perspective.
1
2015
... 由此可见, 植物叶片经济性状呈现出一种普适的快-慢型谱(Wright et al., 2004; Reich, 2014).在谱的一端为“快速投资-收益”型物种, 它们具有光合速率大、呼吸速率快、养分含量高、但寿命短、比叶重小、碳和养分归还快等特点(Wright et al., 2004; Gagliardi et al., 2015); 这种类型的植物因其养分投资高且回收快而支持了高的生长速率和叶片周转速率(Mason & Donovan, 2015), 从而适宜生长在资源丰富的地区.相反, 在谱的另一端为“缓慢投资-收益”型物种, 它们具有叶寿命长、叶片坚硬的特点, 用于叶结构建成的资源较多, 因而减少对光合和呼吸功能的资源投入, 从而使光合速率和呼吸速率都偏低(Wright et al., 2005; Ordoñez et al., 2009; Freschet et al., 2010; Osnas et al., 2013); 这种类型的植物由于养分投资低且回收慢而使其生长速率和叶片周转速率均较慢(Kleyer & Minden, 2015; Mason & Donovan, 2015), 从而适宜生长在低资源环境的地区.自然群落中的植物种均处于“快速投资-收益”型和“缓慢投资-收益”型之间的连续体中. ...
Root hydraulic conductivity measured by pressure clamp is substantially affected by internal unstirred layers.
1
2008
... 叶片失膨点水势或萎蔫水势(Ψtlp)是叶片对干旱胁迫耐性的决定因子, 对植株的生理干旱耐性有重要贡献(Maréchaux et al., 2015).有研究报道Ψtlp与P50、Kleaf呈正相关(Nardini et al., 2012a; Villagra et al., 2013), 表明Ψtlp低(即叶片耐旱性高)的物种, 其叶片组织的供水能力低(Kleaf低), 但抗栓塞阻力强(P50低); 相反, 水分运输效率高(Kleaf高)的物种, 能够保持叶片良好的水分状况, 阻止叶片水势的下降, 从而保持气孔开放和碳同化时间长.P50与Ψtlp的协调关系不仅解释了叶片木质部外导水率下降的原因(Brodribb & Holbrook, 2004b; Knipfer & Steudle, 2008), 还表明了叶片内部非原生质体和共质体之间抗旱阻力的协调关系(Blackman et al., 2010; Vilagrosa et al., 2010; Nardini & Luglio, 2014).此外, Brodribb和Holbrook (2003)的研究表明P50和Ψtlp影响水分胁迫期间的气孔关闭时间.在空穴化发生早期, 首先引起叶片气孔关闭(Choat et al., 2012), 在叶片膨胀体积逐渐损失直至完全丧失之前, 逐渐降低叶片蒸腾速率(Blackman et al., 2010; Scoffoni et al., 2011; Nardini et al., 2012a).因此, 叶片是保护活细胞远离临界水分状况的安全阀门. ...
Leaf morphology of 40 evergreen and deciduous broadleaved subtropical tree species and relationships to functional ecophysiolo- gical traits.
2
2015
... Wright等(2004)的研究表明: 比叶重(LMA)与叶寿命(LL)正相关, 说明LMA高的叶片结构上比较坚硬, 不适于食草动物的咀嚼, 因此能够保持较长时间的叶寿命(Kröber et al., 2015).而LMA与基于叶干质量表达的叶光合速率(Amass)、呼吸速率(Rmass)、叶氮含量(Nmass)和磷含量(Pmass)均呈负相关关系.LMA较大的叶片通常质地较厚或较密实, 从气孔到叶绿体的扩散路径较长, 且扩散阻力较大(Zhang et al., 2015), 从而使Amass下降(Sack et al., 2013).由于氮和磷是光合蛋白(羧化酶)的重要组成成分, 因此Nmass和Pmass增加提高了羧化酶的含量和活性、增加了CO2扩散导度, 进而促进了光合作用(Xiong et al., 2015). ...
... 叶片通过一系列形态-解剖结构调整, 提高植物的抗旱能力, 这是植物生存和竞争的关键(Simonin et al., 2012).在水分胁迫条件下, Kleaf的变化与木质部导管和细胞性状直接相关(Cochard et al., 2004; Nardini et al., 2005).Kleaf与中脉木质部导管的宽度(Aasamaa et al., 2001; Sack et al., 2004)和细胞间隙的比例正相关, 而与叶肉细胞和表皮的体积负相关(Aasamaa et al., 2005; 张志亮等, 2014).导管直径降低会导致叶片水分运输效率下降, 但可通过增加导管数量或叶脉密度来平衡(Givnish, 1987).叶片水力脆弱性(P50)与导管直径有关, 与导管壁厚无关(Nardini et al., 2012a); 而导管直径与壁厚、中脉木质部导管密度呈负相关; 表明直径小且数量多的导管P50较低.因此, 通过调整导管直径和数量可以提高叶片的水力安全(低P50).抗旱植物在提高自身水力安全的同时, 也增加了叶片组织构建上的碳投资, 提高了叶片水分利用的成本(Simonin et al., 2012), 因而, 可能会降低在资源丰富地区的竞争能力(Nardini et al., 2012a).由于叶脉密度、导管大小和数量等随着发育程度的变化, Kleaf在不同生长条件下具有高度的可塑性(Sack & Holbrook, 2006).热带雨林树种的Kleaf与栅栏组织的厚度、栅栏组织/海绵组织的比例有关(Sack & Frole, 2006), 而温带树种的Kleaf则与总的叶片厚度和单位叶片面积的储水能力有关(Sack et al., 2003; Cochard et al., 2004).Kröber等(2015)研究了亚热带地区21个落叶树种和19个常绿树种的叶片形态和生理特性, 结果表明 常绿树种通常有较厚的海绵和栅栏组织.这种水力特性与叶片结构上的相关性可能与结构的协调性(解剖和发育结构)、或与功能协调性有关(为适应某一特定环境而进行的特性的选择) (Sack & Holbrook, 2006).综上所述, 木质部结构的变化为Kleaf与P50之间的关系提供了一种解释. ...
Utilizable water in leaves of 8 arid species as derived from pressure-volume curves and chlorophyll fluorescence.
4
2000
... 叶片水容(Cleaf)表征植物调节根系供水和叶片蒸腾需水之间的不平衡、缓冲叶片水势波动的能力, 是植物避免干旱的水力对策(Lamont & Lamont, 2000; Sack et al., 2003, 2005).Blackman等(2010)测定了20种温带木本植物的叶片体积水容和叶片组织水容, 结果表明Kleaf与叶片组织水容呈正相关, 但与体积水容无关.这说明并不是所有的叶片储存水都可以补充蒸腾的水分损失, 只有叶片组织中的储存水可以与蒸腾流进行自由交换, 支持了叶片水力分区的观点(Zwieniecki et al., 2007).Kleaf与Cleaf功能上的协调性解释了蒸腾速率一定的情况下Kleaf较高的叶片关闭气孔较慢的原因(Tyree et al., 1998; Aasamaa & Sõber, 2001; Hao et al., 2008). ...
... 植物为了容忍叶片机械破坏和导管栓塞, 需要构建稠密的叶脉冗余网络, 以提供额外的水流路径, 但同时也增加了巨大的碳投资(Corson, 2010; Scoffoni et al., 2011; Nardini et al., 2012a; Pantin et al., 2012).近年来, 叶脉功能性状与叶片水力、光合能力、叶片碳构建等关系受到广泛的关注(Lamont & Lamont, 2000; Beerling & Franks, 2010; Blonder et al., 2011; Sack et al., 2012, 2013; Zhang et al., 2012).Brodribb等(2007)提出较高的叶片导水率和光合能力的耦合与叶脉密度有关.Nardini等(2012a)指出, Kleaf_mass和P50与叶脉密度和LMA均存在负相关关系, 表明植物通过提高叶脉密度, 增加其抗旱阻力, 但同时也增加了单位叶面积的碳构建成本(高LMA) (Blonder et al., 2011).Prado和Maurel (2013)的叶片水力模型表明叶片导水率和不同等级的叶脉密度共同促成最优化的木质部网络水力效率.叶片主脉密度对Kleaf的影响微小(Sack & Frole, 2006), 而细脉密度对Kleaf的影响较大.这是因为细脉密度的增加, 不仅增加了叶片木质部的导水率(Cochard et al., 2004), 同时也增加了木质部与周围叶肉组织接触的表面积, 降低了木质部外水流路径的长度(Roth-Nebelsick et al., 2001; Sack & Frole, 2006).由于低等级叶脉具有较高的导水率, 因此, 与碳资源投资相比, 较高的叶脉密度在经济上是可行的. ...
... Sack等(2013)基于大范围物种研究提出了“flux trait network hypothesis”假说, 阐明了叶脉性状对水力性状和气体交换性状的影响机制: 即叶脉密度通过Kleaf对气孔导度和基于叶面积表达的光合速率(Aarea)产生影响, 进而影响基于叶干质量表达的光合速率(Amass)和相对生长速率(Lamont & Lamont, 2000; Brodribb et al., 2007; Boyce et al., 2009; Brodribb et al., 2010; McKown et al., 2010; Walls, 2011).尽管该假说不一定适用于所有物种, 但它阐明了叶脉、水力、气孔和气体交换性状之间的关系.从某种意义上讲, 叶脉密度反映了资源流通能力(Sack et al., 2013).因此, 叶脉密度对叶片碳、水通量的影响可促成“快-慢”经济型物种.叶脉系统的进化, 也支持了高水分流通速率和其他快速叶片经济型谱(Lamont & Lamont, 2000; McDowell et al., 2008; Boyce et al., 2009; Sack & Scoffoni, 2013).因此, 该假说在预测植物地理分布和对气候变化的响应上有潜在的应用前景. ...
Photosynthetically relevant foliar traits correlating better on a mass vs an area basis: Of ecophysiological relevance or just a case of mathematical imperatives and statistical quicksand?
0
2013
Changes in leaf hydraulics and stomatal conductance following drought stress and irrigation in Ceratonia siliqua (Carob tree).
2
2003
... P50值(叶片导水率损失50%对应的叶片水势)是常用于量化叶片水力脆弱性的指标.Kleaf和P50是植物适应不同环境条件的主要生理性状(Sack et al., 2005; Nardini et al., 2012a), 决定植物的竞争能力和生产力(Nardini & Luglio, 2014).一般而言, 植物茎干栓塞阻力(用P50表示)的增加常会引起水力效率(Kstem)的降低(Bucci et al., 2006)或碳消耗的增加(Jacobsen et al., 2007).然而, 由于叶片水平的P50与基于叶面积的叶片导水率(Kleaf_area)缺少相关性, 人们常认为不存在水力效率与安全的权衡(Brodribb & Holbrook, 2003, 2004b; Lo Gullo et al., 2003; Nardini et al., 2003, 2012a; Blackman et al., 2010; Scoffoni et al., 2011).但近期Nardini和Luglio (2014)综合分析了全球130个木本被子植物叶片水力数据发现: P50与Kleaf_area确实没有相关性, 但与基于叶干质量的叶片导水率(Kleaf_mass)存在弱正相关关系, 表明叶片水平可能存在水力安全和水力效率的权衡.Kleaf_mass不仅可以解释叶片水力系统的水分运输效率, 还可估算构建叶片水力系统的碳消耗(Nardini et al., 2012b; Simonin et al., 2012); 抗旱阻力强的叶片(P50低), 其碳消耗较大(Nardini et al., 2012a).至于这种权衡关系是否有普适性, 还需验证. ...
... 叶片水力安全阈值(safety margin)是指叶片气孔关闭所对应的叶片水势(Ψmin)与P50的差值(Brodribb & Holbrook, 2004b; Sperry, 2004), 用于表征植物的水力对策(Apgaua et al., 2015).安全阈值较大的物种, 当Kleaf显著下降时会关闭气孔, 保护木质部安全, 从而最大限度地降低栓塞风险(Cochard, 2002; Cochard et al., 2002; Nardini et al., 2003; Brodribb & Holbrook, 2004b); 相反, 安全阈值较小的物种, 当Kleaf下降时, 气孔并不完全关闭, 从而使植物能够长时间地维持气体交换(Salleo et al., 2000, 2001; Brodribb & Holbrook, 2003, 2004a, 2004b; Bucci et al., 2003; Lo Gullo et al., 2003), 直至Kleaf下降到空穴化不可逆转的水平(Sperry et al., 2002; Brodribb & Holbrook, 2004b).通常, 对空穴化更脆弱的物种, 其水力安全阈值较小, 在非干旱条件下也有可能经历栓塞(Meinzer et al., 2009), 但其修复栓塞的能力较大(Johnson et al., 2009, 2011; Choat et al., 2012; Ambrose et al., 2015), 在修复栓塞的同时也会引起巨大的代谢消耗(Nardini et al., 2011).纵观森林群落中的各种物种, 虽然对干旱发生的响应差异较大, 但总体上被子植物的安全阈值范围很小, 常不能随着水分可利用性的巨大差异而发生变化, 因而在旱季有木质部栓塞的风险(Choat et al., 2012). ...
Diurnal and seasonal variations in leaf hydraulic conductance in evergreen and deciduous trees.
1
2005
... 植物一方面要最大限度地吸收CO2进行光合作用, 另一方面要最小限度地控制水分散失, 这两个生理过程共享同一个气孔路径(Xiong et al., 2015).因此, 植物通过气孔吸收CO2的气孔导度(Gs, 气相水分散失能力指标)与释放水蒸气的水力导度(Kleaf, 液相水分传输能力指标)之间存在权衡机制(Sack & Holbrook, 2006).由于C3植物的Gs与光合速率的耦合关系, 因此Kleaf与光合速率也存在正相关关系(Brodribb et al., 2005, 2007).对系统发生、叶片结构、叶片寿命、地理分布等特性差异较大的大量物种的综合研究发现, Kleaf与Gs和光合速率呈正相关关系(Brodribb et al., 2005, 2007), 表明水力供需能力及碳获取与水分运输能力之间存在平衡关系(Brodribb & Feild, 2000; Meinzer, 2002; Sack et al., 2003, 2005; Brodribb & Holbrook, 2004b; Brodribb et al., 2005; Campanello et al., 2008; Prentice et al., 2014), 也反映叶片水平的气液两相导度的协调性; 有效的叶片水分供应能够最大限度地维持气孔开放(Brodribb et al., 2005, 2007; Lo Gullo et al., 2005; Nardini et al., 2005; Manzoni et al., 2013).Katul等(2003)提出了一个把Gs、光合速率和水分运输效率联系在一起的理论框架模型, 可以直接预测最大羧化能力与土壤-植物水力性状之间的关系. ...
Biological constraints on water transport in the soil-plant-atmosphere system.
1
2013
... 植物一方面要最大限度地吸收CO2进行光合作用, 另一方面要最小限度地控制水分散失, 这两个生理过程共享同一个气孔路径(Xiong et al., 2015).因此, 植物通过气孔吸收CO2的气孔导度(Gs, 气相水分散失能力指标)与释放水蒸气的水力导度(Kleaf, 液相水分传输能力指标)之间存在权衡机制(Sack & Holbrook, 2006).由于C3植物的Gs与光合速率的耦合关系, 因此Kleaf与光合速率也存在正相关关系(Brodribb et al., 2005, 2007).对系统发生、叶片结构、叶片寿命、地理分布等特性差异较大的大量物种的综合研究发现, Kleaf与Gs和光合速率呈正相关关系(Brodribb et al., 2005, 2007), 表明水力供需能力及碳获取与水分运输能力之间存在平衡关系(Brodribb & Feild, 2000; Meinzer, 2002; Sack et al., 2003, 2005; Brodribb & Holbrook, 2004b; Brodribb et al., 2005; Campanello et al., 2008; Prentice et al., 2014), 也反映叶片水平的气液两相导度的协调性; 有效的叶片水分供应能够最大限度地维持气孔开放(Brodribb et al., 2005, 2007; Lo Gullo et al., 2005; Nardini et al., 2005; Manzoni et al., 2013).Katul等(2003)提出了一个把Gs、光合速率和水分运输效率联系在一起的理论框架模型, 可以直接预测最大羧化能力与土壤-植物水力性状之间的关系. ...
Growth dynamics of root and shoot hydraulic conductance in seedlings of five neotropical tree species: Scaling to show possible adaptation to differing light regimes.
1
1998
... 叶片水容(Cleaf)表征植物调节根系供水和叶片蒸腾需水之间的不平衡、缓冲叶片水势波动的能力, 是植物避免干旱的水力对策(Lamont & Lamont, 2000; Sack et al., 2003, 2005).Blackman等(2010)测定了20种温带木本植物的叶片体积水容和叶片组织水容, 结果表明Kleaf与叶片组织水容呈正相关, 但与体积水容无关.这说明并不是所有的叶片储存水都可以补充蒸腾的水分损失, 只有叶片组织中的储存水可以与蒸腾流进行自由交换, 支持了叶片水力分区的观点(Zwieniecki et al., 2007).Kleaf与Cleaf功能上的协调性解释了蒸腾速率一定的情况下Kleaf较高的叶片关闭气孔较慢的原因(Tyree et al., 1998; Aasamaa & Sõber, 2001; Hao et al., 2008). ...
Are symplast tolerance to intense drought conditions and xylem vulnerability to cavitation coordinated? An integrated analysis of photosynthetic, hydraulic and leaf level processes in two Mediterranean drought-resistant species.
1
2010
... 叶片失膨点水势或萎蔫水势(Ψtlp)是叶片对干旱胁迫耐性的决定因子, 对植株的生理干旱耐性有重要贡献(Maréchaux et al., 2015).有研究报道Ψtlp与P50、Kleaf呈正相关(Nardini et al., 2012a; Villagra et al., 2013), 表明Ψtlp低(即叶片耐旱性高)的物种, 其叶片组织的供水能力低(Kleaf低), 但抗栓塞阻力强(P50低); 相反, 水分运输效率高(Kleaf高)的物种, 能够保持叶片良好的水分状况, 阻止叶片水势的下降, 从而保持气孔开放和碳同化时间长.P50与Ψtlp的协调关系不仅解释了叶片木质部外导水率下降的原因(Brodribb & Holbrook, 2004b; Knipfer & Steudle, 2008), 还表明了叶片内部非原生质体和共质体之间抗旱阻力的协调关系(Blackman et al., 2010; Vilagrosa et al., 2010; Nardini & Luglio, 2014).此外, Brodribb和Holbrook (2003)的研究表明P50和Ψtlp影响水分胁迫期间的气孔关闭时间.在空穴化发生早期, 首先引起叶片气孔关闭(Choat et al., 2012), 在叶片膨胀体积逐渐损失直至完全丧失之前, 逐渐降低叶片蒸腾速率(Blackman et al., 2010; Scoffoni et al., 2011; Nardini et al., 2012a).因此, 叶片是保护活细胞远离临界水分状况的安全阀门. ...
Functional relationships between leaf hydraulics and leaf economic traits in response to nutrient addition in subtropical tree species.
5
2013
... LES (Wright et al., 2004)认为, 植物的化学、结构和生理功能性状呈一条连续变化的带谱, 在带谱的两端分别是“快速投资-收益”型和“缓慢投资-收益”型物种, 其间是两种类型的过渡梯度类型.叶片水力性状影响叶片碳同化能力、水分利用效率和植物生长(Sack & Holbrook, 2006), 对植物生产力和进化起着重要的作用(Brodribb et al., 2010).叶片导水率(Kleaf)和叶片水力脆弱性(P50)常用于反映叶片水力效率和水力安全; 两者的权衡关系可限制物种地理分布, 对不同空间尺度的植被组成和分布非常重要(Nardini et al., 2012a).因此, 近期越来越多的研究关注叶片水力性状对叶片经济性状的影响(Sack & Frole, 2006; Sack & Scoffoni, 2013; Sack et al., 2013; 龚容和高琼, 2015).例如, Villagra等(2013)提出LES反映的碳和养分“投资-收益”的平衡与叶片水力性状有关.Kleaf高的物种, 叶片组织密度低、寿命短, 资源获取速率快(Reich, 2014).此外, 由于叶片水平水分流动阻力占整个植株阻力的比例较大(≥30%), 且与叶片气体交换有关, 因此, Kleaf对植物碳经济、水经济及两者之间的耦合关系均至关重要(Brodribb et al., 2005, 2007; Reich, 2014).为此, 本文首先综述植物叶片水力性状、经济性状及两者之间的权衡关系; 然后从结构和功能角度探讨叶片水力性状与经济性状的调节机制; 最后针对植物的水力和经济性状耦合关系提出今后的研究方向. ...
... 叶片失膨点水势或萎蔫水势(Ψtlp)是叶片对干旱胁迫耐性的决定因子, 对植株的生理干旱耐性有重要贡献(Maréchaux et al., 2015).有研究报道Ψtlp与P50、Kleaf呈正相关(Nardini et al., 2012a; Villagra et al., 2013), 表明Ψtlp低(即叶片耐旱性高)的物种, 其叶片组织的供水能力低(Kleaf低), 但抗栓塞阻力强(P50低); 相反, 水分运输效率高(Kleaf高)的物种, 能够保持叶片良好的水分状况, 阻止叶片水势的下降, 从而保持气孔开放和碳同化时间长.P50与Ψtlp的协调关系不仅解释了叶片木质部外导水率下降的原因(Brodribb & Holbrook, 2004b; Knipfer & Steudle, 2008), 还表明了叶片内部非原生质体和共质体之间抗旱阻力的协调关系(Blackman et al., 2010; Vilagrosa et al., 2010; Nardini & Luglio, 2014).此外, Brodribb和Holbrook (2003)的研究表明P50和Ψtlp影响水分胁迫期间的气孔关闭时间.在空穴化发生早期, 首先引起叶片气孔关闭(Choat et al., 2012), 在叶片膨胀体积逐渐损失直至完全丧失之前, 逐渐降低叶片蒸腾速率(Blackman et al., 2010; Scoffoni et al., 2011; Nardini et al., 2012a).因此, 叶片是保护活细胞远离临界水分状况的安全阀门. ...



{kind=link}
{kind=link}