常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 )。目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 )。长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究。然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱。
湖北西南部地区是湖北重要的天然林分布区, 亚热带常绿阔叶林为该地区的地带性植被。随着海拔的升高, 含有落叶成分的亚热带常绿落叶阔叶混交林逐步成为优势植被。亚热带常绿落叶阔叶混交林植物区系成分复杂, 生物多样性丰富, 不仅为我国的经济建设提供了大量的优质木材和林副产品, 而且对我国特有的古老孑遗物种保护、维护区域生态平衡发挥着重要的作用(汤景明等, 2010 )。由于气候、地质和区域物种库等方面的差异, 不同区域的常绿落叶阔叶混交林具有不同的变化特征, 针对不同区域的森林植被开展相关的生态学研究显得尤为必要。对亚热带常绿落叶阔叶混交林的群落类型进行科学划分, 不仅有助于我们对现存森林植被基本特征的认识, 而且是我们进一步深入开展生态系统功能与生态过程研究的基础。了解常绿和落叶物种比例随群落类型的变化规律, 能够揭示不同群落中环境条件对物种分布的影响, 从而为我们阐明生物多样性维持与调控机制奠定基础。亚热带常绿落叶阔叶混交林的群落类型及组成结构的研究是对其进行有效地保育、可持续经营与合理利用的科学基础。
本研究以鄂西南星斗山和木林子自然保护区亚热带常绿落叶阔叶混交林为研究对象, 在建立系列森林动态固定样地的基础上, 对该地区的植物群落进行了数量分类与排序, 探讨植被与环境之间的关系, 并分析常绿和落叶物种随不同群落类型及环境条件的变化, 以期为深入研究亚热带常绿落叶阔叶混交林的生物多样性与生态功能维持机制, 合理保护与利用此类森林植被提供科学依据。
1 研究区概况
研究地点位于湖北恩施的星斗山国家级自然保护区和木林子国家级自然保护区。以上2个保护区是鄂西南天然森林植被保存最为完好的区域, 是鄂西南现存天然森林植被的典型代表。
星斗山国家级自然保护区位于鄂西南恩施、利川与咸丰三县(市、州)交界处, 地理坐标29.95°- 30.17° N, 108.95°-109.45° E。是齐岳山向东北的延伸部分, 因有大巴山系巫山余脉作屏障, 成为第三纪植物的“避难所”。海拔672-1751 m。雨量充沛, 气候温和, 年平均气温12.8℃, 年降水量1200-1400 mm, 无霜期234天, 相对湿度85%左右。土壤以山地黄壤、黄棕壤、棕壤为主(艾训儒和谭建锡, 1999 ; 方元平和刘胜祥, 2006 )。
湖北木林子国家级自然保护区位于湖北省恩施土家族苗族自治州鹤峰县境内, 地理坐标29.21°- 30.17° N, 109.98°-110.81° E, 海拔1100.0-2095.6 m, 属武陵山脉北支余脉, 地势由西北和东南向中间逐渐倾斜。该区属于亚热带季风性湿润气候, 雨热同期, 时空分布不均, 年降水量1733 mm, 春秋多阴雨, 夏季雨量较多, 冬季雨少雾多, 蒸发小, 湿度大, 年平均相对湿度为82%, 无霜期270-279天。年平均气温为15.5 ℃, 最冷月(1月)平均气温为4.6 ℃, 最热月(7月)平均气温为26 ℃。土壤以黄棕壤、棕壤、黄壤为主(葛继稳等, 2009 )。
2 研究方法
2.1 固定样地的建立与调查
2013年9月-2014年6月, 在湖北星斗山自然保护区和湖北木林子自然保护区的典型天然森林植被中, 分别随机设置永久性森林动态样地45个和47个, 每个样地面积均为20 m × 20 m。这些样地分布于海拔1248-1800 m之间。样地建设采用CTFS森林动态样地监测标准(Condit, 1998 )。在每个样地中利用网格法将其分割成16个5 m × 5 m的小样方, 在小样方内对所有胸径≥1 cm的乔木、灌木和藤本进行编号、测量点刷漆标记并开展每木检尺, 记录内容包括物种名称、胸径、坐标、萌生与死亡状况, 同时记录样地所在坡度、海拔、凋落物厚度。
2.2 常绿和落叶物种的确定
常绿和落叶物种的划分主要通过查阅《湖北植物志》(傅书遐, 2002 )及咨询植物分类专家来确定, 对于那些不明确的物种, 由作者在冬季进行实地观测和采访当地林业工作者获得。
2.3 数据处理
坡向以正北为起点(0°), 采用顺时针旋转的角度表示, 数字等级划分为: 1表示阴坡(0°-45°和315°-360°), 2表示半阴坡(45°-90°和270°-315°), 3表示半阳坡(90°-135°和225°-270°), 4表示阳坡(135°-225°), 数值越大, 表示光照越多。土壤因子的测定方法为: 土壤pH值用电位法、土壤有机质用重铬酸钾容量法、土壤碱解N用碱解扩散法、速效P用双酸法浸提-钼锑抗比色法、全N用半微量凯氏定氮法、全P用硫酸-高氯酸消解法。
以样地为单位分别计算物种的重要值, 计算公式为: 重要值(IV ) = (相对多度+相对显著度+相对频度)/3。采用VEAPAN软件包中的TWINSPAN进行群落分类; 通过CANOCO 4.5软件(张金屯, 1995 )对样地和物种的重要值矩阵进行DCA排序。根据个体胸径(DBH )将物种划分为3个等级: 小径级(1 cm ≤ DBH < 5 cm)、中径级(5 cm ≤ DBH < 10 cm)和大径级(DBH ≥ 10 cm)。基于Bray-Curtis相似距离, 采用基于矩阵的多元回归分析(Permutation-based multiple regression on distance matrices, MRM)确定显著影响群落物种组成变化的环境因子(Lichstein, 2007 )。运用多元逐步回归分析(multiple regression analysis)来分析常绿和落叶物种与环境因子之间的关系。
使用R 3.1.1和Origin 8.5进行统计分析和作图。
3 结果和分析
3.1 群落数量分类与排序
应用TWINSPAN分类可将所调查的92个样地分为5个群落类型(图1 )。在DCA排序图(图2 )中, DCA前两轴的贡献率分别为56.7%和29.8%, 累计贡献率达86.5%。结合TWINSPAN分类与DCA排序的结果, 可将鄂西南星斗山和木林子自然保护区亚热带常绿落叶阔叶混交林的森林群落划分为5类, 其中木林子包括I (青冈-翅柃+尖连蕊茶群落(Cycl- obalanopsis glauca-Eurya alata + Camellia cuspidata ) )、II (川陕鹅耳枥+青冈-翅柃群落(Carpinus fargesiana + Cyclobalanopsis glauca-Eurya alata ))和III (川陕鹅耳枥+青冈-水马桑群落(Carpinus fargesiana + Cyclobalanopsis glauca-Weigela japonica ) ) 3个群落, 星斗山包括IV (光叶水青冈+多脉青冈-翅柃群落(Fagus lucida + Cyclobalanopsis multinervis- Eurya alata ) )和V (川陕鹅耳枥+交让木+包果柯-翅柃群落(Carpinus fargesiana+Daphniphyllum macro- podum + Lithocarpus cleistocarpus-Euryaalata )) 2个群落。各群落的主要物种组成见表1 。各样地的物种状况见附录1。
图1 鄂西南两个保护区亚热带常绿落叶阔叶混交林92个样地的TWINSPAN分类树状图。D, 分类次数; N, 样方数。
Fig. 1 Dendrogram of TWINSPAN classification for 92 plots in the subtropical evergreen-deciduous broadleaved mixed forest across two nature reserves in Southwest Hubei, China. D, division number; N, number of plot.
图2 鄂西南两个保护区亚热带常绿落叶阔叶混交林92个样地的DCA二维排序图。罗马数字(I, II, III, IV, V)代表不同的群落类型。
Fig. 2 Two-dimensional DCA ordination diagram for 92 plots in the subtropical evergreen-deciduous broadleaved mixed forest across two nature reserves in Southwest Hubei, China. Roman numerals (I, II, III, IV, V) indicate different community types.
3.2 不同群落类型内常绿和落叶物种丰富度、多度、胸高断面积及重要值的变化
在5个群落类型中, 群落II、群落III和群落V内落叶物种的物种丰富度显著大于常绿物种(p < 0.001) (图3A )。群落I与群落IV内常绿和落叶物种的物种丰富度差异不显著(p > 0.05)。
图3 鄂西南两个保护区亚热带常绿落叶阔叶混交林5种群落类型的常绿和落叶物种丰富度、多度、胸高断面积和重要值(平均值±标准偏差)。不同小写字母(a, b)表示显著差异(p < 0.05)。I、II、III、IV、V分别为青冈-翅柃+尖连蕊茶群落, 川陕鹅耳枥+青冈-翅柃群落, 川陕鹅耳枥+青冈-水马桑群落, 光叶水青冈+多脉青冈-翅柃群落和川陕鹅耳枥+交让木+包果柯-翅柃 群落。
Fig. 3 Species richness, abundance, basal area and importance value of the five forest types in the subtropical evergreen-deciduous broadleaved mixed forest across two nature reserves in Southwest Hubei, China (mean ± SD ). Different lowercase letters (a, b) indicate significant differences at the level of 0.05. I, II, III, IV, V indicate different community types: Cyclobalanopsis glauca-Eurya alata + Camellia cuspidata, Carpinus fargesiana + Cyclobalanopsis glauca-Eurya alata, Carpinus fargesiana + Cyclobalanopsis glauca-Weigela japonica, Fagus lucida + Cyclobalanopsis multinervis-Eurya alata, Carpinus fargesiana + Daphniphyllum macropodum + Lithocarpus cleistocarpus-Eurya alata .
群落I和群落III中常绿与落叶物种的多度存在显著的差异(p < 0.001), 但群落I的落叶物种多度显著小于常绿物种, 而群落III呈现相反的趋势(图3B )。其他3个群落类型中常绿与落叶物种的多度差异不显著(p > 0.05)。
群落II、群落III和群落V中落叶物种的胸高断面积均显著(p < 0.001)大于常绿物种(图3C )。群落I与群落IV的常绿和落叶物种的胸高断面积差异不显著(p > 0.05), 但群落IV中落叶物种的胸高断面积略高于常绿树种。
群落II、群落III和群落V中落叶物种的重要值显著大于常绿物种(p < 0.01) (图3D ), 群落I中的常绿物种重要值显著大于落叶物种, 群落IV的常绿物种重要值大于落叶物种, 但差异并不显著(p > 0.05)。
3.3 不同径级的常绿和落叶物种丰富度、多度、胸高断面积及重要值比例的变化
除群落I外, 所有群落类型内的常绿和落叶物种丰富度比例均随胸径级的增加而下降(图4A )。除群落IV内1-5 cm的径级外, 其他群落类型各径级中的落叶物种丰富度均大于常绿物种。
图4 鄂西南两个保护区亚热带常绿落叶阔叶混交林5种群落类型的常绿和落叶物种不同径级的物种丰富度、多度、胸高断面积和重要值比例。a, 胸径1-5 cm; b, 胸径5-10 cm; c, 胸径≥10 cm。I-V同图3 。
Fig. 4 Evergreen/deciduous ratio of species richness, abundance, basal area and importance value with size class in subtropical evergreen-deciduous broadleaved mixed forest across two nature reserves in Southwest Hubei, China. a, DBH 1-5 cm; b, DBH 5-10 cm; c, DBH ≥10 cm; DBH, diameter at breast height. I-V see Fig. 3 .
除群落I外, 常绿和落叶物种多度在群落内所占的比例随径级的增加而逐渐下降(图4B )。除群落III外, 各群落内1-5 cm径级的常绿物种多度均大于落叶物种, 在5-10 cm的径级内, 群落I和群落IV的常绿物种多度大于落叶物种, 群落II、群落III和群落V的落叶物种多度大于常绿物种。除群落I外, 各群落的落叶物种多度在≥10 cm的径级内均大于常绿物种。
各群落类型内常绿物种和落叶物种的胸高断面积比例均随着径级的增加而升高(图4C )。群落II、群落III和群落V中落叶物种的胸高断面积比例在1-5 cm和5-10 cm的径级内大于常绿物种, 群落I和群落IV中则呈现相反的趋势。各群落胸径≥10 cm的落叶物种比例均大于常绿物种。
除群落III外, 常绿物种的重要值在1-5 cm径级内均大于落叶物种(图4D )。在5-10 cm的径级内, 群落I和群落IV的常绿物种重要值大于落叶物种, 群落II、群落III和群落V的落叶物种重要值大于常绿物种。各群落胸径≥ 10 cm的落叶物种重要值均大于常绿物种。
3.4 群落物种组成与环境因子的关系
基于距离矩阵的多元回归分析(MRM)结果表明: 环境因子可以解释物种组成变化的59%, 海拔、坡度、坡向、土壤全氮、土壤全磷、土壤碱解氮、土壤速效磷是影响研究区群落分布的主要因子(表2 )。
整体来看, 群落I主要分布在坡度大、全磷、全氮含量高、海拔在1538 m左右的阴坡; 群落II主要分布在坡度较大、速效磷含量低、海拔在1470 m左右的阴坡和半阴坡; 群落III主要分布在坡度较小, 全磷含量较高、海拔在1762 m左右的阳坡和半阳坡; 群落IV主要分布在碱解氮和全氮含量高、海拔在1587 m左右的阴坡和半阴坡; 群落V主要分布在坡度小、速效磷含量高、全磷和碱解氮含量低、海拔在1406 m左右的半阴坡。
采用多元逐步回归分析对各群落常绿和落叶物种丰富度与环境因子关系的研究(表3 )表明, 除土壤含水量和有机质之外, 其他环境因子在不同群落类型间与常绿和落叶物种呈现显著的相关性(p < 0.05)。海拔高度与群落III和群落V的落叶物种显著相关。坡度与群落I和群落IV的落叶物种显著负相关。坡位与群落I、群落III和群落IV显著相关。土壤氮与群落I、群落II、群落III和群落IV显著相关。土壤磷与群落II和群落V显著负相关。
4 讨论
4.1 亚热带常绿落叶阔叶混交林群落的数量分类与排序
通过TWINSPAN及DCA方法对鄂西南木林子自然保护区和星斗山自然保护区的亚热带常绿落叶阔叶混交林进行数量分类和排序, 将两个保护区的92个样地分成了5个群落类型。TWINSPAN划分出的群落类型在DCA排序图上能明显地区分开, 说明排序与分类结果基本吻合, 数量化的分类结果是可信的。
植被分类是植被生态学研究中最复杂的一个问题, 直到现在还没有植被学家们认同的统一分类原则和分类系统(宋永昌, 2004 )。而我国亚热带地区植被具有复杂多样、次生性强和过渡类型多等特点, 更增加了该地区植被分类的困难(赖江山等, 2010 )。传统的植物群落分类多根据群落的外貌特征及群落的种类组成划分群落类型, 个人经验对分类结果的影响比较大(刘海江和郭柯, 2003 ; 刘万德等, 2010 )。随着植被研究的精度要求不断提高, 提供客观归类和划分结果的数量分类方法在植被分类中越来越重要(袁秀等, 2013 )。目前数量生态学中的分类和排序方法已成为现代植被生态学研究中必不可少的手段, 双向指示种分析法(TWINSPAN)和除趋势对应分析法(DCA)已广泛应用于森林和草地等植物群落分析(Ozinga et al ., 2005 ; 孙菊等, 2009 ; 廉凯敏等, 2015 )。本研究也证实了TWINSPAN分类与DCA排序这两种方法相结合应用于亚热带常绿落叶阔叶混交林植被数量分析是可行的, 而且可以相互补充和交叉检验。
4.2 常绿和落叶物种比例随群落类型的变化及其环境解释
由于叶片是植物光合作用的主要场所, 因而叶片的物候过程对于群落外貌、林下环境、凋落物分解和生态系统生产力具有重要作用(Bohlman et al ., 1998 ; Quigley & Platt, 2003 )。落叶是物种适应环境胁迫的一种重要策略, 通过落叶可以回避不利生境对树木生长造成的伤害和不利影响(Poorter & Markesteijn, 2008 ), 低温和水分胁迫是阔叶树种落叶的重要原因(Lechowicz, 1984 ; Reich et al ., 2004 )。与热带部分地区树种因土壤水分亏缺而出现旱季落叶的情况不同(Williams et al ., 2008 ; de Faria Lopes et al ., 2012 ), 亚热带常绿落叶阔叶混交林的形成更多受到海拔升高而形成的低温胁迫的影响(Oliveira- Filho et al ., 2006 ; 白坤栋等, 2010 ; 宋坤, 2012 )。回归分析中土壤含水量与常绿和落叶物种分布并无显著的相关性, 这个结果证明了研究区常绿和落叶物种的变化与土壤水分无关。温度是限制植物组成、生长、繁殖和分布的重要因子(Woodward, 1987 )。研究地区冬季气温较低, 一些喜温且生长迅速竞争能力强的常绿阔叶树种受到限制, 但绝对低温尚较高, 因此较耐寒的常绿阔叶树与落叶阔叶树均能生长, 最终在该地段形成了亚热带常绿落叶阔叶混交林(吴征镒, 1980 )。
从物种组成和群落结构来看, 研究区各群落类型常绿和落叶物种存在一定的差异。群落I和群落IV的常绿和落叶物种在群落内的差异多不显著或者常绿物种大于落叶物种, 而群落II、群落III和群落V则表现为落叶物种大于常绿物种。造成这种差异的主要原因可能是环境差异和人为干扰。环境是影响物种组成的决定性因素, 环境因子组成的变化产生了不同的生境, 从而引起不同地带及区域的物种差异(Toledo et al ., 2011 )。而人为干扰能够显著地改变群落结构和物种组成, 从而影响群落的发展方向(Lomolino, 2001 ; Haberl et al ., 2007 ; Nogués-Bravo et al ., 2008 )。群落III位于海拔较高的阳坡, 随着海拔的升高, 气温逐渐降低, 光照强度增加, 较低的气温和阳坡较好的光照条件促进落叶物种的生长(Malhi et al ., 2010 )。充足的光照也促进土壤种子库萌发, 使群落内幼苗和幼树的密度较大。同时, 群落III位于保护区森林防火隔离带附近, 较频繁地受到了人为的干扰, 干扰提高了林下的光照条件, 导致群落内喜光的落叶树种增加和耐阴的常绿树种减少(Hawthorne, 1996 ; Bongers et al ., 2009 )。群落II和群落V分布于低海拔地区, 长期受到人类樵采等活动的影响, 林下的光照条件较好, 群落内的落叶物种大于常绿物种。群落I和群落IV主要位于阴坡, 且人为干扰较少, 林内光照条件较差, 适宜常绿物种的生长。
除地形因子和人为干扰之外, 土壤养分也对常绿和落叶物种比例的变化产生了一定的影响。落叶物种分布区的土壤氮含量常被认为高于常绿物种分布区, 其原因是落叶物种比常绿物种具有更高的氮矿化速率(Aerts & Chapin, 1999 ; Cornwell et al ., 2008 ), 然而也有研究表明常绿和落叶物种的氮矿化速率并无显著差异(Reich et al ., 2001 ; Booth et al ., 2005 )。本研究发现土壤氮含量在多数群落内与常绿物种丰富度显著正相关, 这可能与不同群落类型的物种属性及真菌差异有关(Mueller et al ., 2012 )。总体来看, 研究区常绿物种更易受土壤养分的影响。关于土壤养分对各群落内常绿和落叶物种分布的影响有待进一步深入分析。
4.3 常绿和落叶物种比例随胸径级的变化
除群落III外, 各群落类型小径级(1-5 cm)内落叶物种的物种丰富度高于常绿物种, 但多度、胸高断面积和重要值却表现为落叶物种小于常绿物种, 这主要与常绿和落叶物种的生活型属性有关。研究区群落内小径级个体的生活型主要是小乔木和灌木, 而大乔木个体密度相对较低。这些小乔木和灌木物种以常绿树种为主, 因此出现了小径级内落叶物种丰富度高但多度、胸高断面积和重要值低的现象。较高强度的干扰能够提高落叶树种在群落中的比例(丁易和臧润国, 2008 ), 群落III的小径级在各个指标上都是落叶树种占优势, 这可能说明人为干扰和环境条件显著影响了该群落的更新和发展方向, 导致该群落在未来一段时间内都将以落叶物种为主。大径级(≥10 cm)的落叶物种在多数群落内的物种丰富度、多度、胸高断面积和重要值均大于常绿物种, 表明落叶树种在林冠层占据显著的优势, Chabot和Hicks (1982) 的研究也发现, 光照条件较好的林冠层落叶物种更多。林冠层落叶树种占据优势能够显著影响林下光照条件, 从而影响森林更新和物种多样性(Quigley & Platt, 2003 )。落叶物种具有更高的资源养分利用效率(Kloeppel et al ., 2000 ), 能够利用夏季良好的水热条件迅速获得碳水化合物完成高生长, 在与常绿树种的竞争过程中居于优势地位(Antúnez et al ., 2001 )。同时, 落叶物种采取了落叶的策略来适应研究区的冬季低温胁迫, 而常绿物种则保留叶片越冬, 低温可能损害常绿树种的气孔和光合速率而使常绿植物生长受到严重制约(Wyka & Oleksyn, 2014 ), 且较大的季节性温差所引起的冰凌和雪灾也可能造成常绿植物受到伤害从而限制其在林冠层的优势(白坤栋等, 2010 )。林下的常绿树木由于林冠层的郁闭而受到光强的限制。尽管在林冠层落叶期间(晚秋到早春), 林下光照条件可以由于林冠层的落叶而得到改善, 但低温限制了林下常绿树木的生长。这种竞争最终形成了林冠层以落叶阔叶树为主和林下层以常绿树种为主的格局。
有关常绿、落叶物种随群落类型变化的具体机制还需要进一步深入细致研究, 如群落所处的具体小生境(土壤、地形和小气候等因子)、不同物种的生理生态特性与生态适应性等方面。这些研究将随着鄂西南地区森林动态样地监测平台的建设逐步开展。通过长期定位观测和基础研究, 阐明该地区常绿落叶阔叶混交林的恢复动态规律, 可为该地区基于生态系统功能的恢复实践提供重要的理论依据。
附录
附录1 鄂西南星斗山和木林子自然保护区亚热带常绿落叶阔叶混交林各样地物种名录
Appendix 1 List of species of the subtropical evergreen-deciduous broadleaved mixed forest across two national nature reserves in Southwest Hubei, China
http://www.plant-ecology.com/appendix/CJPE2015-10-A1.pdf
致谢 感谢湖北民族学院林学园艺学院2012级本科生、硕士生冯广、刘峻城、林勇及木林子国家级自然保护区米显齐书记等人在野外调查工作中给予的帮助。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Aerts R Chapin FS III 1999 ). The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns.
Advances in Ecological Research , 30 , 1 -67 .
[本文引用: 1]
[2]
Ai XR Tan XJ 1999 ). On the population structure of Davidia involucrata in Xingdoushan protected region
. Journal of Hubei Institute for Nationalities (Natural Sciences) , 17 , 12 -15 . (in Chinese with English abstract)
[本文引用: 1]
[艾训儒 , 谭建锡 (1999 ). 星斗山自然保护区珙桐种群结构特征研究
. 湖北民族学院学报(自然科学版) , 17 , 12 -15 .]
[本文引用: 1]
[3]
An SQ Zhang JH Tan JK Zhu XL Zhao RL 1998 ). Review of studies on secondary forests in north subtropics, China.
Journal of Wuhan Botanical Research , 16 , 268 -272 . (in Chinese with English abstract)
[本文引用: 2]
[安树青 , 张久海 , 谈健康 , 朱学雷 , 赵儒林 (1998 ). 中国北亚热带次生森林植被研究述评
. 武汉植物学研究 , 16 , 268 -272 .]
[本文引用: 2]
[4]
Antúnez I Retamosa EC Villar R 2001 ). Relative growth rate in phylogenetically related deciduous and evergreen woody species.
Oecologia , 128 , 172 -180 .
[本文引用: 1]
[5]
Bai KD Jiang DB Cao KF Wan XC Liao DB 2010 ). Photosynthetic response to seasonal temperature changes in evergreen and deciduous broad-leaved trees in montane forests of Ailao Mountain and Mao’er Mountain.
Acta Ecologica Sinica , 30 , 905 -913 . (in Chinese with English abstract)
[本文引用: 2]
[白坤栋 , 蒋得斌 , 曹坤芳 , 万贤崇 , 廖德宝 (2010 ). 哀牢山和猫儿山中山常绿和落叶阔叶树光合特性对季节温度变化的响应
. 生态学报 , 30 , 905 -913 .]
[本文引用: 2]
[6]
Ban JD Qi GS 1995 ). A Study of Vegetation of Western Hubei . Press of Huazhong University of Science and Technology , Wuhan . (in Chinese)
[本文引用: 1]
[班继德 , 漆根深 (1995 ). 鄂西植被研究 . 华中理工大学出版社 , 武汉 .]
[本文引用: 1]
[7]
Bohlman SA Adams JB Smith MO Peterson DL 1998 ). Seasonal foliage changes in the Eastern Amazon Basin detected from landsat thematic mapper satellite images.
Biotropica , 30 , 376 -391 .
[本文引用: 1]
[8]
Bongers F Poorter L Hawthorne WD Sheil D 2009 ). The intermediate disturbance hypothesis applies to tropical forests, but disturbance contributes little to tree diversity.
Ecology Letters , 12 , 798 -805 .
[本文引用: 1]
[9]
Booth MS Stark JM Rastetter E 2005 ). Controls on nitrogen cycling in terrestrial ecosystems: A synthetic analysis of literature data.
Ecological Monographs , 75 , 139 -157 .
[本文引用: 1]
[10]
Cao TR Qi CJ Yu XL 1997 ). Studies on species diversity of Fagus lucida communities on the Badagongshan Mountain, Hunan.
Chinese Biodiversity , 5 (2 ), 112 -120 . (in Chinese with English abstract)
[本文引用: 1]
[曹铁如 , 祁承经 , 喻勋林 (1997 ). 湖南八大公山亮叶水青冈群落物种多样性的研究
. 生物多样性 , 5 (2 ), 112 -120 .]
[本文引用: 1]
[11]
Chabot BF Hicks DJ 1982 ). The ecology of leaf life spans.
Annual Review of Ecology and Systematics , 13 , 229 -259 .
[本文引用: 1]
[12]
Condit R 1998 ). Tropical Forest Census Plots: Methods and Results from Barro Colorado Island, Panama and a Comparison with Other Plots
. Springer Science & Business Media, Berlin.
[本文引用: 1]
[13]
Cornwell WK Cornelissen JHC Amatangelo K Dorrepaal E Eviner VT Godoy O Hobbie SE Hoorens B Kurokawa H Pérez-Harguindeguy N Quested HM Santiago LS Wardle DA Wright IJ Aerts R Allison SD van Bodegom P Brovkin V Chatain A Callaghan TV Díaz S Garnier E Gurvich DE Kazakou E Klein JA Read J Reich PB Soudzilovskaia NA Vaieretti MV Westoby M 2008 ). Plant species traits are the predominant control on litter decomposition rates within biomes worldwide.
Ecology Letters , 11 , 1065 -1071 .
[本文引用: 1]
[14]
de Faria Lopes S Schiavini I do Vale VS do Prado Júnior JA de Silvério Arantes C 2012 ). Historical review of studies in seasonal semideciduous forests in Brazil: A perspective for conservation.
Brazilian Geographical Journal: Geosciences and Humanities Research Medium , 3 , 21 -40 .
[本文引用: 1]
[15]
Ding Y Zang RG 2008 ). Changes in deciduous trees during recovery of tropical lowland rain forests on abandoned shifting cultivation lands in Hainan Island, South China.
Biodiversity Science , 16 , 103 -109 . (in Chinese with English abstract)
[本文引用: 1]
[丁易 , 臧润国 (2008 ). 海南岛热带低地雨林刀耕火种弃耕地恢复过程中落叶树种的变化
. 生物多样性 , 16 , 103 -109 .]
[本文引用: 1]
[16]
Fang JY 2001 ). Re-discussion about the forest vegetation zonation in eastern China.
Acta Botanica Sinica , 43 , 522 -533 . (in Chinese with English abstract)
[本文引用: 2]
[方精云 (2001 ). 也论我国东部植被带的划分
. 植物学报 , 43 , 522 -533 .]
[本文引用: 2]
[17]
Fang YP Liu SX 2006 ). Research on the national protected plants in Xingdoushan Nature Reserve in Hubei Province.
Subtropical Plant Science , 34 (3 ), 48 -52 . (in Chinese with English abstract)
[本文引用: 1]
[方元平 , 刘胜祥 (2006 ). 湖北星斗山自然保护区国家重点保护野生植物调查研究
. 亚热带植物科学 , 34 (3 ), 48 -52 .]
[本文引用: 1]
[18]
Fu SX 2002 ). Flora of Hubeiensis . Hubei Science and Technology Press , Wuhan . (in Chinese)
[本文引用: 1]
[傅书遐 (2002 ). 湖北植物志 . 湖北科学技术出版社 , 武汉 .]
[本文引用: 1]
[19]
Ge JW Hu HX Li B 2009 ). Scientific Survey and Study on Forest Biodiversity in Mulinzi Nature Reserve of Hubei, Central China . Science Press , Beijing . (in Chinese)
[本文引用: 1]
[葛继稳 , 胡鸿兴 , 李博 (2009 ). 湖北木林子自然保护区森林生物多样性研究 . 科学出版社 , 北京 .]
[本文引用: 1]
[20]
Haberl H Erb KH Krausmann F Gaube V Bondeau A Plutzar C Gingrich S Lucht W Fischer-Kowalski M 2007 ). Quantifying and mapping the human appropriation of net primary production in earth’s terrestrial ecosystems.
Proceedings of the National Academy of Sciences of the United States of America , 104 , 12942 -12947 .
[本文引用: 1]
[21]
Han WH Li XK Ye D Lü SH Xiang WS Song TQ Cao HL 2009 ). Interspecific association and correlation between dominant woody plant species in an evergreen and deciduous broad-leaved mixed forest of karst area, Northwest Guangxi.
Journal of Mountain Science , 27 , 719 -726 . (in Chinese with English abstract)
[本文引用: 1]
[韩文衡 , 李先琨 , 叶铎 , 吕仕洪 , 向悟生 , 宋同清 , 曹洪麟 (2009 ). 桂西北喀斯特区常绿落叶阔叶混交林种群种间联结性与相关性
. 山地学报 , 27 , 719 -726 .]
[本文引用: 1]
[22]
Hawthorne WD 1996 ). Holes and the sums of parts in Ghanaian forest: Regeneration, scale and sustainable use. Proceedings of the Royal Society of Edinburgh. Section B.
Biological Sciences , 104 , 75 -176 .
[本文引用: 1]
[23]
Kloeppel BD Gower ST Vogel JG Reich PB 2000 ). Leaf- level resource use for evergreen and deciduous conifers along a resource availability gradient.
Functional Ecology , 14 , 281 -292 .
[本文引用: 1]
[24]
Lai JS Mi XC Ren HB Ma KP 2010 ). Numerical classification of associations in subtropical evergreen broad-leaved forest based on multivariate regression trees—a case study of 24 hm2 Gutianshan forest plot in China.
Chinese Journal of Plant Ecology , 34 , 761 -769 . (in Chinese with English abstract)
[本文引用: 1]
[赖江山 , 米湘成 , 任海保 , 马克平 (2010 ). 基于多元回归树的常绿阔叶林群丛数量分类——以古田山24公顷森林样地为例
. 植物生态学报 , 34 , 761 -769 .]
[本文引用: 1]
[25]
Lechowicz MJ 1984 ). Why do temperate deciduous trees leaf out at different times? Adaptation and ecology of forest communities.
The American Naturalist , 124 , 821 -842 .
[本文引用: 1]
[26]
Lian KM Wu YJ Zhang L Zhao LL Zhang F 2015 ). Numerical classification and ordination of wild Castanea mollissima communities in Taikuan River Nature Reserve, Shanxi.
Chinese Journal of Ecology , 34 , 33 -39 . (in Chinese with English abstract)
[本文引用: 1]
[廉凯敏 , 吴应建 , 张丽 , 赵璐璐 , 张峰 (2015 ). 太宽河自然保护区板栗群落数量分类与排序
. 生态学杂志 , 34 , 33 -39 .]
[本文引用: 1]
[27]
Lichstein JW 2007 ). Multiple regression on distance matrices: A multivariate spatial analysis tool.
Plant Ecology , 188 , 117 -131 .
[本文引用: 1]
[28]
Liu HB Wang QG Lu JM Xu YZ Lu ZJ Qiao XJ Bao DC Guo YL Meng HJ Jiang MX 2014 ). Root-sprouting ability in an evergreen and deciduous broad-leaved mixed forest.
Chinese Science Bulletin , 59 , 3491 -3498 . (in Chinese)
[本文引用: 1]
[刘海波 , 王庆刚 , 路俊盟 , 徐耀粘 , 卢志军 , 乔秀娟 , 鲍大川 , 郭屹立 , 孟红杰 , 江明喜 (2014 ). 八大公山常绿落叶阔叶混交林根萌能力
. 科学通报 , 59 , 3491 -3498 .]
[本文引用: 1]
[29]
Liu HJ Guo K 2003 ). Classification and ordination analysis of plant communities in inter-dune lowland in Hunshandak Sandy Land.
Acta Ecologica Sinica , 23 , 2163 -2169 . (in Chinese with English abstract)
[本文引用: 1]
[刘海江 , 郭柯 (2003 ). 浑善达克沙地丘间低地植物群落的分类与排序
. 生态学报 , 23 , 2163 -2169 .]
[本文引用: 1]
[30]
Liu WD Ding Y Zang RG Su JR Yang M Cai DL Li RC Chen SW 2010 ). Quantitative classification and ordination of low-altitudinal tropical forests in Bawangling, Hainan Island.
Chinese Journal of Ecology , 29 , 1526 -1532 . (in Chinese with English abstract)
[本文引用: 1]
[刘万德 , 丁易 , 臧润国 , 苏建荣 , 杨民 , 蔡笃磊 , 李儒财 , 陈少伟 (2010 ). 海南岛霸王岭林区低海拔热带林群落数量分类与排序
. 生态学杂志 , 29 , 1526 -1532 .]
[本文引用: 1]
[31]
Lomolino MV 2001 ). Elevation gradients of species-density: Historical and prospective views.
Global Ecology and Biogeography , 10 , 3 -13 .
[本文引用: 1]
[32]
Lu SF Liu SX Fang YP 2005 ). A preliminary research of invasive species around Xingdoushan National Nature Reserve, Hubei.
Journal of Huanggang Normal University , 25 (3 ), 48 -52 . (in Chinese with English abstract)
[本文引用: 1]
[卢少飞 , 刘胜祥 , 方元平 (2005 ). 星斗山国家级自然保护区外来入侵植物初步研究
. 黄冈师范学院学报 , 25 (3 ), 48 -52 .]
[本文引用: 1]
[33]
Malhi Y Silman M Salinas N Bush M Meir P Saatchi S 2010 ). Introduction: Elevation gradients in the tropics: Laboratories for ecosystem ecology and global change research.
Global Change Biology , 16 , 3171 -3175 .
[本文引用: 1]
[34]
Mueller KE Hobbie SE Oleksyn J Reich PB Eissenstat DM 2012 ). Do evergreen and deciduous trees have different effects on net N mineralization in soil.
Ecology , 93 , 1463 -1472 .
[本文引用: 1]
[35]
Nogués-Bravo D Araújo MB Romdal T Rahbek C 2008 ). Scale effects and human impact on the elevational species richness gradients.
Nature , 453 , 216 -219 .
[本文引用: 1]
[36]
Oliveira-Filho AT Jarenkow JA Rodal MJN 2006 ). Floristic relationships of seasonally dry forests of eastern South America based on tree species distribution patterns. In: Pennington RT, Ratter JA eds. Neotropical Savannas and Seasonally Dry Forests: Plant Diversity, Biogeography, and Conservation . CRC Press , Boca Raton, USA . 151 -184 .
[本文引用: 1]
[37]
Ozinga WA Schaminée JHJ Bekker RM Bonn S Poschlod P Tackenberg O Bakker J van Groenendael JM 2005 ). Predictability of plant species composition from environmental conditions is constrained by dispersal limitation.
Oikos , 108 , 555 -561 .
[本文引用: 1]
[38]
Poorter L Markesteijn L 2008 ). Seedling traits determine drought tolerance of tropical tree species.
Biotropica , 40 , 321 -331 .
[本文引用: 1]
[39]
Quigley MF Platt WJ 2003 ). Composition and structure of seasonally deciduous forests in the Americas.
Ecological Monographs , 73 , 87 -106 .
[本文引用: 2]
[40]
Reich PB Bakken P Carlson D Frelich LE Friedman SK Grigal DF 2001 ). Influence of logging, fire, and forest type on biodiversity and productivity in southern boreal forests.
Ecology , 82 , 2731 -2748 .
[本文引用: 1]
[41]
Reich PB Uhl C Walters MB Prugh L Ellsworth DS 2004 ). Leaf demography and phenology in Amazonian rain forest: A census of 40000 leaves of 23 tree species.
Ecological Monographs , 74 , 3 -23 .
[本文引用: 1]
[42]
Shen ZH Zhao ZE Wu JQ Jin YX Huang HD Jiang MX 1999 ). Spatial pattern and relation with the topographical factors of the rare pare species at Dalaoling in the Three Gorges Region.
Acta Phytoecologica Sinica , 23 , 171 -180 . (in Chinese with English abstract)
[本文引用: 1]
[沈泽昊 , 赵子恩 , 吴金清 , 金义兴 , 黄汉东 , 江明喜 (1999 ). 三峡地区大老岭珍稀植物的分布格局及其与地形因子的关系
. 植物生态学报 , 23 , 171 -180 .]
[本文引用: 1]
[43]
Shen ZH Fang JY 2001 ). Niche comparison of two Fagus species based on the topographic patterns of their populations.
Acta Phytoecologica Sinica , 25 , 392 -398 . (in Chinese with English abstract)
[沈泽昊 , 方精云 (2001 ). 基于种群分布地形格局的两种水青冈生态位比较研究
. 植物生态学报 , 25 , 392 -398 .]
[44]
Song K 2012 ). Vegetation Characteristic of Evergreen- deciduous Broadleaved Forest Ecotone and Its Formation Mechanism, a Study in Anhui
. PhD dissertation, East China Normal University, Shanghai . 50 -60 . (in Chinese with English abstract).
[本文引用: 1]
[宋坤 (2012 ). 安徽常绿阔叶林-落叶阔叶林交错带的森林植被特征及其成因
. 博士学位论文, 华东师范大学, 上海 . 50 -60 .]
[本文引用: 1]
[45]
Song YC 1999 ). Perspective of the vegetation zonation of forest region in eastern China.
Acta Botanica Sinica , 41 , 541 -552 . (in Chinese with English abstract)
[本文引用: 1]
[宋永昌 (1999 ). 中国东部森林植被带划分之我见
. 植物学报 , 41 , 541 -552 .]
[本文引用: 1]
[46]
Song YC 2004 ). Tentative classification scheme of evergreen broad-leaved forests of China.
Acta Phytoecologica Sinica , 28 , 435 -448 . (in Chinese with English abstract)
[本文引用: 1]
[宋永昌 (2004 ). 中国常绿阔叶林分类试行方案
. 植物生态学报 , 28 , 435 -448 .]
[本文引用: 1]
[47]
Sun J Li XZ Hu YM Wang XW Lü JJ Li ZM Chen HW 2009 ). Classification, species diversity, and species distribution gradient of permafrost wetland plant communities in Great Xing’an Mountains valleys of Northeast China.
Chinese Journal of Applied Ecology , 20 , 2049 -2056 . (in Chinese with English abstract)
[本文引用: 1]
[孙菊 , 李秀珍 , 胡远满 , 王宪伟 , 吕久俊 , 李宗梅 , 陈宏伟 (2009 ). 大兴安岭沟谷冻土湿地植物群落分类、物种多样性和物种分布梯度
. 应用生态学报 , 20 , 2049 -2056 .]
[本文引用: 1]
[48]
Tang JM Dai HH Xun JG Zhai MP 2010 ). Effects on regeneration pattern for woodland fertility of mixed evergreen and deciduous broadleaved mountain forests.
Hubei Forestry Science and Technology , (6 ), 7 -10 . (in Chinese with English abstract)
[本文引用: 2]
[汤景明 , 戴汉会 , 荀继刚 , 翟明普 (2010 ). 山地常绿落叶阔叶混交林更新方式对林地肥力的影响
. 湖北林业科技 , (6 ), 7 -10 .]
[本文引用: 2]
[49]
Toledo M Poorter L Peña-Claros M Alarcón A Balcázar J Leaño C Licona JC Bongers F 2011 ). Climate and soil drive forest structure in Bolivian lowland forests.
Journal of Tropical Ecology , 27 , 333 -345 .
[本文引用: 1]
[50]
Wang ZX Lei Y Fujiwara K Liu LH Xue YG 2006 ). Community classification, species composition, and regeneration of Fagus lucida forests in subtropical mountains, China.
Biodiversity Science , 14 , 29 -40 . (in Chinese with English abstract)
[本文引用: 1]
[汪正祥 , 雷耘 , Fujiwara K 刘林翰 , 薛跃规 (2006 ). 亚热带山地亮叶水青冈林的群落分类及物种组成与更新
. 生物多样性 , 14 , 29 -40 .]
[本文引用: 1]
[51]
Williams LJ Bunyavejchewin S Baker PJ 2008 ). Deciduousness in a seasonal tropical forest in western Thailand: Interannual and intraspecific variation in timing, duration and environmental cues.
Oecologia , 155 , 571 -582 .
[本文引用: 1]
[52]
Woodward FI 1987 ). Climate and Plant Distribution . Cambridge University Press , Cambridge, UK .
[本文引用: 1]
[53]
Wu ZL 2000 ). Forest of China . China Forestry Publishing House , Beijing . (in Chinese)
[本文引用: 1]
[吴中伦 (2000 ). 中国森林 . 中国林业出版社 , 北京 .]
[本文引用: 1]
[54]
Wu ZY 1980 ). Chinese Vegetation . Science Press , Beijing . 279 -306 . (in Chinese)
[本文引用: 3]
[吴征镒 (1980 ). 中国植被 . 科学出版社 , 北京 . 279 -306 .]
[本文引用: 3]
[55]
Wyka TP Oleksyn J 2014 ). Photosynthetic ecophysiology of evergreen leaves in the woody angiosperms—A review.
Dendrobiology , 72 , 3 -27 .
[本文引用: 1]
[56]
Xiong XG Xiong GM Xie ZQ 2002 ). The regeneration of tree species in the mixed evergreen-deciduous broad-leaved forests in the Shennongjia Mountains, Hubei Province.
Acta Ecologica Sinica , 22 , 2001 -2005 . (in Chinese with English abstract)
[本文引用: 1]
[熊小刚 , 熊高明 , 谢宗强 (2002 ). 神农架地区常绿落叶阔叶混交林树种更新研究
. 生态学报 , 22 , 2001 -2005 .]
[本文引用: 1]
[57]
Yuan X Ma KM Wang D 2013 ). Optimal number of herb vegetation clusters: A case study on Yellow River Delta.
Acta Ecologica Sinica , 33 , 2514 -2521 . (in Chinese with English abstract)
[本文引用: 1]
[袁秀 , 马克明 , 王德 (2013 ). 草地植物群落最优分类数的确定——以黄河三角洲为例
. 生态学报 , 33 , 2514 -2521 .]
[本文引用: 1]
[58]
Zhang JT 1995 ). Quantitative Vegetation Ecology . Science and Technology Press of China , Beijing . (in Chinese)
[本文引用: 1]
[张金屯 (1995 ). 植被数量生态学方法 . 中国科学技术出版社 , 北京 .]
[本文引用: 1]
[59]
Zhang M Xiong GM Xie ZQ 2004 ). Ecological investigation of the mixed broad leaved deciduous and evergreen forest in Three Gorges Reservoir Area.
Resources and Environment in the Yangtze Basin , 13 , 168 -173 . (in Chinese with English abstract)
[本文引用: 1]
[张谧 , 熊高明 , 谢宗强 (2004 ). 三峡库区常绿落叶阔叶混交林的监测研究
. 长江流域资源与环境 , 13 , 168 -173 .]
[本文引用: 1]
[60]
Zhao LJ Xiang WH Li JX Deng XW Liu C 2013 ). Floristic composition, structure and phytogeographic characteristics in a Lithocarpus glaber-Cyclobalanopsis glauca forest community in the subtropical region.
Scientia Silvae Sinicae , 49 (12 ), 10 -17 . (in Chinese with English abstract)
[本文引用: 1]
[赵丽娟 , 项文化 , 李家湘 , 邓湘雯 , 刘聪 (2013 ). 中亚热带石栎-青冈群落物种组成、结构及区系特征
. 林业科学 , 49 (12 ), 10 -17 .]
[本文引用: 1]
[61]
Zhou GY 1996 ). The deciduous broad-leaved forest of China—Also discussing about the belonging of the evergreen mixed deciduous broad-leaved forest.
Journal of Ningbo University , 9 (3 ), 93 -102 . (in Chinese with English abstract)
[本文引用: 1]
[周光裕 (1996 ). 中国的落叶阔叶林——兼论常绿落叶阔叶混交林的归属问题
. 宁波大学学报 , 9 (3 ), 93 -102 .]
[本文引用: 1]
The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns.
1
1999
... 除地形因子和人为干扰之外, 土壤养分也对常绿和落叶物种比例的变化产生了一定的影响.落叶物种分布区的土壤氮含量常被认为高于常绿物种分布区, 其原因是落叶物种比常绿物种具有更高的氮矿化速率(Aerts & Chapin, 1999 ; Cornwell et al ., 2008 ), 然而也有研究表明常绿和落叶物种的氮矿化速率并无显著差异(Reich et al ., 2001 ; Booth et al ., 2005 ).本研究发现土壤氮含量在多数群落内与常绿物种丰富度显著正相关, 这可能与不同群落类型的物种属性及真菌差异有关(Mueller et al ., 2012 ).总体来看, 研究区常绿物种更易受土壤养分的影响.关于土壤养分对各群落内常绿和落叶物种分布的影响有待进一步深入分析. ...
星斗山自然保护区珙桐种群结构特征研究
1
1999
... 星斗山国家级自然保护区位于鄂西南恩施、利川与咸丰三县(市、州)交界处, 地理坐标29.95°- 30.17° N, 108.95°-109.45° E.是齐岳山向东北的延伸部分, 因有大巴山系巫山余脉作屏障, 成为第三纪植物的“避难所”.海拔672-1751 m.雨量充沛, 气候温和, 年平均气温12.8℃, 年降水量1200-1400 mm, 无霜期234天, 相对湿度85%左右.土壤以山地黄壤、黄棕壤、棕壤为主(艾训儒和谭建锡, 1999 ; 方元平和刘胜祥, 2006 ). ...
星斗山自然保护区珙桐种群结构特征研究
1
1999
... 星斗山国家级自然保护区位于鄂西南恩施、利川与咸丰三县(市、州)交界处, 地理坐标29.95°- 30.17° N, 108.95°-109.45° E.是齐岳山向东北的延伸部分, 因有大巴山系巫山余脉作屏障, 成为第三纪植物的“避难所”.海拔672-1751 m.雨量充沛, 气候温和, 年平均气温12.8℃, 年降水量1200-1400 mm, 无霜期234天, 相对湿度85%左右.土壤以山地黄壤、黄棕壤、棕壤为主(艾训儒和谭建锡, 1999 ; 方元平和刘胜祥, 2006 ). ...
中国北亚热带次生森林植被研究述评
2
1998
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
... )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
中国北亚热带次生森林植被研究述评
2
1998
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
... )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
Relative growth rate in phylogenetically related deciduous and evergreen woody species.
1
2001
... 除群落III外, 各群落类型小径级(1-5 cm)内落叶物种的物种丰富度高于常绿物种, 但多度、胸高断面积和重要值却表现为落叶物种小于常绿物种, 这主要与常绿和落叶物种的生活型属性有关.研究区群落内小径级个体的生活型主要是小乔木和灌木, 而大乔木个体密度相对较低.这些小乔木和灌木物种以常绿树种为主, 因此出现了小径级内落叶物种丰富度高但多度、胸高断面积和重要值低的现象.较高强度的干扰能够提高落叶树种在群落中的比例(丁易和臧润国, 2008 ), 群落III的小径级在各个指标上都是落叶树种占优势, 这可能说明人为干扰和环境条件显著影响了该群落的更新和发展方向, 导致该群落在未来一段时间内都将以落叶物种为主.大径级(≥10 cm)的落叶物种在多数群落内的物种丰富度、多度、胸高断面积和重要值均大于常绿物种, 表明落叶树种在林冠层占据显著的优势, Chabot和Hicks (1982) 的研究也发现, 光照条件较好的林冠层落叶物种更多.林冠层落叶树种占据优势能够显著影响林下光照条件, 从而影响森林更新和物种多样性(Quigley & Platt, 2003 ).落叶物种具有更高的资源养分利用效率(Kloeppel et al ., 2000 ), 能够利用夏季良好的水热条件迅速获得碳水化合物完成高生长, 在与常绿树种的竞争过程中居于优势地位(Antúnez et al ., 2001 ).同时, 落叶物种采取了落叶的策略来适应研究区的冬季低温胁迫, 而常绿物种则保留叶片越冬, 低温可能损害常绿树种的气孔和光合速率而使常绿植物生长受到严重制约(Wyka & Oleksyn, 2014 ), 且较大的季节性温差所引起的冰凌和雪灾也可能造成常绿植物受到伤害从而限制其在林冠层的优势(白坤栋等, 2010 ).林下的常绿树木由于林冠层的郁闭而受到光强的限制.尽管在林冠层落叶期间(晚秋到早春), 林下光照条件可以由于林冠层的落叶而得到改善, 但低温限制了林下常绿树木的生长.这种竞争最终形成了林冠层以落叶阔叶树为主和林下层以常绿树种为主的格局. ...
哀牢山和猫儿山中山常绿和落叶阔叶树光合特性对季节温度变化的响应
2
2010
... 由于叶片是植物光合作用的主要场所, 因而叶片的物候过程对于群落外貌、林下环境、凋落物分解和生态系统生产力具有重要作用(Bohlman et al ., 1998 ; Quigley & Platt, 2003 ).落叶是物种适应环境胁迫的一种重要策略, 通过落叶可以回避不利生境对树木生长造成的伤害和不利影响(Poorter & Markesteijn, 2008 ), 低温和水分胁迫是阔叶树种落叶的重要原因(Lechowicz, 1984 ; Reich et al ., 2004 ).与热带部分地区树种因土壤水分亏缺而出现旱季落叶的情况不同(Williams et al ., 2008 ; de Faria Lopes et al ., 2012 ), 亚热带常绿落叶阔叶混交林的形成更多受到海拔升高而形成的低温胁迫的影响(Oliveira- Filho et al ., 2006 ; 白坤栋等, 2010 ; 宋坤, 2012 ).回归分析中土壤含水量与常绿和落叶物种分布并无显著的相关性, 这个结果证明了研究区常绿和落叶物种的变化与土壤水分无关.温度是限制植物组成、生长、繁殖和分布的重要因子(Woodward, 1987 ).研究地区冬季气温较低, 一些喜温且生长迅速竞争能力强的常绿阔叶树种受到限制, 但绝对低温尚较高, 因此较耐寒的常绿阔叶树与落叶阔叶树均能生长, 最终在该地段形成了亚热带常绿落叶阔叶混交林(吴征镒, 1980 ). ...
... 除群落III外, 各群落类型小径级(1-5 cm)内落叶物种的物种丰富度高于常绿物种, 但多度、胸高断面积和重要值却表现为落叶物种小于常绿物种, 这主要与常绿和落叶物种的生活型属性有关.研究区群落内小径级个体的生活型主要是小乔木和灌木, 而大乔木个体密度相对较低.这些小乔木和灌木物种以常绿树种为主, 因此出现了小径级内落叶物种丰富度高但多度、胸高断面积和重要值低的现象.较高强度的干扰能够提高落叶树种在群落中的比例(丁易和臧润国, 2008 ), 群落III的小径级在各个指标上都是落叶树种占优势, 这可能说明人为干扰和环境条件显著影响了该群落的更新和发展方向, 导致该群落在未来一段时间内都将以落叶物种为主.大径级(≥10 cm)的落叶物种在多数群落内的物种丰富度、多度、胸高断面积和重要值均大于常绿物种, 表明落叶树种在林冠层占据显著的优势, Chabot和Hicks (1982) 的研究也发现, 光照条件较好的林冠层落叶物种更多.林冠层落叶树种占据优势能够显著影响林下光照条件, 从而影响森林更新和物种多样性(Quigley & Platt, 2003 ).落叶物种具有更高的资源养分利用效率(Kloeppel et al ., 2000 ), 能够利用夏季良好的水热条件迅速获得碳水化合物完成高生长, 在与常绿树种的竞争过程中居于优势地位(Antúnez et al ., 2001 ).同时, 落叶物种采取了落叶的策略来适应研究区的冬季低温胁迫, 而常绿物种则保留叶片越冬, 低温可能损害常绿树种的气孔和光合速率而使常绿植物生长受到严重制约(Wyka & Oleksyn, 2014 ), 且较大的季节性温差所引起的冰凌和雪灾也可能造成常绿植物受到伤害从而限制其在林冠层的优势(白坤栋等, 2010 ).林下的常绿树木由于林冠层的郁闭而受到光强的限制.尽管在林冠层落叶期间(晚秋到早春), 林下光照条件可以由于林冠层的落叶而得到改善, 但低温限制了林下常绿树木的生长.这种竞争最终形成了林冠层以落叶阔叶树为主和林下层以常绿树种为主的格局. ...
哀牢山和猫儿山中山常绿和落叶阔叶树光合特性对季节温度变化的响应
2
2010
... 由于叶片是植物光合作用的主要场所, 因而叶片的物候过程对于群落外貌、林下环境、凋落物分解和生态系统生产力具有重要作用(Bohlman et al ., 1998 ; Quigley & Platt, 2003 ).落叶是物种适应环境胁迫的一种重要策略, 通过落叶可以回避不利生境对树木生长造成的伤害和不利影响(Poorter & Markesteijn, 2008 ), 低温和水分胁迫是阔叶树种落叶的重要原因(Lechowicz, 1984 ; Reich et al ., 2004 ).与热带部分地区树种因土壤水分亏缺而出现旱季落叶的情况不同(Williams et al ., 2008 ; de Faria Lopes et al ., 2012 ), 亚热带常绿落叶阔叶混交林的形成更多受到海拔升高而形成的低温胁迫的影响(Oliveira- Filho et al ., 2006 ; 白坤栋等, 2010 ; 宋坤, 2012 ).回归分析中土壤含水量与常绿和落叶物种分布并无显著的相关性, 这个结果证明了研究区常绿和落叶物种的变化与土壤水分无关.温度是限制植物组成、生长、繁殖和分布的重要因子(Woodward, 1987 ).研究地区冬季气温较低, 一些喜温且生长迅速竞争能力强的常绿阔叶树种受到限制, 但绝对低温尚较高, 因此较耐寒的常绿阔叶树与落叶阔叶树均能生长, 最终在该地段形成了亚热带常绿落叶阔叶混交林(吴征镒, 1980 ). ...
... 除群落III外, 各群落类型小径级(1-5 cm)内落叶物种的物种丰富度高于常绿物种, 但多度、胸高断面积和重要值却表现为落叶物种小于常绿物种, 这主要与常绿和落叶物种的生活型属性有关.研究区群落内小径级个体的生活型主要是小乔木和灌木, 而大乔木个体密度相对较低.这些小乔木和灌木物种以常绿树种为主, 因此出现了小径级内落叶物种丰富度高但多度、胸高断面积和重要值低的现象.较高强度的干扰能够提高落叶树种在群落中的比例(丁易和臧润国, 2008 ), 群落III的小径级在各个指标上都是落叶树种占优势, 这可能说明人为干扰和环境条件显著影响了该群落的更新和发展方向, 导致该群落在未来一段时间内都将以落叶物种为主.大径级(≥10 cm)的落叶物种在多数群落内的物种丰富度、多度、胸高断面积和重要值均大于常绿物种, 表明落叶树种在林冠层占据显著的优势, Chabot和Hicks (1982) 的研究也发现, 光照条件较好的林冠层落叶物种更多.林冠层落叶树种占据优势能够显著影响林下光照条件, 从而影响森林更新和物种多样性(Quigley & Platt, 2003 ).落叶物种具有更高的资源养分利用效率(Kloeppel et al ., 2000 ), 能够利用夏季良好的水热条件迅速获得碳水化合物完成高生长, 在与常绿树种的竞争过程中居于优势地位(Antúnez et al ., 2001 ).同时, 落叶物种采取了落叶的策略来适应研究区的冬季低温胁迫, 而常绿物种则保留叶片越冬, 低温可能损害常绿树种的气孔和光合速率而使常绿植物生长受到严重制约(Wyka & Oleksyn, 2014 ), 且较大的季节性温差所引起的冰凌和雪灾也可能造成常绿植物受到伤害从而限制其在林冠层的优势(白坤栋等, 2010 ).林下的常绿树木由于林冠层的郁闭而受到光强的限制.尽管在林冠层落叶期间(晚秋到早春), 林下光照条件可以由于林冠层的落叶而得到改善, 但低温限制了林下常绿树木的生长.这种竞争最终形成了林冠层以落叶阔叶树为主和林下层以常绿树种为主的格局. ...
1
1995
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
1
1995
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
Seasonal foliage changes in the Eastern Amazon Basin detected from landsat thematic mapper satellite images.
1
1998
... 由于叶片是植物光合作用的主要场所, 因而叶片的物候过程对于群落外貌、林下环境、凋落物分解和生态系统生产力具有重要作用(Bohlman et al ., 1998 ; Quigley & Platt, 2003 ).落叶是物种适应环境胁迫的一种重要策略, 通过落叶可以回避不利生境对树木生长造成的伤害和不利影响(Poorter & Markesteijn, 2008 ), 低温和水分胁迫是阔叶树种落叶的重要原因(Lechowicz, 1984 ; Reich et al ., 2004 ).与热带部分地区树种因土壤水分亏缺而出现旱季落叶的情况不同(Williams et al ., 2008 ; de Faria Lopes et al ., 2012 ), 亚热带常绿落叶阔叶混交林的形成更多受到海拔升高而形成的低温胁迫的影响(Oliveira- Filho et al ., 2006 ; 白坤栋等, 2010 ; 宋坤, 2012 ).回归分析中土壤含水量与常绿和落叶物种分布并无显著的相关性, 这个结果证明了研究区常绿和落叶物种的变化与土壤水分无关.温度是限制植物组成、生长、繁殖和分布的重要因子(Woodward, 1987 ).研究地区冬季气温较低, 一些喜温且生长迅速竞争能力强的常绿阔叶树种受到限制, 但绝对低温尚较高, 因此较耐寒的常绿阔叶树与落叶阔叶树均能生长, 最终在该地段形成了亚热带常绿落叶阔叶混交林(吴征镒, 1980 ). ...
The intermediate disturbance hypothesis applies to tropical forests, but disturbance contributes little to tree diversity.
1
2009
... 从物种组成和群落结构来看, 研究区各群落类型常绿和落叶物种存在一定的差异.群落I和群落IV的常绿和落叶物种在群落内的差异多不显著或者常绿物种大于落叶物种, 而群落II、群落III和群落V则表现为落叶物种大于常绿物种.造成这种差异的主要原因可能是环境差异和人为干扰.环境是影响物种组成的决定性因素, 环境因子组成的变化产生了不同的生境, 从而引起不同地带及区域的物种差异(Toledo et al ., 2011 ).而人为干扰能够显著地改变群落结构和物种组成, 从而影响群落的发展方向(Lomolino, 2001 ; Haberl et al ., 2007 ; Nogués-Bravo et al ., 2008 ).群落III位于海拔较高的阳坡, 随着海拔的升高, 气温逐渐降低, 光照强度增加, 较低的气温和阳坡较好的光照条件促进落叶物种的生长(Malhi et al ., 2010 ).充足的光照也促进土壤种子库萌发, 使群落内幼苗和幼树的密度较大.同时, 群落III位于保护区森林防火隔离带附近, 较频繁地受到了人为的干扰, 干扰提高了林下的光照条件, 导致群落内喜光的落叶树种增加和耐阴的常绿树种减少(Hawthorne, 1996 ; Bongers et al ., 2009 ).群落II和群落V分布于低海拔地区, 长期受到人类樵采等活动的影响, 林下的光照条件较好, 群落内的落叶物种大于常绿物种.群落I和群落IV主要位于阴坡, 且人为干扰较少, 林内光照条件较差, 适宜常绿物种的生长. ...
Controls on nitrogen cycling in terrestrial ecosystems: A synthetic analysis of literature data.
1
2005
... 除地形因子和人为干扰之外, 土壤养分也对常绿和落叶物种比例的变化产生了一定的影响.落叶物种分布区的土壤氮含量常被认为高于常绿物种分布区, 其原因是落叶物种比常绿物种具有更高的氮矿化速率(Aerts & Chapin, 1999 ; Cornwell et al ., 2008 ), 然而也有研究表明常绿和落叶物种的氮矿化速率并无显著差异(Reich et al ., 2001 ; Booth et al ., 2005 ).本研究发现土壤氮含量在多数群落内与常绿物种丰富度显著正相关, 这可能与不同群落类型的物种属性及真菌差异有关(Mueller et al ., 2012 ).总体来看, 研究区常绿物种更易受土壤养分的影响.关于土壤养分对各群落内常绿和落叶物种分布的影响有待进一步深入分析. ...
湖南八大公山亮叶水青冈群落物种多样性的研究
1
1997
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
湖南八大公山亮叶水青冈群落物种多样性的研究
1
1997
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
The ecology of leaf life spans.
1
1982
... 除群落III外, 各群落类型小径级(1-5 cm)内落叶物种的物种丰富度高于常绿物种, 但多度、胸高断面积和重要值却表现为落叶物种小于常绿物种, 这主要与常绿和落叶物种的生活型属性有关.研究区群落内小径级个体的生活型主要是小乔木和灌木, 而大乔木个体密度相对较低.这些小乔木和灌木物种以常绿树种为主, 因此出现了小径级内落叶物种丰富度高但多度、胸高断面积和重要值低的现象.较高强度的干扰能够提高落叶树种在群落中的比例(丁易和臧润国, 2008 ), 群落III的小径级在各个指标上都是落叶树种占优势, 这可能说明人为干扰和环境条件显著影响了该群落的更新和发展方向, 导致该群落在未来一段时间内都将以落叶物种为主.大径级(≥10 cm)的落叶物种在多数群落内的物种丰富度、多度、胸高断面积和重要值均大于常绿物种, 表明落叶树种在林冠层占据显著的优势, Chabot和Hicks (1982) 的研究也发现, 光照条件较好的林冠层落叶物种更多.林冠层落叶树种占据优势能够显著影响林下光照条件, 从而影响森林更新和物种多样性(Quigley & Platt, 2003 ).落叶物种具有更高的资源养分利用效率(Kloeppel et al ., 2000 ), 能够利用夏季良好的水热条件迅速获得碳水化合物完成高生长, 在与常绿树种的竞争过程中居于优势地位(Antúnez et al ., 2001 ).同时, 落叶物种采取了落叶的策略来适应研究区的冬季低温胁迫, 而常绿物种则保留叶片越冬, 低温可能损害常绿树种的气孔和光合速率而使常绿植物生长受到严重制约(Wyka & Oleksyn, 2014 ), 且较大的季节性温差所引起的冰凌和雪灾也可能造成常绿植物受到伤害从而限制其在林冠层的优势(白坤栋等, 2010 ).林下的常绿树木由于林冠层的郁闭而受到光强的限制.尽管在林冠层落叶期间(晚秋到早春), 林下光照条件可以由于林冠层的落叶而得到改善, 但低温限制了林下常绿树木的生长.这种竞争最终形成了林冠层以落叶阔叶树为主和林下层以常绿树种为主的格局. ...
Tropical Forest Census Plots: Methods and Results from Barro Colorado Island, Panama and a Comparison with Other Plots
1
1998
... 2013年9月-2014年6月, 在湖北星斗山自然保护区和湖北木林子自然保护区的典型天然森林植被中, 分别随机设置永久性森林动态样地45个和47个, 每个样地面积均为20 m × 20 m.这些样地分布于海拔1248-1800 m之间.样地建设采用CTFS森林动态样地监测标准(Condit, 1998 ).在每个样地中利用网格法将其分割成16个5 m × 5 m的小样方, 在小样方内对所有胸径≥1 cm的乔木、灌木和藤本进行编号、测量点刷漆标记并开展每木检尺, 记录内容包括物种名称、胸径、坐标、萌生与死亡状况, 同时记录样地所在坡度、海拔、凋落物厚度. ...
Plant species traits are the predominant control on litter decomposition rates within biomes worldwide.
1
2008
... 除地形因子和人为干扰之外, 土壤养分也对常绿和落叶物种比例的变化产生了一定的影响.落叶物种分布区的土壤氮含量常被认为高于常绿物种分布区, 其原因是落叶物种比常绿物种具有更高的氮矿化速率(Aerts & Chapin, 1999 ; Cornwell et al ., 2008 ), 然而也有研究表明常绿和落叶物种的氮矿化速率并无显著差异(Reich et al ., 2001 ; Booth et al ., 2005 ).本研究发现土壤氮含量在多数群落内与常绿物种丰富度显著正相关, 这可能与不同群落类型的物种属性及真菌差异有关(Mueller et al ., 2012 ).总体来看, 研究区常绿物种更易受土壤养分的影响.关于土壤养分对各群落内常绿和落叶物种分布的影响有待进一步深入分析. ...
Historical review of studies in seasonal semideciduous forests in Brazil: A perspective for conservation.
1
2012
... 由于叶片是植物光合作用的主要场所, 因而叶片的物候过程对于群落外貌、林下环境、凋落物分解和生态系统生产力具有重要作用(Bohlman et al ., 1998 ; Quigley & Platt, 2003 ).落叶是物种适应环境胁迫的一种重要策略, 通过落叶可以回避不利生境对树木生长造成的伤害和不利影响(Poorter & Markesteijn, 2008 ), 低温和水分胁迫是阔叶树种落叶的重要原因(Lechowicz, 1984 ; Reich et al ., 2004 ).与热带部分地区树种因土壤水分亏缺而出现旱季落叶的情况不同(Williams et al ., 2008 ; de Faria Lopes et al ., 2012 ), 亚热带常绿落叶阔叶混交林的形成更多受到海拔升高而形成的低温胁迫的影响(Oliveira- Filho et al ., 2006 ; 白坤栋等, 2010 ; 宋坤, 2012 ).回归分析中土壤含水量与常绿和落叶物种分布并无显著的相关性, 这个结果证明了研究区常绿和落叶物种的变化与土壤水分无关.温度是限制植物组成、生长、繁殖和分布的重要因子(Woodward, 1987 ).研究地区冬季气温较低, 一些喜温且生长迅速竞争能力强的常绿阔叶树种受到限制, 但绝对低温尚较高, 因此较耐寒的常绿阔叶树与落叶阔叶树均能生长, 最终在该地段形成了亚热带常绿落叶阔叶混交林(吴征镒, 1980 ). ...
海南岛热带低地雨林刀耕火种弃耕地恢复过程中落叶树种的变化
1
2008
... 除群落III外, 各群落类型小径级(1-5 cm)内落叶物种的物种丰富度高于常绿物种, 但多度、胸高断面积和重要值却表现为落叶物种小于常绿物种, 这主要与常绿和落叶物种的生活型属性有关.研究区群落内小径级个体的生活型主要是小乔木和灌木, 而大乔木个体密度相对较低.这些小乔木和灌木物种以常绿树种为主, 因此出现了小径级内落叶物种丰富度高但多度、胸高断面积和重要值低的现象.较高强度的干扰能够提高落叶树种在群落中的比例(丁易和臧润国, 2008 ), 群落III的小径级在各个指标上都是落叶树种占优势, 这可能说明人为干扰和环境条件显著影响了该群落的更新和发展方向, 导致该群落在未来一段时间内都将以落叶物种为主.大径级(≥10 cm)的落叶物种在多数群落内的物种丰富度、多度、胸高断面积和重要值均大于常绿物种, 表明落叶树种在林冠层占据显著的优势, Chabot和Hicks (1982) 的研究也发现, 光照条件较好的林冠层落叶物种更多.林冠层落叶树种占据优势能够显著影响林下光照条件, 从而影响森林更新和物种多样性(Quigley & Platt, 2003 ).落叶物种具有更高的资源养分利用效率(Kloeppel et al ., 2000 ), 能够利用夏季良好的水热条件迅速获得碳水化合物完成高生长, 在与常绿树种的竞争过程中居于优势地位(Antúnez et al ., 2001 ).同时, 落叶物种采取了落叶的策略来适应研究区的冬季低温胁迫, 而常绿物种则保留叶片越冬, 低温可能损害常绿树种的气孔和光合速率而使常绿植物生长受到严重制约(Wyka & Oleksyn, 2014 ), 且较大的季节性温差所引起的冰凌和雪灾也可能造成常绿植物受到伤害从而限制其在林冠层的优势(白坤栋等, 2010 ).林下的常绿树木由于林冠层的郁闭而受到光强的限制.尽管在林冠层落叶期间(晚秋到早春), 林下光照条件可以由于林冠层的落叶而得到改善, 但低温限制了林下常绿树木的生长.这种竞争最终形成了林冠层以落叶阔叶树为主和林下层以常绿树种为主的格局. ...
海南岛热带低地雨林刀耕火种弃耕地恢复过程中落叶树种的变化
1
2008
... 除群落III外, 各群落类型小径级(1-5 cm)内落叶物种的物种丰富度高于常绿物种, 但多度、胸高断面积和重要值却表现为落叶物种小于常绿物种, 这主要与常绿和落叶物种的生活型属性有关.研究区群落内小径级个体的生活型主要是小乔木和灌木, 而大乔木个体密度相对较低.这些小乔木和灌木物种以常绿树种为主, 因此出现了小径级内落叶物种丰富度高但多度、胸高断面积和重要值低的现象.较高强度的干扰能够提高落叶树种在群落中的比例(丁易和臧润国, 2008 ), 群落III的小径级在各个指标上都是落叶树种占优势, 这可能说明人为干扰和环境条件显著影响了该群落的更新和发展方向, 导致该群落在未来一段时间内都将以落叶物种为主.大径级(≥10 cm)的落叶物种在多数群落内的物种丰富度、多度、胸高断面积和重要值均大于常绿物种, 表明落叶树种在林冠层占据显著的优势, Chabot和Hicks (1982) 的研究也发现, 光照条件较好的林冠层落叶物种更多.林冠层落叶树种占据优势能够显著影响林下光照条件, 从而影响森林更新和物种多样性(Quigley & Platt, 2003 ).落叶物种具有更高的资源养分利用效率(Kloeppel et al ., 2000 ), 能够利用夏季良好的水热条件迅速获得碳水化合物完成高生长, 在与常绿树种的竞争过程中居于优势地位(Antúnez et al ., 2001 ).同时, 落叶物种采取了落叶的策略来适应研究区的冬季低温胁迫, 而常绿物种则保留叶片越冬, 低温可能损害常绿树种的气孔和光合速率而使常绿植物生长受到严重制约(Wyka & Oleksyn, 2014 ), 且较大的季节性温差所引起的冰凌和雪灾也可能造成常绿植物受到伤害从而限制其在林冠层的优势(白坤栋等, 2010 ).林下的常绿树木由于林冠层的郁闭而受到光强的限制.尽管在林冠层落叶期间(晚秋到早春), 林下光照条件可以由于林冠层的落叶而得到改善, 但低温限制了林下常绿树木的生长.这种竞争最终形成了林冠层以落叶阔叶树为主和林下层以常绿树种为主的格局. ...
也论我国东部植被带的划分
2
2001
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
... )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
也论我国东部植被带的划分
2
2001
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
... )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
湖北星斗山自然保护区国家重点保护野生植物调查研究
1
2006
... 星斗山国家级自然保护区位于鄂西南恩施、利川与咸丰三县(市、州)交界处, 地理坐标29.95°- 30.17° N, 108.95°-109.45° E.是齐岳山向东北的延伸部分, 因有大巴山系巫山余脉作屏障, 成为第三纪植物的“避难所”.海拔672-1751 m.雨量充沛, 气候温和, 年平均气温12.8℃, 年降水量1200-1400 mm, 无霜期234天, 相对湿度85%左右.土壤以山地黄壤、黄棕壤、棕壤为主(艾训儒和谭建锡, 1999 ; 方元平和刘胜祥, 2006 ). ...
湖北星斗山自然保护区国家重点保护野生植物调查研究
1
2006
... 星斗山国家级自然保护区位于鄂西南恩施、利川与咸丰三县(市、州)交界处, 地理坐标29.95°- 30.17° N, 108.95°-109.45° E.是齐岳山向东北的延伸部分, 因有大巴山系巫山余脉作屏障, 成为第三纪植物的“避难所”.海拔672-1751 m.雨量充沛, 气候温和, 年平均气温12.8℃, 年降水量1200-1400 mm, 无霜期234天, 相对湿度85%左右.土壤以山地黄壤、黄棕壤、棕壤为主(艾训儒和谭建锡, 1999 ; 方元平和刘胜祥, 2006 ). ...
1
2002
... 常绿和落叶物种的划分主要通过查阅《湖北植物志》(傅书遐, 2002 )及咨询植物分类专家来确定, 对于那些不明确的物种, 由作者在冬季进行实地观测和采访当地林业工作者获得. ...
1
2002
... 常绿和落叶物种的划分主要通过查阅《湖北植物志》(傅书遐, 2002 )及咨询植物分类专家来确定, 对于那些不明确的物种, 由作者在冬季进行实地观测和采访当地林业工作者获得. ...
1
2009
... 湖北木林子国家级自然保护区位于湖北省恩施土家族苗族自治州鹤峰县境内, 地理坐标29.21°- 30.17° N, 109.98°-110.81° E, 海拔1100.0-2095.6 m, 属武陵山脉北支余脉, 地势由西北和东南向中间逐渐倾斜.该区属于亚热带季风性湿润气候, 雨热同期, 时空分布不均, 年降水量1733 mm, 春秋多阴雨, 夏季雨量较多, 冬季雨少雾多, 蒸发小, 湿度大, 年平均相对湿度为82%, 无霜期270-279天.年平均气温为15.5 ℃, 最冷月(1月)平均气温为4.6 ℃, 最热月(7月)平均气温为26 ℃.土壤以黄棕壤、棕壤、黄壤为主(葛继稳等, 2009 ). ...
1
2009
... 湖北木林子国家级自然保护区位于湖北省恩施土家族苗族自治州鹤峰县境内, 地理坐标29.21°- 30.17° N, 109.98°-110.81° E, 海拔1100.0-2095.6 m, 属武陵山脉北支余脉, 地势由西北和东南向中间逐渐倾斜.该区属于亚热带季风性湿润气候, 雨热同期, 时空分布不均, 年降水量1733 mm, 春秋多阴雨, 夏季雨量较多, 冬季雨少雾多, 蒸发小, 湿度大, 年平均相对湿度为82%, 无霜期270-279天.年平均气温为15.5 ℃, 最冷月(1月)平均气温为4.6 ℃, 最热月(7月)平均气温为26 ℃.土壤以黄棕壤、棕壤、黄壤为主(葛继稳等, 2009 ). ...
Quantifying and mapping the human appropriation of net primary production in earth’s terrestrial ecosystems.
1
2007
... 从物种组成和群落结构来看, 研究区各群落类型常绿和落叶物种存在一定的差异.群落I和群落IV的常绿和落叶物种在群落内的差异多不显著或者常绿物种大于落叶物种, 而群落II、群落III和群落V则表现为落叶物种大于常绿物种.造成这种差异的主要原因可能是环境差异和人为干扰.环境是影响物种组成的决定性因素, 环境因子组成的变化产生了不同的生境, 从而引起不同地带及区域的物种差异(Toledo et al ., 2011 ).而人为干扰能够显著地改变群落结构和物种组成, 从而影响群落的发展方向(Lomolino, 2001 ; Haberl et al ., 2007 ; Nogués-Bravo et al ., 2008 ).群落III位于海拔较高的阳坡, 随着海拔的升高, 气温逐渐降低, 光照强度增加, 较低的气温和阳坡较好的光照条件促进落叶物种的生长(Malhi et al ., 2010 ).充足的光照也促进土壤种子库萌发, 使群落内幼苗和幼树的密度较大.同时, 群落III位于保护区森林防火隔离带附近, 较频繁地受到了人为的干扰, 干扰提高了林下的光照条件, 导致群落内喜光的落叶树种增加和耐阴的常绿树种减少(Hawthorne, 1996 ; Bongers et al ., 2009 ).群落II和群落V分布于低海拔地区, 长期受到人类樵采等活动的影响, 林下的光照条件较好, 群落内的落叶物种大于常绿物种.群落I和群落IV主要位于阴坡, 且人为干扰较少, 林内光照条件较差, 适宜常绿物种的生长. ...
桂西北喀斯特区常绿落叶阔叶混交林种群种间联结性与相关性
1
2009
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
桂西北喀斯特区常绿落叶阔叶混交林种群种间联结性与相关性
1
2009
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
Holes and the sums of parts in Ghanaian forest: Regeneration, scale and sustainable use. Proceedings of the Royal Society of Edinburgh. Section B.
1
1996
... 从物种组成和群落结构来看, 研究区各群落类型常绿和落叶物种存在一定的差异.群落I和群落IV的常绿和落叶物种在群落内的差异多不显著或者常绿物种大于落叶物种, 而群落II、群落III和群落V则表现为落叶物种大于常绿物种.造成这种差异的主要原因可能是环境差异和人为干扰.环境是影响物种组成的决定性因素, 环境因子组成的变化产生了不同的生境, 从而引起不同地带及区域的物种差异(Toledo et al ., 2011 ).而人为干扰能够显著地改变群落结构和物种组成, 从而影响群落的发展方向(Lomolino, 2001 ; Haberl et al ., 2007 ; Nogués-Bravo et al ., 2008 ).群落III位于海拔较高的阳坡, 随着海拔的升高, 气温逐渐降低, 光照强度增加, 较低的气温和阳坡较好的光照条件促进落叶物种的生长(Malhi et al ., 2010 ).充足的光照也促进土壤种子库萌发, 使群落内幼苗和幼树的密度较大.同时, 群落III位于保护区森林防火隔离带附近, 较频繁地受到了人为的干扰, 干扰提高了林下的光照条件, 导致群落内喜光的落叶树种增加和耐阴的常绿树种减少(Hawthorne, 1996 ; Bongers et al ., 2009 ).群落II和群落V分布于低海拔地区, 长期受到人类樵采等活动的影响, 林下的光照条件较好, 群落内的落叶物种大于常绿物种.群落I和群落IV主要位于阴坡, 且人为干扰较少, 林内光照条件较差, 适宜常绿物种的生长. ...
Leaf- level resource use for evergreen and deciduous conifers along a resource availability gradient.
1
2000
... 除群落III外, 各群落类型小径级(1-5 cm)内落叶物种的物种丰富度高于常绿物种, 但多度、胸高断面积和重要值却表现为落叶物种小于常绿物种, 这主要与常绿和落叶物种的生活型属性有关.研究区群落内小径级个体的生活型主要是小乔木和灌木, 而大乔木个体密度相对较低.这些小乔木和灌木物种以常绿树种为主, 因此出现了小径级内落叶物种丰富度高但多度、胸高断面积和重要值低的现象.较高强度的干扰能够提高落叶树种在群落中的比例(丁易和臧润国, 2008 ), 群落III的小径级在各个指标上都是落叶树种占优势, 这可能说明人为干扰和环境条件显著影响了该群落的更新和发展方向, 导致该群落在未来一段时间内都将以落叶物种为主.大径级(≥10 cm)的落叶物种在多数群落内的物种丰富度、多度、胸高断面积和重要值均大于常绿物种, 表明落叶树种在林冠层占据显著的优势, Chabot和Hicks (1982) 的研究也发现, 光照条件较好的林冠层落叶物种更多.林冠层落叶树种占据优势能够显著影响林下光照条件, 从而影响森林更新和物种多样性(Quigley & Platt, 2003 ).落叶物种具有更高的资源养分利用效率(Kloeppel et al ., 2000 ), 能够利用夏季良好的水热条件迅速获得碳水化合物完成高生长, 在与常绿树种的竞争过程中居于优势地位(Antúnez et al ., 2001 ).同时, 落叶物种采取了落叶的策略来适应研究区的冬季低温胁迫, 而常绿物种则保留叶片越冬, 低温可能损害常绿树种的气孔和光合速率而使常绿植物生长受到严重制约(Wyka & Oleksyn, 2014 ), 且较大的季节性温差所引起的冰凌和雪灾也可能造成常绿植物受到伤害从而限制其在林冠层的优势(白坤栋等, 2010 ).林下的常绿树木由于林冠层的郁闭而受到光强的限制.尽管在林冠层落叶期间(晚秋到早春), 林下光照条件可以由于林冠层的落叶而得到改善, 但低温限制了林下常绿树木的生长.这种竞争最终形成了林冠层以落叶阔叶树为主和林下层以常绿树种为主的格局. ...
基于多元回归树的常绿阔叶林群丛数量分类——以古田山24公顷森林样地为例
1
2010
... 植被分类是植被生态学研究中最复杂的一个问题, 直到现在还没有植被学家们认同的统一分类原则和分类系统(宋永昌, 2004 ).而我国亚热带地区植被具有复杂多样、次生性强和过渡类型多等特点, 更增加了该地区植被分类的困难(赖江山等, 2010 ).传统的植物群落分类多根据群落的外貌特征及群落的种类组成划分群落类型, 个人经验对分类结果的影响比较大(刘海江和郭柯, 2003 ; 刘万德等, 2010 ).随着植被研究的精度要求不断提高, 提供客观归类和划分结果的数量分类方法在植被分类中越来越重要(袁秀等, 2013 ).目前数量生态学中的分类和排序方法已成为现代植被生态学研究中必不可少的手段, 双向指示种分析法(TWINSPAN)和除趋势对应分析法(DCA)已广泛应用于森林和草地等植物群落分析(Ozinga et al ., 2005 ; 孙菊等, 2009 ; 廉凯敏等, 2015 ).本研究也证实了TWINSPAN分类与DCA排序这两种方法相结合应用于亚热带常绿落叶阔叶混交林植被数量分析是可行的, 而且可以相互补充和交叉检验. ...
基于多元回归树的常绿阔叶林群丛数量分类——以古田山24公顷森林样地为例
1
2010
... 植被分类是植被生态学研究中最复杂的一个问题, 直到现在还没有植被学家们认同的统一分类原则和分类系统(宋永昌, 2004 ).而我国亚热带地区植被具有复杂多样、次生性强和过渡类型多等特点, 更增加了该地区植被分类的困难(赖江山等, 2010 ).传统的植物群落分类多根据群落的外貌特征及群落的种类组成划分群落类型, 个人经验对分类结果的影响比较大(刘海江和郭柯, 2003 ; 刘万德等, 2010 ).随着植被研究的精度要求不断提高, 提供客观归类和划分结果的数量分类方法在植被分类中越来越重要(袁秀等, 2013 ).目前数量生态学中的分类和排序方法已成为现代植被生态学研究中必不可少的手段, 双向指示种分析法(TWINSPAN)和除趋势对应分析法(DCA)已广泛应用于森林和草地等植物群落分析(Ozinga et al ., 2005 ; 孙菊等, 2009 ; 廉凯敏等, 2015 ).本研究也证实了TWINSPAN分类与DCA排序这两种方法相结合应用于亚热带常绿落叶阔叶混交林植被数量分析是可行的, 而且可以相互补充和交叉检验. ...
Why do temperate deciduous trees leaf out at different times? Adaptation and ecology of forest communities.
1
1984
... 由于叶片是植物光合作用的主要场所, 因而叶片的物候过程对于群落外貌、林下环境、凋落物分解和生态系统生产力具有重要作用(Bohlman et al ., 1998 ; Quigley & Platt, 2003 ).落叶是物种适应环境胁迫的一种重要策略, 通过落叶可以回避不利生境对树木生长造成的伤害和不利影响(Poorter & Markesteijn, 2008 ), 低温和水分胁迫是阔叶树种落叶的重要原因(Lechowicz, 1984 ; Reich et al ., 2004 ).与热带部分地区树种因土壤水分亏缺而出现旱季落叶的情况不同(Williams et al ., 2008 ; de Faria Lopes et al ., 2012 ), 亚热带常绿落叶阔叶混交林的形成更多受到海拔升高而形成的低温胁迫的影响(Oliveira- Filho et al ., 2006 ; 白坤栋等, 2010 ; 宋坤, 2012 ).回归分析中土壤含水量与常绿和落叶物种分布并无显著的相关性, 这个结果证明了研究区常绿和落叶物种的变化与土壤水分无关.温度是限制植物组成、生长、繁殖和分布的重要因子(Woodward, 1987 ).研究地区冬季气温较低, 一些喜温且生长迅速竞争能力强的常绿阔叶树种受到限制, 但绝对低温尚较高, 因此较耐寒的常绿阔叶树与落叶阔叶树均能生长, 最终在该地段形成了亚热带常绿落叶阔叶混交林(吴征镒, 1980 ). ...
太宽河自然保护区板栗群落数量分类与排序
1
2015
... 植被分类是植被生态学研究中最复杂的一个问题, 直到现在还没有植被学家们认同的统一分类原则和分类系统(宋永昌, 2004 ).而我国亚热带地区植被具有复杂多样、次生性强和过渡类型多等特点, 更增加了该地区植被分类的困难(赖江山等, 2010 ).传统的植物群落分类多根据群落的外貌特征及群落的种类组成划分群落类型, 个人经验对分类结果的影响比较大(刘海江和郭柯, 2003 ; 刘万德等, 2010 ).随着植被研究的精度要求不断提高, 提供客观归类和划分结果的数量分类方法在植被分类中越来越重要(袁秀等, 2013 ).目前数量生态学中的分类和排序方法已成为现代植被生态学研究中必不可少的手段, 双向指示种分析法(TWINSPAN)和除趋势对应分析法(DCA)已广泛应用于森林和草地等植物群落分析(Ozinga et al ., 2005 ; 孙菊等, 2009 ; 廉凯敏等, 2015 ).本研究也证实了TWINSPAN分类与DCA排序这两种方法相结合应用于亚热带常绿落叶阔叶混交林植被数量分析是可行的, 而且可以相互补充和交叉检验. ...
太宽河自然保护区板栗群落数量分类与排序
1
2015
... 植被分类是植被生态学研究中最复杂的一个问题, 直到现在还没有植被学家们认同的统一分类原则和分类系统(宋永昌, 2004 ).而我国亚热带地区植被具有复杂多样、次生性强和过渡类型多等特点, 更增加了该地区植被分类的困难(赖江山等, 2010 ).传统的植物群落分类多根据群落的外貌特征及群落的种类组成划分群落类型, 个人经验对分类结果的影响比较大(刘海江和郭柯, 2003 ; 刘万德等, 2010 ).随着植被研究的精度要求不断提高, 提供客观归类和划分结果的数量分类方法在植被分类中越来越重要(袁秀等, 2013 ).目前数量生态学中的分类和排序方法已成为现代植被生态学研究中必不可少的手段, 双向指示种分析法(TWINSPAN)和除趋势对应分析法(DCA)已广泛应用于森林和草地等植物群落分析(Ozinga et al ., 2005 ; 孙菊等, 2009 ; 廉凯敏等, 2015 ).本研究也证实了TWINSPAN分类与DCA排序这两种方法相结合应用于亚热带常绿落叶阔叶混交林植被数量分析是可行的, 而且可以相互补充和交叉检验. ...
Multiple regression on distance matrices: A multivariate spatial analysis tool.
1
2007
... 以样地为单位分别计算物种的重要值, 计算公式为: 重要值(IV ) = (相对多度+相对显著度+相对频度)/3.采用VEAPAN软件包中的TWINSPAN进行群落分类; 通过CANOCO 4.5软件(张金屯, 1995 )对样地和物种的重要值矩阵进行DCA排序.根据个体胸径(DBH )将物种划分为3个等级: 小径级(1 cm ≤ DBH < 5 cm)、中径级(5 cm ≤ DBH < 10 cm)和大径级(DBH ≥ 10 cm).基于Bray-Curtis相似距离, 采用基于矩阵的多元回归分析(Permutation-based multiple regression on distance matrices, MRM)确定显著影响群落物种组成变化的环境因子(Lichstein, 2007 ).运用多元逐步回归分析(multiple regression analysis)来分析常绿和落叶物种与环境因子之间的关系. ...
八大公山常绿落叶阔叶混交林根萌能力
1
2014
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
八大公山常绿落叶阔叶混交林根萌能力
1
2014
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
浑善达克沙地丘间低地植物群落的分类与排序
1
2003
... 植被分类是植被生态学研究中最复杂的一个问题, 直到现在还没有植被学家们认同的统一分类原则和分类系统(宋永昌, 2004 ).而我国亚热带地区植被具有复杂多样、次生性强和过渡类型多等特点, 更增加了该地区植被分类的困难(赖江山等, 2010 ).传统的植物群落分类多根据群落的外貌特征及群落的种类组成划分群落类型, 个人经验对分类结果的影响比较大(刘海江和郭柯, 2003 ; 刘万德等, 2010 ).随着植被研究的精度要求不断提高, 提供客观归类和划分结果的数量分类方法在植被分类中越来越重要(袁秀等, 2013 ).目前数量生态学中的分类和排序方法已成为现代植被生态学研究中必不可少的手段, 双向指示种分析法(TWINSPAN)和除趋势对应分析法(DCA)已广泛应用于森林和草地等植物群落分析(Ozinga et al ., 2005 ; 孙菊等, 2009 ; 廉凯敏等, 2015 ).本研究也证实了TWINSPAN分类与DCA排序这两种方法相结合应用于亚热带常绿落叶阔叶混交林植被数量分析是可行的, 而且可以相互补充和交叉检验. ...
浑善达克沙地丘间低地植物群落的分类与排序
1
2003
... 植被分类是植被生态学研究中最复杂的一个问题, 直到现在还没有植被学家们认同的统一分类原则和分类系统(宋永昌, 2004 ).而我国亚热带地区植被具有复杂多样、次生性强和过渡类型多等特点, 更增加了该地区植被分类的困难(赖江山等, 2010 ).传统的植物群落分类多根据群落的外貌特征及群落的种类组成划分群落类型, 个人经验对分类结果的影响比较大(刘海江和郭柯, 2003 ; 刘万德等, 2010 ).随着植被研究的精度要求不断提高, 提供客观归类和划分结果的数量分类方法在植被分类中越来越重要(袁秀等, 2013 ).目前数量生态学中的分类和排序方法已成为现代植被生态学研究中必不可少的手段, 双向指示种分析法(TWINSPAN)和除趋势对应分析法(DCA)已广泛应用于森林和草地等植物群落分析(Ozinga et al ., 2005 ; 孙菊等, 2009 ; 廉凯敏等, 2015 ).本研究也证实了TWINSPAN分类与DCA排序这两种方法相结合应用于亚热带常绿落叶阔叶混交林植被数量分析是可行的, 而且可以相互补充和交叉检验. ...
海南岛霸王岭林区低海拔热带林群落数量分类与排序
1
2010
... 植被分类是植被生态学研究中最复杂的一个问题, 直到现在还没有植被学家们认同的统一分类原则和分类系统(宋永昌, 2004 ).而我国亚热带地区植被具有复杂多样、次生性强和过渡类型多等特点, 更增加了该地区植被分类的困难(赖江山等, 2010 ).传统的植物群落分类多根据群落的外貌特征及群落的种类组成划分群落类型, 个人经验对分类结果的影响比较大(刘海江和郭柯, 2003 ; 刘万德等, 2010 ).随着植被研究的精度要求不断提高, 提供客观归类和划分结果的数量分类方法在植被分类中越来越重要(袁秀等, 2013 ).目前数量生态学中的分类和排序方法已成为现代植被生态学研究中必不可少的手段, 双向指示种分析法(TWINSPAN)和除趋势对应分析法(DCA)已广泛应用于森林和草地等植物群落分析(Ozinga et al ., 2005 ; 孙菊等, 2009 ; 廉凯敏等, 2015 ).本研究也证实了TWINSPAN分类与DCA排序这两种方法相结合应用于亚热带常绿落叶阔叶混交林植被数量分析是可行的, 而且可以相互补充和交叉检验. ...
海南岛霸王岭林区低海拔热带林群落数量分类与排序
1
2010
... 植被分类是植被生态学研究中最复杂的一个问题, 直到现在还没有植被学家们认同的统一分类原则和分类系统(宋永昌, 2004 ).而我国亚热带地区植被具有复杂多样、次生性强和过渡类型多等特点, 更增加了该地区植被分类的困难(赖江山等, 2010 ).传统的植物群落分类多根据群落的外貌特征及群落的种类组成划分群落类型, 个人经验对分类结果的影响比较大(刘海江和郭柯, 2003 ; 刘万德等, 2010 ).随着植被研究的精度要求不断提高, 提供客观归类和划分结果的数量分类方法在植被分类中越来越重要(袁秀等, 2013 ).目前数量生态学中的分类和排序方法已成为现代植被生态学研究中必不可少的手段, 双向指示种分析法(TWINSPAN)和除趋势对应分析法(DCA)已广泛应用于森林和草地等植物群落分析(Ozinga et al ., 2005 ; 孙菊等, 2009 ; 廉凯敏等, 2015 ).本研究也证实了TWINSPAN分类与DCA排序这两种方法相结合应用于亚热带常绿落叶阔叶混交林植被数量分析是可行的, 而且可以相互补充和交叉检验. ...
Elevation gradients of species-density: Historical and prospective views.
1
2001
... 从物种组成和群落结构来看, 研究区各群落类型常绿和落叶物种存在一定的差异.群落I和群落IV的常绿和落叶物种在群落内的差异多不显著或者常绿物种大于落叶物种, 而群落II、群落III和群落V则表现为落叶物种大于常绿物种.造成这种差异的主要原因可能是环境差异和人为干扰.环境是影响物种组成的决定性因素, 环境因子组成的变化产生了不同的生境, 从而引起不同地带及区域的物种差异(Toledo et al ., 2011 ).而人为干扰能够显著地改变群落结构和物种组成, 从而影响群落的发展方向(Lomolino, 2001 ; Haberl et al ., 2007 ; Nogués-Bravo et al ., 2008 ).群落III位于海拔较高的阳坡, 随着海拔的升高, 气温逐渐降低, 光照强度增加, 较低的气温和阳坡较好的光照条件促进落叶物种的生长(Malhi et al ., 2010 ).充足的光照也促进土壤种子库萌发, 使群落内幼苗和幼树的密度较大.同时, 群落III位于保护区森林防火隔离带附近, 较频繁地受到了人为的干扰, 干扰提高了林下的光照条件, 导致群落内喜光的落叶树种增加和耐阴的常绿树种减少(Hawthorne, 1996 ; Bongers et al ., 2009 ).群落II和群落V分布于低海拔地区, 长期受到人类樵采等活动的影响, 林下的光照条件较好, 群落内的落叶物种大于常绿物种.群落I和群落IV主要位于阴坡, 且人为干扰较少, 林内光照条件较差, 适宜常绿物种的生长. ...
星斗山国家级自然保护区外来入侵植物初步研究
1
2005
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
星斗山国家级自然保护区外来入侵植物初步研究
1
2005
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
Introduction: Elevation gradients in the tropics: Laboratories for ecosystem ecology and global change research.
1
2010
... 从物种组成和群落结构来看, 研究区各群落类型常绿和落叶物种存在一定的差异.群落I和群落IV的常绿和落叶物种在群落内的差异多不显著或者常绿物种大于落叶物种, 而群落II、群落III和群落V则表现为落叶物种大于常绿物种.造成这种差异的主要原因可能是环境差异和人为干扰.环境是影响物种组成的决定性因素, 环境因子组成的变化产生了不同的生境, 从而引起不同地带及区域的物种差异(Toledo et al ., 2011 ).而人为干扰能够显著地改变群落结构和物种组成, 从而影响群落的发展方向(Lomolino, 2001 ; Haberl et al ., 2007 ; Nogués-Bravo et al ., 2008 ).群落III位于海拔较高的阳坡, 随着海拔的升高, 气温逐渐降低, 光照强度增加, 较低的气温和阳坡较好的光照条件促进落叶物种的生长(Malhi et al ., 2010 ).充足的光照也促进土壤种子库萌发, 使群落内幼苗和幼树的密度较大.同时, 群落III位于保护区森林防火隔离带附近, 较频繁地受到了人为的干扰, 干扰提高了林下的光照条件, 导致群落内喜光的落叶树种增加和耐阴的常绿树种减少(Hawthorne, 1996 ; Bongers et al ., 2009 ).群落II和群落V分布于低海拔地区, 长期受到人类樵采等活动的影响, 林下的光照条件较好, 群落内的落叶物种大于常绿物种.群落I和群落IV主要位于阴坡, 且人为干扰较少, 林内光照条件较差, 适宜常绿物种的生长. ...
Do evergreen and deciduous trees have different effects on net N mineralization in soil.
1
2012
... 除地形因子和人为干扰之外, 土壤养分也对常绿和落叶物种比例的变化产生了一定的影响.落叶物种分布区的土壤氮含量常被认为高于常绿物种分布区, 其原因是落叶物种比常绿物种具有更高的氮矿化速率(Aerts & Chapin, 1999 ; Cornwell et al ., 2008 ), 然而也有研究表明常绿和落叶物种的氮矿化速率并无显著差异(Reich et al ., 2001 ; Booth et al ., 2005 ).本研究发现土壤氮含量在多数群落内与常绿物种丰富度显著正相关, 这可能与不同群落类型的物种属性及真菌差异有关(Mueller et al ., 2012 ).总体来看, 研究区常绿物种更易受土壤养分的影响.关于土壤养分对各群落内常绿和落叶物种分布的影响有待进一步深入分析. ...
Scale effects and human impact on the elevational species richness gradients.
1
2008
... 从物种组成和群落结构来看, 研究区各群落类型常绿和落叶物种存在一定的差异.群落I和群落IV的常绿和落叶物种在群落内的差异多不显著或者常绿物种大于落叶物种, 而群落II、群落III和群落V则表现为落叶物种大于常绿物种.造成这种差异的主要原因可能是环境差异和人为干扰.环境是影响物种组成的决定性因素, 环境因子组成的变化产生了不同的生境, 从而引起不同地带及区域的物种差异(Toledo et al ., 2011 ).而人为干扰能够显著地改变群落结构和物种组成, 从而影响群落的发展方向(Lomolino, 2001 ; Haberl et al ., 2007 ; Nogués-Bravo et al ., 2008 ).群落III位于海拔较高的阳坡, 随着海拔的升高, 气温逐渐降低, 光照强度增加, 较低的气温和阳坡较好的光照条件促进落叶物种的生长(Malhi et al ., 2010 ).充足的光照也促进土壤种子库萌发, 使群落内幼苗和幼树的密度较大.同时, 群落III位于保护区森林防火隔离带附近, 较频繁地受到了人为的干扰, 干扰提高了林下的光照条件, 导致群落内喜光的落叶树种增加和耐阴的常绿树种减少(Hawthorne, 1996 ; Bongers et al ., 2009 ).群落II和群落V分布于低海拔地区, 长期受到人类樵采等活动的影响, 林下的光照条件较好, 群落内的落叶物种大于常绿物种.群落I和群落IV主要位于阴坡, 且人为干扰较少, 林内光照条件较差, 适宜常绿物种的生长. ...
1
2006
... 由于叶片是植物光合作用的主要场所, 因而叶片的物候过程对于群落外貌、林下环境、凋落物分解和生态系统生产力具有重要作用(Bohlman et al ., 1998 ; Quigley & Platt, 2003 ).落叶是物种适应环境胁迫的一种重要策略, 通过落叶可以回避不利生境对树木生长造成的伤害和不利影响(Poorter & Markesteijn, 2008 ), 低温和水分胁迫是阔叶树种落叶的重要原因(Lechowicz, 1984 ; Reich et al ., 2004 ).与热带部分地区树种因土壤水分亏缺而出现旱季落叶的情况不同(Williams et al ., 2008 ; de Faria Lopes et al ., 2012 ), 亚热带常绿落叶阔叶混交林的形成更多受到海拔升高而形成的低温胁迫的影响(Oliveira- Filho et al ., 2006 ; 白坤栋等, 2010 ; 宋坤, 2012 ).回归分析中土壤含水量与常绿和落叶物种分布并无显著的相关性, 这个结果证明了研究区常绿和落叶物种的变化与土壤水分无关.温度是限制植物组成、生长、繁殖和分布的重要因子(Woodward, 1987 ).研究地区冬季气温较低, 一些喜温且生长迅速竞争能力强的常绿阔叶树种受到限制, 但绝对低温尚较高, 因此较耐寒的常绿阔叶树与落叶阔叶树均能生长, 最终在该地段形成了亚热带常绿落叶阔叶混交林(吴征镒, 1980 ). ...
Predictability of plant species composition from environmental conditions is constrained by dispersal limitation.
1
2005
... 植被分类是植被生态学研究中最复杂的一个问题, 直到现在还没有植被学家们认同的统一分类原则和分类系统(宋永昌, 2004 ).而我国亚热带地区植被具有复杂多样、次生性强和过渡类型多等特点, 更增加了该地区植被分类的困难(赖江山等, 2010 ).传统的植物群落分类多根据群落的外貌特征及群落的种类组成划分群落类型, 个人经验对分类结果的影响比较大(刘海江和郭柯, 2003 ; 刘万德等, 2010 ).随着植被研究的精度要求不断提高, 提供客观归类和划分结果的数量分类方法在植被分类中越来越重要(袁秀等, 2013 ).目前数量生态学中的分类和排序方法已成为现代植被生态学研究中必不可少的手段, 双向指示种分析法(TWINSPAN)和除趋势对应分析法(DCA)已广泛应用于森林和草地等植物群落分析(Ozinga et al ., 2005 ; 孙菊等, 2009 ; 廉凯敏等, 2015 ).本研究也证实了TWINSPAN分类与DCA排序这两种方法相结合应用于亚热带常绿落叶阔叶混交林植被数量分析是可行的, 而且可以相互补充和交叉检验. ...
Seedling traits determine drought tolerance of tropical tree species.
1
2008
... 由于叶片是植物光合作用的主要场所, 因而叶片的物候过程对于群落外貌、林下环境、凋落物分解和生态系统生产力具有重要作用(Bohlman et al ., 1998 ; Quigley & Platt, 2003 ).落叶是物种适应环境胁迫的一种重要策略, 通过落叶可以回避不利生境对树木生长造成的伤害和不利影响(Poorter & Markesteijn, 2008 ), 低温和水分胁迫是阔叶树种落叶的重要原因(Lechowicz, 1984 ; Reich et al ., 2004 ).与热带部分地区树种因土壤水分亏缺而出现旱季落叶的情况不同(Williams et al ., 2008 ; de Faria Lopes et al ., 2012 ), 亚热带常绿落叶阔叶混交林的形成更多受到海拔升高而形成的低温胁迫的影响(Oliveira- Filho et al ., 2006 ; 白坤栋等, 2010 ; 宋坤, 2012 ).回归分析中土壤含水量与常绿和落叶物种分布并无显著的相关性, 这个结果证明了研究区常绿和落叶物种的变化与土壤水分无关.温度是限制植物组成、生长、繁殖和分布的重要因子(Woodward, 1987 ).研究地区冬季气温较低, 一些喜温且生长迅速竞争能力强的常绿阔叶树种受到限制, 但绝对低温尚较高, 因此较耐寒的常绿阔叶树与落叶阔叶树均能生长, 最终在该地段形成了亚热带常绿落叶阔叶混交林(吴征镒, 1980 ). ...
Composition and structure of seasonally deciduous forests in the Americas.
2
2003
... 由于叶片是植物光合作用的主要场所, 因而叶片的物候过程对于群落外貌、林下环境、凋落物分解和生态系统生产力具有重要作用(Bohlman et al ., 1998 ; Quigley & Platt, 2003 ).落叶是物种适应环境胁迫的一种重要策略, 通过落叶可以回避不利生境对树木生长造成的伤害和不利影响(Poorter & Markesteijn, 2008 ), 低温和水分胁迫是阔叶树种落叶的重要原因(Lechowicz, 1984 ; Reich et al ., 2004 ).与热带部分地区树种因土壤水分亏缺而出现旱季落叶的情况不同(Williams et al ., 2008 ; de Faria Lopes et al ., 2012 ), 亚热带常绿落叶阔叶混交林的形成更多受到海拔升高而形成的低温胁迫的影响(Oliveira- Filho et al ., 2006 ; 白坤栋等, 2010 ; 宋坤, 2012 ).回归分析中土壤含水量与常绿和落叶物种分布并无显著的相关性, 这个结果证明了研究区常绿和落叶物种的变化与土壤水分无关.温度是限制植物组成、生长、繁殖和分布的重要因子(Woodward, 1987 ).研究地区冬季气温较低, 一些喜温且生长迅速竞争能力强的常绿阔叶树种受到限制, 但绝对低温尚较高, 因此较耐寒的常绿阔叶树与落叶阔叶树均能生长, 最终在该地段形成了亚热带常绿落叶阔叶混交林(吴征镒, 1980 ). ...
... 除群落III外, 各群落类型小径级(1-5 cm)内落叶物种的物种丰富度高于常绿物种, 但多度、胸高断面积和重要值却表现为落叶物种小于常绿物种, 这主要与常绿和落叶物种的生活型属性有关.研究区群落内小径级个体的生活型主要是小乔木和灌木, 而大乔木个体密度相对较低.这些小乔木和灌木物种以常绿树种为主, 因此出现了小径级内落叶物种丰富度高但多度、胸高断面积和重要值低的现象.较高强度的干扰能够提高落叶树种在群落中的比例(丁易和臧润国, 2008 ), 群落III的小径级在各个指标上都是落叶树种占优势, 这可能说明人为干扰和环境条件显著影响了该群落的更新和发展方向, 导致该群落在未来一段时间内都将以落叶物种为主.大径级(≥10 cm)的落叶物种在多数群落内的物种丰富度、多度、胸高断面积和重要值均大于常绿物种, 表明落叶树种在林冠层占据显著的优势, Chabot和Hicks (1982) 的研究也发现, 光照条件较好的林冠层落叶物种更多.林冠层落叶树种占据优势能够显著影响林下光照条件, 从而影响森林更新和物种多样性(Quigley & Platt, 2003 ).落叶物种具有更高的资源养分利用效率(Kloeppel et al ., 2000 ), 能够利用夏季良好的水热条件迅速获得碳水化合物完成高生长, 在与常绿树种的竞争过程中居于优势地位(Antúnez et al ., 2001 ).同时, 落叶物种采取了落叶的策略来适应研究区的冬季低温胁迫, 而常绿物种则保留叶片越冬, 低温可能损害常绿树种的气孔和光合速率而使常绿植物生长受到严重制约(Wyka & Oleksyn, 2014 ), 且较大的季节性温差所引起的冰凌和雪灾也可能造成常绿植物受到伤害从而限制其在林冠层的优势(白坤栋等, 2010 ).林下的常绿树木由于林冠层的郁闭而受到光强的限制.尽管在林冠层落叶期间(晚秋到早春), 林下光照条件可以由于林冠层的落叶而得到改善, 但低温限制了林下常绿树木的生长.这种竞争最终形成了林冠层以落叶阔叶树为主和林下层以常绿树种为主的格局. ...
Influence of logging, fire, and forest type on biodiversity and productivity in southern boreal forests.
1
2001
... 除地形因子和人为干扰之外, 土壤养分也对常绿和落叶物种比例的变化产生了一定的影响.落叶物种分布区的土壤氮含量常被认为高于常绿物种分布区, 其原因是落叶物种比常绿物种具有更高的氮矿化速率(Aerts & Chapin, 1999 ; Cornwell et al ., 2008 ), 然而也有研究表明常绿和落叶物种的氮矿化速率并无显著差异(Reich et al ., 2001 ; Booth et al ., 2005 ).本研究发现土壤氮含量在多数群落内与常绿物种丰富度显著正相关, 这可能与不同群落类型的物种属性及真菌差异有关(Mueller et al ., 2012 ).总体来看, 研究区常绿物种更易受土壤养分的影响.关于土壤养分对各群落内常绿和落叶物种分布的影响有待进一步深入分析. ...
Leaf demography and phenology in Amazonian rain forest: A census of 40000 leaves of 23 tree species.
1
2004
... 由于叶片是植物光合作用的主要场所, 因而叶片的物候过程对于群落外貌、林下环境、凋落物分解和生态系统生产力具有重要作用(Bohlman et al ., 1998 ; Quigley & Platt, 2003 ).落叶是物种适应环境胁迫的一种重要策略, 通过落叶可以回避不利生境对树木生长造成的伤害和不利影响(Poorter & Markesteijn, 2008 ), 低温和水分胁迫是阔叶树种落叶的重要原因(Lechowicz, 1984 ; Reich et al ., 2004 ).与热带部分地区树种因土壤水分亏缺而出现旱季落叶的情况不同(Williams et al ., 2008 ; de Faria Lopes et al ., 2012 ), 亚热带常绿落叶阔叶混交林的形成更多受到海拔升高而形成的低温胁迫的影响(Oliveira- Filho et al ., 2006 ; 白坤栋等, 2010 ; 宋坤, 2012 ).回归分析中土壤含水量与常绿和落叶物种分布并无显著的相关性, 这个结果证明了研究区常绿和落叶物种的变化与土壤水分无关.温度是限制植物组成、生长、繁殖和分布的重要因子(Woodward, 1987 ).研究地区冬季气温较低, 一些喜温且生长迅速竞争能力强的常绿阔叶树种受到限制, 但绝对低温尚较高, 因此较耐寒的常绿阔叶树与落叶阔叶树均能生长, 最终在该地段形成了亚热带常绿落叶阔叶混交林(吴征镒, 1980 ). ...
三峡地区大老岭珍稀植物的分布格局及其与地形因子的关系
1
1999
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
三峡地区大老岭珍稀植物的分布格局及其与地形因子的关系
1
1999
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
基于种群分布地形格局的两种水青冈生态位比较研究
0
2001
基于种群分布地形格局的两种水青冈生态位比较研究
0
2001
安徽常绿阔叶林-落叶阔叶林交错带的森林植被特征及其成因
1
2012
... 由于叶片是植物光合作用的主要场所, 因而叶片的物候过程对于群落外貌、林下环境、凋落物分解和生态系统生产力具有重要作用(Bohlman et al ., 1998 ; Quigley & Platt, 2003 ).落叶是物种适应环境胁迫的一种重要策略, 通过落叶可以回避不利生境对树木生长造成的伤害和不利影响(Poorter & Markesteijn, 2008 ), 低温和水分胁迫是阔叶树种落叶的重要原因(Lechowicz, 1984 ; Reich et al ., 2004 ).与热带部分地区树种因土壤水分亏缺而出现旱季落叶的情况不同(Williams et al ., 2008 ; de Faria Lopes et al ., 2012 ), 亚热带常绿落叶阔叶混交林的形成更多受到海拔升高而形成的低温胁迫的影响(Oliveira- Filho et al ., 2006 ; 白坤栋等, 2010 ; 宋坤, 2012 ).回归分析中土壤含水量与常绿和落叶物种分布并无显著的相关性, 这个结果证明了研究区常绿和落叶物种的变化与土壤水分无关.温度是限制植物组成、生长、繁殖和分布的重要因子(Woodward, 1987 ).研究地区冬季气温较低, 一些喜温且生长迅速竞争能力强的常绿阔叶树种受到限制, 但绝对低温尚较高, 因此较耐寒的常绿阔叶树与落叶阔叶树均能生长, 最终在该地段形成了亚热带常绿落叶阔叶混交林(吴征镒, 1980 ). ...
安徽常绿阔叶林-落叶阔叶林交错带的森林植被特征及其成因
1
2012
... 由于叶片是植物光合作用的主要场所, 因而叶片的物候过程对于群落外貌、林下环境、凋落物分解和生态系统生产力具有重要作用(Bohlman et al ., 1998 ; Quigley & Platt, 2003 ).落叶是物种适应环境胁迫的一种重要策略, 通过落叶可以回避不利生境对树木生长造成的伤害和不利影响(Poorter & Markesteijn, 2008 ), 低温和水分胁迫是阔叶树种落叶的重要原因(Lechowicz, 1984 ; Reich et al ., 2004 ).与热带部分地区树种因土壤水分亏缺而出现旱季落叶的情况不同(Williams et al ., 2008 ; de Faria Lopes et al ., 2012 ), 亚热带常绿落叶阔叶混交林的形成更多受到海拔升高而形成的低温胁迫的影响(Oliveira- Filho et al ., 2006 ; 白坤栋等, 2010 ; 宋坤, 2012 ).回归分析中土壤含水量与常绿和落叶物种分布并无显著的相关性, 这个结果证明了研究区常绿和落叶物种的变化与土壤水分无关.温度是限制植物组成、生长、繁殖和分布的重要因子(Woodward, 1987 ).研究地区冬季气温较低, 一些喜温且生长迅速竞争能力强的常绿阔叶树种受到限制, 但绝对低温尚较高, 因此较耐寒的常绿阔叶树与落叶阔叶树均能生长, 最终在该地段形成了亚热带常绿落叶阔叶混交林(吴征镒, 1980 ). ...
中国东部森林植被带划分之我见
1
1999
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
中国东部森林植被带划分之我见
1
1999
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
中国常绿阔叶林分类试行方案
1
2004
... 植被分类是植被生态学研究中最复杂的一个问题, 直到现在还没有植被学家们认同的统一分类原则和分类系统(宋永昌, 2004 ).而我国亚热带地区植被具有复杂多样、次生性强和过渡类型多等特点, 更增加了该地区植被分类的困难(赖江山等, 2010 ).传统的植物群落分类多根据群落的外貌特征及群落的种类组成划分群落类型, 个人经验对分类结果的影响比较大(刘海江和郭柯, 2003 ; 刘万德等, 2010 ).随着植被研究的精度要求不断提高, 提供客观归类和划分结果的数量分类方法在植被分类中越来越重要(袁秀等, 2013 ).目前数量生态学中的分类和排序方法已成为现代植被生态学研究中必不可少的手段, 双向指示种分析法(TWINSPAN)和除趋势对应分析法(DCA)已广泛应用于森林和草地等植物群落分析(Ozinga et al ., 2005 ; 孙菊等, 2009 ; 廉凯敏等, 2015 ).本研究也证实了TWINSPAN分类与DCA排序这两种方法相结合应用于亚热带常绿落叶阔叶混交林植被数量分析是可行的, 而且可以相互补充和交叉检验. ...
中国常绿阔叶林分类试行方案
1
2004
... 植被分类是植被生态学研究中最复杂的一个问题, 直到现在还没有植被学家们认同的统一分类原则和分类系统(宋永昌, 2004 ).而我国亚热带地区植被具有复杂多样、次生性强和过渡类型多等特点, 更增加了该地区植被分类的困难(赖江山等, 2010 ).传统的植物群落分类多根据群落的外貌特征及群落的种类组成划分群落类型, 个人经验对分类结果的影响比较大(刘海江和郭柯, 2003 ; 刘万德等, 2010 ).随着植被研究的精度要求不断提高, 提供客观归类和划分结果的数量分类方法在植被分类中越来越重要(袁秀等, 2013 ).目前数量生态学中的分类和排序方法已成为现代植被生态学研究中必不可少的手段, 双向指示种分析法(TWINSPAN)和除趋势对应分析法(DCA)已广泛应用于森林和草地等植物群落分析(Ozinga et al ., 2005 ; 孙菊等, 2009 ; 廉凯敏等, 2015 ).本研究也证实了TWINSPAN分类与DCA排序这两种方法相结合应用于亚热带常绿落叶阔叶混交林植被数量分析是可行的, 而且可以相互补充和交叉检验. ...
大兴安岭沟谷冻土湿地植物群落分类、物种多样性和物种分布梯度
1
2009
... 植被分类是植被生态学研究中最复杂的一个问题, 直到现在还没有植被学家们认同的统一分类原则和分类系统(宋永昌, 2004 ).而我国亚热带地区植被具有复杂多样、次生性强和过渡类型多等特点, 更增加了该地区植被分类的困难(赖江山等, 2010 ).传统的植物群落分类多根据群落的外貌特征及群落的种类组成划分群落类型, 个人经验对分类结果的影响比较大(刘海江和郭柯, 2003 ; 刘万德等, 2010 ).随着植被研究的精度要求不断提高, 提供客观归类和划分结果的数量分类方法在植被分类中越来越重要(袁秀等, 2013 ).目前数量生态学中的分类和排序方法已成为现代植被生态学研究中必不可少的手段, 双向指示种分析法(TWINSPAN)和除趋势对应分析法(DCA)已广泛应用于森林和草地等植物群落分析(Ozinga et al ., 2005 ; 孙菊等, 2009 ; 廉凯敏等, 2015 ).本研究也证实了TWINSPAN分类与DCA排序这两种方法相结合应用于亚热带常绿落叶阔叶混交林植被数量分析是可行的, 而且可以相互补充和交叉检验. ...
大兴安岭沟谷冻土湿地植物群落分类、物种多样性和物种分布梯度
1
2009
... 植被分类是植被生态学研究中最复杂的一个问题, 直到现在还没有植被学家们认同的统一分类原则和分类系统(宋永昌, 2004 ).而我国亚热带地区植被具有复杂多样、次生性强和过渡类型多等特点, 更增加了该地区植被分类的困难(赖江山等, 2010 ).传统的植物群落分类多根据群落的外貌特征及群落的种类组成划分群落类型, 个人经验对分类结果的影响比较大(刘海江和郭柯, 2003 ; 刘万德等, 2010 ).随着植被研究的精度要求不断提高, 提供客观归类和划分结果的数量分类方法在植被分类中越来越重要(袁秀等, 2013 ).目前数量生态学中的分类和排序方法已成为现代植被生态学研究中必不可少的手段, 双向指示种分析法(TWINSPAN)和除趋势对应分析法(DCA)已广泛应用于森林和草地等植物群落分析(Ozinga et al ., 2005 ; 孙菊等, 2009 ; 廉凯敏等, 2015 ).本研究也证实了TWINSPAN分类与DCA排序这两种方法相结合应用于亚热带常绿落叶阔叶混交林植被数量分析是可行的, 而且可以相互补充和交叉检验. ...
山地常绿落叶阔叶混交林更新方式对林地肥力的影响
2
2010
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
... 湖北西南部地区是湖北重要的天然林分布区, 亚热带常绿阔叶林为该地区的地带性植被.随着海拔的升高, 含有落叶成分的亚热带常绿落叶阔叶混交林逐步成为优势植被.亚热带常绿落叶阔叶混交林植物区系成分复杂, 生物多样性丰富, 不仅为我国的经济建设提供了大量的优质木材和林副产品, 而且对我国特有的古老孑遗物种保护、维护区域生态平衡发挥着重要的作用(汤景明等, 2010 ).由于气候、地质和区域物种库等方面的差异, 不同区域的常绿落叶阔叶混交林具有不同的变化特征, 针对不同区域的森林植被开展相关的生态学研究显得尤为必要.对亚热带常绿落叶阔叶混交林的群落类型进行科学划分, 不仅有助于我们对现存森林植被基本特征的认识, 而且是我们进一步深入开展生态系统功能与生态过程研究的基础.了解常绿和落叶物种比例随群落类型的变化规律, 能够揭示不同群落中环境条件对物种分布的影响, 从而为我们阐明生物多样性维持与调控机制奠定基础.亚热带常绿落叶阔叶混交林的群落类型及组成结构的研究是对其进行有效地保育、可持续经营与合理利用的科学基础. ...
山地常绿落叶阔叶混交林更新方式对林地肥力的影响
2
2010
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
... 湖北西南部地区是湖北重要的天然林分布区, 亚热带常绿阔叶林为该地区的地带性植被.随着海拔的升高, 含有落叶成分的亚热带常绿落叶阔叶混交林逐步成为优势植被.亚热带常绿落叶阔叶混交林植物区系成分复杂, 生物多样性丰富, 不仅为我国的经济建设提供了大量的优质木材和林副产品, 而且对我国特有的古老孑遗物种保护、维护区域生态平衡发挥着重要的作用(汤景明等, 2010 ).由于气候、地质和区域物种库等方面的差异, 不同区域的常绿落叶阔叶混交林具有不同的变化特征, 针对不同区域的森林植被开展相关的生态学研究显得尤为必要.对亚热带常绿落叶阔叶混交林的群落类型进行科学划分, 不仅有助于我们对现存森林植被基本特征的认识, 而且是我们进一步深入开展生态系统功能与生态过程研究的基础.了解常绿和落叶物种比例随群落类型的变化规律, 能够揭示不同群落中环境条件对物种分布的影响, 从而为我们阐明生物多样性维持与调控机制奠定基础.亚热带常绿落叶阔叶混交林的群落类型及组成结构的研究是对其进行有效地保育、可持续经营与合理利用的科学基础. ...
Climate and soil drive forest structure in Bolivian lowland forests.
1
2011
... 从物种组成和群落结构来看, 研究区各群落类型常绿和落叶物种存在一定的差异.群落I和群落IV的常绿和落叶物种在群落内的差异多不显著或者常绿物种大于落叶物种, 而群落II、群落III和群落V则表现为落叶物种大于常绿物种.造成这种差异的主要原因可能是环境差异和人为干扰.环境是影响物种组成的决定性因素, 环境因子组成的变化产生了不同的生境, 从而引起不同地带及区域的物种差异(Toledo et al ., 2011 ).而人为干扰能够显著地改变群落结构和物种组成, 从而影响群落的发展方向(Lomolino, 2001 ; Haberl et al ., 2007 ; Nogués-Bravo et al ., 2008 ).群落III位于海拔较高的阳坡, 随着海拔的升高, 气温逐渐降低, 光照强度增加, 较低的气温和阳坡较好的光照条件促进落叶物种的生长(Malhi et al ., 2010 ).充足的光照也促进土壤种子库萌发, 使群落内幼苗和幼树的密度较大.同时, 群落III位于保护区森林防火隔离带附近, 较频繁地受到了人为的干扰, 干扰提高了林下的光照条件, 导致群落内喜光的落叶树种增加和耐阴的常绿树种减少(Hawthorne, 1996 ; Bongers et al ., 2009 ).群落II和群落V分布于低海拔地区, 长期受到人类樵采等活动的影响, 林下的光照条件较好, 群落内的落叶物种大于常绿物种.群落I和群落IV主要位于阴坡, 且人为干扰较少, 林内光照条件较差, 适宜常绿物种的生长. ...
亚热带山地亮叶水青冈林的群落分类及物种组成与更新
1
2006
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
亚热带山地亮叶水青冈林的群落分类及物种组成与更新
1
2006
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
Deciduousness in a seasonal tropical forest in western Thailand: Interannual and intraspecific variation in timing, duration and environmental cues.
1
2008
... 由于叶片是植物光合作用的主要场所, 因而叶片的物候过程对于群落外貌、林下环境、凋落物分解和生态系统生产力具有重要作用(Bohlman et al ., 1998 ; Quigley & Platt, 2003 ).落叶是物种适应环境胁迫的一种重要策略, 通过落叶可以回避不利生境对树木生长造成的伤害和不利影响(Poorter & Markesteijn, 2008 ), 低温和水分胁迫是阔叶树种落叶的重要原因(Lechowicz, 1984 ; Reich et al ., 2004 ).与热带部分地区树种因土壤水分亏缺而出现旱季落叶的情况不同(Williams et al ., 2008 ; de Faria Lopes et al ., 2012 ), 亚热带常绿落叶阔叶混交林的形成更多受到海拔升高而形成的低温胁迫的影响(Oliveira- Filho et al ., 2006 ; 白坤栋等, 2010 ; 宋坤, 2012 ).回归分析中土壤含水量与常绿和落叶物种分布并无显著的相关性, 这个结果证明了研究区常绿和落叶物种的变化与土壤水分无关.温度是限制植物组成、生长、繁殖和分布的重要因子(Woodward, 1987 ).研究地区冬季气温较低, 一些喜温且生长迅速竞争能力强的常绿阔叶树种受到限制, 但绝对低温尚较高, 因此较耐寒的常绿阔叶树与落叶阔叶树均能生长, 最终在该地段形成了亚热带常绿落叶阔叶混交林(吴征镒, 1980 ). ...
1
1987
... 由于叶片是植物光合作用的主要场所, 因而叶片的物候过程对于群落外貌、林下环境、凋落物分解和生态系统生产力具有重要作用(Bohlman et al ., 1998 ; Quigley & Platt, 2003 ).落叶是物种适应环境胁迫的一种重要策略, 通过落叶可以回避不利生境对树木生长造成的伤害和不利影响(Poorter & Markesteijn, 2008 ), 低温和水分胁迫是阔叶树种落叶的重要原因(Lechowicz, 1984 ; Reich et al ., 2004 ).与热带部分地区树种因土壤水分亏缺而出现旱季落叶的情况不同(Williams et al ., 2008 ; de Faria Lopes et al ., 2012 ), 亚热带常绿落叶阔叶混交林的形成更多受到海拔升高而形成的低温胁迫的影响(Oliveira- Filho et al ., 2006 ; 白坤栋等, 2010 ; 宋坤, 2012 ).回归分析中土壤含水量与常绿和落叶物种分布并无显著的相关性, 这个结果证明了研究区常绿和落叶物种的变化与土壤水分无关.温度是限制植物组成、生长、繁殖和分布的重要因子(Woodward, 1987 ).研究地区冬季气温较低, 一些喜温且生长迅速竞争能力强的常绿阔叶树种受到限制, 但绝对低温尚较高, 因此较耐寒的常绿阔叶树与落叶阔叶树均能生长, 最终在该地段形成了亚热带常绿落叶阔叶混交林(吴征镒, 1980 ). ...
1
2000
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
1
2000
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
3
1980
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
... ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
... 由于叶片是植物光合作用的主要场所, 因而叶片的物候过程对于群落外貌、林下环境、凋落物分解和生态系统生产力具有重要作用(Bohlman et al ., 1998 ; Quigley & Platt, 2003 ).落叶是物种适应环境胁迫的一种重要策略, 通过落叶可以回避不利生境对树木生长造成的伤害和不利影响(Poorter & Markesteijn, 2008 ), 低温和水分胁迫是阔叶树种落叶的重要原因(Lechowicz, 1984 ; Reich et al ., 2004 ).与热带部分地区树种因土壤水分亏缺而出现旱季落叶的情况不同(Williams et al ., 2008 ; de Faria Lopes et al ., 2012 ), 亚热带常绿落叶阔叶混交林的形成更多受到海拔升高而形成的低温胁迫的影响(Oliveira- Filho et al ., 2006 ; 白坤栋等, 2010 ; 宋坤, 2012 ).回归分析中土壤含水量与常绿和落叶物种分布并无显著的相关性, 这个结果证明了研究区常绿和落叶物种的变化与土壤水分无关.温度是限制植物组成、生长、繁殖和分布的重要因子(Woodward, 1987 ).研究地区冬季气温较低, 一些喜温且生长迅速竞争能力强的常绿阔叶树种受到限制, 但绝对低温尚较高, 因此较耐寒的常绿阔叶树与落叶阔叶树均能生长, 最终在该地段形成了亚热带常绿落叶阔叶混交林(吴征镒, 1980 ). ...
3
1980
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
... ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
... 由于叶片是植物光合作用的主要场所, 因而叶片的物候过程对于群落外貌、林下环境、凋落物分解和生态系统生产力具有重要作用(Bohlman et al ., 1998 ; Quigley & Platt, 2003 ).落叶是物种适应环境胁迫的一种重要策略, 通过落叶可以回避不利生境对树木生长造成的伤害和不利影响(Poorter & Markesteijn, 2008 ), 低温和水分胁迫是阔叶树种落叶的重要原因(Lechowicz, 1984 ; Reich et al ., 2004 ).与热带部分地区树种因土壤水分亏缺而出现旱季落叶的情况不同(Williams et al ., 2008 ; de Faria Lopes et al ., 2012 ), 亚热带常绿落叶阔叶混交林的形成更多受到海拔升高而形成的低温胁迫的影响(Oliveira- Filho et al ., 2006 ; 白坤栋等, 2010 ; 宋坤, 2012 ).回归分析中土壤含水量与常绿和落叶物种分布并无显著的相关性, 这个结果证明了研究区常绿和落叶物种的变化与土壤水分无关.温度是限制植物组成、生长、繁殖和分布的重要因子(Woodward, 1987 ).研究地区冬季气温较低, 一些喜温且生长迅速竞争能力强的常绿阔叶树种受到限制, 但绝对低温尚较高, 因此较耐寒的常绿阔叶树与落叶阔叶树均能生长, 最终在该地段形成了亚热带常绿落叶阔叶混交林(吴征镒, 1980 ). ...
Photosynthetic ecophysiology of evergreen leaves in the woody angiosperms—A review.
1
2014
... 除群落III外, 各群落类型小径级(1-5 cm)内落叶物种的物种丰富度高于常绿物种, 但多度、胸高断面积和重要值却表现为落叶物种小于常绿物种, 这主要与常绿和落叶物种的生活型属性有关.研究区群落内小径级个体的生活型主要是小乔木和灌木, 而大乔木个体密度相对较低.这些小乔木和灌木物种以常绿树种为主, 因此出现了小径级内落叶物种丰富度高但多度、胸高断面积和重要值低的现象.较高强度的干扰能够提高落叶树种在群落中的比例(丁易和臧润国, 2008 ), 群落III的小径级在各个指标上都是落叶树种占优势, 这可能说明人为干扰和环境条件显著影响了该群落的更新和发展方向, 导致该群落在未来一段时间内都将以落叶物种为主.大径级(≥10 cm)的落叶物种在多数群落内的物种丰富度、多度、胸高断面积和重要值均大于常绿物种, 表明落叶树种在林冠层占据显著的优势, Chabot和Hicks (1982) 的研究也发现, 光照条件较好的林冠层落叶物种更多.林冠层落叶树种占据优势能够显著影响林下光照条件, 从而影响森林更新和物种多样性(Quigley & Platt, 2003 ).落叶物种具有更高的资源养分利用效率(Kloeppel et al ., 2000 ), 能够利用夏季良好的水热条件迅速获得碳水化合物完成高生长, 在与常绿树种的竞争过程中居于优势地位(Antúnez et al ., 2001 ).同时, 落叶物种采取了落叶的策略来适应研究区的冬季低温胁迫, 而常绿物种则保留叶片越冬, 低温可能损害常绿树种的气孔和光合速率而使常绿植物生长受到严重制约(Wyka & Oleksyn, 2014 ), 且较大的季节性温差所引起的冰凌和雪灾也可能造成常绿植物受到伤害从而限制其在林冠层的优势(白坤栋等, 2010 ).林下的常绿树木由于林冠层的郁闭而受到光强的限制.尽管在林冠层落叶期间(晚秋到早春), 林下光照条件可以由于林冠层的落叶而得到改善, 但低温限制了林下常绿树木的生长.这种竞争最终形成了林冠层以落叶阔叶树为主和林下层以常绿树种为主的格局. ...
神农架地区常绿落叶阔叶混交林树种更新研究
1
2002
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
神农架地区常绿落叶阔叶混交林树种更新研究
1
2002
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
草地植物群落最优分类数的确定——以黄河三角洲为例
1
2013
... 植被分类是植被生态学研究中最复杂的一个问题, 直到现在还没有植被学家们认同的统一分类原则和分类系统(宋永昌, 2004 ).而我国亚热带地区植被具有复杂多样、次生性强和过渡类型多等特点, 更增加了该地区植被分类的困难(赖江山等, 2010 ).传统的植物群落分类多根据群落的外貌特征及群落的种类组成划分群落类型, 个人经验对分类结果的影响比较大(刘海江和郭柯, 2003 ; 刘万德等, 2010 ).随着植被研究的精度要求不断提高, 提供客观归类和划分结果的数量分类方法在植被分类中越来越重要(袁秀等, 2013 ).目前数量生态学中的分类和排序方法已成为现代植被生态学研究中必不可少的手段, 双向指示种分析法(TWINSPAN)和除趋势对应分析法(DCA)已广泛应用于森林和草地等植物群落分析(Ozinga et al ., 2005 ; 孙菊等, 2009 ; 廉凯敏等, 2015 ).本研究也证实了TWINSPAN分类与DCA排序这两种方法相结合应用于亚热带常绿落叶阔叶混交林植被数量分析是可行的, 而且可以相互补充和交叉检验. ...
草地植物群落最优分类数的确定——以黄河三角洲为例
1
2013
... 植被分类是植被生态学研究中最复杂的一个问题, 直到现在还没有植被学家们认同的统一分类原则和分类系统(宋永昌, 2004 ).而我国亚热带地区植被具有复杂多样、次生性强和过渡类型多等特点, 更增加了该地区植被分类的困难(赖江山等, 2010 ).传统的植物群落分类多根据群落的外貌特征及群落的种类组成划分群落类型, 个人经验对分类结果的影响比较大(刘海江和郭柯, 2003 ; 刘万德等, 2010 ).随着植被研究的精度要求不断提高, 提供客观归类和划分结果的数量分类方法在植被分类中越来越重要(袁秀等, 2013 ).目前数量生态学中的分类和排序方法已成为现代植被生态学研究中必不可少的手段, 双向指示种分析法(TWINSPAN)和除趋势对应分析法(DCA)已广泛应用于森林和草地等植物群落分析(Ozinga et al ., 2005 ; 孙菊等, 2009 ; 廉凯敏等, 2015 ).本研究也证实了TWINSPAN分类与DCA排序这两种方法相结合应用于亚热带常绿落叶阔叶混交林植被数量分析是可行的, 而且可以相互补充和交叉检验. ...
1
1995
... 以样地为单位分别计算物种的重要值, 计算公式为: 重要值(IV ) = (相对多度+相对显著度+相对频度)/3.采用VEAPAN软件包中的TWINSPAN进行群落分类; 通过CANOCO 4.5软件(张金屯, 1995 )对样地和物种的重要值矩阵进行DCA排序.根据个体胸径(DBH )将物种划分为3个等级: 小径级(1 cm ≤ DBH < 5 cm)、中径级(5 cm ≤ DBH < 10 cm)和大径级(DBH ≥ 10 cm).基于Bray-Curtis相似距离, 采用基于矩阵的多元回归分析(Permutation-based multiple regression on distance matrices, MRM)确定显著影响群落物种组成变化的环境因子(Lichstein, 2007 ).运用多元逐步回归分析(multiple regression analysis)来分析常绿和落叶物种与环境因子之间的关系. ...
1
1995
... 以样地为单位分别计算物种的重要值, 计算公式为: 重要值(IV ) = (相对多度+相对显著度+相对频度)/3.采用VEAPAN软件包中的TWINSPAN进行群落分类; 通过CANOCO 4.5软件(张金屯, 1995 )对样地和物种的重要值矩阵进行DCA排序.根据个体胸径(DBH )将物种划分为3个等级: 小径级(1 cm ≤ DBH < 5 cm)、中径级(5 cm ≤ DBH < 10 cm)和大径级(DBH ≥ 10 cm).基于Bray-Curtis相似距离, 采用基于矩阵的多元回归分析(Permutation-based multiple regression on distance matrices, MRM)确定显著影响群落物种组成变化的环境因子(Lichstein, 2007 ).运用多元逐步回归分析(multiple regression analysis)来分析常绿和落叶物种与环境因子之间的关系. ...
三峡库区常绿落叶阔叶混交林的监测研究
1
2004
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
三峡库区常绿落叶阔叶混交林的监测研究
1
2004
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
中亚热带石栎-青冈群落物种组成、结构及区系特征
1
2013
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
中亚热带石栎-青冈群落物种组成、结构及区系特征
1
2013
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
中国的落叶阔叶林——兼论常绿落叶阔叶混交林的归属问题
1
1996
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...
中国的落叶阔叶林——兼论常绿落叶阔叶混交林的归属问题
1
1996
... 常绿落叶阔叶混交林是一种介于落叶阔叶林与常绿阔叶林之间的森林植被类型, 通常被认为是与常绿阔叶林和落叶阔叶林并列的一种植被类型(吴中伦, 2000 ), 并可进一步划分为不同的植被亚型(吴征镒, 1980 ).目前对常绿落叶阔叶混交林自然分布的划定和分类存在不同观点(吴征镒, 1980 ; 安树青等, 1998 ; 宋永昌, 1999 ; 方精云, 2001 )(安树青 et al. 1998 ).长期以来, 我国对常绿落叶阔叶混交林的基本概念(班继德和漆根深, 1995 ; 周光裕, 1996 )、空间分布(沈泽昊和方精云, 2001 ; 张谧等, 2004 )、物种组成(曹铁如等, 1997 ; 汪正祥等, 2006 ; 赵丽娟等, 2013 )、群落更新(熊小刚等, 2002 ; 汤景明等, 2010 ; 刘海波等, 2014 )、种间关系(韩文衡等, 2009 )、外来种入侵(卢少飞等, 2005 )和珍稀物种分布(沈泽昊等, 1999 )等方面有了一定的研究.然而, 相对于典型的常绿阔叶林和落叶阔叶林生态研究来说, 亚热带常绿落叶阔叶混交林及其生态特征的研究还非常薄弱. ...



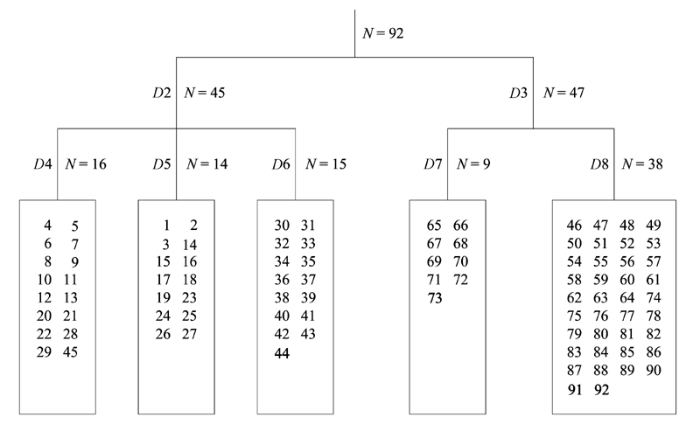
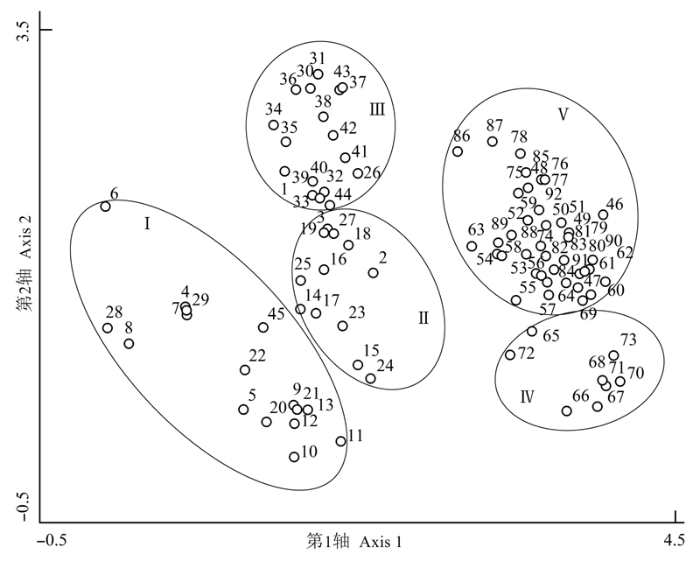
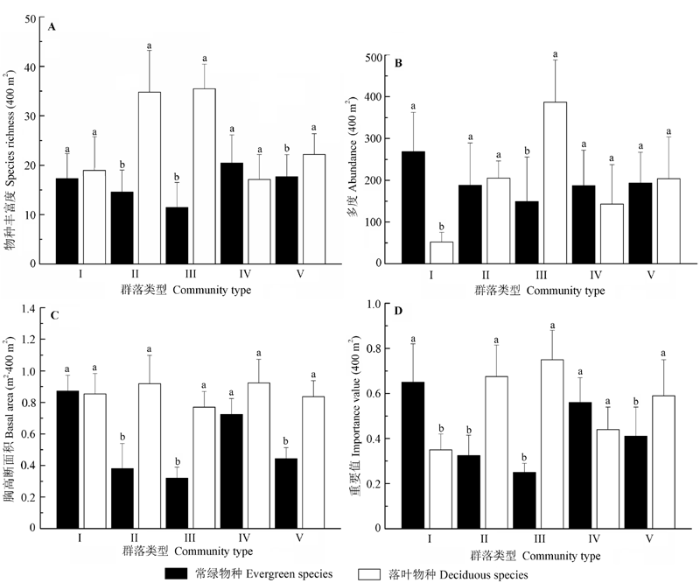
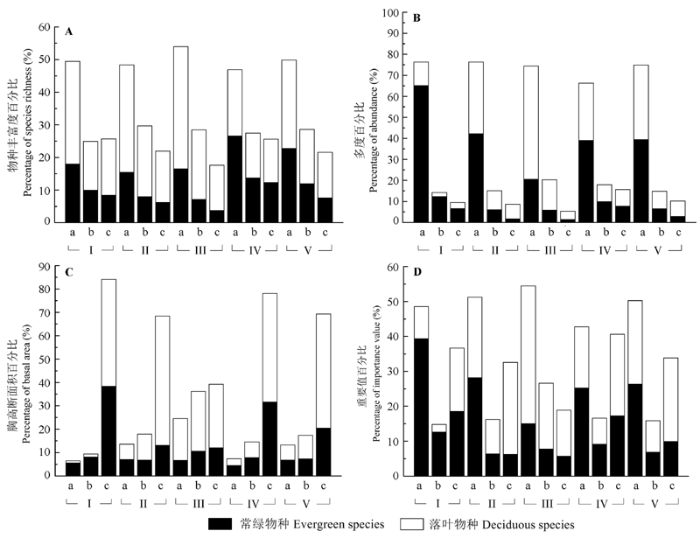
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}