当非土著种进入一个过去不曾分布的地区, 并能存活、繁殖而建立种群, 其种群的进一步扩散已经或将要造成明显的生态环境和经济后果, 这一过程称为生物入侵(biological invasion)。生物入侵强调外来入侵种的地理属性及其造成的影响(Elton, 1958 ; 李博和陈家宽, 2002 )。
纵观生物入侵研究的发展历程, 生物入侵的相关机制与假说向来是入侵生态学研究的难点、热点, 备受争议, 至今没有哪一种机制或假说能解释大部分的生物入侵现象(Ren & Zhang, 2009 ; Lowry et al ., 2013 )。今后的整合研究或许能为研究生物入侵机制与假说提供新的视角(Kueffer et al ., 2013 )。随着人类对自然界改造的加剧, 全球贸易往来过度密切, 全球生物入侵形势复杂多变, 外来种入侵带来的生态环境、社会经济、人类健康及生物安全等问题前所未有地严峻(Bradley et al ., 2012 )。目前生物入侵已成为全球变化的因素, 也是导致全球生物多样性丧失的主要原因, 仅次于生境破坏, 并很可能成为第一因素(Mack et al ., 2000 )。生物入侵将与其他全球变化因素一起左右着地球的未来。
目前与红树林有关的生物入侵研究仍处于起步阶段。我们在ISI Web of Science网站上, 以“外来种” “引入种” “入侵种” “入侵”和“红树林” “红树植物”为关键词, 可检索到文献120篇左右, 近年来每年出版有关文献12篇左右(图1 )。本文应用HistCite软件(http://en.wikipedia.org/wiki/Histcite)对上述文献进行分析, 发现红树林生物入侵研究一直不够活跃。从被引用最多的几篇文献来看, 红树林生物入侵研究热点还只限于红树林入侵生物多样性与分布格局、生物入侵红树林产生的相关效应(主要是对生态系统结构与功能的效应)、入侵风险评价及预测未来入侵; 研究区域集中在中国华南与东南沿海(尤其是珠江口、雷州半岛西侧)和美国东南海岸及夏威夷群岛(表1 ; 图2 )。
基于这些文献资料, 我们统计分析了现有红树林生物入侵研究文献的全球分布格局, 针对产生明显危害的入侵种, 综述了它们的入侵模式、入侵红树林产生的效应, 以及涉及的主要防治方法。基于上述资料, 展望今后红树林生物入侵研究的发展趋势, 为红树林生物入侵的研究和管理提供借鉴, 同时引起学者、公众和决策者对红树林保护和研究的重视。
对全球红树林外来入侵种没有详细的统计调查, 现有的红树林生物入侵研究比较局限, 通常是针对造成明显入侵效应的入侵种开展相应的入侵效应研究。现阶段红树林生物入侵研究的主要研究对象为植物, 动物和微生物研究很少, 植物、动物和微生物研究的文献量分别为78、8、1篇。世界范围内, 红树林生物入侵研究区域集中于中国华南和东南沿海, 其次是美国夏威夷群岛和东南沿海, 在巴西、印度、澳大利亚、尼日利亚及加勒比海区域只有零星研究(附表1 )。
动物和微生物入侵红树林的相关研究更少。一种名为狮子鱼(lionfish)的海洋鱼类入侵红树林后构成危害, 是目前主要的红树林动物入侵研究对象。而有关入侵红树林的微生物研究仅有1篇报道, 是与红树林外来植物小叶海金沙(Lygodium microphyllum )共生的丛枝菌根真菌(arbuscular mycorrhizal fungi, AMF)对红树林的入侵研究。
现在红树林生物入侵效应研究已经从生态系统结构和功能效应, 逐渐转移到生态系统服务效应上。红树林生物入侵效应研究目前还仅仅局限于外来植物入侵红树林导致的生态系统结构和功能效应, 而对社会经济、人类健康、生态系统服务的影响至今没有报道, 今后亟需重视。
互花米草是禾本科米草属(又名绳草属)多年生草本植物, 原产于美洲大西洋沿岸和墨西哥湾。近200年来, 经不同的入侵途径, 互花米草的分布区域已经从其原产地扩展到欧洲、北美西海岸、新西兰与中国沿海, 被公认为是全球海岸滩涂湿地生态系统最成功的入侵植物之一(王卿等, 2006 )。外界条件与互花米草自身特性相辅相成, 促成了互花米草高效的、难以防治的入侵模式:快速扩张呈现出点源扩散和多点爆发的特征, 对乡土植被的侵占表现为交互生长与斑块推进相结合(Xiao et al ., 2010 )。因此, 互花米草一旦成功入侵, 很难根除和控制。
互花米草已经出现在中国所有的红树林区。对福建境内红树林互花米草入侵的研究走在国内前列。互花米草入侵导致红树林生境土壤退化, 漳江口互花米草生境的土壤有机碳、土壤微生物生物量碳、氮含量以及土壤蔗糖酶与磷酸酶活性等相对于红树林生境均有明显下降(张祥霖等, 2008 )。但是, 互花米草群落对碳、氮具有很强的富集能力, 相对于红树林对碳、氮弱的富集能力, 互花米草的存在能更好地拦截和吸附陆源污染物, 并使重金属富集在互花米草群落沉积物中, 增加泉州湾红树林污染富集承受能力(王爱军等, 2008 )。
互花米草入侵还会改变红树林的生物多样性及其行为模式。在九龙江口红树林区, 互花米草入侵区域底栖动物的栖息密度、种类多样性、丰度比红树林区高, 比涂滩要低(洪荣标等, 2005 )。互花米草的入侵还改变着氨氧化古菌与细菌的丰度, 影响氨氧化微生物的群落结构(Zhang et al ., 2011 )。在漳江口红树林区, 红树林生境与互花米草生境之间多毛类动物栖息密度、生物量、丰度指数、均匀度指数和多样性指数均有显著差异(陈昕韡等, 2012 )。红树林与互花米草生境大型底栖动物次级生产力也存在差异, 海榄雌(Avicennia marina )生境大型底栖动物的年次级生产力最高, 而互花米草生境大型底栖动物的P/B值(P: 平均次级生产力; B: 平均生物量)最高, 因其优势类群为个体较小的寡毛类(吴辰等, 2013 )。再有, 互花米草相对较高的根密度显著降低了红树林中蟹类在互花米草生境打洞的深度及洞穴的复杂性(Wang et al ., 2014 )。
上述为目前有关互花米草入侵对红树林微生境及底栖生物产生的效应研究, 零散而浅显, 研究区域局限, 相关机制也不明确。鉴于生物入侵的地域性, 不同地区生物入侵导致的效应可能截然不同。深入研究的同时综合比较不同区域互花米草入侵产生的效应, 全面深入理解互花米草入侵机制, 对于制定合适的防治与管理措施具有积极意义。
微甘菊是菊科假泽兰属多年生草质藤本, 原产于热带南美与中美洲。微甘菊生长迅速且适应性强, 英文称其为“Mile-a-Minute Weed” (一分钟一英里杂草), 极具入侵性(Waterhouse, 1994 )。微甘菊在原产地因有天敌控制不具危害, 但在缺乏天敌的入侵地迅速发展为危害性严重的有害杂草(昝启杰等, 2000 )。微甘菊因其独特的生物学特性及生态适应能力(Tripathi et al ., 2011 ), 传入新生境后能迅速爆发。如今, 微甘菊已在整个亚洲热带及北美地区肆虐, 成为著名的世界性恶草(Zhang et al ., 2004 )。
相对于互花米草对红树林的入侵研究, 微甘菊入侵红树林的研究报道更显不足。而且相关研究还只关注于微甘菊在红树林的入侵途径, 及对红树林群落的胁迫, 而对红树林生态系统其他结构和功能的效应都未见研究报道。整体上, 微甘菊入侵红树林的相关研究极其薄弱, 亟需全面加强。
无瓣海桑是海桑科海桑属乔木, 天然分布于印度、孟加拉国、马来西亚等, 因其速生性而被广泛用于海岸滩涂造林, 1985年引入中国海南岛, 现在常见于中国大部分红树林区(李云等, 1998 )。
虽然在群落或生态系统尺度上, 无瓣海桑确实能改良生境, 增加生物多样性, 但在更大尺度上是否有同样的效应, 目前还不清楚。无瓣海桑对林下乡土红树植物的竞争与化感抑制效应是不容忽视的。长远看, 无瓣海桑的入侵势必降低引入地植物和动物区系的特有性。只有对现有的无瓣海桑入侵与否进行深入探讨, 明确无瓣海桑能否爆发入侵, 才能为未来无瓣海桑的引种种植提供合理建议。反观现在中国红树林区大面积种植无瓣海桑的情况(Ren et al ., 2009 ), 不能不令人担忧。
上述是迄今所有的有关红树林植物入侵研究的具体情况, 所涉及的入侵植物入侵性突出或极富争议, 入侵效应明显, 首当其冲受到关注。中国东南沿海和美国夏威夷群岛是研究红树林植物入侵的集中区域, 而且中国红树林生物入侵研究多于世界其他国家和地区。国内外对红树林生物入侵整体上关注不够, 红树林的入侵研究与保护应受到重视。
综上所述, 现阶段动物入侵红树林的效应研究仅限于狮子鱼的文献报道, 其他入侵动物因未呈现具体效应而没有深入的研究, 此外, 未见其他类似报道。上述红树林动物入侵研究都是国外的报道, 国内仍是空白。相对于植物入侵红树林研究, 动物入侵研究更为严峻, 需加以重视。
微生物入侵红树林的研究寥寥无几。微生物入侵多半是依附在入侵的动植物上, 被动植物带入新生境。与小叶海金沙共生的丛枝菌根真菌(AMF), 在植物入侵红树林时被引入(Soti et al ., 2014 )。一些入侵非土著海洋的微生物是否入侵红树林, 目前仍不明确(Ruiz et al ., 2000 ; 陆琴燕等, 2013 ), 有待关注。而附属于红树植物木材上的一些真菌种类, 未经严格的检疫检验, 通过商业贸易交易到非土著地区, 成为极具危险的潜在入侵病原体(Vannini et al ., 2012 )。研究红树林植物或动物入侵时, 关注与它们共生或寄生的微生物, 是研究微生物入侵与防治的切实可行的途径。
纵观现有生物入侵研究, 红树林的生物入侵研究严重滞后。红树林生物入侵研究的现阶段还主要是关注植物入侵及其产生的效应, 动物入侵研究才刚有涉足, 微生物入侵研究几乎空白。红树林生物入侵的效应、入侵机制研究尚未开展, 而且国内外同类研究严重不成比例, 研究结果可能存在随机性与片面性, 不同地区也无法进行对比与整合。一直以来, 生物入侵的防治往往走在系统研究生物入侵机制的前面, 在没有充分认识入侵机理的前提下, 采取任何防治手段都是盲目的。因此, 亟需启动红树林生物入侵机制研究, 同步于防治工作, 以期最大限度地减轻生物入侵危害, 发挥外来种的积极效应。
针对微甘菊, 通常是人工清除, 先割除地上部分, 再挖出地下根与茎, 放在烈日下曝晒, 但其群落很快就又恢复如初, 毫无效果。而在拔除微甘菊后, 种植人工干预的乔灌草群落, 能在一定程度上控制微甘菊的危害(李小川等, 2003 )。
无瓣海桑繁殖和更新存在一定的局限性, 其扩散能力不是很强, 人为适当的砍伐或拔除就能控制其种群数量。研究还发现对无瓣海桑做低光照处理, 能有效地抑制其生长(Chen et al ., 2013a )。
总之, 物理控制法虽然在短时间内比较有效, 但大多费时费力, 成本也较高。而且外来入侵种本身就对物理胁迫或干扰有很强的抵抗力, 其高繁殖力使得物理控制需年年进行。现在使用的各种物理控制方法在第一个生长季能有效地抑制其生长, 但从更长的时期来看, 并不能彻底有效地根除外来入侵种。
化学控制是利用化学药剂对入侵种进行灭杀, 以达到控制甚至消灭入侵种的目的。相对于物理控制, 化学控制效果更明显, 而且节省人力物力, 是控制外来有害物种的主要方式(Mack et al ., 2000 )。但是, 化学药剂缺乏目标特异性, 在毒杀目标物种的同时, 也毒害了非目标物种, 使用前必须掌握所用化学药剂的安全性(昝启杰等, 2007 )。
对比几种不同的除草剂对微甘菊的灭杀效果及其对其他生物的安全性, 证实一种名为“森草净”的除草剂对红树林区的微甘菊有极好的杀灭效果, 且对施用区域的生物多样性无不良影响, 被认为是微甘菊化学控制的首选除草剂(昝启杰等, 2007 )。
但是, 应用化学药剂控制入侵生物通常只能清除地上部分, 对地下部分效果差。而且化学药剂具有一定的毒性, 且有残留问题, 在杀灭入侵种的同时, 也不可避免地会杀灭其他乡土生物, 容易造成环境污染, 使施药区干扰程度加大, 对生态环境以及人类健康、经济发展等造成影响, 所以必须谨慎使用。
生物防治是以入侵生物与天敌的生态学原理, 依据生物之间相互依存与制约关系, 利用一种或多种生物控制另一种生物种群的消长。具体是指从入侵种原产地引进昆虫、真菌、病原生物或捕食者等天敌来抑制入侵种的生长和繁殖, 从而遏制其种群的爆发。生物控制很好地避免了物理和化学控制的弊端, 是未来生物入侵控制的主要手段(Mack et al ., 2000 )。
生物控制也不是毫无风险, 我们无法确保引进的天敌不会作用于非目标生物而成为新的入侵种。利用天敌对入侵种进行防治, 需要了解二者的生物学、生态学特性, 协调和处理好生物与生物及环境之间的关系。然而, 天敌的作用极其复杂, 生物控制中对引进天敌的效果与后果始终存在争议。因此, 对于大多数入侵生物的生物控制仍处于实验阶段, 没有得到大规模的应用(Mack et al ., 2000 )。
为了有效地控制红树林入侵生物, 现在越来越强调综合控制, 将物理、化学、生物等方法有机地结合起来, 取长补短, 相互协调。成功的生物入侵毕竟是一个小概率事件, 因此在生物入侵过程中, 抓住早期的关键环节, 预防入侵永远是最有效而代价最低的手段。再配以严格而科学的综合防治和监控, 才能有效地防治生物入侵。
入侵生态学是生态学领域一门活跃的学科, 经过150多年的发展, 入侵种的多样性与分布格局、入侵效应、入侵机制与防治等相关研究日趋系统全面。红树林生态系统因其独特性, 其生物入侵相关研究全面滞后于其他类型生态系统。通过对比, 我们总结了红树林生物入侵研究今后应当加强的方面, 具体如下:
当前红树林生物入侵研究多以植物为背景——外来植物入侵红树林, 或是红树植物入侵非土著区域; 与之相关的动物及微生物入侵研究亟需加强。红树林生物入侵效应研究至今仅限于生物入侵对生态系统结构和功能的影响, 对其社会经济、人类健康以及生态系统服务效应应予以重视。再有, 应加快开展红树林生物入侵的相关机制研究, 及更广泛时空尺度上的机制探索, 注重世界范围内不同区域同类对比综合研究。
应用分子生物学技术实现红树林生物入侵微观层面研究, 从分子基因水平探讨红树林生物入侵的遗传学基础。3S技术从景观以上水平展示红树林生物入侵的时空动态。数学模型模拟红树林生物入侵相关过程的物质和能量动态, 追溯或预测红树林生物入侵的轨迹或响应动态。随着相同主题的独立研究持续增多, 多对象、多位点、多尺度的整合研究(如Meta-analysis)逐步实现, 将为寻求红树林生物入侵普遍规律性提供新的视角。
红树林处于海洋—陆地—河口过渡带, 是承受全球气候变化及其引起的海平面上升等影响的最前沿区域, 又是人类活动极为频繁和强烈的地带, 全球变化下的大尺度环境扰动将强烈影响红树林生物入侵。全球变化下, 深入探讨红树林入侵种多样性与格局、入侵效应、入侵机制、防治策略的变化动态及其驱动因素。明确全球变化对红树林生物入侵的整体效应与机制, 对于在剧烈环境变动下预测及应对生物入侵有重要意义。
红树林生物入侵研究的最终目的是对入侵现状的管理控制以及对未来入侵的预测。未来会不断开展红树林外来物种入侵风险及危害评估, 建立外来物种监测与预警体系, 科学、合理、谨慎引种, 引种后实施严格的监管, 增强研究人员、公众与管理决策人员之间信息共享, 交流协作, 增强生物入侵全民意识与参与性。今后, 健全的红树林生物入侵管理体系将在防治生物入侵实践中发挥主导作用。
红树林被誉为海岸线上的“绿色长城”, 保护着海岸带及其附属的一切。在全球化背景下, 人类活动促进了生物入侵的发生, 红树林生态系统结构单一、脆弱, 生物入侵将是其面临的一大威胁。详尽的红树林生物入侵研究是更好地预测和防治未来红树林及其他沿海生态系统生物入侵的基础。迄今, 红树林生物入侵相关研究远远落后于其他生态系统的同类研究, 红树林及其相关的沿海生态系统的保护与管理盲目而无力。红树林生态系统独特而珍稀, 只有对红树林生物入侵做深入透彻的研究, 获得全面的认识理解, 才能更好地实现红树林的保护与可持续利用。这需要对红树林感兴趣与热爱的研究者、公众和政府决策者的共同努力。
[31]
Elton CS 1958 ). The Ecology of Invasions by Animals and Plants
. Methuen, London.
[本文引用: 1]
[32]
Emery HE Fulweiler RW 2014 ). Spartina alterniflora and invasive Phragmites australis stands have similar green- house gas emissions in a New England marsh.
Aquatic Botany , 116 , 83 -92 .
[本文引用: 1]
[33]
Fisher AJ DiTomaso JM Gordon TR 2005 ). Intraspecific groups of Claviceps purpurea associated with grass species in Willapa Bay, Washington, and the prospects for biological control of invasive Spartina alterniflora.
Biological Control , 34 , 170 -179 .
[本文引用: 1]
[34]
Fourqurean JW Smith III TJ Possley J Collins TM Lee D Namoff S 2010 ). Are mangroves in the tropical Atlantic ripe for invasion? Exotic mangrove trees in the forests of South Florida.
Biological Invasions , 12 , 2509 -2522 .
[本文引用: 2]
[35]
Fry B Cormier N 2011 ). Chemical ecology of red mangroves, Rhizophora mangle, in the Hawaiian Islands.
Pacific Science , 65 , 219 -234 .
[本文引用: 1]
[36]
Grevstad FS Strong DR Garcia-Rossi D Switzer RW Wecker MS 2003 ). Biological control of Spartina alterniflora in Willapa Bay, Washington using the planthopper Prokelisia marginata: Agent specificity and early results.
Biological Control , 27 , 32 -42 .
[本文引用: 1]
[37]
Han QY Huang XP Shi P Zhang QM 2006 ). Coastal wetland in South China: Degradation trends, causes and protection countermeasures.
Chinese Science Bulletin , 51 , 121 -128 .
[本文引用: 1]
[38]
Han WD Li DJ Li Y Wu XF 2003 ). The soil dynamic study of restored Sonneratia apetala plantations. Journal of Nanjing Forestry University (
Natural Sciences Edition ), 27 (2 ), 49 -54 . (in Chinese with English abstract)
[本文引用: 1]
[韩维栋 , 凌大炯 , 李燕 , 吴小凤 (2003 ). 人工无瓣海桑林的土壤动态研究
. 南京林业大学学报(自然科学版) , 27 (2 ), 49 -54 .]
[本文引用: 1]
[39]
Holland BS Dawson MN Crow GL Hofmann DK 2004 ). Global phylogeography of Cassiopea (Scyphozoa: Rhizostomeae): Molecular evidence for cryptic species and multiple invasions of the Hawaiian Islands.
Marine Biology , 145 , 1119 -1128 .
[本文引用: 1]
[40]
Hong RB Lü XM Chen L Fang SH 2005 ). Benthos on mangrove wetland and smooth cordgrass (Spartina alterniflora) wetland in Jiulongjiang Estuary.
Journal of Oceanography In Taiwan Strait , 24 , 189 -194 . (in Chinese with English abstract)
[本文引用: 1]
[洪荣标 , 吕小梅 , 陈岚 , 方少华 (2005 ). 九龙江口红树林湿地与米草湿地的底栖生物
. 台湾海峡 , 24 , 189 -194 .]
[本文引用: 1]
[41]
Huang YL Fang XT Lu L Yan YB Chen SH Hu L Zhu CC Ge XJ Shi SH 2012 ). Transcriptome analysis of an invasive weed Mikania micrantha.
Biologia Plantarum , 56 , 111 -116 .
[本文引用: 1]
[42]
Isebor C Ajayi T Anyanwu A 2003 ). The incidence of Nypa fruticans (Wurmb) and its impact on fisheries production in the Niger Delta mangrove ecosystem
. In: 16th Annual Conference of the Fisheries Society of Nigeria (FISON). Maiduguri, Nigeria . 13 -16 .
[本文引用: 1]
[43]
Kathiresan K Bingham BL 2001 ). Biology of mangroves and mangrove ecosystems.
Advances in Marine Biology , 40 , 81 -251 .
[本文引用: 1]
[44]
Kong GH Wu QG Hu QM 2000 ). Exotic weed Mikania micrantha HBK appeared in South China.
Journal of Tropical Subtropical Botany , 8 , 27 .
[45]
Kueffer C Pyšek P Richardson DM 2013 ). Integrative invasion science: Model systems, multi-site studies, focused meta-analysis and invasion syndromes.
New Phytologist , 200 , 615 -633 .
[本文引用: 1]
[46]
Kulbicki M Beets J Chabanet P Cure K Darling E Floeter SR Galzin R Green A Harmelin-Vivien M Hixon M Letourneur Y de Loma TL McClanahan T McIlwain J MouTham G Myers R O’Leary JK Planes S Vigliola L Wantiez L 2012 ). Distributions of Indo-Pacific lionfishes Pterois spp. in their native ranges: Implications for the Atlantic invasion.
Marine Ecology Progress Series , 446 , 189 -205 .
[本文引用: 1]
[47]
Leung JYS Tam NFY 2013 ). Influence of plantation of an exotic mangrove species, Sonneratia caseolaris (L.) Engl., on macrobenthic infaunal community in Futian Mangrove National Nature Reserve, China.
Journal of Experimental Marine Biology and Ecology , 448 , 1 -9 .
[本文引用: 1]
[48]
Li B Chen JK 2002 ). Ecology of biological invasions: Achievements and challenges.
World Sci-Tech R & D , 24 (2 ), 26 -36 . (in Chinese with English abstract)
[本文引用: 2]
[李博 , 陈家宽 (2002 ). 生物入侵生态学: 成就与挑战
. 世界科技研究与发展 , 24 (2 ), 26 -36 .]
[本文引用: 2]
[49]
Li B Liao CH Zhang XD Chen HL Wang Q Cheng ZY Gan XJ Wu JH Zhao B Ma ZJ Chen XL Jiang LF Chen JK 2009 ). Spartina alterniflora invasions in the Yangtze River estuary, China: An overview of current status and ecosystem effects.
Ecological Engineering , 35 , 511 -520 .
[本文引用: 1]
[50]
Li FL Yang Q Zan QJ Tam NFY Shin PKS Vrijmoed LLP Cheung SG 2011 ). Differences in leaf construction cost between alien and native mangrove species in Futian, Shenzhen, China: Implications for invasiveness of alien species.
Marine Pollution Bulletin , 62 , 1957 -1962 .
[本文引用: 1]
[51]
Li FR Chen JQ Chen MR Yu YN Chen LY Li J Peng SL 2007 ). Research progress in the control of Smooth cordgrass.
Ecology and Environment , 16 , 1795 -1800 . (in Chinese with English abstract)
[本文引用: 2]
[1]
Allen JA 1998 ). Mangroves as alien species: The case of Hawaii.
Global Ecology and Biogeography Letters , 7 , 61 -71 .
[本文引用: 2]
[2]
Alvizu A Díaz MC Bastidas C Rützler K Thacker RW Márquez LM 2013 ). A skeleton-less sponge of Caribbean mangroves: Invasive or undescribed?
Invertebrate Biology , 132 , 81 -94 .
[本文引用: 1]
[51]
[李富荣 , 陈俊勤 , 陈沐荣 , 虞依娜 , 陈蕾伊 , 李静 , 彭少麟 (2007 ). 互花米草防治研究进展
. 生态环境 , 16 , 1795 -1800 .]
[本文引用: 2]
[52]
Li J Tian GH Peng SL Yang XB Chen LY Zeng Y Liu WH Li KH 2011 ). Allelopathy of Sonneratia apetala and Spartina alterniflora in soil.
Journal of Ecology and Rural Environment , 27 , 105 -108 . (in Chinese with English abstract)
[本文引用: 1]
[李静 , 田广红 , 彭少麟 , 杨雄邦 , 陈蕾伊 , 曾艳 , 刘文辉 , 何克宏 (2011 ). 无瓣海桑和互花米草群落土壤化感作用比较
. 生态与农村环境学报 , 27 , 105 -108 .]
[本文引用: 1]
[53]
Li M Liao BW Zheng SF 2003 ). Ecological Effect of Sonneratia apetala Plantation.
Shanghai Environmental Sciences , 22 , 540 -543 . (in Chinese with English abstract)
[3]
Arbeláez N Acero A 2011 ). Ocurrence of the lionfish Pterois volitans (Linnaeus) in the mangrove of Bahía de Chengue, Colombian Caribbean.
Boletín de Investigaciones Marinasy Costeras Invemar , 40 , 431 -435 .
[本文引用: 1]
[4]
Baker P Fajans JS Baker SM 2012 ). Habitat dominance of a nonindigenous tropical bivalve, Perna viridis (Linnaeus, 1758), in a subtropical estuary in the Gulf of Mexico.
Journal of Molluscan Studies , 78 , 28 -33 .
[本文引用: 1]
[53]
[李玫 , 廖宝文 , 郑松发 (2003 ). 无瓣海桑海滩人工林的生态影响
. 上海环境科学 , 22 , 540 -543 .]
[54]
Li M Liao BW Zheng SF Chen YJ 2004a ). Disturbance of directly introduction of Sonneratia apetala on the secondary Aegiceras corniculatum community.
Guang- dong Forestry Science and Technology , 20 (3 ), 19 -21 . (in Chinese with English abstract)
[本文引用: 1]
[5]
Barbour AB Allen MS Frazer TK Sherman KD 2011 ). Evaluating the potential efficacy of invasive lionfish (Pterois volitans) removals.
PLoS ONE , 6 , e19666 .
[本文引用: 1]
[6]
Barbour AB Montgomery ML Adamson AA Díaz-Ferguson E Silliman BR 2010 ). Mangrove use by the invasive lionfish Pterois volitans.
Marine Ecology Progress Series , 401 , 291 -294 .
[本文引用: 1]
[54]
[李玫 , 廖宝文 , 郑松发 , 陈玉军 (2004a ). 无瓣海桑的直接引入对次生桐花树群落的扰动
. 广东林业科技 , 20 (3 ), 19 -21 .]
[本文引用: 1]
[55]
Li M Liao BW Zheng SF Chen YJ 2004 b). Allelopathic effects of Sonneratia apetala aqueous extracts on growth performance of some indigenous mangroves.
Forest Research , 17 , 641 -645 . (in Chinese with English abstract)
[本文引用: 1]
[7]
Bishop MJ Krassoi FR Mcpherson RG Brown KR Summerhayes SA Wilkie EM O’connor WA 2010 ). Change in wild-oyster assemblages of Port Stephens, NSW, Australia, since commencement of non-native Pacific oyster (Crassostrea gigas) aquaculture.
Marine and Freshwater Research , 61 , 714 -723 .
[本文引用: 1]
[8]
Biswas SR Choudhury JK Nishat A Rahman MM 2007 ). Do invasive plants threaten the Sundarbans mangrove forest of Bangladesh?
Forest Ecology and Management , 245 , 1 -9 .
[本文引用: 2]
[55]
[李玫 , 廖宝文 , 郑松发 , 陈玉军 (2004 b). 无瓣海桑对乡土红树植物的化感作用
. 林业科学研究 , 17 , 641 -645 .]
[本文引用: 1]
[56]
Li WH Zhang CB Gao GJ Zan QJ Yang ZY 2007 ). Relationship between Mikania micrantha invasion and soil microbial biomass, respiration and functional diversity.
Plant and Soil , 296 , 197 -207 .
[本文引用: 1]
[9]
Bradley BA Blumenthal DM Early R Grosholz ED Lawler JJ Miller LP Sorte CJB D’Antonio CM Diez JM Dukes JS Ibanez I Olden JD 2012 ). Global change, global trade, and the next wave of plant invasions.
Frontiers in Ecology and the Environment , 10 , 20 -28 .
[本文引用: 1]
[10]
Bridgewater PB Cresswell ID 1999 ). Biogeography of mangrove and salt marsh vegetation: implications for conservation and management in Australia.
Mangroves and Salt Marshes , 3 , 117 -125 .
[本文引用: 1]
[57]
Li WH Zhang CB Jiang HB Xin GR Yang ZY 2006 ). Changes in soil microbial community associated with invasion of the exotic weed, Mikania micrantha H. B. K.
Plant and Soil , 281 , 309 -324 .
[本文引用: 1]
[58]
Li XC Yin ZY He LP Huang WH Zhou QW Zhang YA 2003 ). The effect of intervening artificially Mikania micrantha population.
Journal of Fujian College of Forestry , 23 , 214 -218 . (in Chinese with English abstract)
[本文引用: 1]
[11]
Chen BM Peng SL Ni GY 2009 ). Effects of the invasive plant Mikania micrantha H. B. K. on soil nitrogen availability through allelopathy in South China.
Biological Invasions , 11 , 1291 -1299 .
[本文引用: 1]
[12]
Chen L Deng ZF An SQ Zhao CJ Zhou CF Zhi YB 2007 ). Alternate irrigation of fresh and salt water restrains clonal growth and reproduction of Spartina alterniflora. Journal of Plant Ecology (
Chinese Version ), 31 , 645 -651 . (in Chinese with English abstract)
[本文引用: 1]
[58]
[李小川 , 殷作云 , 何立平 , 黄文辉 , 周启文 , 张贻安 (2003 ). 人为干预群落对薇甘菊的影响
. 福建林学院学报 , 23 , 214 -218 .]
[本文引用: 1]
[59]
Li Y Zheng DZ Chen HX Liao BW Zheng SF Chen XR 1998 ). Preliminary study on introduction of mangrove Sonneratia apetala Buch-Ham.
Forest Research , 11 , 39 -44 . (in Chinese with English abstract)
[本文引用: 1]
[12]
[陈琳 , 邓自发 , 安树青 , 赵聪蛟 , 周长芳 , 智颖飙 (2007 ). 淡咸水轮换浇灌抑制互花米草的克隆生长和繁殖
. 植物生态学报 , 31 , 645 -651 .]
[本文引用: 1]
[13]
Chen LY Peng SL Li J Lin ZG Zeng Y 2013 a). Competitive control of an exotic mangrove species: Restoration of native mangrove forests by altering light availability.
Restoration Ecology , 21 , 215 -223 .
[本文引用: 1]
[59]
[李云 , 郑德璋 , 陈焕雄 , 廖宝文 , 郑松发 , 陈相如 (1998 ). 红树植物无瓣海桑引种的初步研究
. 林业科学研究 , 11 , 39 -44 .]
[本文引用: 1]
[60]
Li ZF 2009 ). Preliminary Studies on Control of the Invasive Alien plant, Spartina alterniflora. Master degree dissertation
. Fujian Agriculture and Forestry University, Fuzhou. (in Chinese with English abstract)
[本文引用: 1]
[14]
Chen LZ Tam NFY Wang WQ Zhang YH Lin GH 2013 b). Significant niche overlap between native and exotic Sonneratia mangrove species along a continuum of varying inundation periods.
Estuarine, Coastal and Shelf Science , 117 , 22 -28 .
[本文引用: 1]
[15]
Chen XW Cai LZ Wu C Peng X Cao J Xu P Liu S Fu SJ 2012 ). Polychaete community in mangrove and salt marsh in Zhangjiang River Estuary, Fujian Province of East China.
Chinese Journal of Applied Ecology , 23 , 931 -938 . (in Chinese with English abstract)
[本文引用: 1]
[60]
[李兆飞 (2009 ). 外来入侵植物互花米草控制的初步研究
. 硕士学位论文, 福建农林大学, 福州.]
[本文引用: 1]
[61]
Lian JY Ye WH Cao HL Lai ZM Wang ZM Cai CX 2006 ). Influence of obligate parasite Cuscuta campestris on the community of its host Mikania micrantha.
Weed Research , 46 , 441 -443 .
[本文引用: 1]
[15]
[陈昕韡 , 蔡立哲 , 吴辰 , 彭欣 , 曹婧 , 许鹏 , 刘莎 , 傅素晶 (2012 ). 福建漳江口红树林和盐沼湿地的多毛类动物群落
. 应用生态学报 , 23 , 931 -938 .]
[本文引用: 1]
[16]
Chen ZY Gao H Wu H Li B 2005 ). Effects of simulated canopy shade on seed germination and seedlings growth of Spartina alterniflora and Scirpus mariqueter.
Hubei Agricultural Sciences , (2 ), 82 -84 . (in Chinese with English abstract)
[本文引用: 1]
[62]
Liang SC Liang MZ Wu YL Zan SJ Wang YJ Xie Q 2005 ). Analysis of the spatial structure of natural Sonneratia caseolaris + S. apetala forest in Futian, Shenzhen.
Guihaia , 25 , 393 -398 . (in Chinese with English abstract)
[本文引用: 1]
[梁士楚 , 梁铭忠 , 吴苑玲 , 昝启杰 , 王勇军 , 谢强 (2005 ). 深圳福田海桑+无瓣海桑自然林的空间结构分析
. 广西植物 , 25 , 393 -398 .]
[本文引用: 1]
[16]
[陈中义 , 高慧 , 吴涵 , 李博 (2005 ). 模拟遮荫对互花米草和海三棱草种子萌发及幼苗生长的影响
. 湖北农业科学 , (2 ), 82 -84 .]
[本文引用: 1]
[17]
Chimner RA Fry B Kaneshiro MY Cormier N 2006 ). Current extent and historical expansion of introduced mangroves on O’ahu, Hawai’i.
Pacific Science , 60 , 377 -383 .
[本文引用: 1]
[63]
Liao BW Zheng FS Chen YJ Li M Li YD 2004 ). Biological characteristics and ecological adaptability for non- indigenous mangrove species Sonneratia apetala.
Chinese Journal of Ecology , 23 , 10 -15 . (in Chinese with English abstract)
[本文引用: 1]
[廖宝文 , 郑松发 , 陈玉军 , 李玫 , 李意德 (2004 ). 外来红树植物无瓣海桑生物学特性与生态环境适应性分析
. 生态学杂志 , 23 , 10 -15 .]
[本文引用: 1]
[18]
Claydon JAB Calosso MC Traiger SB 2012 ). Progression of invasive lionfish in seagrass, mangrove and reef habitats.
Marine Ecology Progress Series , 448 , 119 -129 .
[本文引用: 1]
[19]
Cox EF Allen JA 1999 ). Stand structure and productivity of the introduced Rhizophora mangle in Hawaii.
Estuaries , 22 , 276 -284 .
[本文引用: 1]
[64]
Lowry E Rollinson EJ Laybourn AJ Scott TE Aiello- Lammens ME Gray SM Mickley J Gurevitch J 2013 ). Biological invasions: A field synopsis, systematic review, and database of the literature.
Ecology and Evolution , 3 , 182 -196 .
[本文引用: 2]
[65]
Lu QY Liu Y Li CH Wei XL Liu Y 2013 ). Impacts of alien species invasion on the South China Sea ecosystem and related control strategies.
Chinese Journal of Ecology , 32 , 2186 -2193 . (in Chinese with English abstract)
[本文引用: 2]
[20]
Daehler CC Strong DR 1996 ). Status, prediction and prevention of introduced cordgrass Spartina spp. invasions in Pacific estuaries, USA.
Biological Conservation , 78 , 51 -58 .
[本文引用: 1]
[21]
Demopoulos AWJ 2004 ). Aliens in Paradise: A Comparative Assessment of Introduced and Native Mangrove Benthic Community Composition, Food-web Structure, and Litter-fall Production
. PhD dissertation, Hawaii University, Hawaii, USA.
[本文引用: 1]
[65]
[陆琴燕 , 刘永 , 李纯厚 , 魏小岚 , 刘勇 (2013 ). 海洋外来物种入侵对南海生态系统的影响及防控对策
. 生态学杂志 , 32 , 2186 -2193 .]
[本文引用: 2]
[66]
Lu W Yang S Chen L Wang W Du X Wang C Ma Y Lin G Lin G 2014 ). Changes in carbon pool and stand structure of a native subtropical mangrove forest after inter-planting with exotic species Sonneratia apetala.
PLoS ONE , 9 , e91238 .
[本文引用: 1]
[22]
Demopoulos AWJ Fry B Smith CR 2007 ). Food web structure in exotic and native mangroves: A Hawaii-Puerto Rico comparison.
Oecologia , 153 , 675 -686 .
[本文引用: 1]
[23]
Demopoulos AWJ Smith CR 2010 ). Invasive mangroves alter macrofaunal community structure and facilitate opportunistic exotics.
Marine Ecology Progress Series , 404 , 51 -67 .
[本文引用: 1]
[67]
Lugo AE 1998 ). Mangrove forests: A tough system to invade but an easy one to rehabilitate.
Marine Pollution Bulletin , 37 , 427 -430 .
[本文引用: 1]
[68]
Mack RN Simberloff D Mark Lonsdale W Evans H Clout M Bazzaz FA 2000 ). Biotic invasions: causes, epidemiology, global consequences, and control.
Ecological Applications , 10 , 689 -710 .
[本文引用: 8]
[24]
D’iorio MM 2003 ). Mangroves and Shoreline Change on Molokai, Hawaii: Assessing the Role of Introduced Rhizophora Mangle in Sediment Dynamics and Coastal Change Using Remote Sensing and GIS . Harvard University Press , Cambridge, USA .
[本文引用: 1]
[25]
D’iorio MM Jupiter SD Cochran SA Potts DC 2007 ). Optimizing remote sensing and GIS tools for mapping and managing the distribution of an invasive mangrove (Rhizophora mangle) on South Molokai, Hawaii.
Marine Geodesy , 30 , 125 -144 .
[本文引用: 1]
[69]
Maja W Zerbe S Kuo YL 2008 ). Distribution and ecological range of the alien plant species Mikania micrantha Kunth (Asteraceae) in Taiwan.
Journal of Ecology and Environment , 31 , 277 -290 .
[本文引用: 1]
[70]
Mao ZL Lai MD Zhao ZY Yang XM 2011 ). Effect of invasion plants (Mikania micrantha H. B. K.) on carbon stock of mangrove ecosystem in Shenzhen Bay.
Ecology and Environmental Sciences , 20 , 1813 -1818 . (in Chinese with English abstract)
[本文引用: 1]
[毛子龙 , 赖梅东 , 赵振业 , 杨小毛 (2011 ). 薇甘菊入侵对深圳湾红树林生态系统碳储量的影响
. 生态环境 , 20 , 1813 -1818 .]
[本文引用: 1]
[71]
Miao CL Liao BW Zhu NH Guan W Li L He YX 2012 ). Seasonal variation of community of artificial restored mangrove in North Hezhou of Zhuhai invaded by other species.
Journal of Ecologic Science , 31 , 18 -21 . (in Chinese with English abstract)
[本文引用: 1]
[26]
Donnelly MJ Green DM Walters LJ 2008 ). Allelopathic effects of fruits of the Brazilian pepper Schinus terebinthifolius on growth, leaf production and biomass of seedlings of the red mangrove Rhizophora mangle and the black mangrove Avicennia germinans.
Journal of Experimental Marine Biology and Ecology , 357 , 149 -156 .
[本文引用: 1]
[27]
Du WQ Ma LN Liu J Zhang QF Zheng XF 2006 ). Controlling technique of Spartina alterniflora Loisel in mangrove-spartina ecotones.
Chinese Journal of Eco-Agriculture , 14 , 154 -156 . (in Chinese with English abstract)
[本文引用: 1]
[71]
[苗春玲 , 廖宝文 , 朱宁华 , 管伟 , 李琳 , 何奕雄 (2012 ). 珠海鹤洲北其他种入侵人工恢复红树林群落的季节动态
. 生态科学 , 31 , 18 -21 .]
[本文引用: 1]
[72]
Occhipinti-Ambrogi A Savini D 2003 ). Biological invasions as a component of global change in stressed marine ecosystems.
Marine Pollution Bulletin , 46 , 542 -551 .
[本文引用: 1]
[73]
Pan H Xue ZY Chen GR 2006 ). Whether or not the Sonneratla apetala B. Ham plantation caused biological invasion in Jiulong River Estuary.
Wetland Science & Management , 2 , 51 -55 . (in Chinese with English abstract)
[潘辉 , 薛志勇 , 陈国荣 (2006 ). 无瓣海桑造林是否造成九龙江口生物入侵的探讨
. 湿地科学与管理 , 2 , 51 -55 .]
[27]
[杜文琴 , 马丽娜 , 刘建 , 张秋芳 , 郑雪芳 (2006 ). 红树林区内互花米草防除技术研究
. 中国生态农业学报 , 14 , 154 -156 .]
[本文引用: 1]
[28]
Duke NC 1993 ). Mangrove floristics and biogeography. In: Robertson AI, Alongi DM eds. Tropical Mangrove Ecosystems . AGU Press , Washington . 63 -100 .
[本文引用: 1]
[74]
Prakash S Balamurugan J Kumar TTA Balasubramanian T 2012 ). Invasion and abundance of reef-inhabiting fishes in the Vellar estuary, southeast coast of India, especially the lionfish Pterois volitans Linnaeus.
Current Science , 103 , 941 -944 .
[本文引用: 1]
[75]
Puth LM Post DM 2005 ). Studying invasion: Have we missed the boat?
Ecology Letters , 8 , 715 -721 .
[本文引用: 1]
[76]
Ren H Guo QF Liu H Li J Zhang QM Xu HL Xu FH 2014 ). Patterns of alien plant invasion across coastal bay areas in southern China.
Journal of Coastal Research , 30 , 448 -455 .
[本文引用: 3]
[77]
Ren H Jian SG Lu HF Zhang QM Shen WJ Han WD Yin ZY Guo QF 2008 ). Restoration of mangrove plantations and colonisation by native species in Leizhou Bay, South China.
Ecological Research , 23 , 401 -407 .
[本文引用: 1]
[29]
Dukes JS Mooney HA 1999 ). Does global change increase the success of biological invaders?
Trends in Ecology & Evolution , 14 , 135 -139 .
[本文引用: 1]
[30]
Ellison CA Evans HC Djeddour DH Thomas SE 2008 ). Biology and host range of the rust fungus Puccinia spegazzinii: A new classical biological control agent for the invasive, alien weed Mikania micrantha in Asia.
Biological Control , 45 , 133 -145 .
[本文引用: 1]
[78]
Ren H Lu HF Shen WJ Huang C Guo QF Li ZA Jian SG 2009 ). Sonneratia apetala Buch. Ham in the mangrove ecosystems of China: An invasive species or restoration species?
Ecological Engineering , 35 , 1243 -1248 .
[本文引用: 1]
[79]
Ren MX Zhang QG 2009 ). The relative generality of plant invasion mechanisms and predicting future invasive plants.
Weed Research , 49 , 449 -460 .
[本文引用: 1]
[80]
Richardson DM Pyšek P Rejmánek M Barbour MG Panetta FD West CJ 2000 ). Naturalization and invasion of alien plants: Concepts and definitions.
Diversity and Distributions , 6 , 93 -107 .
[本文引用: 1]
[81]
Ruiz GM Fofonoff PW Carlton JT Wonham MJ Hines AH 2000 ). Invasion of coastal marine communities in North America: Apparent patterns, processes, and biases.
Annual Review of Ecology and Systematics , 31 , 481 -531 .
[本文引用: 2]
[82]
Sakai AK Allendorf FW Holt JS Lodge DM Molofsky J With KA Baughman S Cabin RJ Cohen JE Ellstrand NC McCauley DE O’Neil P Parker IM Thompson JN Weller SG 2001 ). The population biology of invasive species.
Annual Review of Ecology and Systematics , 32 , 305 -332 .
[本文引用: 2]
[83]
Salgado-Barragán J Toledano-Granados A 2006 ). The false mussel Mytilopsis adamsi Morrison, 1946 (Mollusca: Bivalvia: Dreissenidae) in the Pacific waters of Mexico: A case of biological invasion.
Hydrobiologia , 563 , 1 -7 .
[本文引用: 1]
[84]
Shao H Peng SL Liu YX Zhang C Xiang YC 2002 ). The biological control and the natural enemy of Mikania micrantha H. B. K in China.
Ecologic Science , 21 , 33 -36 . (in Chinese with English abstract)
[本文引用: 1]
[邵华 , 彭少麟 , 刘运笑 , 张弛 , 向言词 (2002 ). 薇甘菊的生物防治及其天敌在中国的新发现
. 生态科学 , 21 , 33 -36 .]
[本文引用: 1]
[85]
Shen H Hong L Chen H Ye WH Cao HL Wang ZM 2011 ). The response of the invasive weed Mikania micrantha to infection density of the obligate parasite Cuscuta campestris and its implications for biological control of M. micrantha.
Botanical Studies , 52 , 89 -97 .
[本文引用: 1]
[86]
Silliman BR Zieman JC 2001 ). Top-down control of Spartina alterniflora production by periwinkle grazing in a Virginia salt marsh.
Ecology , 82 , 2830 -2845 .
[本文引用: 1]
[87]
Siple MC Donahue MJ 2013 ). Invasive mangrove removal and recovery: Food web effects across a chronosequence.
Journal of Experimental Marine Biology and Ecology , 448 , 128 -135 .
[本文引用: 1]
[88]
Soares M 1999 ). Structure and degree of perturbation of mangroves at Tijuca Lagoon, Rio de Janeiro, RJ, Brazil.
Revista Brasileira de Biologia , 59 , 503 -515 .
[89]
Soti PG Jayachandran K Purcell M Volin JC Kitajima K 2014 ). Mycorrhizal symbiosis and Lygodium microphy- llum invasion in South Florida—A biogeographic compar- ison.
Symbiosis , 62 , 81 -90 .
[本文引用: 1]
[90]
Sweetman AK Middelburg JJ Berle AM Bernardino AF Schander C Demopoulos AWJ Smith CR 2010 ). Impacts of exotic mangrove forests and mangrove deforestation on carbon remineralization and ecosystem functioning in marine sediments.
Biogeosciences , 7 , 2129 -2145 .
[本文引用: 1]
[91]
Tan YY 2009 ). Ecological Study of Spartina Alterniflora in Mangroves Habitats
. Master degree dissertation, Guangxi Normal University, Guilin. (in Chinese with English abstract)
[覃盈盈 (2009 ). 红树林生境中互花米草的生态学研究
. 硕士学位论文, 广西师范大学, 桂林 .]
[92]
Tang GL Shen LH Weng WH Zhang JE Liao BW Liu JL Teng XS 2007 ). Effects of using Sonneratia apetala to control the growth of Spartina alterniflora Loisel.
Journal of South China Agricultural University , 28 , 10 -13 . (in Chinese with English abstract)
[本文引用: 1]
[唐国玲 , 沈禄恒 , 翁伟花 , 章家恩 , 廖宝文 , 刘金苓 , 滕兴顺 (2007 ). 无瓣海桑对互花米草的生态控制效果
. 华南农业大学学报 , 28 , 10 -13 .]
[本文引用: 1]
[93]
Theoharides KA Dukes JS 2007 ). Plant invasion across space and time: Factors affecting nonindigenous species success during four stages of invasion.
New Phytologist , 176 , 256 -273 .
[本文引用: 2]
[94]
Tripathi RS Khan ML Yadav AS 2011 ). Biology of Mikania micrantha H. B. K.: A review. In: Bhatt JR, Singh JS, Singh SP, Tripathi RS, Kohli RK eds. Invasive Alien Plants: An Ecological Appraisal for the Indian Subcontinent . CAB International , Oxford, UK . 99 -107 .
[本文引用: 1]
[95]
Vannini A Franceschini S Vettraino AM 2012 ). Manu- factured wood trade to Europe: A potential uninspected carrier of alien fungi.
Biological Invasions , 14 , 1991 -1997 .
[本文引用: 1]
[96]
Wan HW Wang Q Jiang D Fu JY Yang YP Liu XM 2014 ). Monitoring the invasion of Spartina alterniflora using very high resolution unmanned aerial vehicle imagery in Beihai, Guangxi (China).
The Scientific World Journal , 2014 , 1 -7 .
[本文引用: 1]
[97]
Wang AA Sun X Cai JB Chai XL Liu J Shang X 2014 ). Impacts of invasion plant (Spartina alterniflora) on macro-benthic fauna community in mangroves tidal flat.
Zhejiang Nongye Kexue , (4 ), 572 -577 . (in Chinese)
[本文引用: 1]
[王安安 , 孙雪 , 蔡景波 , 柴雪良 , 刘健 , 商栩 (2014 ). 互花米草入侵对红树林湿地潮滩大型底栖动物群落的影响
. 浙江农业科学 , (4 ), 572 -577 .]
[本文引用: 1]
[98]
Wang AJ Chen J Li DY 2008 ). Impact of Spartina alterniflora on sedimentary environment of coastal wetlands of the Quanzhou Bay.
The Ocean Engineering , 26 (4 ), 60 -69 . (in Chinese with English abstract)
[本文引用: 1]
[王爱军 , 陈坚 , 李东义 (2008 ). 互花米草对福建泉州湾海岸湿地沉积环境影响
. 海洋工程 , 26 (4 ), 60 -69 .]
[本文引用: 1]
[99]
Wang M Gao XQ Wang WQ 2014 ). Differences in burrow morphology of crabs between Spartina alterniflora marsh and mangrove habitats.
Ecological Engineering , 69 , 213 -219 .
[本文引用: 1]
[100]
Wang Q An SQ Ma ZJ Zhan B Chen JK Li B 2006 ). Invasive Spartina alterniflora: Biology, ecology and management.
Acta Phytotaxonomica Sinica , 44 , 559 -588 . (in Chinese with English abstract)
[本文引用: 2]
[王卿 , 安树青 , 马志军 , 赵斌 , 陈家宽 , 李博 (2006 ). 入侵植物互花米草——生物学、生态学及管理
. 植物分类学报 , 44 , 559 -588 .]
[本文引用: 2]
[101]
Wang RL Ding LW Sun QY Li J Xu ZF Peng SL 2008 ). Genome sequence and characterization of a new virus infecting Mikania micrantha H. B. K.
Archives of Virology , 153 , 1765 -1770 .
[本文引用: 1]
[102]
Wang RL Staehelin C Peng SL Wang WT Xie XM Lu HN 2010 ). Responses of Mikania micrantha, an invasive weed to elevated CO2 : Induction of β-caryophyllene synthase, changes in emission capability and allelopathic potential of β-caryophyllene.
Journal of Chemical Ecology , 36 , 1076 -1082 .
[本文引用: 1]
[103]
Wang T Chen GP Zan QJ Wang CB Su YJ 2012 ). AFLP genome scan to detect genetic structure and candidate loci under selection for local adaptation of the invasive weed Mikania micrantha.
PLoS ONE , 7 , e41310 .
[本文引用: 1]
[104]
Wang X Ma ZY Yang H Zhou GY Lou J 2008 ). Structural features of artificial Sonneratia apetala-S. caseolaris community at Dongzhaigang Harbor, Hainan Island.
Chinese Journal of Tropical Crops , 29 , 374 -379 . (in Chinese with English abstract)
[本文引用: 1]
[王旭 , 马宗耀 , 杨怀 , 周光益 , 楼坚 (2008 ). 海南东寨港无瓣海桑——海桑人工红树群落结构特征
. 热带作物学报 , 29 , 374 -379 .]
[本文引用: 1]
[105]
Wang YJ Zan QJ 2001 ). The birds community and ecological evaluation of Sonneration apetala + Sonneration casedaris mangrove plantation.
Ecologic Science , 20 , 41 -46 . (in Chinese with English abstract)
[本文引用: 1]
[王勇军 , 昝启杰 (2001 ). 深圳福田无瓣海桑与海桑人工林鸟类群落研究及生态评价
. 生态科学 , 20 , 41 -46 .]
[本文引用: 1]
[106]
Wang ZC Zhang YM Pan XY Ma ZJ Chen JK Li B 2006 ). Effects of winter burning and cutting on aboveground growth and reproduction of Spartina alterniflora: A field experiment at Chongming Dongtan, Shanghai.
Biodiversity Science , 14 , 275 -283 . (in Chinese with English abstract)
[本文引用: 1]
[王智晨 , 张亦默 , 潘晓云 , 马志军 , 陈家宽 , 李博 (2006 ). 冬季火烧与收割对互花米草地上部分生长与繁殖的影响
. 生物多样性 , 14 , 275 -283 .]
[本文引用: 1]
[107]
Waterhouse DF 1994 ). Biological Control of Weeds: Southeast Asian Prospects
. ACIAR, Canberra.
[本文引用: 3]
[108]
Wen ZZ Liao ZL Li P Shang X 2014 ). Impacts of invasion plant (Spartina alterniflora) on the benthic food-web in Ximen Island, Zhejiang, China.
Zhejiang Nongye Kexue, (5 ), 740 -744 , 752 . (in Chinese)
[本文引用: 1]
[温作柱 , 廖忠鹭 , 李平 , 商栩 (2014 ). 互花米草入侵对西门岛潮滩底栖食物网的影响
. 浙江农业科学 , (5 ), 740 -744 , 752 .]
[本文引用: 1]
[109]
Wester L 1981 ). Introduction and spread of mangroves in the Hawaiian Islands.
Yearbook of the Association of Pacific Coast Geographers , 43 , 125 -137 .
[本文引用: 1]
[110]
Wu C Cai LZ Chen XW Guo T Zhuo Y Fu SJ Peng X Cao J 2013 ). Secondary production of macrofauna in mangrove and salt marsh of Zhangjiang River Estuary, Fujian. Journal of Xiamen University (
Natural Science ), 52 , 259 -266 . (in Chinese with English abstract)
[本文引用: 1]
[吴辰 , 蔡立哲 , 陈昕韡 , 郭涛 , 卓异 , 傅素晶 , 彭欣 , 曹婧 (2013 ). 福建漳江口红树林和盐沼湿地大型底栖动物次级生产力研究
. 厦门大学学报(自然科学版) , 52 , 259 -266 .]
[本文引用: 1]
[111]
Wu H Ding JQ 2014 ). Recent progress in invasion ecology.
Chinese Science Bulletin , 59 , 438 -448 . (in Chinese with English abstract)
[本文引用: 1]
[吴昊 , 丁建清 (2014 ). 入侵生态学最新研究动态
. 科学通报 , 59 , 438 -448 .]
[本文引用: 1]
[112]
Xiao DR Zhang LQ Zhu ZC 2010 ). The range expansion patterns of Spartina alterniflora on salt marshes in the Yangtze Estuary, China.
Estuarine, Coastal and Shelf Science , 88 , 99 -104 .
[本文引用: 1]
[113]
Xin K Zhou Q Arndt SK Yang X 2013 ). Invasive capacity of the mangrove Sonneratia apetala in Hainan Island, China.
Journal of Tropical Forest Science , 25 , 70 -78 .
[本文引用: 1]
[114]
Yao LQ 2010 ). Effects of introduced plant (Sonneratia apetala) on native mangroves in Jiulong River estuary.
Forestry Prospect and Design , (2 ), 76 -78 . (in Chinese)
[本文引用: 1]
[姚历强 (2010 ). 无瓣海桑引入对龙海九龙江口乡土红树林影响的调查
. 林业勘察设计 , (2 ), 76 -78 .]
[本文引用: 1]
[115]
Yin ZY Li XC He LP Huang WH Zhou YW Li RG Xu QH Zhang YA Luo H 2003 ). A preliminary study on ecological control of Mikania micrantha H. B. K.
Guangdong Forestry Science and Technology , 19 , 17 -22 . (in Chinese with English abstract)
[本文引用: 1]
[殷祚云 , 李小川 , 何立平 , 黄文辉 , 周永文 , 黎容根 , 徐庆华 , 张贻安 , 罗航 (2003 ). 薇甘菊生态防除研究初报
. 广东林业科技 , 19 , 17 -22 .]
[本文引用: 1]
[116]
Yu XM Yang FJ 2011 ). Ecological characters and invasion route of Mikania micrantha in Shenzhen Bay.
Journal of Northeast Forestry University , 39 , 51 -52, 88 . (in Chinese with English abstract)
[本文引用: 1]
[于晓梅 , 杨逢建 (2011 ). 薇甘菊在深圳湾的入侵路线及其生态特征
. 东北林业大学学报 , 39 , 51 -52, 88 .]
[本文引用: 1]
[117]
Yuan L Zhang LQ Xiao DR Zhang J Wang RZ Yuan LQ Gu ZQ Chen X Ping Y Zhu ZC 2008 ). A demonstration study using the integrated technique of cutting plus waterlogging for the control of Spartina alterniflora.
Acta Ecologica Sinica , 28 , 5723 -5730 . (in Chinese with English abstract)
[本文引用: 1]
[袁琳 , 张利权 , 肖德荣 , 张杰 , 王睿照 , 袁连奇 , 古志钦 , 陈曦 , 平原 , 祝振昌 (2008 ). 刈割与水位调节集成技术控制互花米草(Spartina alterniflora)
. 生态学报 , 28 , 5723 -5730 .]
[本文引用: 1]
[118]
Zan QJ Sun YJ Liao WB Li MG Wang BS 2007 ). Chemical herbicide salfometuron-Methyl controlling Mikania micrantha and its residue in the soil.
Acta Ecologica Sinica , 27 , 3407 -3416 . (in Chinese with English abstract)
[本文引用: 3]
[昝启杰 , 孙延军 , 廖文波 , 李鸣光 , 王伯荪 (2007 ). 森草净杀灭薇甘菊(Mikania micrantha)及其安全性
. 生态学报 , 27 , 3407 -3416 .]
[本文引用: 3]
[119]
Zan QJ Wang BS Wang YJ Li MG 2003 ). Ecological assessment on the introduced Sonneratia caseolaris and S. apetala at the mangrove forest of Shenzhen Bay, China.
Acta Botanica Sinica , 45 , 544 -551 .
[本文引用: 1]
[120]
Zan QJ Wang BS Wang YJ Zhang JL Liao WB Li MG 2003 ). The harm caused by Mikania micrantha and its control by Cuscuta campestris.
Acta Phytoecologica Sinica , 27 , 822 -828 . (in Chinese with English abstract)
[昝启杰 , 王伯荪 , 王勇军 , 张军丽 , 廖文波 , 李鸣光 (2003 ). 薇甘菊的危害与田野菟丝子的防除作用
. 植物生态学报 , 27 , 822 -828 .]
[121]
Zan QJ Wang YJ Wang BS Liao WB Li MG 2000 ). The distribution and harm of the exotic weed Mikania micrantha.
Chinese Journal of Ecology , 19 (6 ), 58 -61, 77 . (in Chinese with English abstract)
[昝启杰 , 王勇军 , 王伯荪 , 廖文波 , 李鸣光 (2000 ). 外来杂草薇甘菊的分布及危害
. 生态学杂志 , 19 (6 ), 58 -61, 77 .]
[122]
Zeng WJ Liao BW Chen XR Li J Ma SQ Guan W 2008 ). The ecological effect of mangrove Sonneratia apetala mixed with three local mangrove species.
Ecological Science , 27 , 31 -37 . (in Chinese with English abstract)
[本文引用: 1]
[曾雯珺 , 廖宝文 , 陈先仁 , 粟娟 , 马苏全 , 管伟 (2008 ). 无瓣海桑与三种乡土红树植物混交的生态效应
. 生态科学 , 27 , 31 -37 .]
[本文引用: 1]
[123]
Zhang LY Ye WH Cao HL Feng HL 2004 ). Mikania micrantha H. B. K. in China―An overview.
Weed Research , 44 , 42 -49 .
[本文引用: 3]
[124]
Zhang QF Peng JJ Chen Q Li XF Xu CY Yin HB Yu S 2011 ). Impacts of Spartina alterniflora invasion on abundance and composition of ammonia oxidizers in estuarine sediment.
Journal of Soils and Sediments , 11 , 1020 -1031 .
[本文引用: 1]
[125]
Zhang QM Yu HB Chen XS Zheng DZ 1997 ). The relationship between mangrove zone on tidal flats and tidal levels.
Acta Ecologica Sinica , 17 , 258 -265 . (in Chinese with English abstract)
[本文引用: 1]
[张乔民 , 于红兵 , 陈欣树 , 郑德璋 (1997 ). 红树林生长带与潮汐水位关系的研究
. 生态学报 , 17 , 258 -265 .]
[本文引用: 1]
[126]
Zhang XL Shi SL Pan GX Li LQ Zhang XH Li ZP 2008 ). Changes in eco-chemical properties of a mangrove wetland under spartina invasion from Zhangjiangkou, Fujian, China.
Advances in Earth Science , 23 , 974 -981 . (in Chinese with English abstract)
[本文引用: 1]
[张祥霖 , 石盛莉 , 潘根兴 , 李恋卿 , 张旭辉 , 李志鹏 (2008 ). 互花米草入侵下福建漳江口红树林湿地土壤生态化学变化
. 地球科学进展 , 23 , 974 -981 .]
[本文引用: 1]
[127]
Zhang YH Huang GM Wang WQ Chen LZ Lin GH 2012 ). Interactions between mangroves and exotic Spartina in an anthropogenically disturbed estuary in southern China.
Ecology , 93 , 588 -597 .
[本文引用: 1]
[128]
Zhang Y Zhang H 2008 ). The research on spatial patterns and damage of Mikania micrantha at Qi’ao Mangrove Island in Zhuhai City.
Progress in Modern Biomedicine , 8 , 713 -716 . (in Chinese with English abstract)
[张阳 , 张华 (2008 ). 珠海淇澳岛红树林湿地薇甘菊危害情况和发生因素研究
. 现代生物医学进展 , 8 , 713 -716 .]
[129]
Zhou QQ Chen ZL Xin K 2010 ). Research on the alien invasive status of mangrove in China.
Journal of Anhui Agricultural Sciences , 38 , 2662 -2664 . (in Chinese with English abstract)
[本文引用: 2]
[周青青 , 陈志力 , 辛琨 (2010 ). 我国红树林外来入侵现状研究综述
. 安徽农业科学 , 38 , 2662 -2664 .]
[本文引用: 2]
[130]
Zhu HW 2012 ). Sonneratia Apetala Invasion Risk Compreh- ensive Assessment in Qi’ao Island, Zhuhai
. Master degree dissertation, Chinese Academy of Forestry Sciences, Beijing. (in Chinese with English abstract)
[本文引用: 1]
[朱宏伟 (2012 ). 珠海淇澳岛无瓣海桑入侵风险综合评估
. 硕士学位论文, 中国林业科学研究院, 北京.]
[本文引用: 1]
The Ecology of Invasions by Animals and Plants
1
1958
... 当非土著种进入一个过去不曾分布的地区, 并能存活、繁殖而建立种群, 其种群的进一步扩散已经或将要造成明显的生态环境和经济后果, 这一过程称为生物入侵(biological invasion).生物入侵强调外来入侵种的地理属性及其造成的影响(Elton, 1958 ; 李博和陈家宽, 2002 ). ...
Spartina alterniflora and invasive Phragmites australis stands have similar green- house gas emissions in a New England marsh.
1
2014
... 互花米草有200多年的入侵扩张历史.不同国家和地区对互花米草的入侵现状、入侵效应、入侵原因与机制, 以及相关防治措施都有过全面而系统的研究(Daehler & Strong, 1996 ; Li et al ., 2009 ).现阶段对互花米草入侵的研究, 一方面是从分子水平揭示互花米草成功入侵的遗传学基础、耐受性以及表型可塑性的分子机理或原因(吴昊和丁建清, 2014 ); 另一方面是应用3S技术, 从景观以上尺度监测互花米草入侵的动态变化(Wan et al ., 2014 ).互花米草入侵不可避免地要受全球变化影响, 二者的相互反馈在全球变化日益强烈的今天愈加受到重视(Emery & Fulweiler, 2014 ).随着互花米草在海岸滩涂的快速扩散, 海岸带上所有的生态系统不可避免地会受到互花米草入侵的威胁.互花米草对潮间带生境的高度适应性, 使之在入侵地扩散, 并成功地入侵红树林生态系统(Zhang et al ., 2012 ). ...
Intraspecific groups of Claviceps purpurea associated with grass species in Willapa Bay, Washington, and the prospects for biological control of invasive Spartina alterniflora.
1
2005
... 目前研究显示, 玉黍螺(Littoraria irrorata )、麦角菌(Ciavieps purpurea )和光蝉(Prokelisia marginata )在互花米草生物控制中得到应用(王卿等, 2006 ).玉黍螺直接取食互花米草的叶片, 抑制其生长(Silliman & Zieman, 2001 ).麦角菌能使互花米草感染麦角病, 在种子内形成菌核, 降低其种子产量, 限制其扩散(Fisher et al ., 2005 ).但麦角菌对禾本科其他一些植物也有影响, 而且其适应性与变异性强, 因此引用麦角菌作为生物控制途径还有待进一步研究(李富荣等, 2007 ).还有一种从华盛顿引入的光蝉, 对入侵威拉帕湾的互花米草具有很强的杀伤力, 对其他植物无明显影响(Grevstad et al ., 2003 ). ...
Are mangroves in the tropical Atlantic ripe for invasion? Exotic mangrove trees in the forests of South Florida.
2
2010
... 早期研究认为, 由于红树林生态系统极为苛刻的生境条件, 很少有外来物种能适应环境生存并形成入侵(Lugo, 1998 ).但在全球变化下, 尤其在人类活动的参与下, 生物进入到新生境的概率及入侵成功率大幅提升, 几乎没有什么生境能避免生物入侵(Dukes & Mooney, 1999 ; Occhipinti-Ambrogi & Savini, 2003 ).当前研究显示, 日渐增多的外来种已经出现在红树林生态系统中, 有些已造成明显的入侵危害(Bridgewater & Cresswell, 1999 ; Biswas et al ., 2007 ; Ren et al ., 2014 ).红树林生态系统植物物种少, 群落结构单一, 系统稳定性差, 被入侵破坏后难以恢复.与此同时, 红树林生态系统的组成物种也有可能入侵其他非土著地区, 改变这些地区原有的生态系统(Allen, 1998 ; Fourqurean et al ., 2010 ). ...
... 近来研究发现热带大西洋区红树植物物种萎缩, 而其与印度洋-太平洋区红树植物亲缘关系很近, 将来容易被此区域的红树植物入侵(Fourqurean et al ., 2010 ).可以预见, 美国东部沿海红树林区将是以后外来植物入侵红树林的热点区域.今后, 结合红树植物对夏威夷群岛的入侵, 对比红树植物入侵岛屿与入侵大陆的差异性, 对入侵生态学将会有更全面的认识. ...
Chemical ecology of red mangroves, Rhizophora mangle, in the Hawaiian Islands.
1
2011
... 外来红树植物Rhizophora mangle 入侵夏威夷海岛无植被裸滩, 显著改变了这里原有的生态系统结构与功能.红树群落高的生产力大幅增加了整个区域的生物量(Cox & Allen, 1999 ; Fry & Cormier, 2011 ; D’iorio, 2003 ).Rhizophora mangle 对地下部分的凋落物输入, 改善了沉积物的理化特征, 并改变了原有的食物链(网)结构, 丰富了底栖生物群落(Demopoulos, 2004 ; Demopoulos et al ., 2007 ; Siple & Donahue, 2013 ).底栖生物群落的丰富多样, 为进一步的外来生物入侵提供了优良的条件, 形成一个外来生物入侵循环(Demopoulos & Smith, 2010 ).外来红树植物入侵也对底栖生态系统功能产生了深远的影响, 夏威夷群岛海岸的沉积物代谢和碳再矿化动态在移除入侵种Rhizophora mangle 数年后仍没有摆脱其影响(Sweetman et al ., 2010 ). ...
Biological control of Spartina alterniflora in Willapa Bay, Washington using the planthopper Prokelisia marginata: Agent specificity and early results.
1
2003
... 目前研究显示, 玉黍螺(Littoraria irrorata )、麦角菌(Ciavieps purpurea )和光蝉(Prokelisia marginata )在互花米草生物控制中得到应用(王卿等, 2006 ).玉黍螺直接取食互花米草的叶片, 抑制其生长(Silliman & Zieman, 2001 ).麦角菌能使互花米草感染麦角病, 在种子内形成菌核, 降低其种子产量, 限制其扩散(Fisher et al ., 2005 ).但麦角菌对禾本科其他一些植物也有影响, 而且其适应性与变异性强, 因此引用麦角菌作为生物控制途径还有待进一步研究(李富荣等, 2007 ).还有一种从华盛顿引入的光蝉, 对入侵威拉帕湾的互花米草具有很强的杀伤力, 对其他植物无明显影响(Grevstad et al ., 2003 ). ...
Coastal wetland in South China: Degradation trends, causes and protection countermeasures.
1
2006
... 微甘菊入侵后, 通常给当地生态系统带来了毁灭性灾害, 因此对微甘菊的防治需求十分紧迫(Waterhouse, 1994 ).物理和化学防治手段的弊端促使学者寻求生物手段来防治微甘菊入侵.业已证实, 寄生植物野地菟丝子(Cuscuta campestris )能安全有效地防治微甘菊(Lian et al ., 2006 ; Shen et al ., 2011 ), 一些新的微生物或病毒也能用来控制微甘菊(Ellison et al ., 2008 ; Wang et al ., 2008 ).除关注微甘菊对生态系统地上植物群落的危害外, 对生态系统其他结构(如地下生物)和功能的效应的关注也逐步形成(Li et al ., 2006 , 2007 ).微甘菊入侵机制的研究日渐成熟(Zhang et al ., 2004 ; Chen et al ., 2009 ).有关微甘菊入侵的分子手段研究(Huang et al ., 2012 ; Wang et al ., 2012 ), 以及微甘菊对全球气候变化的响应研究(Wang et al ., 2010 )初现端倪.随着微甘菊的持续入侵扩散, 温暖湿润沿海地区的红树林生态系统也难逃其入侵危害(Han et al ., 2006 ). ...
人工无瓣海桑林的土壤动态研究
1
2003
... 无瓣海桑在引入地多用作沿海涂滩造林与恢复树种, 具有明显的改良土壤效应, 可增加土壤有机质和N、P、K含量, 有利于后续乡土红树植物定植(韩维栋等, 2003 ).在乡土红树林群落中混种无瓣海桑, 虽然整个林分生物量或土壤碳库没有显著增加, 但无瓣海桑相对较高的凋落物量和低C/N比, 有利于分解者利用(Lu et al ., 2014 ).无瓣海桑的引入能提高群落结构的复杂性与生物多样性.在深圳福田红树林保护区, 混种无瓣海桑+海桑人工林, 相对于天然红树林, 鸟类群落生物多样性得到提高(王勇军和昝启杰, 2001 ).鉴于上述研究结果, 一些学者认为无瓣海桑并没有表现出入侵危害(Zan et al ., 2003 ; 朱宏伟, 2012 ; Xin et al ., 2013 ). ...
人工无瓣海桑林的土壤动态研究
1
2003
... 无瓣海桑在引入地多用作沿海涂滩造林与恢复树种, 具有明显的改良土壤效应, 可增加土壤有机质和N、P、K含量, 有利于后续乡土红树植物定植(韩维栋等, 2003 ).在乡土红树林群落中混种无瓣海桑, 虽然整个林分生物量或土壤碳库没有显著增加, 但无瓣海桑相对较高的凋落物量和低C/N比, 有利于分解者利用(Lu et al ., 2014 ).无瓣海桑的引入能提高群落结构的复杂性与生物多样性.在深圳福田红树林保护区, 混种无瓣海桑+海桑人工林, 相对于天然红树林, 鸟类群落生物多样性得到提高(王勇军和昝启杰, 2001 ).鉴于上述研究结果, 一些学者认为无瓣海桑并没有表现出入侵危害(Zan et al ., 2003 ; 朱宏伟, 2012 ; Xin et al ., 2013 ). ...
Global phylogeography of Cassiopea (Scyphozoa: Rhizostomeae): Molecular evidence for cryptic species and multiple invasions of the Hawaiian Islands.
1
2004
... 此外, 一些软体、海绵或腔肠动物(如牡蛎、贻贝、海绵等)由于人为活动传入非土著地区, 暂时还没有大肆爆发形成入侵危害(Holland et al ., 2004 ; Salgado-Barragán & Toledano-Granados, 2006 ; Bishop et al ., 2010 ; Baker et al ., 2012 ; Alvizu et al ., 2013 ); 还有一些入侵非土著海洋的动物是否入侵红树林尚不明确(Ruiz et al ., 2000 ; 陆琴燕等, 2013 ), 有待今后持续关注. ...
九龙江口红树林湿地与米草湿地的底栖生物
1
2005
... 互花米草入侵还会改变红树林的生物多样性及其行为模式.在九龙江口红树林区, 互花米草入侵区域底栖动物的栖息密度、种类多样性、丰度比红树林区高, 比涂滩要低(洪荣标等, 2005 ).互花米草的入侵还改变着氨氧化古菌与细菌的丰度, 影响氨氧化微生物的群落结构(Zhang et al ., 2011 ).在漳江口红树林区, 红树林生境与互花米草生境之间多毛类动物栖息密度、生物量、丰度指数、均匀度指数和多样性指数均有显著差异(陈昕韡等, 2012 ).红树林与互花米草生境大型底栖动物次级生产力也存在差异, 海榄雌(Avicennia marina )生境大型底栖动物的年次级生产力最高, 而互花米草生境大型底栖动物的P/B值(P: 平均次级生产力; B: 平均生物量)最高, 因其优势类群为个体较小的寡毛类(吴辰等, 2013 ).再有, 互花米草相对较高的根密度显著降低了红树林中蟹类在互花米草生境打洞的深度及洞穴的复杂性(Wang et al ., 2014 ). ...
九龙江口红树林湿地与米草湿地的底栖生物
1
2005
... 互花米草入侵还会改变红树林的生物多样性及其行为模式.在九龙江口红树林区, 互花米草入侵区域底栖动物的栖息密度、种类多样性、丰度比红树林区高, 比涂滩要低(洪荣标等, 2005 ).互花米草的入侵还改变着氨氧化古菌与细菌的丰度, 影响氨氧化微生物的群落结构(Zhang et al ., 2011 ).在漳江口红树林区, 红树林生境与互花米草生境之间多毛类动物栖息密度、生物量、丰度指数、均匀度指数和多样性指数均有显著差异(陈昕韡等, 2012 ).红树林与互花米草生境大型底栖动物次级生产力也存在差异, 海榄雌(Avicennia marina )生境大型底栖动物的年次级生产力最高, 而互花米草生境大型底栖动物的P/B值(P: 平均次级生产力; B: 平均生物量)最高, 因其优势类群为个体较小的寡毛类(吴辰等, 2013 ).再有, 互花米草相对较高的根密度显著降低了红树林中蟹类在互花米草生境打洞的深度及洞穴的复杂性(Wang et al ., 2014 ). ...
Transcriptome analysis of an invasive weed Mikania micrantha.
1
2012
... 微甘菊入侵后, 通常给当地生态系统带来了毁灭性灾害, 因此对微甘菊的防治需求十分紧迫(Waterhouse, 1994 ).物理和化学防治手段的弊端促使学者寻求生物手段来防治微甘菊入侵.业已证实, 寄生植物野地菟丝子(Cuscuta campestris )能安全有效地防治微甘菊(Lian et al ., 2006 ; Shen et al ., 2011 ), 一些新的微生物或病毒也能用来控制微甘菊(Ellison et al ., 2008 ; Wang et al ., 2008 ).除关注微甘菊对生态系统地上植物群落的危害外, 对生态系统其他结构(如地下生物)和功能的效应的关注也逐步形成(Li et al ., 2006 , 2007 ).微甘菊入侵机制的研究日渐成熟(Zhang et al ., 2004 ; Chen et al ., 2009 ).有关微甘菊入侵的分子手段研究(Huang et al ., 2012 ; Wang et al ., 2012 ), 以及微甘菊对全球气候变化的响应研究(Wang et al ., 2010 )初现端倪.随着微甘菊的持续入侵扩散, 温暖湿润沿海地区的红树林生态系统也难逃其入侵危害(Han et al ., 2006 ). ...
The incidence of Nypa fruticans (Wurmb) and its impact on fisheries production in the Niger Delta mangrove ecosystem
1
2003
... 在植物入侵红树林研究文献中, 无瓣海桑(Sonneratia apetala )因其入侵性存在极大的争议, 相关研究报道最多, 有24篇, 紧随其后的为互花米草(Spartina alterniflora )(17篇)、微甘菊(Mikania micrantha )(14篇)和Rhizophora mangle (11篇).无瓣海桑、互花米草入侵红树林的研究区域为中国华南和东南沿海.微甘菊入侵红树林研究除1篇在美国东海岸外, 其余都在中国华南和东南沿海.Rhizophora mangle 入侵研究则仅限于夏威夷群岛.外来植物Nypa fruticans 由人类活动引入尼日利亚红树林, 繁殖爆发, 对当地渔业生产造成危害(Isebor et al ., 2003 ).外来植物Schinus terebinthifolius 入侵美国佛罗里达州的红树林, 通过化感作用抑制了乡土红树植物幼苗的生长(Donnelly et al ., 2008 ).外来种海桑(Sonneratia caseolaris )在深圳福田红树林保护区引种种植后, 提高了泥滩大型底栖动物的多样性(Leung & Tam, 2013 ).此外, 红树林中还存在许多其他外来植物, 但都仅止于调查记录(Biswas et al ., 2007 ; 周青青等, 2010 ). ...
Biology of mangroves and mangrove ecosystems.
1
2001
... 红树林是指分布在热带、亚热带海岸潮间带, 受海水周期性浸淹的木本植物群落; 上述定义强调红树林分布的非地带性以及与潮汐的关系(Duke, 1993 ; 张乔民等, 1997 ).红树林生态系统生境严苛, 高盐、高温、强风, 以及因受潮汐浸淹而底泥缺氧, 只有少数几种进化出高度适应特征的植物能生存(Kathiresan & Bingham, 2001 ).独特的红树林生态系统, 其生物入侵的表现既与其他生态系统有共同之处, 又有其自身特异性. ...
Exotic weed Mikania micrantha HBK appeared in South China.
0
2000
Integrative invasion science: Model systems, multi-site studies, focused meta-analysis and invasion syndromes.
1
2013
... 纵观生物入侵研究的发展历程, 生物入侵的相关机制与假说向来是入侵生态学研究的难点、热点, 备受争议, 至今没有哪一种机制或假说能解释大部分的生物入侵现象(Ren & Zhang, 2009 ; Lowry et al ., 2013 ).今后的整合研究或许能为研究生物入侵机制与假说提供新的视角(Kueffer et al ., 2013 ).随着人类对自然界改造的加剧, 全球贸易往来过度密切, 全球生物入侵形势复杂多变, 外来种入侵带来的生态环境、社会经济、人类健康及生物安全等问题前所未有地严峻(Bradley et al ., 2012 ).目前生物入侵已成为全球变化的因素, 也是导致全球生物多样性丧失的主要原因, 仅次于生境破坏, 并很可能成为第一因素(Mack et al ., 2000 ).生物入侵将与其他全球变化因素一起左右着地球的未来. ...
Distributions of Indo-Pacific lionfishes Pterois spp. in their native ranges: Implications for the Atlantic invasion.
1
2012
... 狮子鱼原产于印度洋和太平洋海域, 20世纪80年代被引入大西洋与加勒比海域.红树林优越的觅食、繁育和避害条件, 使狮子鱼容易成功定居; 加之, 由于缺乏天敌, 其种群密度迅速增加(Arbeláez & Acero, 2011 ; Kulbicki et al ., 2012 ; Prakash et al ., 2012 ).这种鱼类在红树林的爆发, 明显降低了其他鱼类的种群数量, 严重损害了红树林的生态和经济价值(Barbour et al ., 2010 , 2011 ; Claydon et al ., 2012 ). ...
Influence of plantation of an exotic mangrove species, Sonneratia caseolaris (L.) Engl., on macrobenthic infaunal community in Futian Mangrove National Nature Reserve, China.
1
2013
... 在植物入侵红树林研究文献中, 无瓣海桑(Sonneratia apetala )因其入侵性存在极大的争议, 相关研究报道最多, 有24篇, 紧随其后的为互花米草(Spartina alterniflora )(17篇)、微甘菊(Mikania micrantha )(14篇)和Rhizophora mangle (11篇).无瓣海桑、互花米草入侵红树林的研究区域为中国华南和东南沿海.微甘菊入侵红树林研究除1篇在美国东海岸外, 其余都在中国华南和东南沿海.Rhizophora mangle 入侵研究则仅限于夏威夷群岛.外来植物Nypa fruticans 由人类活动引入尼日利亚红树林, 繁殖爆发, 对当地渔业生产造成危害(Isebor et al ., 2003 ).外来植物Schinus terebinthifolius 入侵美国佛罗里达州的红树林, 通过化感作用抑制了乡土红树植物幼苗的生长(Donnelly et al ., 2008 ).外来种海桑(Sonneratia caseolaris )在深圳福田红树林保护区引种种植后, 提高了泥滩大型底栖动物的多样性(Leung & Tam, 2013 ).此外, 红树林中还存在许多其他外来植物, 但都仅止于调查记录(Biswas et al ., 2007 ; 周青青等, 2010 ). ...
生物入侵生态学: 成就与挑战
2
2002
... 当非土著种进入一个过去不曾分布的地区, 并能存活、繁殖而建立种群, 其种群的进一步扩散已经或将要造成明显的生态环境和经济后果, 这一过程称为生物入侵(biological invasion).生物入侵强调外来入侵种的地理属性及其造成的影响(Elton, 1958 ; 李博和陈家宽, 2002 ). ...
... 迄今, 生物入侵研究已发展成一门活跃的生态学分支学科——入侵生态学.生物入侵研究从入侵现状的调查入手, 体现为入侵种多样性与分布格局、入侵效应的调查与评价(Richardson et al ., 2000 ; 李博和陈家宽, 2002 ).在洞悉生物入侵现状后, 紧随的研究是探索生物入侵发生的原因和机制, 探讨外来种的入侵潜力与入侵性(invasiveness)、繁殖体压力(propagule pressure)、生境或生态系统的可入侵性(invasibility)、入侵种与被侵入系统的相互反馈及诸多假说(Sakai et al ., 2001 ; Theoharides & Dukes, 2007 ; Lowry et al ., 2013 ).最后, 在明确生物入侵发生的原因和机制基础上, 对生物入侵进行防治和管理, 以期减小其危害, 并对未来入侵进行合理的预测(Mack et al ., 2000 ). ...
生物入侵生态学: 成就与挑战
2
2002
... 当非土著种进入一个过去不曾分布的地区, 并能存活、繁殖而建立种群, 其种群的进一步扩散已经或将要造成明显的生态环境和经济后果, 这一过程称为生物入侵(biological invasion).生物入侵强调外来入侵种的地理属性及其造成的影响(Elton, 1958 ; 李博和陈家宽, 2002 ). ...
... 迄今, 生物入侵研究已发展成一门活跃的生态学分支学科——入侵生态学.生物入侵研究从入侵现状的调查入手, 体现为入侵种多样性与分布格局、入侵效应的调查与评价(Richardson et al ., 2000 ; 李博和陈家宽, 2002 ).在洞悉生物入侵现状后, 紧随的研究是探索生物入侵发生的原因和机制, 探讨外来种的入侵潜力与入侵性(invasiveness)、繁殖体压力(propagule pressure)、生境或生态系统的可入侵性(invasibility)、入侵种与被侵入系统的相互反馈及诸多假说(Sakai et al ., 2001 ; Theoharides & Dukes, 2007 ; Lowry et al ., 2013 ).最后, 在明确生物入侵发生的原因和机制基础上, 对生物入侵进行防治和管理, 以期减小其危害, 并对未来入侵进行合理的预测(Mack et al ., 2000 ). ...
Spartina alterniflora invasions in the Yangtze River estuary, China: An overview of current status and ecosystem effects.
1
2009
... 互花米草有200多年的入侵扩张历史.不同国家和地区对互花米草的入侵现状、入侵效应、入侵原因与机制, 以及相关防治措施都有过全面而系统的研究(Daehler & Strong, 1996 ; Li et al ., 2009 ).现阶段对互花米草入侵的研究, 一方面是从分子水平揭示互花米草成功入侵的遗传学基础、耐受性以及表型可塑性的分子机理或原因(吴昊和丁建清, 2014 ); 另一方面是应用3S技术, 从景观以上尺度监测互花米草入侵的动态变化(Wan et al ., 2014 ).互花米草入侵不可避免地要受全球变化影响, 二者的相互反馈在全球变化日益强烈的今天愈加受到重视(Emery & Fulweiler, 2014 ).随着互花米草在海岸滩涂的快速扩散, 海岸带上所有的生态系统不可避免地会受到互花米草入侵的威胁.互花米草对潮间带生境的高度适应性, 使之在入侵地扩散, 并成功地入侵红树林生态系统(Zhang et al ., 2012 ). ...
Differences in leaf construction cost between alien and native mangrove species in Futian, Shenzhen, China: Implications for invasiveness of alien species.
1
2011
... 但是也有学者研究发现, 无瓣海桑对其林下乡土红树植物群落会产生不利影响.在无瓣海桑林下, 秋茄树(Kandelia obovata )受到的抑制最小, 可以正常生长; 木榄(Bruguiera gymnorhiza )、海桑、桐花树(Aegiceras corniculatum )受到的抑制次之; 红海兰(Rhizophora stylosa )的保存率最低(李玫等, 2004a ; 梁士楚等, 2005 ; 王旭等, 2008 ; 曾雯珺等, 2008 ; 姚历强, 2010 ).相对于乡土红树植物, 无瓣海桑具有优异的生物学形状——种子幼苗品质好, 抗逆性强(廖宝文等, 2004 ), 生物量大(Chen et al ., 2013b ), 生态位与乡土红树植物有显著重叠(Li et al ., 2011 ), 有一定的扩散能力以及对乡土红树植物的化感作用(李玫等, 2004b ), 以后有可能破坏甚至取代乡土红树植物群落(Ren et al ., 2008 ).因此, 现阶段研究更多建议决策者谨慎考虑无瓣海桑在红树林保护区引种种植. ...
互花米草防治研究进展
2
2007
... 草甘膦(RodeoTM )是目前国内外互花米草控制中唯一得到实际应用的除草剂, 施用后能被植物迅速吸收, 影响植物关键生理过程而导致植株死亡(李富荣等, 2007 ).但是草甘膦等几种除草剂对互花米草的防除效果并不好.米草克(MCK)在使用两个月后对互花米草的防除效果达到100%, 具有很好的推广价值和应用前景(李兆飞, 2009 ).有的学者还自行研制能杀死互花米草的药剂, 该药剂不影响红树植物生长繁殖, 并对水生生物安全不构成影响(杜文琴等, 2006 ). ...
... 目前研究显示, 玉黍螺(Littoraria irrorata )、麦角菌(Ciavieps purpurea )和光蝉(Prokelisia marginata )在互花米草生物控制中得到应用(王卿等, 2006 ).玉黍螺直接取食互花米草的叶片, 抑制其生长(Silliman & Zieman, 2001 ).麦角菌能使互花米草感染麦角病, 在种子内形成菌核, 降低其种子产量, 限制其扩散(Fisher et al ., 2005 ).但麦角菌对禾本科其他一些植物也有影响, 而且其适应性与变异性强, 因此引用麦角菌作为生物控制途径还有待进一步研究(李富荣等, 2007 ).还有一种从华盛顿引入的光蝉, 对入侵威拉帕湾的互花米草具有很强的杀伤力, 对其他植物无明显影响(Grevstad et al ., 2003 ). ...
Mangroves as alien species: The case of Hawaii.
2
1998
... 早期研究认为, 由于红树林生态系统极为苛刻的生境条件, 很少有外来物种能适应环境生存并形成入侵(Lugo, 1998 ).但在全球变化下, 尤其在人类活动的参与下, 生物进入到新生境的概率及入侵成功率大幅提升, 几乎没有什么生境能避免生物入侵(Dukes & Mooney, 1999 ; Occhipinti-Ambrogi & Savini, 2003 ).当前研究显示, 日渐增多的外来种已经出现在红树林生态系统中, 有些已造成明显的入侵危害(Bridgewater & Cresswell, 1999 ; Biswas et al ., 2007 ; Ren et al ., 2014 ).红树林生态系统植物物种少, 群落结构单一, 系统稳定性差, 被入侵破坏后难以恢复.与此同时, 红树林生态系统的组成物种也有可能入侵其他非土著地区, 改变这些地区原有的生态系统(Allen, 1998 ; Fourqurean et al ., 2010 ). ...
... 红树植物同样可以作为外来种入侵非土著区域, 代表性研究为外来红树植物入侵美国夏威夷群岛的研究(Wester, 1981 ; Allen, 1998 ).佛罗里达州的Rhizophora mangle 被人为引种到夏威夷群岛, 种群爆发扩散, 现已覆盖25%的夏威夷南部海岸(Chimner et al ., 2006 ; D’iorio et al ., 2007 ). ...
A skeleton-less sponge of Caribbean mangroves: Invasive or undescribed?
1
2013
... 此外, 一些软体、海绵或腔肠动物(如牡蛎、贻贝、海绵等)由于人为活动传入非土著地区, 暂时还没有大肆爆发形成入侵危害(Holland et al ., 2004 ; Salgado-Barragán & Toledano-Granados, 2006 ; Bishop et al ., 2010 ; Baker et al ., 2012 ; Alvizu et al ., 2013 ); 还有一些入侵非土著海洋的动物是否入侵红树林尚不明确(Ruiz et al ., 2000 ; 陆琴燕等, 2013 ), 有待今后持续关注. ...
互花米草防治研究进展
2
2007
... 草甘膦(RodeoTM )是目前国内外互花米草控制中唯一得到实际应用的除草剂, 施用后能被植物迅速吸收, 影响植物关键生理过程而导致植株死亡(李富荣等, 2007 ).但是草甘膦等几种除草剂对互花米草的防除效果并不好.米草克(MCK)在使用两个月后对互花米草的防除效果达到100%, 具有很好的推广价值和应用前景(李兆飞, 2009 ).有的学者还自行研制能杀死互花米草的药剂, 该药剂不影响红树植物生长繁殖, 并对水生生物安全不构成影响(杜文琴等, 2006 ). ...
... 目前研究显示, 玉黍螺(Littoraria irrorata )、麦角菌(Ciavieps purpurea )和光蝉(Prokelisia marginata )在互花米草生物控制中得到应用(王卿等, 2006 ).玉黍螺直接取食互花米草的叶片, 抑制其生长(Silliman & Zieman, 2001 ).麦角菌能使互花米草感染麦角病, 在种子内形成菌核, 降低其种子产量, 限制其扩散(Fisher et al ., 2005 ).但麦角菌对禾本科其他一些植物也有影响, 而且其适应性与变异性强, 因此引用麦角菌作为生物控制途径还有待进一步研究(李富荣等, 2007 ).还有一种从华盛顿引入的光蝉, 对入侵威拉帕湾的互花米草具有很强的杀伤力, 对其他植物无明显影响(Grevstad et al ., 2003 ). ...
无瓣海桑和互花米草群落土壤化感作用比较
1
2011
... 一些研究认为无瓣海桑通过竞争或化感作用能有效地控制红树林区互花米草的滋生蔓延, 最后取代互花米草(唐国玲等, 2007 ; 李静等, 2011 ).这种植物替代的生物控制方法虽有很好的应用前景, 但考虑到无瓣海桑的引入对土著红树林也存在一定的危害, 须谨慎采用. ...
无瓣海桑和互花米草群落土壤化感作用比较
1
2011
... 一些研究认为无瓣海桑通过竞争或化感作用能有效地控制红树林区互花米草的滋生蔓延, 最后取代互花米草(唐国玲等, 2007 ; 李静等, 2011 ).这种植物替代的生物控制方法虽有很好的应用前景, 但考虑到无瓣海桑的引入对土著红树林也存在一定的危害, 须谨慎采用. ...
Ocurrence of the lionfish Pterois volitans (Linnaeus) in the mangrove of Bahía de Chengue, Colombian Caribbean.
1
2011
... 狮子鱼原产于印度洋和太平洋海域, 20世纪80年代被引入大西洋与加勒比海域.红树林优越的觅食、繁育和避害条件, 使狮子鱼容易成功定居; 加之, 由于缺乏天敌, 其种群密度迅速增加(Arbeláez & Acero, 2011 ; Kulbicki et al ., 2012 ; Prakash et al ., 2012 ).这种鱼类在红树林的爆发, 明显降低了其他鱼类的种群数量, 严重损害了红树林的生态和经济价值(Barbour et al ., 2010 , 2011 ; Claydon et al ., 2012 ). ...
Habitat dominance of a nonindigenous tropical bivalve, Perna viridis (Linnaeus, 1758), in a subtropical estuary in the Gulf of Mexico.
1
2012
... 此外, 一些软体、海绵或腔肠动物(如牡蛎、贻贝、海绵等)由于人为活动传入非土著地区, 暂时还没有大肆爆发形成入侵危害(Holland et al ., 2004 ; Salgado-Barragán & Toledano-Granados, 2006 ; Bishop et al ., 2010 ; Baker et al ., 2012 ; Alvizu et al ., 2013 ); 还有一些入侵非土著海洋的动物是否入侵红树林尚不明确(Ruiz et al ., 2000 ; 陆琴燕等, 2013 ), 有待今后持续关注. ...
无瓣海桑的直接引入对次生桐花树群落的扰动
1
2004a
... 但是也有学者研究发现, 无瓣海桑对其林下乡土红树植物群落会产生不利影响.在无瓣海桑林下, 秋茄树(Kandelia obovata )受到的抑制最小, 可以正常生长; 木榄(Bruguiera gymnorhiza )、海桑、桐花树(Aegiceras corniculatum )受到的抑制次之; 红海兰(Rhizophora stylosa )的保存率最低(李玫等, 2004a ; 梁士楚等, 2005 ; 王旭等, 2008 ; 曾雯珺等, 2008 ; 姚历强, 2010 ).相对于乡土红树植物, 无瓣海桑具有优异的生物学形状——种子幼苗品质好, 抗逆性强(廖宝文等, 2004 ), 生物量大(Chen et al ., 2013b ), 生态位与乡土红树植物有显著重叠(Li et al ., 2011 ), 有一定的扩散能力以及对乡土红树植物的化感作用(李玫等, 2004b ), 以后有可能破坏甚至取代乡土红树植物群落(Ren et al ., 2008 ).因此, 现阶段研究更多建议决策者谨慎考虑无瓣海桑在红树林保护区引种种植. ...
Evaluating the potential efficacy of invasive lionfish (Pterois volitans) removals.
1
2011
... 狮子鱼原产于印度洋和太平洋海域, 20世纪80年代被引入大西洋与加勒比海域.红树林优越的觅食、繁育和避害条件, 使狮子鱼容易成功定居; 加之, 由于缺乏天敌, 其种群密度迅速增加(Arbeláez & Acero, 2011 ; Kulbicki et al ., 2012 ; Prakash et al ., 2012 ).这种鱼类在红树林的爆发, 明显降低了其他鱼类的种群数量, 严重损害了红树林的生态和经济价值(Barbour et al ., 2010 , 2011 ; Claydon et al ., 2012 ). ...
Mangrove use by the invasive lionfish Pterois volitans.
1
2010
... 狮子鱼原产于印度洋和太平洋海域, 20世纪80年代被引入大西洋与加勒比海域.红树林优越的觅食、繁育和避害条件, 使狮子鱼容易成功定居; 加之, 由于缺乏天敌, 其种群密度迅速增加(Arbeláez & Acero, 2011 ; Kulbicki et al ., 2012 ; Prakash et al ., 2012 ).这种鱼类在红树林的爆发, 明显降低了其他鱼类的种群数量, 严重损害了红树林的生态和经济价值(Barbour et al ., 2010 , 2011 ; Claydon et al ., 2012 ). ...
无瓣海桑的直接引入对次生桐花树群落的扰动
1
2004a
... 但是也有学者研究发现, 无瓣海桑对其林下乡土红树植物群落会产生不利影响.在无瓣海桑林下, 秋茄树(Kandelia obovata )受到的抑制最小, 可以正常生长; 木榄(Bruguiera gymnorhiza )、海桑、桐花树(Aegiceras corniculatum )受到的抑制次之; 红海兰(Rhizophora stylosa )的保存率最低(李玫等, 2004a ; 梁士楚等, 2005 ; 王旭等, 2008 ; 曾雯珺等, 2008 ; 姚历强, 2010 ).相对于乡土红树植物, 无瓣海桑具有优异的生物学形状——种子幼苗品质好, 抗逆性强(廖宝文等, 2004 ), 生物量大(Chen et al ., 2013b ), 生态位与乡土红树植物有显著重叠(Li et al ., 2011 ), 有一定的扩散能力以及对乡土红树植物的化感作用(李玫等, 2004b ), 以后有可能破坏甚至取代乡土红树植物群落(Ren et al ., 2008 ).因此, 现阶段研究更多建议决策者谨慎考虑无瓣海桑在红树林保护区引种种植. ...
b). 无瓣海桑对乡土红树植物的化感作用
1
2004
... 但是也有学者研究发现, 无瓣海桑对其林下乡土红树植物群落会产生不利影响.在无瓣海桑林下, 秋茄树(Kandelia obovata )受到的抑制最小, 可以正常生长; 木榄(Bruguiera gymnorhiza )、海桑、桐花树(Aegiceras corniculatum )受到的抑制次之; 红海兰(Rhizophora stylosa )的保存率最低(李玫等, 2004a ; 梁士楚等, 2005 ; 王旭等, 2008 ; 曾雯珺等, 2008 ; 姚历强, 2010 ).相对于乡土红树植物, 无瓣海桑具有优异的生物学形状——种子幼苗品质好, 抗逆性强(廖宝文等, 2004 ), 生物量大(Chen et al ., 2013b ), 生态位与乡土红树植物有显著重叠(Li et al ., 2011 ), 有一定的扩散能力以及对乡土红树植物的化感作用(李玫等, 2004b ), 以后有可能破坏甚至取代乡土红树植物群落(Ren et al ., 2008 ).因此, 现阶段研究更多建议决策者谨慎考虑无瓣海桑在红树林保护区引种种植. ...
Change in wild-oyster assemblages of Port Stephens, NSW, Australia, since commencement of non-native Pacific oyster (Crassostrea gigas) aquaculture.
1
2010
... 此外, 一些软体、海绵或腔肠动物(如牡蛎、贻贝、海绵等)由于人为活动传入非土著地区, 暂时还没有大肆爆发形成入侵危害(Holland et al ., 2004 ; Salgado-Barragán & Toledano-Granados, 2006 ; Bishop et al ., 2010 ; Baker et al ., 2012 ; Alvizu et al ., 2013 ); 还有一些入侵非土著海洋的动物是否入侵红树林尚不明确(Ruiz et al ., 2000 ; 陆琴燕等, 2013 ), 有待今后持续关注. ...
Do invasive plants threaten the Sundarbans mangrove forest of Bangladesh?
2
2007
... 早期研究认为, 由于红树林生态系统极为苛刻的生境条件, 很少有外来物种能适应环境生存并形成入侵(Lugo, 1998 ).但在全球变化下, 尤其在人类活动的参与下, 生物进入到新生境的概率及入侵成功率大幅提升, 几乎没有什么生境能避免生物入侵(Dukes & Mooney, 1999 ; Occhipinti-Ambrogi & Savini, 2003 ).当前研究显示, 日渐增多的外来种已经出现在红树林生态系统中, 有些已造成明显的入侵危害(Bridgewater & Cresswell, 1999 ; Biswas et al ., 2007 ; Ren et al ., 2014 ).红树林生态系统植物物种少, 群落结构单一, 系统稳定性差, 被入侵破坏后难以恢复.与此同时, 红树林生态系统的组成物种也有可能入侵其他非土著地区, 改变这些地区原有的生态系统(Allen, 1998 ; Fourqurean et al ., 2010 ). ...
... 在植物入侵红树林研究文献中, 无瓣海桑(Sonneratia apetala )因其入侵性存在极大的争议, 相关研究报道最多, 有24篇, 紧随其后的为互花米草(Spartina alterniflora )(17篇)、微甘菊(Mikania micrantha )(14篇)和Rhizophora mangle (11篇).无瓣海桑、互花米草入侵红树林的研究区域为中国华南和东南沿海.微甘菊入侵红树林研究除1篇在美国东海岸外, 其余都在中国华南和东南沿海.Rhizophora mangle 入侵研究则仅限于夏威夷群岛.外来植物Nypa fruticans 由人类活动引入尼日利亚红树林, 繁殖爆发, 对当地渔业生产造成危害(Isebor et al ., 2003 ).外来植物Schinus terebinthifolius 入侵美国佛罗里达州的红树林, 通过化感作用抑制了乡土红树植物幼苗的生长(Donnelly et al ., 2008 ).外来种海桑(Sonneratia caseolaris )在深圳福田红树林保护区引种种植后, 提高了泥滩大型底栖动物的多样性(Leung & Tam, 2013 ).此外, 红树林中还存在许多其他外来植物, 但都仅止于调查记录(Biswas et al ., 2007 ; 周青青等, 2010 ). ...
b). 无瓣海桑对乡土红树植物的化感作用
1
2004
... 但是也有学者研究发现, 无瓣海桑对其林下乡土红树植物群落会产生不利影响.在无瓣海桑林下, 秋茄树(Kandelia obovata )受到的抑制最小, 可以正常生长; 木榄(Bruguiera gymnorhiza )、海桑、桐花树(Aegiceras corniculatum )受到的抑制次之; 红海兰(Rhizophora stylosa )的保存率最低(李玫等, 2004a ; 梁士楚等, 2005 ; 王旭等, 2008 ; 曾雯珺等, 2008 ; 姚历强, 2010 ).相对于乡土红树植物, 无瓣海桑具有优异的生物学形状——种子幼苗品质好, 抗逆性强(廖宝文等, 2004 ), 生物量大(Chen et al ., 2013b ), 生态位与乡土红树植物有显著重叠(Li et al ., 2011 ), 有一定的扩散能力以及对乡土红树植物的化感作用(李玫等, 2004b ), 以后有可能破坏甚至取代乡土红树植物群落(Ren et al ., 2008 ).因此, 现阶段研究更多建议决策者谨慎考虑无瓣海桑在红树林保护区引种种植. ...
Relationship between Mikania micrantha invasion and soil microbial biomass, respiration and functional diversity.
1
2007
... 微甘菊入侵后, 通常给当地生态系统带来了毁灭性灾害, 因此对微甘菊的防治需求十分紧迫(Waterhouse, 1994 ).物理和化学防治手段的弊端促使学者寻求生物手段来防治微甘菊入侵.业已证实, 寄生植物野地菟丝子(Cuscuta campestris )能安全有效地防治微甘菊(Lian et al ., 2006 ; Shen et al ., 2011 ), 一些新的微生物或病毒也能用来控制微甘菊(Ellison et al ., 2008 ; Wang et al ., 2008 ).除关注微甘菊对生态系统地上植物群落的危害外, 对生态系统其他结构(如地下生物)和功能的效应的关注也逐步形成(Li et al ., 2006 , 2007 ).微甘菊入侵机制的研究日渐成熟(Zhang et al ., 2004 ; Chen et al ., 2009 ).有关微甘菊入侵的分子手段研究(Huang et al ., 2012 ; Wang et al ., 2012 ), 以及微甘菊对全球气候变化的响应研究(Wang et al ., 2010 )初现端倪.随着微甘菊的持续入侵扩散, 温暖湿润沿海地区的红树林生态系统也难逃其入侵危害(Han et al ., 2006 ). ...
Global change, global trade, and the next wave of plant invasions.
1
2012
... 纵观生物入侵研究的发展历程, 生物入侵的相关机制与假说向来是入侵生态学研究的难点、热点, 备受争议, 至今没有哪一种机制或假说能解释大部分的生物入侵现象(Ren & Zhang, 2009 ; Lowry et al ., 2013 ).今后的整合研究或许能为研究生物入侵机制与假说提供新的视角(Kueffer et al ., 2013 ).随着人类对自然界改造的加剧, 全球贸易往来过度密切, 全球生物入侵形势复杂多变, 外来种入侵带来的生态环境、社会经济、人类健康及生物安全等问题前所未有地严峻(Bradley et al ., 2012 ).目前生物入侵已成为全球变化的因素, 也是导致全球生物多样性丧失的主要原因, 仅次于生境破坏, 并很可能成为第一因素(Mack et al ., 2000 ).生物入侵将与其他全球变化因素一起左右着地球的未来. ...
Biogeography of mangrove and salt marsh vegetation: implications for conservation and management in Australia.
1
1999
... 早期研究认为, 由于红树林生态系统极为苛刻的生境条件, 很少有外来物种能适应环境生存并形成入侵(Lugo, 1998 ).但在全球变化下, 尤其在人类活动的参与下, 生物进入到新生境的概率及入侵成功率大幅提升, 几乎没有什么生境能避免生物入侵(Dukes & Mooney, 1999 ; Occhipinti-Ambrogi & Savini, 2003 ).当前研究显示, 日渐增多的外来种已经出现在红树林生态系统中, 有些已造成明显的入侵危害(Bridgewater & Cresswell, 1999 ; Biswas et al ., 2007 ; Ren et al ., 2014 ).红树林生态系统植物物种少, 群落结构单一, 系统稳定性差, 被入侵破坏后难以恢复.与此同时, 红树林生态系统的组成物种也有可能入侵其他非土著地区, 改变这些地区原有的生态系统(Allen, 1998 ; Fourqurean et al ., 2010 ). ...
Changes in soil microbial community associated with invasion of the exotic weed, Mikania micrantha H. B. K.
1
2006
... 微甘菊入侵后, 通常给当地生态系统带来了毁灭性灾害, 因此对微甘菊的防治需求十分紧迫(Waterhouse, 1994 ).物理和化学防治手段的弊端促使学者寻求生物手段来防治微甘菊入侵.业已证实, 寄生植物野地菟丝子(Cuscuta campestris )能安全有效地防治微甘菊(Lian et al ., 2006 ; Shen et al ., 2011 ), 一些新的微生物或病毒也能用来控制微甘菊(Ellison et al ., 2008 ; Wang et al ., 2008 ).除关注微甘菊对生态系统地上植物群落的危害外, 对生态系统其他结构(如地下生物)和功能的效应的关注也逐步形成(Li et al ., 2006 , 2007 ).微甘菊入侵机制的研究日渐成熟(Zhang et al ., 2004 ; Chen et al ., 2009 ).有关微甘菊入侵的分子手段研究(Huang et al ., 2012 ; Wang et al ., 2012 ), 以及微甘菊对全球气候变化的响应研究(Wang et al ., 2010 )初现端倪.随着微甘菊的持续入侵扩散, 温暖湿润沿海地区的红树林生态系统也难逃其入侵危害(Han et al ., 2006 ). ...
人为干预群落对薇甘菊的影响
1
2003
... 针对微甘菊, 通常是人工清除, 先割除地上部分, 再挖出地下根与茎, 放在烈日下曝晒, 但其群落很快就又恢复如初, 毫无效果.而在拔除微甘菊后, 种植人工干预的乔灌草群落, 能在一定程度上控制微甘菊的危害(李小川等, 2003 ). ...
Effects of the invasive plant Mikania micrantha H. B. K. on soil nitrogen availability through allelopathy in South China.
1
2009
... 微甘菊入侵后, 通常给当地生态系统带来了毁灭性灾害, 因此对微甘菊的防治需求十分紧迫(Waterhouse, 1994 ).物理和化学防治手段的弊端促使学者寻求生物手段来防治微甘菊入侵.业已证实, 寄生植物野地菟丝子(Cuscuta campestris )能安全有效地防治微甘菊(Lian et al ., 2006 ; Shen et al ., 2011 ), 一些新的微生物或病毒也能用来控制微甘菊(Ellison et al ., 2008 ; Wang et al ., 2008 ).除关注微甘菊对生态系统地上植物群落的危害外, 对生态系统其他结构(如地下生物)和功能的效应的关注也逐步形成(Li et al ., 2006 , 2007 ).微甘菊入侵机制的研究日渐成熟(Zhang et al ., 2004 ; Chen et al ., 2009 ).有关微甘菊入侵的分子手段研究(Huang et al ., 2012 ; Wang et al ., 2012 ), 以及微甘菊对全球气候变化的响应研究(Wang et al ., 2010 )初现端倪.随着微甘菊的持续入侵扩散, 温暖湿润沿海地区的红树林生态系统也难逃其入侵危害(Han et al ., 2006 ). ...
淡咸水轮换浇灌抑制互花米草的克隆生长和繁殖
1
2007
... 持续的淡咸水轮换浇灌, 互花米草的生物量积累、无性与有性繁殖能力都会受到抑制(陈琳等, 2007 ).而且选择关键季节, 刈割互花米草地上部分, 同时配合一定水位的持续淹水, 更能有效地发挥治理互花米草的效果(袁琳等, 2008 ).有些物理控制措施, 如遮阴或冬季火烧, 对互花米草影响不大, 反而会影响乡土植物生长, 促进互花米草的入侵, 应当避免 (陈中义等, 2005 ; 王智晨等, 2006 ). ...
人为干预群落对薇甘菊的影响
1
2003
... 针对微甘菊, 通常是人工清除, 先割除地上部分, 再挖出地下根与茎, 放在烈日下曝晒, 但其群落很快就又恢复如初, 毫无效果.而在拔除微甘菊后, 种植人工干预的乔灌草群落, 能在一定程度上控制微甘菊的危害(李小川等, 2003 ). ...
红树植物无瓣海桑引种的初步研究
1
1998
... 无瓣海桑是海桑科海桑属乔木, 天然分布于印度、孟加拉国、马来西亚等, 因其速生性而被广泛用于海岸滩涂造林, 1985年引入中国海南岛, 现在常见于中国大部分红树林区(李云等, 1998 ). ...
淡咸水轮换浇灌抑制互花米草的克隆生长和繁殖
1
2007
... 持续的淡咸水轮换浇灌, 互花米草的生物量积累、无性与有性繁殖能力都会受到抑制(陈琳等, 2007 ).而且选择关键季节, 刈割互花米草地上部分, 同时配合一定水位的持续淹水, 更能有效地发挥治理互花米草的效果(袁琳等, 2008 ).有些物理控制措施, 如遮阴或冬季火烧, 对互花米草影响不大, 反而会影响乡土植物生长, 促进互花米草的入侵, 应当避免 (陈中义等, 2005 ; 王智晨等, 2006 ). ...
a). Competitive control of an exotic mangrove species: Restoration of native mangrove forests by altering light availability.
1
2013
... 无瓣海桑繁殖和更新存在一定的局限性, 其扩散能力不是很强, 人为适当的砍伐或拔除就能控制其种群数量.研究还发现对无瓣海桑做低光照处理, 能有效地抑制其生长(Chen et al ., 2013a ). ...
红树植物无瓣海桑引种的初步研究
1
1998
... 无瓣海桑是海桑科海桑属乔木, 天然分布于印度、孟加拉国、马来西亚等, 因其速生性而被广泛用于海岸滩涂造林, 1985年引入中国海南岛, 现在常见于中国大部分红树林区(李云等, 1998 ). ...
外来入侵植物互花米草控制的初步研究
1
2009
... 草甘膦(RodeoTM )是目前国内外互花米草控制中唯一得到实际应用的除草剂, 施用后能被植物迅速吸收, 影响植物关键生理过程而导致植株死亡(李富荣等, 2007 ).但是草甘膦等几种除草剂对互花米草的防除效果并不好.米草克(MCK)在使用两个月后对互花米草的防除效果达到100%, 具有很好的推广价值和应用前景(李兆飞, 2009 ).有的学者还自行研制能杀死互花米草的药剂, 该药剂不影响红树植物生长繁殖, 并对水生生物安全不构成影响(杜文琴等, 2006 ). ...
b). Significant niche overlap between native and exotic Sonneratia mangrove species along a continuum of varying inundation periods.
1
2013
... 但是也有学者研究发现, 无瓣海桑对其林下乡土红树植物群落会产生不利影响.在无瓣海桑林下, 秋茄树(Kandelia obovata )受到的抑制最小, 可以正常生长; 木榄(Bruguiera gymnorhiza )、海桑、桐花树(Aegiceras corniculatum )受到的抑制次之; 红海兰(Rhizophora stylosa )的保存率最低(李玫等, 2004a ; 梁士楚等, 2005 ; 王旭等, 2008 ; 曾雯珺等, 2008 ; 姚历强, 2010 ).相对于乡土红树植物, 无瓣海桑具有优异的生物学形状——种子幼苗品质好, 抗逆性强(廖宝文等, 2004 ), 生物量大(Chen et al ., 2013b ), 生态位与乡土红树植物有显著重叠(Li et al ., 2011 ), 有一定的扩散能力以及对乡土红树植物的化感作用(李玫等, 2004b ), 以后有可能破坏甚至取代乡土红树植物群落(Ren et al ., 2008 ).因此, 现阶段研究更多建议决策者谨慎考虑无瓣海桑在红树林保护区引种种植. ...
福建漳江口红树林和盐沼湿地的多毛类动物群落
1
2012
... 互花米草入侵还会改变红树林的生物多样性及其行为模式.在九龙江口红树林区, 互花米草入侵区域底栖动物的栖息密度、种类多样性、丰度比红树林区高, 比涂滩要低(洪荣标等, 2005 ).互花米草的入侵还改变着氨氧化古菌与细菌的丰度, 影响氨氧化微生物的群落结构(Zhang et al ., 2011 ).在漳江口红树林区, 红树林生境与互花米草生境之间多毛类动物栖息密度、生物量、丰度指数、均匀度指数和多样性指数均有显著差异(陈昕韡等, 2012 ).红树林与互花米草生境大型底栖动物次级生产力也存在差异, 海榄雌(Avicennia marina )生境大型底栖动物的年次级生产力最高, 而互花米草生境大型底栖动物的P/B值(P: 平均次级生产力; B: 平均生物量)最高, 因其优势类群为个体较小的寡毛类(吴辰等, 2013 ).再有, 互花米草相对较高的根密度显著降低了红树林中蟹类在互花米草生境打洞的深度及洞穴的复杂性(Wang et al ., 2014 ). ...
外来入侵植物互花米草控制的初步研究
1
2009
... 草甘膦(RodeoTM )是目前国内外互花米草控制中唯一得到实际应用的除草剂, 施用后能被植物迅速吸收, 影响植物关键生理过程而导致植株死亡(李富荣等, 2007 ).但是草甘膦等几种除草剂对互花米草的防除效果并不好.米草克(MCK)在使用两个月后对互花米草的防除效果达到100%, 具有很好的推广价值和应用前景(李兆飞, 2009 ).有的学者还自行研制能杀死互花米草的药剂, 该药剂不影响红树植物生长繁殖, 并对水生生物安全不构成影响(杜文琴等, 2006 ). ...
Influence of obligate parasite Cuscuta campestris on the community of its host Mikania micrantha.
1
2006
... 微甘菊入侵后, 通常给当地生态系统带来了毁灭性灾害, 因此对微甘菊的防治需求十分紧迫(Waterhouse, 1994 ).物理和化学防治手段的弊端促使学者寻求生物手段来防治微甘菊入侵.业已证实, 寄生植物野地菟丝子(Cuscuta campestris )能安全有效地防治微甘菊(Lian et al ., 2006 ; Shen et al ., 2011 ), 一些新的微生物或病毒也能用来控制微甘菊(Ellison et al ., 2008 ; Wang et al ., 2008 ).除关注微甘菊对生态系统地上植物群落的危害外, 对生态系统其他结构(如地下生物)和功能的效应的关注也逐步形成(Li et al ., 2006 , 2007 ).微甘菊入侵机制的研究日渐成熟(Zhang et al ., 2004 ; Chen et al ., 2009 ).有关微甘菊入侵的分子手段研究(Huang et al ., 2012 ; Wang et al ., 2012 ), 以及微甘菊对全球气候变化的响应研究(Wang et al ., 2010 )初现端倪.随着微甘菊的持续入侵扩散, 温暖湿润沿海地区的红树林生态系统也难逃其入侵危害(Han et al ., 2006 ). ...
福建漳江口红树林和盐沼湿地的多毛类动物群落
1
2012
... 互花米草入侵还会改变红树林的生物多样性及其行为模式.在九龙江口红树林区, 互花米草入侵区域底栖动物的栖息密度、种类多样性、丰度比红树林区高, 比涂滩要低(洪荣标等, 2005 ).互花米草的入侵还改变着氨氧化古菌与细菌的丰度, 影响氨氧化微生物的群落结构(Zhang et al ., 2011 ).在漳江口红树林区, 红树林生境与互花米草生境之间多毛类动物栖息密度、生物量、丰度指数、均匀度指数和多样性指数均有显著差异(陈昕韡等, 2012 ).红树林与互花米草生境大型底栖动物次级生产力也存在差异, 海榄雌(Avicennia marina )生境大型底栖动物的年次级生产力最高, 而互花米草生境大型底栖动物的P/B值(P: 平均次级生产力; B: 平均生物量)最高, 因其优势类群为个体较小的寡毛类(吴辰等, 2013 ).再有, 互花米草相对较高的根密度显著降低了红树林中蟹类在互花米草生境打洞的深度及洞穴的复杂性(Wang et al ., 2014 ). ...
模拟遮荫对互花米草和海三棱草种子萌发及幼苗生长的影响
1
2005
... 持续的淡咸水轮换浇灌, 互花米草的生物量积累、无性与有性繁殖能力都会受到抑制(陈琳等, 2007 ).而且选择关键季节, 刈割互花米草地上部分, 同时配合一定水位的持续淹水, 更能有效地发挥治理互花米草的效果(袁琳等, 2008 ).有些物理控制措施, 如遮阴或冬季火烧, 对互花米草影响不大, 反而会影响乡土植物生长, 促进互花米草的入侵, 应当避免 (陈中义等, 2005 ; 王智晨等, 2006 ). ...
深圳福田海桑+无瓣海桑自然林的空间结构分析
1
2005
... 但是也有学者研究发现, 无瓣海桑对其林下乡土红树植物群落会产生不利影响.在无瓣海桑林下, 秋茄树(Kandelia obovata )受到的抑制最小, 可以正常生长; 木榄(Bruguiera gymnorhiza )、海桑、桐花树(Aegiceras corniculatum )受到的抑制次之; 红海兰(Rhizophora stylosa )的保存率最低(李玫等, 2004a ; 梁士楚等, 2005 ; 王旭等, 2008 ; 曾雯珺等, 2008 ; 姚历强, 2010 ).相对于乡土红树植物, 无瓣海桑具有优异的生物学形状——种子幼苗品质好, 抗逆性强(廖宝文等, 2004 ), 生物量大(Chen et al ., 2013b ), 生态位与乡土红树植物有显著重叠(Li et al ., 2011 ), 有一定的扩散能力以及对乡土红树植物的化感作用(李玫等, 2004b ), 以后有可能破坏甚至取代乡土红树植物群落(Ren et al ., 2008 ).因此, 现阶段研究更多建议决策者谨慎考虑无瓣海桑在红树林保护区引种种植. ...
深圳福田海桑+无瓣海桑自然林的空间结构分析
1
2005
... 但是也有学者研究发现, 无瓣海桑对其林下乡土红树植物群落会产生不利影响.在无瓣海桑林下, 秋茄树(Kandelia obovata )受到的抑制最小, 可以正常生长; 木榄(Bruguiera gymnorhiza )、海桑、桐花树(Aegiceras corniculatum )受到的抑制次之; 红海兰(Rhizophora stylosa )的保存率最低(李玫等, 2004a ; 梁士楚等, 2005 ; 王旭等, 2008 ; 曾雯珺等, 2008 ; 姚历强, 2010 ).相对于乡土红树植物, 无瓣海桑具有优异的生物学形状——种子幼苗品质好, 抗逆性强(廖宝文等, 2004 ), 生物量大(Chen et al ., 2013b ), 生态位与乡土红树植物有显著重叠(Li et al ., 2011 ), 有一定的扩散能力以及对乡土红树植物的化感作用(李玫等, 2004b ), 以后有可能破坏甚至取代乡土红树植物群落(Ren et al ., 2008 ).因此, 现阶段研究更多建议决策者谨慎考虑无瓣海桑在红树林保护区引种种植. ...
模拟遮荫对互花米草和海三棱草种子萌发及幼苗生长的影响
1
2005
... 持续的淡咸水轮换浇灌, 互花米草的生物量积累、无性与有性繁殖能力都会受到抑制(陈琳等, 2007 ).而且选择关键季节, 刈割互花米草地上部分, 同时配合一定水位的持续淹水, 更能有效地发挥治理互花米草的效果(袁琳等, 2008 ).有些物理控制措施, 如遮阴或冬季火烧, 对互花米草影响不大, 反而会影响乡土植物生长, 促进互花米草的入侵, 应当避免 (陈中义等, 2005 ; 王智晨等, 2006 ). ...
Current extent and historical expansion of introduced mangroves on O’ahu, Hawai’i.
1
2006
... 红树植物同样可以作为外来种入侵非土著区域, 代表性研究为外来红树植物入侵美国夏威夷群岛的研究(Wester, 1981 ; Allen, 1998 ).佛罗里达州的Rhizophora mangle 被人为引种到夏威夷群岛, 种群爆发扩散, 现已覆盖25%的夏威夷南部海岸(Chimner et al ., 2006 ; D’iorio et al ., 2007 ). ...
外来红树植物无瓣海桑生物学特性与生态环境适应性分析
1
2004
... 但是也有学者研究发现, 无瓣海桑对其林下乡土红树植物群落会产生不利影响.在无瓣海桑林下, 秋茄树(Kandelia obovata )受到的抑制最小, 可以正常生长; 木榄(Bruguiera gymnorhiza )、海桑、桐花树(Aegiceras corniculatum )受到的抑制次之; 红海兰(Rhizophora stylosa )的保存率最低(李玫等, 2004a ; 梁士楚等, 2005 ; 王旭等, 2008 ; 曾雯珺等, 2008 ; 姚历强, 2010 ).相对于乡土红树植物, 无瓣海桑具有优异的生物学形状——种子幼苗品质好, 抗逆性强(廖宝文等, 2004 ), 生物量大(Chen et al ., 2013b ), 生态位与乡土红树植物有显著重叠(Li et al ., 2011 ), 有一定的扩散能力以及对乡土红树植物的化感作用(李玫等, 2004b ), 以后有可能破坏甚至取代乡土红树植物群落(Ren et al ., 2008 ).因此, 现阶段研究更多建议决策者谨慎考虑无瓣海桑在红树林保护区引种种植. ...
外来红树植物无瓣海桑生物学特性与生态环境适应性分析
1
2004
... 但是也有学者研究发现, 无瓣海桑对其林下乡土红树植物群落会产生不利影响.在无瓣海桑林下, 秋茄树(Kandelia obovata )受到的抑制最小, 可以正常生长; 木榄(Bruguiera gymnorhiza )、海桑、桐花树(Aegiceras corniculatum )受到的抑制次之; 红海兰(Rhizophora stylosa )的保存率最低(李玫等, 2004a ; 梁士楚等, 2005 ; 王旭等, 2008 ; 曾雯珺等, 2008 ; 姚历强, 2010 ).相对于乡土红树植物, 无瓣海桑具有优异的生物学形状——种子幼苗品质好, 抗逆性强(廖宝文等, 2004 ), 生物量大(Chen et al ., 2013b ), 生态位与乡土红树植物有显著重叠(Li et al ., 2011 ), 有一定的扩散能力以及对乡土红树植物的化感作用(李玫等, 2004b ), 以后有可能破坏甚至取代乡土红树植物群落(Ren et al ., 2008 ).因此, 现阶段研究更多建议决策者谨慎考虑无瓣海桑在红树林保护区引种种植. ...
Progression of invasive lionfish in seagrass, mangrove and reef habitats.
1
2012
... 狮子鱼原产于印度洋和太平洋海域, 20世纪80年代被引入大西洋与加勒比海域.红树林优越的觅食、繁育和避害条件, 使狮子鱼容易成功定居; 加之, 由于缺乏天敌, 其种群密度迅速增加(Arbeláez & Acero, 2011 ; Kulbicki et al ., 2012 ; Prakash et al ., 2012 ).这种鱼类在红树林的爆发, 明显降低了其他鱼类的种群数量, 严重损害了红树林的生态和经济价值(Barbour et al ., 2010 , 2011 ; Claydon et al ., 2012 ). ...
Stand structure and productivity of the introduced Rhizophora mangle in Hawaii.
1
1999
... 外来红树植物Rhizophora mangle 入侵夏威夷海岛无植被裸滩, 显著改变了这里原有的生态系统结构与功能.红树群落高的生产力大幅增加了整个区域的生物量(Cox & Allen, 1999 ; Fry & Cormier, 2011 ; D’iorio, 2003 ).Rhizophora mangle 对地下部分的凋落物输入, 改善了沉积物的理化特征, 并改变了原有的食物链(网)结构, 丰富了底栖生物群落(Demopoulos, 2004 ; Demopoulos et al ., 2007 ; Siple & Donahue, 2013 ).底栖生物群落的丰富多样, 为进一步的外来生物入侵提供了优良的条件, 形成一个外来生物入侵循环(Demopoulos & Smith, 2010 ).外来红树植物入侵也对底栖生态系统功能产生了深远的影响, 夏威夷群岛海岸的沉积物代谢和碳再矿化动态在移除入侵种Rhizophora mangle 数年后仍没有摆脱其影响(Sweetman et al ., 2010 ). ...
Biological invasions: A field synopsis, systematic review, and database of the literature.
2
2013
... 迄今, 生物入侵研究已发展成一门活跃的生态学分支学科——入侵生态学.生物入侵研究从入侵现状的调查入手, 体现为入侵种多样性与分布格局、入侵效应的调查与评价(Richardson et al ., 2000 ; 李博和陈家宽, 2002 ).在洞悉生物入侵现状后, 紧随的研究是探索生物入侵发生的原因和机制, 探讨外来种的入侵潜力与入侵性(invasiveness)、繁殖体压力(propagule pressure)、生境或生态系统的可入侵性(invasibility)、入侵种与被侵入系统的相互反馈及诸多假说(Sakai et al ., 2001 ; Theoharides & Dukes, 2007 ; Lowry et al ., 2013 ).最后, 在明确生物入侵发生的原因和机制基础上, 对生物入侵进行防治和管理, 以期减小其危害, 并对未来入侵进行合理的预测(Mack et al ., 2000 ). ...
... 纵观生物入侵研究的发展历程, 生物入侵的相关机制与假说向来是入侵生态学研究的难点、热点, 备受争议, 至今没有哪一种机制或假说能解释大部分的生物入侵现象(Ren & Zhang, 2009 ; Lowry et al ., 2013 ).今后的整合研究或许能为研究生物入侵机制与假说提供新的视角(Kueffer et al ., 2013 ).随着人类对自然界改造的加剧, 全球贸易往来过度密切, 全球生物入侵形势复杂多变, 外来种入侵带来的生态环境、社会经济、人类健康及生物安全等问题前所未有地严峻(Bradley et al ., 2012 ).目前生物入侵已成为全球变化的因素, 也是导致全球生物多样性丧失的主要原因, 仅次于生境破坏, 并很可能成为第一因素(Mack et al ., 2000 ).生物入侵将与其他全球变化因素一起左右着地球的未来. ...
海洋外来物种入侵对南海生态系统的影响及防控对策
2
2013
... 此外, 一些软体、海绵或腔肠动物(如牡蛎、贻贝、海绵等)由于人为活动传入非土著地区, 暂时还没有大肆爆发形成入侵危害(Holland et al ., 2004 ; Salgado-Barragán & Toledano-Granados, 2006 ; Bishop et al ., 2010 ; Baker et al ., 2012 ; Alvizu et al ., 2013 ); 还有一些入侵非土著海洋的动物是否入侵红树林尚不明确(Ruiz et al ., 2000 ; 陆琴燕等, 2013 ), 有待今后持续关注. ...
... 微生物入侵红树林的研究寥寥无几.微生物入侵多半是依附在入侵的动植物上, 被动植物带入新生境.与小叶海金沙共生的丛枝菌根真菌(AMF), 在植物入侵红树林时被引入(Soti et al ., 2014 ).一些入侵非土著海洋的微生物是否入侵红树林, 目前仍不明确(Ruiz et al ., 2000 ; 陆琴燕等, 2013 ), 有待关注.而附属于红树植物木材上的一些真菌种类, 未经严格的检疫检验, 通过商业贸易交易到非土著地区, 成为极具危险的潜在入侵病原体(Vannini et al ., 2012 ).研究红树林植物或动物入侵时, 关注与它们共生或寄生的微生物, 是研究微生物入侵与防治的切实可行的途径. ...
Status, prediction and prevention of introduced cordgrass Spartina spp. invasions in Pacific estuaries, USA.
1
1996
... 互花米草有200多年的入侵扩张历史.不同国家和地区对互花米草的入侵现状、入侵效应、入侵原因与机制, 以及相关防治措施都有过全面而系统的研究(Daehler & Strong, 1996 ; Li et al ., 2009 ).现阶段对互花米草入侵的研究, 一方面是从分子水平揭示互花米草成功入侵的遗传学基础、耐受性以及表型可塑性的分子机理或原因(吴昊和丁建清, 2014 ); 另一方面是应用3S技术, 从景观以上尺度监测互花米草入侵的动态变化(Wan et al ., 2014 ).互花米草入侵不可避免地要受全球变化影响, 二者的相互反馈在全球变化日益强烈的今天愈加受到重视(Emery & Fulweiler, 2014 ).随着互花米草在海岸滩涂的快速扩散, 海岸带上所有的生态系统不可避免地会受到互花米草入侵的威胁.互花米草对潮间带生境的高度适应性, 使之在入侵地扩散, 并成功地入侵红树林生态系统(Zhang et al ., 2012 ). ...
Aliens in Paradise: A Comparative Assessment of Introduced and Native Mangrove Benthic Community Composition, Food-web Structure, and Litter-fall Production
1
2004
... 外来红树植物Rhizophora mangle 入侵夏威夷海岛无植被裸滩, 显著改变了这里原有的生态系统结构与功能.红树群落高的生产力大幅增加了整个区域的生物量(Cox & Allen, 1999 ; Fry & Cormier, 2011 ; D’iorio, 2003 ).Rhizophora mangle 对地下部分的凋落物输入, 改善了沉积物的理化特征, 并改变了原有的食物链(网)结构, 丰富了底栖生物群落(Demopoulos, 2004 ; Demopoulos et al ., 2007 ; Siple & Donahue, 2013 ).底栖生物群落的丰富多样, 为进一步的外来生物入侵提供了优良的条件, 形成一个外来生物入侵循环(Demopoulos & Smith, 2010 ).外来红树植物入侵也对底栖生态系统功能产生了深远的影响, 夏威夷群岛海岸的沉积物代谢和碳再矿化动态在移除入侵种Rhizophora mangle 数年后仍没有摆脱其影响(Sweetman et al ., 2010 ). ...
海洋外来物种入侵对南海生态系统的影响及防控对策
2
2013
... 此外, 一些软体、海绵或腔肠动物(如牡蛎、贻贝、海绵等)由于人为活动传入非土著地区, 暂时还没有大肆爆发形成入侵危害(Holland et al ., 2004 ; Salgado-Barragán & Toledano-Granados, 2006 ; Bishop et al ., 2010 ; Baker et al ., 2012 ; Alvizu et al ., 2013 ); 还有一些入侵非土著海洋的动物是否入侵红树林尚不明确(Ruiz et al ., 2000 ; 陆琴燕等, 2013 ), 有待今后持续关注. ...
... 微生物入侵红树林的研究寥寥无几.微生物入侵多半是依附在入侵的动植物上, 被动植物带入新生境.与小叶海金沙共生的丛枝菌根真菌(AMF), 在植物入侵红树林时被引入(Soti et al ., 2014 ).一些入侵非土著海洋的微生物是否入侵红树林, 目前仍不明确(Ruiz et al ., 2000 ; 陆琴燕等, 2013 ), 有待关注.而附属于红树植物木材上的一些真菌种类, 未经严格的检疫检验, 通过商业贸易交易到非土著地区, 成为极具危险的潜在入侵病原体(Vannini et al ., 2012 ).研究红树林植物或动物入侵时, 关注与它们共生或寄生的微生物, 是研究微生物入侵与防治的切实可行的途径. ...
Changes in carbon pool and stand structure of a native subtropical mangrove forest after inter-planting with exotic species Sonneratia apetala.
1
2014
... 无瓣海桑在引入地多用作沿海涂滩造林与恢复树种, 具有明显的改良土壤效应, 可增加土壤有机质和N、P、K含量, 有利于后续乡土红树植物定植(韩维栋等, 2003 ).在乡土红树林群落中混种无瓣海桑, 虽然整个林分生物量或土壤碳库没有显著增加, 但无瓣海桑相对较高的凋落物量和低C/N比, 有利于分解者利用(Lu et al ., 2014 ).无瓣海桑的引入能提高群落结构的复杂性与生物多样性.在深圳福田红树林保护区, 混种无瓣海桑+海桑人工林, 相对于天然红树林, 鸟类群落生物多样性得到提高(王勇军和昝启杰, 2001 ).鉴于上述研究结果, 一些学者认为无瓣海桑并没有表现出入侵危害(Zan et al ., 2003 ; 朱宏伟, 2012 ; Xin et al ., 2013 ). ...
Food web structure in exotic and native mangroves: A Hawaii-Puerto Rico comparison.
1
2007
... 外来红树植物Rhizophora mangle 入侵夏威夷海岛无植被裸滩, 显著改变了这里原有的生态系统结构与功能.红树群落高的生产力大幅增加了整个区域的生物量(Cox & Allen, 1999 ; Fry & Cormier, 2011 ; D’iorio, 2003 ).Rhizophora mangle 对地下部分的凋落物输入, 改善了沉积物的理化特征, 并改变了原有的食物链(网)结构, 丰富了底栖生物群落(Demopoulos, 2004 ; Demopoulos et al ., 2007 ; Siple & Donahue, 2013 ).底栖生物群落的丰富多样, 为进一步的外来生物入侵提供了优良的条件, 形成一个外来生物入侵循环(Demopoulos & Smith, 2010 ).外来红树植物入侵也对底栖生态系统功能产生了深远的影响, 夏威夷群岛海岸的沉积物代谢和碳再矿化动态在移除入侵种Rhizophora mangle 数年后仍没有摆脱其影响(Sweetman et al ., 2010 ). ...
Invasive mangroves alter macrofaunal community structure and facilitate opportunistic exotics.
1
2010
... 外来红树植物Rhizophora mangle 入侵夏威夷海岛无植被裸滩, 显著改变了这里原有的生态系统结构与功能.红树群落高的生产力大幅增加了整个区域的生物量(Cox & Allen, 1999 ; Fry & Cormier, 2011 ; D’iorio, 2003 ).Rhizophora mangle 对地下部分的凋落物输入, 改善了沉积物的理化特征, 并改变了原有的食物链(网)结构, 丰富了底栖生物群落(Demopoulos, 2004 ; Demopoulos et al ., 2007 ; Siple & Donahue, 2013 ).底栖生物群落的丰富多样, 为进一步的外来生物入侵提供了优良的条件, 形成一个外来生物入侵循环(Demopoulos & Smith, 2010 ).外来红树植物入侵也对底栖生态系统功能产生了深远的影响, 夏威夷群岛海岸的沉积物代谢和碳再矿化动态在移除入侵种Rhizophora mangle 数年后仍没有摆脱其影响(Sweetman et al ., 2010 ). ...
Mangrove forests: A tough system to invade but an easy one to rehabilitate.
1
1998
... 早期研究认为, 由于红树林生态系统极为苛刻的生境条件, 很少有外来物种能适应环境生存并形成入侵(Lugo, 1998 ).但在全球变化下, 尤其在人类活动的参与下, 生物进入到新生境的概率及入侵成功率大幅提升, 几乎没有什么生境能避免生物入侵(Dukes & Mooney, 1999 ; Occhipinti-Ambrogi & Savini, 2003 ).当前研究显示, 日渐增多的外来种已经出现在红树林生态系统中, 有些已造成明显的入侵危害(Bridgewater & Cresswell, 1999 ; Biswas et al ., 2007 ; Ren et al ., 2014 ).红树林生态系统植物物种少, 群落结构单一, 系统稳定性差, 被入侵破坏后难以恢复.与此同时, 红树林生态系统的组成物种也有可能入侵其他非土著地区, 改变这些地区原有的生态系统(Allen, 1998 ; Fourqurean et al ., 2010 ). ...
Biotic invasions: causes, epidemiology, global consequences, and control.
8
2000
... 迄今, 生物入侵研究已发展成一门活跃的生态学分支学科——入侵生态学.生物入侵研究从入侵现状的调查入手, 体现为入侵种多样性与分布格局、入侵效应的调查与评价(Richardson et al ., 2000 ; 李博和陈家宽, 2002 ).在洞悉生物入侵现状后, 紧随的研究是探索生物入侵发生的原因和机制, 探讨外来种的入侵潜力与入侵性(invasiveness)、繁殖体压力(propagule pressure)、生境或生态系统的可入侵性(invasibility)、入侵种与被侵入系统的相互反馈及诸多假说(Sakai et al ., 2001 ; Theoharides & Dukes, 2007 ; Lowry et al ., 2013 ).最后, 在明确生物入侵发生的原因和机制基础上, 对生物入侵进行防治和管理, 以期减小其危害, 并对未来入侵进行合理的预测(Mack et al ., 2000 ). ...
... 纵观生物入侵研究的发展历程, 生物入侵的相关机制与假说向来是入侵生态学研究的难点、热点, 备受争议, 至今没有哪一种机制或假说能解释大部分的生物入侵现象(Ren & Zhang, 2009 ; Lowry et al ., 2013 ).今后的整合研究或许能为研究生物入侵机制与假说提供新的视角(Kueffer et al ., 2013 ).随着人类对自然界改造的加剧, 全球贸易往来过度密切, 全球生物入侵形势复杂多变, 外来种入侵带来的生态环境、社会经济、人类健康及生物安全等问题前所未有地严峻(Bradley et al ., 2012 ).目前生物入侵已成为全球变化的因素, 也是导致全球生物多样性丧失的主要原因, 仅次于生境破坏, 并很可能成为第一因素(Mack et al ., 2000 ).生物入侵将与其他全球变化因素一起左右着地球的未来. ...
... 针对生物入侵的不同阶段, 应采取不同的防治手段.在外来种还没有传入新生境前, 主要是防御.对最初传入阶段的防御是代价最小且最有效的防治生物入侵手段(Puth & Post, 2005 ).强化检疫检验, 慎重引种, 以及引种后规范管理, 能在很大程度上减小外来物种入侵的可能性(Mack et al ., 2000 ).若防御失败, 外来物种成功传入新生境, 对外来物种的防治目标转变为根除外来种, 通常很难彻底根除.最后, 外来种成功入侵并造成严重的生态环境或经济后果, 防治目的变为控制入侵种的危害在一个可接受水平(Mack et al ., 2000 ; Sakai et al ., 2001 ; Theoharides & Dukes, 2007 ). ...
... ).若防御失败, 外来物种成功传入新生境, 对外来物种的防治目标转变为根除外来种, 通常很难彻底根除.最后, 外来种成功入侵并造成严重的生态环境或经济后果, 防治目的变为控制入侵种的危害在一个可接受水平(Mack et al ., 2000 ; Sakai et al ., 2001 ; Theoharides & Dukes, 2007 ). ...
... 常见的控制外来生物入侵爆发方法主要有物理、化学和生物三大类(Waterhouse, 1994 ; Mack et al ., 2000 ).针对红树林生态系统现有的主要成功 入侵且危害严重的入侵种, 我们总结了以下控制方法. ...
... 化学控制是利用化学药剂对入侵种进行灭杀, 以达到控制甚至消灭入侵种的目的.相对于物理控制, 化学控制效果更明显, 而且节省人力物力, 是控制外来有害物种的主要方式(Mack et al ., 2000 ).但是, 化学药剂缺乏目标特异性, 在毒杀目标物种的同时, 也毒害了非目标物种, 使用前必须掌握所用化学药剂的安全性(昝启杰等, 2007 ). ...
... 生物防治是以入侵生物与天敌的生态学原理, 依据生物之间相互依存与制约关系, 利用一种或多种生物控制另一种生物种群的消长.具体是指从入侵种原产地引进昆虫、真菌、病原生物或捕食者等天敌来抑制入侵种的生长和繁殖, 从而遏制其种群的爆发.生物控制很好地避免了物理和化学控制的弊端, 是未来生物入侵控制的主要手段(Mack et al ., 2000 ). ...
... 生物控制也不是毫无风险, 我们无法确保引进的天敌不会作用于非目标生物而成为新的入侵种.利用天敌对入侵种进行防治, 需要了解二者的生物学、生态学特性, 协调和处理好生物与生物及环境之间的关系.然而, 天敌的作用极其复杂, 生物控制中对引进天敌的效果与后果始终存在争议.因此, 对于大多数入侵生物的生物控制仍处于实验阶段, 没有得到大规模的应用(Mack et al ., 2000 ). ...
1
2003
... 外来红树植物Rhizophora mangle 入侵夏威夷海岛无植被裸滩, 显著改变了这里原有的生态系统结构与功能.红树群落高的生产力大幅增加了整个区域的生物量(Cox & Allen, 1999 ; Fry & Cormier, 2011 ; D’iorio, 2003 ).Rhizophora mangle 对地下部分的凋落物输入, 改善了沉积物的理化特征, 并改变了原有的食物链(网)结构, 丰富了底栖生物群落(Demopoulos, 2004 ; Demopoulos et al ., 2007 ; Siple & Donahue, 2013 ).底栖生物群落的丰富多样, 为进一步的外来生物入侵提供了优良的条件, 形成一个外来生物入侵循环(Demopoulos & Smith, 2010 ).外来红树植物入侵也对底栖生态系统功能产生了深远的影响, 夏威夷群岛海岸的沉积物代谢和碳再矿化动态在移除入侵种Rhizophora mangle 数年后仍没有摆脱其影响(Sweetman et al ., 2010 ). ...
Optimizing remote sensing and GIS tools for mapping and managing the distribution of an invasive mangrove (Rhizophora mangle) on South Molokai, Hawaii.
1
2007
... 红树植物同样可以作为外来种入侵非土著区域, 代表性研究为外来红树植物入侵美国夏威夷群岛的研究(Wester, 1981 ; Allen, 1998 ).佛罗里达州的Rhizophora mangle 被人为引种到夏威夷群岛, 种群爆发扩散, 现已覆盖25%的夏威夷南部海岸(Chimner et al ., 2006 ; D’iorio et al ., 2007 ). ...
Distribution and ecological range of the alien plant species Mikania micrantha Kunth (Asteraceae) in Taiwan.
1
2008
... 现有的微甘菊入侵研究主要集中于陆地森林生态系统(Zhang et al ., 2004 ), 红树林同类研究比较少, 而且都集中于中国华南沿海红树林(Ren et al ., 2014 ).微甘菊进入红树林后, 迅速生长, 在红树植物上形成一层茂密的覆盖层, 强烈争夺资源(尤其是光照)及通过化感作用抑制或杀死红树植物(Maja et al ., 2008 ).张阳和张华(2008)研究过珠海淇澳岛红树林中微甘菊的分布模式与危害, 强调自然条件和人为因素在影响红树林中微甘菊的分布上起着重要作用; 推测当地微甘菊爆发主要是由于人类不适当的开发活动破坏了当地红树林. ...
薇甘菊入侵对深圳湾红树林生态系统碳储量的影响
1
2011
... 微甘菊在中国华南沿海的另一个重灾区是深圳湾(Ren et al ., 2014 ), 拦海造田使海水退去, 红树林底泥裸露, 微甘菊种子得以萌发生长, 严重危害红树林(于晓梅和杨逢建, 2011 ).微甘菊入侵深圳湾红树林后, 一方面导致红树植物枯萎, 减少了植被生物量; 另一方面增加了凋落物量, 促进了土壤微生物活动, 加速了土壤有机质分解释放, 降低了土壤碳储量, 削弱了红树林碳汇(毛子龙等, 2011 ).微甘菊的入侵, 还会影响人工恢复红树林的季节动态, 降低红树植物的成活率, 妨害红树林生态恢复(苗春玲等, 2012 ). ...
薇甘菊入侵对深圳湾红树林生态系统碳储量的影响
1
2011
... 微甘菊在中国华南沿海的另一个重灾区是深圳湾(Ren et al ., 2014 ), 拦海造田使海水退去, 红树林底泥裸露, 微甘菊种子得以萌发生长, 严重危害红树林(于晓梅和杨逢建, 2011 ).微甘菊入侵深圳湾红树林后, 一方面导致红树植物枯萎, 减少了植被生物量; 另一方面增加了凋落物量, 促进了土壤微生物活动, 加速了土壤有机质分解释放, 降低了土壤碳储量, 削弱了红树林碳汇(毛子龙等, 2011 ).微甘菊的入侵, 还会影响人工恢复红树林的季节动态, 降低红树植物的成活率, 妨害红树林生态恢复(苗春玲等, 2012 ). ...
珠海鹤洲北其他种入侵人工恢复红树林群落的季节动态
1
2012
... 微甘菊在中国华南沿海的另一个重灾区是深圳湾(Ren et al ., 2014 ), 拦海造田使海水退去, 红树林底泥裸露, 微甘菊种子得以萌发生长, 严重危害红树林(于晓梅和杨逢建, 2011 ).微甘菊入侵深圳湾红树林后, 一方面导致红树植物枯萎, 减少了植被生物量; 另一方面增加了凋落物量, 促进了土壤微生物活动, 加速了土壤有机质分解释放, 降低了土壤碳储量, 削弱了红树林碳汇(毛子龙等, 2011 ).微甘菊的入侵, 还会影响人工恢复红树林的季节动态, 降低红树植物的成活率, 妨害红树林生态恢复(苗春玲等, 2012 ). ...
Allelopathic effects of fruits of the Brazilian pepper Schinus terebinthifolius on growth, leaf production and biomass of seedlings of the red mangrove Rhizophora mangle and the black mangrove Avicennia germinans.
1
2008
... 在植物入侵红树林研究文献中, 无瓣海桑(Sonneratia apetala )因其入侵性存在极大的争议, 相关研究报道最多, 有24篇, 紧随其后的为互花米草(Spartina alterniflora )(17篇)、微甘菊(Mikania micrantha )(14篇)和Rhizophora mangle (11篇).无瓣海桑、互花米草入侵红树林的研究区域为中国华南和东南沿海.微甘菊入侵红树林研究除1篇在美国东海岸外, 其余都在中国华南和东南沿海.Rhizophora mangle 入侵研究则仅限于夏威夷群岛.外来植物Nypa fruticans 由人类活动引入尼日利亚红树林, 繁殖爆发, 对当地渔业生产造成危害(Isebor et al ., 2003 ).外来植物Schinus terebinthifolius 入侵美国佛罗里达州的红树林, 通过化感作用抑制了乡土红树植物幼苗的生长(Donnelly et al ., 2008 ).外来种海桑(Sonneratia caseolaris )在深圳福田红树林保护区引种种植后, 提高了泥滩大型底栖动物的多样性(Leung & Tam, 2013 ).此外, 红树林中还存在许多其他外来植物, 但都仅止于调查记录(Biswas et al ., 2007 ; 周青青等, 2010 ). ...
红树林区内互花米草防除技术研究
1
2006
... 草甘膦(RodeoTM )是目前国内外互花米草控制中唯一得到实际应用的除草剂, 施用后能被植物迅速吸收, 影响植物关键生理过程而导致植株死亡(李富荣等, 2007 ).但是草甘膦等几种除草剂对互花米草的防除效果并不好.米草克(MCK)在使用两个月后对互花米草的防除效果达到100%, 具有很好的推广价值和应用前景(李兆飞, 2009 ).有的学者还自行研制能杀死互花米草的药剂, 该药剂不影响红树植物生长繁殖, 并对水生生物安全不构成影响(杜文琴等, 2006 ). ...
珠海鹤洲北其他种入侵人工恢复红树林群落的季节动态
1
2012
... 微甘菊在中国华南沿海的另一个重灾区是深圳湾(Ren et al ., 2014 ), 拦海造田使海水退去, 红树林底泥裸露, 微甘菊种子得以萌发生长, 严重危害红树林(于晓梅和杨逢建, 2011 ).微甘菊入侵深圳湾红树林后, 一方面导致红树植物枯萎, 减少了植被生物量; 另一方面增加了凋落物量, 促进了土壤微生物活动, 加速了土壤有机质分解释放, 降低了土壤碳储量, 削弱了红树林碳汇(毛子龙等, 2011 ).微甘菊的入侵, 还会影响人工恢复红树林的季节动态, 降低红树植物的成活率, 妨害红树林生态恢复(苗春玲等, 2012 ). ...
Biological invasions as a component of global change in stressed marine ecosystems.
1
2003
... 早期研究认为, 由于红树林生态系统极为苛刻的生境条件, 很少有外来物种能适应环境生存并形成入侵(Lugo, 1998 ).但在全球变化下, 尤其在人类活动的参与下, 生物进入到新生境的概率及入侵成功率大幅提升, 几乎没有什么生境能避免生物入侵(Dukes & Mooney, 1999 ; Occhipinti-Ambrogi & Savini, 2003 ).当前研究显示, 日渐增多的外来种已经出现在红树林生态系统中, 有些已造成明显的入侵危害(Bridgewater & Cresswell, 1999 ; Biswas et al ., 2007 ; Ren et al ., 2014 ).红树林生态系统植物物种少, 群落结构单一, 系统稳定性差, 被入侵破坏后难以恢复.与此同时, 红树林生态系统的组成物种也有可能入侵其他非土著地区, 改变这些地区原有的生态系统(Allen, 1998 ; Fourqurean et al ., 2010 ). ...
无瓣海桑造林是否造成九龙江口生物入侵的探讨
0
2006
无瓣海桑造林是否造成九龙江口生物入侵的探讨
0
2006
红树林区内互花米草防除技术研究
1
2006
... 草甘膦(RodeoTM )是目前国内外互花米草控制中唯一得到实际应用的除草剂, 施用后能被植物迅速吸收, 影响植物关键生理过程而导致植株死亡(李富荣等, 2007 ).但是草甘膦等几种除草剂对互花米草的防除效果并不好.米草克(MCK)在使用两个月后对互花米草的防除效果达到100%, 具有很好的推广价值和应用前景(李兆飞, 2009 ).有的学者还自行研制能杀死互花米草的药剂, 该药剂不影响红树植物生长繁殖, 并对水生生物安全不构成影响(杜文琴等, 2006 ). ...
1
1993
... 红树林是指分布在热带、亚热带海岸潮间带, 受海水周期性浸淹的木本植物群落; 上述定义强调红树林分布的非地带性以及与潮汐的关系(Duke, 1993 ; 张乔民等, 1997 ).红树林生态系统生境严苛, 高盐、高温、强风, 以及因受潮汐浸淹而底泥缺氧, 只有少数几种进化出高度适应特征的植物能生存(Kathiresan & Bingham, 2001 ).独特的红树林生态系统, 其生物入侵的表现既与其他生态系统有共同之处, 又有其自身特异性. ...
Invasion and abundance of reef-inhabiting fishes in the Vellar estuary, southeast coast of India, especially the lionfish Pterois volitans Linnaeus.
1
2012
... 狮子鱼原产于印度洋和太平洋海域, 20世纪80年代被引入大西洋与加勒比海域.红树林优越的觅食、繁育和避害条件, 使狮子鱼容易成功定居; 加之, 由于缺乏天敌, 其种群密度迅速增加(Arbeláez & Acero, 2011 ; Kulbicki et al ., 2012 ; Prakash et al ., 2012 ).这种鱼类在红树林的爆发, 明显降低了其他鱼类的种群数量, 严重损害了红树林的生态和经济价值(Barbour et al ., 2010 , 2011 ; Claydon et al ., 2012 ). ...
Studying invasion: Have we missed the boat?
1
2005
... 针对生物入侵的不同阶段, 应采取不同的防治手段.在外来种还没有传入新生境前, 主要是防御.对最初传入阶段的防御是代价最小且最有效的防治生物入侵手段(Puth & Post, 2005 ).强化检疫检验, 慎重引种, 以及引种后规范管理, 能在很大程度上减小外来物种入侵的可能性(Mack et al ., 2000 ).若防御失败, 外来物种成功传入新生境, 对外来物种的防治目标转变为根除外来种, 通常很难彻底根除.最后, 外来种成功入侵并造成严重的生态环境或经济后果, 防治目的变为控制入侵种的危害在一个可接受水平(Mack et al ., 2000 ; Sakai et al ., 2001 ; Theoharides & Dukes, 2007 ). ...
Patterns of alien plant invasion across coastal bay areas in southern China.
3
2014
... 早期研究认为, 由于红树林生态系统极为苛刻的生境条件, 很少有外来物种能适应环境生存并形成入侵(Lugo, 1998 ).但在全球变化下, 尤其在人类活动的参与下, 生物进入到新生境的概率及入侵成功率大幅提升, 几乎没有什么生境能避免生物入侵(Dukes & Mooney, 1999 ; Occhipinti-Ambrogi & Savini, 2003 ).当前研究显示, 日渐增多的外来种已经出现在红树林生态系统中, 有些已造成明显的入侵危害(Bridgewater & Cresswell, 1999 ; Biswas et al ., 2007 ; Ren et al ., 2014 ).红树林生态系统植物物种少, 群落结构单一, 系统稳定性差, 被入侵破坏后难以恢复.与此同时, 红树林生态系统的组成物种也有可能入侵其他非土著地区, 改变这些地区原有的生态系统(Allen, 1998 ; Fourqurean et al ., 2010 ). ...
... 现有的微甘菊入侵研究主要集中于陆地森林生态系统(Zhang et al ., 2004 ), 红树林同类研究比较少, 而且都集中于中国华南沿海红树林(Ren et al ., 2014 ).微甘菊进入红树林后, 迅速生长, 在红树植物上形成一层茂密的覆盖层, 强烈争夺资源(尤其是光照)及通过化感作用抑制或杀死红树植物(Maja et al ., 2008 ).张阳和张华(2008)研究过珠海淇澳岛红树林中微甘菊的分布模式与危害, 强调自然条件和人为因素在影响红树林中微甘菊的分布上起着重要作用; 推测当地微甘菊爆发主要是由于人类不适当的开发活动破坏了当地红树林. ...
... 微甘菊在中国华南沿海的另一个重灾区是深圳湾(Ren et al ., 2014 ), 拦海造田使海水退去, 红树林底泥裸露, 微甘菊种子得以萌发生长, 严重危害红树林(于晓梅和杨逢建, 2011 ).微甘菊入侵深圳湾红树林后, 一方面导致红树植物枯萎, 减少了植被生物量; 另一方面增加了凋落物量, 促进了土壤微生物活动, 加速了土壤有机质分解释放, 降低了土壤碳储量, 削弱了红树林碳汇(毛子龙等, 2011 ).微甘菊的入侵, 还会影响人工恢复红树林的季节动态, 降低红树植物的成活率, 妨害红树林生态恢复(苗春玲等, 2012 ). ...
Restoration of mangrove plantations and colonisation by native species in Leizhou Bay, South China.
1
2008
... 但是也有学者研究发现, 无瓣海桑对其林下乡土红树植物群落会产生不利影响.在无瓣海桑林下, 秋茄树(Kandelia obovata )受到的抑制最小, 可以正常生长; 木榄(Bruguiera gymnorhiza )、海桑、桐花树(Aegiceras corniculatum )受到的抑制次之; 红海兰(Rhizophora stylosa )的保存率最低(李玫等, 2004a ; 梁士楚等, 2005 ; 王旭等, 2008 ; 曾雯珺等, 2008 ; 姚历强, 2010 ).相对于乡土红树植物, 无瓣海桑具有优异的生物学形状——种子幼苗品质好, 抗逆性强(廖宝文等, 2004 ), 生物量大(Chen et al ., 2013b ), 生态位与乡土红树植物有显著重叠(Li et al ., 2011 ), 有一定的扩散能力以及对乡土红树植物的化感作用(李玫等, 2004b ), 以后有可能破坏甚至取代乡土红树植物群落(Ren et al ., 2008 ).因此, 现阶段研究更多建议决策者谨慎考虑无瓣海桑在红树林保护区引种种植. ...
Does global change increase the success of biological invaders?
1
1999
... 早期研究认为, 由于红树林生态系统极为苛刻的生境条件, 很少有外来物种能适应环境生存并形成入侵(Lugo, 1998 ).但在全球变化下, 尤其在人类活动的参与下, 生物进入到新生境的概率及入侵成功率大幅提升, 几乎没有什么生境能避免生物入侵(Dukes & Mooney, 1999 ; Occhipinti-Ambrogi & Savini, 2003 ).当前研究显示, 日渐增多的外来种已经出现在红树林生态系统中, 有些已造成明显的入侵危害(Bridgewater & Cresswell, 1999 ; Biswas et al ., 2007 ; Ren et al ., 2014 ).红树林生态系统植物物种少, 群落结构单一, 系统稳定性差, 被入侵破坏后难以恢复.与此同时, 红树林生态系统的组成物种也有可能入侵其他非土著地区, 改变这些地区原有的生态系统(Allen, 1998 ; Fourqurean et al ., 2010 ). ...
Biology and host range of the rust fungus Puccinia spegazzinii: A new classical biological control agent for the invasive, alien weed Mikania micrantha in Asia.
1
2008
... 微甘菊入侵后, 通常给当地生态系统带来了毁灭性灾害, 因此对微甘菊的防治需求十分紧迫(Waterhouse, 1994 ).物理和化学防治手段的弊端促使学者寻求生物手段来防治微甘菊入侵.业已证实, 寄生植物野地菟丝子(Cuscuta campestris )能安全有效地防治微甘菊(Lian et al ., 2006 ; Shen et al ., 2011 ), 一些新的微生物或病毒也能用来控制微甘菊(Ellison et al ., 2008 ; Wang et al ., 2008 ).除关注微甘菊对生态系统地上植物群落的危害外, 对生态系统其他结构(如地下生物)和功能的效应的关注也逐步形成(Li et al ., 2006 , 2007 ).微甘菊入侵机制的研究日渐成熟(Zhang et al ., 2004 ; Chen et al ., 2009 ).有关微甘菊入侵的分子手段研究(Huang et al ., 2012 ; Wang et al ., 2012 ), 以及微甘菊对全球气候变化的响应研究(Wang et al ., 2010 )初现端倪.随着微甘菊的持续入侵扩散, 温暖湿润沿海地区的红树林生态系统也难逃其入侵危害(Han et al ., 2006 ). ...
Sonneratia apetala Buch. Ham in the mangrove ecosystems of China: An invasive species or restoration species?
1
2009
... 虽然在群落或生态系统尺度上, 无瓣海桑确实能改良生境, 增加生物多样性, 但在更大尺度上是否有同样的效应, 目前还不清楚.无瓣海桑对林下乡土红树植物的竞争与化感抑制效应是不容忽视的.长远看, 无瓣海桑的入侵势必降低引入地植物和动物区系的特有性.只有对现有的无瓣海桑入侵与否进行深入探讨, 明确无瓣海桑能否爆发入侵, 才能为未来无瓣海桑的引种种植提供合理建议.反观现在中国红树林区大面积种植无瓣海桑的情况(Ren et al ., 2009 ), 不能不令人担忧. ...
The relative generality of plant invasion mechanisms and predicting future invasive plants.
1
2009
... 纵观生物入侵研究的发展历程, 生物入侵的相关机制与假说向来是入侵生态学研究的难点、热点, 备受争议, 至今没有哪一种机制或假说能解释大部分的生物入侵现象(Ren & Zhang, 2009 ; Lowry et al ., 2013 ).今后的整合研究或许能为研究生物入侵机制与假说提供新的视角(Kueffer et al ., 2013 ).随着人类对自然界改造的加剧, 全球贸易往来过度密切, 全球生物入侵形势复杂多变, 外来种入侵带来的生态环境、社会经济、人类健康及生物安全等问题前所未有地严峻(Bradley et al ., 2012 ).目前生物入侵已成为全球变化的因素, 也是导致全球生物多样性丧失的主要原因, 仅次于生境破坏, 并很可能成为第一因素(Mack et al ., 2000 ).生物入侵将与其他全球变化因素一起左右着地球的未来. ...
Naturalization and invasion of alien plants: Concepts and definitions.
1
2000
... 迄今, 生物入侵研究已发展成一门活跃的生态学分支学科——入侵生态学.生物入侵研究从入侵现状的调查入手, 体现为入侵种多样性与分布格局、入侵效应的调查与评价(Richardson et al ., 2000 ; 李博和陈家宽, 2002 ).在洞悉生物入侵现状后, 紧随的研究是探索生物入侵发生的原因和机制, 探讨外来种的入侵潜力与入侵性(invasiveness)、繁殖体压力(propagule pressure)、生境或生态系统的可入侵性(invasibility)、入侵种与被侵入系统的相互反馈及诸多假说(Sakai et al ., 2001 ; Theoharides & Dukes, 2007 ; Lowry et al ., 2013 ).最后, 在明确生物入侵发生的原因和机制基础上, 对生物入侵进行防治和管理, 以期减小其危害, 并对未来入侵进行合理的预测(Mack et al ., 2000 ). ...
Invasion of coastal marine communities in North America: Apparent patterns, processes, and biases.
2
2000
... 此外, 一些软体、海绵或腔肠动物(如牡蛎、贻贝、海绵等)由于人为活动传入非土著地区, 暂时还没有大肆爆发形成入侵危害(Holland et al ., 2004 ; Salgado-Barragán & Toledano-Granados, 2006 ; Bishop et al ., 2010 ; Baker et al ., 2012 ; Alvizu et al ., 2013 ); 还有一些入侵非土著海洋的动物是否入侵红树林尚不明确(Ruiz et al ., 2000 ; 陆琴燕等, 2013 ), 有待今后持续关注. ...
... 微生物入侵红树林的研究寥寥无几.微生物入侵多半是依附在入侵的动植物上, 被动植物带入新生境.与小叶海金沙共生的丛枝菌根真菌(AMF), 在植物入侵红树林时被引入(Soti et al ., 2014 ).一些入侵非土著海洋的微生物是否入侵红树林, 目前仍不明确(Ruiz et al ., 2000 ; 陆琴燕等, 2013 ), 有待关注.而附属于红树植物木材上的一些真菌种类, 未经严格的检疫检验, 通过商业贸易交易到非土著地区, 成为极具危险的潜在入侵病原体(Vannini et al ., 2012 ).研究红树林植物或动物入侵时, 关注与它们共生或寄生的微生物, 是研究微生物入侵与防治的切实可行的途径. ...
The population biology of invasive species.
2
2001
... 迄今, 生物入侵研究已发展成一门活跃的生态学分支学科——入侵生态学.生物入侵研究从入侵现状的调查入手, 体现为入侵种多样性与分布格局、入侵效应的调查与评价(Richardson et al ., 2000 ; 李博和陈家宽, 2002 ).在洞悉生物入侵现状后, 紧随的研究是探索生物入侵发生的原因和机制, 探讨外来种的入侵潜力与入侵性(invasiveness)、繁殖体压力(propagule pressure)、生境或生态系统的可入侵性(invasibility)、入侵种与被侵入系统的相互反馈及诸多假说(Sakai et al ., 2001 ; Theoharides & Dukes, 2007 ; Lowry et al ., 2013 ).最后, 在明确生物入侵发生的原因和机制基础上, 对生物入侵进行防治和管理, 以期减小其危害, 并对未来入侵进行合理的预测(Mack et al ., 2000 ). ...
... 针对生物入侵的不同阶段, 应采取不同的防治手段.在外来种还没有传入新生境前, 主要是防御.对最初传入阶段的防御是代价最小且最有效的防治生物入侵手段(Puth & Post, 2005 ).强化检疫检验, 慎重引种, 以及引种后规范管理, 能在很大程度上减小外来物种入侵的可能性(Mack et al ., 2000 ).若防御失败, 外来物种成功传入新生境, 对外来物种的防治目标转变为根除外来种, 通常很难彻底根除.最后, 外来种成功入侵并造成严重的生态环境或经济后果, 防治目的变为控制入侵种的危害在一个可接受水平(Mack et al ., 2000 ; Sakai et al ., 2001 ; Theoharides & Dukes, 2007 ). ...
The false mussel Mytilopsis adamsi Morrison, 1946 (Mollusca: Bivalvia: Dreissenidae) in the Pacific waters of Mexico: A case of biological invasion.
1
2006
... 此外, 一些软体、海绵或腔肠动物(如牡蛎、贻贝、海绵等)由于人为活动传入非土著地区, 暂时还没有大肆爆发形成入侵危害(Holland et al ., 2004 ; Salgado-Barragán & Toledano-Granados, 2006 ; Bishop et al ., 2010 ; Baker et al ., 2012 ; Alvizu et al ., 2013 ); 还有一些入侵非土著海洋的动物是否入侵红树林尚不明确(Ruiz et al ., 2000 ; 陆琴燕等, 2013 ), 有待今后持续关注. ...
薇甘菊的生物防治及其天敌在中国的新发现
1
2002
... 很多研究证实寄生植物野地菟丝子能严重影响微甘菊茎叶生长及开花结实, 但不影响其他植物正常的生长繁殖, 用菟丝子控制微甘菊安全可行(昝启杰等, 2003 ).在入侵地发掘一些微甘菊的天敌, 像昆虫、微生物等, 也能控制微甘菊(邵华等, 2002 ).通过植物搭配种植, 种植当地适生的乔灌草植物或促进野生植物生长, 建立稳定的复层群落结构, 也可持续有效地控制微甘菊(殷祚云等, 2003 ). ...
薇甘菊的生物防治及其天敌在中国的新发现
1
2002
... 很多研究证实寄生植物野地菟丝子能严重影响微甘菊茎叶生长及开花结实, 但不影响其他植物正常的生长繁殖, 用菟丝子控制微甘菊安全可行(昝启杰等, 2003 ).在入侵地发掘一些微甘菊的天敌, 像昆虫、微生物等, 也能控制微甘菊(邵华等, 2002 ).通过植物搭配种植, 种植当地适生的乔灌草植物或促进野生植物生长, 建立稳定的复层群落结构, 也可持续有效地控制微甘菊(殷祚云等, 2003 ). ...
The response of the invasive weed Mikania micrantha to infection density of the obligate parasite Cuscuta campestris and its implications for biological control of M. micrantha.
1
2011
... 微甘菊入侵后, 通常给当地生态系统带来了毁灭性灾害, 因此对微甘菊的防治需求十分紧迫(Waterhouse, 1994 ).物理和化学防治手段的弊端促使学者寻求生物手段来防治微甘菊入侵.业已证实, 寄生植物野地菟丝子(Cuscuta campestris )能安全有效地防治微甘菊(Lian et al ., 2006 ; Shen et al ., 2011 ), 一些新的微生物或病毒也能用来控制微甘菊(Ellison et al ., 2008 ; Wang et al ., 2008 ).除关注微甘菊对生态系统地上植物群落的危害外, 对生态系统其他结构(如地下生物)和功能的效应的关注也逐步形成(Li et al ., 2006 , 2007 ).微甘菊入侵机制的研究日渐成熟(Zhang et al ., 2004 ; Chen et al ., 2009 ).有关微甘菊入侵的分子手段研究(Huang et al ., 2012 ; Wang et al ., 2012 ), 以及微甘菊对全球气候变化的响应研究(Wang et al ., 2010 )初现端倪.随着微甘菊的持续入侵扩散, 温暖湿润沿海地区的红树林生态系统也难逃其入侵危害(Han et al ., 2006 ). ...
Top-down control of Spartina alterniflora production by periwinkle grazing in a Virginia salt marsh.
1
2001
... 目前研究显示, 玉黍螺(Littoraria irrorata )、麦角菌(Ciavieps purpurea )和光蝉(Prokelisia marginata )在互花米草生物控制中得到应用(王卿等, 2006 ).玉黍螺直接取食互花米草的叶片, 抑制其生长(Silliman & Zieman, 2001 ).麦角菌能使互花米草感染麦角病, 在种子内形成菌核, 降低其种子产量, 限制其扩散(Fisher et al ., 2005 ).但麦角菌对禾本科其他一些植物也有影响, 而且其适应性与变异性强, 因此引用麦角菌作为生物控制途径还有待进一步研究(李富荣等, 2007 ).还有一种从华盛顿引入的光蝉, 对入侵威拉帕湾的互花米草具有很强的杀伤力, 对其他植物无明显影响(Grevstad et al ., 2003 ). ...
Invasive mangrove removal and recovery: Food web effects across a chronosequence.
1
2013
... 外来红树植物Rhizophora mangle 入侵夏威夷海岛无植被裸滩, 显著改变了这里原有的生态系统结构与功能.红树群落高的生产力大幅增加了整个区域的生物量(Cox & Allen, 1999 ; Fry & Cormier, 2011 ; D’iorio, 2003 ).Rhizophora mangle 对地下部分的凋落物输入, 改善了沉积物的理化特征, 并改变了原有的食物链(网)结构, 丰富了底栖生物群落(Demopoulos, 2004 ; Demopoulos et al ., 2007 ; Siple & Donahue, 2013 ).底栖生物群落的丰富多样, 为进一步的外来生物入侵提供了优良的条件, 形成一个外来生物入侵循环(Demopoulos & Smith, 2010 ).外来红树植物入侵也对底栖生态系统功能产生了深远的影响, 夏威夷群岛海岸的沉积物代谢和碳再矿化动态在移除入侵种Rhizophora mangle 数年后仍没有摆脱其影响(Sweetman et al ., 2010 ). ...
Structure and degree of perturbation of mangroves at Tijuca Lagoon, Rio de Janeiro, RJ, Brazil.
0
1999
Mycorrhizal symbiosis and Lygodium microphy- llum invasion in South Florida—A biogeographic compar- ison.
1
2014
... 微生物入侵红树林的研究寥寥无几.微生物入侵多半是依附在入侵的动植物上, 被动植物带入新生境.与小叶海金沙共生的丛枝菌根真菌(AMF), 在植物入侵红树林时被引入(Soti et al ., 2014 ).一些入侵非土著海洋的微生物是否入侵红树林, 目前仍不明确(Ruiz et al ., 2000 ; 陆琴燕等, 2013 ), 有待关注.而附属于红树植物木材上的一些真菌种类, 未经严格的检疫检验, 通过商业贸易交易到非土著地区, 成为极具危险的潜在入侵病原体(Vannini et al ., 2012 ).研究红树林植物或动物入侵时, 关注与它们共生或寄生的微生物, 是研究微生物入侵与防治的切实可行的途径. ...
Impacts of exotic mangrove forests and mangrove deforestation on carbon remineralization and ecosystem functioning in marine sediments.
1
2010
... 外来红树植物Rhizophora mangle 入侵夏威夷海岛无植被裸滩, 显著改变了这里原有的生态系统结构与功能.红树群落高的生产力大幅增加了整个区域的生物量(Cox & Allen, 1999 ; Fry & Cormier, 2011 ; D’iorio, 2003 ).Rhizophora mangle 对地下部分的凋落物输入, 改善了沉积物的理化特征, 并改变了原有的食物链(网)结构, 丰富了底栖生物群落(Demopoulos, 2004 ; Demopoulos et al ., 2007 ; Siple & Donahue, 2013 ).底栖生物群落的丰富多样, 为进一步的外来生物入侵提供了优良的条件, 形成一个外来生物入侵循环(Demopoulos & Smith, 2010 ).外来红树植物入侵也对底栖生态系统功能产生了深远的影响, 夏威夷群岛海岸的沉积物代谢和碳再矿化动态在移除入侵种Rhizophora mangle 数年后仍没有摆脱其影响(Sweetman et al ., 2010 ). ...
无瓣海桑对互花米草的生态控制效果
1
2007
... 一些研究认为无瓣海桑通过竞争或化感作用能有效地控制红树林区互花米草的滋生蔓延, 最后取代互花米草(唐国玲等, 2007 ; 李静等, 2011 ).这种植物替代的生物控制方法虽有很好的应用前景, 但考虑到无瓣海桑的引入对土著红树林也存在一定的危害, 须谨慎采用. ...
无瓣海桑对互花米草的生态控制效果
1
2007
... 一些研究认为无瓣海桑通过竞争或化感作用能有效地控制红树林区互花米草的滋生蔓延, 最后取代互花米草(唐国玲等, 2007 ; 李静等, 2011 ).这种植物替代的生物控制方法虽有很好的应用前景, 但考虑到无瓣海桑的引入对土著红树林也存在一定的危害, 须谨慎采用. ...
Plant invasion across space and time: Factors affecting nonindigenous species success during four stages of invasion.
2
2007
... 迄今, 生物入侵研究已发展成一门活跃的生态学分支学科——入侵生态学.生物入侵研究从入侵现状的调查入手, 体现为入侵种多样性与分布格局、入侵效应的调查与评价(Richardson et al ., 2000 ; 李博和陈家宽, 2002 ).在洞悉生物入侵现状后, 紧随的研究是探索生物入侵发生的原因和机制, 探讨外来种的入侵潜力与入侵性(invasiveness)、繁殖体压力(propagule pressure)、生境或生态系统的可入侵性(invasibility)、入侵种与被侵入系统的相互反馈及诸多假说(Sakai et al ., 2001 ; Theoharides & Dukes, 2007 ; Lowry et al ., 2013 ).最后, 在明确生物入侵发生的原因和机制基础上, 对生物入侵进行防治和管理, 以期减小其危害, 并对未来入侵进行合理的预测(Mack et al ., 2000 ). ...
... 针对生物入侵的不同阶段, 应采取不同的防治手段.在外来种还没有传入新生境前, 主要是防御.对最初传入阶段的防御是代价最小且最有效的防治生物入侵手段(Puth & Post, 2005 ).强化检疫检验, 慎重引种, 以及引种后规范管理, 能在很大程度上减小外来物种入侵的可能性(Mack et al ., 2000 ).若防御失败, 外来物种成功传入新生境, 对外来物种的防治目标转变为根除外来种, 通常很难彻底根除.最后, 外来种成功入侵并造成严重的生态环境或经济后果, 防治目的变为控制入侵种的危害在一个可接受水平(Mack et al ., 2000 ; Sakai et al ., 2001 ; Theoharides & Dukes, 2007 ). ...
1
2011
... 微甘菊是菊科假泽兰属多年生草质藤本, 原产于热带南美与中美洲.微甘菊生长迅速且适应性强, 英文称其为“Mile-a-Minute Weed” (一分钟一英里杂草), 极具入侵性(Waterhouse, 1994 ).微甘菊在原产地因有天敌控制不具危害, 但在缺乏天敌的入侵地迅速发展为危害性严重的有害杂草(昝启杰等, 2000 ).微甘菊因其独特的生物学特性及生态适应能力(Tripathi et al ., 2011 ), 传入新生境后能迅速爆发.如今, 微甘菊已在整个亚洲热带及北美地区肆虐, 成为著名的世界性恶草(Zhang et al ., 2004 ). ...
Manu- factured wood trade to Europe: A potential uninspected carrier of alien fungi.
1
2012
... 微生物入侵红树林的研究寥寥无几.微生物入侵多半是依附在入侵的动植物上, 被动植物带入新生境.与小叶海金沙共生的丛枝菌根真菌(AMF), 在植物入侵红树林时被引入(Soti et al ., 2014 ).一些入侵非土著海洋的微生物是否入侵红树林, 目前仍不明确(Ruiz et al ., 2000 ; 陆琴燕等, 2013 ), 有待关注.而附属于红树植物木材上的一些真菌种类, 未经严格的检疫检验, 通过商业贸易交易到非土著地区, 成为极具危险的潜在入侵病原体(Vannini et al ., 2012 ).研究红树林植物或动物入侵时, 关注与它们共生或寄生的微生物, 是研究微生物入侵与防治的切实可行的途径. ...
Monitoring the invasion of Spartina alterniflora using very high resolution unmanned aerial vehicle imagery in Beihai, Guangxi (China).
1
2014
... 互花米草有200多年的入侵扩张历史.不同国家和地区对互花米草的入侵现状、入侵效应、入侵原因与机制, 以及相关防治措施都有过全面而系统的研究(Daehler & Strong, 1996 ; Li et al ., 2009 ).现阶段对互花米草入侵的研究, 一方面是从分子水平揭示互花米草成功入侵的遗传学基础、耐受性以及表型可塑性的分子机理或原因(吴昊和丁建清, 2014 ); 另一方面是应用3S技术, 从景观以上尺度监测互花米草入侵的动态变化(Wan et al ., 2014 ).互花米草入侵不可避免地要受全球变化影响, 二者的相互反馈在全球变化日益强烈的今天愈加受到重视(Emery & Fulweiler, 2014 ).随着互花米草在海岸滩涂的快速扩散, 海岸带上所有的生态系统不可避免地会受到互花米草入侵的威胁.互花米草对潮间带生境的高度适应性, 使之在入侵地扩散, 并成功地入侵红树林生态系统(Zhang et al ., 2012 ). ...
互花米草入侵对红树林湿地潮滩大型底栖动物群落的影响
1
2014
... 另外, 浙江乐清西门岛红树林区, 互花米草侵占的滩涂与原有潮滩相比, 大型底栖动物群落多样性指数并没有下降, 但种类组成和群落结构显著不同(王安安等, 2014 ).而且营养价值高的互花米草会成为入侵区域底栖生物的主要食物来源, 改变着底栖营养关系(温作柱等, 2014 ). ...
互花米草入侵对红树林湿地潮滩大型底栖动物群落的影响
1
2014
... 另外, 浙江乐清西门岛红树林区, 互花米草侵占的滩涂与原有潮滩相比, 大型底栖动物群落多样性指数并没有下降, 但种类组成和群落结构显著不同(王安安等, 2014 ).而且营养价值高的互花米草会成为入侵区域底栖生物的主要食物来源, 改变着底栖营养关系(温作柱等, 2014 ). ...
互花米草对福建泉州湾海岸湿地沉积环境影响
1
2008
... 互花米草已经出现在中国所有的红树林区.对福建境内红树林互花米草入侵的研究走在国内前列.互花米草入侵导致红树林生境土壤退化, 漳江口互花米草生境的土壤有机碳、土壤微生物生物量碳、氮含量以及土壤蔗糖酶与磷酸酶活性等相对于红树林生境均有明显下降(张祥霖等, 2008 ).但是, 互花米草群落对碳、氮具有很强的富集能力, 相对于红树林对碳、氮弱的富集能力, 互花米草的存在能更好地拦截和吸附陆源污染物, 并使重金属富集在互花米草群落沉积物中, 增加泉州湾红树林污染富集承受能力(王爱军等, 2008 ). ...
互花米草对福建泉州湾海岸湿地沉积环境影响
1
2008
... 互花米草已经出现在中国所有的红树林区.对福建境内红树林互花米草入侵的研究走在国内前列.互花米草入侵导致红树林生境土壤退化, 漳江口互花米草生境的土壤有机碳、土壤微生物生物量碳、氮含量以及土壤蔗糖酶与磷酸酶活性等相对于红树林生境均有明显下降(张祥霖等, 2008 ).但是, 互花米草群落对碳、氮具有很强的富集能力, 相对于红树林对碳、氮弱的富集能力, 互花米草的存在能更好地拦截和吸附陆源污染物, 并使重金属富集在互花米草群落沉积物中, 增加泉州湾红树林污染富集承受能力(王爱军等, 2008 ). ...
Differences in burrow morphology of crabs between Spartina alterniflora marsh and mangrove habitats.
1
2014
... 互花米草入侵还会改变红树林的生物多样性及其行为模式.在九龙江口红树林区, 互花米草入侵区域底栖动物的栖息密度、种类多样性、丰度比红树林区高, 比涂滩要低(洪荣标等, 2005 ).互花米草的入侵还改变着氨氧化古菌与细菌的丰度, 影响氨氧化微生物的群落结构(Zhang et al ., 2011 ).在漳江口红树林区, 红树林生境与互花米草生境之间多毛类动物栖息密度、生物量、丰度指数、均匀度指数和多样性指数均有显著差异(陈昕韡等, 2012 ).红树林与互花米草生境大型底栖动物次级生产力也存在差异, 海榄雌(Avicennia marina )生境大型底栖动物的年次级生产力最高, 而互花米草生境大型底栖动物的P/B值(P: 平均次级生产力; B: 平均生物量)最高, 因其优势类群为个体较小的寡毛类(吴辰等, 2013 ).再有, 互花米草相对较高的根密度显著降低了红树林中蟹类在互花米草生境打洞的深度及洞穴的复杂性(Wang et al ., 2014 ). ...
入侵植物互花米草——生物学、生态学及管理
2
2006
... 互花米草是禾本科米草属(又名绳草属)多年生草本植物, 原产于美洲大西洋沿岸和墨西哥湾.近200年来, 经不同的入侵途径, 互花米草的分布区域已经从其原产地扩展到欧洲、北美西海岸、新西兰与中国沿海, 被公认为是全球海岸滩涂湿地生态系统最成功的入侵植物之一(王卿等, 2006 ).外界条件与互花米草自身特性相辅相成, 促成了互花米草高效的、难以防治的入侵模式:快速扩张呈现出点源扩散和多点爆发的特征, 对乡土植被的侵占表现为交互生长与斑块推进相结合(Xiao et al ., 2010 ).因此, 互花米草一旦成功入侵, 很难根除和控制. ...
... 目前研究显示, 玉黍螺(Littoraria irrorata )、麦角菌(Ciavieps purpurea )和光蝉(Prokelisia marginata )在互花米草生物控制中得到应用(王卿等, 2006 ).玉黍螺直接取食互花米草的叶片, 抑制其生长(Silliman & Zieman, 2001 ).麦角菌能使互花米草感染麦角病, 在种子内形成菌核, 降低其种子产量, 限制其扩散(Fisher et al ., 2005 ).但麦角菌对禾本科其他一些植物也有影响, 而且其适应性与变异性强, 因此引用麦角菌作为生物控制途径还有待进一步研究(李富荣等, 2007 ).还有一种从华盛顿引入的光蝉, 对入侵威拉帕湾的互花米草具有很强的杀伤力, 对其他植物无明显影响(Grevstad et al ., 2003 ). ...
入侵植物互花米草——生物学、生态学及管理
2
2006
... 互花米草是禾本科米草属(又名绳草属)多年生草本植物, 原产于美洲大西洋沿岸和墨西哥湾.近200年来, 经不同的入侵途径, 互花米草的分布区域已经从其原产地扩展到欧洲、北美西海岸、新西兰与中国沿海, 被公认为是全球海岸滩涂湿地生态系统最成功的入侵植物之一(王卿等, 2006 ).外界条件与互花米草自身特性相辅相成, 促成了互花米草高效的、难以防治的入侵模式:快速扩张呈现出点源扩散和多点爆发的特征, 对乡土植被的侵占表现为交互生长与斑块推进相结合(Xiao et al ., 2010 ).因此, 互花米草一旦成功入侵, 很难根除和控制. ...
... 目前研究显示, 玉黍螺(Littoraria irrorata )、麦角菌(Ciavieps purpurea )和光蝉(Prokelisia marginata )在互花米草生物控制中得到应用(王卿等, 2006 ).玉黍螺直接取食互花米草的叶片, 抑制其生长(Silliman & Zieman, 2001 ).麦角菌能使互花米草感染麦角病, 在种子内形成菌核, 降低其种子产量, 限制其扩散(Fisher et al ., 2005 ).但麦角菌对禾本科其他一些植物也有影响, 而且其适应性与变异性强, 因此引用麦角菌作为生物控制途径还有待进一步研究(李富荣等, 2007 ).还有一种从华盛顿引入的光蝉, 对入侵威拉帕湾的互花米草具有很强的杀伤力, 对其他植物无明显影响(Grevstad et al ., 2003 ). ...
Genome sequence and characterization of a new virus infecting Mikania micrantha H. B. K.
1
2008
... 微甘菊入侵后, 通常给当地生态系统带来了毁灭性灾害, 因此对微甘菊的防治需求十分紧迫(Waterhouse, 1994 ).物理和化学防治手段的弊端促使学者寻求生物手段来防治微甘菊入侵.业已证实, 寄生植物野地菟丝子(Cuscuta campestris )能安全有效地防治微甘菊(Lian et al ., 2006 ; Shen et al ., 2011 ), 一些新的微生物或病毒也能用来控制微甘菊(Ellison et al ., 2008 ; Wang et al ., 2008 ).除关注微甘菊对生态系统地上植物群落的危害外, 对生态系统其他结构(如地下生物)和功能的效应的关注也逐步形成(Li et al ., 2006 , 2007 ).微甘菊入侵机制的研究日渐成熟(Zhang et al ., 2004 ; Chen et al ., 2009 ).有关微甘菊入侵的分子手段研究(Huang et al ., 2012 ; Wang et al ., 2012 ), 以及微甘菊对全球气候变化的响应研究(Wang et al ., 2010 )初现端倪.随着微甘菊的持续入侵扩散, 温暖湿润沿海地区的红树林生态系统也难逃其入侵危害(Han et al ., 2006 ). ...
Responses of Mikania micrantha, an invasive weed to elevated CO2 : Induction of β-caryophyllene synthase, changes in emission capability and allelopathic potential of β-caryophyllene.
1
2010
... 微甘菊入侵后, 通常给当地生态系统带来了毁灭性灾害, 因此对微甘菊的防治需求十分紧迫(Waterhouse, 1994 ).物理和化学防治手段的弊端促使学者寻求生物手段来防治微甘菊入侵.业已证实, 寄生植物野地菟丝子(Cuscuta campestris )能安全有效地防治微甘菊(Lian et al ., 2006 ; Shen et al ., 2011 ), 一些新的微生物或病毒也能用来控制微甘菊(Ellison et al ., 2008 ; Wang et al ., 2008 ).除关注微甘菊对生态系统地上植物群落的危害外, 对生态系统其他结构(如地下生物)和功能的效应的关注也逐步形成(Li et al ., 2006 , 2007 ).微甘菊入侵机制的研究日渐成熟(Zhang et al ., 2004 ; Chen et al ., 2009 ).有关微甘菊入侵的分子手段研究(Huang et al ., 2012 ; Wang et al ., 2012 ), 以及微甘菊对全球气候变化的响应研究(Wang et al ., 2010 )初现端倪.随着微甘菊的持续入侵扩散, 温暖湿润沿海地区的红树林生态系统也难逃其入侵危害(Han et al ., 2006 ). ...
AFLP genome scan to detect genetic structure and candidate loci under selection for local adaptation of the invasive weed Mikania micrantha.
1
2012
... 微甘菊入侵后, 通常给当地生态系统带来了毁灭性灾害, 因此对微甘菊的防治需求十分紧迫(Waterhouse, 1994 ).物理和化学防治手段的弊端促使学者寻求生物手段来防治微甘菊入侵.业已证实, 寄生植物野地菟丝子(Cuscuta campestris )能安全有效地防治微甘菊(Lian et al ., 2006 ; Shen et al ., 2011 ), 一些新的微生物或病毒也能用来控制微甘菊(Ellison et al ., 2008 ; Wang et al ., 2008 ).除关注微甘菊对生态系统地上植物群落的危害外, 对生态系统其他结构(如地下生物)和功能的效应的关注也逐步形成(Li et al ., 2006 , 2007 ).微甘菊入侵机制的研究日渐成熟(Zhang et al ., 2004 ; Chen et al ., 2009 ).有关微甘菊入侵的分子手段研究(Huang et al ., 2012 ; Wang et al ., 2012 ), 以及微甘菊对全球气候变化的响应研究(Wang et al ., 2010 )初现端倪.随着微甘菊的持续入侵扩散, 温暖湿润沿海地区的红树林生态系统也难逃其入侵危害(Han et al ., 2006 ). ...
海南东寨港无瓣海桑——海桑人工红树群落结构特征
1
2008
... 但是也有学者研究发现, 无瓣海桑对其林下乡土红树植物群落会产生不利影响.在无瓣海桑林下, 秋茄树(Kandelia obovata )受到的抑制最小, 可以正常生长; 木榄(Bruguiera gymnorhiza )、海桑、桐花树(Aegiceras corniculatum )受到的抑制次之; 红海兰(Rhizophora stylosa )的保存率最低(李玫等, 2004a ; 梁士楚等, 2005 ; 王旭等, 2008 ; 曾雯珺等, 2008 ; 姚历强, 2010 ).相对于乡土红树植物, 无瓣海桑具有优异的生物学形状——种子幼苗品质好, 抗逆性强(廖宝文等, 2004 ), 生物量大(Chen et al ., 2013b ), 生态位与乡土红树植物有显著重叠(Li et al ., 2011 ), 有一定的扩散能力以及对乡土红树植物的化感作用(李玫等, 2004b ), 以后有可能破坏甚至取代乡土红树植物群落(Ren et al ., 2008 ).因此, 现阶段研究更多建议决策者谨慎考虑无瓣海桑在红树林保护区引种种植. ...
海南东寨港无瓣海桑——海桑人工红树群落结构特征
1
2008
... 但是也有学者研究发现, 无瓣海桑对其林下乡土红树植物群落会产生不利影响.在无瓣海桑林下, 秋茄树(Kandelia obovata )受到的抑制最小, 可以正常生长; 木榄(Bruguiera gymnorhiza )、海桑、桐花树(Aegiceras corniculatum )受到的抑制次之; 红海兰(Rhizophora stylosa )的保存率最低(李玫等, 2004a ; 梁士楚等, 2005 ; 王旭等, 2008 ; 曾雯珺等, 2008 ; 姚历强, 2010 ).相对于乡土红树植物, 无瓣海桑具有优异的生物学形状——种子幼苗品质好, 抗逆性强(廖宝文等, 2004 ), 生物量大(Chen et al ., 2013b ), 生态位与乡土红树植物有显著重叠(Li et al ., 2011 ), 有一定的扩散能力以及对乡土红树植物的化感作用(李玫等, 2004b ), 以后有可能破坏甚至取代乡土红树植物群落(Ren et al ., 2008 ).因此, 现阶段研究更多建议决策者谨慎考虑无瓣海桑在红树林保护区引种种植. ...
深圳福田无瓣海桑与海桑人工林鸟类群落研究及生态评价
1
2001
... 无瓣海桑在引入地多用作沿海涂滩造林与恢复树种, 具有明显的改良土壤效应, 可增加土壤有机质和N、P、K含量, 有利于后续乡土红树植物定植(韩维栋等, 2003 ).在乡土红树林群落中混种无瓣海桑, 虽然整个林分生物量或土壤碳库没有显著增加, 但无瓣海桑相对较高的凋落物量和低C/N比, 有利于分解者利用(Lu et al ., 2014 ).无瓣海桑的引入能提高群落结构的复杂性与生物多样性.在深圳福田红树林保护区, 混种无瓣海桑+海桑人工林, 相对于天然红树林, 鸟类群落生物多样性得到提高(王勇军和昝启杰, 2001 ).鉴于上述研究结果, 一些学者认为无瓣海桑并没有表现出入侵危害(Zan et al ., 2003 ; 朱宏伟, 2012 ; Xin et al ., 2013 ). ...
深圳福田无瓣海桑与海桑人工林鸟类群落研究及生态评价
1
2001
... 无瓣海桑在引入地多用作沿海涂滩造林与恢复树种, 具有明显的改良土壤效应, 可增加土壤有机质和N、P、K含量, 有利于后续乡土红树植物定植(韩维栋等, 2003 ).在乡土红树林群落中混种无瓣海桑, 虽然整个林分生物量或土壤碳库没有显著增加, 但无瓣海桑相对较高的凋落物量和低C/N比, 有利于分解者利用(Lu et al ., 2014 ).无瓣海桑的引入能提高群落结构的复杂性与生物多样性.在深圳福田红树林保护区, 混种无瓣海桑+海桑人工林, 相对于天然红树林, 鸟类群落生物多样性得到提高(王勇军和昝启杰, 2001 ).鉴于上述研究结果, 一些学者认为无瓣海桑并没有表现出入侵危害(Zan et al ., 2003 ; 朱宏伟, 2012 ; Xin et al ., 2013 ). ...
冬季火烧与收割对互花米草地上部分生长与繁殖的影响
1
2006
... 持续的淡咸水轮换浇灌, 互花米草的生物量积累、无性与有性繁殖能力都会受到抑制(陈琳等, 2007 ).而且选择关键季节, 刈割互花米草地上部分, 同时配合一定水位的持续淹水, 更能有效地发挥治理互花米草的效果(袁琳等, 2008 ).有些物理控制措施, 如遮阴或冬季火烧, 对互花米草影响不大, 反而会影响乡土植物生长, 促进互花米草的入侵, 应当避免 (陈中义等, 2005 ; 王智晨等, 2006 ). ...
冬季火烧与收割对互花米草地上部分生长与繁殖的影响
1
2006
... 持续的淡咸水轮换浇灌, 互花米草的生物量积累、无性与有性繁殖能力都会受到抑制(陈琳等, 2007 ).而且选择关键季节, 刈割互花米草地上部分, 同时配合一定水位的持续淹水, 更能有效地发挥治理互花米草的效果(袁琳等, 2008 ).有些物理控制措施, 如遮阴或冬季火烧, 对互花米草影响不大, 反而会影响乡土植物生长, 促进互花米草的入侵, 应当避免 (陈中义等, 2005 ; 王智晨等, 2006 ). ...
Biological Control of Weeds: Southeast Asian Prospects
3
1994
... 微甘菊是菊科假泽兰属多年生草质藤本, 原产于热带南美与中美洲.微甘菊生长迅速且适应性强, 英文称其为“Mile-a-Minute Weed” (一分钟一英里杂草), 极具入侵性(Waterhouse, 1994 ).微甘菊在原产地因有天敌控制不具危害, 但在缺乏天敌的入侵地迅速发展为危害性严重的有害杂草(昝启杰等, 2000 ).微甘菊因其独特的生物学特性及生态适应能力(Tripathi et al ., 2011 ), 传入新生境后能迅速爆发.如今, 微甘菊已在整个亚洲热带及北美地区肆虐, 成为著名的世界性恶草(Zhang et al ., 2004 ). ...
... 微甘菊入侵后, 通常给当地生态系统带来了毁灭性灾害, 因此对微甘菊的防治需求十分紧迫(Waterhouse, 1994 ).物理和化学防治手段的弊端促使学者寻求生物手段来防治微甘菊入侵.业已证实, 寄生植物野地菟丝子(Cuscuta campestris )能安全有效地防治微甘菊(Lian et al ., 2006 ; Shen et al ., 2011 ), 一些新的微生物或病毒也能用来控制微甘菊(Ellison et al ., 2008 ; Wang et al ., 2008 ).除关注微甘菊对生态系统地上植物群落的危害外, 对生态系统其他结构(如地下生物)和功能的效应的关注也逐步形成(Li et al ., 2006 , 2007 ).微甘菊入侵机制的研究日渐成熟(Zhang et al ., 2004 ; Chen et al ., 2009 ).有关微甘菊入侵的分子手段研究(Huang et al ., 2012 ; Wang et al ., 2012 ), 以及微甘菊对全球气候变化的响应研究(Wang et al ., 2010 )初现端倪.随着微甘菊的持续入侵扩散, 温暖湿润沿海地区的红树林生态系统也难逃其入侵危害(Han et al ., 2006 ). ...
... 常见的控制外来生物入侵爆发方法主要有物理、化学和生物三大类(Waterhouse, 1994 ; Mack et al ., 2000 ).针对红树林生态系统现有的主要成功 入侵且危害严重的入侵种, 我们总结了以下控制方法. ...
互花米草入侵对西门岛潮滩底栖食物网的影响
1
2014
... 另外, 浙江乐清西门岛红树林区, 互花米草侵占的滩涂与原有潮滩相比, 大型底栖动物群落多样性指数并没有下降, 但种类组成和群落结构显著不同(王安安等, 2014 ).而且营养价值高的互花米草会成为入侵区域底栖生物的主要食物来源, 改变着底栖营养关系(温作柱等, 2014 ). ...
互花米草入侵对西门岛潮滩底栖食物网的影响
1
2014
... 另外, 浙江乐清西门岛红树林区, 互花米草侵占的滩涂与原有潮滩相比, 大型底栖动物群落多样性指数并没有下降, 但种类组成和群落结构显著不同(王安安等, 2014 ).而且营养价值高的互花米草会成为入侵区域底栖生物的主要食物来源, 改变着底栖营养关系(温作柱等, 2014 ). ...
Introduction and spread of mangroves in the Hawaiian Islands.
1
1981
... 红树植物同样可以作为外来种入侵非土著区域, 代表性研究为外来红树植物入侵美国夏威夷群岛的研究(Wester, 1981 ; Allen, 1998 ).佛罗里达州的Rhizophora mangle 被人为引种到夏威夷群岛, 种群爆发扩散, 现已覆盖25%的夏威夷南部海岸(Chimner et al ., 2006 ; D’iorio et al ., 2007 ). ...
福建漳江口红树林和盐沼湿地大型底栖动物次级生产力研究
1
2013
... 互花米草入侵还会改变红树林的生物多样性及其行为模式.在九龙江口红树林区, 互花米草入侵区域底栖动物的栖息密度、种类多样性、丰度比红树林区高, 比涂滩要低(洪荣标等, 2005 ).互花米草的入侵还改变着氨氧化古菌与细菌的丰度, 影响氨氧化微生物的群落结构(Zhang et al ., 2011 ).在漳江口红树林区, 红树林生境与互花米草生境之间多毛类动物栖息密度、生物量、丰度指数、均匀度指数和多样性指数均有显著差异(陈昕韡等, 2012 ).红树林与互花米草生境大型底栖动物次级生产力也存在差异, 海榄雌(Avicennia marina )生境大型底栖动物的年次级生产力最高, 而互花米草生境大型底栖动物的P/B值(P: 平均次级生产力; B: 平均生物量)最高, 因其优势类群为个体较小的寡毛类(吴辰等, 2013 ).再有, 互花米草相对较高的根密度显著降低了红树林中蟹类在互花米草生境打洞的深度及洞穴的复杂性(Wang et al ., 2014 ). ...
福建漳江口红树林和盐沼湿地大型底栖动物次级生产力研究
1
2013
... 互花米草入侵还会改变红树林的生物多样性及其行为模式.在九龙江口红树林区, 互花米草入侵区域底栖动物的栖息密度、种类多样性、丰度比红树林区高, 比涂滩要低(洪荣标等, 2005 ).互花米草的入侵还改变着氨氧化古菌与细菌的丰度, 影响氨氧化微生物的群落结构(Zhang et al ., 2011 ).在漳江口红树林区, 红树林生境与互花米草生境之间多毛类动物栖息密度、生物量、丰度指数、均匀度指数和多样性指数均有显著差异(陈昕韡等, 2012 ).红树林与互花米草生境大型底栖动物次级生产力也存在差异, 海榄雌(Avicennia marina )生境大型底栖动物的年次级生产力最高, 而互花米草生境大型底栖动物的P/B值(P: 平均次级生产力; B: 平均生物量)最高, 因其优势类群为个体较小的寡毛类(吴辰等, 2013 ).再有, 互花米草相对较高的根密度显著降低了红树林中蟹类在互花米草生境打洞的深度及洞穴的复杂性(Wang et al ., 2014 ). ...
入侵生态学最新研究动态
1
2014
... 互花米草有200多年的入侵扩张历史.不同国家和地区对互花米草的入侵现状、入侵效应、入侵原因与机制, 以及相关防治措施都有过全面而系统的研究(Daehler & Strong, 1996 ; Li et al ., 2009 ).现阶段对互花米草入侵的研究, 一方面是从分子水平揭示互花米草成功入侵的遗传学基础、耐受性以及表型可塑性的分子机理或原因(吴昊和丁建清, 2014 ); 另一方面是应用3S技术, 从景观以上尺度监测互花米草入侵的动态变化(Wan et al ., 2014 ).互花米草入侵不可避免地要受全球变化影响, 二者的相互反馈在全球变化日益强烈的今天愈加受到重视(Emery & Fulweiler, 2014 ).随着互花米草在海岸滩涂的快速扩散, 海岸带上所有的生态系统不可避免地会受到互花米草入侵的威胁.互花米草对潮间带生境的高度适应性, 使之在入侵地扩散, 并成功地入侵红树林生态系统(Zhang et al ., 2012 ). ...
入侵生态学最新研究动态
1
2014
... 互花米草有200多年的入侵扩张历史.不同国家和地区对互花米草的入侵现状、入侵效应、入侵原因与机制, 以及相关防治措施都有过全面而系统的研究(Daehler & Strong, 1996 ; Li et al ., 2009 ).现阶段对互花米草入侵的研究, 一方面是从分子水平揭示互花米草成功入侵的遗传学基础、耐受性以及表型可塑性的分子机理或原因(吴昊和丁建清, 2014 ); 另一方面是应用3S技术, 从景观以上尺度监测互花米草入侵的动态变化(Wan et al ., 2014 ).互花米草入侵不可避免地要受全球变化影响, 二者的相互反馈在全球变化日益强烈的今天愈加受到重视(Emery & Fulweiler, 2014 ).随着互花米草在海岸滩涂的快速扩散, 海岸带上所有的生态系统不可避免地会受到互花米草入侵的威胁.互花米草对潮间带生境的高度适应性, 使之在入侵地扩散, 并成功地入侵红树林生态系统(Zhang et al ., 2012 ). ...
The range expansion patterns of Spartina alterniflora on salt marshes in the Yangtze Estuary, China.
1
2010
... 互花米草是禾本科米草属(又名绳草属)多年生草本植物, 原产于美洲大西洋沿岸和墨西哥湾.近200年来, 经不同的入侵途径, 互花米草的分布区域已经从其原产地扩展到欧洲、北美西海岸、新西兰与中国沿海, 被公认为是全球海岸滩涂湿地生态系统最成功的入侵植物之一(王卿等, 2006 ).外界条件与互花米草自身特性相辅相成, 促成了互花米草高效的、难以防治的入侵模式:快速扩张呈现出点源扩散和多点爆发的特征, 对乡土植被的侵占表现为交互生长与斑块推进相结合(Xiao et al ., 2010 ).因此, 互花米草一旦成功入侵, 很难根除和控制. ...
Invasive capacity of the mangrove Sonneratia apetala in Hainan Island, China.
1
2013
... 无瓣海桑在引入地多用作沿海涂滩造林与恢复树种, 具有明显的改良土壤效应, 可增加土壤有机质和N、P、K含量, 有利于后续乡土红树植物定植(韩维栋等, 2003 ).在乡土红树林群落中混种无瓣海桑, 虽然整个林分生物量或土壤碳库没有显著增加, 但无瓣海桑相对较高的凋落物量和低C/N比, 有利于分解者利用(Lu et al ., 2014 ).无瓣海桑的引入能提高群落结构的复杂性与生物多样性.在深圳福田红树林保护区, 混种无瓣海桑+海桑人工林, 相对于天然红树林, 鸟类群落生物多样性得到提高(王勇军和昝启杰, 2001 ).鉴于上述研究结果, 一些学者认为无瓣海桑并没有表现出入侵危害(Zan et al ., 2003 ; 朱宏伟, 2012 ; Xin et al ., 2013 ). ...
无瓣海桑引入对龙海九龙江口乡土红树林影响的调查
1
2010
... 但是也有学者研究发现, 无瓣海桑对其林下乡土红树植物群落会产生不利影响.在无瓣海桑林下, 秋茄树(Kandelia obovata )受到的抑制最小, 可以正常生长; 木榄(Bruguiera gymnorhiza )、海桑、桐花树(Aegiceras corniculatum )受到的抑制次之; 红海兰(Rhizophora stylosa )的保存率最低(李玫等, 2004a ; 梁士楚等, 2005 ; 王旭等, 2008 ; 曾雯珺等, 2008 ; 姚历强, 2010 ).相对于乡土红树植物, 无瓣海桑具有优异的生物学形状——种子幼苗品质好, 抗逆性强(廖宝文等, 2004 ), 生物量大(Chen et al ., 2013b ), 生态位与乡土红树植物有显著重叠(Li et al ., 2011 ), 有一定的扩散能力以及对乡土红树植物的化感作用(李玫等, 2004b ), 以后有可能破坏甚至取代乡土红树植物群落(Ren et al ., 2008 ).因此, 现阶段研究更多建议决策者谨慎考虑无瓣海桑在红树林保护区引种种植. ...
无瓣海桑引入对龙海九龙江口乡土红树林影响的调查
1
2010
... 但是也有学者研究发现, 无瓣海桑对其林下乡土红树植物群落会产生不利影响.在无瓣海桑林下, 秋茄树(Kandelia obovata )受到的抑制最小, 可以正常生长; 木榄(Bruguiera gymnorhiza )、海桑、桐花树(Aegiceras corniculatum )受到的抑制次之; 红海兰(Rhizophora stylosa )的保存率最低(李玫等, 2004a ; 梁士楚等, 2005 ; 王旭等, 2008 ; 曾雯珺等, 2008 ; 姚历强, 2010 ).相对于乡土红树植物, 无瓣海桑具有优异的生物学形状——种子幼苗品质好, 抗逆性强(廖宝文等, 2004 ), 生物量大(Chen et al ., 2013b ), 生态位与乡土红树植物有显著重叠(Li et al ., 2011 ), 有一定的扩散能力以及对乡土红树植物的化感作用(李玫等, 2004b ), 以后有可能破坏甚至取代乡土红树植物群落(Ren et al ., 2008 ).因此, 现阶段研究更多建议决策者谨慎考虑无瓣海桑在红树林保护区引种种植. ...
薇甘菊生态防除研究初报
1
2003
... 很多研究证实寄生植物野地菟丝子能严重影响微甘菊茎叶生长及开花结实, 但不影响其他植物正常的生长繁殖, 用菟丝子控制微甘菊安全可行(昝启杰等, 2003 ).在入侵地发掘一些微甘菊的天敌, 像昆虫、微生物等, 也能控制微甘菊(邵华等, 2002 ).通过植物搭配种植, 种植当地适生的乔灌草植物或促进野生植物生长, 建立稳定的复层群落结构, 也可持续有效地控制微甘菊(殷祚云等, 2003 ). ...
薇甘菊生态防除研究初报
1
2003
... 很多研究证实寄生植物野地菟丝子能严重影响微甘菊茎叶生长及开花结实, 但不影响其他植物正常的生长繁殖, 用菟丝子控制微甘菊安全可行(昝启杰等, 2003 ).在入侵地发掘一些微甘菊的天敌, 像昆虫、微生物等, 也能控制微甘菊(邵华等, 2002 ).通过植物搭配种植, 种植当地适生的乔灌草植物或促进野生植物生长, 建立稳定的复层群落结构, 也可持续有效地控制微甘菊(殷祚云等, 2003 ). ...
薇甘菊在深圳湾的入侵路线及其生态特征
1
2011
... 微甘菊在中国华南沿海的另一个重灾区是深圳湾(Ren et al ., 2014 ), 拦海造田使海水退去, 红树林底泥裸露, 微甘菊种子得以萌发生长, 严重危害红树林(于晓梅和杨逢建, 2011 ).微甘菊入侵深圳湾红树林后, 一方面导致红树植物枯萎, 减少了植被生物量; 另一方面增加了凋落物量, 促进了土壤微生物活动, 加速了土壤有机质分解释放, 降低了土壤碳储量, 削弱了红树林碳汇(毛子龙等, 2011 ).微甘菊的入侵, 还会影响人工恢复红树林的季节动态, 降低红树植物的成活率, 妨害红树林生态恢复(苗春玲等, 2012 ). ...
薇甘菊在深圳湾的入侵路线及其生态特征
1
2011
... 微甘菊在中国华南沿海的另一个重灾区是深圳湾(Ren et al ., 2014 ), 拦海造田使海水退去, 红树林底泥裸露, 微甘菊种子得以萌发生长, 严重危害红树林(于晓梅和杨逢建, 2011 ).微甘菊入侵深圳湾红树林后, 一方面导致红树植物枯萎, 减少了植被生物量; 另一方面增加了凋落物量, 促进了土壤微生物活动, 加速了土壤有机质分解释放, 降低了土壤碳储量, 削弱了红树林碳汇(毛子龙等, 2011 ).微甘菊的入侵, 还会影响人工恢复红树林的季节动态, 降低红树植物的成活率, 妨害红树林生态恢复(苗春玲等, 2012 ). ...
刈割与水位调节集成技术控制互花米草(Spartina alterniflora)
1
2008
... 持续的淡咸水轮换浇灌, 互花米草的生物量积累、无性与有性繁殖能力都会受到抑制(陈琳等, 2007 ).而且选择关键季节, 刈割互花米草地上部分, 同时配合一定水位的持续淹水, 更能有效地发挥治理互花米草的效果(袁琳等, 2008 ).有些物理控制措施, 如遮阴或冬季火烧, 对互花米草影响不大, 反而会影响乡土植物生长, 促进互花米草的入侵, 应当避免 (陈中义等, 2005 ; 王智晨等, 2006 ). ...
刈割与水位调节集成技术控制互花米草(Spartina alterniflora)
1
2008
... 持续的淡咸水轮换浇灌, 互花米草的生物量积累、无性与有性繁殖能力都会受到抑制(陈琳等, 2007 ).而且选择关键季节, 刈割互花米草地上部分, 同时配合一定水位的持续淹水, 更能有效地发挥治理互花米草的效果(袁琳等, 2008 ).有些物理控制措施, 如遮阴或冬季火烧, 对互花米草影响不大, 反而会影响乡土植物生长, 促进互花米草的入侵, 应当避免 (陈中义等, 2005 ; 王智晨等, 2006 ). ...
森草净杀灭薇甘菊(Mikania micrantha)及其安全性
3
2007
... 化学控制是利用化学药剂对入侵种进行灭杀, 以达到控制甚至消灭入侵种的目的.相对于物理控制, 化学控制效果更明显, 而且节省人力物力, 是控制外来有害物种的主要方式(Mack et al ., 2000 ).但是, 化学药剂缺乏目标特异性, 在毒杀目标物种的同时, 也毒害了非目标物种, 使用前必须掌握所用化学药剂的安全性(昝启杰等, 2007 ). ...
... 对比几种不同的除草剂对微甘菊的灭杀效果及其对其他生物的安全性, 证实一种名为“森草净”的除草剂对红树林区的微甘菊有极好的杀灭效果, 且对施用区域的生物多样性无不良影响, 被认为是微甘菊化学控制的首选除草剂(昝启杰等, 2007 ). ...
... 很多研究证实寄生植物野地菟丝子能严重影响微甘菊茎叶生长及开花结实, 但不影响其他植物正常的生长繁殖, 用菟丝子控制微甘菊安全可行(昝启杰等, 2003 ).在入侵地发掘一些微甘菊的天敌, 像昆虫、微生物等, 也能控制微甘菊(邵华等, 2002 ).通过植物搭配种植, 种植当地适生的乔灌草植物或促进野生植物生长, 建立稳定的复层群落结构, 也可持续有效地控制微甘菊(殷祚云等, 2003 ). ...
森草净杀灭薇甘菊(Mikania micrantha)及其安全性
3
2007
... 化学控制是利用化学药剂对入侵种进行灭杀, 以达到控制甚至消灭入侵种的目的.相对于物理控制, 化学控制效果更明显, 而且节省人力物力, 是控制外来有害物种的主要方式(Mack et al ., 2000 ).但是, 化学药剂缺乏目标特异性, 在毒杀目标物种的同时, 也毒害了非目标物种, 使用前必须掌握所用化学药剂的安全性(昝启杰等, 2007 ). ...
... 对比几种不同的除草剂对微甘菊的灭杀效果及其对其他生物的安全性, 证实一种名为“森草净”的除草剂对红树林区的微甘菊有极好的杀灭效果, 且对施用区域的生物多样性无不良影响, 被认为是微甘菊化学控制的首选除草剂(昝启杰等, 2007 ). ...
... 很多研究证实寄生植物野地菟丝子能严重影响微甘菊茎叶生长及开花结实, 但不影响其他植物正常的生长繁殖, 用菟丝子控制微甘菊安全可行(昝启杰等, 2003 ).在入侵地发掘一些微甘菊的天敌, 像昆虫、微生物等, 也能控制微甘菊(邵华等, 2002 ).通过植物搭配种植, 种植当地适生的乔灌草植物或促进野生植物生长, 建立稳定的复层群落结构, 也可持续有效地控制微甘菊(殷祚云等, 2003 ). ...
Ecological assessment on the introduced Sonneratia caseolaris and S. apetala at the mangrove forest of Shenzhen Bay, China.
1
2003
... 无瓣海桑在引入地多用作沿海涂滩造林与恢复树种, 具有明显的改良土壤效应, 可增加土壤有机质和N、P、K含量, 有利于后续乡土红树植物定植(韩维栋等, 2003 ).在乡土红树林群落中混种无瓣海桑, 虽然整个林分生物量或土壤碳库没有显著增加, 但无瓣海桑相对较高的凋落物量和低C/N比, 有利于分解者利用(Lu et al ., 2014 ).无瓣海桑的引入能提高群落结构的复杂性与生物多样性.在深圳福田红树林保护区, 混种无瓣海桑+海桑人工林, 相对于天然红树林, 鸟类群落生物多样性得到提高(王勇军和昝启杰, 2001 ).鉴于上述研究结果, 一些学者认为无瓣海桑并没有表现出入侵危害(Zan et al ., 2003 ; 朱宏伟, 2012 ; Xin et al ., 2013 ). ...
无瓣海桑与三种乡土红树植物混交的生态效应
1
2008
... 但是也有学者研究发现, 无瓣海桑对其林下乡土红树植物群落会产生不利影响.在无瓣海桑林下, 秋茄树(Kandelia obovata )受到的抑制最小, 可以正常生长; 木榄(Bruguiera gymnorhiza )、海桑、桐花树(Aegiceras corniculatum )受到的抑制次之; 红海兰(Rhizophora stylosa )的保存率最低(李玫等, 2004a ; 梁士楚等, 2005 ; 王旭等, 2008 ; 曾雯珺等, 2008 ; 姚历强, 2010 ).相对于乡土红树植物, 无瓣海桑具有优异的生物学形状——种子幼苗品质好, 抗逆性强(廖宝文等, 2004 ), 生物量大(Chen et al ., 2013b ), 生态位与乡土红树植物有显著重叠(Li et al ., 2011 ), 有一定的扩散能力以及对乡土红树植物的化感作用(李玫等, 2004b ), 以后有可能破坏甚至取代乡土红树植物群落(Ren et al ., 2008 ).因此, 现阶段研究更多建议决策者谨慎考虑无瓣海桑在红树林保护区引种种植. ...
无瓣海桑与三种乡土红树植物混交的生态效应
1
2008
... 但是也有学者研究发现, 无瓣海桑对其林下乡土红树植物群落会产生不利影响.在无瓣海桑林下, 秋茄树(Kandelia obovata )受到的抑制最小, 可以正常生长; 木榄(Bruguiera gymnorhiza )、海桑、桐花树(Aegiceras corniculatum )受到的抑制次之; 红海兰(Rhizophora stylosa )的保存率最低(李玫等, 2004a ; 梁士楚等, 2005 ; 王旭等, 2008 ; 曾雯珺等, 2008 ; 姚历强, 2010 ).相对于乡土红树植物, 无瓣海桑具有优异的生物学形状——种子幼苗品质好, 抗逆性强(廖宝文等, 2004 ), 生物量大(Chen et al ., 2013b ), 生态位与乡土红树植物有显著重叠(Li et al ., 2011 ), 有一定的扩散能力以及对乡土红树植物的化感作用(李玫等, 2004b ), 以后有可能破坏甚至取代乡土红树植物群落(Ren et al ., 2008 ).因此, 现阶段研究更多建议决策者谨慎考虑无瓣海桑在红树林保护区引种种植. ...
Mikania micrantha H. B. K. in China―An overview.
3
2004
... 微甘菊是菊科假泽兰属多年生草质藤本, 原产于热带南美与中美洲.微甘菊生长迅速且适应性强, 英文称其为“Mile-a-Minute Weed” (一分钟一英里杂草), 极具入侵性(Waterhouse, 1994 ).微甘菊在原产地因有天敌控制不具危害, 但在缺乏天敌的入侵地迅速发展为危害性严重的有害杂草(昝启杰等, 2000 ).微甘菊因其独特的生物学特性及生态适应能力(Tripathi et al ., 2011 ), 传入新生境后能迅速爆发.如今, 微甘菊已在整个亚洲热带及北美地区肆虐, 成为著名的世界性恶草(Zhang et al ., 2004 ). ...
... 微甘菊入侵后, 通常给当地生态系统带来了毁灭性灾害, 因此对微甘菊的防治需求十分紧迫(Waterhouse, 1994 ).物理和化学防治手段的弊端促使学者寻求生物手段来防治微甘菊入侵.业已证实, 寄生植物野地菟丝子(Cuscuta campestris )能安全有效地防治微甘菊(Lian et al ., 2006 ; Shen et al ., 2011 ), 一些新的微生物或病毒也能用来控制微甘菊(Ellison et al ., 2008 ; Wang et al ., 2008 ).除关注微甘菊对生态系统地上植物群落的危害外, 对生态系统其他结构(如地下生物)和功能的效应的关注也逐步形成(Li et al ., 2006 , 2007 ).微甘菊入侵机制的研究日渐成熟(Zhang et al ., 2004 ; Chen et al ., 2009 ).有关微甘菊入侵的分子手段研究(Huang et al ., 2012 ; Wang et al ., 2012 ), 以及微甘菊对全球气候变化的响应研究(Wang et al ., 2010 )初现端倪.随着微甘菊的持续入侵扩散, 温暖湿润沿海地区的红树林生态系统也难逃其入侵危害(Han et al ., 2006 ). ...
... 现有的微甘菊入侵研究主要集中于陆地森林生态系统(Zhang et al ., 2004 ), 红树林同类研究比较少, 而且都集中于中国华南沿海红树林(Ren et al ., 2014 ).微甘菊进入红树林后, 迅速生长, 在红树植物上形成一层茂密的覆盖层, 强烈争夺资源(尤其是光照)及通过化感作用抑制或杀死红树植物(Maja et al ., 2008 ).张阳和张华(2008)研究过珠海淇澳岛红树林中微甘菊的分布模式与危害, 强调自然条件和人为因素在影响红树林中微甘菊的分布上起着重要作用; 推测当地微甘菊爆发主要是由于人类不适当的开发活动破坏了当地红树林. ...
Impacts of Spartina alterniflora invasion on abundance and composition of ammonia oxidizers in estuarine sediment.
1
2011
... 互花米草入侵还会改变红树林的生物多样性及其行为模式.在九龙江口红树林区, 互花米草入侵区域底栖动物的栖息密度、种类多样性、丰度比红树林区高, 比涂滩要低(洪荣标等, 2005 ).互花米草的入侵还改变着氨氧化古菌与细菌的丰度, 影响氨氧化微生物的群落结构(Zhang et al ., 2011 ).在漳江口红树林区, 红树林生境与互花米草生境之间多毛类动物栖息密度、生物量、丰度指数、均匀度指数和多样性指数均有显著差异(陈昕韡等, 2012 ).红树林与互花米草生境大型底栖动物次级生产力也存在差异, 海榄雌(Avicennia marina )生境大型底栖动物的年次级生产力最高, 而互花米草生境大型底栖动物的P/B值(P: 平均次级生产力; B: 平均生物量)最高, 因其优势类群为个体较小的寡毛类(吴辰等, 2013 ).再有, 互花米草相对较高的根密度显著降低了红树林中蟹类在互花米草生境打洞的深度及洞穴的复杂性(Wang et al ., 2014 ). ...
红树林生长带与潮汐水位关系的研究
1
1997
... 红树林是指分布在热带、亚热带海岸潮间带, 受海水周期性浸淹的木本植物群落; 上述定义强调红树林分布的非地带性以及与潮汐的关系(Duke, 1993 ; 张乔民等, 1997 ).红树林生态系统生境严苛, 高盐、高温、强风, 以及因受潮汐浸淹而底泥缺氧, 只有少数几种进化出高度适应特征的植物能生存(Kathiresan & Bingham, 2001 ).独特的红树林生态系统, 其生物入侵的表现既与其他生态系统有共同之处, 又有其自身特异性. ...
红树林生长带与潮汐水位关系的研究
1
1997
... 红树林是指分布在热带、亚热带海岸潮间带, 受海水周期性浸淹的木本植物群落; 上述定义强调红树林分布的非地带性以及与潮汐的关系(Duke, 1993 ; 张乔民等, 1997 ).红树林生态系统生境严苛, 高盐、高温、强风, 以及因受潮汐浸淹而底泥缺氧, 只有少数几种进化出高度适应特征的植物能生存(Kathiresan & Bingham, 2001 ).独特的红树林生态系统, 其生物入侵的表现既与其他生态系统有共同之处, 又有其自身特异性. ...
互花米草入侵下福建漳江口红树林湿地土壤生态化学变化
1
2008
... 互花米草已经出现在中国所有的红树林区.对福建境内红树林互花米草入侵的研究走在国内前列.互花米草入侵导致红树林生境土壤退化, 漳江口互花米草生境的土壤有机碳、土壤微生物生物量碳、氮含量以及土壤蔗糖酶与磷酸酶活性等相对于红树林生境均有明显下降(张祥霖等, 2008 ).但是, 互花米草群落对碳、氮具有很强的富集能力, 相对于红树林对碳、氮弱的富集能力, 互花米草的存在能更好地拦截和吸附陆源污染物, 并使重金属富集在互花米草群落沉积物中, 增加泉州湾红树林污染富集承受能力(王爱军等, 2008 ). ...
互花米草入侵下福建漳江口红树林湿地土壤生态化学变化
1
2008
... 互花米草已经出现在中国所有的红树林区.对福建境内红树林互花米草入侵的研究走在国内前列.互花米草入侵导致红树林生境土壤退化, 漳江口互花米草生境的土壤有机碳、土壤微生物生物量碳、氮含量以及土壤蔗糖酶与磷酸酶活性等相对于红树林生境均有明显下降(张祥霖等, 2008 ).但是, 互花米草群落对碳、氮具有很强的富集能力, 相对于红树林对碳、氮弱的富集能力, 互花米草的存在能更好地拦截和吸附陆源污染物, 并使重金属富集在互花米草群落沉积物中, 增加泉州湾红树林污染富集承受能力(王爱军等, 2008 ). ...
Interactions between mangroves and exotic Spartina in an anthropogenically disturbed estuary in southern China.
1
2012
... 互花米草有200多年的入侵扩张历史.不同国家和地区对互花米草的入侵现状、入侵效应、入侵原因与机制, 以及相关防治措施都有过全面而系统的研究(Daehler & Strong, 1996 ; Li et al ., 2009 ).现阶段对互花米草入侵的研究, 一方面是从分子水平揭示互花米草成功入侵的遗传学基础、耐受性以及表型可塑性的分子机理或原因(吴昊和丁建清, 2014 ); 另一方面是应用3S技术, 从景观以上尺度监测互花米草入侵的动态变化(Wan et al ., 2014 ).互花米草入侵不可避免地要受全球变化影响, 二者的相互反馈在全球变化日益强烈的今天愈加受到重视(Emery & Fulweiler, 2014 ).随着互花米草在海岸滩涂的快速扩散, 海岸带上所有的生态系统不可避免地会受到互花米草入侵的威胁.互花米草对潮间带生境的高度适应性, 使之在入侵地扩散, 并成功地入侵红树林生态系统(Zhang et al ., 2012 ). ...
珠海淇澳岛红树林湿地薇甘菊危害情况和发生因素研究
0
2008
珠海淇澳岛红树林湿地薇甘菊危害情况和发生因素研究
0
2008
我国红树林外来入侵现状研究综述
2
2010
... 在植物入侵红树林研究文献中, 无瓣海桑(Sonneratia apetala )因其入侵性存在极大的争议, 相关研究报道最多, 有24篇, 紧随其后的为互花米草(Spartina alterniflora )(17篇)、微甘菊(Mikania micrantha )(14篇)和Rhizophora mangle (11篇).无瓣海桑、互花米草入侵红树林的研究区域为中国华南和东南沿海.微甘菊入侵红树林研究除1篇在美国东海岸外, 其余都在中国华南和东南沿海.Rhizophora mangle 入侵研究则仅限于夏威夷群岛.外来植物Nypa fruticans 由人类活动引入尼日利亚红树林, 繁殖爆发, 对当地渔业生产造成危害(Isebor et al ., 2003 ).外来植物Schinus terebinthifolius 入侵美国佛罗里达州的红树林, 通过化感作用抑制了乡土红树植物幼苗的生长(Donnelly et al ., 2008 ).外来种海桑(Sonneratia caseolaris )在深圳福田红树林保护区引种种植后, 提高了泥滩大型底栖动物的多样性(Leung & Tam, 2013 ).此外, 红树林中还存在许多其他外来植物, 但都仅止于调查记录(Biswas et al ., 2007 ; 周青青等, 2010 ). ...
... 微甘菊是菊科假泽兰属多年生草质藤本, 原产于热带南美与中美洲.微甘菊生长迅速且适应性强, 英文称其为“Mile-a-Minute Weed” (一分钟一英里杂草), 极具入侵性(Waterhouse, 1994 ).微甘菊在原产地因有天敌控制不具危害, 但在缺乏天敌的入侵地迅速发展为危害性严重的有害杂草(昝启杰等, 2000 ).微甘菊因其独特的生物学特性及生态适应能力(Tripathi et al ., 2011 ), 传入新生境后能迅速爆发.如今, 微甘菊已在整个亚洲热带及北美地区肆虐, 成为著名的世界性恶草(Zhang et al ., 2004 ). ...
我国红树林外来入侵现状研究综述
2
2010
... 在植物入侵红树林研究文献中, 无瓣海桑(Sonneratia apetala )因其入侵性存在极大的争议, 相关研究报道最多, 有24篇, 紧随其后的为互花米草(Spartina alterniflora )(17篇)、微甘菊(Mikania micrantha )(14篇)和Rhizophora mangle (11篇).无瓣海桑、互花米草入侵红树林的研究区域为中国华南和东南沿海.微甘菊入侵红树林研究除1篇在美国东海岸外, 其余都在中国华南和东南沿海.Rhizophora mangle 入侵研究则仅限于夏威夷群岛.外来植物Nypa fruticans 由人类活动引入尼日利亚红树林, 繁殖爆发, 对当地渔业生产造成危害(Isebor et al ., 2003 ).外来植物Schinus terebinthifolius 入侵美国佛罗里达州的红树林, 通过化感作用抑制了乡土红树植物幼苗的生长(Donnelly et al ., 2008 ).外来种海桑(Sonneratia caseolaris )在深圳福田红树林保护区引种种植后, 提高了泥滩大型底栖动物的多样性(Leung & Tam, 2013 ).此外, 红树林中还存在许多其他外来植物, 但都仅止于调查记录(Biswas et al ., 2007 ; 周青青等, 2010 ). ...
... 微甘菊是菊科假泽兰属多年生草质藤本, 原产于热带南美与中美洲.微甘菊生长迅速且适应性强, 英文称其为“Mile-a-Minute Weed” (一分钟一英里杂草), 极具入侵性(Waterhouse, 1994 ).微甘菊在原产地因有天敌控制不具危害, 但在缺乏天敌的入侵地迅速发展为危害性严重的有害杂草(昝启杰等, 2000 ).微甘菊因其独特的生物学特性及生态适应能力(Tripathi et al ., 2011 ), 传入新生境后能迅速爆发.如今, 微甘菊已在整个亚洲热带及北美地区肆虐, 成为著名的世界性恶草(Zhang et al ., 2004 ). ...
珠海淇澳岛无瓣海桑入侵风险综合评估
1
2012
... 无瓣海桑在引入地多用作沿海涂滩造林与恢复树种, 具有明显的改良土壤效应, 可增加土壤有机质和N、P、K含量, 有利于后续乡土红树植物定植(韩维栋等, 2003 ).在乡土红树林群落中混种无瓣海桑, 虽然整个林分生物量或土壤碳库没有显著增加, 但无瓣海桑相对较高的凋落物量和低C/N比, 有利于分解者利用(Lu et al ., 2014 ).无瓣海桑的引入能提高群落结构的复杂性与生物多样性.在深圳福田红树林保护区, 混种无瓣海桑+海桑人工林, 相对于天然红树林, 鸟类群落生物多样性得到提高(王勇军和昝启杰, 2001 ).鉴于上述研究结果, 一些学者认为无瓣海桑并没有表现出入侵危害(Zan et al ., 2003 ; 朱宏伟, 2012 ; Xin et al ., 2013 ). ...
珠海淇澳岛无瓣海桑入侵风险综合评估
1
2012
... 无瓣海桑在引入地多用作沿海涂滩造林与恢复树种, 具有明显的改良土壤效应, 可增加土壤有机质和N、P、K含量, 有利于后续乡土红树植物定植(韩维栋等, 2003 ).在乡土红树林群落中混种无瓣海桑, 虽然整个林分生物量或土壤碳库没有显著增加, 但无瓣海桑相对较高的凋落物量和低C/N比, 有利于分解者利用(Lu et al ., 2014 ).无瓣海桑的引入能提高群落结构的复杂性与生物多样性.在深圳福田红树林保护区, 混种无瓣海桑+海桑人工林, 相对于天然红树林, 鸟类群落生物多样性得到提高(王勇军和昝启杰, 2001 ).鉴于上述研究结果, 一些学者认为无瓣海桑并没有表现出入侵危害(Zan et al ., 2003 ; 朱宏伟, 2012 ; Xin et al ., 2013 ). ...



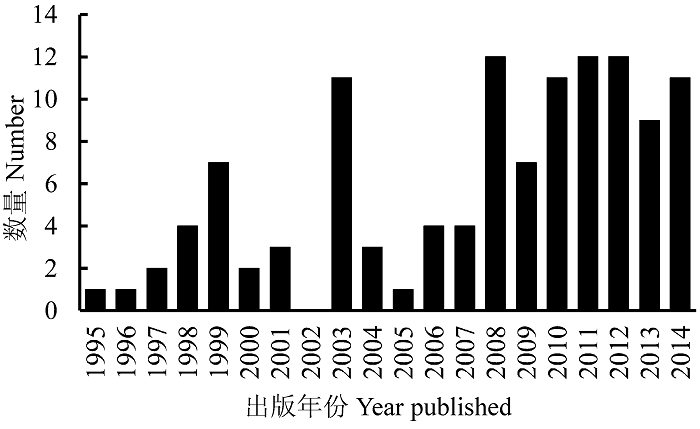
{kind=link}
{kind=link}
{kind=link}
{kind=link}