植物光合生理特征能体现植物对不同生境的适应, 反映植物对不同环境变化的响应(Jiang & He, 1999 )。不同物种的光合生理特征间有显著的差异(Bassow & Bazzaz, 1997 ), 这正是不同植物利用不同的生态策略获得资源利用的反映(Niu et al ., 2003 )。一般而言, C4 光合途径或具有固氮能力的物种比C3 途径和无固氮能力的植物具备较高的光合能力和光合氮利用效率(Jiang & He, 1999 ), 因为氮含量与植物光合能力有非常密切的联系(Evans, 1989 )。双子叶植物、非禾本科植物、豆科植物和中生型植物的叶片氮含量分别高于相应的单子叶植物、禾本科植物、非豆科植物和旱生型植物(张文彦等, 2010 ), 草本植物比乔木和灌丛具有更高的净光合速率和光合氮利用效率(Wright et al ., 2005 )。Wright等(2005)在对全球植物叶片普遍特征的关系研究中还发现, 一般而言, 草本植物比灌木具有更低的基于面积的氮含量和比叶质量。另外, 植物光合生理特征也会随着叶片生活史长短(Hikosaka & Hirose, 2000 )和植株的大小而变化(Liu et al ., 2010 )。
影响植物光合能力的环境因子包括光照、水分、温度、营养物质和CO2 浓度等(于贵瑞和王秋凤, 2010 ), 而水分和氮是植物光合和生长必不可少的因子(Novriyanti et al ., 2012 ), Shangguan等(2000) 研究发现, 水分和氮素亏缺均会强烈地抑制植物的光合活动, 且氮素和水分对光合的影响是相互制约的。干旱大大降低了单叶和群落的净光合速率(崔骁勇等, 2000 ), 而提高了植物的光合水分利用效率(严昌荣等, 2001 )。An和Shangguan (2008)对不同光照和不同浓度氮素下白车轴草(Trifolium repens )幼苗光合生理特征的研究发现, 低浓度氮下基于面积的CO2 同化速率(A area )、光合氮利用效率(PNUE )和叶绿素含量随着氮素浓度的升高而增加; 高浓度氮下, 这些光合特征值会下降, 而比叶面积(SLA )和基于质量的叶片氮含量(N mass )会随着氮素浓度的升高持续增加。另外, 海拔通过影响CO2 分压、温度、水分等环境因子而影响植物光合生理特征, Ran等(2013) 将岷江冷杉(Abies faxoniana )移植到低海拔处, 降低了岷江冷杉的叶氮含量和水分利用效率, 而增加了净光合速率和PNUE 。
高寒草甸是青藏高原最主要的生态系统类型(周华坤等, 2008 ), 由于长期忽略了对草地资源的科学管理, 粗放经营、超载放牧以及对草地资源不合理的开发利用, 使得高寒草地在结构上沙化, 功能上生产力降低, 生物多样性和生态系统功能严重丧失(武高林和杜国祯, 2007 )。关于高寒草甸恢复演替的研究已有很多, 放牧干扰及种间竞争对草地群落演替具有最大的影响(孙海群等, 1999 )。围封消除了放牧干扰, 围封地植物光合生理特征可以很好地反映退化草地自然恢复过程各阶段群落光合动态及其对环境因子的响应。尽管植物光合生理特征对环境变化的响应在国内外有大量详细的研究, 但由于技术设备等原因的限制, 高寒草甸植物光合作用对环境适应机理的研究依然较少, 尤其缺乏从物种到群落水平上的研究, 而这方面的研究对于植物适应日益剧增的放牧、围栏封育等土地利用方式极其重要。本文在判定亚高山草甸围封地各群落演替阶段的基础上, 通过对植物光合生理特征和表层土壤养分的探讨, 掌握了围封地恢复演替不同阶段的光合生理特征动态及其主要驱动因子, 对亚高寒退化草甸围封恢复具有一定的理论指导意义。
1 材料和方法
1.1 研究地自然概况
实验地是一块围封多年的草地, 位于甘肃省甘南藏族自治州合作市附近的兰州大学高寒草甸与湿地生态系统定位研究站, 地理位置34.93° N, 102.88° E, 海拔2900-3000 m。年平均气温2.0 ℃, 最冷月(12-2月)平均气温-8.9 ℃, 最热月(6-8月)平均气温11.5 ℃, 0 ℃以上年积温1730 ℃。年降水量560 mm, 50%以上集中在6-8月份, 雨热同季。土壤类型为亚高山草甸土, 以密丛型禾本科和莎草科植物为主, 混以其他杂草类(张大勇等, 1988 )。
1.2 研究方法
通过试验地调查, 在围封地内选取5个优势种不同的群落样地, 分别记为A、B、C、D、E。它们分别是以多年生草本植物为优势种的刺儿菜(Cirsium setosum )-甘青蒿(Artemisia tangutica )群落(A)、披碱草(Elymus dahuricus )-刺儿菜群落(B)、紫苜蓿(Medicago sativa )-甘青蒿群落(C)和以半灌木和灌木为优势种的沙蒿(Artemisia desertorum )-羊茅(Festuca ovina )群落(D)和金露梅(Potentilla fruticosa )群落(E)。在每个群落样带中随机选取8个0.5 m × 0.5 m的样方, 对样方信息进行调查, 记录样方内物种及其多度、代表物种高度, 估测各物种盖度及样地盖度。根据各群落物种丰富度、多样性指数及植物生活型组成判断5个群落的演替序列。对各群落优势种及主要物种的光合生理特征值及相应的叶片氮含量进行测定, 同时分别对各群落样地表层(0-20 cm)土壤含水量及全氮含量进行测定。
1.3 光合特征值测定
2013年7月下旬到8月上旬, 选择晴朗天气, 利用便携式光合作用测定仪(LI-6400, LI-COR, Lincoln, USA), 于9:30-11:30测定植物叶片的气体交换参数。测定时, 选用开放气路, 空气流速为500 cm3 ·min-1 , 在外界CO2 浓度下(大约为350 μmol·mol-1 ), 用红蓝光源提供1000 μmol·m-2 ·s-1 光强进行测定, 外界温度在20 ℃左右。选取各群落的优势种及主要物种(尽量是各群落共有种)进行测定, 每个群落测3组, 每一组相同物种3-5株, 选取植株中端展开的成熟健康叶片进行测定, 待数据稳定后读取数据。野外实验可以直接测得基于叶面积的CO2 同化速率(A area )、气孔导度(G s )和蒸腾速率(T r ), 而光合氮利用效率(PNUE )用A area 跟基于面积的叶片氮含量(N area )的比率表示, 光合水分利用效率(WUE )用A area /T r 表示。将测定叶片装入信封带回实验室进行叶面积扫描及氮素测定。
1.4 叶绿素SPAD 值测定
使用便携式叶绿素仪(SPAD-502, Minolta Camera, Osaka, Japan)测定物种叶片的SPAD 值, 选择植株中端成熟健康的叶片进行测定, 测定物种及重复数与测定光合特征值时的一致。SPAD-502是在比较650 nm和940 nm波长光线投射率的基础上测定的(Manetas et al ., 1998 ), 所测叶绿素含量是一个相对值。
1.5 比叶面积测定
将测定光合作用的叶片带到实验室, 用扫描仪扫描叶面积图片, 利用ImageJ软件算出扫描的叶面积, 将这些叶片装入信封, 在75 ℃条件下烘干48 h至恒重, 用精度为1/10000 g的电子秤称量, 比叶面积(SLA ) =叶面积/叶片干质量。
1.6 叶片全氮含量测定
将烘干叶片研磨过筛, 称取一定量的叶片样品于消煮管中, 利用H2 SO4 -HClO4 消煮法对样品进行处理, 然后在化学分析仪(Smartchem 200, WestCo Scientific Instruments, Brookfield, USA)上测定叶片全氮含量。
1.7 土壤水分及土壤全氮测定
将土壤样品一部分在105 ℃下烘干测量其土壤含水量, 另一部分自然风干后研磨、过筛, 用半微量凯氏定氮法测定土壤全氮含量。
1.8 数据处理
不同物种及不同群落间的差异利用SPSS 19.0进行one-way ANOVA差异性检验, 利用SPSS 19.0对不同物种及不同群落中共有种的光合生理特征做主成分分析(PCA), 并在Origin 8.0中作图。
2 结果和分析
2.1 植物群落组成
5个群落中共调查到25科59属90种物种, 其中A群落15科28属32种, B群落14科26属28种, C群落18科39属57种, D群落19科41属57种, E群落23科51属75种。除沙蒿-羊茅群落(D)坡度较大外, 其他群落所处的坡度、坡向没有明显的差异。
A群落是由刺儿菜为优势种的多年生杂草群落; B群落是由禾本科披碱草为优势种并伴生多年生杂类草的群落; C群落豆科植物重要值及种数有了明显的升高, 除紫苜蓿、少花米口袋(Gueldenstaedtia verna )外出现了豆科植物野豌豆(Vicia sepium )、斜茎黄耆(Astragalus adsurgens ), 且从C群落开始物种丰富度及多样性指数大幅度增加, 分别从20、3.55增加到32、4.00 (表1 ); D群落的优势种沙蒿属于半灌木, 并有少量的灌木金露梅出现, 群落生活型有所增加; 而E群落优势种为灌木金露梅。丰富度及多样性指数从小到大可以将群落排列为B < A < C < D < E (表1 )。A、B群落间差异不大, 一般而言, 杂生类草本植物比禾本科植物先侵入群落成为优势种, 因此A群落应该在B群落的演替前期。综合上面分析, 可以将5个群落演替进程表示为A→B→C→D→E, 形成一个以草本植物→半灌木→灌木的演替序列, 其中A→C杂生草本植物所占比例有所下降, 禾本科跟豆科植物重要值逐渐增加。
2.2 不同演替阶段优势种的光合生理特征
不同演替阶段群落优势种的光合生理特征间存在显著差异(表2 )。A area 、PNUE 、WUE 的最大值分别是最小值的4.1倍(17.41 vs 4.29 μmol CO2 ·m-2 ·
s-1 )、6.0倍(362.17 vs 60.3 μmol CO2 ·mol-1 ·s-1 )和2.6倍(3.88 vs 1.51 μmol CO2 ·mmol-1 H2 O)。
披碱草具有最大的A area , 为17.41 μmol CO2 ·m-2 ·s-1 , 其他植物的A area 依次是紫苜蓿>老鹳草>刺儿菜>少花米口袋>甘青蒿>沙蒿>圆穗蓼, 灌丛植物金露梅的A area 最小, 为4.29 μmol CO2 ·m-2 ·s-1 。豆科植物紫苜蓿具有最大的SPAD 值, 其他依次为刺儿菜>少花米口袋>披碱草>圆穗蓼>沙蒿>甘青蒿>金露梅>老鹳草; 紫苜蓿、少花米口袋因自身具有的固氮能力具有较大的N mass , 其他依次为甘青蒿>金露梅>刺儿菜>沙蒿>老鹳草>圆穗蓼, 禾本科披碱草的N mass 最小; 甘青蒿、紫苜蓿具有最大的SLA, 其他依次为沙蒿>金露梅>少花米口袋>圆穗蓼>披碱草>老鹳草, 刺儿菜的SLA 值最小(表2 )。
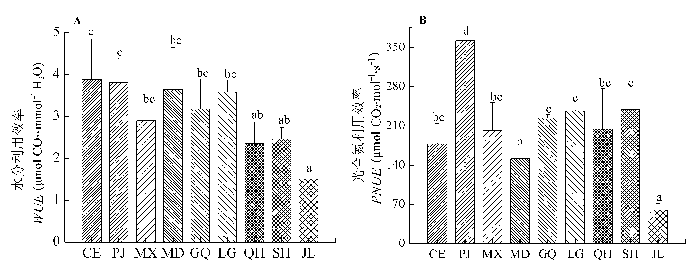
演替前期的刺儿菜和后期的金露梅分别具有最大和最小的WUE, 具有较大A area 的紫苜蓿表现出低的WUE, 这可能是其具有较大的蒸腾速率, 光合能力与水分利用效率权衡的结果, 其他依次为披碱草>少花米口袋>老鹳草>甘青蒿>沙蒿>圆穗蓼(图1A )。披碱 草具有最大的PNUE, 为362.17 μmol CO2 ·mol-1 ·s-1 ,其他依次为沙蒿>老鹳草>甘青蒿>圆穗蓼>紫苜蓿>刺儿菜>少花米口袋, 金露梅的PNUE 最小, 为60.3 μmol CO2 ·mol-1 ·s-1 (图1B )。
图1 不同优势种资源利用效率(平均值±标准偏差)。CE,刺儿菜; GQ, 甘青蒿; JL, 金露梅; LG, 老鹳草; MD, 少花米口袋; MX, 紫苜蓿; PJ, 披碱草; QH, 圆穗蓼; SH, 沙蒿。不同小写字母表示同一测定指标差异显著(p < 0.05)。
Fig. 1 Resource-use efficiency of different dominant species (mean ± SD ). CE, Cirsium setosum; GQ, Artemisia tangutica; JL, Potentilla fruticosa; LG, Geranium wilfordii; MD, Gueldenstaedtia verna; MX, Medicago sativa; PJ, Elymus dahuricus; QH, Polygonum macrophyllum; SH, Artemisia desertorum . PNUE, photosynthetic nitrogen-use efficiency; WUE, water-use efficiency. Different letters in the same column are significantly different (p < 0.05).
2.3 不同物种光合生理特征间的相关性及其PCA分析
从表3 可以看出, A area 与PNUE 和WUE 均极显著正相关(p < 0.001); N mass 与PNUE 极显著负相关(p < 0.01), 而与SLA 极显著正相关(p < 0.01); SPAD 与A area 极显著正相关, 与其他生理特征的正相关性不显著; G s 与A area 和PNUE 显著正相关(p < 0.05), 而与WUE 显著负相关(p < 0.05); 另外, SLA 与WUE 显著负相关(p < 0.05), WUE 与PNUE 间显著正相关(p < 0.05)。
PCA分析显示, 物种的第一主成分PC1 (component 1)和第二主成分PC2 (component 2)的方差贡献率分别是43.17%和25.94% (图2 )。其中A area 、WUE 、PNUE 在第一主成分上的载荷较大, 其载荷值分别是0.97、0.78、0.70, 而N mass 、SPAD 在第二主成分上的载荷较大, 其载荷值分别是0.95、0.64。灌丛金露梅处在第一主成分的最左端, 是因为其A area 、WUE 及PNUE 在所研究的物种中均最小; 豆科植物紫苜蓿和少花米口袋由于自身的固氮能力具有较大的N mass , 处在PC2的上端, 图2 显示, 9种植物在主成分坐标轴中大致分为3个部分, 分别是以豆科、杂草和灌木聚集的。这些结果表明: 不同物种对光合生理特征有不同的权衡, 相同功能群植物间的这种权衡关系较近, 且禾本科的披碱草与杂生草本植物的权衡策略较近。
图2 9种植物光合生理特征主成分分析。CE,刺儿菜; GQ, 甘青蒿; JL, 金露梅; LG, 老鹳草; MD, 少花米口袋; MX, 紫苜蓿; PJ, 披碱草; QH, 圆穗蓼; SH, 沙蒿。
Fig. 2 Principal components (PC) analysis of photosynthetic physiological characteristics of nine different species. CE, Cirsium setosum; GQ, Artemisia tangutica; JL, Potentilla fruticosa; LG, Geranium wilfordii; MD, Gueldenstaedtia verna; MX, Medicago sativa; PJ, Elymus dahuricus; QH, Polygonum macrophyllum; SH, Artemisia desertorum .
2.4 同一物种在不同演替群落中光合生理特征的PCA分析
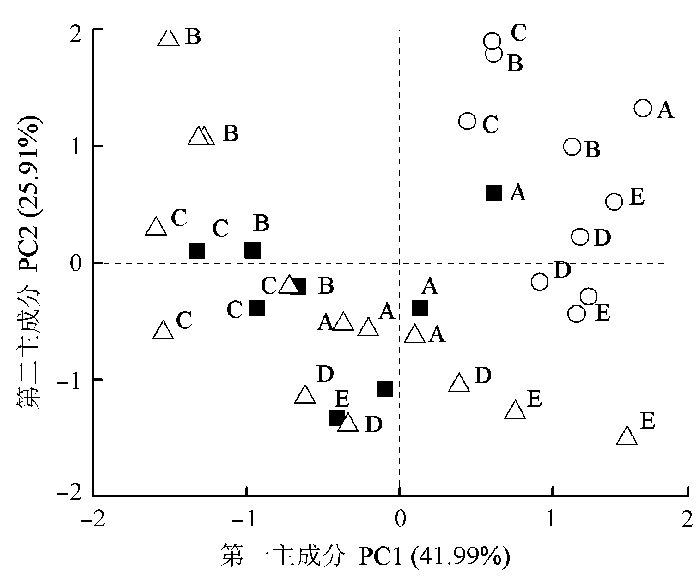
不同群落3种共有种光合生理特征的PCA分析显示: 第一主成分PC1和第二主成分PC2的方差贡献率分别是41.99%和25.91%; PC1载荷较大的因子是SLA 、PUNE 、N mass 和WUE, 其载荷值分别为0.84、-0.78、0.70和-0.69, PC2载荷较大的因子是A area 和SPAD, 其载荷值为0.71和0.70。图3 显示, 所测样品有按物种聚集的趋势, 老鹳草聚集区与禾本科披碱草交叉在一起, 而豆科紫苜蓿形成独自的聚集区, 另外, 3个物种的聚集区有按群落往外延伸的趋势, 反映了植物对不同演替环境变化的响应, 杂生草老鹳草与禾本科披碱草在PC1和PC2上均有变化, 说明它们对于群落变化的响应是同时改变两个主成分上的因子, 而紫苜蓿沿PC2轴的正负轴延伸, 在PC1上基本没有变化, 说明紫苜蓿主要是通过改变PC2来适应群落变化的。从图3 也可以看出群落水平上的这种变化小于物种之间的差异。
图3 5个群落共有种光合生理特征的主成分分析。■, 老鹳草; ○, 紫苜蓿; △, 披碱草; A, 刺儿菜-甘青蒿群落; B, 披碱草-刺儿菜群落; C, 紫苜蓿-甘青蒿群落; D, 沙蒿-羊茅群落; E, 金露梅群落。
Fig. 3 Principal components (PC) analysis of photosynthetic physiological characteristics of common species in five communities. ■, Geranium wilfordii; ○, Medicago sativa; △, Elymus dahuricus ; A, Cirsium setosum -Artemisia tangutica community; B, Elymus dahuricus -Cirsium setosum community; C, Medicago sativa -Artemisia tangutica community; D, Artemisia desertorum -Festuca ovina community; E, Potentilla fruticosa community.
3 讨论
3.1 优势种光合生理特征对演替的响应
光合作用决定着植物物质积累能力的高低, 一定程度上也决定了植物生长的快慢。演替前期, 优势种光合作用速率较高, 随着演替的进行, 其光合作用速率逐渐下降(丁圣彦和宋永昌, 1999 )。本文所研究的群落从A到E顺次形成了由低到高的演替序列, 表2 所选优势种从上到下(图1 由左到右)反映了由低到高演替群落优势种的光合生理特征, 从演替前期的披碱草到中期的甘青蒿、沙蒿到顶级群落的灌丛金露梅A area 逐渐降低, 与前人的研究一致。Bazzaz (1979)的研究也表明, 一般情况下, 植物光合速率随着演替的进行而降低, 草本植物比灌丛植物具有更高的光合速率, 这是因为演替早期优势种具有阳生植物的特性, 而演替后期植物具有阴生植物的特性(李庆康和马克平, 2002 ), 一般而言, 阳生植物的净光合速率高于阴生植物(Bassow & Bazzaz, 1997 ), 较高光合速率的物种也被认为在能量获取和竞争力上有优势(Givnish, 1988 )。A area 与叶绿素含量呈显著正相关(表3 ), SPAD 值从演替前期到演替后期也呈降低趋势(表2 )。随着演替时间尺度的延伸,安慧和上官周平(2007) 发现优势种叶片氮含量呈增加—降低—增加的双峰曲线, 灌丛群落优势种叶片氮含量明显高于其他群落优势种, Ellsworth和Reich (1996) 发现植物叶片氮含量随演替进行逐渐降低, 本研究中, 优势种叶片氮含量没有按演替变化表现出明显规律, 而是按植物功能型表现为豆科植物>灌丛植物和非豆科杂生类植物>禾本科植物。
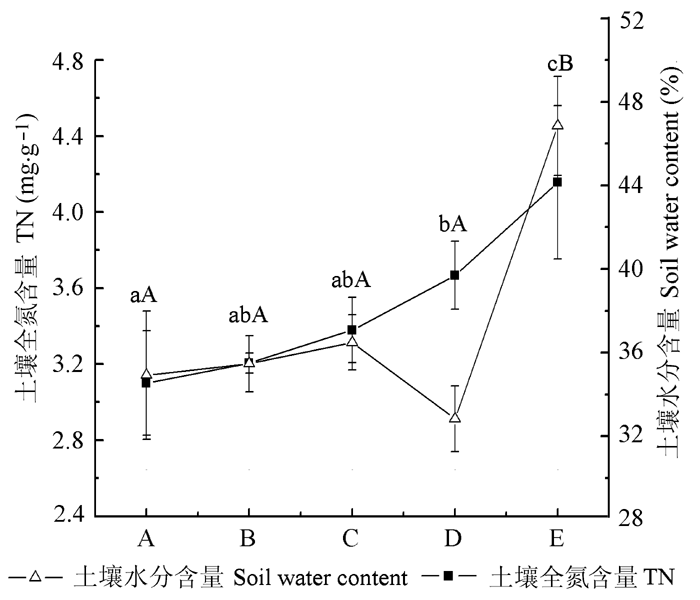
氮和水分是植物光合作用和生长必不可少的因子(Novriyanti et al ., 2012 ), 光合氮利用效率和水分利用效率可以预测氮素和水分是如何最优化地增加净光合速率的(Castellanos et al ., 2005 ), 资源匮乏的环境中物种会采用资源保护的特征如高的资源利用效率(RUE )来增加自身的竞争力(Funk & Vitousek, 2007 )。当土壤供水不足时, 植物通过降低G s 控制气体交换及调节光化学过程等生理活动降低净光合速率, 以此来减少自身对水分的需求(Mahajan & Narendra, 2005 ), 本研究中, 随着演替的进行, 群落表层土壤水分及全氮呈增长趋势(D群落由于坡度较大, 土壤含水量较低)(图4 ), 优势种WUE 逐渐降低(图1A ), 尽管D群落土壤含水量较低, 该群落优势种沙蒿、圆穗蓼的WUE 却没有因此升高, 崔骁勇等(2000) 的研究表明沙蒿通过对土壤干旱和高温的忍耐机制而保持长时间较高的光能利用效率, 依靠高蒸腾速率和强的水分吸收能力来提高自己在群落中的竞争力, 这种靠增加G s 获得高的光合速率和PNUE 的方式必然会导致WUE 的下降(Huang et al ., 2008 ); PNUE 没有随演替变化表现出一致的规律(图1B ), 而是与优势种叶片氮含量呈显著负相关(表3 ), Lambers等(2008) 的研究表明, 在能够支持植物生长的光强下, 氮含量较低的植物叶片的PNUE 较大, 这是因为这些植物合理地将叶片氮分配到光合中, 以达到最优的光合氮利用效率(Novriyanti et al ., 2012 ), 优势种披碱草正是利用这种策略占据B群落, 并在该群落中具有很高的重要值(表2 )。总之, 演替不同阶段优势种光合生理特征体现了其对不同环境的适应策略。
图4 五个群落样地土壤水分与全氮含量(平均值±标准偏差)。A, 刺儿菜-甘青蒿群落; B, 披碱草-刺儿菜群落; C, 紫苜蓿-甘青蒿群落; D, 沙蒿-羊茅群落; E, 金露梅群落。不同大写字母表示土壤水分含量间的差异显著; 不同小写字母表示土壤全氮含量间差异显著(p < 0.05)。
Fig. 4 Soil water content and total nitrogen content (TN) of five different community plots (mean ± SD ). A, Cirsium setosum -Artemisia tangutica community; B, Elymus dahuricus - Cirsium setosum community; C, Medicago sativa -Artemisia tangutica community; D, Artemisia desertorum -Festuca ovina community; E, Potentilla fruticosa community. Different capital letters indicate significant differences among soil water contents; different small letters indicate significant differences among soil total nitrogen contents (p < 0.05).
3.2 同一植物在不同演替阶段的光合生理特征权衡及其影响因子
排除了不同种的差异后, 同种植物在不同演替阶段光合生理特征的研究更能体现植物对演替的响应。植被演替不同阶段由于群落组成及结构不同而导致各种环境因子的变化。首先, 植物群落演替对土壤理化性质产生一定的影响, 如土壤全氮随植被演替的进行呈增加趋势(Wang, 2002 ); 其次, 不同演替阶段群落的结构不同, 导致垂直方向上光照的空间异质性, 演替早期是一个阳性的开放系统, 植物能够获得更高的相对光量子通量而具有较大的光合速率(Anten et al ., 1998a )。所有这些环境因子的改变都会导致植物光合生理特征的变化。本研究中, 披碱草和老鹳草在演替前期(A)光合能力较弱(图3 ), 可能是受氮限制的原因, 安慧和上官周平(2007) 发现在低氮含量下白车轴草幼苗的光合能力会随着氮素的添加而增加, 而在高氮素含量下光合能力又会下降。在一个土壤水分、全氮含量逐渐升高的演替序列中(图4 ), 氮和水分对植物的限制作用逐渐减小, 非豆科植物披碱草、老鹳草的PNUE 、WUE 逐渐降低, 叶片N mass 逐渐升高(图3 ), 反映了不同物种通过对营养物质的不同分配策略来保护匮乏资源, 而豆科植物紫苜蓿的这些特征值却没有明显的变化, 在整个演替进程中紫苜蓿保持了较大的SLA (图3 ), 低SLA 植物能够更好地适应资源贫瘠和干旱的环境, 而高SLA 的植物保持体内营养物质的能力较强(李玉霖等, 2005 ), 这可能就是演替中养分、水分对紫苜蓿没有显著影响的原因, 紫苜蓿与其他两个非豆科植物一样, 随着演替的进行A area 、SPAD 有所下降。在养分和水分影响不大的情况下, 光照可能是导致其光合能力发生变化的主要原因, 光照会控制叶氮的分布(Anten et al ., 1998b ), 进而影响叶片的光合能力。
综上所述, 本文对甘南亚高寒草甸围封地典型群落样地进行样方调查, 通过各群落丰富度、Shannon-Wiener指数、植物生活型及主要物种重要值判定该围封地内形成一个以草本植物→半灌木→灌木的演替序列, 群落表层土壤含水量及全氮含量随演替逐渐增加。通过对各演替阶段优势种及共有种的光合生理特征分析发现, 演替前期, 光合能力较强及资源利用效率较高的物种占据了群落的优势地位; 随着演替的进行, 这些物种的优势地位逐渐被光合能力及资源利用效率低的物种取代; 同种植物在演替前期也表现出比后期更强的光合能力及资源利用效率, 显示了沿着演替梯度由物质获取能力较强的群落向物质保持能力强的群落的转变, 而水分及氮素含量是导致这种转变发生的主要影响因子。这些研究结果对亚高寒退化草甸围封恢复具有一定的理论指导意义。另外, 在演替梯度上, 群落垂直方向光照的空间异质性对植物光合生理特征也有重要的影响, 尚需进一步的研究。
致谢 感谢兰州大学生命科学学院干旱与草地生态教育部重点实验室在实验中给予的帮助。
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
An H Shangguan ZP 2007 ). Photosynthetic characteristics of dominant plant species at different succession stages of vegetation on Loess Plateau.
Chinese Journal of Applied Ecology , 18 , 1175 -1180 .(in Chinese with English abstract)
[安慧 , 上官周平 (2007 ). 黄土高原植被不同演替阶段优势种的光合生理特性
. 应用生态学报 , 18 , 1175 -1180 .]
[2]
An H Shangguan ZP 2008 ). Specific leaf area, leaf nitrogen content, and photosynthetic acclimation of Trifolium repens L. seedlings grown at different irradiances and nitrogen concentrations.
Photosynthetica , 46 , 143 -147 .
[3]
Anten NPR Miyazawa K Hikosaka K Nagashima H Hirose T 1998 a). Leaf nitrogen distribution in relation to leaf age and photon flux density in dominant and subordinate plants in dense stands of a dicotyledonous herb.
Oecologia , 113 , 314 -324 .
[本文引用: 1]
[4]
Anten NPR Werger MJA Medina E 1998 b). Nitrogen distribution and leaf area indices in relation to photosynthetic nitrogen use efficiency in savanna grasses.
Plant Ecology , 138 , 63 -75 .
[本文引用: 1]
[5]
Bassow SL Bazzaz FA 1997 ). Intra-and inter-specific variation in canopy photosynthesis in a mixed deciduous forest.
Oecologia , 109 , 507 -515 .
[本文引用: 2]
[6]
Bazzaz FA 1979 ). The physiological ecology of plant succession.
Annual Review of Ecology and Systematics , 10 , 351 -371 .
[7]
Castellanos AE Martinez MJ Llano JM Halvorson WL Espiricueta M Espejel I 2005 ). Successional trends in Sonoran Desert abandoned agricultural fields in northern Mexico.
Journal of Arid Environments , 60 , 437 -455 .
[本文引用: 1]
[8]
Cui XY Du ZC Wang YF 2000 ). Photosynthetic characteristics of a semi-arid sandy grassland community in inner mongolia.
Acta Phytoecologica Sinica , 24 , 541 -546 .(in Chinese with English abstract)
[本文引用: 1]
[崔骁勇 , 杜占池 , 王艳芬 (2000 ). 内蒙古半干旱草原区沙地植物群落光合特征的动态研究
. 植物生态学报 , 24 , 541 -546 .]
[本文引用: 1]
[9]
Ding SY Song YC 1999 ). The comparation of photosynthesis physi-ecology of evergreen broad-leaved forest of Tiantong national forest park in Zhejiang Province, China.
Acta Ecologica Sinica , 19 , 318 -323 .(in Chinese with English abstract)
[本文引用: 1]
[丁圣彦 , 宋永昌 (1999 ). 浙江天童常绿阔叶林演替系列优势种光合生理生态的比较
. 生态学报 , 19 , 318 -323 .]
[本文引用: 1]
[10]
Ellsworth DS Reich PB 1996 ). Photosynthesis and leaf nitrogen in five Amazonian tree species during early secondary succession.
Ecology , 77 , 581 -594 .
[11]
Evans JR 1989 ). Photosynthesis and nitrogen relationships in leaves of C3 plants.
Oecologia , 78 , 9 -19 .
[本文引用: 1]
[12]
Funk JL Vitousek PM 2007 ). Resource-use efficiency and plant invasion in low-resource systems.
Nature , 446 , 1079 -1081 .
[本文引用: 1]
[13]
Givnish TJ 1988 ). Adaptation to sun and shade: A whole-plant perspective.
Australian Journal of Plant Physiology , 15 , 63 -92 .
[本文引用: 1]
[14]
Hikosaka K Hirose T 2000 ). Photosynthetic nitrogen-use efficiency in evergreen broad-leaved woody species coexisting in a warm-temperate forest.
Tree Physiology , 20 , 1249 -1254 .
[本文引用: 1]
[15]
Huang ZQ Xu ZH Blumfield TJ Bubb K 2008 ). Effects of mulching on growth, foliar photosynthetic nitrogen and water use efficiency of hardwood plantations in subtropical Australia.
Forest Ecology and Management , 255 , 3447 -3454 .
[本文引用: 1]
[16]
Jiang GM He WM 1999 ). Species-and habitat-variability of photosynthesis, transpiration and water use efficiency of different plant species in Maowusu Sand Area.
Acta Botanica Sinica , 41 , 114 -1124 .
[本文引用: 2]
[17]
Lambers H Chapin III FS Pons TL 2008 ). Plant Physiological Ecology. 2nd edn. Springer
, New York.
[18]
Li QK Ma KP 2002 ). Advances in plant succession ecophysiology.
Acta Phytoecologica Sinica , 26 (S1 ), 9 -19 .(in Chinese with English abstract)
[本文引用: 1]
[李庆康 , 马克平 (2002 ). 植物群落演替过程中植物生理生态学特性及其主要环境因子的变化
. 植物生态学报 , 26 (S1 ), 9 -19 .]
[本文引用: 1]
[19]
Li YL Cui JY Su YZ 2005 ). Specific leaf area and leaf dry matter content of some plants in different dune habitats.
Acta Ecologica Sinica , 25 , 304 -311 .(in Chinese with English abstract)
[本文引用: 1]
[李玉霖 , 崔建垣 , 苏永中 (2005 ). 不同沙丘生境主要植物比叶面积和叶干物质含量的比较
. 生态学报 , 25 , 304 -311 .]
[本文引用: 1]
[20]
Liu FD Yang WJ Wang ZS Xu Z Liu H Zhang M Liu YH An SQ Sun SC 2010 ). Plant size effects on the relationships among specific leaf area, leaf nutrient content, and photosynthetic capacity in tropical woody species.
Acta Oecologica , 36 , 149 -159 .
[本文引用: 1]
[21]
Mahajan S Narendra T 2005 ). Cold, salinity and drought stresses: An overview.
Archives of Biochemistry and Biophysis , 444 , 139 -158 .
[本文引用: 1]
[22]
Manetas Y Grammatikopoulos G Kyparissis A 1998 ). The use of the portable, non-destructive, SPAD-502 (Minolta) chlorophyll meter with leaves of varying trichome density and anthocyanin content.
Journal of Plant Physiology , 153 , 513 -516 .
[本文引用: 1]
[23]
Niu SL Jiang GM Gao LM Li YG Liu MZ 2003 ). Comparison of gas exchange traits of different plant species in Hunshandak sand area.
Acta Phytoecologica Sinica , 27 , 318 -324 .
[本文引用: 1]
[24]
Novriyanti E Watanabe M Makoto K Takeda T Hashidoko Y Koike T 2012 ). Photosynthetic nitrogen and water use efficiency of acacia and eucalypt seedlings as afforestation species.
Photosynthetica , 50 , 273 -281 .
[本文引用: 3]
[25]
Ran F Zhang XL Zhang YB Korpelainen H Li CY 2013 ). Altitudinal variation in growth, photosynthetic capacity and water use efficiency of Abies faxoniana Rehd. et Wils. seedlings as revealed by reciprocal transplantations.
Trees , 27 , 1405 -1416 .
[26]
Shangguan ZP Shao MG Dyckmans J 2000 ). Effects of nitrogen nutrition and water deficit on net photosynthetic rate and chlorophyll fluorescence in winter wheat.
Journal of Plant Physiology , 156 , 46 -51 .
[27]
Sun HQ Zhou H Wang P 1999 ). Progress on grassland degenerated succession.
Grassland of China , 21 (1 ), 51 -56 .(in Chinese with English abstract)
[本文引用: 1]
[孙海群 , 周禾 , 王培 (1999 ). 草地退化演替研究进展
. 中国草地 , 21 (1 ), 51 -56 .]
[本文引用: 1]
[28]
Wang GH 2002 ). Plant traits and soil chemical variables during a secondary vegetation succession in abandoned fields on the loess plateau.
Acta Botanica Sinica , 44 , 990 -998 .
[本文引用: 1]
[29]
Wright LJ Reich PB Cornelissen JHC Falster DS Garnier E Hikosaka K Lamont BB Lee W Oleksyn J Osada N Poorter H Villar R Warton DI Westoby M 2005 ). Assessing the generality of global leaf trait relationships.
New Phytologist , 166 , 485 -496 .
[本文引用: 1]
[30]
Wu GL Du GZ 2007 ). Discussion on ecological construction and sustainable development of degraded alpine grassland ecosystem of the Qinghai-Tibetan Plateau.
Chinese Journal of Nature , 29 , 159 -164 .(in Chinese)
[本文引用: 1]
[武高林 , 杜国祯 (2007 ). 青藏高原退化高寒草地生态系统恢复和可持续发展探讨
. 自然杂志 , 29 , 159 -164 .]
[本文引用: 1]
[31]
Yan CR Han XG Chen LZ 2001 ). Water use efficiency of six woody species in relation to micro-environmental factors of different habitats.
Acta Ecologica Sinica , 21 , 1952 -1956 .(in Chinese with English abstract)
[本文引用: 1]
[严昌荣 , 韩兴国 , 陈灵芝 (2001 ). 六种木本植物水分利用效率和其小生境关系研究
. 生态学报 , 21 , 1952 -1956 .]
[本文引用: 1]
[32]
Yu GR Wang QF 2010 ). Ecophysiology of Plant Photosynthesis, Transpiration, and Water Use . Science Press , Beijing . 152 .(in Chinese)
[本文引用: 1]
[于贵瑞 , 王秋凤 (2010 ). 植物光合、蒸腾与水分利用的生理生态学 . 科学出版社 , 北京 . 152 .]
[本文引用: 1]
[33]
Zhang DY Wang G Zhao SL 1988 ). A quantitative study of the vegetation succession on the abandoned arable lands of the subalpine meadows in Gannan Prefecture of Gansu Province I. Analysis of community comosition.
Acta Phytoecological et Geobotanica Sinica , 12 , 283 -291 .(in Chinese with English abstract)
[本文引用: 1]
[张大勇 , 王刚 , 赵松岭 (1988 ). 亚高山草甸弃耕地植物群落演替的数量研究I. 群落组成分析
. 植物生态学与地植物学学报 , 12 , 283 -291 .]
[本文引用: 1]
[34]
Zhang WY Fan JW Zhong HP Hu ZM Song LL Wang N 2010 ). The nitrogen: phosphorus stoichiometry of different plant functional groups for dominant species of typical steppes in China.
Acta Agrestia Sinica , 18 , 503 -509 .(in Chinese with English abstract)
[本文引用: 1]
[张文彦 , 樊江文 , 钟华平 , 胡中民 , 宋璐璐 , 王宁 (2010 ). 中国典型草原优势植物功能群氮磷化学计量学特征研究
. 草地学报 , 18 , 503 -509 .]
[本文引用: 1]
[35]
Zhou HK Zhao XQ Zhao L Li YN Wang SP Xu SX Zhou L 2008 ). Restoration capability of alpine meadow ecosystem on Qinghai-Tibetan Plateau.
Chinese Journal of Ecology , 27 , 697 -704 .(in Chinese with English abstract)
[本文引用: 1]
[周华坤 , 赵新全 , 赵亮 , 李英年 , 汪诗平 , 徐世晓 , 周立 (2008 ). 青藏高原高寒草甸生态系统的恢复能力
. 生态学杂志 , 27 , 697 -704 .]
[本文引用: 1]
黄土高原植被不同演替阶段优势种的光合生理特性
0
2007
黄土高原植被不同演替阶段优势种的光合生理特性
0
2007
Specific leaf area, leaf nitrogen content, and photosynthetic acclimation of Trifolium repens L. seedlings grown at different irradiances and nitrogen concentrations.
0
2008
a). Leaf nitrogen distribution in relation to leaf age and photon flux density in dominant and subordinate plants in dense stands of a dicotyledonous herb.
1
1998
... 排除了不同种的差异后, 同种植物在不同演替阶段光合生理特征的研究更能体现植物对演替的响应.植被演替不同阶段由于群落组成及结构不同而导致各种环境因子的变化.首先, 植物群落演替对土壤理化性质产生一定的影响, 如土壤全氮随植被演替的进行呈增加趋势(Wang, 2002 ); 其次, 不同演替阶段群落的结构不同, 导致垂直方向上光照的空间异质性, 演替早期是一个阳性的开放系统, 植物能够获得更高的相对光量子通量而具有较大的光合速率(Anten et al ., 1998a ).所有这些环境因子的改变都会导致植物光合生理特征的变化.本研究中, 披碱草和老鹳草在演替前期(A)光合能力较弱(图3 ), 可能是受氮限制的原因, 安慧和上官周平(2007) 发现在低氮含量下白车轴草幼苗的光合能力会随着氮素的添加而增加, 而在高氮素含量下光合能力又会下降.在一个土壤水分、全氮含量逐渐升高的演替序列中(图4 ), 氮和水分对植物的限制作用逐渐减小, 非豆科植物披碱草、老鹳草的PNUE 、WUE 逐渐降低, 叶片N mass 逐渐升高(图3 ), 反映了不同物种通过对营养物质的不同分配策略来保护匮乏资源, 而豆科植物紫苜蓿的这些特征值却没有明显的变化, 在整个演替进程中紫苜蓿保持了较大的SLA (图3 ), 低SLA 植物能够更好地适应资源贫瘠和干旱的环境, 而高SLA 的植物保持体内营养物质的能力较强(李玉霖等, 2005 ), 这可能就是演替中养分、水分对紫苜蓿没有显著影响的原因, 紫苜蓿与其他两个非豆科植物一样, 随着演替的进行A area 、SPAD 有所下降.在养分和水分影响不大的情况下, 光照可能是导致其光合能力发生变化的主要原因, 光照会控制叶氮的分布(Anten et al ., 1998b ), 进而影响叶片的光合能力. ...
b). Nitrogen distribution and leaf area indices in relation to photosynthetic nitrogen use efficiency in savanna grasses.
1
1998
... 排除了不同种的差异后, 同种植物在不同演替阶段光合生理特征的研究更能体现植物对演替的响应.植被演替不同阶段由于群落组成及结构不同而导致各种环境因子的变化.首先, 植物群落演替对土壤理化性质产生一定的影响, 如土壤全氮随植被演替的进行呈增加趋势(Wang, 2002 ); 其次, 不同演替阶段群落的结构不同, 导致垂直方向上光照的空间异质性, 演替早期是一个阳性的开放系统, 植物能够获得更高的相对光量子通量而具有较大的光合速率(Anten et al ., 1998a ).所有这些环境因子的改变都会导致植物光合生理特征的变化.本研究中, 披碱草和老鹳草在演替前期(A)光合能力较弱(图3 ), 可能是受氮限制的原因, 安慧和上官周平(2007) 发现在低氮含量下白车轴草幼苗的光合能力会随着氮素的添加而增加, 而在高氮素含量下光合能力又会下降.在一个土壤水分、全氮含量逐渐升高的演替序列中(图4 ), 氮和水分对植物的限制作用逐渐减小, 非豆科植物披碱草、老鹳草的PNUE 、WUE 逐渐降低, 叶片N mass 逐渐升高(图3 ), 反映了不同物种通过对营养物质的不同分配策略来保护匮乏资源, 而豆科植物紫苜蓿的这些特征值却没有明显的变化, 在整个演替进程中紫苜蓿保持了较大的SLA (图3 ), 低SLA 植物能够更好地适应资源贫瘠和干旱的环境, 而高SLA 的植物保持体内营养物质的能力较强(李玉霖等, 2005 ), 这可能就是演替中养分、水分对紫苜蓿没有显著影响的原因, 紫苜蓿与其他两个非豆科植物一样, 随着演替的进行A area 、SPAD 有所下降.在养分和水分影响不大的情况下, 光照可能是导致其光合能力发生变化的主要原因, 光照会控制叶氮的分布(Anten et al ., 1998b ), 进而影响叶片的光合能力. ...
Intra-and inter-specific variation in canopy photosynthesis in a mixed deciduous forest.
2
1997
... 植物光合生理特征能体现植物对不同生境的适应, 反映植物对不同环境变化的响应(Jiang & He, 1999 ).不同物种的光合生理特征间有显著的差异(Bassow & Bazzaz, 1997 ), 这正是不同植物利用不同的生态策略获得资源利用的反映(Niu et al ., 2003 ).一般而言, C4 光合途径或具有固氮能力的物种比C3 途径和无固氮能力的植物具备较高的光合能力和光合氮利用效率(Jiang & He, 1999 ), 因为氮含量与植物光合能力有非常密切的联系(Evans, 1989 ).双子叶植物、非禾本科植物、豆科植物和中生型植物的叶片氮含量分别高于相应的单子叶植物、禾本科植物、非豆科植物和旱生型植物(张文彦等, 2010 ), 草本植物比乔木和灌丛具有更高的净光合速率和光合氮利用效率(Wright et al ., 2005 ).Wright等(2005)在对全球植物叶片普遍特征的关系研究中还发现, 一般而言, 草本植物比灌木具有更低的基于面积的氮含量和比叶质量.另外, 植物光合生理特征也会随着叶片生活史长短(Hikosaka & Hirose, 2000 )和植株的大小而变化(Liu et al ., 2010 ). ...
... 光合作用决定着植物物质积累能力的高低, 一定程度上也决定了植物生长的快慢.演替前期, 优势种光合作用速率较高, 随着演替的进行, 其光合作用速率逐渐下降(丁圣彦和宋永昌, 1999 ).本文所研究的群落从A到E顺次形成了由低到高的演替序列, 表2 所选优势种从上到下(图1 由左到右)反映了由低到高演替群落优势种的光合生理特征, 从演替前期的披碱草到中期的甘青蒿、沙蒿到顶级群落的灌丛金露梅A area 逐渐降低, 与前人的研究一致.Bazzaz (1979)的研究也表明, 一般情况下, 植物光合速率随着演替的进行而降低, 草本植物比灌丛植物具有更高的光合速率, 这是因为演替早期优势种具有阳生植物的特性, 而演替后期植物具有阴生植物的特性(李庆康和马克平, 2002 ), 一般而言, 阳生植物的净光合速率高于阴生植物(Bassow & Bazzaz, 1997 ), 较高光合速率的物种也被认为在能量获取和竞争力上有优势(Givnish, 1988 ).A area 与叶绿素含量呈显著正相关(表3 ), SPAD 值从演替前期到演替后期也呈降低趋势(表2 ).随着演替时间尺度的延伸,安慧和上官周平(2007) 发现优势种叶片氮含量呈增加—降低—增加的双峰曲线, 灌丛群落优势种叶片氮含量明显高于其他群落优势种, Ellsworth和Reich (1996) 发现植物叶片氮含量随演替进行逐渐降低, 本研究中, 优势种叶片氮含量没有按演替变化表现出明显规律, 而是按植物功能型表现为豆科植物>灌丛植物和非豆科杂生类植物>禾本科植物. ...
The physiological ecology of plant succession.
0
1979
Successional trends in Sonoran Desert abandoned agricultural fields in northern Mexico.
1
2005
... 氮和水分是植物光合作用和生长必不可少的因子(Novriyanti et al ., 2012 ), 光合氮利用效率和水分利用效率可以预测氮素和水分是如何最优化地增加净光合速率的(Castellanos et al ., 2005 ), 资源匮乏的环境中物种会采用资源保护的特征如高的资源利用效率(RUE )来增加自身的竞争力(Funk & Vitousek, 2007 ).当土壤供水不足时, 植物通过降低G s 控制气体交换及调节光化学过程等生理活动降低净光合速率, 以此来减少自身对水分的需求(Mahajan & Narendra, 2005 ), 本研究中, 随着演替的进行, 群落表层土壤水分及全氮呈增长趋势(D群落由于坡度较大, 土壤含水量较低)(图4 ), 优势种WUE 逐渐降低(图1A ), 尽管D群落土壤含水量较低, 该群落优势种沙蒿、圆穗蓼的WUE 却没有因此升高, 崔骁勇等(2000) 的研究表明沙蒿通过对土壤干旱和高温的忍耐机制而保持长时间较高的光能利用效率, 依靠高蒸腾速率和强的水分吸收能力来提高自己在群落中的竞争力, 这种靠增加G s 获得高的光合速率和PNUE 的方式必然会导致WUE 的下降(Huang et al ., 2008 ); PNUE 没有随演替变化表现出一致的规律(图1B ), 而是与优势种叶片氮含量呈显著负相关(表3 ), Lambers等(2008) 的研究表明, 在能够支持植物生长的光强下, 氮含量较低的植物叶片的PNUE 较大, 这是因为这些植物合理地将叶片氮分配到光合中, 以达到最优的光合氮利用效率(Novriyanti et al ., 2012 ), 优势种披碱草正是利用这种策略占据B群落, 并在该群落中具有很高的重要值(表2 ).总之, 演替不同阶段优势种光合生理特征体现了其对不同环境的适应策略. ...
内蒙古半干旱草原区沙地植物群落光合特征的动态研究
1
2000
... 影响植物光合能力的环境因子包括光照、水分、温度、营养物质和CO2 浓度等(于贵瑞和王秋凤, 2010 ), 而水分和氮是植物光合和生长必不可少的因子(Novriyanti et al ., 2012 ), Shangguan等(2000) 研究发现, 水分和氮素亏缺均会强烈地抑制植物的光合活动, 且氮素和水分对光合的影响是相互制约的.干旱大大降低了单叶和群落的净光合速率(崔骁勇等, 2000 ), 而提高了植物的光合水分利用效率(严昌荣等, 2001 ).An和Shangguan (2008)对不同光照和不同浓度氮素下白车轴草(Trifolium repens )幼苗光合生理特征的研究发现, 低浓度氮下基于面积的CO2 同化速率(A area )、光合氮利用效率(PNUE )和叶绿素含量随着氮素浓度的升高而增加; 高浓度氮下, 这些光合特征值会下降, 而比叶面积(SLA )和基于质量的叶片氮含量(N mass )会随着氮素浓度的升高持续增加.另外, 海拔通过影响CO2 分压、温度、水分等环境因子而影响植物光合生理特征, Ran等(2013) 将岷江冷杉(Abies faxoniana )移植到低海拔处, 降低了岷江冷杉的叶氮含量和水分利用效率, 而增加了净光合速率和PNUE . ...
内蒙古半干旱草原区沙地植物群落光合特征的动态研究
1
2000
... 影响植物光合能力的环境因子包括光照、水分、温度、营养物质和CO2 浓度等(于贵瑞和王秋凤, 2010 ), 而水分和氮是植物光合和生长必不可少的因子(Novriyanti et al ., 2012 ), Shangguan等(2000) 研究发现, 水分和氮素亏缺均会强烈地抑制植物的光合活动, 且氮素和水分对光合的影响是相互制约的.干旱大大降低了单叶和群落的净光合速率(崔骁勇等, 2000 ), 而提高了植物的光合水分利用效率(严昌荣等, 2001 ).An和Shangguan (2008)对不同光照和不同浓度氮素下白车轴草(Trifolium repens )幼苗光合生理特征的研究发现, 低浓度氮下基于面积的CO2 同化速率(A area )、光合氮利用效率(PNUE )和叶绿素含量随着氮素浓度的升高而增加; 高浓度氮下, 这些光合特征值会下降, 而比叶面积(SLA )和基于质量的叶片氮含量(N mass )会随着氮素浓度的升高持续增加.另外, 海拔通过影响CO2 分压、温度、水分等环境因子而影响植物光合生理特征, Ran等(2013) 将岷江冷杉(Abies faxoniana )移植到低海拔处, 降低了岷江冷杉的叶氮含量和水分利用效率, 而增加了净光合速率和PNUE . ...
浙江天童常绿阔叶林演替系列优势种光合生理生态的比较
1
1999
... 光合作用决定着植物物质积累能力的高低, 一定程度上也决定了植物生长的快慢.演替前期, 优势种光合作用速率较高, 随着演替的进行, 其光合作用速率逐渐下降(丁圣彦和宋永昌, 1999 ).本文所研究的群落从A到E顺次形成了由低到高的演替序列, 表2 所选优势种从上到下(图1 由左到右)反映了由低到高演替群落优势种的光合生理特征, 从演替前期的披碱草到中期的甘青蒿、沙蒿到顶级群落的灌丛金露梅A area 逐渐降低, 与前人的研究一致.Bazzaz (1979)的研究也表明, 一般情况下, 植物光合速率随着演替的进行而降低, 草本植物比灌丛植物具有更高的光合速率, 这是因为演替早期优势种具有阳生植物的特性, 而演替后期植物具有阴生植物的特性(李庆康和马克平, 2002 ), 一般而言, 阳生植物的净光合速率高于阴生植物(Bassow & Bazzaz, 1997 ), 较高光合速率的物种也被认为在能量获取和竞争力上有优势(Givnish, 1988 ).A area 与叶绿素含量呈显著正相关(表3 ), SPAD 值从演替前期到演替后期也呈降低趋势(表2 ).随着演替时间尺度的延伸,安慧和上官周平(2007) 发现优势种叶片氮含量呈增加—降低—增加的双峰曲线, 灌丛群落优势种叶片氮含量明显高于其他群落优势种, Ellsworth和Reich (1996) 发现植物叶片氮含量随演替进行逐渐降低, 本研究中, 优势种叶片氮含量没有按演替变化表现出明显规律, 而是按植物功能型表现为豆科植物>灌丛植物和非豆科杂生类植物>禾本科植物. ...
浙江天童常绿阔叶林演替系列优势种光合生理生态的比较
1
1999
... 光合作用决定着植物物质积累能力的高低, 一定程度上也决定了植物生长的快慢.演替前期, 优势种光合作用速率较高, 随着演替的进行, 其光合作用速率逐渐下降(丁圣彦和宋永昌, 1999 ).本文所研究的群落从A到E顺次形成了由低到高的演替序列, 表2 所选优势种从上到下(图1 由左到右)反映了由低到高演替群落优势种的光合生理特征, 从演替前期的披碱草到中期的甘青蒿、沙蒿到顶级群落的灌丛金露梅A area 逐渐降低, 与前人的研究一致.Bazzaz (1979)的研究也表明, 一般情况下, 植物光合速率随着演替的进行而降低, 草本植物比灌丛植物具有更高的光合速率, 这是因为演替早期优势种具有阳生植物的特性, 而演替后期植物具有阴生植物的特性(李庆康和马克平, 2002 ), 一般而言, 阳生植物的净光合速率高于阴生植物(Bassow & Bazzaz, 1997 ), 较高光合速率的物种也被认为在能量获取和竞争力上有优势(Givnish, 1988 ).A area 与叶绿素含量呈显著正相关(表3 ), SPAD 值从演替前期到演替后期也呈降低趋势(表2 ).随着演替时间尺度的延伸,安慧和上官周平(2007) 发现优势种叶片氮含量呈增加—降低—增加的双峰曲线, 灌丛群落优势种叶片氮含量明显高于其他群落优势种, Ellsworth和Reich (1996) 发现植物叶片氮含量随演替进行逐渐降低, 本研究中, 优势种叶片氮含量没有按演替变化表现出明显规律, 而是按植物功能型表现为豆科植物>灌丛植物和非豆科杂生类植物>禾本科植物. ...
Photosynthesis and leaf nitrogen in five Amazonian tree species during early secondary succession.
0
1996
Photosynthesis and nitrogen relationships in leaves of C3 plants.
1
1989
... 植物光合生理特征能体现植物对不同生境的适应, 反映植物对不同环境变化的响应(Jiang & He, 1999 ).不同物种的光合生理特征间有显著的差异(Bassow & Bazzaz, 1997 ), 这正是不同植物利用不同的生态策略获得资源利用的反映(Niu et al ., 2003 ).一般而言, C4 光合途径或具有固氮能力的物种比C3 途径和无固氮能力的植物具备较高的光合能力和光合氮利用效率(Jiang & He, 1999 ), 因为氮含量与植物光合能力有非常密切的联系(Evans, 1989 ).双子叶植物、非禾本科植物、豆科植物和中生型植物的叶片氮含量分别高于相应的单子叶植物、禾本科植物、非豆科植物和旱生型植物(张文彦等, 2010 ), 草本植物比乔木和灌丛具有更高的净光合速率和光合氮利用效率(Wright et al ., 2005 ).Wright等(2005)在对全球植物叶片普遍特征的关系研究中还发现, 一般而言, 草本植物比灌木具有更低的基于面积的氮含量和比叶质量.另外, 植物光合生理特征也会随着叶片生活史长短(Hikosaka & Hirose, 2000 )和植株的大小而变化(Liu et al ., 2010 ). ...
Resource-use efficiency and plant invasion in low-resource systems.
1
2007
... 氮和水分是植物光合作用和生长必不可少的因子(Novriyanti et al ., 2012 ), 光合氮利用效率和水分利用效率可以预测氮素和水分是如何最优化地增加净光合速率的(Castellanos et al ., 2005 ), 资源匮乏的环境中物种会采用资源保护的特征如高的资源利用效率(RUE )来增加自身的竞争力(Funk & Vitousek, 2007 ).当土壤供水不足时, 植物通过降低G s 控制气体交换及调节光化学过程等生理活动降低净光合速率, 以此来减少自身对水分的需求(Mahajan & Narendra, 2005 ), 本研究中, 随着演替的进行, 群落表层土壤水分及全氮呈增长趋势(D群落由于坡度较大, 土壤含水量较低)(图4 ), 优势种WUE 逐渐降低(图1A ), 尽管D群落土壤含水量较低, 该群落优势种沙蒿、圆穗蓼的WUE 却没有因此升高, 崔骁勇等(2000) 的研究表明沙蒿通过对土壤干旱和高温的忍耐机制而保持长时间较高的光能利用效率, 依靠高蒸腾速率和强的水分吸收能力来提高自己在群落中的竞争力, 这种靠增加G s 获得高的光合速率和PNUE 的方式必然会导致WUE 的下降(Huang et al ., 2008 ); PNUE 没有随演替变化表现出一致的规律(图1B ), 而是与优势种叶片氮含量呈显著负相关(表3 ), Lambers等(2008) 的研究表明, 在能够支持植物生长的光强下, 氮含量较低的植物叶片的PNUE 较大, 这是因为这些植物合理地将叶片氮分配到光合中, 以达到最优的光合氮利用效率(Novriyanti et al ., 2012 ), 优势种披碱草正是利用这种策略占据B群落, 并在该群落中具有很高的重要值(表2 ).总之, 演替不同阶段优势种光合生理特征体现了其对不同环境的适应策略. ...
Adaptation to sun and shade: A whole-plant perspective.
1
1988
... 光合作用决定着植物物质积累能力的高低, 一定程度上也决定了植物生长的快慢.演替前期, 优势种光合作用速率较高, 随着演替的进行, 其光合作用速率逐渐下降(丁圣彦和宋永昌, 1999 ).本文所研究的群落从A到E顺次形成了由低到高的演替序列, 表2 所选优势种从上到下(图1 由左到右)反映了由低到高演替群落优势种的光合生理特征, 从演替前期的披碱草到中期的甘青蒿、沙蒿到顶级群落的灌丛金露梅A area 逐渐降低, 与前人的研究一致.Bazzaz (1979)的研究也表明, 一般情况下, 植物光合速率随着演替的进行而降低, 草本植物比灌丛植物具有更高的光合速率, 这是因为演替早期优势种具有阳生植物的特性, 而演替后期植物具有阴生植物的特性(李庆康和马克平, 2002 ), 一般而言, 阳生植物的净光合速率高于阴生植物(Bassow & Bazzaz, 1997 ), 较高光合速率的物种也被认为在能量获取和竞争力上有优势(Givnish, 1988 ).A area 与叶绿素含量呈显著正相关(表3 ), SPAD 值从演替前期到演替后期也呈降低趋势(表2 ).随着演替时间尺度的延伸,安慧和上官周平(2007) 发现优势种叶片氮含量呈增加—降低—增加的双峰曲线, 灌丛群落优势种叶片氮含量明显高于其他群落优势种, Ellsworth和Reich (1996) 发现植物叶片氮含量随演替进行逐渐降低, 本研究中, 优势种叶片氮含量没有按演替变化表现出明显规律, 而是按植物功能型表现为豆科植物>灌丛植物和非豆科杂生类植物>禾本科植物. ...
Photosynthetic nitrogen-use efficiency in evergreen broad-leaved woody species coexisting in a warm-temperate forest.
1
2000
... 植物光合生理特征能体现植物对不同生境的适应, 反映植物对不同环境变化的响应(Jiang & He, 1999 ).不同物种的光合生理特征间有显著的差异(Bassow & Bazzaz, 1997 ), 这正是不同植物利用不同的生态策略获得资源利用的反映(Niu et al ., 2003 ).一般而言, C4 光合途径或具有固氮能力的物种比C3 途径和无固氮能力的植物具备较高的光合能力和光合氮利用效率(Jiang & He, 1999 ), 因为氮含量与植物光合能力有非常密切的联系(Evans, 1989 ).双子叶植物、非禾本科植物、豆科植物和中生型植物的叶片氮含量分别高于相应的单子叶植物、禾本科植物、非豆科植物和旱生型植物(张文彦等, 2010 ), 草本植物比乔木和灌丛具有更高的净光合速率和光合氮利用效率(Wright et al ., 2005 ).Wright等(2005)在对全球植物叶片普遍特征的关系研究中还发现, 一般而言, 草本植物比灌木具有更低的基于面积的氮含量和比叶质量.另外, 植物光合生理特征也会随着叶片生活史长短(Hikosaka & Hirose, 2000 )和植株的大小而变化(Liu et al ., 2010 ). ...
Effects of mulching on growth, foliar photosynthetic nitrogen and water use efficiency of hardwood plantations in subtropical Australia.
1
2008
... 氮和水分是植物光合作用和生长必不可少的因子(Novriyanti et al ., 2012 ), 光合氮利用效率和水分利用效率可以预测氮素和水分是如何最优化地增加净光合速率的(Castellanos et al ., 2005 ), 资源匮乏的环境中物种会采用资源保护的特征如高的资源利用效率(RUE )来增加自身的竞争力(Funk & Vitousek, 2007 ).当土壤供水不足时, 植物通过降低G s 控制气体交换及调节光化学过程等生理活动降低净光合速率, 以此来减少自身对水分的需求(Mahajan & Narendra, 2005 ), 本研究中, 随着演替的进行, 群落表层土壤水分及全氮呈增长趋势(D群落由于坡度较大, 土壤含水量较低)(图4 ), 优势种WUE 逐渐降低(图1A ), 尽管D群落土壤含水量较低, 该群落优势种沙蒿、圆穗蓼的WUE 却没有因此升高, 崔骁勇等(2000) 的研究表明沙蒿通过对土壤干旱和高温的忍耐机制而保持长时间较高的光能利用效率, 依靠高蒸腾速率和强的水分吸收能力来提高自己在群落中的竞争力, 这种靠增加G s 获得高的光合速率和PNUE 的方式必然会导致WUE 的下降(Huang et al ., 2008 ); PNUE 没有随演替变化表现出一致的规律(图1B ), 而是与优势种叶片氮含量呈显著负相关(表3 ), Lambers等(2008) 的研究表明, 在能够支持植物生长的光强下, 氮含量较低的植物叶片的PNUE 较大, 这是因为这些植物合理地将叶片氮分配到光合中, 以达到最优的光合氮利用效率(Novriyanti et al ., 2012 ), 优势种披碱草正是利用这种策略占据B群落, 并在该群落中具有很高的重要值(表2 ).总之, 演替不同阶段优势种光合生理特征体现了其对不同环境的适应策略. ...
Species-and habitat-variability of photosynthesis, transpiration and water use efficiency of different plant species in Maowusu Sand Area.
2
1999
... 植物光合生理特征能体现植物对不同生境的适应, 反映植物对不同环境变化的响应(Jiang & He, 1999 ).不同物种的光合生理特征间有显著的差异(Bassow & Bazzaz, 1997 ), 这正是不同植物利用不同的生态策略获得资源利用的反映(Niu et al ., 2003 ).一般而言, C4 光合途径或具有固氮能力的物种比C3 途径和无固氮能力的植物具备较高的光合能力和光合氮利用效率(Jiang & He, 1999 ), 因为氮含量与植物光合能力有非常密切的联系(Evans, 1989 ).双子叶植物、非禾本科植物、豆科植物和中生型植物的叶片氮含量分别高于相应的单子叶植物、禾本科植物、非豆科植物和旱生型植物(张文彦等, 2010 ), 草本植物比乔木和灌丛具有更高的净光合速率和光合氮利用效率(Wright et al ., 2005 ).Wright等(2005)在对全球植物叶片普遍特征的关系研究中还发现, 一般而言, 草本植物比灌木具有更低的基于面积的氮含量和比叶质量.另外, 植物光合生理特征也会随着叶片生活史长短(Hikosaka & Hirose, 2000 )和植株的大小而变化(Liu et al ., 2010 ). ...
... 途径和无固氮能力的植物具备较高的光合能力和光合氮利用效率(Jiang & He, 1999 ), 因为氮含量与植物光合能力有非常密切的联系(Evans, 1989 ).双子叶植物、非禾本科植物、豆科植物和中生型植物的叶片氮含量分别高于相应的单子叶植物、禾本科植物、非豆科植物和旱生型植物(张文彦等, 2010 ), 草本植物比乔木和灌丛具有更高的净光合速率和光合氮利用效率(Wright et al ., 2005 ).Wright等(2005)在对全球植物叶片普遍特征的关系研究中还发现, 一般而言, 草本植物比灌木具有更低的基于面积的氮含量和比叶质量.另外, 植物光合生理特征也会随着叶片生活史长短(Hikosaka & Hirose, 2000 )和植株的大小而变化(Liu et al ., 2010 ). ...
Plant Physiological Ecology. 2nd edn. Springer
0
2008
植物群落演替过程中植物生理生态学特性及其主要环境因子的变化
1
2002
... 光合作用决定着植物物质积累能力的高低, 一定程度上也决定了植物生长的快慢.演替前期, 优势种光合作用速率较高, 随着演替的进行, 其光合作用速率逐渐下降(丁圣彦和宋永昌, 1999 ).本文所研究的群落从A到E顺次形成了由低到高的演替序列, 表2 所选优势种从上到下(图1 由左到右)反映了由低到高演替群落优势种的光合生理特征, 从演替前期的披碱草到中期的甘青蒿、沙蒿到顶级群落的灌丛金露梅A area 逐渐降低, 与前人的研究一致.Bazzaz (1979)的研究也表明, 一般情况下, 植物光合速率随着演替的进行而降低, 草本植物比灌丛植物具有更高的光合速率, 这是因为演替早期优势种具有阳生植物的特性, 而演替后期植物具有阴生植物的特性(李庆康和马克平, 2002 ), 一般而言, 阳生植物的净光合速率高于阴生植物(Bassow & Bazzaz, 1997 ), 较高光合速率的物种也被认为在能量获取和竞争力上有优势(Givnish, 1988 ).A area 与叶绿素含量呈显著正相关(表3 ), SPAD 值从演替前期到演替后期也呈降低趋势(表2 ).随着演替时间尺度的延伸,安慧和上官周平(2007) 发现优势种叶片氮含量呈增加—降低—增加的双峰曲线, 灌丛群落优势种叶片氮含量明显高于其他群落优势种, Ellsworth和Reich (1996) 发现植物叶片氮含量随演替进行逐渐降低, 本研究中, 优势种叶片氮含量没有按演替变化表现出明显规律, 而是按植物功能型表现为豆科植物>灌丛植物和非豆科杂生类植物>禾本科植物. ...
植物群落演替过程中植物生理生态学特性及其主要环境因子的变化
1
2002
... 光合作用决定着植物物质积累能力的高低, 一定程度上也决定了植物生长的快慢.演替前期, 优势种光合作用速率较高, 随着演替的进行, 其光合作用速率逐渐下降(丁圣彦和宋永昌, 1999 ).本文所研究的群落从A到E顺次形成了由低到高的演替序列, 表2 所选优势种从上到下(图1 由左到右)反映了由低到高演替群落优势种的光合生理特征, 从演替前期的披碱草到中期的甘青蒿、沙蒿到顶级群落的灌丛金露梅A area 逐渐降低, 与前人的研究一致.Bazzaz (1979)的研究也表明, 一般情况下, 植物光合速率随着演替的进行而降低, 草本植物比灌丛植物具有更高的光合速率, 这是因为演替早期优势种具有阳生植物的特性, 而演替后期植物具有阴生植物的特性(李庆康和马克平, 2002 ), 一般而言, 阳生植物的净光合速率高于阴生植物(Bassow & Bazzaz, 1997 ), 较高光合速率的物种也被认为在能量获取和竞争力上有优势(Givnish, 1988 ).A area 与叶绿素含量呈显著正相关(表3 ), SPAD 值从演替前期到演替后期也呈降低趋势(表2 ).随着演替时间尺度的延伸,安慧和上官周平(2007) 发现优势种叶片氮含量呈增加—降低—增加的双峰曲线, 灌丛群落优势种叶片氮含量明显高于其他群落优势种, Ellsworth和Reich (1996) 发现植物叶片氮含量随演替进行逐渐降低, 本研究中, 优势种叶片氮含量没有按演替变化表现出明显规律, 而是按植物功能型表现为豆科植物>灌丛植物和非豆科杂生类植物>禾本科植物. ...
不同沙丘生境主要植物比叶面积和叶干物质含量的比较
1
2005
... 排除了不同种的差异后, 同种植物在不同演替阶段光合生理特征的研究更能体现植物对演替的响应.植被演替不同阶段由于群落组成及结构不同而导致各种环境因子的变化.首先, 植物群落演替对土壤理化性质产生一定的影响, 如土壤全氮随植被演替的进行呈增加趋势(Wang, 2002 ); 其次, 不同演替阶段群落的结构不同, 导致垂直方向上光照的空间异质性, 演替早期是一个阳性的开放系统, 植物能够获得更高的相对光量子通量而具有较大的光合速率(Anten et al ., 1998a ).所有这些环境因子的改变都会导致植物光合生理特征的变化.本研究中, 披碱草和老鹳草在演替前期(A)光合能力较弱(图3 ), 可能是受氮限制的原因, 安慧和上官周平(2007) 发现在低氮含量下白车轴草幼苗的光合能力会随着氮素的添加而增加, 而在高氮素含量下光合能力又会下降.在一个土壤水分、全氮含量逐渐升高的演替序列中(图4 ), 氮和水分对植物的限制作用逐渐减小, 非豆科植物披碱草、老鹳草的PNUE 、WUE 逐渐降低, 叶片N mass 逐渐升高(图3 ), 反映了不同物种通过对营养物质的不同分配策略来保护匮乏资源, 而豆科植物紫苜蓿的这些特征值却没有明显的变化, 在整个演替进程中紫苜蓿保持了较大的SLA (图3 ), 低SLA 植物能够更好地适应资源贫瘠和干旱的环境, 而高SLA 的植物保持体内营养物质的能力较强(李玉霖等, 2005 ), 这可能就是演替中养分、水分对紫苜蓿没有显著影响的原因, 紫苜蓿与其他两个非豆科植物一样, 随着演替的进行A area 、SPAD 有所下降.在养分和水分影响不大的情况下, 光照可能是导致其光合能力发生变化的主要原因, 光照会控制叶氮的分布(Anten et al ., 1998b ), 进而影响叶片的光合能力. ...
不同沙丘生境主要植物比叶面积和叶干物质含量的比较
1
2005
... 排除了不同种的差异后, 同种植物在不同演替阶段光合生理特征的研究更能体现植物对演替的响应.植被演替不同阶段由于群落组成及结构不同而导致各种环境因子的变化.首先, 植物群落演替对土壤理化性质产生一定的影响, 如土壤全氮随植被演替的进行呈增加趋势(Wang, 2002 ); 其次, 不同演替阶段群落的结构不同, 导致垂直方向上光照的空间异质性, 演替早期是一个阳性的开放系统, 植物能够获得更高的相对光量子通量而具有较大的光合速率(Anten et al ., 1998a ).所有这些环境因子的改变都会导致植物光合生理特征的变化.本研究中, 披碱草和老鹳草在演替前期(A)光合能力较弱(图3 ), 可能是受氮限制的原因, 安慧和上官周平(2007) 发现在低氮含量下白车轴草幼苗的光合能力会随着氮素的添加而增加, 而在高氮素含量下光合能力又会下降.在一个土壤水分、全氮含量逐渐升高的演替序列中(图4 ), 氮和水分对植物的限制作用逐渐减小, 非豆科植物披碱草、老鹳草的PNUE 、WUE 逐渐降低, 叶片N mass 逐渐升高(图3 ), 反映了不同物种通过对营养物质的不同分配策略来保护匮乏资源, 而豆科植物紫苜蓿的这些特征值却没有明显的变化, 在整个演替进程中紫苜蓿保持了较大的SLA (图3 ), 低SLA 植物能够更好地适应资源贫瘠和干旱的环境, 而高SLA 的植物保持体内营养物质的能力较强(李玉霖等, 2005 ), 这可能就是演替中养分、水分对紫苜蓿没有显著影响的原因, 紫苜蓿与其他两个非豆科植物一样, 随着演替的进行A area 、SPAD 有所下降.在养分和水分影响不大的情况下, 光照可能是导致其光合能力发生变化的主要原因, 光照会控制叶氮的分布(Anten et al ., 1998b ), 进而影响叶片的光合能力. ...
Plant size effects on the relationships among specific leaf area, leaf nutrient content, and photosynthetic capacity in tropical woody species.
1
2010
... 植物光合生理特征能体现植物对不同生境的适应, 反映植物对不同环境变化的响应(Jiang & He, 1999 ).不同物种的光合生理特征间有显著的差异(Bassow & Bazzaz, 1997 ), 这正是不同植物利用不同的生态策略获得资源利用的反映(Niu et al ., 2003 ).一般而言, C4 光合途径或具有固氮能力的物种比C3 途径和无固氮能力的植物具备较高的光合能力和光合氮利用效率(Jiang & He, 1999 ), 因为氮含量与植物光合能力有非常密切的联系(Evans, 1989 ).双子叶植物、非禾本科植物、豆科植物和中生型植物的叶片氮含量分别高于相应的单子叶植物、禾本科植物、非豆科植物和旱生型植物(张文彦等, 2010 ), 草本植物比乔木和灌丛具有更高的净光合速率和光合氮利用效率(Wright et al ., 2005 ).Wright等(2005)在对全球植物叶片普遍特征的关系研究中还发现, 一般而言, 草本植物比灌木具有更低的基于面积的氮含量和比叶质量.另外, 植物光合生理特征也会随着叶片生活史长短(Hikosaka & Hirose, 2000 )和植株的大小而变化(Liu et al ., 2010 ). ...
Cold, salinity and drought stresses: An overview.
1
2005
... 氮和水分是植物光合作用和生长必不可少的因子(Novriyanti et al ., 2012 ), 光合氮利用效率和水分利用效率可以预测氮素和水分是如何最优化地增加净光合速率的(Castellanos et al ., 2005 ), 资源匮乏的环境中物种会采用资源保护的特征如高的资源利用效率(RUE )来增加自身的竞争力(Funk & Vitousek, 2007 ).当土壤供水不足时, 植物通过降低G s 控制气体交换及调节光化学过程等生理活动降低净光合速率, 以此来减少自身对水分的需求(Mahajan & Narendra, 2005 ), 本研究中, 随着演替的进行, 群落表层土壤水分及全氮呈增长趋势(D群落由于坡度较大, 土壤含水量较低)(图4 ), 优势种WUE 逐渐降低(图1A ), 尽管D群落土壤含水量较低, 该群落优势种沙蒿、圆穗蓼的WUE 却没有因此升高, 崔骁勇等(2000) 的研究表明沙蒿通过对土壤干旱和高温的忍耐机制而保持长时间较高的光能利用效率, 依靠高蒸腾速率和强的水分吸收能力来提高自己在群落中的竞争力, 这种靠增加G s 获得高的光合速率和PNUE 的方式必然会导致WUE 的下降(Huang et al ., 2008 ); PNUE 没有随演替变化表现出一致的规律(图1B ), 而是与优势种叶片氮含量呈显著负相关(表3 ), Lambers等(2008) 的研究表明, 在能够支持植物生长的光强下, 氮含量较低的植物叶片的PNUE 较大, 这是因为这些植物合理地将叶片氮分配到光合中, 以达到最优的光合氮利用效率(Novriyanti et al ., 2012 ), 优势种披碱草正是利用这种策略占据B群落, 并在该群落中具有很高的重要值(表2 ).总之, 演替不同阶段优势种光合生理特征体现了其对不同环境的适应策略. ...
The use of the portable, non-destructive, SPAD-502 (Minolta) chlorophyll meter with leaves of varying trichome density and anthocyanin content.
1
1998
... 使用便携式叶绿素仪(SPAD-502, Minolta Camera, Osaka, Japan)测定物种叶片的SPAD 值, 选择植株中端成熟健康的叶片进行测定, 测定物种及重复数与测定光合特征值时的一致.SPAD-502是在比较650 nm和940 nm波长光线投射率的基础上测定的(Manetas et al ., 1998 ), 所测叶绿素含量是一个相对值. ...
Comparison of gas exchange traits of different plant species in Hunshandak sand area.
1
2003
... 植物光合生理特征能体现植物对不同生境的适应, 反映植物对不同环境变化的响应(Jiang & He, 1999 ).不同物种的光合生理特征间有显著的差异(Bassow & Bazzaz, 1997 ), 这正是不同植物利用不同的生态策略获得资源利用的反映(Niu et al ., 2003 ).一般而言, C4 光合途径或具有固氮能力的物种比C3 途径和无固氮能力的植物具备较高的光合能力和光合氮利用效率(Jiang & He, 1999 ), 因为氮含量与植物光合能力有非常密切的联系(Evans, 1989 ).双子叶植物、非禾本科植物、豆科植物和中生型植物的叶片氮含量分别高于相应的单子叶植物、禾本科植物、非豆科植物和旱生型植物(张文彦等, 2010 ), 草本植物比乔木和灌丛具有更高的净光合速率和光合氮利用效率(Wright et al ., 2005 ).Wright等(2005)在对全球植物叶片普遍特征的关系研究中还发现, 一般而言, 草本植物比灌木具有更低的基于面积的氮含量和比叶质量.另外, 植物光合生理特征也会随着叶片生活史长短(Hikosaka & Hirose, 2000 )和植株的大小而变化(Liu et al ., 2010 ). ...
Photosynthetic nitrogen and water use efficiency of acacia and eucalypt seedlings as afforestation species.
3
2012
... 影响植物光合能力的环境因子包括光照、水分、温度、营养物质和CO2 浓度等(于贵瑞和王秋凤, 2010 ), 而水分和氮是植物光合和生长必不可少的因子(Novriyanti et al ., 2012 ), Shangguan等(2000) 研究发现, 水分和氮素亏缺均会强烈地抑制植物的光合活动, 且氮素和水分对光合的影响是相互制约的.干旱大大降低了单叶和群落的净光合速率(崔骁勇等, 2000 ), 而提高了植物的光合水分利用效率(严昌荣等, 2001 ).An和Shangguan (2008)对不同光照和不同浓度氮素下白车轴草(Trifolium repens )幼苗光合生理特征的研究发现, 低浓度氮下基于面积的CO2 同化速率(A area )、光合氮利用效率(PNUE )和叶绿素含量随着氮素浓度的升高而增加; 高浓度氮下, 这些光合特征值会下降, 而比叶面积(SLA )和基于质量的叶片氮含量(N mass )会随着氮素浓度的升高持续增加.另外, 海拔通过影响CO2 分压、温度、水分等环境因子而影响植物光合生理特征, Ran等(2013) 将岷江冷杉(Abies faxoniana )移植到低海拔处, 降低了岷江冷杉的叶氮含量和水分利用效率, 而增加了净光合速率和PNUE . ...
... 氮和水分是植物光合作用和生长必不可少的因子(Novriyanti et al ., 2012 ), 光合氮利用效率和水分利用效率可以预测氮素和水分是如何最优化地增加净光合速率的(Castellanos et al ., 2005 ), 资源匮乏的环境中物种会采用资源保护的特征如高的资源利用效率(RUE )来增加自身的竞争力(Funk & Vitousek, 2007 ).当土壤供水不足时, 植物通过降低G s 控制气体交换及调节光化学过程等生理活动降低净光合速率, 以此来减少自身对水分的需求(Mahajan & Narendra, 2005 ), 本研究中, 随着演替的进行, 群落表层土壤水分及全氮呈增长趋势(D群落由于坡度较大, 土壤含水量较低)(图4 ), 优势种WUE 逐渐降低(图1A ), 尽管D群落土壤含水量较低, 该群落优势种沙蒿、圆穗蓼的WUE 却没有因此升高, 崔骁勇等(2000) 的研究表明沙蒿通过对土壤干旱和高温的忍耐机制而保持长时间较高的光能利用效率, 依靠高蒸腾速率和强的水分吸收能力来提高自己在群落中的竞争力, 这种靠增加G s 获得高的光合速率和PNUE 的方式必然会导致WUE 的下降(Huang et al ., 2008 ); PNUE 没有随演替变化表现出一致的规律(图1B ), 而是与优势种叶片氮含量呈显著负相关(表3 ), Lambers等(2008) 的研究表明, 在能够支持植物生长的光强下, 氮含量较低的植物叶片的PNUE 较大, 这是因为这些植物合理地将叶片氮分配到光合中, 以达到最优的光合氮利用效率(Novriyanti et al ., 2012 ), 优势种披碱草正是利用这种策略占据B群落, 并在该群落中具有很高的重要值(表2 ).总之, 演替不同阶段优势种光合生理特征体现了其对不同环境的适应策略. ...
... 较大, 这是因为这些植物合理地将叶片氮分配到光合中, 以达到最优的光合氮利用效率(Novriyanti et al ., 2012 ), 优势种披碱草正是利用这种策略占据B群落, 并在该群落中具有很高的重要值(表2 ).总之, 演替不同阶段优势种光合生理特征体现了其对不同环境的适应策略. ...
Altitudinal variation in growth, photosynthetic capacity and water use efficiency of Abies faxoniana Rehd. et Wils. seedlings as revealed by reciprocal transplantations.
0
2013
Effects of nitrogen nutrition and water deficit on net photosynthetic rate and chlorophyll fluorescence in winter wheat.
0
2000
草地退化演替研究进展
1
1999
... 高寒草甸是青藏高原最主要的生态系统类型(周华坤等, 2008 ), 由于长期忽略了对草地资源的科学管理, 粗放经营、超载放牧以及对草地资源不合理的开发利用, 使得高寒草地在结构上沙化, 功能上生产力降低, 生物多样性和生态系统功能严重丧失(武高林和杜国祯, 2007 ).关于高寒草甸恢复演替的研究已有很多, 放牧干扰及种间竞争对草地群落演替具有最大的影响(孙海群等, 1999 ).围封消除了放牧干扰, 围封地植物光合生理特征可以很好地反映退化草地自然恢复过程各阶段群落光合动态及其对环境因子的响应.尽管植物光合生理特征对环境变化的响应在国内外有大量详细的研究, 但由于技术设备等原因的限制, 高寒草甸植物光合作用对环境适应机理的研究依然较少, 尤其缺乏从物种到群落水平上的研究, 而这方面的研究对于植物适应日益剧增的放牧、围栏封育等土地利用方式极其重要.本文在判定亚高山草甸围封地各群落演替阶段的基础上, 通过对植物光合生理特征和表层土壤养分的探讨, 掌握了围封地恢复演替不同阶段的光合生理特征动态及其主要驱动因子, 对亚高寒退化草甸围封恢复具有一定的理论指导意义. ...
草地退化演替研究进展
1
1999
... 高寒草甸是青藏高原最主要的生态系统类型(周华坤等, 2008 ), 由于长期忽略了对草地资源的科学管理, 粗放经营、超载放牧以及对草地资源不合理的开发利用, 使得高寒草地在结构上沙化, 功能上生产力降低, 生物多样性和生态系统功能严重丧失(武高林和杜国祯, 2007 ).关于高寒草甸恢复演替的研究已有很多, 放牧干扰及种间竞争对草地群落演替具有最大的影响(孙海群等, 1999 ).围封消除了放牧干扰, 围封地植物光合生理特征可以很好地反映退化草地自然恢复过程各阶段群落光合动态及其对环境因子的响应.尽管植物光合生理特征对环境变化的响应在国内外有大量详细的研究, 但由于技术设备等原因的限制, 高寒草甸植物光合作用对环境适应机理的研究依然较少, 尤其缺乏从物种到群落水平上的研究, 而这方面的研究对于植物适应日益剧增的放牧、围栏封育等土地利用方式极其重要.本文在判定亚高山草甸围封地各群落演替阶段的基础上, 通过对植物光合生理特征和表层土壤养分的探讨, 掌握了围封地恢复演替不同阶段的光合生理特征动态及其主要驱动因子, 对亚高寒退化草甸围封恢复具有一定的理论指导意义. ...
Plant traits and soil chemical variables during a secondary vegetation succession in abandoned fields on the loess plateau.
1
2002
... 排除了不同种的差异后, 同种植物在不同演替阶段光合生理特征的研究更能体现植物对演替的响应.植被演替不同阶段由于群落组成及结构不同而导致各种环境因子的变化.首先, 植物群落演替对土壤理化性质产生一定的影响, 如土壤全氮随植被演替的进行呈增加趋势(Wang, 2002 ); 其次, 不同演替阶段群落的结构不同, 导致垂直方向上光照的空间异质性, 演替早期是一个阳性的开放系统, 植物能够获得更高的相对光量子通量而具有较大的光合速率(Anten et al ., 1998a ).所有这些环境因子的改变都会导致植物光合生理特征的变化.本研究中, 披碱草和老鹳草在演替前期(A)光合能力较弱(图3 ), 可能是受氮限制的原因, 安慧和上官周平(2007) 发现在低氮含量下白车轴草幼苗的光合能力会随着氮素的添加而增加, 而在高氮素含量下光合能力又会下降.在一个土壤水分、全氮含量逐渐升高的演替序列中(图4 ), 氮和水分对植物的限制作用逐渐减小, 非豆科植物披碱草、老鹳草的PNUE 、WUE 逐渐降低, 叶片N mass 逐渐升高(图3 ), 反映了不同物种通过对营养物质的不同分配策略来保护匮乏资源, 而豆科植物紫苜蓿的这些特征值却没有明显的变化, 在整个演替进程中紫苜蓿保持了较大的SLA (图3 ), 低SLA 植物能够更好地适应资源贫瘠和干旱的环境, 而高SLA 的植物保持体内营养物质的能力较强(李玉霖等, 2005 ), 这可能就是演替中养分、水分对紫苜蓿没有显著影响的原因, 紫苜蓿与其他两个非豆科植物一样, 随着演替的进行A area 、SPAD 有所下降.在养分和水分影响不大的情况下, 光照可能是导致其光合能力发生变化的主要原因, 光照会控制叶氮的分布(Anten et al ., 1998b ), 进而影响叶片的光合能力. ...
Assessing the generality of global leaf trait relationships.
1
2005
... 植物光合生理特征能体现植物对不同生境的适应, 反映植物对不同环境变化的响应(Jiang & He, 1999 ).不同物种的光合生理特征间有显著的差异(Bassow & Bazzaz, 1997 ), 这正是不同植物利用不同的生态策略获得资源利用的反映(Niu et al ., 2003 ).一般而言, C4 光合途径或具有固氮能力的物种比C3 途径和无固氮能力的植物具备较高的光合能力和光合氮利用效率(Jiang & He, 1999 ), 因为氮含量与植物光合能力有非常密切的联系(Evans, 1989 ).双子叶植物、非禾本科植物、豆科植物和中生型植物的叶片氮含量分别高于相应的单子叶植物、禾本科植物、非豆科植物和旱生型植物(张文彦等, 2010 ), 草本植物比乔木和灌丛具有更高的净光合速率和光合氮利用效率(Wright et al ., 2005 ).Wright等(2005)在对全球植物叶片普遍特征的关系研究中还发现, 一般而言, 草本植物比灌木具有更低的基于面积的氮含量和比叶质量.另外, 植物光合生理特征也会随着叶片生活史长短(Hikosaka & Hirose, 2000 )和植株的大小而变化(Liu et al ., 2010 ). ...
青藏高原退化高寒草地生态系统恢复和可持续发展探讨
1
2007
... 高寒草甸是青藏高原最主要的生态系统类型(周华坤等, 2008 ), 由于长期忽略了对草地资源的科学管理, 粗放经营、超载放牧以及对草地资源不合理的开发利用, 使得高寒草地在结构上沙化, 功能上生产力降低, 生物多样性和生态系统功能严重丧失(武高林和杜国祯, 2007 ).关于高寒草甸恢复演替的研究已有很多, 放牧干扰及种间竞争对草地群落演替具有最大的影响(孙海群等, 1999 ).围封消除了放牧干扰, 围封地植物光合生理特征可以很好地反映退化草地自然恢复过程各阶段群落光合动态及其对环境因子的响应.尽管植物光合生理特征对环境变化的响应在国内外有大量详细的研究, 但由于技术设备等原因的限制, 高寒草甸植物光合作用对环境适应机理的研究依然较少, 尤其缺乏从物种到群落水平上的研究, 而这方面的研究对于植物适应日益剧增的放牧、围栏封育等土地利用方式极其重要.本文在判定亚高山草甸围封地各群落演替阶段的基础上, 通过对植物光合生理特征和表层土壤养分的探讨, 掌握了围封地恢复演替不同阶段的光合生理特征动态及其主要驱动因子, 对亚高寒退化草甸围封恢复具有一定的理论指导意义. ...
青藏高原退化高寒草地生态系统恢复和可持续发展探讨
1
2007
... 高寒草甸是青藏高原最主要的生态系统类型(周华坤等, 2008 ), 由于长期忽略了对草地资源的科学管理, 粗放经营、超载放牧以及对草地资源不合理的开发利用, 使得高寒草地在结构上沙化, 功能上生产力降低, 生物多样性和生态系统功能严重丧失(武高林和杜国祯, 2007 ).关于高寒草甸恢复演替的研究已有很多, 放牧干扰及种间竞争对草地群落演替具有最大的影响(孙海群等, 1999 ).围封消除了放牧干扰, 围封地植物光合生理特征可以很好地反映退化草地自然恢复过程各阶段群落光合动态及其对环境因子的响应.尽管植物光合生理特征对环境变化的响应在国内外有大量详细的研究, 但由于技术设备等原因的限制, 高寒草甸植物光合作用对环境适应机理的研究依然较少, 尤其缺乏从物种到群落水平上的研究, 而这方面的研究对于植物适应日益剧增的放牧、围栏封育等土地利用方式极其重要.本文在判定亚高山草甸围封地各群落演替阶段的基础上, 通过对植物光合生理特征和表层土壤养分的探讨, 掌握了围封地恢复演替不同阶段的光合生理特征动态及其主要驱动因子, 对亚高寒退化草甸围封恢复具有一定的理论指导意义. ...
六种木本植物水分利用效率和其小生境关系研究
1
2001
... 影响植物光合能力的环境因子包括光照、水分、温度、营养物质和CO2 浓度等(于贵瑞和王秋凤, 2010 ), 而水分和氮是植物光合和生长必不可少的因子(Novriyanti et al ., 2012 ), Shangguan等(2000) 研究发现, 水分和氮素亏缺均会强烈地抑制植物的光合活动, 且氮素和水分对光合的影响是相互制约的.干旱大大降低了单叶和群落的净光合速率(崔骁勇等, 2000 ), 而提高了植物的光合水分利用效率(严昌荣等, 2001 ).An和Shangguan (2008)对不同光照和不同浓度氮素下白车轴草(Trifolium repens )幼苗光合生理特征的研究发现, 低浓度氮下基于面积的CO2 同化速率(A area )、光合氮利用效率(PNUE )和叶绿素含量随着氮素浓度的升高而增加; 高浓度氮下, 这些光合特征值会下降, 而比叶面积(SLA )和基于质量的叶片氮含量(N mass )会随着氮素浓度的升高持续增加.另外, 海拔通过影响CO2 分压、温度、水分等环境因子而影响植物光合生理特征, Ran等(2013) 将岷江冷杉(Abies faxoniana )移植到低海拔处, 降低了岷江冷杉的叶氮含量和水分利用效率, 而增加了净光合速率和PNUE . ...
六种木本植物水分利用效率和其小生境关系研究
1
2001
... 影响植物光合能力的环境因子包括光照、水分、温度、营养物质和CO2 浓度等(于贵瑞和王秋凤, 2010 ), 而水分和氮是植物光合和生长必不可少的因子(Novriyanti et al ., 2012 ), Shangguan等(2000) 研究发现, 水分和氮素亏缺均会强烈地抑制植物的光合活动, 且氮素和水分对光合的影响是相互制约的.干旱大大降低了单叶和群落的净光合速率(崔骁勇等, 2000 ), 而提高了植物的光合水分利用效率(严昌荣等, 2001 ).An和Shangguan (2008)对不同光照和不同浓度氮素下白车轴草(Trifolium repens )幼苗光合生理特征的研究发现, 低浓度氮下基于面积的CO2 同化速率(A area )、光合氮利用效率(PNUE )和叶绿素含量随着氮素浓度的升高而增加; 高浓度氮下, 这些光合特征值会下降, 而比叶面积(SLA )和基于质量的叶片氮含量(N mass )会随着氮素浓度的升高持续增加.另外, 海拔通过影响CO2 分压、温度、水分等环境因子而影响植物光合生理特征, Ran等(2013) 将岷江冷杉(Abies faxoniana )移植到低海拔处, 降低了岷江冷杉的叶氮含量和水分利用效率, 而增加了净光合速率和PNUE . ...
1
2010
... 影响植物光合能力的环境因子包括光照、水分、温度、营养物质和CO2 浓度等(于贵瑞和王秋凤, 2010 ), 而水分和氮是植物光合和生长必不可少的因子(Novriyanti et al ., 2012 ), Shangguan等(2000) 研究发现, 水分和氮素亏缺均会强烈地抑制植物的光合活动, 且氮素和水分对光合的影响是相互制约的.干旱大大降低了单叶和群落的净光合速率(崔骁勇等, 2000 ), 而提高了植物的光合水分利用效率(严昌荣等, 2001 ).An和Shangguan (2008)对不同光照和不同浓度氮素下白车轴草(Trifolium repens )幼苗光合生理特征的研究发现, 低浓度氮下基于面积的CO2 同化速率(A area )、光合氮利用效率(PNUE )和叶绿素含量随着氮素浓度的升高而增加; 高浓度氮下, 这些光合特征值会下降, 而比叶面积(SLA )和基于质量的叶片氮含量(N mass )会随着氮素浓度的升高持续增加.另外, 海拔通过影响CO2 分压、温度、水分等环境因子而影响植物光合生理特征, Ran等(2013) 将岷江冷杉(Abies faxoniana )移植到低海拔处, 降低了岷江冷杉的叶氮含量和水分利用效率, 而增加了净光合速率和PNUE . ...
1
2010
... 影响植物光合能力的环境因子包括光照、水分、温度、营养物质和CO2 浓度等(于贵瑞和王秋凤, 2010 ), 而水分和氮是植物光合和生长必不可少的因子(Novriyanti et al ., 2012 ), Shangguan等(2000) 研究发现, 水分和氮素亏缺均会强烈地抑制植物的光合活动, 且氮素和水分对光合的影响是相互制约的.干旱大大降低了单叶和群落的净光合速率(崔骁勇等, 2000 ), 而提高了植物的光合水分利用效率(严昌荣等, 2001 ).An和Shangguan (2008)对不同光照和不同浓度氮素下白车轴草(Trifolium repens )幼苗光合生理特征的研究发现, 低浓度氮下基于面积的CO2 同化速率(A area )、光合氮利用效率(PNUE )和叶绿素含量随着氮素浓度的升高而增加; 高浓度氮下, 这些光合特征值会下降, 而比叶面积(SLA )和基于质量的叶片氮含量(N mass )会随着氮素浓度的升高持续增加.另外, 海拔通过影响CO2 分压、温度、水分等环境因子而影响植物光合生理特征, Ran等(2013) 将岷江冷杉(Abies faxoniana )移植到低海拔处, 降低了岷江冷杉的叶氮含量和水分利用效率, 而增加了净光合速率和PNUE . ...
亚高山草甸弃耕地植物群落演替的数量研究I. 群落组成分析
1
1988
... 实验地是一块围封多年的草地, 位于甘肃省甘南藏族自治州合作市附近的兰州大学高寒草甸与湿地生态系统定位研究站, 地理位置34.93° N, 102.88° E, 海拔2900-3000 m.年平均气温2.0 ℃, 最冷月(12-2月)平均气温-8.9 ℃, 最热月(6-8月)平均气温11.5 ℃, 0 ℃以上年积温1730 ℃.年降水量560 mm, 50%以上集中在6-8月份, 雨热同季.土壤类型为亚高山草甸土, 以密丛型禾本科和莎草科植物为主, 混以其他杂草类(张大勇等, 1988 ). ...
亚高山草甸弃耕地植物群落演替的数量研究I. 群落组成分析
1
1988
... 实验地是一块围封多年的草地, 位于甘肃省甘南藏族自治州合作市附近的兰州大学高寒草甸与湿地生态系统定位研究站, 地理位置34.93° N, 102.88° E, 海拔2900-3000 m.年平均气温2.0 ℃, 最冷月(12-2月)平均气温-8.9 ℃, 最热月(6-8月)平均气温11.5 ℃, 0 ℃以上年积温1730 ℃.年降水量560 mm, 50%以上集中在6-8月份, 雨热同季.土壤类型为亚高山草甸土, 以密丛型禾本科和莎草科植物为主, 混以其他杂草类(张大勇等, 1988 ). ...
中国典型草原优势植物功能群氮磷化学计量学特征研究
1
2010
... 植物光合生理特征能体现植物对不同生境的适应, 反映植物对不同环境变化的响应(Jiang & He, 1999 ).不同物种的光合生理特征间有显著的差异(Bassow & Bazzaz, 1997 ), 这正是不同植物利用不同的生态策略获得资源利用的反映(Niu et al ., 2003 ).一般而言, C4 光合途径或具有固氮能力的物种比C3 途径和无固氮能力的植物具备较高的光合能力和光合氮利用效率(Jiang & He, 1999 ), 因为氮含量与植物光合能力有非常密切的联系(Evans, 1989 ).双子叶植物、非禾本科植物、豆科植物和中生型植物的叶片氮含量分别高于相应的单子叶植物、禾本科植物、非豆科植物和旱生型植物(张文彦等, 2010 ), 草本植物比乔木和灌丛具有更高的净光合速率和光合氮利用效率(Wright et al ., 2005 ).Wright等(2005)在对全球植物叶片普遍特征的关系研究中还发现, 一般而言, 草本植物比灌木具有更低的基于面积的氮含量和比叶质量.另外, 植物光合生理特征也会随着叶片生活史长短(Hikosaka & Hirose, 2000 )和植株的大小而变化(Liu et al ., 2010 ). ...
中国典型草原优势植物功能群氮磷化学计量学特征研究
1
2010
... 植物光合生理特征能体现植物对不同生境的适应, 反映植物对不同环境变化的响应(Jiang & He, 1999 ).不同物种的光合生理特征间有显著的差异(Bassow & Bazzaz, 1997 ), 这正是不同植物利用不同的生态策略获得资源利用的反映(Niu et al ., 2003 ).一般而言, C4 光合途径或具有固氮能力的物种比C3 途径和无固氮能力的植物具备较高的光合能力和光合氮利用效率(Jiang & He, 1999 ), 因为氮含量与植物光合能力有非常密切的联系(Evans, 1989 ).双子叶植物、非禾本科植物、豆科植物和中生型植物的叶片氮含量分别高于相应的单子叶植物、禾本科植物、非豆科植物和旱生型植物(张文彦等, 2010 ), 草本植物比乔木和灌丛具有更高的净光合速率和光合氮利用效率(Wright et al ., 2005 ).Wright等(2005)在对全球植物叶片普遍特征的关系研究中还发现, 一般而言, 草本植物比灌木具有更低的基于面积的氮含量和比叶质量.另外, 植物光合生理特征也会随着叶片生活史长短(Hikosaka & Hirose, 2000 )和植株的大小而变化(Liu et al ., 2010 ). ...
青藏高原高寒草甸生态系统的恢复能力
1
2008
... 高寒草甸是青藏高原最主要的生态系统类型(周华坤等, 2008 ), 由于长期忽略了对草地资源的科学管理, 粗放经营、超载放牧以及对草地资源不合理的开发利用, 使得高寒草地在结构上沙化, 功能上生产力降低, 生物多样性和生态系统功能严重丧失(武高林和杜国祯, 2007 ).关于高寒草甸恢复演替的研究已有很多, 放牧干扰及种间竞争对草地群落演替具有最大的影响(孙海群等, 1999 ).围封消除了放牧干扰, 围封地植物光合生理特征可以很好地反映退化草地自然恢复过程各阶段群落光合动态及其对环境因子的响应.尽管植物光合生理特征对环境变化的响应在国内外有大量详细的研究, 但由于技术设备等原因的限制, 高寒草甸植物光合作用对环境适应机理的研究依然较少, 尤其缺乏从物种到群落水平上的研究, 而这方面的研究对于植物适应日益剧增的放牧、围栏封育等土地利用方式极其重要.本文在判定亚高山草甸围封地各群落演替阶段的基础上, 通过对植物光合生理特征和表层土壤养分的探讨, 掌握了围封地恢复演替不同阶段的光合生理特征动态及其主要驱动因子, 对亚高寒退化草甸围封恢复具有一定的理论指导意义. ...
青藏高原高寒草甸生态系统的恢复能力
1
2008
... 高寒草甸是青藏高原最主要的生态系统类型(周华坤等, 2008 ), 由于长期忽略了对草地资源的科学管理, 粗放经营、超载放牧以及对草地资源不合理的开发利用, 使得高寒草地在结构上沙化, 功能上生产力降低, 生物多样性和生态系统功能严重丧失(武高林和杜国祯, 2007 ).关于高寒草甸恢复演替的研究已有很多, 放牧干扰及种间竞争对草地群落演替具有最大的影响(孙海群等, 1999 ).围封消除了放牧干扰, 围封地植物光合生理特征可以很好地反映退化草地自然恢复过程各阶段群落光合动态及其对环境因子的响应.尽管植物光合生理特征对环境变化的响应在国内外有大量详细的研究, 但由于技术设备等原因的限制, 高寒草甸植物光合作用对环境适应机理的研究依然较少, 尤其缺乏从物种到群落水平上的研究, 而这方面的研究对于植物适应日益剧增的放牧、围栏封育等土地利用方式极其重要.本文在判定亚高山草甸围封地各群落演替阶段的基础上, 通过对植物光合生理特征和表层土壤养分的探讨, 掌握了围封地恢复演替不同阶段的光合生理特征动态及其主要驱动因子, 对亚高寒退化草甸围封恢复具有一定的理论指导意义. ...
 , 贾芸, 安琪, 安嫣菲
, 贾芸, 安琪, 安嫣菲


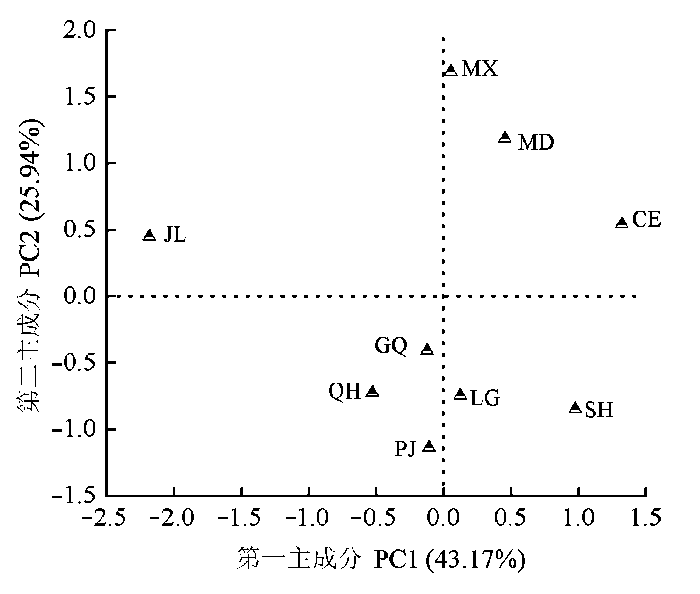
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}