Investigating tree water use efficiency (WUE) is important for understanding the coupling of carbon and water cycles in terrestrial ecosystems and its responses and adaptation to climatic change. Dahurian larch (Larix gmelinii), the dominant tree species in the Chinese boreal forest, plays an important role in the regional carbon budget. In this study, we measured the foliar WUE and associated physiological parameters of 30-year-old Dahurian larch trees from 17 provenances with divergent climatic conditions in a common garden. Our specific aims were to compare differences in WUE among the provenances and explore whether the observed differences are attributed to potential adaptation or acclimation to local habitats.
Methods
We have measured net photosynthetic rate (Pn), transpiration rate (Tr), stomatal conductance (Gs), and leaf nitrogen concentration (NL) of three replicate trees per provenance from mid June to mid September of 2010. WUE was calculated as the ratio of Pn to Tr. Specific leaf area (SLA) was calculated as one-side projected leaf area divided by the leaf dry mass. The aridity index (AI) of each provenance was calculated as the ratio of mean annual evaporation to mean annual precipitation.
Important findings
WUE, Pn, Gs, Tr, SLA and NL all differed significantly among the provenances. WUE showed a significant exponential relationship with Gs. WUE increased significantly with the increase of Gs when the Gs was less than 0.2, and was relatively stable when the Gs was greater than 0.2. WUE was significantly and positively correlated with NL, but negatively with SLA. The degree of these correlations increased with increasing AI values of the tree origins. WUE was negatively correlated with the mean annual temperature and mean annual precipitation of the tree origins, but positively correlated with the AI of the tree origins. Tr and Pn were positively correlated with both mean annual temperature and AI of the tree origins, these results suggest that the trees may adapt to the local climatic conditions of their origins, which results in the significant difference in the needle morphological and physiological properties, and thus WUE among the provenances of Dahurian larch trees.
Keywords:Larix gmelinii
;
water use efficiency
;
photosynthetic physiology
;
aridity index
;
climatic change
QUANXian-Kui, WANGChuan-Kuan. Comparison of foliar water use efficiency among 17 provenances of Larix gmelinii in the Mao’ershan area. Chinese Journal of Plant Ecology, 2015, 39(4): 352-361 https://doi.org/10.17521/cjpe.2015.0034
本实验所在的同质种植园位于东北林业大学帽儿山森林生态系统研究站内(45.40° N, 127.67° E)。在国内落叶松的自然分布范围内选取代表性种源17个(表1), 于1980年秋采种, 1981年育苗, 1983年春将17个种源的二年实生苗栽种在生态站试验样地内。样地设计按完全随机区组设计, 重复5次, 80株小区设计, 双行排列, 按株行距1.5 m × 2.0 m定植, 四周设有保护行(杨传平等, 2002)。1997年和2001年间伐两次, 株行距现为4.5 m × 2.5 m。该地区气候属于大陆性温带季风气候, 1989-2009年平均年降水量为629 mm, 平均年蒸发量为864 mm, 年平均气温3.1 ℃, 1月和7月份平均气温分别为-18.5 ℃和22 ℃。同质园内土壤、地形、气候条件一致, 平均海拔300 m, 平均坡度10°-15°, 土壤为暗棕色森林土。
Table 1
表1
表1 落叶松17个种源原地的地理和气候条件
Table 1 The geographic and climatic conditions of the origins of the 17 provenances of Larix gmelinii
种源 Provenance
代码 Code
纬度Latitude (N)
经度Longitude (E)
年平均气温 MAT (℃)
平均年降水量 MAP (mm)
平均年蒸发量 MAE (mm)
干燥度 AI
≥10 ℃积温CT (℃)
相对湿度 RH (%)
友好 Youhao
1
47.80°
128.83°
-1.05
622.0
955.8
1.54
2 147.2
78
乌伊岭 Wuyiling
2
48.67°
129.42°
-1.19
584.6
1 038.0
1.78
1 851.2
73
新林 Xinlin
3
51.70°
124.33°
-3.35
507.3
912.0
1.80
1 535.3
69
桦皮窑 Huapiyao
4
50.70°
126.72°
-1.50
539.0
1 100.0
2.04
1 900.0
68
三站 Sanzhan
5
49.62°
126.80°
-1.30
527.2
1 076.0
2.04
1 650.0
70
十八站 Shibazhan
6
52.42°
125.27°
-2.03
480.0
999.1
2.08
1 680.0
70
沾河 Zhanhe
7
48.45°
126.50°
0.12
501.7
1 057.0
2.11
2 442.3
70
塔河 Tahe
8
52.32°
124.72°
-2.74
487.9
1 028.0
2.11
1 671.4
68
莫尔道嘎 Moerdaoga
9
51.25°
120.58°
-4.50
471.0
999.4
2.12
1 485.0
70
甘河 Ganhe
10
50.58°
123.22°
-2.50
470.0
1 059.0
2.25
1 616.8
68
根河 Genhe
11
50.68°
121.95°
-5.00
436.3
991.4
2.27
1 297.6
70
鹤北 Hebei
12
47.55°
130.42°
1.85
530.0
1 235.0
2.33
2 456.0
74
满归 Mangui
13
52.05°
122.18°
-5.83
466.0
1 100.0
2.36
1 550.0
72
绰尔 Chuoer
14
48.17°
121.25°
-3.40
462.3
1 122.0
2.43
1 238.4
70
库都尔 Kuerdu
15
49.78°
121.88°
-4.00
500.0
1 280.0
2.56
1 739.5
67
阿尔山 Aershan
16
47.17°
119.95°
-3.30
425.1
1 100.0
2.59
1 354.3
70
中央站 Zhongyangzhan
17
50.75°
125.20°
-2.20
484.4
1 387.0
2.86
1 780.0
69
Meteorological data (1976-1985 average) of original sites were obtained from Yang et al. (2002) and China Meteorological Administration. AI, aridity index; CT, cumulative temperature above 10 °C; MAE, mean annual evaporation; MAP, mean annual precipitation; MAT, mean annual temperature; RH, relative humility.气象数据(1976-1985)来自杨传平等(2002)和国家气象局资料。
图1 落叶松针叶净光合速率(Pn)、蒸腾速率(Tr)、水分利用效率(WUE)、气孔导度(Gs)、比叶面积(SLA)和叶氮含量(NL)的种源比较(平均值±标准误差, n = 9)。不同字母表示种源间差异显著(p < 0.05)。种源代码同表1。
Fig. 1 Comparisons of foliar net photosynthetic rate (Pn), transpiration rate (Tr), water use efficiency (WUE), stomatal conductance (Gs), special leaf area (SLA), and leaf nitrogen concentration (NL) of Larix gmelinii trees originating from 17 provenances (mean ± SE, n = 9). Different letters indicate significant differences among provenances (p < 0.05). Provenance codes see Table 1.
Fig. 2 Relationships between foliar water use efficiency (WUE) and stomatal conductance (Gs), special leaf area (SLA), and leaf nitrogen concentration (NL) for the Larix gmelinii trees within different groups of the aridity index (AI).
Fig. 3 Foliar water use efficiency (WUE), transpiration rate (Tr), and net photosynthetic rate (Pn) for 17 Larix gmelinii provenances in relation to mean annual temperature (MAT), mean annual precipitation (MAP) and aridity index (AI) of the origins.
de MiguelM, Sánchez-GómezD, CerveraMT, ArandaI (2012).
Functional and genetic characterization of gas exchange and intrinsic water use efficiency in a full-sib family of Pinus pinaster Ait. in response to drought.
Evidence of changing intrinsic water-use efficiency under rising atmospheric CO2 concentrations in boreal fennoscandia from subfossil leaves and tree ring δ13C ratios.
High leaf mass per area of related species assemblages may reflect low rainfall and carbon isotope discrimination rather than low phosphorus and nitrogen concentrations.
Productivity, water-use efficiency and tolerance to moderate water deficit correlate in 33 poplar genotypes from a Populus deltoides × Populus trichocarpa F1 progeny.
The characteristics of foliar δ13C values of plants and plant water use efficiency indicated by δ13C values in two fragmented rain forests in Xishuangbanna, Yunnan.
Acta Botanica Sinica, 43, 186-192.(in Chinese with English abstract)
Altitudinal variation in growth, photosynthetic capacity and water use efficiency of Abies faxoniana Rehd. et Wils. seedlings as revealed by reciprocal transplantations.
Photosynthesis and respiration rates depend on leaf and root morphology and nitrogen concentration in nine boreal tree species differing in relative growth rate.
Enhanced assimilation rate and water use efficiency with latitude through increased photosynthetic capacity and internal conductance in balsam poplar (Populus balsamifera L.).
Coupling of respiration, nitrogen, and sugars underlies convergent temperature acclimation in Pinus banksiana across wide-ranging sites and populations.
Does rainfall explain variation in leaf morphology and physiology among populations of red ironbark (Eucalyptus sideroxylon subsp. tricarpa) grown in a common garden?
Climate warming and increasing atmospheric CO2 have contributed to increased intrinsic water-use efficiency on the northeastern Tibetan Plateau since 1850.
Interactions between CO2 enhancement and N addition on net primary productivity and water-use efficiency in a mesocosm with multiple subtropical tree species.
A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests.
1
2010
... 树木在长期的进化和选择过程中已形成通过改变生理、分子以及遗传等方面特征去响应环境改变的能力, 然而在目前全球气候变化加剧的背景下, 树木能否通过这种能力的改变去适应未来快速的气候变化已成为研究的热点问题(Saxe et al., 2001; Allen et al., 2010; Kremer et al., 2012; Shaw & Etterson, 2012; Reyer et al., 2013).研究同质种植园内同一树种不同种源树木的差异及其和种源原地气候条件的关系是一种验证树木对气候适应/驯化的重要方法(Reich et al., 1996; Warren et al., 2005; Martin et al., 2007; Frei et al., 2012). ...
Eucalypts forming a canopy functional type in dry sclerophyll forests respond differentially to environment.
1
2000
... 与本文假设相同的是, 来自于低温少雨地区的种源具有较高的WUE, 并且和种源原地年平均气温及平均年降水量均呈线性负相关关系(图3G、3H).这是因为随着温度的升高叶片光合作用酶的活性增强、Gs增大, 进而影响CO2同化速率和Tr, 导致WUE的差异.本研究中17个种源落叶松的Pn和Tr均存在显著的种间差异, 但随着种源原地温度的升高, Tr明显增大, 而Pn总体上并没有表现出明显的增大趋势, 仅在一定的降水区间内和温度存在一定的线性相关(图3A、3D).因此, 本研究中树木Tr的增大是导致WUE和种源原地年平均气温负相关的主要原因.温度和树木WUE间这种负相关性在相近的研究中也有所体现(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).大量种源试验表明树木在干旱环境条件下具有更高的WUE (Anderson et al., 2000; Warren & Adams, 2000; Gouveia & Freitas, 2009; Ran et al., 2013), 这是由于干旱环境下空气湿度和土壤含水量降低、叶片Gs和Tr减小, 而Pn的变化相对较小, 从而导致WUE增大(Farquhar et al., 1989; Zhu et al., 2011; Liu et al., 2013).然而, 本研究结果显示Pn和Tr均与种源原地平均年降水量不相关(图3B、3E), 这是因为降水只考虑了水分收入, 未考虑水分的支出等情况, 不能完全表征一个地方的干旱程度.Warren等(2005)对同质园内来自于水分条件差异明显的15个种源Eucalyptus sideroxylon树木研究表明叶片WUE存在种源差异, 但和降水没有明显相关性, 相似的结果也在对不同种源Eucalyptus microtheca的研究中出现(Li, 2000).因此选用一个较合理的代表气候干旱程度的指标尤为重要, AI作为表征一个地区干湿程度的指标, 近来成为全球变化研究中经常涉及的气候指标之一(Arora, 2002; 孟猛等, 2004).本研究结果表明落叶松针叶WUE和AI显著正相关且相关性高于降水和温度(图3), 这与Warren和Adams (2000)对海岸松4个种源树木的研究结果相一致.此外, AI和Pn也存在线性正相关关系, 并且和不同水分蒸发范围内的Tr存在显著相关关系.这表明干旱情况下Pn的增加和Tr的减小是WUE提高的主要原因(Ramírez-Valiente et al., 2011). ...
The use of the aridity index to assess climate change effect on annual runoff.
1
2002
... 与本文假设相同的是, 来自于低温少雨地区的种源具有较高的WUE, 并且和种源原地年平均气温及平均年降水量均呈线性负相关关系(图3G、3H).这是因为随着温度的升高叶片光合作用酶的活性增强、Gs增大, 进而影响CO2同化速率和Tr, 导致WUE的差异.本研究中17个种源落叶松的Pn和Tr均存在显著的种间差异, 但随着种源原地温度的升高, Tr明显增大, 而Pn总体上并没有表现出明显的增大趋势, 仅在一定的降水区间内和温度存在一定的线性相关(图3A、3D).因此, 本研究中树木Tr的增大是导致WUE和种源原地年平均气温负相关的主要原因.温度和树木WUE间这种负相关性在相近的研究中也有所体现(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).大量种源试验表明树木在干旱环境条件下具有更高的WUE (Anderson et al., 2000; Warren & Adams, 2000; Gouveia & Freitas, 2009; Ran et al., 2013), 这是由于干旱环境下空气湿度和土壤含水量降低、叶片Gs和Tr减小, 而Pn的变化相对较小, 从而导致WUE增大(Farquhar et al., 1989; Zhu et al., 2011; Liu et al., 2013).然而, 本研究结果显示Pn和Tr均与种源原地平均年降水量不相关(图3B、3E), 这是因为降水只考虑了水分收入, 未考虑水分的支出等情况, 不能完全表征一个地方的干旱程度.Warren等(2005)对同质园内来自于水分条件差异明显的15个种源Eucalyptus sideroxylon树木研究表明叶片WUE存在种源差异, 但和降水没有明显相关性, 相似的结果也在对不同种源Eucalyptus microtheca的研究中出现(Li, 2000).因此选用一个较合理的代表气候干旱程度的指标尤为重要, AI作为表征一个地区干湿程度的指标, 近来成为全球变化研究中经常涉及的气候指标之一(Arora, 2002; 孟猛等, 2004).本研究结果表明落叶松针叶WUE和AI显著正相关且相关性高于降水和温度(图3), 这与Warren和Adams (2000)对海岸松4个种源树木的研究结果相一致.此外, AI和Pn也存在线性正相关关系, 并且和不同水分蒸发范围内的Tr存在显著相关关系.这表明干旱情况下Pn的增加和Tr的减小是WUE提高的主要原因(Ramírez-Valiente et al., 2011). ...
To what extent is altitudinal variation of functional traits driven by genetic adaptation in european oak and beech?
1
2011
... 本研究结果表明落叶松NL和WUE间存在显著的正相关关系, 并且来自于高AI地区的种源树木WUE对NL具有更高的敏感性(图2C).有研究表明: 来自低温少雨地区种源的树木具有较高的NL, 以此提高光合能力获取更多的光合产物, 从而适应低温干燥的环境并且从贫瘠的土壤中获取更多的养分和水分(Reich et al., 1996; Weih & Karlsson, 2001; Oleksyn et al., 2003; Tjoelker et al., 2008; Bresson et al., 2011).因此, 树木NL同WUE间多表现为正相关关系(Sparks & Ehleringer, 1997; Monclus et al., 2009; Yan et al., 2014).相同NL水平下来自于高AI种源树木具有更高的WUE, 这对该树种能够在多样的环境中生存具有重要的现实意义.这是因为在干旱贫瘠的环境中土壤氮和水是树木生长的限制因子, 通过提高叶片氮的利用效率获得更多的光合产物、提高WUE, 从而有助于其生长和生存(Kenney et al., 2014; Yan et al., 2014).此外, 在模拟未来气候变化对树木生长乃至生存的影响时, 研究者认为气候的变化将导致落叶松分布区内永冻层融解萎缩, 土壤干燥, 湿润度下降, 最终使得落叶松林不适应新环境而逐步北移, 直至出境(张新时, 1993; 蒋延玲和周广胜, 2001; 李峰等, 2006).本研究发现落叶松树木可以通过调节叶片结构、养分含量、光合生理等形态和生理特征来调节WUE而适应干旱的环境, 这对研究该树种在气候变化下的应对策略和对新环境的适应能力具有重要的参考价值. ...
Carbon isotope composition of boreal plants: Functional grouping of life forms.
1
1997
... 水分利用效率(WUE)是反映植物单位耗水量生产出的同化量的一个重要生理生态参数(Farquhar & Richards, 1984), 是耦合植物叶片光合作用和蒸腾作用的重要指标, 对研究生态系统水循环和碳循环相互作用、揭示植物对全球变化响应和适应对策具有重要意义(Saxe et al., 2001; Gao et al., 2014).气候变化条件下的树木WUE的响应及其机理已成为全球生态学研究的热点(Monclus et al., 2009; de Miguel et al., 2012; Klein et al., 2013).温度和降水是影响WUE的两个重要环境因子.有研究认为当温度升高后光合酶活性提高, 气孔导度(Gs)减小, 从而导致WUE增大(Morecroft et al., 1992; Xiao et al., 2013; Xu et al., 2013); 也有研究认为温度升高会导致胞间CO2浓度(Ci)减小、蒸腾速率(Tr)增大, WUE反而减小(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).降水通过改变土壤水分可利用性和空气湿度, 从而影响树木的气孔关闭和Tr, 进而改变树木的WUE.然而降水和WUE之间关系的研究结果也存在一些不确定性.有研究结果表明二者呈正相关关系(Schulze et al., 1998; Picotte et al., 2007; Niu et al., 2008; Klein et al., 2013); 也有研究认为二者呈负相关关系(Gimenez et al., 1992; Schulze et al., 2006; Monclus et al., 2009; Zhu et al., 2011).此外, 树种间由于生物学特性的差异导致其对环境的适应方式有所不同, 从而引起WUE种间差异.有研究表明, 相同环境下常绿乔木的WUE大于落叶乔木(Brooks et al., 1997; Kloeppel et al., 1998), 而有些研究却得出相反的结果(Damesin et al., 1997; 渠春梅等, 2001).因此, 环境变化对树木WUE的影响以及树木WUE对环境变化响应的内在机理还有待于深入研究. ...
Between-tree variations in leaf δ13C of Quercus pubescens and Quercus ilex among Mediterranean habitats with different water availability.
1
1997
... 水分利用效率(WUE)是反映植物单位耗水量生产出的同化量的一个重要生理生态参数(Farquhar & Richards, 1984), 是耦合植物叶片光合作用和蒸腾作用的重要指标, 对研究生态系统水循环和碳循环相互作用、揭示植物对全球变化响应和适应对策具有重要意义(Saxe et al., 2001; Gao et al., 2014).气候变化条件下的树木WUE的响应及其机理已成为全球生态学研究的热点(Monclus et al., 2009; de Miguel et al., 2012; Klein et al., 2013).温度和降水是影响WUE的两个重要环境因子.有研究认为当温度升高后光合酶活性提高, 气孔导度(Gs)减小, 从而导致WUE增大(Morecroft et al., 1992; Xiao et al., 2013; Xu et al., 2013); 也有研究认为温度升高会导致胞间CO2浓度(Ci)减小、蒸腾速率(Tr)增大, WUE反而减小(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).降水通过改变土壤水分可利用性和空气湿度, 从而影响树木的气孔关闭和Tr, 进而改变树木的WUE.然而降水和WUE之间关系的研究结果也存在一些不确定性.有研究结果表明二者呈正相关关系(Schulze et al., 1998; Picotte et al., 2007; Niu et al., 2008; Klein et al., 2013); 也有研究认为二者呈负相关关系(Gimenez et al., 1992; Schulze et al., 2006; Monclus et al., 2009; Zhu et al., 2011).此外, 树种间由于生物学特性的差异导致其对环境的适应方式有所不同, 从而引起WUE种间差异.有研究表明, 相同环境下常绿乔木的WUE大于落叶乔木(Brooks et al., 1997; Kloeppel et al., 1998), 而有些研究却得出相反的结果(Damesin et al., 1997; 渠春梅等, 2001).因此, 环境变化对树木WUE的影响以及树木WUE对环境变化响应的内在机理还有待于深入研究. ...
Functional and genetic characterization of gas exchange and intrinsic water use efficiency in a full-sib family of Pinus pinaster Ait. in response to drought.
2
2012
... 水分利用效率(WUE)是反映植物单位耗水量生产出的同化量的一个重要生理生态参数(Farquhar & Richards, 1984), 是耦合植物叶片光合作用和蒸腾作用的重要指标, 对研究生态系统水循环和碳循环相互作用、揭示植物对全球变化响应和适应对策具有重要意义(Saxe et al., 2001; Gao et al., 2014).气候变化条件下的树木WUE的响应及其机理已成为全球生态学研究的热点(Monclus et al., 2009; de Miguel et al., 2012; Klein et al., 2013).温度和降水是影响WUE的两个重要环境因子.有研究认为当温度升高后光合酶活性提高, 气孔导度(Gs)减小, 从而导致WUE增大(Morecroft et al., 1992; Xiao et al., 2013; Xu et al., 2013); 也有研究认为温度升高会导致胞间CO2浓度(Ci)减小、蒸腾速率(Tr)增大, WUE反而减小(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).降水通过改变土壤水分可利用性和空气湿度, 从而影响树木的气孔关闭和Tr, 进而改变树木的WUE.然而降水和WUE之间关系的研究结果也存在一些不确定性.有研究结果表明二者呈正相关关系(Schulze et al., 1998; Picotte et al., 2007; Niu et al., 2008; Klein et al., 2013); 也有研究认为二者呈负相关关系(Gimenez et al., 1992; Schulze et al., 2006; Monclus et al., 2009; Zhu et al., 2011).此外, 树种间由于生物学特性的差异导致其对环境的适应方式有所不同, 从而引起WUE种间差异.有研究表明, 相同环境下常绿乔木的WUE大于落叶乔木(Brooks et al., 1997; Kloeppel et al., 1998), 而有些研究却得出相反的结果(Damesin et al., 1997; 渠春梅等, 2001).因此, 环境变化对树木WUE的影响以及树木WUE对环境变化响应的内在机理还有待于深入研究. ...
... SLA作为叶片结构的重要特征之一也是影响WUE的重要因素, 有研究表明随着SLA的减小, 叶片厚度增加、光合组织密度增大, 导致水分的蒸腾减小和固碳能力提高, 从而WUE增大, 因此两者间多表现为负相关关系(Reich et al., 1998; Lamont et al., 2002; Warren et al., 2005; Gago et al., 2014).目前对于两者间关系的研究多集中在树种间(Lamont et al., 2002; Holtum & Winter, 2005)、同一树种不同种源幼苗间(Fernández et al., 1999; Warren et al., 2005)、同一种源不同幼苗个体间(Monclus et al., 2009; Roussel et al., 2009; de Miguel et al., 2012)等方面的表型差异, 而对环境变化下两者间的关系及其调控机理研究较少.de Miguel等(2012)对全同胞家系海岸松幼苗研究发现SLA和WUE存在负相关关系, 并认为这是基因调控的结果并能通过子代遗传, 相似的结果在其他研究中也有所体现(Fernández et al., 1999; Ramírez-Valiente et al., 2011).和我们假设相一致的是, 17个种源落叶松的30年树木SLA具有明显的种源差异并和WUE间存在显著的负相关关系(p < 0.05), 且随着种源原地AI的增大SLA对WUE的影响程度增强(图2B).这种在同质环境中生活多年仍存在显著的种源间差异且和种源原地气候条件有较好关系的结果进一步支持了这种差异是一种基因调控下的对种源原地环境长期适应的结果. ...
Carbon isotope discrimination and photosynthesis.
2
1989
... Gs作为叶片同外界进行水分和气体交换的主要通道, 调节着碳同化速率和水分消耗间的平衡(Gagen et al., 2011).在一定范围内, 当Gs升高时叶片Ci增加, 导致Pn增大, 虽然Tr也增大, 但光合速率增大的幅度较大, 从而Gs和WUE呈一定的正相关关系(Picotte et al., 2007; Gago et al., 2014), 但随着Gs的继续增大, 叶片在通过气孔得到CO2和失去水分间达到平衡, 此时WUE的限制因素不再是Gs, 而转为光合相关的酶含量和活性等其他生理因素(Farquhar et al., 1989; Gimenez et al., 1992).本试验结果表明在起始阶段WUE和Gs呈明显线性相关, 而当Gs增大到0.2左右时WUE趋于稳定, 但来自于较干旱地方的种源树木在相同WUE水平时需要较低的Gs (图2A), 这验证了我们假设干旱条件下树木通过减小Gs提高WUE的假设.通过较小的Gs获得较大的WUE可以使树木在干旱的条件下减少水分的蒸腾损失, 调节树木水势变化, 避免脱水, 从而提高植物对土壤水分的有效利用(Kenney et al., 2014).来自不同水分条件地区种源树木在同质园内生长30年后, 还存有与原地气候相适应的Gs对WUE的凋节方式, 这表现出明显的子代遗传效应.de Miguel等(2012)通过对来自全胞家系海岸松(Pinus pinaster)无性系苗木的研究也取得了相似的结果. ...
... 与本文假设相同的是, 来自于低温少雨地区的种源具有较高的WUE, 并且和种源原地年平均气温及平均年降水量均呈线性负相关关系(图3G、3H).这是因为随着温度的升高叶片光合作用酶的活性增强、Gs增大, 进而影响CO2同化速率和Tr, 导致WUE的差异.本研究中17个种源落叶松的Pn和Tr均存在显著的种间差异, 但随着种源原地温度的升高, Tr明显增大, 而Pn总体上并没有表现出明显的增大趋势, 仅在一定的降水区间内和温度存在一定的线性相关(图3A、3D).因此, 本研究中树木Tr的增大是导致WUE和种源原地年平均气温负相关的主要原因.温度和树木WUE间这种负相关性在相近的研究中也有所体现(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).大量种源试验表明树木在干旱环境条件下具有更高的WUE (Anderson et al., 2000; Warren & Adams, 2000; Gouveia & Freitas, 2009; Ran et al., 2013), 这是由于干旱环境下空气湿度和土壤含水量降低、叶片Gs和Tr减小, 而Pn的变化相对较小, 从而导致WUE增大(Farquhar et al., 1989; Zhu et al., 2011; Liu et al., 2013).然而, 本研究结果显示Pn和Tr均与种源原地平均年降水量不相关(图3B、3E), 这是因为降水只考虑了水分收入, 未考虑水分的支出等情况, 不能完全表征一个地方的干旱程度.Warren等(2005)对同质园内来自于水分条件差异明显的15个种源Eucalyptus sideroxylon树木研究表明叶片WUE存在种源差异, 但和降水没有明显相关性, 相似的结果也在对不同种源Eucalyptus microtheca的研究中出现(Li, 2000).因此选用一个较合理的代表气候干旱程度的指标尤为重要, AI作为表征一个地区干湿程度的指标, 近来成为全球变化研究中经常涉及的气候指标之一(Arora, 2002; 孟猛等, 2004).本研究结果表明落叶松针叶WUE和AI显著正相关且相关性高于降水和温度(图3), 这与Warren和Adams (2000)对海岸松4个种源树木的研究结果相一致.此外, AI和Pn也存在线性正相关关系, 并且和不同水分蒸发范围内的Tr存在显著相关关系.这表明干旱情况下Pn的增加和Tr的减小是WUE提高的主要原因(Ramírez-Valiente et al., 2011). ...
Isotopic composition of plant carbon correlates with water-use efficiency of wheat genotypes.
1
1984
... 水分利用效率(WUE)是反映植物单位耗水量生产出的同化量的一个重要生理生态参数(Farquhar & Richards, 1984), 是耦合植物叶片光合作用和蒸腾作用的重要指标, 对研究生态系统水循环和碳循环相互作用、揭示植物对全球变化响应和适应对策具有重要意义(Saxe et al., 2001; Gao et al., 2014).气候变化条件下的树木WUE的响应及其机理已成为全球生态学研究的热点(Monclus et al., 2009; de Miguel et al., 2012; Klein et al., 2013).温度和降水是影响WUE的两个重要环境因子.有研究认为当温度升高后光合酶活性提高, 气孔导度(Gs)减小, 从而导致WUE增大(Morecroft et al., 1992; Xiao et al., 2013; Xu et al., 2013); 也有研究认为温度升高会导致胞间CO2浓度(Ci)减小、蒸腾速率(Tr)增大, WUE反而减小(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).降水通过改变土壤水分可利用性和空气湿度, 从而影响树木的气孔关闭和Tr, 进而改变树木的WUE.然而降水和WUE之间关系的研究结果也存在一些不确定性.有研究结果表明二者呈正相关关系(Schulze et al., 1998; Picotte et al., 2007; Niu et al., 2008; Klein et al., 2013); 也有研究认为二者呈负相关关系(Gimenez et al., 1992; Schulze et al., 2006; Monclus et al., 2009; Zhu et al., 2011).此外, 树种间由于生物学特性的差异导致其对环境的适应方式有所不同, 从而引起WUE种间差异.有研究表明, 相同环境下常绿乔木的WUE大于落叶乔木(Brooks et al., 1997; Kloeppel et al., 1998), 而有些研究却得出相反的结果(Damesin et al., 1997; 渠春梅等, 2001).因此, 环境变化对树木WUE的影响以及树木WUE对环境变化响应的内在机理还有待于深入研究. ...
Response of Pinus pinaster Ait. provenances at early age to water supply. I. Water relation parameters.
2
1999
... SLA作为叶片结构的重要特征之一也是影响WUE的重要因素, 有研究表明随着SLA的减小, 叶片厚度增加、光合组织密度增大, 导致水分的蒸腾减小和固碳能力提高, 从而WUE增大, 因此两者间多表现为负相关关系(Reich et al., 1998; Lamont et al., 2002; Warren et al., 2005; Gago et al., 2014).目前对于两者间关系的研究多集中在树种间(Lamont et al., 2002; Holtum & Winter, 2005)、同一树种不同种源幼苗间(Fernández et al., 1999; Warren et al., 2005)、同一种源不同幼苗个体间(Monclus et al., 2009; Roussel et al., 2009; de Miguel et al., 2012)等方面的表型差异, 而对环境变化下两者间的关系及其调控机理研究较少.de Miguel等(2012)对全同胞家系海岸松幼苗研究发现SLA和WUE存在负相关关系, 并认为这是基因调控的结果并能通过子代遗传, 相似的结果在其他研究中也有所体现(Fernández et al., 1999; Ramírez-Valiente et al., 2011).和我们假设相一致的是, 17个种源落叶松的30年树木SLA具有明显的种源差异并和WUE间存在显著的负相关关系(p < 0.05), 且随着种源原地AI的增大SLA对WUE的影响程度增强(图2B).这种在同质环境中生活多年仍存在显著的种源间差异且和种源原地气候条件有较好关系的结果进一步支持了这种差异是一种基因调控下的对种源原地环境长期适应的结果. ...
... 存在负相关关系, 并认为这是基因调控的结果并能通过子代遗传, 相似的结果在其他研究中也有所体现(Fernández et al., 1999; Ramírez-Valiente et al., 2011).和我们假设相一致的是, 17个种源落叶松的30年树木SLA具有明显的种源差异并和WUE间存在显著的负相关关系(p < 0.05), 且随着种源原地AI的增大SLA对WUE的影响程度增强(图2B).这种在同质环境中生活多年仍存在显著的种源间差异且和种源原地气候条件有较好关系的结果进一步支持了这种差异是一种基因调控下的对种源原地环境长期适应的结果. ...
Phenotypic differentiation in a common garden reflects the phylogeography of a widespread alpine plant.
2
2012
... 作为WUE的两大组分, Pn的增大或Tr的减小都会导致WUE的增大.本研究中落叶松叶片Pn存在显著的种源差异, 并且与种源原地的AI有显著的正相关性(表1; 图1, 图3), 这和以往对来自不同纬度种源树木研究结果相似(Soolanayakanahally et al., 2009; Robson et al., 2012).较高的叶Gs、氮含量和较小的SLA导致了较高的Pn, 并且Pn和种源原地的气候条件具有明显的相关关系, 这是基因调控下的Pn对种源原地环境长期适应的结果(Martin et al., 2007; Savolainen et al., 2007; Soolanayakanahally et al., 2009; Frei et al., 2012).除友好种源外, 随着Pn的增大, WUE表现出相近的变化趋势; 虽然叶片Tr也存在显著的种源差异, 然而WUE并没有随着Tr的变化表现出相应的变化趋势(图1), 这表明WUE受 Pn的影响较大.从研究结果变异程度而言, Pn的变异系数为15.95%, 最高值是最低值的1.60倍, 而Tr的变异系数为9.6%, 最高值较最低值高出44.99%, 由此使得WUE的变异趋势和Pn相近.这是因为叶片的其他生理和结构特征, 如叶片的Gs、NL、SLA等直接影响Pn和Tr, 进而引起了WUE的差异(Gago et al., 2014). ...
... 树木在长期的进化和选择过程中已形成通过改变生理、分子以及遗传等方面特征去响应环境改变的能力, 然而在目前全球气候变化加剧的背景下, 树木能否通过这种能力的改变去适应未来快速的气候变化已成为研究的热点问题(Saxe et al., 2001; Allen et al., 2010; Kremer et al., 2012; Shaw & Etterson, 2012; Reyer et al., 2013).研究同质种植园内同一树种不同种源树木的差异及其和种源原地气候条件的关系是一种验证树木对气候适应/驯化的重要方法(Reich et al., 1996; Warren et al., 2005; Martin et al., 2007; Frei et al., 2012). ...
Evidence of changing intrinsic water-use efficiency under rising atmospheric CO2 concentrations in boreal fennoscandia from subfossil leaves and tree ring δ13C ratios.
1
2011
... Gs作为叶片同外界进行水分和气体交换的主要通道, 调节着碳同化速率和水分消耗间的平衡(Gagen et al., 2011).在一定范围内, 当Gs升高时叶片Ci增加, 导致Pn增大, 虽然Tr也增大, 但光合速率增大的幅度较大, 从而Gs和WUE呈一定的正相关关系(Picotte et al., 2007; Gago et al., 2014), 但随着Gs的继续增大, 叶片在通过气孔得到CO2和失去水分间达到平衡, 此时WUE的限制因素不再是Gs, 而转为光合相关的酶含量和活性等其他生理因素(Farquhar et al., 1989; Gimenez et al., 1992).本试验结果表明在起始阶段WUE和Gs呈明显线性相关, 而当Gs增大到0.2左右时WUE趋于稳定, 但来自于较干旱地方的种源树木在相同WUE水平时需要较低的Gs (图2A), 这验证了我们假设干旱条件下树木通过减小Gs提高WUE的假设.通过较小的Gs获得较大的WUE可以使树木在干旱的条件下减少水分的蒸腾损失, 调节树木水势变化, 避免脱水, 从而提高植物对土壤水分的有效利用(Kenney et al., 2014).来自不同水分条件地区种源树木在同质园内生长30年后, 还存有与原地气候相适应的Gs对WUE的凋节方式, 这表现出明显的子代遗传效应.de Miguel等(2012)通过对来自全胞家系海岸松(Pinus pinaster)无性系苗木的研究也取得了相似的结果. ...
Opportunities for improving leaf water use efficiency under climate change conditions.
3
2014
... 作为WUE的两大组分, Pn的增大或Tr的减小都会导致WUE的增大.本研究中落叶松叶片Pn存在显著的种源差异, 并且与种源原地的AI有显著的正相关性(表1; 图1, 图3), 这和以往对来自不同纬度种源树木研究结果相似(Soolanayakanahally et al., 2009; Robson et al., 2012).较高的叶Gs、氮含量和较小的SLA导致了较高的Pn, 并且Pn和种源原地的气候条件具有明显的相关关系, 这是基因调控下的Pn对种源原地环境长期适应的结果(Martin et al., 2007; Savolainen et al., 2007; Soolanayakanahally et al., 2009; Frei et al., 2012).除友好种源外, 随着Pn的增大, WUE表现出相近的变化趋势; 虽然叶片Tr也存在显著的种源差异, 然而WUE并没有随着Tr的变化表现出相应的变化趋势(图1), 这表明WUE受 Pn的影响较大.从研究结果变异程度而言, Pn的变异系数为15.95%, 最高值是最低值的1.60倍, 而Tr的变异系数为9.6%, 最高值较最低值高出44.99%, 由此使得WUE的变异趋势和Pn相近.这是因为叶片的其他生理和结构特征, 如叶片的Gs、NL、SLA等直接影响Pn和Tr, 进而引起了WUE的差异(Gago et al., 2014). ...
... Gs作为叶片同外界进行水分和气体交换的主要通道, 调节着碳同化速率和水分消耗间的平衡(Gagen et al., 2011).在一定范围内, 当Gs升高时叶片Ci增加, 导致Pn增大, 虽然Tr也增大, 但光合速率增大的幅度较大, 从而Gs和WUE呈一定的正相关关系(Picotte et al., 2007; Gago et al., 2014), 但随着Gs的继续增大, 叶片在通过气孔得到CO2和失去水分间达到平衡, 此时WUE的限制因素不再是Gs, 而转为光合相关的酶含量和活性等其他生理因素(Farquhar et al., 1989; Gimenez et al., 1992).本试验结果表明在起始阶段WUE和Gs呈明显线性相关, 而当Gs增大到0.2左右时WUE趋于稳定, 但来自于较干旱地方的种源树木在相同WUE水平时需要较低的Gs (图2A), 这验证了我们假设干旱条件下树木通过减小Gs提高WUE的假设.通过较小的Gs获得较大的WUE可以使树木在干旱的条件下减少水分的蒸腾损失, 调节树木水势变化, 避免脱水, 从而提高植物对土壤水分的有效利用(Kenney et al., 2014).来自不同水分条件地区种源树木在同质园内生长30年后, 还存有与原地气候相适应的Gs对WUE的凋节方式, 这表现出明显的子代遗传效应.de Miguel等(2012)通过对来自全胞家系海岸松(Pinus pinaster)无性系苗木的研究也取得了相似的结果. ...
... SLA作为叶片结构的重要特征之一也是影响WUE的重要因素, 有研究表明随着SLA的减小, 叶片厚度增加、光合组织密度增大, 导致水分的蒸腾减小和固碳能力提高, 从而WUE增大, 因此两者间多表现为负相关关系(Reich et al., 1998; Lamont et al., 2002; Warren et al., 2005; Gago et al., 2014).目前对于两者间关系的研究多集中在树种间(Lamont et al., 2002; Holtum & Winter, 2005)、同一树种不同种源幼苗间(Fernández et al., 1999; Warren et al., 2005)、同一种源不同幼苗个体间(Monclus et al., 2009; Roussel et al., 2009; de Miguel et al., 2012)等方面的表型差异, 而对环境变化下两者间的关系及其调控机理研究较少.de Miguel等(2012)对全同胞家系海岸松幼苗研究发现SLA和WUE存在负相关关系, 并认为这是基因调控的结果并能通过子代遗传, 相似的结果在其他研究中也有所体现(Fernández et al., 1999; Ramírez-Valiente et al., 2011).和我们假设相一致的是, 17个种源落叶松的30年树木SLA具有明显的种源差异并和WUE间存在显著的负相关关系(p < 0.05), 且随着种源原地AI的增大SLA对WUE的影响程度增强(图2B).这种在同质环境中生活多年仍存在显著的种源间差异且和种源原地气候条件有较好关系的结果进一步支持了这种差异是一种基因调控下的对种源原地环境长期适应的结果. ...
Water use efficiency threshold for terrestrial ecosystem carbon sequestration in China under afforestation.
1
2014
... 水分利用效率(WUE)是反映植物单位耗水量生产出的同化量的一个重要生理生态参数(Farquhar & Richards, 1984), 是耦合植物叶片光合作用和蒸腾作用的重要指标, 对研究生态系统水循环和碳循环相互作用、揭示植物对全球变化响应和适应对策具有重要意义(Saxe et al., 2001; Gao et al., 2014).气候变化条件下的树木WUE的响应及其机理已成为全球生态学研究的热点(Monclus et al., 2009; de Miguel et al., 2012; Klein et al., 2013).温度和降水是影响WUE的两个重要环境因子.有研究认为当温度升高后光合酶活性提高, 气孔导度(Gs)减小, 从而导致WUE增大(Morecroft et al., 1992; Xiao et al., 2013; Xu et al., 2013); 也有研究认为温度升高会导致胞间CO2浓度(Ci)减小、蒸腾速率(Tr)增大, WUE反而减小(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).降水通过改变土壤水分可利用性和空气湿度, 从而影响树木的气孔关闭和Tr, 进而改变树木的WUE.然而降水和WUE之间关系的研究结果也存在一些不确定性.有研究结果表明二者呈正相关关系(Schulze et al., 1998; Picotte et al., 2007; Niu et al., 2008; Klein et al., 2013); 也有研究认为二者呈负相关关系(Gimenez et al., 1992; Schulze et al., 2006; Monclus et al., 2009; Zhu et al., 2011).此外, 树种间由于生物学特性的差异导致其对环境的适应方式有所不同, 从而引起WUE种间差异.有研究表明, 相同环境下常绿乔木的WUE大于落叶乔木(Brooks et al., 1997; Kloeppel et al., 1998), 而有些研究却得出相反的结果(Damesin et al., 1997; 渠春梅等, 2001).因此, 环境变化对树木WUE的影响以及树木WUE对环境变化响应的内在机理还有待于深入研究. ...
Regulation of photosynthetic rate of two sunflower hybrids under water stress.
2
1992
... 水分利用效率(WUE)是反映植物单位耗水量生产出的同化量的一个重要生理生态参数(Farquhar & Richards, 1984), 是耦合植物叶片光合作用和蒸腾作用的重要指标, 对研究生态系统水循环和碳循环相互作用、揭示植物对全球变化响应和适应对策具有重要意义(Saxe et al., 2001; Gao et al., 2014).气候变化条件下的树木WUE的响应及其机理已成为全球生态学研究的热点(Monclus et al., 2009; de Miguel et al., 2012; Klein et al., 2013).温度和降水是影响WUE的两个重要环境因子.有研究认为当温度升高后光合酶活性提高, 气孔导度(Gs)减小, 从而导致WUE增大(Morecroft et al., 1992; Xiao et al., 2013; Xu et al., 2013); 也有研究认为温度升高会导致胞间CO2浓度(Ci)减小、蒸腾速率(Tr)增大, WUE反而减小(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).降水通过改变土壤水分可利用性和空气湿度, 从而影响树木的气孔关闭和Tr, 进而改变树木的WUE.然而降水和WUE之间关系的研究结果也存在一些不确定性.有研究结果表明二者呈正相关关系(Schulze et al., 1998; Picotte et al., 2007; Niu et al., 2008; Klein et al., 2013); 也有研究认为二者呈负相关关系(Gimenez et al., 1992; Schulze et al., 2006; Monclus et al., 2009; Zhu et al., 2011).此外, 树种间由于生物学特性的差异导致其对环境的适应方式有所不同, 从而引起WUE种间差异.有研究表明, 相同环境下常绿乔木的WUE大于落叶乔木(Brooks et al., 1997; Kloeppel et al., 1998), 而有些研究却得出相反的结果(Damesin et al., 1997; 渠春梅等, 2001).因此, 环境变化对树木WUE的影响以及树木WUE对环境变化响应的内在机理还有待于深入研究. ...
... Gs作为叶片同外界进行水分和气体交换的主要通道, 调节着碳同化速率和水分消耗间的平衡(Gagen et al., 2011).在一定范围内, 当Gs升高时叶片Ci增加, 导致Pn增大, 虽然Tr也增大, 但光合速率增大的幅度较大, 从而Gs和WUE呈一定的正相关关系(Picotte et al., 2007; Gago et al., 2014), 但随着Gs的继续增大, 叶片在通过气孔得到CO2和失去水分间达到平衡, 此时WUE的限制因素不再是Gs, 而转为光合相关的酶含量和活性等其他生理因素(Farquhar et al., 1989; Gimenez et al., 1992).本试验结果表明在起始阶段WUE和Gs呈明显线性相关, 而当Gs增大到0.2左右时WUE趋于稳定, 但来自于较干旱地方的种源树木在相同WUE水平时需要较低的Gs (图2A), 这验证了我们假设干旱条件下树木通过减小Gs提高WUE的假设.通过较小的Gs获得较大的WUE可以使树木在干旱的条件下减少水分的蒸腾损失, 调节树木水势变化, 避免脱水, 从而提高植物对土壤水分的有效利用(Kenney et al., 2014).来自不同水分条件地区种源树木在同质园内生长30年后, 还存有与原地气候相适应的Gs对WUE的凋节方式, 这表现出明显的子代遗传效应.de Miguel等(2012)通过对来自全胞家系海岸松(Pinus pinaster)无性系苗木的研究也取得了相似的结果. ...
Modulation of leaf attributes and water use efficiency in quercus suber along a rainfall gradient.
1
2009
... 与本文假设相同的是, 来自于低温少雨地区的种源具有较高的WUE, 并且和种源原地年平均气温及平均年降水量均呈线性负相关关系(图3G、3H).这是因为随着温度的升高叶片光合作用酶的活性增强、Gs增大, 进而影响CO2同化速率和Tr, 导致WUE的差异.本研究中17个种源落叶松的Pn和Tr均存在显著的种间差异, 但随着种源原地温度的升高, Tr明显增大, 而Pn总体上并没有表现出明显的增大趋势, 仅在一定的降水区间内和温度存在一定的线性相关(图3A、3D).因此, 本研究中树木Tr的增大是导致WUE和种源原地年平均气温负相关的主要原因.温度和树木WUE间这种负相关性在相近的研究中也有所体现(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).大量种源试验表明树木在干旱环境条件下具有更高的WUE (Anderson et al., 2000; Warren & Adams, 2000; Gouveia & Freitas, 2009; Ran et al., 2013), 这是由于干旱环境下空气湿度和土壤含水量降低、叶片Gs和Tr减小, 而Pn的变化相对较小, 从而导致WUE增大(Farquhar et al., 1989; Zhu et al., 2011; Liu et al., 2013).然而, 本研究结果显示Pn和Tr均与种源原地平均年降水量不相关(图3B、3E), 这是因为降水只考虑了水分收入, 未考虑水分的支出等情况, 不能完全表征一个地方的干旱程度.Warren等(2005)对同质园内来自于水分条件差异明显的15个种源Eucalyptus sideroxylon树木研究表明叶片WUE存在种源差异, 但和降水没有明显相关性, 相似的结果也在对不同种源Eucalyptus microtheca的研究中出现(Li, 2000).因此选用一个较合理的代表气候干旱程度的指标尤为重要, AI作为表征一个地区干湿程度的指标, 近来成为全球变化研究中经常涉及的气候指标之一(Arora, 2002; 孟猛等, 2004).本研究结果表明落叶松针叶WUE和AI显著正相关且相关性高于降水和温度(图3), 这与Warren和Adams (2000)对海岸松4个种源树木的研究结果相一致.此外, AI和Pn也存在线性正相关关系, 并且和不同水分蒸发范围内的Tr存在显著相关关系.这表明干旱情况下Pn的增加和Tr的减小是WUE提高的主要原因(Ramírez-Valiente et al., 2011). ...
Carbon isotope composition of canopy leaves in a tropical forest in panama throughout a seasonal cycle.
1
2005
... SLA作为叶片结构的重要特征之一也是影响WUE的重要因素, 有研究表明随着SLA的减小, 叶片厚度增加、光合组织密度增大, 导致水分的蒸腾减小和固碳能力提高, 从而WUE增大, 因此两者间多表现为负相关关系(Reich et al., 1998; Lamont et al., 2002; Warren et al., 2005; Gago et al., 2014).目前对于两者间关系的研究多集中在树种间(Lamont et al., 2002; Holtum & Winter, 2005)、同一树种不同种源幼苗间(Fernández et al., 1999; Warren et al., 2005)、同一种源不同幼苗个体间(Monclus et al., 2009; Roussel et al., 2009; de Miguel et al., 2012)等方面的表型差异, 而对环境变化下两者间的关系及其调控机理研究较少.de Miguel等(2012)对全同胞家系海岸松幼苗研究发现SLA和WUE存在负相关关系, 并认为这是基因调控的结果并能通过子代遗传, 相似的结果在其他研究中也有所体现(Fernández et al., 1999; Ramírez-Valiente et al., 2011).和我们假设相一致的是, 17个种源落叶松的30年树木SLA具有明显的种源差异并和WUE间存在显著的负相关关系(p < 0.05), 且随着种源原地AI的增大SLA对WUE的影响程度增强(图2B).这种在同质环境中生活多年仍存在显著的种源间差异且和种源原地气候条件有较好关系的结果进一步支持了这种差异是一种基因调控下的对种源原地环境长期适应的结果. ...
... 本研究结果表明落叶松NL和WUE间存在显著的正相关关系, 并且来自于高AI地区的种源树木WUE对NL具有更高的敏感性(图2C).有研究表明: 来自低温少雨地区种源的树木具有较高的NL, 以此提高光合能力获取更多的光合产物, 从而适应低温干燥的环境并且从贫瘠的土壤中获取更多的养分和水分(Reich et al., 1996; Weih & Karlsson, 2001; Oleksyn et al., 2003; Tjoelker et al., 2008; Bresson et al., 2011).因此, 树木NL同WUE间多表现为正相关关系(Sparks & Ehleringer, 1997; Monclus et al., 2009; Yan et al., 2014).相同NL水平下来自于高AI种源树木具有更高的WUE, 这对该树种能够在多样的环境中生存具有重要的现实意义.这是因为在干旱贫瘠的环境中土壤氮和水是树木生长的限制因子, 通过提高叶片氮的利用效率获得更多的光合产物、提高WUE, 从而有助于其生长和生存(Kenney et al., 2014; Yan et al., 2014).此外, 在模拟未来气候变化对树木生长乃至生存的影响时, 研究者认为气候的变化将导致落叶松分布区内永冻层融解萎缩, 土壤干燥, 湿润度下降, 最终使得落叶松林不适应新环境而逐步北移, 直至出境(张新时, 1993; 蒋延玲和周广胜, 2001; 李峰等, 2006).本研究发现落叶松树木可以通过调节叶片结构、养分含量、光合生理等形态和生理特征来调节WUE而适应干旱的环境, 这对研究该树种在气候变化下的应对策略和对新环境的适应能力具有重要的参考价值. ...
Direct and indirect selection on flowering time, water-use efficiency (WUE, δ13C), and WUE plasticity to drought in Arabidopsis thaliana.
2
2014
... Gs作为叶片同外界进行水分和气体交换的主要通道, 调节着碳同化速率和水分消耗间的平衡(Gagen et al., 2011).在一定范围内, 当Gs升高时叶片Ci增加, 导致Pn增大, 虽然Tr也增大, 但光合速率增大的幅度较大, 从而Gs和WUE呈一定的正相关关系(Picotte et al., 2007; Gago et al., 2014), 但随着Gs的继续增大, 叶片在通过气孔得到CO2和失去水分间达到平衡, 此时WUE的限制因素不再是Gs, 而转为光合相关的酶含量和活性等其他生理因素(Farquhar et al., 1989; Gimenez et al., 1992).本试验结果表明在起始阶段WUE和Gs呈明显线性相关, 而当Gs增大到0.2左右时WUE趋于稳定, 但来自于较干旱地方的种源树木在相同WUE水平时需要较低的Gs (图2A), 这验证了我们假设干旱条件下树木通过减小Gs提高WUE的假设.通过较小的Gs获得较大的WUE可以使树木在干旱的条件下减少水分的蒸腾损失, 调节树木水势变化, 避免脱水, 从而提高植物对土壤水分的有效利用(Kenney et al., 2014).来自不同水分条件地区种源树木在同质园内生长30年后, 还存有与原地气候相适应的Gs对WUE的凋节方式, 这表现出明显的子代遗传效应.de Miguel等(2012)通过对来自全胞家系海岸松(Pinus pinaster)无性系苗木的研究也取得了相似的结果. ...
... 本研究结果表明落叶松NL和WUE间存在显著的正相关关系, 并且来自于高AI地区的种源树木WUE对NL具有更高的敏感性(图2C).有研究表明: 来自低温少雨地区种源的树木具有较高的NL, 以此提高光合能力获取更多的光合产物, 从而适应低温干燥的环境并且从贫瘠的土壤中获取更多的养分和水分(Reich et al., 1996; Weih & Karlsson, 2001; Oleksyn et al., 2003; Tjoelker et al., 2008; Bresson et al., 2011).因此, 树木NL同WUE间多表现为正相关关系(Sparks & Ehleringer, 1997; Monclus et al., 2009; Yan et al., 2014).相同NL水平下来自于高AI种源树木具有更高的WUE, 这对该树种能够在多样的环境中生存具有重要的现实意义.这是因为在干旱贫瘠的环境中土壤氮和水是树木生长的限制因子, 通过提高叶片氮的利用效率获得更多的光合产物、提高WUE, 从而有助于其生长和生存(Kenney et al., 2014; Yan et al., 2014).此外, 在模拟未来气候变化对树木生长乃至生存的影响时, 研究者认为气候的变化将导致落叶松分布区内永冻层融解萎缩, 土壤干燥, 湿润度下降, 最终使得落叶松林不适应新环境而逐步北移, 直至出境(张新时, 1993; 蒋延玲和周广胜, 2001; 李峰等, 2006).本研究发现落叶松树木可以通过调节叶片结构、养分含量、光合生理等形态和生理特征来调节WUE而适应干旱的环境, 这对研究该树种在气候变化下的应对策略和对新环境的适应能力具有重要的参考价值. ...
Differential ecophysiological response of a major mediterranean pine species across a climatic gradient.
2
2013
... 水分利用效率(WUE)是反映植物单位耗水量生产出的同化量的一个重要生理生态参数(Farquhar & Richards, 1984), 是耦合植物叶片光合作用和蒸腾作用的重要指标, 对研究生态系统水循环和碳循环相互作用、揭示植物对全球变化响应和适应对策具有重要意义(Saxe et al., 2001; Gao et al., 2014).气候变化条件下的树木WUE的响应及其机理已成为全球生态学研究的热点(Monclus et al., 2009; de Miguel et al., 2012; Klein et al., 2013).温度和降水是影响WUE的两个重要环境因子.有研究认为当温度升高后光合酶活性提高, 气孔导度(Gs)减小, 从而导致WUE增大(Morecroft et al., 1992; Xiao et al., 2013; Xu et al., 2013); 也有研究认为温度升高会导致胞间CO2浓度(Ci)减小、蒸腾速率(Tr)增大, WUE反而减小(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).降水通过改变土壤水分可利用性和空气湿度, 从而影响树木的气孔关闭和Tr, 进而改变树木的WUE.然而降水和WUE之间关系的研究结果也存在一些不确定性.有研究结果表明二者呈正相关关系(Schulze et al., 1998; Picotte et al., 2007; Niu et al., 2008; Klein et al., 2013); 也有研究认为二者呈负相关关系(Gimenez et al., 1992; Schulze et al., 2006; Monclus et al., 2009; Zhu et al., 2011).此外, 树种间由于生物学特性的差异导致其对环境的适应方式有所不同, 从而引起WUE种间差异.有研究表明, 相同环境下常绿乔木的WUE大于落叶乔木(Brooks et al., 1997; Kloeppel et al., 1998), 而有些研究却得出相反的结果(Damesin et al., 1997; 渠春梅等, 2001).因此, 环境变化对树木WUE的影响以及树木WUE对环境变化响应的内在机理还有待于深入研究. ...
... ; Klein et al., 2013); 也有研究认为二者呈负相关关系(Gimenez et al., 1992; Schulze et al., 2006; Monclus et al., 2009; Zhu et al., 2011).此外, 树种间由于生物学特性的差异导致其对环境的适应方式有所不同, 从而引起WUE种间差异.有研究表明, 相同环境下常绿乔木的WUE大于落叶乔木(Brooks et al., 1997; Kloeppel et al., 1998), 而有些研究却得出相反的结果(Damesin et al., 1997; 渠春梅等, 2001).因此, 环境变化对树木WUE的影响以及树木WUE对环境变化响应的内在机理还有待于深入研究. ...
Foliar carbon isotope discrimination in Larix species and sympatric evergreen conifers: A global comparison.
1
1998
... 水分利用效率(WUE)是反映植物单位耗水量生产出的同化量的一个重要生理生态参数(Farquhar & Richards, 1984), 是耦合植物叶片光合作用和蒸腾作用的重要指标, 对研究生态系统水循环和碳循环相互作用、揭示植物对全球变化响应和适应对策具有重要意义(Saxe et al., 2001; Gao et al., 2014).气候变化条件下的树木WUE的响应及其机理已成为全球生态学研究的热点(Monclus et al., 2009; de Miguel et al., 2012; Klein et al., 2013).温度和降水是影响WUE的两个重要环境因子.有研究认为当温度升高后光合酶活性提高, 气孔导度(Gs)减小, 从而导致WUE增大(Morecroft et al., 1992; Xiao et al., 2013; Xu et al., 2013); 也有研究认为温度升高会导致胞间CO2浓度(Ci)减小、蒸腾速率(Tr)增大, WUE反而减小(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).降水通过改变土壤水分可利用性和空气湿度, 从而影响树木的气孔关闭和Tr, 进而改变树木的WUE.然而降水和WUE之间关系的研究结果也存在一些不确定性.有研究结果表明二者呈正相关关系(Schulze et al., 1998; Picotte et al., 2007; Niu et al., 2008; Klein et al., 2013); 也有研究认为二者呈负相关关系(Gimenez et al., 1992; Schulze et al., 2006; Monclus et al., 2009; Zhu et al., 2011).此外, 树种间由于生物学特性的差异导致其对环境的适应方式有所不同, 从而引起WUE种间差异.有研究表明, 相同环境下常绿乔木的WUE大于落叶乔木(Brooks et al., 1997; Kloeppel et al., 1998), 而有些研究却得出相反的结果(Damesin et al., 1997; 渠春梅等, 2001).因此, 环境变化对树木WUE的影响以及树木WUE对环境变化响应的内在机理还有待于深入研究. ...
Long-distance gene flow and adaptation of forest trees to rapid climate change.
1
2012
... 树木在长期的进化和选择过程中已形成通过改变生理、分子以及遗传等方面特征去响应环境改变的能力, 然而在目前全球气候变化加剧的背景下, 树木能否通过这种能力的改变去适应未来快速的气候变化已成为研究的热点问题(Saxe et al., 2001; Allen et al., 2010; Kremer et al., 2012; Shaw & Etterson, 2012; Reyer et al., 2013).研究同质种植园内同一树种不同种源树木的差异及其和种源原地气候条件的关系是一种验证树木对气候适应/驯化的重要方法(Reich et al., 1996; Warren et al., 2005; Martin et al., 2007; Frei et al., 2012). ...
High leaf mass per area of related species assemblages may reflect low rainfall and carbon isotope discrimination rather than low phosphorus and nitrogen concentrations.
2
2002
... SLA作为叶片结构的重要特征之一也是影响WUE的重要因素, 有研究表明随着SLA的减小, 叶片厚度增加、光合组织密度增大, 导致水分的蒸腾减小和固碳能力提高, 从而WUE增大, 因此两者间多表现为负相关关系(Reich et al., 1998; Lamont et al., 2002; Warren et al., 2005; Gago et al., 2014).目前对于两者间关系的研究多集中在树种间(Lamont et al., 2002; Holtum & Winter, 2005)、同一树种不同种源幼苗间(Fernández et al., 1999; Warren et al., 2005)、同一种源不同幼苗个体间(Monclus et al., 2009; Roussel et al., 2009; de Miguel et al., 2012)等方面的表型差异, 而对环境变化下两者间的关系及其调控机理研究较少.de Miguel等(2012)对全同胞家系海岸松幼苗研究发现SLA和WUE存在负相关关系, 并认为这是基因调控的结果并能通过子代遗传, 相似的结果在其他研究中也有所体现(Fernández et al., 1999; Ramírez-Valiente et al., 2011).和我们假设相一致的是, 17个种源落叶松的30年树木SLA具有明显的种源差异并和WUE间存在显著的负相关关系(p < 0.05), 且随着种源原地AI的增大SLA对WUE的影响程度增强(图2B).这种在同质环境中生活多年仍存在显著的种源间差异且和种源原地气候条件有较好关系的结果进一步支持了这种差异是一种基因调控下的对种源原地环境长期适应的结果. ...
... ).目前对于两者间关系的研究多集中在树种间(Lamont et al., 2002; Holtum & Winter, 2005)、同一树种不同种源幼苗间(Fernández et al., 1999; Warren et al., 2005)、同一种源不同幼苗个体间(Monclus et al., 2009; Roussel et al., 2009; de Miguel et al., 2012)等方面的表型差异, 而对环境变化下两者间的关系及其调控机理研究较少.de Miguel等(2012)对全同胞家系海岸松幼苗研究发现SLA和WUE存在负相关关系, 并认为这是基因调控的结果并能通过子代遗传, 相似的结果在其他研究中也有所体现(Fernández et al., 1999; Ramírez-Valiente et al., 2011).和我们假设相一致的是, 17个种源落叶松的30年树木SLA具有明显的种源差异并和WUE间存在显著的负相关关系(p < 0.05), 且随着种源原地AI的增大SLA对WUE的影响程度增强(图2B).这种在同质环境中生活多年仍存在显著的种源间差异且和种源原地气候条件有较好关系的结果进一步支持了这种差异是一种基因调控下的对种源原地环境长期适应的结果. ...
Population differences in water-use efficiency of Eucalyptus microtheca seedlings under different watering regimes.
1
2000
... 与本文假设相同的是, 来自于低温少雨地区的种源具有较高的WUE, 并且和种源原地年平均气温及平均年降水量均呈线性负相关关系(图3G、3H).这是因为随着温度的升高叶片光合作用酶的活性增强、Gs增大, 进而影响CO2同化速率和Tr, 导致WUE的差异.本研究中17个种源落叶松的Pn和Tr均存在显著的种间差异, 但随着种源原地温度的升高, Tr明显增大, 而Pn总体上并没有表现出明显的增大趋势, 仅在一定的降水区间内和温度存在一定的线性相关(图3A、3D).因此, 本研究中树木Tr的增大是导致WUE和种源原地年平均气温负相关的主要原因.温度和树木WUE间这种负相关性在相近的研究中也有所体现(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).大量种源试验表明树木在干旱环境条件下具有更高的WUE (Anderson et al., 2000; Warren & Adams, 2000; Gouveia & Freitas, 2009; Ran et al., 2013), 这是由于干旱环境下空气湿度和土壤含水量降低、叶片Gs和Tr减小, 而Pn的变化相对较小, 从而导致WUE增大(Farquhar et al., 1989; Zhu et al., 2011; Liu et al., 2013).然而, 本研究结果显示Pn和Tr均与种源原地平均年降水量不相关(图3B、3E), 这是因为降水只考虑了水分收入, 未考虑水分的支出等情况, 不能完全表征一个地方的干旱程度.Warren等(2005)对同质园内来自于水分条件差异明显的15个种源Eucalyptus sideroxylon树木研究表明叶片WUE存在种源差异, 但和降水没有明显相关性, 相似的结果也在对不同种源Eucalyptus microtheca的研究中出现(Li, 2000).因此选用一个较合理的代表气候干旱程度的指标尤为重要, AI作为表征一个地区干湿程度的指标, 近来成为全球变化研究中经常涉及的气候指标之一(Arora, 2002; 孟猛等, 2004).本研究结果表明落叶松针叶WUE和AI显著正相关且相关性高于降水和温度(图3), 这与Warren和Adams (2000)对海岸松4个种源树木的研究结果相一致.此外, AI和Pn也存在线性正相关关系, 并且和不同水分蒸发范围内的Tr存在显著相关关系.这表明干旱情况下Pn的增加和Tr的减小是WUE提高的主要原因(Ramírez-Valiente et al., 2011). ...
... 本研究结果表明落叶松NL和WUE间存在显著的正相关关系, 并且来自于高AI地区的种源树木WUE对NL具有更高的敏感性(图2C).有研究表明: 来自低温少雨地区种源的树木具有较高的NL, 以此提高光合能力获取更多的光合产物, 从而适应低温干燥的环境并且从贫瘠的土壤中获取更多的养分和水分(Reich et al., 1996; Weih & Karlsson, 2001; Oleksyn et al., 2003; Tjoelker et al., 2008; Bresson et al., 2011).因此, 树木NL同WUE间多表现为正相关关系(Sparks & Ehleringer, 1997; Monclus et al., 2009; Yan et al., 2014).相同NL水平下来自于高AI种源树木具有更高的WUE, 这对该树种能够在多样的环境中生存具有重要的现实意义.这是因为在干旱贫瘠的环境中土壤氮和水是树木生长的限制因子, 通过提高叶片氮的利用效率获得更多的光合产物、提高WUE, 从而有助于其生长和生存(Kenney et al., 2014; Yan et al., 2014).此外, 在模拟未来气候变化对树木生长乃至生存的影响时, 研究者认为气候的变化将导致落叶松分布区内永冻层融解萎缩, 土壤干燥, 湿润度下降, 最终使得落叶松林不适应新环境而逐步北移, 直至出境(张新时, 1993; 蒋延玲和周广胜, 2001; 李峰等, 2006).本研究发现落叶松树木可以通过调节叶片结构、养分含量、光合生理等形态和生理特征来调节WUE而适应干旱的环境, 这对研究该树种在气候变化下的应对策略和对新环境的适应能力具有重要的参考价值. ...
Effects of soil water and nitrogen availability on photosynthesis and water use efficiency of Robinia pseudoacacia seedlings.
1
2013
... 与本文假设相同的是, 来自于低温少雨地区的种源具有较高的WUE, 并且和种源原地年平均气温及平均年降水量均呈线性负相关关系(图3G、3H).这是因为随着温度的升高叶片光合作用酶的活性增强、Gs增大, 进而影响CO2同化速率和Tr, 导致WUE的差异.本研究中17个种源落叶松的Pn和Tr均存在显著的种间差异, 但随着种源原地温度的升高, Tr明显增大, 而Pn总体上并没有表现出明显的增大趋势, 仅在一定的降水区间内和温度存在一定的线性相关(图3A、3D).因此, 本研究中树木Tr的增大是导致WUE和种源原地年平均气温负相关的主要原因.温度和树木WUE间这种负相关性在相近的研究中也有所体现(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).大量种源试验表明树木在干旱环境条件下具有更高的WUE (Anderson et al., 2000; Warren & Adams, 2000; Gouveia & Freitas, 2009; Ran et al., 2013), 这是由于干旱环境下空气湿度和土壤含水量降低、叶片Gs和Tr减小, 而Pn的变化相对较小, 从而导致WUE增大(Farquhar et al., 1989; Zhu et al., 2011; Liu et al., 2013).然而, 本研究结果显示Pn和Tr均与种源原地平均年降水量不相关(图3B、3E), 这是因为降水只考虑了水分收入, 未考虑水分的支出等情况, 不能完全表征一个地方的干旱程度.Warren等(2005)对同质园内来自于水分条件差异明显的15个种源Eucalyptus sideroxylon树木研究表明叶片WUE存在种源差异, 但和降水没有明显相关性, 相似的结果也在对不同种源Eucalyptus microtheca的研究中出现(Li, 2000).因此选用一个较合理的代表气候干旱程度的指标尤为重要, AI作为表征一个地区干湿程度的指标, 近来成为全球变化研究中经常涉及的气候指标之一(Arora, 2002; 孟猛等, 2004).本研究结果表明落叶松针叶WUE和AI显著正相关且相关性高于降水和温度(图3), 这与Warren和Adams (2000)对海岸松4个种源树木的研究结果相一致.此外, AI和Pn也存在线性正相关关系, 并且和不同水分蒸发范围内的Tr存在显著相关关系.这表明干旱情况下Pn的增加和Tr的减小是WUE提高的主要原因(Ramírez-Valiente et al., 2011). ...
Genetic variation in leaf pigment, optical and photosynthetic function among diverse phenotypes of Metrosideros polymorpha grown in a common garden.
2
2007
... 作为WUE的两大组分, Pn的增大或Tr的减小都会导致WUE的增大.本研究中落叶松叶片Pn存在显著的种源差异, 并且与种源原地的AI有显著的正相关性(表1; 图1, 图3), 这和以往对来自不同纬度种源树木研究结果相似(Soolanayakanahally et al., 2009; Robson et al., 2012).较高的叶Gs、氮含量和较小的SLA导致了较高的Pn, 并且Pn和种源原地的气候条件具有明显的相关关系, 这是基因调控下的Pn对种源原地环境长期适应的结果(Martin et al., 2007; Savolainen et al., 2007; Soolanayakanahally et al., 2009; Frei et al., 2012).除友好种源外, 随着Pn的增大, WUE表现出相近的变化趋势; 虽然叶片Tr也存在显著的种源差异, 然而WUE并没有随着Tr的变化表现出相应的变化趋势(图1), 这表明WUE受 Pn的影响较大.从研究结果变异程度而言, Pn的变异系数为15.95%, 最高值是最低值的1.60倍, 而Tr的变异系数为9.6%, 最高值较最低值高出44.99%, 由此使得WUE的变异趋势和Pn相近.这是因为叶片的其他生理和结构特征, 如叶片的Gs、NL、SLA等直接影响Pn和Tr, 进而引起了WUE的差异(Gago et al., 2014). ...
... 树木在长期的进化和选择过程中已形成通过改变生理、分子以及遗传等方面特征去响应环境改变的能力, 然而在目前全球气候变化加剧的背景下, 树木能否通过这种能力的改变去适应未来快速的气候变化已成为研究的热点问题(Saxe et al., 2001; Allen et al., 2010; Kremer et al., 2012; Shaw & Etterson, 2012; Reyer et al., 2013).研究同质种植园内同一树种不同种源树木的差异及其和种源原地气候条件的关系是一种验证树木对气候适应/驯化的重要方法(Reich et al., 1996; Warren et al., 2005; Martin et al., 2007; Frei et al., 2012). ...
地理生态学的干燥度指数及其应用评述
1
2004
... 与本文假设相同的是, 来自于低温少雨地区的种源具有较高的WUE, 并且和种源原地年平均气温及平均年降水量均呈线性负相关关系(图3G、3H).这是因为随着温度的升高叶片光合作用酶的活性增强、Gs增大, 进而影响CO2同化速率和Tr, 导致WUE的差异.本研究中17个种源落叶松的Pn和Tr均存在显著的种间差异, 但随着种源原地温度的升高, Tr明显增大, 而Pn总体上并没有表现出明显的增大趋势, 仅在一定的降水区间内和温度存在一定的线性相关(图3A、3D).因此, 本研究中树木Tr的增大是导致WUE和种源原地年平均气温负相关的主要原因.温度和树木WUE间这种负相关性在相近的研究中也有所体现(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).大量种源试验表明树木在干旱环境条件下具有更高的WUE (Anderson et al., 2000; Warren & Adams, 2000; Gouveia & Freitas, 2009; Ran et al., 2013), 这是由于干旱环境下空气湿度和土壤含水量降低、叶片Gs和Tr减小, 而Pn的变化相对较小, 从而导致WUE增大(Farquhar et al., 1989; Zhu et al., 2011; Liu et al., 2013).然而, 本研究结果显示Pn和Tr均与种源原地平均年降水量不相关(图3B、3E), 这是因为降水只考虑了水分收入, 未考虑水分的支出等情况, 不能完全表征一个地方的干旱程度.Warren等(2005)对同质园内来自于水分条件差异明显的15个种源Eucalyptus sideroxylon树木研究表明叶片WUE存在种源差异, 但和降水没有明显相关性, 相似的结果也在对不同种源Eucalyptus microtheca的研究中出现(Li, 2000).因此选用一个较合理的代表气候干旱程度的指标尤为重要, AI作为表征一个地区干湿程度的指标, 近来成为全球变化研究中经常涉及的气候指标之一(Arora, 2002; 孟猛等, 2004).本研究结果表明落叶松针叶WUE和AI显著正相关且相关性高于降水和温度(图3), 这与Warren和Adams (2000)对海岸松4个种源树木的研究结果相一致.此外, AI和Pn也存在线性正相关关系, 并且和不同水分蒸发范围内的Tr存在显著相关关系.这表明干旱情况下Pn的增加和Tr的减小是WUE提高的主要原因(Ramírez-Valiente et al., 2011). ...
地理生态学的干燥度指数及其应用评述
1
2004
... 与本文假设相同的是, 来自于低温少雨地区的种源具有较高的WUE, 并且和种源原地年平均气温及平均年降水量均呈线性负相关关系(图3G、3H).这是因为随着温度的升高叶片光合作用酶的活性增强、Gs增大, 进而影响CO2同化速率和Tr, 导致WUE的差异.本研究中17个种源落叶松的Pn和Tr均存在显著的种间差异, 但随着种源原地温度的升高, Tr明显增大, 而Pn总体上并没有表现出明显的增大趋势, 仅在一定的降水区间内和温度存在一定的线性相关(图3A、3D).因此, 本研究中树木Tr的增大是导致WUE和种源原地年平均气温负相关的主要原因.温度和树木WUE间这种负相关性在相近的研究中也有所体现(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).大量种源试验表明树木在干旱环境条件下具有更高的WUE (Anderson et al., 2000; Warren & Adams, 2000; Gouveia & Freitas, 2009; Ran et al., 2013), 这是由于干旱环境下空气湿度和土壤含水量降低、叶片Gs和Tr减小, 而Pn的变化相对较小, 从而导致WUE增大(Farquhar et al., 1989; Zhu et al., 2011; Liu et al., 2013).然而, 本研究结果显示Pn和Tr均与种源原地平均年降水量不相关(图3B、3E), 这是因为降水只考虑了水分收入, 未考虑水分的支出等情况, 不能完全表征一个地方的干旱程度.Warren等(2005)对同质园内来自于水分条件差异明显的15个种源Eucalyptus sideroxylon树木研究表明叶片WUE存在种源差异, 但和降水没有明显相关性, 相似的结果也在对不同种源Eucalyptus microtheca的研究中出现(Li, 2000).因此选用一个较合理的代表气候干旱程度的指标尤为重要, AI作为表征一个地区干湿程度的指标, 近来成为全球变化研究中经常涉及的气候指标之一(Arora, 2002; 孟猛等, 2004).本研究结果表明落叶松针叶WUE和AI显著正相关且相关性高于降水和温度(图3), 这与Warren和Adams (2000)对海岸松4个种源树木的研究结果相一致.此外, AI和Pn也存在线性正相关关系, 并且和不同水分蒸发范围内的Tr存在显著相关关系.这表明干旱情况下Pn的增加和Tr的减小是WUE提高的主要原因(Ramírez-Valiente et al., 2011). ...
Productivity, water-use efficiency and tolerance to moderate water deficit correlate in 33 poplar genotypes from a Populus deltoides × Populus trichocarpa F1 progeny.
4
2009
... 水分利用效率(WUE)是反映植物单位耗水量生产出的同化量的一个重要生理生态参数(Farquhar & Richards, 1984), 是耦合植物叶片光合作用和蒸腾作用的重要指标, 对研究生态系统水循环和碳循环相互作用、揭示植物对全球变化响应和适应对策具有重要意义(Saxe et al., 2001; Gao et al., 2014).气候变化条件下的树木WUE的响应及其机理已成为全球生态学研究的热点(Monclus et al., 2009; de Miguel et al., 2012; Klein et al., 2013).温度和降水是影响WUE的两个重要环境因子.有研究认为当温度升高后光合酶活性提高, 气孔导度(Gs)减小, 从而导致WUE增大(Morecroft et al., 1992; Xiao et al., 2013; Xu et al., 2013); 也有研究认为温度升高会导致胞间CO2浓度(Ci)减小、蒸腾速率(Tr)增大, WUE反而减小(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).降水通过改变土壤水分可利用性和空气湿度, 从而影响树木的气孔关闭和Tr, 进而改变树木的WUE.然而降水和WUE之间关系的研究结果也存在一些不确定性.有研究结果表明二者呈正相关关系(Schulze et al., 1998; Picotte et al., 2007; Niu et al., 2008; Klein et al., 2013); 也有研究认为二者呈负相关关系(Gimenez et al., 1992; Schulze et al., 2006; Monclus et al., 2009; Zhu et al., 2011).此外, 树种间由于生物学特性的差异导致其对环境的适应方式有所不同, 从而引起WUE种间差异.有研究表明, 相同环境下常绿乔木的WUE大于落叶乔木(Brooks et al., 1997; Kloeppel et al., 1998), 而有些研究却得出相反的结果(Damesin et al., 1997; 渠春梅等, 2001).因此, 环境变化对树木WUE的影响以及树木WUE对环境变化响应的内在机理还有待于深入研究. ...
... ; Monclus et al., 2009; Zhu et al., 2011).此外, 树种间由于生物学特性的差异导致其对环境的适应方式有所不同, 从而引起WUE种间差异.有研究表明, 相同环境下常绿乔木的WUE大于落叶乔木(Brooks et al., 1997; Kloeppel et al., 1998), 而有些研究却得出相反的结果(Damesin et al., 1997; 渠春梅等, 2001).因此, 环境变化对树木WUE的影响以及树木WUE对环境变化响应的内在机理还有待于深入研究. ...
... SLA作为叶片结构的重要特征之一也是影响WUE的重要因素, 有研究表明随着SLA的减小, 叶片厚度增加、光合组织密度增大, 导致水分的蒸腾减小和固碳能力提高, 从而WUE增大, 因此两者间多表现为负相关关系(Reich et al., 1998; Lamont et al., 2002; Warren et al., 2005; Gago et al., 2014).目前对于两者间关系的研究多集中在树种间(Lamont et al., 2002; Holtum & Winter, 2005)、同一树种不同种源幼苗间(Fernández et al., 1999; Warren et al., 2005)、同一种源不同幼苗个体间(Monclus et al., 2009; Roussel et al., 2009; de Miguel et al., 2012)等方面的表型差异, 而对环境变化下两者间的关系及其调控机理研究较少.de Miguel等(2012)对全同胞家系海岸松幼苗研究发现SLA和WUE存在负相关关系, 并认为这是基因调控的结果并能通过子代遗传, 相似的结果在其他研究中也有所体现(Fernández et al., 1999; Ramírez-Valiente et al., 2011).和我们假设相一致的是, 17个种源落叶松的30年树木SLA具有明显的种源差异并和WUE间存在显著的负相关关系(p < 0.05), 且随着种源原地AI的增大SLA对WUE的影响程度增强(图2B).这种在同质环境中生活多年仍存在显著的种源间差异且和种源原地气候条件有较好关系的结果进一步支持了这种差异是一种基因调控下的对种源原地环境长期适应的结果. ...
... 本研究结果表明落叶松NL和WUE间存在显著的正相关关系, 并且来自于高AI地区的种源树木WUE对NL具有更高的敏感性(图2C).有研究表明: 来自低温少雨地区种源的树木具有较高的NL, 以此提高光合能力获取更多的光合产物, 从而适应低温干燥的环境并且从贫瘠的土壤中获取更多的养分和水分(Reich et al., 1996; Weih & Karlsson, 2001; Oleksyn et al., 2003; Tjoelker et al., 2008; Bresson et al., 2011).因此, 树木NL同WUE间多表现为正相关关系(Sparks & Ehleringer, 1997; Monclus et al., 2009; Yan et al., 2014).相同NL水平下来自于高AI种源树木具有更高的WUE, 这对该树种能够在多样的环境中生存具有重要的现实意义.这是因为在干旱贫瘠的环境中土壤氮和水是树木生长的限制因子, 通过提高叶片氮的利用效率获得更多的光合产物、提高WUE, 从而有助于其生长和生存(Kenney et al., 2014; Yan et al., 2014).此外, 在模拟未来气候变化对树木生长乃至生存的影响时, 研究者认为气候的变化将导致落叶松分布区内永冻层融解萎缩, 土壤干燥, 湿润度下降, 最终使得落叶松林不适应新环境而逐步北移, 直至出境(张新时, 1993; 蒋延玲和周广胜, 2001; 李峰等, 2006).本研究发现落叶松树木可以通过调节叶片结构、养分含量、光合生理等形态和生理特征来调节WUE而适应干旱的环境, 这对研究该树种在气候变化下的应对策略和对新环境的适应能力具有重要的参考价值. ...
Altitudinal trends in leaf nutrient contents, leaf size and δ13C of Alchemilla alpina.
3
1992
... 水分利用效率(WUE)是反映植物单位耗水量生产出的同化量的一个重要生理生态参数(Farquhar & Richards, 1984), 是耦合植物叶片光合作用和蒸腾作用的重要指标, 对研究生态系统水循环和碳循环相互作用、揭示植物对全球变化响应和适应对策具有重要意义(Saxe et al., 2001; Gao et al., 2014).气候变化条件下的树木WUE的响应及其机理已成为全球生态学研究的热点(Monclus et al., 2009; de Miguel et al., 2012; Klein et al., 2013).温度和降水是影响WUE的两个重要环境因子.有研究认为当温度升高后光合酶活性提高, 气孔导度(Gs)减小, 从而导致WUE增大(Morecroft et al., 1992; Xiao et al., 2013; Xu et al., 2013); 也有研究认为温度升高会导致胞间CO2浓度(Ci)减小、蒸腾速率(Tr)增大, WUE反而减小(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).降水通过改变土壤水分可利用性和空气湿度, 从而影响树木的气孔关闭和Tr, 进而改变树木的WUE.然而降水和WUE之间关系的研究结果也存在一些不确定性.有研究结果表明二者呈正相关关系(Schulze et al., 1998; Picotte et al., 2007; Niu et al., 2008; Klein et al., 2013); 也有研究认为二者呈负相关关系(Gimenez et al., 1992; Schulze et al., 2006; Monclus et al., 2009; Zhu et al., 2011).此外, 树种间由于生物学特性的差异导致其对环境的适应方式有所不同, 从而引起WUE种间差异.有研究表明, 相同环境下常绿乔木的WUE大于落叶乔木(Brooks et al., 1997; Kloeppel et al., 1998), 而有些研究却得出相反的结果(Damesin et al., 1997; 渠春梅等, 2001).因此, 环境变化对树木WUE的影响以及树木WUE对环境变化响应的内在机理还有待于深入研究. ...
... 反而减小(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).降水通过改变土壤水分可利用性和空气湿度, 从而影响树木的气孔关闭和Tr, 进而改变树木的WUE.然而降水和WUE之间关系的研究结果也存在一些不确定性.有研究结果表明二者呈正相关关系(Schulze et al., 1998; Picotte et al., 2007; Niu et al., 2008; Klein et al., 2013); 也有研究认为二者呈负相关关系(Gimenez et al., 1992; Schulze et al., 2006; Monclus et al., 2009; Zhu et al., 2011).此外, 树种间由于生物学特性的差异导致其对环境的适应方式有所不同, 从而引起WUE种间差异.有研究表明, 相同环境下常绿乔木的WUE大于落叶乔木(Brooks et al., 1997; Kloeppel et al., 1998), 而有些研究却得出相反的结果(Damesin et al., 1997; 渠春梅等, 2001).因此, 环境变化对树木WUE的影响以及树木WUE对环境变化响应的内在机理还有待于深入研究. ...
... 与本文假设相同的是, 来自于低温少雨地区的种源具有较高的WUE, 并且和种源原地年平均气温及平均年降水量均呈线性负相关关系(图3G、3H).这是因为随着温度的升高叶片光合作用酶的活性增强、Gs增大, 进而影响CO2同化速率和Tr, 导致WUE的差异.本研究中17个种源落叶松的Pn和Tr均存在显著的种间差异, 但随着种源原地温度的升高, Tr明显增大, 而Pn总体上并没有表现出明显的增大趋势, 仅在一定的降水区间内和温度存在一定的线性相关(图3A、3D).因此, 本研究中树木Tr的增大是导致WUE和种源原地年平均气温负相关的主要原因.温度和树木WUE间这种负相关性在相近的研究中也有所体现(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).大量种源试验表明树木在干旱环境条件下具有更高的WUE (Anderson et al., 2000; Warren & Adams, 2000; Gouveia & Freitas, 2009; Ran et al., 2013), 这是由于干旱环境下空气湿度和土壤含水量降低、叶片Gs和Tr减小, 而Pn的变化相对较小, 从而导致WUE增大(Farquhar et al., 1989; Zhu et al., 2011; Liu et al., 2013).然而, 本研究结果显示Pn和Tr均与种源原地平均年降水量不相关(图3B、3E), 这是因为降水只考虑了水分收入, 未考虑水分的支出等情况, 不能完全表征一个地方的干旱程度.Warren等(2005)对同质园内来自于水分条件差异明显的15个种源Eucalyptus sideroxylon树木研究表明叶片WUE存在种源差异, 但和降水没有明显相关性, 相似的结果也在对不同种源Eucalyptus microtheca的研究中出现(Li, 2000).因此选用一个较合理的代表气候干旱程度的指标尤为重要, AI作为表征一个地区干湿程度的指标, 近来成为全球变化研究中经常涉及的气候指标之一(Arora, 2002; 孟猛等, 2004).本研究结果表明落叶松针叶WUE和AI显著正相关且相关性高于降水和温度(图3), 这与Warren和Adams (2000)对海岸松4个种源树木的研究结果相一致.此外, AI和Pn也存在线性正相关关系, 并且和不同水分蒸发范围内的Tr存在显著相关关系.这表明干旱情况下Pn的增加和Tr的减小是WUE提高的主要原因(Ramírez-Valiente et al., 2011). ...
Leaf shape linked to photosynthetic rates and temperature optima in South African Pelargonium species.
2
2008
... 水分利用效率(WUE)是反映植物单位耗水量生产出的同化量的一个重要生理生态参数(Farquhar & Richards, 1984), 是耦合植物叶片光合作用和蒸腾作用的重要指标, 对研究生态系统水循环和碳循环相互作用、揭示植物对全球变化响应和适应对策具有重要意义(Saxe et al., 2001; Gao et al., 2014).气候变化条件下的树木WUE的响应及其机理已成为全球生态学研究的热点(Monclus et al., 2009; de Miguel et al., 2012; Klein et al., 2013).温度和降水是影响WUE的两个重要环境因子.有研究认为当温度升高后光合酶活性提高, 气孔导度(Gs)减小, 从而导致WUE增大(Morecroft et al., 1992; Xiao et al., 2013; Xu et al., 2013); 也有研究认为温度升高会导致胞间CO2浓度(Ci)减小、蒸腾速率(Tr)增大, WUE反而减小(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).降水通过改变土壤水分可利用性和空气湿度, 从而影响树木的气孔关闭和Tr, 进而改变树木的WUE.然而降水和WUE之间关系的研究结果也存在一些不确定性.有研究结果表明二者呈正相关关系(Schulze et al., 1998; Picotte et al., 2007; Niu et al., 2008; Klein et al., 2013); 也有研究认为二者呈负相关关系(Gimenez et al., 1992; Schulze et al., 2006; Monclus et al., 2009; Zhu et al., 2011).此外, 树种间由于生物学特性的差异导致其对环境的适应方式有所不同, 从而引起WUE种间差异.有研究表明, 相同环境下常绿乔木的WUE大于落叶乔木(Brooks et al., 1997; Kloeppel et al., 1998), 而有些研究却得出相反的结果(Damesin et al., 1997; 渠春梅等, 2001).因此, 环境变化对树木WUE的影响以及树木WUE对环境变化响应的内在机理还有待于深入研究. ...
... 与本文假设相同的是, 来自于低温少雨地区的种源具有较高的WUE, 并且和种源原地年平均气温及平均年降水量均呈线性负相关关系(图3G、3H).这是因为随着温度的升高叶片光合作用酶的活性增强、Gs增大, 进而影响CO2同化速率和Tr, 导致WUE的差异.本研究中17个种源落叶松的Pn和Tr均存在显著的种间差异, 但随着种源原地温度的升高, Tr明显增大, 而Pn总体上并没有表现出明显的增大趋势, 仅在一定的降水区间内和温度存在一定的线性相关(图3A、3D).因此, 本研究中树木Tr的增大是导致WUE和种源原地年平均气温负相关的主要原因.温度和树木WUE间这种负相关性在相近的研究中也有所体现(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).大量种源试验表明树木在干旱环境条件下具有更高的WUE (Anderson et al., 2000; Warren & Adams, 2000; Gouveia & Freitas, 2009; Ran et al., 2013), 这是由于干旱环境下空气湿度和土壤含水量降低、叶片Gs和Tr减小, 而Pn的变化相对较小, 从而导致WUE增大(Farquhar et al., 1989; Zhu et al., 2011; Liu et al., 2013).然而, 本研究结果显示Pn和Tr均与种源原地平均年降水量不相关(图3B、3E), 这是因为降水只考虑了水分收入, 未考虑水分的支出等情况, 不能完全表征一个地方的干旱程度.Warren等(2005)对同质园内来自于水分条件差异明显的15个种源Eucalyptus sideroxylon树木研究表明叶片WUE存在种源差异, 但和降水没有明显相关性, 相似的结果也在对不同种源Eucalyptus microtheca的研究中出现(Li, 2000).因此选用一个较合理的代表气候干旱程度的指标尤为重要, AI作为表征一个地区干湿程度的指标, 近来成为全球变化研究中经常涉及的气候指标之一(Arora, 2002; 孟猛等, 2004).本研究结果表明落叶松针叶WUE和AI显著正相关且相关性高于降水和温度(图3), 这与Warren和Adams (2000)对海岸松4个种源树木的研究结果相一致.此外, AI和Pn也存在线性正相关关系, 并且和不同水分蒸发范围内的Tr存在显著相关关系.这表明干旱情况下Pn的增加和Tr的减小是WUE提高的主要原因(Ramírez-Valiente et al., 2011). ...
Water-mediated responses of ecosystem carbon fluxes to climatic change in a temperate steppe.
1
2008
... 水分利用效率(WUE)是反映植物单位耗水量生产出的同化量的一个重要生理生态参数(Farquhar & Richards, 1984), 是耦合植物叶片光合作用和蒸腾作用的重要指标, 对研究生态系统水循环和碳循环相互作用、揭示植物对全球变化响应和适应对策具有重要意义(Saxe et al., 2001; Gao et al., 2014).气候变化条件下的树木WUE的响应及其机理已成为全球生态学研究的热点(Monclus et al., 2009; de Miguel et al., 2012; Klein et al., 2013).温度和降水是影响WUE的两个重要环境因子.有研究认为当温度升高后光合酶活性提高, 气孔导度(Gs)减小, 从而导致WUE增大(Morecroft et al., 1992; Xiao et al., 2013; Xu et al., 2013); 也有研究认为温度升高会导致胞间CO2浓度(Ci)减小、蒸腾速率(Tr)增大, WUE反而减小(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).降水通过改变土壤水分可利用性和空气湿度, 从而影响树木的气孔关闭和Tr, 进而改变树木的WUE.然而降水和WUE之间关系的研究结果也存在一些不确定性.有研究结果表明二者呈正相关关系(Schulze et al., 1998; Picotte et al., 2007; Niu et al., 2008; Klein et al., 2013); 也有研究认为二者呈负相关关系(Gimenez et al., 1992; Schulze et al., 2006; Monclus et al., 2009; Zhu et al., 2011).此外, 树种间由于生物学特性的差异导致其对环境的适应方式有所不同, 从而引起WUE种间差异.有研究表明, 相同环境下常绿乔木的WUE大于落叶乔木(Brooks et al., 1997; Kloeppel et al., 1998), 而有些研究却得出相反的结果(Damesin et al., 1997; 渠春梅等, 2001).因此, 环境变化对树木WUE的影响以及树木WUE对环境变化响应的内在机理还有待于深入研究. ...
Nutrient conservation increases with latitude of origin in european Pinus sylvestris populations.
1
2003
... 本研究结果表明落叶松NL和WUE间存在显著的正相关关系, 并且来自于高AI地区的种源树木WUE对NL具有更高的敏感性(图2C).有研究表明: 来自低温少雨地区种源的树木具有较高的NL, 以此提高光合能力获取更多的光合产物, 从而适应低温干燥的环境并且从贫瘠的土壤中获取更多的养分和水分(Reich et al., 1996; Weih & Karlsson, 2001; Oleksyn et al., 2003; Tjoelker et al., 2008; Bresson et al., 2011).因此, 树木NL同WUE间多表现为正相关关系(Sparks & Ehleringer, 1997; Monclus et al., 2009; Yan et al., 2014).相同NL水平下来自于高AI种源树木具有更高的WUE, 这对该树种能够在多样的环境中生存具有重要的现实意义.这是因为在干旱贫瘠的环境中土壤氮和水是树木生长的限制因子, 通过提高叶片氮的利用效率获得更多的光合产物、提高WUE, 从而有助于其生长和生存(Kenney et al., 2014; Yan et al., 2014).此外, 在模拟未来气候变化对树木生长乃至生存的影响时, 研究者认为气候的变化将导致落叶松分布区内永冻层融解萎缩, 土壤干燥, 湿润度下降, 最终使得落叶松林不适应新环境而逐步北移, 直至出境(张新时, 1993; 蒋延玲和周广胜, 2001; 李峰等, 2006).本研究发现落叶松树木可以通过调节叶片结构、养分含量、光合生理等形态和生理特征来调节WUE而适应干旱的环境, 这对研究该树种在气候变化下的应对策略和对新环境的适应能力具有重要的参考价值. ...
Plastic responses to temporal variation in moisture availability: Consequences for water use efficiency and plant performance.
2
2007
... 水分利用效率(WUE)是反映植物单位耗水量生产出的同化量的一个重要生理生态参数(Farquhar & Richards, 1984), 是耦合植物叶片光合作用和蒸腾作用的重要指标, 对研究生态系统水循环和碳循环相互作用、揭示植物对全球变化响应和适应对策具有重要意义(Saxe et al., 2001; Gao et al., 2014).气候变化条件下的树木WUE的响应及其机理已成为全球生态学研究的热点(Monclus et al., 2009; de Miguel et al., 2012; Klein et al., 2013).温度和降水是影响WUE的两个重要环境因子.有研究认为当温度升高后光合酶活性提高, 气孔导度(Gs)减小, 从而导致WUE增大(Morecroft et al., 1992; Xiao et al., 2013; Xu et al., 2013); 也有研究认为温度升高会导致胞间CO2浓度(Ci)减小、蒸腾速率(Tr)增大, WUE反而减小(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).降水通过改变土壤水分可利用性和空气湿度, 从而影响树木的气孔关闭和Tr, 进而改变树木的WUE.然而降水和WUE之间关系的研究结果也存在一些不确定性.有研究结果表明二者呈正相关关系(Schulze et al., 1998; Picotte et al., 2007; Niu et al., 2008; Klein et al., 2013); 也有研究认为二者呈负相关关系(Gimenez et al., 1992; Schulze et al., 2006; Monclus et al., 2009; Zhu et al., 2011).此外, 树种间由于生物学特性的差异导致其对环境的适应方式有所不同, 从而引起WUE种间差异.有研究表明, 相同环境下常绿乔木的WUE大于落叶乔木(Brooks et al., 1997; Kloeppel et al., 1998), 而有些研究却得出相反的结果(Damesin et al., 1997; 渠春梅等, 2001).因此, 环境变化对树木WUE的影响以及树木WUE对环境变化响应的内在机理还有待于深入研究. ...
... Gs作为叶片同外界进行水分和气体交换的主要通道, 调节着碳同化速率和水分消耗间的平衡(Gagen et al., 2011).在一定范围内, 当Gs升高时叶片Ci增加, 导致Pn增大, 虽然Tr也增大, 但光合速率增大的幅度较大, 从而Gs和WUE呈一定的正相关关系(Picotte et al., 2007; Gago et al., 2014), 但随着Gs的继续增大, 叶片在通过气孔得到CO2和失去水分间达到平衡, 此时WUE的限制因素不再是Gs, 而转为光合相关的酶含量和活性等其他生理因素(Farquhar et al., 1989; Gimenez et al., 1992).本试验结果表明在起始阶段WUE和Gs呈明显线性相关, 而当Gs增大到0.2左右时WUE趋于稳定, 但来自于较干旱地方的种源树木在相同WUE水平时需要较低的Gs (图2A), 这验证了我们假设干旱条件下树木通过减小Gs提高WUE的假设.通过较小的Gs获得较大的WUE可以使树木在干旱的条件下减少水分的蒸腾损失, 调节树木水势变化, 避免脱水, 从而提高植物对土壤水分的有效利用(Kenney et al., 2014).来自不同水分条件地区种源树木在同质园内生长30年后, 还存有与原地气候相适应的Gs对WUE的凋节方式, 这表现出明显的子代遗传效应.de Miguel等(2012)通过对来自全胞家系海岸松(Pinus pinaster)无性系苗木的研究也取得了相似的结果. ...
云南西双版纳片断化热带雨林植物叶片δ13C值的特点及其对水分利用效率的指示
1
2001
... 水分利用效率(WUE)是反映植物单位耗水量生产出的同化量的一个重要生理生态参数(Farquhar & Richards, 1984), 是耦合植物叶片光合作用和蒸腾作用的重要指标, 对研究生态系统水循环和碳循环相互作用、揭示植物对全球变化响应和适应对策具有重要意义(Saxe et al., 2001; Gao et al., 2014).气候变化条件下的树木WUE的响应及其机理已成为全球生态学研究的热点(Monclus et al., 2009; de Miguel et al., 2012; Klein et al., 2013).温度和降水是影响WUE的两个重要环境因子.有研究认为当温度升高后光合酶活性提高, 气孔导度(Gs)减小, 从而导致WUE增大(Morecroft et al., 1992; Xiao et al., 2013; Xu et al., 2013); 也有研究认为温度升高会导致胞间CO2浓度(Ci)减小、蒸腾速率(Tr)增大, WUE反而减小(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).降水通过改变土壤水分可利用性和空气湿度, 从而影响树木的气孔关闭和Tr, 进而改变树木的WUE.然而降水和WUE之间关系的研究结果也存在一些不确定性.有研究结果表明二者呈正相关关系(Schulze et al., 1998; Picotte et al., 2007; Niu et al., 2008; Klein et al., 2013); 也有研究认为二者呈负相关关系(Gimenez et al., 1992; Schulze et al., 2006; Monclus et al., 2009; Zhu et al., 2011).此外, 树种间由于生物学特性的差异导致其对环境的适应方式有所不同, 从而引起WUE种间差异.有研究表明, 相同环境下常绿乔木的WUE大于落叶乔木(Brooks et al., 1997; Kloeppel et al., 1998), 而有些研究却得出相反的结果(Damesin et al., 1997; 渠春梅等, 2001).因此, 环境变化对树木WUE的影响以及树木WUE对环境变化响应的内在机理还有待于深入研究. ...
云南西双版纳片断化热带雨林植物叶片δ13C值的特点及其对水分利用效率的指示
1
2001
... 水分利用效率(WUE)是反映植物单位耗水量生产出的同化量的一个重要生理生态参数(Farquhar & Richards, 1984), 是耦合植物叶片光合作用和蒸腾作用的重要指标, 对研究生态系统水循环和碳循环相互作用、揭示植物对全球变化响应和适应对策具有重要意义(Saxe et al., 2001; Gao et al., 2014).气候变化条件下的树木WUE的响应及其机理已成为全球生态学研究的热点(Monclus et al., 2009; de Miguel et al., 2012; Klein et al., 2013).温度和降水是影响WUE的两个重要环境因子.有研究认为当温度升高后光合酶活性提高, 气孔导度(Gs)减小, 从而导致WUE增大(Morecroft et al., 1992; Xiao et al., 2013; Xu et al., 2013); 也有研究认为温度升高会导致胞间CO2浓度(Ci)减小、蒸腾速率(Tr)增大, WUE反而减小(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).降水通过改变土壤水分可利用性和空气湿度, 从而影响树木的气孔关闭和Tr, 进而改变树木的WUE.然而降水和WUE之间关系的研究结果也存在一些不确定性.有研究结果表明二者呈正相关关系(Schulze et al., 1998; Picotte et al., 2007; Niu et al., 2008; Klein et al., 2013); 也有研究认为二者呈负相关关系(Gimenez et al., 1992; Schulze et al., 2006; Monclus et al., 2009; Zhu et al., 2011).此外, 树种间由于生物学特性的差异导致其对环境的适应方式有所不同, 从而引起WUE种间差异.有研究表明, 相同环境下常绿乔木的WUE大于落叶乔木(Brooks et al., 1997; Kloeppel et al., 1998), 而有些研究却得出相反的结果(Damesin et al., 1997; 渠春梅等, 2001).因此, 环境变化对树木WUE的影响以及树木WUE对环境变化响应的内在机理还有待于深入研究. ...
Factors affecting cork oak growth under dry conditions: Local adaptation and contrasting additive genetic variance within populations.
2
2011
... SLA作为叶片结构的重要特征之一也是影响WUE的重要因素, 有研究表明随着SLA的减小, 叶片厚度增加、光合组织密度增大, 导致水分的蒸腾减小和固碳能力提高, 从而WUE增大, 因此两者间多表现为负相关关系(Reich et al., 1998; Lamont et al., 2002; Warren et al., 2005; Gago et al., 2014).目前对于两者间关系的研究多集中在树种间(Lamont et al., 2002; Holtum & Winter, 2005)、同一树种不同种源幼苗间(Fernández et al., 1999; Warren et al., 2005)、同一种源不同幼苗个体间(Monclus et al., 2009; Roussel et al., 2009; de Miguel et al., 2012)等方面的表型差异, 而对环境变化下两者间的关系及其调控机理研究较少.de Miguel等(2012)对全同胞家系海岸松幼苗研究发现SLA和WUE存在负相关关系, 并认为这是基因调控的结果并能通过子代遗传, 相似的结果在其他研究中也有所体现(Fernández et al., 1999; Ramírez-Valiente et al., 2011).和我们假设相一致的是, 17个种源落叶松的30年树木SLA具有明显的种源差异并和WUE间存在显著的负相关关系(p < 0.05), 且随着种源原地AI的增大SLA对WUE的影响程度增强(图2B).这种在同质环境中生活多年仍存在显著的种源间差异且和种源原地气候条件有较好关系的结果进一步支持了这种差异是一种基因调控下的对种源原地环境长期适应的结果. ...
... 与本文假设相同的是, 来自于低温少雨地区的种源具有较高的WUE, 并且和种源原地年平均气温及平均年降水量均呈线性负相关关系(图3G、3H).这是因为随着温度的升高叶片光合作用酶的活性增强、Gs增大, 进而影响CO2同化速率和Tr, 导致WUE的差异.本研究中17个种源落叶松的Pn和Tr均存在显著的种间差异, 但随着种源原地温度的升高, Tr明显增大, 而Pn总体上并没有表现出明显的增大趋势, 仅在一定的降水区间内和温度存在一定的线性相关(图3A、3D).因此, 本研究中树木Tr的增大是导致WUE和种源原地年平均气温负相关的主要原因.温度和树木WUE间这种负相关性在相近的研究中也有所体现(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).大量种源试验表明树木在干旱环境条件下具有更高的WUE (Anderson et al., 2000; Warren & Adams, 2000; Gouveia & Freitas, 2009; Ran et al., 2013), 这是由于干旱环境下空气湿度和土壤含水量降低、叶片Gs和Tr减小, 而Pn的变化相对较小, 从而导致WUE增大(Farquhar et al., 1989; Zhu et al., 2011; Liu et al., 2013).然而, 本研究结果显示Pn和Tr均与种源原地平均年降水量不相关(图3B、3E), 这是因为降水只考虑了水分收入, 未考虑水分的支出等情况, 不能完全表征一个地方的干旱程度.Warren等(2005)对同质园内来自于水分条件差异明显的15个种源Eucalyptus sideroxylon树木研究表明叶片WUE存在种源差异, 但和降水没有明显相关性, 相似的结果也在对不同种源Eucalyptus microtheca的研究中出现(Li, 2000).因此选用一个较合理的代表气候干旱程度的指标尤为重要, AI作为表征一个地区干湿程度的指标, 近来成为全球变化研究中经常涉及的气候指标之一(Arora, 2002; 孟猛等, 2004).本研究结果表明落叶松针叶WUE和AI显著正相关且相关性高于降水和温度(图3), 这与Warren和Adams (2000)对海岸松4个种源树木的研究结果相一致.此外, AI和Pn也存在线性正相关关系, 并且和不同水分蒸发范围内的Tr存在显著相关关系.这表明干旱情况下Pn的增加和Tr的减小是WUE提高的主要原因(Ramírez-Valiente et al., 2011). ...
Altitudinal variation in growth, photosynthetic capacity and water use efficiency of Abies faxoniana Rehd. et Wils. seedlings as revealed by reciprocal transplantations.
1
2013
... 与本文假设相同的是, 来自于低温少雨地区的种源具有较高的WUE, 并且和种源原地年平均气温及平均年降水量均呈线性负相关关系(图3G、3H).这是因为随着温度的升高叶片光合作用酶的活性增强、Gs增大, 进而影响CO2同化速率和Tr, 导致WUE的差异.本研究中17个种源落叶松的Pn和Tr均存在显著的种间差异, 但随着种源原地温度的升高, Tr明显增大, 而Pn总体上并没有表现出明显的增大趋势, 仅在一定的降水区间内和温度存在一定的线性相关(图3A、3D).因此, 本研究中树木Tr的增大是导致WUE和种源原地年平均气温负相关的主要原因.温度和树木WUE间这种负相关性在相近的研究中也有所体现(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).大量种源试验表明树木在干旱环境条件下具有更高的WUE (Anderson et al., 2000; Warren & Adams, 2000; Gouveia & Freitas, 2009; Ran et al., 2013), 这是由于干旱环境下空气湿度和土壤含水量降低、叶片Gs和Tr减小, 而Pn的变化相对较小, 从而导致WUE增大(Farquhar et al., 1989; Zhu et al., 2011; Liu et al., 2013).然而, 本研究结果显示Pn和Tr均与种源原地平均年降水量不相关(图3B、3E), 这是因为降水只考虑了水分收入, 未考虑水分的支出等情况, 不能完全表征一个地方的干旱程度.Warren等(2005)对同质园内来自于水分条件差异明显的15个种源Eucalyptus sideroxylon树木研究表明叶片WUE存在种源差异, 但和降水没有明显相关性, 相似的结果也在对不同种源Eucalyptus microtheca的研究中出现(Li, 2000).因此选用一个较合理的代表气候干旱程度的指标尤为重要, AI作为表征一个地区干湿程度的指标, 近来成为全球变化研究中经常涉及的气候指标之一(Arora, 2002; 孟猛等, 2004).本研究结果表明落叶松针叶WUE和AI显著正相关且相关性高于降水和温度(图3), 这与Warren和Adams (2000)对海岸松4个种源树木的研究结果相一致.此外, AI和Pn也存在线性正相关关系, 并且和不同水分蒸发范围内的Tr存在显著相关关系.这表明干旱情况下Pn的增加和Tr的减小是WUE提高的主要原因(Ramírez-Valiente et al., 2011). ...
Needle respiration and nitrogen concentration in scots pine populations from a broad latitudinal range: A common garden test with field-grown trees.
2
1996
... 本研究结果表明落叶松NL和WUE间存在显著的正相关关系, 并且来自于高AI地区的种源树木WUE对NL具有更高的敏感性(图2C).有研究表明: 来自低温少雨地区种源的树木具有较高的NL, 以此提高光合能力获取更多的光合产物, 从而适应低温干燥的环境并且从贫瘠的土壤中获取更多的养分和水分(Reich et al., 1996; Weih & Karlsson, 2001; Oleksyn et al., 2003; Tjoelker et al., 2008; Bresson et al., 2011).因此, 树木NL同WUE间多表现为正相关关系(Sparks & Ehleringer, 1997; Monclus et al., 2009; Yan et al., 2014).相同NL水平下来自于高AI种源树木具有更高的WUE, 这对该树种能够在多样的环境中生存具有重要的现实意义.这是因为在干旱贫瘠的环境中土壤氮和水是树木生长的限制因子, 通过提高叶片氮的利用效率获得更多的光合产物、提高WUE, 从而有助于其生长和生存(Kenney et al., 2014; Yan et al., 2014).此外, 在模拟未来气候变化对树木生长乃至生存的影响时, 研究者认为气候的变化将导致落叶松分布区内永冻层融解萎缩, 土壤干燥, 湿润度下降, 最终使得落叶松林不适应新环境而逐步北移, 直至出境(张新时, 1993; 蒋延玲和周广胜, 2001; 李峰等, 2006).本研究发现落叶松树木可以通过调节叶片结构、养分含量、光合生理等形态和生理特征来调节WUE而适应干旱的环境, 这对研究该树种在气候变化下的应对策略和对新环境的适应能力具有重要的参考价值. ...
... 树木在长期的进化和选择过程中已形成通过改变生理、分子以及遗传等方面特征去响应环境改变的能力, 然而在目前全球气候变化加剧的背景下, 树木能否通过这种能力的改变去适应未来快速的气候变化已成为研究的热点问题(Saxe et al., 2001; Allen et al., 2010; Kremer et al., 2012; Shaw & Etterson, 2012; Reyer et al., 2013).研究同质种植园内同一树种不同种源树木的差异及其和种源原地气候条件的关系是一种验证树木对气候适应/驯化的重要方法(Reich et al., 1996; Warren et al., 2005; Martin et al., 2007; Frei et al., 2012). ...
Photosynthesis and respiration rates depend on leaf and root morphology and nitrogen concentration in nine boreal tree species differing in relative growth rate.
1
1998
... SLA作为叶片结构的重要特征之一也是影响WUE的重要因素, 有研究表明随着SLA的减小, 叶片厚度增加、光合组织密度增大, 导致水分的蒸腾减小和固碳能力提高, 从而WUE增大, 因此两者间多表现为负相关关系(Reich et al., 1998; Lamont et al., 2002; Warren et al., 2005; Gago et al., 2014).目前对于两者间关系的研究多集中在树种间(Lamont et al., 2002; Holtum & Winter, 2005)、同一树种不同种源幼苗间(Fernández et al., 1999; Warren et al., 2005)、同一种源不同幼苗个体间(Monclus et al., 2009; Roussel et al., 2009; de Miguel et al., 2012)等方面的表型差异, 而对环境变化下两者间的关系及其调控机理研究较少.de Miguel等(2012)对全同胞家系海岸松幼苗研究发现SLA和WUE存在负相关关系, 并认为这是基因调控的结果并能通过子代遗传, 相似的结果在其他研究中也有所体现(Fernández et al., 1999; Ramírez-Valiente et al., 2011).和我们假设相一致的是, 17个种源落叶松的30年树木SLA具有明显的种源差异并和WUE间存在显著的负相关关系(p < 0.05), 且随着种源原地AI的增大SLA对WUE的影响程度增强(图2B).这种在同质环境中生活多年仍存在显著的种源间差异且和种源原地气候条件有较好关系的结果进一步支持了这种差异是一种基因调控下的对种源原地环境长期适应的结果. ...
A plant’s perspective of extremes: Terrestrial plant responses to changing climatic variability.
1
2013
... 树木在长期的进化和选择过程中已形成通过改变生理、分子以及遗传等方面特征去响应环境改变的能力, 然而在目前全球气候变化加剧的背景下, 树木能否通过这种能力的改变去适应未来快速的气候变化已成为研究的热点问题(Saxe et al., 2001; Allen et al., 2010; Kremer et al., 2012; Shaw & Etterson, 2012; Reyer et al., 2013).研究同质种植园内同一树种不同种源树木的差异及其和种源原地气候条件的关系是一种验证树木对气候适应/驯化的重要方法(Reich et al., 1996; Warren et al., 2005; Martin et al., 2007; Frei et al., 2012). ...
Variation in functional leaf traits among beech provenances during a spanish summer reflects the differences in their origin.
1
2012
... 作为WUE的两大组分, Pn的增大或Tr的减小都会导致WUE的增大.本研究中落叶松叶片Pn存在显著的种源差异, 并且与种源原地的AI有显著的正相关性(表1; 图1, 图3), 这和以往对来自不同纬度种源树木研究结果相似(Soolanayakanahally et al., 2009; Robson et al., 2012).较高的叶Gs、氮含量和较小的SLA导致了较高的Pn, 并且Pn和种源原地的气候条件具有明显的相关关系, 这是基因调控下的Pn对种源原地环境长期适应的结果(Martin et al., 2007; Savolainen et al., 2007; Soolanayakanahally et al., 2009; Frei et al., 2012).除友好种源外, 随着Pn的增大, WUE表现出相近的变化趋势; 虽然叶片Tr也存在显著的种源差异, 然而WUE并没有随着Tr的变化表现出相应的变化趋势(图1), 这表明WUE受 Pn的影响较大.从研究结果变异程度而言, Pn的变异系数为15.95%, 最高值是最低值的1.60倍, 而Tr的变异系数为9.6%, 最高值较最低值高出44.99%, 由此使得WUE的变异趋势和Pn相近.这是因为叶片的其他生理和结构特征, 如叶片的Gs、NL、SLA等直接影响Pn和Tr, 进而引起了WUE的差异(Gago et al., 2014). ...
Diversity of water use efficiency among Quercus robur genotypes: Contribution of related leaf traits.
1
2009
... SLA作为叶片结构的重要特征之一也是影响WUE的重要因素, 有研究表明随着SLA的减小, 叶片厚度增加、光合组织密度增大, 导致水分的蒸腾减小和固碳能力提高, 从而WUE增大, 因此两者间多表现为负相关关系(Reich et al., 1998; Lamont et al., 2002; Warren et al., 2005; Gago et al., 2014).目前对于两者间关系的研究多集中在树种间(Lamont et al., 2002; Holtum & Winter, 2005)、同一树种不同种源幼苗间(Fernández et al., 1999; Warren et al., 2005)、同一种源不同幼苗个体间(Monclus et al., 2009; Roussel et al., 2009; de Miguel et al., 2012)等方面的表型差异, 而对环境变化下两者间的关系及其调控机理研究较少.de Miguel等(2012)对全同胞家系海岸松幼苗研究发现SLA和WUE存在负相关关系, 并认为这是基因调控的结果并能通过子代遗传, 相似的结果在其他研究中也有所体现(Fernández et al., 1999; Ramírez-Valiente et al., 2011).和我们假设相一致的是, 17个种源落叶松的30年树木SLA具有明显的种源差异并和WUE间存在显著的负相关关系(p < 0.05), 且随着种源原地AI的增大SLA对WUE的影响程度增强(图2B).这种在同质环境中生活多年仍存在显著的种源间差异且和种源原地气候条件有较好关系的结果进一步支持了这种差异是一种基因调控下的对种源原地环境长期适应的结果. ...
Gene flow and local adaptation in trees.
1
2007
... 作为WUE的两大组分, Pn的增大或Tr的减小都会导致WUE的增大.本研究中落叶松叶片Pn存在显著的种源差异, 并且与种源原地的AI有显著的正相关性(表1; 图1, 图3), 这和以往对来自不同纬度种源树木研究结果相似(Soolanayakanahally et al., 2009; Robson et al., 2012).较高的叶Gs、氮含量和较小的SLA导致了较高的Pn, 并且Pn和种源原地的气候条件具有明显的相关关系, 这是基因调控下的Pn对种源原地环境长期适应的结果(Martin et al., 2007; Savolainen et al., 2007; Soolanayakanahally et al., 2009; Frei et al., 2012).除友好种源外, 随着Pn的增大, WUE表现出相近的变化趋势; 虽然叶片Tr也存在显著的种源差异, 然而WUE并没有随着Tr的变化表现出相应的变化趋势(图1), 这表明WUE受 Pn的影响较大.从研究结果变异程度而言, Pn的变异系数为15.95%, 最高值是最低值的1.60倍, 而Tr的变异系数为9.6%, 最高值较最低值高出44.99%, 由此使得WUE的变异趋势和Pn相近.这是因为叶片的其他生理和结构特征, 如叶片的Gs、NL、SLA等直接影响Pn和Tr, 进而引起了WUE的差异(Gago et al., 2014). ...
Tree and forest functioning in response to global warming.
2
2001
... 水分利用效率(WUE)是反映植物单位耗水量生产出的同化量的一个重要生理生态参数(Farquhar & Richards, 1984), 是耦合植物叶片光合作用和蒸腾作用的重要指标, 对研究生态系统水循环和碳循环相互作用、揭示植物对全球变化响应和适应对策具有重要意义(Saxe et al., 2001; Gao et al., 2014).气候变化条件下的树木WUE的响应及其机理已成为全球生态学研究的热点(Monclus et al., 2009; de Miguel et al., 2012; Klein et al., 2013).温度和降水是影响WUE的两个重要环境因子.有研究认为当温度升高后光合酶活性提高, 气孔导度(Gs)减小, 从而导致WUE增大(Morecroft et al., 1992; Xiao et al., 2013; Xu et al., 2013); 也有研究认为温度升高会导致胞间CO2浓度(Ci)减小、蒸腾速率(Tr)增大, WUE反而减小(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).降水通过改变土壤水分可利用性和空气湿度, 从而影响树木的气孔关闭和Tr, 进而改变树木的WUE.然而降水和WUE之间关系的研究结果也存在一些不确定性.有研究结果表明二者呈正相关关系(Schulze et al., 1998; Picotte et al., 2007; Niu et al., 2008; Klein et al., 2013); 也有研究认为二者呈负相关关系(Gimenez et al., 1992; Schulze et al., 2006; Monclus et al., 2009; Zhu et al., 2011).此外, 树种间由于生物学特性的差异导致其对环境的适应方式有所不同, 从而引起WUE种间差异.有研究表明, 相同环境下常绿乔木的WUE大于落叶乔木(Brooks et al., 1997; Kloeppel et al., 1998), 而有些研究却得出相反的结果(Damesin et al., 1997; 渠春梅等, 2001).因此, 环境变化对树木WUE的影响以及树木WUE对环境变化响应的内在机理还有待于深入研究. ...
... 树木在长期的进化和选择过程中已形成通过改变生理、分子以及遗传等方面特征去响应环境改变的能力, 然而在目前全球气候变化加剧的背景下, 树木能否通过这种能力的改变去适应未来快速的气候变化已成为研究的热点问题(Saxe et al., 2001; Allen et al., 2010; Kremer et al., 2012; Shaw & Etterson, 2012; Reyer et al., 2013).研究同质种植园内同一树种不同种源树木的差异及其和种源原地气候条件的关系是一种验证树木对气候适应/驯化的重要方法(Reich et al., 1996; Warren et al., 2005; Martin et al., 2007; Frei et al., 2012). ...
Leaf and wood carbon isotope ratios, specific leaf areas and wood growth of Eucalyptus species across a rainfall gradient in Australia.
1
2006
... 水分利用效率(WUE)是反映植物单位耗水量生产出的同化量的一个重要生理生态参数(Farquhar & Richards, 1984), 是耦合植物叶片光合作用和蒸腾作用的重要指标, 对研究生态系统水循环和碳循环相互作用、揭示植物对全球变化响应和适应对策具有重要意义(Saxe et al., 2001; Gao et al., 2014).气候变化条件下的树木WUE的响应及其机理已成为全球生态学研究的热点(Monclus et al., 2009; de Miguel et al., 2012; Klein et al., 2013).温度和降水是影响WUE的两个重要环境因子.有研究认为当温度升高后光合酶活性提高, 气孔导度(Gs)减小, 从而导致WUE增大(Morecroft et al., 1992; Xiao et al., 2013; Xu et al., 2013); 也有研究认为温度升高会导致胞间CO2浓度(Ci)减小、蒸腾速率(Tr)增大, WUE反而减小(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).降水通过改变土壤水分可利用性和空气湿度, 从而影响树木的气孔关闭和Tr, 进而改变树木的WUE.然而降水和WUE之间关系的研究结果也存在一些不确定性.有研究结果表明二者呈正相关关系(Schulze et al., 1998; Picotte et al., 2007; Niu et al., 2008; Klein et al., 2013); 也有研究认为二者呈负相关关系(Gimenez et al., 1992; Schulze et al., 2006; Monclus et al., 2009; Zhu et al., 2011).此外, 树种间由于生物学特性的差异导致其对环境的适应方式有所不同, 从而引起WUE种间差异.有研究表明, 相同环境下常绿乔木的WUE大于落叶乔木(Brooks et al., 1997; Kloeppel et al., 1998), 而有些研究却得出相反的结果(Damesin et al., 1997; 渠春梅等, 2001).因此, 环境变化对树木WUE的影响以及树木WUE对环境变化响应的内在机理还有待于深入研究. ...
Carbon and nitrogen isotope discrimination and nitrogen nutrition of trees along a rainfall gradient in northern Australia.
1
1998
... 水分利用效率(WUE)是反映植物单位耗水量生产出的同化量的一个重要生理生态参数(Farquhar & Richards, 1984), 是耦合植物叶片光合作用和蒸腾作用的重要指标, 对研究生态系统水循环和碳循环相互作用、揭示植物对全球变化响应和适应对策具有重要意义(Saxe et al., 2001; Gao et al., 2014).气候变化条件下的树木WUE的响应及其机理已成为全球生态学研究的热点(Monclus et al., 2009; de Miguel et al., 2012; Klein et al., 2013).温度和降水是影响WUE的两个重要环境因子.有研究认为当温度升高后光合酶活性提高, 气孔导度(Gs)减小, 从而导致WUE增大(Morecroft et al., 1992; Xiao et al., 2013; Xu et al., 2013); 也有研究认为温度升高会导致胞间CO2浓度(Ci)减小、蒸腾速率(Tr)增大, WUE反而减小(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).降水通过改变土壤水分可利用性和空气湿度, 从而影响树木的气孔关闭和Tr, 进而改变树木的WUE.然而降水和WUE之间关系的研究结果也存在一些不确定性.有研究结果表明二者呈正相关关系(Schulze et al., 1998; Picotte et al., 2007; Niu et al., 2008; Klein et al., 2013); 也有研究认为二者呈负相关关系(Gimenez et al., 1992; Schulze et al., 2006; Monclus et al., 2009; Zhu et al., 2011).此外, 树种间由于生物学特性的差异导致其对环境的适应方式有所不同, 从而引起WUE种间差异.有研究表明, 相同环境下常绿乔木的WUE大于落叶乔木(Brooks et al., 1997; Kloeppel et al., 1998), 而有些研究却得出相反的结果(Damesin et al., 1997; 渠春梅等, 2001).因此, 环境变化对树木WUE的影响以及树木WUE对环境变化响应的内在机理还有待于深入研究. ...
Rapid climate change and the rate of adaptation: Insight from experimental quantitative genetics.
1
2012
... 树木在长期的进化和选择过程中已形成通过改变生理、分子以及遗传等方面特征去响应环境改变的能力, 然而在目前全球气候变化加剧的背景下, 树木能否通过这种能力的改变去适应未来快速的气候变化已成为研究的热点问题(Saxe et al., 2001; Allen et al., 2010; Kremer et al., 2012; Shaw & Etterson, 2012; Reyer et al., 2013).研究同质种植园内同一树种不同种源树木的差异及其和种源原地气候条件的关系是一种验证树木对气候适应/驯化的重要方法(Reich et al., 1996; Warren et al., 2005; Martin et al., 2007; Frei et al., 2012). ...
Enhanced assimilation rate and water use efficiency with latitude through increased photosynthetic capacity and internal conductance in balsam poplar (Populus balsamifera L.).
2
2009
... 作为WUE的两大组分, Pn的增大或Tr的减小都会导致WUE的增大.本研究中落叶松叶片Pn存在显著的种源差异, 并且与种源原地的AI有显著的正相关性(表1; 图1, 图3), 这和以往对来自不同纬度种源树木研究结果相似(Soolanayakanahally et al., 2009; Robson et al., 2012).较高的叶Gs、氮含量和较小的SLA导致了较高的Pn, 并且Pn和种源原地的气候条件具有明显的相关关系, 这是基因调控下的Pn对种源原地环境长期适应的结果(Martin et al., 2007; Savolainen et al., 2007; Soolanayakanahally et al., 2009; Frei et al., 2012).除友好种源外, 随着Pn的增大, WUE表现出相近的变化趋势; 虽然叶片Tr也存在显著的种源差异, 然而WUE并没有随着Tr的变化表现出相应的变化趋势(图1), 这表明WUE受 Pn的影响较大.从研究结果变异程度而言, Pn的变异系数为15.95%, 最高值是最低值的1.60倍, 而Tr的变异系数为9.6%, 最高值较最低值高出44.99%, 由此使得WUE的变异趋势和Pn相近.这是因为叶片的其他生理和结构特征, 如叶片的Gs、NL、SLA等直接影响Pn和Tr, 进而引起了WUE的差异(Gago et al., 2014). ...
... ; Soolanayakanahally et al., 2009; Frei et al., 2012).除友好种源外, 随着Pn的增大, WUE表现出相近的变化趋势; 虽然叶片Tr也存在显著的种源差异, 然而WUE并没有随着Tr的变化表现出相应的变化趋势(图1), 这表明WUE受 Pn的影响较大.从研究结果变异程度而言, Pn的变异系数为15.95%, 最高值是最低值的1.60倍, 而Tr的变异系数为9.6%, 最高值较最低值高出44.99%, 由此使得WUE的变异趋势和Pn相近.这是因为叶片的其他生理和结构特征, 如叶片的Gs、NL、SLA等直接影响Pn和Tr, 进而引起了WUE的差异(Gago et al., 2014). ...
Leaf carbon isotope discrimination and nitrogen content for riparian trees along elevational transects.
1
1997
... 本研究结果表明落叶松NL和WUE间存在显著的正相关关系, 并且来自于高AI地区的种源树木WUE对NL具有更高的敏感性(图2C).有研究表明: 来自低温少雨地区种源的树木具有较高的NL, 以此提高光合能力获取更多的光合产物, 从而适应低温干燥的环境并且从贫瘠的土壤中获取更多的养分和水分(Reich et al., 1996; Weih & Karlsson, 2001; Oleksyn et al., 2003; Tjoelker et al., 2008; Bresson et al., 2011).因此, 树木NL同WUE间多表现为正相关关系(Sparks & Ehleringer, 1997; Monclus et al., 2009; Yan et al., 2014).相同NL水平下来自于高AI种源树木具有更高的WUE, 这对该树种能够在多样的环境中生存具有重要的现实意义.这是因为在干旱贫瘠的环境中土壤氮和水是树木生长的限制因子, 通过提高叶片氮的利用效率获得更多的光合产物、提高WUE, 从而有助于其生长和生存(Kenney et al., 2014; Yan et al., 2014).此外, 在模拟未来气候变化对树木生长乃至生存的影响时, 研究者认为气候的变化将导致落叶松分布区内永冻层融解萎缩, 土壤干燥, 湿润度下降, 最终使得落叶松林不适应新环境而逐步北移, 直至出境(张新时, 1993; 蒋延玲和周广胜, 2001; 李峰等, 2006).本研究发现落叶松树木可以通过调节叶片结构、养分含量、光合生理等形态和生理特征来调节WUE而适应干旱的环境, 这对研究该树种在气候变化下的应对策略和对新环境的适应能力具有重要的参考价值. ...
Coupling of respiration, nitrogen, and sugars underlies convergent temperature acclimation in Pinus banksiana across wide-ranging sites and populations.
1
2008
... 本研究结果表明落叶松NL和WUE间存在显著的正相关关系, 并且来自于高AI地区的种源树木WUE对NL具有更高的敏感性(图2C).有研究表明: 来自低温少雨地区种源的树木具有较高的NL, 以此提高光合能力获取更多的光合产物, 从而适应低温干燥的环境并且从贫瘠的土壤中获取更多的养分和水分(Reich et al., 1996; Weih & Karlsson, 2001; Oleksyn et al., 2003; Tjoelker et al., 2008; Bresson et al., 2011).因此, 树木NL同WUE间多表现为正相关关系(Sparks & Ehleringer, 1997; Monclus et al., 2009; Yan et al., 2014).相同NL水平下来自于高AI种源树木具有更高的WUE, 这对该树种能够在多样的环境中生存具有重要的现实意义.这是因为在干旱贫瘠的环境中土壤氮和水是树木生长的限制因子, 通过提高叶片氮的利用效率获得更多的光合产物、提高WUE, 从而有助于其生长和生存(Kenney et al., 2014; Yan et al., 2014).此外, 在模拟未来气候变化对树木生长乃至生存的影响时, 研究者认为气候的变化将导致落叶松分布区内永冻层融解萎缩, 土壤干燥, 湿润度下降, 最终使得落叶松林不适应新环境而逐步北移, 直至出境(张新时, 1993; 蒋延玲和周广胜, 2001; 李峰等, 2006).本研究发现落叶松树木可以通过调节叶片结构、养分含量、光合生理等形态和生理特征来调节WUE而适应干旱的环境, 这对研究该树种在气候变化下的应对策略和对新环境的适应能力具有重要的参考价值. ...
The influence of fire on carbon distribution and net primary production of boreal Larix gmelinii forests in Northeastern China.
Water availability and branch length determine δ13C in foliage of Pinus pinaster.
1
2000
... 与本文假设相同的是, 来自于低温少雨地区的种源具有较高的WUE, 并且和种源原地年平均气温及平均年降水量均呈线性负相关关系(图3G、3H).这是因为随着温度的升高叶片光合作用酶的活性增强、Gs增大, 进而影响CO2同化速率和Tr, 导致WUE的差异.本研究中17个种源落叶松的Pn和Tr均存在显著的种间差异, 但随着种源原地温度的升高, Tr明显增大, 而Pn总体上并没有表现出明显的增大趋势, 仅在一定的降水区间内和温度存在一定的线性相关(图3A、3D).因此, 本研究中树木Tr的增大是导致WUE和种源原地年平均气温负相关的主要原因.温度和树木WUE间这种负相关性在相近的研究中也有所体现(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).大量种源试验表明树木在干旱环境条件下具有更高的WUE (Anderson et al., 2000; Warren & Adams, 2000; Gouveia & Freitas, 2009; Ran et al., 2013), 这是由于干旱环境下空气湿度和土壤含水量降低、叶片Gs和Tr减小, 而Pn的变化相对较小, 从而导致WUE增大(Farquhar et al., 1989; Zhu et al., 2011; Liu et al., 2013).然而, 本研究结果显示Pn和Tr均与种源原地平均年降水量不相关(图3B、3E), 这是因为降水只考虑了水分收入, 未考虑水分的支出等情况, 不能完全表征一个地方的干旱程度.Warren等(2005)对同质园内来自于水分条件差异明显的15个种源Eucalyptus sideroxylon树木研究表明叶片WUE存在种源差异, 但和降水没有明显相关性, 相似的结果也在对不同种源Eucalyptus microtheca的研究中出现(Li, 2000).因此选用一个较合理的代表气候干旱程度的指标尤为重要, AI作为表征一个地区干湿程度的指标, 近来成为全球变化研究中经常涉及的气候指标之一(Arora, 2002; 孟猛等, 2004).本研究结果表明落叶松针叶WUE和AI显著正相关且相关性高于降水和温度(图3), 这与Warren和Adams (2000)对海岸松4个种源树木的研究结果相一致.此外, AI和Pn也存在线性正相关关系, 并且和不同水分蒸发范围内的Tr存在显著相关关系.这表明干旱情况下Pn的增加和Tr的减小是WUE提高的主要原因(Ramírez-Valiente et al., 2011). ...
Does rainfall explain variation in leaf morphology and physiology among populations of red ironbark (Eucalyptus sideroxylon subsp. tricarpa) grown in a common garden?
3
2005
... SLA作为叶片结构的重要特征之一也是影响WUE的重要因素, 有研究表明随着SLA的减小, 叶片厚度增加、光合组织密度增大, 导致水分的蒸腾减小和固碳能力提高, 从而WUE增大, 因此两者间多表现为负相关关系(Reich et al., 1998; Lamont et al., 2002; Warren et al., 2005; Gago et al., 2014).目前对于两者间关系的研究多集中在树种间(Lamont et al., 2002; Holtum & Winter, 2005)、同一树种不同种源幼苗间(Fernández et al., 1999; Warren et al., 2005)、同一种源不同幼苗个体间(Monclus et al., 2009; Roussel et al., 2009; de Miguel et al., 2012)等方面的表型差异, 而对环境变化下两者间的关系及其调控机理研究较少.de Miguel等(2012)对全同胞家系海岸松幼苗研究发现SLA和WUE存在负相关关系, 并认为这是基因调控的结果并能通过子代遗传, 相似的结果在其他研究中也有所体现(Fernández et al., 1999; Ramírez-Valiente et al., 2011).和我们假设相一致的是, 17个种源落叶松的30年树木SLA具有明显的种源差异并和WUE间存在显著的负相关关系(p < 0.05), 且随着种源原地AI的增大SLA对WUE的影响程度增强(图2B).这种在同质环境中生活多年仍存在显著的种源间差异且和种源原地气候条件有较好关系的结果进一步支持了这种差异是一种基因调控下的对种源原地环境长期适应的结果. ...
... ; Warren et al., 2005)、同一种源不同幼苗个体间(Monclus et al., 2009; Roussel et al., 2009; de Miguel et al., 2012)等方面的表型差异, 而对环境变化下两者间的关系及其调控机理研究较少.de Miguel等(2012)对全同胞家系海岸松幼苗研究发现SLA和WUE存在负相关关系, 并认为这是基因调控的结果并能通过子代遗传, 相似的结果在其他研究中也有所体现(Fernández et al., 1999; Ramírez-Valiente et al., 2011).和我们假设相一致的是, 17个种源落叶松的30年树木SLA具有明显的种源差异并和WUE间存在显著的负相关关系(p < 0.05), 且随着种源原地AI的增大SLA对WUE的影响程度增强(图2B).这种在同质环境中生活多年仍存在显著的种源间差异且和种源原地气候条件有较好关系的结果进一步支持了这种差异是一种基因调控下的对种源原地环境长期适应的结果. ...
... 树木在长期的进化和选择过程中已形成通过改变生理、分子以及遗传等方面特征去响应环境改变的能力, 然而在目前全球气候变化加剧的背景下, 树木能否通过这种能力的改变去适应未来快速的气候变化已成为研究的热点问题(Saxe et al., 2001; Allen et al., 2010; Kremer et al., 2012; Shaw & Etterson, 2012; Reyer et al., 2013).研究同质种植园内同一树种不同种源树木的差异及其和种源原地气候条件的关系是一种验证树木对气候适应/驯化的重要方法(Reich et al., 1996; Warren et al., 2005; Martin et al., 2007; Frei et al., 2012). ...
Growth response of mountain birch to air and soil temperature: Is increasing leaf- nitrogen content an acclimation to lower air temperature?
1
2001
... 本研究结果表明落叶松NL和WUE间存在显著的正相关关系, 并且来自于高AI地区的种源树木WUE对NL具有更高的敏感性(图2C).有研究表明: 来自低温少雨地区种源的树木具有较高的NL, 以此提高光合能力获取更多的光合产物, 从而适应低温干燥的环境并且从贫瘠的土壤中获取更多的养分和水分(Reich et al., 1996; Weih & Karlsson, 2001; Oleksyn et al., 2003; Tjoelker et al., 2008; Bresson et al., 2011).因此, 树木NL同WUE间多表现为正相关关系(Sparks & Ehleringer, 1997; Monclus et al., 2009; Yan et al., 2014).相同NL水平下来自于高AI种源树木具有更高的WUE, 这对该树种能够在多样的环境中生存具有重要的现实意义.这是因为在干旱贫瘠的环境中土壤氮和水是树木生长的限制因子, 通过提高叶片氮的利用效率获得更多的光合产物、提高WUE, 从而有助于其生长和生存(Kenney et al., 2014; Yan et al., 2014).此外, 在模拟未来气候变化对树木生长乃至生存的影响时, 研究者认为气候的变化将导致落叶松分布区内永冻层融解萎缩, 土壤干燥, 湿润度下降, 最终使得落叶松林不适应新环境而逐步北移, 直至出境(张新时, 1993; 蒋延玲和周广胜, 2001; 李峰等, 2006).本研究发现落叶松树木可以通过调节叶片结构、养分含量、光合生理等形态和生理特征来调节WUE而适应干旱的环境, 这对研究该树种在气候变化下的应对策略和对新环境的适应能力具有重要的参考价值. ...
Carbon fluxes, evapotranspiration, and water use efficiency of terrestrial ecosystems in China.
1
2013
... 水分利用效率(WUE)是反映植物单位耗水量生产出的同化量的一个重要生理生态参数(Farquhar & Richards, 1984), 是耦合植物叶片光合作用和蒸腾作用的重要指标, 对研究生态系统水循环和碳循环相互作用、揭示植物对全球变化响应和适应对策具有重要意义(Saxe et al., 2001; Gao et al., 2014).气候变化条件下的树木WUE的响应及其机理已成为全球生态学研究的热点(Monclus et al., 2009; de Miguel et al., 2012; Klein et al., 2013).温度和降水是影响WUE的两个重要环境因子.有研究认为当温度升高后光合酶活性提高, 气孔导度(Gs)减小, 从而导致WUE增大(Morecroft et al., 1992; Xiao et al., 2013; Xu et al., 2013); 也有研究认为温度升高会导致胞间CO2浓度(Ci)减小、蒸腾速率(Tr)增大, WUE反而减小(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).降水通过改变土壤水分可利用性和空气湿度, 从而影响树木的气孔关闭和Tr, 进而改变树木的WUE.然而降水和WUE之间关系的研究结果也存在一些不确定性.有研究结果表明二者呈正相关关系(Schulze et al., 1998; Picotte et al., 2007; Niu et al., 2008; Klein et al., 2013); 也有研究认为二者呈负相关关系(Gimenez et al., 1992; Schulze et al., 2006; Monclus et al., 2009; Zhu et al., 2011).此外, 树种间由于生物学特性的差异导致其对环境的适应方式有所不同, 从而引起WUE种间差异.有研究表明, 相同环境下常绿乔木的WUE大于落叶乔木(Brooks et al., 1997; Kloeppel et al., 1998), 而有些研究却得出相反的结果(Damesin et al., 1997; 渠春梅等, 2001).因此, 环境变化对树木WUE的影响以及树木WUE对环境变化响应的内在机理还有待于深入研究. ...
Climate warming and increasing atmospheric CO2 have contributed to increased intrinsic water-use efficiency on the northeastern Tibetan Plateau since 1850.
1
2013
... 水分利用效率(WUE)是反映植物单位耗水量生产出的同化量的一个重要生理生态参数(Farquhar & Richards, 1984), 是耦合植物叶片光合作用和蒸腾作用的重要指标, 对研究生态系统水循环和碳循环相互作用、揭示植物对全球变化响应和适应对策具有重要意义(Saxe et al., 2001; Gao et al., 2014).气候变化条件下的树木WUE的响应及其机理已成为全球生态学研究的热点(Monclus et al., 2009; de Miguel et al., 2012; Klein et al., 2013).温度和降水是影响WUE的两个重要环境因子.有研究认为当温度升高后光合酶活性提高, 气孔导度(Gs)减小, 从而导致WUE增大(Morecroft et al., 1992; Xiao et al., 2013; Xu et al., 2013); 也有研究认为温度升高会导致胞间CO2浓度(Ci)减小、蒸腾速率(Tr)增大, WUE反而减小(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).降水通过改变土壤水分可利用性和空气湿度, 从而影响树木的气孔关闭和Tr, 进而改变树木的WUE.然而降水和WUE之间关系的研究结果也存在一些不确定性.有研究结果表明二者呈正相关关系(Schulze et al., 1998; Picotte et al., 2007; Niu et al., 2008; Klein et al., 2013); 也有研究认为二者呈负相关关系(Gimenez et al., 1992; Schulze et al., 2006; Monclus et al., 2009; Zhu et al., 2011).此外, 树种间由于生物学特性的差异导致其对环境的适应方式有所不同, 从而引起WUE种间差异.有研究表明, 相同环境下常绿乔木的WUE大于落叶乔木(Brooks et al., 1997; Kloeppel et al., 1998), 而有些研究却得出相反的结果(Damesin et al., 1997; 渠春梅等, 2001).因此, 环境变化对树木WUE的影响以及树木WUE对环境变化响应的内在机理还有待于深入研究. ...
Interactions between CO2 enhancement and N addition on net primary productivity and water-use efficiency in a mesocosm with multiple subtropical tree species.
1
2014
... 本研究结果表明落叶松NL和WUE间存在显著的正相关关系, 并且来自于高AI地区的种源树木WUE对NL具有更高的敏感性(图2C).有研究表明: 来自低温少雨地区种源的树木具有较高的NL, 以此提高光合能力获取更多的光合产物, 从而适应低温干燥的环境并且从贫瘠的土壤中获取更多的养分和水分(Reich et al., 1996; Weih & Karlsson, 2001; Oleksyn et al., 2003; Tjoelker et al., 2008; Bresson et al., 2011).因此, 树木NL同WUE间多表现为正相关关系(Sparks & Ehleringer, 1997; Monclus et al., 2009; Yan et al., 2014).相同NL水平下来自于高AI种源树木具有更高的WUE, 这对该树种能够在多样的环境中生存具有重要的现实意义.这是因为在干旱贫瘠的环境中土壤氮和水是树木生长的限制因子, 通过提高叶片氮的利用效率获得更多的光合产物、提高WUE, 从而有助于其生长和生存(Kenney et al., 2014; Yan et al., 2014).此外, 在模拟未来气候变化对树木生长乃至生存的影响时, 研究者认为气候的变化将导致落叶松分布区内永冻层融解萎缩, 土壤干燥, 湿润度下降, 最终使得落叶松林不适应新环境而逐步北移, 直至出境(张新时, 1993; 蒋延玲和周广胜, 2001; 李峰等, 2006).本研究发现落叶松树木可以通过调节叶片结构、养分含量、光合生理等形态和生理特征来调节WUE而适应干旱的环境, 这对研究该树种在气候变化下的应对策略和对新环境的适应能力具有重要的参考价值. ...
帽儿山地区21年生兴安落叶松种源试验
1
2002
... 本实验所在的同质种植园位于东北林业大学帽儿山森林生态系统研究站内(45.40° N, 127.67° E).在国内落叶松的自然分布范围内选取代表性种源17个(表1), 于1980年秋采种, 1981年育苗, 1983年春将17个种源的二年实生苗栽种在生态站试验样地内.样地设计按完全随机区组设计, 重复5次, 80株小区设计, 双行排列, 按株行距1.5 m × 2.0 m定植, 四周设有保护行(杨传平等, 2002).1997年和2001年间伐两次, 株行距现为4.5 m × 2.5 m.该地区气候属于大陆性温带季风气候, 1989-2009年平均年降水量为629 mm, 平均年蒸发量为864 mm, 年平均气温3.1 ℃, 1月和7月份平均气温分别为-18.5 ℃和22 ℃.同质园内土壤、地形、气候条件一致, 平均海拔300 m, 平均坡度10°-15°, 土壤为暗棕色森林土. ...
帽儿山地区21年生兴安落叶松种源试验
1
2002
... 本实验所在的同质种植园位于东北林业大学帽儿山森林生态系统研究站内(45.40° N, 127.67° E).在国内落叶松的自然分布范围内选取代表性种源17个(表1), 于1980年秋采种, 1981年育苗, 1983年春将17个种源的二年实生苗栽种在生态站试验样地内.样地设计按完全随机区组设计, 重复5次, 80株小区设计, 双行排列, 按株行距1.5 m × 2.0 m定植, 四周设有保护行(杨传平等, 2002).1997年和2001年间伐两次, 株行距现为4.5 m × 2.5 m.该地区气候属于大陆性温带季风气候, 1989-2009年平均年降水量为629 mm, 平均年蒸发量为864 mm, 年平均气温3.1 ℃, 1月和7月份平均气温分别为-18.5 ℃和22 ℃.同质园内土壤、地形、气候条件一致, 平均海拔300 m, 平均坡度10°-15°, 土壤为暗棕色森林土. ...
... 本研究结果表明落叶松NL和WUE间存在显著的正相关关系, 并且来自于高AI地区的种源树木WUE对NL具有更高的敏感性(图2C).有研究表明: 来自低温少雨地区种源的树木具有较高的NL, 以此提高光合能力获取更多的光合产物, 从而适应低温干燥的环境并且从贫瘠的土壤中获取更多的养分和水分(Reich et al., 1996; Weih & Karlsson, 2001; Oleksyn et al., 2003; Tjoelker et al., 2008; Bresson et al., 2011).因此, 树木NL同WUE间多表现为正相关关系(Sparks & Ehleringer, 1997; Monclus et al., 2009; Yan et al., 2014).相同NL水平下来自于高AI种源树木具有更高的WUE, 这对该树种能够在多样的环境中生存具有重要的现实意义.这是因为在干旱贫瘠的环境中土壤氮和水是树木生长的限制因子, 通过提高叶片氮的利用效率获得更多的光合产物、提高WUE, 从而有助于其生长和生存(Kenney et al., 2014; Yan et al., 2014).此外, 在模拟未来气候变化对树木生长乃至生存的影响时, 研究者认为气候的变化将导致落叶松分布区内永冻层融解萎缩, 土壤干燥, 湿润度下降, 最终使得落叶松林不适应新环境而逐步北移, 直至出境(张新时, 1993; 蒋延玲和周广胜, 2001; 李峰等, 2006).本研究发现落叶松树木可以通过调节叶片结构、养分含量、光合生理等形态和生理特征来调节WUE而适应干旱的环境, 这对研究该树种在气候变化下的应对策略和对新环境的适应能力具有重要的参考价值. ...
Evaluating the effects of future climate change and elevated CO2 on the water use efficiency in terrestrial ecosystems of China.
4
2011
... 水分利用效率(WUE)是反映植物单位耗水量生产出的同化量的一个重要生理生态参数(Farquhar & Richards, 1984), 是耦合植物叶片光合作用和蒸腾作用的重要指标, 对研究生态系统水循环和碳循环相互作用、揭示植物对全球变化响应和适应对策具有重要意义(Saxe et al., 2001; Gao et al., 2014).气候变化条件下的树木WUE的响应及其机理已成为全球生态学研究的热点(Monclus et al., 2009; de Miguel et al., 2012; Klein et al., 2013).温度和降水是影响WUE的两个重要环境因子.有研究认为当温度升高后光合酶活性提高, 气孔导度(Gs)减小, 从而导致WUE增大(Morecroft et al., 1992; Xiao et al., 2013; Xu et al., 2013); 也有研究认为温度升高会导致胞间CO2浓度(Ci)减小、蒸腾速率(Tr)增大, WUE反而减小(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).降水通过改变土壤水分可利用性和空气湿度, 从而影响树木的气孔关闭和Tr, 进而改变树木的WUE.然而降水和WUE之间关系的研究结果也存在一些不确定性.有研究结果表明二者呈正相关关系(Schulze et al., 1998; Picotte et al., 2007; Niu et al., 2008; Klein et al., 2013); 也有研究认为二者呈负相关关系(Gimenez et al., 1992; Schulze et al., 2006; Monclus et al., 2009; Zhu et al., 2011).此外, 树种间由于生物学特性的差异导致其对环境的适应方式有所不同, 从而引起WUE种间差异.有研究表明, 相同环境下常绿乔木的WUE大于落叶乔木(Brooks et al., 1997; Kloeppel et al., 1998), 而有些研究却得出相反的结果(Damesin et al., 1997; 渠春梅等, 2001).因此, 环境变化对树木WUE的影响以及树木WUE对环境变化响应的内在机理还有待于深入研究. ...
... ; Zhu et al., 2011).此外, 树种间由于生物学特性的差异导致其对环境的适应方式有所不同, 从而引起WUE种间差异.有研究表明, 相同环境下常绿乔木的WUE大于落叶乔木(Brooks et al., 1997; Kloeppel et al., 1998), 而有些研究却得出相反的结果(Damesin et al., 1997; 渠春梅等, 2001).因此, 环境变化对树木WUE的影响以及树木WUE对环境变化响应的内在机理还有待于深入研究. ...
... 与本文假设相同的是, 来自于低温少雨地区的种源具有较高的WUE, 并且和种源原地年平均气温及平均年降水量均呈线性负相关关系(图3G、3H).这是因为随着温度的升高叶片光合作用酶的活性增强、Gs增大, 进而影响CO2同化速率和Tr, 导致WUE的差异.本研究中17个种源落叶松的Pn和Tr均存在显著的种间差异, 但随着种源原地温度的升高, Tr明显增大, 而Pn总体上并没有表现出明显的增大趋势, 仅在一定的降水区间内和温度存在一定的线性相关(图3A、3D).因此, 本研究中树木Tr的增大是导致WUE和种源原地年平均气温负相关的主要原因.温度和树木WUE间这种负相关性在相近的研究中也有所体现(Morecroft et al., 1992; Nicotra et al., 2008; Zhu et al., 2011).大量种源试验表明树木在干旱环境条件下具有更高的WUE (Anderson et al., 2000; Warren & Adams, 2000; Gouveia & Freitas, 2009; Ran et al., 2013), 这是由于干旱环境下空气湿度和土壤含水量降低、叶片Gs和Tr减小, 而Pn的变化相对较小, 从而导致WUE增大(Farquhar et al., 1989; Zhu et al., 2011; Liu et al., 2013).然而, 本研究结果显示Pn和Tr均与种源原地平均年降水量不相关(图3B、3E), 这是因为降水只考虑了水分收入, 未考虑水分的支出等情况, 不能完全表征一个地方的干旱程度.Warren等(2005)对同质园内来自于水分条件差异明显的15个种源Eucalyptus sideroxylon树木研究表明叶片WUE存在种源差异, 但和降水没有明显相关性, 相似的结果也在对不同种源Eucalyptus microtheca的研究中出现(Li, 2000).因此选用一个较合理的代表气候干旱程度的指标尤为重要, AI作为表征一个地区干湿程度的指标, 近来成为全球变化研究中经常涉及的气候指标之一(Arora, 2002; 孟猛等, 2004).本研究结果表明落叶松针叶WUE和AI显著正相关且相关性高于降水和温度(图3), 这与Warren和Adams (2000)对海岸松4个种源树木的研究结果相一致.此外, AI和Pn也存在线性正相关关系, 并且和不同水分蒸发范围内的Tr存在显著相关关系.这表明干旱情况下Pn的增加和Tr的减小是WUE提高的主要原因(Ramírez-Valiente et al., 2011). ...
... ; Zhu et al., 2011; Liu et al., 2013).然而, 本研究结果显示Pn和Tr均与种源原地平均年降水量不相关(图3B、3E), 这是因为降水只考虑了水分收入, 未考虑水分的支出等情况, 不能完全表征一个地方的干旱程度.Warren等(2005)对同质园内来自于水分条件差异明显的15个种源Eucalyptus sideroxylon树木研究表明叶片WUE存在种源差异, 但和降水没有明显相关性, 相似的结果也在对不同种源Eucalyptus microtheca的研究中出现(Li, 2000).因此选用一个较合理的代表气候干旱程度的指标尤为重要, AI作为表征一个地区干湿程度的指标, 近来成为全球变化研究中经常涉及的气候指标之一(Arora, 2002; 孟猛等, 2004).本研究结果表明落叶松针叶WUE和AI显著正相关且相关性高于降水和温度(图3), 这与Warren和Adams (2000)对海岸松4个种源树木的研究结果相一致.此外, AI和Pn也存在线性正相关关系, 并且和不同水分蒸发范围内的Tr存在显著相关关系.这表明干旱情况下Pn的增加和Tr的减小是WUE提高的主要原因(Ramírez-Valiente et al., 2011). ...



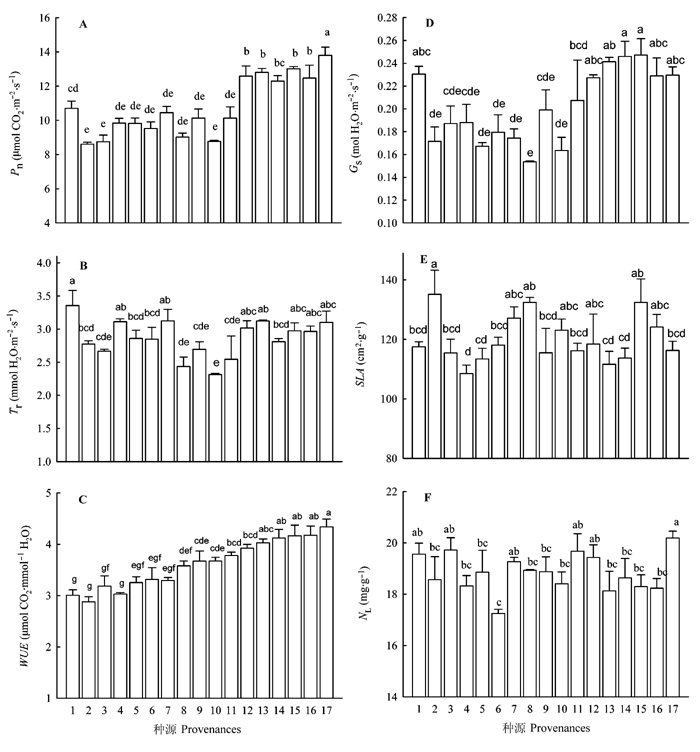
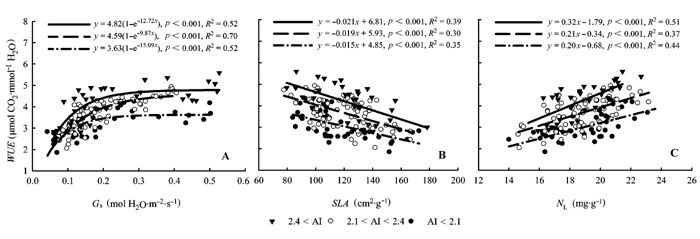
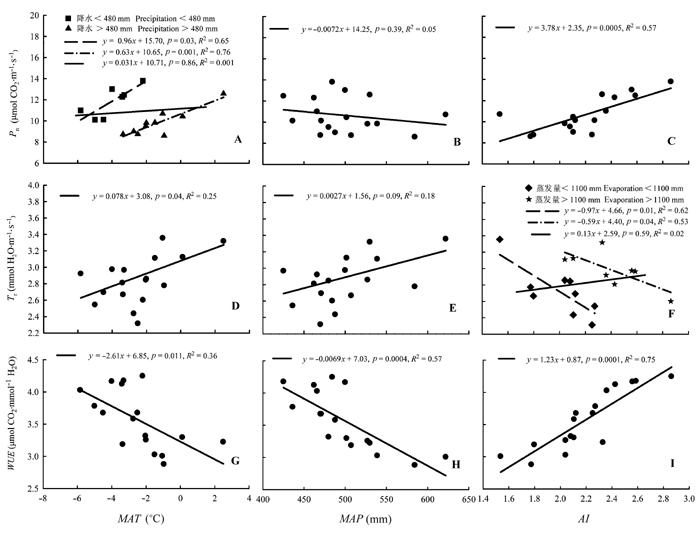
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}