植物功能性状与环境和生态系统功能的关系是近年来生态学和全球变化研究的热点(Harrison et al ., 2010 ; Bernard-Verdier et al ., 2012 ), 各功能性状之间的权衡和组合决定了植物的生活史对策(Vandermeer, 1972 ; Silvertown, 2004 )。在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ), 叶片是植物进行光合作用的主要器官(张林和罗天祥, 2004 ), 两者大小和数量的配置关系直接影响着植物的机械力学和水力结构特征, 从而决定了植物的生活史策略, 并可能影响物种生态位的利用(Givnish, 1987 ; Enquist et al ., 1999 ; West et al ., 1999 ; Westoby et al ., 2002 )。植物单位大小枝条上支撑的叶面积必须与支撑枝的横截面积成一定的比例, 才能符合植物器官间的机械力学和水力结构需求(Niklas, 1992 ; Enquist et al ., 1999 ; Normand et al ., 2008 ; Olson et al ., 2009 )。叶片大小和叶片数量是决定树冠紧密程度的两个重要因素, 直接影响到植株的冠层结构和发育模式, 进而影响到植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 )。在某一给定的叶生物量分配或某一个给定的总叶面积下, 植物可能有较少的大叶子或者较多的小叶子(杨冬梅等, 2012a )。坡向作为一种重要的地形要素, 往往对局部区域的太阳辐射和降水进行二次分配(Cantón et al ., 2004 ), 在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 能够对植物的形态产生影响(Bazzaz, 1996 ; 韦莉莉等, 2005 )。当自然资源的丰富度低于或超过植物光合系统所能利用的最低或最高限度时, 植物的光合作用将被抑制(王玉萍等, 2013 )。为提高植物构型构建的生境适应性和风险规避能力, 植物通过功能性状等表型的可塑性调整提高自身的生存适合度, 体现了全球气候变化背景下植物的资源配置策略。因此, 研究坡向梯度上植物枝叶关系规律及其成因, 对于理解植物的环境适应机制有着重要意义。
刺槐(Robinia pseudoacacia )是豆科刺槐属的温带落叶乔木, 具有生长速度快、根系发达、对环境适应性强、固碳氮能力较好、成活率高、萌芽力和根蘖性都很强等优点, 已成为我国黄土丘陵区的主要造林树种和人工林面积最大的树种(于占辉等, 2009 ; 李军等, 2010 )。近年来, 干旱地区许多刺槐人工林出现了生长减慢、枝梢干枯甚至整株死亡的衰败现象, 引起了学术界的广泛关注(单长卷等, 2004 ; 李军等, 2010 )。目前, 许多学者研究了刺槐叶面积生长与林分密度的关系(尹婧等, 2008 ), 土层厚度对刺槐旱季水分状况和生长的影响(王林等, 2013 ), 不同林龄和退耕年限刺槐人工林碳、氮储量及分配规律(申家朋和张文辉, 2014 ; 艾泽民等, 2014 ), 揭示了刺槐生长以及生物量积累和分配格局等一系列规律, 但关于微地形影响下刺槐当年生小枝内枝叶关系的研究较为少见, 更鲜有不同坡向茎截面积与总叶面积、出叶强度与单叶面积关系的报道, 限制了对异质生境中刺槐适应机制的解析。本研究选择兰州北山九州台西北师范大学绿化基地的刺槐人工林为研究对象, 探讨刺槐当年生小枝内的茎截面积-总叶面积、出叶强度-单叶面积关系, 为进一步了解刺槐的环境适应机制提供理论依据, 对提高干旱半干旱地区人工林的可持续性和稳定性具有重要意义。
1 研究地区和研究方法
1.1 研究区和样地概况
研究区位于兰州市北山九州台区西北师范大学绿化基地, 地理位置为36.17°-36.23° N, 103.21°- 103.25° E, 海拔1536-1914 m, 面积857.04 hm2 , 属北温带半干旱大陆性季风气候, 年平均气温5-9 ℃, ≥10 ℃的积温3385.4 ℃, 年降水量250-360 mm, 主要集中在6-9月。土壤为黄土母质上发育起来的灰钙土, 植被群落以刺槐人工林和侧柏人工林为主体, 其中刺槐人工林主要位于海拔1545-1782 m的各个坡向上, 主要林下植物有红砂(Reaumuria soongarica )、枸杞(Lycium chinense )、柽柳(Tamarix chinensis )、柠条锦鸡儿(Caragana korshinskii )等。供试的刺槐林建植于2002年, 造林时沿等高线挖间距2 m左右的水平沟, 在水平沟内以2 m的株距栽植刺槐, 造林平均密度为2500株·hm-2 ; 人工林每年用提灌的黄河水灌溉4次, 并不定期进行病虫害防治, 期间不再进行补种。
1.2 实验设计
在地形图上将研究区划分为4个坡向, 各坡向沿垂直海拔梯度(1550-1750 m)间隔50 m布置长180 m、宽10 m的水平样带1条, 总计20条水平样带。在多次现场踏勘的基础上, 每条水平样带上间隔5 m设置12个10 m × 10 m的样方, GPS测量每个样方的经纬度和海拔。针对各样方内胸径≥1 cm的每木个体进行群落学特征调查, 记录株高、胸径、冠幅等, 测定各样方的群落高度和郁闭度。
1.3 野外取样和测量
野外取样和室内测量于2014年8-9月的刺槐生长旺季完成。首先进行土壤水分采样, 每个样方中用土钻(直径= 4 cm)在0-30 cm土层范围分3层间隔10 cm取土样各1份, 重复3次, 同时取0-30 cm混合土样1份, 重复3次, 所有样品均剔除明显的植物根段和凋落物等杂质, 装入编号的铝盒中, 带回实验室; 然后对样方内所有刺槐进行枝叶取样, 株高2 m以下的刺槐, 直接从冠层中部外层4个方位剪取4个无明显叶片损失的当年生小枝, 株高2 m以上的刺槐, 用高枝剪获取树冠中部外层4个方位的枝条各1枝, 放入塑封袋, 编码后带回实验室进一步 处理。
样品带回实验室后, 在6 h内完成以下测量: 称量土壤样品的质量, 然后在105 ℃的烘箱内烘干12 h, 取出称量, 计算出各层及0-30 cm土层的土壤质量含水量; 对于每个当年生小枝, 首先分别用直尺和游标卡尺测量其长度和最粗直径, 然后清点记录叶片数并摘除, 待分别称量完枝条和叶片的鲜质量后, 再测量当年生小枝上每个叶片的叶面积(通过扫描每一个分枝上的所有叶片, 用MapInfo软件进行计算)。最后, 各样品置于烘箱中烘干后称量, 以备其他分析。
1.4 空间数据
利用ArcGIS对研究区1:5万地形图进行数字化, 得到研究区域10 m分辨率的数字高程模型(digital elevation mode, DEM)。在DEM的基础上利用Spatial Analyst模块中的Surface Analysis命令提取坡向, 它表示地表面上一点的切平面的法线矢量n 在水平面的投影nxoy 与过该点的正北方向的夹角, 其中坡向组别以国际划分法确定(陈瑶等, 2006 ), 具体方法参见汤国安等(2010) 文献。
1.5 数据分析
将坡向划分为北坡(315°-360°和0°-45°)、东坡(45°-135°)、南坡(135°-225°)和西坡(225°-315°)。在所研究的4个功能性状中, 茎截面积指当年生小枝上1级枝的横截面积, 总叶面积指当年生小枝上所支撑的所有叶片面积之和, 出叶强度指单位体积枝条所支撑的叶片数, 单叶面积指当年生小枝上所支撑的所有叶片单叶面积的平均值, 枝条体积由其横截面积乘以长度获得(许月等, 2012 )。对每个样方刺槐枝叶属性的平均值进行对数(以10为底)转换后再进行分析, 对数转换使之符合正态分布。对枝大小-叶大小以及叶数量-叶大小两组功能性状关系的研究, 均采用y = axb , 线性转换成log(y ) = log(a ) + b log(x ), 式中x 和y 表示两个特征参数, a 为性状关系的截距, b 为斜率, 即异速生长参数或相对生长的指数, 当|b | = 1时, 表示两者是等速变化关系; 当|b| 显著偏离1时, 两者间为异速变化关系(Harvey & Pagel, 1991 )。数据分析主要采用标准化主轴估计(standardized major axis estimation, SMA)的方法(Warton et al ., 2006 ), 由软件(S)MATR Version 2.0 (Falster et al ., 2006 )计算完成。每一个回归斜率的置信区间根据Pitman (1939)方法计算。回归斜率与1或-1的显著性检验根据Warton和Weber (2002)的方法判断。在完成线性回归后, 对回归斜率进行异质性测试, 如果组间没有异质性, 则表示有共同斜率(Warton & Weber, 2002 )。此后, 不同坡向梯度间的功能性状的平均值的比较采用单因素方差分析(one-way ANOVA), 利用成对比较检验分析各坡向刺槐枝叶关系线性回归方程在y 轴上截距的差异。
2 结果分析
2.1 不同坡向刺槐林样地群落和土壤水分特征
不同坡向上刺槐林样带群落的郁闭度、高度和土壤水分的变化如表1 所示。随着坡向由北坡向东坡、南坡和西坡转变, 刺槐林样带群落的郁闭度、高度和土壤水分特征呈现先减小后增大的趋势, 且在东坡和西坡之间无显著差异(p > 0.05)。从北坡到南坡, 群落郁闭度减小了22.68%, 植株高度减小了24.29%, 土壤水分含量减小了22.12%。表明在4个坡向中, 北坡生境条件最适宜植株的生长。
2.2 不同坡向刺槐枝叶性状
不同坡向上刺槐枝与叶性状的变化如表2 所示。随着坡向由北坡向东坡、南坡和西坡转变, 刺槐小枝茎截面积、总叶面积和单叶面积呈现先减小后增大的趋势, 北坡比南坡分别减小了9.92%、33.51%和30.69%, 刺槐出叶强度则呈现先增大后减小的趋势, 北坡比南坡增大了50.00%。这些性状值在东坡和西坡之间无显著差异(p > 0.05)。
2.3 不同坡向刺槐茎截面积-总叶面积关系
在各坡向上, 刺槐小枝茎截面积和总叶面积间均显著正相关, 回归斜率显著大于1 (p < 0.05), 且在所有坡向上存在共同斜率1.17 (95%的置信区间CI = (1.15, 1.21), p = 0.51)(图1 ), 表明总叶面积的增加比率大于茎截面积的增加比率。北坡刺槐的茎截面积和总叶面积回归方程的截距显著大于南坡刺槐(p < 0.05)(图1 ), 东坡、西坡刺槐的茎截面积和总叶面积回归方程的截距之间无显著差异(p > 0.05), 表明北坡刺槐比南坡刺槐在单位茎截面积上会支撑更多的叶面积(表2 )。
图1 刺槐茎截面积与总叶面积的关系。A, 北坡; B, 东坡; C, 南坡; D, 西坡。
Fig. 1 Relationship between the cross-sectional area of twigs and total leaf area in Robinia pseudoacacia . A, north slope; B, east slope; C, south slope; D, west slope.
2.4 不同坡向刺槐出叶强度-单叶面积关系
在各坡向上, 刺槐小枝出叶强度和单叶面积间均显著负相关, 回归斜率与-1.00无显著差异(p > 0.05), 且在所有坡向上存在共同斜率-0.99 (95%的置信区间CI = (-1.09, -0.92), p = 0.57)(图2 ), 表明刺槐当年生小枝上支持的叶片存在大小和数量间的等速权衡关系。北坡刺槐的出叶强度和单叶面积回归方程的截距显著大于南坡刺槐(p < 0.05)(图2 ), 东坡、西坡刺槐的出叶强度和单叶面积回归方程的截距之间无显著差异(p > 0.05), 表明北坡刺槐比南坡刺槐具有较大的单叶面积和较小的出叶强度(表2 )。
图2 刺槐出叶强度与单叶面积的关系。A, 北坡; B, 东坡; C, 南坡; D, 西坡。
Fig. 2 Relationship between leafing intensity and individual leaf area in Robinia pseudoacacia . A, north slope; B, east slope; C, south slope; D, west slope.
3 讨论
植物在长期的生长发育过程中, 各构件在异质生境条件下会表现出不同的生态对策(Yang et al ., 2008 ), 植物外部形态具有高度的可塑性特征, 使其能够把资源最佳地分配到不同的器官, 以规避环境风险。本研究发现, 在各坡向间, 刺槐茎截面积和总叶面积均维持显著的正相关关系(p < 0.05), 回归方程间存在共同斜率且异速生长斜率大于1, 此结果与其他枝叶大小关系研究的结果基本一致(Westoby & Wright, 2003 ; Normand et al ., 2008 ; 史青茹等, 2014 ), 出叶强度和单叶面积则表现出显著的等速负相关关系(p < 0.05)。两组关系的回归方程截距均为北坡最大、南坡最小, 这主要与坡向引起环境条件的变化和刺槐枝叶形态在异质生境中的适应机制有关。
3.1 刺槐枝截面积与总叶面积的关系
植物功能性状与生存环境之间是一种相互协同进化与适应的关系(侯兆疆等, 2014 )。枝叶大小关系是植物的重要生活史策略(Westoby et al ., 2002 ), 其比例不仅反映枝叶生物量的分配和叶片在生化和水力生物学方面的功能需求(Brouat et al ., 1998 ; Sun et al ., 2006 ; Normand et al ., 2008 ), 也体现了植物适应特定环境的构型塑造(Enquist & Niklas, 2001 , 2002 )。各坡向上刺槐茎截面积和总叶面积均呈正的异速生长关系, 回归方程间具有共同的SMA斜率且显著大于1 (p < 0.05)(图1 ), 二者的相关关系始终符合Corner法则(Corner, 1949 ), 不随微地形特征的变化而显著改变, 植物对总叶面积的投入显著高于对枝横截面积的投入, 这说明微地形差异无法改变植物在枝叶大小上的协调, 枝大小和叶大小间始终需要维持几何大小和重力的平衡(史青茹等, 2014 )。对于此现象, Westoby等(2002) 曾从枝叶的形态和生理关联性方面进行了解释, 他们认为: 一定大小的叶面积必须要由一定大小的枝来支撑, 两者间必须保持大小的匹配, 满足机械力学和水分动力学的要求, 才能保持机械支撑的平衡以及水分运输和蒸腾的协调; 管道模型理论(Niklas, 1992 )则提出, 为满足叶片对水分的需求, 木质部的横截面积与它们所支持的叶面积成正比。除此之外, West等(1997) 提出了结构分形自相似假说解释这种现象, 即: 所有生物的各器官间具有自相似的分形网络结构, 自然选择的压力必然使其向最优化的方向进化, 使得与外界环境的交换面积和新陈代谢效率最大化, 并使营养物质运输距离和时间最小化, 因此, 枝叶的大小必然按一定的比例来协调。而茎截面积-总叶面积关系的SMA斜率显著大于1 (p < 0.05), 主要是因为叶片是植株碳获取的主要功能构件, 所以在进化过程中为提高光合收益, 植株将尽可能地使分配到叶片中的生物量最大化, 而在叶片支撑结构中的投资最小化。
坡向虽然对刺槐枝叶大小回归方程的斜率没有显著影响, 但使回归方程的截距产生了显著改变。在茎截面积-总叶面积关系中, 其回归方程的截距在北坡显著大于南坡(p < 0.05), 表明在茎截面积一定时, 位于北坡的刺槐在单位大小枝条上支持的总叶面积大于位于南坡的刺槐。叶片大小是植物适应环境中热量和水分交换平衡的结果, 其变化主要是通过影响叶边缘对热量、CO2 和水分的传导能力来影响整个植株的生长速率(杨冬梅等, 2012b )。南坡光照时间长, 水分蒸发大(殷秀琴, 2004 ), 土壤水分含量低(表1 ), 在西北年降水量小的干旱半干旱地区, 由于高温常引起蒸腾作用加强而使植物体水分大量丧失, 因而这种高温和干旱环境严重影响了植物的新陈代谢活动, 细胞的分裂和生长受到限制, 植物生长缓慢, 叶片较小; 然而北坡气温降低, 地面蒸发量减少, 土壤水分增加(表1 ), 良好的水热条件有利于细胞体积和数目增加, 从而促进了叶面积的扩展(李钰等, 2013 )。反过来, 与小叶相比, 大叶具有更厚的叶边缘, 热量和物质通过叶边缘进行转运的阻力更大, 往往会阻止叶片与周围的空气进行热量交换, 减慢CO2 和水蒸气进出叶片的扩散速率, 使大叶比小叶更容易过热, 植物的呼吸和蒸腾成本更高(Givnish, 1978 ), 因此在冷凉、湿润或者荫蔽的环境下大叶由于能够使光合收益最大化而占有优势(Givnish, 1979 ), 而相对小的叶片因为具有较高的热交换能力, 被认为在热、干、高光和低养分的环境下较有优势(Ackerly & Preston, 2003 ; McDonald et al ., 2003 )。另外, 大叶片植物在水分较少的生境中不具有选择优势, 与植物的水力结构特征有关。随着土壤湿度的下降, 水分供应不足会增加水分从土壤运送到叶片的连续水柱断裂的风险, 当大叶片植物的蒸腾速率增大时, 空穴现象和栓塞现象更易发生(McCulloh & Sperry, 2005 ), 从而使得大叶片植物在水分稀缺的生境下不具有水分运输方面的选择优势, 这与Callaway等(1994) 的研究结果一致, 即: 一定大小枝条输导面积支持的总叶面积随着大气蒸腾强度和干旱强度的增加而减小。
3.2 刺槐出叶强度与单叶大小的关系
在各坡向上, 刺槐出叶强度与单叶面积均表现出负的等速生长关系, 说明刺槐的叶存在大小和数量上的生态权衡, 这与杨冬梅等(2012a) 的研究结果一致。位于北坡的刺槐出叶强度小于南坡, 单叶面积大于南坡(表2 ), 如前所述, 叶大小-叶数量的权衡关系是植物对外部环境选择压力的功能性适应结果。北坡光照资源最弱, 且该坡向上群落郁闭程度和拥挤程度大于阳坡(表1 ), 邻体干扰增强, 密度制约改变了植物间可获得性资源的数量, 为使光合效益最大化, 刺槐当年生小枝上长出较大的叶片, 用以增加光合同化面积; 与此同时, 过多的叶片会造成上部叶片对下部叶片的遮挡(李钰等, 2013 ), 使得叶片间互相拥挤的程度加剧, 不利于光资源的充分利用, 因此北坡刺槐形成了“增大单叶面积, 减小出叶强度”的生长策略, 南坡刺槐则形成了“减小单叶面积, 增大出叶强度”的生长策略, 有利于植株本身的光拦截能力和光合碳获取能力最大化。
除此之外, 叶大小-叶数量权衡关系的坡向差异性还可以从以下两个方面解释: (1)植物内在生物量分配限制的结果。由表2 可知, 北坡(阴坡)刺槐单叶面积大于南坡(阳坡), 在植物体上, 在某一给定的叶生物量或某一给定的总叶面积下, 植物要么是有较少的大叶子, 要么是有较多的小叶子, 增加单个叶片的投资意味着叶片数量的减少(杨冬梅等, 2012a ); (2)单叶面积的差异是出叶强度变化的副产物。每个叶片通常都会有一个腋芽分生组织(Bell, 1993 ), 叶片更多的物种, 单位个体大小的腋芽分生组织的密度将会更高(杨冬梅等, 2012b ), 更大的芽库由于有更多的部位具有再生能力, 既可以生长为植株的营养结构或繁殖结构, 也可以保持无活性状态, 进而影响到植株的大小构建和繁殖产出(Bonser & Aarssen, 1996 ), 使植株具有更大的可塑性和更新复壮能力, 因此在南坡干、热、高光和低养分的环境下, 刺槐倾向于具有更高的出叶强度, 叶大小与出叶强度紧密相关, 因此小叶片可能是选择了高的出叶强度的结果。东坡和西坡枝叶功能性状及其关系之间没有明显差异, 主要是因为东坡和西坡属于半阴坡和半阳坡, 皆处于阴坡和阳坡之间的过渡区域, 群落组成也较为复杂, 因此刺槐采取相同的生长策略来适应生境的变化。
本研究发现, 生境对枝叶大小关系有显著的影响, 不同坡向上刺槐枝叶功能性状存在差异。在给定的枝大小下, 处于有利生境的刺槐选择较大的叶面积和较小的出叶强度, 处于不利生境的刺槐则选择较小的叶面积和较大的出叶强度, 这种结果较好地验证了植物通过功能性状的可塑性调整响应环境变异的协同适应性机制。建议在干旱半干旱地区营造人工林时, 根据造林树种在不同坡向的枝叶生长适应性策略, 提高林分对水、热和养分资源的利用效率, 这对干旱半干旱地区人工林的可持续性和稳定性具有重要意义。本文仅从坡向角度分析了枝截面积-总叶面积以及出叶强度-单叶面积关系的变化特征, 它们的关系可能还受到坡度等其他微地形因子以及各物种遗传特性的影响, 并且枝大小-枝数量和其他叶内功能性状对环境的响应等问题, 还需要进一步的实验验证。
致谢 感谢西北师范大学绿化基地胡正军工程师在野外工作中给予的指导和帮助。
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Ackerly DD Preston KA 2003 ). Hydraulic architecture and the evolution of shoot allometry in contrasting climates.
American Journal of Botany , 90 , 1502 -1512 .
[本文引用: 1]
[2]
Ai ZM Chen YM Cao Y 2014 ). Storage and allocation of carbon and nitrogen in Robinia pseudoacacia plantation at different ages in the loess hilly region, China.
Chinese Journal of Applied Ecology , 25 , 333 -341 .(in Chinese with English abstract)
[本文引用: 1]
[艾泽民 , 陈云明 , 曹扬 (2014 ). 黄土丘陵区不同林龄刺槐人工林碳、氮储量及分配格局
. 应用生态学报 , 25 , 333 -341 .]
[本文引用: 1]
[3]
Bazzaz FA 1996 ). Plants in Changing Environments: Linking Physiological, Population, and Community Ecology . Cambridge University Press , Cambridge, UK .
[本文引用: 1]
[4]
Bell AD 1993 ). Plant Form: An Illustrated Guide to Flowering Plant Morphology . Oxford University Press , New York .
[本文引用: 1]
[5]
Bernard-Verdier M Navas ML Vellend M 2012 ). Community assembly along a soil depth gradient: Contrasting patterns of plant trait convergence and divergence in a Mediterranean rangeland.
Journal of Ecology , 100 , 1422 -1433 .
[本文引用: 1]
[6]
Bonser SP Aarssen LW 1996 ). Meristem allocation: A new classification theory for adaptive strategies in herbaceous plants.
Oikos , 77 , 347 -352 .
[本文引用: 1]
[7]
Brouat C Gibernau M Amsellem L McKey D 1998 ). Corner’s rules revisited: Ontogenetic and interspecific patterns in leaf-stem allometry.
New Phytologist , 139 , 459 -470 .
[本文引用: 1]
[8]
Callaway RM Delucia EH Schlesinger WH 1994 ). Biomass allocation of montane and desert ponderosa pine: An analog for response to climate change.
Ecology , 75 , 1474 -1481 .
[9]
Cantón Y del Barrio G Solé Benet A 2004 ). Topographic controls on the spatial distribution of ground cover in the Tabernas badlands of SE Spain.
Catena , 55 , 341 -365 .
[本文引用: 1]
[10]
Chen Y Xu X Zhang DR Wei Y 2006 ). Correlations between vegetation distribution and topographical factors in the northwest of Longmen Mountain, Sichuan Province.
Chinese Journal of Ecology , 25 , 1052 -1055 .(in Chinese with English abstract)
[本文引用: 1]
[陈瑶 , 胥晓 , 张德然 , 魏勇 (2006 ). 四川龙门山西北部植被分布与地形因子的相关性
. 生态学杂志 , 25 , 1052 -1055 .]
[本文引用: 1]
[11]
Corner EJH 1949 ). The durian theory or the origin of the modern tree.
Annals of Botany , 13 , 367 -414 .
[本文引用: 1]
[12]
Enquist BJ Niklas KJ 2001 ). Invariant scaling relations across tree-dominated communities.
Nature , 410 , 655 -660 .
[本文引用: 1]
[13]
Enquist BJ Niklas KJ 2002 ). Global allocation rules for patterns of biomass partitioning in seed plants.
Science , 295 , 1517 -1520 .
[本文引用: 1]
[14]
Enquist BJ West GB Charnov EL Brown JH 1999 ). Allometric scaling of production and life-history variation in vascular plants.
Nature , 401 , 907 -911 .
[本文引用: 2]
[15]
Falster DS Warton DI Wright IJ 2006 ). User’s Guide to SMATR: Standardised Major Axis Tests & Routines Version 2.0
. . Cited 2014-11-20.
URL
[本文引用: 2]
[16]
Givnish TJ 1978 ). Ecological aspects of plant morphology: Leaf form in relation to environment.
Acta Biotheoretica , 27 , 83 -142 .
[17]
Givnish TJ 1979 ). On the adaptive significance of leaf form. In: Solbrig OT, Jain S, Johnson GB eds. Topics in Plant Population Biology . Columbia University Press , New York . 375 -407 .
[本文引用: 2]
[18]
Givnish TJ 1987 ). Comparative studies of leaf form: Assessing the relative roles of selective pressures and phylogenetic constraints.
New Phytologist , 106 (Suppl. ), 131 -160 .
[19]
Givnish TJ Vermeij GJ 1976 ). Sizes and shapes of liane leaves.
The American Naturalist , 110 , 743 -778 .
[本文引用: 2]
[20]
Harrison SP Prentice IC Barboni D Kohfeld KE Ni J Sutra JP 2010 ). Ecophysiological and bioclimatic foundations for a global plant functional classification.
Journal of Vegetation Science , 21 , 300 -317 .
[21]
Harvey PH Pagel MD 1991 ). The Comparative Method in Evolutionary Biology . Oxford University Press , Oxford, UK .
[本文引用: 1]
[22]
Hou ZJ Zhao CZ Li Y Zhang Q Ma XL 2014 ). Trade-off between height and branch numbers in Stellera chamaejasme on slopes of different aspects in a degraded alpine grassland.
Chinese Journal of Plant Ecology , 38 , 281 -288 .(in Chinese with English abstract)
[本文引用: 1]
[侯兆疆 , 赵成章 , 李钰 , 张茜 , 马小丽 (2014 ). 不同坡向高寒退化草地狼毒株高和枝条数的权衡关系
. 植物生态学报 , 38 , 281 -288 .]
[本文引用: 1]
[23]
Li J Wang XC Shao MA Zhao YJ Li XF 2010 ). Simulation of biomass and soil desiccation of Robinia pseudoacacia forestlands on semi-arid and semi-humid regions of China’s Loess Plateau.
Chinese Journal of Plant Ecology , 34 , 330 -339 .(in Chinese with English abstract)
[本文引用: 2]
[李军 , 王学春 , 邵明安 , 赵玉娟 , 李小芳 (2010 ). 黄土高原半干旱和半湿润地区刺槐林地生物量与土壤干燥化效应的模拟
. 植物生态学报 , 34 , 330 -339 .]
[本文引用: 2]
[24]
Li Y Zhao CZ Dong XG Hou ZJ Ma XL Zhang Q 2013 ). Responses of Stellera chamaejasme twig and leaf traits to slope aspect in alpine grassland of Northwest China.
Chinese Journal of Ecology , 32 , 3145 -3151 .(in Chinese with English abstract)
[本文引用: 2]
[李钰 , 赵成章 , 董小刚 , 侯兆疆 , 马小丽 , 张茜 (2013 ). 高寒草地狼毒枝-叶性状对坡向的响应
. 生态学杂志 , 32 , 3145 -3151 .]
[本文引用: 2]
[25]
Li YN Yang DM Sun SC Gao XM 2008 ). Effects of twig size on biomass allocation within twigs and on lamina area supporting efficiency in Rhododendron: Allometric scaling analyses.
Journal of Plant Ecology , 32 , 1175 -1183 .(in Chinese with English abstract)
[本文引用: 1]
[李亚男 , 杨冬梅 , 孙书存 , 高贤明 (2008 ). 杜鹃花属植物小枝大小对小枝生物量分配及叶面积支持效率的影响: 异速生长分析
. 植物生态学报 , 32 , 1175 -1183 .]
[本文引用: 1]
[26]
McCulloh KA Sperry JS 2005 ). Patterns in hydraulic architecture and their implications for transport efficiency.
Tree Physiology , 25 , 257 -267 .
[本文引用: 2]
[27]
McDonald PG Fonseca CR McCoverton J 2003 ). Leaf-size divergence along rainfall and soil-nutrient gradients: Is the method of size reduction common among clades?
Functional Ecology , 17 , 50 -57 .
[28]
Niklas KJ 1992 ). Plant Biomechanics: An Engineering Approach to Plant Form and Function . University of Chicago Press , Chicago .
[本文引用: 5]
[29]
Normand F Bissery C Damour G Lauri PÉ 2008 ). Hydraulic and mechanical stem properties affect leaf-stem allometry in mango cultivars.
New Phytologist , 178 , 590 -602 .
[本文引用: 1]
[30]
Olson ME Aguirre-Hernández R Rosell JA 2009 ). Universal foliage-stem scaling across environments and species in dicot trees: Plasticity, biomechanics and Corner’s rules.
Ecology Letters , 12 , 210 -219 .
[31]
Parkhurst DF Loucks OL 1972 ). Optimal leaf size in relation to environment.
The Journal of Ecology , 60 , 505 -537 .
[本文引用: 1]
[32]
Pitman EJG 1939 ). A note on normal correlation.
Biometrika , 31 , 9 -12 .
[33]
Shan CJ Liang ZS Hao WF Liu SM 2004 ). Relationship between growth of locust and soil water in the different habitats on the Loess Plateau.
Journal of Northwest Forestry University , 19 (2 ), 9 -14 .(in Chinese with English abstract)
[本文引用: 1]
[单长卷 , 梁宗锁 , 郝文芳 , 刘淑明 (2004 ). 黄土高原不同立地条件下刺槐生长与水分关系研究
. 西北林学院学报 , 19 (2 ), 9 -14 .]
[本文引用: 1]
[34]
Shen JP Zhang WH 2014 ). Characteristics of carbon storage and sequestration of Robinia pseudoacacia forest land converted by farmland in the Hilly Loess Plateau Region.
Acta Ecologica Sinica , 34 , 2746 -2754 .(in Chinese with English abstract)
[本文引用: 1]
[申家朋 , 张文辉 (2014 ). 黄土丘陵区退耕还林地刺槐人工林碳储量及分配规律
. 生态学报 , 34 , 2746 -2754 .]
[本文引用: 1]
[35]
Shi QR Xu MS Zhao YT Zhou LL Zhang QQ Ma WJ Zhao Q Yan ER 2014 ). Testing of corner’s rules across woody plants in Tiantong region, Zhejiang Province: Effects of micro-topography.
Chinese Journal of Plant Ecology , 38 , 665 -674 .(in Chinese with English abstract)
[本文引用: 3]
[史青茹 , 许洺山 , 赵延涛 , 周刘丽 , 张晴晴 , 马文济 , 赵绮 , 阎恩荣 (2014 ). 浙江天童木本植物Corner法则的检验: 微地形的影响
. 植物生态学报 , 38 , 665 -674 .]
[本文引用: 3]
[36]
Silvertown J 2004 ). Plant coexistence and the niche.
Trends in Ecology and Evolution , 19 , 605 -611 .
[本文引用: 1]
[37]
Sun SC Jin DM Shi PL 2006 ). The leaf size-twig size spectrum of temperate woody species along an altitudinal gradient: An invariant allometric scaling relationship.
Annals of Botany , 97 , 97 -107 .
[38]
Tang GA Li FY Liu XJ 2010 ). Tutorial of Digital Elevation Model . Science Press , Beijing . 149 .(in Chinese)
[本文引用: 1]
[汤国安 , 李发源 , 刘学军 (2010 ). 数字高程模型教程 . 科学出版社 , 北京 . 149 .]
[本文引用: 1]
[39]
Vandermeer JH 1972 ). Niche theory.
Annual Review of Ecology and Systematics , 3 , 107 -132 .
[40]
Wang L Feng JX Wang SX Jia CR Wan XC 2013 ). The interaction of drought and slope aspect on growth of Quercus variabilis and Platycladus orientalis.
Acta Ecologica Sinica , 33 , 2425 -2433 .(in Chinese with English abstract)
[本文引用: 1]
[王林 , 冯锦霞 , 王双霞 , 贾长荣 , 万贤崇 (2013 ). 干旱和坡向互作对栓皮栎和侧柏生长的影响
. 生态学报 , 33 , 2425 -2433 .]
[本文引用: 1]
[41]
Wang YP Gao HH Liu YS Mu P Yu XP An LZ Zhang F 2013 ). Adaptation mechanisms of alpine plants photosynthetic apparatus against adverse stress: A review.
Chinese Journal of Applied Ecology , 24 , 2049 -2055 .(in Chinese with English abstract)
[本文引用: 1]
[王玉萍 , 高会会 , 刘悦善 , 慕平 , 鱼小军 , 安黎哲 , 张峰 (2013 ). 高山植物光合机构耐受胁迫的适应机制
. 应用生态学报 , 24 , 2049 -2055 .]
[本文引用: 1]
[42]
Warton DI Weber NC 2002 ). Common slope tests for bivariate errors-in-variables models.
Biometrical Journal , 44 , 161 -174 .
[本文引用: 2]
[43]
Warton DI Wright IJ Falster DS 2006 ). Bivariate line fitting methods for allometry.
Biological Reviews , 81 , 259 -291 .
[44]
Wei LL Zhang XQ Hou ZH Xu DY Yu XB 2005 ). Effects of water stress on photosynthesis and carbon allocation in Cunninghamia lancealata seedlings.
Acta Phytoecologica Sinica , 29 , 394 -402 .(in Chinese with English abstract)
[本文引用: 1]
[韦莉莉 , 张小全 , 侯振宏 , 徐德应 , 余雪标 (2005 ). 杉木苗木光合作用及其产物分配对水分胁迫的响应
. 植物生态学报 , 29 , 394 -402 .]
[本文引用: 1]
[45]
West GB Brown JH Enquist BJ 1997 ). A general model for the origin of allometric scaling laws in biology.
Science , 276 , 122 -126 .
[本文引用: 1]
[46]
West GB Brown JH Enquist BJ 1999 ). A general model for the structure and allometry of plant vascular systems.
Nature , 400 , 664 -667 .
[本文引用: 2]
[47]
Westoby M Falster DS Moles AT Vesk PA Wright IJ 2002 ). Plant ecological strategies: Some leading dimensions of variation between species.
Annual Review of Ecology and Systematics , 33 , 125 -159 .
[48]
Westoby M Wright IJ 2003 ). The leaf size-twig size spectrum and its relationship to other important spectra of variation among species.
Oecologia , 135 , 621 -628 .
[本文引用: 1]
[49]
Xu Y Yang XD Xie YM Xu YL Chang SX Yan ER 2012 ). Twig size-number trade-off among woody plants in Tiantong region, Zhejiang Province of China.
Chinese Journal of Plant Ecology , 36 , 1268 -1276 .(in Chinese with English abstract)
[本文引用: 2]
[许月 , 杨晓东 , 谢一鸣 , 徐艺露 , Chang SX 阎恩荣 (2012 ). 浙江天童木本植物小枝的“大小-数量”权衡
. 植物生态学报 , 36 , 1268 -1276 .]
[本文引用: 2]
[50]
Yang DM Li GY Sun SC 2008 ). The generality of leaf size versus number trade-off in temperate woody species.
Annals of Botany , 102 , 623 -629 .
[51]
Yang DM Zhan F Zhang HW 2012 a). Trade-off between leaf size and number in current-year twigs of deciduous broad-leaved woody species at different altitudes on Qingliang Mountain, southeastern China.
Chinese Journal of Plant Ecology , 36 , 281 -291 .(in Chinese with English abstract)
[本文引用: 2]
[杨冬梅 , 占峰 , 张宏伟 (2012 a). 清凉峰不同海拔木本植物小枝内叶大小-数量权衡关系
, 植物生态学报 , 36 , 281 -291 .]
[本文引用: 2]
[52]
Yang DM Zhang JJ Zhou D Qian MJ Zheng Y Jin L M 2012 b). Leaf and twig functional traits of woody plants and their relationships with environmental change: A review.
Chinese Journal of Ecology , 31 , 702 -713 .(in Chinese with English abstract)
[本文引用: 2]
[杨冬梅 , 章佳佳 , 周丹 , 钱敏杰 , 郑瑶 , 金灵妙 (2012 b). 木本植物茎叶功能性状及其关系随环境变化的研究进展
. 生态学杂志 , 31 , 702 -713 .]
[本文引用: 2]
[53]
Yin J Qiu GY He F He KN Tian JH Zhang WQ Xiong YJ Zhao SH Liu JX 2008 ). Leaf area characteristics of plantation stands in semi-arid loess hill-gully region of China
. Journal of Plant Ecology (Chinese Version) , 32 , 440 -447 .(in Chinese with English abstract)
[本文引用: 1]
[尹婧 , 邱国玉 , 何凡 , 贺康宁 , 田晶会 , 张卫强 , 熊育久 , 赵少华 , 刘建新 (2008 ). 半干旱黄土丘陵区人工林叶面积特征
. 植物生态学报 , 32 , 440 -447 .]
[本文引用: 1]
[54]
Yin XQ 2004 ). Biogeography . Higher Education Press , Beijing . 26 -28 .(in Chinese)
[本文引用: 1]
[殷秀琴 (2004 ). 生物地理学 . 高等教育出版社 , 北京 . 26 -28 .]
[本文引用: 1]
[55]
Yu ZH Chen YM Du S 2009 ). Sap flow dynamics in the leaf-flushing period of a Robinia pseudoacacia plantation in semi-arid region of loess plateau.
Scientia Silvae Sinicae , 45 (4 ), 53 -59 .(in Chinese with English abstract)
[本文引用: 1]
[于占辉 , 陈云明 , 杜盛 (2009 ). 黄土高原半干旱区人工林刺槐展叶期树干液流动态分析
. 林业科学 , 45 (4 ), 53 -59 .]
[本文引用: 1]
[56]
Zhang L Luo TX 2004 ). Advances in ecological studies on leaf lifespan and associated leaf traits.
Acta Phytoecologica Sinica , 28 , 844 -852 .(in Chinese with English abstract)
[本文引用: 1]
[张林 , 罗天祥 (2004 ). 植物叶寿命及其相关叶性状的生态学研究进展
. 植物生态学报 , 28 , 844 -852 .]
[本文引用: 1]
Hydraulic architecture and the evolution of shoot allometry in contrasting climates.
1
2003
... 坡向虽然对刺槐枝叶大小回归方程的斜率没有显著影响, 但使回归方程的截距产生了显著改变.在茎截面积-总叶面积关系中, 其回归方程的截距在北坡显著大于南坡(p < 0.05), 表明在茎截面积一定时, 位于北坡的刺槐在单位大小枝条上支持的总叶面积大于位于南坡的刺槐.叶片大小是植物适应环境中热量和水分交换平衡的结果, 其变化主要是通过影响叶边缘对热量、CO2 和水分的传导能力来影响整个植株的生长速率(杨冬梅等, 2012b ).南坡光照时间长, 水分蒸发大(殷秀琴, 2004 ), 土壤水分含量低(表1 ), 在西北年降水量小的干旱半干旱地区, 由于高温常引起蒸腾作用加强而使植物体水分大量丧失, 因而这种高温和干旱环境严重影响了植物的新陈代谢活动, 细胞的分裂和生长受到限制, 植物生长缓慢, 叶片较小; 然而北坡气温降低, 地面蒸发量减少, 土壤水分增加(表1 ), 良好的水热条件有利于细胞体积和数目增加, 从而促进了叶面积的扩展(李钰等, 2013 ).反过来, 与小叶相比, 大叶具有更厚的叶边缘, 热量和物质通过叶边缘进行转运的阻力更大, 往往会阻止叶片与周围的空气进行热量交换, 减慢CO2 和水蒸气进出叶片的扩散速率, 使大叶比小叶更容易过热, 植物的呼吸和蒸腾成本更高(Givnish, 1978 ), 因此在冷凉、湿润或者荫蔽的环境下大叶由于能够使光合收益最大化而占有优势(Givnish, 1979 ), 而相对小的叶片因为具有较高的热交换能力, 被认为在热、干、高光和低养分的环境下较有优势(Ackerly & Preston, 2003 ; McDonald et al ., 2003 ).另外, 大叶片植物在水分较少的生境中不具有选择优势, 与植物的水力结构特征有关.随着土壤湿度的下降, 水分供应不足会增加水分从土壤运送到叶片的连续水柱断裂的风险, 当大叶片植物的蒸腾速率增大时, 空穴现象和栓塞现象更易发生(McCulloh & Sperry, 2005 ), 从而使得大叶片植物在水分稀缺的生境下不具有水分运输方面的选择优势, 这与Callaway等(1994) 的研究结果一致, 即: 一定大小枝条输导面积支持的总叶面积随着大气蒸腾强度和干旱强度的增加而减小. ...
黄土丘陵区不同林龄刺槐人工林碳、氮储量及分配格局
1
2014
... 刺槐(Robinia pseudoacacia )是豆科刺槐属的温带落叶乔木, 具有生长速度快、根系发达、对环境适应性强、固碳氮能力较好、成活率高、萌芽力和根蘖性都很强等优点, 已成为我国黄土丘陵区的主要造林树种和人工林面积最大的树种(于占辉等, 2009 ; 李军等, 2010 ).近年来, 干旱地区许多刺槐人工林出现了生长减慢、枝梢干枯甚至整株死亡的衰败现象, 引起了学术界的广泛关注(单长卷等, 2004 ; 李军等, 2010 ).目前, 许多学者研究了刺槐叶面积生长与林分密度的关系(尹婧等, 2008 ), 土层厚度对刺槐旱季水分状况和生长的影响(王林等, 2013 ), 不同林龄和退耕年限刺槐人工林碳、氮储量及分配规律(申家朋和张文辉, 2014 ; 艾泽民等, 2014 ), 揭示了刺槐生长以及生物量积累和分配格局等一系列规律, 但关于微地形影响下刺槐当年生小枝内枝叶关系的研究较为少见, 更鲜有不同坡向茎截面积与总叶面积、出叶强度与单叶面积关系的报道, 限制了对异质生境中刺槐适应机制的解析.本研究选择兰州北山九州台西北师范大学绿化基地的刺槐人工林为研究对象, 探讨刺槐当年生小枝内的茎截面积-总叶面积、出叶强度-单叶面积关系, 为进一步了解刺槐的环境适应机制提供理论依据, 对提高干旱半干旱地区人工林的可持续性和稳定性具有重要意义. ...
黄土丘陵区不同林龄刺槐人工林碳、氮储量及分配格局
1
2014
... 刺槐(Robinia pseudoacacia )是豆科刺槐属的温带落叶乔木, 具有生长速度快、根系发达、对环境适应性强、固碳氮能力较好、成活率高、萌芽力和根蘖性都很强等优点, 已成为我国黄土丘陵区的主要造林树种和人工林面积最大的树种(于占辉等, 2009 ; 李军等, 2010 ).近年来, 干旱地区许多刺槐人工林出现了生长减慢、枝梢干枯甚至整株死亡的衰败现象, 引起了学术界的广泛关注(单长卷等, 2004 ; 李军等, 2010 ).目前, 许多学者研究了刺槐叶面积生长与林分密度的关系(尹婧等, 2008 ), 土层厚度对刺槐旱季水分状况和生长的影响(王林等, 2013 ), 不同林龄和退耕年限刺槐人工林碳、氮储量及分配规律(申家朋和张文辉, 2014 ; 艾泽民等, 2014 ), 揭示了刺槐生长以及生物量积累和分配格局等一系列规律, 但关于微地形影响下刺槐当年生小枝内枝叶关系的研究较为少见, 更鲜有不同坡向茎截面积与总叶面积、出叶强度与单叶面积关系的报道, 限制了对异质生境中刺槐适应机制的解析.本研究选择兰州北山九州台西北师范大学绿化基地的刺槐人工林为研究对象, 探讨刺槐当年生小枝内的茎截面积-总叶面积、出叶强度-单叶面积关系, 为进一步了解刺槐的环境适应机制提供理论依据, 对提高干旱半干旱地区人工林的可持续性和稳定性具有重要意义. ...
1
1996
... 植物功能性状与环境和生态系统功能的关系是近年来生态学和全球变化研究的热点(Harrison et al ., 2010 ; Bernard-Verdier et al ., 2012 ), 各功能性状之间的权衡和组合决定了植物的生活史对策(Vandermeer, 1972 ; Silvertown, 2004 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ), 叶片是植物进行光合作用的主要器官(张林和罗天祥, 2004 ), 两者大小和数量的配置关系直接影响着植物的机械力学和水力结构特征, 从而决定了植物的生活史策略, 并可能影响物种生态位的利用(Givnish, 1987 ; Enquist et al ., 1999 ; West et al ., 1999 ; Westoby et al ., 2002 ).植物单位大小枝条上支撑的叶面积必须与支撑枝的横截面积成一定的比例, 才能符合植物器官间的机械力学和水力结构需求(Niklas, 1992 ; Enquist et al ., 1999 ; Normand et al ., 2008 ; Olson et al ., 2009 ).叶片大小和叶片数量是决定树冠紧密程度的两个重要因素, 直接影响到植株的冠层结构和发育模式, 进而影响到植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ).在某一给定的叶生物量分配或某一个给定的总叶面积下, 植物可能有较少的大叶子或者较多的小叶子(杨冬梅等, 2012a ).坡向作为一种重要的地形要素, 往往对局部区域的太阳辐射和降水进行二次分配(Cantón et al ., 2004 ), 在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 能够对植物的形态产生影响(Bazzaz, 1996 ; 韦莉莉等, 2005 ).当自然资源的丰富度低于或超过植物光合系统所能利用的最低或最高限度时, 植物的光合作用将被抑制(王玉萍等, 2013 ).为提高植物构型构建的生境适应性和风险规避能力, 植物通过功能性状等表型的可塑性调整提高自身的生存适合度, 体现了全球气候变化背景下植物的资源配置策略.因此, 研究坡向梯度上植物枝叶关系规律及其成因, 对于理解植物的环境适应机制有着重要意义. ...
1
1993
... 除此之外, 叶大小-叶数量权衡关系的坡向差异性还可以从以下两个方面解释: (1)植物内在生物量分配限制的结果.由表2 可知, 北坡(阴坡)刺槐单叶面积大于南坡(阳坡), 在植物体上, 在某一给定的叶生物量或某一给定的总叶面积下, 植物要么是有较少的大叶子, 要么是有较多的小叶子, 增加单个叶片的投资意味着叶片数量的减少(杨冬梅等, 2012a ); (2)单叶面积的差异是出叶强度变化的副产物.每个叶片通常都会有一个腋芽分生组织(Bell, 1993 ), 叶片更多的物种, 单位个体大小的腋芽分生组织的密度将会更高(杨冬梅等, 2012b ), 更大的芽库由于有更多的部位具有再生能力, 既可以生长为植株的营养结构或繁殖结构, 也可以保持无活性状态, 进而影响到植株的大小构建和繁殖产出(Bonser & Aarssen, 1996 ), 使植株具有更大的可塑性和更新复壮能力, 因此在南坡干、热、高光和低养分的环境下, 刺槐倾向于具有更高的出叶强度, 叶大小与出叶强度紧密相关, 因此小叶片可能是选择了高的出叶强度的结果.东坡和西坡枝叶功能性状及其关系之间没有明显差异, 主要是因为东坡和西坡属于半阴坡和半阳坡, 皆处于阴坡和阳坡之间的过渡区域, 群落组成也较为复杂, 因此刺槐采取相同的生长策略来适应生境的变化. ...
Community assembly along a soil depth gradient: Contrasting patterns of plant trait convergence and divergence in a Mediterranean rangeland.
1
2012
... 植物功能性状与环境和生态系统功能的关系是近年来生态学和全球变化研究的热点(Harrison et al ., 2010 ; Bernard-Verdier et al ., 2012 ), 各功能性状之间的权衡和组合决定了植物的生活史对策(Vandermeer, 1972 ; Silvertown, 2004 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ), 叶片是植物进行光合作用的主要器官(张林和罗天祥, 2004 ), 两者大小和数量的配置关系直接影响着植物的机械力学和水力结构特征, 从而决定了植物的生活史策略, 并可能影响物种生态位的利用(Givnish, 1987 ; Enquist et al ., 1999 ; West et al ., 1999 ; Westoby et al ., 2002 ).植物单位大小枝条上支撑的叶面积必须与支撑枝的横截面积成一定的比例, 才能符合植物器官间的机械力学和水力结构需求(Niklas, 1992 ; Enquist et al ., 1999 ; Normand et al ., 2008 ; Olson et al ., 2009 ).叶片大小和叶片数量是决定树冠紧密程度的两个重要因素, 直接影响到植株的冠层结构和发育模式, 进而影响到植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ).在某一给定的叶生物量分配或某一个给定的总叶面积下, 植物可能有较少的大叶子或者较多的小叶子(杨冬梅等, 2012a ).坡向作为一种重要的地形要素, 往往对局部区域的太阳辐射和降水进行二次分配(Cantón et al ., 2004 ), 在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 能够对植物的形态产生影响(Bazzaz, 1996 ; 韦莉莉等, 2005 ).当自然资源的丰富度低于或超过植物光合系统所能利用的最低或最高限度时, 植物的光合作用将被抑制(王玉萍等, 2013 ).为提高植物构型构建的生境适应性和风险规避能力, 植物通过功能性状等表型的可塑性调整提高自身的生存适合度, 体现了全球气候变化背景下植物的资源配置策略.因此, 研究坡向梯度上植物枝叶关系规律及其成因, 对于理解植物的环境适应机制有着重要意义. ...
Meristem allocation: A new classification theory for adaptive strategies in herbaceous plants.
1
1996
... 除此之外, 叶大小-叶数量权衡关系的坡向差异性还可以从以下两个方面解释: (1)植物内在生物量分配限制的结果.由表2 可知, 北坡(阴坡)刺槐单叶面积大于南坡(阳坡), 在植物体上, 在某一给定的叶生物量或某一给定的总叶面积下, 植物要么是有较少的大叶子, 要么是有较多的小叶子, 增加单个叶片的投资意味着叶片数量的减少(杨冬梅等, 2012a ); (2)单叶面积的差异是出叶强度变化的副产物.每个叶片通常都会有一个腋芽分生组织(Bell, 1993 ), 叶片更多的物种, 单位个体大小的腋芽分生组织的密度将会更高(杨冬梅等, 2012b ), 更大的芽库由于有更多的部位具有再生能力, 既可以生长为植株的营养结构或繁殖结构, 也可以保持无活性状态, 进而影响到植株的大小构建和繁殖产出(Bonser & Aarssen, 1996 ), 使植株具有更大的可塑性和更新复壮能力, 因此在南坡干、热、高光和低养分的环境下, 刺槐倾向于具有更高的出叶强度, 叶大小与出叶强度紧密相关, 因此小叶片可能是选择了高的出叶强度的结果.东坡和西坡枝叶功能性状及其关系之间没有明显差异, 主要是因为东坡和西坡属于半阴坡和半阳坡, 皆处于阴坡和阳坡之间的过渡区域, 群落组成也较为复杂, 因此刺槐采取相同的生长策略来适应生境的变化. ...
Corner’s rules revisited: Ontogenetic and interspecific patterns in leaf-stem allometry.
1
1998
... 植物功能性状与生存环境之间是一种相互协同进化与适应的关系(侯兆疆等, 2014 ).枝叶大小关系是植物的重要生活史策略(Westoby et al ., 2002 ), 其比例不仅反映枝叶生物量的分配和叶片在生化和水力生物学方面的功能需求(Brouat et al ., 1998 ; Sun et al ., 2006 ; Normand et al ., 2008 ), 也体现了植物适应特定环境的构型塑造(Enquist & Niklas, 2001 , 2002 ).各坡向上刺槐茎截面积和总叶面积均呈正的异速生长关系, 回归方程间具有共同的SMA斜率且显著大于1 (p < 0.05)(图1 ), 二者的相关关系始终符合Corner法则(Corner, 1949 ), 不随微地形特征的变化而显著改变, 植物对总叶面积的投入显著高于对枝横截面积的投入, 这说明微地形差异无法改变植物在枝叶大小上的协调, 枝大小和叶大小间始终需要维持几何大小和重力的平衡(史青茹等, 2014 ).对于此现象, Westoby等(2002) 曾从枝叶的形态和生理关联性方面进行了解释, 他们认为: 一定大小的叶面积必须要由一定大小的枝来支撑, 两者间必须保持大小的匹配, 满足机械力学和水分动力学的要求, 才能保持机械支撑的平衡以及水分运输和蒸腾的协调; 管道模型理论(Niklas, 1992 )则提出, 为满足叶片对水分的需求, 木质部的横截面积与它们所支持的叶面积成正比.除此之外, West等(1997) 提出了结构分形自相似假说解释这种现象, 即: 所有生物的各器官间具有自相似的分形网络结构, 自然选择的压力必然使其向最优化的方向进化, 使得与外界环境的交换面积和新陈代谢效率最大化, 并使营养物质运输距离和时间最小化, 因此, 枝叶的大小必然按一定的比例来协调.而茎截面积-总叶面积关系的SMA斜率显著大于1 (p < 0.05), 主要是因为叶片是植株碳获取的主要功能构件, 所以在进化过程中为提高光合收益, 植株将尽可能地使分配到叶片中的生物量最大化, 而在叶片支撑结构中的投资最小化. ...
Biomass allocation of montane and desert ponderosa pine: An analog for response to climate change.
0
1994
Topographic controls on the spatial distribution of ground cover in the Tabernas badlands of SE Spain.
1
2004
... 植物功能性状与环境和生态系统功能的关系是近年来生态学和全球变化研究的热点(Harrison et al ., 2010 ; Bernard-Verdier et al ., 2012 ), 各功能性状之间的权衡和组合决定了植物的生活史对策(Vandermeer, 1972 ; Silvertown, 2004 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ), 叶片是植物进行光合作用的主要器官(张林和罗天祥, 2004 ), 两者大小和数量的配置关系直接影响着植物的机械力学和水力结构特征, 从而决定了植物的生活史策略, 并可能影响物种生态位的利用(Givnish, 1987 ; Enquist et al ., 1999 ; West et al ., 1999 ; Westoby et al ., 2002 ).植物单位大小枝条上支撑的叶面积必须与支撑枝的横截面积成一定的比例, 才能符合植物器官间的机械力学和水力结构需求(Niklas, 1992 ; Enquist et al ., 1999 ; Normand et al ., 2008 ; Olson et al ., 2009 ).叶片大小和叶片数量是决定树冠紧密程度的两个重要因素, 直接影响到植株的冠层结构和发育模式, 进而影响到植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ).在某一给定的叶生物量分配或某一个给定的总叶面积下, 植物可能有较少的大叶子或者较多的小叶子(杨冬梅等, 2012a ).坡向作为一种重要的地形要素, 往往对局部区域的太阳辐射和降水进行二次分配(Cantón et al ., 2004 ), 在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 能够对植物的形态产生影响(Bazzaz, 1996 ; 韦莉莉等, 2005 ).当自然资源的丰富度低于或超过植物光合系统所能利用的最低或最高限度时, 植物的光合作用将被抑制(王玉萍等, 2013 ).为提高植物构型构建的生境适应性和风险规避能力, 植物通过功能性状等表型的可塑性调整提高自身的生存适合度, 体现了全球气候变化背景下植物的资源配置策略.因此, 研究坡向梯度上植物枝叶关系规律及其成因, 对于理解植物的环境适应机制有着重要意义. ...
四川龙门山西北部植被分布与地形因子的相关性
1
2006
... 利用ArcGIS对研究区1:5万地形图进行数字化, 得到研究区域10 m分辨率的数字高程模型(digital elevation mode, DEM).在DEM的基础上利用Spatial Analyst模块中的Surface Analysis命令提取坡向, 它表示地表面上一点的切平面的法线矢量n 在水平面的投影nxoy 与过该点的正北方向的夹角, 其中坡向组别以国际划分法确定(陈瑶等, 2006 ), 具体方法参见汤国安等(2010) 文献. ...
四川龙门山西北部植被分布与地形因子的相关性
1
2006
... 利用ArcGIS对研究区1:5万地形图进行数字化, 得到研究区域10 m分辨率的数字高程模型(digital elevation mode, DEM).在DEM的基础上利用Spatial Analyst模块中的Surface Analysis命令提取坡向, 它表示地表面上一点的切平面的法线矢量n 在水平面的投影nxoy 与过该点的正北方向的夹角, 其中坡向组别以国际划分法确定(陈瑶等, 2006 ), 具体方法参见汤国安等(2010) 文献. ...
The durian theory or the origin of the modern tree.
1
1949
... 植物功能性状与生存环境之间是一种相互协同进化与适应的关系(侯兆疆等, 2014 ).枝叶大小关系是植物的重要生活史策略(Westoby et al ., 2002 ), 其比例不仅反映枝叶生物量的分配和叶片在生化和水力生物学方面的功能需求(Brouat et al ., 1998 ; Sun et al ., 2006 ; Normand et al ., 2008 ), 也体现了植物适应特定环境的构型塑造(Enquist & Niklas, 2001 , 2002 ).各坡向上刺槐茎截面积和总叶面积均呈正的异速生长关系, 回归方程间具有共同的SMA斜率且显著大于1 (p < 0.05)(图1 ), 二者的相关关系始终符合Corner法则(Corner, 1949 ), 不随微地形特征的变化而显著改变, 植物对总叶面积的投入显著高于对枝横截面积的投入, 这说明微地形差异无法改变植物在枝叶大小上的协调, 枝大小和叶大小间始终需要维持几何大小和重力的平衡(史青茹等, 2014 ).对于此现象, Westoby等(2002) 曾从枝叶的形态和生理关联性方面进行了解释, 他们认为: 一定大小的叶面积必须要由一定大小的枝来支撑, 两者间必须保持大小的匹配, 满足机械力学和水分动力学的要求, 才能保持机械支撑的平衡以及水分运输和蒸腾的协调; 管道模型理论(Niklas, 1992 )则提出, 为满足叶片对水分的需求, 木质部的横截面积与它们所支持的叶面积成正比.除此之外, West等(1997) 提出了结构分形自相似假说解释这种现象, 即: 所有生物的各器官间具有自相似的分形网络结构, 自然选择的压力必然使其向最优化的方向进化, 使得与外界环境的交换面积和新陈代谢效率最大化, 并使营养物质运输距离和时间最小化, 因此, 枝叶的大小必然按一定的比例来协调.而茎截面积-总叶面积关系的SMA斜率显著大于1 (p < 0.05), 主要是因为叶片是植株碳获取的主要功能构件, 所以在进化过程中为提高光合收益, 植株将尽可能地使分配到叶片中的生物量最大化, 而在叶片支撑结构中的投资最小化. ...
Invariant scaling relations across tree-dominated communities.
1
2001
... 植物功能性状与生存环境之间是一种相互协同进化与适应的关系(侯兆疆等, 2014 ).枝叶大小关系是植物的重要生活史策略(Westoby et al ., 2002 ), 其比例不仅反映枝叶生物量的分配和叶片在生化和水力生物学方面的功能需求(Brouat et al ., 1998 ; Sun et al ., 2006 ; Normand et al ., 2008 ), 也体现了植物适应特定环境的构型塑造(Enquist & Niklas, 2001 , 2002 ).各坡向上刺槐茎截面积和总叶面积均呈正的异速生长关系, 回归方程间具有共同的SMA斜率且显著大于1 (p < 0.05)(图1 ), 二者的相关关系始终符合Corner法则(Corner, 1949 ), 不随微地形特征的变化而显著改变, 植物对总叶面积的投入显著高于对枝横截面积的投入, 这说明微地形差异无法改变植物在枝叶大小上的协调, 枝大小和叶大小间始终需要维持几何大小和重力的平衡(史青茹等, 2014 ).对于此现象, Westoby等(2002) 曾从枝叶的形态和生理关联性方面进行了解释, 他们认为: 一定大小的叶面积必须要由一定大小的枝来支撑, 两者间必须保持大小的匹配, 满足机械力学和水分动力学的要求, 才能保持机械支撑的平衡以及水分运输和蒸腾的协调; 管道模型理论(Niklas, 1992 )则提出, 为满足叶片对水分的需求, 木质部的横截面积与它们所支持的叶面积成正比.除此之外, West等(1997) 提出了结构分形自相似假说解释这种现象, 即: 所有生物的各器官间具有自相似的分形网络结构, 自然选择的压力必然使其向最优化的方向进化, 使得与外界环境的交换面积和新陈代谢效率最大化, 并使营养物质运输距离和时间最小化, 因此, 枝叶的大小必然按一定的比例来协调.而茎截面积-总叶面积关系的SMA斜率显著大于1 (p < 0.05), 主要是因为叶片是植株碳获取的主要功能构件, 所以在进化过程中为提高光合收益, 植株将尽可能地使分配到叶片中的生物量最大化, 而在叶片支撑结构中的投资最小化. ...
Global allocation rules for patterns of biomass partitioning in seed plants.
1
2002
... 植物功能性状与生存环境之间是一种相互协同进化与适应的关系(侯兆疆等, 2014 ).枝叶大小关系是植物的重要生活史策略(Westoby et al ., 2002 ), 其比例不仅反映枝叶生物量的分配和叶片在生化和水力生物学方面的功能需求(Brouat et al ., 1998 ; Sun et al ., 2006 ; Normand et al ., 2008 ), 也体现了植物适应特定环境的构型塑造(Enquist & Niklas, 2001 , 2002 ).各坡向上刺槐茎截面积和总叶面积均呈正的异速生长关系, 回归方程间具有共同的SMA斜率且显著大于1 (p < 0.05)(图1 ), 二者的相关关系始终符合Corner法则(Corner, 1949 ), 不随微地形特征的变化而显著改变, 植物对总叶面积的投入显著高于对枝横截面积的投入, 这说明微地形差异无法改变植物在枝叶大小上的协调, 枝大小和叶大小间始终需要维持几何大小和重力的平衡(史青茹等, 2014 ).对于此现象, Westoby等(2002) 曾从枝叶的形态和生理关联性方面进行了解释, 他们认为: 一定大小的叶面积必须要由一定大小的枝来支撑, 两者间必须保持大小的匹配, 满足机械力学和水分动力学的要求, 才能保持机械支撑的平衡以及水分运输和蒸腾的协调; 管道模型理论(Niklas, 1992 )则提出, 为满足叶片对水分的需求, 木质部的横截面积与它们所支持的叶面积成正比.除此之外, West等(1997) 提出了结构分形自相似假说解释这种现象, 即: 所有生物的各器官间具有自相似的分形网络结构, 自然选择的压力必然使其向最优化的方向进化, 使得与外界环境的交换面积和新陈代谢效率最大化, 并使营养物质运输距离和时间最小化, 因此, 枝叶的大小必然按一定的比例来协调.而茎截面积-总叶面积关系的SMA斜率显著大于1 (p < 0.05), 主要是因为叶片是植株碳获取的主要功能构件, 所以在进化过程中为提高光合收益, 植株将尽可能地使分配到叶片中的生物量最大化, 而在叶片支撑结构中的投资最小化. ...
Allometric scaling of production and life-history variation in vascular plants.
2
1999
... 植物功能性状与环境和生态系统功能的关系是近年来生态学和全球变化研究的热点(Harrison et al ., 2010 ; Bernard-Verdier et al ., 2012 ), 各功能性状之间的权衡和组合决定了植物的生活史对策(Vandermeer, 1972 ; Silvertown, 2004 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ), 叶片是植物进行光合作用的主要器官(张林和罗天祥, 2004 ), 两者大小和数量的配置关系直接影响着植物的机械力学和水力结构特征, 从而决定了植物的生活史策略, 并可能影响物种生态位的利用(Givnish, 1987 ; Enquist et al ., 1999 ; West et al ., 1999 ; Westoby et al ., 2002 ).植物单位大小枝条上支撑的叶面积必须与支撑枝的横截面积成一定的比例, 才能符合植物器官间的机械力学和水力结构需求(Niklas, 1992 ; Enquist et al ., 1999 ; Normand et al ., 2008 ; Olson et al ., 2009 ).叶片大小和叶片数量是决定树冠紧密程度的两个重要因素, 直接影响到植株的冠层结构和发育模式, 进而影响到植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ).在某一给定的叶生物量分配或某一个给定的总叶面积下, 植物可能有较少的大叶子或者较多的小叶子(杨冬梅等, 2012a ).坡向作为一种重要的地形要素, 往往对局部区域的太阳辐射和降水进行二次分配(Cantón et al ., 2004 ), 在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 能够对植物的形态产生影响(Bazzaz, 1996 ; 韦莉莉等, 2005 ).当自然资源的丰富度低于或超过植物光合系统所能利用的最低或最高限度时, 植物的光合作用将被抑制(王玉萍等, 2013 ).为提高植物构型构建的生境适应性和风险规避能力, 植物通过功能性状等表型的可塑性调整提高自身的生存适合度, 体现了全球气候变化背景下植物的资源配置策略.因此, 研究坡向梯度上植物枝叶关系规律及其成因, 对于理解植物的环境适应机制有着重要意义. ...
... ; Enquist et al ., 1999 ; Normand et al ., 2008 ; Olson et al ., 2009 ).叶片大小和叶片数量是决定树冠紧密程度的两个重要因素, 直接影响到植株的冠层结构和发育模式, 进而影响到植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ).在某一给定的叶生物量分配或某一个给定的总叶面积下, 植物可能有较少的大叶子或者较多的小叶子(杨冬梅等, 2012a ).坡向作为一种重要的地形要素, 往往对局部区域的太阳辐射和降水进行二次分配(Cantón et al ., 2004 ), 在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 能够对植物的形态产生影响(Bazzaz, 1996 ; 韦莉莉等, 2005 ).当自然资源的丰富度低于或超过植物光合系统所能利用的最低或最高限度时, 植物的光合作用将被抑制(王玉萍等, 2013 ).为提高植物构型构建的生境适应性和风险规避能力, 植物通过功能性状等表型的可塑性调整提高自身的生存适合度, 体现了全球气候变化背景下植物的资源配置策略.因此, 研究坡向梯度上植物枝叶关系规律及其成因, 对于理解植物的环境适应机制有着重要意义. ...
User’s Guide to SMATR: Standardised Major Axis Tests & Routines Version 2.0
2
2006
... 将坡向划分为北坡(315°-360°和0°-45°)、东坡(45°-135°)、南坡(135°-225°)和西坡(225°-315°).在所研究的4个功能性状中, 茎截面积指当年生小枝上1级枝的横截面积, 总叶面积指当年生小枝上所支撑的所有叶片面积之和, 出叶强度指单位体积枝条所支撑的叶片数, 单叶面积指当年生小枝上所支撑的所有叶片单叶面积的平均值, 枝条体积由其横截面积乘以长度获得(许月等, 2012 ).对每个样方刺槐枝叶属性的平均值进行对数(以10为底)转换后再进行分析, 对数转换使之符合正态分布.对枝大小-叶大小以及叶数量-叶大小两组功能性状关系的研究, 均采用y = axb , 线性转换成log(y ) = log(a ) + b log(x ), 式中x 和y 表示两个特征参数, a 为性状关系的截距, b 为斜率, 即异速生长参数或相对生长的指数, 当|b | = 1时, 表示两者是等速变化关系; 当|b| 显著偏离1时, 两者间为异速变化关系(Harvey & Pagel, 1991 ).数据分析主要采用标准化主轴估计(standardized major axis estimation, SMA)的方法(Warton et al ., 2006 ), 由软件(S)MATR Version 2.0 (Falster et al ., 2006 )计算完成.每一个回归斜率的置信区间根据Pitman (1939)方法计算.回归斜率与1或-1的显著性检验根据Warton和Weber (2002)的方法判断.在完成线性回归后, 对回归斜率进行异质性测试, 如果组间没有异质性, 则表示有共同斜率(Warton & Weber, 2002 ).此后, 不同坡向梯度间的功能性状的平均值的比较采用单因素方差分析(one-way ANOVA), 利用成对比较检验分析各坡向刺槐枝叶关系线性回归方程在y 轴上截距的差异. ...
... 坡向虽然对刺槐枝叶大小回归方程的斜率没有显著影响, 但使回归方程的截距产生了显著改变.在茎截面积-总叶面积关系中, 其回归方程的截距在北坡显著大于南坡(p < 0.05), 表明在茎截面积一定时, 位于北坡的刺槐在单位大小枝条上支持的总叶面积大于位于南坡的刺槐.叶片大小是植物适应环境中热量和水分交换平衡的结果, 其变化主要是通过影响叶边缘对热量、CO2 和水分的传导能力来影响整个植株的生长速率(杨冬梅等, 2012b ).南坡光照时间长, 水分蒸发大(殷秀琴, 2004 ), 土壤水分含量低(表1 ), 在西北年降水量小的干旱半干旱地区, 由于高温常引起蒸腾作用加强而使植物体水分大量丧失, 因而这种高温和干旱环境严重影响了植物的新陈代谢活动, 细胞的分裂和生长受到限制, 植物生长缓慢, 叶片较小; 然而北坡气温降低, 地面蒸发量减少, 土壤水分增加(表1 ), 良好的水热条件有利于细胞体积和数目增加, 从而促进了叶面积的扩展(李钰等, 2013 ).反过来, 与小叶相比, 大叶具有更厚的叶边缘, 热量和物质通过叶边缘进行转运的阻力更大, 往往会阻止叶片与周围的空气进行热量交换, 减慢CO2 和水蒸气进出叶片的扩散速率, 使大叶比小叶更容易过热, 植物的呼吸和蒸腾成本更高(Givnish, 1978 ), 因此在冷凉、湿润或者荫蔽的环境下大叶由于能够使光合收益最大化而占有优势(Givnish, 1979 ), 而相对小的叶片因为具有较高的热交换能力, 被认为在热、干、高光和低养分的环境下较有优势(Ackerly & Preston, 2003 ; McDonald et al ., 2003 ).另外, 大叶片植物在水分较少的生境中不具有选择优势, 与植物的水力结构特征有关.随着土壤湿度的下降, 水分供应不足会增加水分从土壤运送到叶片的连续水柱断裂的风险, 当大叶片植物的蒸腾速率增大时, 空穴现象和栓塞现象更易发生(McCulloh & Sperry, 2005 ), 从而使得大叶片植物在水分稀缺的生境下不具有水分运输方面的选择优势, 这与Callaway等(1994) 的研究结果一致, 即: 一定大小枝条输导面积支持的总叶面积随着大气蒸腾强度和干旱强度的增加而减小. ...
Ecological aspects of plant morphology: Leaf form in relation to environment.
0
1978
2
1979
... 植物功能性状与环境和生态系统功能的关系是近年来生态学和全球变化研究的热点(Harrison et al ., 2010 ; Bernard-Verdier et al ., 2012 ), 各功能性状之间的权衡和组合决定了植物的生活史对策(Vandermeer, 1972 ; Silvertown, 2004 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ), 叶片是植物进行光合作用的主要器官(张林和罗天祥, 2004 ), 两者大小和数量的配置关系直接影响着植物的机械力学和水力结构特征, 从而决定了植物的生活史策略, 并可能影响物种生态位的利用(Givnish, 1987 ; Enquist et al ., 1999 ; West et al ., 1999 ; Westoby et al ., 2002 ).植物单位大小枝条上支撑的叶面积必须与支撑枝的横截面积成一定的比例, 才能符合植物器官间的机械力学和水力结构需求(Niklas, 1992 ; Enquist et al ., 1999 ; Normand et al ., 2008 ; Olson et al ., 2009 ).叶片大小和叶片数量是决定树冠紧密程度的两个重要因素, 直接影响到植株的冠层结构和发育模式, 进而影响到植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ).在某一给定的叶生物量分配或某一个给定的总叶面积下, 植物可能有较少的大叶子或者较多的小叶子(杨冬梅等, 2012a ).坡向作为一种重要的地形要素, 往往对局部区域的太阳辐射和降水进行二次分配(Cantón et al ., 2004 ), 在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 能够对植物的形态产生影响(Bazzaz, 1996 ; 韦莉莉等, 2005 ).当自然资源的丰富度低于或超过植物光合系统所能利用的最低或最高限度时, 植物的光合作用将被抑制(王玉萍等, 2013 ).为提高植物构型构建的生境适应性和风险规避能力, 植物通过功能性状等表型的可塑性调整提高自身的生存适合度, 体现了全球气候变化背景下植物的资源配置策略.因此, 研究坡向梯度上植物枝叶关系规律及其成因, 对于理解植物的环境适应机制有着重要意义. ...
... 坡向虽然对刺槐枝叶大小回归方程的斜率没有显著影响, 但使回归方程的截距产生了显著改变.在茎截面积-总叶面积关系中, 其回归方程的截距在北坡显著大于南坡(p < 0.05), 表明在茎截面积一定时, 位于北坡的刺槐在单位大小枝条上支持的总叶面积大于位于南坡的刺槐.叶片大小是植物适应环境中热量和水分交换平衡的结果, 其变化主要是通过影响叶边缘对热量、CO2 和水分的传导能力来影响整个植株的生长速率(杨冬梅等, 2012b ).南坡光照时间长, 水分蒸发大(殷秀琴, 2004 ), 土壤水分含量低(表1 ), 在西北年降水量小的干旱半干旱地区, 由于高温常引起蒸腾作用加强而使植物体水分大量丧失, 因而这种高温和干旱环境严重影响了植物的新陈代谢活动, 细胞的分裂和生长受到限制, 植物生长缓慢, 叶片较小; 然而北坡气温降低, 地面蒸发量减少, 土壤水分增加(表1 ), 良好的水热条件有利于细胞体积和数目增加, 从而促进了叶面积的扩展(李钰等, 2013 ).反过来, 与小叶相比, 大叶具有更厚的叶边缘, 热量和物质通过叶边缘进行转运的阻力更大, 往往会阻止叶片与周围的空气进行热量交换, 减慢CO2 和水蒸气进出叶片的扩散速率, 使大叶比小叶更容易过热, 植物的呼吸和蒸腾成本更高(Givnish, 1978 ), 因此在冷凉、湿润或者荫蔽的环境下大叶由于能够使光合收益最大化而占有优势(Givnish, 1979 ), 而相对小的叶片因为具有较高的热交换能力, 被认为在热、干、高光和低养分的环境下较有优势(Ackerly & Preston, 2003 ; McDonald et al ., 2003 ).另外, 大叶片植物在水分较少的生境中不具有选择优势, 与植物的水力结构特征有关.随着土壤湿度的下降, 水分供应不足会增加水分从土壤运送到叶片的连续水柱断裂的风险, 当大叶片植物的蒸腾速率增大时, 空穴现象和栓塞现象更易发生(McCulloh & Sperry, 2005 ), 从而使得大叶片植物在水分稀缺的生境下不具有水分运输方面的选择优势, 这与Callaway等(1994) 的研究结果一致, 即: 一定大小枝条输导面积支持的总叶面积随着大气蒸腾强度和干旱强度的增加而减小. ...
Comparative studies of leaf form: Assessing the relative roles of selective pressures and phylogenetic constraints.
0
1987
Sizes and shapes of liane leaves.
2
1976
... 植物功能性状与环境和生态系统功能的关系是近年来生态学和全球变化研究的热点(Harrison et al ., 2010 ; Bernard-Verdier et al ., 2012 ), 各功能性状之间的权衡和组合决定了植物的生活史对策(Vandermeer, 1972 ; Silvertown, 2004 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ), 叶片是植物进行光合作用的主要器官(张林和罗天祥, 2004 ), 两者大小和数量的配置关系直接影响着植物的机械力学和水力结构特征, 从而决定了植物的生活史策略, 并可能影响物种生态位的利用(Givnish, 1987 ; Enquist et al ., 1999 ; West et al ., 1999 ; Westoby et al ., 2002 ).植物单位大小枝条上支撑的叶面积必须与支撑枝的横截面积成一定的比例, 才能符合植物器官间的机械力学和水力结构需求(Niklas, 1992 ; Enquist et al ., 1999 ; Normand et al ., 2008 ; Olson et al ., 2009 ).叶片大小和叶片数量是决定树冠紧密程度的两个重要因素, 直接影响到植株的冠层结构和发育模式, 进而影响到植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ).在某一给定的叶生物量分配或某一个给定的总叶面积下, 植物可能有较少的大叶子或者较多的小叶子(杨冬梅等, 2012a ).坡向作为一种重要的地形要素, 往往对局部区域的太阳辐射和降水进行二次分配(Cantón et al ., 2004 ), 在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 能够对植物的形态产生影响(Bazzaz, 1996 ; 韦莉莉等, 2005 ).当自然资源的丰富度低于或超过植物光合系统所能利用的最低或最高限度时, 植物的光合作用将被抑制(王玉萍等, 2013 ).为提高植物构型构建的生境适应性和风险规避能力, 植物通过功能性状等表型的可塑性调整提高自身的生存适合度, 体现了全球气候变化背景下植物的资源配置策略.因此, 研究坡向梯度上植物枝叶关系规律及其成因, 对于理解植物的环境适应机制有着重要意义. ...
... ; Givnish & Vermeij, 1976 ).在某一给定的叶生物量分配或某一个给定的总叶面积下, 植物可能有较少的大叶子或者较多的小叶子(杨冬梅等, 2012a ).坡向作为一种重要的地形要素, 往往对局部区域的太阳辐射和降水进行二次分配(Cantón et al ., 2004 ), 在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 能够对植物的形态产生影响(Bazzaz, 1996 ; 韦莉莉等, 2005 ).当自然资源的丰富度低于或超过植物光合系统所能利用的最低或最高限度时, 植物的光合作用将被抑制(王玉萍等, 2013 ).为提高植物构型构建的生境适应性和风险规避能力, 植物通过功能性状等表型的可塑性调整提高自身的生存适合度, 体现了全球气候变化背景下植物的资源配置策略.因此, 研究坡向梯度上植物枝叶关系规律及其成因, 对于理解植物的环境适应机制有着重要意义. ...
Ecophysiological and bioclimatic foundations for a global plant functional classification.
0
2010
1
1991
... 将坡向划分为北坡(315°-360°和0°-45°)、东坡(45°-135°)、南坡(135°-225°)和西坡(225°-315°).在所研究的4个功能性状中, 茎截面积指当年生小枝上1级枝的横截面积, 总叶面积指当年生小枝上所支撑的所有叶片面积之和, 出叶强度指单位体积枝条所支撑的叶片数, 单叶面积指当年生小枝上所支撑的所有叶片单叶面积的平均值, 枝条体积由其横截面积乘以长度获得(许月等, 2012 ).对每个样方刺槐枝叶属性的平均值进行对数(以10为底)转换后再进行分析, 对数转换使之符合正态分布.对枝大小-叶大小以及叶数量-叶大小两组功能性状关系的研究, 均采用y = axb , 线性转换成log(y ) = log(a ) + b log(x ), 式中x 和y 表示两个特征参数, a 为性状关系的截距, b 为斜率, 即异速生长参数或相对生长的指数, 当|b | = 1时, 表示两者是等速变化关系; 当|b| 显著偏离1时, 两者间为异速变化关系(Harvey & Pagel, 1991 ).数据分析主要采用标准化主轴估计(standardized major axis estimation, SMA)的方法(Warton et al ., 2006 ), 由软件(S)MATR Version 2.0 (Falster et al ., 2006 )计算完成.每一个回归斜率的置信区间根据Pitman (1939)方法计算.回归斜率与1或-1的显著性检验根据Warton和Weber (2002)的方法判断.在完成线性回归后, 对回归斜率进行异质性测试, 如果组间没有异质性, 则表示有共同斜率(Warton & Weber, 2002 ).此后, 不同坡向梯度间的功能性状的平均值的比较采用单因素方差分析(one-way ANOVA), 利用成对比较检验分析各坡向刺槐枝叶关系线性回归方程在y 轴上截距的差异. ...
不同坡向高寒退化草地狼毒株高和枝条数的权衡关系
1
2014
... 植物功能性状与生存环境之间是一种相互协同进化与适应的关系(侯兆疆等, 2014 ).枝叶大小关系是植物的重要生活史策略(Westoby et al ., 2002 ), 其比例不仅反映枝叶生物量的分配和叶片在生化和水力生物学方面的功能需求(Brouat et al ., 1998 ; Sun et al ., 2006 ; Normand et al ., 2008 ), 也体现了植物适应特定环境的构型塑造(Enquist & Niklas, 2001 , 2002 ).各坡向上刺槐茎截面积和总叶面积均呈正的异速生长关系, 回归方程间具有共同的SMA斜率且显著大于1 (p < 0.05)(图1 ), 二者的相关关系始终符合Corner法则(Corner, 1949 ), 不随微地形特征的变化而显著改变, 植物对总叶面积的投入显著高于对枝横截面积的投入, 这说明微地形差异无法改变植物在枝叶大小上的协调, 枝大小和叶大小间始终需要维持几何大小和重力的平衡(史青茹等, 2014 ).对于此现象, Westoby等(2002) 曾从枝叶的形态和生理关联性方面进行了解释, 他们认为: 一定大小的叶面积必须要由一定大小的枝来支撑, 两者间必须保持大小的匹配, 满足机械力学和水分动力学的要求, 才能保持机械支撑的平衡以及水分运输和蒸腾的协调; 管道模型理论(Niklas, 1992 )则提出, 为满足叶片对水分的需求, 木质部的横截面积与它们所支持的叶面积成正比.除此之外, West等(1997) 提出了结构分形自相似假说解释这种现象, 即: 所有生物的各器官间具有自相似的分形网络结构, 自然选择的压力必然使其向最优化的方向进化, 使得与外界环境的交换面积和新陈代谢效率最大化, 并使营养物质运输距离和时间最小化, 因此, 枝叶的大小必然按一定的比例来协调.而茎截面积-总叶面积关系的SMA斜率显著大于1 (p < 0.05), 主要是因为叶片是植株碳获取的主要功能构件, 所以在进化过程中为提高光合收益, 植株将尽可能地使分配到叶片中的生物量最大化, 而在叶片支撑结构中的投资最小化. ...
不同坡向高寒退化草地狼毒株高和枝条数的权衡关系
1
2014
... 植物功能性状与生存环境之间是一种相互协同进化与适应的关系(侯兆疆等, 2014 ).枝叶大小关系是植物的重要生活史策略(Westoby et al ., 2002 ), 其比例不仅反映枝叶生物量的分配和叶片在生化和水力生物学方面的功能需求(Brouat et al ., 1998 ; Sun et al ., 2006 ; Normand et al ., 2008 ), 也体现了植物适应特定环境的构型塑造(Enquist & Niklas, 2001 , 2002 ).各坡向上刺槐茎截面积和总叶面积均呈正的异速生长关系, 回归方程间具有共同的SMA斜率且显著大于1 (p < 0.05)(图1 ), 二者的相关关系始终符合Corner法则(Corner, 1949 ), 不随微地形特征的变化而显著改变, 植物对总叶面积的投入显著高于对枝横截面积的投入, 这说明微地形差异无法改变植物在枝叶大小上的协调, 枝大小和叶大小间始终需要维持几何大小和重力的平衡(史青茹等, 2014 ).对于此现象, Westoby等(2002) 曾从枝叶的形态和生理关联性方面进行了解释, 他们认为: 一定大小的叶面积必须要由一定大小的枝来支撑, 两者间必须保持大小的匹配, 满足机械力学和水分动力学的要求, 才能保持机械支撑的平衡以及水分运输和蒸腾的协调; 管道模型理论(Niklas, 1992 )则提出, 为满足叶片对水分的需求, 木质部的横截面积与它们所支持的叶面积成正比.除此之外, West等(1997) 提出了结构分形自相似假说解释这种现象, 即: 所有生物的各器官间具有自相似的分形网络结构, 自然选择的压力必然使其向最优化的方向进化, 使得与外界环境的交换面积和新陈代谢效率最大化, 并使营养物质运输距离和时间最小化, 因此, 枝叶的大小必然按一定的比例来协调.而茎截面积-总叶面积关系的SMA斜率显著大于1 (p < 0.05), 主要是因为叶片是植株碳获取的主要功能构件, 所以在进化过程中为提高光合收益, 植株将尽可能地使分配到叶片中的生物量最大化, 而在叶片支撑结构中的投资最小化. ...
黄土高原半干旱和半湿润地区刺槐林地生物量与土壤干燥化效应的模拟
2
2010
... 刺槐(Robinia pseudoacacia )是豆科刺槐属的温带落叶乔木, 具有生长速度快、根系发达、对环境适应性强、固碳氮能力较好、成活率高、萌芽力和根蘖性都很强等优点, 已成为我国黄土丘陵区的主要造林树种和人工林面积最大的树种(于占辉等, 2009 ; 李军等, 2010 ).近年来, 干旱地区许多刺槐人工林出现了生长减慢、枝梢干枯甚至整株死亡的衰败现象, 引起了学术界的广泛关注(单长卷等, 2004 ; 李军等, 2010 ).目前, 许多学者研究了刺槐叶面积生长与林分密度的关系(尹婧等, 2008 ), 土层厚度对刺槐旱季水分状况和生长的影响(王林等, 2013 ), 不同林龄和退耕年限刺槐人工林碳、氮储量及分配规律(申家朋和张文辉, 2014 ; 艾泽民等, 2014 ), 揭示了刺槐生长以及生物量积累和分配格局等一系列规律, 但关于微地形影响下刺槐当年生小枝内枝叶关系的研究较为少见, 更鲜有不同坡向茎截面积与总叶面积、出叶强度与单叶面积关系的报道, 限制了对异质生境中刺槐适应机制的解析.本研究选择兰州北山九州台西北师范大学绿化基地的刺槐人工林为研究对象, 探讨刺槐当年生小枝内的茎截面积-总叶面积、出叶强度-单叶面积关系, 为进一步了解刺槐的环境适应机制提供理论依据, 对提高干旱半干旱地区人工林的可持续性和稳定性具有重要意义. ...
... ; 李军等, 2010 ).目前, 许多学者研究了刺槐叶面积生长与林分密度的关系(尹婧等, 2008 ), 土层厚度对刺槐旱季水分状况和生长的影响(王林等, 2013 ), 不同林龄和退耕年限刺槐人工林碳、氮储量及分配规律(申家朋和张文辉, 2014 ; 艾泽民等, 2014 ), 揭示了刺槐生长以及生物量积累和分配格局等一系列规律, 但关于微地形影响下刺槐当年生小枝内枝叶关系的研究较为少见, 更鲜有不同坡向茎截面积与总叶面积、出叶强度与单叶面积关系的报道, 限制了对异质生境中刺槐适应机制的解析.本研究选择兰州北山九州台西北师范大学绿化基地的刺槐人工林为研究对象, 探讨刺槐当年生小枝内的茎截面积-总叶面积、出叶强度-单叶面积关系, 为进一步了解刺槐的环境适应机制提供理论依据, 对提高干旱半干旱地区人工林的可持续性和稳定性具有重要意义. ...
黄土高原半干旱和半湿润地区刺槐林地生物量与土壤干燥化效应的模拟
2
2010
... 刺槐(Robinia pseudoacacia )是豆科刺槐属的温带落叶乔木, 具有生长速度快、根系发达、对环境适应性强、固碳氮能力较好、成活率高、萌芽力和根蘖性都很强等优点, 已成为我国黄土丘陵区的主要造林树种和人工林面积最大的树种(于占辉等, 2009 ; 李军等, 2010 ).近年来, 干旱地区许多刺槐人工林出现了生长减慢、枝梢干枯甚至整株死亡的衰败现象, 引起了学术界的广泛关注(单长卷等, 2004 ; 李军等, 2010 ).目前, 许多学者研究了刺槐叶面积生长与林分密度的关系(尹婧等, 2008 ), 土层厚度对刺槐旱季水分状况和生长的影响(王林等, 2013 ), 不同林龄和退耕年限刺槐人工林碳、氮储量及分配规律(申家朋和张文辉, 2014 ; 艾泽民等, 2014 ), 揭示了刺槐生长以及生物量积累和分配格局等一系列规律, 但关于微地形影响下刺槐当年生小枝内枝叶关系的研究较为少见, 更鲜有不同坡向茎截面积与总叶面积、出叶强度与单叶面积关系的报道, 限制了对异质生境中刺槐适应机制的解析.本研究选择兰州北山九州台西北师范大学绿化基地的刺槐人工林为研究对象, 探讨刺槐当年生小枝内的茎截面积-总叶面积、出叶强度-单叶面积关系, 为进一步了解刺槐的环境适应机制提供理论依据, 对提高干旱半干旱地区人工林的可持续性和稳定性具有重要意义. ...
... ; 李军等, 2010 ).目前, 许多学者研究了刺槐叶面积生长与林分密度的关系(尹婧等, 2008 ), 土层厚度对刺槐旱季水分状况和生长的影响(王林等, 2013 ), 不同林龄和退耕年限刺槐人工林碳、氮储量及分配规律(申家朋和张文辉, 2014 ; 艾泽民等, 2014 ), 揭示了刺槐生长以及生物量积累和分配格局等一系列规律, 但关于微地形影响下刺槐当年生小枝内枝叶关系的研究较为少见, 更鲜有不同坡向茎截面积与总叶面积、出叶强度与单叶面积关系的报道, 限制了对异质生境中刺槐适应机制的解析.本研究选择兰州北山九州台西北师范大学绿化基地的刺槐人工林为研究对象, 探讨刺槐当年生小枝内的茎截面积-总叶面积、出叶强度-单叶面积关系, 为进一步了解刺槐的环境适应机制提供理论依据, 对提高干旱半干旱地区人工林的可持续性和稳定性具有重要意义. ...
高寒草地狼毒枝-叶性状对坡向的响应
2
2013
... 坡向虽然对刺槐枝叶大小回归方程的斜率没有显著影响, 但使回归方程的截距产生了显著改变.在茎截面积-总叶面积关系中, 其回归方程的截距在北坡显著大于南坡(p < 0.05), 表明在茎截面积一定时, 位于北坡的刺槐在单位大小枝条上支持的总叶面积大于位于南坡的刺槐.叶片大小是植物适应环境中热量和水分交换平衡的结果, 其变化主要是通过影响叶边缘对热量、CO2 和水分的传导能力来影响整个植株的生长速率(杨冬梅等, 2012b ).南坡光照时间长, 水分蒸发大(殷秀琴, 2004 ), 土壤水分含量低(表1 ), 在西北年降水量小的干旱半干旱地区, 由于高温常引起蒸腾作用加强而使植物体水分大量丧失, 因而这种高温和干旱环境严重影响了植物的新陈代谢活动, 细胞的分裂和生长受到限制, 植物生长缓慢, 叶片较小; 然而北坡气温降低, 地面蒸发量减少, 土壤水分增加(表1 ), 良好的水热条件有利于细胞体积和数目增加, 从而促进了叶面积的扩展(李钰等, 2013 ).反过来, 与小叶相比, 大叶具有更厚的叶边缘, 热量和物质通过叶边缘进行转运的阻力更大, 往往会阻止叶片与周围的空气进行热量交换, 减慢CO2 和水蒸气进出叶片的扩散速率, 使大叶比小叶更容易过热, 植物的呼吸和蒸腾成本更高(Givnish, 1978 ), 因此在冷凉、湿润或者荫蔽的环境下大叶由于能够使光合收益最大化而占有优势(Givnish, 1979 ), 而相对小的叶片因为具有较高的热交换能力, 被认为在热、干、高光和低养分的环境下较有优势(Ackerly & Preston, 2003 ; McDonald et al ., 2003 ).另外, 大叶片植物在水分较少的生境中不具有选择优势, 与植物的水力结构特征有关.随着土壤湿度的下降, 水分供应不足会增加水分从土壤运送到叶片的连续水柱断裂的风险, 当大叶片植物的蒸腾速率增大时, 空穴现象和栓塞现象更易发生(McCulloh & Sperry, 2005 ), 从而使得大叶片植物在水分稀缺的生境下不具有水分运输方面的选择优势, 这与Callaway等(1994) 的研究结果一致, 即: 一定大小枝条输导面积支持的总叶面积随着大气蒸腾强度和干旱强度的增加而减小. ...
... 在各坡向上, 刺槐出叶强度与单叶面积均表现出负的等速生长关系, 说明刺槐的叶存在大小和数量上的生态权衡, 这与杨冬梅等(2012a) 的研究结果一致.位于北坡的刺槐出叶强度小于南坡, 单叶面积大于南坡(表2 ), 如前所述, 叶大小-叶数量的权衡关系是植物对外部环境选择压力的功能性适应结果.北坡光照资源最弱, 且该坡向上群落郁闭程度和拥挤程度大于阳坡(表1 ), 邻体干扰增强, 密度制约改变了植物间可获得性资源的数量, 为使光合效益最大化, 刺槐当年生小枝上长出较大的叶片, 用以增加光合同化面积; 与此同时, 过多的叶片会造成上部叶片对下部叶片的遮挡(李钰等, 2013 ), 使得叶片间互相拥挤的程度加剧, 不利于光资源的充分利用, 因此北坡刺槐形成了“增大单叶面积, 减小出叶强度”的生长策略, 南坡刺槐则形成了“减小单叶面积, 增大出叶强度”的生长策略, 有利于植株本身的光拦截能力和光合碳获取能力最大化. ...
高寒草地狼毒枝-叶性状对坡向的响应
2
2013
... 坡向虽然对刺槐枝叶大小回归方程的斜率没有显著影响, 但使回归方程的截距产生了显著改变.在茎截面积-总叶面积关系中, 其回归方程的截距在北坡显著大于南坡(p < 0.05), 表明在茎截面积一定时, 位于北坡的刺槐在单位大小枝条上支持的总叶面积大于位于南坡的刺槐.叶片大小是植物适应环境中热量和水分交换平衡的结果, 其变化主要是通过影响叶边缘对热量、CO2 和水分的传导能力来影响整个植株的生长速率(杨冬梅等, 2012b ).南坡光照时间长, 水分蒸发大(殷秀琴, 2004 ), 土壤水分含量低(表1 ), 在西北年降水量小的干旱半干旱地区, 由于高温常引起蒸腾作用加强而使植物体水分大量丧失, 因而这种高温和干旱环境严重影响了植物的新陈代谢活动, 细胞的分裂和生长受到限制, 植物生长缓慢, 叶片较小; 然而北坡气温降低, 地面蒸发量减少, 土壤水分增加(表1 ), 良好的水热条件有利于细胞体积和数目增加, 从而促进了叶面积的扩展(李钰等, 2013 ).反过来, 与小叶相比, 大叶具有更厚的叶边缘, 热量和物质通过叶边缘进行转运的阻力更大, 往往会阻止叶片与周围的空气进行热量交换, 减慢CO2 和水蒸气进出叶片的扩散速率, 使大叶比小叶更容易过热, 植物的呼吸和蒸腾成本更高(Givnish, 1978 ), 因此在冷凉、湿润或者荫蔽的环境下大叶由于能够使光合收益最大化而占有优势(Givnish, 1979 ), 而相对小的叶片因为具有较高的热交换能力, 被认为在热、干、高光和低养分的环境下较有优势(Ackerly & Preston, 2003 ; McDonald et al ., 2003 ).另外, 大叶片植物在水分较少的生境中不具有选择优势, 与植物的水力结构特征有关.随着土壤湿度的下降, 水分供应不足会增加水分从土壤运送到叶片的连续水柱断裂的风险, 当大叶片植物的蒸腾速率增大时, 空穴现象和栓塞现象更易发生(McCulloh & Sperry, 2005 ), 从而使得大叶片植物在水分稀缺的生境下不具有水分运输方面的选择优势, 这与Callaway等(1994) 的研究结果一致, 即: 一定大小枝条输导面积支持的总叶面积随着大气蒸腾强度和干旱强度的增加而减小. ...
... 在各坡向上, 刺槐出叶强度与单叶面积均表现出负的等速生长关系, 说明刺槐的叶存在大小和数量上的生态权衡, 这与杨冬梅等(2012a) 的研究结果一致.位于北坡的刺槐出叶强度小于南坡, 单叶面积大于南坡(表2 ), 如前所述, 叶大小-叶数量的权衡关系是植物对外部环境选择压力的功能性适应结果.北坡光照资源最弱, 且该坡向上群落郁闭程度和拥挤程度大于阳坡(表1 ), 邻体干扰增强, 密度制约改变了植物间可获得性资源的数量, 为使光合效益最大化, 刺槐当年生小枝上长出较大的叶片, 用以增加光合同化面积; 与此同时, 过多的叶片会造成上部叶片对下部叶片的遮挡(李钰等, 2013 ), 使得叶片间互相拥挤的程度加剧, 不利于光资源的充分利用, 因此北坡刺槐形成了“增大单叶面积, 减小出叶强度”的生长策略, 南坡刺槐则形成了“减小单叶面积, 增大出叶强度”的生长策略, 有利于植株本身的光拦截能力和光合碳获取能力最大化. ...
杜鹃花属植物小枝大小对小枝生物量分配及叶面积支持效率的影响: 异速生长分析
1
2008
... 植物功能性状与环境和生态系统功能的关系是近年来生态学和全球变化研究的热点(Harrison et al ., 2010 ; Bernard-Verdier et al ., 2012 ), 各功能性状之间的权衡和组合决定了植物的生活史对策(Vandermeer, 1972 ; Silvertown, 2004 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ), 叶片是植物进行光合作用的主要器官(张林和罗天祥, 2004 ), 两者大小和数量的配置关系直接影响着植物的机械力学和水力结构特征, 从而决定了植物的生活史策略, 并可能影响物种生态位的利用(Givnish, 1987 ; Enquist et al ., 1999 ; West et al ., 1999 ; Westoby et al ., 2002 ).植物单位大小枝条上支撑的叶面积必须与支撑枝的横截面积成一定的比例, 才能符合植物器官间的机械力学和水力结构需求(Niklas, 1992 ; Enquist et al ., 1999 ; Normand et al ., 2008 ; Olson et al ., 2009 ).叶片大小和叶片数量是决定树冠紧密程度的两个重要因素, 直接影响到植株的冠层结构和发育模式, 进而影响到植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ).在某一给定的叶生物量分配或某一个给定的总叶面积下, 植物可能有较少的大叶子或者较多的小叶子(杨冬梅等, 2012a ).坡向作为一种重要的地形要素, 往往对局部区域的太阳辐射和降水进行二次分配(Cantón et al ., 2004 ), 在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 能够对植物的形态产生影响(Bazzaz, 1996 ; 韦莉莉等, 2005 ).当自然资源的丰富度低于或超过植物光合系统所能利用的最低或最高限度时, 植物的光合作用将被抑制(王玉萍等, 2013 ).为提高植物构型构建的生境适应性和风险规避能力, 植物通过功能性状等表型的可塑性调整提高自身的生存适合度, 体现了全球气候变化背景下植物的资源配置策略.因此, 研究坡向梯度上植物枝叶关系规律及其成因, 对于理解植物的环境适应机制有着重要意义. ...
杜鹃花属植物小枝大小对小枝生物量分配及叶面积支持效率的影响: 异速生长分析
1
2008
... 植物功能性状与环境和生态系统功能的关系是近年来生态学和全球变化研究的热点(Harrison et al ., 2010 ; Bernard-Verdier et al ., 2012 ), 各功能性状之间的权衡和组合决定了植物的生活史对策(Vandermeer, 1972 ; Silvertown, 2004 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ), 叶片是植物进行光合作用的主要器官(张林和罗天祥, 2004 ), 两者大小和数量的配置关系直接影响着植物的机械力学和水力结构特征, 从而决定了植物的生活史策略, 并可能影响物种生态位的利用(Givnish, 1987 ; Enquist et al ., 1999 ; West et al ., 1999 ; Westoby et al ., 2002 ).植物单位大小枝条上支撑的叶面积必须与支撑枝的横截面积成一定的比例, 才能符合植物器官间的机械力学和水力结构需求(Niklas, 1992 ; Enquist et al ., 1999 ; Normand et al ., 2008 ; Olson et al ., 2009 ).叶片大小和叶片数量是决定树冠紧密程度的两个重要因素, 直接影响到植株的冠层结构和发育模式, 进而影响到植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ).在某一给定的叶生物量分配或某一个给定的总叶面积下, 植物可能有较少的大叶子或者较多的小叶子(杨冬梅等, 2012a ).坡向作为一种重要的地形要素, 往往对局部区域的太阳辐射和降水进行二次分配(Cantón et al ., 2004 ), 在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 能够对植物的形态产生影响(Bazzaz, 1996 ; 韦莉莉等, 2005 ).当自然资源的丰富度低于或超过植物光合系统所能利用的最低或最高限度时, 植物的光合作用将被抑制(王玉萍等, 2013 ).为提高植物构型构建的生境适应性和风险规避能力, 植物通过功能性状等表型的可塑性调整提高自身的生存适合度, 体现了全球气候变化背景下植物的资源配置策略.因此, 研究坡向梯度上植物枝叶关系规律及其成因, 对于理解植物的环境适应机制有着重要意义. ...
Patterns in hydraulic architecture and their implications for transport efficiency.
2
2005
... 坡向虽然对刺槐枝叶大小回归方程的斜率没有显著影响, 但使回归方程的截距产生了显著改变.在茎截面积-总叶面积关系中, 其回归方程的截距在北坡显著大于南坡(p < 0.05), 表明在茎截面积一定时, 位于北坡的刺槐在单位大小枝条上支持的总叶面积大于位于南坡的刺槐.叶片大小是植物适应环境中热量和水分交换平衡的结果, 其变化主要是通过影响叶边缘对热量、CO2 和水分的传导能力来影响整个植株的生长速率(杨冬梅等, 2012b ).南坡光照时间长, 水分蒸发大(殷秀琴, 2004 ), 土壤水分含量低(表1 ), 在西北年降水量小的干旱半干旱地区, 由于高温常引起蒸腾作用加强而使植物体水分大量丧失, 因而这种高温和干旱环境严重影响了植物的新陈代谢活动, 细胞的分裂和生长受到限制, 植物生长缓慢, 叶片较小; 然而北坡气温降低, 地面蒸发量减少, 土壤水分增加(表1 ), 良好的水热条件有利于细胞体积和数目增加, 从而促进了叶面积的扩展(李钰等, 2013 ).反过来, 与小叶相比, 大叶具有更厚的叶边缘, 热量和物质通过叶边缘进行转运的阻力更大, 往往会阻止叶片与周围的空气进行热量交换, 减慢CO2 和水蒸气进出叶片的扩散速率, 使大叶比小叶更容易过热, 植物的呼吸和蒸腾成本更高(Givnish, 1978 ), 因此在冷凉、湿润或者荫蔽的环境下大叶由于能够使光合收益最大化而占有优势(Givnish, 1979 ), 而相对小的叶片因为具有较高的热交换能力, 被认为在热、干、高光和低养分的环境下较有优势(Ackerly & Preston, 2003 ; McDonald et al ., 2003 ).另外, 大叶片植物在水分较少的生境中不具有选择优势, 与植物的水力结构特征有关.随着土壤湿度的下降, 水分供应不足会增加水分从土壤运送到叶片的连续水柱断裂的风险, 当大叶片植物的蒸腾速率增大时, 空穴现象和栓塞现象更易发生(McCulloh & Sperry, 2005 ), 从而使得大叶片植物在水分稀缺的生境下不具有水分运输方面的选择优势, 这与Callaway等(1994) 的研究结果一致, 即: 一定大小枝条输导面积支持的总叶面积随着大气蒸腾强度和干旱强度的增加而减小. ...
... ).另外, 大叶片植物在水分较少的生境中不具有选择优势, 与植物的水力结构特征有关.随着土壤湿度的下降, 水分供应不足会增加水分从土壤运送到叶片的连续水柱断裂的风险, 当大叶片植物的蒸腾速率增大时, 空穴现象和栓塞现象更易发生(McCulloh & Sperry, 2005 ), 从而使得大叶片植物在水分稀缺的生境下不具有水分运输方面的选择优势, 这与Callaway等(1994) 的研究结果一致, 即: 一定大小枝条输导面积支持的总叶面积随着大气蒸腾强度和干旱强度的增加而减小. ...
Leaf-size divergence along rainfall and soil-nutrient gradients: Is the method of size reduction common among clades?
0
2003
5
1992
... 植物功能性状与环境和生态系统功能的关系是近年来生态学和全球变化研究的热点(Harrison et al ., 2010 ; Bernard-Verdier et al ., 2012 ), 各功能性状之间的权衡和组合决定了植物的生活史对策(Vandermeer, 1972 ; Silvertown, 2004 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ), 叶片是植物进行光合作用的主要器官(张林和罗天祥, 2004 ), 两者大小和数量的配置关系直接影响着植物的机械力学和水力结构特征, 从而决定了植物的生活史策略, 并可能影响物种生态位的利用(Givnish, 1987 ; Enquist et al ., 1999 ; West et al ., 1999 ; Westoby et al ., 2002 ).植物单位大小枝条上支撑的叶面积必须与支撑枝的横截面积成一定的比例, 才能符合植物器官间的机械力学和水力结构需求(Niklas, 1992 ; Enquist et al ., 1999 ; Normand et al ., 2008 ; Olson et al ., 2009 ).叶片大小和叶片数量是决定树冠紧密程度的两个重要因素, 直接影响到植株的冠层结构和发育模式, 进而影响到植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ).在某一给定的叶生物量分配或某一个给定的总叶面积下, 植物可能有较少的大叶子或者较多的小叶子(杨冬梅等, 2012a ).坡向作为一种重要的地形要素, 往往对局部区域的太阳辐射和降水进行二次分配(Cantón et al ., 2004 ), 在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 能够对植物的形态产生影响(Bazzaz, 1996 ; 韦莉莉等, 2005 ).当自然资源的丰富度低于或超过植物光合系统所能利用的最低或最高限度时, 植物的光合作用将被抑制(王玉萍等, 2013 ).为提高植物构型构建的生境适应性和风险规避能力, 植物通过功能性状等表型的可塑性调整提高自身的生存适合度, 体现了全球气候变化背景下植物的资源配置策略.因此, 研究坡向梯度上植物枝叶关系规律及其成因, 对于理解植物的环境适应机制有着重要意义. ...
... ; Normand et al ., 2008 ; Olson et al ., 2009 ).叶片大小和叶片数量是决定树冠紧密程度的两个重要因素, 直接影响到植株的冠层结构和发育模式, 进而影响到植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ).在某一给定的叶生物量分配或某一个给定的总叶面积下, 植物可能有较少的大叶子或者较多的小叶子(杨冬梅等, 2012a ).坡向作为一种重要的地形要素, 往往对局部区域的太阳辐射和降水进行二次分配(Cantón et al ., 2004 ), 在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 能够对植物的形态产生影响(Bazzaz, 1996 ; 韦莉莉等, 2005 ).当自然资源的丰富度低于或超过植物光合系统所能利用的最低或最高限度时, 植物的光合作用将被抑制(王玉萍等, 2013 ).为提高植物构型构建的生境适应性和风险规避能力, 植物通过功能性状等表型的可塑性调整提高自身的生存适合度, 体现了全球气候变化背景下植物的资源配置策略.因此, 研究坡向梯度上植物枝叶关系规律及其成因, 对于理解植物的环境适应机制有着重要意义. ...
... 植物在长期的生长发育过程中, 各构件在异质生境条件下会表现出不同的生态对策(Yang et al ., 2008 ), 植物外部形态具有高度的可塑性特征, 使其能够把资源最佳地分配到不同的器官, 以规避环境风险.本研究发现, 在各坡向间, 刺槐茎截面积和总叶面积均维持显著的正相关关系(p < 0.05), 回归方程间存在共同斜率且异速生长斜率大于1, 此结果与其他枝叶大小关系研究的结果基本一致(Westoby & Wright, 2003 ; Normand et al ., 2008 ; 史青茹等, 2014 ), 出叶强度和单叶面积则表现出显著的等速负相关关系(p < 0.05).两组关系的回归方程截距均为北坡最大、南坡最小, 这主要与坡向引起环境条件的变化和刺槐枝叶形态在异质生境中的适应机制有关. ...
... 植物功能性状与生存环境之间是一种相互协同进化与适应的关系(侯兆疆等, 2014 ).枝叶大小关系是植物的重要生活史策略(Westoby et al ., 2002 ), 其比例不仅反映枝叶生物量的分配和叶片在生化和水力生物学方面的功能需求(Brouat et al ., 1998 ; Sun et al ., 2006 ; Normand et al ., 2008 ), 也体现了植物适应特定环境的构型塑造(Enquist & Niklas, 2001 , 2002 ).各坡向上刺槐茎截面积和总叶面积均呈正的异速生长关系, 回归方程间具有共同的SMA斜率且显著大于1 (p < 0.05)(图1 ), 二者的相关关系始终符合Corner法则(Corner, 1949 ), 不随微地形特征的变化而显著改变, 植物对总叶面积的投入显著高于对枝横截面积的投入, 这说明微地形差异无法改变植物在枝叶大小上的协调, 枝大小和叶大小间始终需要维持几何大小和重力的平衡(史青茹等, 2014 ).对于此现象, Westoby等(2002) 曾从枝叶的形态和生理关联性方面进行了解释, 他们认为: 一定大小的叶面积必须要由一定大小的枝来支撑, 两者间必须保持大小的匹配, 满足机械力学和水分动力学的要求, 才能保持机械支撑的平衡以及水分运输和蒸腾的协调; 管道模型理论(Niklas, 1992 )则提出, 为满足叶片对水分的需求, 木质部的横截面积与它们所支持的叶面积成正比.除此之外, West等(1997) 提出了结构分形自相似假说解释这种现象, 即: 所有生物的各器官间具有自相似的分形网络结构, 自然选择的压力必然使其向最优化的方向进化, 使得与外界环境的交换面积和新陈代谢效率最大化, 并使营养物质运输距离和时间最小化, 因此, 枝叶的大小必然按一定的比例来协调.而茎截面积-总叶面积关系的SMA斜率显著大于1 (p < 0.05), 主要是因为叶片是植株碳获取的主要功能构件, 所以在进化过程中为提高光合收益, 植株将尽可能地使分配到叶片中的生物量最大化, 而在叶片支撑结构中的投资最小化. ...
... 曾从枝叶的形态和生理关联性方面进行了解释, 他们认为: 一定大小的叶面积必须要由一定大小的枝来支撑, 两者间必须保持大小的匹配, 满足机械力学和水分动力学的要求, 才能保持机械支撑的平衡以及水分运输和蒸腾的协调; 管道模型理论(Niklas, 1992 )则提出, 为满足叶片对水分的需求, 木质部的横截面积与它们所支持的叶面积成正比.除此之外, West等(1997) 提出了结构分形自相似假说解释这种现象, 即: 所有生物的各器官间具有自相似的分形网络结构, 自然选择的压力必然使其向最优化的方向进化, 使得与外界环境的交换面积和新陈代谢效率最大化, 并使营养物质运输距离和时间最小化, 因此, 枝叶的大小必然按一定的比例来协调.而茎截面积-总叶面积关系的SMA斜率显著大于1 (p < 0.05), 主要是因为叶片是植株碳获取的主要功能构件, 所以在进化过程中为提高光合收益, 植株将尽可能地使分配到叶片中的生物量最大化, 而在叶片支撑结构中的投资最小化. ...
Hydraulic and mechanical stem properties affect leaf-stem allometry in mango cultivars.
1
2008
... 植物功能性状与环境和生态系统功能的关系是近年来生态学和全球变化研究的热点(Harrison et al ., 2010 ; Bernard-Verdier et al ., 2012 ), 各功能性状之间的权衡和组合决定了植物的生活史对策(Vandermeer, 1972 ; Silvertown, 2004 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ), 叶片是植物进行光合作用的主要器官(张林和罗天祥, 2004 ), 两者大小和数量的配置关系直接影响着植物的机械力学和水力结构特征, 从而决定了植物的生活史策略, 并可能影响物种生态位的利用(Givnish, 1987 ; Enquist et al ., 1999 ; West et al ., 1999 ; Westoby et al ., 2002 ).植物单位大小枝条上支撑的叶面积必须与支撑枝的横截面积成一定的比例, 才能符合植物器官间的机械力学和水力结构需求(Niklas, 1992 ; Enquist et al ., 1999 ; Normand et al ., 2008 ; Olson et al ., 2009 ).叶片大小和叶片数量是决定树冠紧密程度的两个重要因素, 直接影响到植株的冠层结构和发育模式, 进而影响到植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ).在某一给定的叶生物量分配或某一个给定的总叶面积下, 植物可能有较少的大叶子或者较多的小叶子(杨冬梅等, 2012a ).坡向作为一种重要的地形要素, 往往对局部区域的太阳辐射和降水进行二次分配(Cantón et al ., 2004 ), 在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 能够对植物的形态产生影响(Bazzaz, 1996 ; 韦莉莉等, 2005 ).当自然资源的丰富度低于或超过植物光合系统所能利用的最低或最高限度时, 植物的光合作用将被抑制(王玉萍等, 2013 ).为提高植物构型构建的生境适应性和风险规避能力, 植物通过功能性状等表型的可塑性调整提高自身的生存适合度, 体现了全球气候变化背景下植物的资源配置策略.因此, 研究坡向梯度上植物枝叶关系规律及其成因, 对于理解植物的环境适应机制有着重要意义. ...
Universal foliage-stem scaling across environments and species in dicot trees: Plasticity, biomechanics and Corner’s rules.
0
2009
Optimal leaf size in relation to environment.
1
1972
... 植物功能性状与环境和生态系统功能的关系是近年来生态学和全球变化研究的热点(Harrison et al ., 2010 ; Bernard-Verdier et al ., 2012 ), 各功能性状之间的权衡和组合决定了植物的生活史对策(Vandermeer, 1972 ; Silvertown, 2004 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ), 叶片是植物进行光合作用的主要器官(张林和罗天祥, 2004 ), 两者大小和数量的配置关系直接影响着植物的机械力学和水力结构特征, 从而决定了植物的生活史策略, 并可能影响物种生态位的利用(Givnish, 1987 ; Enquist et al ., 1999 ; West et al ., 1999 ; Westoby et al ., 2002 ).植物单位大小枝条上支撑的叶面积必须与支撑枝的横截面积成一定的比例, 才能符合植物器官间的机械力学和水力结构需求(Niklas, 1992 ; Enquist et al ., 1999 ; Normand et al ., 2008 ; Olson et al ., 2009 ).叶片大小和叶片数量是决定树冠紧密程度的两个重要因素, 直接影响到植株的冠层结构和发育模式, 进而影响到植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ).在某一给定的叶生物量分配或某一个给定的总叶面积下, 植物可能有较少的大叶子或者较多的小叶子(杨冬梅等, 2012a ).坡向作为一种重要的地形要素, 往往对局部区域的太阳辐射和降水进行二次分配(Cantón et al ., 2004 ), 在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 能够对植物的形态产生影响(Bazzaz, 1996 ; 韦莉莉等, 2005 ).当自然资源的丰富度低于或超过植物光合系统所能利用的最低或最高限度时, 植物的光合作用将被抑制(王玉萍等, 2013 ).为提高植物构型构建的生境适应性和风险规避能力, 植物通过功能性状等表型的可塑性调整提高自身的生存适合度, 体现了全球气候变化背景下植物的资源配置策略.因此, 研究坡向梯度上植物枝叶关系规律及其成因, 对于理解植物的环境适应机制有着重要意义. ...
A note on normal correlation.
0
1939
黄土高原不同立地条件下刺槐生长与水分关系研究
1
2004
... 刺槐(Robinia pseudoacacia )是豆科刺槐属的温带落叶乔木, 具有生长速度快、根系发达、对环境适应性强、固碳氮能力较好、成活率高、萌芽力和根蘖性都很强等优点, 已成为我国黄土丘陵区的主要造林树种和人工林面积最大的树种(于占辉等, 2009 ; 李军等, 2010 ).近年来, 干旱地区许多刺槐人工林出现了生长减慢、枝梢干枯甚至整株死亡的衰败现象, 引起了学术界的广泛关注(单长卷等, 2004 ; 李军等, 2010 ).目前, 许多学者研究了刺槐叶面积生长与林分密度的关系(尹婧等, 2008 ), 土层厚度对刺槐旱季水分状况和生长的影响(王林等, 2013 ), 不同林龄和退耕年限刺槐人工林碳、氮储量及分配规律(申家朋和张文辉, 2014 ; 艾泽民等, 2014 ), 揭示了刺槐生长以及生物量积累和分配格局等一系列规律, 但关于微地形影响下刺槐当年生小枝内枝叶关系的研究较为少见, 更鲜有不同坡向茎截面积与总叶面积、出叶强度与单叶面积关系的报道, 限制了对异质生境中刺槐适应机制的解析.本研究选择兰州北山九州台西北师范大学绿化基地的刺槐人工林为研究对象, 探讨刺槐当年生小枝内的茎截面积-总叶面积、出叶强度-单叶面积关系, 为进一步了解刺槐的环境适应机制提供理论依据, 对提高干旱半干旱地区人工林的可持续性和稳定性具有重要意义. ...
黄土高原不同立地条件下刺槐生长与水分关系研究
1
2004
... 刺槐(Robinia pseudoacacia )是豆科刺槐属的温带落叶乔木, 具有生长速度快、根系发达、对环境适应性强、固碳氮能力较好、成活率高、萌芽力和根蘖性都很强等优点, 已成为我国黄土丘陵区的主要造林树种和人工林面积最大的树种(于占辉等, 2009 ; 李军等, 2010 ).近年来, 干旱地区许多刺槐人工林出现了生长减慢、枝梢干枯甚至整株死亡的衰败现象, 引起了学术界的广泛关注(单长卷等, 2004 ; 李军等, 2010 ).目前, 许多学者研究了刺槐叶面积生长与林分密度的关系(尹婧等, 2008 ), 土层厚度对刺槐旱季水分状况和生长的影响(王林等, 2013 ), 不同林龄和退耕年限刺槐人工林碳、氮储量及分配规律(申家朋和张文辉, 2014 ; 艾泽民等, 2014 ), 揭示了刺槐生长以及生物量积累和分配格局等一系列规律, 但关于微地形影响下刺槐当年生小枝内枝叶关系的研究较为少见, 更鲜有不同坡向茎截面积与总叶面积、出叶强度与单叶面积关系的报道, 限制了对异质生境中刺槐适应机制的解析.本研究选择兰州北山九州台西北师范大学绿化基地的刺槐人工林为研究对象, 探讨刺槐当年生小枝内的茎截面积-总叶面积、出叶强度-单叶面积关系, 为进一步了解刺槐的环境适应机制提供理论依据, 对提高干旱半干旱地区人工林的可持续性和稳定性具有重要意义. ...
黄土丘陵区退耕还林地刺槐人工林碳储量及分配规律
1
2014
... 刺槐(Robinia pseudoacacia )是豆科刺槐属的温带落叶乔木, 具有生长速度快、根系发达、对环境适应性强、固碳氮能力较好、成活率高、萌芽力和根蘖性都很强等优点, 已成为我国黄土丘陵区的主要造林树种和人工林面积最大的树种(于占辉等, 2009 ; 李军等, 2010 ).近年来, 干旱地区许多刺槐人工林出现了生长减慢、枝梢干枯甚至整株死亡的衰败现象, 引起了学术界的广泛关注(单长卷等, 2004 ; 李军等, 2010 ).目前, 许多学者研究了刺槐叶面积生长与林分密度的关系(尹婧等, 2008 ), 土层厚度对刺槐旱季水分状况和生长的影响(王林等, 2013 ), 不同林龄和退耕年限刺槐人工林碳、氮储量及分配规律(申家朋和张文辉, 2014 ; 艾泽民等, 2014 ), 揭示了刺槐生长以及生物量积累和分配格局等一系列规律, 但关于微地形影响下刺槐当年生小枝内枝叶关系的研究较为少见, 更鲜有不同坡向茎截面积与总叶面积、出叶强度与单叶面积关系的报道, 限制了对异质生境中刺槐适应机制的解析.本研究选择兰州北山九州台西北师范大学绿化基地的刺槐人工林为研究对象, 探讨刺槐当年生小枝内的茎截面积-总叶面积、出叶强度-单叶面积关系, 为进一步了解刺槐的环境适应机制提供理论依据, 对提高干旱半干旱地区人工林的可持续性和稳定性具有重要意义. ...
黄土丘陵区退耕还林地刺槐人工林碳储量及分配规律
1
2014
... 刺槐(Robinia pseudoacacia )是豆科刺槐属的温带落叶乔木, 具有生长速度快、根系发达、对环境适应性强、固碳氮能力较好、成活率高、萌芽力和根蘖性都很强等优点, 已成为我国黄土丘陵区的主要造林树种和人工林面积最大的树种(于占辉等, 2009 ; 李军等, 2010 ).近年来, 干旱地区许多刺槐人工林出现了生长减慢、枝梢干枯甚至整株死亡的衰败现象, 引起了学术界的广泛关注(单长卷等, 2004 ; 李军等, 2010 ).目前, 许多学者研究了刺槐叶面积生长与林分密度的关系(尹婧等, 2008 ), 土层厚度对刺槐旱季水分状况和生长的影响(王林等, 2013 ), 不同林龄和退耕年限刺槐人工林碳、氮储量及分配规律(申家朋和张文辉, 2014 ; 艾泽民等, 2014 ), 揭示了刺槐生长以及生物量积累和分配格局等一系列规律, 但关于微地形影响下刺槐当年生小枝内枝叶关系的研究较为少见, 更鲜有不同坡向茎截面积与总叶面积、出叶强度与单叶面积关系的报道, 限制了对异质生境中刺槐适应机制的解析.本研究选择兰州北山九州台西北师范大学绿化基地的刺槐人工林为研究对象, 探讨刺槐当年生小枝内的茎截面积-总叶面积、出叶强度-单叶面积关系, 为进一步了解刺槐的环境适应机制提供理论依据, 对提高干旱半干旱地区人工林的可持续性和稳定性具有重要意义. ...
浙江天童木本植物Corner法则的检验: 微地形的影响
3
2014
... 植物功能性状与环境和生态系统功能的关系是近年来生态学和全球变化研究的热点(Harrison et al ., 2010 ; Bernard-Verdier et al ., 2012 ), 各功能性状之间的权衡和组合决定了植物的生活史对策(Vandermeer, 1972 ; Silvertown, 2004 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ), 叶片是植物进行光合作用的主要器官(张林和罗天祥, 2004 ), 两者大小和数量的配置关系直接影响着植物的机械力学和水力结构特征, 从而决定了植物的生活史策略, 并可能影响物种生态位的利用(Givnish, 1987 ; Enquist et al ., 1999 ; West et al ., 1999 ; Westoby et al ., 2002 ).植物单位大小枝条上支撑的叶面积必须与支撑枝的横截面积成一定的比例, 才能符合植物器官间的机械力学和水力结构需求(Niklas, 1992 ; Enquist et al ., 1999 ; Normand et al ., 2008 ; Olson et al ., 2009 ).叶片大小和叶片数量是决定树冠紧密程度的两个重要因素, 直接影响到植株的冠层结构和发育模式, 进而影响到植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ).在某一给定的叶生物量分配或某一个给定的总叶面积下, 植物可能有较少的大叶子或者较多的小叶子(杨冬梅等, 2012a ).坡向作为一种重要的地形要素, 往往对局部区域的太阳辐射和降水进行二次分配(Cantón et al ., 2004 ), 在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 能够对植物的形态产生影响(Bazzaz, 1996 ; 韦莉莉等, 2005 ).当自然资源的丰富度低于或超过植物光合系统所能利用的最低或最高限度时, 植物的光合作用将被抑制(王玉萍等, 2013 ).为提高植物构型构建的生境适应性和风险规避能力, 植物通过功能性状等表型的可塑性调整提高自身的生存适合度, 体现了全球气候变化背景下植物的资源配置策略.因此, 研究坡向梯度上植物枝叶关系规律及其成因, 对于理解植物的环境适应机制有着重要意义. ...
... 植物在长期的生长发育过程中, 各构件在异质生境条件下会表现出不同的生态对策(Yang et al ., 2008 ), 植物外部形态具有高度的可塑性特征, 使其能够把资源最佳地分配到不同的器官, 以规避环境风险.本研究发现, 在各坡向间, 刺槐茎截面积和总叶面积均维持显著的正相关关系(p < 0.05), 回归方程间存在共同斜率且异速生长斜率大于1, 此结果与其他枝叶大小关系研究的结果基本一致(Westoby & Wright, 2003 ; Normand et al ., 2008 ; 史青茹等, 2014 ), 出叶强度和单叶面积则表现出显著的等速负相关关系(p < 0.05).两组关系的回归方程截距均为北坡最大、南坡最小, 这主要与坡向引起环境条件的变化和刺槐枝叶形态在异质生境中的适应机制有关. ...
... 植物功能性状与生存环境之间是一种相互协同进化与适应的关系(侯兆疆等, 2014 ).枝叶大小关系是植物的重要生活史策略(Westoby et al ., 2002 ), 其比例不仅反映枝叶生物量的分配和叶片在生化和水力生物学方面的功能需求(Brouat et al ., 1998 ; Sun et al ., 2006 ; Normand et al ., 2008 ), 也体现了植物适应特定环境的构型塑造(Enquist & Niklas, 2001 , 2002 ).各坡向上刺槐茎截面积和总叶面积均呈正的异速生长关系, 回归方程间具有共同的SMA斜率且显著大于1 (p < 0.05)(图1 ), 二者的相关关系始终符合Corner法则(Corner, 1949 ), 不随微地形特征的变化而显著改变, 植物对总叶面积的投入显著高于对枝横截面积的投入, 这说明微地形差异无法改变植物在枝叶大小上的协调, 枝大小和叶大小间始终需要维持几何大小和重力的平衡(史青茹等, 2014 ).对于此现象, Westoby等(2002) 曾从枝叶的形态和生理关联性方面进行了解释, 他们认为: 一定大小的叶面积必须要由一定大小的枝来支撑, 两者间必须保持大小的匹配, 满足机械力学和水分动力学的要求, 才能保持机械支撑的平衡以及水分运输和蒸腾的协调; 管道模型理论(Niklas, 1992 )则提出, 为满足叶片对水分的需求, 木质部的横截面积与它们所支持的叶面积成正比.除此之外, West等(1997) 提出了结构分形自相似假说解释这种现象, 即: 所有生物的各器官间具有自相似的分形网络结构, 自然选择的压力必然使其向最优化的方向进化, 使得与外界环境的交换面积和新陈代谢效率最大化, 并使营养物质运输距离和时间最小化, 因此, 枝叶的大小必然按一定的比例来协调.而茎截面积-总叶面积关系的SMA斜率显著大于1 (p < 0.05), 主要是因为叶片是植株碳获取的主要功能构件, 所以在进化过程中为提高光合收益, 植株将尽可能地使分配到叶片中的生物量最大化, 而在叶片支撑结构中的投资最小化. ...
浙江天童木本植物Corner法则的检验: 微地形的影响
3
2014
... 植物功能性状与环境和生态系统功能的关系是近年来生态学和全球变化研究的热点(Harrison et al ., 2010 ; Bernard-Verdier et al ., 2012 ), 各功能性状之间的权衡和组合决定了植物的生活史对策(Vandermeer, 1972 ; Silvertown, 2004 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ), 叶片是植物进行光合作用的主要器官(张林和罗天祥, 2004 ), 两者大小和数量的配置关系直接影响着植物的机械力学和水力结构特征, 从而决定了植物的生活史策略, 并可能影响物种生态位的利用(Givnish, 1987 ; Enquist et al ., 1999 ; West et al ., 1999 ; Westoby et al ., 2002 ).植物单位大小枝条上支撑的叶面积必须与支撑枝的横截面积成一定的比例, 才能符合植物器官间的机械力学和水力结构需求(Niklas, 1992 ; Enquist et al ., 1999 ; Normand et al ., 2008 ; Olson et al ., 2009 ).叶片大小和叶片数量是决定树冠紧密程度的两个重要因素, 直接影响到植株的冠层结构和发育模式, 进而影响到植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ).在某一给定的叶生物量分配或某一个给定的总叶面积下, 植物可能有较少的大叶子或者较多的小叶子(杨冬梅等, 2012a ).坡向作为一种重要的地形要素, 往往对局部区域的太阳辐射和降水进行二次分配(Cantón et al ., 2004 ), 在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 能够对植物的形态产生影响(Bazzaz, 1996 ; 韦莉莉等, 2005 ).当自然资源的丰富度低于或超过植物光合系统所能利用的最低或最高限度时, 植物的光合作用将被抑制(王玉萍等, 2013 ).为提高植物构型构建的生境适应性和风险规避能力, 植物通过功能性状等表型的可塑性调整提高自身的生存适合度, 体现了全球气候变化背景下植物的资源配置策略.因此, 研究坡向梯度上植物枝叶关系规律及其成因, 对于理解植物的环境适应机制有着重要意义. ...
... 植物在长期的生长发育过程中, 各构件在异质生境条件下会表现出不同的生态对策(Yang et al ., 2008 ), 植物外部形态具有高度的可塑性特征, 使其能够把资源最佳地分配到不同的器官, 以规避环境风险.本研究发现, 在各坡向间, 刺槐茎截面积和总叶面积均维持显著的正相关关系(p < 0.05), 回归方程间存在共同斜率且异速生长斜率大于1, 此结果与其他枝叶大小关系研究的结果基本一致(Westoby & Wright, 2003 ; Normand et al ., 2008 ; 史青茹等, 2014 ), 出叶强度和单叶面积则表现出显著的等速负相关关系(p < 0.05).两组关系的回归方程截距均为北坡最大、南坡最小, 这主要与坡向引起环境条件的变化和刺槐枝叶形态在异质生境中的适应机制有关. ...
... 植物功能性状与生存环境之间是一种相互协同进化与适应的关系(侯兆疆等, 2014 ).枝叶大小关系是植物的重要生活史策略(Westoby et al ., 2002 ), 其比例不仅反映枝叶生物量的分配和叶片在生化和水力生物学方面的功能需求(Brouat et al ., 1998 ; Sun et al ., 2006 ; Normand et al ., 2008 ), 也体现了植物适应特定环境的构型塑造(Enquist & Niklas, 2001 , 2002 ).各坡向上刺槐茎截面积和总叶面积均呈正的异速生长关系, 回归方程间具有共同的SMA斜率且显著大于1 (p < 0.05)(图1 ), 二者的相关关系始终符合Corner法则(Corner, 1949 ), 不随微地形特征的变化而显著改变, 植物对总叶面积的投入显著高于对枝横截面积的投入, 这说明微地形差异无法改变植物在枝叶大小上的协调, 枝大小和叶大小间始终需要维持几何大小和重力的平衡(史青茹等, 2014 ).对于此现象, Westoby等(2002) 曾从枝叶的形态和生理关联性方面进行了解释, 他们认为: 一定大小的叶面积必须要由一定大小的枝来支撑, 两者间必须保持大小的匹配, 满足机械力学和水分动力学的要求, 才能保持机械支撑的平衡以及水分运输和蒸腾的协调; 管道模型理论(Niklas, 1992 )则提出, 为满足叶片对水分的需求, 木质部的横截面积与它们所支持的叶面积成正比.除此之外, West等(1997) 提出了结构分形自相似假说解释这种现象, 即: 所有生物的各器官间具有自相似的分形网络结构, 自然选择的压力必然使其向最优化的方向进化, 使得与外界环境的交换面积和新陈代谢效率最大化, 并使营养物质运输距离和时间最小化, 因此, 枝叶的大小必然按一定的比例来协调.而茎截面积-总叶面积关系的SMA斜率显著大于1 (p < 0.05), 主要是因为叶片是植株碳获取的主要功能构件, 所以在进化过程中为提高光合收益, 植株将尽可能地使分配到叶片中的生物量最大化, 而在叶片支撑结构中的投资最小化. ...
Plant coexistence and the niche.
1
2004
... 植物功能性状与生存环境之间是一种相互协同进化与适应的关系(侯兆疆等, 2014 ).枝叶大小关系是植物的重要生活史策略(Westoby et al ., 2002 ), 其比例不仅反映枝叶生物量的分配和叶片在生化和水力生物学方面的功能需求(Brouat et al ., 1998 ; Sun et al ., 2006 ; Normand et al ., 2008 ), 也体现了植物适应特定环境的构型塑造(Enquist & Niklas, 2001 , 2002 ).各坡向上刺槐茎截面积和总叶面积均呈正的异速生长关系, 回归方程间具有共同的SMA斜率且显著大于1 (p < 0.05)(图1 ), 二者的相关关系始终符合Corner法则(Corner, 1949 ), 不随微地形特征的变化而显著改变, 植物对总叶面积的投入显著高于对枝横截面积的投入, 这说明微地形差异无法改变植物在枝叶大小上的协调, 枝大小和叶大小间始终需要维持几何大小和重力的平衡(史青茹等, 2014 ).对于此现象, Westoby等(2002) 曾从枝叶的形态和生理关联性方面进行了解释, 他们认为: 一定大小的叶面积必须要由一定大小的枝来支撑, 两者间必须保持大小的匹配, 满足机械力学和水分动力学的要求, 才能保持机械支撑的平衡以及水分运输和蒸腾的协调; 管道模型理论(Niklas, 1992 )则提出, 为满足叶片对水分的需求, 木质部的横截面积与它们所支持的叶面积成正比.除此之外, West等(1997) 提出了结构分形自相似假说解释这种现象, 即: 所有生物的各器官间具有自相似的分形网络结构, 自然选择的压力必然使其向最优化的方向进化, 使得与外界环境的交换面积和新陈代谢效率最大化, 并使营养物质运输距离和时间最小化, 因此, 枝叶的大小必然按一定的比例来协调.而茎截面积-总叶面积关系的SMA斜率显著大于1 (p < 0.05), 主要是因为叶片是植株碳获取的主要功能构件, 所以在进化过程中为提高光合收益, 植株将尽可能地使分配到叶片中的生物量最大化, 而在叶片支撑结构中的投资最小化. ...
The leaf size-twig size spectrum of temperate woody species along an altitudinal gradient: An invariant allometric scaling relationship.
0
2006
1
2010
... 植物功能性状与环境和生态系统功能的关系是近年来生态学和全球变化研究的热点(Harrison et al ., 2010 ; Bernard-Verdier et al ., 2012 ), 各功能性状之间的权衡和组合决定了植物的生活史对策(Vandermeer, 1972 ; Silvertown, 2004 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ), 叶片是植物进行光合作用的主要器官(张林和罗天祥, 2004 ), 两者大小和数量的配置关系直接影响着植物的机械力学和水力结构特征, 从而决定了植物的生活史策略, 并可能影响物种生态位的利用(Givnish, 1987 ; Enquist et al ., 1999 ; West et al ., 1999 ; Westoby et al ., 2002 ).植物单位大小枝条上支撑的叶面积必须与支撑枝的横截面积成一定的比例, 才能符合植物器官间的机械力学和水力结构需求(Niklas, 1992 ; Enquist et al ., 1999 ; Normand et al ., 2008 ; Olson et al ., 2009 ).叶片大小和叶片数量是决定树冠紧密程度的两个重要因素, 直接影响到植株的冠层结构和发育模式, 进而影响到植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ).在某一给定的叶生物量分配或某一个给定的总叶面积下, 植物可能有较少的大叶子或者较多的小叶子(杨冬梅等, 2012a ).坡向作为一种重要的地形要素, 往往对局部区域的太阳辐射和降水进行二次分配(Cantón et al ., 2004 ), 在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 能够对植物的形态产生影响(Bazzaz, 1996 ; 韦莉莉等, 2005 ).当自然资源的丰富度低于或超过植物光合系统所能利用的最低或最高限度时, 植物的光合作用将被抑制(王玉萍等, 2013 ).为提高植物构型构建的生境适应性和风险规避能力, 植物通过功能性状等表型的可塑性调整提高自身的生存适合度, 体现了全球气候变化背景下植物的资源配置策略.因此, 研究坡向梯度上植物枝叶关系规律及其成因, 对于理解植物的环境适应机制有着重要意义. ...
1
2010
... 植物功能性状与环境和生态系统功能的关系是近年来生态学和全球变化研究的热点(Harrison et al ., 2010 ; Bernard-Verdier et al ., 2012 ), 各功能性状之间的权衡和组合决定了植物的生活史对策(Vandermeer, 1972 ; Silvertown, 2004 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ), 叶片是植物进行光合作用的主要器官(张林和罗天祥, 2004 ), 两者大小和数量的配置关系直接影响着植物的机械力学和水力结构特征, 从而决定了植物的生活史策略, 并可能影响物种生态位的利用(Givnish, 1987 ; Enquist et al ., 1999 ; West et al ., 1999 ; Westoby et al ., 2002 ).植物单位大小枝条上支撑的叶面积必须与支撑枝的横截面积成一定的比例, 才能符合植物器官间的机械力学和水力结构需求(Niklas, 1992 ; Enquist et al ., 1999 ; Normand et al ., 2008 ; Olson et al ., 2009 ).叶片大小和叶片数量是决定树冠紧密程度的两个重要因素, 直接影响到植株的冠层结构和发育模式, 进而影响到植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ).在某一给定的叶生物量分配或某一个给定的总叶面积下, 植物可能有较少的大叶子或者较多的小叶子(杨冬梅等, 2012a ).坡向作为一种重要的地形要素, 往往对局部区域的太阳辐射和降水进行二次分配(Cantón et al ., 2004 ), 在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 能够对植物的形态产生影响(Bazzaz, 1996 ; 韦莉莉等, 2005 ).当自然资源的丰富度低于或超过植物光合系统所能利用的最低或最高限度时, 植物的光合作用将被抑制(王玉萍等, 2013 ).为提高植物构型构建的生境适应性和风险规避能力, 植物通过功能性状等表型的可塑性调整提高自身的生存适合度, 体现了全球气候变化背景下植物的资源配置策略.因此, 研究坡向梯度上植物枝叶关系规律及其成因, 对于理解植物的环境适应机制有着重要意义. ...
干旱和坡向互作对栓皮栎和侧柏生长的影响
1
2013
... 刺槐(Robinia pseudoacacia )是豆科刺槐属的温带落叶乔木, 具有生长速度快、根系发达、对环境适应性强、固碳氮能力较好、成活率高、萌芽力和根蘖性都很强等优点, 已成为我国黄土丘陵区的主要造林树种和人工林面积最大的树种(于占辉等, 2009 ; 李军等, 2010 ).近年来, 干旱地区许多刺槐人工林出现了生长减慢、枝梢干枯甚至整株死亡的衰败现象, 引起了学术界的广泛关注(单长卷等, 2004 ; 李军等, 2010 ).目前, 许多学者研究了刺槐叶面积生长与林分密度的关系(尹婧等, 2008 ), 土层厚度对刺槐旱季水分状况和生长的影响(王林等, 2013 ), 不同林龄和退耕年限刺槐人工林碳、氮储量及分配规律(申家朋和张文辉, 2014 ; 艾泽民等, 2014 ), 揭示了刺槐生长以及生物量积累和分配格局等一系列规律, 但关于微地形影响下刺槐当年生小枝内枝叶关系的研究较为少见, 更鲜有不同坡向茎截面积与总叶面积、出叶强度与单叶面积关系的报道, 限制了对异质生境中刺槐适应机制的解析.本研究选择兰州北山九州台西北师范大学绿化基地的刺槐人工林为研究对象, 探讨刺槐当年生小枝内的茎截面积-总叶面积、出叶强度-单叶面积关系, 为进一步了解刺槐的环境适应机制提供理论依据, 对提高干旱半干旱地区人工林的可持续性和稳定性具有重要意义. ...
干旱和坡向互作对栓皮栎和侧柏生长的影响
1
2013
... 刺槐(Robinia pseudoacacia )是豆科刺槐属的温带落叶乔木, 具有生长速度快、根系发达、对环境适应性强、固碳氮能力较好、成活率高、萌芽力和根蘖性都很强等优点, 已成为我国黄土丘陵区的主要造林树种和人工林面积最大的树种(于占辉等, 2009 ; 李军等, 2010 ).近年来, 干旱地区许多刺槐人工林出现了生长减慢、枝梢干枯甚至整株死亡的衰败现象, 引起了学术界的广泛关注(单长卷等, 2004 ; 李军等, 2010 ).目前, 许多学者研究了刺槐叶面积生长与林分密度的关系(尹婧等, 2008 ), 土层厚度对刺槐旱季水分状况和生长的影响(王林等, 2013 ), 不同林龄和退耕年限刺槐人工林碳、氮储量及分配规律(申家朋和张文辉, 2014 ; 艾泽民等, 2014 ), 揭示了刺槐生长以及生物量积累和分配格局等一系列规律, 但关于微地形影响下刺槐当年生小枝内枝叶关系的研究较为少见, 更鲜有不同坡向茎截面积与总叶面积、出叶强度与单叶面积关系的报道, 限制了对异质生境中刺槐适应机制的解析.本研究选择兰州北山九州台西北师范大学绿化基地的刺槐人工林为研究对象, 探讨刺槐当年生小枝内的茎截面积-总叶面积、出叶强度-单叶面积关系, 为进一步了解刺槐的环境适应机制提供理论依据, 对提高干旱半干旱地区人工林的可持续性和稳定性具有重要意义. ...
高山植物光合机构耐受胁迫的适应机制
1
2013
... 植物功能性状与环境和生态系统功能的关系是近年来生态学和全球变化研究的热点(Harrison et al ., 2010 ; Bernard-Verdier et al ., 2012 ), 各功能性状之间的权衡和组合决定了植物的生活史对策(Vandermeer, 1972 ; Silvertown, 2004 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ), 叶片是植物进行光合作用的主要器官(张林和罗天祥, 2004 ), 两者大小和数量的配置关系直接影响着植物的机械力学和水力结构特征, 从而决定了植物的生活史策略, 并可能影响物种生态位的利用(Givnish, 1987 ; Enquist et al ., 1999 ; West et al ., 1999 ; Westoby et al ., 2002 ).植物单位大小枝条上支撑的叶面积必须与支撑枝的横截面积成一定的比例, 才能符合植物器官间的机械力学和水力结构需求(Niklas, 1992 ; Enquist et al ., 1999 ; Normand et al ., 2008 ; Olson et al ., 2009 ).叶片大小和叶片数量是决定树冠紧密程度的两个重要因素, 直接影响到植株的冠层结构和发育模式, 进而影响到植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ).在某一给定的叶生物量分配或某一个给定的总叶面积下, 植物可能有较少的大叶子或者较多的小叶子(杨冬梅等, 2012a ).坡向作为一种重要的地形要素, 往往对局部区域的太阳辐射和降水进行二次分配(Cantón et al ., 2004 ), 在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 能够对植物的形态产生影响(Bazzaz, 1996 ; 韦莉莉等, 2005 ).当自然资源的丰富度低于或超过植物光合系统所能利用的最低或最高限度时, 植物的光合作用将被抑制(王玉萍等, 2013 ).为提高植物构型构建的生境适应性和风险规避能力, 植物通过功能性状等表型的可塑性调整提高自身的生存适合度, 体现了全球气候变化背景下植物的资源配置策略.因此, 研究坡向梯度上植物枝叶关系规律及其成因, 对于理解植物的环境适应机制有着重要意义. ...
高山植物光合机构耐受胁迫的适应机制
1
2013
... 植物功能性状与环境和生态系统功能的关系是近年来生态学和全球变化研究的热点(Harrison et al ., 2010 ; Bernard-Verdier et al ., 2012 ), 各功能性状之间的权衡和组合决定了植物的生活史对策(Vandermeer, 1972 ; Silvertown, 2004 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ), 叶片是植物进行光合作用的主要器官(张林和罗天祥, 2004 ), 两者大小和数量的配置关系直接影响着植物的机械力学和水力结构特征, 从而决定了植物的生活史策略, 并可能影响物种生态位的利用(Givnish, 1987 ; Enquist et al ., 1999 ; West et al ., 1999 ; Westoby et al ., 2002 ).植物单位大小枝条上支撑的叶面积必须与支撑枝的横截面积成一定的比例, 才能符合植物器官间的机械力学和水力结构需求(Niklas, 1992 ; Enquist et al ., 1999 ; Normand et al ., 2008 ; Olson et al ., 2009 ).叶片大小和叶片数量是决定树冠紧密程度的两个重要因素, 直接影响到植株的冠层结构和发育模式, 进而影响到植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ).在某一给定的叶生物量分配或某一个给定的总叶面积下, 植物可能有较少的大叶子或者较多的小叶子(杨冬梅等, 2012a ).坡向作为一种重要的地形要素, 往往对局部区域的太阳辐射和降水进行二次分配(Cantón et al ., 2004 ), 在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 能够对植物的形态产生影响(Bazzaz, 1996 ; 韦莉莉等, 2005 ).当自然资源的丰富度低于或超过植物光合系统所能利用的最低或最高限度时, 植物的光合作用将被抑制(王玉萍等, 2013 ).为提高植物构型构建的生境适应性和风险规避能力, 植物通过功能性状等表型的可塑性调整提高自身的生存适合度, 体现了全球气候变化背景下植物的资源配置策略.因此, 研究坡向梯度上植物枝叶关系规律及其成因, 对于理解植物的环境适应机制有着重要意义. ...
Common slope tests for bivariate errors-in-variables models.
2
2002
... 将坡向划分为北坡(315°-360°和0°-45°)、东坡(45°-135°)、南坡(135°-225°)和西坡(225°-315°).在所研究的4个功能性状中, 茎截面积指当年生小枝上1级枝的横截面积, 总叶面积指当年生小枝上所支撑的所有叶片面积之和, 出叶强度指单位体积枝条所支撑的叶片数, 单叶面积指当年生小枝上所支撑的所有叶片单叶面积的平均值, 枝条体积由其横截面积乘以长度获得(许月等, 2012 ).对每个样方刺槐枝叶属性的平均值进行对数(以10为底)转换后再进行分析, 对数转换使之符合正态分布.对枝大小-叶大小以及叶数量-叶大小两组功能性状关系的研究, 均采用y = axb , 线性转换成log(y ) = log(a ) + b log(x ), 式中x 和y 表示两个特征参数, a 为性状关系的截距, b 为斜率, 即异速生长参数或相对生长的指数, 当|b | = 1时, 表示两者是等速变化关系; 当|b| 显著偏离1时, 两者间为异速变化关系(Harvey & Pagel, 1991 ).数据分析主要采用标准化主轴估计(standardized major axis estimation, SMA)的方法(Warton et al ., 2006 ), 由软件(S)MATR Version 2.0 (Falster et al ., 2006 )计算完成.每一个回归斜率的置信区间根据Pitman (1939)方法计算.回归斜率与1或-1的显著性检验根据Warton和Weber (2002)的方法判断.在完成线性回归后, 对回归斜率进行异质性测试, 如果组间没有异质性, 则表示有共同斜率(Warton & Weber, 2002 ).此后, 不同坡向梯度间的功能性状的平均值的比较采用单因素方差分析(one-way ANOVA), 利用成对比较检验分析各坡向刺槐枝叶关系线性回归方程在y 轴上截距的差异. ...
... )计算完成.每一个回归斜率的置信区间根据Pitman (1939)方法计算.回归斜率与1或-1的显著性检验根据Warton和Weber (2002)的方法判断.在完成线性回归后, 对回归斜率进行异质性测试, 如果组间没有异质性, 则表示有共同斜率(Warton & Weber, 2002 ).此后, 不同坡向梯度间的功能性状的平均值的比较采用单因素方差分析(one-way ANOVA), 利用成对比较检验分析各坡向刺槐枝叶关系线性回归方程在y 轴上截距的差异. ...
Bivariate line fitting methods for allometry.
0
2006
杉木苗木光合作用及其产物分配对水分胁迫的响应
1
2005
... 植物功能性状与环境和生态系统功能的关系是近年来生态学和全球变化研究的热点(Harrison et al ., 2010 ; Bernard-Verdier et al ., 2012 ), 各功能性状之间的权衡和组合决定了植物的生活史对策(Vandermeer, 1972 ; Silvertown, 2004 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ), 叶片是植物进行光合作用的主要器官(张林和罗天祥, 2004 ), 两者大小和数量的配置关系直接影响着植物的机械力学和水力结构特征, 从而决定了植物的生活史策略, 并可能影响物种生态位的利用(Givnish, 1987 ; Enquist et al ., 1999 ; West et al ., 1999 ; Westoby et al ., 2002 ).植物单位大小枝条上支撑的叶面积必须与支撑枝的横截面积成一定的比例, 才能符合植物器官间的机械力学和水力结构需求(Niklas, 1992 ; Enquist et al ., 1999 ; Normand et al ., 2008 ; Olson et al ., 2009 ).叶片大小和叶片数量是决定树冠紧密程度的两个重要因素, 直接影响到植株的冠层结构和发育模式, 进而影响到植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ).在某一给定的叶生物量分配或某一个给定的总叶面积下, 植物可能有较少的大叶子或者较多的小叶子(杨冬梅等, 2012a ).坡向作为一种重要的地形要素, 往往对局部区域的太阳辐射和降水进行二次分配(Cantón et al ., 2004 ), 在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 能够对植物的形态产生影响(Bazzaz, 1996 ; 韦莉莉等, 2005 ).当自然资源的丰富度低于或超过植物光合系统所能利用的最低或最高限度时, 植物的光合作用将被抑制(王玉萍等, 2013 ).为提高植物构型构建的生境适应性和风险规避能力, 植物通过功能性状等表型的可塑性调整提高自身的生存适合度, 体现了全球气候变化背景下植物的资源配置策略.因此, 研究坡向梯度上植物枝叶关系规律及其成因, 对于理解植物的环境适应机制有着重要意义. ...
杉木苗木光合作用及其产物分配对水分胁迫的响应
1
2005
... 植物功能性状与环境和生态系统功能的关系是近年来生态学和全球变化研究的热点(Harrison et al ., 2010 ; Bernard-Verdier et al ., 2012 ), 各功能性状之间的权衡和组合决定了植物的生活史对策(Vandermeer, 1972 ; Silvertown, 2004 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ), 叶片是植物进行光合作用的主要器官(张林和罗天祥, 2004 ), 两者大小和数量的配置关系直接影响着植物的机械力学和水力结构特征, 从而决定了植物的生活史策略, 并可能影响物种生态位的利用(Givnish, 1987 ; Enquist et al ., 1999 ; West et al ., 1999 ; Westoby et al ., 2002 ).植物单位大小枝条上支撑的叶面积必须与支撑枝的横截面积成一定的比例, 才能符合植物器官间的机械力学和水力结构需求(Niklas, 1992 ; Enquist et al ., 1999 ; Normand et al ., 2008 ; Olson et al ., 2009 ).叶片大小和叶片数量是决定树冠紧密程度的两个重要因素, 直接影响到植株的冠层结构和发育模式, 进而影响到植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ).在某一给定的叶生物量分配或某一个给定的总叶面积下, 植物可能有较少的大叶子或者较多的小叶子(杨冬梅等, 2012a ).坡向作为一种重要的地形要素, 往往对局部区域的太阳辐射和降水进行二次分配(Cantón et al ., 2004 ), 在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 能够对植物的形态产生影响(Bazzaz, 1996 ; 韦莉莉等, 2005 ).当自然资源的丰富度低于或超过植物光合系统所能利用的最低或最高限度时, 植物的光合作用将被抑制(王玉萍等, 2013 ).为提高植物构型构建的生境适应性和风险规避能力, 植物通过功能性状等表型的可塑性调整提高自身的生存适合度, 体现了全球气候变化背景下植物的资源配置策略.因此, 研究坡向梯度上植物枝叶关系规律及其成因, 对于理解植物的环境适应机制有着重要意义. ...
A general model for the origin of allometric scaling laws in biology.
1
1997
... 植物功能性状与环境和生态系统功能的关系是近年来生态学和全球变化研究的热点(Harrison et al ., 2010 ; Bernard-Verdier et al ., 2012 ), 各功能性状之间的权衡和组合决定了植物的生活史对策(Vandermeer, 1972 ; Silvertown, 2004 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ), 叶片是植物进行光合作用的主要器官(张林和罗天祥, 2004 ), 两者大小和数量的配置关系直接影响着植物的机械力学和水力结构特征, 从而决定了植物的生活史策略, 并可能影响物种生态位的利用(Givnish, 1987 ; Enquist et al ., 1999 ; West et al ., 1999 ; Westoby et al ., 2002 ).植物单位大小枝条上支撑的叶面积必须与支撑枝的横截面积成一定的比例, 才能符合植物器官间的机械力学和水力结构需求(Niklas, 1992 ; Enquist et al ., 1999 ; Normand et al ., 2008 ; Olson et al ., 2009 ).叶片大小和叶片数量是决定树冠紧密程度的两个重要因素, 直接影响到植株的冠层结构和发育模式, 进而影响到植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ).在某一给定的叶生物量分配或某一个给定的总叶面积下, 植物可能有较少的大叶子或者较多的小叶子(杨冬梅等, 2012a ).坡向作为一种重要的地形要素, 往往对局部区域的太阳辐射和降水进行二次分配(Cantón et al ., 2004 ), 在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 能够对植物的形态产生影响(Bazzaz, 1996 ; 韦莉莉等, 2005 ).当自然资源的丰富度低于或超过植物光合系统所能利用的最低或最高限度时, 植物的光合作用将被抑制(王玉萍等, 2013 ).为提高植物构型构建的生境适应性和风险规避能力, 植物通过功能性状等表型的可塑性调整提高自身的生存适合度, 体现了全球气候变化背景下植物的资源配置策略.因此, 研究坡向梯度上植物枝叶关系规律及其成因, 对于理解植物的环境适应机制有着重要意义. ...
A general model for the structure and allometry of plant vascular systems.
2
1999
... 植物功能性状与环境和生态系统功能的关系是近年来生态学和全球变化研究的热点(Harrison et al ., 2010 ; Bernard-Verdier et al ., 2012 ), 各功能性状之间的权衡和组合决定了植物的生活史对策(Vandermeer, 1972 ; Silvertown, 2004 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ), 叶片是植物进行光合作用的主要器官(张林和罗天祥, 2004 ), 两者大小和数量的配置关系直接影响着植物的机械力学和水力结构特征, 从而决定了植物的生活史策略, 并可能影响物种生态位的利用(Givnish, 1987 ; Enquist et al ., 1999 ; West et al ., 1999 ; Westoby et al ., 2002 ).植物单位大小枝条上支撑的叶面积必须与支撑枝的横截面积成一定的比例, 才能符合植物器官间的机械力学和水力结构需求(Niklas, 1992 ; Enquist et al ., 1999 ; Normand et al ., 2008 ; Olson et al ., 2009 ).叶片大小和叶片数量是决定树冠紧密程度的两个重要因素, 直接影响到植株的冠层结构和发育模式, 进而影响到植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ).在某一给定的叶生物量分配或某一个给定的总叶面积下, 植物可能有较少的大叶子或者较多的小叶子(杨冬梅等, 2012a ).坡向作为一种重要的地形要素, 往往对局部区域的太阳辐射和降水进行二次分配(Cantón et al ., 2004 ), 在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 能够对植物的形态产生影响(Bazzaz, 1996 ; 韦莉莉等, 2005 ).当自然资源的丰富度低于或超过植物光合系统所能利用的最低或最高限度时, 植物的光合作用将被抑制(王玉萍等, 2013 ).为提高植物构型构建的生境适应性和风险规避能力, 植物通过功能性状等表型的可塑性调整提高自身的生存适合度, 体现了全球气候变化背景下植物的资源配置策略.因此, 研究坡向梯度上植物枝叶关系规律及其成因, 对于理解植物的环境适应机制有着重要意义. ...
... 植物功能性状与生存环境之间是一种相互协同进化与适应的关系(侯兆疆等, 2014 ).枝叶大小关系是植物的重要生活史策略(Westoby et al ., 2002 ), 其比例不仅反映枝叶生物量的分配和叶片在生化和水力生物学方面的功能需求(Brouat et al ., 1998 ; Sun et al ., 2006 ; Normand et al ., 2008 ), 也体现了植物适应特定环境的构型塑造(Enquist & Niklas, 2001 , 2002 ).各坡向上刺槐茎截面积和总叶面积均呈正的异速生长关系, 回归方程间具有共同的SMA斜率且显著大于1 (p < 0.05)(图1 ), 二者的相关关系始终符合Corner法则(Corner, 1949 ), 不随微地形特征的变化而显著改变, 植物对总叶面积的投入显著高于对枝横截面积的投入, 这说明微地形差异无法改变植物在枝叶大小上的协调, 枝大小和叶大小间始终需要维持几何大小和重力的平衡(史青茹等, 2014 ).对于此现象, Westoby等(2002) 曾从枝叶的形态和生理关联性方面进行了解释, 他们认为: 一定大小的叶面积必须要由一定大小的枝来支撑, 两者间必须保持大小的匹配, 满足机械力学和水分动力学的要求, 才能保持机械支撑的平衡以及水分运输和蒸腾的协调; 管道模型理论(Niklas, 1992 )则提出, 为满足叶片对水分的需求, 木质部的横截面积与它们所支持的叶面积成正比.除此之外, West等(1997) 提出了结构分形自相似假说解释这种现象, 即: 所有生物的各器官间具有自相似的分形网络结构, 自然选择的压力必然使其向最优化的方向进化, 使得与外界环境的交换面积和新陈代谢效率最大化, 并使营养物质运输距离和时间最小化, 因此, 枝叶的大小必然按一定的比例来协调.而茎截面积-总叶面积关系的SMA斜率显著大于1 (p < 0.05), 主要是因为叶片是植株碳获取的主要功能构件, 所以在进化过程中为提高光合收益, 植株将尽可能地使分配到叶片中的生物量最大化, 而在叶片支撑结构中的投资最小化. ...
Plant ecological strategies: Some leading dimensions of variation between species.
0
2002
The leaf size-twig size spectrum and its relationship to other important spectra of variation among species.
1
2003
... 植物在长期的生长发育过程中, 各构件在异质生境条件下会表现出不同的生态对策(Yang et al ., 2008 ), 植物外部形态具有高度的可塑性特征, 使其能够把资源最佳地分配到不同的器官, 以规避环境风险.本研究发现, 在各坡向间, 刺槐茎截面积和总叶面积均维持显著的正相关关系(p < 0.05), 回归方程间存在共同斜率且异速生长斜率大于1, 此结果与其他枝叶大小关系研究的结果基本一致(Westoby & Wright, 2003 ; Normand et al ., 2008 ; 史青茹等, 2014 ), 出叶强度和单叶面积则表现出显著的等速负相关关系(p < 0.05).两组关系的回归方程截距均为北坡最大、南坡最小, 这主要与坡向引起环境条件的变化和刺槐枝叶形态在异质生境中的适应机制有关. ...
浙江天童木本植物小枝的“大小-数量”权衡
2
2012
... 将坡向划分为北坡(315°-360°和0°-45°)、东坡(45°-135°)、南坡(135°-225°)和西坡(225°-315°).在所研究的4个功能性状中, 茎截面积指当年生小枝上1级枝的横截面积, 总叶面积指当年生小枝上所支撑的所有叶片面积之和, 出叶强度指单位体积枝条所支撑的叶片数, 单叶面积指当年生小枝上所支撑的所有叶片单叶面积的平均值, 枝条体积由其横截面积乘以长度获得(许月等, 2012 ).对每个样方刺槐枝叶属性的平均值进行对数(以10为底)转换后再进行分析, 对数转换使之符合正态分布.对枝大小-叶大小以及叶数量-叶大小两组功能性状关系的研究, 均采用y = axb , 线性转换成log(y ) = log(a ) + b log(x ), 式中x 和y 表示两个特征参数, a 为性状关系的截距, b 为斜率, 即异速生长参数或相对生长的指数, 当|b | = 1时, 表示两者是等速变化关系; 当|b| 显著偏离1时, 两者间为异速变化关系(Harvey & Pagel, 1991 ).数据分析主要采用标准化主轴估计(standardized major axis estimation, SMA)的方法(Warton et al ., 2006 ), 由软件(S)MATR Version 2.0 (Falster et al ., 2006 )计算完成.每一个回归斜率的置信区间根据Pitman (1939)方法计算.回归斜率与1或-1的显著性检验根据Warton和Weber (2002)的方法判断.在完成线性回归后, 对回归斜率进行异质性测试, 如果组间没有异质性, 则表示有共同斜率(Warton & Weber, 2002 ).此后, 不同坡向梯度间的功能性状的平均值的比较采用单因素方差分析(one-way ANOVA), 利用成对比较检验分析各坡向刺槐枝叶关系线性回归方程在y 轴上截距的差异. ...
... 植物在长期的生长发育过程中, 各构件在异质生境条件下会表现出不同的生态对策(Yang et al ., 2008 ), 植物外部形态具有高度的可塑性特征, 使其能够把资源最佳地分配到不同的器官, 以规避环境风险.本研究发现, 在各坡向间, 刺槐茎截面积和总叶面积均维持显著的正相关关系(p < 0.05), 回归方程间存在共同斜率且异速生长斜率大于1, 此结果与其他枝叶大小关系研究的结果基本一致(Westoby & Wright, 2003 ; Normand et al ., 2008 ; 史青茹等, 2014 ), 出叶强度和单叶面积则表现出显著的等速负相关关系(p < 0.05).两组关系的回归方程截距均为北坡最大、南坡最小, 这主要与坡向引起环境条件的变化和刺槐枝叶形态在异质生境中的适应机制有关. ...
浙江天童木本植物小枝的“大小-数量”权衡
2
2012
... 将坡向划分为北坡(315°-360°和0°-45°)、东坡(45°-135°)、南坡(135°-225°)和西坡(225°-315°).在所研究的4个功能性状中, 茎截面积指当年生小枝上1级枝的横截面积, 总叶面积指当年生小枝上所支撑的所有叶片面积之和, 出叶强度指单位体积枝条所支撑的叶片数, 单叶面积指当年生小枝上所支撑的所有叶片单叶面积的平均值, 枝条体积由其横截面积乘以长度获得(许月等, 2012 ).对每个样方刺槐枝叶属性的平均值进行对数(以10为底)转换后再进行分析, 对数转换使之符合正态分布.对枝大小-叶大小以及叶数量-叶大小两组功能性状关系的研究, 均采用y = axb , 线性转换成log(y ) = log(a ) + b log(x ), 式中x 和y 表示两个特征参数, a 为性状关系的截距, b 为斜率, 即异速生长参数或相对生长的指数, 当|b | = 1时, 表示两者是等速变化关系; 当|b| 显著偏离1时, 两者间为异速变化关系(Harvey & Pagel, 1991 ).数据分析主要采用标准化主轴估计(standardized major axis estimation, SMA)的方法(Warton et al ., 2006 ), 由软件(S)MATR Version 2.0 (Falster et al ., 2006 )计算完成.每一个回归斜率的置信区间根据Pitman (1939)方法计算.回归斜率与1或-1的显著性检验根据Warton和Weber (2002)的方法判断.在完成线性回归后, 对回归斜率进行异质性测试, 如果组间没有异质性, 则表示有共同斜率(Warton & Weber, 2002 ).此后, 不同坡向梯度间的功能性状的平均值的比较采用单因素方差分析(one-way ANOVA), 利用成对比较检验分析各坡向刺槐枝叶关系线性回归方程在y 轴上截距的差异. ...
... 植物在长期的生长发育过程中, 各构件在异质生境条件下会表现出不同的生态对策(Yang et al ., 2008 ), 植物外部形态具有高度的可塑性特征, 使其能够把资源最佳地分配到不同的器官, 以规避环境风险.本研究发现, 在各坡向间, 刺槐茎截面积和总叶面积均维持显著的正相关关系(p < 0.05), 回归方程间存在共同斜率且异速生长斜率大于1, 此结果与其他枝叶大小关系研究的结果基本一致(Westoby & Wright, 2003 ; Normand et al ., 2008 ; 史青茹等, 2014 ), 出叶强度和单叶面积则表现出显著的等速负相关关系(p < 0.05).两组关系的回归方程截距均为北坡最大、南坡最小, 这主要与坡向引起环境条件的变化和刺槐枝叶形态在异质生境中的适应机制有关. ...
The generality of leaf size versus number trade-off in temperate woody species.
0
2008
清凉峰不同海拔木本植物小枝内叶大小-数量权衡关系
2
2012
... 植物功能性状与环境和生态系统功能的关系是近年来生态学和全球变化研究的热点(Harrison et al ., 2010 ; Bernard-Verdier et al ., 2012 ), 各功能性状之间的权衡和组合决定了植物的生活史对策(Vandermeer, 1972 ; Silvertown, 2004 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ), 叶片是植物进行光合作用的主要器官(张林和罗天祥, 2004 ), 两者大小和数量的配置关系直接影响着植物的机械力学和水力结构特征, 从而决定了植物的生活史策略, 并可能影响物种生态位的利用(Givnish, 1987 ; Enquist et al ., 1999 ; West et al ., 1999 ; Westoby et al ., 2002 ).植物单位大小枝条上支撑的叶面积必须与支撑枝的横截面积成一定的比例, 才能符合植物器官间的机械力学和水力结构需求(Niklas, 1992 ; Enquist et al ., 1999 ; Normand et al ., 2008 ; Olson et al ., 2009 ).叶片大小和叶片数量是决定树冠紧密程度的两个重要因素, 直接影响到植株的冠层结构和发育模式, 进而影响到植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ).在某一给定的叶生物量分配或某一个给定的总叶面积下, 植物可能有较少的大叶子或者较多的小叶子(杨冬梅等, 2012a ).坡向作为一种重要的地形要素, 往往对局部区域的太阳辐射和降水进行二次分配(Cantón et al ., 2004 ), 在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 能够对植物的形态产生影响(Bazzaz, 1996 ; 韦莉莉等, 2005 ).当自然资源的丰富度低于或超过植物光合系统所能利用的最低或最高限度时, 植物的光合作用将被抑制(王玉萍等, 2013 ).为提高植物构型构建的生境适应性和风险规避能力, 植物通过功能性状等表型的可塑性调整提高自身的生存适合度, 体现了全球气候变化背景下植物的资源配置策略.因此, 研究坡向梯度上植物枝叶关系规律及其成因, 对于理解植物的环境适应机制有着重要意义. ...
... 除此之外, 叶大小-叶数量权衡关系的坡向差异性还可以从以下两个方面解释: (1)植物内在生物量分配限制的结果.由表2 可知, 北坡(阴坡)刺槐单叶面积大于南坡(阳坡), 在植物体上, 在某一给定的叶生物量或某一给定的总叶面积下, 植物要么是有较少的大叶子, 要么是有较多的小叶子, 增加单个叶片的投资意味着叶片数量的减少(杨冬梅等, 2012a ); (2)单叶面积的差异是出叶强度变化的副产物.每个叶片通常都会有一个腋芽分生组织(Bell, 1993 ), 叶片更多的物种, 单位个体大小的腋芽分生组织的密度将会更高(杨冬梅等, 2012b ), 更大的芽库由于有更多的部位具有再生能力, 既可以生长为植株的营养结构或繁殖结构, 也可以保持无活性状态, 进而影响到植株的大小构建和繁殖产出(Bonser & Aarssen, 1996 ), 使植株具有更大的可塑性和更新复壮能力, 因此在南坡干、热、高光和低养分的环境下, 刺槐倾向于具有更高的出叶强度, 叶大小与出叶强度紧密相关, 因此小叶片可能是选择了高的出叶强度的结果.东坡和西坡枝叶功能性状及其关系之间没有明显差异, 主要是因为东坡和西坡属于半阴坡和半阳坡, 皆处于阴坡和阳坡之间的过渡区域, 群落组成也较为复杂, 因此刺槐采取相同的生长策略来适应生境的变化. ...
清凉峰不同海拔木本植物小枝内叶大小-数量权衡关系
2
2012
... 植物功能性状与环境和生态系统功能的关系是近年来生态学和全球变化研究的热点(Harrison et al ., 2010 ; Bernard-Verdier et al ., 2012 ), 各功能性状之间的权衡和组合决定了植物的生活史对策(Vandermeer, 1972 ; Silvertown, 2004 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ), 叶片是植物进行光合作用的主要器官(张林和罗天祥, 2004 ), 两者大小和数量的配置关系直接影响着植物的机械力学和水力结构特征, 从而决定了植物的生活史策略, 并可能影响物种生态位的利用(Givnish, 1987 ; Enquist et al ., 1999 ; West et al ., 1999 ; Westoby et al ., 2002 ).植物单位大小枝条上支撑的叶面积必须与支撑枝的横截面积成一定的比例, 才能符合植物器官间的机械力学和水力结构需求(Niklas, 1992 ; Enquist et al ., 1999 ; Normand et al ., 2008 ; Olson et al ., 2009 ).叶片大小和叶片数量是决定树冠紧密程度的两个重要因素, 直接影响到植株的冠层结构和发育模式, 进而影响到植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ).在某一给定的叶生物量分配或某一个给定的总叶面积下, 植物可能有较少的大叶子或者较多的小叶子(杨冬梅等, 2012a ).坡向作为一种重要的地形要素, 往往对局部区域的太阳辐射和降水进行二次分配(Cantón et al ., 2004 ), 在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 能够对植物的形态产生影响(Bazzaz, 1996 ; 韦莉莉等, 2005 ).当自然资源的丰富度低于或超过植物光合系统所能利用的最低或最高限度时, 植物的光合作用将被抑制(王玉萍等, 2013 ).为提高植物构型构建的生境适应性和风险规避能力, 植物通过功能性状等表型的可塑性调整提高自身的生存适合度, 体现了全球气候变化背景下植物的资源配置策略.因此, 研究坡向梯度上植物枝叶关系规律及其成因, 对于理解植物的环境适应机制有着重要意义. ...
... 除此之外, 叶大小-叶数量权衡关系的坡向差异性还可以从以下两个方面解释: (1)植物内在生物量分配限制的结果.由表2 可知, 北坡(阴坡)刺槐单叶面积大于南坡(阳坡), 在植物体上, 在某一给定的叶生物量或某一给定的总叶面积下, 植物要么是有较少的大叶子, 要么是有较多的小叶子, 增加单个叶片的投资意味着叶片数量的减少(杨冬梅等, 2012a ); (2)单叶面积的差异是出叶强度变化的副产物.每个叶片通常都会有一个腋芽分生组织(Bell, 1993 ), 叶片更多的物种, 单位个体大小的腋芽分生组织的密度将会更高(杨冬梅等, 2012b ), 更大的芽库由于有更多的部位具有再生能力, 既可以生长为植株的营养结构或繁殖结构, 也可以保持无活性状态, 进而影响到植株的大小构建和繁殖产出(Bonser & Aarssen, 1996 ), 使植株具有更大的可塑性和更新复壮能力, 因此在南坡干、热、高光和低养分的环境下, 刺槐倾向于具有更高的出叶强度, 叶大小与出叶强度紧密相关, 因此小叶片可能是选择了高的出叶强度的结果.东坡和西坡枝叶功能性状及其关系之间没有明显差异, 主要是因为东坡和西坡属于半阴坡和半阳坡, 皆处于阴坡和阳坡之间的过渡区域, 群落组成也较为复杂, 因此刺槐采取相同的生长策略来适应生境的变化. ...
木本植物茎叶功能性状及其关系随环境变化的研究进展
2
2012
... 坡向虽然对刺槐枝叶大小回归方程的斜率没有显著影响, 但使回归方程的截距产生了显著改变.在茎截面积-总叶面积关系中, 其回归方程的截距在北坡显著大于南坡(p < 0.05), 表明在茎截面积一定时, 位于北坡的刺槐在单位大小枝条上支持的总叶面积大于位于南坡的刺槐.叶片大小是植物适应环境中热量和水分交换平衡的结果, 其变化主要是通过影响叶边缘对热量、CO2 和水分的传导能力来影响整个植株的生长速率(杨冬梅等, 2012b ).南坡光照时间长, 水分蒸发大(殷秀琴, 2004 ), 土壤水分含量低(表1 ), 在西北年降水量小的干旱半干旱地区, 由于高温常引起蒸腾作用加强而使植物体水分大量丧失, 因而这种高温和干旱环境严重影响了植物的新陈代谢活动, 细胞的分裂和生长受到限制, 植物生长缓慢, 叶片较小; 然而北坡气温降低, 地面蒸发量减少, 土壤水分增加(表1 ), 良好的水热条件有利于细胞体积和数目增加, 从而促进了叶面积的扩展(李钰等, 2013 ).反过来, 与小叶相比, 大叶具有更厚的叶边缘, 热量和物质通过叶边缘进行转运的阻力更大, 往往会阻止叶片与周围的空气进行热量交换, 减慢CO2 和水蒸气进出叶片的扩散速率, 使大叶比小叶更容易过热, 植物的呼吸和蒸腾成本更高(Givnish, 1978 ), 因此在冷凉、湿润或者荫蔽的环境下大叶由于能够使光合收益最大化而占有优势(Givnish, 1979 ), 而相对小的叶片因为具有较高的热交换能力, 被认为在热、干、高光和低养分的环境下较有优势(Ackerly & Preston, 2003 ; McDonald et al ., 2003 ).另外, 大叶片植物在水分较少的生境中不具有选择优势, 与植物的水力结构特征有关.随着土壤湿度的下降, 水分供应不足会增加水分从土壤运送到叶片的连续水柱断裂的风险, 当大叶片植物的蒸腾速率增大时, 空穴现象和栓塞现象更易发生(McCulloh & Sperry, 2005 ), 从而使得大叶片植物在水分稀缺的生境下不具有水分运输方面的选择优势, 这与Callaway等(1994) 的研究结果一致, 即: 一定大小枝条输导面积支持的总叶面积随着大气蒸腾强度和干旱强度的增加而减小. ...
... 除此之外, 叶大小-叶数量权衡关系的坡向差异性还可以从以下两个方面解释: (1)植物内在生物量分配限制的结果.由表2 可知, 北坡(阴坡)刺槐单叶面积大于南坡(阳坡), 在植物体上, 在某一给定的叶生物量或某一给定的总叶面积下, 植物要么是有较少的大叶子, 要么是有较多的小叶子, 增加单个叶片的投资意味着叶片数量的减少(杨冬梅等, 2012a ); (2)单叶面积的差异是出叶强度变化的副产物.每个叶片通常都会有一个腋芽分生组织(Bell, 1993 ), 叶片更多的物种, 单位个体大小的腋芽分生组织的密度将会更高(杨冬梅等, 2012b ), 更大的芽库由于有更多的部位具有再生能力, 既可以生长为植株的营养结构或繁殖结构, 也可以保持无活性状态, 进而影响到植株的大小构建和繁殖产出(Bonser & Aarssen, 1996 ), 使植株具有更大的可塑性和更新复壮能力, 因此在南坡干、热、高光和低养分的环境下, 刺槐倾向于具有更高的出叶强度, 叶大小与出叶强度紧密相关, 因此小叶片可能是选择了高的出叶强度的结果.东坡和西坡枝叶功能性状及其关系之间没有明显差异, 主要是因为东坡和西坡属于半阴坡和半阳坡, 皆处于阴坡和阳坡之间的过渡区域, 群落组成也较为复杂, 因此刺槐采取相同的生长策略来适应生境的变化. ...
木本植物茎叶功能性状及其关系随环境变化的研究进展
2
2012
... 坡向虽然对刺槐枝叶大小回归方程的斜率没有显著影响, 但使回归方程的截距产生了显著改变.在茎截面积-总叶面积关系中, 其回归方程的截距在北坡显著大于南坡(p < 0.05), 表明在茎截面积一定时, 位于北坡的刺槐在单位大小枝条上支持的总叶面积大于位于南坡的刺槐.叶片大小是植物适应环境中热量和水分交换平衡的结果, 其变化主要是通过影响叶边缘对热量、CO2 和水分的传导能力来影响整个植株的生长速率(杨冬梅等, 2012b ).南坡光照时间长, 水分蒸发大(殷秀琴, 2004 ), 土壤水分含量低(表1 ), 在西北年降水量小的干旱半干旱地区, 由于高温常引起蒸腾作用加强而使植物体水分大量丧失, 因而这种高温和干旱环境严重影响了植物的新陈代谢活动, 细胞的分裂和生长受到限制, 植物生长缓慢, 叶片较小; 然而北坡气温降低, 地面蒸发量减少, 土壤水分增加(表1 ), 良好的水热条件有利于细胞体积和数目增加, 从而促进了叶面积的扩展(李钰等, 2013 ).反过来, 与小叶相比, 大叶具有更厚的叶边缘, 热量和物质通过叶边缘进行转运的阻力更大, 往往会阻止叶片与周围的空气进行热量交换, 减慢CO2 和水蒸气进出叶片的扩散速率, 使大叶比小叶更容易过热, 植物的呼吸和蒸腾成本更高(Givnish, 1978 ), 因此在冷凉、湿润或者荫蔽的环境下大叶由于能够使光合收益最大化而占有优势(Givnish, 1979 ), 而相对小的叶片因为具有较高的热交换能力, 被认为在热、干、高光和低养分的环境下较有优势(Ackerly & Preston, 2003 ; McDonald et al ., 2003 ).另外, 大叶片植物在水分较少的生境中不具有选择优势, 与植物的水力结构特征有关.随着土壤湿度的下降, 水分供应不足会增加水分从土壤运送到叶片的连续水柱断裂的风险, 当大叶片植物的蒸腾速率增大时, 空穴现象和栓塞现象更易发生(McCulloh & Sperry, 2005 ), 从而使得大叶片植物在水分稀缺的生境下不具有水分运输方面的选择优势, 这与Callaway等(1994) 的研究结果一致, 即: 一定大小枝条输导面积支持的总叶面积随着大气蒸腾强度和干旱强度的增加而减小. ...
... 除此之外, 叶大小-叶数量权衡关系的坡向差异性还可以从以下两个方面解释: (1)植物内在生物量分配限制的结果.由表2 可知, 北坡(阴坡)刺槐单叶面积大于南坡(阳坡), 在植物体上, 在某一给定的叶生物量或某一给定的总叶面积下, 植物要么是有较少的大叶子, 要么是有较多的小叶子, 增加单个叶片的投资意味着叶片数量的减少(杨冬梅等, 2012a ); (2)单叶面积的差异是出叶强度变化的副产物.每个叶片通常都会有一个腋芽分生组织(Bell, 1993 ), 叶片更多的物种, 单位个体大小的腋芽分生组织的密度将会更高(杨冬梅等, 2012b ), 更大的芽库由于有更多的部位具有再生能力, 既可以生长为植株的营养结构或繁殖结构, 也可以保持无活性状态, 进而影响到植株的大小构建和繁殖产出(Bonser & Aarssen, 1996 ), 使植株具有更大的可塑性和更新复壮能力, 因此在南坡干、热、高光和低养分的环境下, 刺槐倾向于具有更高的出叶强度, 叶大小与出叶强度紧密相关, 因此小叶片可能是选择了高的出叶强度的结果.东坡和西坡枝叶功能性状及其关系之间没有明显差异, 主要是因为东坡和西坡属于半阴坡和半阳坡, 皆处于阴坡和阳坡之间的过渡区域, 群落组成也较为复杂, 因此刺槐采取相同的生长策略来适应生境的变化. ...
半干旱黄土丘陵区人工林叶面积特征
1
2008
... 刺槐(Robinia pseudoacacia )是豆科刺槐属的温带落叶乔木, 具有生长速度快、根系发达、对环境适应性强、固碳氮能力较好、成活率高、萌芽力和根蘖性都很强等优点, 已成为我国黄土丘陵区的主要造林树种和人工林面积最大的树种(于占辉等, 2009 ; 李军等, 2010 ).近年来, 干旱地区许多刺槐人工林出现了生长减慢、枝梢干枯甚至整株死亡的衰败现象, 引起了学术界的广泛关注(单长卷等, 2004 ; 李军等, 2010 ).目前, 许多学者研究了刺槐叶面积生长与林分密度的关系(尹婧等, 2008 ), 土层厚度对刺槐旱季水分状况和生长的影响(王林等, 2013 ), 不同林龄和退耕年限刺槐人工林碳、氮储量及分配规律(申家朋和张文辉, 2014 ; 艾泽民等, 2014 ), 揭示了刺槐生长以及生物量积累和分配格局等一系列规律, 但关于微地形影响下刺槐当年生小枝内枝叶关系的研究较为少见, 更鲜有不同坡向茎截面积与总叶面积、出叶强度与单叶面积关系的报道, 限制了对异质生境中刺槐适应机制的解析.本研究选择兰州北山九州台西北师范大学绿化基地的刺槐人工林为研究对象, 探讨刺槐当年生小枝内的茎截面积-总叶面积、出叶强度-单叶面积关系, 为进一步了解刺槐的环境适应机制提供理论依据, 对提高干旱半干旱地区人工林的可持续性和稳定性具有重要意义. ...
半干旱黄土丘陵区人工林叶面积特征
1
2008
... 刺槐(Robinia pseudoacacia )是豆科刺槐属的温带落叶乔木, 具有生长速度快、根系发达、对环境适应性强、固碳氮能力较好、成活率高、萌芽力和根蘖性都很强等优点, 已成为我国黄土丘陵区的主要造林树种和人工林面积最大的树种(于占辉等, 2009 ; 李军等, 2010 ).近年来, 干旱地区许多刺槐人工林出现了生长减慢、枝梢干枯甚至整株死亡的衰败现象, 引起了学术界的广泛关注(单长卷等, 2004 ; 李军等, 2010 ).目前, 许多学者研究了刺槐叶面积生长与林分密度的关系(尹婧等, 2008 ), 土层厚度对刺槐旱季水分状况和生长的影响(王林等, 2013 ), 不同林龄和退耕年限刺槐人工林碳、氮储量及分配规律(申家朋和张文辉, 2014 ; 艾泽民等, 2014 ), 揭示了刺槐生长以及生物量积累和分配格局等一系列规律, 但关于微地形影响下刺槐当年生小枝内枝叶关系的研究较为少见, 更鲜有不同坡向茎截面积与总叶面积、出叶强度与单叶面积关系的报道, 限制了对异质生境中刺槐适应机制的解析.本研究选择兰州北山九州台西北师范大学绿化基地的刺槐人工林为研究对象, 探讨刺槐当年生小枝内的茎截面积-总叶面积、出叶强度-单叶面积关系, 为进一步了解刺槐的环境适应机制提供理论依据, 对提高干旱半干旱地区人工林的可持续性和稳定性具有重要意义. ...
1
2004
... 坡向虽然对刺槐枝叶大小回归方程的斜率没有显著影响, 但使回归方程的截距产生了显著改变.在茎截面积-总叶面积关系中, 其回归方程的截距在北坡显著大于南坡(p < 0.05), 表明在茎截面积一定时, 位于北坡的刺槐在单位大小枝条上支持的总叶面积大于位于南坡的刺槐.叶片大小是植物适应环境中热量和水分交换平衡的结果, 其变化主要是通过影响叶边缘对热量、CO2 和水分的传导能力来影响整个植株的生长速率(杨冬梅等, 2012b ).南坡光照时间长, 水分蒸发大(殷秀琴, 2004 ), 土壤水分含量低(表1 ), 在西北年降水量小的干旱半干旱地区, 由于高温常引起蒸腾作用加强而使植物体水分大量丧失, 因而这种高温和干旱环境严重影响了植物的新陈代谢活动, 细胞的分裂和生长受到限制, 植物生长缓慢, 叶片较小; 然而北坡气温降低, 地面蒸发量减少, 土壤水分增加(表1 ), 良好的水热条件有利于细胞体积和数目增加, 从而促进了叶面积的扩展(李钰等, 2013 ).反过来, 与小叶相比, 大叶具有更厚的叶边缘, 热量和物质通过叶边缘进行转运的阻力更大, 往往会阻止叶片与周围的空气进行热量交换, 减慢CO2 和水蒸气进出叶片的扩散速率, 使大叶比小叶更容易过热, 植物的呼吸和蒸腾成本更高(Givnish, 1978 ), 因此在冷凉、湿润或者荫蔽的环境下大叶由于能够使光合收益最大化而占有优势(Givnish, 1979 ), 而相对小的叶片因为具有较高的热交换能力, 被认为在热、干、高光和低养分的环境下较有优势(Ackerly & Preston, 2003 ; McDonald et al ., 2003 ).另外, 大叶片植物在水分较少的生境中不具有选择优势, 与植物的水力结构特征有关.随着土壤湿度的下降, 水分供应不足会增加水分从土壤运送到叶片的连续水柱断裂的风险, 当大叶片植物的蒸腾速率增大时, 空穴现象和栓塞现象更易发生(McCulloh & Sperry, 2005 ), 从而使得大叶片植物在水分稀缺的生境下不具有水分运输方面的选择优势, 这与Callaway等(1994) 的研究结果一致, 即: 一定大小枝条输导面积支持的总叶面积随着大气蒸腾强度和干旱强度的增加而减小. ...
1
2004
... 坡向虽然对刺槐枝叶大小回归方程的斜率没有显著影响, 但使回归方程的截距产生了显著改变.在茎截面积-总叶面积关系中, 其回归方程的截距在北坡显著大于南坡(p < 0.05), 表明在茎截面积一定时, 位于北坡的刺槐在单位大小枝条上支持的总叶面积大于位于南坡的刺槐.叶片大小是植物适应环境中热量和水分交换平衡的结果, 其变化主要是通过影响叶边缘对热量、CO2 和水分的传导能力来影响整个植株的生长速率(杨冬梅等, 2012b ).南坡光照时间长, 水分蒸发大(殷秀琴, 2004 ), 土壤水分含量低(表1 ), 在西北年降水量小的干旱半干旱地区, 由于高温常引起蒸腾作用加强而使植物体水分大量丧失, 因而这种高温和干旱环境严重影响了植物的新陈代谢活动, 细胞的分裂和生长受到限制, 植物生长缓慢, 叶片较小; 然而北坡气温降低, 地面蒸发量减少, 土壤水分增加(表1 ), 良好的水热条件有利于细胞体积和数目增加, 从而促进了叶面积的扩展(李钰等, 2013 ).反过来, 与小叶相比, 大叶具有更厚的叶边缘, 热量和物质通过叶边缘进行转运的阻力更大, 往往会阻止叶片与周围的空气进行热量交换, 减慢CO2 和水蒸气进出叶片的扩散速率, 使大叶比小叶更容易过热, 植物的呼吸和蒸腾成本更高(Givnish, 1978 ), 因此在冷凉、湿润或者荫蔽的环境下大叶由于能够使光合收益最大化而占有优势(Givnish, 1979 ), 而相对小的叶片因为具有较高的热交换能力, 被认为在热、干、高光和低养分的环境下较有优势(Ackerly & Preston, 2003 ; McDonald et al ., 2003 ).另外, 大叶片植物在水分较少的生境中不具有选择优势, 与植物的水力结构特征有关.随着土壤湿度的下降, 水分供应不足会增加水分从土壤运送到叶片的连续水柱断裂的风险, 当大叶片植物的蒸腾速率增大时, 空穴现象和栓塞现象更易发生(McCulloh & Sperry, 2005 ), 从而使得大叶片植物在水分稀缺的生境下不具有水分运输方面的选择优势, 这与Callaway等(1994) 的研究结果一致, 即: 一定大小枝条输导面积支持的总叶面积随着大气蒸腾强度和干旱强度的增加而减小. ...
黄土高原半干旱区人工林刺槐展叶期树干液流动态分析
1
2009
... 刺槐(Robinia pseudoacacia )是豆科刺槐属的温带落叶乔木, 具有生长速度快、根系发达、对环境适应性强、固碳氮能力较好、成活率高、萌芽力和根蘖性都很强等优点, 已成为我国黄土丘陵区的主要造林树种和人工林面积最大的树种(于占辉等, 2009 ; 李军等, 2010 ).近年来, 干旱地区许多刺槐人工林出现了生长减慢、枝梢干枯甚至整株死亡的衰败现象, 引起了学术界的广泛关注(单长卷等, 2004 ; 李军等, 2010 ).目前, 许多学者研究了刺槐叶面积生长与林分密度的关系(尹婧等, 2008 ), 土层厚度对刺槐旱季水分状况和生长的影响(王林等, 2013 ), 不同林龄和退耕年限刺槐人工林碳、氮储量及分配规律(申家朋和张文辉, 2014 ; 艾泽民等, 2014 ), 揭示了刺槐生长以及生物量积累和分配格局等一系列规律, 但关于微地形影响下刺槐当年生小枝内枝叶关系的研究较为少见, 更鲜有不同坡向茎截面积与总叶面积、出叶强度与单叶面积关系的报道, 限制了对异质生境中刺槐适应机制的解析.本研究选择兰州北山九州台西北师范大学绿化基地的刺槐人工林为研究对象, 探讨刺槐当年生小枝内的茎截面积-总叶面积、出叶强度-单叶面积关系, 为进一步了解刺槐的环境适应机制提供理论依据, 对提高干旱半干旱地区人工林的可持续性和稳定性具有重要意义. ...
黄土高原半干旱区人工林刺槐展叶期树干液流动态分析
1
2009
... 刺槐(Robinia pseudoacacia )是豆科刺槐属的温带落叶乔木, 具有生长速度快、根系发达、对环境适应性强、固碳氮能力较好、成活率高、萌芽力和根蘖性都很强等优点, 已成为我国黄土丘陵区的主要造林树种和人工林面积最大的树种(于占辉等, 2009 ; 李军等, 2010 ).近年来, 干旱地区许多刺槐人工林出现了生长减慢、枝梢干枯甚至整株死亡的衰败现象, 引起了学术界的广泛关注(单长卷等, 2004 ; 李军等, 2010 ).目前, 许多学者研究了刺槐叶面积生长与林分密度的关系(尹婧等, 2008 ), 土层厚度对刺槐旱季水分状况和生长的影响(王林等, 2013 ), 不同林龄和退耕年限刺槐人工林碳、氮储量及分配规律(申家朋和张文辉, 2014 ; 艾泽民等, 2014 ), 揭示了刺槐生长以及生物量积累和分配格局等一系列规律, 但关于微地形影响下刺槐当年生小枝内枝叶关系的研究较为少见, 更鲜有不同坡向茎截面积与总叶面积、出叶强度与单叶面积关系的报道, 限制了对异质生境中刺槐适应机制的解析.本研究选择兰州北山九州台西北师范大学绿化基地的刺槐人工林为研究对象, 探讨刺槐当年生小枝内的茎截面积-总叶面积、出叶强度-单叶面积关系, 为进一步了解刺槐的环境适应机制提供理论依据, 对提高干旱半干旱地区人工林的可持续性和稳定性具有重要意义. ...
植物叶寿命及其相关叶性状的生态学研究进展
1
2004
... 植物功能性状与环境和生态系统功能的关系是近年来生态学和全球变化研究的热点(Harrison et al ., 2010 ; Bernard-Verdier et al ., 2012 ), 各功能性状之间的权衡和组合决定了植物的生活史对策(Vandermeer, 1972 ; Silvertown, 2004 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ), 叶片是植物进行光合作用的主要器官(张林和罗天祥, 2004 ), 两者大小和数量的配置关系直接影响着植物的机械力学和水力结构特征, 从而决定了植物的生活史策略, 并可能影响物种生态位的利用(Givnish, 1987 ; Enquist et al ., 1999 ; West et al ., 1999 ; Westoby et al ., 2002 ).植物单位大小枝条上支撑的叶面积必须与支撑枝的横截面积成一定的比例, 才能符合植物器官间的机械力学和水力结构需求(Niklas, 1992 ; Enquist et al ., 1999 ; Normand et al ., 2008 ; Olson et al ., 2009 ).叶片大小和叶片数量是决定树冠紧密程度的两个重要因素, 直接影响到植株的冠层结构和发育模式, 进而影响到植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ).在某一给定的叶生物量分配或某一个给定的总叶面积下, 植物可能有较少的大叶子或者较多的小叶子(杨冬梅等, 2012a ).坡向作为一种重要的地形要素, 往往对局部区域的太阳辐射和降水进行二次分配(Cantón et al ., 2004 ), 在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 能够对植物的形态产生影响(Bazzaz, 1996 ; 韦莉莉等, 2005 ).当自然资源的丰富度低于或超过植物光合系统所能利用的最低或最高限度时, 植物的光合作用将被抑制(王玉萍等, 2013 ).为提高植物构型构建的生境适应性和风险规避能力, 植物通过功能性状等表型的可塑性调整提高自身的生存适合度, 体现了全球气候变化背景下植物的资源配置策略.因此, 研究坡向梯度上植物枝叶关系规律及其成因, 对于理解植物的环境适应机制有着重要意义. ...
植物叶寿命及其相关叶性状的生态学研究进展
1
2004
... 植物功能性状与环境和生态系统功能的关系是近年来生态学和全球变化研究的热点(Harrison et al ., 2010 ; Bernard-Verdier et al ., 2012 ), 各功能性状之间的权衡和组合决定了植物的生活史对策(Vandermeer, 1972 ; Silvertown, 2004 ).在植物器官中, 当年生小枝是植物分枝系统最具有活力的部分(李亚男等, 2008 ), 叶片是植物进行光合作用的主要器官(张林和罗天祥, 2004 ), 两者大小和数量的配置关系直接影响着植物的机械力学和水力结构特征, 从而决定了植物的生活史策略, 并可能影响物种生态位的利用(Givnish, 1987 ; Enquist et al ., 1999 ; West et al ., 1999 ; Westoby et al ., 2002 ).植物单位大小枝条上支撑的叶面积必须与支撑枝的横截面积成一定的比例, 才能符合植物器官间的机械力学和水力结构需求(Niklas, 1992 ; Enquist et al ., 1999 ; Normand et al ., 2008 ; Olson et al ., 2009 ).叶片大小和叶片数量是决定树冠紧密程度的两个重要因素, 直接影响到植株的冠层结构和发育模式, 进而影响到植物对光的截取和对碳的获取能力(Parkhurst & Loucks, 1972 ; Givnish & Vermeij, 1976 ).在某一给定的叶生物量分配或某一个给定的总叶面积下, 植物可能有较少的大叶子或者较多的小叶子(杨冬梅等, 2012a ).坡向作为一种重要的地形要素, 往往对局部区域的太阳辐射和降水进行二次分配(Cantón et al ., 2004 ), 在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 能够对植物的形态产生影响(Bazzaz, 1996 ; 韦莉莉等, 2005 ).当自然资源的丰富度低于或超过植物光合系统所能利用的最低或最高限度时, 植物的光合作用将被抑制(王玉萍等, 2013 ).为提高植物构型构建的生境适应性和风险规避能力, 植物通过功能性状等表型的可塑性调整提高自身的生存适合度, 体现了全球气候变化背景下植物的资源配置策略.因此, 研究坡向梯度上植物枝叶关系规律及其成因, 对于理解植物的环境适应机制有着重要意义. ...
 , 宋清华, 杜晶, 陈静, 王继伟
, 宋清华, 杜晶, 陈静, 王继伟


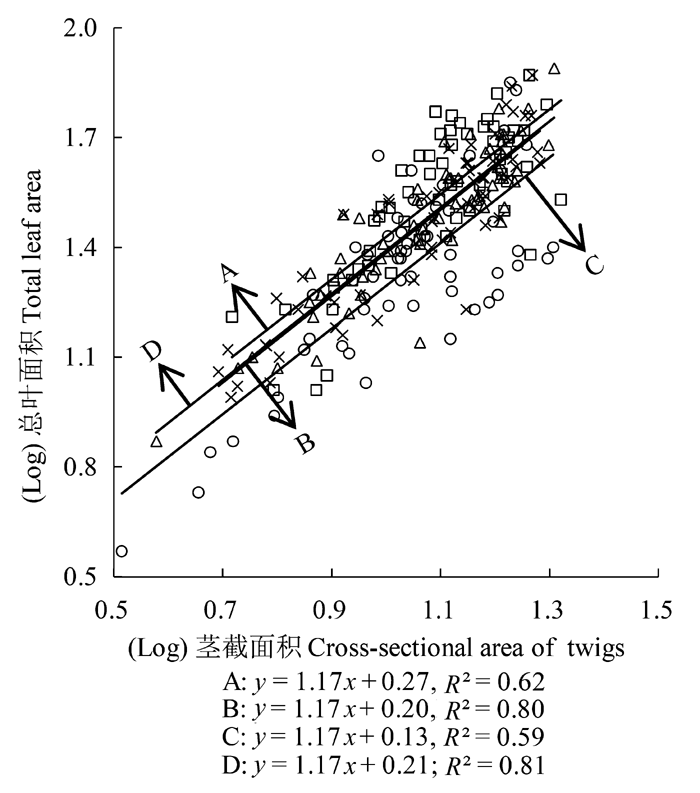
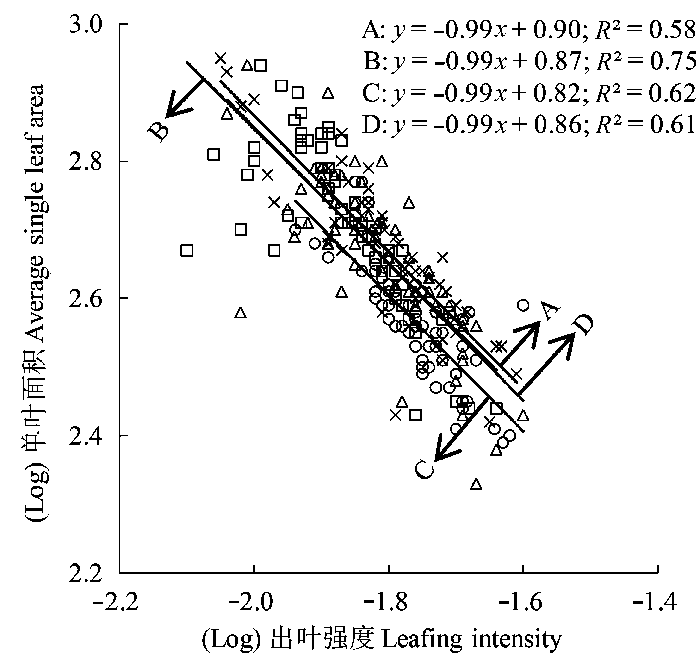
{kind=link}
{kind=link}
{kind=link}
{kind=link}