毛竹(Phyllostachys heterocycla ‘Pubescens’)原产于我国暖温带和亚热带地区, 是我国南方重要的笋、材两用竹种(郑郁善和洪伟, 1998 )。毛竹经济和生态效益极高, 栽培历史悠久(杨怀等, 2010 ), 对毛竹的研究主要集中在如何获取更大的经济利益上(刘烁, 2010 ), 而对其生态方面的研究较少。近年发现, 许多地区的毛竹林扩张并取代了周围的针、阔叶森林(洪伟等, 2004 ; 丁丽霞等, 2006 ; 朱长龙和上官林平, 2009 ; 杨清培等, 2011 )。毛竹具有强大的克隆性生长繁育及扩鞭能力, 极易向周围的常绿阔叶林、针阔混交林蔓延扩张, 蚕食其他针阔树种, 导致周边林木失活枯死并抑制林下幼苗更新, 最终转变为毛竹纯林。
国内外一些学者从人类活动(蔡亮等, 2003 ; Suzuki & Nakagoshi, 2008 )、生长特性(董鸣, 1996 ; Okutomi et al ., 1996 )、土壤资源(张春生等, 2007 ; 黄启堂, 2008 ; 吴家森等, 2008 ; 杨怀等, 2010 ; 宋庆妮等, 2013 )、光环境(Suzuki & Nakatsubo, 2001 ; 刘烁, 2010 )和化感作用(Chou & Yang, 1982 ; 梁倩倩等, 2012 ; 白尚斌等, 2013 )等方面对毛竹林扩张形成纯林的原因进行了分析探讨, 但有关毛竹林扩张导致其他林木失活枯死并抑制林下幼苗更新的机制, 目前尚有争议。
生物或非生物因素对外来入侵种的正反馈作用对外来生物的定植和传播有不可忽视的作用。其中, 植物和土壤的相互反馈在促进或抑制植物个体的更新上具有重要意义(Vogelsang & Bever, 2009 )。植物与土壤微生物形成强共生关系, 如菌根(mycorr- hiza), 干扰往往会削弱或破坏这种共生关系, 从而可能促进外来种的定植, 使原植物群落发生改变。如果退化的土壤微生物群落进一步抑制本地种的更新, 群落更替会加速(Vogelsang & Bever, 2009 )。菌根与植物共生现象的普遍性与其生态学功能的重要性, 使其容易成为寄主植物反馈调节的媒介。例如有研究发现, 在亚高山森林植被和高山草甸的过渡带, 柳树(Salix spp.)可以通过自身的枯落物及其外生菌根间接降低周围草本植物群落的丛枝菌根侵染, 进而可能改变菌根真菌在这一地区的分布, 这一改变又反过来影响草本植物群落的分布和生长(Becklin et al ., 2012 )。
综上所述, 人们有如下的共识: 1)干扰会削弱或破坏植物与土壤微生物的共生关系; 2)植物和土壤的相互反馈在促进或抑制植物个体的更新上具有重要意义; 3)菌根的养分吸收和抗逆境胁迫功能对针阔混交林及常绿阔叶林中树种的正常生长很重要。此外, 根据我们预实验观察, 毛竹是弱菌根共生植物, 而菌根共生对天目山针阔混交林及常绿阔叶林中的绝大部分树种都更为重要。基于上述共识和观察, 我们提出了菌根减弱假说, 即毛竹扩张进入周边针阔混交林导致林木死亡并抑制林下幼苗更新的机制可能是: 毛竹扩张引起森林群落的菌根系统紊乱, 使宿主植物与菌根真菌的共生关系受创, 进而影响宿主植物的分布与更新, 最终导致林木枯萎或死亡并阻碍非毛竹树种的林下更新。
为检验菌根减弱假说, 我们选择了位于浙江天目山自然保护区林场的一块缓坡, 毛竹从山脚向山上的针阔混交林渗透扩张, 形成明显的毛竹纯林-针阔混交林连续带, 其间有较窄的竹-林过渡带, 我们以此作为研究对象。我们选取了在针阔混交林、竹-林过渡带同时存在的优势乔、灌木物种, 包括杉木(Cunninghamia lanceolata )、枫香树(Liquidambar formosana )、青冈(Cyclobalanopsis glauca )、柳杉(Cryptomeria fortunei )、江浙山胡椒(Lindera chienii )、毛柄连蕊茶(Camellia fraterna )进行调查。 通过测定比较毛竹在毛竹纯林、竹-林过渡带的丛枝菌根(arbuscular mycorrhiza, AM)侵染频率和强度, 内生菌根树种(杉木、毛柄连蕊茶、江浙山胡椒)在竹-林过渡带、针阔混交林的菌根侵染频率和强度, 外生菌根树种(青冈、柳杉、枫香树)在竹-林过渡带、针阔混交林的菌根侵染频率, 检验我们的菌根减弱假说。根据菌根减弱假说, 我们预计: 毛竹林扩张进入周边针阔混交林引起林木菌根真菌根系统紊乱, 表现为过渡带中菌根侵染频率和侵染强度急剧减少。
1 材料和方法
1.1 研究地点
本研究地点位于浙江省西北部临安市境内的天目山国家级自然保护区(30.31°-30.36° N, 119.40- 119.45° E), 属于亚热带季风气侯, 冬暖夏凉, 雨量充沛。自山麓禅源寺至主峰山顶仙人顶, 年平均气温为8.8-14.8 ℃, 最冷月平均气温-2.6-3.4 ℃, 极端最低气温-20.2 ℃, 最热月平均气温19.9-28.1 ℃, 极端最高气温38.2 ℃, ≥10 ℃积温2500-5100 ℃, 无霜期209-235天, 年降水量1390-1870 mm, 年太阳总辐射3770-4460 MJ·m-2 。土壤以红壤、黄壤、黄红壤、棕黄壤为主(丁丽霞等, 2006 )。近年来毛竹扩张并取代针阔混交林的现象大量出现, 大多发生于该保护区内低、中山区。
1.2 样带设计与样品采集
实验样带位于保护区林场有毛竹扩张并取代针阔混交林的地区。试验地山坡基本南向, 坡度在30°-35°之间, 海拔390-590 m, 土壤为黄壤、红黄壤, 类似于美国土壤分类的Ultisols (极育土)。针阔混交林的主要乔木树种为苦槠栲(Castanopsis sclerophylla )、青冈、柳杉、木荷(Schima superba )、杉木、枫香树、马尾松(Pinus massoniana )等。灌木有毛柄连蕊茶、江浙山胡椒、马银花(Rhododendron ovatum )、山矾(Symplocos caudata )、山胡椒(Lindera glauca )、豆腐柴(Premna microphylla )、菝葜(Smilax china )、油茶(Camellia oleifera )和南天竹(Nandina domestica )等。草本植物主要为菊科、禾本科、鳞毛蕨科和莎草科植物。
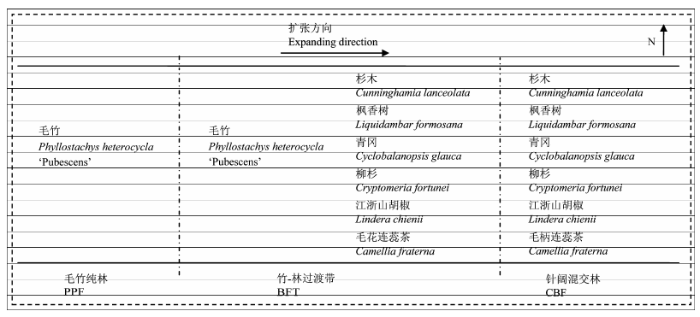
2011年7月中旬, 我们在垂直于毛竹扩张方向设置毛竹纯林、竹-林过渡带、针阔混交林3个样带(图1 ), 其中竹-林过渡带宽10-20 m。我们选取毛竹纯林和竹-林过渡带的毛竹和针阔混交林、竹-林过渡带同时存在的主要树种: 杉木、枫香树、青冈、柳杉、江浙山胡椒、毛柄连蕊茶(图1 )。由于竹-林过渡带空间逼仄, 各树种个体数目有限, 我们在每个样带中心处每树种随机选择3株成年树木, 以避免边缘影响。我们从每株树周围取0-30 cm浅层细根(确认属于该树后)装入标记的封口袋, 置于冷箱中(4 ℃左右)运回实验室。
图1 试验地样带布设示意图。从毛竹纯林到针阔混交林, 海拔逐渐升高。
Fig. 1 Diagram of belt transects in the experimental area. The altitude gradually increased from PPF to CBF. BFT, bamboo-forest transition; CBF, coniferous and broad-leaved mixed forest; PPF, Phyllostachys heterocycla ‘Pubescens’ forest.
以往的菌根资源调查中, 对毛竹的菌根类型有内生(弓明钦等, 1997 ; 陈祥欣和陈连庆, 1983 )和外生(苏琍英等, 1992 )两种不同的说法。柳杉的菌根类型有内生(陈祥欣和陈连庆, 1983 )和外生(苏琍英等, 1992 )不同的描述。枫香树的菌根类型也有内外生(苏琍英等, 1992 )或外生的报道(陈祥欣和陈连庆, 1983 )。也有调查显示青冈是内外生菌根树种(苏琍英等, 1992 )。但在我们本研究区域的预实验研究中发现, 经根切片染色确定, 毛竹、杉木、毛柄连蕊茶、江浙山胡椒菌根是内生菌根(endomycorrhiza, EM)类型, 青冈、柳杉、枫香树菌根是外生菌根(ectomycorrhiza, ECM)类型。
1.3 内生菌根侵染频率与侵染强度的测定
内生菌根的染色采用醋酸墨水染色法(Vierh- eilig et al ., 1998 )。根系样品用去离子水冲洗至无土壤杂物粘附, 在解剖镜下将根尖部无次生生长的0.5-1.0 cm根尖段剪下, 放入70%的乙醇中保存待用。每样品包括至少500个合格根尖段。染色时, 随机选取30-50条根尖段, 用去离子水冲洗3遍后, 置于10% KOH溶液中放入高压锅(121 ℃)内处理10-30 min, 使根尖段脱色透明。然后再用去离子水冲洗3次, 置于3%的碱性H2 O2 (25%-28% NH4 OH 8 mL, 3% H2 O2 100 mL)溶液中漂白15-45 min, 再用去离子水冲洗3次。随后将这些根尖段放入5%醋酸(CH3 COOH)中酸化30 min后移至5%醋酸墨水染色液(5%冰醋酸95 mL, 派克牌纯黑墨水5 mL)中煮沸3-30 min染色。染色后, 用5%醋酸冲洗根尖段20 min。最后置染色根尖段于去离子水中浸泡脱色24 h以上。
挑取脱色处理后的根尖段于载玻片上, 一块载玻片上平行放置8条根尖段, 将其拉直, 用甘油明胶固封(Widden, 2001 ), 加盖洁净盖玻片将根尖段压平后, 在Zeiss Axio Imager M1显微镜(Carl Zeiss AG, Heidenheim, Germany)放大100-400倍条件下观察丛枝菌根形态并记录、拍照(图2 )。丛枝菌根的侵染频率和强度采用侵染分级法(Trouvelot et al ., 1986 )计算。丛枝菌根侵染频率为侵染的根尖段数占总根尖段数的比率, 侵染强度为丛枝菌根结构(丛枝、泡囊及菌丝)占根尖段的比率。
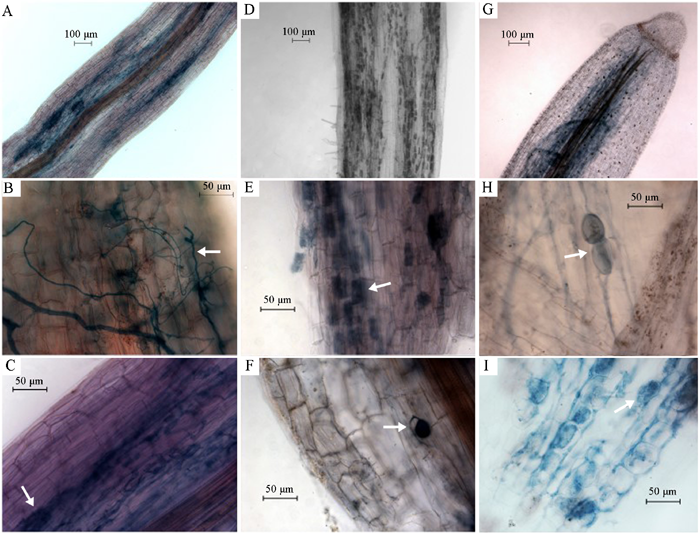
图2 丛枝菌根形态(墨水染色)。A, 杉木侵染根尖段。B, 杉木菌根内菌丝(箭头所示)。C, 杉木菌根丛枝结构(箭头所示)。D, 毛柄连蕊茶侵染根尖段。E, 毛柄连蕊茶菌根丛枝结构(箭头所示)。F, 毛柄连蕊茶菌根孢子(箭头所示)。G, 江浙山胡椒侵染根尖段。H, 江浙山胡椒菌根孢子(箭头所示)。I, 江浙山胡椒丛枝结构(箭头所示)。
Fig. 2 Morphology of arbuscular mycorrhiza, stained with ink. A, An infected root tip of Cunninghamia lanceolata . B, Arrow indicates intraradical hyphae in a root of Cunninghamia lanceolata . C, Arrow indicates arbuscules in a root of Cunninghamia lanceolata . D, A infected root tip of Camellia fraterna . E, Arrow indicates arbuscules in a root of Camellia fraterna . F, Arrow indicates a fungal spore in a root of Camellia fraterna . G, An infected root tip of Lindera chienii . H, Arrow indicates two fungal spores in a root of Lindera chienii . I, Arrow indicates arbuscules in a root of Lindera chienii .
1.4 外生菌根侵染频率测定
实验室中, 用去离子水将细根冲洗干净, 去除根系表面的土壤颗粒及动植物残体, 选出完整的活根样品, 放入70%的乙醇中保存待用。每样品包括至少500个合格根尖段。测定时, 从每一样品中随机选择300个根尖段(约1.0 cm), 在Stemi SV 11 10-50倍体视镜(Carl Zeiss AG, Heidenheim Germany)下直接镜检, 外生菌根的典型辨别特征: 1级根(距离根轴最远端且没有分枝的细根)根尖粗短肥大, 或有各种形态的分枝, 表面或有菌套包裹, 无根冠和根毛(图3 )。外生菌根侵染频率为侵染的根尖段数占总根尖段数的比率。
图3 枫香树(A-C)、柳杉(D-F)、青冈(G-I)外生菌根形态。
Fig. 3 Morphology of ectomycorrhiza in Liquidambar formosana (A-C), Cryptomeria fortunei (D-F), and Cyclobalanopsis glauca (G-I).
1.5 土壤样品采集和理化性质测定
在毛竹纯林和竹-林过渡带, 在靠近毛竹茎基部30 cm处和离毛竹茎基部2 m以外分别随机选择5个样点, 用小铲取表层(0-10 cm)土壤50 g, 装入标记的自封袋。在针阔混交林样带随机采取表层土(0-10 cm)土样5份, 装袋后均放入便携冷藏箱(4 ℃左右)带回实验室进行土壤pH值、含水率和有效氮(铵态氮、硝态氮)含量等理化指标测定(鲍士旦, 2000 )。
1.6 数据分析
为检验毛竹纯林和竹-林过渡带中土壤与毛竹的距离远近是否会影响土壤养分含量, 我们用所采土样距离毛竹的位置(近距/远距毛竹)为因变量, 对两个样带的土壤因子(土壤含水率、pH值、有效氮(铵态氮NH4 + -N、硝态氮NO3 - -N)含量)分别进行t 检验(Student’s t -test)。为检测毛竹林扩张对周边林木丛枝菌根的影响, 我们采用以样地林分类型(针阔混交林、竹-林过渡带)为固定因子的一元方差分析(one-way ANOVA), 比较3种感染内生菌根真菌的树木(杉木、毛柄连蕊茶、江浙山胡椒)的丛枝菌根侵染频率和强度在两个林分类型间的差异。毛竹林扩张对周边林木外生菌根的影响的检验方法与检验内生菌根的方法相同, 但外生菌根只测定了菌根侵染频率, 故只对菌根侵染频率进行一元方差分析(one-way ANOVA), 以比较其在3个林分类型间的差异。
由于本实验中实验单元的重复数较少, 因此数据分析中的α 值设为0.1。所有的数据分析都在SPSS 16.0软件中完成(SPSS, Chicago, USA)。
2 结果
2.1 毛竹侵入对林分土壤养分有效性的影响
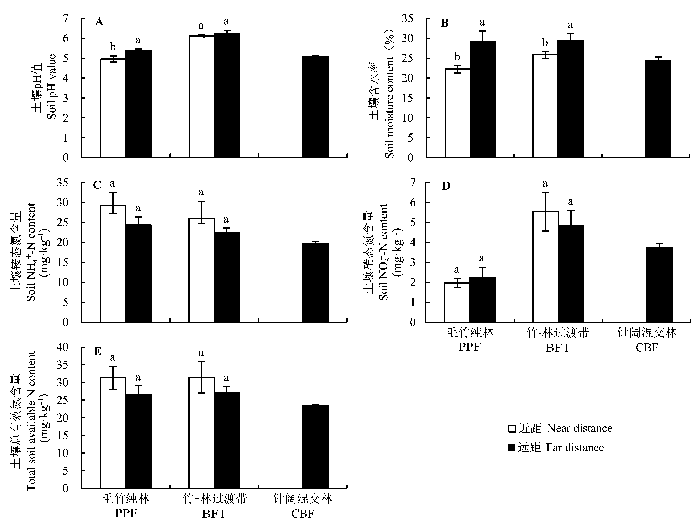
在竹-林过渡带和毛竹纯林中, 因距离毛竹的远近不同, 土壤含水率有显著差异, 远距毛竹的土壤含水率比毛竹附近土壤显著为高。在毛竹纯林中, 远距毛竹的土壤具有显著高的pH值(图4A , 4B)。
图4 试验地3种林分的土壤pH值(A)、含水量(B)、铵态氮(C)、硝态氮(D)、总有效氮(E)对比(平均值±标准误差)。BFT, 竹-林过渡带; CBF, 针阔混交林; PPF, 毛竹纯林。不同小写字母表示同一林型中离毛竹根远近距离的土壤养分指标差异显著(p < 0.1)。
Fig. 4 Comparison of soil pH value (A), soil water content (B), NH4 + -N (C), NO3 - -N (D), total available N (NH4 + -N + NO3 - -N) (E) contents in three stands in the study site (mean ± SE ). BFT, bamboo-forest transition; CBF, coniferous and broad-leaved mixed forest; PPF, Phyllostachys heterocycla ‘Pubescens’ forest. Different lowercase letters denote significant difference among soil nutrients due to the different distance to bamboo roots in the same forest stand (p < 0.1).
同一样地类型的土壤铵态氮、硝态氮和总有效氮与土样距毛竹距离(近距/远距)没有显著差异(图4C , 4D, 4E)。但在不同的样带类型中, 铵态氮含量差异显著, 似乎是随着毛竹林扩张进程而升高, 毛竹纯林的铵态氮含量显著高于针阔混交林, 竹-林过渡带的铵态氮含量居于二者之间(图4C ; 表1 )。毛竹纯林土壤中的硝态氮含量显著低于竹-林过渡带和针阔混交林土壤, 而土壤硝态氮含量在后二者间无显著差异(图4D ; 表1 )。
2.2 毛竹侵入对丛枝菌根侵染频率与侵染强度的影响
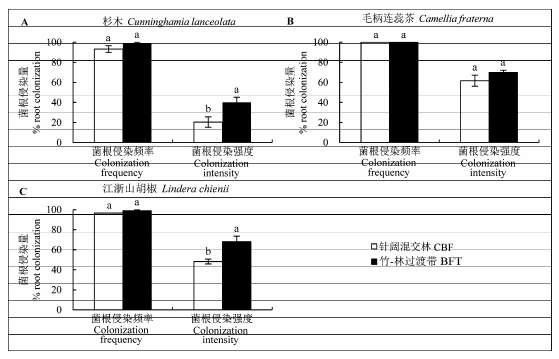
对针阔混交林和竹-林过渡带中杉木、毛柄连蕊茶、江浙山胡椒丛枝菌根侵染频率、强度的观察中, 我们发现3个树种在两个样带中均有很高的侵染频率(>95%, 图5 ), 几乎每条根尖段上都发现有内生菌根侵染结构(菌丝、泡囊、丛枝, 图2 ), 树木的菌根侵染频率在两个森林类型中没有明显差异(图5 )。竹-林过渡带内杉木、江浙山胡椒的菌根侵染强度显著高于针阔混交林, 毛柄连蕊茶的菌根侵染强度在竹-林过渡带略高于针阔混交林, 但差异不显著。
图5 试验地丛枝菌根树种杉木、毛柄连蕊茶、江浙山胡椒在毛竹-针阔混交林竞争界面两侧的菌根侵染频率和强度对比(平均值±标准误差)。BFT, 竹-林过渡带; CBF, 针阔混交林。不同小写字母表示不同样带的差异显著(p < 0.1)。
Fig. 5 Comparison of frequency and intensity of the arbuscular mycorrhizal fungi root colonization of Cunninghamia lanceolata (A), Camellia fraternal (B), Lindera chienii (C) at two sides of bamboo and broad-leaved forest interface in the study site (mean ± SE ). BFT, bamboo-forest transition; CBF, coniferous and broad-leaved mixed forest. Different lowercase letters denote significant difference among different forest stands (p < 0.1).
2.3 毛竹侵入对外生菌根侵染频率的影响
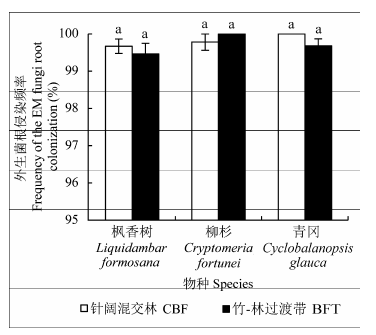
在对针阔混交林和竹-林过渡带的枫香树、柳杉、青冈外生菌根观测中, 我们发现3个树种在两个样带中菌根侵染频率都达到了95%以上(图6 ), 几乎所有的营养根均被外生菌根侵染(图3 ), 没有观测到这3个树种的外生菌根侵染频率在两个样带中有显著差异。
图6 试验地外生菌根树种枫香树、柳杉、青冈在竹-针阔竞争界面两侧的菌根侵染频率对比(平均值±标准误差)。BFT, 竹-林过渡带; CBF, 针阔混交林。相同小写字母表示同一树种在不同样带的侵染频率差异不显著(p < 0.1)。
Fig. 6 Comparison of frequency of the ectomycorrhizal fungi root colonization in Liquidambar formosana, Cryptomeria fortune, Cyclobalanopsis glauca at two sides of bamboo and broad-leaved forest interface in the study site (mean ± SE ). BFT, bamboo-forest transition; CBF, coniferous and broad- leaved mixed forest. Same lowercase letter denotes insignificant difference between the same species in different stands (p < 0.1).
2.4 毛竹侵入过程中其自身内生菌根量的变化
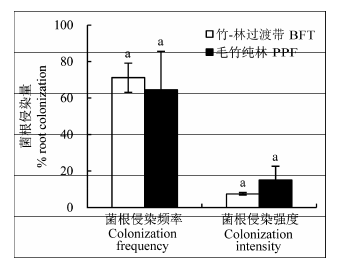
对毛竹的丛枝菌根侵染情况的观测中, 我们发现毛竹丛枝菌根侵染频率较高, 达到60%-70% (图7 ), 但显著低于其他树种。丛枝菌根真菌侵染强度弱, 表现在受侵染的根皮层细胞比例低, 且多为菌丝(图8 ), 总体明显弱于其他树种, 这也许表明毛竹对菌根的依赖程度较低。在竹-林过渡带和毛竹纯林两个类型的样带中, 毛竹根的丛枝菌根真菌侵染频率和侵染强度没有显著差异(图7 )。
图7 试验地毛竹在竹-林过渡带、毛竹纯林的菌根侵染频率和强度对比(平均值±标准误差)。BFT, 竹-林过渡带; PPF, 毛竹纯林。相同小写字母表示同一树种在不同样带的侵染频率差异不显著(p < 0.1)。
Fig. 7 Comparison of frequency and intensity of the arbuscular mycorrhizal fungi root colonization of Phyllostachys heterocycla ‘Pubescens’ at coniferous and broad-leaved mixed forest and coniferous and broad-leaved mixed forest in the study site (mean ± SE ). BFT, bamboo-forest transition; PPF, Phyllostachys heterocycla ‘Pubescens’ forest. Same lowercase letter denotes insignificant difference between the same species in different stands (p < 0.1).
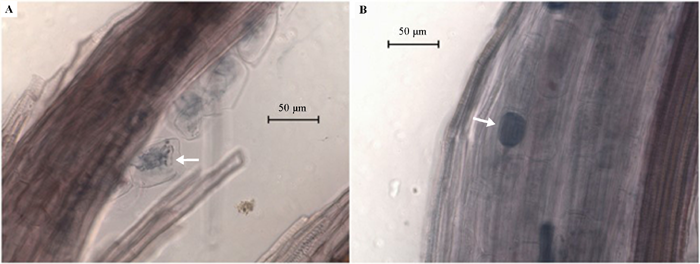
图8 毛竹丛枝菌根形态(墨水染色)。A, 箭头所示为胞内的丛枝结构。B, 箭头所示为根内孢子。
Fig. 8 Morphology of arbuscular mycorrhiza of Phyllostachys heterocycla ‘Pubescens’ stained with ink. A, Arrow indicates arbuscules in the root cells. B, Arrow indicates a fungal spore in a bamboo root.
3 讨论
3.1 土壤参数的比较
在毛竹纯林和竹-林过渡带, 近距毛竹的土壤含水率显著低于远离毛竹的土壤含水率。有研究认为毛竹高强度增殖和生长需要吸收大量的水(周文伟, 1991 ); 此外, 野外观测到毛竹根系密集于植株附近, 我们推断毛竹对土壤水的影响应多限于根系范围内及附近, 我们的数据支持这一推论。毛竹纯林、竹-林过渡带和天然针阔混交林样带的土壤参数中, 虽然土壤铵态氮和硝态氮含量在不同的样带中表现出一定的差异, 但竹-林过渡带和针阔混交林的土壤铵态氮和硝态氮含量没有显著差异, 有效氮总量在3个样带类型中也并未表现出显著差异, 变化趋势似乎不足以成为毛竹扩张导致其取代天然次生针阔混交林的主要原因或机制。或者说, 可能是由于植被变化而导致土壤养分变化。孰因孰果, 需要进一步研究探讨。
土壤无机氮的组分比例及其含量反映了土壤氮矿化与植物吸收的净结果。植被物种组成变化和植被类型的转变会通过不同的凋落物质量和养分利用效率而影响土壤氮的“矿化-固化”以及植被对氮的吸收效力, 从而反映到土壤无机氮的组分比例及其含量(李贵才等, 2001 )。有研究表明毛竹林向常绿阔叶林扩张会增加土壤总碳、总氮含量(吴家森等, 2008 ; 宋庆妮等, 2013 )。本研究结果显示, 毛竹林扩张使针阔混交林迅速转变为竹-林过渡带、毛竹纯林, 土壤有效氮总量有一定程度的增加。宋庆妮等(2013)研究发现, 毛竹林向常绿阔叶林扩张, 改变了土壤氮素矿化过程和格局, 导致竹-林过渡带土壤氨化作用增强, 硝化和总矿化作用减弱。本研究中竹-林过渡带硝态氮含量高于天然针阔混交林的结果(图4 )支持 毛竹林扩张将改变原有林分土壤氮素矿化过程和 格局。这也许是由于上层林木的衰亡而导致的林下土壤有机物腐解条件(光、温、水)改善, 利于土壤硝化过程。这在林冠郁闭的森林生态系统受到干扰后(林冠天窗下)常常出现(Bormann & Likens, 1979 )。然而, 毛竹纯林内的光、温条件亦优于林冠郁闭的次生针阔混交林, 而土壤硝态氮含量甚至显著低于针阔混交林(图4 )。我们认为毛竹林内土壤水含量的降低可能有部分原因, 但也有可能是毛竹在不同生长时期对无机氮的吸收偏爱不同。有研究认为植物生长早期一般喜好铵态氮, 而后期则喜好硝态氮(李生秀, 2008 )。竹-林过渡带的竹株是毛竹林扩张的分布前沿, 多处于幼竹阶段, 可能对铵态氮需求量大, 导致硝态氮积累。毛竹纯林以成熟毛竹居多, 对硝态氮的较大需求量使毛竹纯林土壤中硝态氮显著低于竹-林过渡带和针阔混交林。此外, 竹-林过渡带有最高的土壤pH值(竹-林过渡带>毛竹纯林>针阔混交林, 图4 ), 一般认为, 高pH值促进氮矿化, 尤其是硝化作用随pH值增加而线性增加(李贵才等, 2001 ), 本研究得出的竹-林过渡带土壤硝态氮积累的结果支持此说法。但也有一些研究显示竹-林过渡带的土壤pH值要低于常绿阔叶林的(宋庆妮等, 2013 )。
三个林带所处的地形条件有较大差异, 竹-林过渡带处于谷底, 不可否认存在由于地形差异所导致的土壤参数的差异。由于本研究地条件有限, 难以找到相同地形条件下的不同林带进行对照采样比较, 本研究无法对此进行验证。
3.2 菌根侵染频率与侵染强度
本研究中, 不论是有毛竹在近旁生长, 还是毛竹在一定距离之外, 竹-林过渡带的林木菌根感染率均未见有显著变化; 但是在感染强度方面, 较之针阔混交林, 具丛枝菌根的各树种在竹-林过渡带均表现出一定程度的增强(图5 )。这与基于毛竹扩张到天然林会导致原有林木与菌根真菌共生关系变弱的假定所推论的林木菌根真菌感染频率下降、感染强度降低的假设有相当大的距离。因而, 毛竹扩张过程中林木的菌根共生关系减弱, 进而使林木失活、林下幼苗无法更新的推理未得到验证。这里需要讨论两方面的问题, 尽管目前的数据并不足以完全回答它们: 其一, 毛竹扩张到天然混交林是否会导致原有的菌根系统紊乱?其二, 竹-林过渡带内林木丛枝菌根真菌感染强度增加的原因是什么?是否是病理性或是代偿性增生?
菌根真菌向宿主植物转运限制性资源(如磷)以获取有机碳源的回报(Fitter & Moyersoen, 1996 ; Johnson & Graham, 2013 ), 因此, 相对于快速生长的植物, 生长缓慢的植物有可能表现出更多的菌根共生效应, 因为后者更容易遇到局部养分枯竭的问题(Smith et al ., 2003 )。过去的很多研究都发现菌根真菌在低磷的土壤中增殖更快(Smith & Smith, 2013 )。在低磷条件下, 丛枝菌根真菌为维持其生存, 可以抑制根系对磷的直接吸收途径, 提高丛枝菌根途径提供磷占总吸收磷的比例(Smith & Smith, 2013 )。有关毛竹林扩张界面土壤资源的有效性的研究结果显示, 毛竹在侵入过程中会从土壤中吸收大量的磷、钾(吴家森等, 2008 ), 导致周边植物可利用的有效磷和钾含量大幅降低, 其中土壤中有效磷含量对毛竹粗生长有重要的影响(黎祖尧, 1993 )。在根系可吸收磷越来越少的情况下, 植物可能会通过调节资源分配, 投入更多的资源于菌根上, 此时菌根真菌感染强度的增加属于代偿性增加。本研究中没有测量土壤速效磷含量及其他相关指标, 因此还需要在增加土壤数据的基础上进行深入研究。
菌根真菌和宿主植物互利共生形成菌根, 两者都可以从矿质养分和有机碳源的相互交换中获益, 但菌根共生体并非总是对植物有利(Aerts, 2002 )。菌根对植物的作用存在积极(共生)、中立、消极(寄生) 3种, 且这3种作用被认为是取决于菌根真菌在共生关系中的成本-收益(提供给植物的土壤资源-分配给真菌的光合产物)的权衡。在共生收益高于成本时, 菌根真菌和宿主植物是积极的共生关系; 在共生收益低于成本时, 菌根真菌和植物之间可以是消极的寄生关系(Johnson et al ., 1997 )。毛竹林的扩张进入导致周边植物逐渐失活枯萎, 植物的光合能力也可能受到极大的影响, 植物从自身适合度考虑, 会减少对真菌的光合产物供给, 碳成为比土壤养分更重要的限制资源, 这可能打破植物-菌根真菌间原来的成本-收益格局, 使得菌根真菌付出的成本上升, 当成本高于收益时, 菌根可能会采取“欺骗者(cheater)”策略(Johnson et al ., 1997 ), 变共生为寄生, 此时菌根真菌感染强度的增加可能属于病理性增加。在实验过程中我们发现, 相比针阔混交林健康的根系, 竹-林过渡带的林木根系变衰弱, 甚至出现病态, 这可能是菌根真菌寄生后产生的反应。至于在本研究系统中这一过程是否是主导机制, 尚需进一步研究探讨。
3.3 毛竹扩张取代天然混交林林木机制的探讨
本研究结果不支持菌根减少假说, 因此关于毛竹林扩张进入周边常绿阔叶林及针阔混交林, 扩张导致周边林木死亡并抑制林下幼苗更新的机制可能需要从其他方面寻找。目前, 在化感方面尚无新的发现。现有的研究结果和数据支持在生长的局部范围内毛竹是一个“超级竞争者”的观点。毛竹扩张通过无性繁殖, 母株为子株生长提供大量养分、碳水化合物的支持, 使得子竹在短期内从竹笋伸长至林冠层而不被遮阴(《中国森林》编辑委员会, 2000)。子竹的根系同时迅速生长, 在局域土壤内形成高密度的吸收根。新子竹长成后产生新的竹鞭向外扩张。因此, 一片毛竹林实际上可能就是由一株竹衍生而来的, 竹鞭为其树干, 竹林为其树枝。旺盛的生理活动和繁殖, 高速度的生长, 使地上竹林形成良好的郁闭, 控制了地上的空间; 又使地下的竹鞭形成了网络状的水平郁闭。有研究表明毛竹林的细根生物量可达常绿阔叶林的5倍以上(刘骏等, 2013 ), 且毛竹细根分布趋于水、肥、热、气条件良好的上、中层土壤(92%以上的竹鞭分布在0-40 cm土层间)(蔡亮等, 2003 ), 毛竹林细根的比根长、生长速率和周转率均高于阔叶林(刘骏等, 2013 )。竹鞭穿行能力极强, 毛竹林逐年外扩, 而其他植物很难扩入其内。即使毛竹林立竹全部砍光, 残留在土壤中的竹鞭仍能自我复壮, 抽出新株, 使竹林逐步恢复, 而不被其他植物所更替(张春生等, 2007 )。这种超强的生长特点是其他任何木本植物所不具备的, 这让毛竹在自然成林方面具有了无法超越的优势。我们认为毛竹这种独特的生态特性值得关注和深入研究。
致谢 感谢北京师范大学生命科学学院陈红梅、刘晓雪同学, 浙江农林大学林业与生物技术学院陈娟同学在本研究工作中给予的帮助。
作者声明没有竞争性利益冲突.
参考文献
文献选项
[35]
Wu JS Jiang PK Wang ZL 2008 ). The effects of Phyllostachys pubescens expansion on soil fertility in national nature reserve of Mount Tianmu.
Acta Agriculturae Universitatis Jiangxiensis , 30 , 689 -692 .(in Chinese with English abstract)
[本文引用: 3]
[吴家森 , 姜培坤 , 王祖良 (2008 ). 天目山国家级自然保护区毛竹扩张对林地土壤肥力的影响
. 江西农业大学学报 , 30 , 689 -692 .]
[本文引用: 3]
[36]
Yang H Li PX Dai HT Liu D Yao XS 2010 ). Effects of Phyllostachys pubescens expansion on plant species diversity in Jigong Mountain and discussion of control measures.
Journal of Xinyang Normal University Natural Science Edition , 23 , 553 -557 .(in Chinese with English abstract)
[本文引用: 2]
[杨怀 , 李培学 , 戴慧堂 , 刘丹 , 姚贤胜 (2010 ). 鸡公山毛竹扩张对植物多样性的影响及控制措施
. 信阳师范学院学报(自然科学版) , 23 , 553 -557 .]
[本文引用: 2]
[37]
Yang QP Wang B Guo QR Zhao GD Fang K Liu YQ 2011 ). Effects of Phyllostachys edulis expansion on carbon storage of evergreen broad-leaved forest in Dagangshan Mountain, Jiangxi.
Acta Agriculturae Universitatis Jiangxiensis , 33 , 529 -536 .(in Chinese with English abstract)
[本文引用: 1]
[杨清培 , 王兵 , 郭起荣 , 赵广东 , 方楷 , 刘苑秋 (2011 ). 大岗山毛竹扩张对常绿阔叶林生态系统碳储特征的影响
. 江西农业大学学报 , 33 , 529 -536 .]
[本文引用: 1]
[38]
Zhang CS Chen JH Zhu F 2007 ). Survey and analysis on development law in Phyllostachys heterocycla cv. Pubescens.
Nonwood Forest Research , 25 (4 ), 74 -76 .(in Chinese with English abstract)
[本文引用: 2]
[张春生 , 陈建华 , 朱凡 (2007 ). 毛竹生长发育规律的调查分析
. 经济林研究 , 25 (4 ), 74 -76 .]
[本文引用: 2]
[39]
Zheng YS Hong W 1998 ). A study on age structure model of bamboo stand and its application.
Scientia Silvae Sinicae , 34 (3 ), 34 -41 .(in Chinese with English abstract)
[本文引用: 1]
[郑郁善 , 洪伟 (1998 ). 毛竹林丰产年龄结构模型与应用研究
. 林业科学 , 34 (3 ), 34 -41 .]
[本文引用: 1]
[40]
Zhou WW 1991 ). An analysis of the influence of precipitation on the growth of bamboo forest.
Journal of Bamboo Research , 10 (2 ), 33 -39 .(in Chinese with English abstract)
[本文引用: 1]
[周文伟 (1991 ). 降水对毛竹林生长的影响分析
. 竹子研究汇刊 , 10 (2 ), 33 -39 .
[本文引用: 1]
[41]
Zhu CL Shangguan LP 2009 ). Preliminary study on the influence of the expansion edge on biodiversity of moso bamboo forest in Jinggang Mountain.
Territory & Natural Resources Study , (3 ), 45 -46 .(in Chinese with English abstract)
[本文引用: 1]
[朱长龙 , 上官林平 (2009 ). 井冈山毛竹林扩边对生物多样性的影响初探
. 国土与自然资源研究 , (3 ), 45 -46 .]
[本文引用: 1]
[1]
Aerts R 2002). The role of various types of mycorrhizal fungi in nutrient cycling and plant competition. In: van der Heijden MGA, Sanders IR eds. Mycorrhizal Ecology . Springer-Verlag , Berlin . 117 -133 .
[本文引用: 1]
[2]
Bai SB Zhou GM Wang YX Liang QQ Chen J Cheng YY Shen R 2013 ). Allelopathic potential of Phyllostachys edulis on two dominant tree species of evergreen broad-leaved forest in its invasive process.
Environmental Science , 34 , 4066 -4072 .(in Chinese with English abstract)
[本文引用: 1]
[白尚斌 , 周国模 , 王懿祥 , 梁倩倩 , 陈娟 , 程艳艳 , 沈蕊 (2013 ). 毛竹入侵对常绿阔叶林主要树种的化感作用研究
. 环境科学 , 34 , 4066 -4072 .]
[本文引用: 1]
[3]
Bao SD 2000 ). Soil and Agricultural Chemistry Analysis. 3rd edn . China Agriculture Press , Beijing .(in Chinese)
[本文引用: 1]
[鲍士旦 (2000 ). 土壤农化分析. 第三版 . 中国农业出版社 , 北京 .]
[本文引用: 1]
[4]
Becklin KM Pallo ML Galen C 2012 ). Willows indirectly reduce arbuscular mycorrhizal fungal colonization in understorey communities.
Journal of Ecology , 100 , 343 -351 .
[本文引用: 1]
[5]
Bormann FH Likens GE 1979 ). Pattern and Process in a Forested Ecosystem
. Springer-Verlag, New York.
[本文引用: 1]
[6]
Cai L Zhang RL Li CF Ding Y 2003 ). A method to inhibit the expansion of Phyllostachys pubescens stands based on the analysis of underground rhizome.
Journal of Northeast Forestry University , 31 (5 ), 68 -70 .(in Chinese with English abstract)
[本文引用: 2]
[蔡亮 , 张瑞霖 , 李春福 , 丁滪 (2003 ). 基于竹鞭状态分析的抑制毛竹林扩散的方法
. 东北林业大学学报 , 31 (5 ), 68 -70 .]
[本文引用: 2]
[7]
Chen XX Chen LQ 1983 ). Investigation on mycorrhizae of main tree species my in subtropical area of China.
Forest Science and Technology , (5 ), 8 -12 .(in Chinese)
[本文引用: 3]
[陈祥欣 , 陈连庆 (1983 ). 我国亚热带主要树种菌根调查
. 林业科技通讯 , (5 ), 8 -12 .]
[本文引用: 3]
[8]
Chou CH Yang CM 1982 ). Allelopathic research of subtropical vegetation in Taiwan II. Comparative exclusion of understory by Phyllostachys edulis and Cryptomeria japonica.
Journal of Chemical Ecology , 8 , 1489 -1507 .
[本文引用: 1]
[9]
Smith FA Smith SE Timonen (2003 ). Mycorrhizas. In: de Kroon H, Visser EJW eds. Root Ecology . Springer-Verlag , Berlin . 257 -295 .
[10]
Ding LX Wang ZL Zhou GM Du QZ 2006 ). Monitoring Phyllostachys pubescens stands expansion in national nature reserve of Mount Tianmu by remote sensing.
Journal of Zhejiang Forestry College , 23 , 297 -300 .(in Chinese with English abstract)
[本文引用: 2]
[丁丽霞 , 王祖良 , 周国模 , 杜晴洲 (2006 ). 天目山国家级自然保护区毛竹林扩张遥感监测
. 浙江林学院学报 , 23 , 297 -300 .]
[本文引用: 2]
[11]
Dong M 1996 ). Plant clonal growth in heterogeneous habitats: risk-spreading.
Acta Phytoecologica Sinica , 20 , 543 -548 .(in Chinese with English abstract)
[本文引用: 1]
[董鸣 (1996 ). 异质性生境中的植物克隆生长: 风险分摊
. 植物生态学报 , 20 , 543 -548 .]
[本文引用: 1]
[12]
Fitter AH Moyersoen B 1996 ). Evolutionary trends in root-microbe symbioses.
Philosophical Transactions of the Royal Society of London Series B-Biological Sciences , 351 , 1367 -1375 .
[本文引用: 1]
[13]
Gong MQ Chen YL Zhong CL 1997 ). Mycorrhizal Research and Application . China Forestry Publishing House , Beijing .(in Chinese)
[本文引用: 1]
[弓明钦 , 陈应龙 , 仲崇禄 (1997 ). 菌根研究及应用 . 中国林业出版社 , 北京 .]
[本文引用: 1]
[14]
Hong W Hu XS Wu CZ Yan SJ Feng L Lin YM 2004 ). Comparison study on community structure features of the mixed forest of Phyllostachys pubescences in Fujian Province.
Journal of Plant Resources and Environment , 13 (1 ), 37 -42 .(in Chinese with English abstract)
[本文引用: 1]
[洪伟 , 胡喜生 , 吴承祯 , 闫淑君 , 封磊 , 林勇明 (2004 ). 福建省毛竹混交林群落结构特征的比较
. 植物资源与环境学报 , 13 (1 ), 37 -42 .]
[本文引用: 1]
[15]
Huang QT 2008 ). Effect of leaf and root allelochemicals of Phyllostachys heterocycla cv. pubescens on germination of Chinese fir seed.
Journal of Fujian Forestry Science and Technology , 35 (2 ), 75 -77 .(in Chinese with English abstract)
[本文引用: 1]
[黄启堂 (2008 ). 毛竹叶及其竹鞭生化物质对杉木种子的发芽效应
. 福建林业科技 , 35 (2 ), 75 -77 .]
[本文引用: 1]
[16]
Johnson NC Graham JH 2013 ). The continuum concept remains a useful framework for studying mycorrhizal functioning.
Plant and Soil , 363 , 411 -419 .
[本文引用: 1]
[17]
Johnson NC Graham JH Smith FA 1997 ). Functioning of mycorrhizal associations along the mutualism-parasitism continuum.
New Phytologist , 135 , 575 -585 .
[本文引用: 2]
[18]
Li GC Han XG Huang JH Tang JW 2001 ). A review of affecting factors of soil nitrogen mineralization in forest ecosystems.
Acta Ecologica Sinica , 21 , 1187 -1195 .(in Chinese with English abstract)
[本文引用: 2]
[李贵才 , 韩兴国 , 黄建辉 , 唐建维 (2001 ). 森林生态系统土壤氮矿化影响因素研究进展
. 生态学报 , 21 , 1187 -1195 .]
[本文引用: 2]
[19]
Li SX 2008 ). Soil and Plant Nitrogen in Dryland Areas of China . Science Press , Beijing .(in Chinese)
[本文引用: 1]
[李生秀 (2008 ). 中国旱地土壤植物氮素 . 科学出版社 , 北京 .]
[本文引用: 1]
[20]
Li ZY 1993 ). Study on effect of soil on diameter eye-high growth of Phyllostachys pubescens.
Journal of Bamboo Research , 12 (3 ), 29 -36 .(in Chinese with English abstract)
[本文引用: 1]
[黎祖尧 (1993 ). 土壤对毛竹眉径生长影响的研究
. 竹子研究汇刊 , 12 (3 ), 29 -36 .]
[本文引用: 1]
[21]
Liang QQ Bai SB Zhou GM Wang YX Liao J 2012 ). Effect of aqueous extracts of Phyllostachys heterocycla cv. Pubescens on seed germination and seedling growth of Festuca arundinacea.
Acta Agriculturae Zhejiangensis , 24 , 434 -439 .(in Chinese with English abstract)
[本文引用: 1]
[梁倩倩 , 白尚斌 , 周国模 , 王懿祥 , 廖娟 (2012 ). 毛竹浸提液对高羊茅种子萌发及幼苗生长的影响
. 浙江农业学报 , 24 , 434 -439 .]
[本文引用: 1]
[22]
Liu J Yang QP Song QN Yu DK Yang GY Qi HY Shi JM 2013 ). Strategy of fine root expansion of Phyllostachys pubescens population into evergreen broad-leaved forest.
Chinese Journal of Plant Ecology , 37 , 230 -238 .(in Chinese with English abstract)
[本文引用: 2]
[刘骏 , 杨清培 , 宋庆妮 , 余定坤 , 杨光耀 , 祁红艳 , 施建敏 (2013 ). 毛竹种群向常绿阔叶林扩张的细根策略
. 植物生态学报 , 37 , 230 -238 .]
[本文引用: 2]
[23]
Liu S 2010 ). The Influence of the Changes of Light Environment on the Other Species in Phyllostachys edulis Stands. Master degree dissertation, Zhejiang A&F University,
Lin’an , Zhejiang . 24 -51 .(in Chinese with English abstract)
[本文引用: 2]
[刘烁 (2010 ). 毛竹蔓延过程中林内光环境变化对其他树种的影响
. 硕士学位论文, 浙江农林大学, 浙江临安 . 24 -51 .]
[本文引用: 2]
[24]
Okutomi K Shinoda S Fukuda H 1996 ). Causal analysis of the invasion of broad-leaved forest by bamboo in Japan.
Journal of Vegetation Science , 7 , 723 -728 .
[本文引用: 1]
[25]
Smith FA Smith SE 2013 ). How useful is the mutualism- parasitism continuum of arbuscular mycorrhizal functioning?
Plant and Soil , 363 , 7 -18 .
[本文引用: 3]
[26]
Song QN Yang QP Liu J Yu DK Fang K Xu P He YJ 2013 ). Effects of Phyllostachys edulis expansion on soil nitrogen mineralization and its availability in evergreen broadleaf forest.
Chinese Journal of Applied Ecology , 24 , 338 -344 .(in Chinese with English abstract)
[本文引用: 3]
[宋庆妮 , 杨清培 , 刘骏 , 余定坤 , 方楷 , 徐佩 , 何宇娟 (2013 ). 毛竹扩张对常绿阔叶林土壤氮素矿化及有效性的影响
. 应用生态学报 , 24 , 338 -344 .]
[本文引用: 3]
[27]
Su LY Cheng AX Yu AL Fu WQ Zheng PY 1992 ). Investigation on mycorrhizae of forest trees in Natural Reserve of Mount Tianmu.
Journal of Zhejiang Forestry College , 9 , 263 -276 .(in Chinese with English abstract)
[本文引用: 4]
[苏琍英 , 程爱兴 , 喻爱林 , 傅卫庆 , 郑平谣 (1992 ). 天目山自然保护区林木菌根调查
. 浙江林学院学报 , 9 , 263 -276 .]
[本文引用: 4]
[28]
Suzuki S Nakagoshi N 2008 ). Expansion of bamboo forests caused by reduced bamboo-shoot harvest under different natural and artificial conditions.
Ecological Research , 23 , 641 -647 .
[本文引用: 1]
[29]
Suzuki T Nakatsubo T 2001 ). Impact of the bamboo Phyllostachys bambusoides on the light environment and plant communities on riverbanks.
Journal of Forest Research , 6 , 81 -86 .
[本文引用: 1]
[30]
The Editorial Board of Forest in China (2000 ). Forest in China Vol. 4 . China Forestry Publishing House , Beijing .(in Chinese)
[《中国森林》编辑委员会 (2000 ). 中国森林. 第4卷 . 中国林业出版社出版 , 北京.]
[31]
Trouvelot A Kough JL Gianinazzi-Pearson V 1986). Mesure du taux de mycorhization VA d’un systeme radiculaire. Recherche de methodes d’estimation ayant une signification fonctionnelle. In: Gianinazzi-Pearson V, Gianinazzi S eds. Physiological and Genetical Aspects of Mycorrhizae . INRA Press , Paris . 217 -221 .
[本文引用: 1]
[32]
Vierheilig H Coughlan AP Wyss U Piché Y 1998 ). Ink and vinegar, a simple staining technique for arbuscular- mycorrhizal fungi.
Applied and Environmental Microbiology , 64 , 5004 -5007 .
[本文引用: 1]
[33]
Vogelsang KM Bever JD 2009 ). Mycorrhizal densities decline in association with nonnative plants and contribute to plant invasion.
Ecology , 90 , 399 -407 .
[本文引用: 2]
[34]
Widden P 2001 ). The use of glycerin jelly for mounting stained roots for the observation and quantification of endomycorrhizal fungi.
Mycologia , 93 , 1026 -1027 .
[本文引用: 1]
天目山国家级自然保护区毛竹扩张对林地土壤肥力的影响
3
2008
... 国内外一些学者从人类活动(蔡亮等, 2003 ; Suzuki & Nakagoshi, 2008 )、生长特性(董鸣, 1996 ; Okutomi et al ., 1996 )、土壤资源(张春生等, 2007 ; 黄启堂, 2008 ; 吴家森等, 2008 ; 杨怀等, 2010 ; 宋庆妮等, 2013 )、光环境(Suzuki & Nakatsubo, 2001 ; 刘烁, 2010 )和化感作用(Chou & Yang, 1982 ; 梁倩倩等, 2012 ; 白尚斌等, 2013 )等方面对毛竹林扩张形成纯林的原因进行了分析探讨, 但有关毛竹林扩张导致其他林木失活枯死并抑制林下幼苗更新的机制, 目前尚有争议. ...
... 土壤无机氮的组分比例及其含量反映了土壤氮矿化与植物吸收的净结果.植被物种组成变化和植被类型的转变会通过不同的凋落物质量和养分利用效率而影响土壤氮的“矿化-固化”以及植被对氮的吸收效力, 从而反映到土壤无机氮的组分比例及其含量(李贵才等, 2001 ).有研究表明毛竹林向常绿阔叶林扩张会增加土壤总碳、总氮含量(吴家森等, 2008 ; 宋庆妮等, 2013 ).本研究结果显示, 毛竹林扩张使针阔混交林迅速转变为竹-林过渡带、毛竹纯林, 土壤有效氮总量有一定程度的增加.宋庆妮等(2013)研究发现, 毛竹林向常绿阔叶林扩张, 改变了土壤氮素矿化过程和格局, 导致竹-林过渡带土壤氨化作用增强, 硝化和总矿化作用减弱.本研究中竹-林过渡带硝态氮含量高于天然针阔混交林的结果(图4 )支持 毛竹林扩张将改变原有林分土壤氮素矿化过程和 格局.这也许是由于上层林木的衰亡而导致的林下土壤有机物腐解条件(光、温、水)改善, 利于土壤硝化过程.这在林冠郁闭的森林生态系统受到干扰后(林冠天窗下)常常出现(Bormann & Likens, 1979 ).然而, 毛竹纯林内的光、温条件亦优于林冠郁闭的次生针阔混交林, 而土壤硝态氮含量甚至显著低于针阔混交林(图4 ).我们认为毛竹林内土壤水含量的降低可能有部分原因, 但也有可能是毛竹在不同生长时期对无机氮的吸收偏爱不同.有研究认为植物生长早期一般喜好铵态氮, 而后期则喜好硝态氮(李生秀, 2008 ).竹-林过渡带的竹株是毛竹林扩张的分布前沿, 多处于幼竹阶段, 可能对铵态氮需求量大, 导致硝态氮积累.毛竹纯林以成熟毛竹居多, 对硝态氮的较大需求量使毛竹纯林土壤中硝态氮显著低于竹-林过渡带和针阔混交林.此外, 竹-林过渡带有最高的土壤pH值(竹-林过渡带>毛竹纯林>针阔混交林, 图4 ), 一般认为, 高pH值促进氮矿化, 尤其是硝化作用随pH值增加而线性增加(李贵才等, 2001 ), 本研究得出的竹-林过渡带土壤硝态氮积累的结果支持此说法.但也有一些研究显示竹-林过渡带的土壤pH值要低于常绿阔叶林的(宋庆妮等, 2013 ). ...
... 菌根真菌向宿主植物转运限制性资源(如磷)以获取有机碳源的回报(Fitter & Moyersoen, 1996 ; Johnson & Graham, 2013 ), 因此, 相对于快速生长的植物, 生长缓慢的植物有可能表现出更多的菌根共生效应, 因为后者更容易遇到局部养分枯竭的问题(Smith et al ., 2003 ).过去的很多研究都发现菌根真菌在低磷的土壤中增殖更快(Smith & Smith, 2013 ).在低磷条件下, 丛枝菌根真菌为维持其生存, 可以抑制根系对磷的直接吸收途径, 提高丛枝菌根途径提供磷占总吸收磷的比例(Smith & Smith, 2013 ).有关毛竹林扩张界面土壤资源的有效性的研究结果显示, 毛竹在侵入过程中会从土壤中吸收大量的磷、钾(吴家森等, 2008 ), 导致周边植物可利用的有效磷和钾含量大幅降低, 其中土壤中有效磷含量对毛竹粗生长有重要的影响(黎祖尧, 1993 ).在根系可吸收磷越来越少的情况下, 植物可能会通过调节资源分配, 投入更多的资源于菌根上, 此时菌根真菌感染强度的增加属于代偿性增加.本研究中没有测量土壤速效磷含量及其他相关指标, 因此还需要在增加土壤数据的基础上进行深入研究. ...
天目山国家级自然保护区毛竹扩张对林地土壤肥力的影响
3
2008
... 国内外一些学者从人类活动(蔡亮等, 2003 ; Suzuki & Nakagoshi, 2008 )、生长特性(董鸣, 1996 ; Okutomi et al ., 1996 )、土壤资源(张春生等, 2007 ; 黄启堂, 2008 ; 吴家森等, 2008 ; 杨怀等, 2010 ; 宋庆妮等, 2013 )、光环境(Suzuki & Nakatsubo, 2001 ; 刘烁, 2010 )和化感作用(Chou & Yang, 1982 ; 梁倩倩等, 2012 ; 白尚斌等, 2013 )等方面对毛竹林扩张形成纯林的原因进行了分析探讨, 但有关毛竹林扩张导致其他林木失活枯死并抑制林下幼苗更新的机制, 目前尚有争议. ...
... 土壤无机氮的组分比例及其含量反映了土壤氮矿化与植物吸收的净结果.植被物种组成变化和植被类型的转变会通过不同的凋落物质量和养分利用效率而影响土壤氮的“矿化-固化”以及植被对氮的吸收效力, 从而反映到土壤无机氮的组分比例及其含量(李贵才等, 2001 ).有研究表明毛竹林向常绿阔叶林扩张会增加土壤总碳、总氮含量(吴家森等, 2008 ; 宋庆妮等, 2013 ).本研究结果显示, 毛竹林扩张使针阔混交林迅速转变为竹-林过渡带、毛竹纯林, 土壤有效氮总量有一定程度的增加.宋庆妮等(2013)研究发现, 毛竹林向常绿阔叶林扩张, 改变了土壤氮素矿化过程和格局, 导致竹-林过渡带土壤氨化作用增强, 硝化和总矿化作用减弱.本研究中竹-林过渡带硝态氮含量高于天然针阔混交林的结果(图4 )支持 毛竹林扩张将改变原有林分土壤氮素矿化过程和 格局.这也许是由于上层林木的衰亡而导致的林下土壤有机物腐解条件(光、温、水)改善, 利于土壤硝化过程.这在林冠郁闭的森林生态系统受到干扰后(林冠天窗下)常常出现(Bormann & Likens, 1979 ).然而, 毛竹纯林内的光、温条件亦优于林冠郁闭的次生针阔混交林, 而土壤硝态氮含量甚至显著低于针阔混交林(图4 ).我们认为毛竹林内土壤水含量的降低可能有部分原因, 但也有可能是毛竹在不同生长时期对无机氮的吸收偏爱不同.有研究认为植物生长早期一般喜好铵态氮, 而后期则喜好硝态氮(李生秀, 2008 ).竹-林过渡带的竹株是毛竹林扩张的分布前沿, 多处于幼竹阶段, 可能对铵态氮需求量大, 导致硝态氮积累.毛竹纯林以成熟毛竹居多, 对硝态氮的较大需求量使毛竹纯林土壤中硝态氮显著低于竹-林过渡带和针阔混交林.此外, 竹-林过渡带有最高的土壤pH值(竹-林过渡带>毛竹纯林>针阔混交林, 图4 ), 一般认为, 高pH值促进氮矿化, 尤其是硝化作用随pH值增加而线性增加(李贵才等, 2001 ), 本研究得出的竹-林过渡带土壤硝态氮积累的结果支持此说法.但也有一些研究显示竹-林过渡带的土壤pH值要低于常绿阔叶林的(宋庆妮等, 2013 ). ...
... 菌根真菌向宿主植物转运限制性资源(如磷)以获取有机碳源的回报(Fitter & Moyersoen, 1996 ; Johnson & Graham, 2013 ), 因此, 相对于快速生长的植物, 生长缓慢的植物有可能表现出更多的菌根共生效应, 因为后者更容易遇到局部养分枯竭的问题(Smith et al ., 2003 ).过去的很多研究都发现菌根真菌在低磷的土壤中增殖更快(Smith & Smith, 2013 ).在低磷条件下, 丛枝菌根真菌为维持其生存, 可以抑制根系对磷的直接吸收途径, 提高丛枝菌根途径提供磷占总吸收磷的比例(Smith & Smith, 2013 ).有关毛竹林扩张界面土壤资源的有效性的研究结果显示, 毛竹在侵入过程中会从土壤中吸收大量的磷、钾(吴家森等, 2008 ), 导致周边植物可利用的有效磷和钾含量大幅降低, 其中土壤中有效磷含量对毛竹粗生长有重要的影响(黎祖尧, 1993 ).在根系可吸收磷越来越少的情况下, 植物可能会通过调节资源分配, 投入更多的资源于菌根上, 此时菌根真菌感染强度的增加属于代偿性增加.本研究中没有测量土壤速效磷含量及其他相关指标, 因此还需要在增加土壤数据的基础上进行深入研究. ...
鸡公山毛竹扩张对植物多样性的影响及控制措施
2
2010
... 毛竹(Phyllostachys heterocycla ‘Pubescens’)原产于我国暖温带和亚热带地区, 是我国南方重要的笋、材两用竹种(郑郁善和洪伟, 1998 ).毛竹经济和生态效益极高, 栽培历史悠久(杨怀等, 2010 ), 对毛竹的研究主要集中在如何获取更大的经济利益上(刘烁, 2010 ), 而对其生态方面的研究较少.近年发现, 许多地区的毛竹林扩张并取代了周围的针、阔叶森林(洪伟等, 2004 ; 丁丽霞等, 2006 ; 朱长龙和上官林平, 2009 ; 杨清培等, 2011 ).毛竹具有强大的克隆性生长繁育及扩鞭能力, 极易向周围的常绿阔叶林、针阔混交林蔓延扩张, 蚕食其他针阔树种, 导致周边林木失活枯死并抑制林下幼苗更新, 最终转变为毛竹纯林. ...
... 国内外一些学者从人类活动(蔡亮等, 2003 ; Suzuki & Nakagoshi, 2008 )、生长特性(董鸣, 1996 ; Okutomi et al ., 1996 )、土壤资源(张春生等, 2007 ; 黄启堂, 2008 ; 吴家森等, 2008 ; 杨怀等, 2010 ; 宋庆妮等, 2013 )、光环境(Suzuki & Nakatsubo, 2001 ; 刘烁, 2010 )和化感作用(Chou & Yang, 1982 ; 梁倩倩等, 2012 ; 白尚斌等, 2013 )等方面对毛竹林扩张形成纯林的原因进行了分析探讨, 但有关毛竹林扩张导致其他林木失活枯死并抑制林下幼苗更新的机制, 目前尚有争议. ...
鸡公山毛竹扩张对植物多样性的影响及控制措施
2
2010
... 毛竹(Phyllostachys heterocycla ‘Pubescens’)原产于我国暖温带和亚热带地区, 是我国南方重要的笋、材两用竹种(郑郁善和洪伟, 1998 ).毛竹经济和生态效益极高, 栽培历史悠久(杨怀等, 2010 ), 对毛竹的研究主要集中在如何获取更大的经济利益上(刘烁, 2010 ), 而对其生态方面的研究较少.近年发现, 许多地区的毛竹林扩张并取代了周围的针、阔叶森林(洪伟等, 2004 ; 丁丽霞等, 2006 ; 朱长龙和上官林平, 2009 ; 杨清培等, 2011 ).毛竹具有强大的克隆性生长繁育及扩鞭能力, 极易向周围的常绿阔叶林、针阔混交林蔓延扩张, 蚕食其他针阔树种, 导致周边林木失活枯死并抑制林下幼苗更新, 最终转变为毛竹纯林. ...
... 国内外一些学者从人类活动(蔡亮等, 2003 ; Suzuki & Nakagoshi, 2008 )、生长特性(董鸣, 1996 ; Okutomi et al ., 1996 )、土壤资源(张春生等, 2007 ; 黄启堂, 2008 ; 吴家森等, 2008 ; 杨怀等, 2010 ; 宋庆妮等, 2013 )、光环境(Suzuki & Nakatsubo, 2001 ; 刘烁, 2010 )和化感作用(Chou & Yang, 1982 ; 梁倩倩等, 2012 ; 白尚斌等, 2013 )等方面对毛竹林扩张形成纯林的原因进行了分析探讨, 但有关毛竹林扩张导致其他林木失活枯死并抑制林下幼苗更新的机制, 目前尚有争议. ...
大岗山毛竹扩张对常绿阔叶林生态系统碳储特征的影响
1
2011
... 毛竹(Phyllostachys heterocycla ‘Pubescens’)原产于我国暖温带和亚热带地区, 是我国南方重要的笋、材两用竹种(郑郁善和洪伟, 1998 ).毛竹经济和生态效益极高, 栽培历史悠久(杨怀等, 2010 ), 对毛竹的研究主要集中在如何获取更大的经济利益上(刘烁, 2010 ), 而对其生态方面的研究较少.近年发现, 许多地区的毛竹林扩张并取代了周围的针、阔叶森林(洪伟等, 2004 ; 丁丽霞等, 2006 ; 朱长龙和上官林平, 2009 ; 杨清培等, 2011 ).毛竹具有强大的克隆性生长繁育及扩鞭能力, 极易向周围的常绿阔叶林、针阔混交林蔓延扩张, 蚕食其他针阔树种, 导致周边林木失活枯死并抑制林下幼苗更新, 最终转变为毛竹纯林. ...
大岗山毛竹扩张对常绿阔叶林生态系统碳储特征的影响
1
2011
... 毛竹(Phyllostachys heterocycla ‘Pubescens’)原产于我国暖温带和亚热带地区, 是我国南方重要的笋、材两用竹种(郑郁善和洪伟, 1998 ).毛竹经济和生态效益极高, 栽培历史悠久(杨怀等, 2010 ), 对毛竹的研究主要集中在如何获取更大的经济利益上(刘烁, 2010 ), 而对其生态方面的研究较少.近年发现, 许多地区的毛竹林扩张并取代了周围的针、阔叶森林(洪伟等, 2004 ; 丁丽霞等, 2006 ; 朱长龙和上官林平, 2009 ; 杨清培等, 2011 ).毛竹具有强大的克隆性生长繁育及扩鞭能力, 极易向周围的常绿阔叶林、针阔混交林蔓延扩张, 蚕食其他针阔树种, 导致周边林木失活枯死并抑制林下幼苗更新, 最终转变为毛竹纯林. ...
毛竹生长发育规律的调查分析
2
2007
... 国内外一些学者从人类活动(蔡亮等, 2003 ; Suzuki & Nakagoshi, 2008 )、生长特性(董鸣, 1996 ; Okutomi et al ., 1996 )、土壤资源(张春生等, 2007 ; 黄启堂, 2008 ; 吴家森等, 2008 ; 杨怀等, 2010 ; 宋庆妮等, 2013 )、光环境(Suzuki & Nakatsubo, 2001 ; 刘烁, 2010 )和化感作用(Chou & Yang, 1982 ; 梁倩倩等, 2012 ; 白尚斌等, 2013 )等方面对毛竹林扩张形成纯林的原因进行了分析探讨, 但有关毛竹林扩张导致其他林木失活枯死并抑制林下幼苗更新的机制, 目前尚有争议. ...
... 本研究结果不支持菌根减少假说, 因此关于毛竹林扩张进入周边常绿阔叶林及针阔混交林, 扩张导致周边林木死亡并抑制林下幼苗更新的机制可能需要从其他方面寻找.目前, 在化感方面尚无新的发现.现有的研究结果和数据支持在生长的局部范围内毛竹是一个“超级竞争者”的观点.毛竹扩张通过无性繁殖, 母株为子株生长提供大量养分、碳水化合物的支持, 使得子竹在短期内从竹笋伸长至林冠层而不被遮阴(《中国森林》编辑委员会, 2000).子竹的根系同时迅速生长, 在局域土壤内形成高密度的吸收根.新子竹长成后产生新的竹鞭向外扩张.因此, 一片毛竹林实际上可能就是由一株竹衍生而来的, 竹鞭为其树干, 竹林为其树枝.旺盛的生理活动和繁殖, 高速度的生长, 使地上竹林形成良好的郁闭, 控制了地上的空间; 又使地下的竹鞭形成了网络状的水平郁闭.有研究表明毛竹林的细根生物量可达常绿阔叶林的5倍以上(刘骏等, 2013 ), 且毛竹细根分布趋于水、肥、热、气条件良好的上、中层土壤(92%以上的竹鞭分布在0-40 cm土层间)(蔡亮等, 2003 ), 毛竹林细根的比根长、生长速率和周转率均高于阔叶林(刘骏等, 2013 ).竹鞭穿行能力极强, 毛竹林逐年外扩, 而其他植物很难扩入其内.即使毛竹林立竹全部砍光, 残留在土壤中的竹鞭仍能自我复壮, 抽出新株, 使竹林逐步恢复, 而不被其他植物所更替(张春生等, 2007 ).这种超强的生长特点是其他任何木本植物所不具备的, 这让毛竹在自然成林方面具有了无法超越的优势.我们认为毛竹这种独特的生态特性值得关注和深入研究. ...
毛竹生长发育规律的调查分析
2
2007
... 国内外一些学者从人类活动(蔡亮等, 2003 ; Suzuki & Nakagoshi, 2008 )、生长特性(董鸣, 1996 ; Okutomi et al ., 1996 )、土壤资源(张春生等, 2007 ; 黄启堂, 2008 ; 吴家森等, 2008 ; 杨怀等, 2010 ; 宋庆妮等, 2013 )、光环境(Suzuki & Nakatsubo, 2001 ; 刘烁, 2010 )和化感作用(Chou & Yang, 1982 ; 梁倩倩等, 2012 ; 白尚斌等, 2013 )等方面对毛竹林扩张形成纯林的原因进行了分析探讨, 但有关毛竹林扩张导致其他林木失活枯死并抑制林下幼苗更新的机制, 目前尚有争议. ...
... 本研究结果不支持菌根减少假说, 因此关于毛竹林扩张进入周边常绿阔叶林及针阔混交林, 扩张导致周边林木死亡并抑制林下幼苗更新的机制可能需要从其他方面寻找.目前, 在化感方面尚无新的发现.现有的研究结果和数据支持在生长的局部范围内毛竹是一个“超级竞争者”的观点.毛竹扩张通过无性繁殖, 母株为子株生长提供大量养分、碳水化合物的支持, 使得子竹在短期内从竹笋伸长至林冠层而不被遮阴(《中国森林》编辑委员会, 2000).子竹的根系同时迅速生长, 在局域土壤内形成高密度的吸收根.新子竹长成后产生新的竹鞭向外扩张.因此, 一片毛竹林实际上可能就是由一株竹衍生而来的, 竹鞭为其树干, 竹林为其树枝.旺盛的生理活动和繁殖, 高速度的生长, 使地上竹林形成良好的郁闭, 控制了地上的空间; 又使地下的竹鞭形成了网络状的水平郁闭.有研究表明毛竹林的细根生物量可达常绿阔叶林的5倍以上(刘骏等, 2013 ), 且毛竹细根分布趋于水、肥、热、气条件良好的上、中层土壤(92%以上的竹鞭分布在0-40 cm土层间)(蔡亮等, 2003 ), 毛竹林细根的比根长、生长速率和周转率均高于阔叶林(刘骏等, 2013 ).竹鞭穿行能力极强, 毛竹林逐年外扩, 而其他植物很难扩入其内.即使毛竹林立竹全部砍光, 残留在土壤中的竹鞭仍能自我复壮, 抽出新株, 使竹林逐步恢复, 而不被其他植物所更替(张春生等, 2007 ).这种超强的生长特点是其他任何木本植物所不具备的, 这让毛竹在自然成林方面具有了无法超越的优势.我们认为毛竹这种独特的生态特性值得关注和深入研究. ...
毛竹林丰产年龄结构模型与应用研究
1
1998
... 毛竹(Phyllostachys heterocycla ‘Pubescens’)原产于我国暖温带和亚热带地区, 是我国南方重要的笋、材两用竹种(郑郁善和洪伟, 1998 ).毛竹经济和生态效益极高, 栽培历史悠久(杨怀等, 2010 ), 对毛竹的研究主要集中在如何获取更大的经济利益上(刘烁, 2010 ), 而对其生态方面的研究较少.近年发现, 许多地区的毛竹林扩张并取代了周围的针、阔叶森林(洪伟等, 2004 ; 丁丽霞等, 2006 ; 朱长龙和上官林平, 2009 ; 杨清培等, 2011 ).毛竹具有强大的克隆性生长繁育及扩鞭能力, 极易向周围的常绿阔叶林、针阔混交林蔓延扩张, 蚕食其他针阔树种, 导致周边林木失活枯死并抑制林下幼苗更新, 最终转变为毛竹纯林. ...
毛竹林丰产年龄结构模型与应用研究
1
1998
... 毛竹(Phyllostachys heterocycla ‘Pubescens’)原产于我国暖温带和亚热带地区, 是我国南方重要的笋、材两用竹种(郑郁善和洪伟, 1998 ).毛竹经济和生态效益极高, 栽培历史悠久(杨怀等, 2010 ), 对毛竹的研究主要集中在如何获取更大的经济利益上(刘烁, 2010 ), 而对其生态方面的研究较少.近年发现, 许多地区的毛竹林扩张并取代了周围的针、阔叶森林(洪伟等, 2004 ; 丁丽霞等, 2006 ; 朱长龙和上官林平, 2009 ; 杨清培等, 2011 ).毛竹具有强大的克隆性生长繁育及扩鞭能力, 极易向周围的常绿阔叶林、针阔混交林蔓延扩张, 蚕食其他针阔树种, 导致周边林木失活枯死并抑制林下幼苗更新, 最终转变为毛竹纯林. ...
降水对毛竹林生长的影响分析
1
1991
... 在毛竹纯林和竹-林过渡带, 近距毛竹的土壤含水率显著低于远离毛竹的土壤含水率.有研究认为毛竹高强度增殖和生长需要吸收大量的水(周文伟, 1991 ); 此外, 野外观测到毛竹根系密集于植株附近, 我们推断毛竹对土壤水的影响应多限于根系范围内及附近, 我们的数据支持这一推论.毛竹纯林、竹-林过渡带和天然针阔混交林样带的土壤参数中, 虽然土壤铵态氮和硝态氮含量在不同的样带中表现出一定的差异, 但竹-林过渡带和针阔混交林的土壤铵态氮和硝态氮含量没有显著差异, 有效氮总量在3个样带类型中也并未表现出显著差异, 变化趋势似乎不足以成为毛竹扩张导致其取代天然次生针阔混交林的主要原因或机制.或者说, 可能是由于植被变化而导致土壤养分变化.孰因孰果, 需要进一步研究探讨. ...
降水对毛竹林生长的影响分析
1
1991
... 在毛竹纯林和竹-林过渡带, 近距毛竹的土壤含水率显著低于远离毛竹的土壤含水率.有研究认为毛竹高强度增殖和生长需要吸收大量的水(周文伟, 1991 ); 此外, 野外观测到毛竹根系密集于植株附近, 我们推断毛竹对土壤水的影响应多限于根系范围内及附近, 我们的数据支持这一推论.毛竹纯林、竹-林过渡带和天然针阔混交林样带的土壤参数中, 虽然土壤铵态氮和硝态氮含量在不同的样带中表现出一定的差异, 但竹-林过渡带和针阔混交林的土壤铵态氮和硝态氮含量没有显著差异, 有效氮总量在3个样带类型中也并未表现出显著差异, 变化趋势似乎不足以成为毛竹扩张导致其取代天然次生针阔混交林的主要原因或机制.或者说, 可能是由于植被变化而导致土壤养分变化.孰因孰果, 需要进一步研究探讨. ...
井冈山毛竹林扩边对生物多样性的影响初探
1
2009
... 毛竹(Phyllostachys heterocycla ‘Pubescens’)原产于我国暖温带和亚热带地区, 是我国南方重要的笋、材两用竹种(郑郁善和洪伟, 1998 ).毛竹经济和生态效益极高, 栽培历史悠久(杨怀等, 2010 ), 对毛竹的研究主要集中在如何获取更大的经济利益上(刘烁, 2010 ), 而对其生态方面的研究较少.近年发现, 许多地区的毛竹林扩张并取代了周围的针、阔叶森林(洪伟等, 2004 ; 丁丽霞等, 2006 ; 朱长龙和上官林平, 2009 ; 杨清培等, 2011 ).毛竹具有强大的克隆性生长繁育及扩鞭能力, 极易向周围的常绿阔叶林、针阔混交林蔓延扩张, 蚕食其他针阔树种, 导致周边林木失活枯死并抑制林下幼苗更新, 最终转变为毛竹纯林. ...
井冈山毛竹林扩边对生物多样性的影响初探
1
2009
... 毛竹(Phyllostachys heterocycla ‘Pubescens’)原产于我国暖温带和亚热带地区, 是我国南方重要的笋、材两用竹种(郑郁善和洪伟, 1998 ).毛竹经济和生态效益极高, 栽培历史悠久(杨怀等, 2010 ), 对毛竹的研究主要集中在如何获取更大的经济利益上(刘烁, 2010 ), 而对其生态方面的研究较少.近年发现, 许多地区的毛竹林扩张并取代了周围的针、阔叶森林(洪伟等, 2004 ; 丁丽霞等, 2006 ; 朱长龙和上官林平, 2009 ; 杨清培等, 2011 ).毛竹具有强大的克隆性生长繁育及扩鞭能力, 极易向周围的常绿阔叶林、针阔混交林蔓延扩张, 蚕食其他针阔树种, 导致周边林木失活枯死并抑制林下幼苗更新, 最终转变为毛竹纯林. ...
1
... 菌根真菌和宿主植物互利共生形成菌根, 两者都可以从矿质养分和有机碳源的相互交换中获益, 但菌根共生体并非总是对植物有利(Aerts, 2002 ).菌根对植物的作用存在积极(共生)、中立、消极(寄生) 3种, 且这3种作用被认为是取决于菌根真菌在共生关系中的成本-收益(提供给植物的土壤资源-分配给真菌的光合产物)的权衡.在共生收益高于成本时, 菌根真菌和宿主植物是积极的共生关系; 在共生收益低于成本时, 菌根真菌和植物之间可以是消极的寄生关系(Johnson et al ., 1997 ).毛竹林的扩张进入导致周边植物逐渐失活枯萎, 植物的光合能力也可能受到极大的影响, 植物从自身适合度考虑, 会减少对真菌的光合产物供给, 碳成为比土壤养分更重要的限制资源, 这可能打破植物-菌根真菌间原来的成本-收益格局, 使得菌根真菌付出的成本上升, 当成本高于收益时, 菌根可能会采取“欺骗者(cheater)”策略(Johnson et al ., 1997 ), 变共生为寄生, 此时菌根真菌感染强度的增加可能属于病理性增加.在实验过程中我们发现, 相比针阔混交林健康的根系, 竹-林过渡带的林木根系变衰弱, 甚至出现病态, 这可能是菌根真菌寄生后产生的反应.至于在本研究系统中这一过程是否是主导机制, 尚需进一步研究探讨. ...
毛竹入侵对常绿阔叶林主要树种的化感作用研究
1
2013
... 国内外一些学者从人类活动(蔡亮等, 2003 ; Suzuki & Nakagoshi, 2008 )、生长特性(董鸣, 1996 ; Okutomi et al ., 1996 )、土壤资源(张春生等, 2007 ; 黄启堂, 2008 ; 吴家森等, 2008 ; 杨怀等, 2010 ; 宋庆妮等, 2013 )、光环境(Suzuki & Nakatsubo, 2001 ; 刘烁, 2010 )和化感作用(Chou & Yang, 1982 ; 梁倩倩等, 2012 ; 白尚斌等, 2013 )等方面对毛竹林扩张形成纯林的原因进行了分析探讨, 但有关毛竹林扩张导致其他林木失活枯死并抑制林下幼苗更新的机制, 目前尚有争议. ...
毛竹入侵对常绿阔叶林主要树种的化感作用研究
1
2013
... 国内外一些学者从人类活动(蔡亮等, 2003 ; Suzuki & Nakagoshi, 2008 )、生长特性(董鸣, 1996 ; Okutomi et al ., 1996 )、土壤资源(张春生等, 2007 ; 黄启堂, 2008 ; 吴家森等, 2008 ; 杨怀等, 2010 ; 宋庆妮等, 2013 )、光环境(Suzuki & Nakatsubo, 2001 ; 刘烁, 2010 )和化感作用(Chou & Yang, 1982 ; 梁倩倩等, 2012 ; 白尚斌等, 2013 )等方面对毛竹林扩张形成纯林的原因进行了分析探讨, 但有关毛竹林扩张导致其他林木失活枯死并抑制林下幼苗更新的机制, 目前尚有争议. ...
1
2000
... 在毛竹纯林和竹-林过渡带, 在靠近毛竹茎基部30 cm处和离毛竹茎基部2 m以外分别随机选择5个样点, 用小铲取表层(0-10 cm)土壤50 g, 装入标记的自封袋.在针阔混交林样带随机采取表层土(0-10 cm)土样5份, 装袋后均放入便携冷藏箱(4 ℃左右)带回实验室进行土壤pH值、含水率和有效氮(铵态氮、硝态氮)含量等理化指标测定(鲍士旦, 2000 ). ...
1
2000
... 在毛竹纯林和竹-林过渡带, 在靠近毛竹茎基部30 cm处和离毛竹茎基部2 m以外分别随机选择5个样点, 用小铲取表层(0-10 cm)土壤50 g, 装入标记的自封袋.在针阔混交林样带随机采取表层土(0-10 cm)土样5份, 装袋后均放入便携冷藏箱(4 ℃左右)带回实验室进行土壤pH值、含水率和有效氮(铵态氮、硝态氮)含量等理化指标测定(鲍士旦, 2000 ). ...
Willows indirectly reduce arbuscular mycorrhizal fungal colonization in understorey communities.
1
2012
... 生物或非生物因素对外来入侵种的正反馈作用对外来生物的定植和传播有不可忽视的作用.其中, 植物和土壤的相互反馈在促进或抑制植物个体的更新上具有重要意义(Vogelsang & Bever, 2009 ).植物与土壤微生物形成强共生关系, 如菌根(mycorr- hiza), 干扰往往会削弱或破坏这种共生关系, 从而可能促进外来种的定植, 使原植物群落发生改变.如果退化的土壤微生物群落进一步抑制本地种的更新, 群落更替会加速(Vogelsang & Bever, 2009 ).菌根与植物共生现象的普遍性与其生态学功能的重要性, 使其容易成为寄主植物反馈调节的媒介.例如有研究发现, 在亚高山森林植被和高山草甸的过渡带, 柳树(Salix spp.)可以通过自身的枯落物及其外生菌根间接降低周围草本植物群落的丛枝菌根侵染, 进而可能改变菌根真菌在这一地区的分布, 这一改变又反过来影响草本植物群落的分布和生长(Becklin et al ., 2012 ). ...
Pattern and Process in a Forested Ecosystem
1
1979
... 土壤无机氮的组分比例及其含量反映了土壤氮矿化与植物吸收的净结果.植被物种组成变化和植被类型的转变会通过不同的凋落物质量和养分利用效率而影响土壤氮的“矿化-固化”以及植被对氮的吸收效力, 从而反映到土壤无机氮的组分比例及其含量(李贵才等, 2001 ).有研究表明毛竹林向常绿阔叶林扩张会增加土壤总碳、总氮含量(吴家森等, 2008 ; 宋庆妮等, 2013 ).本研究结果显示, 毛竹林扩张使针阔混交林迅速转变为竹-林过渡带、毛竹纯林, 土壤有效氮总量有一定程度的增加.宋庆妮等(2013)研究发现, 毛竹林向常绿阔叶林扩张, 改变了土壤氮素矿化过程和格局, 导致竹-林过渡带土壤氨化作用增强, 硝化和总矿化作用减弱.本研究中竹-林过渡带硝态氮含量高于天然针阔混交林的结果(图4 )支持 毛竹林扩张将改变原有林分土壤氮素矿化过程和 格局.这也许是由于上层林木的衰亡而导致的林下土壤有机物腐解条件(光、温、水)改善, 利于土壤硝化过程.这在林冠郁闭的森林生态系统受到干扰后(林冠天窗下)常常出现(Bormann & Likens, 1979 ).然而, 毛竹纯林内的光、温条件亦优于林冠郁闭的次生针阔混交林, 而土壤硝态氮含量甚至显著低于针阔混交林(图4 ).我们认为毛竹林内土壤水含量的降低可能有部分原因, 但也有可能是毛竹在不同生长时期对无机氮的吸收偏爱不同.有研究认为植物生长早期一般喜好铵态氮, 而后期则喜好硝态氮(李生秀, 2008 ).竹-林过渡带的竹株是毛竹林扩张的分布前沿, 多处于幼竹阶段, 可能对铵态氮需求量大, 导致硝态氮积累.毛竹纯林以成熟毛竹居多, 对硝态氮的较大需求量使毛竹纯林土壤中硝态氮显著低于竹-林过渡带和针阔混交林.此外, 竹-林过渡带有最高的土壤pH值(竹-林过渡带>毛竹纯林>针阔混交林, 图4 ), 一般认为, 高pH值促进氮矿化, 尤其是硝化作用随pH值增加而线性增加(李贵才等, 2001 ), 本研究得出的竹-林过渡带土壤硝态氮积累的结果支持此说法.但也有一些研究显示竹-林过渡带的土壤pH值要低于常绿阔叶林的(宋庆妮等, 2013 ). ...
基于竹鞭状态分析的抑制毛竹林扩散的方法
2
2003
... 国内外一些学者从人类活动(蔡亮等, 2003 ; Suzuki & Nakagoshi, 2008 )、生长特性(董鸣, 1996 ; Okutomi et al ., 1996 )、土壤资源(张春生等, 2007 ; 黄启堂, 2008 ; 吴家森等, 2008 ; 杨怀等, 2010 ; 宋庆妮等, 2013 )、光环境(Suzuki & Nakatsubo, 2001 ; 刘烁, 2010 )和化感作用(Chou & Yang, 1982 ; 梁倩倩等, 2012 ; 白尚斌等, 2013 )等方面对毛竹林扩张形成纯林的原因进行了分析探讨, 但有关毛竹林扩张导致其他林木失活枯死并抑制林下幼苗更新的机制, 目前尚有争议. ...
... 本研究结果不支持菌根减少假说, 因此关于毛竹林扩张进入周边常绿阔叶林及针阔混交林, 扩张导致周边林木死亡并抑制林下幼苗更新的机制可能需要从其他方面寻找.目前, 在化感方面尚无新的发现.现有的研究结果和数据支持在生长的局部范围内毛竹是一个“超级竞争者”的观点.毛竹扩张通过无性繁殖, 母株为子株生长提供大量养分、碳水化合物的支持, 使得子竹在短期内从竹笋伸长至林冠层而不被遮阴(《中国森林》编辑委员会, 2000).子竹的根系同时迅速生长, 在局域土壤内形成高密度的吸收根.新子竹长成后产生新的竹鞭向外扩张.因此, 一片毛竹林实际上可能就是由一株竹衍生而来的, 竹鞭为其树干, 竹林为其树枝.旺盛的生理活动和繁殖, 高速度的生长, 使地上竹林形成良好的郁闭, 控制了地上的空间; 又使地下的竹鞭形成了网络状的水平郁闭.有研究表明毛竹林的细根生物量可达常绿阔叶林的5倍以上(刘骏等, 2013 ), 且毛竹细根分布趋于水、肥、热、气条件良好的上、中层土壤(92%以上的竹鞭分布在0-40 cm土层间)(蔡亮等, 2003 ), 毛竹林细根的比根长、生长速率和周转率均高于阔叶林(刘骏等, 2013 ).竹鞭穿行能力极强, 毛竹林逐年外扩, 而其他植物很难扩入其内.即使毛竹林立竹全部砍光, 残留在土壤中的竹鞭仍能自我复壮, 抽出新株, 使竹林逐步恢复, 而不被其他植物所更替(张春生等, 2007 ).这种超强的生长特点是其他任何木本植物所不具备的, 这让毛竹在自然成林方面具有了无法超越的优势.我们认为毛竹这种独特的生态特性值得关注和深入研究. ...
基于竹鞭状态分析的抑制毛竹林扩散的方法
2
2003
... 国内外一些学者从人类活动(蔡亮等, 2003 ; Suzuki & Nakagoshi, 2008 )、生长特性(董鸣, 1996 ; Okutomi et al ., 1996 )、土壤资源(张春生等, 2007 ; 黄启堂, 2008 ; 吴家森等, 2008 ; 杨怀等, 2010 ; 宋庆妮等, 2013 )、光环境(Suzuki & Nakatsubo, 2001 ; 刘烁, 2010 )和化感作用(Chou & Yang, 1982 ; 梁倩倩等, 2012 ; 白尚斌等, 2013 )等方面对毛竹林扩张形成纯林的原因进行了分析探讨, 但有关毛竹林扩张导致其他林木失活枯死并抑制林下幼苗更新的机制, 目前尚有争议. ...
... 本研究结果不支持菌根减少假说, 因此关于毛竹林扩张进入周边常绿阔叶林及针阔混交林, 扩张导致周边林木死亡并抑制林下幼苗更新的机制可能需要从其他方面寻找.目前, 在化感方面尚无新的发现.现有的研究结果和数据支持在生长的局部范围内毛竹是一个“超级竞争者”的观点.毛竹扩张通过无性繁殖, 母株为子株生长提供大量养分、碳水化合物的支持, 使得子竹在短期内从竹笋伸长至林冠层而不被遮阴(《中国森林》编辑委员会, 2000).子竹的根系同时迅速生长, 在局域土壤内形成高密度的吸收根.新子竹长成后产生新的竹鞭向外扩张.因此, 一片毛竹林实际上可能就是由一株竹衍生而来的, 竹鞭为其树干, 竹林为其树枝.旺盛的生理活动和繁殖, 高速度的生长, 使地上竹林形成良好的郁闭, 控制了地上的空间; 又使地下的竹鞭形成了网络状的水平郁闭.有研究表明毛竹林的细根生物量可达常绿阔叶林的5倍以上(刘骏等, 2013 ), 且毛竹细根分布趋于水、肥、热、气条件良好的上、中层土壤(92%以上的竹鞭分布在0-40 cm土层间)(蔡亮等, 2003 ), 毛竹林细根的比根长、生长速率和周转率均高于阔叶林(刘骏等, 2013 ).竹鞭穿行能力极强, 毛竹林逐年外扩, 而其他植物很难扩入其内.即使毛竹林立竹全部砍光, 残留在土壤中的竹鞭仍能自我复壮, 抽出新株, 使竹林逐步恢复, 而不被其他植物所更替(张春生等, 2007 ).这种超强的生长特点是其他任何木本植物所不具备的, 这让毛竹在自然成林方面具有了无法超越的优势.我们认为毛竹这种独特的生态特性值得关注和深入研究. ...
我国亚热带主要树种菌根调查
3
1983
... 以往的菌根资源调查中, 对毛竹的菌根类型有内生(弓明钦等, 1997 ; 陈祥欣和陈连庆, 1983 )和外生(苏琍英等, 1992 )两种不同的说法.柳杉的菌根类型有内生(陈祥欣和陈连庆, 1983 )和外生(苏琍英等, 1992 )不同的描述.枫香树的菌根类型也有内外生(苏琍英等, 1992 )或外生的报道(陈祥欣和陈连庆, 1983 ).也有调查显示青冈是内外生菌根树种(苏琍英等, 1992 ).但在我们本研究区域的预实验研究中发现, 经根切片染色确定, 毛竹、杉木、毛柄连蕊茶、江浙山胡椒菌根是内生菌根(endomycorrhiza, EM)类型, 青冈、柳杉、枫香树菌根是外生菌根(ectomycorrhiza, ECM)类型. ...
... )两种不同的说法.柳杉的菌根类型有内生(陈祥欣和陈连庆, 1983 )和外生(苏琍英等, 1992 )不同的描述.枫香树的菌根类型也有内外生(苏琍英等, 1992 )或外生的报道(陈祥欣和陈连庆, 1983 ).也有调查显示青冈是内外生菌根树种(苏琍英等, 1992 ).但在我们本研究区域的预实验研究中发现, 经根切片染色确定, 毛竹、杉木、毛柄连蕊茶、江浙山胡椒菌根是内生菌根(endomycorrhiza, EM)类型, 青冈、柳杉、枫香树菌根是外生菌根(ectomycorrhiza, ECM)类型. ...
... )或外生的报道(陈祥欣和陈连庆, 1983 ).也有调查显示青冈是内外生菌根树种(苏琍英等, 1992 ).但在我们本研究区域的预实验研究中发现, 经根切片染色确定, 毛竹、杉木、毛柄连蕊茶、江浙山胡椒菌根是内生菌根(endomycorrhiza, EM)类型, 青冈、柳杉、枫香树菌根是外生菌根(ectomycorrhiza, ECM)类型. ...
我国亚热带主要树种菌根调查
3
1983
... 以往的菌根资源调查中, 对毛竹的菌根类型有内生(弓明钦等, 1997 ; 陈祥欣和陈连庆, 1983 )和外生(苏琍英等, 1992 )两种不同的说法.柳杉的菌根类型有内生(陈祥欣和陈连庆, 1983 )和外生(苏琍英等, 1992 )不同的描述.枫香树的菌根类型也有内外生(苏琍英等, 1992 )或外生的报道(陈祥欣和陈连庆, 1983 ).也有调查显示青冈是内外生菌根树种(苏琍英等, 1992 ).但在我们本研究区域的预实验研究中发现, 经根切片染色确定, 毛竹、杉木、毛柄连蕊茶、江浙山胡椒菌根是内生菌根(endomycorrhiza, EM)类型, 青冈、柳杉、枫香树菌根是外生菌根(ectomycorrhiza, ECM)类型. ...
... )两种不同的说法.柳杉的菌根类型有内生(陈祥欣和陈连庆, 1983 )和外生(苏琍英等, 1992 )不同的描述.枫香树的菌根类型也有内外生(苏琍英等, 1992 )或外生的报道(陈祥欣和陈连庆, 1983 ).也有调查显示青冈是内外生菌根树种(苏琍英等, 1992 ).但在我们本研究区域的预实验研究中发现, 经根切片染色确定, 毛竹、杉木、毛柄连蕊茶、江浙山胡椒菌根是内生菌根(endomycorrhiza, EM)类型, 青冈、柳杉、枫香树菌根是外生菌根(ectomycorrhiza, ECM)类型. ...
... )或外生的报道(陈祥欣和陈连庆, 1983 ).也有调查显示青冈是内外生菌根树种(苏琍英等, 1992 ).但在我们本研究区域的预实验研究中发现, 经根切片染色确定, 毛竹、杉木、毛柄连蕊茶、江浙山胡椒菌根是内生菌根(endomycorrhiza, EM)类型, 青冈、柳杉、枫香树菌根是外生菌根(ectomycorrhiza, ECM)类型. ...
Allelopathic research of subtropical vegetation in Taiwan II. Comparative exclusion of understory by Phyllostachys edulis and Cryptomeria japonica.
1
1982
... 国内外一些学者从人类活动(蔡亮等, 2003 ; Suzuki & Nakagoshi, 2008 )、生长特性(董鸣, 1996 ; Okutomi et al ., 1996 )、土壤资源(张春生等, 2007 ; 黄启堂, 2008 ; 吴家森等, 2008 ; 杨怀等, 2010 ; 宋庆妮等, 2013 )、光环境(Suzuki & Nakatsubo, 2001 ; 刘烁, 2010 )和化感作用(Chou & Yang, 1982 ; 梁倩倩等, 2012 ; 白尚斌等, 2013 )等方面对毛竹林扩张形成纯林的原因进行了分析探讨, 但有关毛竹林扩张导致其他林木失活枯死并抑制林下幼苗更新的机制, 目前尚有争议. ...
天目山国家级自然保护区毛竹林扩张遥感监测
2
2006
... 毛竹(Phyllostachys heterocycla ‘Pubescens’)原产于我国暖温带和亚热带地区, 是我国南方重要的笋、材两用竹种(郑郁善和洪伟, 1998 ).毛竹经济和生态效益极高, 栽培历史悠久(杨怀等, 2010 ), 对毛竹的研究主要集中在如何获取更大的经济利益上(刘烁, 2010 ), 而对其生态方面的研究较少.近年发现, 许多地区的毛竹林扩张并取代了周围的针、阔叶森林(洪伟等, 2004 ; 丁丽霞等, 2006 ; 朱长龙和上官林平, 2009 ; 杨清培等, 2011 ).毛竹具有强大的克隆性生长繁育及扩鞭能力, 极易向周围的常绿阔叶林、针阔混交林蔓延扩张, 蚕食其他针阔树种, 导致周边林木失活枯死并抑制林下幼苗更新, 最终转变为毛竹纯林. ...
... 本研究地点位于浙江省西北部临安市境内的天目山国家级自然保护区(30.31°-30.36° N, 119.40- 119.45° E), 属于亚热带季风气侯, 冬暖夏凉, 雨量充沛.自山麓禅源寺至主峰山顶仙人顶, 年平均气温为8.8-14.8 ℃, 最冷月平均气温-2.6-3.4 ℃, 极端最低气温-20.2 ℃, 最热月平均气温19.9-28.1 ℃, 极端最高气温38.2 ℃, ≥10 ℃积温2500-5100 ℃, 无霜期209-235天, 年降水量1390-1870 mm, 年太阳总辐射3770-4460 MJ·m-2 .土壤以红壤、黄壤、黄红壤、棕黄壤为主(丁丽霞等, 2006 ).近年来毛竹扩张并取代针阔混交林的现象大量出现, 大多发生于该保护区内低、中山区. ...
天目山国家级自然保护区毛竹林扩张遥感监测
2
2006
... 毛竹(Phyllostachys heterocycla ‘Pubescens’)原产于我国暖温带和亚热带地区, 是我国南方重要的笋、材两用竹种(郑郁善和洪伟, 1998 ).毛竹经济和生态效益极高, 栽培历史悠久(杨怀等, 2010 ), 对毛竹的研究主要集中在如何获取更大的经济利益上(刘烁, 2010 ), 而对其生态方面的研究较少.近年发现, 许多地区的毛竹林扩张并取代了周围的针、阔叶森林(洪伟等, 2004 ; 丁丽霞等, 2006 ; 朱长龙和上官林平, 2009 ; 杨清培等, 2011 ).毛竹具有强大的克隆性生长繁育及扩鞭能力, 极易向周围的常绿阔叶林、针阔混交林蔓延扩张, 蚕食其他针阔树种, 导致周边林木失活枯死并抑制林下幼苗更新, 最终转变为毛竹纯林. ...
... 本研究地点位于浙江省西北部临安市境内的天目山国家级自然保护区(30.31°-30.36° N, 119.40- 119.45° E), 属于亚热带季风气侯, 冬暖夏凉, 雨量充沛.自山麓禅源寺至主峰山顶仙人顶, 年平均气温为8.8-14.8 ℃, 最冷月平均气温-2.6-3.4 ℃, 极端最低气温-20.2 ℃, 最热月平均气温19.9-28.1 ℃, 极端最高气温38.2 ℃, ≥10 ℃积温2500-5100 ℃, 无霜期209-235天, 年降水量1390-1870 mm, 年太阳总辐射3770-4460 MJ·m-2 .土壤以红壤、黄壤、黄红壤、棕黄壤为主(丁丽霞等, 2006 ).近年来毛竹扩张并取代针阔混交林的现象大量出现, 大多发生于该保护区内低、中山区. ...
异质性生境中的植物克隆生长: 风险分摊
1
1996
... 国内外一些学者从人类活动(蔡亮等, 2003 ; Suzuki & Nakagoshi, 2008 )、生长特性(董鸣, 1996 ; Okutomi et al ., 1996 )、土壤资源(张春生等, 2007 ; 黄启堂, 2008 ; 吴家森等, 2008 ; 杨怀等, 2010 ; 宋庆妮等, 2013 )、光环境(Suzuki & Nakatsubo, 2001 ; 刘烁, 2010 )和化感作用(Chou & Yang, 1982 ; 梁倩倩等, 2012 ; 白尚斌等, 2013 )等方面对毛竹林扩张形成纯林的原因进行了分析探讨, 但有关毛竹林扩张导致其他林木失活枯死并抑制林下幼苗更新的机制, 目前尚有争议. ...
异质性生境中的植物克隆生长: 风险分摊
1
1996
... 国内外一些学者从人类活动(蔡亮等, 2003 ; Suzuki & Nakagoshi, 2008 )、生长特性(董鸣, 1996 ; Okutomi et al ., 1996 )、土壤资源(张春生等, 2007 ; 黄启堂, 2008 ; 吴家森等, 2008 ; 杨怀等, 2010 ; 宋庆妮等, 2013 )、光环境(Suzuki & Nakatsubo, 2001 ; 刘烁, 2010 )和化感作用(Chou & Yang, 1982 ; 梁倩倩等, 2012 ; 白尚斌等, 2013 )等方面对毛竹林扩张形成纯林的原因进行了分析探讨, 但有关毛竹林扩张导致其他林木失活枯死并抑制林下幼苗更新的机制, 目前尚有争议. ...
Evolutionary trends in root-microbe symbioses.
1
1996
... 菌根真菌向宿主植物转运限制性资源(如磷)以获取有机碳源的回报(Fitter & Moyersoen, 1996 ; Johnson & Graham, 2013 ), 因此, 相对于快速生长的植物, 生长缓慢的植物有可能表现出更多的菌根共生效应, 因为后者更容易遇到局部养分枯竭的问题(Smith et al ., 2003 ).过去的很多研究都发现菌根真菌在低磷的土壤中增殖更快(Smith & Smith, 2013 ).在低磷条件下, 丛枝菌根真菌为维持其生存, 可以抑制根系对磷的直接吸收途径, 提高丛枝菌根途径提供磷占总吸收磷的比例(Smith & Smith, 2013 ).有关毛竹林扩张界面土壤资源的有效性的研究结果显示, 毛竹在侵入过程中会从土壤中吸收大量的磷、钾(吴家森等, 2008 ), 导致周边植物可利用的有效磷和钾含量大幅降低, 其中土壤中有效磷含量对毛竹粗生长有重要的影响(黎祖尧, 1993 ).在根系可吸收磷越来越少的情况下, 植物可能会通过调节资源分配, 投入更多的资源于菌根上, 此时菌根真菌感染强度的增加属于代偿性增加.本研究中没有测量土壤速效磷含量及其他相关指标, 因此还需要在增加土壤数据的基础上进行深入研究. ...
1
1997
... 以往的菌根资源调查中, 对毛竹的菌根类型有内生(弓明钦等, 1997 ; 陈祥欣和陈连庆, 1983 )和外生(苏琍英等, 1992 )两种不同的说法.柳杉的菌根类型有内生(陈祥欣和陈连庆, 1983 )和外生(苏琍英等, 1992 )不同的描述.枫香树的菌根类型也有内外生(苏琍英等, 1992 )或外生的报道(陈祥欣和陈连庆, 1983 ).也有调查显示青冈是内外生菌根树种(苏琍英等, 1992 ).但在我们本研究区域的预实验研究中发现, 经根切片染色确定, 毛竹、杉木、毛柄连蕊茶、江浙山胡椒菌根是内生菌根(endomycorrhiza, EM)类型, 青冈、柳杉、枫香树菌根是外生菌根(ectomycorrhiza, ECM)类型. ...
1
1997
... 以往的菌根资源调查中, 对毛竹的菌根类型有内生(弓明钦等, 1997 ; 陈祥欣和陈连庆, 1983 )和外生(苏琍英等, 1992 )两种不同的说法.柳杉的菌根类型有内生(陈祥欣和陈连庆, 1983 )和外生(苏琍英等, 1992 )不同的描述.枫香树的菌根类型也有内外生(苏琍英等, 1992 )或外生的报道(陈祥欣和陈连庆, 1983 ).也有调查显示青冈是内外生菌根树种(苏琍英等, 1992 ).但在我们本研究区域的预实验研究中发现, 经根切片染色确定, 毛竹、杉木、毛柄连蕊茶、江浙山胡椒菌根是内生菌根(endomycorrhiza, EM)类型, 青冈、柳杉、枫香树菌根是外生菌根(ectomycorrhiza, ECM)类型. ...
福建省毛竹混交林群落结构特征的比较
1
2004
... 毛竹(Phyllostachys heterocycla ‘Pubescens’)原产于我国暖温带和亚热带地区, 是我国南方重要的笋、材两用竹种(郑郁善和洪伟, 1998 ).毛竹经济和生态效益极高, 栽培历史悠久(杨怀等, 2010 ), 对毛竹的研究主要集中在如何获取更大的经济利益上(刘烁, 2010 ), 而对其生态方面的研究较少.近年发现, 许多地区的毛竹林扩张并取代了周围的针、阔叶森林(洪伟等, 2004 ; 丁丽霞等, 2006 ; 朱长龙和上官林平, 2009 ; 杨清培等, 2011 ).毛竹具有强大的克隆性生长繁育及扩鞭能力, 极易向周围的常绿阔叶林、针阔混交林蔓延扩张, 蚕食其他针阔树种, 导致周边林木失活枯死并抑制林下幼苗更新, 最终转变为毛竹纯林. ...
福建省毛竹混交林群落结构特征的比较
1
2004
... 毛竹(Phyllostachys heterocycla ‘Pubescens’)原产于我国暖温带和亚热带地区, 是我国南方重要的笋、材两用竹种(郑郁善和洪伟, 1998 ).毛竹经济和生态效益极高, 栽培历史悠久(杨怀等, 2010 ), 对毛竹的研究主要集中在如何获取更大的经济利益上(刘烁, 2010 ), 而对其生态方面的研究较少.近年发现, 许多地区的毛竹林扩张并取代了周围的针、阔叶森林(洪伟等, 2004 ; 丁丽霞等, 2006 ; 朱长龙和上官林平, 2009 ; 杨清培等, 2011 ).毛竹具有强大的克隆性生长繁育及扩鞭能力, 极易向周围的常绿阔叶林、针阔混交林蔓延扩张, 蚕食其他针阔树种, 导致周边林木失活枯死并抑制林下幼苗更新, 最终转变为毛竹纯林. ...
毛竹叶及其竹鞭生化物质对杉木种子的发芽效应
1
2008
... 国内外一些学者从人类活动(蔡亮等, 2003 ; Suzuki & Nakagoshi, 2008 )、生长特性(董鸣, 1996 ; Okutomi et al ., 1996 )、土壤资源(张春生等, 2007 ; 黄启堂, 2008 ; 吴家森等, 2008 ; 杨怀等, 2010 ; 宋庆妮等, 2013 )、光环境(Suzuki & Nakatsubo, 2001 ; 刘烁, 2010 )和化感作用(Chou & Yang, 1982 ; 梁倩倩等, 2012 ; 白尚斌等, 2013 )等方面对毛竹林扩张形成纯林的原因进行了分析探讨, 但有关毛竹林扩张导致其他林木失活枯死并抑制林下幼苗更新的机制, 目前尚有争议. ...
毛竹叶及其竹鞭生化物质对杉木种子的发芽效应
1
2008
... 国内外一些学者从人类活动(蔡亮等, 2003 ; Suzuki & Nakagoshi, 2008 )、生长特性(董鸣, 1996 ; Okutomi et al ., 1996 )、土壤资源(张春生等, 2007 ; 黄启堂, 2008 ; 吴家森等, 2008 ; 杨怀等, 2010 ; 宋庆妮等, 2013 )、光环境(Suzuki & Nakatsubo, 2001 ; 刘烁, 2010 )和化感作用(Chou & Yang, 1982 ; 梁倩倩等, 2012 ; 白尚斌等, 2013 )等方面对毛竹林扩张形成纯林的原因进行了分析探讨, 但有关毛竹林扩张导致其他林木失活枯死并抑制林下幼苗更新的机制, 目前尚有争议. ...
The continuum concept remains a useful framework for studying mycorrhizal functioning.
1
2013
... 菌根真菌向宿主植物转运限制性资源(如磷)以获取有机碳源的回报(Fitter & Moyersoen, 1996 ; Johnson & Graham, 2013 ), 因此, 相对于快速生长的植物, 生长缓慢的植物有可能表现出更多的菌根共生效应, 因为后者更容易遇到局部养分枯竭的问题(Smith et al ., 2003 ).过去的很多研究都发现菌根真菌在低磷的土壤中增殖更快(Smith & Smith, 2013 ).在低磷条件下, 丛枝菌根真菌为维持其生存, 可以抑制根系对磷的直接吸收途径, 提高丛枝菌根途径提供磷占总吸收磷的比例(Smith & Smith, 2013 ).有关毛竹林扩张界面土壤资源的有效性的研究结果显示, 毛竹在侵入过程中会从土壤中吸收大量的磷、钾(吴家森等, 2008 ), 导致周边植物可利用的有效磷和钾含量大幅降低, 其中土壤中有效磷含量对毛竹粗生长有重要的影响(黎祖尧, 1993 ).在根系可吸收磷越来越少的情况下, 植物可能会通过调节资源分配, 投入更多的资源于菌根上, 此时菌根真菌感染强度的增加属于代偿性增加.本研究中没有测量土壤速效磷含量及其他相关指标, 因此还需要在增加土壤数据的基础上进行深入研究. ...
Functioning of mycorrhizal associations along the mutualism-parasitism continuum.
2
1997
... 菌根真菌和宿主植物互利共生形成菌根, 两者都可以从矿质养分和有机碳源的相互交换中获益, 但菌根共生体并非总是对植物有利(Aerts, 2002 ).菌根对植物的作用存在积极(共生)、中立、消极(寄生) 3种, 且这3种作用被认为是取决于菌根真菌在共生关系中的成本-收益(提供给植物的土壤资源-分配给真菌的光合产物)的权衡.在共生收益高于成本时, 菌根真菌和宿主植物是积极的共生关系; 在共生收益低于成本时, 菌根真菌和植物之间可以是消极的寄生关系(Johnson et al ., 1997 ).毛竹林的扩张进入导致周边植物逐渐失活枯萎, 植物的光合能力也可能受到极大的影响, 植物从自身适合度考虑, 会减少对真菌的光合产物供给, 碳成为比土壤养分更重要的限制资源, 这可能打破植物-菌根真菌间原来的成本-收益格局, 使得菌根真菌付出的成本上升, 当成本高于收益时, 菌根可能会采取“欺骗者(cheater)”策略(Johnson et al ., 1997 ), 变共生为寄生, 此时菌根真菌感染强度的增加可能属于病理性增加.在实验过程中我们发现, 相比针阔混交林健康的根系, 竹-林过渡带的林木根系变衰弱, 甚至出现病态, 这可能是菌根真菌寄生后产生的反应.至于在本研究系统中这一过程是否是主导机制, 尚需进一步研究探讨. ...
... ).毛竹林的扩张进入导致周边植物逐渐失活枯萎, 植物的光合能力也可能受到极大的影响, 植物从自身适合度考虑, 会减少对真菌的光合产物供给, 碳成为比土壤养分更重要的限制资源, 这可能打破植物-菌根真菌间原来的成本-收益格局, 使得菌根真菌付出的成本上升, 当成本高于收益时, 菌根可能会采取“欺骗者(cheater)”策略(Johnson et al ., 1997 ), 变共生为寄生, 此时菌根真菌感染强度的增加可能属于病理性增加.在实验过程中我们发现, 相比针阔混交林健康的根系, 竹-林过渡带的林木根系变衰弱, 甚至出现病态, 这可能是菌根真菌寄生后产生的反应.至于在本研究系统中这一过程是否是主导机制, 尚需进一步研究探讨. ...
森林生态系统土壤氮矿化影响因素研究进展
2
2001
... 土壤无机氮的组分比例及其含量反映了土壤氮矿化与植物吸收的净结果.植被物种组成变化和植被类型的转变会通过不同的凋落物质量和养分利用效率而影响土壤氮的“矿化-固化”以及植被对氮的吸收效力, 从而反映到土壤无机氮的组分比例及其含量(李贵才等, 2001 ).有研究表明毛竹林向常绿阔叶林扩张会增加土壤总碳、总氮含量(吴家森等, 2008 ; 宋庆妮等, 2013 ).本研究结果显示, 毛竹林扩张使针阔混交林迅速转变为竹-林过渡带、毛竹纯林, 土壤有效氮总量有一定程度的增加.宋庆妮等(2013)研究发现, 毛竹林向常绿阔叶林扩张, 改变了土壤氮素矿化过程和格局, 导致竹-林过渡带土壤氨化作用增强, 硝化和总矿化作用减弱.本研究中竹-林过渡带硝态氮含量高于天然针阔混交林的结果(图4 )支持 毛竹林扩张将改变原有林分土壤氮素矿化过程和 格局.这也许是由于上层林木的衰亡而导致的林下土壤有机物腐解条件(光、温、水)改善, 利于土壤硝化过程.这在林冠郁闭的森林生态系统受到干扰后(林冠天窗下)常常出现(Bormann & Likens, 1979 ).然而, 毛竹纯林内的光、温条件亦优于林冠郁闭的次生针阔混交林, 而土壤硝态氮含量甚至显著低于针阔混交林(图4 ).我们认为毛竹林内土壤水含量的降低可能有部分原因, 但也有可能是毛竹在不同生长时期对无机氮的吸收偏爱不同.有研究认为植物生长早期一般喜好铵态氮, 而后期则喜好硝态氮(李生秀, 2008 ).竹-林过渡带的竹株是毛竹林扩张的分布前沿, 多处于幼竹阶段, 可能对铵态氮需求量大, 导致硝态氮积累.毛竹纯林以成熟毛竹居多, 对硝态氮的较大需求量使毛竹纯林土壤中硝态氮显著低于竹-林过渡带和针阔混交林.此外, 竹-林过渡带有最高的土壤pH值(竹-林过渡带>毛竹纯林>针阔混交林, 图4 ), 一般认为, 高pH值促进氮矿化, 尤其是硝化作用随pH值增加而线性增加(李贵才等, 2001 ), 本研究得出的竹-林过渡带土壤硝态氮积累的结果支持此说法.但也有一些研究显示竹-林过渡带的土壤pH值要低于常绿阔叶林的(宋庆妮等, 2013 ). ...
... ), 一般认为, 高pH值促进氮矿化, 尤其是硝化作用随pH值增加而线性增加(李贵才等, 2001 ), 本研究得出的竹-林过渡带土壤硝态氮积累的结果支持此说法.但也有一些研究显示竹-林过渡带的土壤pH值要低于常绿阔叶林的(宋庆妮等, 2013 ). ...
森林生态系统土壤氮矿化影响因素研究进展
2
2001
... 土壤无机氮的组分比例及其含量反映了土壤氮矿化与植物吸收的净结果.植被物种组成变化和植被类型的转变会通过不同的凋落物质量和养分利用效率而影响土壤氮的“矿化-固化”以及植被对氮的吸收效力, 从而反映到土壤无机氮的组分比例及其含量(李贵才等, 2001 ).有研究表明毛竹林向常绿阔叶林扩张会增加土壤总碳、总氮含量(吴家森等, 2008 ; 宋庆妮等, 2013 ).本研究结果显示, 毛竹林扩张使针阔混交林迅速转变为竹-林过渡带、毛竹纯林, 土壤有效氮总量有一定程度的增加.宋庆妮等(2013)研究发现, 毛竹林向常绿阔叶林扩张, 改变了土壤氮素矿化过程和格局, 导致竹-林过渡带土壤氨化作用增强, 硝化和总矿化作用减弱.本研究中竹-林过渡带硝态氮含量高于天然针阔混交林的结果(图4 )支持 毛竹林扩张将改变原有林分土壤氮素矿化过程和 格局.这也许是由于上层林木的衰亡而导致的林下土壤有机物腐解条件(光、温、水)改善, 利于土壤硝化过程.这在林冠郁闭的森林生态系统受到干扰后(林冠天窗下)常常出现(Bormann & Likens, 1979 ).然而, 毛竹纯林内的光、温条件亦优于林冠郁闭的次生针阔混交林, 而土壤硝态氮含量甚至显著低于针阔混交林(图4 ).我们认为毛竹林内土壤水含量的降低可能有部分原因, 但也有可能是毛竹在不同生长时期对无机氮的吸收偏爱不同.有研究认为植物生长早期一般喜好铵态氮, 而后期则喜好硝态氮(李生秀, 2008 ).竹-林过渡带的竹株是毛竹林扩张的分布前沿, 多处于幼竹阶段, 可能对铵态氮需求量大, 导致硝态氮积累.毛竹纯林以成熟毛竹居多, 对硝态氮的较大需求量使毛竹纯林土壤中硝态氮显著低于竹-林过渡带和针阔混交林.此外, 竹-林过渡带有最高的土壤pH值(竹-林过渡带>毛竹纯林>针阔混交林, 图4 ), 一般认为, 高pH值促进氮矿化, 尤其是硝化作用随pH值增加而线性增加(李贵才等, 2001 ), 本研究得出的竹-林过渡带土壤硝态氮积累的结果支持此说法.但也有一些研究显示竹-林过渡带的土壤pH值要低于常绿阔叶林的(宋庆妮等, 2013 ). ...
... ), 一般认为, 高pH值促进氮矿化, 尤其是硝化作用随pH值增加而线性增加(李贵才等, 2001 ), 本研究得出的竹-林过渡带土壤硝态氮积累的结果支持此说法.但也有一些研究显示竹-林过渡带的土壤pH值要低于常绿阔叶林的(宋庆妮等, 2013 ). ...
1
2008
... 土壤无机氮的组分比例及其含量反映了土壤氮矿化与植物吸收的净结果.植被物种组成变化和植被类型的转变会通过不同的凋落物质量和养分利用效率而影响土壤氮的“矿化-固化”以及植被对氮的吸收效力, 从而反映到土壤无机氮的组分比例及其含量(李贵才等, 2001 ).有研究表明毛竹林向常绿阔叶林扩张会增加土壤总碳、总氮含量(吴家森等, 2008 ; 宋庆妮等, 2013 ).本研究结果显示, 毛竹林扩张使针阔混交林迅速转变为竹-林过渡带、毛竹纯林, 土壤有效氮总量有一定程度的增加.宋庆妮等(2013)研究发现, 毛竹林向常绿阔叶林扩张, 改变了土壤氮素矿化过程和格局, 导致竹-林过渡带土壤氨化作用增强, 硝化和总矿化作用减弱.本研究中竹-林过渡带硝态氮含量高于天然针阔混交林的结果(图4 )支持 毛竹林扩张将改变原有林分土壤氮素矿化过程和 格局.这也许是由于上层林木的衰亡而导致的林下土壤有机物腐解条件(光、温、水)改善, 利于土壤硝化过程.这在林冠郁闭的森林生态系统受到干扰后(林冠天窗下)常常出现(Bormann & Likens, 1979 ).然而, 毛竹纯林内的光、温条件亦优于林冠郁闭的次生针阔混交林, 而土壤硝态氮含量甚至显著低于针阔混交林(图4 ).我们认为毛竹林内土壤水含量的降低可能有部分原因, 但也有可能是毛竹在不同生长时期对无机氮的吸收偏爱不同.有研究认为植物生长早期一般喜好铵态氮, 而后期则喜好硝态氮(李生秀, 2008 ).竹-林过渡带的竹株是毛竹林扩张的分布前沿, 多处于幼竹阶段, 可能对铵态氮需求量大, 导致硝态氮积累.毛竹纯林以成熟毛竹居多, 对硝态氮的较大需求量使毛竹纯林土壤中硝态氮显著低于竹-林过渡带和针阔混交林.此外, 竹-林过渡带有最高的土壤pH值(竹-林过渡带>毛竹纯林>针阔混交林, 图4 ), 一般认为, 高pH值促进氮矿化, 尤其是硝化作用随pH值增加而线性增加(李贵才等, 2001 ), 本研究得出的竹-林过渡带土壤硝态氮积累的结果支持此说法.但也有一些研究显示竹-林过渡带的土壤pH值要低于常绿阔叶林的(宋庆妮等, 2013 ). ...
1
2008
... 土壤无机氮的组分比例及其含量反映了土壤氮矿化与植物吸收的净结果.植被物种组成变化和植被类型的转变会通过不同的凋落物质量和养分利用效率而影响土壤氮的“矿化-固化”以及植被对氮的吸收效力, 从而反映到土壤无机氮的组分比例及其含量(李贵才等, 2001 ).有研究表明毛竹林向常绿阔叶林扩张会增加土壤总碳、总氮含量(吴家森等, 2008 ; 宋庆妮等, 2013 ).本研究结果显示, 毛竹林扩张使针阔混交林迅速转变为竹-林过渡带、毛竹纯林, 土壤有效氮总量有一定程度的增加.宋庆妮等(2013)研究发现, 毛竹林向常绿阔叶林扩张, 改变了土壤氮素矿化过程和格局, 导致竹-林过渡带土壤氨化作用增强, 硝化和总矿化作用减弱.本研究中竹-林过渡带硝态氮含量高于天然针阔混交林的结果(图4 )支持 毛竹林扩张将改变原有林分土壤氮素矿化过程和 格局.这也许是由于上层林木的衰亡而导致的林下土壤有机物腐解条件(光、温、水)改善, 利于土壤硝化过程.这在林冠郁闭的森林生态系统受到干扰后(林冠天窗下)常常出现(Bormann & Likens, 1979 ).然而, 毛竹纯林内的光、温条件亦优于林冠郁闭的次生针阔混交林, 而土壤硝态氮含量甚至显著低于针阔混交林(图4 ).我们认为毛竹林内土壤水含量的降低可能有部分原因, 但也有可能是毛竹在不同生长时期对无机氮的吸收偏爱不同.有研究认为植物生长早期一般喜好铵态氮, 而后期则喜好硝态氮(李生秀, 2008 ).竹-林过渡带的竹株是毛竹林扩张的分布前沿, 多处于幼竹阶段, 可能对铵态氮需求量大, 导致硝态氮积累.毛竹纯林以成熟毛竹居多, 对硝态氮的较大需求量使毛竹纯林土壤中硝态氮显著低于竹-林过渡带和针阔混交林.此外, 竹-林过渡带有最高的土壤pH值(竹-林过渡带>毛竹纯林>针阔混交林, 图4 ), 一般认为, 高pH值促进氮矿化, 尤其是硝化作用随pH值增加而线性增加(李贵才等, 2001 ), 本研究得出的竹-林过渡带土壤硝态氮积累的结果支持此说法.但也有一些研究显示竹-林过渡带的土壤pH值要低于常绿阔叶林的(宋庆妮等, 2013 ). ...
土壤对毛竹眉径生长影响的研究
1
1993
... 菌根真菌向宿主植物转运限制性资源(如磷)以获取有机碳源的回报(Fitter & Moyersoen, 1996 ; Johnson & Graham, 2013 ), 因此, 相对于快速生长的植物, 生长缓慢的植物有可能表现出更多的菌根共生效应, 因为后者更容易遇到局部养分枯竭的问题(Smith et al ., 2003 ).过去的很多研究都发现菌根真菌在低磷的土壤中增殖更快(Smith & Smith, 2013 ).在低磷条件下, 丛枝菌根真菌为维持其生存, 可以抑制根系对磷的直接吸收途径, 提高丛枝菌根途径提供磷占总吸收磷的比例(Smith & Smith, 2013 ).有关毛竹林扩张界面土壤资源的有效性的研究结果显示, 毛竹在侵入过程中会从土壤中吸收大量的磷、钾(吴家森等, 2008 ), 导致周边植物可利用的有效磷和钾含量大幅降低, 其中土壤中有效磷含量对毛竹粗生长有重要的影响(黎祖尧, 1993 ).在根系可吸收磷越来越少的情况下, 植物可能会通过调节资源分配, 投入更多的资源于菌根上, 此时菌根真菌感染强度的增加属于代偿性增加.本研究中没有测量土壤速效磷含量及其他相关指标, 因此还需要在增加土壤数据的基础上进行深入研究. ...
土壤对毛竹眉径生长影响的研究
1
1993
... 菌根真菌向宿主植物转运限制性资源(如磷)以获取有机碳源的回报(Fitter & Moyersoen, 1996 ; Johnson & Graham, 2013 ), 因此, 相对于快速生长的植物, 生长缓慢的植物有可能表现出更多的菌根共生效应, 因为后者更容易遇到局部养分枯竭的问题(Smith et al ., 2003 ).过去的很多研究都发现菌根真菌在低磷的土壤中增殖更快(Smith & Smith, 2013 ).在低磷条件下, 丛枝菌根真菌为维持其生存, 可以抑制根系对磷的直接吸收途径, 提高丛枝菌根途径提供磷占总吸收磷的比例(Smith & Smith, 2013 ).有关毛竹林扩张界面土壤资源的有效性的研究结果显示, 毛竹在侵入过程中会从土壤中吸收大量的磷、钾(吴家森等, 2008 ), 导致周边植物可利用的有效磷和钾含量大幅降低, 其中土壤中有效磷含量对毛竹粗生长有重要的影响(黎祖尧, 1993 ).在根系可吸收磷越来越少的情况下, 植物可能会通过调节资源分配, 投入更多的资源于菌根上, 此时菌根真菌感染强度的增加属于代偿性增加.本研究中没有测量土壤速效磷含量及其他相关指标, 因此还需要在增加土壤数据的基础上进行深入研究. ...
毛竹浸提液对高羊茅种子萌发及幼苗生长的影响
1
2012
... 国内外一些学者从人类活动(蔡亮等, 2003 ; Suzuki & Nakagoshi, 2008 )、生长特性(董鸣, 1996 ; Okutomi et al ., 1996 )、土壤资源(张春生等, 2007 ; 黄启堂, 2008 ; 吴家森等, 2008 ; 杨怀等, 2010 ; 宋庆妮等, 2013 )、光环境(Suzuki & Nakatsubo, 2001 ; 刘烁, 2010 )和化感作用(Chou & Yang, 1982 ; 梁倩倩等, 2012 ; 白尚斌等, 2013 )等方面对毛竹林扩张形成纯林的原因进行了分析探讨, 但有关毛竹林扩张导致其他林木失活枯死并抑制林下幼苗更新的机制, 目前尚有争议. ...
毛竹浸提液对高羊茅种子萌发及幼苗生长的影响
1
2012
... 国内外一些学者从人类活动(蔡亮等, 2003 ; Suzuki & Nakagoshi, 2008 )、生长特性(董鸣, 1996 ; Okutomi et al ., 1996 )、土壤资源(张春生等, 2007 ; 黄启堂, 2008 ; 吴家森等, 2008 ; 杨怀等, 2010 ; 宋庆妮等, 2013 )、光环境(Suzuki & Nakatsubo, 2001 ; 刘烁, 2010 )和化感作用(Chou & Yang, 1982 ; 梁倩倩等, 2012 ; 白尚斌等, 2013 )等方面对毛竹林扩张形成纯林的原因进行了分析探讨, 但有关毛竹林扩张导致其他林木失活枯死并抑制林下幼苗更新的机制, 目前尚有争议. ...
毛竹种群向常绿阔叶林扩张的细根策略
2
2013
... 本研究结果不支持菌根减少假说, 因此关于毛竹林扩张进入周边常绿阔叶林及针阔混交林, 扩张导致周边林木死亡并抑制林下幼苗更新的机制可能需要从其他方面寻找.目前, 在化感方面尚无新的发现.现有的研究结果和数据支持在生长的局部范围内毛竹是一个“超级竞争者”的观点.毛竹扩张通过无性繁殖, 母株为子株生长提供大量养分、碳水化合物的支持, 使得子竹在短期内从竹笋伸长至林冠层而不被遮阴(《中国森林》编辑委员会, 2000).子竹的根系同时迅速生长, 在局域土壤内形成高密度的吸收根.新子竹长成后产生新的竹鞭向外扩张.因此, 一片毛竹林实际上可能就是由一株竹衍生而来的, 竹鞭为其树干, 竹林为其树枝.旺盛的生理活动和繁殖, 高速度的生长, 使地上竹林形成良好的郁闭, 控制了地上的空间; 又使地下的竹鞭形成了网络状的水平郁闭.有研究表明毛竹林的细根生物量可达常绿阔叶林的5倍以上(刘骏等, 2013 ), 且毛竹细根分布趋于水、肥、热、气条件良好的上、中层土壤(92%以上的竹鞭分布在0-40 cm土层间)(蔡亮等, 2003 ), 毛竹林细根的比根长、生长速率和周转率均高于阔叶林(刘骏等, 2013 ).竹鞭穿行能力极强, 毛竹林逐年外扩, 而其他植物很难扩入其内.即使毛竹林立竹全部砍光, 残留在土壤中的竹鞭仍能自我复壮, 抽出新株, 使竹林逐步恢复, 而不被其他植物所更替(张春生等, 2007 ).这种超强的生长特点是其他任何木本植物所不具备的, 这让毛竹在自然成林方面具有了无法超越的优势.我们认为毛竹这种独特的生态特性值得关注和深入研究. ...
... ), 毛竹林细根的比根长、生长速率和周转率均高于阔叶林(刘骏等, 2013 ).竹鞭穿行能力极强, 毛竹林逐年外扩, 而其他植物很难扩入其内.即使毛竹林立竹全部砍光, 残留在土壤中的竹鞭仍能自我复壮, 抽出新株, 使竹林逐步恢复, 而不被其他植物所更替(张春生等, 2007 ).这种超强的生长特点是其他任何木本植物所不具备的, 这让毛竹在自然成林方面具有了无法超越的优势.我们认为毛竹这种独特的生态特性值得关注和深入研究. ...
毛竹种群向常绿阔叶林扩张的细根策略
2
2013
... 本研究结果不支持菌根减少假说, 因此关于毛竹林扩张进入周边常绿阔叶林及针阔混交林, 扩张导致周边林木死亡并抑制林下幼苗更新的机制可能需要从其他方面寻找.目前, 在化感方面尚无新的发现.现有的研究结果和数据支持在生长的局部范围内毛竹是一个“超级竞争者”的观点.毛竹扩张通过无性繁殖, 母株为子株生长提供大量养分、碳水化合物的支持, 使得子竹在短期内从竹笋伸长至林冠层而不被遮阴(《中国森林》编辑委员会, 2000).子竹的根系同时迅速生长, 在局域土壤内形成高密度的吸收根.新子竹长成后产生新的竹鞭向外扩张.因此, 一片毛竹林实际上可能就是由一株竹衍生而来的, 竹鞭为其树干, 竹林为其树枝.旺盛的生理活动和繁殖, 高速度的生长, 使地上竹林形成良好的郁闭, 控制了地上的空间; 又使地下的竹鞭形成了网络状的水平郁闭.有研究表明毛竹林的细根生物量可达常绿阔叶林的5倍以上(刘骏等, 2013 ), 且毛竹细根分布趋于水、肥、热、气条件良好的上、中层土壤(92%以上的竹鞭分布在0-40 cm土层间)(蔡亮等, 2003 ), 毛竹林细根的比根长、生长速率和周转率均高于阔叶林(刘骏等, 2013 ).竹鞭穿行能力极强, 毛竹林逐年外扩, 而其他植物很难扩入其内.即使毛竹林立竹全部砍光, 残留在土壤中的竹鞭仍能自我复壮, 抽出新株, 使竹林逐步恢复, 而不被其他植物所更替(张春生等, 2007 ).这种超强的生长特点是其他任何木本植物所不具备的, 这让毛竹在自然成林方面具有了无法超越的优势.我们认为毛竹这种独特的生态特性值得关注和深入研究. ...
... ), 毛竹林细根的比根长、生长速率和周转率均高于阔叶林(刘骏等, 2013 ).竹鞭穿行能力极强, 毛竹林逐年外扩, 而其他植物很难扩入其内.即使毛竹林立竹全部砍光, 残留在土壤中的竹鞭仍能自我复壮, 抽出新株, 使竹林逐步恢复, 而不被其他植物所更替(张春生等, 2007 ).这种超强的生长特点是其他任何木本植物所不具备的, 这让毛竹在自然成林方面具有了无法超越的优势.我们认为毛竹这种独特的生态特性值得关注和深入研究. ...
毛竹蔓延过程中林内光环境变化对其他树种的影响
2
2010
... 毛竹(Phyllostachys heterocycla ‘Pubescens’)原产于我国暖温带和亚热带地区, 是我国南方重要的笋、材两用竹种(郑郁善和洪伟, 1998 ).毛竹经济和生态效益极高, 栽培历史悠久(杨怀等, 2010 ), 对毛竹的研究主要集中在如何获取更大的经济利益上(刘烁, 2010 ), 而对其生态方面的研究较少.近年发现, 许多地区的毛竹林扩张并取代了周围的针、阔叶森林(洪伟等, 2004 ; 丁丽霞等, 2006 ; 朱长龙和上官林平, 2009 ; 杨清培等, 2011 ).毛竹具有强大的克隆性生长繁育及扩鞭能力, 极易向周围的常绿阔叶林、针阔混交林蔓延扩张, 蚕食其他针阔树种, 导致周边林木失活枯死并抑制林下幼苗更新, 最终转变为毛竹纯林. ...
... 国内外一些学者从人类活动(蔡亮等, 2003 ; Suzuki & Nakagoshi, 2008 )、生长特性(董鸣, 1996 ; Okutomi et al ., 1996 )、土壤资源(张春生等, 2007 ; 黄启堂, 2008 ; 吴家森等, 2008 ; 杨怀等, 2010 ; 宋庆妮等, 2013 )、光环境(Suzuki & Nakatsubo, 2001 ; 刘烁, 2010 )和化感作用(Chou & Yang, 1982 ; 梁倩倩等, 2012 ; 白尚斌等, 2013 )等方面对毛竹林扩张形成纯林的原因进行了分析探讨, 但有关毛竹林扩张导致其他林木失活枯死并抑制林下幼苗更新的机制, 目前尚有争议. ...
毛竹蔓延过程中林内光环境变化对其他树种的影响
2
2010
... 毛竹(Phyllostachys heterocycla ‘Pubescens’)原产于我国暖温带和亚热带地区, 是我国南方重要的笋、材两用竹种(郑郁善和洪伟, 1998 ).毛竹经济和生态效益极高, 栽培历史悠久(杨怀等, 2010 ), 对毛竹的研究主要集中在如何获取更大的经济利益上(刘烁, 2010 ), 而对其生态方面的研究较少.近年发现, 许多地区的毛竹林扩张并取代了周围的针、阔叶森林(洪伟等, 2004 ; 丁丽霞等, 2006 ; 朱长龙和上官林平, 2009 ; 杨清培等, 2011 ).毛竹具有强大的克隆性生长繁育及扩鞭能力, 极易向周围的常绿阔叶林、针阔混交林蔓延扩张, 蚕食其他针阔树种, 导致周边林木失活枯死并抑制林下幼苗更新, 最终转变为毛竹纯林. ...
... 国内外一些学者从人类活动(蔡亮等, 2003 ; Suzuki & Nakagoshi, 2008 )、生长特性(董鸣, 1996 ; Okutomi et al ., 1996 )、土壤资源(张春生等, 2007 ; 黄启堂, 2008 ; 吴家森等, 2008 ; 杨怀等, 2010 ; 宋庆妮等, 2013 )、光环境(Suzuki & Nakatsubo, 2001 ; 刘烁, 2010 )和化感作用(Chou & Yang, 1982 ; 梁倩倩等, 2012 ; 白尚斌等, 2013 )等方面对毛竹林扩张形成纯林的原因进行了分析探讨, 但有关毛竹林扩张导致其他林木失活枯死并抑制林下幼苗更新的机制, 目前尚有争议. ...
Causal analysis of the invasion of broad-leaved forest by bamboo in Japan.
1
1996
... 国内外一些学者从人类活动(蔡亮等, 2003 ; Suzuki & Nakagoshi, 2008 )、生长特性(董鸣, 1996 ; Okutomi et al ., 1996 )、土壤资源(张春生等, 2007 ; 黄启堂, 2008 ; 吴家森等, 2008 ; 杨怀等, 2010 ; 宋庆妮等, 2013 )、光环境(Suzuki & Nakatsubo, 2001 ; 刘烁, 2010 )和化感作用(Chou & Yang, 1982 ; 梁倩倩等, 2012 ; 白尚斌等, 2013 )等方面对毛竹林扩张形成纯林的原因进行了分析探讨, 但有关毛竹林扩张导致其他林木失活枯死并抑制林下幼苗更新的机制, 目前尚有争议. ...
How useful is the mutualism- parasitism continuum of arbuscular mycorrhizal functioning?
3
2013
... 菌根真菌向宿主植物转运限制性资源(如磷)以获取有机碳源的回报(Fitter & Moyersoen, 1996 ; Johnson & Graham, 2013 ), 因此, 相对于快速生长的植物, 生长缓慢的植物有可能表现出更多的菌根共生效应, 因为后者更容易遇到局部养分枯竭的问题(Smith et al ., 2003 ).过去的很多研究都发现菌根真菌在低磷的土壤中增殖更快(Smith & Smith, 2013 ).在低磷条件下, 丛枝菌根真菌为维持其生存, 可以抑制根系对磷的直接吸收途径, 提高丛枝菌根途径提供磷占总吸收磷的比例(Smith & Smith, 2013 ).有关毛竹林扩张界面土壤资源的有效性的研究结果显示, 毛竹在侵入过程中会从土壤中吸收大量的磷、钾(吴家森等, 2008 ), 导致周边植物可利用的有效磷和钾含量大幅降低, 其中土壤中有效磷含量对毛竹粗生长有重要的影响(黎祖尧, 1993 ).在根系可吸收磷越来越少的情况下, 植物可能会通过调节资源分配, 投入更多的资源于菌根上, 此时菌根真菌感染强度的增加属于代偿性增加.本研究中没有测量土壤速效磷含量及其他相关指标, 因此还需要在增加土壤数据的基础上进行深入研究. ...
... ).过去的很多研究都发现菌根真菌在低磷的土壤中增殖更快(Smith & Smith, 2013 ).在低磷条件下, 丛枝菌根真菌为维持其生存, 可以抑制根系对磷的直接吸收途径, 提高丛枝菌根途径提供磷占总吸收磷的比例(Smith & Smith, 2013 ).有关毛竹林扩张界面土壤资源的有效性的研究结果显示, 毛竹在侵入过程中会从土壤中吸收大量的磷、钾(吴家森等, 2008 ), 导致周边植物可利用的有效磷和钾含量大幅降低, 其中土壤中有效磷含量对毛竹粗生长有重要的影响(黎祖尧, 1993 ).在根系可吸收磷越来越少的情况下, 植物可能会通过调节资源分配, 投入更多的资源于菌根上, 此时菌根真菌感染强度的增加属于代偿性增加.本研究中没有测量土壤速效磷含量及其他相关指标, 因此还需要在增加土壤数据的基础上进行深入研究. ...
... ).在低磷条件下, 丛枝菌根真菌为维持其生存, 可以抑制根系对磷的直接吸收途径, 提高丛枝菌根途径提供磷占总吸收磷的比例(Smith & Smith, 2013 ).有关毛竹林扩张界面土壤资源的有效性的研究结果显示, 毛竹在侵入过程中会从土壤中吸收大量的磷、钾(吴家森等, 2008 ), 导致周边植物可利用的有效磷和钾含量大幅降低, 其中土壤中有效磷含量对毛竹粗生长有重要的影响(黎祖尧, 1993 ).在根系可吸收磷越来越少的情况下, 植物可能会通过调节资源分配, 投入更多的资源于菌根上, 此时菌根真菌感染强度的增加属于代偿性增加.本研究中没有测量土壤速效磷含量及其他相关指标, 因此还需要在增加土壤数据的基础上进行深入研究. ...
毛竹扩张对常绿阔叶林土壤氮素矿化及有效性的影响
3
2013
... 国内外一些学者从人类活动(蔡亮等, 2003 ; Suzuki & Nakagoshi, 2008 )、生长特性(董鸣, 1996 ; Okutomi et al ., 1996 )、土壤资源(张春生等, 2007 ; 黄启堂, 2008 ; 吴家森等, 2008 ; 杨怀等, 2010 ; 宋庆妮等, 2013 )、光环境(Suzuki & Nakatsubo, 2001 ; 刘烁, 2010 )和化感作用(Chou & Yang, 1982 ; 梁倩倩等, 2012 ; 白尚斌等, 2013 )等方面对毛竹林扩张形成纯林的原因进行了分析探讨, 但有关毛竹林扩张导致其他林木失活枯死并抑制林下幼苗更新的机制, 目前尚有争议. ...
... 土壤无机氮的组分比例及其含量反映了土壤氮矿化与植物吸收的净结果.植被物种组成变化和植被类型的转变会通过不同的凋落物质量和养分利用效率而影响土壤氮的“矿化-固化”以及植被对氮的吸收效力, 从而反映到土壤无机氮的组分比例及其含量(李贵才等, 2001 ).有研究表明毛竹林向常绿阔叶林扩张会增加土壤总碳、总氮含量(吴家森等, 2008 ; 宋庆妮等, 2013 ).本研究结果显示, 毛竹林扩张使针阔混交林迅速转变为竹-林过渡带、毛竹纯林, 土壤有效氮总量有一定程度的增加.宋庆妮等(2013)研究发现, 毛竹林向常绿阔叶林扩张, 改变了土壤氮素矿化过程和格局, 导致竹-林过渡带土壤氨化作用增强, 硝化和总矿化作用减弱.本研究中竹-林过渡带硝态氮含量高于天然针阔混交林的结果(图4 )支持 毛竹林扩张将改变原有林分土壤氮素矿化过程和 格局.这也许是由于上层林木的衰亡而导致的林下土壤有机物腐解条件(光、温、水)改善, 利于土壤硝化过程.这在林冠郁闭的森林生态系统受到干扰后(林冠天窗下)常常出现(Bormann & Likens, 1979 ).然而, 毛竹纯林内的光、温条件亦优于林冠郁闭的次生针阔混交林, 而土壤硝态氮含量甚至显著低于针阔混交林(图4 ).我们认为毛竹林内土壤水含量的降低可能有部分原因, 但也有可能是毛竹在不同生长时期对无机氮的吸收偏爱不同.有研究认为植物生长早期一般喜好铵态氮, 而后期则喜好硝态氮(李生秀, 2008 ).竹-林过渡带的竹株是毛竹林扩张的分布前沿, 多处于幼竹阶段, 可能对铵态氮需求量大, 导致硝态氮积累.毛竹纯林以成熟毛竹居多, 对硝态氮的较大需求量使毛竹纯林土壤中硝态氮显著低于竹-林过渡带和针阔混交林.此外, 竹-林过渡带有最高的土壤pH值(竹-林过渡带>毛竹纯林>针阔混交林, 图4 ), 一般认为, 高pH值促进氮矿化, 尤其是硝化作用随pH值增加而线性增加(李贵才等, 2001 ), 本研究得出的竹-林过渡带土壤硝态氮积累的结果支持此说法.但也有一些研究显示竹-林过渡带的土壤pH值要低于常绿阔叶林的(宋庆妮等, 2013 ). ...
... ), 本研究得出的竹-林过渡带土壤硝态氮积累的结果支持此说法.但也有一些研究显示竹-林过渡带的土壤pH值要低于常绿阔叶林的(宋庆妮等, 2013 ). ...
毛竹扩张对常绿阔叶林土壤氮素矿化及有效性的影响
3
2013
... 国内外一些学者从人类活动(蔡亮等, 2003 ; Suzuki & Nakagoshi, 2008 )、生长特性(董鸣, 1996 ; Okutomi et al ., 1996 )、土壤资源(张春生等, 2007 ; 黄启堂, 2008 ; 吴家森等, 2008 ; 杨怀等, 2010 ; 宋庆妮等, 2013 )、光环境(Suzuki & Nakatsubo, 2001 ; 刘烁, 2010 )和化感作用(Chou & Yang, 1982 ; 梁倩倩等, 2012 ; 白尚斌等, 2013 )等方面对毛竹林扩张形成纯林的原因进行了分析探讨, 但有关毛竹林扩张导致其他林木失活枯死并抑制林下幼苗更新的机制, 目前尚有争议. ...
... 土壤无机氮的组分比例及其含量反映了土壤氮矿化与植物吸收的净结果.植被物种组成变化和植被类型的转变会通过不同的凋落物质量和养分利用效率而影响土壤氮的“矿化-固化”以及植被对氮的吸收效力, 从而反映到土壤无机氮的组分比例及其含量(李贵才等, 2001 ).有研究表明毛竹林向常绿阔叶林扩张会增加土壤总碳、总氮含量(吴家森等, 2008 ; 宋庆妮等, 2013 ).本研究结果显示, 毛竹林扩张使针阔混交林迅速转变为竹-林过渡带、毛竹纯林, 土壤有效氮总量有一定程度的增加.宋庆妮等(2013)研究发现, 毛竹林向常绿阔叶林扩张, 改变了土壤氮素矿化过程和格局, 导致竹-林过渡带土壤氨化作用增强, 硝化和总矿化作用减弱.本研究中竹-林过渡带硝态氮含量高于天然针阔混交林的结果(图4 )支持 毛竹林扩张将改变原有林分土壤氮素矿化过程和 格局.这也许是由于上层林木的衰亡而导致的林下土壤有机物腐解条件(光、温、水)改善, 利于土壤硝化过程.这在林冠郁闭的森林生态系统受到干扰后(林冠天窗下)常常出现(Bormann & Likens, 1979 ).然而, 毛竹纯林内的光、温条件亦优于林冠郁闭的次生针阔混交林, 而土壤硝态氮含量甚至显著低于针阔混交林(图4 ).我们认为毛竹林内土壤水含量的降低可能有部分原因, 但也有可能是毛竹在不同生长时期对无机氮的吸收偏爱不同.有研究认为植物生长早期一般喜好铵态氮, 而后期则喜好硝态氮(李生秀, 2008 ).竹-林过渡带的竹株是毛竹林扩张的分布前沿, 多处于幼竹阶段, 可能对铵态氮需求量大, 导致硝态氮积累.毛竹纯林以成熟毛竹居多, 对硝态氮的较大需求量使毛竹纯林土壤中硝态氮显著低于竹-林过渡带和针阔混交林.此外, 竹-林过渡带有最高的土壤pH值(竹-林过渡带>毛竹纯林>针阔混交林, 图4 ), 一般认为, 高pH值促进氮矿化, 尤其是硝化作用随pH值增加而线性增加(李贵才等, 2001 ), 本研究得出的竹-林过渡带土壤硝态氮积累的结果支持此说法.但也有一些研究显示竹-林过渡带的土壤pH值要低于常绿阔叶林的(宋庆妮等, 2013 ). ...
... ), 本研究得出的竹-林过渡带土壤硝态氮积累的结果支持此说法.但也有一些研究显示竹-林过渡带的土壤pH值要低于常绿阔叶林的(宋庆妮等, 2013 ). ...
天目山自然保护区林木菌根调查
4
1992
... 以往的菌根资源调查中, 对毛竹的菌根类型有内生(弓明钦等, 1997 ; 陈祥欣和陈连庆, 1983 )和外生(苏琍英等, 1992 )两种不同的说法.柳杉的菌根类型有内生(陈祥欣和陈连庆, 1983 )和外生(苏琍英等, 1992 )不同的描述.枫香树的菌根类型也有内外生(苏琍英等, 1992 )或外生的报道(陈祥欣和陈连庆, 1983 ).也有调查显示青冈是内外生菌根树种(苏琍英等, 1992 ).但在我们本研究区域的预实验研究中发现, 经根切片染色确定, 毛竹、杉木、毛柄连蕊茶、江浙山胡椒菌根是内生菌根(endomycorrhiza, EM)类型, 青冈、柳杉、枫香树菌根是外生菌根(ectomycorrhiza, ECM)类型. ...
... )和外生(苏琍英等, 1992 )不同的描述.枫香树的菌根类型也有内外生(苏琍英等, 1992 )或外生的报道(陈祥欣和陈连庆, 1983 ).也有调查显示青冈是内外生菌根树种(苏琍英等, 1992 ).但在我们本研究区域的预实验研究中发现, 经根切片染色确定, 毛竹、杉木、毛柄连蕊茶、江浙山胡椒菌根是内生菌根(endomycorrhiza, EM)类型, 青冈、柳杉、枫香树菌根是外生菌根(ectomycorrhiza, ECM)类型. ...
... )不同的描述.枫香树的菌根类型也有内外生(苏琍英等, 1992 )或外生的报道(陈祥欣和陈连庆, 1983 ).也有调查显示青冈是内外生菌根树种(苏琍英等, 1992 ).但在我们本研究区域的预实验研究中发现, 经根切片染色确定, 毛竹、杉木、毛柄连蕊茶、江浙山胡椒菌根是内生菌根(endomycorrhiza, EM)类型, 青冈、柳杉、枫香树菌根是外生菌根(ectomycorrhiza, ECM)类型. ...
... ).也有调查显示青冈是内外生菌根树种(苏琍英等, 1992 ).但在我们本研究区域的预实验研究中发现, 经根切片染色确定, 毛竹、杉木、毛柄连蕊茶、江浙山胡椒菌根是内生菌根(endomycorrhiza, EM)类型, 青冈、柳杉、枫香树菌根是外生菌根(ectomycorrhiza, ECM)类型. ...
天目山自然保护区林木菌根调查
4
1992
... 以往的菌根资源调查中, 对毛竹的菌根类型有内生(弓明钦等, 1997 ; 陈祥欣和陈连庆, 1983 )和外生(苏琍英等, 1992 )两种不同的说法.柳杉的菌根类型有内生(陈祥欣和陈连庆, 1983 )和外生(苏琍英等, 1992 )不同的描述.枫香树的菌根类型也有内外生(苏琍英等, 1992 )或外生的报道(陈祥欣和陈连庆, 1983 ).也有调查显示青冈是内外生菌根树种(苏琍英等, 1992 ).但在我们本研究区域的预实验研究中发现, 经根切片染色确定, 毛竹、杉木、毛柄连蕊茶、江浙山胡椒菌根是内生菌根(endomycorrhiza, EM)类型, 青冈、柳杉、枫香树菌根是外生菌根(ectomycorrhiza, ECM)类型. ...
... )和外生(苏琍英等, 1992 )不同的描述.枫香树的菌根类型也有内外生(苏琍英等, 1992 )或外生的报道(陈祥欣和陈连庆, 1983 ).也有调查显示青冈是内外生菌根树种(苏琍英等, 1992 ).但在我们本研究区域的预实验研究中发现, 经根切片染色确定, 毛竹、杉木、毛柄连蕊茶、江浙山胡椒菌根是内生菌根(endomycorrhiza, EM)类型, 青冈、柳杉、枫香树菌根是外生菌根(ectomycorrhiza, ECM)类型. ...
... )不同的描述.枫香树的菌根类型也有内外生(苏琍英等, 1992 )或外生的报道(陈祥欣和陈连庆, 1983 ).也有调查显示青冈是内外生菌根树种(苏琍英等, 1992 ).但在我们本研究区域的预实验研究中发现, 经根切片染色确定, 毛竹、杉木、毛柄连蕊茶、江浙山胡椒菌根是内生菌根(endomycorrhiza, EM)类型, 青冈、柳杉、枫香树菌根是外生菌根(ectomycorrhiza, ECM)类型. ...
... ).也有调查显示青冈是内外生菌根树种(苏琍英等, 1992 ).但在我们本研究区域的预实验研究中发现, 经根切片染色确定, 毛竹、杉木、毛柄连蕊茶、江浙山胡椒菌根是内生菌根(endomycorrhiza, EM)类型, 青冈、柳杉、枫香树菌根是外生菌根(ectomycorrhiza, ECM)类型. ...
Expansion of bamboo forests caused by reduced bamboo-shoot harvest under different natural and artificial conditions.
1
2008
... 国内外一些学者从人类活动(蔡亮等, 2003 ; Suzuki & Nakagoshi, 2008 )、生长特性(董鸣, 1996 ; Okutomi et al ., 1996 )、土壤资源(张春生等, 2007 ; 黄启堂, 2008 ; 吴家森等, 2008 ; 杨怀等, 2010 ; 宋庆妮等, 2013 )、光环境(Suzuki & Nakatsubo, 2001 ; 刘烁, 2010 )和化感作用(Chou & Yang, 1982 ; 梁倩倩等, 2012 ; 白尚斌等, 2013 )等方面对毛竹林扩张形成纯林的原因进行了分析探讨, 但有关毛竹林扩张导致其他林木失活枯死并抑制林下幼苗更新的机制, 目前尚有争议. ...
Impact of the bamboo Phyllostachys bambusoides on the light environment and plant communities on riverbanks.
1
2001
... 国内外一些学者从人类活动(蔡亮等, 2003 ; Suzuki & Nakagoshi, 2008 )、生长特性(董鸣, 1996 ; Okutomi et al ., 1996 )、土壤资源(张春生等, 2007 ; 黄启堂, 2008 ; 吴家森等, 2008 ; 杨怀等, 2010 ; 宋庆妮等, 2013 )、光环境(Suzuki & Nakatsubo, 2001 ; 刘烁, 2010 )和化感作用(Chou & Yang, 1982 ; 梁倩倩等, 2012 ; 白尚斌等, 2013 )等方面对毛竹林扩张形成纯林的原因进行了分析探讨, 但有关毛竹林扩张导致其他林木失活枯死并抑制林下幼苗更新的机制, 目前尚有争议. ...
1
... 挑取脱色处理后的根尖段于载玻片上, 一块载玻片上平行放置8条根尖段, 将其拉直, 用甘油明胶固封(Widden, 2001 ), 加盖洁净盖玻片将根尖段压平后, 在Zeiss Axio Imager M1显微镜(Carl Zeiss AG, Heidenheim, Germany)放大100-400倍条件下观察丛枝菌根形态并记录、拍照(图2 ).丛枝菌根的侵染频率和强度采用侵染分级法(Trouvelot et al ., 1986 )计算.丛枝菌根侵染频率为侵染的根尖段数占总根尖段数的比率, 侵染强度为丛枝菌根结构(丛枝、泡囊及菌丝)占根尖段的比率. ...
Ink and vinegar, a simple staining technique for arbuscular- mycorrhizal fungi.
1
1998
... 内生菌根的染色采用醋酸墨水染色法(Vierh- eilig et al ., 1998 ).根系样品用去离子水冲洗至无土壤杂物粘附, 在解剖镜下将根尖部无次生生长的0.5-1.0 cm根尖段剪下, 放入70%的乙醇中保存待用.每样品包括至少500个合格根尖段.染色时, 随机选取30-50条根尖段, 用去离子水冲洗3遍后, 置于10% KOH溶液中放入高压锅(121 ℃)内处理10-30 min, 使根尖段脱色透明.然后再用去离子水冲洗3次, 置于3%的碱性H2 O2 (25%-28% NH4 OH 8 mL, 3% H2 O2 100 mL)溶液中漂白15-45 min, 再用去离子水冲洗3次.随后将这些根尖段放入5%醋酸(CH3 COOH)中酸化30 min后移至5%醋酸墨水染色液(5%冰醋酸95 mL, 派克牌纯黑墨水5 mL)中煮沸3-30 min染色.染色后, 用5%醋酸冲洗根尖段20 min.最后置染色根尖段于去离子水中浸泡脱色24 h以上. ...
Mycorrhizal densities decline in association with nonnative plants and contribute to plant invasion.
2
2009
... 生物或非生物因素对外来入侵种的正反馈作用对外来生物的定植和传播有不可忽视的作用.其中, 植物和土壤的相互反馈在促进或抑制植物个体的更新上具有重要意义(Vogelsang & Bever, 2009 ).植物与土壤微生物形成强共生关系, 如菌根(mycorr- hiza), 干扰往往会削弱或破坏这种共生关系, 从而可能促进外来种的定植, 使原植物群落发生改变.如果退化的土壤微生物群落进一步抑制本地种的更新, 群落更替会加速(Vogelsang & Bever, 2009 ).菌根与植物共生现象的普遍性与其生态学功能的重要性, 使其容易成为寄主植物反馈调节的媒介.例如有研究发现, 在亚高山森林植被和高山草甸的过渡带, 柳树(Salix spp.)可以通过自身的枯落物及其外生菌根间接降低周围草本植物群落的丛枝菌根侵染, 进而可能改变菌根真菌在这一地区的分布, 这一改变又反过来影响草本植物群落的分布和生长(Becklin et al ., 2012 ). ...
... ).植物与土壤微生物形成强共生关系, 如菌根(mycorr- hiza), 干扰往往会削弱或破坏这种共生关系, 从而可能促进外来种的定植, 使原植物群落发生改变.如果退化的土壤微生物群落进一步抑制本地种的更新, 群落更替会加速(Vogelsang & Bever, 2009 ).菌根与植物共生现象的普遍性与其生态学功能的重要性, 使其容易成为寄主植物反馈调节的媒介.例如有研究发现, 在亚高山森林植被和高山草甸的过渡带, 柳树(Salix spp.)可以通过自身的枯落物及其外生菌根间接降低周围草本植物群落的丛枝菌根侵染, 进而可能改变菌根真菌在这一地区的分布, 这一改变又反过来影响草本植物群落的分布和生长(Becklin et al ., 2012 ). ...
The use of glycerin jelly for mounting stained roots for the observation and quantification of endomycorrhizal fungi.
1
2001
... 挑取脱色处理后的根尖段于载玻片上, 一块载玻片上平行放置8条根尖段, 将其拉直, 用甘油明胶固封(Widden, 2001 ), 加盖洁净盖玻片将根尖段压平后, 在Zeiss Axio Imager M1显微镜(Carl Zeiss AG, Heidenheim, Germany)放大100-400倍条件下观察丛枝菌根形态并记录、拍照(图2 ).丛枝菌根的侵染频率和强度采用侵染分级法(Trouvelot et al ., 1986 )计算.丛枝菌根侵染频率为侵染的根尖段数占总根尖段数的比率, 侵染强度为丛枝菌根结构(丛枝、泡囊及菌丝)占根尖段的比率. ...
 , 白尚斌
, 白尚斌


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}