The objective of this study was to understand how arbuscular mycorrhizal (AM) fungal community would vary with plant evolution.
Methods
Data-mining was carried out against MaarjAM database with ribosomal small subunit (SSU) gene. We predicted that the richness of root-associated AM fungal species increases from lower to higher modern land plants. Totally, 188 host plant species were selected in this analysis.
Important findings
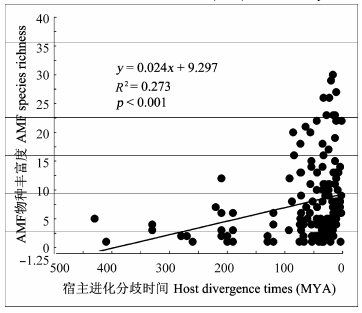
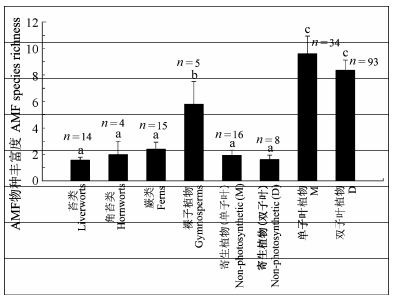
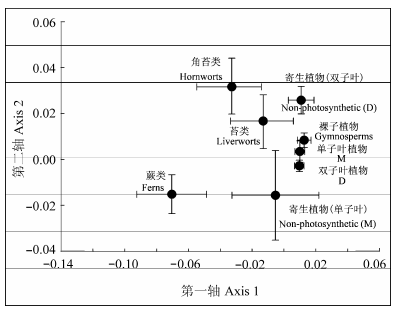
The AM fungal species richness increased with the divergence of host plant species, and significantly differed among plant phylogenetic groups. The more recently diverged host plants (i.e. gymnosperms and angiosperms) harbored higher AM fungal species richness, whereas the earlier-originated plants (liverworts, hornworts and ferns) possessed lower AM fungal species richness. The myco-heterotrophytes in angiosperms showed similarly low richness of AM fungal species to the lower plants. In addition, the AM fungal community composition significantly differed among different plant phylogenetic groups. Findings in this study provided some indirect evidence that AM fungal communities varied with plant evolution. It is suggested that plants might maintain the most effective AM fungi but discard those inefficient ones during evolution.
Fig. 1 Relationship between the time of divergence of hosts (MYA, million year) and the richness of arbuscular mycorrhizal fungal (AMF) species in roots.
Fig. 2 Distribution of species richness of arbuscular mycorrhizal fungi (AMF) in different plant phylogenetic groups (mean ± SE). Different lowercase letters indicate significant differences among different plant phylogenetic groups (p < 0.05). D, dicotyledons; M, monocotyledons.
Fig. 3 Non-metric multidimensional scaling (NMDS) analysis of arbuscular mycorrhizal fungal community composition in different phylogenetic groups. D, dicotyledons; M, monocotyledons.
Table 1
表1
表1 不同植物进化类群根内丛枝菌根真菌群落组成的单因素非参数MANOVA分析
Table 1 One-way NPMANOVA of arbuscular mycorrhizal fungal community composition in different phylogenetic groups of host plants
Molecular study of arbuscular mycorrhizal fungi colonizing the sporophyte of the eusporangiate rattlesnake fern (Botrychium virginian- um, Ophioglossaceae)
Effects of understory removal and tree girdling on soil microbial community composition and litter decomposition in two Eucalyptus plantations in South China.
Epiparasitic plants specialized on arbuscular mycorrhizal fungi.
1
2002
... AM真菌是植物进化过程中不可缺少的部分, 例如, 植物从水体向陆地的拓殖, 孢子植物向被子植物的进化以及菌根异养植物的分化等(Bidartondo et al., 2002; Wang et al., 2010; Field et al., 2012).本研究发现, 从早期起源植物到晚期起源植物, AM真菌物种丰富度依次增加.除裸子植物之外, 不同寄主植物进化类群中, AM真菌群落组成显著不同. ...
Do arbuscular mycorrhizal fungi alter plant-pathogen relations?
1
2001
... 植物与土壤微生物互作对生态系统功能具有重要的调控作用(Wu et al., 2011).丛枝菌根(arbuscular mycorrhiza, AM)真菌是一类与陆生植物根系共生的土壤功能微生物类群(Smith & Read, 2008).AM真菌在陆地生态系统广泛分布, 并且具有重要的生态功能.在植物-AM真菌共生体中, 植物为AM真菌提供光合作用产生的碳源, 而AM真菌为寄主植物提供诸多益处, 包括促进养分吸收、增加寄主植物对非生物胁迫的耐性和对生物胁迫的抗性(Borowicz, 2001; Smith & Read, 2008).通过菌丝桥, AM真菌能够调节植物间的相互作用(Wagg et al., 2011), 进而调控植物的种群动态和群落过程(van der Heijden et al., 1998; Koide & Dickie, 2002). ...
Contrasting arbuscular mycorrhizal responses of vascular and non-vascular plants to a simulated palaeozoic CO2 decline.
4
2012
... 研究发现, AM真菌是最为古老的一种微生物菌群.AM真菌可能起源于奥陶纪(至今约4.6亿年), 在泥盆纪(至今约4.0亿年)之前, 已与早期植物根系形成共生体(Remy et al., 1994; Redecker et al., 2000).在早期植物登陆过程中, AM真菌帮助其适应恶劣环境, 如在水肥供应不足时(Pirozynski & Malloch, 1975; Wang et al., 2010).在植物成功登陆之后, AM真菌在驱动植物进化方面可能起着更为重要的作用.近年来, 这一研究领域取得了较为丰富的成果.例如, Read等(2000)通过扫描电镜检测发现, 苔类植物类群中起源最早的分支能够形成典型的菌根结构, 因此, AM真菌被认为是苔类植物根内的“古老遗产” (Kottke & Nebel, 2005).在维管植物和苔类植物中, AM真菌的侵染策略和结构保守(Read et al., 2000; Ligrone et al., 2007).Fonseca和Berbara (2008)发现AM真菌能将磷转移给苔类植物.Wang等(2010)发现菌根共生基因在不同的植物系统类群里功能保守.通过模拟中生代中后期大气CO2浓度变化的研究发现, 在高CO2浓度条件下, AM真菌能够促进起源较早的苔类植物的生长与繁殖; 但在CO2浓度降低时, 苔类植物的菌根响应降低, 而维管植物的菌根响应增加(Humphreys et al., 2010; Field et al., 2012).以上研究表明, AM真菌可能是驱动植物进化的关键组分. ...
... AM真菌是植物进化过程中不可缺少的部分, 例如, 植物从水体向陆地的拓殖, 孢子植物向被子植物的进化以及菌根异养植物的分化等(Bidartondo et al., 2002; Wang et al., 2010; Field et al., 2012).本研究发现, 从早期起源植物到晚期起源植物, AM真菌物种丰富度依次增加.除裸子植物之外, 不同寄主植物进化类群中, AM真菌群落组成显著不同. ...
Mutualistic mycorrhiza-like symbiosis in the most ancient group of land plants.
1
2010
... 研究发现, AM真菌是最为古老的一种微生物菌群.AM真菌可能起源于奥陶纪(至今约4.6亿年), 在泥盆纪(至今约4.0亿年)之前, 已与早期植物根系形成共生体(Remy et al., 1994; Redecker et al., 2000).在早期植物登陆过程中, AM真菌帮助其适应恶劣环境, 如在水肥供应不足时(Pirozynski & Malloch, 1975; Wang et al., 2010).在植物成功登陆之后, AM真菌在驱动植物进化方面可能起着更为重要的作用.近年来, 这一研究领域取得了较为丰富的成果.例如, Read等(2000)通过扫描电镜检测发现, 苔类植物类群中起源最早的分支能够形成典型的菌根结构, 因此, AM真菌被认为是苔类植物根内的“古老遗产” (Kottke & Nebel, 2005).在维管植物和苔类植物中, AM真菌的侵染策略和结构保守(Read et al., 2000; Ligrone et al., 2007).Fonseca和Berbara (2008)发现AM真菌能将磷转移给苔类植物.Wang等(2010)发现菌根共生基因在不同的植物系统类群里功能保守.通过模拟中生代中后期大气CO2浓度变化的研究发现, 在高CO2浓度条件下, AM真菌能够促进起源较早的苔类植物的生长与繁殖; 但在CO2浓度降低时, 苔类植物的菌根响应降低, 而维管植物的菌根响应增加(Humphreys et al., 2010; Field et al., 2012).以上研究表明, AM真菌可能是驱动植物进化的关键组分. ...
Reciprocal rewards stabilize cooperation in the mycorrhizal symbiosis.
0
2011
Effects of mycorrhizal fungi on plant populations.
1
2002
... 植物与土壤微生物互作对生态系统功能具有重要的调控作用(Wu et al., 2011).丛枝菌根(arbuscular mycorrhiza, AM)真菌是一类与陆生植物根系共生的土壤功能微生物类群(Smith & Read, 2008).AM真菌在陆地生态系统广泛分布, 并且具有重要的生态功能.在植物-AM真菌共生体中, 植物为AM真菌提供光合作用产生的碳源, 而AM真菌为寄主植物提供诸多益处, 包括促进养分吸收、增加寄主植物对非生物胁迫的耐性和对生物胁迫的抗性(Borowicz, 2001; Smith & Read, 2008).通过菌丝桥, AM真菌能够调节植物间的相互作用(Wagg et al., 2011), 进而调控植物的种群动态和群落过程(van der Heijden et al., 1998; Koide & Dickie, 2002). ...
The evolution of mycorrhiza-like associations in liverworts: An update.
1
2005
... 研究发现, AM真菌是最为古老的一种微生物菌群.AM真菌可能起源于奥陶纪(至今约4.6亿年), 在泥盆纪(至今约4.0亿年)之前, 已与早期植物根系形成共生体(Remy et al., 1994; Redecker et al., 2000).在早期植物登陆过程中, AM真菌帮助其适应恶劣环境, 如在水肥供应不足时(Pirozynski & Malloch, 1975; Wang et al., 2010).在植物成功登陆之后, AM真菌在驱动植物进化方面可能起着更为重要的作用.近年来, 这一研究领域取得了较为丰富的成果.例如, Read等(2000)通过扫描电镜检测发现, 苔类植物类群中起源最早的分支能够形成典型的菌根结构, 因此, AM真菌被认为是苔类植物根内的“古老遗产” (Kottke & Nebel, 2005).在维管植物和苔类植物中, AM真菌的侵染策略和结构保守(Read et al., 2000; Ligrone et al., 2007).Fonseca和Berbara (2008)发现AM真菌能将磷转移给苔类植物.Wang等(2010)发现菌根共生基因在不同的植物系统类群里功能保守.通过模拟中生代中后期大气CO2浓度变化的研究发现, 在高CO2浓度条件下, AM真菌能够促进起源较早的苔类植物的生长与繁殖; 但在CO2浓度降低时, 苔类植物的菌根响应降低, 而维管植物的菌根响应增加(Humphreys et al., 2010; Field et al., 2012).以上研究表明, AM真菌可能是驱动植物进化的关键组分. ...
Molecular study of arbuscular mycorrhizal fungi colonizing the sporophyte of the eusporangiate rattlesnake fern (Botrychium virginian- um, Ophioglossaceae)
1
2007
... AM真菌群落在不同寄主植物进化类群中的组成差异可能是由双方共生功能相互选择造成的.Kiers等(2011)报道了菌根共生双方的“相互奖励”机制: 寄主植物将碳源分配给能提供更多磷素的AM真菌.由于AM真菌具有较高的功能多样性, 不同寄主植物进化类群的功能需求不同, 在进化过程中, 寄主植物可能会保留一些具有特定功能的AM真菌与之共生.例如, 苔藓类、蕨类和寄生植物都要经过一段菌根异养时期.期间, 这些植物所需要的碳源与养分完全依赖于与之共生的AM真菌.研究发现, 这些植物对Glomus group A类真菌几乎具有专一性(Russell & Bulman, 2005; Franke et al., 2006; Kovács et al., 2007).然而, 裸子植物和被子植物不但需要AM真菌提供养分, 还需要AM真菌帮助它们抵御胁迫环境, 如高温、干旱和病原菌等.因此, 基于更多的功能需求, 裸子植物和被子植物可能会与更多的AM真菌共生. ...
Glomeromycotean associations in liverworts: A molecular, cellular, and taxonomic analysis.
2
2007
... 研究发现, AM真菌是最为古老的一种微生物菌群.AM真菌可能起源于奥陶纪(至今约4.6亿年), 在泥盆纪(至今约4.0亿年)之前, 已与早期植物根系形成共生体(Remy et al., 1994; Redecker et al., 2000).在早期植物登陆过程中, AM真菌帮助其适应恶劣环境, 如在水肥供应不足时(Pirozynski & Malloch, 1975; Wang et al., 2010).在植物成功登陆之后, AM真菌在驱动植物进化方面可能起着更为重要的作用.近年来, 这一研究领域取得了较为丰富的成果.例如, Read等(2000)通过扫描电镜检测发现, 苔类植物类群中起源最早的分支能够形成典型的菌根结构, 因此, AM真菌被认为是苔类植物根内的“古老遗产” (Kottke & Nebel, 2005).在维管植物和苔类植物中, AM真菌的侵染策略和结构保守(Read et al., 2000; Ligrone et al., 2007).Fonseca和Berbara (2008)发现AM真菌能将磷转移给苔类植物.Wang等(2010)发现菌根共生基因在不同的植物系统类群里功能保守.通过模拟中生代中后期大气CO2浓度变化的研究发现, 在高CO2浓度条件下, AM真菌能够促进起源较早的苔类植物的生长与繁殖; 但在CO2浓度降低时, 苔类植物的菌根响应降低, 而维管植物的菌根响应增加(Humphreys et al., 2010; Field et al., 2012).以上研究表明, AM真菌可能是驱动植物进化的关键组分. ...
Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity.
1
1998
... 植物与土壤微生物互作对生态系统功能具有重要的调控作用(Wu et al., 2011).丛枝菌根(arbuscular mycorrhiza, AM)真菌是一类与陆生植物根系共生的土壤功能微生物类群(Smith & Read, 2008).AM真菌在陆地生态系统广泛分布, 并且具有重要的生态功能.在植物-AM真菌共生体中, 植物为AM真菌提供光合作用产生的碳源, 而AM真菌为寄主植物提供诸多益处, 包括促进养分吸收、增加寄主植物对非生物胁迫的耐性和对生物胁迫的抗性(Borowicz, 2001; Smith & Read, 2008).通过菌丝桥, AM真菌能够调节植物间的相互作用(Wagg et al., 2011), 进而调控植物的种群动态和群落过程(van der Heijden et al., 1998; Koide & Dickie, 2002). ...
Taxon- omic and functional diversity in arbuscular mycorrhizal fungi―Is there any relationship?
1
2004
... 然而, 目前我们并不清楚AM真菌群落如何随着寄主植物的进化而变化.根内AM真菌的物种丰富度不仅反映了AM真菌与寄主植物的兼容性, 而且反映了根内有具有活性的AM真菌.由于AM真菌具有较高功能多样性(van der Heijden et al., 2004), AM真菌与寄主植物的兼容性高低可能反映了寄主植物的一些功能需求.低兼容性表明寄主植物可能仅仅选择了很少的几种AM真菌与之共生来满足其特定的功能需求; 而高兼容性可能反映了寄主植物选择了更多的AM真菌与之共生以期提供更多的功能.根内AM真菌的寄主特异性反映了寄主植物对AM真菌的选择性变异.本文通过对MaarjAM数据库进行数据挖掘, 总结分析了不同植物类群的AM真菌物种丰富度和群落组成, 以期揭示AM真菌群落与寄主植物进化的关系. ...
Mycorrhizal fungal identity and diversity relaxes plant-plant competition.
1
2011
... 植物与土壤微生物互作对生态系统功能具有重要的调控作用(Wu et al., 2011).丛枝菌根(arbuscular mycorrhiza, AM)真菌是一类与陆生植物根系共生的土壤功能微生物类群(Smith & Read, 2008).AM真菌在陆地生态系统广泛分布, 并且具有重要的生态功能.在植物-AM真菌共生体中, 植物为AM真菌提供光合作用产生的碳源, 而AM真菌为寄主植物提供诸多益处, 包括促进养分吸收、增加寄主植物对非生物胁迫的耐性和对生物胁迫的抗性(Borowicz, 2001; Smith & Read, 2008).通过菌丝桥, AM真菌能够调节植物间的相互作用(Wagg et al., 2011), 进而调控植物的种群动态和群落过程(van der Heijden et al., 1998; Koide & Dickie, 2002). ...
Presence of three mycorrhizal genes in the common ancestor of land plants suggests a key role of mycorrhizas in the colonization of land by plants.
2
2010
... 研究发现, AM真菌是最为古老的一种微生物菌群.AM真菌可能起源于奥陶纪(至今约4.6亿年), 在泥盆纪(至今约4.0亿年)之前, 已与早期植物根系形成共生体(Remy et al., 1994; Redecker et al., 2000).在早期植物登陆过程中, AM真菌帮助其适应恶劣环境, 如在水肥供应不足时(Pirozynski & Malloch, 1975; Wang et al., 2010).在植物成功登陆之后, AM真菌在驱动植物进化方面可能起着更为重要的作用.近年来, 这一研究领域取得了较为丰富的成果.例如, Read等(2000)通过扫描电镜检测发现, 苔类植物类群中起源最早的分支能够形成典型的菌根结构, 因此, AM真菌被认为是苔类植物根内的“古老遗产” (Kottke & Nebel, 2005).在维管植物和苔类植物中, AM真菌的侵染策略和结构保守(Read et al., 2000; Ligrone et al., 2007).Fonseca和Berbara (2008)发现AM真菌能将磷转移给苔类植物.Wang等(2010)发现菌根共生基因在不同的植物系统类群里功能保守.通过模拟中生代中后期大气CO2浓度变化的研究发现, 在高CO2浓度条件下, AM真菌能够促进起源较早的苔类植物的生长与繁殖; 但在CO2浓度降低时, 苔类植物的菌根响应降低, 而维管植物的菌根响应增加(Humphreys et al., 2010; Field et al., 2012).以上研究表明, AM真菌可能是驱动植物进化的关键组分. ...
... AM真菌是植物进化过程中不可缺少的部分, 例如, 植物从水体向陆地的拓殖, 孢子植物向被子植物的进化以及菌根异养植物的分化等(Bidartondo et al., 2002; Wang et al., 2010; Field et al., 2012).本研究发现, 从早期起源植物到晚期起源植物, AM真菌物种丰富度依次增加.除裸子植物之外, 不同寄主植物进化类群中, AM真菌群落组成显著不同. ...
Arbuscular mycorrhizal symbionts in Botrychium (Ophioglossaceae).
Effects of understory removal and tree girdling on soil microbial community composition and litter decomposition in two Eucalyptus plantations in South China.
1
2011
... 植物与土壤微生物互作对生态系统功能具有重要的调控作用(Wu et al., 2011).丛枝菌根(arbuscular mycorrhiza, AM)真菌是一类与陆生植物根系共生的土壤功能微生物类群(Smith & Read, 2008).AM真菌在陆地生态系统广泛分布, 并且具有重要的生态功能.在植物-AM真菌共生体中, 植物为AM真菌提供光合作用产生的碳源, 而AM真菌为寄主植物提供诸多益处, 包括促进养分吸收、增加寄主植物对非生物胁迫的耐性和对生物胁迫的抗性(Borowicz, 2001; Smith & Read, 2008).通过菌丝桥, AM真菌能够调节植物间的相互作用(Wagg et al., 2011), 进而调控植物的种群动态和群落过程(van der Heijden et al., 1998; Koide & Dickie, 2002). ...
 , 王琪, 郭伊, 熊艳琴, 许明敏, 戴亚军
, 王琪, 郭伊, 熊艳琴, 许明敏, 戴亚军


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}