植物的生态策略和适应性、响应性功能要通过植物内部与外部不同功能性状来实现(Bernard- Verdier et al ., 2012 ), 性状的构建、稳固和性状之间的强弱变化与替补转换则靠植物自身的资源权衡和补偿机制来调控(Fortunel et al ., 2012 ; 陈莹婷和许振柱, 2014 ), 是植物在特定环境下生存策略的重要表现(Mooney et al ., 2010 )。根系构型决定了根系在土壤空间中的位置和资源获取方式, 是植物与资源环境异质性相互适应的结果(Malamy, 2005 ; Rogers & Benfey, 2015 )。根系分叉数和连接长度作为根系构型的重要特征指标, 受土壤空间异质性(Villordon et al ., 2014 )、群落环境(单立山等, 2012 )、遗传特性(Kiswara et al ., 2009 ; Bingham & Wu, 2011 )等诸多因素的影响, 在其生存和发展的过程中, 会根据所处生境的资源条件, 调整、转换或补偿自身功能, 实现根系功能性状间的资源权衡。根系分叉数的变化会影响根系的空间分布属性、营养吸收能力和固定作用; 根系连接长度的大小直接决定了根系在土壤中的空间拓展和营养物质吸收能力(Walk et al ., 2004 ), 二者的权衡关系直接反映了植物根系吸收效率和根系构建过程中所需的碳量(Fitter & Stickland, 1991 )。当根系处于水分状态良好的土层中时, 为了提高水分利用率, 会形成更多的侧根, 增大根分叉数(周艳松和王立群, 2011 ); 在干旱胁迫环境中, 较大的分叉数会引起根系内部对营养物质的竞争(杨小林等, 2008 ), 此时植物倾向于增大连接长度, 以减少根系之间的交叠重复, 占据较大的水分和养分资源空间(郭京衡等, 2014 ), 这是植物为适应不同生境条件而形成的获取资源策略, 以达到“生存、生长、繁殖”之三大目的。地形作为生境条件的一种综合指示, 往往能营造局部小气候(Cantón et al ., 2004 )。坡向作为一种重要的地形要素(Fekedulegn et al ., 2003 ), 可以对局部区域的太阳辐射和降水进行二次分配, 其空间变化是群落组成、物种多样性、生态系统过程和植物功能性状的重要决定因素(de Bello et al ., 2006 ; Bennie et al ., 2008 ; Carletti et al ., 2009 )。在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 在各个水平上影响了植物生长及其对水分利用的有效性(Sardans et al ., 2008 ; Huang et al ., 2015 ), 进而引起植物碳资源分配格局和构型构建策略的改变, 为提高生境适应性和风险规避能力, 植物对连接长度和分叉数等主要根系功能性状进行资源优化配置, 构建与生境相适应的根系构型, 优化根系营养吸收和传输系统。因此, 研究坡向梯度上植物根系构型的生态适应策略, 对于理解植物的环境适应机制有重要意义。
近年来, 多数学者对植物根系的研究主要集中在异质生境中根系功能性状可塑性(Grossman & Rice, 2012 ; Pacheco-Villalobos & Hardtk, 2012 )、不同群落环境下根系构型的适应性调整(Rogers & Benfey, 2015 )、根系生长与土壤环境的关系(Forde, 2014 ; Szoboszlay et al ., 2015 )、根系结构和功能(Hodge et al ., 2009 )等方面, 关于退化草地中植物构件生物量分配模式(周兵等, 2015 )、影响根系构型的主要功能性状(周艳松和王立群, 2011 ), 以及不同入侵阶段植物功能性状表型可塑性(Hulme, 2008 ; Pyšek et al ., 2009 )等方面也进行了系统研究, 而从根系构型构建策略的角度出发, 对生境梯度植物根系功能性状间资源权衡机制的研究仍显不足。甘肃臭草(Melica przewalskyi )是禾本科臭草属多年生根茎植物, 通过根状茎的快速横向生长, 长出幼芽和根系, 形成新植株进行无性繁殖, 在地下建立庞大的根茎系统, 形成密集的圆形株丛, 其分株间的间隔子短, 分枝角度小, 能够在竞争过程中占据优势地位。近年来, 在祁连山北坡中山区荒漠草原与典型草原过渡带, 甘肃臭草替代了原生地带性植被的优势种西北针茅(Stipa sareptana var. krylovii ), 形成大量面积不等、繁茂生长的单一优势种群落斑块(赵成章和龙瑞军, 2008 ), 表现出较高的形态可塑性和对逆境的适应能力, 其强大的入侵能力使众多学者对其资源利用策略和生长扩散过程产生了兴趣。目前, 对甘肃臭草型退化草地优势物种空间格局分析及其对水分的响应(赵成章等, 2011 ; 高福元和赵成章, 2012 )、甘肃臭草斑块土壤水分空间异质性(杨泉等, 2014 ), 以及甘肃臭草茎-叶性状、叶内性状(党晶晶等, 2014 ,2015 )等方面进行了研究, 而生境梯度上甘肃臭草根系构型的可塑性变化尚未见报道。鉴于此, 本文在野外调查的基础上, 试图弄清以下问题: (1)不同坡向甘肃臭草根系分叉数和连接长度的权衡关系如何?(2)导致甘肃臭草根系分叉数和连接长度变化的原因有哪些?旨在探讨祁连山地甘肃臭草种群根系构型构建的资源配置模式, 进一步认识甘肃臭草单一优势种群斑块形成机制和扩散机理。
1 材料和方法
1.1 研究区域和样地概况
研究区位于祁连山北坡中山区的甘肃省肃南县皇城草原, 海拔2534-2818 m, 属于典型的大陆性气候, 同时又具有水热垂直地带性变化的山地气候特点, 年平均气温1.5 ℃, 7月平均气温14 ℃, 1月平均气温-12.5 ℃, >0 ℃积温为1400-1688 ℃, 年降水量270-350 mm, 降水主要集中在6-8月。土壤以栗钙土和黑钙土为主, 受地势起伏的影响, 植被分布具有明显的垂直分异性, 植被以旱生多年生禾本科植物和湿中生灌木为主。天然草地植物主要有狼毒(Stellera chamaejasme )、西北针茅、冷蒿(Artemisia frigida )、甘肃臭草、冰草(Agropyron cristatum )、草地早熟禾(Poa pratensis )、赖草(Leymus secalinus )、星毛委陵菜(Potentilla acaulis )、碱韭(Allium polyrhizum )等。供试草地为当地村落一块有围栏的种公羊放牧草场, 每年11月至次年6月放牧, 期间羊群可以在这块草场中自由采食; 每年的6月至11月为禁牧期, 因此, 供试草地各个坡向上羊群的放牧时间和强度基本一致, 能够消除放牧干扰的因素, 实验数据具有可比性, 能够反映不同坡向对植物生长的影响。各个坡向草地群落特征和优势种存在差异, 南坡甘肃臭草为优势种, 东坡、西坡甘肃臭草和西北针茅为共同优势种, 北坡西北针茅为优势种、甘肃臭草为伴生种。
1.2 群落调查和植物取样
2013年8月, 在研究区选择一座相对独立的小山(101.80°-101.83° E, 37.91°-37.93° N), 山脚海拔2540 m, 山高约200 m, 山坡包含所有标准8个坡向。在地形图上将这座小山按照坡向划分, 沿8个方向从山脚向山顶海拔每上升20 m设置一块10 m × 10 m样地, 每个方向10块样地, 总共为80块样地, 各样地的坡度介于10°-21°之间。取样时用手持GPS记录每块样地的经纬度与海拔。取样过程如下: 第一, 进行草地群落学调查, 每块样地用1 m × 1 m的样方框, 测定草地群落所有植物的密度、高度和地上生物量, 重复6次。第二, 每块样地设置3个30 cm × 30 cm的小样方, 每个坡向共计30个小样方, 在保持地上植株完整的情况下挖取土柱(30 cm × 30 cm × 50 cm), 之后用孔径为0.25 mm的网孔筛把土柱上的泥土在附近的河边全部冲洗干净, 将甘肃臭草同其他植物仔细分开, 整株挑出, 然后把地上和地下部分分别装入自封袋中带回实验室。第三, 进行土壤水分采样, 每块样地用土钻(直径 = 4 cm)在0-50 cm土层范围分5层间隔10 cm取土样, 重复3次, 同时在每块样地用土钻(直径 = 4 cm)取0-50 cm混合土样, 重复3次, 所有样品剔除明显的植物根段和枯落物等杂质, 装入编号的铝盒中, 带回实验室。
在根系扫描仪的根盘里放少量水, 把甘肃臭草根系放在根盘里, 并用毛刷仔细梳理根系使其在水中分布均匀, 经扫描仪扫描根系并以图形文件格式存储到计算机中, 再用Win-RHIZO (Pro 2009a, Regent Instruments, Quebec, Canada)软件对之进行分析, 获得各坡向甘肃臭草分叉数和平均连接长度。将采集的甘肃臭草地下和地上部分分别包装, 在80 ℃烘箱里烘干24 h后, 用电子天平(0.0001 g)称取质量, 并计算地上和地下的生物量分配比例; 土壤样品在105 ℃的烘箱内烘干12 h, 取出称量质量, 计算出各样地土壤质量含水量。
1.3 空间数据采集
利用ArcGIS对研究区1:5万地形图进行数字化, 得到研究区域10 m分辨率的数字高程模型(digital elevation model, DEM)。在DEM基础上利用Spatial Analyst模块中的Surface Analysis命令提取坡向, 它表示地表面上一点的切平面的法线矢量n 在水平面的投影nxoy 与过该点的正北方向的夹角, 其中坡向组别以国际划分法确定(陈瑶等, 2006 ), 具体方法参见汤国安等(2010) 文献。
1.4 数据处理
将80个样地划分为北坡(315°-360°和0-45°)、东坡(45°-135°)、南坡(135°-225°)和西坡(225°-315°) 4个坡向组。利用Win-RHIZO根系分析软件分析提取的根系分叉的数量, 计算出各坡向甘肃臭草的根系分叉数。分叉数是指单位根长上分叉的数量, 在相同的单位长度内, 分叉数越高, 表明根系吸收养分或水分的能力越强(刘佳等, 2010 )。计算公式为:
分叉(No.·cm-1 ) =分叉数(No.)/根长(cm)
实验所用数据采用SPSS 16.0软件进行处理分析, 用SigmaPlot 10.0和Excel软件绘图。实验数据均进行以10为底的对数转换, 使之符合正态分布, 进行单因素方差分析(one-way ANOVA), 显著性水平设为0.05。利用线性回归分析法对甘肃臭草根系分叉数和连接长度间的关系进行分析。
2 结果
2.1 不同坡向甘肃臭草根系分叉数和连接长度的变化
单因素方差分析表明, 不同坡向草地甘肃臭草的根系分叉数与连接长度均存在显著差异(p < 0.05, 图1 )。随着坡向由北坡转向东坡、西坡和南坡, 甘肃臭草根系分叉数呈逐渐减小的趋势, 北坡分叉数是南坡的2.14倍; 根系连接长度呈逐渐增大的趋势, 从北坡的0.06 cm增加到南坡的0.12 cm, 其中东坡和西坡无显著差异(p > 0.05)。根系分叉数和连接长度的变率均呈先减小后增加的趋势, 其中南坡根系分叉数和根系连接长度的变率最大, 表明生长在南坡的甘肃臭草根系构型变化较大。
图1 甘肃臭草根系分叉数和连接长度沿坡向梯度的变化(平均值±标准误差)。不同小写字母表示坡向间差异显著(p < 0.05)。
Fig. 1 Changes in root forks and link length of Melica przewalskyi along an aspect gradient (mean ± SE ). Different lowercase letters indicate significant differences among slope aspects (p < 0.05).
2.2 不同坡向甘肃臭草根系分叉数和连接长度的关系
Pearson相关性分析表明, 南坡和北坡甘肃臭草根系分叉数和连接长度间均存在极显著的负相关关系(p < 0.01), 而东坡和西坡存在显著的负相关关系(p < 0.05)。在坡向梯度上, 随着甘肃臭草根系连接长度的增加, 根系分叉数均呈减小趋势, 表明甘肃臭草根系分叉数和连接长度之间存在权衡关系(图2 )。
图2 不同坡向甘肃臭草根系分叉数和连接长度的关系。A, 北坡。B, 东坡。C, 西坡。D, 南坡。
Fig. 2 The relationship between root forks and link length of Melica przewalskyi in different aspects of slopes. A, North slope. B, East slope. C, West slope. D, South slope.
2.3 不同坡向甘肃臭草生物学特征
不同坡向梯度分布的甘肃臭草其生物学特征差异均显著(p < 0.05, 表1 )。随着坡向由北坡转向东坡、西坡和南坡, 甘肃臭草种群的密度和高度呈逐渐增大的趋势, 其中南坡甘肃臭草种群密度是北坡的2.52倍, 南坡甘肃臭草种群高度相对于北坡增加了40%; 根冠比在东、西坡具有最大值, 北坡具有最小值, 其中东西与南北之间无显著差异(p > 0.05)。
2.4 不同坡向草地群落及土壤水分特征
各个坡向上草地群落的盖度、高度和地上生物量和土壤水分均存在显著差异(p < 0.05, 表2 )。草地群落的高度、盖度和地上生物量按照北坡、东坡、西坡、南坡的顺序依次递减; 群落土壤水分在南坡具有最小值, 北坡具有最大值, 东坡和西坡之间差异不显著(p > 0.05)。
3 讨论
植物在不同功能和性状之间对有限资源的分配存在着互相牵制的权衡作用(Shipley et al ., 2006 ), 这就需要植物在生长发育过程中, 通过不断优化自身的资源分配来提高适合度以适应环境的变化(Fabbro & Körner, 2004)。根系作为植物地上部分与土壤的连接器官, 在较大程度上影响着植物的水分和养分吸收、运移和存储功能(Dannowski & Block, 2005 ; Kong et al ., 2014 ), 根系分叉数和连接长度能够反映根系不同部位构建过程中的碳消耗特点, 影响根系拓展空间、获取营养的代价与效率, 体现了植物性状和构型构建对生境变化的响应, 从中可以理解和阐释植物的物种分布、种间关系、权衡及其作用机制(Westoby & Wright, 2006 )。本研究发现, 随着坡向由北坡向东坡、西坡、南坡转变, 甘肃臭草根系分叉数呈逐渐减小的趋势, 而根系连接长度呈相反的变化趋势(图1 ), 即甘肃臭草分配给根系分叉数和连接长度的资源间存在着“此消彼长”的权衡关系(图2 )。经Pearson相关性分析得出, 群落密度与根系分叉数、连接长度呈极显著相关关系(p < 0.01), 群落高度、土壤含水量与二者显著相关(p < 0.05), 地上生物量与根系分叉数显著相关(p < 0.05, 表3 ), 表明坡向梯度上甘肃臭草根系分叉数和连接长度权衡关系的差异性, 是植物根系构建的资源配置模式对高寒草地生境异质性适应的结果。
植物在环境胁迫下具有权衡性状关系的适应策略(Mooney et al ., 2010 ), 通过生理和发育调节形成与环境相适应的表型特征, 有利于提高植物水分、养分利用效率等生物学功能(李西良等, 2014 )。植物根系构型对环境的适应是通过根系分支结构、空间分布格局等构型特征的改变表现出来的, 是植物长期适应环境的进化结果(Guswa, 2010 )。研究区位于北半球中纬度地区, 山地北坡光照资源最弱(殷秀琴, 2004 )、土壤含水量较高(表2 ), 草地群落的高度、盖度和生物量最高(表2 ), 甘肃臭草处于斑块形成的初期阶段, 其种群密度较小, 也没有高度优势(表1 , 表2 ), 与草地群落的优势种群西北针茅相比, 处于种间竞争的劣势地位。在种间胁迫较大的情况下, 生长在北坡的甘肃臭草采取了增大根系分叉数、减小根系连接长度的根系构型构建策略; 根系分叉数的增加能够增强根系重叠程度、构建密集的根系网络系统, 有利于甘肃臭草地下根系系统和地上株丛的密集生长, 增强种群的密集分布强度, 提高根系对土壤资源的吸收和利用效率, 以斑块式抱团生长实现种群内个体间的高度协同; 减小根系连接长度可能会缩短甘肃臭草克隆分株间的资源传输距离, 有利于增强甘肃臭草克隆分株个体间的相互联系, 保证种群内分株间的资源连接和共享, 从而使种群能够在激烈的资源竞争中存活下来, 与相关学者得出一致的结论, 即“分叉数较大的植物能够迅速占据大量土壤空间快速生长, 适宜生长在土壤资源丰富的环境中” (周艳松和王立群, 2011 ; 郭京衡等, 2014 )。因此, 增大分叉数、减小连接长度是分布于北坡的甘肃臭草增大斑块密度, 提升竞争能力和空间拓殖能力的一种生活史对策, 使根系分叉数和连接长度之间的权衡关系达到极显著水平(p < 0.01, 图2 )。
植物根系构型构建策略的差异可以用根系碳投入与土壤水分养分收益的权衡关系来解释(Oppelt et al ., 2001 )。根系分叉数的增加会导致根系重叠加剧, 增加根系内部的资源竞争态势(杨小林等, 2008 )。根系连接长度的变化会影响根系空间拓展能力和利用土壤资源的有效性(Schenk & Jackson, 2002a , 2002b ), 这与植物自身根系碳投入与土壤水分养分的收益权衡相契合, 是提高植物资源获取能力的一个重要策略(杨小林等, 2008 )。相对于北坡, 山地南坡日照时间长、水分蒸发量大(殷秀琴, 2004 ), 生长在南坡的甘肃臭草多呈直径不等的单一优势种群斑块, 占据了草地群落的绝对资源优势(表1 , 表2 )。甘肃臭草采用克隆繁殖模式形成了庞大的互相连通交织的根茎系统, 构建了高度聚集的根系网络(高福元和赵成章, 2012 ), 斑块内部种群聚集程度较高, 斑块中心土壤含水量明显低于斑块边缘的土壤含水量(赵成章等, 2011 ), 体现了甘肃臭草向斑块周围扩张的迫切需求。在地上和地下生存空间拥挤、土壤含水量分布不均衡状态下, 为了避免不必要的种内竞争, 甘肃臭草限制了地下生物量的分配比例, 以减缓营养繁殖速度, 降低种群聚集强度, 此时甘肃臭草选择了增加根系连接长度、减少分叉数的根系构型构建模式。根系分叉数的减小能够降低种群内部根系重叠程度, 避免不必要的种内竞争; 连接长度的增大可以增强根系空间拓展能力, 扩大单位根系所占的土壤体空间, 使根系从斑块中心向土壤水分较好、生长发育空间较大的边缘区域拓殖, 是南坡甘肃臭草单优种群斑块扩张的主要动力, 与赵成章等(2011) 关于甘肃臭草通过营养繁殖以类似“同心圆”的方式不断由斑块中心向边缘扩大种群领地的观点一致。这符合相关学者“当植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 增加根系连接长度等策略, 以寻求更多的水分和养分, 保证自身的生存和生长”的结论(单立山等, 2013 ; 杜建会等, 2014 )。因此, 南坡甘肃臭草根系分叉数和连接长度间呈极显著负相关关系(p < 0.01, 图2 ), 减小分叉数、增加连接长度是甘肃臭草斑块扩张的根系构型构建策略, 降低了甘肃臭草单一优势种群斑块内部的根系拥挤程度。
环境异质性是植物群落结构特征差异形成的主要原因之一, 随着资源环境的改变, 植物群落的地上生物量、总盖度和平均高度都会发生明显变化, 必然会改变种群内每块植株可获得性资源的数量。根系构型会对周围环境产生适应性的改变(Oppelt et al ., 2005 ), 通过权衡各功能性状指标间的生物量分配模式, 以维持个体生长率最大。东坡和西坡属于半阴坡和半阳坡, 是植被类型的交错带, 植被多样性较高, 群落组成也较为复杂, 草地群落密度和地上生物量都相对较大(表2 ), 邻体干扰较强。虽然分布于东坡和西坡的甘肃臭草斑块已经形成, 但斑块内聚集程度不高, 而且斑块面积较小, 与西北针茅共同成为草地群落的优势种群, 在种间竞争中没有绝对的优势地位。作为一种禾本科多年生密集型根茎克隆植物, 甘肃臭草通过根状茎的生长实现大量营养繁殖, 增大了地下生物量的分配比例, 导致东坡和西坡的根冠比显著大于北坡和南坡(表1 ), 有助于提升种间竞争地位。此时, 为了增强种间聚集程度、扩大分布区域, 甘肃臭草选择了既有利于提高地上株丛和地下根系的分布密度, 增强种群空间分布聚集强度和斑块稳固性, 又有助于提升根系空间拓展能力, 逐步扩大种群空间生态位的根系分叉数和连接长度优化配置模式, 导致生长在东坡和西坡的甘肃臭草根系分叉数和连接长度间呈显著负相关关系(p < 0.05, 图2 ), 二者大小介于北坡和南坡之间, 体现了植物根系对高寒草地生境异质性的环境适应性特征。
4 结论
根系构型通过权衡碳分配、碳消耗采取了相应的生态适应策略, 表现出不一样的适应特性。本研究发现, 生境对根系分叉数和连接长度的权衡关系具有显著影响, 随着坡向由北坡向东坡、西坡、南坡转变, 高寒退化草地甘肃臭草根系分叉数逐渐减小, 而连接长度逐渐增大, 虽然不同坡向甘肃臭草种群的根系分叉数与连接长度之间存在权衡关系, 但是北坡和南坡甘肃臭草给予根系分叉数与连接长度的投资模式表现出相反的趋势。这种结果较好地验证了植物根系通过构型的可塑性调整响应环境变异的协同适应性机制, 自然选择使甘肃臭草以最“经济”的方式构建根系系统, 找到最佳的收益/代价关系, 以适应不同资源环境, 基于植物对环境修饰的可逆机制, 为甘肃臭草适应异质生境提供了一个快速反应机制。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[49]
Yin XQ 2004 ). Biogeography . Higher Education Press , Beijing . 26 -28 . (in Chinese)
[本文引用: 2]
[殷秀琴 (2004 ). 生物地理学 . 高等教育出版社 , 北京 . 26 -28 .]
[本文引用: 2]
[50]
Zhao CZ Gao FY Shi FX Ren H Sheng YP 2011 ). Melica przewalskyi population spatial pattern and response to soil moisture in degraded alpine grassland.
Acta Ecologica Sinica , 31 , 6688 -6695 . (in Chinese with English abstract)
[本文引用: 3]
[赵成章 , 高福元 , 石福习 , 任珩 , 盛亚萍 (2011 ). 高寒退化草地甘肃臭草种群分布格局及其对土壤水分的响应
. 生态学报 , 31 , 6688 -6695 .]
[本文引用: 3]
[51]
Zhao CZ Long RJ 2008 ). Rehabilitation process of degraded Melica przewalskyi grassland in the upper reaches of Shiyang River.
Journal of Mountain Science , 26 , 286 -292 . (in Chinese with English abstract)
[本文引用: 1]
[赵成章 , 龙瑞军 (2008 ). 石羊河上游甘肃臭草型退化草地植被恢复过程
. 山地学报 , 26 , 286 -292 .]
[本文引用: 1]
[52]
Zhou B Yan XH Xiao YA Wang N Kuang ZQ 2015 ). Module biomass of Ageratum conyzoides populations in different habitats.
Acta Ecologicy Sinica , 35 , 2602 -2608 . (in Chinese with English abstract)
[本文引用: 1]
[周兵 , 闫小红 , 肖宜安 , 王宁 , 旷志强 (2015 ). 不同生境下入侵植物胜红蓟种群构件生物量分配特性
. 生态学报 , 35 , 2602 -2608 .]
[本文引用: 1]
[53]
Zhou YS Wang LQ 2011 ). Ecological adaptation of root architecture to grassland degradation in Potentilla acaulis.
Chinese Journal of Plant Ecology , 35 , 490 -499 . (in Chinese with English abstract)
[本文引用: 3]
[周艳松 , 王立群 (2011 ). 星毛委陵菜根系构型对草原退化的生态适应
. 植物生态学报 , 35 , 490 -499 .]
[本文引用: 3]
[1]
Bennie J Huntley B Wiltshire A Hill MO Baxter R 2008 ). Slope, aspect and climate: Spatially explicit and implicit models of topographic microclimate in chalk grassland.
Ecological Modelling , 216 , 47 -59 .
[本文引用: 1]
[2]
Bernard-Verdier M Navas M-L Vellend M Violle C Fayolle A Garnier E 2012 ). Community assembly along a soil depth gradient: Contrasting patterns of plant trait convergence and divergence in a Mediterranean rangeland.
Journal of Ecology , 100 , 1422 -1433 .
[本文引用: 1]
[3]
Bingham IJ Wu LH 2011 ). Simulation of wheat growth using the 3D root architecture model SPACSYS: Validation and sensitivity analysis.
European Journal of Agronomy , 34 , 181 -189 .
[本文引用: 1]
[4]
Cantón Y Del Barrio G Solé-Benet A Lázaro R 2004 ). Topographic controls on the spatial distribution of ground cover in the Tabernas badlands of SE Spain.
CATENA , 55 , 341 -365 .
[本文引用: 1]
[5]
Carletti P Vendramin E Pizzeghello D Concheri G Zanella A Nardi S Squartini A 2009 ). Soil humic compounds and microbial communities in six spruce forests as function of parent material, slope aspect and stand age.
Plant and Soil , 315 , 47 -65 .
[本文引用: 1]
[6]
Chen Y Xu X Zhang DR Wei Y 2006 ). Correlations between vegetation distribution and topographical factors in the northwest of Longmen Mountain, Sichuan Province.
Chinese Journal of Ecology , 25 , 1052 -1055 . (in Chinese with English abstract)
[本文引用: 1]
[陈瑶 , 胥晓 , 张德然 , 魏勇 (2006 ). 四川龙门山西北部植被分布与地形因子的相关性
. 生态学杂志 , 25 , 1052 -1055 .]
[本文引用: 1]
[7]
Chen YT Xu ZZ 2014 ). Review on research of leaf economics spectrum.
Chinese Journal of Plant Ecology , 38 , 1135 -1153 . (in Chinese with English abstract)
[本文引用: 1]
[陈莹婷 , 许振柱 (2014 ). 植物叶经济谱的研究进展
. 植物生态学报 , 38 , 1135 -1153 .]
[本文引用: 1]
[8]
Dang JJ Zhao CZ Li Y Hou ZJ Dong XG 2014 ). Variations with slope in stem and leaf traits of Melica przewalskyi in alpine grassland.
Chinese Journal of Plant Ecology , 38 , 1307 -1314 . (in Chinese with English abstract)
[本文引用: 1]
[党晶晶 , 赵成章 , 李钰 , 侯兆疆 , 董小刚 (2014 ). 高寒草地甘肃臭草茎——叶性状的坡度差异性
. 植物生态学报 , 38 , 1307 -1314 .]
[本文引用: 1]
[9]
Dang JJ Zhao CZ Li Y Hou ZJ Dong XG 2015 ). Relationship between leaf traits of Melica przewalskyi and slope aspects in alpine grassland of Qilian Mountains, China.
Chinese Journal of Plant Ecology , 39 , 23 -31 . (in Chinese with English abstract)
[本文引用: 1]
[党晶晶 , 赵成章 , 李钰 , 侯兆疆 , 董小刚 (2015 ). 祁连山高寒草地甘肃臭草叶性状与坡向间的关系
. 植物生态学报 , 39 , 23 -31 .]
[本文引用: 1]
[10]
Dannowski M Block A 2005 ). Fractal geometry and root system structures of heterogeneous plant communities.
Plant and Soil , 272 , 61 -76 .
[本文引用: 1]
[11]
de Bello F Lepš J Sebastia M-T 2006 ). Variations in species and functional plant diversity along climatic and grazing gradients.
Ecography , 29 , 801 -810 .
[本文引用: 1]
[12]
Du JH Liu AL Dong YX Hu MY Liang J Li W 2014 ). Architectural characteristics of roots in typical coastal psammophytes of South China.
Chinese Journal of Plant Ecology , 38 , 888 -896 . (in Chinese with English abstract)
[本文引用: 1]
[杜建会 , 刘安隆 , 董玉祥 , 胡绵友 , 梁杰 , 李薇 (2014 ). 华南海岸典型沙生植物根系构型特征
. 植物生态学报 , 38 , 888 -896 .]
[本文引用: 1]
[13]
Fabbro T Körner C 2004 ). Altitudinal differences in flower traits and reproductive allocation.
Flora-Morphology, Distribution, Functional Ecology of Plants , 199 , 70 -81 .
[14]
Fekedulegn D Hicks RR Colbert JJ 2003 ). Influence of topographic aspect, precipitation and drought on radial growth of four major tree species in an Appalachian watershed.
Forest Ecology and Management , 177 , 409 -425 .
[本文引用: 1]
[15]
Fitter AH Sticklabd TR 1991 ). Architectural analysis of plant root systems 2. Influence of nutrient supply on architecture in contrasting plant species.
New Phytologist , 118 , 383 -389 .
[本文引用: 1]
[16]
Forde BG 2014 ). Nitrogen signalling pathways shaping root system architecture: An update.
Current Opinion in Plant Biology , 21 , 30 -36 .
[本文引用: 1]
[17]
Fortunel C Fine PVA Baraloto C 2012 ). Leaf, stem and root tissue strategies across 758 neotropical tree species.
Functional Ecology , 26 , 1153 -1161 .
[本文引用: 1]
[18]
Gao FY Zhao CZ 2012 ). In the process of grassland degradation the spatial pattern and spatial association of dominant species.
Acta Ecologica Sinica , 32 , 6661 -6669 . (in Chinese with English abstract)
[本文引用: 2]
[高福元 , 赵成章 (2012 ). 甘肃臭草型退化草地优势种群空间格局及其关联性
. 生态学报 , 32 , 6661 -6669 .]
[本文引用: 2]
[19]
Grossman JD Rice KJ 2012 ). Evolution of root plasticity responses to variation in soil nutrient distribution and concentration.
Evolutionary Applications , 5 , 850 -857 .
[本文引用: 1]
[20]
Guo JH Zeng FJ Li CJ Zhang B 2014 ). Root architecture and ecological adaptation strategies in three shelterbelt plant species in the southern Taklimakan Desert.
Chinese Journal of Plant Ecology , 38 , 36 -44 . (in Chinese with English abstract)
[本文引用: 2]
[郭京衡 , 曾凡江 , 李尝君 , 张波 (2014 ). 塔克拉玛干沙漠南缘三种防护林植物根系构型及其生态适应策略
. 植物生态学报 , 38 , 36 -44 .]
[本文引用: 2]
[21]
Guswa AJ 2010 ). Effect of plant uptake strategy on the water- optimal root depth.
Water Resources Research , 46 , doi: 10.1029/2010WR009122 .
[本文引用: 1]
[22]
Hodge A Berta G Doussan C Merchan F Crespi M 2009 ). Plant root growth, architecture and function.
Plant and Soil , 321 , 153 -187 .
[本文引用: 1]
[23]
Huang YM Liu D An SS 2015 ). Effects of slope aspect on soil nitrogen and microbial properties in the Chinese Loess region.
CATENA , 125 , 135 -145 .
[本文引用: 1]
[24]
Hulme PE 2008 ). Phenotypic plasticity and plant invasions: Is it all jack?
Functional Ecology , 22 , 3 -7 .
[本文引用: 1]
[25]
Kiswara W Behnke N van Avesaath P Huiskes AHL Erftemeijer PLA Bouma TJ 2009 ). Root architecture of six tropical seagrass species, growing in three contrasting habitats in Indonesian waters.
Aquatic Botany , 90 , 235 -245 .
[本文引用: 1]
[26]
Kong XP Zhang ML de Smet I Ding ZJ 2014 ). Designer crops: Optimal root system architecture for nutrient acquisition.
Trends in Biotechnology , 32 , 597 -598 .
[本文引用: 1]
[27]
Li XL Hou XY Wu XH Sa RL Ji L Chen HJ Liu ZY Ding Y 2014 ). Plastic responses of stem and leaf functional traits in Leymus chinensis to long-term grazing in a meadow steppe.
Chinese Journal of Plant Ecology , 38 , 440 -451 . (in Chinese with English abstract)
[本文引用: 1]
[李西良 , 侯向阳 , 吴新宏 , 萨茹拉 , 纪磊 , 陈海军 , 刘志英 , 丁勇 (2014 ). 草甸草原羊草茎叶功能性状对长期过度放牧的可塑性响应
. 植物生态学报 , 38 , 440 -451 .]
[本文引用: 1]
[28]
Liu J Xiang WH Xu X Chen R Tian DL Peng CH Fang X 2010 ). Analysis of architecture and functions of fine roots of five subtropical tree species in Huitong, Hunan Province, China.
Chinese Journal of Plant Ecology , 34 , 938 -945 . (in Chinese with English abstract)
[本文引用: 1]
[刘佳 , 项文化 , 徐晓 , 陈瑞 , 田大伦 , 彭长辉 , 方晰 (2010 ). 湖南会同5个亚热带树种的细根构型及功能特征分析
. 植物生态学报 , 34 , 938 -945 .]
[本文引用: 1]
[29]
Malamy JE 2005 ). Intrinsic and environmental response pathways that regulate root system architecture.
Plant, Cell & Environment , 28 , 67 -77 .
[本文引用: 1]
[30]
Mooney KA Halitschke R Kessler A Agrawal AA 2010 ). Evolutionary trade-offs in plants mediate the strength of trophic cascades.
Science , 327 , 1642 -1644 .
[本文引用: 2]
[31]
Oppelt AL Kurth W Godbold DL 2001 ). Topology, scaling relations and Leonardo’s rule in root systems from African tree species.
Tree Physiology , 21 , 117 -128 .
[本文引用: 1]
[32]
Oppelt AL Kurth W Godbold DL 2005 ). Contrasting rooting patterns of some arid-zone fruit tree species from Botswana. II. Coarse root distribution.
Agroforestry Systems , 64 , 13 -24 .
[本文引用: 1]
[33]
Pacheco-Villalobos D Hardtke CS 2012 ). Natural genetic variation of root system architecture from Arabidopsis to Brachypodium: Towards adaptive value.
Philosophical Transactions of the Royal Society B: Biological Sciences , 367 , 1552 -1558 .
[本文引用: 1]
[34]
Pyšek P Křivánek PM Jarošík V 2009 ). Planting intensity, residence time, and species traits determine invasion success of alien woody species.
Ecology , 90 , 2734 -2744 .
[本文引用: 1]
[35]
Rogers ED Benfey PN 2015 ). Regulation of plant root system architecture: Implications for crop advancement.
Current Opinion in Biotechnology , 32 , 93 -98 .
[本文引用: 2]
[36]
Sardans J Peñuelas J Estiarte M 2008 ). Changes in soil enzymes related to C and N cycle and in soil C and N content under prolonged warming and drought in a Mediterranean shrubland.
Applied Soil Ecology , 39 , 223 -235 .
[本文引用: 1]
[37]
Schenk HJ Jackson RB 2002 a). Rooting depths, lateral root spreads and below-ground/above-ground allometries of plants in water-limited ecosystems.
Journal of Ecology , 90 , 480 -494 .
[本文引用: 1]
[38]
Schenk HJ Jackson RB 2002 b). The global biogeography of roots.
Ecological Monographs , 73 , 311 -328 .
[本文引用: 1]
[39]
Shan LS Li Y Dong QL Geng DM 2012 ). Ecological adaptation of Reaumuria soongorica root system architecture to arid environment.
Journal of Desert Research , 32 , 1283 -1290 . (in Chinese with English abstract)
[本文引用: 1]
[单立山 , 李毅 , 董秋莲 , 耿东梅 (2012 ). 红砂根系构型对干旱的生态适应
. 中国沙漠 , 32 , 1283 -1290 .]
[本文引用: 1]
[40]
Shan LS Li Y Ren W Su SP Dong QL Geng DM 2013 ). Root architecture of two desert plants in central Hexi Corridor of Northwest China.
Chinese Journal of Applied Ecology , 24 , 25 -31 . (in Chinese with English abstract)
[本文引用: 1]
[单立山 , 李毅 , 任伟 , 苏世平 , 董秋莲 , 耿东梅 (2013 ). 河西走廊中部两种荒漠植物根系构型特征
. 应用生态学报 , 24 , 25 -31 .]
[本文引用: 1]
[41]
Shipley B Lechowicz MJ Wright I Reich PB 2006 ). Fundamental trade-offs generating the worldwide leaf economics spectrum.
Ecology , 87 , 535 -541 .
[本文引用: 1]
[42]
Szoboszlay M Lambers J Chappell J Kupper JV Moe LA McNear Jr DH 2015 ). Comparison of root system architecture and rhizosphere microbial communities of Balsas teosinte and domesticated corn cultivars.
Soil Biology & Biochemistry , 80 , 34 -44 .
[本文引用: 1]
[43]
Tang GA Li FY Liu XJ 2010 ). Tutorial of Digital Elevation Model . Science Press , Beijing . 149 . (in Chinese)
[本文引用: 1]
[汤国安 , 李发源 , 刘学军 (2010 ). 数字高程模型教程 . 科学出版社 , 北京 . 149 .]
[本文引用: 1]
[44]
Villordon AQ Ginzberg I Firon N 2014 ). Root architecture and root and tuber crop productivity.
Trends in Plant Science , 19 , 419 -425 .
[本文引用: 1]
[45]
Walk TC van Erp E Lynch JP 2004 ). Modelling applicability of fractal analysis to efficiency of soil exploration by roots.
Annals of Botany , 94 , 119 -128 .
[本文引用: 1]
[46]
Westoby M Wright IJ 2006 ). Land-plant ecology on the basis of functional traits.
Trends in Ecology & Evolution , 21 , 261 -268 .
[本文引用: 1]
[47]
Yang Q Zhao CZ Shi LL Dang JJ Zha GD 2014 ). Spatial autocorrelation analysis on soil moisture of Melica przewalskyi patch in a degraded alpine grassland of Qilian Mountains, Northwest China.
Chinese Journal of Ecology , 33 , 716 -722 . (in Chinese with English abstract)
[本文引用: 1]
[杨泉 , 赵成章 , 史丽丽 , 党晶晶 , 查高德 (2014 ). 祁连山地甘肃臭草斑块土壤水分的空间自相关分析
. 生态学杂志 , 33 , 716 -722 .]
[本文引用: 1]
[48]
Yang XL Zhang XM Li YL Li SC Sun HL 2008 ). Analysis of root architecture and root adaptive strategy in the Taklimakan desert area of China
. Journal of Plant Ecology (Chinese Version) , 32 , 1268 -1276 . (in Chinese with English abstract)
[本文引用: 3]
[杨小林 , 张希明 , 李义玲 , 李绍才 , 孙海龙 (2008 ). 塔克拉玛干沙漠腹地3种植物根系构型及其生境适应策略
. 植物生态学报 , 32 , 1268 -1276 .]
[本文引用: 3]
2
2004
... 植物在环境胁迫下具有权衡性状关系的适应策略(Mooney et al ., 2010 ), 通过生理和发育调节形成与环境相适应的表型特征, 有利于提高植物水分、养分利用效率等生物学功能(李西良等, 2014 ).植物根系构型对环境的适应是通过根系分支结构、空间分布格局等构型特征的改变表现出来的, 是植物长期适应环境的进化结果(Guswa, 2010 ).研究区位于北半球中纬度地区, 山地北坡光照资源最弱(殷秀琴, 2004 )、土壤含水量较高(表2 ), 草地群落的高度、盖度和生物量最高(表2 ), 甘肃臭草处于斑块形成的初期阶段, 其种群密度较小, 也没有高度优势(表1 , 表2 ), 与草地群落的优势种群西北针茅相比, 处于种间竞争的劣势地位.在种间胁迫较大的情况下, 生长在北坡的甘肃臭草采取了增大根系分叉数、减小根系连接长度的根系构型构建策略; 根系分叉数的增加能够增强根系重叠程度、构建密集的根系网络系统, 有利于甘肃臭草地下根系系统和地上株丛的密集生长, 增强种群的密集分布强度, 提高根系对土壤资源的吸收和利用效率, 以斑块式抱团生长实现种群内个体间的高度协同; 减小根系连接长度可能会缩短甘肃臭草克隆分株间的资源传输距离, 有利于增强甘肃臭草克隆分株个体间的相互联系, 保证种群内分株间的资源连接和共享, 从而使种群能够在激烈的资源竞争中存活下来, 与相关学者得出一致的结论, 即“分叉数较大的植物能够迅速占据大量土壤空间快速生长, 适宜生长在土壤资源丰富的环境中” (周艳松和王立群, 2011 ; 郭京衡等, 2014 ).因此, 增大分叉数、减小连接长度是分布于北坡的甘肃臭草增大斑块密度, 提升竞争能力和空间拓殖能力的一种生活史对策, 使根系分叉数和连接长度之间的权衡关系达到极显著水平(p < 0.01, 图2 ). ...
... 植物根系构型构建策略的差异可以用根系碳投入与土壤水分养分收益的权衡关系来解释(Oppelt et al ., 2001 ).根系分叉数的增加会导致根系重叠加剧, 增加根系内部的资源竞争态势(杨小林等, 2008 ).根系连接长度的变化会影响根系空间拓展能力和利用土壤资源的有效性(Schenk & Jackson, 2002a , 2002b ), 这与植物自身根系碳投入与土壤水分养分的收益权衡相契合, 是提高植物资源获取能力的一个重要策略(杨小林等, 2008 ).相对于北坡, 山地南坡日照时间长、水分蒸发量大(殷秀琴, 2004 ), 生长在南坡的甘肃臭草多呈直径不等的单一优势种群斑块, 占据了草地群落的绝对资源优势(表1 , 表2 ).甘肃臭草采用克隆繁殖模式形成了庞大的互相连通交织的根茎系统, 构建了高度聚集的根系网络(高福元和赵成章, 2012 ), 斑块内部种群聚集程度较高, 斑块中心土壤含水量明显低于斑块边缘的土壤含水量(赵成章等, 2011 ), 体现了甘肃臭草向斑块周围扩张的迫切需求.在地上和地下生存空间拥挤、土壤含水量分布不均衡状态下, 为了避免不必要的种内竞争, 甘肃臭草限制了地下生物量的分配比例, 以减缓营养繁殖速度, 降低种群聚集强度, 此时甘肃臭草选择了增加根系连接长度、减少分叉数的根系构型构建模式.根系分叉数的减小能够降低种群内部根系重叠程度, 避免不必要的种内竞争; 连接长度的增大可以增强根系空间拓展能力, 扩大单位根系所占的土壤体空间, 使根系从斑块中心向土壤水分较好、生长发育空间较大的边缘区域拓殖, 是南坡甘肃臭草单优种群斑块扩张的主要动力, 与赵成章等(2011) 关于甘肃臭草通过营养繁殖以类似“同心圆”的方式不断由斑块中心向边缘扩大种群领地的观点一致.这符合相关学者“当植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 增加根系连接长度等策略, 以寻求更多的水分和养分, 保证自身的生存和生长”的结论(单立山等, 2013 ; 杜建会等, 2014 ).因此, 南坡甘肃臭草根系分叉数和连接长度间呈极显著负相关关系(p < 0.01, 图2 ), 减小分叉数、增加连接长度是甘肃臭草斑块扩张的根系构型构建策略, 降低了甘肃臭草单一优势种群斑块内部的根系拥挤程度. ...
2
2004
... 植物在环境胁迫下具有权衡性状关系的适应策略(Mooney et al ., 2010 ), 通过生理和发育调节形成与环境相适应的表型特征, 有利于提高植物水分、养分利用效率等生物学功能(李西良等, 2014 ).植物根系构型对环境的适应是通过根系分支结构、空间分布格局等构型特征的改变表现出来的, 是植物长期适应环境的进化结果(Guswa, 2010 ).研究区位于北半球中纬度地区, 山地北坡光照资源最弱(殷秀琴, 2004 )、土壤含水量较高(表2 ), 草地群落的高度、盖度和生物量最高(表2 ), 甘肃臭草处于斑块形成的初期阶段, 其种群密度较小, 也没有高度优势(表1 , 表2 ), 与草地群落的优势种群西北针茅相比, 处于种间竞争的劣势地位.在种间胁迫较大的情况下, 生长在北坡的甘肃臭草采取了增大根系分叉数、减小根系连接长度的根系构型构建策略; 根系分叉数的增加能够增强根系重叠程度、构建密集的根系网络系统, 有利于甘肃臭草地下根系系统和地上株丛的密集生长, 增强种群的密集分布强度, 提高根系对土壤资源的吸收和利用效率, 以斑块式抱团生长实现种群内个体间的高度协同; 减小根系连接长度可能会缩短甘肃臭草克隆分株间的资源传输距离, 有利于增强甘肃臭草克隆分株个体间的相互联系, 保证种群内分株间的资源连接和共享, 从而使种群能够在激烈的资源竞争中存活下来, 与相关学者得出一致的结论, 即“分叉数较大的植物能够迅速占据大量土壤空间快速生长, 适宜生长在土壤资源丰富的环境中” (周艳松和王立群, 2011 ; 郭京衡等, 2014 ).因此, 增大分叉数、减小连接长度是分布于北坡的甘肃臭草增大斑块密度, 提升竞争能力和空间拓殖能力的一种生活史对策, 使根系分叉数和连接长度之间的权衡关系达到极显著水平(p < 0.01, 图2 ). ...
... 植物根系构型构建策略的差异可以用根系碳投入与土壤水分养分收益的权衡关系来解释(Oppelt et al ., 2001 ).根系分叉数的增加会导致根系重叠加剧, 增加根系内部的资源竞争态势(杨小林等, 2008 ).根系连接长度的变化会影响根系空间拓展能力和利用土壤资源的有效性(Schenk & Jackson, 2002a , 2002b ), 这与植物自身根系碳投入与土壤水分养分的收益权衡相契合, 是提高植物资源获取能力的一个重要策略(杨小林等, 2008 ).相对于北坡, 山地南坡日照时间长、水分蒸发量大(殷秀琴, 2004 ), 生长在南坡的甘肃臭草多呈直径不等的单一优势种群斑块, 占据了草地群落的绝对资源优势(表1 , 表2 ).甘肃臭草采用克隆繁殖模式形成了庞大的互相连通交织的根茎系统, 构建了高度聚集的根系网络(高福元和赵成章, 2012 ), 斑块内部种群聚集程度较高, 斑块中心土壤含水量明显低于斑块边缘的土壤含水量(赵成章等, 2011 ), 体现了甘肃臭草向斑块周围扩张的迫切需求.在地上和地下生存空间拥挤、土壤含水量分布不均衡状态下, 为了避免不必要的种内竞争, 甘肃臭草限制了地下生物量的分配比例, 以减缓营养繁殖速度, 降低种群聚集强度, 此时甘肃臭草选择了增加根系连接长度、减少分叉数的根系构型构建模式.根系分叉数的减小能够降低种群内部根系重叠程度, 避免不必要的种内竞争; 连接长度的增大可以增强根系空间拓展能力, 扩大单位根系所占的土壤体空间, 使根系从斑块中心向土壤水分较好、生长发育空间较大的边缘区域拓殖, 是南坡甘肃臭草单优种群斑块扩张的主要动力, 与赵成章等(2011) 关于甘肃臭草通过营养繁殖以类似“同心圆”的方式不断由斑块中心向边缘扩大种群领地的观点一致.这符合相关学者“当植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 增加根系连接长度等策略, 以寻求更多的水分和养分, 保证自身的生存和生长”的结论(单立山等, 2013 ; 杜建会等, 2014 ).因此, 南坡甘肃臭草根系分叉数和连接长度间呈极显著负相关关系(p < 0.01, 图2 ), 减小分叉数、增加连接长度是甘肃臭草斑块扩张的根系构型构建策略, 降低了甘肃臭草单一优势种群斑块内部的根系拥挤程度. ...
高寒退化草地甘肃臭草种群分布格局及其对土壤水分的响应
3
2011
... 近年来, 多数学者对植物根系的研究主要集中在异质生境中根系功能性状可塑性(Grossman & Rice, 2012 ; Pacheco-Villalobos & Hardtk, 2012 )、不同群落环境下根系构型的适应性调整(Rogers & Benfey, 2015 )、根系生长与土壤环境的关系(Forde, 2014 ; Szoboszlay et al ., 2015 )、根系结构和功能(Hodge et al ., 2009 )等方面, 关于退化草地中植物构件生物量分配模式(周兵等, 2015 )、影响根系构型的主要功能性状(周艳松和王立群, 2011 ), 以及不同入侵阶段植物功能性状表型可塑性(Hulme, 2008 ; Pyšek et al ., 2009 )等方面也进行了系统研究, 而从根系构型构建策略的角度出发, 对生境梯度植物根系功能性状间资源权衡机制的研究仍显不足.甘肃臭草(Melica przewalskyi )是禾本科臭草属多年生根茎植物, 通过根状茎的快速横向生长, 长出幼芽和根系, 形成新植株进行无性繁殖, 在地下建立庞大的根茎系统, 形成密集的圆形株丛, 其分株间的间隔子短, 分枝角度小, 能够在竞争过程中占据优势地位.近年来, 在祁连山北坡中山区荒漠草原与典型草原过渡带, 甘肃臭草替代了原生地带性植被的优势种西北针茅(Stipa sareptana var. krylovii ), 形成大量面积不等、繁茂生长的单一优势种群落斑块(赵成章和龙瑞军, 2008 ), 表现出较高的形态可塑性和对逆境的适应能力, 其强大的入侵能力使众多学者对其资源利用策略和生长扩散过程产生了兴趣.目前, 对甘肃臭草型退化草地优势物种空间格局分析及其对水分的响应(赵成章等, 2011 ; 高福元和赵成章, 2012 )、甘肃臭草斑块土壤水分空间异质性(杨泉等, 2014 ), 以及甘肃臭草茎-叶性状、叶内性状(党晶晶等, 2014 ,2015 )等方面进行了研究, 而生境梯度上甘肃臭草根系构型的可塑性变化尚未见报道.鉴于此, 本文在野外调查的基础上, 试图弄清以下问题: (1)不同坡向甘肃臭草根系分叉数和连接长度的权衡关系如何?(2)导致甘肃臭草根系分叉数和连接长度变化的原因有哪些?旨在探讨祁连山地甘肃臭草种群根系构型构建的资源配置模式, 进一步认识甘肃臭草单一优势种群斑块形成机制和扩散机理. ...
... 植物根系构型构建策略的差异可以用根系碳投入与土壤水分养分收益的权衡关系来解释(Oppelt et al ., 2001 ).根系分叉数的增加会导致根系重叠加剧, 增加根系内部的资源竞争态势(杨小林等, 2008 ).根系连接长度的变化会影响根系空间拓展能力和利用土壤资源的有效性(Schenk & Jackson, 2002a , 2002b ), 这与植物自身根系碳投入与土壤水分养分的收益权衡相契合, 是提高植物资源获取能力的一个重要策略(杨小林等, 2008 ).相对于北坡, 山地南坡日照时间长、水分蒸发量大(殷秀琴, 2004 ), 生长在南坡的甘肃臭草多呈直径不等的单一优势种群斑块, 占据了草地群落的绝对资源优势(表1 , 表2 ).甘肃臭草采用克隆繁殖模式形成了庞大的互相连通交织的根茎系统, 构建了高度聚集的根系网络(高福元和赵成章, 2012 ), 斑块内部种群聚集程度较高, 斑块中心土壤含水量明显低于斑块边缘的土壤含水量(赵成章等, 2011 ), 体现了甘肃臭草向斑块周围扩张的迫切需求.在地上和地下生存空间拥挤、土壤含水量分布不均衡状态下, 为了避免不必要的种内竞争, 甘肃臭草限制了地下生物量的分配比例, 以减缓营养繁殖速度, 降低种群聚集强度, 此时甘肃臭草选择了增加根系连接长度、减少分叉数的根系构型构建模式.根系分叉数的减小能够降低种群内部根系重叠程度, 避免不必要的种内竞争; 连接长度的增大可以增强根系空间拓展能力, 扩大单位根系所占的土壤体空间, 使根系从斑块中心向土壤水分较好、生长发育空间较大的边缘区域拓殖, 是南坡甘肃臭草单优种群斑块扩张的主要动力, 与赵成章等(2011) 关于甘肃臭草通过营养繁殖以类似“同心圆”的方式不断由斑块中心向边缘扩大种群领地的观点一致.这符合相关学者“当植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 增加根系连接长度等策略, 以寻求更多的水分和养分, 保证自身的生存和生长”的结论(单立山等, 2013 ; 杜建会等, 2014 ).因此, 南坡甘肃臭草根系分叉数和连接长度间呈极显著负相关关系(p < 0.01, 图2 ), 减小分叉数、增加连接长度是甘肃臭草斑块扩张的根系构型构建策略, 降低了甘肃臭草单一优势种群斑块内部的根系拥挤程度. ...
... ), 体现了甘肃臭草向斑块周围扩张的迫切需求.在地上和地下生存空间拥挤、土壤含水量分布不均衡状态下, 为了避免不必要的种内竞争, 甘肃臭草限制了地下生物量的分配比例, 以减缓营养繁殖速度, 降低种群聚集强度, 此时甘肃臭草选择了增加根系连接长度、减少分叉数的根系构型构建模式.根系分叉数的减小能够降低种群内部根系重叠程度, 避免不必要的种内竞争; 连接长度的增大可以增强根系空间拓展能力, 扩大单位根系所占的土壤体空间, 使根系从斑块中心向土壤水分较好、生长发育空间较大的边缘区域拓殖, 是南坡甘肃臭草单优种群斑块扩张的主要动力, 与赵成章等(2011) 关于甘肃臭草通过营养繁殖以类似“同心圆”的方式不断由斑块中心向边缘扩大种群领地的观点一致.这符合相关学者“当植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 增加根系连接长度等策略, 以寻求更多的水分和养分, 保证自身的生存和生长”的结论(单立山等, 2013 ; 杜建会等, 2014 ).因此, 南坡甘肃臭草根系分叉数和连接长度间呈极显著负相关关系(p < 0.01, 图2 ), 减小分叉数、增加连接长度是甘肃臭草斑块扩张的根系构型构建策略, 降低了甘肃臭草单一优势种群斑块内部的根系拥挤程度. ...
高寒退化草地甘肃臭草种群分布格局及其对土壤水分的响应
3
2011
... 近年来, 多数学者对植物根系的研究主要集中在异质生境中根系功能性状可塑性(Grossman & Rice, 2012 ; Pacheco-Villalobos & Hardtk, 2012 )、不同群落环境下根系构型的适应性调整(Rogers & Benfey, 2015 )、根系生长与土壤环境的关系(Forde, 2014 ; Szoboszlay et al ., 2015 )、根系结构和功能(Hodge et al ., 2009 )等方面, 关于退化草地中植物构件生物量分配模式(周兵等, 2015 )、影响根系构型的主要功能性状(周艳松和王立群, 2011 ), 以及不同入侵阶段植物功能性状表型可塑性(Hulme, 2008 ; Pyšek et al ., 2009 )等方面也进行了系统研究, 而从根系构型构建策略的角度出发, 对生境梯度植物根系功能性状间资源权衡机制的研究仍显不足.甘肃臭草(Melica przewalskyi )是禾本科臭草属多年生根茎植物, 通过根状茎的快速横向生长, 长出幼芽和根系, 形成新植株进行无性繁殖, 在地下建立庞大的根茎系统, 形成密集的圆形株丛, 其分株间的间隔子短, 分枝角度小, 能够在竞争过程中占据优势地位.近年来, 在祁连山北坡中山区荒漠草原与典型草原过渡带, 甘肃臭草替代了原生地带性植被的优势种西北针茅(Stipa sareptana var. krylovii ), 形成大量面积不等、繁茂生长的单一优势种群落斑块(赵成章和龙瑞军, 2008 ), 表现出较高的形态可塑性和对逆境的适应能力, 其强大的入侵能力使众多学者对其资源利用策略和生长扩散过程产生了兴趣.目前, 对甘肃臭草型退化草地优势物种空间格局分析及其对水分的响应(赵成章等, 2011 ; 高福元和赵成章, 2012 )、甘肃臭草斑块土壤水分空间异质性(杨泉等, 2014 ), 以及甘肃臭草茎-叶性状、叶内性状(党晶晶等, 2014 ,2015 )等方面进行了研究, 而生境梯度上甘肃臭草根系构型的可塑性变化尚未见报道.鉴于此, 本文在野外调查的基础上, 试图弄清以下问题: (1)不同坡向甘肃臭草根系分叉数和连接长度的权衡关系如何?(2)导致甘肃臭草根系分叉数和连接长度变化的原因有哪些?旨在探讨祁连山地甘肃臭草种群根系构型构建的资源配置模式, 进一步认识甘肃臭草单一优势种群斑块形成机制和扩散机理. ...
... 植物根系构型构建策略的差异可以用根系碳投入与土壤水分养分收益的权衡关系来解释(Oppelt et al ., 2001 ).根系分叉数的增加会导致根系重叠加剧, 增加根系内部的资源竞争态势(杨小林等, 2008 ).根系连接长度的变化会影响根系空间拓展能力和利用土壤资源的有效性(Schenk & Jackson, 2002a , 2002b ), 这与植物自身根系碳投入与土壤水分养分的收益权衡相契合, 是提高植物资源获取能力的一个重要策略(杨小林等, 2008 ).相对于北坡, 山地南坡日照时间长、水分蒸发量大(殷秀琴, 2004 ), 生长在南坡的甘肃臭草多呈直径不等的单一优势种群斑块, 占据了草地群落的绝对资源优势(表1 , 表2 ).甘肃臭草采用克隆繁殖模式形成了庞大的互相连通交织的根茎系统, 构建了高度聚集的根系网络(高福元和赵成章, 2012 ), 斑块内部种群聚集程度较高, 斑块中心土壤含水量明显低于斑块边缘的土壤含水量(赵成章等, 2011 ), 体现了甘肃臭草向斑块周围扩张的迫切需求.在地上和地下生存空间拥挤、土壤含水量分布不均衡状态下, 为了避免不必要的种内竞争, 甘肃臭草限制了地下生物量的分配比例, 以减缓营养繁殖速度, 降低种群聚集强度, 此时甘肃臭草选择了增加根系连接长度、减少分叉数的根系构型构建模式.根系分叉数的减小能够降低种群内部根系重叠程度, 避免不必要的种内竞争; 连接长度的增大可以增强根系空间拓展能力, 扩大单位根系所占的土壤体空间, 使根系从斑块中心向土壤水分较好、生长发育空间较大的边缘区域拓殖, 是南坡甘肃臭草单优种群斑块扩张的主要动力, 与赵成章等(2011) 关于甘肃臭草通过营养繁殖以类似“同心圆”的方式不断由斑块中心向边缘扩大种群领地的观点一致.这符合相关学者“当植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 增加根系连接长度等策略, 以寻求更多的水分和养分, 保证自身的生存和生长”的结论(单立山等, 2013 ; 杜建会等, 2014 ).因此, 南坡甘肃臭草根系分叉数和连接长度间呈极显著负相关关系(p < 0.01, 图2 ), 减小分叉数、增加连接长度是甘肃臭草斑块扩张的根系构型构建策略, 降低了甘肃臭草单一优势种群斑块内部的根系拥挤程度. ...
... ), 体现了甘肃臭草向斑块周围扩张的迫切需求.在地上和地下生存空间拥挤、土壤含水量分布不均衡状态下, 为了避免不必要的种内竞争, 甘肃臭草限制了地下生物量的分配比例, 以减缓营养繁殖速度, 降低种群聚集强度, 此时甘肃臭草选择了增加根系连接长度、减少分叉数的根系构型构建模式.根系分叉数的减小能够降低种群内部根系重叠程度, 避免不必要的种内竞争; 连接长度的增大可以增强根系空间拓展能力, 扩大单位根系所占的土壤体空间, 使根系从斑块中心向土壤水分较好、生长发育空间较大的边缘区域拓殖, 是南坡甘肃臭草单优种群斑块扩张的主要动力, 与赵成章等(2011) 关于甘肃臭草通过营养繁殖以类似“同心圆”的方式不断由斑块中心向边缘扩大种群领地的观点一致.这符合相关学者“当植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 增加根系连接长度等策略, 以寻求更多的水分和养分, 保证自身的生存和生长”的结论(单立山等, 2013 ; 杜建会等, 2014 ).因此, 南坡甘肃臭草根系分叉数和连接长度间呈极显著负相关关系(p < 0.01, 图2 ), 减小分叉数、增加连接长度是甘肃臭草斑块扩张的根系构型构建策略, 降低了甘肃臭草单一优势种群斑块内部的根系拥挤程度. ...
石羊河上游甘肃臭草型退化草地植被恢复过程
1
2008
... 近年来, 多数学者对植物根系的研究主要集中在异质生境中根系功能性状可塑性(Grossman & Rice, 2012 ; Pacheco-Villalobos & Hardtk, 2012 )、不同群落环境下根系构型的适应性调整(Rogers & Benfey, 2015 )、根系生长与土壤环境的关系(Forde, 2014 ; Szoboszlay et al ., 2015 )、根系结构和功能(Hodge et al ., 2009 )等方面, 关于退化草地中植物构件生物量分配模式(周兵等, 2015 )、影响根系构型的主要功能性状(周艳松和王立群, 2011 ), 以及不同入侵阶段植物功能性状表型可塑性(Hulme, 2008 ; Pyšek et al ., 2009 )等方面也进行了系统研究, 而从根系构型构建策略的角度出发, 对生境梯度植物根系功能性状间资源权衡机制的研究仍显不足.甘肃臭草(Melica przewalskyi )是禾本科臭草属多年生根茎植物, 通过根状茎的快速横向生长, 长出幼芽和根系, 形成新植株进行无性繁殖, 在地下建立庞大的根茎系统, 形成密集的圆形株丛, 其分株间的间隔子短, 分枝角度小, 能够在竞争过程中占据优势地位.近年来, 在祁连山北坡中山区荒漠草原与典型草原过渡带, 甘肃臭草替代了原生地带性植被的优势种西北针茅(Stipa sareptana var. krylovii ), 形成大量面积不等、繁茂生长的单一优势种群落斑块(赵成章和龙瑞军, 2008 ), 表现出较高的形态可塑性和对逆境的适应能力, 其强大的入侵能力使众多学者对其资源利用策略和生长扩散过程产生了兴趣.目前, 对甘肃臭草型退化草地优势物种空间格局分析及其对水分的响应(赵成章等, 2011 ; 高福元和赵成章, 2012 )、甘肃臭草斑块土壤水分空间异质性(杨泉等, 2014 ), 以及甘肃臭草茎-叶性状、叶内性状(党晶晶等, 2014 ,2015 )等方面进行了研究, 而生境梯度上甘肃臭草根系构型的可塑性变化尚未见报道.鉴于此, 本文在野外调查的基础上, 试图弄清以下问题: (1)不同坡向甘肃臭草根系分叉数和连接长度的权衡关系如何?(2)导致甘肃臭草根系分叉数和连接长度变化的原因有哪些?旨在探讨祁连山地甘肃臭草种群根系构型构建的资源配置模式, 进一步认识甘肃臭草单一优势种群斑块形成机制和扩散机理. ...
石羊河上游甘肃臭草型退化草地植被恢复过程
1
2008
... 近年来, 多数学者对植物根系的研究主要集中在异质生境中根系功能性状可塑性(Grossman & Rice, 2012 ; Pacheco-Villalobos & Hardtk, 2012 )、不同群落环境下根系构型的适应性调整(Rogers & Benfey, 2015 )、根系生长与土壤环境的关系(Forde, 2014 ; Szoboszlay et al ., 2015 )、根系结构和功能(Hodge et al ., 2009 )等方面, 关于退化草地中植物构件生物量分配模式(周兵等, 2015 )、影响根系构型的主要功能性状(周艳松和王立群, 2011 ), 以及不同入侵阶段植物功能性状表型可塑性(Hulme, 2008 ; Pyšek et al ., 2009 )等方面也进行了系统研究, 而从根系构型构建策略的角度出发, 对生境梯度植物根系功能性状间资源权衡机制的研究仍显不足.甘肃臭草(Melica przewalskyi )是禾本科臭草属多年生根茎植物, 通过根状茎的快速横向生长, 长出幼芽和根系, 形成新植株进行无性繁殖, 在地下建立庞大的根茎系统, 形成密集的圆形株丛, 其分株间的间隔子短, 分枝角度小, 能够在竞争过程中占据优势地位.近年来, 在祁连山北坡中山区荒漠草原与典型草原过渡带, 甘肃臭草替代了原生地带性植被的优势种西北针茅(Stipa sareptana var. krylovii ), 形成大量面积不等、繁茂生长的单一优势种群落斑块(赵成章和龙瑞军, 2008 ), 表现出较高的形态可塑性和对逆境的适应能力, 其强大的入侵能力使众多学者对其资源利用策略和生长扩散过程产生了兴趣.目前, 对甘肃臭草型退化草地优势物种空间格局分析及其对水分的响应(赵成章等, 2011 ; 高福元和赵成章, 2012 )、甘肃臭草斑块土壤水分空间异质性(杨泉等, 2014 ), 以及甘肃臭草茎-叶性状、叶内性状(党晶晶等, 2014 ,2015 )等方面进行了研究, 而生境梯度上甘肃臭草根系构型的可塑性变化尚未见报道.鉴于此, 本文在野外调查的基础上, 试图弄清以下问题: (1)不同坡向甘肃臭草根系分叉数和连接长度的权衡关系如何?(2)导致甘肃臭草根系分叉数和连接长度变化的原因有哪些?旨在探讨祁连山地甘肃臭草种群根系构型构建的资源配置模式, 进一步认识甘肃臭草单一优势种群斑块形成机制和扩散机理. ...
不同生境下入侵植物胜红蓟种群构件生物量分配特性
1
2015
... 近年来, 多数学者对植物根系的研究主要集中在异质生境中根系功能性状可塑性(Grossman & Rice, 2012 ; Pacheco-Villalobos & Hardtk, 2012 )、不同群落环境下根系构型的适应性调整(Rogers & Benfey, 2015 )、根系生长与土壤环境的关系(Forde, 2014 ; Szoboszlay et al ., 2015 )、根系结构和功能(Hodge et al ., 2009 )等方面, 关于退化草地中植物构件生物量分配模式(周兵等, 2015 )、影响根系构型的主要功能性状(周艳松和王立群, 2011 ), 以及不同入侵阶段植物功能性状表型可塑性(Hulme, 2008 ; Pyšek et al ., 2009 )等方面也进行了系统研究, 而从根系构型构建策略的角度出发, 对生境梯度植物根系功能性状间资源权衡机制的研究仍显不足.甘肃臭草(Melica przewalskyi )是禾本科臭草属多年生根茎植物, 通过根状茎的快速横向生长, 长出幼芽和根系, 形成新植株进行无性繁殖, 在地下建立庞大的根茎系统, 形成密集的圆形株丛, 其分株间的间隔子短, 分枝角度小, 能够在竞争过程中占据优势地位.近年来, 在祁连山北坡中山区荒漠草原与典型草原过渡带, 甘肃臭草替代了原生地带性植被的优势种西北针茅(Stipa sareptana var. krylovii ), 形成大量面积不等、繁茂生长的单一优势种群落斑块(赵成章和龙瑞军, 2008 ), 表现出较高的形态可塑性和对逆境的适应能力, 其强大的入侵能力使众多学者对其资源利用策略和生长扩散过程产生了兴趣.目前, 对甘肃臭草型退化草地优势物种空间格局分析及其对水分的响应(赵成章等, 2011 ; 高福元和赵成章, 2012 )、甘肃臭草斑块土壤水分空间异质性(杨泉等, 2014 ), 以及甘肃臭草茎-叶性状、叶内性状(党晶晶等, 2014 ,2015 )等方面进行了研究, 而生境梯度上甘肃臭草根系构型的可塑性变化尚未见报道.鉴于此, 本文在野外调查的基础上, 试图弄清以下问题: (1)不同坡向甘肃臭草根系分叉数和连接长度的权衡关系如何?(2)导致甘肃臭草根系分叉数和连接长度变化的原因有哪些?旨在探讨祁连山地甘肃臭草种群根系构型构建的资源配置模式, 进一步认识甘肃臭草单一优势种群斑块形成机制和扩散机理. ...
不同生境下入侵植物胜红蓟种群构件生物量分配特性
1
2015
... 近年来, 多数学者对植物根系的研究主要集中在异质生境中根系功能性状可塑性(Grossman & Rice, 2012 ; Pacheco-Villalobos & Hardtk, 2012 )、不同群落环境下根系构型的适应性调整(Rogers & Benfey, 2015 )、根系生长与土壤环境的关系(Forde, 2014 ; Szoboszlay et al ., 2015 )、根系结构和功能(Hodge et al ., 2009 )等方面, 关于退化草地中植物构件生物量分配模式(周兵等, 2015 )、影响根系构型的主要功能性状(周艳松和王立群, 2011 ), 以及不同入侵阶段植物功能性状表型可塑性(Hulme, 2008 ; Pyšek et al ., 2009 )等方面也进行了系统研究, 而从根系构型构建策略的角度出发, 对生境梯度植物根系功能性状间资源权衡机制的研究仍显不足.甘肃臭草(Melica przewalskyi )是禾本科臭草属多年生根茎植物, 通过根状茎的快速横向生长, 长出幼芽和根系, 形成新植株进行无性繁殖, 在地下建立庞大的根茎系统, 形成密集的圆形株丛, 其分株间的间隔子短, 分枝角度小, 能够在竞争过程中占据优势地位.近年来, 在祁连山北坡中山区荒漠草原与典型草原过渡带, 甘肃臭草替代了原生地带性植被的优势种西北针茅(Stipa sareptana var. krylovii ), 形成大量面积不等、繁茂生长的单一优势种群落斑块(赵成章和龙瑞军, 2008 ), 表现出较高的形态可塑性和对逆境的适应能力, 其强大的入侵能力使众多学者对其资源利用策略和生长扩散过程产生了兴趣.目前, 对甘肃臭草型退化草地优势物种空间格局分析及其对水分的响应(赵成章等, 2011 ; 高福元和赵成章, 2012 )、甘肃臭草斑块土壤水分空间异质性(杨泉等, 2014 ), 以及甘肃臭草茎-叶性状、叶内性状(党晶晶等, 2014 ,2015 )等方面进行了研究, 而生境梯度上甘肃臭草根系构型的可塑性变化尚未见报道.鉴于此, 本文在野外调查的基础上, 试图弄清以下问题: (1)不同坡向甘肃臭草根系分叉数和连接长度的权衡关系如何?(2)导致甘肃臭草根系分叉数和连接长度变化的原因有哪些?旨在探讨祁连山地甘肃臭草种群根系构型构建的资源配置模式, 进一步认识甘肃臭草单一优势种群斑块形成机制和扩散机理. ...
星毛委陵菜根系构型对草原退化的生态适应
3
2011
... 植物的生态策略和适应性、响应性功能要通过植物内部与外部不同功能性状来实现(Bernard- Verdier et al ., 2012 ), 性状的构建、稳固和性状之间的强弱变化与替补转换则靠植物自身的资源权衡和补偿机制来调控(Fortunel et al ., 2012 ; 陈莹婷和许振柱, 2014 ), 是植物在特定环境下生存策略的重要表现(Mooney et al ., 2010 ).根系构型决定了根系在土壤空间中的位置和资源获取方式, 是植物与资源环境异质性相互适应的结果(Malamy, 2005 ; Rogers & Benfey, 2015 ).根系分叉数和连接长度作为根系构型的重要特征指标, 受土壤空间异质性(Villordon et al ., 2014 )、群落环境(单立山等, 2012 )、遗传特性(Kiswara et al ., 2009 ; Bingham & Wu, 2011 )等诸多因素的影响, 在其生存和发展的过程中, 会根据所处生境的资源条件, 调整、转换或补偿自身功能, 实现根系功能性状间的资源权衡.根系分叉数的变化会影响根系的空间分布属性、营养吸收能力和固定作用; 根系连接长度的大小直接决定了根系在土壤中的空间拓展和营养物质吸收能力(Walk et al ., 2004 ), 二者的权衡关系直接反映了植物根系吸收效率和根系构建过程中所需的碳量(Fitter & Stickland, 1991 ).当根系处于水分状态良好的土层中时, 为了提高水分利用率, 会形成更多的侧根, 增大根分叉数(周艳松和王立群, 2011 ); 在干旱胁迫环境中, 较大的分叉数会引起根系内部对营养物质的竞争(杨小林等, 2008 ), 此时植物倾向于增大连接长度, 以减少根系之间的交叠重复, 占据较大的水分和养分资源空间(郭京衡等, 2014 ), 这是植物为适应不同生境条件而形成的获取资源策略, 以达到“生存、生长、繁殖”之三大目的.地形作为生境条件的一种综合指示, 往往能营造局部小气候(Cantón et al ., 2004 ).坡向作为一种重要的地形要素(Fekedulegn et al ., 2003 ), 可以对局部区域的太阳辐射和降水进行二次分配, 其空间变化是群落组成、物种多样性、生态系统过程和植物功能性状的重要决定因素(de Bello et al ., 2006 ; Bennie et al ., 2008 ; Carletti et al ., 2009 ).在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 在各个水平上影响了植物生长及其对水分利用的有效性(Sardans et al ., 2008 ; Huang et al ., 2015 ), 进而引起植物碳资源分配格局和构型构建策略的改变, 为提高生境适应性和风险规避能力, 植物对连接长度和分叉数等主要根系功能性状进行资源优化配置, 构建与生境相适应的根系构型, 优化根系营养吸收和传输系统.因此, 研究坡向梯度上植物根系构型的生态适应策略, 对于理解植物的环境适应机制有重要意义. ...
... 近年来, 多数学者对植物根系的研究主要集中在异质生境中根系功能性状可塑性(Grossman & Rice, 2012 ; Pacheco-Villalobos & Hardtk, 2012 )、不同群落环境下根系构型的适应性调整(Rogers & Benfey, 2015 )、根系生长与土壤环境的关系(Forde, 2014 ; Szoboszlay et al ., 2015 )、根系结构和功能(Hodge et al ., 2009 )等方面, 关于退化草地中植物构件生物量分配模式(周兵等, 2015 )、影响根系构型的主要功能性状(周艳松和王立群, 2011 ), 以及不同入侵阶段植物功能性状表型可塑性(Hulme, 2008 ; Pyšek et al ., 2009 )等方面也进行了系统研究, 而从根系构型构建策略的角度出发, 对生境梯度植物根系功能性状间资源权衡机制的研究仍显不足.甘肃臭草(Melica przewalskyi )是禾本科臭草属多年生根茎植物, 通过根状茎的快速横向生长, 长出幼芽和根系, 形成新植株进行无性繁殖, 在地下建立庞大的根茎系统, 形成密集的圆形株丛, 其分株间的间隔子短, 分枝角度小, 能够在竞争过程中占据优势地位.近年来, 在祁连山北坡中山区荒漠草原与典型草原过渡带, 甘肃臭草替代了原生地带性植被的优势种西北针茅(Stipa sareptana var. krylovii ), 形成大量面积不等、繁茂生长的单一优势种群落斑块(赵成章和龙瑞军, 2008 ), 表现出较高的形态可塑性和对逆境的适应能力, 其强大的入侵能力使众多学者对其资源利用策略和生长扩散过程产生了兴趣.目前, 对甘肃臭草型退化草地优势物种空间格局分析及其对水分的响应(赵成章等, 2011 ; 高福元和赵成章, 2012 )、甘肃臭草斑块土壤水分空间异质性(杨泉等, 2014 ), 以及甘肃臭草茎-叶性状、叶内性状(党晶晶等, 2014 ,2015 )等方面进行了研究, 而生境梯度上甘肃臭草根系构型的可塑性变化尚未见报道.鉴于此, 本文在野外调查的基础上, 试图弄清以下问题: (1)不同坡向甘肃臭草根系分叉数和连接长度的权衡关系如何?(2)导致甘肃臭草根系分叉数和连接长度变化的原因有哪些?旨在探讨祁连山地甘肃臭草种群根系构型构建的资源配置模式, 进一步认识甘肃臭草单一优势种群斑块形成机制和扩散机理. ...
... 植物在环境胁迫下具有权衡性状关系的适应策略(Mooney et al ., 2010 ), 通过生理和发育调节形成与环境相适应的表型特征, 有利于提高植物水分、养分利用效率等生物学功能(李西良等, 2014 ).植物根系构型对环境的适应是通过根系分支结构、空间分布格局等构型特征的改变表现出来的, 是植物长期适应环境的进化结果(Guswa, 2010 ).研究区位于北半球中纬度地区, 山地北坡光照资源最弱(殷秀琴, 2004 )、土壤含水量较高(表2 ), 草地群落的高度、盖度和生物量最高(表2 ), 甘肃臭草处于斑块形成的初期阶段, 其种群密度较小, 也没有高度优势(表1 , 表2 ), 与草地群落的优势种群西北针茅相比, 处于种间竞争的劣势地位.在种间胁迫较大的情况下, 生长在北坡的甘肃臭草采取了增大根系分叉数、减小根系连接长度的根系构型构建策略; 根系分叉数的增加能够增强根系重叠程度、构建密集的根系网络系统, 有利于甘肃臭草地下根系系统和地上株丛的密集生长, 增强种群的密集分布强度, 提高根系对土壤资源的吸收和利用效率, 以斑块式抱团生长实现种群内个体间的高度协同; 减小根系连接长度可能会缩短甘肃臭草克隆分株间的资源传输距离, 有利于增强甘肃臭草克隆分株个体间的相互联系, 保证种群内分株间的资源连接和共享, 从而使种群能够在激烈的资源竞争中存活下来, 与相关学者得出一致的结论, 即“分叉数较大的植物能够迅速占据大量土壤空间快速生长, 适宜生长在土壤资源丰富的环境中” (周艳松和王立群, 2011 ; 郭京衡等, 2014 ).因此, 增大分叉数、减小连接长度是分布于北坡的甘肃臭草增大斑块密度, 提升竞争能力和空间拓殖能力的一种生活史对策, 使根系分叉数和连接长度之间的权衡关系达到极显著水平(p < 0.01, 图2 ). ...
星毛委陵菜根系构型对草原退化的生态适应
3
2011
... 植物的生态策略和适应性、响应性功能要通过植物内部与外部不同功能性状来实现(Bernard- Verdier et al ., 2012 ), 性状的构建、稳固和性状之间的强弱变化与替补转换则靠植物自身的资源权衡和补偿机制来调控(Fortunel et al ., 2012 ; 陈莹婷和许振柱, 2014 ), 是植物在特定环境下生存策略的重要表现(Mooney et al ., 2010 ).根系构型决定了根系在土壤空间中的位置和资源获取方式, 是植物与资源环境异质性相互适应的结果(Malamy, 2005 ; Rogers & Benfey, 2015 ).根系分叉数和连接长度作为根系构型的重要特征指标, 受土壤空间异质性(Villordon et al ., 2014 )、群落环境(单立山等, 2012 )、遗传特性(Kiswara et al ., 2009 ; Bingham & Wu, 2011 )等诸多因素的影响, 在其生存和发展的过程中, 会根据所处生境的资源条件, 调整、转换或补偿自身功能, 实现根系功能性状间的资源权衡.根系分叉数的变化会影响根系的空间分布属性、营养吸收能力和固定作用; 根系连接长度的大小直接决定了根系在土壤中的空间拓展和营养物质吸收能力(Walk et al ., 2004 ), 二者的权衡关系直接反映了植物根系吸收效率和根系构建过程中所需的碳量(Fitter & Stickland, 1991 ).当根系处于水分状态良好的土层中时, 为了提高水分利用率, 会形成更多的侧根, 增大根分叉数(周艳松和王立群, 2011 ); 在干旱胁迫环境中, 较大的分叉数会引起根系内部对营养物质的竞争(杨小林等, 2008 ), 此时植物倾向于增大连接长度, 以减少根系之间的交叠重复, 占据较大的水分和养分资源空间(郭京衡等, 2014 ), 这是植物为适应不同生境条件而形成的获取资源策略, 以达到“生存、生长、繁殖”之三大目的.地形作为生境条件的一种综合指示, 往往能营造局部小气候(Cantón et al ., 2004 ).坡向作为一种重要的地形要素(Fekedulegn et al ., 2003 ), 可以对局部区域的太阳辐射和降水进行二次分配, 其空间变化是群落组成、物种多样性、生态系统过程和植物功能性状的重要决定因素(de Bello et al ., 2006 ; Bennie et al ., 2008 ; Carletti et al ., 2009 ).在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 在各个水平上影响了植物生长及其对水分利用的有效性(Sardans et al ., 2008 ; Huang et al ., 2015 ), 进而引起植物碳资源分配格局和构型构建策略的改变, 为提高生境适应性和风险规避能力, 植物对连接长度和分叉数等主要根系功能性状进行资源优化配置, 构建与生境相适应的根系构型, 优化根系营养吸收和传输系统.因此, 研究坡向梯度上植物根系构型的生态适应策略, 对于理解植物的环境适应机制有重要意义. ...
... 近年来, 多数学者对植物根系的研究主要集中在异质生境中根系功能性状可塑性(Grossman & Rice, 2012 ; Pacheco-Villalobos & Hardtk, 2012 )、不同群落环境下根系构型的适应性调整(Rogers & Benfey, 2015 )、根系生长与土壤环境的关系(Forde, 2014 ; Szoboszlay et al ., 2015 )、根系结构和功能(Hodge et al ., 2009 )等方面, 关于退化草地中植物构件生物量分配模式(周兵等, 2015 )、影响根系构型的主要功能性状(周艳松和王立群, 2011 ), 以及不同入侵阶段植物功能性状表型可塑性(Hulme, 2008 ; Pyšek et al ., 2009 )等方面也进行了系统研究, 而从根系构型构建策略的角度出发, 对生境梯度植物根系功能性状间资源权衡机制的研究仍显不足.甘肃臭草(Melica przewalskyi )是禾本科臭草属多年生根茎植物, 通过根状茎的快速横向生长, 长出幼芽和根系, 形成新植株进行无性繁殖, 在地下建立庞大的根茎系统, 形成密集的圆形株丛, 其分株间的间隔子短, 分枝角度小, 能够在竞争过程中占据优势地位.近年来, 在祁连山北坡中山区荒漠草原与典型草原过渡带, 甘肃臭草替代了原生地带性植被的优势种西北针茅(Stipa sareptana var. krylovii ), 形成大量面积不等、繁茂生长的单一优势种群落斑块(赵成章和龙瑞军, 2008 ), 表现出较高的形态可塑性和对逆境的适应能力, 其强大的入侵能力使众多学者对其资源利用策略和生长扩散过程产生了兴趣.目前, 对甘肃臭草型退化草地优势物种空间格局分析及其对水分的响应(赵成章等, 2011 ; 高福元和赵成章, 2012 )、甘肃臭草斑块土壤水分空间异质性(杨泉等, 2014 ), 以及甘肃臭草茎-叶性状、叶内性状(党晶晶等, 2014 ,2015 )等方面进行了研究, 而生境梯度上甘肃臭草根系构型的可塑性变化尚未见报道.鉴于此, 本文在野外调查的基础上, 试图弄清以下问题: (1)不同坡向甘肃臭草根系分叉数和连接长度的权衡关系如何?(2)导致甘肃臭草根系分叉数和连接长度变化的原因有哪些?旨在探讨祁连山地甘肃臭草种群根系构型构建的资源配置模式, 进一步认识甘肃臭草单一优势种群斑块形成机制和扩散机理. ...
... 植物在环境胁迫下具有权衡性状关系的适应策略(Mooney et al ., 2010 ), 通过生理和发育调节形成与环境相适应的表型特征, 有利于提高植物水分、养分利用效率等生物学功能(李西良等, 2014 ).植物根系构型对环境的适应是通过根系分支结构、空间分布格局等构型特征的改变表现出来的, 是植物长期适应环境的进化结果(Guswa, 2010 ).研究区位于北半球中纬度地区, 山地北坡光照资源最弱(殷秀琴, 2004 )、土壤含水量较高(表2 ), 草地群落的高度、盖度和生物量最高(表2 ), 甘肃臭草处于斑块形成的初期阶段, 其种群密度较小, 也没有高度优势(表1 , 表2 ), 与草地群落的优势种群西北针茅相比, 处于种间竞争的劣势地位.在种间胁迫较大的情况下, 生长在北坡的甘肃臭草采取了增大根系分叉数、减小根系连接长度的根系构型构建策略; 根系分叉数的增加能够增强根系重叠程度、构建密集的根系网络系统, 有利于甘肃臭草地下根系系统和地上株丛的密集生长, 增强种群的密集分布强度, 提高根系对土壤资源的吸收和利用效率, 以斑块式抱团生长实现种群内个体间的高度协同; 减小根系连接长度可能会缩短甘肃臭草克隆分株间的资源传输距离, 有利于增强甘肃臭草克隆分株个体间的相互联系, 保证种群内分株间的资源连接和共享, 从而使种群能够在激烈的资源竞争中存活下来, 与相关学者得出一致的结论, 即“分叉数较大的植物能够迅速占据大量土壤空间快速生长, 适宜生长在土壤资源丰富的环境中” (周艳松和王立群, 2011 ; 郭京衡等, 2014 ).因此, 增大分叉数、减小连接长度是分布于北坡的甘肃臭草增大斑块密度, 提升竞争能力和空间拓殖能力的一种生活史对策, 使根系分叉数和连接长度之间的权衡关系达到极显著水平(p < 0.01, 图2 ). ...
Slope, aspect and climate: Spatially explicit and implicit models of topographic microclimate in chalk grassland.
1
2008
... 植物的生态策略和适应性、响应性功能要通过植物内部与外部不同功能性状来实现(Bernard- Verdier et al ., 2012 ), 性状的构建、稳固和性状之间的强弱变化与替补转换则靠植物自身的资源权衡和补偿机制来调控(Fortunel et al ., 2012 ; 陈莹婷和许振柱, 2014 ), 是植物在特定环境下生存策略的重要表现(Mooney et al ., 2010 ).根系构型决定了根系在土壤空间中的位置和资源获取方式, 是植物与资源环境异质性相互适应的结果(Malamy, 2005 ; Rogers & Benfey, 2015 ).根系分叉数和连接长度作为根系构型的重要特征指标, 受土壤空间异质性(Villordon et al ., 2014 )、群落环境(单立山等, 2012 )、遗传特性(Kiswara et al ., 2009 ; Bingham & Wu, 2011 )等诸多因素的影响, 在其生存和发展的过程中, 会根据所处生境的资源条件, 调整、转换或补偿自身功能, 实现根系功能性状间的资源权衡.根系分叉数的变化会影响根系的空间分布属性、营养吸收能力和固定作用; 根系连接长度的大小直接决定了根系在土壤中的空间拓展和营养物质吸收能力(Walk et al ., 2004 ), 二者的权衡关系直接反映了植物根系吸收效率和根系构建过程中所需的碳量(Fitter & Stickland, 1991 ).当根系处于水分状态良好的土层中时, 为了提高水分利用率, 会形成更多的侧根, 增大根分叉数(周艳松和王立群, 2011 ); 在干旱胁迫环境中, 较大的分叉数会引起根系内部对营养物质的竞争(杨小林等, 2008 ), 此时植物倾向于增大连接长度, 以减少根系之间的交叠重复, 占据较大的水分和养分资源空间(郭京衡等, 2014 ), 这是植物为适应不同生境条件而形成的获取资源策略, 以达到“生存、生长、繁殖”之三大目的.地形作为生境条件的一种综合指示, 往往能营造局部小气候(Cantón et al ., 2004 ).坡向作为一种重要的地形要素(Fekedulegn et al ., 2003 ), 可以对局部区域的太阳辐射和降水进行二次分配, 其空间变化是群落组成、物种多样性、生态系统过程和植物功能性状的重要决定因素(de Bello et al ., 2006 ; Bennie et al ., 2008 ; Carletti et al ., 2009 ).在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 在各个水平上影响了植物生长及其对水分利用的有效性(Sardans et al ., 2008 ; Huang et al ., 2015 ), 进而引起植物碳资源分配格局和构型构建策略的改变, 为提高生境适应性和风险规避能力, 植物对连接长度和分叉数等主要根系功能性状进行资源优化配置, 构建与生境相适应的根系构型, 优化根系营养吸收和传输系统.因此, 研究坡向梯度上植物根系构型的生态适应策略, 对于理解植物的环境适应机制有重要意义. ...
Community assembly along a soil depth gradient: Contrasting patterns of plant trait convergence and divergence in a Mediterranean rangeland.
1
2012
... 植物的生态策略和适应性、响应性功能要通过植物内部与外部不同功能性状来实现(Bernard- Verdier et al ., 2012 ), 性状的构建、稳固和性状之间的强弱变化与替补转换则靠植物自身的资源权衡和补偿机制来调控(Fortunel et al ., 2012 ; 陈莹婷和许振柱, 2014 ), 是植物在特定环境下生存策略的重要表现(Mooney et al ., 2010 ).根系构型决定了根系在土壤空间中的位置和资源获取方式, 是植物与资源环境异质性相互适应的结果(Malamy, 2005 ; Rogers & Benfey, 2015 ).根系分叉数和连接长度作为根系构型的重要特征指标, 受土壤空间异质性(Villordon et al ., 2014 )、群落环境(单立山等, 2012 )、遗传特性(Kiswara et al ., 2009 ; Bingham & Wu, 2011 )等诸多因素的影响, 在其生存和发展的过程中, 会根据所处生境的资源条件, 调整、转换或补偿自身功能, 实现根系功能性状间的资源权衡.根系分叉数的变化会影响根系的空间分布属性、营养吸收能力和固定作用; 根系连接长度的大小直接决定了根系在土壤中的空间拓展和营养物质吸收能力(Walk et al ., 2004 ), 二者的权衡关系直接反映了植物根系吸收效率和根系构建过程中所需的碳量(Fitter & Stickland, 1991 ).当根系处于水分状态良好的土层中时, 为了提高水分利用率, 会形成更多的侧根, 增大根分叉数(周艳松和王立群, 2011 ); 在干旱胁迫环境中, 较大的分叉数会引起根系内部对营养物质的竞争(杨小林等, 2008 ), 此时植物倾向于增大连接长度, 以减少根系之间的交叠重复, 占据较大的水分和养分资源空间(郭京衡等, 2014 ), 这是植物为适应不同生境条件而形成的获取资源策略, 以达到“生存、生长、繁殖”之三大目的.地形作为生境条件的一种综合指示, 往往能营造局部小气候(Cantón et al ., 2004 ).坡向作为一种重要的地形要素(Fekedulegn et al ., 2003 ), 可以对局部区域的太阳辐射和降水进行二次分配, 其空间变化是群落组成、物种多样性、生态系统过程和植物功能性状的重要决定因素(de Bello et al ., 2006 ; Bennie et al ., 2008 ; Carletti et al ., 2009 ).在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 在各个水平上影响了植物生长及其对水分利用的有效性(Sardans et al ., 2008 ; Huang et al ., 2015 ), 进而引起植物碳资源分配格局和构型构建策略的改变, 为提高生境适应性和风险规避能力, 植物对连接长度和分叉数等主要根系功能性状进行资源优化配置, 构建与生境相适应的根系构型, 优化根系营养吸收和传输系统.因此, 研究坡向梯度上植物根系构型的生态适应策略, 对于理解植物的环境适应机制有重要意义. ...
Simulation of wheat growth using the 3D root architecture model SPACSYS: Validation and sensitivity analysis.
1
2011
... 植物的生态策略和适应性、响应性功能要通过植物内部与外部不同功能性状来实现(Bernard- Verdier et al ., 2012 ), 性状的构建、稳固和性状之间的强弱变化与替补转换则靠植物自身的资源权衡和补偿机制来调控(Fortunel et al ., 2012 ; 陈莹婷和许振柱, 2014 ), 是植物在特定环境下生存策略的重要表现(Mooney et al ., 2010 ).根系构型决定了根系在土壤空间中的位置和资源获取方式, 是植物与资源环境异质性相互适应的结果(Malamy, 2005 ; Rogers & Benfey, 2015 ).根系分叉数和连接长度作为根系构型的重要特征指标, 受土壤空间异质性(Villordon et al ., 2014 )、群落环境(单立山等, 2012 )、遗传特性(Kiswara et al ., 2009 ; Bingham & Wu, 2011 )等诸多因素的影响, 在其生存和发展的过程中, 会根据所处生境的资源条件, 调整、转换或补偿自身功能, 实现根系功能性状间的资源权衡.根系分叉数的变化会影响根系的空间分布属性、营养吸收能力和固定作用; 根系连接长度的大小直接决定了根系在土壤中的空间拓展和营养物质吸收能力(Walk et al ., 2004 ), 二者的权衡关系直接反映了植物根系吸收效率和根系构建过程中所需的碳量(Fitter & Stickland, 1991 ).当根系处于水分状态良好的土层中时, 为了提高水分利用率, 会形成更多的侧根, 增大根分叉数(周艳松和王立群, 2011 ); 在干旱胁迫环境中, 较大的分叉数会引起根系内部对营养物质的竞争(杨小林等, 2008 ), 此时植物倾向于增大连接长度, 以减少根系之间的交叠重复, 占据较大的水分和养分资源空间(郭京衡等, 2014 ), 这是植物为适应不同生境条件而形成的获取资源策略, 以达到“生存、生长、繁殖”之三大目的.地形作为生境条件的一种综合指示, 往往能营造局部小气候(Cantón et al ., 2004 ).坡向作为一种重要的地形要素(Fekedulegn et al ., 2003 ), 可以对局部区域的太阳辐射和降水进行二次分配, 其空间变化是群落组成、物种多样性、生态系统过程和植物功能性状的重要决定因素(de Bello et al ., 2006 ; Bennie et al ., 2008 ; Carletti et al ., 2009 ).在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 在各个水平上影响了植物生长及其对水分利用的有效性(Sardans et al ., 2008 ; Huang et al ., 2015 ), 进而引起植物碳资源分配格局和构型构建策略的改变, 为提高生境适应性和风险规避能力, 植物对连接长度和分叉数等主要根系功能性状进行资源优化配置, 构建与生境相适应的根系构型, 优化根系营养吸收和传输系统.因此, 研究坡向梯度上植物根系构型的生态适应策略, 对于理解植物的环境适应机制有重要意义. ...
Topographic controls on the spatial distribution of ground cover in the Tabernas badlands of SE Spain.
1
2004
... 植物的生态策略和适应性、响应性功能要通过植物内部与外部不同功能性状来实现(Bernard- Verdier et al ., 2012 ), 性状的构建、稳固和性状之间的强弱变化与替补转换则靠植物自身的资源权衡和补偿机制来调控(Fortunel et al ., 2012 ; 陈莹婷和许振柱, 2014 ), 是植物在特定环境下生存策略的重要表现(Mooney et al ., 2010 ).根系构型决定了根系在土壤空间中的位置和资源获取方式, 是植物与资源环境异质性相互适应的结果(Malamy, 2005 ; Rogers & Benfey, 2015 ).根系分叉数和连接长度作为根系构型的重要特征指标, 受土壤空间异质性(Villordon et al ., 2014 )、群落环境(单立山等, 2012 )、遗传特性(Kiswara et al ., 2009 ; Bingham & Wu, 2011 )等诸多因素的影响, 在其生存和发展的过程中, 会根据所处生境的资源条件, 调整、转换或补偿自身功能, 实现根系功能性状间的资源权衡.根系分叉数的变化会影响根系的空间分布属性、营养吸收能力和固定作用; 根系连接长度的大小直接决定了根系在土壤中的空间拓展和营养物质吸收能力(Walk et al ., 2004 ), 二者的权衡关系直接反映了植物根系吸收效率和根系构建过程中所需的碳量(Fitter & Stickland, 1991 ).当根系处于水分状态良好的土层中时, 为了提高水分利用率, 会形成更多的侧根, 增大根分叉数(周艳松和王立群, 2011 ); 在干旱胁迫环境中, 较大的分叉数会引起根系内部对营养物质的竞争(杨小林等, 2008 ), 此时植物倾向于增大连接长度, 以减少根系之间的交叠重复, 占据较大的水分和养分资源空间(郭京衡等, 2014 ), 这是植物为适应不同生境条件而形成的获取资源策略, 以达到“生存、生长、繁殖”之三大目的.地形作为生境条件的一种综合指示, 往往能营造局部小气候(Cantón et al ., 2004 ).坡向作为一种重要的地形要素(Fekedulegn et al ., 2003 ), 可以对局部区域的太阳辐射和降水进行二次分配, 其空间变化是群落组成、物种多样性、生态系统过程和植物功能性状的重要决定因素(de Bello et al ., 2006 ; Bennie et al ., 2008 ; Carletti et al ., 2009 ).在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 在各个水平上影响了植物生长及其对水分利用的有效性(Sardans et al ., 2008 ; Huang et al ., 2015 ), 进而引起植物碳资源分配格局和构型构建策略的改变, 为提高生境适应性和风险规避能力, 植物对连接长度和分叉数等主要根系功能性状进行资源优化配置, 构建与生境相适应的根系构型, 优化根系营养吸收和传输系统.因此, 研究坡向梯度上植物根系构型的生态适应策略, 对于理解植物的环境适应机制有重要意义. ...
Soil humic compounds and microbial communities in six spruce forests as function of parent material, slope aspect and stand age.
1
2009
... 植物的生态策略和适应性、响应性功能要通过植物内部与外部不同功能性状来实现(Bernard- Verdier et al ., 2012 ), 性状的构建、稳固和性状之间的强弱变化与替补转换则靠植物自身的资源权衡和补偿机制来调控(Fortunel et al ., 2012 ; 陈莹婷和许振柱, 2014 ), 是植物在特定环境下生存策略的重要表现(Mooney et al ., 2010 ).根系构型决定了根系在土壤空间中的位置和资源获取方式, 是植物与资源环境异质性相互适应的结果(Malamy, 2005 ; Rogers & Benfey, 2015 ).根系分叉数和连接长度作为根系构型的重要特征指标, 受土壤空间异质性(Villordon et al ., 2014 )、群落环境(单立山等, 2012 )、遗传特性(Kiswara et al ., 2009 ; Bingham & Wu, 2011 )等诸多因素的影响, 在其生存和发展的过程中, 会根据所处生境的资源条件, 调整、转换或补偿自身功能, 实现根系功能性状间的资源权衡.根系分叉数的变化会影响根系的空间分布属性、营养吸收能力和固定作用; 根系连接长度的大小直接决定了根系在土壤中的空间拓展和营养物质吸收能力(Walk et al ., 2004 ), 二者的权衡关系直接反映了植物根系吸收效率和根系构建过程中所需的碳量(Fitter & Stickland, 1991 ).当根系处于水分状态良好的土层中时, 为了提高水分利用率, 会形成更多的侧根, 增大根分叉数(周艳松和王立群, 2011 ); 在干旱胁迫环境中, 较大的分叉数会引起根系内部对营养物质的竞争(杨小林等, 2008 ), 此时植物倾向于增大连接长度, 以减少根系之间的交叠重复, 占据较大的水分和养分资源空间(郭京衡等, 2014 ), 这是植物为适应不同生境条件而形成的获取资源策略, 以达到“生存、生长、繁殖”之三大目的.地形作为生境条件的一种综合指示, 往往能营造局部小气候(Cantón et al ., 2004 ).坡向作为一种重要的地形要素(Fekedulegn et al ., 2003 ), 可以对局部区域的太阳辐射和降水进行二次分配, 其空间变化是群落组成、物种多样性、生态系统过程和植物功能性状的重要决定因素(de Bello et al ., 2006 ; Bennie et al ., 2008 ; Carletti et al ., 2009 ).在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 在各个水平上影响了植物生长及其对水分利用的有效性(Sardans et al ., 2008 ; Huang et al ., 2015 ), 进而引起植物碳资源分配格局和构型构建策略的改变, 为提高生境适应性和风险规避能力, 植物对连接长度和分叉数等主要根系功能性状进行资源优化配置, 构建与生境相适应的根系构型, 优化根系营养吸收和传输系统.因此, 研究坡向梯度上植物根系构型的生态适应策略, 对于理解植物的环境适应机制有重要意义. ...
四川龙门山西北部植被分布与地形因子的相关性
1
2006
... 利用ArcGIS对研究区1:5万地形图进行数字化, 得到研究区域10 m分辨率的数字高程模型(digital elevation model, DEM).在DEM基础上利用Spatial Analyst模块中的Surface Analysis命令提取坡向, 它表示地表面上一点的切平面的法线矢量n 在水平面的投影nxoy 与过该点的正北方向的夹角, 其中坡向组别以国际划分法确定(陈瑶等, 2006 ), 具体方法参见汤国安等(2010) 文献. ...
四川龙门山西北部植被分布与地形因子的相关性
1
2006
... 利用ArcGIS对研究区1:5万地形图进行数字化, 得到研究区域10 m分辨率的数字高程模型(digital elevation model, DEM).在DEM基础上利用Spatial Analyst模块中的Surface Analysis命令提取坡向, 它表示地表面上一点的切平面的法线矢量n 在水平面的投影nxoy 与过该点的正北方向的夹角, 其中坡向组别以国际划分法确定(陈瑶等, 2006 ), 具体方法参见汤国安等(2010) 文献. ...
植物叶经济谱的研究进展
1
2014
... 植物的生态策略和适应性、响应性功能要通过植物内部与外部不同功能性状来实现(Bernard- Verdier et al ., 2012 ), 性状的构建、稳固和性状之间的强弱变化与替补转换则靠植物自身的资源权衡和补偿机制来调控(Fortunel et al ., 2012 ; 陈莹婷和许振柱, 2014 ), 是植物在特定环境下生存策略的重要表现(Mooney et al ., 2010 ).根系构型决定了根系在土壤空间中的位置和资源获取方式, 是植物与资源环境异质性相互适应的结果(Malamy, 2005 ; Rogers & Benfey, 2015 ).根系分叉数和连接长度作为根系构型的重要特征指标, 受土壤空间异质性(Villordon et al ., 2014 )、群落环境(单立山等, 2012 )、遗传特性(Kiswara et al ., 2009 ; Bingham & Wu, 2011 )等诸多因素的影响, 在其生存和发展的过程中, 会根据所处生境的资源条件, 调整、转换或补偿自身功能, 实现根系功能性状间的资源权衡.根系分叉数的变化会影响根系的空间分布属性、营养吸收能力和固定作用; 根系连接长度的大小直接决定了根系在土壤中的空间拓展和营养物质吸收能力(Walk et al ., 2004 ), 二者的权衡关系直接反映了植物根系吸收效率和根系构建过程中所需的碳量(Fitter & Stickland, 1991 ).当根系处于水分状态良好的土层中时, 为了提高水分利用率, 会形成更多的侧根, 增大根分叉数(周艳松和王立群, 2011 ); 在干旱胁迫环境中, 较大的分叉数会引起根系内部对营养物质的竞争(杨小林等, 2008 ), 此时植物倾向于增大连接长度, 以减少根系之间的交叠重复, 占据较大的水分和养分资源空间(郭京衡等, 2014 ), 这是植物为适应不同生境条件而形成的获取资源策略, 以达到“生存、生长、繁殖”之三大目的.地形作为生境条件的一种综合指示, 往往能营造局部小气候(Cantón et al ., 2004 ).坡向作为一种重要的地形要素(Fekedulegn et al ., 2003 ), 可以对局部区域的太阳辐射和降水进行二次分配, 其空间变化是群落组成、物种多样性、生态系统过程和植物功能性状的重要决定因素(de Bello et al ., 2006 ; Bennie et al ., 2008 ; Carletti et al ., 2009 ).在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 在各个水平上影响了植物生长及其对水分利用的有效性(Sardans et al ., 2008 ; Huang et al ., 2015 ), 进而引起植物碳资源分配格局和构型构建策略的改变, 为提高生境适应性和风险规避能力, 植物对连接长度和分叉数等主要根系功能性状进行资源优化配置, 构建与生境相适应的根系构型, 优化根系营养吸收和传输系统.因此, 研究坡向梯度上植物根系构型的生态适应策略, 对于理解植物的环境适应机制有重要意义. ...
植物叶经济谱的研究进展
1
2014
... 植物的生态策略和适应性、响应性功能要通过植物内部与外部不同功能性状来实现(Bernard- Verdier et al ., 2012 ), 性状的构建、稳固和性状之间的强弱变化与替补转换则靠植物自身的资源权衡和补偿机制来调控(Fortunel et al ., 2012 ; 陈莹婷和许振柱, 2014 ), 是植物在特定环境下生存策略的重要表现(Mooney et al ., 2010 ).根系构型决定了根系在土壤空间中的位置和资源获取方式, 是植物与资源环境异质性相互适应的结果(Malamy, 2005 ; Rogers & Benfey, 2015 ).根系分叉数和连接长度作为根系构型的重要特征指标, 受土壤空间异质性(Villordon et al ., 2014 )、群落环境(单立山等, 2012 )、遗传特性(Kiswara et al ., 2009 ; Bingham & Wu, 2011 )等诸多因素的影响, 在其生存和发展的过程中, 会根据所处生境的资源条件, 调整、转换或补偿自身功能, 实现根系功能性状间的资源权衡.根系分叉数的变化会影响根系的空间分布属性、营养吸收能力和固定作用; 根系连接长度的大小直接决定了根系在土壤中的空间拓展和营养物质吸收能力(Walk et al ., 2004 ), 二者的权衡关系直接反映了植物根系吸收效率和根系构建过程中所需的碳量(Fitter & Stickland, 1991 ).当根系处于水分状态良好的土层中时, 为了提高水分利用率, 会形成更多的侧根, 增大根分叉数(周艳松和王立群, 2011 ); 在干旱胁迫环境中, 较大的分叉数会引起根系内部对营养物质的竞争(杨小林等, 2008 ), 此时植物倾向于增大连接长度, 以减少根系之间的交叠重复, 占据较大的水分和养分资源空间(郭京衡等, 2014 ), 这是植物为适应不同生境条件而形成的获取资源策略, 以达到“生存、生长、繁殖”之三大目的.地形作为生境条件的一种综合指示, 往往能营造局部小气候(Cantón et al ., 2004 ).坡向作为一种重要的地形要素(Fekedulegn et al ., 2003 ), 可以对局部区域的太阳辐射和降水进行二次分配, 其空间变化是群落组成、物种多样性、生态系统过程和植物功能性状的重要决定因素(de Bello et al ., 2006 ; Bennie et al ., 2008 ; Carletti et al ., 2009 ).在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 在各个水平上影响了植物生长及其对水分利用的有效性(Sardans et al ., 2008 ; Huang et al ., 2015 ), 进而引起植物碳资源分配格局和构型构建策略的改变, 为提高生境适应性和风险规避能力, 植物对连接长度和分叉数等主要根系功能性状进行资源优化配置, 构建与生境相适应的根系构型, 优化根系营养吸收和传输系统.因此, 研究坡向梯度上植物根系构型的生态适应策略, 对于理解植物的环境适应机制有重要意义. ...
高寒草地甘肃臭草茎——叶性状的坡度差异性
1
2014
... 近年来, 多数学者对植物根系的研究主要集中在异质生境中根系功能性状可塑性(Grossman & Rice, 2012 ; Pacheco-Villalobos & Hardtk, 2012 )、不同群落环境下根系构型的适应性调整(Rogers & Benfey, 2015 )、根系生长与土壤环境的关系(Forde, 2014 ; Szoboszlay et al ., 2015 )、根系结构和功能(Hodge et al ., 2009 )等方面, 关于退化草地中植物构件生物量分配模式(周兵等, 2015 )、影响根系构型的主要功能性状(周艳松和王立群, 2011 ), 以及不同入侵阶段植物功能性状表型可塑性(Hulme, 2008 ; Pyšek et al ., 2009 )等方面也进行了系统研究, 而从根系构型构建策略的角度出发, 对生境梯度植物根系功能性状间资源权衡机制的研究仍显不足.甘肃臭草(Melica przewalskyi )是禾本科臭草属多年生根茎植物, 通过根状茎的快速横向生长, 长出幼芽和根系, 形成新植株进行无性繁殖, 在地下建立庞大的根茎系统, 形成密集的圆形株丛, 其分株间的间隔子短, 分枝角度小, 能够在竞争过程中占据优势地位.近年来, 在祁连山北坡中山区荒漠草原与典型草原过渡带, 甘肃臭草替代了原生地带性植被的优势种西北针茅(Stipa sareptana var. krylovii ), 形成大量面积不等、繁茂生长的单一优势种群落斑块(赵成章和龙瑞军, 2008 ), 表现出较高的形态可塑性和对逆境的适应能力, 其强大的入侵能力使众多学者对其资源利用策略和生长扩散过程产生了兴趣.目前, 对甘肃臭草型退化草地优势物种空间格局分析及其对水分的响应(赵成章等, 2011 ; 高福元和赵成章, 2012 )、甘肃臭草斑块土壤水分空间异质性(杨泉等, 2014 ), 以及甘肃臭草茎-叶性状、叶内性状(党晶晶等, 2014 ,2015 )等方面进行了研究, 而生境梯度上甘肃臭草根系构型的可塑性变化尚未见报道.鉴于此, 本文在野外调查的基础上, 试图弄清以下问题: (1)不同坡向甘肃臭草根系分叉数和连接长度的权衡关系如何?(2)导致甘肃臭草根系分叉数和连接长度变化的原因有哪些?旨在探讨祁连山地甘肃臭草种群根系构型构建的资源配置模式, 进一步认识甘肃臭草单一优势种群斑块形成机制和扩散机理. ...
高寒草地甘肃臭草茎——叶性状的坡度差异性
1
2014
... 近年来, 多数学者对植物根系的研究主要集中在异质生境中根系功能性状可塑性(Grossman & Rice, 2012 ; Pacheco-Villalobos & Hardtk, 2012 )、不同群落环境下根系构型的适应性调整(Rogers & Benfey, 2015 )、根系生长与土壤环境的关系(Forde, 2014 ; Szoboszlay et al ., 2015 )、根系结构和功能(Hodge et al ., 2009 )等方面, 关于退化草地中植物构件生物量分配模式(周兵等, 2015 )、影响根系构型的主要功能性状(周艳松和王立群, 2011 ), 以及不同入侵阶段植物功能性状表型可塑性(Hulme, 2008 ; Pyšek et al ., 2009 )等方面也进行了系统研究, 而从根系构型构建策略的角度出发, 对生境梯度植物根系功能性状间资源权衡机制的研究仍显不足.甘肃臭草(Melica przewalskyi )是禾本科臭草属多年生根茎植物, 通过根状茎的快速横向生长, 长出幼芽和根系, 形成新植株进行无性繁殖, 在地下建立庞大的根茎系统, 形成密集的圆形株丛, 其分株间的间隔子短, 分枝角度小, 能够在竞争过程中占据优势地位.近年来, 在祁连山北坡中山区荒漠草原与典型草原过渡带, 甘肃臭草替代了原生地带性植被的优势种西北针茅(Stipa sareptana var. krylovii ), 形成大量面积不等、繁茂生长的单一优势种群落斑块(赵成章和龙瑞军, 2008 ), 表现出较高的形态可塑性和对逆境的适应能力, 其强大的入侵能力使众多学者对其资源利用策略和生长扩散过程产生了兴趣.目前, 对甘肃臭草型退化草地优势物种空间格局分析及其对水分的响应(赵成章等, 2011 ; 高福元和赵成章, 2012 )、甘肃臭草斑块土壤水分空间异质性(杨泉等, 2014 ), 以及甘肃臭草茎-叶性状、叶内性状(党晶晶等, 2014 ,2015 )等方面进行了研究, 而生境梯度上甘肃臭草根系构型的可塑性变化尚未见报道.鉴于此, 本文在野外调查的基础上, 试图弄清以下问题: (1)不同坡向甘肃臭草根系分叉数和连接长度的权衡关系如何?(2)导致甘肃臭草根系分叉数和连接长度变化的原因有哪些?旨在探讨祁连山地甘肃臭草种群根系构型构建的资源配置模式, 进一步认识甘肃臭草单一优势种群斑块形成机制和扩散机理. ...
祁连山高寒草地甘肃臭草叶性状与坡向间的关系
1
2015
... 近年来, 多数学者对植物根系的研究主要集中在异质生境中根系功能性状可塑性(Grossman & Rice, 2012 ; Pacheco-Villalobos & Hardtk, 2012 )、不同群落环境下根系构型的适应性调整(Rogers & Benfey, 2015 )、根系生长与土壤环境的关系(Forde, 2014 ; Szoboszlay et al ., 2015 )、根系结构和功能(Hodge et al ., 2009 )等方面, 关于退化草地中植物构件生物量分配模式(周兵等, 2015 )、影响根系构型的主要功能性状(周艳松和王立群, 2011 ), 以及不同入侵阶段植物功能性状表型可塑性(Hulme, 2008 ; Pyšek et al ., 2009 )等方面也进行了系统研究, 而从根系构型构建策略的角度出发, 对生境梯度植物根系功能性状间资源权衡机制的研究仍显不足.甘肃臭草(Melica przewalskyi )是禾本科臭草属多年生根茎植物, 通过根状茎的快速横向生长, 长出幼芽和根系, 形成新植株进行无性繁殖, 在地下建立庞大的根茎系统, 形成密集的圆形株丛, 其分株间的间隔子短, 分枝角度小, 能够在竞争过程中占据优势地位.近年来, 在祁连山北坡中山区荒漠草原与典型草原过渡带, 甘肃臭草替代了原生地带性植被的优势种西北针茅(Stipa sareptana var. krylovii ), 形成大量面积不等、繁茂生长的单一优势种群落斑块(赵成章和龙瑞军, 2008 ), 表现出较高的形态可塑性和对逆境的适应能力, 其强大的入侵能力使众多学者对其资源利用策略和生长扩散过程产生了兴趣.目前, 对甘肃臭草型退化草地优势物种空间格局分析及其对水分的响应(赵成章等, 2011 ; 高福元和赵成章, 2012 )、甘肃臭草斑块土壤水分空间异质性(杨泉等, 2014 ), 以及甘肃臭草茎-叶性状、叶内性状(党晶晶等, 2014 ,2015 )等方面进行了研究, 而生境梯度上甘肃臭草根系构型的可塑性变化尚未见报道.鉴于此, 本文在野外调查的基础上, 试图弄清以下问题: (1)不同坡向甘肃臭草根系分叉数和连接长度的权衡关系如何?(2)导致甘肃臭草根系分叉数和连接长度变化的原因有哪些?旨在探讨祁连山地甘肃臭草种群根系构型构建的资源配置模式, 进一步认识甘肃臭草单一优势种群斑块形成机制和扩散机理. ...
祁连山高寒草地甘肃臭草叶性状与坡向间的关系
1
2015
... 近年来, 多数学者对植物根系的研究主要集中在异质生境中根系功能性状可塑性(Grossman & Rice, 2012 ; Pacheco-Villalobos & Hardtk, 2012 )、不同群落环境下根系构型的适应性调整(Rogers & Benfey, 2015 )、根系生长与土壤环境的关系(Forde, 2014 ; Szoboszlay et al ., 2015 )、根系结构和功能(Hodge et al ., 2009 )等方面, 关于退化草地中植物构件生物量分配模式(周兵等, 2015 )、影响根系构型的主要功能性状(周艳松和王立群, 2011 ), 以及不同入侵阶段植物功能性状表型可塑性(Hulme, 2008 ; Pyšek et al ., 2009 )等方面也进行了系统研究, 而从根系构型构建策略的角度出发, 对生境梯度植物根系功能性状间资源权衡机制的研究仍显不足.甘肃臭草(Melica przewalskyi )是禾本科臭草属多年生根茎植物, 通过根状茎的快速横向生长, 长出幼芽和根系, 形成新植株进行无性繁殖, 在地下建立庞大的根茎系统, 形成密集的圆形株丛, 其分株间的间隔子短, 分枝角度小, 能够在竞争过程中占据优势地位.近年来, 在祁连山北坡中山区荒漠草原与典型草原过渡带, 甘肃臭草替代了原生地带性植被的优势种西北针茅(Stipa sareptana var. krylovii ), 形成大量面积不等、繁茂生长的单一优势种群落斑块(赵成章和龙瑞军, 2008 ), 表现出较高的形态可塑性和对逆境的适应能力, 其强大的入侵能力使众多学者对其资源利用策略和生长扩散过程产生了兴趣.目前, 对甘肃臭草型退化草地优势物种空间格局分析及其对水分的响应(赵成章等, 2011 ; 高福元和赵成章, 2012 )、甘肃臭草斑块土壤水分空间异质性(杨泉等, 2014 ), 以及甘肃臭草茎-叶性状、叶内性状(党晶晶等, 2014 ,2015 )等方面进行了研究, 而生境梯度上甘肃臭草根系构型的可塑性变化尚未见报道.鉴于此, 本文在野外调查的基础上, 试图弄清以下问题: (1)不同坡向甘肃臭草根系分叉数和连接长度的权衡关系如何?(2)导致甘肃臭草根系分叉数和连接长度变化的原因有哪些?旨在探讨祁连山地甘肃臭草种群根系构型构建的资源配置模式, 进一步认识甘肃臭草单一优势种群斑块形成机制和扩散机理. ...
Fractal geometry and root system structures of heterogeneous plant communities.
1
2005
... 植物在不同功能和性状之间对有限资源的分配存在着互相牵制的权衡作用(Shipley et al ., 2006 ), 这就需要植物在生长发育过程中, 通过不断优化自身的资源分配来提高适合度以适应环境的变化(Fabbro & Körner, 2004).根系作为植物地上部分与土壤的连接器官, 在较大程度上影响着植物的水分和养分吸收、运移和存储功能(Dannowski & Block, 2005 ; Kong et al ., 2014 ), 根系分叉数和连接长度能够反映根系不同部位构建过程中的碳消耗特点, 影响根系拓展空间、获取营养的代价与效率, 体现了植物性状和构型构建对生境变化的响应, 从中可以理解和阐释植物的物种分布、种间关系、权衡及其作用机制(Westoby & Wright, 2006 ).本研究发现, 随着坡向由北坡向东坡、西坡、南坡转变, 甘肃臭草根系分叉数呈逐渐减小的趋势, 而根系连接长度呈相反的变化趋势(图1 ), 即甘肃臭草分配给根系分叉数和连接长度的资源间存在着“此消彼长”的权衡关系(图2 ).经Pearson相关性分析得出, 群落密度与根系分叉数、连接长度呈极显著相关关系(p < 0.01), 群落高度、土壤含水量与二者显著相关(p < 0.05), 地上生物量与根系分叉数显著相关(p < 0.05, 表3 ), 表明坡向梯度上甘肃臭草根系分叉数和连接长度权衡关系的差异性, 是植物根系构建的资源配置模式对高寒草地生境异质性适应的结果. ...
Variations in species and functional plant diversity along climatic and grazing gradients.
1
2006
... 植物的生态策略和适应性、响应性功能要通过植物内部与外部不同功能性状来实现(Bernard- Verdier et al ., 2012 ), 性状的构建、稳固和性状之间的强弱变化与替补转换则靠植物自身的资源权衡和补偿机制来调控(Fortunel et al ., 2012 ; 陈莹婷和许振柱, 2014 ), 是植物在特定环境下生存策略的重要表现(Mooney et al ., 2010 ).根系构型决定了根系在土壤空间中的位置和资源获取方式, 是植物与资源环境异质性相互适应的结果(Malamy, 2005 ; Rogers & Benfey, 2015 ).根系分叉数和连接长度作为根系构型的重要特征指标, 受土壤空间异质性(Villordon et al ., 2014 )、群落环境(单立山等, 2012 )、遗传特性(Kiswara et al ., 2009 ; Bingham & Wu, 2011 )等诸多因素的影响, 在其生存和发展的过程中, 会根据所处生境的资源条件, 调整、转换或补偿自身功能, 实现根系功能性状间的资源权衡.根系分叉数的变化会影响根系的空间分布属性、营养吸收能力和固定作用; 根系连接长度的大小直接决定了根系在土壤中的空间拓展和营养物质吸收能力(Walk et al ., 2004 ), 二者的权衡关系直接反映了植物根系吸收效率和根系构建过程中所需的碳量(Fitter & Stickland, 1991 ).当根系处于水分状态良好的土层中时, 为了提高水分利用率, 会形成更多的侧根, 增大根分叉数(周艳松和王立群, 2011 ); 在干旱胁迫环境中, 较大的分叉数会引起根系内部对营养物质的竞争(杨小林等, 2008 ), 此时植物倾向于增大连接长度, 以减少根系之间的交叠重复, 占据较大的水分和养分资源空间(郭京衡等, 2014 ), 这是植物为适应不同生境条件而形成的获取资源策略, 以达到“生存、生长、繁殖”之三大目的.地形作为生境条件的一种综合指示, 往往能营造局部小气候(Cantón et al ., 2004 ).坡向作为一种重要的地形要素(Fekedulegn et al ., 2003 ), 可以对局部区域的太阳辐射和降水进行二次分配, 其空间变化是群落组成、物种多样性、生态系统过程和植物功能性状的重要决定因素(de Bello et al ., 2006 ; Bennie et al ., 2008 ; Carletti et al ., 2009 ).在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 在各个水平上影响了植物生长及其对水分利用的有效性(Sardans et al ., 2008 ; Huang et al ., 2015 ), 进而引起植物碳资源分配格局和构型构建策略的改变, 为提高生境适应性和风险规避能力, 植物对连接长度和分叉数等主要根系功能性状进行资源优化配置, 构建与生境相适应的根系构型, 优化根系营养吸收和传输系统.因此, 研究坡向梯度上植物根系构型的生态适应策略, 对于理解植物的环境适应机制有重要意义. ...
华南海岸典型沙生植物根系构型特征
1
2014
... 植物根系构型构建策略的差异可以用根系碳投入与土壤水分养分收益的权衡关系来解释(Oppelt et al ., 2001 ).根系分叉数的增加会导致根系重叠加剧, 增加根系内部的资源竞争态势(杨小林等, 2008 ).根系连接长度的变化会影响根系空间拓展能力和利用土壤资源的有效性(Schenk & Jackson, 2002a , 2002b ), 这与植物自身根系碳投入与土壤水分养分的收益权衡相契合, 是提高植物资源获取能力的一个重要策略(杨小林等, 2008 ).相对于北坡, 山地南坡日照时间长、水分蒸发量大(殷秀琴, 2004 ), 生长在南坡的甘肃臭草多呈直径不等的单一优势种群斑块, 占据了草地群落的绝对资源优势(表1 , 表2 ).甘肃臭草采用克隆繁殖模式形成了庞大的互相连通交织的根茎系统, 构建了高度聚集的根系网络(高福元和赵成章, 2012 ), 斑块内部种群聚集程度较高, 斑块中心土壤含水量明显低于斑块边缘的土壤含水量(赵成章等, 2011 ), 体现了甘肃臭草向斑块周围扩张的迫切需求.在地上和地下生存空间拥挤、土壤含水量分布不均衡状态下, 为了避免不必要的种内竞争, 甘肃臭草限制了地下生物量的分配比例, 以减缓营养繁殖速度, 降低种群聚集强度, 此时甘肃臭草选择了增加根系连接长度、减少分叉数的根系构型构建模式.根系分叉数的减小能够降低种群内部根系重叠程度, 避免不必要的种内竞争; 连接长度的增大可以增强根系空间拓展能力, 扩大单位根系所占的土壤体空间, 使根系从斑块中心向土壤水分较好、生长发育空间较大的边缘区域拓殖, 是南坡甘肃臭草单优种群斑块扩张的主要动力, 与赵成章等(2011) 关于甘肃臭草通过营养繁殖以类似“同心圆”的方式不断由斑块中心向边缘扩大种群领地的观点一致.这符合相关学者“当植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 增加根系连接长度等策略, 以寻求更多的水分和养分, 保证自身的生存和生长”的结论(单立山等, 2013 ; 杜建会等, 2014 ).因此, 南坡甘肃臭草根系分叉数和连接长度间呈极显著负相关关系(p < 0.01, 图2 ), 减小分叉数、增加连接长度是甘肃臭草斑块扩张的根系构型构建策略, 降低了甘肃臭草单一优势种群斑块内部的根系拥挤程度. ...
华南海岸典型沙生植物根系构型特征
1
2014
... 植物根系构型构建策略的差异可以用根系碳投入与土壤水分养分收益的权衡关系来解释(Oppelt et al ., 2001 ).根系分叉数的增加会导致根系重叠加剧, 增加根系内部的资源竞争态势(杨小林等, 2008 ).根系连接长度的变化会影响根系空间拓展能力和利用土壤资源的有效性(Schenk & Jackson, 2002a , 2002b ), 这与植物自身根系碳投入与土壤水分养分的收益权衡相契合, 是提高植物资源获取能力的一个重要策略(杨小林等, 2008 ).相对于北坡, 山地南坡日照时间长、水分蒸发量大(殷秀琴, 2004 ), 生长在南坡的甘肃臭草多呈直径不等的单一优势种群斑块, 占据了草地群落的绝对资源优势(表1 , 表2 ).甘肃臭草采用克隆繁殖模式形成了庞大的互相连通交织的根茎系统, 构建了高度聚集的根系网络(高福元和赵成章, 2012 ), 斑块内部种群聚集程度较高, 斑块中心土壤含水量明显低于斑块边缘的土壤含水量(赵成章等, 2011 ), 体现了甘肃臭草向斑块周围扩张的迫切需求.在地上和地下生存空间拥挤、土壤含水量分布不均衡状态下, 为了避免不必要的种内竞争, 甘肃臭草限制了地下生物量的分配比例, 以减缓营养繁殖速度, 降低种群聚集强度, 此时甘肃臭草选择了增加根系连接长度、减少分叉数的根系构型构建模式.根系分叉数的减小能够降低种群内部根系重叠程度, 避免不必要的种内竞争; 连接长度的增大可以增强根系空间拓展能力, 扩大单位根系所占的土壤体空间, 使根系从斑块中心向土壤水分较好、生长发育空间较大的边缘区域拓殖, 是南坡甘肃臭草单优种群斑块扩张的主要动力, 与赵成章等(2011) 关于甘肃臭草通过营养繁殖以类似“同心圆”的方式不断由斑块中心向边缘扩大种群领地的观点一致.这符合相关学者“当植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 增加根系连接长度等策略, 以寻求更多的水分和养分, 保证自身的生存和生长”的结论(单立山等, 2013 ; 杜建会等, 2014 ).因此, 南坡甘肃臭草根系分叉数和连接长度间呈极显著负相关关系(p < 0.01, 图2 ), 减小分叉数、增加连接长度是甘肃臭草斑块扩张的根系构型构建策略, 降低了甘肃臭草单一优势种群斑块内部的根系拥挤程度. ...
Altitudinal differences in flower traits and reproductive allocation.
0
2004
Influence of topographic aspect, precipitation and drought on radial growth of four major tree species in an Appalachian watershed.
1
2003
... 植物的生态策略和适应性、响应性功能要通过植物内部与外部不同功能性状来实现(Bernard- Verdier et al ., 2012 ), 性状的构建、稳固和性状之间的强弱变化与替补转换则靠植物自身的资源权衡和补偿机制来调控(Fortunel et al ., 2012 ; 陈莹婷和许振柱, 2014 ), 是植物在特定环境下生存策略的重要表现(Mooney et al ., 2010 ).根系构型决定了根系在土壤空间中的位置和资源获取方式, 是植物与资源环境异质性相互适应的结果(Malamy, 2005 ; Rogers & Benfey, 2015 ).根系分叉数和连接长度作为根系构型的重要特征指标, 受土壤空间异质性(Villordon et al ., 2014 )、群落环境(单立山等, 2012 )、遗传特性(Kiswara et al ., 2009 ; Bingham & Wu, 2011 )等诸多因素的影响, 在其生存和发展的过程中, 会根据所处生境的资源条件, 调整、转换或补偿自身功能, 实现根系功能性状间的资源权衡.根系分叉数的变化会影响根系的空间分布属性、营养吸收能力和固定作用; 根系连接长度的大小直接决定了根系在土壤中的空间拓展和营养物质吸收能力(Walk et al ., 2004 ), 二者的权衡关系直接反映了植物根系吸收效率和根系构建过程中所需的碳量(Fitter & Stickland, 1991 ).当根系处于水分状态良好的土层中时, 为了提高水分利用率, 会形成更多的侧根, 增大根分叉数(周艳松和王立群, 2011 ); 在干旱胁迫环境中, 较大的分叉数会引起根系内部对营养物质的竞争(杨小林等, 2008 ), 此时植物倾向于增大连接长度, 以减少根系之间的交叠重复, 占据较大的水分和养分资源空间(郭京衡等, 2014 ), 这是植物为适应不同生境条件而形成的获取资源策略, 以达到“生存、生长、繁殖”之三大目的.地形作为生境条件的一种综合指示, 往往能营造局部小气候(Cantón et al ., 2004 ).坡向作为一种重要的地形要素(Fekedulegn et al ., 2003 ), 可以对局部区域的太阳辐射和降水进行二次分配, 其空间变化是群落组成、物种多样性、生态系统过程和植物功能性状的重要决定因素(de Bello et al ., 2006 ; Bennie et al ., 2008 ; Carletti et al ., 2009 ).在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 在各个水平上影响了植物生长及其对水分利用的有效性(Sardans et al ., 2008 ; Huang et al ., 2015 ), 进而引起植物碳资源分配格局和构型构建策略的改变, 为提高生境适应性和风险规避能力, 植物对连接长度和分叉数等主要根系功能性状进行资源优化配置, 构建与生境相适应的根系构型, 优化根系营养吸收和传输系统.因此, 研究坡向梯度上植物根系构型的生态适应策略, 对于理解植物的环境适应机制有重要意义. ...
Architectural analysis of plant root systems 2. Influence of nutrient supply on architecture in contrasting plant species.
1
1991
... 植物的生态策略和适应性、响应性功能要通过植物内部与外部不同功能性状来实现(Bernard- Verdier et al ., 2012 ), 性状的构建、稳固和性状之间的强弱变化与替补转换则靠植物自身的资源权衡和补偿机制来调控(Fortunel et al ., 2012 ; 陈莹婷和许振柱, 2014 ), 是植物在特定环境下生存策略的重要表现(Mooney et al ., 2010 ).根系构型决定了根系在土壤空间中的位置和资源获取方式, 是植物与资源环境异质性相互适应的结果(Malamy, 2005 ; Rogers & Benfey, 2015 ).根系分叉数和连接长度作为根系构型的重要特征指标, 受土壤空间异质性(Villordon et al ., 2014 )、群落环境(单立山等, 2012 )、遗传特性(Kiswara et al ., 2009 ; Bingham & Wu, 2011 )等诸多因素的影响, 在其生存和发展的过程中, 会根据所处生境的资源条件, 调整、转换或补偿自身功能, 实现根系功能性状间的资源权衡.根系分叉数的变化会影响根系的空间分布属性、营养吸收能力和固定作用; 根系连接长度的大小直接决定了根系在土壤中的空间拓展和营养物质吸收能力(Walk et al ., 2004 ), 二者的权衡关系直接反映了植物根系吸收效率和根系构建过程中所需的碳量(Fitter & Stickland, 1991 ).当根系处于水分状态良好的土层中时, 为了提高水分利用率, 会形成更多的侧根, 增大根分叉数(周艳松和王立群, 2011 ); 在干旱胁迫环境中, 较大的分叉数会引起根系内部对营养物质的竞争(杨小林等, 2008 ), 此时植物倾向于增大连接长度, 以减少根系之间的交叠重复, 占据较大的水分和养分资源空间(郭京衡等, 2014 ), 这是植物为适应不同生境条件而形成的获取资源策略, 以达到“生存、生长、繁殖”之三大目的.地形作为生境条件的一种综合指示, 往往能营造局部小气候(Cantón et al ., 2004 ).坡向作为一种重要的地形要素(Fekedulegn et al ., 2003 ), 可以对局部区域的太阳辐射和降水进行二次分配, 其空间变化是群落组成、物种多样性、生态系统过程和植物功能性状的重要决定因素(de Bello et al ., 2006 ; Bennie et al ., 2008 ; Carletti et al ., 2009 ).在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 在各个水平上影响了植物生长及其对水分利用的有效性(Sardans et al ., 2008 ; Huang et al ., 2015 ), 进而引起植物碳资源分配格局和构型构建策略的改变, 为提高生境适应性和风险规避能力, 植物对连接长度和分叉数等主要根系功能性状进行资源优化配置, 构建与生境相适应的根系构型, 优化根系营养吸收和传输系统.因此, 研究坡向梯度上植物根系构型的生态适应策略, 对于理解植物的环境适应机制有重要意义. ...
Nitrogen signalling pathways shaping root system architecture: An update.
1
2014
... 近年来, 多数学者对植物根系的研究主要集中在异质生境中根系功能性状可塑性(Grossman & Rice, 2012 ; Pacheco-Villalobos & Hardtk, 2012 )、不同群落环境下根系构型的适应性调整(Rogers & Benfey, 2015 )、根系生长与土壤环境的关系(Forde, 2014 ; Szoboszlay et al ., 2015 )、根系结构和功能(Hodge et al ., 2009 )等方面, 关于退化草地中植物构件生物量分配模式(周兵等, 2015 )、影响根系构型的主要功能性状(周艳松和王立群, 2011 ), 以及不同入侵阶段植物功能性状表型可塑性(Hulme, 2008 ; Pyšek et al ., 2009 )等方面也进行了系统研究, 而从根系构型构建策略的角度出发, 对生境梯度植物根系功能性状间资源权衡机制的研究仍显不足.甘肃臭草(Melica przewalskyi )是禾本科臭草属多年生根茎植物, 通过根状茎的快速横向生长, 长出幼芽和根系, 形成新植株进行无性繁殖, 在地下建立庞大的根茎系统, 形成密集的圆形株丛, 其分株间的间隔子短, 分枝角度小, 能够在竞争过程中占据优势地位.近年来, 在祁连山北坡中山区荒漠草原与典型草原过渡带, 甘肃臭草替代了原生地带性植被的优势种西北针茅(Stipa sareptana var. krylovii ), 形成大量面积不等、繁茂生长的单一优势种群落斑块(赵成章和龙瑞军, 2008 ), 表现出较高的形态可塑性和对逆境的适应能力, 其强大的入侵能力使众多学者对其资源利用策略和生长扩散过程产生了兴趣.目前, 对甘肃臭草型退化草地优势物种空间格局分析及其对水分的响应(赵成章等, 2011 ; 高福元和赵成章, 2012 )、甘肃臭草斑块土壤水分空间异质性(杨泉等, 2014 ), 以及甘肃臭草茎-叶性状、叶内性状(党晶晶等, 2014 ,2015 )等方面进行了研究, 而生境梯度上甘肃臭草根系构型的可塑性变化尚未见报道.鉴于此, 本文在野外调查的基础上, 试图弄清以下问题: (1)不同坡向甘肃臭草根系分叉数和连接长度的权衡关系如何?(2)导致甘肃臭草根系分叉数和连接长度变化的原因有哪些?旨在探讨祁连山地甘肃臭草种群根系构型构建的资源配置模式, 进一步认识甘肃臭草单一优势种群斑块形成机制和扩散机理. ...
Leaf, stem and root tissue strategies across 758 neotropical tree species.
1
2012
... 植物的生态策略和适应性、响应性功能要通过植物内部与外部不同功能性状来实现(Bernard- Verdier et al ., 2012 ), 性状的构建、稳固和性状之间的强弱变化与替补转换则靠植物自身的资源权衡和补偿机制来调控(Fortunel et al ., 2012 ; 陈莹婷和许振柱, 2014 ), 是植物在特定环境下生存策略的重要表现(Mooney et al ., 2010 ).根系构型决定了根系在土壤空间中的位置和资源获取方式, 是植物与资源环境异质性相互适应的结果(Malamy, 2005 ; Rogers & Benfey, 2015 ).根系分叉数和连接长度作为根系构型的重要特征指标, 受土壤空间异质性(Villordon et al ., 2014 )、群落环境(单立山等, 2012 )、遗传特性(Kiswara et al ., 2009 ; Bingham & Wu, 2011 )等诸多因素的影响, 在其生存和发展的过程中, 会根据所处生境的资源条件, 调整、转换或补偿自身功能, 实现根系功能性状间的资源权衡.根系分叉数的变化会影响根系的空间分布属性、营养吸收能力和固定作用; 根系连接长度的大小直接决定了根系在土壤中的空间拓展和营养物质吸收能力(Walk et al ., 2004 ), 二者的权衡关系直接反映了植物根系吸收效率和根系构建过程中所需的碳量(Fitter & Stickland, 1991 ).当根系处于水分状态良好的土层中时, 为了提高水分利用率, 会形成更多的侧根, 增大根分叉数(周艳松和王立群, 2011 ); 在干旱胁迫环境中, 较大的分叉数会引起根系内部对营养物质的竞争(杨小林等, 2008 ), 此时植物倾向于增大连接长度, 以减少根系之间的交叠重复, 占据较大的水分和养分资源空间(郭京衡等, 2014 ), 这是植物为适应不同生境条件而形成的获取资源策略, 以达到“生存、生长、繁殖”之三大目的.地形作为生境条件的一种综合指示, 往往能营造局部小气候(Cantón et al ., 2004 ).坡向作为一种重要的地形要素(Fekedulegn et al ., 2003 ), 可以对局部区域的太阳辐射和降水进行二次分配, 其空间变化是群落组成、物种多样性、生态系统过程和植物功能性状的重要决定因素(de Bello et al ., 2006 ; Bennie et al ., 2008 ; Carletti et al ., 2009 ).在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 在各个水平上影响了植物生长及其对水分利用的有效性(Sardans et al ., 2008 ; Huang et al ., 2015 ), 进而引起植物碳资源分配格局和构型构建策略的改变, 为提高生境适应性和风险规避能力, 植物对连接长度和分叉数等主要根系功能性状进行资源优化配置, 构建与生境相适应的根系构型, 优化根系营养吸收和传输系统.因此, 研究坡向梯度上植物根系构型的生态适应策略, 对于理解植物的环境适应机制有重要意义. ...
甘肃臭草型退化草地优势种群空间格局及其关联性
2
2012
... 近年来, 多数学者对植物根系的研究主要集中在异质生境中根系功能性状可塑性(Grossman & Rice, 2012 ; Pacheco-Villalobos & Hardtk, 2012 )、不同群落环境下根系构型的适应性调整(Rogers & Benfey, 2015 )、根系生长与土壤环境的关系(Forde, 2014 ; Szoboszlay et al ., 2015 )、根系结构和功能(Hodge et al ., 2009 )等方面, 关于退化草地中植物构件生物量分配模式(周兵等, 2015 )、影响根系构型的主要功能性状(周艳松和王立群, 2011 ), 以及不同入侵阶段植物功能性状表型可塑性(Hulme, 2008 ; Pyšek et al ., 2009 )等方面也进行了系统研究, 而从根系构型构建策略的角度出发, 对生境梯度植物根系功能性状间资源权衡机制的研究仍显不足.甘肃臭草(Melica przewalskyi )是禾本科臭草属多年生根茎植物, 通过根状茎的快速横向生长, 长出幼芽和根系, 形成新植株进行无性繁殖, 在地下建立庞大的根茎系统, 形成密集的圆形株丛, 其分株间的间隔子短, 分枝角度小, 能够在竞争过程中占据优势地位.近年来, 在祁连山北坡中山区荒漠草原与典型草原过渡带, 甘肃臭草替代了原生地带性植被的优势种西北针茅(Stipa sareptana var. krylovii ), 形成大量面积不等、繁茂生长的单一优势种群落斑块(赵成章和龙瑞军, 2008 ), 表现出较高的形态可塑性和对逆境的适应能力, 其强大的入侵能力使众多学者对其资源利用策略和生长扩散过程产生了兴趣.目前, 对甘肃臭草型退化草地优势物种空间格局分析及其对水分的响应(赵成章等, 2011 ; 高福元和赵成章, 2012 )、甘肃臭草斑块土壤水分空间异质性(杨泉等, 2014 ), 以及甘肃臭草茎-叶性状、叶内性状(党晶晶等, 2014 ,2015 )等方面进行了研究, 而生境梯度上甘肃臭草根系构型的可塑性变化尚未见报道.鉴于此, 本文在野外调查的基础上, 试图弄清以下问题: (1)不同坡向甘肃臭草根系分叉数和连接长度的权衡关系如何?(2)导致甘肃臭草根系分叉数和连接长度变化的原因有哪些?旨在探讨祁连山地甘肃臭草种群根系构型构建的资源配置模式, 进一步认识甘肃臭草单一优势种群斑块形成机制和扩散机理. ...
... 植物根系构型构建策略的差异可以用根系碳投入与土壤水分养分收益的权衡关系来解释(Oppelt et al ., 2001 ).根系分叉数的增加会导致根系重叠加剧, 增加根系内部的资源竞争态势(杨小林等, 2008 ).根系连接长度的变化会影响根系空间拓展能力和利用土壤资源的有效性(Schenk & Jackson, 2002a , 2002b ), 这与植物自身根系碳投入与土壤水分养分的收益权衡相契合, 是提高植物资源获取能力的一个重要策略(杨小林等, 2008 ).相对于北坡, 山地南坡日照时间长、水分蒸发量大(殷秀琴, 2004 ), 生长在南坡的甘肃臭草多呈直径不等的单一优势种群斑块, 占据了草地群落的绝对资源优势(表1 , 表2 ).甘肃臭草采用克隆繁殖模式形成了庞大的互相连通交织的根茎系统, 构建了高度聚集的根系网络(高福元和赵成章, 2012 ), 斑块内部种群聚集程度较高, 斑块中心土壤含水量明显低于斑块边缘的土壤含水量(赵成章等, 2011 ), 体现了甘肃臭草向斑块周围扩张的迫切需求.在地上和地下生存空间拥挤、土壤含水量分布不均衡状态下, 为了避免不必要的种内竞争, 甘肃臭草限制了地下生物量的分配比例, 以减缓营养繁殖速度, 降低种群聚集强度, 此时甘肃臭草选择了增加根系连接长度、减少分叉数的根系构型构建模式.根系分叉数的减小能够降低种群内部根系重叠程度, 避免不必要的种内竞争; 连接长度的增大可以增强根系空间拓展能力, 扩大单位根系所占的土壤体空间, 使根系从斑块中心向土壤水分较好、生长发育空间较大的边缘区域拓殖, 是南坡甘肃臭草单优种群斑块扩张的主要动力, 与赵成章等(2011) 关于甘肃臭草通过营养繁殖以类似“同心圆”的方式不断由斑块中心向边缘扩大种群领地的观点一致.这符合相关学者“当植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 增加根系连接长度等策略, 以寻求更多的水分和养分, 保证自身的生存和生长”的结论(单立山等, 2013 ; 杜建会等, 2014 ).因此, 南坡甘肃臭草根系分叉数和连接长度间呈极显著负相关关系(p < 0.01, 图2 ), 减小分叉数、增加连接长度是甘肃臭草斑块扩张的根系构型构建策略, 降低了甘肃臭草单一优势种群斑块内部的根系拥挤程度. ...
甘肃臭草型退化草地优势种群空间格局及其关联性
2
2012
... 近年来, 多数学者对植物根系的研究主要集中在异质生境中根系功能性状可塑性(Grossman & Rice, 2012 ; Pacheco-Villalobos & Hardtk, 2012 )、不同群落环境下根系构型的适应性调整(Rogers & Benfey, 2015 )、根系生长与土壤环境的关系(Forde, 2014 ; Szoboszlay et al ., 2015 )、根系结构和功能(Hodge et al ., 2009 )等方面, 关于退化草地中植物构件生物量分配模式(周兵等, 2015 )、影响根系构型的主要功能性状(周艳松和王立群, 2011 ), 以及不同入侵阶段植物功能性状表型可塑性(Hulme, 2008 ; Pyšek et al ., 2009 )等方面也进行了系统研究, 而从根系构型构建策略的角度出发, 对生境梯度植物根系功能性状间资源权衡机制的研究仍显不足.甘肃臭草(Melica przewalskyi )是禾本科臭草属多年生根茎植物, 通过根状茎的快速横向生长, 长出幼芽和根系, 形成新植株进行无性繁殖, 在地下建立庞大的根茎系统, 形成密集的圆形株丛, 其分株间的间隔子短, 分枝角度小, 能够在竞争过程中占据优势地位.近年来, 在祁连山北坡中山区荒漠草原与典型草原过渡带, 甘肃臭草替代了原生地带性植被的优势种西北针茅(Stipa sareptana var. krylovii ), 形成大量面积不等、繁茂生长的单一优势种群落斑块(赵成章和龙瑞军, 2008 ), 表现出较高的形态可塑性和对逆境的适应能力, 其强大的入侵能力使众多学者对其资源利用策略和生长扩散过程产生了兴趣.目前, 对甘肃臭草型退化草地优势物种空间格局分析及其对水分的响应(赵成章等, 2011 ; 高福元和赵成章, 2012 )、甘肃臭草斑块土壤水分空间异质性(杨泉等, 2014 ), 以及甘肃臭草茎-叶性状、叶内性状(党晶晶等, 2014 ,2015 )等方面进行了研究, 而生境梯度上甘肃臭草根系构型的可塑性变化尚未见报道.鉴于此, 本文在野外调查的基础上, 试图弄清以下问题: (1)不同坡向甘肃臭草根系分叉数和连接长度的权衡关系如何?(2)导致甘肃臭草根系分叉数和连接长度变化的原因有哪些?旨在探讨祁连山地甘肃臭草种群根系构型构建的资源配置模式, 进一步认识甘肃臭草单一优势种群斑块形成机制和扩散机理. ...
... 植物根系构型构建策略的差异可以用根系碳投入与土壤水分养分收益的权衡关系来解释(Oppelt et al ., 2001 ).根系分叉数的增加会导致根系重叠加剧, 增加根系内部的资源竞争态势(杨小林等, 2008 ).根系连接长度的变化会影响根系空间拓展能力和利用土壤资源的有效性(Schenk & Jackson, 2002a , 2002b ), 这与植物自身根系碳投入与土壤水分养分的收益权衡相契合, 是提高植物资源获取能力的一个重要策略(杨小林等, 2008 ).相对于北坡, 山地南坡日照时间长、水分蒸发量大(殷秀琴, 2004 ), 生长在南坡的甘肃臭草多呈直径不等的单一优势种群斑块, 占据了草地群落的绝对资源优势(表1 , 表2 ).甘肃臭草采用克隆繁殖模式形成了庞大的互相连通交织的根茎系统, 构建了高度聚集的根系网络(高福元和赵成章, 2012 ), 斑块内部种群聚集程度较高, 斑块中心土壤含水量明显低于斑块边缘的土壤含水量(赵成章等, 2011 ), 体现了甘肃臭草向斑块周围扩张的迫切需求.在地上和地下生存空间拥挤、土壤含水量分布不均衡状态下, 为了避免不必要的种内竞争, 甘肃臭草限制了地下生物量的分配比例, 以减缓营养繁殖速度, 降低种群聚集强度, 此时甘肃臭草选择了增加根系连接长度、减少分叉数的根系构型构建模式.根系分叉数的减小能够降低种群内部根系重叠程度, 避免不必要的种内竞争; 连接长度的增大可以增强根系空间拓展能力, 扩大单位根系所占的土壤体空间, 使根系从斑块中心向土壤水分较好、生长发育空间较大的边缘区域拓殖, 是南坡甘肃臭草单优种群斑块扩张的主要动力, 与赵成章等(2011) 关于甘肃臭草通过营养繁殖以类似“同心圆”的方式不断由斑块中心向边缘扩大种群领地的观点一致.这符合相关学者“当植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 增加根系连接长度等策略, 以寻求更多的水分和养分, 保证自身的生存和生长”的结论(单立山等, 2013 ; 杜建会等, 2014 ).因此, 南坡甘肃臭草根系分叉数和连接长度间呈极显著负相关关系(p < 0.01, 图2 ), 减小分叉数、增加连接长度是甘肃臭草斑块扩张的根系构型构建策略, 降低了甘肃臭草单一优势种群斑块内部的根系拥挤程度. ...
Evolution of root plasticity responses to variation in soil nutrient distribution and concentration.
1
2012
... 近年来, 多数学者对植物根系的研究主要集中在异质生境中根系功能性状可塑性(Grossman & Rice, 2012 ; Pacheco-Villalobos & Hardtk, 2012 )、不同群落环境下根系构型的适应性调整(Rogers & Benfey, 2015 )、根系生长与土壤环境的关系(Forde, 2014 ; Szoboszlay et al ., 2015 )、根系结构和功能(Hodge et al ., 2009 )等方面, 关于退化草地中植物构件生物量分配模式(周兵等, 2015 )、影响根系构型的主要功能性状(周艳松和王立群, 2011 ), 以及不同入侵阶段植物功能性状表型可塑性(Hulme, 2008 ; Pyšek et al ., 2009 )等方面也进行了系统研究, 而从根系构型构建策略的角度出发, 对生境梯度植物根系功能性状间资源权衡机制的研究仍显不足.甘肃臭草(Melica przewalskyi )是禾本科臭草属多年生根茎植物, 通过根状茎的快速横向生长, 长出幼芽和根系, 形成新植株进行无性繁殖, 在地下建立庞大的根茎系统, 形成密集的圆形株丛, 其分株间的间隔子短, 分枝角度小, 能够在竞争过程中占据优势地位.近年来, 在祁连山北坡中山区荒漠草原与典型草原过渡带, 甘肃臭草替代了原生地带性植被的优势种西北针茅(Stipa sareptana var. krylovii ), 形成大量面积不等、繁茂生长的单一优势种群落斑块(赵成章和龙瑞军, 2008 ), 表现出较高的形态可塑性和对逆境的适应能力, 其强大的入侵能力使众多学者对其资源利用策略和生长扩散过程产生了兴趣.目前, 对甘肃臭草型退化草地优势物种空间格局分析及其对水分的响应(赵成章等, 2011 ; 高福元和赵成章, 2012 )、甘肃臭草斑块土壤水分空间异质性(杨泉等, 2014 ), 以及甘肃臭草茎-叶性状、叶内性状(党晶晶等, 2014 ,2015 )等方面进行了研究, 而生境梯度上甘肃臭草根系构型的可塑性变化尚未见报道.鉴于此, 本文在野外调查的基础上, 试图弄清以下问题: (1)不同坡向甘肃臭草根系分叉数和连接长度的权衡关系如何?(2)导致甘肃臭草根系分叉数和连接长度变化的原因有哪些?旨在探讨祁连山地甘肃臭草种群根系构型构建的资源配置模式, 进一步认识甘肃臭草单一优势种群斑块形成机制和扩散机理. ...
塔克拉玛干沙漠南缘三种防护林植物根系构型及其生态适应策略
2
2014
... 植物的生态策略和适应性、响应性功能要通过植物内部与外部不同功能性状来实现(Bernard- Verdier et al ., 2012 ), 性状的构建、稳固和性状之间的强弱变化与替补转换则靠植物自身的资源权衡和补偿机制来调控(Fortunel et al ., 2012 ; 陈莹婷和许振柱, 2014 ), 是植物在特定环境下生存策略的重要表现(Mooney et al ., 2010 ).根系构型决定了根系在土壤空间中的位置和资源获取方式, 是植物与资源环境异质性相互适应的结果(Malamy, 2005 ; Rogers & Benfey, 2015 ).根系分叉数和连接长度作为根系构型的重要特征指标, 受土壤空间异质性(Villordon et al ., 2014 )、群落环境(单立山等, 2012 )、遗传特性(Kiswara et al ., 2009 ; Bingham & Wu, 2011 )等诸多因素的影响, 在其生存和发展的过程中, 会根据所处生境的资源条件, 调整、转换或补偿自身功能, 实现根系功能性状间的资源权衡.根系分叉数的变化会影响根系的空间分布属性、营养吸收能力和固定作用; 根系连接长度的大小直接决定了根系在土壤中的空间拓展和营养物质吸收能力(Walk et al ., 2004 ), 二者的权衡关系直接反映了植物根系吸收效率和根系构建过程中所需的碳量(Fitter & Stickland, 1991 ).当根系处于水分状态良好的土层中时, 为了提高水分利用率, 会形成更多的侧根, 增大根分叉数(周艳松和王立群, 2011 ); 在干旱胁迫环境中, 较大的分叉数会引起根系内部对营养物质的竞争(杨小林等, 2008 ), 此时植物倾向于增大连接长度, 以减少根系之间的交叠重复, 占据较大的水分和养分资源空间(郭京衡等, 2014 ), 这是植物为适应不同生境条件而形成的获取资源策略, 以达到“生存、生长、繁殖”之三大目的.地形作为生境条件的一种综合指示, 往往能营造局部小气候(Cantón et al ., 2004 ).坡向作为一种重要的地形要素(Fekedulegn et al ., 2003 ), 可以对局部区域的太阳辐射和降水进行二次分配, 其空间变化是群落组成、物种多样性、生态系统过程和植物功能性状的重要决定因素(de Bello et al ., 2006 ; Bennie et al ., 2008 ; Carletti et al ., 2009 ).在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 在各个水平上影响了植物生长及其对水分利用的有效性(Sardans et al ., 2008 ; Huang et al ., 2015 ), 进而引起植物碳资源分配格局和构型构建策略的改变, 为提高生境适应性和风险规避能力, 植物对连接长度和分叉数等主要根系功能性状进行资源优化配置, 构建与生境相适应的根系构型, 优化根系营养吸收和传输系统.因此, 研究坡向梯度上植物根系构型的生态适应策略, 对于理解植物的环境适应机制有重要意义. ...
... 植物在环境胁迫下具有权衡性状关系的适应策略(Mooney et al ., 2010 ), 通过生理和发育调节形成与环境相适应的表型特征, 有利于提高植物水分、养分利用效率等生物学功能(李西良等, 2014 ).植物根系构型对环境的适应是通过根系分支结构、空间分布格局等构型特征的改变表现出来的, 是植物长期适应环境的进化结果(Guswa, 2010 ).研究区位于北半球中纬度地区, 山地北坡光照资源最弱(殷秀琴, 2004 )、土壤含水量较高(表2 ), 草地群落的高度、盖度和生物量最高(表2 ), 甘肃臭草处于斑块形成的初期阶段, 其种群密度较小, 也没有高度优势(表1 , 表2 ), 与草地群落的优势种群西北针茅相比, 处于种间竞争的劣势地位.在种间胁迫较大的情况下, 生长在北坡的甘肃臭草采取了增大根系分叉数、减小根系连接长度的根系构型构建策略; 根系分叉数的增加能够增强根系重叠程度、构建密集的根系网络系统, 有利于甘肃臭草地下根系系统和地上株丛的密集生长, 增强种群的密集分布强度, 提高根系对土壤资源的吸收和利用效率, 以斑块式抱团生长实现种群内个体间的高度协同; 减小根系连接长度可能会缩短甘肃臭草克隆分株间的资源传输距离, 有利于增强甘肃臭草克隆分株个体间的相互联系, 保证种群内分株间的资源连接和共享, 从而使种群能够在激烈的资源竞争中存活下来, 与相关学者得出一致的结论, 即“分叉数较大的植物能够迅速占据大量土壤空间快速生长, 适宜生长在土壤资源丰富的环境中” (周艳松和王立群, 2011 ; 郭京衡等, 2014 ).因此, 增大分叉数、减小连接长度是分布于北坡的甘肃臭草增大斑块密度, 提升竞争能力和空间拓殖能力的一种生活史对策, 使根系分叉数和连接长度之间的权衡关系达到极显著水平(p < 0.01, 图2 ). ...
塔克拉玛干沙漠南缘三种防护林植物根系构型及其生态适应策略
2
2014
... 植物的生态策略和适应性、响应性功能要通过植物内部与外部不同功能性状来实现(Bernard- Verdier et al ., 2012 ), 性状的构建、稳固和性状之间的强弱变化与替补转换则靠植物自身的资源权衡和补偿机制来调控(Fortunel et al ., 2012 ; 陈莹婷和许振柱, 2014 ), 是植物在特定环境下生存策略的重要表现(Mooney et al ., 2010 ).根系构型决定了根系在土壤空间中的位置和资源获取方式, 是植物与资源环境异质性相互适应的结果(Malamy, 2005 ; Rogers & Benfey, 2015 ).根系分叉数和连接长度作为根系构型的重要特征指标, 受土壤空间异质性(Villordon et al ., 2014 )、群落环境(单立山等, 2012 )、遗传特性(Kiswara et al ., 2009 ; Bingham & Wu, 2011 )等诸多因素的影响, 在其生存和发展的过程中, 会根据所处生境的资源条件, 调整、转换或补偿自身功能, 实现根系功能性状间的资源权衡.根系分叉数的变化会影响根系的空间分布属性、营养吸收能力和固定作用; 根系连接长度的大小直接决定了根系在土壤中的空间拓展和营养物质吸收能力(Walk et al ., 2004 ), 二者的权衡关系直接反映了植物根系吸收效率和根系构建过程中所需的碳量(Fitter & Stickland, 1991 ).当根系处于水分状态良好的土层中时, 为了提高水分利用率, 会形成更多的侧根, 增大根分叉数(周艳松和王立群, 2011 ); 在干旱胁迫环境中, 较大的分叉数会引起根系内部对营养物质的竞争(杨小林等, 2008 ), 此时植物倾向于增大连接长度, 以减少根系之间的交叠重复, 占据较大的水分和养分资源空间(郭京衡等, 2014 ), 这是植物为适应不同生境条件而形成的获取资源策略, 以达到“生存、生长、繁殖”之三大目的.地形作为生境条件的一种综合指示, 往往能营造局部小气候(Cantón et al ., 2004 ).坡向作为一种重要的地形要素(Fekedulegn et al ., 2003 ), 可以对局部区域的太阳辐射和降水进行二次分配, 其空间变化是群落组成、物种多样性、生态系统过程和植物功能性状的重要决定因素(de Bello et al ., 2006 ; Bennie et al ., 2008 ; Carletti et al ., 2009 ).在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 在各个水平上影响了植物生长及其对水分利用的有效性(Sardans et al ., 2008 ; Huang et al ., 2015 ), 进而引起植物碳资源分配格局和构型构建策略的改变, 为提高生境适应性和风险规避能力, 植物对连接长度和分叉数等主要根系功能性状进行资源优化配置, 构建与生境相适应的根系构型, 优化根系营养吸收和传输系统.因此, 研究坡向梯度上植物根系构型的生态适应策略, 对于理解植物的环境适应机制有重要意义. ...
... 植物在环境胁迫下具有权衡性状关系的适应策略(Mooney et al ., 2010 ), 通过生理和发育调节形成与环境相适应的表型特征, 有利于提高植物水分、养分利用效率等生物学功能(李西良等, 2014 ).植物根系构型对环境的适应是通过根系分支结构、空间分布格局等构型特征的改变表现出来的, 是植物长期适应环境的进化结果(Guswa, 2010 ).研究区位于北半球中纬度地区, 山地北坡光照资源最弱(殷秀琴, 2004 )、土壤含水量较高(表2 ), 草地群落的高度、盖度和生物量最高(表2 ), 甘肃臭草处于斑块形成的初期阶段, 其种群密度较小, 也没有高度优势(表1 , 表2 ), 与草地群落的优势种群西北针茅相比, 处于种间竞争的劣势地位.在种间胁迫较大的情况下, 生长在北坡的甘肃臭草采取了增大根系分叉数、减小根系连接长度的根系构型构建策略; 根系分叉数的增加能够增强根系重叠程度、构建密集的根系网络系统, 有利于甘肃臭草地下根系系统和地上株丛的密集生长, 增强种群的密集分布强度, 提高根系对土壤资源的吸收和利用效率, 以斑块式抱团生长实现种群内个体间的高度协同; 减小根系连接长度可能会缩短甘肃臭草克隆分株间的资源传输距离, 有利于增强甘肃臭草克隆分株个体间的相互联系, 保证种群内分株间的资源连接和共享, 从而使种群能够在激烈的资源竞争中存活下来, 与相关学者得出一致的结论, 即“分叉数较大的植物能够迅速占据大量土壤空间快速生长, 适宜生长在土壤资源丰富的环境中” (周艳松和王立群, 2011 ; 郭京衡等, 2014 ).因此, 增大分叉数、减小连接长度是分布于北坡的甘肃臭草增大斑块密度, 提升竞争能力和空间拓殖能力的一种生活史对策, 使根系分叉数和连接长度之间的权衡关系达到极显著水平(p < 0.01, 图2 ). ...
Effect of plant uptake strategy on the water- optimal root depth.
1
2010
... 植物在环境胁迫下具有权衡性状关系的适应策略(Mooney et al ., 2010 ), 通过生理和发育调节形成与环境相适应的表型特征, 有利于提高植物水分、养分利用效率等生物学功能(李西良等, 2014 ).植物根系构型对环境的适应是通过根系分支结构、空间分布格局等构型特征的改变表现出来的, 是植物长期适应环境的进化结果(Guswa, 2010 ).研究区位于北半球中纬度地区, 山地北坡光照资源最弱(殷秀琴, 2004 )、土壤含水量较高(表2 ), 草地群落的高度、盖度和生物量最高(表2 ), 甘肃臭草处于斑块形成的初期阶段, 其种群密度较小, 也没有高度优势(表1 , 表2 ), 与草地群落的优势种群西北针茅相比, 处于种间竞争的劣势地位.在种间胁迫较大的情况下, 生长在北坡的甘肃臭草采取了增大根系分叉数、减小根系连接长度的根系构型构建策略; 根系分叉数的增加能够增强根系重叠程度、构建密集的根系网络系统, 有利于甘肃臭草地下根系系统和地上株丛的密集生长, 增强种群的密集分布强度, 提高根系对土壤资源的吸收和利用效率, 以斑块式抱团生长实现种群内个体间的高度协同; 减小根系连接长度可能会缩短甘肃臭草克隆分株间的资源传输距离, 有利于增强甘肃臭草克隆分株个体间的相互联系, 保证种群内分株间的资源连接和共享, 从而使种群能够在激烈的资源竞争中存活下来, 与相关学者得出一致的结论, 即“分叉数较大的植物能够迅速占据大量土壤空间快速生长, 适宜生长在土壤资源丰富的环境中” (周艳松和王立群, 2011 ; 郭京衡等, 2014 ).因此, 增大分叉数、减小连接长度是分布于北坡的甘肃臭草增大斑块密度, 提升竞争能力和空间拓殖能力的一种生活史对策, 使根系分叉数和连接长度之间的权衡关系达到极显著水平(p < 0.01, 图2 ). ...
Plant root growth, architecture and function.
1
2009
... 近年来, 多数学者对植物根系的研究主要集中在异质生境中根系功能性状可塑性(Grossman & Rice, 2012 ; Pacheco-Villalobos & Hardtk, 2012 )、不同群落环境下根系构型的适应性调整(Rogers & Benfey, 2015 )、根系生长与土壤环境的关系(Forde, 2014 ; Szoboszlay et al ., 2015 )、根系结构和功能(Hodge et al ., 2009 )等方面, 关于退化草地中植物构件生物量分配模式(周兵等, 2015 )、影响根系构型的主要功能性状(周艳松和王立群, 2011 ), 以及不同入侵阶段植物功能性状表型可塑性(Hulme, 2008 ; Pyšek et al ., 2009 )等方面也进行了系统研究, 而从根系构型构建策略的角度出发, 对生境梯度植物根系功能性状间资源权衡机制的研究仍显不足.甘肃臭草(Melica przewalskyi )是禾本科臭草属多年生根茎植物, 通过根状茎的快速横向生长, 长出幼芽和根系, 形成新植株进行无性繁殖, 在地下建立庞大的根茎系统, 形成密集的圆形株丛, 其分株间的间隔子短, 分枝角度小, 能够在竞争过程中占据优势地位.近年来, 在祁连山北坡中山区荒漠草原与典型草原过渡带, 甘肃臭草替代了原生地带性植被的优势种西北针茅(Stipa sareptana var. krylovii ), 形成大量面积不等、繁茂生长的单一优势种群落斑块(赵成章和龙瑞军, 2008 ), 表现出较高的形态可塑性和对逆境的适应能力, 其强大的入侵能力使众多学者对其资源利用策略和生长扩散过程产生了兴趣.目前, 对甘肃臭草型退化草地优势物种空间格局分析及其对水分的响应(赵成章等, 2011 ; 高福元和赵成章, 2012 )、甘肃臭草斑块土壤水分空间异质性(杨泉等, 2014 ), 以及甘肃臭草茎-叶性状、叶内性状(党晶晶等, 2014 ,2015 )等方面进行了研究, 而生境梯度上甘肃臭草根系构型的可塑性变化尚未见报道.鉴于此, 本文在野外调查的基础上, 试图弄清以下问题: (1)不同坡向甘肃臭草根系分叉数和连接长度的权衡关系如何?(2)导致甘肃臭草根系分叉数和连接长度变化的原因有哪些?旨在探讨祁连山地甘肃臭草种群根系构型构建的资源配置模式, 进一步认识甘肃臭草单一优势种群斑块形成机制和扩散机理. ...
Effects of slope aspect on soil nitrogen and microbial properties in the Chinese Loess region.
1
2015
... 植物的生态策略和适应性、响应性功能要通过植物内部与外部不同功能性状来实现(Bernard- Verdier et al ., 2012 ), 性状的构建、稳固和性状之间的强弱变化与替补转换则靠植物自身的资源权衡和补偿机制来调控(Fortunel et al ., 2012 ; 陈莹婷和许振柱, 2014 ), 是植物在特定环境下生存策略的重要表现(Mooney et al ., 2010 ).根系构型决定了根系在土壤空间中的位置和资源获取方式, 是植物与资源环境异质性相互适应的结果(Malamy, 2005 ; Rogers & Benfey, 2015 ).根系分叉数和连接长度作为根系构型的重要特征指标, 受土壤空间异质性(Villordon et al ., 2014 )、群落环境(单立山等, 2012 )、遗传特性(Kiswara et al ., 2009 ; Bingham & Wu, 2011 )等诸多因素的影响, 在其生存和发展的过程中, 会根据所处生境的资源条件, 调整、转换或补偿自身功能, 实现根系功能性状间的资源权衡.根系分叉数的变化会影响根系的空间分布属性、营养吸收能力和固定作用; 根系连接长度的大小直接决定了根系在土壤中的空间拓展和营养物质吸收能力(Walk et al ., 2004 ), 二者的权衡关系直接反映了植物根系吸收效率和根系构建过程中所需的碳量(Fitter & Stickland, 1991 ).当根系处于水分状态良好的土层中时, 为了提高水分利用率, 会形成更多的侧根, 增大根分叉数(周艳松和王立群, 2011 ); 在干旱胁迫环境中, 较大的分叉数会引起根系内部对营养物质的竞争(杨小林等, 2008 ), 此时植物倾向于增大连接长度, 以减少根系之间的交叠重复, 占据较大的水分和养分资源空间(郭京衡等, 2014 ), 这是植物为适应不同生境条件而形成的获取资源策略, 以达到“生存、生长、繁殖”之三大目的.地形作为生境条件的一种综合指示, 往往能营造局部小气候(Cantón et al ., 2004 ).坡向作为一种重要的地形要素(Fekedulegn et al ., 2003 ), 可以对局部区域的太阳辐射和降水进行二次分配, 其空间变化是群落组成、物种多样性、生态系统过程和植物功能性状的重要决定因素(de Bello et al ., 2006 ; Bennie et al ., 2008 ; Carletti et al ., 2009 ).在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 在各个水平上影响了植物生长及其对水分利用的有效性(Sardans et al ., 2008 ; Huang et al ., 2015 ), 进而引起植物碳资源分配格局和构型构建策略的改变, 为提高生境适应性和风险规避能力, 植物对连接长度和分叉数等主要根系功能性状进行资源优化配置, 构建与生境相适应的根系构型, 优化根系营养吸收和传输系统.因此, 研究坡向梯度上植物根系构型的生态适应策略, 对于理解植物的环境适应机制有重要意义. ...
Phenotypic plasticity and plant invasions: Is it all jack?
1
2008
... 近年来, 多数学者对植物根系的研究主要集中在异质生境中根系功能性状可塑性(Grossman & Rice, 2012 ; Pacheco-Villalobos & Hardtk, 2012 )、不同群落环境下根系构型的适应性调整(Rogers & Benfey, 2015 )、根系生长与土壤环境的关系(Forde, 2014 ; Szoboszlay et al ., 2015 )、根系结构和功能(Hodge et al ., 2009 )等方面, 关于退化草地中植物构件生物量分配模式(周兵等, 2015 )、影响根系构型的主要功能性状(周艳松和王立群, 2011 ), 以及不同入侵阶段植物功能性状表型可塑性(Hulme, 2008 ; Pyšek et al ., 2009 )等方面也进行了系统研究, 而从根系构型构建策略的角度出发, 对生境梯度植物根系功能性状间资源权衡机制的研究仍显不足.甘肃臭草(Melica przewalskyi )是禾本科臭草属多年生根茎植物, 通过根状茎的快速横向生长, 长出幼芽和根系, 形成新植株进行无性繁殖, 在地下建立庞大的根茎系统, 形成密集的圆形株丛, 其分株间的间隔子短, 分枝角度小, 能够在竞争过程中占据优势地位.近年来, 在祁连山北坡中山区荒漠草原与典型草原过渡带, 甘肃臭草替代了原生地带性植被的优势种西北针茅(Stipa sareptana var. krylovii ), 形成大量面积不等、繁茂生长的单一优势种群落斑块(赵成章和龙瑞军, 2008 ), 表现出较高的形态可塑性和对逆境的适应能力, 其强大的入侵能力使众多学者对其资源利用策略和生长扩散过程产生了兴趣.目前, 对甘肃臭草型退化草地优势物种空间格局分析及其对水分的响应(赵成章等, 2011 ; 高福元和赵成章, 2012 )、甘肃臭草斑块土壤水分空间异质性(杨泉等, 2014 ), 以及甘肃臭草茎-叶性状、叶内性状(党晶晶等, 2014 ,2015 )等方面进行了研究, 而生境梯度上甘肃臭草根系构型的可塑性变化尚未见报道.鉴于此, 本文在野外调查的基础上, 试图弄清以下问题: (1)不同坡向甘肃臭草根系分叉数和连接长度的权衡关系如何?(2)导致甘肃臭草根系分叉数和连接长度变化的原因有哪些?旨在探讨祁连山地甘肃臭草种群根系构型构建的资源配置模式, 进一步认识甘肃臭草单一优势种群斑块形成机制和扩散机理. ...
Root architecture of six tropical seagrass species, growing in three contrasting habitats in Indonesian waters.
1
2009
... 植物的生态策略和适应性、响应性功能要通过植物内部与外部不同功能性状来实现(Bernard- Verdier et al ., 2012 ), 性状的构建、稳固和性状之间的强弱变化与替补转换则靠植物自身的资源权衡和补偿机制来调控(Fortunel et al ., 2012 ; 陈莹婷和许振柱, 2014 ), 是植物在特定环境下生存策略的重要表现(Mooney et al ., 2010 ).根系构型决定了根系在土壤空间中的位置和资源获取方式, 是植物与资源环境异质性相互适应的结果(Malamy, 2005 ; Rogers & Benfey, 2015 ).根系分叉数和连接长度作为根系构型的重要特征指标, 受土壤空间异质性(Villordon et al ., 2014 )、群落环境(单立山等, 2012 )、遗传特性(Kiswara et al ., 2009 ; Bingham & Wu, 2011 )等诸多因素的影响, 在其生存和发展的过程中, 会根据所处生境的资源条件, 调整、转换或补偿自身功能, 实现根系功能性状间的资源权衡.根系分叉数的变化会影响根系的空间分布属性、营养吸收能力和固定作用; 根系连接长度的大小直接决定了根系在土壤中的空间拓展和营养物质吸收能力(Walk et al ., 2004 ), 二者的权衡关系直接反映了植物根系吸收效率和根系构建过程中所需的碳量(Fitter & Stickland, 1991 ).当根系处于水分状态良好的土层中时, 为了提高水分利用率, 会形成更多的侧根, 增大根分叉数(周艳松和王立群, 2011 ); 在干旱胁迫环境中, 较大的分叉数会引起根系内部对营养物质的竞争(杨小林等, 2008 ), 此时植物倾向于增大连接长度, 以减少根系之间的交叠重复, 占据较大的水分和养分资源空间(郭京衡等, 2014 ), 这是植物为适应不同生境条件而形成的获取资源策略, 以达到“生存、生长、繁殖”之三大目的.地形作为生境条件的一种综合指示, 往往能营造局部小气候(Cantón et al ., 2004 ).坡向作为一种重要的地形要素(Fekedulegn et al ., 2003 ), 可以对局部区域的太阳辐射和降水进行二次分配, 其空间变化是群落组成、物种多样性、生态系统过程和植物功能性状的重要决定因素(de Bello et al ., 2006 ; Bennie et al ., 2008 ; Carletti et al ., 2009 ).在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 在各个水平上影响了植物生长及其对水分利用的有效性(Sardans et al ., 2008 ; Huang et al ., 2015 ), 进而引起植物碳资源分配格局和构型构建策略的改变, 为提高生境适应性和风险规避能力, 植物对连接长度和分叉数等主要根系功能性状进行资源优化配置, 构建与生境相适应的根系构型, 优化根系营养吸收和传输系统.因此, 研究坡向梯度上植物根系构型的生态适应策略, 对于理解植物的环境适应机制有重要意义. ...
Designer crops: Optimal root system architecture for nutrient acquisition.
1
2014
... 植物在不同功能和性状之间对有限资源的分配存在着互相牵制的权衡作用(Shipley et al ., 2006 ), 这就需要植物在生长发育过程中, 通过不断优化自身的资源分配来提高适合度以适应环境的变化(Fabbro & Körner, 2004).根系作为植物地上部分与土壤的连接器官, 在较大程度上影响着植物的水分和养分吸收、运移和存储功能(Dannowski & Block, 2005 ; Kong et al ., 2014 ), 根系分叉数和连接长度能够反映根系不同部位构建过程中的碳消耗特点, 影响根系拓展空间、获取营养的代价与效率, 体现了植物性状和构型构建对生境变化的响应, 从中可以理解和阐释植物的物种分布、种间关系、权衡及其作用机制(Westoby & Wright, 2006 ).本研究发现, 随着坡向由北坡向东坡、西坡、南坡转变, 甘肃臭草根系分叉数呈逐渐减小的趋势, 而根系连接长度呈相反的变化趋势(图1 ), 即甘肃臭草分配给根系分叉数和连接长度的资源间存在着“此消彼长”的权衡关系(图2 ).经Pearson相关性分析得出, 群落密度与根系分叉数、连接长度呈极显著相关关系(p < 0.01), 群落高度、土壤含水量与二者显著相关(p < 0.05), 地上生物量与根系分叉数显著相关(p < 0.05, 表3 ), 表明坡向梯度上甘肃臭草根系分叉数和连接长度权衡关系的差异性, 是植物根系构建的资源配置模式对高寒草地生境异质性适应的结果. ...
草甸草原羊草茎叶功能性状对长期过度放牧的可塑性响应
1
2014
... 植物在环境胁迫下具有权衡性状关系的适应策略(Mooney et al ., 2010 ), 通过生理和发育调节形成与环境相适应的表型特征, 有利于提高植物水分、养分利用效率等生物学功能(李西良等, 2014 ).植物根系构型对环境的适应是通过根系分支结构、空间分布格局等构型特征的改变表现出来的, 是植物长期适应环境的进化结果(Guswa, 2010 ).研究区位于北半球中纬度地区, 山地北坡光照资源最弱(殷秀琴, 2004 )、土壤含水量较高(表2 ), 草地群落的高度、盖度和生物量最高(表2 ), 甘肃臭草处于斑块形成的初期阶段, 其种群密度较小, 也没有高度优势(表1 , 表2 ), 与草地群落的优势种群西北针茅相比, 处于种间竞争的劣势地位.在种间胁迫较大的情况下, 生长在北坡的甘肃臭草采取了增大根系分叉数、减小根系连接长度的根系构型构建策略; 根系分叉数的增加能够增强根系重叠程度、构建密集的根系网络系统, 有利于甘肃臭草地下根系系统和地上株丛的密集生长, 增强种群的密集分布强度, 提高根系对土壤资源的吸收和利用效率, 以斑块式抱团生长实现种群内个体间的高度协同; 减小根系连接长度可能会缩短甘肃臭草克隆分株间的资源传输距离, 有利于增强甘肃臭草克隆分株个体间的相互联系, 保证种群内分株间的资源连接和共享, 从而使种群能够在激烈的资源竞争中存活下来, 与相关学者得出一致的结论, 即“分叉数较大的植物能够迅速占据大量土壤空间快速生长, 适宜生长在土壤资源丰富的环境中” (周艳松和王立群, 2011 ; 郭京衡等, 2014 ).因此, 增大分叉数、减小连接长度是分布于北坡的甘肃臭草增大斑块密度, 提升竞争能力和空间拓殖能力的一种生活史对策, 使根系分叉数和连接长度之间的权衡关系达到极显著水平(p < 0.01, 图2 ). ...
草甸草原羊草茎叶功能性状对长期过度放牧的可塑性响应
1
2014
... 植物在环境胁迫下具有权衡性状关系的适应策略(Mooney et al ., 2010 ), 通过生理和发育调节形成与环境相适应的表型特征, 有利于提高植物水分、养分利用效率等生物学功能(李西良等, 2014 ).植物根系构型对环境的适应是通过根系分支结构、空间分布格局等构型特征的改变表现出来的, 是植物长期适应环境的进化结果(Guswa, 2010 ).研究区位于北半球中纬度地区, 山地北坡光照资源最弱(殷秀琴, 2004 )、土壤含水量较高(表2 ), 草地群落的高度、盖度和生物量最高(表2 ), 甘肃臭草处于斑块形成的初期阶段, 其种群密度较小, 也没有高度优势(表1 , 表2 ), 与草地群落的优势种群西北针茅相比, 处于种间竞争的劣势地位.在种间胁迫较大的情况下, 生长在北坡的甘肃臭草采取了增大根系分叉数、减小根系连接长度的根系构型构建策略; 根系分叉数的增加能够增强根系重叠程度、构建密集的根系网络系统, 有利于甘肃臭草地下根系系统和地上株丛的密集生长, 增强种群的密集分布强度, 提高根系对土壤资源的吸收和利用效率, 以斑块式抱团生长实现种群内个体间的高度协同; 减小根系连接长度可能会缩短甘肃臭草克隆分株间的资源传输距离, 有利于增强甘肃臭草克隆分株个体间的相互联系, 保证种群内分株间的资源连接和共享, 从而使种群能够在激烈的资源竞争中存活下来, 与相关学者得出一致的结论, 即“分叉数较大的植物能够迅速占据大量土壤空间快速生长, 适宜生长在土壤资源丰富的环境中” (周艳松和王立群, 2011 ; 郭京衡等, 2014 ).因此, 增大分叉数、减小连接长度是分布于北坡的甘肃臭草增大斑块密度, 提升竞争能力和空间拓殖能力的一种生活史对策, 使根系分叉数和连接长度之间的权衡关系达到极显著水平(p < 0.01, 图2 ). ...
湖南会同5个亚热带树种的细根构型及功能特征分析
1
2010
... 将80个样地划分为北坡(315°-360°和0-45°)、东坡(45°-135°)、南坡(135°-225°)和西坡(225°-315°) 4个坡向组.利用Win-RHIZO根系分析软件分析提取的根系分叉的数量, 计算出各坡向甘肃臭草的根系分叉数.分叉数是指单位根长上分叉的数量, 在相同的单位长度内, 分叉数越高, 表明根系吸收养分或水分的能力越强(刘佳等, 2010 ).计算公式为: ...
湖南会同5个亚热带树种的细根构型及功能特征分析
1
2010
... 将80个样地划分为北坡(315°-360°和0-45°)、东坡(45°-135°)、南坡(135°-225°)和西坡(225°-315°) 4个坡向组.利用Win-RHIZO根系分析软件分析提取的根系分叉的数量, 计算出各坡向甘肃臭草的根系分叉数.分叉数是指单位根长上分叉的数量, 在相同的单位长度内, 分叉数越高, 表明根系吸收养分或水分的能力越强(刘佳等, 2010 ).计算公式为: ...
Intrinsic and environmental response pathways that regulate root system architecture.
1
2005
... 植物的生态策略和适应性、响应性功能要通过植物内部与外部不同功能性状来实现(Bernard- Verdier et al ., 2012 ), 性状的构建、稳固和性状之间的强弱变化与替补转换则靠植物自身的资源权衡和补偿机制来调控(Fortunel et al ., 2012 ; 陈莹婷和许振柱, 2014 ), 是植物在特定环境下生存策略的重要表现(Mooney et al ., 2010 ).根系构型决定了根系在土壤空间中的位置和资源获取方式, 是植物与资源环境异质性相互适应的结果(Malamy, 2005 ; Rogers & Benfey, 2015 ).根系分叉数和连接长度作为根系构型的重要特征指标, 受土壤空间异质性(Villordon et al ., 2014 )、群落环境(单立山等, 2012 )、遗传特性(Kiswara et al ., 2009 ; Bingham & Wu, 2011 )等诸多因素的影响, 在其生存和发展的过程中, 会根据所处生境的资源条件, 调整、转换或补偿自身功能, 实现根系功能性状间的资源权衡.根系分叉数的变化会影响根系的空间分布属性、营养吸收能力和固定作用; 根系连接长度的大小直接决定了根系在土壤中的空间拓展和营养物质吸收能力(Walk et al ., 2004 ), 二者的权衡关系直接反映了植物根系吸收效率和根系构建过程中所需的碳量(Fitter & Stickland, 1991 ).当根系处于水分状态良好的土层中时, 为了提高水分利用率, 会形成更多的侧根, 增大根分叉数(周艳松和王立群, 2011 ); 在干旱胁迫环境中, 较大的分叉数会引起根系内部对营养物质的竞争(杨小林等, 2008 ), 此时植物倾向于增大连接长度, 以减少根系之间的交叠重复, 占据较大的水分和养分资源空间(郭京衡等, 2014 ), 这是植物为适应不同生境条件而形成的获取资源策略, 以达到“生存、生长、繁殖”之三大目的.地形作为生境条件的一种综合指示, 往往能营造局部小气候(Cantón et al ., 2004 ).坡向作为一种重要的地形要素(Fekedulegn et al ., 2003 ), 可以对局部区域的太阳辐射和降水进行二次分配, 其空间变化是群落组成、物种多样性、生态系统过程和植物功能性状的重要决定因素(de Bello et al ., 2006 ; Bennie et al ., 2008 ; Carletti et al ., 2009 ).在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 在各个水平上影响了植物生长及其对水分利用的有效性(Sardans et al ., 2008 ; Huang et al ., 2015 ), 进而引起植物碳资源分配格局和构型构建策略的改变, 为提高生境适应性和风险规避能力, 植物对连接长度和分叉数等主要根系功能性状进行资源优化配置, 构建与生境相适应的根系构型, 优化根系营养吸收和传输系统.因此, 研究坡向梯度上植物根系构型的生态适应策略, 对于理解植物的环境适应机制有重要意义. ...
Evolutionary trade-offs in plants mediate the strength of trophic cascades.
2
2010
... 植物的生态策略和适应性、响应性功能要通过植物内部与外部不同功能性状来实现(Bernard- Verdier et al ., 2012 ), 性状的构建、稳固和性状之间的强弱变化与替补转换则靠植物自身的资源权衡和补偿机制来调控(Fortunel et al ., 2012 ; 陈莹婷和许振柱, 2014 ), 是植物在特定环境下生存策略的重要表现(Mooney et al ., 2010 ).根系构型决定了根系在土壤空间中的位置和资源获取方式, 是植物与资源环境异质性相互适应的结果(Malamy, 2005 ; Rogers & Benfey, 2015 ).根系分叉数和连接长度作为根系构型的重要特征指标, 受土壤空间异质性(Villordon et al ., 2014 )、群落环境(单立山等, 2012 )、遗传特性(Kiswara et al ., 2009 ; Bingham & Wu, 2011 )等诸多因素的影响, 在其生存和发展的过程中, 会根据所处生境的资源条件, 调整、转换或补偿自身功能, 实现根系功能性状间的资源权衡.根系分叉数的变化会影响根系的空间分布属性、营养吸收能力和固定作用; 根系连接长度的大小直接决定了根系在土壤中的空间拓展和营养物质吸收能力(Walk et al ., 2004 ), 二者的权衡关系直接反映了植物根系吸收效率和根系构建过程中所需的碳量(Fitter & Stickland, 1991 ).当根系处于水分状态良好的土层中时, 为了提高水分利用率, 会形成更多的侧根, 增大根分叉数(周艳松和王立群, 2011 ); 在干旱胁迫环境中, 较大的分叉数会引起根系内部对营养物质的竞争(杨小林等, 2008 ), 此时植物倾向于增大连接长度, 以减少根系之间的交叠重复, 占据较大的水分和养分资源空间(郭京衡等, 2014 ), 这是植物为适应不同生境条件而形成的获取资源策略, 以达到“生存、生长、繁殖”之三大目的.地形作为生境条件的一种综合指示, 往往能营造局部小气候(Cantón et al ., 2004 ).坡向作为一种重要的地形要素(Fekedulegn et al ., 2003 ), 可以对局部区域的太阳辐射和降水进行二次分配, 其空间变化是群落组成、物种多样性、生态系统过程和植物功能性状的重要决定因素(de Bello et al ., 2006 ; Bennie et al ., 2008 ; Carletti et al ., 2009 ).在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 在各个水平上影响了植物生长及其对水分利用的有效性(Sardans et al ., 2008 ; Huang et al ., 2015 ), 进而引起植物碳资源分配格局和构型构建策略的改变, 为提高生境适应性和风险规避能力, 植物对连接长度和分叉数等主要根系功能性状进行资源优化配置, 构建与生境相适应的根系构型, 优化根系营养吸收和传输系统.因此, 研究坡向梯度上植物根系构型的生态适应策略, 对于理解植物的环境适应机制有重要意义. ...
... 植物在环境胁迫下具有权衡性状关系的适应策略(Mooney et al ., 2010 ), 通过生理和发育调节形成与环境相适应的表型特征, 有利于提高植物水分、养分利用效率等生物学功能(李西良等, 2014 ).植物根系构型对环境的适应是通过根系分支结构、空间分布格局等构型特征的改变表现出来的, 是植物长期适应环境的进化结果(Guswa, 2010 ).研究区位于北半球中纬度地区, 山地北坡光照资源最弱(殷秀琴, 2004 )、土壤含水量较高(表2 ), 草地群落的高度、盖度和生物量最高(表2 ), 甘肃臭草处于斑块形成的初期阶段, 其种群密度较小, 也没有高度优势(表1 , 表2 ), 与草地群落的优势种群西北针茅相比, 处于种间竞争的劣势地位.在种间胁迫较大的情况下, 生长在北坡的甘肃臭草采取了增大根系分叉数、减小根系连接长度的根系构型构建策略; 根系分叉数的增加能够增强根系重叠程度、构建密集的根系网络系统, 有利于甘肃臭草地下根系系统和地上株丛的密集生长, 增强种群的密集分布强度, 提高根系对土壤资源的吸收和利用效率, 以斑块式抱团生长实现种群内个体间的高度协同; 减小根系连接长度可能会缩短甘肃臭草克隆分株间的资源传输距离, 有利于增强甘肃臭草克隆分株个体间的相互联系, 保证种群内分株间的资源连接和共享, 从而使种群能够在激烈的资源竞争中存活下来, 与相关学者得出一致的结论, 即“分叉数较大的植物能够迅速占据大量土壤空间快速生长, 适宜生长在土壤资源丰富的环境中” (周艳松和王立群, 2011 ; 郭京衡等, 2014 ).因此, 增大分叉数、减小连接长度是分布于北坡的甘肃臭草增大斑块密度, 提升竞争能力和空间拓殖能力的一种生活史对策, 使根系分叉数和连接长度之间的权衡关系达到极显著水平(p < 0.01, 图2 ). ...
Topology, scaling relations and Leonardo’s rule in root systems from African tree species.
1
2001
... 植物根系构型构建策略的差异可以用根系碳投入与土壤水分养分收益的权衡关系来解释(Oppelt et al ., 2001 ).根系分叉数的增加会导致根系重叠加剧, 增加根系内部的资源竞争态势(杨小林等, 2008 ).根系连接长度的变化会影响根系空间拓展能力和利用土壤资源的有效性(Schenk & Jackson, 2002a , 2002b ), 这与植物自身根系碳投入与土壤水分养分的收益权衡相契合, 是提高植物资源获取能力的一个重要策略(杨小林等, 2008 ).相对于北坡, 山地南坡日照时间长、水分蒸发量大(殷秀琴, 2004 ), 生长在南坡的甘肃臭草多呈直径不等的单一优势种群斑块, 占据了草地群落的绝对资源优势(表1 , 表2 ).甘肃臭草采用克隆繁殖模式形成了庞大的互相连通交织的根茎系统, 构建了高度聚集的根系网络(高福元和赵成章, 2012 ), 斑块内部种群聚集程度较高, 斑块中心土壤含水量明显低于斑块边缘的土壤含水量(赵成章等, 2011 ), 体现了甘肃臭草向斑块周围扩张的迫切需求.在地上和地下生存空间拥挤、土壤含水量分布不均衡状态下, 为了避免不必要的种内竞争, 甘肃臭草限制了地下生物量的分配比例, 以减缓营养繁殖速度, 降低种群聚集强度, 此时甘肃臭草选择了增加根系连接长度、减少分叉数的根系构型构建模式.根系分叉数的减小能够降低种群内部根系重叠程度, 避免不必要的种内竞争; 连接长度的增大可以增强根系空间拓展能力, 扩大单位根系所占的土壤体空间, 使根系从斑块中心向土壤水分较好、生长发育空间较大的边缘区域拓殖, 是南坡甘肃臭草单优种群斑块扩张的主要动力, 与赵成章等(2011) 关于甘肃臭草通过营养繁殖以类似“同心圆”的方式不断由斑块中心向边缘扩大种群领地的观点一致.这符合相关学者“当植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 增加根系连接长度等策略, 以寻求更多的水分和养分, 保证自身的生存和生长”的结论(单立山等, 2013 ; 杜建会等, 2014 ).因此, 南坡甘肃臭草根系分叉数和连接长度间呈极显著负相关关系(p < 0.01, 图2 ), 减小分叉数、增加连接长度是甘肃臭草斑块扩张的根系构型构建策略, 降低了甘肃臭草单一优势种群斑块内部的根系拥挤程度. ...
Contrasting rooting patterns of some arid-zone fruit tree species from Botswana. II. Coarse root distribution.
1
2005
... 环境异质性是植物群落结构特征差异形成的主要原因之一, 随着资源环境的改变, 植物群落的地上生物量、总盖度和平均高度都会发生明显变化, 必然会改变种群内每块植株可获得性资源的数量.根系构型会对周围环境产生适应性的改变(Oppelt et al ., 2005 ), 通过权衡各功能性状指标间的生物量分配模式, 以维持个体生长率最大.东坡和西坡属于半阴坡和半阳坡, 是植被类型的交错带, 植被多样性较高, 群落组成也较为复杂, 草地群落密度和地上生物量都相对较大(表2 ), 邻体干扰较强.虽然分布于东坡和西坡的甘肃臭草斑块已经形成, 但斑块内聚集程度不高, 而且斑块面积较小, 与西北针茅共同成为草地群落的优势种群, 在种间竞争中没有绝对的优势地位.作为一种禾本科多年生密集型根茎克隆植物, 甘肃臭草通过根状茎的生长实现大量营养繁殖, 增大了地下生物量的分配比例, 导致东坡和西坡的根冠比显著大于北坡和南坡(表1 ), 有助于提升种间竞争地位.此时, 为了增强种间聚集程度、扩大分布区域, 甘肃臭草选择了既有利于提高地上株丛和地下根系的分布密度, 增强种群空间分布聚集强度和斑块稳固性, 又有助于提升根系空间拓展能力, 逐步扩大种群空间生态位的根系分叉数和连接长度优化配置模式, 导致生长在东坡和西坡的甘肃臭草根系分叉数和连接长度间呈显著负相关关系(p < 0.05, 图2 ), 二者大小介于北坡和南坡之间, 体现了植物根系对高寒草地生境异质性的环境适应性特征. ...
Natural genetic variation of root system architecture from Arabidopsis to Brachypodium: Towards adaptive value.
1
2012
... 近年来, 多数学者对植物根系的研究主要集中在异质生境中根系功能性状可塑性(Grossman & Rice, 2012 ; Pacheco-Villalobos & Hardtk, 2012 )、不同群落环境下根系构型的适应性调整(Rogers & Benfey, 2015 )、根系生长与土壤环境的关系(Forde, 2014 ; Szoboszlay et al ., 2015 )、根系结构和功能(Hodge et al ., 2009 )等方面, 关于退化草地中植物构件生物量分配模式(周兵等, 2015 )、影响根系构型的主要功能性状(周艳松和王立群, 2011 ), 以及不同入侵阶段植物功能性状表型可塑性(Hulme, 2008 ; Pyšek et al ., 2009 )等方面也进行了系统研究, 而从根系构型构建策略的角度出发, 对生境梯度植物根系功能性状间资源权衡机制的研究仍显不足.甘肃臭草(Melica przewalskyi )是禾本科臭草属多年生根茎植物, 通过根状茎的快速横向生长, 长出幼芽和根系, 形成新植株进行无性繁殖, 在地下建立庞大的根茎系统, 形成密集的圆形株丛, 其分株间的间隔子短, 分枝角度小, 能够在竞争过程中占据优势地位.近年来, 在祁连山北坡中山区荒漠草原与典型草原过渡带, 甘肃臭草替代了原生地带性植被的优势种西北针茅(Stipa sareptana var. krylovii ), 形成大量面积不等、繁茂生长的单一优势种群落斑块(赵成章和龙瑞军, 2008 ), 表现出较高的形态可塑性和对逆境的适应能力, 其强大的入侵能力使众多学者对其资源利用策略和生长扩散过程产生了兴趣.目前, 对甘肃臭草型退化草地优势物种空间格局分析及其对水分的响应(赵成章等, 2011 ; 高福元和赵成章, 2012 )、甘肃臭草斑块土壤水分空间异质性(杨泉等, 2014 ), 以及甘肃臭草茎-叶性状、叶内性状(党晶晶等, 2014 ,2015 )等方面进行了研究, 而生境梯度上甘肃臭草根系构型的可塑性变化尚未见报道.鉴于此, 本文在野外调查的基础上, 试图弄清以下问题: (1)不同坡向甘肃臭草根系分叉数和连接长度的权衡关系如何?(2)导致甘肃臭草根系分叉数和连接长度变化的原因有哪些?旨在探讨祁连山地甘肃臭草种群根系构型构建的资源配置模式, 进一步认识甘肃臭草单一优势种群斑块形成机制和扩散机理. ...
Planting intensity, residence time, and species traits determine invasion success of alien woody species.
1
2009
... 近年来, 多数学者对植物根系的研究主要集中在异质生境中根系功能性状可塑性(Grossman & Rice, 2012 ; Pacheco-Villalobos & Hardtk, 2012 )、不同群落环境下根系构型的适应性调整(Rogers & Benfey, 2015 )、根系生长与土壤环境的关系(Forde, 2014 ; Szoboszlay et al ., 2015 )、根系结构和功能(Hodge et al ., 2009 )等方面, 关于退化草地中植物构件生物量分配模式(周兵等, 2015 )、影响根系构型的主要功能性状(周艳松和王立群, 2011 ), 以及不同入侵阶段植物功能性状表型可塑性(Hulme, 2008 ; Pyšek et al ., 2009 )等方面也进行了系统研究, 而从根系构型构建策略的角度出发, 对生境梯度植物根系功能性状间资源权衡机制的研究仍显不足.甘肃臭草(Melica przewalskyi )是禾本科臭草属多年生根茎植物, 通过根状茎的快速横向生长, 长出幼芽和根系, 形成新植株进行无性繁殖, 在地下建立庞大的根茎系统, 形成密集的圆形株丛, 其分株间的间隔子短, 分枝角度小, 能够在竞争过程中占据优势地位.近年来, 在祁连山北坡中山区荒漠草原与典型草原过渡带, 甘肃臭草替代了原生地带性植被的优势种西北针茅(Stipa sareptana var. krylovii ), 形成大量面积不等、繁茂生长的单一优势种群落斑块(赵成章和龙瑞军, 2008 ), 表现出较高的形态可塑性和对逆境的适应能力, 其强大的入侵能力使众多学者对其资源利用策略和生长扩散过程产生了兴趣.目前, 对甘肃臭草型退化草地优势物种空间格局分析及其对水分的响应(赵成章等, 2011 ; 高福元和赵成章, 2012 )、甘肃臭草斑块土壤水分空间异质性(杨泉等, 2014 ), 以及甘肃臭草茎-叶性状、叶内性状(党晶晶等, 2014 ,2015 )等方面进行了研究, 而生境梯度上甘肃臭草根系构型的可塑性变化尚未见报道.鉴于此, 本文在野外调查的基础上, 试图弄清以下问题: (1)不同坡向甘肃臭草根系分叉数和连接长度的权衡关系如何?(2)导致甘肃臭草根系分叉数和连接长度变化的原因有哪些?旨在探讨祁连山地甘肃臭草种群根系构型构建的资源配置模式, 进一步认识甘肃臭草单一优势种群斑块形成机制和扩散机理. ...
Regulation of plant root system architecture: Implications for crop advancement.
2
2015
... 植物的生态策略和适应性、响应性功能要通过植物内部与外部不同功能性状来实现(Bernard- Verdier et al ., 2012 ), 性状的构建、稳固和性状之间的强弱变化与替补转换则靠植物自身的资源权衡和补偿机制来调控(Fortunel et al ., 2012 ; 陈莹婷和许振柱, 2014 ), 是植物在特定环境下生存策略的重要表现(Mooney et al ., 2010 ).根系构型决定了根系在土壤空间中的位置和资源获取方式, 是植物与资源环境异质性相互适应的结果(Malamy, 2005 ; Rogers & Benfey, 2015 ).根系分叉数和连接长度作为根系构型的重要特征指标, 受土壤空间异质性(Villordon et al ., 2014 )、群落环境(单立山等, 2012 )、遗传特性(Kiswara et al ., 2009 ; Bingham & Wu, 2011 )等诸多因素的影响, 在其生存和发展的过程中, 会根据所处生境的资源条件, 调整、转换或补偿自身功能, 实现根系功能性状间的资源权衡.根系分叉数的变化会影响根系的空间分布属性、营养吸收能力和固定作用; 根系连接长度的大小直接决定了根系在土壤中的空间拓展和营养物质吸收能力(Walk et al ., 2004 ), 二者的权衡关系直接反映了植物根系吸收效率和根系构建过程中所需的碳量(Fitter & Stickland, 1991 ).当根系处于水分状态良好的土层中时, 为了提高水分利用率, 会形成更多的侧根, 增大根分叉数(周艳松和王立群, 2011 ); 在干旱胁迫环境中, 较大的分叉数会引起根系内部对营养物质的竞争(杨小林等, 2008 ), 此时植物倾向于增大连接长度, 以减少根系之间的交叠重复, 占据较大的水分和养分资源空间(郭京衡等, 2014 ), 这是植物为适应不同生境条件而形成的获取资源策略, 以达到“生存、生长、繁殖”之三大目的.地形作为生境条件的一种综合指示, 往往能营造局部小气候(Cantón et al ., 2004 ).坡向作为一种重要的地形要素(Fekedulegn et al ., 2003 ), 可以对局部区域的太阳辐射和降水进行二次分配, 其空间变化是群落组成、物种多样性、生态系统过程和植物功能性状的重要决定因素(de Bello et al ., 2006 ; Bennie et al ., 2008 ; Carletti et al ., 2009 ).在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 在各个水平上影响了植物生长及其对水分利用的有效性(Sardans et al ., 2008 ; Huang et al ., 2015 ), 进而引起植物碳资源分配格局和构型构建策略的改变, 为提高生境适应性和风险规避能力, 植物对连接长度和分叉数等主要根系功能性状进行资源优化配置, 构建与生境相适应的根系构型, 优化根系营养吸收和传输系统.因此, 研究坡向梯度上植物根系构型的生态适应策略, 对于理解植物的环境适应机制有重要意义. ...
... 近年来, 多数学者对植物根系的研究主要集中在异质生境中根系功能性状可塑性(Grossman & Rice, 2012 ; Pacheco-Villalobos & Hardtk, 2012 )、不同群落环境下根系构型的适应性调整(Rogers & Benfey, 2015 )、根系生长与土壤环境的关系(Forde, 2014 ; Szoboszlay et al ., 2015 )、根系结构和功能(Hodge et al ., 2009 )等方面, 关于退化草地中植物构件生物量分配模式(周兵等, 2015 )、影响根系构型的主要功能性状(周艳松和王立群, 2011 ), 以及不同入侵阶段植物功能性状表型可塑性(Hulme, 2008 ; Pyšek et al ., 2009 )等方面也进行了系统研究, 而从根系构型构建策略的角度出发, 对生境梯度植物根系功能性状间资源权衡机制的研究仍显不足.甘肃臭草(Melica przewalskyi )是禾本科臭草属多年生根茎植物, 通过根状茎的快速横向生长, 长出幼芽和根系, 形成新植株进行无性繁殖, 在地下建立庞大的根茎系统, 形成密集的圆形株丛, 其分株间的间隔子短, 分枝角度小, 能够在竞争过程中占据优势地位.近年来, 在祁连山北坡中山区荒漠草原与典型草原过渡带, 甘肃臭草替代了原生地带性植被的优势种西北针茅(Stipa sareptana var. krylovii ), 形成大量面积不等、繁茂生长的单一优势种群落斑块(赵成章和龙瑞军, 2008 ), 表现出较高的形态可塑性和对逆境的适应能力, 其强大的入侵能力使众多学者对其资源利用策略和生长扩散过程产生了兴趣.目前, 对甘肃臭草型退化草地优势物种空间格局分析及其对水分的响应(赵成章等, 2011 ; 高福元和赵成章, 2012 )、甘肃臭草斑块土壤水分空间异质性(杨泉等, 2014 ), 以及甘肃臭草茎-叶性状、叶内性状(党晶晶等, 2014 ,2015 )等方面进行了研究, 而生境梯度上甘肃臭草根系构型的可塑性变化尚未见报道.鉴于此, 本文在野外调查的基础上, 试图弄清以下问题: (1)不同坡向甘肃臭草根系分叉数和连接长度的权衡关系如何?(2)导致甘肃臭草根系分叉数和连接长度变化的原因有哪些?旨在探讨祁连山地甘肃臭草种群根系构型构建的资源配置模式, 进一步认识甘肃臭草单一优势种群斑块形成机制和扩散机理. ...
Changes in soil enzymes related to C and N cycle and in soil C and N content under prolonged warming and drought in a Mediterranean shrubland.
1
2008
... 植物的生态策略和适应性、响应性功能要通过植物内部与外部不同功能性状来实现(Bernard- Verdier et al ., 2012 ), 性状的构建、稳固和性状之间的强弱变化与替补转换则靠植物自身的资源权衡和补偿机制来调控(Fortunel et al ., 2012 ; 陈莹婷和许振柱, 2014 ), 是植物在特定环境下生存策略的重要表现(Mooney et al ., 2010 ).根系构型决定了根系在土壤空间中的位置和资源获取方式, 是植物与资源环境异质性相互适应的结果(Malamy, 2005 ; Rogers & Benfey, 2015 ).根系分叉数和连接长度作为根系构型的重要特征指标, 受土壤空间异质性(Villordon et al ., 2014 )、群落环境(单立山等, 2012 )、遗传特性(Kiswara et al ., 2009 ; Bingham & Wu, 2011 )等诸多因素的影响, 在其生存和发展的过程中, 会根据所处生境的资源条件, 调整、转换或补偿自身功能, 实现根系功能性状间的资源权衡.根系分叉数的变化会影响根系的空间分布属性、营养吸收能力和固定作用; 根系连接长度的大小直接决定了根系在土壤中的空间拓展和营养物质吸收能力(Walk et al ., 2004 ), 二者的权衡关系直接反映了植物根系吸收效率和根系构建过程中所需的碳量(Fitter & Stickland, 1991 ).当根系处于水分状态良好的土层中时, 为了提高水分利用率, 会形成更多的侧根, 增大根分叉数(周艳松和王立群, 2011 ); 在干旱胁迫环境中, 较大的分叉数会引起根系内部对营养物质的竞争(杨小林等, 2008 ), 此时植物倾向于增大连接长度, 以减少根系之间的交叠重复, 占据较大的水分和养分资源空间(郭京衡等, 2014 ), 这是植物为适应不同生境条件而形成的获取资源策略, 以达到“生存、生长、繁殖”之三大目的.地形作为生境条件的一种综合指示, 往往能营造局部小气候(Cantón et al ., 2004 ).坡向作为一种重要的地形要素(Fekedulegn et al ., 2003 ), 可以对局部区域的太阳辐射和降水进行二次分配, 其空间变化是群落组成、物种多样性、生态系统过程和植物功能性状的重要决定因素(de Bello et al ., 2006 ; Bennie et al ., 2008 ; Carletti et al ., 2009 ).在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 在各个水平上影响了植物生长及其对水分利用的有效性(Sardans et al ., 2008 ; Huang et al ., 2015 ), 进而引起植物碳资源分配格局和构型构建策略的改变, 为提高生境适应性和风险规避能力, 植物对连接长度和分叉数等主要根系功能性状进行资源优化配置, 构建与生境相适应的根系构型, 优化根系营养吸收和传输系统.因此, 研究坡向梯度上植物根系构型的生态适应策略, 对于理解植物的环境适应机制有重要意义. ...
Rooting depths, lateral root spreads and below-ground/above-ground allometries of plants in water-limited ecosystems.
1
2002
... 植物根系构型构建策略的差异可以用根系碳投入与土壤水分养分收益的权衡关系来解释(Oppelt et al ., 2001 ).根系分叉数的增加会导致根系重叠加剧, 增加根系内部的资源竞争态势(杨小林等, 2008 ).根系连接长度的变化会影响根系空间拓展能力和利用土壤资源的有效性(Schenk & Jackson, 2002a , 2002b ), 这与植物自身根系碳投入与土壤水分养分的收益权衡相契合, 是提高植物资源获取能力的一个重要策略(杨小林等, 2008 ).相对于北坡, 山地南坡日照时间长、水分蒸发量大(殷秀琴, 2004 ), 生长在南坡的甘肃臭草多呈直径不等的单一优势种群斑块, 占据了草地群落的绝对资源优势(表1 , 表2 ).甘肃臭草采用克隆繁殖模式形成了庞大的互相连通交织的根茎系统, 构建了高度聚集的根系网络(高福元和赵成章, 2012 ), 斑块内部种群聚集程度较高, 斑块中心土壤含水量明显低于斑块边缘的土壤含水量(赵成章等, 2011 ), 体现了甘肃臭草向斑块周围扩张的迫切需求.在地上和地下生存空间拥挤、土壤含水量分布不均衡状态下, 为了避免不必要的种内竞争, 甘肃臭草限制了地下生物量的分配比例, 以减缓营养繁殖速度, 降低种群聚集强度, 此时甘肃臭草选择了增加根系连接长度、减少分叉数的根系构型构建模式.根系分叉数的减小能够降低种群内部根系重叠程度, 避免不必要的种内竞争; 连接长度的增大可以增强根系空间拓展能力, 扩大单位根系所占的土壤体空间, 使根系从斑块中心向土壤水分较好、生长发育空间较大的边缘区域拓殖, 是南坡甘肃臭草单优种群斑块扩张的主要动力, 与赵成章等(2011) 关于甘肃臭草通过营养繁殖以类似“同心圆”的方式不断由斑块中心向边缘扩大种群领地的观点一致.这符合相关学者“当植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 增加根系连接长度等策略, 以寻求更多的水分和养分, 保证自身的生存和生长”的结论(单立山等, 2013 ; 杜建会等, 2014 ).因此, 南坡甘肃臭草根系分叉数和连接长度间呈极显著负相关关系(p < 0.01, 图2 ), 减小分叉数、增加连接长度是甘肃臭草斑块扩张的根系构型构建策略, 降低了甘肃臭草单一优势种群斑块内部的根系拥挤程度. ...
The global biogeography of roots.
1
2002
... 植物根系构型构建策略的差异可以用根系碳投入与土壤水分养分收益的权衡关系来解释(Oppelt et al ., 2001 ).根系分叉数的增加会导致根系重叠加剧, 增加根系内部的资源竞争态势(杨小林等, 2008 ).根系连接长度的变化会影响根系空间拓展能力和利用土壤资源的有效性(Schenk & Jackson, 2002a , 2002b ), 这与植物自身根系碳投入与土壤水分养分的收益权衡相契合, 是提高植物资源获取能力的一个重要策略(杨小林等, 2008 ).相对于北坡, 山地南坡日照时间长、水分蒸发量大(殷秀琴, 2004 ), 生长在南坡的甘肃臭草多呈直径不等的单一优势种群斑块, 占据了草地群落的绝对资源优势(表1 , 表2 ).甘肃臭草采用克隆繁殖模式形成了庞大的互相连通交织的根茎系统, 构建了高度聚集的根系网络(高福元和赵成章, 2012 ), 斑块内部种群聚集程度较高, 斑块中心土壤含水量明显低于斑块边缘的土壤含水量(赵成章等, 2011 ), 体现了甘肃臭草向斑块周围扩张的迫切需求.在地上和地下生存空间拥挤、土壤含水量分布不均衡状态下, 为了避免不必要的种内竞争, 甘肃臭草限制了地下生物量的分配比例, 以减缓营养繁殖速度, 降低种群聚集强度, 此时甘肃臭草选择了增加根系连接长度、减少分叉数的根系构型构建模式.根系分叉数的减小能够降低种群内部根系重叠程度, 避免不必要的种内竞争; 连接长度的增大可以增强根系空间拓展能力, 扩大单位根系所占的土壤体空间, 使根系从斑块中心向土壤水分较好、生长发育空间较大的边缘区域拓殖, 是南坡甘肃臭草单优种群斑块扩张的主要动力, 与赵成章等(2011) 关于甘肃臭草通过营养繁殖以类似“同心圆”的方式不断由斑块中心向边缘扩大种群领地的观点一致.这符合相关学者“当植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 增加根系连接长度等策略, 以寻求更多的水分和养分, 保证自身的生存和生长”的结论(单立山等, 2013 ; 杜建会等, 2014 ).因此, 南坡甘肃臭草根系分叉数和连接长度间呈极显著负相关关系(p < 0.01, 图2 ), 减小分叉数、增加连接长度是甘肃臭草斑块扩张的根系构型构建策略, 降低了甘肃臭草单一优势种群斑块内部的根系拥挤程度. ...
红砂根系构型对干旱的生态适应
1
2012
... 植物的生态策略和适应性、响应性功能要通过植物内部与外部不同功能性状来实现(Bernard- Verdier et al ., 2012 ), 性状的构建、稳固和性状之间的强弱变化与替补转换则靠植物自身的资源权衡和补偿机制来调控(Fortunel et al ., 2012 ; 陈莹婷和许振柱, 2014 ), 是植物在特定环境下生存策略的重要表现(Mooney et al ., 2010 ).根系构型决定了根系在土壤空间中的位置和资源获取方式, 是植物与资源环境异质性相互适应的结果(Malamy, 2005 ; Rogers & Benfey, 2015 ).根系分叉数和连接长度作为根系构型的重要特征指标, 受土壤空间异质性(Villordon et al ., 2014 )、群落环境(单立山等, 2012 )、遗传特性(Kiswara et al ., 2009 ; Bingham & Wu, 2011 )等诸多因素的影响, 在其生存和发展的过程中, 会根据所处生境的资源条件, 调整、转换或补偿自身功能, 实现根系功能性状间的资源权衡.根系分叉数的变化会影响根系的空间分布属性、营养吸收能力和固定作用; 根系连接长度的大小直接决定了根系在土壤中的空间拓展和营养物质吸收能力(Walk et al ., 2004 ), 二者的权衡关系直接反映了植物根系吸收效率和根系构建过程中所需的碳量(Fitter & Stickland, 1991 ).当根系处于水分状态良好的土层中时, 为了提高水分利用率, 会形成更多的侧根, 增大根分叉数(周艳松和王立群, 2011 ); 在干旱胁迫环境中, 较大的分叉数会引起根系内部对营养物质的竞争(杨小林等, 2008 ), 此时植物倾向于增大连接长度, 以减少根系之间的交叠重复, 占据较大的水分和养分资源空间(郭京衡等, 2014 ), 这是植物为适应不同生境条件而形成的获取资源策略, 以达到“生存、生长、繁殖”之三大目的.地形作为生境条件的一种综合指示, 往往能营造局部小气候(Cantón et al ., 2004 ).坡向作为一种重要的地形要素(Fekedulegn et al ., 2003 ), 可以对局部区域的太阳辐射和降水进行二次分配, 其空间变化是群落组成、物种多样性、生态系统过程和植物功能性状的重要决定因素(de Bello et al ., 2006 ; Bennie et al ., 2008 ; Carletti et al ., 2009 ).在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 在各个水平上影响了植物生长及其对水分利用的有效性(Sardans et al ., 2008 ; Huang et al ., 2015 ), 进而引起植物碳资源分配格局和构型构建策略的改变, 为提高生境适应性和风险规避能力, 植物对连接长度和分叉数等主要根系功能性状进行资源优化配置, 构建与生境相适应的根系构型, 优化根系营养吸收和传输系统.因此, 研究坡向梯度上植物根系构型的生态适应策略, 对于理解植物的环境适应机制有重要意义. ...
红砂根系构型对干旱的生态适应
1
2012
... 植物的生态策略和适应性、响应性功能要通过植物内部与外部不同功能性状来实现(Bernard- Verdier et al ., 2012 ), 性状的构建、稳固和性状之间的强弱变化与替补转换则靠植物自身的资源权衡和补偿机制来调控(Fortunel et al ., 2012 ; 陈莹婷和许振柱, 2014 ), 是植物在特定环境下生存策略的重要表现(Mooney et al ., 2010 ).根系构型决定了根系在土壤空间中的位置和资源获取方式, 是植物与资源环境异质性相互适应的结果(Malamy, 2005 ; Rogers & Benfey, 2015 ).根系分叉数和连接长度作为根系构型的重要特征指标, 受土壤空间异质性(Villordon et al ., 2014 )、群落环境(单立山等, 2012 )、遗传特性(Kiswara et al ., 2009 ; Bingham & Wu, 2011 )等诸多因素的影响, 在其生存和发展的过程中, 会根据所处生境的资源条件, 调整、转换或补偿自身功能, 实现根系功能性状间的资源权衡.根系分叉数的变化会影响根系的空间分布属性、营养吸收能力和固定作用; 根系连接长度的大小直接决定了根系在土壤中的空间拓展和营养物质吸收能力(Walk et al ., 2004 ), 二者的权衡关系直接反映了植物根系吸收效率和根系构建过程中所需的碳量(Fitter & Stickland, 1991 ).当根系处于水分状态良好的土层中时, 为了提高水分利用率, 会形成更多的侧根, 增大根分叉数(周艳松和王立群, 2011 ); 在干旱胁迫环境中, 较大的分叉数会引起根系内部对营养物质的竞争(杨小林等, 2008 ), 此时植物倾向于增大连接长度, 以减少根系之间的交叠重复, 占据较大的水分和养分资源空间(郭京衡等, 2014 ), 这是植物为适应不同生境条件而形成的获取资源策略, 以达到“生存、生长、繁殖”之三大目的.地形作为生境条件的一种综合指示, 往往能营造局部小气候(Cantón et al ., 2004 ).坡向作为一种重要的地形要素(Fekedulegn et al ., 2003 ), 可以对局部区域的太阳辐射和降水进行二次分配, 其空间变化是群落组成、物种多样性、生态系统过程和植物功能性状的重要决定因素(de Bello et al ., 2006 ; Bennie et al ., 2008 ; Carletti et al ., 2009 ).在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 在各个水平上影响了植物生长及其对水分利用的有效性(Sardans et al ., 2008 ; Huang et al ., 2015 ), 进而引起植物碳资源分配格局和构型构建策略的改变, 为提高生境适应性和风险规避能力, 植物对连接长度和分叉数等主要根系功能性状进行资源优化配置, 构建与生境相适应的根系构型, 优化根系营养吸收和传输系统.因此, 研究坡向梯度上植物根系构型的生态适应策略, 对于理解植物的环境适应机制有重要意义. ...
河西走廊中部两种荒漠植物根系构型特征
1
2013
... 植物根系构型构建策略的差异可以用根系碳投入与土壤水分养分收益的权衡关系来解释(Oppelt et al ., 2001 ).根系分叉数的增加会导致根系重叠加剧, 增加根系内部的资源竞争态势(杨小林等, 2008 ).根系连接长度的变化会影响根系空间拓展能力和利用土壤资源的有效性(Schenk & Jackson, 2002a , 2002b ), 这与植物自身根系碳投入与土壤水分养分的收益权衡相契合, 是提高植物资源获取能力的一个重要策略(杨小林等, 2008 ).相对于北坡, 山地南坡日照时间长、水分蒸发量大(殷秀琴, 2004 ), 生长在南坡的甘肃臭草多呈直径不等的单一优势种群斑块, 占据了草地群落的绝对资源优势(表1 , 表2 ).甘肃臭草采用克隆繁殖模式形成了庞大的互相连通交织的根茎系统, 构建了高度聚集的根系网络(高福元和赵成章, 2012 ), 斑块内部种群聚集程度较高, 斑块中心土壤含水量明显低于斑块边缘的土壤含水量(赵成章等, 2011 ), 体现了甘肃臭草向斑块周围扩张的迫切需求.在地上和地下生存空间拥挤、土壤含水量分布不均衡状态下, 为了避免不必要的种内竞争, 甘肃臭草限制了地下生物量的分配比例, 以减缓营养繁殖速度, 降低种群聚集强度, 此时甘肃臭草选择了增加根系连接长度、减少分叉数的根系构型构建模式.根系分叉数的减小能够降低种群内部根系重叠程度, 避免不必要的种内竞争; 连接长度的增大可以增强根系空间拓展能力, 扩大单位根系所占的土壤体空间, 使根系从斑块中心向土壤水分较好、生长发育空间较大的边缘区域拓殖, 是南坡甘肃臭草单优种群斑块扩张的主要动力, 与赵成章等(2011) 关于甘肃臭草通过营养繁殖以类似“同心圆”的方式不断由斑块中心向边缘扩大种群领地的观点一致.这符合相关学者“当植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 增加根系连接长度等策略, 以寻求更多的水分和养分, 保证自身的生存和生长”的结论(单立山等, 2013 ; 杜建会等, 2014 ).因此, 南坡甘肃臭草根系分叉数和连接长度间呈极显著负相关关系(p < 0.01, 图2 ), 减小分叉数、增加连接长度是甘肃臭草斑块扩张的根系构型构建策略, 降低了甘肃臭草单一优势种群斑块内部的根系拥挤程度. ...
河西走廊中部两种荒漠植物根系构型特征
1
2013
... 植物根系构型构建策略的差异可以用根系碳投入与土壤水分养分收益的权衡关系来解释(Oppelt et al ., 2001 ).根系分叉数的增加会导致根系重叠加剧, 增加根系内部的资源竞争态势(杨小林等, 2008 ).根系连接长度的变化会影响根系空间拓展能力和利用土壤资源的有效性(Schenk & Jackson, 2002a , 2002b ), 这与植物自身根系碳投入与土壤水分养分的收益权衡相契合, 是提高植物资源获取能力的一个重要策略(杨小林等, 2008 ).相对于北坡, 山地南坡日照时间长、水分蒸发量大(殷秀琴, 2004 ), 生长在南坡的甘肃臭草多呈直径不等的单一优势种群斑块, 占据了草地群落的绝对资源优势(表1 , 表2 ).甘肃臭草采用克隆繁殖模式形成了庞大的互相连通交织的根茎系统, 构建了高度聚集的根系网络(高福元和赵成章, 2012 ), 斑块内部种群聚集程度较高, 斑块中心土壤含水量明显低于斑块边缘的土壤含水量(赵成章等, 2011 ), 体现了甘肃臭草向斑块周围扩张的迫切需求.在地上和地下生存空间拥挤、土壤含水量分布不均衡状态下, 为了避免不必要的种内竞争, 甘肃臭草限制了地下生物量的分配比例, 以减缓营养繁殖速度, 降低种群聚集强度, 此时甘肃臭草选择了增加根系连接长度、减少分叉数的根系构型构建模式.根系分叉数的减小能够降低种群内部根系重叠程度, 避免不必要的种内竞争; 连接长度的增大可以增强根系空间拓展能力, 扩大单位根系所占的土壤体空间, 使根系从斑块中心向土壤水分较好、生长发育空间较大的边缘区域拓殖, 是南坡甘肃臭草单优种群斑块扩张的主要动力, 与赵成章等(2011) 关于甘肃臭草通过营养繁殖以类似“同心圆”的方式不断由斑块中心向边缘扩大种群领地的观点一致.这符合相关学者“当植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 增加根系连接长度等策略, 以寻求更多的水分和养分, 保证自身的生存和生长”的结论(单立山等, 2013 ; 杜建会等, 2014 ).因此, 南坡甘肃臭草根系分叉数和连接长度间呈极显著负相关关系(p < 0.01, 图2 ), 减小分叉数、增加连接长度是甘肃臭草斑块扩张的根系构型构建策略, 降低了甘肃臭草单一优势种群斑块内部的根系拥挤程度. ...
Fundamental trade-offs generating the worldwide leaf economics spectrum.
1
2006
... 植物在不同功能和性状之间对有限资源的分配存在着互相牵制的权衡作用(Shipley et al ., 2006 ), 这就需要植物在生长发育过程中, 通过不断优化自身的资源分配来提高适合度以适应环境的变化(Fabbro & Körner, 2004).根系作为植物地上部分与土壤的连接器官, 在较大程度上影响着植物的水分和养分吸收、运移和存储功能(Dannowski & Block, 2005 ; Kong et al ., 2014 ), 根系分叉数和连接长度能够反映根系不同部位构建过程中的碳消耗特点, 影响根系拓展空间、获取营养的代价与效率, 体现了植物性状和构型构建对生境变化的响应, 从中可以理解和阐释植物的物种分布、种间关系、权衡及其作用机制(Westoby & Wright, 2006 ).本研究发现, 随着坡向由北坡向东坡、西坡、南坡转变, 甘肃臭草根系分叉数呈逐渐减小的趋势, 而根系连接长度呈相反的变化趋势(图1 ), 即甘肃臭草分配给根系分叉数和连接长度的资源间存在着“此消彼长”的权衡关系(图2 ).经Pearson相关性分析得出, 群落密度与根系分叉数、连接长度呈极显著相关关系(p < 0.01), 群落高度、土壤含水量与二者显著相关(p < 0.05), 地上生物量与根系分叉数显著相关(p < 0.05, 表3 ), 表明坡向梯度上甘肃臭草根系分叉数和连接长度权衡关系的差异性, 是植物根系构建的资源配置模式对高寒草地生境异质性适应的结果. ...
Comparison of root system architecture and rhizosphere microbial communities of Balsas teosinte and domesticated corn cultivars.
1
2015
... 近年来, 多数学者对植物根系的研究主要集中在异质生境中根系功能性状可塑性(Grossman & Rice, 2012 ; Pacheco-Villalobos & Hardtk, 2012 )、不同群落环境下根系构型的适应性调整(Rogers & Benfey, 2015 )、根系生长与土壤环境的关系(Forde, 2014 ; Szoboszlay et al ., 2015 )、根系结构和功能(Hodge et al ., 2009 )等方面, 关于退化草地中植物构件生物量分配模式(周兵等, 2015 )、影响根系构型的主要功能性状(周艳松和王立群, 2011 ), 以及不同入侵阶段植物功能性状表型可塑性(Hulme, 2008 ; Pyšek et al ., 2009 )等方面也进行了系统研究, 而从根系构型构建策略的角度出发, 对生境梯度植物根系功能性状间资源权衡机制的研究仍显不足.甘肃臭草(Melica przewalskyi )是禾本科臭草属多年生根茎植物, 通过根状茎的快速横向生长, 长出幼芽和根系, 形成新植株进行无性繁殖, 在地下建立庞大的根茎系统, 形成密集的圆形株丛, 其分株间的间隔子短, 分枝角度小, 能够在竞争过程中占据优势地位.近年来, 在祁连山北坡中山区荒漠草原与典型草原过渡带, 甘肃臭草替代了原生地带性植被的优势种西北针茅(Stipa sareptana var. krylovii ), 形成大量面积不等、繁茂生长的单一优势种群落斑块(赵成章和龙瑞军, 2008 ), 表现出较高的形态可塑性和对逆境的适应能力, 其强大的入侵能力使众多学者对其资源利用策略和生长扩散过程产生了兴趣.目前, 对甘肃臭草型退化草地优势物种空间格局分析及其对水分的响应(赵成章等, 2011 ; 高福元和赵成章, 2012 )、甘肃臭草斑块土壤水分空间异质性(杨泉等, 2014 ), 以及甘肃臭草茎-叶性状、叶内性状(党晶晶等, 2014 ,2015 )等方面进行了研究, 而生境梯度上甘肃臭草根系构型的可塑性变化尚未见报道.鉴于此, 本文在野外调查的基础上, 试图弄清以下问题: (1)不同坡向甘肃臭草根系分叉数和连接长度的权衡关系如何?(2)导致甘肃臭草根系分叉数和连接长度变化的原因有哪些?旨在探讨祁连山地甘肃臭草种群根系构型构建的资源配置模式, 进一步认识甘肃臭草单一优势种群斑块形成机制和扩散机理. ...
1
2010
... 利用ArcGIS对研究区1:5万地形图进行数字化, 得到研究区域10 m分辨率的数字高程模型(digital elevation model, DEM).在DEM基础上利用Spatial Analyst模块中的Surface Analysis命令提取坡向, 它表示地表面上一点的切平面的法线矢量n 在水平面的投影nxoy 与过该点的正北方向的夹角, 其中坡向组别以国际划分法确定(陈瑶等, 2006 ), 具体方法参见汤国安等(2010) 文献. ...
1
2010
... 利用ArcGIS对研究区1:5万地形图进行数字化, 得到研究区域10 m分辨率的数字高程模型(digital elevation model, DEM).在DEM基础上利用Spatial Analyst模块中的Surface Analysis命令提取坡向, 它表示地表面上一点的切平面的法线矢量n 在水平面的投影nxoy 与过该点的正北方向的夹角, 其中坡向组别以国际划分法确定(陈瑶等, 2006 ), 具体方法参见汤国安等(2010) 文献. ...
Root architecture and root and tuber crop productivity.
1
2014
... 植物的生态策略和适应性、响应性功能要通过植物内部与外部不同功能性状来实现(Bernard- Verdier et al ., 2012 ), 性状的构建、稳固和性状之间的强弱变化与替补转换则靠植物自身的资源权衡和补偿机制来调控(Fortunel et al ., 2012 ; 陈莹婷和许振柱, 2014 ), 是植物在特定环境下生存策略的重要表现(Mooney et al ., 2010 ).根系构型决定了根系在土壤空间中的位置和资源获取方式, 是植物与资源环境异质性相互适应的结果(Malamy, 2005 ; Rogers & Benfey, 2015 ).根系分叉数和连接长度作为根系构型的重要特征指标, 受土壤空间异质性(Villordon et al ., 2014 )、群落环境(单立山等, 2012 )、遗传特性(Kiswara et al ., 2009 ; Bingham & Wu, 2011 )等诸多因素的影响, 在其生存和发展的过程中, 会根据所处生境的资源条件, 调整、转换或补偿自身功能, 实现根系功能性状间的资源权衡.根系分叉数的变化会影响根系的空间分布属性、营养吸收能力和固定作用; 根系连接长度的大小直接决定了根系在土壤中的空间拓展和营养物质吸收能力(Walk et al ., 2004 ), 二者的权衡关系直接反映了植物根系吸收效率和根系构建过程中所需的碳量(Fitter & Stickland, 1991 ).当根系处于水分状态良好的土层中时, 为了提高水分利用率, 会形成更多的侧根, 增大根分叉数(周艳松和王立群, 2011 ); 在干旱胁迫环境中, 较大的分叉数会引起根系内部对营养物质的竞争(杨小林等, 2008 ), 此时植物倾向于增大连接长度, 以减少根系之间的交叠重复, 占据较大的水分和养分资源空间(郭京衡等, 2014 ), 这是植物为适应不同生境条件而形成的获取资源策略, 以达到“生存、生长、繁殖”之三大目的.地形作为生境条件的一种综合指示, 往往能营造局部小气候(Cantón et al ., 2004 ).坡向作为一种重要的地形要素(Fekedulegn et al ., 2003 ), 可以对局部区域的太阳辐射和降水进行二次分配, 其空间变化是群落组成、物种多样性、生态系统过程和植物功能性状的重要决定因素(de Bello et al ., 2006 ; Bennie et al ., 2008 ; Carletti et al ., 2009 ).在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 在各个水平上影响了植物生长及其对水分利用的有效性(Sardans et al ., 2008 ; Huang et al ., 2015 ), 进而引起植物碳资源分配格局和构型构建策略的改变, 为提高生境适应性和风险规避能力, 植物对连接长度和分叉数等主要根系功能性状进行资源优化配置, 构建与生境相适应的根系构型, 优化根系营养吸收和传输系统.因此, 研究坡向梯度上植物根系构型的生态适应策略, 对于理解植物的环境适应机制有重要意义. ...
Modelling applicability of fractal analysis to efficiency of soil exploration by roots.
1
2004
... 植物的生态策略和适应性、响应性功能要通过植物内部与外部不同功能性状来实现(Bernard- Verdier et al ., 2012 ), 性状的构建、稳固和性状之间的强弱变化与替补转换则靠植物自身的资源权衡和补偿机制来调控(Fortunel et al ., 2012 ; 陈莹婷和许振柱, 2014 ), 是植物在特定环境下生存策略的重要表现(Mooney et al ., 2010 ).根系构型决定了根系在土壤空间中的位置和资源获取方式, 是植物与资源环境异质性相互适应的结果(Malamy, 2005 ; Rogers & Benfey, 2015 ).根系分叉数和连接长度作为根系构型的重要特征指标, 受土壤空间异质性(Villordon et al ., 2014 )、群落环境(单立山等, 2012 )、遗传特性(Kiswara et al ., 2009 ; Bingham & Wu, 2011 )等诸多因素的影响, 在其生存和发展的过程中, 会根据所处生境的资源条件, 调整、转换或补偿自身功能, 实现根系功能性状间的资源权衡.根系分叉数的变化会影响根系的空间分布属性、营养吸收能力和固定作用; 根系连接长度的大小直接决定了根系在土壤中的空间拓展和营养物质吸收能力(Walk et al ., 2004 ), 二者的权衡关系直接反映了植物根系吸收效率和根系构建过程中所需的碳量(Fitter & Stickland, 1991 ).当根系处于水分状态良好的土层中时, 为了提高水分利用率, 会形成更多的侧根, 增大根分叉数(周艳松和王立群, 2011 ); 在干旱胁迫环境中, 较大的分叉数会引起根系内部对营养物质的竞争(杨小林等, 2008 ), 此时植物倾向于增大连接长度, 以减少根系之间的交叠重复, 占据较大的水分和养分资源空间(郭京衡等, 2014 ), 这是植物为适应不同生境条件而形成的获取资源策略, 以达到“生存、生长、繁殖”之三大目的.地形作为生境条件的一种综合指示, 往往能营造局部小气候(Cantón et al ., 2004 ).坡向作为一种重要的地形要素(Fekedulegn et al ., 2003 ), 可以对局部区域的太阳辐射和降水进行二次分配, 其空间变化是群落组成、物种多样性、生态系统过程和植物功能性状的重要决定因素(de Bello et al ., 2006 ; Bennie et al ., 2008 ; Carletti et al ., 2009 ).在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 在各个水平上影响了植物生长及其对水分利用的有效性(Sardans et al ., 2008 ; Huang et al ., 2015 ), 进而引起植物碳资源分配格局和构型构建策略的改变, 为提高生境适应性和风险规避能力, 植物对连接长度和分叉数等主要根系功能性状进行资源优化配置, 构建与生境相适应的根系构型, 优化根系营养吸收和传输系统.因此, 研究坡向梯度上植物根系构型的生态适应策略, 对于理解植物的环境适应机制有重要意义. ...
Land-plant ecology on the basis of functional traits.
1
2006
... 植物在不同功能和性状之间对有限资源的分配存在着互相牵制的权衡作用(Shipley et al ., 2006 ), 这就需要植物在生长发育过程中, 通过不断优化自身的资源分配来提高适合度以适应环境的变化(Fabbro & Körner, 2004).根系作为植物地上部分与土壤的连接器官, 在较大程度上影响着植物的水分和养分吸收、运移和存储功能(Dannowski & Block, 2005 ; Kong et al ., 2014 ), 根系分叉数和连接长度能够反映根系不同部位构建过程中的碳消耗特点, 影响根系拓展空间、获取营养的代价与效率, 体现了植物性状和构型构建对生境变化的响应, 从中可以理解和阐释植物的物种分布、种间关系、权衡及其作用机制(Westoby & Wright, 2006 ).本研究发现, 随着坡向由北坡向东坡、西坡、南坡转变, 甘肃臭草根系分叉数呈逐渐减小的趋势, 而根系连接长度呈相反的变化趋势(图1 ), 即甘肃臭草分配给根系分叉数和连接长度的资源间存在着“此消彼长”的权衡关系(图2 ).经Pearson相关性分析得出, 群落密度与根系分叉数、连接长度呈极显著相关关系(p < 0.01), 群落高度、土壤含水量与二者显著相关(p < 0.05), 地上生物量与根系分叉数显著相关(p < 0.05, 表3 ), 表明坡向梯度上甘肃臭草根系分叉数和连接长度权衡关系的差异性, 是植物根系构建的资源配置模式对高寒草地生境异质性适应的结果. ...
祁连山地甘肃臭草斑块土壤水分的空间自相关分析
1
2014
... 近年来, 多数学者对植物根系的研究主要集中在异质生境中根系功能性状可塑性(Grossman & Rice, 2012 ; Pacheco-Villalobos & Hardtk, 2012 )、不同群落环境下根系构型的适应性调整(Rogers & Benfey, 2015 )、根系生长与土壤环境的关系(Forde, 2014 ; Szoboszlay et al ., 2015 )、根系结构和功能(Hodge et al ., 2009 )等方面, 关于退化草地中植物构件生物量分配模式(周兵等, 2015 )、影响根系构型的主要功能性状(周艳松和王立群, 2011 ), 以及不同入侵阶段植物功能性状表型可塑性(Hulme, 2008 ; Pyšek et al ., 2009 )等方面也进行了系统研究, 而从根系构型构建策略的角度出发, 对生境梯度植物根系功能性状间资源权衡机制的研究仍显不足.甘肃臭草(Melica przewalskyi )是禾本科臭草属多年生根茎植物, 通过根状茎的快速横向生长, 长出幼芽和根系, 形成新植株进行无性繁殖, 在地下建立庞大的根茎系统, 形成密集的圆形株丛, 其分株间的间隔子短, 分枝角度小, 能够在竞争过程中占据优势地位.近年来, 在祁连山北坡中山区荒漠草原与典型草原过渡带, 甘肃臭草替代了原生地带性植被的优势种西北针茅(Stipa sareptana var. krylovii ), 形成大量面积不等、繁茂生长的单一优势种群落斑块(赵成章和龙瑞军, 2008 ), 表现出较高的形态可塑性和对逆境的适应能力, 其强大的入侵能力使众多学者对其资源利用策略和生长扩散过程产生了兴趣.目前, 对甘肃臭草型退化草地优势物种空间格局分析及其对水分的响应(赵成章等, 2011 ; 高福元和赵成章, 2012 )、甘肃臭草斑块土壤水分空间异质性(杨泉等, 2014 ), 以及甘肃臭草茎-叶性状、叶内性状(党晶晶等, 2014 ,2015 )等方面进行了研究, 而生境梯度上甘肃臭草根系构型的可塑性变化尚未见报道.鉴于此, 本文在野外调查的基础上, 试图弄清以下问题: (1)不同坡向甘肃臭草根系分叉数和连接长度的权衡关系如何?(2)导致甘肃臭草根系分叉数和连接长度变化的原因有哪些?旨在探讨祁连山地甘肃臭草种群根系构型构建的资源配置模式, 进一步认识甘肃臭草单一优势种群斑块形成机制和扩散机理. ...
祁连山地甘肃臭草斑块土壤水分的空间自相关分析
1
2014
... 近年来, 多数学者对植物根系的研究主要集中在异质生境中根系功能性状可塑性(Grossman & Rice, 2012 ; Pacheco-Villalobos & Hardtk, 2012 )、不同群落环境下根系构型的适应性调整(Rogers & Benfey, 2015 )、根系生长与土壤环境的关系(Forde, 2014 ; Szoboszlay et al ., 2015 )、根系结构和功能(Hodge et al ., 2009 )等方面, 关于退化草地中植物构件生物量分配模式(周兵等, 2015 )、影响根系构型的主要功能性状(周艳松和王立群, 2011 ), 以及不同入侵阶段植物功能性状表型可塑性(Hulme, 2008 ; Pyšek et al ., 2009 )等方面也进行了系统研究, 而从根系构型构建策略的角度出发, 对生境梯度植物根系功能性状间资源权衡机制的研究仍显不足.甘肃臭草(Melica przewalskyi )是禾本科臭草属多年生根茎植物, 通过根状茎的快速横向生长, 长出幼芽和根系, 形成新植株进行无性繁殖, 在地下建立庞大的根茎系统, 形成密集的圆形株丛, 其分株间的间隔子短, 分枝角度小, 能够在竞争过程中占据优势地位.近年来, 在祁连山北坡中山区荒漠草原与典型草原过渡带, 甘肃臭草替代了原生地带性植被的优势种西北针茅(Stipa sareptana var. krylovii ), 形成大量面积不等、繁茂生长的单一优势种群落斑块(赵成章和龙瑞军, 2008 ), 表现出较高的形态可塑性和对逆境的适应能力, 其强大的入侵能力使众多学者对其资源利用策略和生长扩散过程产生了兴趣.目前, 对甘肃臭草型退化草地优势物种空间格局分析及其对水分的响应(赵成章等, 2011 ; 高福元和赵成章, 2012 )、甘肃臭草斑块土壤水分空间异质性(杨泉等, 2014 ), 以及甘肃臭草茎-叶性状、叶内性状(党晶晶等, 2014 ,2015 )等方面进行了研究, 而生境梯度上甘肃臭草根系构型的可塑性变化尚未见报道.鉴于此, 本文在野外调查的基础上, 试图弄清以下问题: (1)不同坡向甘肃臭草根系分叉数和连接长度的权衡关系如何?(2)导致甘肃臭草根系分叉数和连接长度变化的原因有哪些?旨在探讨祁连山地甘肃臭草种群根系构型构建的资源配置模式, 进一步认识甘肃臭草单一优势种群斑块形成机制和扩散机理. ...
塔克拉玛干沙漠腹地3种植物根系构型及其生境适应策略
3
2008
... 植物的生态策略和适应性、响应性功能要通过植物内部与外部不同功能性状来实现(Bernard- Verdier et al ., 2012 ), 性状的构建、稳固和性状之间的强弱变化与替补转换则靠植物自身的资源权衡和补偿机制来调控(Fortunel et al ., 2012 ; 陈莹婷和许振柱, 2014 ), 是植物在特定环境下生存策略的重要表现(Mooney et al ., 2010 ).根系构型决定了根系在土壤空间中的位置和资源获取方式, 是植物与资源环境异质性相互适应的结果(Malamy, 2005 ; Rogers & Benfey, 2015 ).根系分叉数和连接长度作为根系构型的重要特征指标, 受土壤空间异质性(Villordon et al ., 2014 )、群落环境(单立山等, 2012 )、遗传特性(Kiswara et al ., 2009 ; Bingham & Wu, 2011 )等诸多因素的影响, 在其生存和发展的过程中, 会根据所处生境的资源条件, 调整、转换或补偿自身功能, 实现根系功能性状间的资源权衡.根系分叉数的变化会影响根系的空间分布属性、营养吸收能力和固定作用; 根系连接长度的大小直接决定了根系在土壤中的空间拓展和营养物质吸收能力(Walk et al ., 2004 ), 二者的权衡关系直接反映了植物根系吸收效率和根系构建过程中所需的碳量(Fitter & Stickland, 1991 ).当根系处于水分状态良好的土层中时, 为了提高水分利用率, 会形成更多的侧根, 增大根分叉数(周艳松和王立群, 2011 ); 在干旱胁迫环境中, 较大的分叉数会引起根系内部对营养物质的竞争(杨小林等, 2008 ), 此时植物倾向于增大连接长度, 以减少根系之间的交叠重复, 占据较大的水分和养分资源空间(郭京衡等, 2014 ), 这是植物为适应不同生境条件而形成的获取资源策略, 以达到“生存、生长、繁殖”之三大目的.地形作为生境条件的一种综合指示, 往往能营造局部小气候(Cantón et al ., 2004 ).坡向作为一种重要的地形要素(Fekedulegn et al ., 2003 ), 可以对局部区域的太阳辐射和降水进行二次分配, 其空间变化是群落组成、物种多样性、生态系统过程和植物功能性状的重要决定因素(de Bello et al ., 2006 ; Bennie et al ., 2008 ; Carletti et al ., 2009 ).在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 在各个水平上影响了植物生长及其对水分利用的有效性(Sardans et al ., 2008 ; Huang et al ., 2015 ), 进而引起植物碳资源分配格局和构型构建策略的改变, 为提高生境适应性和风险规避能力, 植物对连接长度和分叉数等主要根系功能性状进行资源优化配置, 构建与生境相适应的根系构型, 优化根系营养吸收和传输系统.因此, 研究坡向梯度上植物根系构型的生态适应策略, 对于理解植物的环境适应机制有重要意义. ...
... 植物根系构型构建策略的差异可以用根系碳投入与土壤水分养分收益的权衡关系来解释(Oppelt et al ., 2001 ).根系分叉数的增加会导致根系重叠加剧, 增加根系内部的资源竞争态势(杨小林等, 2008 ).根系连接长度的变化会影响根系空间拓展能力和利用土壤资源的有效性(Schenk & Jackson, 2002a , 2002b ), 这与植物自身根系碳投入与土壤水分养分的收益权衡相契合, 是提高植物资源获取能力的一个重要策略(杨小林等, 2008 ).相对于北坡, 山地南坡日照时间长、水分蒸发量大(殷秀琴, 2004 ), 生长在南坡的甘肃臭草多呈直径不等的单一优势种群斑块, 占据了草地群落的绝对资源优势(表1 , 表2 ).甘肃臭草采用克隆繁殖模式形成了庞大的互相连通交织的根茎系统, 构建了高度聚集的根系网络(高福元和赵成章, 2012 ), 斑块内部种群聚集程度较高, 斑块中心土壤含水量明显低于斑块边缘的土壤含水量(赵成章等, 2011 ), 体现了甘肃臭草向斑块周围扩张的迫切需求.在地上和地下生存空间拥挤、土壤含水量分布不均衡状态下, 为了避免不必要的种内竞争, 甘肃臭草限制了地下生物量的分配比例, 以减缓营养繁殖速度, 降低种群聚集强度, 此时甘肃臭草选择了增加根系连接长度、减少分叉数的根系构型构建模式.根系分叉数的减小能够降低种群内部根系重叠程度, 避免不必要的种内竞争; 连接长度的增大可以增强根系空间拓展能力, 扩大单位根系所占的土壤体空间, 使根系从斑块中心向土壤水分较好、生长发育空间较大的边缘区域拓殖, 是南坡甘肃臭草单优种群斑块扩张的主要动力, 与赵成章等(2011) 关于甘肃臭草通过营养繁殖以类似“同心圆”的方式不断由斑块中心向边缘扩大种群领地的观点一致.这符合相关学者“当植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 增加根系连接长度等策略, 以寻求更多的水分和养分, 保证自身的生存和生长”的结论(单立山等, 2013 ; 杜建会等, 2014 ).因此, 南坡甘肃臭草根系分叉数和连接长度间呈极显著负相关关系(p < 0.01, 图2 ), 减小分叉数、增加连接长度是甘肃臭草斑块扩张的根系构型构建策略, 降低了甘肃臭草单一优势种群斑块内部的根系拥挤程度. ...
... ), 这与植物自身根系碳投入与土壤水分养分的收益权衡相契合, 是提高植物资源获取能力的一个重要策略(杨小林等, 2008 ).相对于北坡, 山地南坡日照时间长、水分蒸发量大(殷秀琴, 2004 ), 生长在南坡的甘肃臭草多呈直径不等的单一优势种群斑块, 占据了草地群落的绝对资源优势(表1 , 表2 ).甘肃臭草采用克隆繁殖模式形成了庞大的互相连通交织的根茎系统, 构建了高度聚集的根系网络(高福元和赵成章, 2012 ), 斑块内部种群聚集程度较高, 斑块中心土壤含水量明显低于斑块边缘的土壤含水量(赵成章等, 2011 ), 体现了甘肃臭草向斑块周围扩张的迫切需求.在地上和地下生存空间拥挤、土壤含水量分布不均衡状态下, 为了避免不必要的种内竞争, 甘肃臭草限制了地下生物量的分配比例, 以减缓营养繁殖速度, 降低种群聚集强度, 此时甘肃臭草选择了增加根系连接长度、减少分叉数的根系构型构建模式.根系分叉数的减小能够降低种群内部根系重叠程度, 避免不必要的种内竞争; 连接长度的增大可以增强根系空间拓展能力, 扩大单位根系所占的土壤体空间, 使根系从斑块中心向土壤水分较好、生长发育空间较大的边缘区域拓殖, 是南坡甘肃臭草单优种群斑块扩张的主要动力, 与赵成章等(2011) 关于甘肃臭草通过营养繁殖以类似“同心圆”的方式不断由斑块中心向边缘扩大种群领地的观点一致.这符合相关学者“当植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 增加根系连接长度等策略, 以寻求更多的水分和养分, 保证自身的生存和生长”的结论(单立山等, 2013 ; 杜建会等, 2014 ).因此, 南坡甘肃臭草根系分叉数和连接长度间呈极显著负相关关系(p < 0.01, 图2 ), 减小分叉数、增加连接长度是甘肃臭草斑块扩张的根系构型构建策略, 降低了甘肃臭草单一优势种群斑块内部的根系拥挤程度. ...
塔克拉玛干沙漠腹地3种植物根系构型及其生境适应策略
3
2008
... 植物的生态策略和适应性、响应性功能要通过植物内部与外部不同功能性状来实现(Bernard- Verdier et al ., 2012 ), 性状的构建、稳固和性状之间的强弱变化与替补转换则靠植物自身的资源权衡和补偿机制来调控(Fortunel et al ., 2012 ; 陈莹婷和许振柱, 2014 ), 是植物在特定环境下生存策略的重要表现(Mooney et al ., 2010 ).根系构型决定了根系在土壤空间中的位置和资源获取方式, 是植物与资源环境异质性相互适应的结果(Malamy, 2005 ; Rogers & Benfey, 2015 ).根系分叉数和连接长度作为根系构型的重要特征指标, 受土壤空间异质性(Villordon et al ., 2014 )、群落环境(单立山等, 2012 )、遗传特性(Kiswara et al ., 2009 ; Bingham & Wu, 2011 )等诸多因素的影响, 在其生存和发展的过程中, 会根据所处生境的资源条件, 调整、转换或补偿自身功能, 实现根系功能性状间的资源权衡.根系分叉数的变化会影响根系的空间分布属性、营养吸收能力和固定作用; 根系连接长度的大小直接决定了根系在土壤中的空间拓展和营养物质吸收能力(Walk et al ., 2004 ), 二者的权衡关系直接反映了植物根系吸收效率和根系构建过程中所需的碳量(Fitter & Stickland, 1991 ).当根系处于水分状态良好的土层中时, 为了提高水分利用率, 会形成更多的侧根, 增大根分叉数(周艳松和王立群, 2011 ); 在干旱胁迫环境中, 较大的分叉数会引起根系内部对营养物质的竞争(杨小林等, 2008 ), 此时植物倾向于增大连接长度, 以减少根系之间的交叠重复, 占据较大的水分和养分资源空间(郭京衡等, 2014 ), 这是植物为适应不同生境条件而形成的获取资源策略, 以达到“生存、生长、繁殖”之三大目的.地形作为生境条件的一种综合指示, 往往能营造局部小气候(Cantón et al ., 2004 ).坡向作为一种重要的地形要素(Fekedulegn et al ., 2003 ), 可以对局部区域的太阳辐射和降水进行二次分配, 其空间变化是群落组成、物种多样性、生态系统过程和植物功能性状的重要决定因素(de Bello et al ., 2006 ; Bennie et al ., 2008 ; Carletti et al ., 2009 ).在半干旱山地, 不同坡向的光照强度、大气温度和土壤水分等环境因子的规律性变化所形成的生境梯度, 在各个水平上影响了植物生长及其对水分利用的有效性(Sardans et al ., 2008 ; Huang et al ., 2015 ), 进而引起植物碳资源分配格局和构型构建策略的改变, 为提高生境适应性和风险规避能力, 植物对连接长度和分叉数等主要根系功能性状进行资源优化配置, 构建与生境相适应的根系构型, 优化根系营养吸收和传输系统.因此, 研究坡向梯度上植物根系构型的生态适应策略, 对于理解植物的环境适应机制有重要意义. ...
... 植物根系构型构建策略的差异可以用根系碳投入与土壤水分养分收益的权衡关系来解释(Oppelt et al ., 2001 ).根系分叉数的增加会导致根系重叠加剧, 增加根系内部的资源竞争态势(杨小林等, 2008 ).根系连接长度的变化会影响根系空间拓展能力和利用土壤资源的有效性(Schenk & Jackson, 2002a , 2002b ), 这与植物自身根系碳投入与土壤水分养分的收益权衡相契合, 是提高植物资源获取能力的一个重要策略(杨小林等, 2008 ).相对于北坡, 山地南坡日照时间长、水分蒸发量大(殷秀琴, 2004 ), 生长在南坡的甘肃臭草多呈直径不等的单一优势种群斑块, 占据了草地群落的绝对资源优势(表1 , 表2 ).甘肃臭草采用克隆繁殖模式形成了庞大的互相连通交织的根茎系统, 构建了高度聚集的根系网络(高福元和赵成章, 2012 ), 斑块内部种群聚集程度较高, 斑块中心土壤含水量明显低于斑块边缘的土壤含水量(赵成章等, 2011 ), 体现了甘肃臭草向斑块周围扩张的迫切需求.在地上和地下生存空间拥挤、土壤含水量分布不均衡状态下, 为了避免不必要的种内竞争, 甘肃臭草限制了地下生物量的分配比例, 以减缓营养繁殖速度, 降低种群聚集强度, 此时甘肃臭草选择了增加根系连接长度、减少分叉数的根系构型构建模式.根系分叉数的减小能够降低种群内部根系重叠程度, 避免不必要的种内竞争; 连接长度的增大可以增强根系空间拓展能力, 扩大单位根系所占的土壤体空间, 使根系从斑块中心向土壤水分较好、生长发育空间较大的边缘区域拓殖, 是南坡甘肃臭草单优种群斑块扩张的主要动力, 与赵成章等(2011) 关于甘肃臭草通过营养繁殖以类似“同心圆”的方式不断由斑块中心向边缘扩大种群领地的观点一致.这符合相关学者“当植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 增加根系连接长度等策略, 以寻求更多的水分和养分, 保证自身的生存和生长”的结论(单立山等, 2013 ; 杜建会等, 2014 ).因此, 南坡甘肃臭草根系分叉数和连接长度间呈极显著负相关关系(p < 0.01, 图2 ), 减小分叉数、增加连接长度是甘肃臭草斑块扩张的根系构型构建策略, 降低了甘肃臭草单一优势种群斑块内部的根系拥挤程度. ...
... ), 这与植物自身根系碳投入与土壤水分养分的收益权衡相契合, 是提高植物资源获取能力的一个重要策略(杨小林等, 2008 ).相对于北坡, 山地南坡日照时间长、水分蒸发量大(殷秀琴, 2004 ), 生长在南坡的甘肃臭草多呈直径不等的单一优势种群斑块, 占据了草地群落的绝对资源优势(表1 , 表2 ).甘肃臭草采用克隆繁殖模式形成了庞大的互相连通交织的根茎系统, 构建了高度聚集的根系网络(高福元和赵成章, 2012 ), 斑块内部种群聚集程度较高, 斑块中心土壤含水量明显低于斑块边缘的土壤含水量(赵成章等, 2011 ), 体现了甘肃臭草向斑块周围扩张的迫切需求.在地上和地下生存空间拥挤、土壤含水量分布不均衡状态下, 为了避免不必要的种内竞争, 甘肃臭草限制了地下生物量的分配比例, 以减缓营养繁殖速度, 降低种群聚集强度, 此时甘肃臭草选择了增加根系连接长度、减少分叉数的根系构型构建模式.根系分叉数的减小能够降低种群内部根系重叠程度, 避免不必要的种内竞争; 连接长度的增大可以增强根系空间拓展能力, 扩大单位根系所占的土壤体空间, 使根系从斑块中心向土壤水分较好、生长发育空间较大的边缘区域拓殖, 是南坡甘肃臭草单优种群斑块扩张的主要动力, 与赵成章等(2011) 关于甘肃臭草通过营养繁殖以类似“同心圆”的方式不断由斑块中心向边缘扩大种群领地的观点一致.这符合相关学者“当植物根系处于干旱或贫瘠的土壤环境中时, 会采取降低分支强度, 增加根系连接长度等策略, 以寻求更多的水分和养分, 保证自身的生存和生长”的结论(单立山等, 2013 ; 杜建会等, 2014 ).因此, 南坡甘肃臭草根系分叉数和连接长度间呈极显著负相关关系(p < 0.01, 图2 ), 减小分叉数、增加连接长度是甘肃臭草斑块扩张的根系构型构建策略, 降低了甘肃臭草单一优势种群斑块内部的根系拥挤程度. ...
 , 史元春, 杜晶, 王继伟, 陈静
, 史元春, 杜晶, 王继伟, 陈静


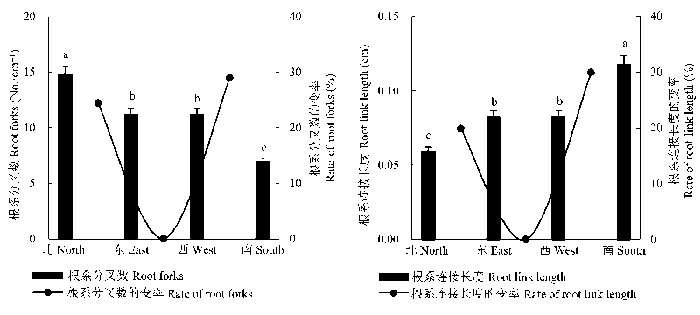
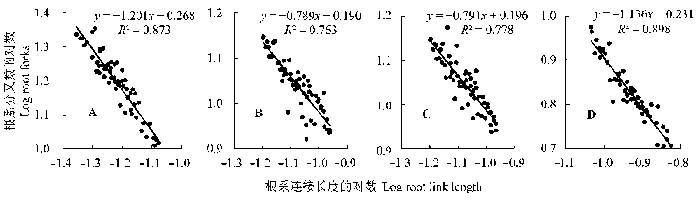
{kind=link}
{kind=link}
{kind=link}
{kind=link}