Comparison of physiological and leaf morphological traits for photosynthesis of the 51 plant species in the Maqu alpine swamp meadow
REN Qing-Ji1, LI Hong-Lin2, *, , BU Hai-Yan2
1Grassland Workstation of Tibetan Autonomous Prefecture of Gannan, Hezuo, Gansu 747000, China2State Key Laboratory of Grassland Agro- ecosystems, School of Life Science, Lanzhou University, Lanzhou 730000, China
Trait-based approaches are often widely used in ecological research to predict or explain the complex ecological processes at both individual and ecosystem levels. Leaf function with morphological and physiological traits can play important roles in plan growth, survival, reproduction in natural environments. The aim of this study is 1) to determine the differences of leaf traits between both the species and the functional groups in a swamp meadow; 2) to explore the relationship between different leaf morphological traits and physiological traits.
Methods
Gas exchanges of 51 species (in 14 families) were measured on six fully expand health leaves (from different individual plants) using a portable photosynthesis system in the field during the peak of growing season. The leaf morphological traits was measured based on 6 single leaves form different individuals, include the net photosynthesis rate (Pn), transpiration rate (Tr), specific leaf area (SLA), relative leaf water content (LWC), leaf area (LA) and the water use efficiency (WUE = Pn/Tr).
Important findings
Result showed that there were significant interspecific differences in the investigated traits as described in above methods. Among the traits, the LWC (coefficient of variation, CV = 0.11) was ranged from 58.12% to 81.83%, 0.0088-0.0278 m2·g-1 for the SLA (CV = 0.27), and 0.51 cm2 to 100.90 cm2 for the LA (CV = 1.73), while the range of 4.25-19.23 μmol CO2·m-2·s-1, 2.89-12.81 mmol H2O·m-2·s-1 and 0.56-3.76 μmol CO2·mmol-1 H2O for Pn (CV = 0.33), Tr (CV = 0.33), and WUE (CV = 0.36), respectively. There were also significant differences between the functional groups (sedge, grass and forbs) for these traits. Forbs have larger LA and higher LWC than sedge and grass; Grass have higher WUE and SLA than those of others; while Sedge have higher Pn. Our result also showed there were high correlation between Pn and SLA, WUE and LWC, indicated the strong impacts of leaf morphology on the gas exchange physiology. The SLA was also related to gas exchange traits both among species and functional groups, while the LWC was only among species and LA among functional groups. In conclusion, significant differences in these functional traits among species suggest that these species could vary in resource use and growth form in their community ecosystem. Also the difference among the functional groups in these traits indicates the resource use of the community would be largely influenced by its species composition, especially the differences of functional groups. The research finding will help to better understanding of the ecosystem function in alpine swamp meadow with related management implication.
Keywords:alpine swamp meadow
;
functional traits
;
leaf morphology
;
gas exchange
;
water use efficiency
;
functional group
RENQing-Ji, LIHong-Lin, BUHai-Yan. Comparison of physiological and leaf morphological traits for photosynthesis of the 51 plant species in the Maqu alpine swamp meadow. Chinese Journal of Plant Ecology, 2015, 39(6): 593-603 https://doi.org/10.17521/cjpe.2015.0057
基于功能性状(functional traits)的研究方法被广泛地应用于生态学和进化生物学的研究中, 功能性状通过影响个体的生存、生长、生殖来间接地影响其适合度, 可以作为预测有机体生长表现的重要指标(Violle et al., 2007)。个体或物种间在功能性状间的差异既反映了其所处的生境特征, 也体现了它们不同的资源利用方式。例如与生长于水分充足的生境的植物相比, 生长于水分不足生境中的植物一般具有较小的比叶面积(specific leaf area, SLA), 较低的净光合速率(net photosynthesis rate, Pn)和蒸腾速率(transpiration rate, Tr), 体现了在控制水分散失和提高生长速率之间的权衡(Reich et al., 1997; Nicotra et al., 2007)。
Table 1 Relative leaf water content (LWC), specific leaf area (SLA), leaf area (LA), net photosynthesis rate (Pn), transpiration rate (Tr) and water use efficiency (WUE) of the 51 species from the alpine meadow swamp in Maqu County, and their coefficient of variation (CV) among the species (mean ± SE)
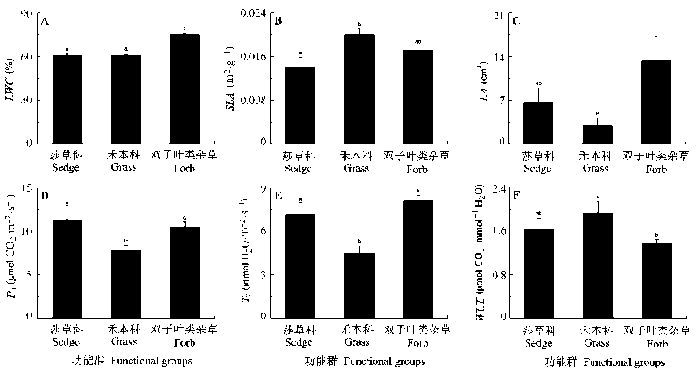
Fig. 1 Comparison of mean valus for relative leaf water content (LWC), specific leaf area (SLA), leaf area (LA), net photosynthesis rate (Pn), transpiration rate (Tr) and water use efficiency (WUE) in the functional groups (mean ± SE). Different small letters indicate significant difference between functional groups for given trait (p < 0.05).
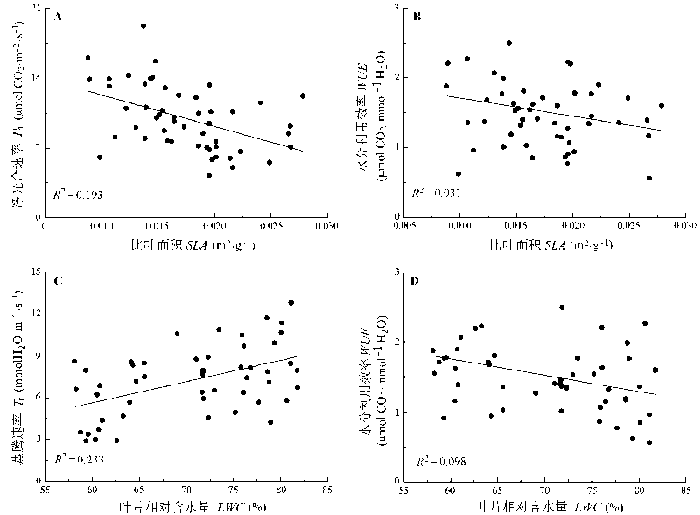
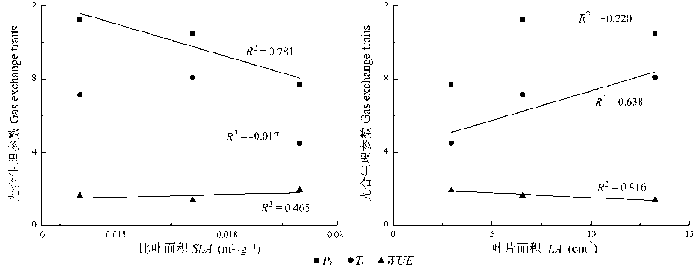
Fig. 2 Results of the regression analysis between the leaf physiological and morphological traits of photosynthesis for the studied species. LWC, Pn, SLA, Tr, WUE, see Fig. 1.
Effect of change of plant community composition along degradation gradients on water conditions in an alpine swamp wetland on the Qinghai- Tibetan Plateau of China.
Chinese Journal of Plant Ecology, 36, 403-410. (in Chinese with English abstract)
Reconciling apparent discrepancies among studies relating life span, structure and function of leaves in contrasting plant forms and climates: ‘The blind men and the elephant retold’.
The effects of plant composition and diversity on ecosystem processes.
1
1997
... 植物功能群的多样性被认为是比物种多样性更重要的生态指标(Walker, 1992; Power et al., 1996; Hooper & Vitousek, 1997; Tilman et al., 1997; Mack & D’Antonio, 1998).有研究认为功能群可以将物种水平的生活史策略和生态系统水平的进程联系起来(Chapin, 1993).根据同一功能群的物种应该具有相似的形态或生理性状(Lauenroth et al., 1978; Pokorny et al., 2005), 我们将玛曲高寒沼泽化草甸的物种划分为3个功能群: 莎草科、禾本科和双子叶类杂草(李伟, 2011).总体上, 不同功能群之间无论在叶片形态特征还是在光合生理特征上都存在显著的差异.莎草科和禾本科植物具有相似且较小的LA和LWC, 但是莎草科植物在Pn和Tr上却显著地高于禾本科植物, 而禾本科植物的SLA显著高于莎草科植物和双子叶类杂草, 最终在叶片水平的WUE指标上, 禾本科和莎草科植物远高于双子叶类杂草.叶片形态和光合生理指标在不同功能群之间的不同显示了不同功能群物种在资源利用上可能的差异, 而这种差异将有助于植物物种对环境资源的充分利用, 进而提高整个系统的稳定性(Naeem et al., 1994; Anderson & Inouye, 2001).由于叶片水平WUE的差异, 如果双子叶类杂草的多度进一步增加, 则可能导致整个群落WUE的降低, 相反, 禾本科和莎草科植物的增加则会提高群落的WUE (Niu et al., 2011). ...
黄河首曲湿地退化过程中植物群落组分及物种多样性的变化
1
2009
... 玛曲高寒湿地位于平均海拔3500 m的青藏高原东缘(Niu et al., 2014), 是黄河上游重要的水源补给区, 具有极其重要的生态和经济地位(张晓云等, 2005).人为和自然原因导致的湿地退化普遍存在于该地区(王根绪等, 2007; Pan et al., 2011).湿地退化导致了在退化梯度上土壤水分减少和植物群落的物种组成改变(后源等, 2009).作为湿地退化演替的过渡性阶段, 沼泽化草甸处在高寒湿地和高寒草甸的中间地带, 其物种组成具有明显的过渡特点(Xiang et al., 2009; Ma et al., 2011), 因此, 研究高寒沼泽化草甸物种在光合和叶片形态特征上的种间差异和相互关系, 是了解湿地退化演替序列的关键. ...
黄河首曲湿地退化过程中植物群落组分及物种多样性的变化
1
2009
... 玛曲高寒湿地位于平均海拔3500 m的青藏高原东缘(Niu et al., 2014), 是黄河上游重要的水源补给区, 具有极其重要的生态和经济地位(张晓云等, 2005).人为和自然原因导致的湿地退化普遍存在于该地区(王根绪等, 2007; Pan et al., 2011).湿地退化导致了在退化梯度上土壤水分减少和植物群落的物种组成改变(后源等, 2009).作为湿地退化演替的过渡性阶段, 沼泽化草甸处在高寒湿地和高寒草甸的中间地带, 其物种组成具有明显的过渡特点(Xiang et al., 2009; Ma et al., 2011), 因此, 研究高寒沼泽化草甸物种在光合和叶片形态特征上的种间差异和相互关系, 是了解湿地退化演替序列的关键. ...
毛乌素沙地若干植物光合作用、蒸腾作用和水分利用效率种间及生境间差异
4
1999
... 作为生态系统的初级生产者, 绿色植物为整个生物圈提供能量, 是其他有机体赖以生存的基础(Atwell et al., 1999) , 而叶片则是绿色植物获取能量的最重要的组织结构, 因此叶片水平功能性状的研究是植物生态学长期以来研究的重点之一(Sultan, 1987; Smith et al., 1998; Ackerly et al., 2000; Wright et al., 2004).植物叶片的光合生理和形态特征(例如SLA、Pn、Tr等)体现了植物的生长策略和资源利用方式, 是植物与环境长期相互作用的结果, 也是植物完成生活史周期和适应环境的重要生理生态特性(Cunningham et al., 1999; 蒋高明和何维明, 1999; Niinemets, 2001; 牛书丽等, 2003).植物的光合生理和叶片形态特征除了受到诸多自然环境因子的影响(Larcher, 1995; Lawlor, 1995; Valladares et al., 1997; 蒋高明和朱桂杰, 2001; Nicotra et al., 2007) 外, 在不同物种、功能群以及生长类型之间也有很大差异(蒋高明和何维明, 1999; 牛书丽等, 2003; 李宏林等, 2012), 例如禾草类植物一般具有较高的水分利用效率(WUE), 而杂草类植物具有较大的叶片面积和较低的WUE; 一年生植物较多年生植物具有高的Pn等.因此, 研究在同一地域不同植物物种光合和叶片形态特征在不同物种间、功能群之间以及生长类型之间的差异, 以及这些差异之间的相关关系, 对于理解和预测整个群落的资源利用及群落结构动态具有重要的意义(Kimenov et al., 1989; Schwarz & Redmann, 1989; Bowman & Turner, 1993; Wang & Yuan, 2001). ...
... 作为生态系统的初级生产者, 绿色植物为整个生物圈提供能量, 是其他有机体赖以生存的基础(Atwell et al., 1999) , 而叶片则是绿色植物获取能量的最重要的组织结构, 因此叶片水平功能性状的研究是植物生态学长期以来研究的重点之一(Sultan, 1987; Smith et al., 1998; Ackerly et al., 2000; Wright et al., 2004).植物叶片的光合生理和形态特征(例如SLA、Pn、Tr等)体现了植物的生长策略和资源利用方式, 是植物与环境长期相互作用的结果, 也是植物完成生活史周期和适应环境的重要生理生态特性(Cunningham et al., 1999; 蒋高明和何维明, 1999; Niinemets, 2001; 牛书丽等, 2003).植物的光合生理和叶片形态特征除了受到诸多自然环境因子的影响(Larcher, 1995; Lawlor, 1995; Valladares et al., 1997; 蒋高明和朱桂杰, 2001; Nicotra et al., 2007) 外, 在不同物种、功能群以及生长类型之间也有很大差异(蒋高明和何维明, 1999; 牛书丽等, 2003; 李宏林等, 2012), 例如禾草类植物一般具有较高的水分利用效率(WUE), 而杂草类植物具有较大的叶片面积和较低的WUE; 一年生植物较多年生植物具有高的Pn等.因此, 研究在同一地域不同植物物种光合和叶片形态特征在不同物种间、功能群之间以及生长类型之间的差异, 以及这些差异之间的相关关系, 对于理解和预测整个群落的资源利用及群落结构动态具有重要的意义(Kimenov et al., 1989; Schwarz & Redmann, 1989; Bowman & Turner, 1993; Wang & Yuan, 2001). ...
The effects of water- and nitrogen-induced stresses on plant community structure in a semiarid grassland.
1
1978
... 植物功能群的多样性被认为是比物种多样性更重要的生态指标(Walker, 1992; Power et al., 1996; Hooper & Vitousek, 1997; Tilman et al., 1997; Mack & D’Antonio, 1998).有研究认为功能群可以将物种水平的生活史策略和生态系统水平的进程联系起来(Chapin, 1993).根据同一功能群的物种应该具有相似的形态或生理性状(Lauenroth et al., 1978; Pokorny et al., 2005), 我们将玛曲高寒沼泽化草甸的物种划分为3个功能群: 莎草科、禾本科和双子叶类杂草(李伟, 2011).总体上, 不同功能群之间无论在叶片形态特征还是在光合生理特征上都存在显著的差异.莎草科和禾本科植物具有相似且较小的LA和LWC, 但是莎草科植物在Pn和Tr上却显著地高于禾本科植物, 而禾本科植物的SLA显著高于莎草科植物和双子叶类杂草, 最终在叶片水平的WUE指标上, 禾本科和莎草科植物远高于双子叶类杂草.叶片形态和光合生理指标在不同功能群之间的不同显示了不同功能群物种在资源利用上可能的差异, 而这种差异将有助于植物物种对环境资源的充分利用, 进而提高整个系统的稳定性(Naeem et al., 1994; Anderson & Inouye, 2001).由于叶片水平WUE的差异, 如果双子叶类杂草的多度进一步增加, 则可能导致整个群落WUE的降低, 相反, 禾本科和莎草科植物的增加则会提高群落的WUE (Niu et al., 2011). ...
Photosynthesis, productivity and environment.
1
1995
... 作为生态系统的初级生产者, 绿色植物为整个生物圈提供能量, 是其他有机体赖以生存的基础(Atwell et al., 1999) , 而叶片则是绿色植物获取能量的最重要的组织结构, 因此叶片水平功能性状的研究是植物生态学长期以来研究的重点之一(Sultan, 1987; Smith et al., 1998; Ackerly et al., 2000; Wright et al., 2004).植物叶片的光合生理和形态特征(例如SLA、Pn、Tr等)体现了植物的生长策略和资源利用方式, 是植物与环境长期相互作用的结果, 也是植物完成生活史周期和适应环境的重要生理生态特性(Cunningham et al., 1999; 蒋高明和何维明, 1999; Niinemets, 2001; 牛书丽等, 2003).植物的光合生理和叶片形态特征除了受到诸多自然环境因子的影响(Larcher, 1995; Lawlor, 1995; Valladares et al., 1997; 蒋高明和朱桂杰, 2001; Nicotra et al., 2007) 外, 在不同物种、功能群以及生长类型之间也有很大差异(蒋高明和何维明, 1999; 牛书丽等, 2003; 李宏林等, 2012), 例如禾草类植物一般具有较高的水分利用效率(WUE), 而杂草类植物具有较大的叶片面积和较低的WUE; 一年生植物较多年生植物具有高的Pn等.因此, 研究在同一地域不同植物物种光合和叶片形态特征在不同物种间、功能群之间以及生长类型之间的差异, 以及这些差异之间的相关关系, 对于理解和预测整个群落的资源利用及群落结构动态具有重要的意义(Kimenov et al., 1989; Schwarz & Redmann, 1989; Bowman & Turner, 1993; Wang & Yuan, 2001). ...
青藏高原高寒沼泽湿地在退化梯度上植物群落组成的改变对湿地水分状况的影响
1
2012
... 作为生态系统的初级生产者, 绿色植物为整个生物圈提供能量, 是其他有机体赖以生存的基础(Atwell et al., 1999) , 而叶片则是绿色植物获取能量的最重要的组织结构, 因此叶片水平功能性状的研究是植物生态学长期以来研究的重点之一(Sultan, 1987; Smith et al., 1998; Ackerly et al., 2000; Wright et al., 2004).植物叶片的光合生理和形态特征(例如SLA、Pn、Tr等)体现了植物的生长策略和资源利用方式, 是植物与环境长期相互作用的结果, 也是植物完成生活史周期和适应环境的重要生理生态特性(Cunningham et al., 1999; 蒋高明和何维明, 1999; Niinemets, 2001; 牛书丽等, 2003).植物的光合生理和叶片形态特征除了受到诸多自然环境因子的影响(Larcher, 1995; Lawlor, 1995; Valladares et al., 1997; 蒋高明和朱桂杰, 2001; Nicotra et al., 2007) 外, 在不同物种、功能群以及生长类型之间也有很大差异(蒋高明和何维明, 1999; 牛书丽等, 2003; 李宏林等, 2012), 例如禾草类植物一般具有较高的水分利用效率(WUE), 而杂草类植物具有较大的叶片面积和较低的WUE; 一年生植物较多年生植物具有高的Pn等.因此, 研究在同一地域不同植物物种光合和叶片形态特征在不同物种间、功能群之间以及生长类型之间的差异, 以及这些差异之间的相关关系, 对于理解和预测整个群落的资源利用及群落结构动态具有重要的意义(Kimenov et al., 1989; Schwarz & Redmann, 1989; Bowman & Turner, 1993; Wang & Yuan, 2001). ...
青藏高原高寒沼泽湿地在退化梯度上植物群落组成的改变对湿地水分状况的影响
1
2012
... 作为生态系统的初级生产者, 绿色植物为整个生物圈提供能量, 是其他有机体赖以生存的基础(Atwell et al., 1999) , 而叶片则是绿色植物获取能量的最重要的组织结构, 因此叶片水平功能性状的研究是植物生态学长期以来研究的重点之一(Sultan, 1987; Smith et al., 1998; Ackerly et al., 2000; Wright et al., 2004).植物叶片的光合生理和形态特征(例如SLA、Pn、Tr等)体现了植物的生长策略和资源利用方式, 是植物与环境长期相互作用的结果, 也是植物完成生活史周期和适应环境的重要生理生态特性(Cunningham et al., 1999; 蒋高明和何维明, 1999; Niinemets, 2001; 牛书丽等, 2003).植物的光合生理和叶片形态特征除了受到诸多自然环境因子的影响(Larcher, 1995; Lawlor, 1995; Valladares et al., 1997; 蒋高明和朱桂杰, 2001; Nicotra et al., 2007) 外, 在不同物种、功能群以及生长类型之间也有很大差异(蒋高明和何维明, 1999; 牛书丽等, 2003; 李宏林等, 2012), 例如禾草类植物一般具有较高的水分利用效率(WUE), 而杂草类植物具有较大的叶片面积和较低的WUE; 一年生植物较多年生植物具有高的Pn等.因此, 研究在同一地域不同植物物种光合和叶片形态特征在不同物种间、功能群之间以及生长类型之间的差异, 以及这些差异之间的相关关系, 对于理解和预测整个群落的资源利用及群落结构动态具有重要的意义(Kimenov et al., 1989; Schwarz & Redmann, 1989; Bowman & Turner, 1993; Wang & Yuan, 2001). ...
施肥导致高寒草甸物种多样性丧失机制研究
1
2011
... 植物功能群的多样性被认为是比物种多样性更重要的生态指标(Walker, 1992; Power et al., 1996; Hooper & Vitousek, 1997; Tilman et al., 1997; Mack & D’Antonio, 1998).有研究认为功能群可以将物种水平的生活史策略和生态系统水平的进程联系起来(Chapin, 1993).根据同一功能群的物种应该具有相似的形态或生理性状(Lauenroth et al., 1978; Pokorny et al., 2005), 我们将玛曲高寒沼泽化草甸的物种划分为3个功能群: 莎草科、禾本科和双子叶类杂草(李伟, 2011).总体上, 不同功能群之间无论在叶片形态特征还是在光合生理特征上都存在显著的差异.莎草科和禾本科植物具有相似且较小的LA和LWC, 但是莎草科植物在Pn和Tr上却显著地高于禾本科植物, 而禾本科植物的SLA显著高于莎草科植物和双子叶类杂草, 最终在叶片水平的WUE指标上, 禾本科和莎草科植物远高于双子叶类杂草.叶片形态和光合生理指标在不同功能群之间的不同显示了不同功能群物种在资源利用上可能的差异, 而这种差异将有助于植物物种对环境资源的充分利用, 进而提高整个系统的稳定性(Naeem et al., 1994; Anderson & Inouye, 2001).由于叶片水平WUE的差异, 如果双子叶类杂草的多度进一步增加, 则可能导致整个群落WUE的降低, 相反, 禾本科和莎草科植物的增加则会提高群落的WUE (Niu et al., 2011). ...
施肥导致高寒草甸物种多样性丧失机制研究
1
2011
... 植物功能群的多样性被认为是比物种多样性更重要的生态指标(Walker, 1992; Power et al., 1996; Hooper & Vitousek, 1997; Tilman et al., 1997; Mack & D’Antonio, 1998).有研究认为功能群可以将物种水平的生活史策略和生态系统水平的进程联系起来(Chapin, 1993).根据同一功能群的物种应该具有相似的形态或生理性状(Lauenroth et al., 1978; Pokorny et al., 2005), 我们将玛曲高寒沼泽化草甸的物种划分为3个功能群: 莎草科、禾本科和双子叶类杂草(李伟, 2011).总体上, 不同功能群之间无论在叶片形态特征还是在光合生理特征上都存在显著的差异.莎草科和禾本科植物具有相似且较小的LA和LWC, 但是莎草科植物在Pn和Tr上却显著地高于禾本科植物, 而禾本科植物的SLA显著高于莎草科植物和双子叶类杂草, 最终在叶片水平的WUE指标上, 禾本科和莎草科植物远高于双子叶类杂草.叶片形态和光合生理指标在不同功能群之间的不同显示了不同功能群物种在资源利用上可能的差异, 而这种差异将有助于植物物种对环境资源的充分利用, 进而提高整个系统的稳定性(Naeem et al., 1994; Anderson & Inouye, 2001).由于叶片水平WUE的差异, 如果双子叶类杂草的多度进一步增加, 则可能导致整个群落WUE的降低, 相反, 禾本科和莎草科植物的增加则会提高群落的WUE (Niu et al., 2011). ...
Soil seed bank dynamics in alpine wetland succession on the Tibetan Plateau.
1
2011
... 玛曲高寒湿地位于平均海拔3500 m的青藏高原东缘(Niu et al., 2014), 是黄河上游重要的水源补给区, 具有极其重要的生态和经济地位(张晓云等, 2005).人为和自然原因导致的湿地退化普遍存在于该地区(王根绪等, 2007; Pan et al., 2011).湿地退化导致了在退化梯度上土壤水分减少和植物群落的物种组成改变(后源等, 2009).作为湿地退化演替的过渡性阶段, 沼泽化草甸处在高寒湿地和高寒草甸的中间地带, 其物种组成具有明显的过渡特点(Xiang et al., 2009; Ma et al., 2011), 因此, 研究高寒沼泽化草甸物种在光合和叶片形态特征上的种间差异和相互关系, 是了解湿地退化演替序列的关键. ...
Impacts of biological invasions on disturbance regimes.
1
1998
... 植物功能群的多样性被认为是比物种多样性更重要的生态指标(Walker, 1992; Power et al., 1996; Hooper & Vitousek, 1997; Tilman et al., 1997; Mack & D’Antonio, 1998).有研究认为功能群可以将物种水平的生活史策略和生态系统水平的进程联系起来(Chapin, 1993).根据同一功能群的物种应该具有相似的形态或生理性状(Lauenroth et al., 1978; Pokorny et al., 2005), 我们将玛曲高寒沼泽化草甸的物种划分为3个功能群: 莎草科、禾本科和双子叶类杂草(李伟, 2011).总体上, 不同功能群之间无论在叶片形态特征还是在光合生理特征上都存在显著的差异.莎草科和禾本科植物具有相似且较小的LA和LWC, 但是莎草科植物在Pn和Tr上却显著地高于禾本科植物, 而禾本科植物的SLA显著高于莎草科植物和双子叶类杂草, 最终在叶片水平的WUE指标上, 禾本科和莎草科植物远高于双子叶类杂草.叶片形态和光合生理指标在不同功能群之间的不同显示了不同功能群物种在资源利用上可能的差异, 而这种差异将有助于植物物种对环境资源的充分利用, 进而提高整个系统的稳定性(Naeem et al., 1994; Anderson & Inouye, 2001).由于叶片水平WUE的差异, 如果双子叶类杂草的多度进一步增加, 则可能导致整个群落WUE的降低, 相反, 禾本科和莎草科植物的增加则会提高群落的WUE (Niu et al., 2011). ...
Declining biodiversity can alter the performance of ecosystems.
1
1994
... 植物功能群的多样性被认为是比物种多样性更重要的生态指标(Walker, 1992; Power et al., 1996; Hooper & Vitousek, 1997; Tilman et al., 1997; Mack & D’Antonio, 1998).有研究认为功能群可以将物种水平的生活史策略和生态系统水平的进程联系起来(Chapin, 1993).根据同一功能群的物种应该具有相似的形态或生理性状(Lauenroth et al., 1978; Pokorny et al., 2005), 我们将玛曲高寒沼泽化草甸的物种划分为3个功能群: 莎草科、禾本科和双子叶类杂草(李伟, 2011).总体上, 不同功能群之间无论在叶片形态特征还是在光合生理特征上都存在显著的差异.莎草科和禾本科植物具有相似且较小的LA和LWC, 但是莎草科植物在Pn和Tr上却显著地高于禾本科植物, 而禾本科植物的SLA显著高于莎草科植物和双子叶类杂草, 最终在叶片水平的WUE指标上, 禾本科和莎草科植物远高于双子叶类杂草.叶片形态和光合生理指标在不同功能群之间的不同显示了不同功能群物种在资源利用上可能的差异, 而这种差异将有助于植物物种对环境资源的充分利用, 进而提高整个系统的稳定性(Naeem et al., 1994; Anderson & Inouye, 2001).由于叶片水平WUE的差异, 如果双子叶类杂草的多度进一步增加, 则可能导致整个群落WUE的降低, 相反, 禾本科和莎草科植物的增加则会提高群落的WUE (Niu et al., 2011). ...
Geographic variation and plasticity to water and nutrients in
4
2007
... 基于功能性状(functional traits)的研究方法被广泛地应用于生态学和进化生物学的研究中, 功能性状通过影响个体的生存、生长、生殖来间接地影响其适合度, 可以作为预测有机体生长表现的重要指标(Violle et al., 2007).个体或物种间在功能性状间的差异既反映了其所处的生境特征, 也体现了它们不同的资源利用方式.例如与生长于水分充足的生境的植物相比, 生长于水分不足生境中的植物一般具有较小的比叶面积(specific leaf area, SLA), 较低的净光合速率(net photosynthesis rate, Pn)和蒸腾速率(transpiration rate, Tr), 体现了在控制水分散失和提高生长速率之间的权衡(Reich et al., 1997; Nicotra et al., 2007). ...
... 作为生态系统的初级生产者, 绿色植物为整个生物圈提供能量, 是其他有机体赖以生存的基础(Atwell et al., 1999) , 而叶片则是绿色植物获取能量的最重要的组织结构, 因此叶片水平功能性状的研究是植物生态学长期以来研究的重点之一(Sultan, 1987; Smith et al., 1998; Ackerly et al., 2000; Wright et al., 2004).植物叶片的光合生理和形态特征(例如SLA、Pn、Tr等)体现了植物的生长策略和资源利用方式, 是植物与环境长期相互作用的结果, 也是植物完成生活史周期和适应环境的重要生理生态特性(Cunningham et al., 1999; 蒋高明和何维明, 1999; Niinemets, 2001; 牛书丽等, 2003).植物的光合生理和叶片形态特征除了受到诸多自然环境因子的影响(Larcher, 1995; Lawlor, 1995; Valladares et al., 1997; 蒋高明和朱桂杰, 2001; Nicotra et al., 2007) 外, 在不同物种、功能群以及生长类型之间也有很大差异(蒋高明和何维明, 1999; 牛书丽等, 2003; 李宏林等, 2012), 例如禾草类植物一般具有较高的水分利用效率(WUE), 而杂草类植物具有较大的叶片面积和较低的WUE; 一年生植物较多年生植物具有高的Pn等.因此, 研究在同一地域不同植物物种光合和叶片形态特征在不同物种间、功能群之间以及生长类型之间的差异, 以及这些差异之间的相关关系, 对于理解和预测整个群落的资源利用及群落结构动态具有重要的意义(Kimenov et al., 1989; Schwarz & Redmann, 1989; Bowman & Turner, 1993; Wang & Yuan, 2001). ...
... 作为植物整体其中一部分的功能性状之间存在非常紧密的关系, 比如光合能力和叶片结构之间(Niinemets, 2001; Nicotra et al., 2007).这种相关关系在对全球尺度的跨物种、跨生境的研究中得到了证实(Wright et al., 2004).我们对高寒沼泽化草甸51个物种功能性状之间相互关系的分析(光合生理特征和叶片形态特征之间)也显示了功能性状之间存在着显著的相关性(图2, 图3).在整体上, LA上分配的生物量的提高(SLA减小)使得LA上的Pn增加, 这与之前所报道的结果是一致的(Cunningham et al., 1999; Niinemets, 2001).一般而言, Pn的提高伴随的是Tr的增加, 而我们的结果显示Tr的增加跟叶片的LWC成正比(图2C).Pn和Tr决定着植物叶片水平的WUE, 比如Pn的减小和Tr的增加都使得WUE减小, 这是叶片形态性状SLA和LWC的增加导致WUE减小的可能原因(图2B, 2D), SLA和LWC通过影响Pn和Tr间接地影响WUE. ...
Fertilization decreases species diversity but increases functional diversity: A three-year experiment in a Tibetan alpine meadow.
2
2014
... 玛曲高寒湿地位于平均海拔3500 m的青藏高原东缘(Niu et al., 2014), 是黄河上游重要的水源补给区, 具有极其重要的生态和经济地位(张晓云等, 2005).人为和自然原因导致的湿地退化普遍存在于该地区(王根绪等, 2007; Pan et al., 2011).湿地退化导致了在退化梯度上土壤水分减少和植物群落的物种组成改变(后源等, 2009).作为湿地退化演替的过渡性阶段, 沼泽化草甸处在高寒湿地和高寒草甸的中间地带, 其物种组成具有明显的过渡特点(Xiang et al., 2009; Ma et al., 2011), 因此, 研究高寒沼泽化草甸物种在光合和叶片形态特征上的种间差异和相互关系, 是了解湿地退化演替序列的关键. ...
... 实验地位于甘肃省甘南藏族自治州玛曲县境内的高寒湿地(33.65° N, 101.88° E), 平均海拔3650 m, 多年观测的年降水量和气温分别为672 mm和2.2 ℃, 年日照时间约为2580 h, 平均无霜期每年少于100天(Chu et al., 2008).该地区属于高寒半湿润半干旱气候, 降水主要集中于短暂的夏季(Niu et al., 2014), 土壤以亚高寒草甸土和泥炭土为主, 主要植被类型为亚高寒草甸(葛庆征等, 2012). ...
内蒙古浑善达克沙地97种植物的光合生理特征
3
2003
... 作为生态系统的初级生产者, 绿色植物为整个生物圈提供能量, 是其他有机体赖以生存的基础(Atwell et al., 1999) , 而叶片则是绿色植物获取能量的最重要的组织结构, 因此叶片水平功能性状的研究是植物生态学长期以来研究的重点之一(Sultan, 1987; Smith et al., 1998; Ackerly et al., 2000; Wright et al., 2004).植物叶片的光合生理和形态特征(例如SLA、Pn、Tr等)体现了植物的生长策略和资源利用方式, 是植物与环境长期相互作用的结果, 也是植物完成生活史周期和适应环境的重要生理生态特性(Cunningham et al., 1999; 蒋高明和何维明, 1999; Niinemets, 2001; 牛书丽等, 2003).植物的光合生理和叶片形态特征除了受到诸多自然环境因子的影响(Larcher, 1995; Lawlor, 1995; Valladares et al., 1997; 蒋高明和朱桂杰, 2001; Nicotra et al., 2007) 外, 在不同物种、功能群以及生长类型之间也有很大差异(蒋高明和何维明, 1999; 牛书丽等, 2003; 李宏林等, 2012), 例如禾草类植物一般具有较高的水分利用效率(WUE), 而杂草类植物具有较大的叶片面积和较低的WUE; 一年生植物较多年生植物具有高的Pn等.因此, 研究在同一地域不同植物物种光合和叶片形态特征在不同物种间、功能群之间以及生长类型之间的差异, 以及这些差异之间的相关关系, 对于理解和预测整个群落的资源利用及群落结构动态具有重要的意义(Kimenov et al., 1989; Schwarz & Redmann, 1989; Bowman & Turner, 1993; Wang & Yuan, 2001). ...
Water-use efficiency in response to climate change: From leaf to ecosystem in a temperate steppe.
1
2011
... 植物功能群的多样性被认为是比物种多样性更重要的生态指标(Walker, 1992; Power et al., 1996; Hooper & Vitousek, 1997; Tilman et al., 1997; Mack & D’Antonio, 1998).有研究认为功能群可以将物种水平的生活史策略和生态系统水平的进程联系起来(Chapin, 1993).根据同一功能群的物种应该具有相似的形态或生理性状(Lauenroth et al., 1978; Pokorny et al., 2005), 我们将玛曲高寒沼泽化草甸的物种划分为3个功能群: 莎草科、禾本科和双子叶类杂草(李伟, 2011).总体上, 不同功能群之间无论在叶片形态特征还是在光合生理特征上都存在显著的差异.莎草科和禾本科植物具有相似且较小的LA和LWC, 但是莎草科植物在Pn和Tr上却显著地高于禾本科植物, 而禾本科植物的SLA显著高于莎草科植物和双子叶类杂草, 最终在叶片水平的WUE指标上, 禾本科和莎草科植物远高于双子叶类杂草.叶片形态和光合生理指标在不同功能群之间的不同显示了不同功能群物种在资源利用上可能的差异, 而这种差异将有助于植物物种对环境资源的充分利用, 进而提高整个系统的稳定性(Naeem et al., 1994; Anderson & Inouye, 2001).由于叶片水平WUE的差异, 如果双子叶类杂草的多度进一步增加, 则可能导致整个群落WUE的降低, 相反, 禾本科和莎草科植物的增加则会提高群落的WUE (Niu et al., 2011). ...
Natural wetland in China.
1
2011
... 玛曲高寒湿地位于平均海拔3500 m的青藏高原东缘(Niu et al., 2014), 是黄河上游重要的水源补给区, 具有极其重要的生态和经济地位(张晓云等, 2005).人为和自然原因导致的湿地退化普遍存在于该地区(王根绪等, 2007; Pan et al., 2011).湿地退化导致了在退化梯度上土壤水分减少和植物群落的物种组成改变(后源等, 2009).作为湿地退化演替的过渡性阶段, 沼泽化草甸处在高寒湿地和高寒草甸的中间地带, 其物种组成具有明显的过渡特点(Xiang et al., 2009; Ma et al., 2011), 因此, 研究高寒沼泽化草甸物种在光合和叶片形态特征上的种间差异和相互关系, 是了解湿地退化演替序列的关键. ...
Decoupled evolution of foliar freezing resistance, temperature niche and morphological leaf traits in Chilean Myrceugenia.
... O, 这些值高于我们在高寒沼泽化草甸中观测到的结果, 可以看到早春短命植物具有相对较高的光合和蒸腾速率, 说明这些植物与多年生的高寒沼泽化草甸植物采取的是不同的生长策略, 这可能和早春短命植物需要在相对较短的时间内完成生活史周期有关(邱娟等, 2007). ...
Reconciling apparent discrepancies among studies relating life span, structure and function of leaves in contrasting plant forms and climates: ‘The blind men and the elephant retold’.
Photosynthetic properties of C4 grass (Spartina gracilis Trin.) from northern environment.
1
1989
... 作为生态系统的初级生产者, 绿色植物为整个生物圈提供能量, 是其他有机体赖以生存的基础(Atwell et al., 1999) , 而叶片则是绿色植物获取能量的最重要的组织结构, 因此叶片水平功能性状的研究是植物生态学长期以来研究的重点之一(Sultan, 1987; Smith et al., 1998; Ackerly et al., 2000; Wright et al., 2004).植物叶片的光合生理和形态特征(例如SLA、Pn、Tr等)体现了植物的生长策略和资源利用方式, 是植物与环境长期相互作用的结果, 也是植物完成生活史周期和适应环境的重要生理生态特性(Cunningham et al., 1999; 蒋高明和何维明, 1999; Niinemets, 2001; 牛书丽等, 2003).植物的光合生理和叶片形态特征除了受到诸多自然环境因子的影响(Larcher, 1995; Lawlor, 1995; Valladares et al., 1997; 蒋高明和朱桂杰, 2001; Nicotra et al., 2007) 外, 在不同物种、功能群以及生长类型之间也有很大差异(蒋高明和何维明, 1999; 牛书丽等, 2003; 李宏林等, 2012), 例如禾草类植物一般具有较高的水分利用效率(WUE), 而杂草类植物具有较大的叶片面积和较低的WUE; 一年生植物较多年生植物具有高的Pn等.因此, 研究在同一地域不同植物物种光合和叶片形态特征在不同物种间、功能群之间以及生长类型之间的差异, 以及这些差异之间的相关关系, 对于理解和预测整个群落的资源利用及群落结构动态具有重要的意义(Kimenov et al., 1989; Schwarz & Redmann, 1989; Bowman & Turner, 1993; Wang & Yuan, 2001). ...
Intra-specific variation in the photosynthetic responses of cyanobiont lichens from contrasting habitats.
3
1998
... 作为生态系统的初级生产者, 绿色植物为整个生物圈提供能量, 是其他有机体赖以生存的基础(Atwell et al., 1999) , 而叶片则是绿色植物获取能量的最重要的组织结构, 因此叶片水平功能性状的研究是植物生态学长期以来研究的重点之一(Sultan, 1987; Smith et al., 1998; Ackerly et al., 2000; Wright et al., 2004).植物叶片的光合生理和形态特征(例如SLA、Pn、Tr等)体现了植物的生长策略和资源利用方式, 是植物与环境长期相互作用的结果, 也是植物完成生活史周期和适应环境的重要生理生态特性(Cunningham et al., 1999; 蒋高明和何维明, 1999; Niinemets, 2001; 牛书丽等, 2003).植物的光合生理和叶片形态特征除了受到诸多自然环境因子的影响(Larcher, 1995; Lawlor, 1995; Valladares et al., 1997; 蒋高明和朱桂杰, 2001; Nicotra et al., 2007) 外, 在不同物种、功能群以及生长类型之间也有很大差异(蒋高明和何维明, 1999; 牛书丽等, 2003; 李宏林等, 2012), 例如禾草类植物一般具有较高的水分利用效率(WUE), 而杂草类植物具有较大的叶片面积和较低的WUE; 一年生植物较多年生植物具有高的Pn等.因此, 研究在同一地域不同植物物种光合和叶片形态特征在不同物种间、功能群之间以及生长类型之间的差异, 以及这些差异之间的相关关系, 对于理解和预测整个群落的资源利用及群落结构动态具有重要的意义(Kimenov et al., 1989; Schwarz & Redmann, 1989; Bowman & Turner, 1993; Wang & Yuan, 2001). ...
... 许多研究表明, 植物在叶片形态特征(如SLA、叶片含水量等)和光合生理特征(如Pn、Tr、气孔导度等)上存在着显著的种间和种内差异(Smith et al., 1998; Wright et al., 2004), 即使它们生长在非常相似的环境中(Bassow & Bazzaz, 1997), 被认为这是长期自然选择的结果(蒋高明和何维明, 1999).我们对51个组成玛曲高寒沼泽化草甸的物种的叶片形态和光合生理特征的分析表明, 不同物种之间在LWC、SLA、LA、Pn、Tr和WUE之间有着非常明显的差异, 说明这些物种虽然生长在非常相似的环境中, 但是在叶片形态和光合策略上有着显著的不同(Smith et al., 1998), 以不同的方式适应相似的生境条件. ...
... 之间有着非常明显的差异, 说明这些物种虽然生长在非常相似的环境中, 但是在叶片形态和光合策略上有着显著的不同(Smith et al., 1998), 以不同的方式适应相似的生境条件. ...
Evolutionary implications of phenotypic plasticity in plants.
1
1987
... 作为生态系统的初级生产者, 绿色植物为整个生物圈提供能量, 是其他有机体赖以生存的基础(Atwell et al., 1999) , 而叶片则是绿色植物获取能量的最重要的组织结构, 因此叶片水平功能性状的研究是植物生态学长期以来研究的重点之一(Sultan, 1987; Smith et al., 1998; Ackerly et al., 2000; Wright et al., 2004).植物叶片的光合生理和形态特征(例如SLA、Pn、Tr等)体现了植物的生长策略和资源利用方式, 是植物与环境长期相互作用的结果, 也是植物完成生活史周期和适应环境的重要生理生态特性(Cunningham et al., 1999; 蒋高明和何维明, 1999; Niinemets, 2001; 牛书丽等, 2003).植物的光合生理和叶片形态特征除了受到诸多自然环境因子的影响(Larcher, 1995; Lawlor, 1995; Valladares et al., 1997; 蒋高明和朱桂杰, 2001; Nicotra et al., 2007) 外, 在不同物种、功能群以及生长类型之间也有很大差异(蒋高明和何维明, 1999; 牛书丽等, 2003; 李宏林等, 2012), 例如禾草类植物一般具有较高的水分利用效率(WUE), 而杂草类植物具有较大的叶片面积和较低的WUE; 一年生植物较多年生植物具有高的Pn等.因此, 研究在同一地域不同植物物种光合和叶片形态特征在不同物种间、功能群之间以及生长类型之间的差异, 以及这些差异之间的相关关系, 对于理解和预测整个群落的资源利用及群落结构动态具有重要的意义(Kimenov et al., 1989; Schwarz & Redmann, 1989; Bowman & Turner, 1993; Wang & Yuan, 2001). ...
The influence of functional diversity and composition on ecosystem processes.
1
1997
... 植物功能群的多样性被认为是比物种多样性更重要的生态指标(Walker, 1992; Power et al., 1996; Hooper & Vitousek, 1997; Tilman et al., 1997; Mack & D’Antonio, 1998).有研究认为功能群可以将物种水平的生活史策略和生态系统水平的进程联系起来(Chapin, 1993).根据同一功能群的物种应该具有相似的形态或生理性状(Lauenroth et al., 1978; Pokorny et al., 2005), 我们将玛曲高寒沼泽化草甸的物种划分为3个功能群: 莎草科、禾本科和双子叶类杂草(李伟, 2011).总体上, 不同功能群之间无论在叶片形态特征还是在光合生理特征上都存在显著的差异.莎草科和禾本科植物具有相似且较小的LA和LWC, 但是莎草科植物在Pn和Tr上却显著地高于禾本科植物, 而禾本科植物的SLA显著高于莎草科植物和双子叶类杂草, 最终在叶片水平的WUE指标上, 禾本科和莎草科植物远高于双子叶类杂草.叶片形态和光合生理指标在不同功能群之间的不同显示了不同功能群物种在资源利用上可能的差异, 而这种差异将有助于植物物种对环境资源的充分利用, 进而提高整个系统的稳定性(Naeem et al., 1994; Anderson & Inouye, 2001).由于叶片水平WUE的差异, 如果双子叶类杂草的多度进一步增加, 则可能导致整个群落WUE的降低, 相反, 禾本科和莎草科植物的增加则会提高群落的WUE (Niu et al., 2011). ...
Photosynthetic responses to dynamic light under field conditions in six tropical rainforest shrubs occuring along a light gradient.
1
1997
... 作为生态系统的初级生产者, 绿色植物为整个生物圈提供能量, 是其他有机体赖以生存的基础(Atwell et al., 1999) , 而叶片则是绿色植物获取能量的最重要的组织结构, 因此叶片水平功能性状的研究是植物生态学长期以来研究的重点之一(Sultan, 1987; Smith et al., 1998; Ackerly et al., 2000; Wright et al., 2004).植物叶片的光合生理和形态特征(例如SLA、Pn、Tr等)体现了植物的生长策略和资源利用方式, 是植物与环境长期相互作用的结果, 也是植物完成生活史周期和适应环境的重要生理生态特性(Cunningham et al., 1999; 蒋高明和何维明, 1999; Niinemets, 2001; 牛书丽等, 2003).植物的光合生理和叶片形态特征除了受到诸多自然环境因子的影响(Larcher, 1995; Lawlor, 1995; Valladares et al., 1997; 蒋高明和朱桂杰, 2001; Nicotra et al., 2007) 外, 在不同物种、功能群以及生长类型之间也有很大差异(蒋高明和何维明, 1999; 牛书丽等, 2003; 李宏林等, 2012), 例如禾草类植物一般具有较高的水分利用效率(WUE), 而杂草类植物具有较大的叶片面积和较低的WUE; 一年生植物较多年生植物具有高的Pn等.因此, 研究在同一地域不同植物物种光合和叶片形态特征在不同物种间、功能群之间以及生长类型之间的差异, 以及这些差异之间的相关关系, 对于理解和预测整个群落的资源利用及群落结构动态具有重要的意义(Kimenov et al., 1989; Schwarz & Redmann, 1989; Bowman & Turner, 1993; Wang & Yuan, 2001). ...
Let the concept of trait be functional!
1
2007
... 基于功能性状(functional traits)的研究方法被广泛地应用于生态学和进化生物学的研究中, 功能性状通过影响个体的生存、生长、生殖来间接地影响其适合度, 可以作为预测有机体生长表现的重要指标(Violle et al., 2007).个体或物种间在功能性状间的差异既反映了其所处的生境特征, 也体现了它们不同的资源利用方式.例如与生长于水分充足的生境的植物相比, 生长于水分不足生境中的植物一般具有较小的比叶面积(specific leaf area, SLA), 较低的净光合速率(net photosynthesis rate, Pn)和蒸腾速率(transpiration rate, Tr), 体现了在控制水分散失和提高生长速率之间的权衡(Reich et al., 1997; Nicotra et al., 2007). ...
Biodiversity and ecological redundancy.
1
1992
... 植物功能群的多样性被认为是比物种多样性更重要的生态指标(Walker, 1992; Power et al., 1996; Hooper & Vitousek, 1997; Tilman et al., 1997; Mack & D’Antonio, 1998).有研究认为功能群可以将物种水平的生活史策略和生态系统水平的进程联系起来(Chapin, 1993).根据同一功能群的物种应该具有相似的形态或生理性状(Lauenroth et al., 1978; Pokorny et al., 2005), 我们将玛曲高寒沼泽化草甸的物种划分为3个功能群: 莎草科、禾本科和双子叶类杂草(李伟, 2011).总体上, 不同功能群之间无论在叶片形态特征还是在光合生理特征上都存在显著的差异.莎草科和禾本科植物具有相似且较小的LA和LWC, 但是莎草科植物在Pn和Tr上却显著地高于禾本科植物, 而禾本科植物的SLA显著高于莎草科植物和双子叶类杂草, 最终在叶片水平的WUE指标上, 禾本科和莎草科植物远高于双子叶类杂草.叶片形态和光合生理指标在不同功能群之间的不同显示了不同功能群物种在资源利用上可能的差异, 而这种差异将有助于植物物种对环境资源的充分利用, 进而提高整个系统的稳定性(Naeem et al., 1994; Anderson & Inouye, 2001).由于叶片水平WUE的差异, 如果双子叶类杂草的多度进一步增加, 则可能导致整个群落WUE的降低, 相反, 禾本科和莎草科植物的增加则会提高群落的WUE (Niu et al., 2011). ...
近40年来青藏高原典型高寒湿地系统的动态变化
1
2007
... 玛曲高寒湿地位于平均海拔3500 m的青藏高原东缘(Niu et al., 2014), 是黄河上游重要的水源补给区, 具有极其重要的生态和经济地位(张晓云等, 2005).人为和自然原因导致的湿地退化普遍存在于该地区(王根绪等, 2007; Pan et al., 2011).湿地退化导致了在退化梯度上土壤水分减少和植物群落的物种组成改变(后源等, 2009).作为湿地退化演替的过渡性阶段, 沼泽化草甸处在高寒湿地和高寒草甸的中间地带, 其物种组成具有明显的过渡特点(Xiang et al., 2009; Ma et al., 2011), 因此, 研究高寒沼泽化草甸物种在光合和叶片形态特征上的种间差异和相互关系, 是了解湿地退化演替序列的关键. ...
近40年来青藏高原典型高寒湿地系统的动态变化
1
2007
... 玛曲高寒湿地位于平均海拔3500 m的青藏高原东缘(Niu et al., 2014), 是黄河上游重要的水源补给区, 具有极其重要的生态和经济地位(张晓云等, 2005).人为和自然原因导致的湿地退化普遍存在于该地区(王根绪等, 2007; Pan et al., 2011).湿地退化导致了在退化梯度上土壤水分减少和植物群落的物种组成改变(后源等, 2009).作为湿地退化演替的过渡性阶段, 沼泽化草甸处在高寒湿地和高寒草甸的中间地带, 其物种组成具有明显的过渡特点(Xiang et al., 2009; Ma et al., 2011), 因此, 研究高寒沼泽化草甸物种在光合和叶片形态特征上的种间差异和相互关系, 是了解湿地退化演替序列的关键. ...
Photosynthesis, transpiration, and water use efficiency in two divergent Leymus chinensis populations from Northeast China.
1
2001
... 作为生态系统的初级生产者, 绿色植物为整个生物圈提供能量, 是其他有机体赖以生存的基础(Atwell et al., 1999) , 而叶片则是绿色植物获取能量的最重要的组织结构, 因此叶片水平功能性状的研究是植物生态学长期以来研究的重点之一(Sultan, 1987; Smith et al., 1998; Ackerly et al., 2000; Wright et al., 2004).植物叶片的光合生理和形态特征(例如SLA、Pn、Tr等)体现了植物的生长策略和资源利用方式, 是植物与环境长期相互作用的结果, 也是植物完成生活史周期和适应环境的重要生理生态特性(Cunningham et al., 1999; 蒋高明和何维明, 1999; Niinemets, 2001; 牛书丽等, 2003).植物的光合生理和叶片形态特征除了受到诸多自然环境因子的影响(Larcher, 1995; Lawlor, 1995; Valladares et al., 1997; 蒋高明和朱桂杰, 2001; Nicotra et al., 2007) 外, 在不同物种、功能群以及生长类型之间也有很大差异(蒋高明和何维明, 1999; 牛书丽等, 2003; 李宏林等, 2012), 例如禾草类植物一般具有较高的水分利用效率(WUE), 而杂草类植物具有较大的叶片面积和较低的WUE; 一年生植物较多年生植物具有高的Pn等.因此, 研究在同一地域不同植物物种光合和叶片形态特征在不同物种间、功能群之间以及生长类型之间的差异, 以及这些差异之间的相关关系, 对于理解和预测整个群落的资源利用及群落结构动态具有重要的意义(Kimenov et al., 1989; Schwarz & Redmann, 1989; Bowman & Turner, 1993; Wang & Yuan, 2001). ...
Photosynthesis, transpiration, and water use efficiency of two Puccinellia species on the Songnen grassland, Northeastern China.
... 作为生态系统的初级生产者, 绿色植物为整个生物圈提供能量, 是其他有机体赖以生存的基础(Atwell et al., 1999) , 而叶片则是绿色植物获取能量的最重要的组织结构, 因此叶片水平功能性状的研究是植物生态学长期以来研究的重点之一(Sultan, 1987; Smith et al., 1998; Ackerly et al., 2000; Wright et al., 2004).植物叶片的光合生理和形态特征(例如SLA、Pn、Tr等)体现了植物的生长策略和资源利用方式, 是植物与环境长期相互作用的结果, 也是植物完成生活史周期和适应环境的重要生理生态特性(Cunningham et al., 1999; 蒋高明和何维明, 1999; Niinemets, 2001; 牛书丽等, 2003).植物的光合生理和叶片形态特征除了受到诸多自然环境因子的影响(Larcher, 1995; Lawlor, 1995; Valladares et al., 1997; 蒋高明和朱桂杰, 2001; Nicotra et al., 2007) 外, 在不同物种、功能群以及生长类型之间也有很大差异(蒋高明和何维明, 1999; 牛书丽等, 2003; 李宏林等, 2012), 例如禾草类植物一般具有较高的水分利用效率(WUE), 而杂草类植物具有较大的叶片面积和较低的WUE; 一年生植物较多年生植物具有高的Pn等.因此, 研究在同一地域不同植物物种光合和叶片形态特征在不同物种间、功能群之间以及生长类型之间的差异, 以及这些差异之间的相关关系, 对于理解和预测整个群落的资源利用及群落结构动态具有重要的意义(Kimenov et al., 1989; Schwarz & Redmann, 1989; Bowman & Turner, 1993; Wang & Yuan, 2001). ...
... 许多研究表明, 植物在叶片形态特征(如SLA、叶片含水量等)和光合生理特征(如Pn、Tr、气孔导度等)上存在着显著的种间和种内差异(Smith et al., 1998; Wright et al., 2004), 即使它们生长在非常相似的环境中(Bassow & Bazzaz, 1997), 被认为这是长期自然选择的结果(蒋高明和何维明, 1999).我们对51个组成玛曲高寒沼泽化草甸的物种的叶片形态和光合生理特征的分析表明, 不同物种之间在LWC、SLA、LA、Pn、Tr和WUE之间有着非常明显的差异, 说明这些物种虽然生长在非常相似的环境中, 但是在叶片形态和光合策略上有着显著的不同(Smith et al., 1998), 以不同的方式适应相似的生境条件. ...
... 作为植物整体其中一部分的功能性状之间存在非常紧密的关系, 比如光合能力和叶片结构之间(Niinemets, 2001; Nicotra et al., 2007).这种相关关系在对全球尺度的跨物种、跨生境的研究中得到了证实(Wright et al., 2004).我们对高寒沼泽化草甸51个物种功能性状之间相互关系的分析(光合生理特征和叶片形态特征之间)也显示了功能性状之间存在着显著的相关性(图2, 图3).在整体上, LA上分配的生物量的提高(SLA减小)使得LA上的Pn增加, 这与之前所报道的结果是一致的(Cunningham et al., 1999; Niinemets, 2001).一般而言, Pn的提高伴随的是Tr的增加, 而我们的结果显示Tr的增加跟叶片的LWC成正比(图2C).Pn和Tr决定着植物叶片水平的WUE, 比如Pn的减小和Tr的增加都使得WUE减小, 这是叶片形态性状SLA和LWC的增加导致WUE减小的可能原因(图2B, 2D), SLA和LWC通过影响Pn和Tr间接地影响WUE. ...
Current status and future prospects of Zoige Marsh in Eastern Qinghai-Tibet Plateau.
1
2009
... 玛曲高寒湿地位于平均海拔3500 m的青藏高原东缘(Niu et al., 2014), 是黄河上游重要的水源补给区, 具有极其重要的生态和经济地位(张晓云等, 2005).人为和自然原因导致的湿地退化普遍存在于该地区(王根绪等, 2007; Pan et al., 2011).湿地退化导致了在退化梯度上土壤水分减少和植物群落的物种组成改变(后源等, 2009).作为湿地退化演替的过渡性阶段, 沼泽化草甸处在高寒湿地和高寒草甸的中间地带, 其物种组成具有明显的过渡特点(Xiang et al., 2009; Ma et al., 2011), 因此, 研究高寒沼泽化草甸物种在光合和叶片形态特征上的种间差异和相互关系, 是了解湿地退化演替序列的关键. ...
... 玛曲高寒湿地位于平均海拔3500 m的青藏高原东缘(Niu et al., 2014), 是黄河上游重要的水源补给区, 具有极其重要的生态和经济地位(张晓云等, 2005).人为和自然原因导致的湿地退化普遍存在于该地区(王根绪等, 2007; Pan et al., 2011).湿地退化导致了在退化梯度上土壤水分减少和植物群落的物种组成改变(后源等, 2009).作为湿地退化演替的过渡性阶段, 沼泽化草甸处在高寒湿地和高寒草甸的中间地带, 其物种组成具有明显的过渡特点(Xiang et al., 2009; Ma et al., 2011), 因此, 研究高寒沼泽化草甸物种在光合和叶片形态特征上的种间差异和相互关系, 是了解湿地退化演替序列的关键. ...
若尔盖湿地面临的威胁、保护现状及对策分析
1
2005
... 玛曲高寒湿地位于平均海拔3500 m的青藏高原东缘(Niu et al., 2014), 是黄河上游重要的水源补给区, 具有极其重要的生态和经济地位(张晓云等, 2005).人为和自然原因导致的湿地退化普遍存在于该地区(王根绪等, 2007; Pan et al., 2011).湿地退化导致了在退化梯度上土壤水分减少和植物群落的物种组成改变(后源等, 2009).作为湿地退化演替的过渡性阶段, 沼泽化草甸处在高寒湿地和高寒草甸的中间地带, 其物种组成具有明显的过渡特点(Xiang et al., 2009; Ma et al., 2011), 因此, 研究高寒沼泽化草甸物种在光合和叶片形态特征上的种间差异和相互关系, 是了解湿地退化演替序列的关键. ...
 , 卜海燕
, 卜海燕

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}