红树林是生长于热带亚热带海岸潮间带、受到海水周期性浸淹的木本植物群落, 兼具陆地生态和海洋生态特性, 是海岸重要生态关键区, 在维护生物多样性和沿海地区生态安全等方面发挥着重要作用。红海榄(Rhizophora stylosa )主要分布于亚洲和大洋洲, 我国的海南、广东、广西和台湾有自然分布, 作为红树林生态系统的建群种之一, 红海榄群落多存在于泥滩深厚的中内滩, 形成致密而纯一的群落。红海榄独特的胎生现象和形态结构对独特生境(潮汐淹水和高盐生境)的适应已经有很多研究(Lin, 1999 )。植物茎干和枝条易受到环境诸因子变化带来的影响(费松林等, 1999 ; 史刚荣等, 2006 ; 史刚荣和刘蕾, 2006 ), 这种表型可塑性是植物克服环境异质性、适应环境的重要途径(Strauss-Debenedetti & Bazzaz, 1991 ), 因此, 生态解剖学方法可以揭示植物结构与环境相适应的关系。独特的红树林海岸潮间带生境为研究植物生态解剖提供了很好的样地和材料, 关于红树植物木材结构的生态解剖研究已有许多报道, 主要集中在盐度与水淹频度对木材结构的影响。最近红树植物木材结构的生态解剖研究表明, 一些红树植物在干旱季节形成层产生的导管比雨季产生的导管管径略小, 能增进水分输导的安全性, 小的纹孔膜孔径和(或)小的管间纹孔表面积能增进水分输导系统的栓塞阻力。红树植物一些特殊的木材形态结构, 如附物纹孔的结构与功能也有新报道(Sun & Lin, 1997 ; Yáñez-Espinosa et al ., 2001 ; 邓传远等, 2004a , 2004b ; Verheyden et al ., 2005 ; Schmitz et al ., 2006 , 2007 ; Robert et al ., 2009 ), 因此, 木材结构与生境相适应的研究还有广阔的空间, 特别是红树植物生长在陆海交界的生态脆弱带, 其土壤理化因子呈高度动态变化(Lin, 1999 ), 应用更多土壤理化指标探讨其对木材结构的影响很有必要。本研究运用生态解剖学的方法和原理, 探讨了红海榄木材解剖特征与土壤理化因子的关系, 揭示了次生木质部解剖特征可塑性的生态适应意义。
1 材料和方法
1.1 研究区概况
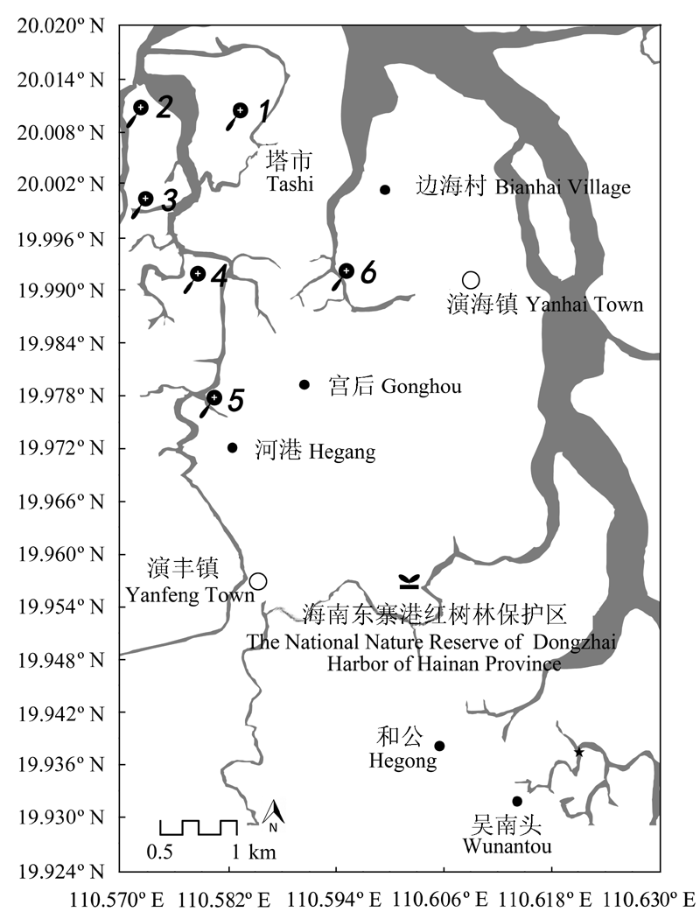
东寨港红树林国家自然保护区位于海南省东北部海口美兰区与文昌市交界处, 地理位置110.57°- 110.63° E、19.63°-20.02° N。保护面积达3337.6 hm2 , 红树林面积2065.0 hm2 。区内海拔高度10- 80 m, 坡度3°-7°, 总地势为南高北低。保护区处于热带北缘, 属热带季风气候, 年平均气温23.8 ℃, 极端最高温度38.9 ℃, 极端最低温度3.0 ℃, 海水表层年平均温度25.0 ℃; 全年平均日照2200 h; 年降水量1676. 4 mm, 80%以上降水集中在5-10月, 形成明显的干湿季; 太阳年辐射总量约为462 kJ·cm-2 。该处潮汐属不正规半日潮, 平均潮差约1 m (Lin, 1999 )。
该保护区自然分布的红树和半红树植物有木榄(Bruguiera gymnorrhiza )、海莲(Bruguiera sexangula )、红海榄、角果木(Ceriops tagal )、秋茄树(Kandelia candel )、榄李(Lumnitzera racemosa )、海漆(Excoecaria agallocha )、蜡烛果(Aegiceras corniculatum )、海榄雌(Avicennia marina )、水椰(Nypa fructicans )、老鼠簕(Acanthus ilicifolius )、小花老鼠簕(Acanthus ebracteatus )、卤蕨(Acrostichum aur- eum )、尖叶卤蕨(Acrostichum speciosum )、海杧果(Cerbera manghas )、黄槿(Hibiscus tiliaceus )、玉蕊(Barringtonia racemosa )等17种, 引种植物包括红树(Rhizophora apiculata )、木果楝(Xylocarpus grana- tum )、海南海桑(Sonneratia × hainanensis )、杯萼海桑(Sonneratia alba )、海桑(Sonneratia caseolaris )、银叶树(Heritiera littoralis )、瓶花木(Scyphiphora hydrophyllacea )、红榄李(Lumnitzera littorea )、桐棉(Thespesia populnea ), 孟加拉的无瓣海桑(Sonner- atia apetala )也被成功地引种到这里, 成为海岸滩涂和河口地带速生快长的一种红树植物。东寨港自然保护区的植物群落有7个主要群系: (1)木榄群系(Formation Bruguiera gymnorrhiza ); (2)红树群系(Formation Rhizophora apiculata ); (3)秋茄群系(Formation Kandelia candel ); (4)桐花树群系(Formation Aegiceras corniculatum ); (5)白骨壤群系(Formation Avicennia marina ); (6)海桑群系(Formation Sonneratia caseolaris ); (7)水椰群系(Formation Nypa fructicans ) (Lin, 1999 )。
1.2 材料
实验材料2008年6月取自海南省东寨港红树林自然保护区6个不连续带状分布的种群, 其种群发育时间大致相同(根据保护区管理处的资料确定), 每一种群取3株标准木的成熟茎干, 用砂纸磨光茎干的横断面, 根据生长轮粗略判断树龄, 截取生长轮一致的茎段用FAA固定。实验材料均取自潮汐浸水时间较为一致的中潮滩(依保护区的观测资料确定)。土壤样品采集方法是: 用内径为5 cm的PPT管钻取20-40 cm深的土样(20-40 cm层土样能较好代表红树林土壤理化因子的时空变动(Sun & Lin, 1997 ; Yáñez-Espinosa et al ., 2001 )), 在每一采样植物的四周采集4个土壤样品混合作为测试样, 采集后装入塑料袋, 带回实验室。2006-2008年间不同季节每采样植物附近如前述方法采集土壤测试样, 共8次采集土壤测试样, 取其平均值作为采样植物生长的土壤理化因子值。
1.3 材料处理及试验方法
1.3.1 土壤处理及试验方法
土壤样品自然风干、研细、拣去根系, 过18号筛, 贮存供各种分析用。土壤理化指标测定方法按照刘光崧(1996)和鲁如坤(2000)的方法进行: 土壤有机质用K2 Cr2 O7 容量法, 土壤全氮用纳氏试剂比色法, 土壤全磷用磷钼蓝比色法, Cl- 用硝酸银滴定法, 全盐含量用电导法, 土壤pH值用电位法, 土壤Na、Ca、Mg、Al、Mn、K含量用原子吸收分光光度法测定。取样植物所处生境条件的土壤理化因子测定数据见表1 。
图1 海南省东寨港红树林自然保护区研究样地分布图。1至6为采集土壤和木材的6个红海榄种群。
Fig. 1 Map of the study sites in the National Nature Mangrove Reserve of Dongzhai Harbor, Hainan Province. Numbers 1 to 6 represent study sites where wood and soil samples were collected.
1.3.2 木材处理及试验方法
按照标准方法进行制片(Berlyn & Miksche, 1976 )。制片材料取自离形成层约1 cm的木材, 不受潮水冲击的那一面木材作为制片的材料。材料软化后用滑走切片机切取横、径、弦三向切片, 制成永久装片。另取一部分木材用冰醋酸-过氧化氢离析液离析, 制成永久装片。永久装片在光学显微镜(Olympus BHS, OLYMPUS, Tokyo, Japan)及激光共聚焦显微镜(MRC-1024ES, BioRad, Hercules, USA)下观察次生木质部形态特征并拍照, 同时应用Motic Image Advanced 3.2软件测定次生木质部数量特征指标17项。其中, 导管频率、单孔率、输导面积等每项指标测量30组数据, 其余14项指标测量200组以上的数据后计算平均值和标准偏差。
取材料软化后用滑走切片机切取横、径、弦三向切片, 直接用酒精系列脱水, 自然干燥后喷金, 在扫描电镜(S520, Hitachi, Tokyo, Japan)下观察各样地次生木质部形态特征并拍照。
材料软化后木材横向切面依标准方法(Berlyn & Miksche, 1976 )超薄切片后在透射电镜(JSM- 1010, JEOL, Tokyo, Japan)下观察。
本文所用术语及各数量特征的描述和计算依据文献IAWA Committee (1989) 和Carlquist (2001) 提出的标准和计算方法。
1.4 数据计算及统计分析
次生木质部的可塑性指数(PI )参考Ashton等(1998)的计算公式: PI = 1 - x /X , 其中x 为最小平均值, X 为最大平均值。应用SPSS for windows 19.0软件进行逐步回归分析, 揭示土壤理化因子与次生木质部数量特征的关系。
2 结果和分析
2.1 不同样地红海榄次生木质部的形态解剖特征
6个样地红海榄次生木质部的形态解剖特征较为一致, 光学显微镜、激光共聚焦显微镜、扫描电镜观察结果如下: (1)可观察到以窄导管为界的生长轮(图2A )。样地6采样植株次生木质部可明显观察到以窄导管为界的生长轮(图2B )。导管在横切面上由单管孔和管孔群构成(包括复管孔和管孔团)。短径列复管孔、倾斜复管孔和管孔团是横切面上常见的管孔群类型(图2A -2C), 横切面材料和离析材料综合观察表明, 长径列复管孔和管孔团由宽、窄导管或环管管胞组成(图2D ), 窄导管含部分纤维状导管分子(图2E ); (2)导管分子端壁甚为倾斜, 导管具梯型穿孔, 梯型穿孔板具厚横隔(图2F ); (3)管间纹孔梯状排列(图2F ), 导管-射线间纹孔为半缘纹孔对, 导管这边的纹孔缘较退化, 纹孔形状圆形, 有些横向或轴向加长呈栅栏状排列, 有些加长纹孔呈一侧复开口(图2G -2J); (4)具环管管胞或分隔环管管胞, 大量不具穿孔的管状分子为韧型纤维, 部分韧型纤维具分隔, 韧型纤维具单纹孔, 胶质纤维易见(图2K ); (5)射线薄壁细胞具晶体(图2L )、淀粉粒(图2M ), 可见射线穿孔细胞(图2N ); 轴向薄壁组织为稀疏傍管薄壁组织, 具淀粉粒; (6)切片材料的扫描电镜观察结果表明, 导管侧壁的微观结构包括纹孔口沟、纹孔口连接沟和螺旋增厚等螺旋雕纹结构(图2O -2P)以及附物结构, 包括附物纹孔、管壁具附物, 穿孔板附物等(图2Q -2R); (7)大部分纹孔膜没有细小的孔洞(图2S ), 少数纹孔膜突起具孔洞, 可能是样品制备时造成的(图2T )。
图2 红海榄次生木质部的形态解剖特征。A, 横切面(LM), 示窄导管为界的生长轮结构。B, 横切面(LSCM), 示窄导管为界的生长轮结构。C, 横切面(LSCM), 示短径列复管孔。D, 离析的环管管胞(SEM)。E, 离析的纤维状导管分子(SEM)。F, 离析的导管分子(SEM), 示导管分子端壁倾斜。G, 横切面(TEM), 示半缘纹孔对和单纹孔对。H, 离析的导管分子(SEM), 示圆形的具缘纹孔或栅栏状排列的纹孔。I, 径切面(SEM), 示圆形的极度退化的具缘纹孔。J, 径切面(SEM), 示一侧复开口的纹孔。K, 横切面(LM), 示胶质纤维。L, 径切面(SEM), 示射线细胞内的晶体。M, 弦切面(SEM), 示射线细胞内的淀粉粒。N, 径切面(SEM), 示射线穿孔细胞。O, 径切面(SEM), 示螺旋增厚。P, 径切面(SEM), 示纹孔口沟和纹孔口连接沟。Q, 弦切面(SEM), 示导管壁附物。R, 弦切面(SEM), 示附物纹孔。S, 弦切面(SEM), 示致密没有孔隙管间纹孔膜结构。T, 径切面(SEM), 示个别纹孔膜具有缝隙。LM, 光学显微镜; SEM, 扫描电子显微镜; TEM, 透射电子显微镜; LSCM, 激光共聚焦显微镜。
Fig. 2 Anatomical characteristics of secondary xylem in Rhizophora stylosa . A, Transverse section (LM), showing ring border delimited by narrower vessels. B, Transverse section (LSCM), showing ring border delimited by significantly narrower vessels. C, Transverse section (LSCM), showing vessels in short radial multiple. D, Macerated vasicentric tracheid (SEM). E, Macerated fibriform vessel element (SEM). F, Macerated vessel element with oblique end wall (SEM). G, Transverse section (TEM), showing haft-bordered pit-pair and simple pit pair. H, Macerated vessel element with round border pits or fence-like pits (SEM). I, Radial section (SEM), showing round remarkably reduced border pits. J, Radial section (SEM), showing long scalariform pits on one side contacted with alternate laternal wall pits (SEM). K, Transverse section (LM), showing gelatinous fibres. L, Radial section (SEM), showing crystals in the ray cells. M, Longitudinal section (SEM), showing starch grains in the ray cells. N, Radial section (SEM), showing perforated ray cell. O, Radial section (SEM), showing helical thickenings. P, Radial section (SEM), showing grooves and grooves interconnecting pit apertures. Q, Longitudinal section (SEM), showing vestured vessel wall. R, Longitudinal section (SEM), showing vestured pits. S, Longitudinal section (SEM), showing non-porous pit membrane. T, Radial section (SEM), an artefactual slit occurs in restricted pit field areas damaged by sample preparation. LM, light microscopy; SEM, scanning electron microscopy; TEM, transmission electron microscopy; LSCM, laser scanning confocal microscopy.
2.2 不同样地红海榄次生木质部数量解剖特征的变动及其与土壤理化因子的关系
表2 为不同样地红海榄次生木质部17个数量解剖特征。由表2 可以看出, 不同样地次生木质部数量解剖特征均表现出一定的可塑性, 单孔率的可塑性最大, 可塑性指数(PI )值为0.54, 管孔密度次之, PI 值为0.45, 导管分子长度的可塑性最小, PI 值为0.20。但同一样地的3株不同标准木的可塑性较小(表2 )。
多重回归分析表明, 土壤全盐含量、Mn2+ 含量、Na+ 含量、Cl- 含量、Ca2+ 含量、有机质含量、全磷含量和土壤pH值显著影响红海榄次生木质部数量解剖特征(表3 )。随着土壤全盐含量的提高, 导管分子长度和PD 呈增大的趋势; 随着土壤Mn2+ 含量的提高, 导管径向直径、50个最大导管径向直径、导管弦向直径、管孔平均面积、50个最大管孔平均面积、纤维长度和纤维壁厚呈减少趋势, 射线宽度呈增大趋势; 随着土壤Na+ 含量的提高, 导管径向直径、50个最大导管径向直径、导管弦向直径、管孔平均面积、50个最大管孔平均面积呈减少的趋势, 射线宽度呈增大趋势; 随着土壤Cl- 含量的提高, 导管弦向直径呈增大趋势; 随着土壤Ca2+ 含量的提高, 管孔密度、射线宽度、射线密度、纤维长度有增大的趋势; 随着土壤有机质含量的提高, 导管弦向直径、管孔平均面积、50个最大管孔平均面积有减少的趋势; 随着土壤全磷含量的提高, 导管弦向直径、管孔平均面积、50个最大管孔平均面积有增大的趋势, 射线宽度和射线密度有减少趋势; 随着土壤pH值的提高, 输导面积有增大的趋势。
3 讨论
3.1 红海榄木材结构的特化对潮间带生境的适应
红海榄是热带或亚热带海岸潮间带生长的木本植物, 中国红树林生态系统的建群种之一, 高盐生境易产生渗透胁迫, 植物木质部必须在强负压下运输水分。植物对特殊生境的适应常表现为具有一些特化的特征, 本研究结果表明, 红海榄不同样地植物次生木质部具有一些共同的形态特征, 能促进水分输导的安全性, 对潮间带生境有高度的适应能力。具体表现在: (1)不同样地的研究植物都存在纤维状导管和环管管胞。环管管胞与导管相伴随, 当导管形成栓塞时, 环管管胞(抗负压, 不易产生空穴而形成栓塞)提供一个旁道使水分向上运输的功能不至于丧失(Carlquist, 2001 )。同时不同样地的红海榄还有纤维状导管, Carlquist (2001) 认为: 纤维状导管在促进水分输导安全性方面与管胞一样有效; (2) 许多导管壁的微观结构有利于水分输导的安全性。Carlquist (2001) 及Kohonen和Helland (2009)认为: 螺旋雕纹、附物等导管壁结构能增进水分的聚合力, 防止栓塞, 不同样地的研究植物都具有较显著的附物和螺旋雕纹, 可能是保证水分输导安全性的结构; (3)许多红树植物(杯萼海桑、银叶树、 角果木、木榄、木果楝和榄李)具有生长轮已有报道, 研究表明红树植物生长轮的形成是土壤水溶液盐度在一年内周期性变动的结果(Robert et al ., 2011 )。本研究观察表明, 红海榄具有生长轮, 且生长轮是以密度大、管孔小的导管为界, 使红海榄1个生长轮内具宽、窄导管并存的特点, Tyree和Zimmermann (2002) 认为: 宽窄导管并存可以同时兼顾水分输导的安全性和有效性。同时, 也可观察到红海榄的复管孔由宽导管和窄导管共同组成(图2A -2C)。因此, 生长轮以窄导管为界, 主要是时间序列上对土壤水溶液盐度波动(如一年中的旱雨季)的适应, 而复管孔宽窄导管并存及比例变动则更可能是空间序列上(不同生长环境土壤水溶液盐度波动)的适应; (4)薄壁细胞(包括射线细胞和轴向薄壁细胞)内具大量淀粉粒。已有研究表明木质部薄壁细胞内淀粉粒水解有助于高负压下栓塞导管的修复(Salleo et al ., 2009 ; Zwieniecki & Holbrook, 2009 ; Nardini et al ., 2011 ; Secchi & Zwieniecki, 2011 )。
综上所述, 红树植物生长在高盐、水淹的独特的红树林生境, 红树植物在适应异质生境的过程中, 其形态结构必然会发生特化。本研究中红海榄木材结构受高盐等土壤理化因子影响严重, 遭受“生理干旱”胁迫明显, 依据Turner (1979) 对植物耐旱方式的分类, 红海榄既是高水势下耐旱型植物, 也是低水势下耐旱型植物, 上述的(1)、(2)和(3)表明红海榄具有与抗“生理干旱” (盐适应性相关)的形态结构, 在高负压下能持续吸收水分以延迟脱水的发生, 保持高的组织水势; 而上述的(4)倾向于支持红海榄为低水势下耐旱型植物, 具有效的溶质积累能力, 使红海榄忍耐脱水能力强。
3.2 不同样地红海榄次生木质部数量特征的变动及生态适应意义
植物生态解剖研究常因取样数量不足而丢失解剖特征变异幅度的全部信息, 或因生态因子的获取不全面而影响分析解剖特征与环境变量之间的关系(费松林等, 1999 )。红树植物许多物种常沿入海口呈带状分布, 取样数量可以很丰富。但生态因子除浸淹频度外, 常以海水或土壤的全盐量为指标表示盐度梯度的变化, 本研究表明, 随土壤全盐量的升高, 管孔密度增大, 而管孔径向直径与弦向直径等指标不受土壤全盐量的影响, 这与Schmitz等(2006) 以土壤水全盐量的变化对红茄苳(Rhizophora mucronata )次生木质部数量特征变动的影响的研究结果是一致的。但是, 与笼统的土壤全盐量相比, 土壤的离子含量更能影响红海榄次生木质部结构的数量特征, 如: 逐步回归分析揭示土壤Mn2+ 含量显著影响导管、射线和纤维组织的多项数量特征指标, 土壤Na+ 含量显著影响导管和射线组织的多项数量特征指标, 其他土壤理化指标, 如土壤全盐量、土壤Cl- 含量、土壤Ca2+ 含量、土壤有机质含量、土壤全磷含量和土壤pH值也显著影响次生木质部结构的数量特征(表3 )。因此, 以往用全盐量为指标探讨红树植物生境因子与木材结构的关系可能存在生态因子获取不全面的问题, 以后的研究应注意避免生态因子信息获取不足的倾向。
一定范围内随着土壤Na+ 和Mn2+ 含量的提高, 木质部导管分子有“小型化”的趋势(导管径向直径、50个最大导管径向直径、导管弦向直径、管孔平均面积、50个最大管孔平均面积呈减少的趋势)。潮间带生境的最主要特点是含有大量的盐离子, 特别是Na+ 含量非常高(表1 ), 高浓度盐离子易造成低水势, 因此红海榄木质部导管内有较强的负压, 且随生境盐分含量的增加, 木质部导管内的负压增强, “小型化”导管抗负压、抗栓塞能力强, 有利于输导的安全性(Carlquist, 2001 ; Tyree & Zimmermann, 2002 )。Mn2+ 与木质部分子形成关系的个体发育研究主要集中在控制实验, 烟草属(Nicotiana )植物体外培养实验表明: Mn2+ 在愈伤组织形成和茎再生的过程中影响木质部分子的数量(导管密度)和木质化的程度(Santandrea et al ., 1998 )。本研究表明土壤Mn2+ 含量影响纤维壁厚, 但逐步回归分析没有揭示土壤Mn2+ 含量与导管密度具有显著相关性。
一定范围内随着土壤Ca2+ 含量的提高, 管孔密度有增大的趋势。功能解剖学的研究表明气体栓塞(embolism)引起导管输水率的丧失与导管密度的大小成线性负相关(Tyree et al ., 1994 )。红树林生境土壤盐度高, 且随着土壤盐度的增大, 植物遭受“生理干旱”越严重, 而随着盐度的提高, 导管密度提高有利于水分输导的安全性。外源Ca2+ 在促进植物体内水分运输, 防止栓塞的功能方面已有许多研究(Plavcová & Hacke, 2011 ), 对杨树个体发育研究表明, 土壤盐胁迫影响植物对Ca2+ 、Mg2+ 和K+ 等矿质营养的吸收, 进而影响植物体内生长素的浓度, 导致导管密度降低(Junghans et al ., 2006 )。综合本研究和前人的研究可以推测, 外源Ca2+ 在次生木质部建成中对形成导管分子的多少有影响, 这种对次生木质部结构数量特征的影响可能是在高盐生境中的适应途径之一。随着土壤Ca2+ 含量的提高, 射线宽度、射线密度有增大的趋势, 射线是次生木质部中的贮藏组织, 射线细胞内的淀粉粒水解有助于高负压下栓塞导管的修复(Salleo et al ., 2009 ; Zwieniecki & Holbrook, 2009 ; Nardini et al ., 2011 ; Secchi & Zwieniecki, 2011 )。因此, Ca2+ 含量的提高, 射线宽度、射线密度有增大的趋势是红海榄在高盐生境下促进水分上升的另一结构适应途径。
在木质部的形成过程中, 季节性的外在因素如阳光、水分、养分及温度会大大修饰组织水平的木质部表型(Alongi et al ., 2005 )。对土壤全磷含量与红海榄木材解剖数量特征的回归分析表明, 随着土壤全磷含量的提高, 导管弦向直径、管孔平均面积、50个最大管孔平均面积有增大的趋势, 这些结果表明一定范围内土壤全磷含量升高, 导管有大型化的趋势。其生态学意义表现为在生境条件较为优越的条件下(如盐度较低、有机营养较丰富的生境中), 大型化的导管有利于提高输导效率, 加快生长。逐步回归分析表明土壤有机质含量与次生木质部数量特征有显著相关性, 土壤有机质包含各种养分元素, 其与红海榄导管数量特征的关系有待于进一步 研究。
对土壤pH值与红海榄木材解剖数量特征的回归分析表明, 输导面积随研究样地内土壤pH值的升高有增大的趋势。宽导管直径和输导面积大是高导管输导效率的基础, 从分析数据及结果可以推测, 红海榄适合生长在低盐度、pH呈中性的生境。
红树林生境有较高的土壤Cl- 含量和土壤K+ 含量。本研究表明随土壤Cl- 含量的升高, 仅导管弦向直径有增大的趋势, 而红海榄次生木质部数量特征指标不受样地土壤K+ 含量变动的影响。形成这种趋势的原因可能是: (1)研究样地土壤Cl- 含量和K+ 含量的变动范围不够大, 不足以反映红海榄次生木质部数量特征的变化; (2)土壤Cl- 和K+ 主要是作为渗透胁迫物质, 没有参与次生形成层的活动, 不影响次生木质部的建成。
4 结论和展望
红海榄生长在高盐生境中, 在适应异质生境的过程中其形态结构发生特化。如红海榄次生木质部具有环管管胞、附物、射线穿孔细胞和分隔木纤维, 从生态-系统演化的观点看, 这是植物在长期适应异质生境中形成的与系统演化相关的性状。尽管红树植物总体上处于高盐和水淹生境, 但红树林生境变动很大, 如沿入海口方向土壤理化因子变动很大, 不同潮间带水淹频次和土壤理化因子变动也很大, 因此, 红树林生境即使在小区域范围内, 也可区划为不同的异质生境。次生木质部形态特征的有无可能是适应大生境的结果, 而次生木质部数量特征的变动(可塑性)则是与小生境相适应的, 因此, 一些形态特征的量化可能更能说明红海榄对异质小生境的适应, 但这些形态特征目前还缺乏量化指标, 如环管管胞、纤维状导管、附物在不同小生境中数量特征的种内变动, 值得利用计算机影像学方法进一步研究(Fonti et al ., 2010 )。不同潮间带小生境中红海榄次生木质部数量特征可塑性大, 对不同盐度的土壤适应范围广, 同时潮间带小生境中红海榄次生木质部数量特征的变动也是受土壤理化因子影响的。如土壤Na+ 和Mn2+ 含量影响导管的宽窄, 土壤Ca2+ 含量影响导管密度, 这些土壤理化因子影响形成层活动, 进而影响次生木质部组成分子的建成的生理机理机制和生态适应意义值得进一步研究, 特别是外源Ca2+ 含量对纹孔膜结构和功能的影响, 在防止植物栓塞方面的机理机制已有许多研究(Herbette & Cochard, 2010 ), 但影响次生木质部组成分子的数量特征, 适应高盐生境, 防止植物栓塞方面的机理机制还没有研究。本研究表明导管壁厚、单孔率、导管聚合度等数量特征的变动与土壤理化因子的变动没有显著相关性, 但值得进一步研究, 因为许多报道表明这些数量特征指标在高盐生境下有调整水分输导的安全性和有效性的作用(Carlquist, 2001 )。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献
文献选项
[1]
Alongi DM Clough BF Robertson AI 2005 ). Nutrient-use efficiency in arid-zone forests of the mangroves Rhizophora stylosa and Avicennia marina.
Aquatic Botany , 82 , 121 -131 .
[本文引用: 1]
[2]
Ashton PMS Olander LP Berlyn GP Thadani R Cameron IR 1998 ). Changes in leaf structure in relation to crown position and tree size of Betula papyrifera within fire-origin stands of interior cedar-hemlock.
Canadian Journal of Botany , 76 , 1180 -1187 .
[3]
Berlyn GP Miksche JP 1976 ). Botanical Microtechnique and Cytochemistry . The Iowa State University Press , Iowa .
[本文引用: 2]
[4]
Carlquist S 2001 ). Comparative Wood Anatomy. 2nd edn
. Springer, Berlin.
[本文引用: 6]
[5]
Deng CY Lin P Guo SZ 2004 a). Study on wood anatomy of Lumnitzera species.
Journal of Xiamen University (Natural Science) , 43 , 406 -411 . (in Chinese with English abstract)
[本文引用: 1]
[邓传远 , 林鹏 , 郭素枝 (2004 a). 榄李属(Lumnitzera)红树植物的木材解剖学研究
. 厦门大学学报(自然科学版) , 43 , 406 -411 . ]
[本文引用: 1]
[6]
Deng CY Lin P Guo SZ 2004 b). Wood structures of some Sonneratia species and their adaptation to intertidal habitats.
Acta Phytoecologica Sinica , 28 , 392 -399 . (in Chinese with English abstract)
[本文引用: 1]
[邓传远 , 林鹏 , 郭素枝 (2004 b). 海桑属红树植物次生木质部解剖特征及其对潮间带生境的适应
. 植物生态学报 , 28 , 392 -399 . ]
[本文引用: 1]
[7]
Fei SL Fang JY Fan YJ Zhao K Liu XJ Cui KM 1999 ). Anatomical characteristics of leaves and woods of Fagus lucida and their relationship to ecological factors in Mountain Fanjingshan, Guizhou, China.
Acta Botanica Sinica , 41 , 1002 -1009 . (in Chinese with English abstract)
[本文引用: 2]
[费松林 , 方精云 , 樊拥军 , 赵坤 , 刘雪皎 , 崔克明 (1999 ). 贵州梵净山亮叶水青冈叶片和木材的解剖学特征及其与生态因子的关系
. 植物学报 , 41 , 1002 -1009 . ]
[本文引用: 2]
[8]
Fonti P von Arx G García-González I Eilmann B Sass- Klaassen U Gärtner H Eckstein D 2010 ). Studying glob- al change through investigation of the plastic responses of xylem anatomy in tree rings.
New Phytologist , 185 , 42 -53 .
[本文引用: 1]
[9]
Herbette S Cochard H 2010 ). Calcium is a major determinant of xylem vulnerability to cavitation.
Plant Physiology , 153 , 1932 -1939 .
[本文引用: 1]
[10]
IAWA Committee 1989 ). IAWA list of microscopic features for hardwood identification: With an appendix on non-anatomical information.
IAWA Bulletin New Series , 10 , 219 -332 .
[本文引用: 1]
[11]
Junghans U Polle A Düchtig P Weiler E Kuhlman B Gruber F Teichmann T 2006 ). Adaptation to high salinity in poplar involves changes in xylem anatomy and auxin physiology.
Plant, Cell & Environment , 29 , 1519 -1531 .
[本文引用: 1]
[12]
Kohonen MM Helland Å 2009 ). On the function of wall sculpturing in xylem conduits.
Journal of Bionic Engineering , 6 , 324 -329 .
[13]
Lin P 1999 ). Mangrove Ecosystem in China . Science Press , Beijing .
[本文引用: 4]
[14]
Liu GS 1996 ). Soil Physical and Chemical Analysis & Description of Soil Profiles . Standards Press of China , Beijing . (in Chinese)
[刘光崧 (1996 ). 土壤理化分析与剖面描述 . 中国标准出版社 , 北京 . ]
[15]
Lu RK 2000 ). Analytical Methods for Soil and Agricultural Chemistry . China Agricultural Science and Technology Press , Beijing . (in Chinese)
[鲁如坤 (2000 ). 土壤农业化学分析方法 . 中国农业科技出版社 , 北京 .]
[16]
Nardini A Lo Gullo MA Salleo S 2011 ). Refilling embolized xylem conduits: Is it a matter of phloem unloading?
Plant Science , 180 , 604 -611 .
[本文引用: 2]
[17]
Plavcová L Hacke UG 2011 ). Heterogeneous distribution of pectin epitopes and calcium in different pit types of four angiosperm species.
New Phytologist , 192 , 885 -897 .
[本文引用: 1]
[18]
Robert EMR Koedam N Beeckman H Schmitz N 2009 ). A safe hydraulic architecture as wood anatomical explanation for the difference in distribution of the mangroves Avicennia and Rhizophora.
Functional Ecology , 23 , 649 -657 .
[本文引用: 1]
[19]
Robert EMR Schmitz N Okello JA Boeren I Beeckman H Koedam N 2011 ). Mangrove growth rings: Fact or fiction?
Trees , 25 , 49 -58 .
[本文引用: 1]
[20]
Salleo S Trifilò P Nardini A Lo Gullo MA 2009 ). Starch-to- sugar conversion in wood parenchyma of field-growing Laurus nobilis plants: A component of the signal pathway for embolism repair?
Functional Plant Biology , 36 , 815 -825 .
[本文引用: 2]
[21]
Santandrea G Schiff S Bennici A 1998 ). Effects of manganese on Nicotiana species cultivated in vitro and characterization of regenerated Mn-tolerant tobacco plants.
Plant Science , 132 , 71 -82 .
[本文引用: 1]
[22]
Schmitz N Jansen S Verheyden A Kairo JG Beeckman H Koedam N 2007 ). Comparative anatomy of intervessel pits in two mangrove species growing along a natural salinity gradient in Gazi Bay, Kenya.
Annals of Botany , 100 , 271 -281 .
[本文引用: 1]
[23]
Schmitz N Verheyden A Beeckman H Kairo JG Koedam N 2006 ). Influence of a salinity gradient on the vessel characters of the mangrove species Rhizophora mucronata.
Annals of Botany , 98 , 1321 -1330 .
[本文引用: 2]
[24]
Secchi F Zwieniecki MA 2011 ). Sensing embolism in xylem vessels: The role of sucrose as a trigger for refilling.
Plant, Cell & Environment , 34 , 514 -524 .
[本文引用: 2]
[25]
Shi GR Cheng XL Liu L Ma CC 2006 ). Anatomical and water physiological plasticity of Grewia biloba var. parviflora leaf and secondary xylem.
Chinese Journal of Applied Ecology , 17 , 1801 -1806 . (in Chinese with English abstract)
[本文引用: 1]
[史刚荣 , 程雪莲 , 刘蕾 , 马成仓 (2006 ). 扁担木叶片和次生木质部解剖和水分生理特征的可塑性
. 应用生态学报 , 17 , 1801 -1806 . ]
[本文引用: 1]
[26]
Shi GR Liu L 2006 ). Secondary xylem anatomic characteristics of dominant plant species in three communities in Xiangshan Mountain, Huaibei, China.
Acta Botanica Yunnanica , 28 , 363 -370 . (in Chinese with English abstract)
[本文引用: 1]
[史刚荣 , 刘蕾 (2006 ). 淮北相山三种群落中优势树种次生木质部的解剖学特征
. 云南植物研究 , 28 , 363 -370 . ]
[本文引用: 1]
[27]
Strauss-Debenedetti S Bazzaz FA 1991 ). Plasticity and acclimation to light in tropical Moraceae of different sucessional positions.
Oecologia , 87 , 377 -387 .
[本文引用: 1]
[28]
Sun Q Lin P 1997 ). Wood structure of Aegiceras corniculatum and its ecological adaptations to salinities.
Hydrobiologia , 352 , 61 -65 .
[本文引用: 2]
[29]
Turner NC 1979 ). Drought resistance and adaptation to water deficits in crop plants. In: Mussell H, Staples RC eds. Stress Physiology in Crop Plants . John Wiley & Sons , New York .
[本文引用: 1]
[30]
Tyree MT Davis SD Cochard H 1994 ). Biophysical perspectives of xylem evolution: Is there a tradeoff of hydraulic efficiency for vulnerability to dysfunction?
IAWA Journal , 15 , 335 -360 .
[本文引用: 1]
[31]
Tyree MT Zimmermann MH 2002 ). Xylem Structure and the Ascent of Sap. 2nd edn
. Springer, Berlin.
[本文引用: 2]
[32]
Verheyden A de Ridder F Schmitz N Beeckman H Koedam N 2005 ). High-resolution time series of vessel density in Kenyan mangrove trees reveal a link with climate.
New Phytologist , 167 , 425 -435 .
[本文引用: 1]
[33]
Yáñez-Espinosa L Terrazas T López-Mata L 2001 ). Effects of flooding on wood and bark anatomy of four species in a mangrove forest community.
Trees , 15 , 91 -97 .
[本文引用: 2]
[34]
Zwieniecki MA Holbrook NM. 2009 ). Confronting Maxwell’s demon: Biophysics of xylem embolism repair.
Trends in Plant Science , 14 , 530 -534 .
[本文引用: 2]
Nutrient-use efficiency in arid-zone forests of the mangroves Rhizophora stylosa and Avicennia marina.
1
2005
... 在木质部的形成过程中, 季节性的外在因素如阳光、水分、养分及温度会大大修饰组织水平的木质部表型(Alongi et al ., 2005 ).对土壤全磷含量与红海榄木材解剖数量特征的回归分析表明, 随着土壤全磷含量的提高, 导管弦向直径、管孔平均面积、50个最大管孔平均面积有增大的趋势, 这些结果表明一定范围内土壤全磷含量升高, 导管有大型化的趋势.其生态学意义表现为在生境条件较为优越的条件下(如盐度较低、有机营养较丰富的生境中), 大型化的导管有利于提高输导效率, 加快生长.逐步回归分析表明土壤有机质含量与次生木质部数量特征有显著相关性, 土壤有机质包含各种养分元素, 其与红海榄导管数量特征的关系有待于进一步 研究. ...
Changes in leaf structure in relation to crown position and tree size of Betula papyrifera within fire-origin stands of interior cedar-hemlock.
0
1998
2
1976
... 按照标准方法进行制片(Berlyn & Miksche, 1976 ).制片材料取自离形成层约1 cm的木材, 不受潮水冲击的那一面木材作为制片的材料.材料软化后用滑走切片机切取横、径、弦三向切片, 制成永久装片.另取一部分木材用冰醋酸-过氧化氢离析液离析, 制成永久装片.永久装片在光学显微镜(Olympus BHS, OLYMPUS, Tokyo, Japan)及激光共聚焦显微镜(MRC-1024ES, BioRad, Hercules, USA)下观察次生木质部形态特征并拍照, 同时应用Motic Image Advanced 3.2软件测定次生木质部数量特征指标17项.其中, 导管频率、单孔率、输导面积等每项指标测量30组数据, 其余14项指标测量200组以上的数据后计算平均值和标准偏差. ...
... 材料软化后木材横向切面依标准方法(Berlyn & Miksche, 1976 )超薄切片后在透射电镜(JSM- 1010, JEOL, Tokyo, Japan)下观察. ...
Comparative Wood Anatomy. 2nd edn
6
2001
... 本文所用术语及各数量特征的描述和计算依据文献IAWA Committee (1989) 和Carlquist (2001) 提出的标准和计算方法. ...
... 红海榄是热带或亚热带海岸潮间带生长的木本植物, 中国红树林生态系统的建群种之一, 高盐生境易产生渗透胁迫, 植物木质部必须在强负压下运输水分.植物对特殊生境的适应常表现为具有一些特化的特征, 本研究结果表明, 红海榄不同样地植物次生木质部具有一些共同的形态特征, 能促进水分输导的安全性, 对潮间带生境有高度的适应能力.具体表现在: (1)不同样地的研究植物都存在纤维状导管和环管管胞.环管管胞与导管相伴随, 当导管形成栓塞时, 环管管胞(抗负压, 不易产生空穴而形成栓塞)提供一个旁道使水分向上运输的功能不至于丧失(Carlquist, 2001 ).同时不同样地的红海榄还有纤维状导管, Carlquist (2001) 认为: 纤维状导管在促进水分输导安全性方面与管胞一样有效; (2) 许多导管壁的微观结构有利于水分输导的安全性.Carlquist (2001) 及Kohonen和Helland (2009)认为: 螺旋雕纹、附物等导管壁结构能增进水分的聚合力, 防止栓塞, 不同样地的研究植物都具有较显著的附物和螺旋雕纹, 可能是保证水分输导安全性的结构; (3)许多红树植物(杯萼海桑、银叶树、 角果木、木榄、木果楝和榄李)具有生长轮已有报道, 研究表明红树植物生长轮的形成是土壤水溶液盐度在一年内周期性变动的结果(Robert et al ., 2011 ).本研究观察表明, 红海榄具有生长轮, 且生长轮是以密度大、管孔小的导管为界, 使红海榄1个生长轮内具宽、窄导管并存的特点, Tyree和Zimmermann (2002) 认为: 宽窄导管并存可以同时兼顾水分输导的安全性和有效性.同时, 也可观察到红海榄的复管孔由宽导管和窄导管共同组成(图2A -2C).因此, 生长轮以窄导管为界, 主要是时间序列上对土壤水溶液盐度波动(如一年中的旱雨季)的适应, 而复管孔宽窄导管并存及比例变动则更可能是空间序列上(不同生长环境土壤水溶液盐度波动)的适应; (4)薄壁细胞(包括射线细胞和轴向薄壁细胞)内具大量淀粉粒.已有研究表明木质部薄壁细胞内淀粉粒水解有助于高负压下栓塞导管的修复(Salleo et al ., 2009 ; Zwieniecki & Holbrook, 2009 ; Nardini et al ., 2011 ; Secchi & Zwieniecki, 2011 ). ...
... ).同时不同样地的红海榄还有纤维状导管, Carlquist (2001) 认为: 纤维状导管在促进水分输导安全性方面与管胞一样有效; (2) 许多导管壁的微观结构有利于水分输导的安全性.Carlquist (2001) 及Kohonen和Helland (2009)认为: 螺旋雕纹、附物等导管壁结构能增进水分的聚合力, 防止栓塞, 不同样地的研究植物都具有较显著的附物和螺旋雕纹, 可能是保证水分输导安全性的结构; (3)许多红树植物(杯萼海桑、银叶树、 角果木、木榄、木果楝和榄李)具有生长轮已有报道, 研究表明红树植物生长轮的形成是土壤水溶液盐度在一年内周期性变动的结果(Robert et al ., 2011 ).本研究观察表明, 红海榄具有生长轮, 且生长轮是以密度大、管孔小的导管为界, 使红海榄1个生长轮内具宽、窄导管并存的特点, Tyree和Zimmermann (2002) 认为: 宽窄导管并存可以同时兼顾水分输导的安全性和有效性.同时, 也可观察到红海榄的复管孔由宽导管和窄导管共同组成(图2A -2C).因此, 生长轮以窄导管为界, 主要是时间序列上对土壤水溶液盐度波动(如一年中的旱雨季)的适应, 而复管孔宽窄导管并存及比例变动则更可能是空间序列上(不同生长环境土壤水溶液盐度波动)的适应; (4)薄壁细胞(包括射线细胞和轴向薄壁细胞)内具大量淀粉粒.已有研究表明木质部薄壁细胞内淀粉粒水解有助于高负压下栓塞导管的修复(Salleo et al ., 2009 ; Zwieniecki & Holbrook, 2009 ; Nardini et al ., 2011 ; Secchi & Zwieniecki, 2011 ). ...
... 认为: 纤维状导管在促进水分输导安全性方面与管胞一样有效; (2) 许多导管壁的微观结构有利于水分输导的安全性.Carlquist (2001) 及Kohonen和Helland (2009)认为: 螺旋雕纹、附物等导管壁结构能增进水分的聚合力, 防止栓塞, 不同样地的研究植物都具有较显著的附物和螺旋雕纹, 可能是保证水分输导安全性的结构; (3)许多红树植物(杯萼海桑、银叶树、 角果木、木榄、木果楝和榄李)具有生长轮已有报道, 研究表明红树植物生长轮的形成是土壤水溶液盐度在一年内周期性变动的结果(Robert et al ., 2011 ).本研究观察表明, 红海榄具有生长轮, 且生长轮是以密度大、管孔小的导管为界, 使红海榄1个生长轮内具宽、窄导管并存的特点, Tyree和Zimmermann (2002) 认为: 宽窄导管并存可以同时兼顾水分输导的安全性和有效性.同时, 也可观察到红海榄的复管孔由宽导管和窄导管共同组成(图2A -2C).因此, 生长轮以窄导管为界, 主要是时间序列上对土壤水溶液盐度波动(如一年中的旱雨季)的适应, 而复管孔宽窄导管并存及比例变动则更可能是空间序列上(不同生长环境土壤水溶液盐度波动)的适应; (4)薄壁细胞(包括射线细胞和轴向薄壁细胞)内具大量淀粉粒.已有研究表明木质部薄壁细胞内淀粉粒水解有助于高负压下栓塞导管的修复(Salleo et al ., 2009 ; Zwieniecki & Holbrook, 2009 ; Nardini et al ., 2011 ; Secchi & Zwieniecki, 2011 ). ...
... 一定范围内随着土壤Na+ 和Mn2+ 含量的提高, 木质部导管分子有“小型化”的趋势(导管径向直径、50个最大导管径向直径、导管弦向直径、管孔平均面积、50个最大管孔平均面积呈减少的趋势).潮间带生境的最主要特点是含有大量的盐离子, 特别是Na+ 含量非常高(表1 ), 高浓度盐离子易造成低水势, 因此红海榄木质部导管内有较强的负压, 且随生境盐分含量的增加, 木质部导管内的负压增强, “小型化”导管抗负压、抗栓塞能力强, 有利于输导的安全性(Carlquist, 2001 ; Tyree & Zimmermann, 2002 ).Mn2+ 与木质部分子形成关系的个体发育研究主要集中在控制实验, 烟草属(Nicotiana )植物体外培养实验表明: Mn2+ 在愈伤组织形成和茎再生的过程中影响木质部分子的数量(导管密度)和木质化的程度(Santandrea et al ., 1998 ).本研究表明土壤Mn2+ 含量影响纤维壁厚, 但逐步回归分析没有揭示土壤Mn2+ 含量与导管密度具有显著相关性. ...
... 红海榄生长在高盐生境中, 在适应异质生境的过程中其形态结构发生特化.如红海榄次生木质部具有环管管胞、附物、射线穿孔细胞和分隔木纤维, 从生态-系统演化的观点看, 这是植物在长期适应异质生境中形成的与系统演化相关的性状.尽管红树植物总体上处于高盐和水淹生境, 但红树林生境变动很大, 如沿入海口方向土壤理化因子变动很大, 不同潮间带水淹频次和土壤理化因子变动也很大, 因此, 红树林生境即使在小区域范围内, 也可区划为不同的异质生境.次生木质部形态特征的有无可能是适应大生境的结果, 而次生木质部数量特征的变动(可塑性)则是与小生境相适应的, 因此, 一些形态特征的量化可能更能说明红海榄对异质小生境的适应, 但这些形态特征目前还缺乏量化指标, 如环管管胞、纤维状导管、附物在不同小生境中数量特征的种内变动, 值得利用计算机影像学方法进一步研究(Fonti et al ., 2010 ).不同潮间带小生境中红海榄次生木质部数量特征可塑性大, 对不同盐度的土壤适应范围广, 同时潮间带小生境中红海榄次生木质部数量特征的变动也是受土壤理化因子影响的.如土壤Na+ 和Mn2+ 含量影响导管的宽窄, 土壤Ca2+ 含量影响导管密度, 这些土壤理化因子影响形成层活动, 进而影响次生木质部组成分子的建成的生理机理机制和生态适应意义值得进一步研究, 特别是外源Ca2+ 含量对纹孔膜结构和功能的影响, 在防止植物栓塞方面的机理机制已有许多研究(Herbette & Cochard, 2010 ), 但影响次生木质部组成分子的数量特征, 适应高盐生境, 防止植物栓塞方面的机理机制还没有研究.本研究表明导管壁厚、单孔率、导管聚合度等数量特征的变动与土壤理化因子的变动没有显著相关性, 但值得进一步研究, 因为许多报道表明这些数量特征指标在高盐生境下有调整水分输导的安全性和有效性的作用(Carlquist, 2001 ). ...
榄李属(Lumnitzera)红树植物的木材解剖学研究
1
2004
... 红树林是生长于热带亚热带海岸潮间带、受到海水周期性浸淹的木本植物群落, 兼具陆地生态和海洋生态特性, 是海岸重要生态关键区, 在维护生物多样性和沿海地区生态安全等方面发挥着重要作用.红海榄(Rhizophora stylosa )主要分布于亚洲和大洋洲, 我国的海南、广东、广西和台湾有自然分布, 作为红树林生态系统的建群种之一, 红海榄群落多存在于泥滩深厚的中内滩, 形成致密而纯一的群落.红海榄独特的胎生现象和形态结构对独特生境(潮汐淹水和高盐生境)的适应已经有很多研究(Lin, 1999 ).植物茎干和枝条易受到环境诸因子变化带来的影响(费松林等, 1999 ; 史刚荣等, 2006 ; 史刚荣和刘蕾, 2006 ), 这种表型可塑性是植物克服环境异质性、适应环境的重要途径(Strauss-Debenedetti & Bazzaz, 1991 ), 因此, 生态解剖学方法可以揭示植物结构与环境相适应的关系.独特的红树林海岸潮间带生境为研究植物生态解剖提供了很好的样地和材料, 关于红树植物木材结构的生态解剖研究已有许多报道, 主要集中在盐度与水淹频度对木材结构的影响.最近红树植物木材结构的生态解剖研究表明, 一些红树植物在干旱季节形成层产生的导管比雨季产生的导管管径略小, 能增进水分输导的安全性, 小的纹孔膜孔径和(或)小的管间纹孔表面积能增进水分输导系统的栓塞阻力.红树植物一些特殊的木材形态结构, 如附物纹孔的结构与功能也有新报道(Sun & Lin, 1997 ; Yáñez-Espinosa et al ., 2001 ; 邓传远等, 2004a , 2004b ; Verheyden et al ., 2005 ; Schmitz et al ., 2006 , 2007 ; Robert et al ., 2009 ), 因此, 木材结构与生境相适应的研究还有广阔的空间, 特别是红树植物生长在陆海交界的生态脆弱带, 其土壤理化因子呈高度动态变化(Lin, 1999 ), 应用更多土壤理化指标探讨其对木材结构的影响很有必要.本研究运用生态解剖学的方法和原理, 探讨了红海榄木材解剖特征与土壤理化因子的关系, 揭示了次生木质部解剖特征可塑性的生态适应意义. ...
榄李属(Lumnitzera)红树植物的木材解剖学研究
1
2004
... 红树林是生长于热带亚热带海岸潮间带、受到海水周期性浸淹的木本植物群落, 兼具陆地生态和海洋生态特性, 是海岸重要生态关键区, 在维护生物多样性和沿海地区生态安全等方面发挥着重要作用.红海榄(Rhizophora stylosa )主要分布于亚洲和大洋洲, 我国的海南、广东、广西和台湾有自然分布, 作为红树林生态系统的建群种之一, 红海榄群落多存在于泥滩深厚的中内滩, 形成致密而纯一的群落.红海榄独特的胎生现象和形态结构对独特生境(潮汐淹水和高盐生境)的适应已经有很多研究(Lin, 1999 ).植物茎干和枝条易受到环境诸因子变化带来的影响(费松林等, 1999 ; 史刚荣等, 2006 ; 史刚荣和刘蕾, 2006 ), 这种表型可塑性是植物克服环境异质性、适应环境的重要途径(Strauss-Debenedetti & Bazzaz, 1991 ), 因此, 生态解剖学方法可以揭示植物结构与环境相适应的关系.独特的红树林海岸潮间带生境为研究植物生态解剖提供了很好的样地和材料, 关于红树植物木材结构的生态解剖研究已有许多报道, 主要集中在盐度与水淹频度对木材结构的影响.最近红树植物木材结构的生态解剖研究表明, 一些红树植物在干旱季节形成层产生的导管比雨季产生的导管管径略小, 能增进水分输导的安全性, 小的纹孔膜孔径和(或)小的管间纹孔表面积能增进水分输导系统的栓塞阻力.红树植物一些特殊的木材形态结构, 如附物纹孔的结构与功能也有新报道(Sun & Lin, 1997 ; Yáñez-Espinosa et al ., 2001 ; 邓传远等, 2004a , 2004b ; Verheyden et al ., 2005 ; Schmitz et al ., 2006 , 2007 ; Robert et al ., 2009 ), 因此, 木材结构与生境相适应的研究还有广阔的空间, 特别是红树植物生长在陆海交界的生态脆弱带, 其土壤理化因子呈高度动态变化(Lin, 1999 ), 应用更多土壤理化指标探讨其对木材结构的影响很有必要.本研究运用生态解剖学的方法和原理, 探讨了红海榄木材解剖特征与土壤理化因子的关系, 揭示了次生木质部解剖特征可塑性的生态适应意义. ...
海桑属红树植物次生木质部解剖特征及其对潮间带生境的适应
1
2004
... 红树林是生长于热带亚热带海岸潮间带、受到海水周期性浸淹的木本植物群落, 兼具陆地生态和海洋生态特性, 是海岸重要生态关键区, 在维护生物多样性和沿海地区生态安全等方面发挥着重要作用.红海榄(Rhizophora stylosa )主要分布于亚洲和大洋洲, 我国的海南、广东、广西和台湾有自然分布, 作为红树林生态系统的建群种之一, 红海榄群落多存在于泥滩深厚的中内滩, 形成致密而纯一的群落.红海榄独特的胎生现象和形态结构对独特生境(潮汐淹水和高盐生境)的适应已经有很多研究(Lin, 1999 ).植物茎干和枝条易受到环境诸因子变化带来的影响(费松林等, 1999 ; 史刚荣等, 2006 ; 史刚荣和刘蕾, 2006 ), 这种表型可塑性是植物克服环境异质性、适应环境的重要途径(Strauss-Debenedetti & Bazzaz, 1991 ), 因此, 生态解剖学方法可以揭示植物结构与环境相适应的关系.独特的红树林海岸潮间带生境为研究植物生态解剖提供了很好的样地和材料, 关于红树植物木材结构的生态解剖研究已有许多报道, 主要集中在盐度与水淹频度对木材结构的影响.最近红树植物木材结构的生态解剖研究表明, 一些红树植物在干旱季节形成层产生的导管比雨季产生的导管管径略小, 能增进水分输导的安全性, 小的纹孔膜孔径和(或)小的管间纹孔表面积能增进水分输导系统的栓塞阻力.红树植物一些特殊的木材形态结构, 如附物纹孔的结构与功能也有新报道(Sun & Lin, 1997 ; Yáñez-Espinosa et al ., 2001 ; 邓传远等, 2004a , 2004b ; Verheyden et al ., 2005 ; Schmitz et al ., 2006 , 2007 ; Robert et al ., 2009 ), 因此, 木材结构与生境相适应的研究还有广阔的空间, 特别是红树植物生长在陆海交界的生态脆弱带, 其土壤理化因子呈高度动态变化(Lin, 1999 ), 应用更多土壤理化指标探讨其对木材结构的影响很有必要.本研究运用生态解剖学的方法和原理, 探讨了红海榄木材解剖特征与土壤理化因子的关系, 揭示了次生木质部解剖特征可塑性的生态适应意义. ...
海桑属红树植物次生木质部解剖特征及其对潮间带生境的适应
1
2004
... 红树林是生长于热带亚热带海岸潮间带、受到海水周期性浸淹的木本植物群落, 兼具陆地生态和海洋生态特性, 是海岸重要生态关键区, 在维护生物多样性和沿海地区生态安全等方面发挥着重要作用.红海榄(Rhizophora stylosa )主要分布于亚洲和大洋洲, 我国的海南、广东、广西和台湾有自然分布, 作为红树林生态系统的建群种之一, 红海榄群落多存在于泥滩深厚的中内滩, 形成致密而纯一的群落.红海榄独特的胎生现象和形态结构对独特生境(潮汐淹水和高盐生境)的适应已经有很多研究(Lin, 1999 ).植物茎干和枝条易受到环境诸因子变化带来的影响(费松林等, 1999 ; 史刚荣等, 2006 ; 史刚荣和刘蕾, 2006 ), 这种表型可塑性是植物克服环境异质性、适应环境的重要途径(Strauss-Debenedetti & Bazzaz, 1991 ), 因此, 生态解剖学方法可以揭示植物结构与环境相适应的关系.独特的红树林海岸潮间带生境为研究植物生态解剖提供了很好的样地和材料, 关于红树植物木材结构的生态解剖研究已有许多报道, 主要集中在盐度与水淹频度对木材结构的影响.最近红树植物木材结构的生态解剖研究表明, 一些红树植物在干旱季节形成层产生的导管比雨季产生的导管管径略小, 能增进水分输导的安全性, 小的纹孔膜孔径和(或)小的管间纹孔表面积能增进水分输导系统的栓塞阻力.红树植物一些特殊的木材形态结构, 如附物纹孔的结构与功能也有新报道(Sun & Lin, 1997 ; Yáñez-Espinosa et al ., 2001 ; 邓传远等, 2004a , 2004b ; Verheyden et al ., 2005 ; Schmitz et al ., 2006 , 2007 ; Robert et al ., 2009 ), 因此, 木材结构与生境相适应的研究还有广阔的空间, 特别是红树植物生长在陆海交界的生态脆弱带, 其土壤理化因子呈高度动态变化(Lin, 1999 ), 应用更多土壤理化指标探讨其对木材结构的影响很有必要.本研究运用生态解剖学的方法和原理, 探讨了红海榄木材解剖特征与土壤理化因子的关系, 揭示了次生木质部解剖特征可塑性的生态适应意义. ...
贵州梵净山亮叶水青冈叶片和木材的解剖学特征及其与生态因子的关系
2
1999
... 红树林是生长于热带亚热带海岸潮间带、受到海水周期性浸淹的木本植物群落, 兼具陆地生态和海洋生态特性, 是海岸重要生态关键区, 在维护生物多样性和沿海地区生态安全等方面发挥着重要作用.红海榄(Rhizophora stylosa )主要分布于亚洲和大洋洲, 我国的海南、广东、广西和台湾有自然分布, 作为红树林生态系统的建群种之一, 红海榄群落多存在于泥滩深厚的中内滩, 形成致密而纯一的群落.红海榄独特的胎生现象和形态结构对独特生境(潮汐淹水和高盐生境)的适应已经有很多研究(Lin, 1999 ).植物茎干和枝条易受到环境诸因子变化带来的影响(费松林等, 1999 ; 史刚荣等, 2006 ; 史刚荣和刘蕾, 2006 ), 这种表型可塑性是植物克服环境异质性、适应环境的重要途径(Strauss-Debenedetti & Bazzaz, 1991 ), 因此, 生态解剖学方法可以揭示植物结构与环境相适应的关系.独特的红树林海岸潮间带生境为研究植物生态解剖提供了很好的样地和材料, 关于红树植物木材结构的生态解剖研究已有许多报道, 主要集中在盐度与水淹频度对木材结构的影响.最近红树植物木材结构的生态解剖研究表明, 一些红树植物在干旱季节形成层产生的导管比雨季产生的导管管径略小, 能增进水分输导的安全性, 小的纹孔膜孔径和(或)小的管间纹孔表面积能增进水分输导系统的栓塞阻力.红树植物一些特殊的木材形态结构, 如附物纹孔的结构与功能也有新报道(Sun & Lin, 1997 ; Yáñez-Espinosa et al ., 2001 ; 邓传远等, 2004a , 2004b ; Verheyden et al ., 2005 ; Schmitz et al ., 2006 , 2007 ; Robert et al ., 2009 ), 因此, 木材结构与生境相适应的研究还有广阔的空间, 特别是红树植物生长在陆海交界的生态脆弱带, 其土壤理化因子呈高度动态变化(Lin, 1999 ), 应用更多土壤理化指标探讨其对木材结构的影响很有必要.本研究运用生态解剖学的方法和原理, 探讨了红海榄木材解剖特征与土壤理化因子的关系, 揭示了次生木质部解剖特征可塑性的生态适应意义. ...
... 植物生态解剖研究常因取样数量不足而丢失解剖特征变异幅度的全部信息, 或因生态因子的获取不全面而影响分析解剖特征与环境变量之间的关系(费松林等, 1999 ).红树植物许多物种常沿入海口呈带状分布, 取样数量可以很丰富.但生态因子除浸淹频度外, 常以海水或土壤的全盐量为指标表示盐度梯度的变化, 本研究表明, 随土壤全盐量的升高, 管孔密度增大, 而管孔径向直径与弦向直径等指标不受土壤全盐量的影响, 这与Schmitz等(2006) 以土壤水全盐量的变化对红茄苳(Rhizophora mucronata )次生木质部数量特征变动的影响的研究结果是一致的.但是, 与笼统的土壤全盐量相比, 土壤的离子含量更能影响红海榄次生木质部结构的数量特征, 如: 逐步回归分析揭示土壤Mn2+ 含量显著影响导管、射线和纤维组织的多项数量特征指标, 土壤Na+ 含量显著影响导管和射线组织的多项数量特征指标, 其他土壤理化指标, 如土壤全盐量、土壤Cl- 含量、土壤Ca2+ 含量、土壤有机质含量、土壤全磷含量和土壤pH值也显著影响次生木质部结构的数量特征(表3 ).因此, 以往用全盐量为指标探讨红树植物生境因子与木材结构的关系可能存在生态因子获取不全面的问题, 以后的研究应注意避免生态因子信息获取不足的倾向. ...
贵州梵净山亮叶水青冈叶片和木材的解剖学特征及其与生态因子的关系
2
1999
... 红树林是生长于热带亚热带海岸潮间带、受到海水周期性浸淹的木本植物群落, 兼具陆地生态和海洋生态特性, 是海岸重要生态关键区, 在维护生物多样性和沿海地区生态安全等方面发挥着重要作用.红海榄(Rhizophora stylosa )主要分布于亚洲和大洋洲, 我国的海南、广东、广西和台湾有自然分布, 作为红树林生态系统的建群种之一, 红海榄群落多存在于泥滩深厚的中内滩, 形成致密而纯一的群落.红海榄独特的胎生现象和形态结构对独特生境(潮汐淹水和高盐生境)的适应已经有很多研究(Lin, 1999 ).植物茎干和枝条易受到环境诸因子变化带来的影响(费松林等, 1999 ; 史刚荣等, 2006 ; 史刚荣和刘蕾, 2006 ), 这种表型可塑性是植物克服环境异质性、适应环境的重要途径(Strauss-Debenedetti & Bazzaz, 1991 ), 因此, 生态解剖学方法可以揭示植物结构与环境相适应的关系.独特的红树林海岸潮间带生境为研究植物生态解剖提供了很好的样地和材料, 关于红树植物木材结构的生态解剖研究已有许多报道, 主要集中在盐度与水淹频度对木材结构的影响.最近红树植物木材结构的生态解剖研究表明, 一些红树植物在干旱季节形成层产生的导管比雨季产生的导管管径略小, 能增进水分输导的安全性, 小的纹孔膜孔径和(或)小的管间纹孔表面积能增进水分输导系统的栓塞阻力.红树植物一些特殊的木材形态结构, 如附物纹孔的结构与功能也有新报道(Sun & Lin, 1997 ; Yáñez-Espinosa et al ., 2001 ; 邓传远等, 2004a , 2004b ; Verheyden et al ., 2005 ; Schmitz et al ., 2006 , 2007 ; Robert et al ., 2009 ), 因此, 木材结构与生境相适应的研究还有广阔的空间, 特别是红树植物生长在陆海交界的生态脆弱带, 其土壤理化因子呈高度动态变化(Lin, 1999 ), 应用更多土壤理化指标探讨其对木材结构的影响很有必要.本研究运用生态解剖学的方法和原理, 探讨了红海榄木材解剖特征与土壤理化因子的关系, 揭示了次生木质部解剖特征可塑性的生态适应意义. ...
... 植物生态解剖研究常因取样数量不足而丢失解剖特征变异幅度的全部信息, 或因生态因子的获取不全面而影响分析解剖特征与环境变量之间的关系(费松林等, 1999 ).红树植物许多物种常沿入海口呈带状分布, 取样数量可以很丰富.但生态因子除浸淹频度外, 常以海水或土壤的全盐量为指标表示盐度梯度的变化, 本研究表明, 随土壤全盐量的升高, 管孔密度增大, 而管孔径向直径与弦向直径等指标不受土壤全盐量的影响, 这与Schmitz等(2006) 以土壤水全盐量的变化对红茄苳(Rhizophora mucronata )次生木质部数量特征变动的影响的研究结果是一致的.但是, 与笼统的土壤全盐量相比, 土壤的离子含量更能影响红海榄次生木质部结构的数量特征, 如: 逐步回归分析揭示土壤Mn2+ 含量显著影响导管、射线和纤维组织的多项数量特征指标, 土壤Na+ 含量显著影响导管和射线组织的多项数量特征指标, 其他土壤理化指标, 如土壤全盐量、土壤Cl- 含量、土壤Ca2+ 含量、土壤有机质含量、土壤全磷含量和土壤pH值也显著影响次生木质部结构的数量特征(表3 ).因此, 以往用全盐量为指标探讨红树植物生境因子与木材结构的关系可能存在生态因子获取不全面的问题, 以后的研究应注意避免生态因子信息获取不足的倾向. ...
Studying glob- al change through investigation of the plastic responses of xylem anatomy in tree rings.
1
2010
... 红海榄生长在高盐生境中, 在适应异质生境的过程中其形态结构发生特化.如红海榄次生木质部具有环管管胞、附物、射线穿孔细胞和分隔木纤维, 从生态-系统演化的观点看, 这是植物在长期适应异质生境中形成的与系统演化相关的性状.尽管红树植物总体上处于高盐和水淹生境, 但红树林生境变动很大, 如沿入海口方向土壤理化因子变动很大, 不同潮间带水淹频次和土壤理化因子变动也很大, 因此, 红树林生境即使在小区域范围内, 也可区划为不同的异质生境.次生木质部形态特征的有无可能是适应大生境的结果, 而次生木质部数量特征的变动(可塑性)则是与小生境相适应的, 因此, 一些形态特征的量化可能更能说明红海榄对异质小生境的适应, 但这些形态特征目前还缺乏量化指标, 如环管管胞、纤维状导管、附物在不同小生境中数量特征的种内变动, 值得利用计算机影像学方法进一步研究(Fonti et al ., 2010 ).不同潮间带小生境中红海榄次生木质部数量特征可塑性大, 对不同盐度的土壤适应范围广, 同时潮间带小生境中红海榄次生木质部数量特征的变动也是受土壤理化因子影响的.如土壤Na+ 和Mn2+ 含量影响导管的宽窄, 土壤Ca2+ 含量影响导管密度, 这些土壤理化因子影响形成层活动, 进而影响次生木质部组成分子的建成的生理机理机制和生态适应意义值得进一步研究, 特别是外源Ca2+ 含量对纹孔膜结构和功能的影响, 在防止植物栓塞方面的机理机制已有许多研究(Herbette & Cochard, 2010 ), 但影响次生木质部组成分子的数量特征, 适应高盐生境, 防止植物栓塞方面的机理机制还没有研究.本研究表明导管壁厚、单孔率、导管聚合度等数量特征的变动与土壤理化因子的变动没有显著相关性, 但值得进一步研究, 因为许多报道表明这些数量特征指标在高盐生境下有调整水分输导的安全性和有效性的作用(Carlquist, 2001 ). ...
Calcium is a major determinant of xylem vulnerability to cavitation.
1
2010
... 红海榄生长在高盐生境中, 在适应异质生境的过程中其形态结构发生特化.如红海榄次生木质部具有环管管胞、附物、射线穿孔细胞和分隔木纤维, 从生态-系统演化的观点看, 这是植物在长期适应异质生境中形成的与系统演化相关的性状.尽管红树植物总体上处于高盐和水淹生境, 但红树林生境变动很大, 如沿入海口方向土壤理化因子变动很大, 不同潮间带水淹频次和土壤理化因子变动也很大, 因此, 红树林生境即使在小区域范围内, 也可区划为不同的异质生境.次生木质部形态特征的有无可能是适应大生境的结果, 而次生木质部数量特征的变动(可塑性)则是与小生境相适应的, 因此, 一些形态特征的量化可能更能说明红海榄对异质小生境的适应, 但这些形态特征目前还缺乏量化指标, 如环管管胞、纤维状导管、附物在不同小生境中数量特征的种内变动, 值得利用计算机影像学方法进一步研究(Fonti et al ., 2010 ).不同潮间带小生境中红海榄次生木质部数量特征可塑性大, 对不同盐度的土壤适应范围广, 同时潮间带小生境中红海榄次生木质部数量特征的变动也是受土壤理化因子影响的.如土壤Na+ 和Mn2+ 含量影响导管的宽窄, 土壤Ca2+ 含量影响导管密度, 这些土壤理化因子影响形成层活动, 进而影响次生木质部组成分子的建成的生理机理机制和生态适应意义值得进一步研究, 特别是外源Ca2+ 含量对纹孔膜结构和功能的影响, 在防止植物栓塞方面的机理机制已有许多研究(Herbette & Cochard, 2010 ), 但影响次生木质部组成分子的数量特征, 适应高盐生境, 防止植物栓塞方面的机理机制还没有研究.本研究表明导管壁厚、单孔率、导管聚合度等数量特征的变动与土壤理化因子的变动没有显著相关性, 但值得进一步研究, 因为许多报道表明这些数量特征指标在高盐生境下有调整水分输导的安全性和有效性的作用(Carlquist, 2001 ). ...
IAWA list of microscopic features for hardwood identification: With an appendix on non-anatomical information.
1
1989
... 本文所用术语及各数量特征的描述和计算依据文献IAWA Committee (1989) 和Carlquist (2001) 提出的标准和计算方法. ...
Adaptation to high salinity in poplar involves changes in xylem anatomy and auxin physiology.
1
2006
... 一定范围内随着土壤Ca2+ 含量的提高, 管孔密度有增大的趋势.功能解剖学的研究表明气体栓塞(embolism)引起导管输水率的丧失与导管密度的大小成线性负相关(Tyree et al ., 1994 ).红树林生境土壤盐度高, 且随着土壤盐度的增大, 植物遭受“生理干旱”越严重, 而随着盐度的提高, 导管密度提高有利于水分输导的安全性.外源Ca2+ 在促进植物体内水分运输, 防止栓塞的功能方面已有许多研究(Plavcová & Hacke, 2011 ), 对杨树个体发育研究表明, 土壤盐胁迫影响植物对Ca2+ 、Mg2+ 和K+ 等矿质营养的吸收, 进而影响植物体内生长素的浓度, 导致导管密度降低(Junghans et al ., 2006 ).综合本研究和前人的研究可以推测, 外源Ca2+ 在次生木质部建成中对形成导管分子的多少有影响, 这种对次生木质部结构数量特征的影响可能是在高盐生境中的适应途径之一.随着土壤Ca2+ 含量的提高, 射线宽度、射线密度有增大的趋势, 射线是次生木质部中的贮藏组织, 射线细胞内的淀粉粒水解有助于高负压下栓塞导管的修复(Salleo et al ., 2009 ; Zwieniecki & Holbrook, 2009 ; Nardini et al ., 2011 ; Secchi & Zwieniecki, 2011 ).因此, Ca2+ 含量的提高, 射线宽度、射线密度有增大的趋势是红海榄在高盐生境下促进水分上升的另一结构适应途径. ...
On the function of wall sculpturing in xylem conduits.
0
2009
4
1999
... 红树林是生长于热带亚热带海岸潮间带、受到海水周期性浸淹的木本植物群落, 兼具陆地生态和海洋生态特性, 是海岸重要生态关键区, 在维护生物多样性和沿海地区生态安全等方面发挥着重要作用.红海榄(Rhizophora stylosa )主要分布于亚洲和大洋洲, 我国的海南、广东、广西和台湾有自然分布, 作为红树林生态系统的建群种之一, 红海榄群落多存在于泥滩深厚的中内滩, 形成致密而纯一的群落.红海榄独特的胎生现象和形态结构对独特生境(潮汐淹水和高盐生境)的适应已经有很多研究(Lin, 1999 ).植物茎干和枝条易受到环境诸因子变化带来的影响(费松林等, 1999 ; 史刚荣等, 2006 ; 史刚荣和刘蕾, 2006 ), 这种表型可塑性是植物克服环境异质性、适应环境的重要途径(Strauss-Debenedetti & Bazzaz, 1991 ), 因此, 生态解剖学方法可以揭示植物结构与环境相适应的关系.独特的红树林海岸潮间带生境为研究植物生态解剖提供了很好的样地和材料, 关于红树植物木材结构的生态解剖研究已有许多报道, 主要集中在盐度与水淹频度对木材结构的影响.最近红树植物木材结构的生态解剖研究表明, 一些红树植物在干旱季节形成层产生的导管比雨季产生的导管管径略小, 能增进水分输导的安全性, 小的纹孔膜孔径和(或)小的管间纹孔表面积能增进水分输导系统的栓塞阻力.红树植物一些特殊的木材形态结构, 如附物纹孔的结构与功能也有新报道(Sun & Lin, 1997 ; Yáñez-Espinosa et al ., 2001 ; 邓传远等, 2004a , 2004b ; Verheyden et al ., 2005 ; Schmitz et al ., 2006 , 2007 ; Robert et al ., 2009 ), 因此, 木材结构与生境相适应的研究还有广阔的空间, 特别是红树植物生长在陆海交界的生态脆弱带, 其土壤理化因子呈高度动态变化(Lin, 1999 ), 应用更多土壤理化指标探讨其对木材结构的影响很有必要.本研究运用生态解剖学的方法和原理, 探讨了红海榄木材解剖特征与土壤理化因子的关系, 揭示了次生木质部解剖特征可塑性的生态适应意义. ...
... ), 因此, 木材结构与生境相适应的研究还有广阔的空间, 特别是红树植物生长在陆海交界的生态脆弱带, 其土壤理化因子呈高度动态变化(Lin, 1999 ), 应用更多土壤理化指标探讨其对木材结构的影响很有必要.本研究运用生态解剖学的方法和原理, 探讨了红海榄木材解剖特征与土壤理化因子的关系, 揭示了次生木质部解剖特征可塑性的生态适应意义. ...
... 东寨港红树林国家自然保护区位于海南省东北部海口美兰区与文昌市交界处, 地理位置110.57°- 110.63° E、19.63°-20.02° N.保护面积达3337.6 hm2 , 红树林面积2065.0 hm2 .区内海拔高度10- 80 m, 坡度3°-7°, 总地势为南高北低.保护区处于热带北缘, 属热带季风气候, 年平均气温23.8 ℃, 极端最高温度38.9 ℃, 极端最低温度3.0 ℃, 海水表层年平均温度25.0 ℃; 全年平均日照2200 h; 年降水量1676. 4 mm, 80%以上降水集中在5-10月, 形成明显的干湿季; 太阳年辐射总量约为462 kJ·cm-2 .该处潮汐属不正规半日潮, 平均潮差约1 m (Lin, 1999 ). ...
... 该保护区自然分布的红树和半红树植物有木榄(Bruguiera gymnorrhiza )、海莲(Bruguiera sexangula )、红海榄、角果木(Ceriops tagal )、秋茄树(Kandelia candel )、榄李(Lumnitzera racemosa )、海漆(Excoecaria agallocha )、蜡烛果(Aegiceras corniculatum )、海榄雌(Avicennia marina )、水椰(Nypa fructicans )、老鼠簕(Acanthus ilicifolius )、小花老鼠簕(Acanthus ebracteatus )、卤蕨(Acrostichum aur- eum )、尖叶卤蕨(Acrostichum speciosum )、海杧果(Cerbera manghas )、黄槿(Hibiscus tiliaceus )、玉蕊(Barringtonia racemosa )等17种, 引种植物包括红树(Rhizophora apiculata )、木果楝(Xylocarpus grana- tum )、海南海桑(Sonneratia × hainanensis )、杯萼海桑(Sonneratia alba )、海桑(Sonneratia caseolaris )、银叶树(Heritiera littoralis )、瓶花木(Scyphiphora hydrophyllacea )、红榄李(Lumnitzera littorea )、桐棉(Thespesia populnea ), 孟加拉的无瓣海桑(Sonner- atia apetala )也被成功地引种到这里, 成为海岸滩涂和河口地带速生快长的一种红树植物.东寨港自然保护区的植物群落有7个主要群系: (1)木榄群系(Formation Bruguiera gymnorrhiza ); (2)红树群系(Formation Rhizophora apiculata ); (3)秋茄群系(Formation Kandelia candel ); (4)桐花树群系(Formation Aegiceras corniculatum ); (5)白骨壤群系(Formation Avicennia marina ); (6)海桑群系(Formation Sonneratia caseolaris ); (7)水椰群系(Formation Nypa fructicans ) (Lin, 1999 ). ...
Refilling embolized xylem conduits: Is it a matter of phloem unloading?
2
2011
... 红海榄是热带或亚热带海岸潮间带生长的木本植物, 中国红树林生态系统的建群种之一, 高盐生境易产生渗透胁迫, 植物木质部必须在强负压下运输水分.植物对特殊生境的适应常表现为具有一些特化的特征, 本研究结果表明, 红海榄不同样地植物次生木质部具有一些共同的形态特征, 能促进水分输导的安全性, 对潮间带生境有高度的适应能力.具体表现在: (1)不同样地的研究植物都存在纤维状导管和环管管胞.环管管胞与导管相伴随, 当导管形成栓塞时, 环管管胞(抗负压, 不易产生空穴而形成栓塞)提供一个旁道使水分向上运输的功能不至于丧失(Carlquist, 2001 ).同时不同样地的红海榄还有纤维状导管, Carlquist (2001) 认为: 纤维状导管在促进水分输导安全性方面与管胞一样有效; (2) 许多导管壁的微观结构有利于水分输导的安全性.Carlquist (2001) 及Kohonen和Helland (2009)认为: 螺旋雕纹、附物等导管壁结构能增进水分的聚合力, 防止栓塞, 不同样地的研究植物都具有较显著的附物和螺旋雕纹, 可能是保证水分输导安全性的结构; (3)许多红树植物(杯萼海桑、银叶树、 角果木、木榄、木果楝和榄李)具有生长轮已有报道, 研究表明红树植物生长轮的形成是土壤水溶液盐度在一年内周期性变动的结果(Robert et al ., 2011 ).本研究观察表明, 红海榄具有生长轮, 且生长轮是以密度大、管孔小的导管为界, 使红海榄1个生长轮内具宽、窄导管并存的特点, Tyree和Zimmermann (2002) 认为: 宽窄导管并存可以同时兼顾水分输导的安全性和有效性.同时, 也可观察到红海榄的复管孔由宽导管和窄导管共同组成(图2A -2C).因此, 生长轮以窄导管为界, 主要是时间序列上对土壤水溶液盐度波动(如一年中的旱雨季)的适应, 而复管孔宽窄导管并存及比例变动则更可能是空间序列上(不同生长环境土壤水溶液盐度波动)的适应; (4)薄壁细胞(包括射线细胞和轴向薄壁细胞)内具大量淀粉粒.已有研究表明木质部薄壁细胞内淀粉粒水解有助于高负压下栓塞导管的修复(Salleo et al ., 2009 ; Zwieniecki & Holbrook, 2009 ; Nardini et al ., 2011 ; Secchi & Zwieniecki, 2011 ). ...
... 一定范围内随着土壤Ca2+ 含量的提高, 管孔密度有增大的趋势.功能解剖学的研究表明气体栓塞(embolism)引起导管输水率的丧失与导管密度的大小成线性负相关(Tyree et al ., 1994 ).红树林生境土壤盐度高, 且随着土壤盐度的增大, 植物遭受“生理干旱”越严重, 而随着盐度的提高, 导管密度提高有利于水分输导的安全性.外源Ca2+ 在促进植物体内水分运输, 防止栓塞的功能方面已有许多研究(Plavcová & Hacke, 2011 ), 对杨树个体发育研究表明, 土壤盐胁迫影响植物对Ca2+ 、Mg2+ 和K+ 等矿质营养的吸收, 进而影响植物体内生长素的浓度, 导致导管密度降低(Junghans et al ., 2006 ).综合本研究和前人的研究可以推测, 外源Ca2+ 在次生木质部建成中对形成导管分子的多少有影响, 这种对次生木质部结构数量特征的影响可能是在高盐生境中的适应途径之一.随着土壤Ca2+ 含量的提高, 射线宽度、射线密度有增大的趋势, 射线是次生木质部中的贮藏组织, 射线细胞内的淀粉粒水解有助于高负压下栓塞导管的修复(Salleo et al ., 2009 ; Zwieniecki & Holbrook, 2009 ; Nardini et al ., 2011 ; Secchi & Zwieniecki, 2011 ).因此, Ca2+ 含量的提高, 射线宽度、射线密度有增大的趋势是红海榄在高盐生境下促进水分上升的另一结构适应途径. ...
Heterogeneous distribution of pectin epitopes and calcium in different pit types of four angiosperm species.
1
2011
... 一定范围内随着土壤Ca2+ 含量的提高, 管孔密度有增大的趋势.功能解剖学的研究表明气体栓塞(embolism)引起导管输水率的丧失与导管密度的大小成线性负相关(Tyree et al ., 1994 ).红树林生境土壤盐度高, 且随着土壤盐度的增大, 植物遭受“生理干旱”越严重, 而随着盐度的提高, 导管密度提高有利于水分输导的安全性.外源Ca2+ 在促进植物体内水分运输, 防止栓塞的功能方面已有许多研究(Plavcová & Hacke, 2011 ), 对杨树个体发育研究表明, 土壤盐胁迫影响植物对Ca2+ 、Mg2+ 和K+ 等矿质营养的吸收, 进而影响植物体内生长素的浓度, 导致导管密度降低(Junghans et al ., 2006 ).综合本研究和前人的研究可以推测, 外源Ca2+ 在次生木质部建成中对形成导管分子的多少有影响, 这种对次生木质部结构数量特征的影响可能是在高盐生境中的适应途径之一.随着土壤Ca2+ 含量的提高, 射线宽度、射线密度有增大的趋势, 射线是次生木质部中的贮藏组织, 射线细胞内的淀粉粒水解有助于高负压下栓塞导管的修复(Salleo et al ., 2009 ; Zwieniecki & Holbrook, 2009 ; Nardini et al ., 2011 ; Secchi & Zwieniecki, 2011 ).因此, Ca2+ 含量的提高, 射线宽度、射线密度有增大的趋势是红海榄在高盐生境下促进水分上升的另一结构适应途径. ...
A safe hydraulic architecture as wood anatomical explanation for the difference in distribution of the mangroves Avicennia and Rhizophora.
1
2009
... 红树林是生长于热带亚热带海岸潮间带、受到海水周期性浸淹的木本植物群落, 兼具陆地生态和海洋生态特性, 是海岸重要生态关键区, 在维护生物多样性和沿海地区生态安全等方面发挥着重要作用.红海榄(Rhizophora stylosa )主要分布于亚洲和大洋洲, 我国的海南、广东、广西和台湾有自然分布, 作为红树林生态系统的建群种之一, 红海榄群落多存在于泥滩深厚的中内滩, 形成致密而纯一的群落.红海榄独特的胎生现象和形态结构对独特生境(潮汐淹水和高盐生境)的适应已经有很多研究(Lin, 1999 ).植物茎干和枝条易受到环境诸因子变化带来的影响(费松林等, 1999 ; 史刚荣等, 2006 ; 史刚荣和刘蕾, 2006 ), 这种表型可塑性是植物克服环境异质性、适应环境的重要途径(Strauss-Debenedetti & Bazzaz, 1991 ), 因此, 生态解剖学方法可以揭示植物结构与环境相适应的关系.独特的红树林海岸潮间带生境为研究植物生态解剖提供了很好的样地和材料, 关于红树植物木材结构的生态解剖研究已有许多报道, 主要集中在盐度与水淹频度对木材结构的影响.最近红树植物木材结构的生态解剖研究表明, 一些红树植物在干旱季节形成层产生的导管比雨季产生的导管管径略小, 能增进水分输导的安全性, 小的纹孔膜孔径和(或)小的管间纹孔表面积能增进水分输导系统的栓塞阻力.红树植物一些特殊的木材形态结构, 如附物纹孔的结构与功能也有新报道(Sun & Lin, 1997 ; Yáñez-Espinosa et al ., 2001 ; 邓传远等, 2004a , 2004b ; Verheyden et al ., 2005 ; Schmitz et al ., 2006 , 2007 ; Robert et al ., 2009 ), 因此, 木材结构与生境相适应的研究还有广阔的空间, 特别是红树植物生长在陆海交界的生态脆弱带, 其土壤理化因子呈高度动态变化(Lin, 1999 ), 应用更多土壤理化指标探讨其对木材结构的影响很有必要.本研究运用生态解剖学的方法和原理, 探讨了红海榄木材解剖特征与土壤理化因子的关系, 揭示了次生木质部解剖特征可塑性的生态适应意义. ...
Mangrove growth rings: Fact or fiction?
1
2011
... 红海榄是热带或亚热带海岸潮间带生长的木本植物, 中国红树林生态系统的建群种之一, 高盐生境易产生渗透胁迫, 植物木质部必须在强负压下运输水分.植物对特殊生境的适应常表现为具有一些特化的特征, 本研究结果表明, 红海榄不同样地植物次生木质部具有一些共同的形态特征, 能促进水分输导的安全性, 对潮间带生境有高度的适应能力.具体表现在: (1)不同样地的研究植物都存在纤维状导管和环管管胞.环管管胞与导管相伴随, 当导管形成栓塞时, 环管管胞(抗负压, 不易产生空穴而形成栓塞)提供一个旁道使水分向上运输的功能不至于丧失(Carlquist, 2001 ).同时不同样地的红海榄还有纤维状导管, Carlquist (2001) 认为: 纤维状导管在促进水分输导安全性方面与管胞一样有效; (2) 许多导管壁的微观结构有利于水分输导的安全性.Carlquist (2001) 及Kohonen和Helland (2009)认为: 螺旋雕纹、附物等导管壁结构能增进水分的聚合力, 防止栓塞, 不同样地的研究植物都具有较显著的附物和螺旋雕纹, 可能是保证水分输导安全性的结构; (3)许多红树植物(杯萼海桑、银叶树、 角果木、木榄、木果楝和榄李)具有生长轮已有报道, 研究表明红树植物生长轮的形成是土壤水溶液盐度在一年内周期性变动的结果(Robert et al ., 2011 ).本研究观察表明, 红海榄具有生长轮, 且生长轮是以密度大、管孔小的导管为界, 使红海榄1个生长轮内具宽、窄导管并存的特点, Tyree和Zimmermann (2002) 认为: 宽窄导管并存可以同时兼顾水分输导的安全性和有效性.同时, 也可观察到红海榄的复管孔由宽导管和窄导管共同组成(图2A -2C).因此, 生长轮以窄导管为界, 主要是时间序列上对土壤水溶液盐度波动(如一年中的旱雨季)的适应, 而复管孔宽窄导管并存及比例变动则更可能是空间序列上(不同生长环境土壤水溶液盐度波动)的适应; (4)薄壁细胞(包括射线细胞和轴向薄壁细胞)内具大量淀粉粒.已有研究表明木质部薄壁细胞内淀粉粒水解有助于高负压下栓塞导管的修复(Salleo et al ., 2009 ; Zwieniecki & Holbrook, 2009 ; Nardini et al ., 2011 ; Secchi & Zwieniecki, 2011 ). ...
Starch-to- sugar conversion in wood parenchyma of field-growing Laurus nobilis plants: A component of the signal pathway for embolism repair?
2
2009
... 红海榄是热带或亚热带海岸潮间带生长的木本植物, 中国红树林生态系统的建群种之一, 高盐生境易产生渗透胁迫, 植物木质部必须在强负压下运输水分.植物对特殊生境的适应常表现为具有一些特化的特征, 本研究结果表明, 红海榄不同样地植物次生木质部具有一些共同的形态特征, 能促进水分输导的安全性, 对潮间带生境有高度的适应能力.具体表现在: (1)不同样地的研究植物都存在纤维状导管和环管管胞.环管管胞与导管相伴随, 当导管形成栓塞时, 环管管胞(抗负压, 不易产生空穴而形成栓塞)提供一个旁道使水分向上运输的功能不至于丧失(Carlquist, 2001 ).同时不同样地的红海榄还有纤维状导管, Carlquist (2001) 认为: 纤维状导管在促进水分输导安全性方面与管胞一样有效; (2) 许多导管壁的微观结构有利于水分输导的安全性.Carlquist (2001) 及Kohonen和Helland (2009)认为: 螺旋雕纹、附物等导管壁结构能增进水分的聚合力, 防止栓塞, 不同样地的研究植物都具有较显著的附物和螺旋雕纹, 可能是保证水分输导安全性的结构; (3)许多红树植物(杯萼海桑、银叶树、 角果木、木榄、木果楝和榄李)具有生长轮已有报道, 研究表明红树植物生长轮的形成是土壤水溶液盐度在一年内周期性变动的结果(Robert et al ., 2011 ).本研究观察表明, 红海榄具有生长轮, 且生长轮是以密度大、管孔小的导管为界, 使红海榄1个生长轮内具宽、窄导管并存的特点, Tyree和Zimmermann (2002) 认为: 宽窄导管并存可以同时兼顾水分输导的安全性和有效性.同时, 也可观察到红海榄的复管孔由宽导管和窄导管共同组成(图2A -2C).因此, 生长轮以窄导管为界, 主要是时间序列上对土壤水溶液盐度波动(如一年中的旱雨季)的适应, 而复管孔宽窄导管并存及比例变动则更可能是空间序列上(不同生长环境土壤水溶液盐度波动)的适应; (4)薄壁细胞(包括射线细胞和轴向薄壁细胞)内具大量淀粉粒.已有研究表明木质部薄壁细胞内淀粉粒水解有助于高负压下栓塞导管的修复(Salleo et al ., 2009 ; Zwieniecki & Holbrook, 2009 ; Nardini et al ., 2011 ; Secchi & Zwieniecki, 2011 ). ...
... 一定范围内随着土壤Ca2+ 含量的提高, 管孔密度有增大的趋势.功能解剖学的研究表明气体栓塞(embolism)引起导管输水率的丧失与导管密度的大小成线性负相关(Tyree et al ., 1994 ).红树林生境土壤盐度高, 且随着土壤盐度的增大, 植物遭受“生理干旱”越严重, 而随着盐度的提高, 导管密度提高有利于水分输导的安全性.外源Ca2+ 在促进植物体内水分运输, 防止栓塞的功能方面已有许多研究(Plavcová & Hacke, 2011 ), 对杨树个体发育研究表明, 土壤盐胁迫影响植物对Ca2+ 、Mg2+ 和K+ 等矿质营养的吸收, 进而影响植物体内生长素的浓度, 导致导管密度降低(Junghans et al ., 2006 ).综合本研究和前人的研究可以推测, 外源Ca2+ 在次生木质部建成中对形成导管分子的多少有影响, 这种对次生木质部结构数量特征的影响可能是在高盐生境中的适应途径之一.随着土壤Ca2+ 含量的提高, 射线宽度、射线密度有增大的趋势, 射线是次生木质部中的贮藏组织, 射线细胞内的淀粉粒水解有助于高负压下栓塞导管的修复(Salleo et al ., 2009 ; Zwieniecki & Holbrook, 2009 ; Nardini et al ., 2011 ; Secchi & Zwieniecki, 2011 ).因此, Ca2+ 含量的提高, 射线宽度、射线密度有增大的趋势是红海榄在高盐生境下促进水分上升的另一结构适应途径. ...
Effects of manganese on Nicotiana species cultivated in vitro and characterization of regenerated Mn-tolerant tobacco plants.
1
1998
... 一定范围内随着土壤Na+ 和Mn2+ 含量的提高, 木质部导管分子有“小型化”的趋势(导管径向直径、50个最大导管径向直径、导管弦向直径、管孔平均面积、50个最大管孔平均面积呈减少的趋势).潮间带生境的最主要特点是含有大量的盐离子, 特别是Na+ 含量非常高(表1 ), 高浓度盐离子易造成低水势, 因此红海榄木质部导管内有较强的负压, 且随生境盐分含量的增加, 木质部导管内的负压增强, “小型化”导管抗负压、抗栓塞能力强, 有利于输导的安全性(Carlquist, 2001 ; Tyree & Zimmermann, 2002 ).Mn2+ 与木质部分子形成关系的个体发育研究主要集中在控制实验, 烟草属(Nicotiana )植物体外培养实验表明: Mn2+ 在愈伤组织形成和茎再生的过程中影响木质部分子的数量(导管密度)和木质化的程度(Santandrea et al ., 1998 ).本研究表明土壤Mn2+ 含量影响纤维壁厚, 但逐步回归分析没有揭示土壤Mn2+ 含量与导管密度具有显著相关性. ...
Comparative anatomy of intervessel pits in two mangrove species growing along a natural salinity gradient in Gazi Bay, Kenya.
1
2007
... 红树林是生长于热带亚热带海岸潮间带、受到海水周期性浸淹的木本植物群落, 兼具陆地生态和海洋生态特性, 是海岸重要生态关键区, 在维护生物多样性和沿海地区生态安全等方面发挥着重要作用.红海榄(Rhizophora stylosa )主要分布于亚洲和大洋洲, 我国的海南、广东、广西和台湾有自然分布, 作为红树林生态系统的建群种之一, 红海榄群落多存在于泥滩深厚的中内滩, 形成致密而纯一的群落.红海榄独特的胎生现象和形态结构对独特生境(潮汐淹水和高盐生境)的适应已经有很多研究(Lin, 1999 ).植物茎干和枝条易受到环境诸因子变化带来的影响(费松林等, 1999 ; 史刚荣等, 2006 ; 史刚荣和刘蕾, 2006 ), 这种表型可塑性是植物克服环境异质性、适应环境的重要途径(Strauss-Debenedetti & Bazzaz, 1991 ), 因此, 生态解剖学方法可以揭示植物结构与环境相适应的关系.独特的红树林海岸潮间带生境为研究植物生态解剖提供了很好的样地和材料, 关于红树植物木材结构的生态解剖研究已有许多报道, 主要集中在盐度与水淹频度对木材结构的影响.最近红树植物木材结构的生态解剖研究表明, 一些红树植物在干旱季节形成层产生的导管比雨季产生的导管管径略小, 能增进水分输导的安全性, 小的纹孔膜孔径和(或)小的管间纹孔表面积能增进水分输导系统的栓塞阻力.红树植物一些特殊的木材形态结构, 如附物纹孔的结构与功能也有新报道(Sun & Lin, 1997 ; Yáñez-Espinosa et al ., 2001 ; 邓传远等, 2004a , 2004b ; Verheyden et al ., 2005 ; Schmitz et al ., 2006 , 2007 ; Robert et al ., 2009 ), 因此, 木材结构与生境相适应的研究还有广阔的空间, 特别是红树植物生长在陆海交界的生态脆弱带, 其土壤理化因子呈高度动态变化(Lin, 1999 ), 应用更多土壤理化指标探讨其对木材结构的影响很有必要.本研究运用生态解剖学的方法和原理, 探讨了红海榄木材解剖特征与土壤理化因子的关系, 揭示了次生木质部解剖特征可塑性的生态适应意义. ...
Influence of a salinity gradient on the vessel characters of the mangrove species Rhizophora mucronata.
2
2006
... 红树林是生长于热带亚热带海岸潮间带、受到海水周期性浸淹的木本植物群落, 兼具陆地生态和海洋生态特性, 是海岸重要生态关键区, 在维护生物多样性和沿海地区生态安全等方面发挥着重要作用.红海榄(Rhizophora stylosa )主要分布于亚洲和大洋洲, 我国的海南、广东、广西和台湾有自然分布, 作为红树林生态系统的建群种之一, 红海榄群落多存在于泥滩深厚的中内滩, 形成致密而纯一的群落.红海榄独特的胎生现象和形态结构对独特生境(潮汐淹水和高盐生境)的适应已经有很多研究(Lin, 1999 ).植物茎干和枝条易受到环境诸因子变化带来的影响(费松林等, 1999 ; 史刚荣等, 2006 ; 史刚荣和刘蕾, 2006 ), 这种表型可塑性是植物克服环境异质性、适应环境的重要途径(Strauss-Debenedetti & Bazzaz, 1991 ), 因此, 生态解剖学方法可以揭示植物结构与环境相适应的关系.独特的红树林海岸潮间带生境为研究植物生态解剖提供了很好的样地和材料, 关于红树植物木材结构的生态解剖研究已有许多报道, 主要集中在盐度与水淹频度对木材结构的影响.最近红树植物木材结构的生态解剖研究表明, 一些红树植物在干旱季节形成层产生的导管比雨季产生的导管管径略小, 能增进水分输导的安全性, 小的纹孔膜孔径和(或)小的管间纹孔表面积能增进水分输导系统的栓塞阻力.红树植物一些特殊的木材形态结构, 如附物纹孔的结构与功能也有新报道(Sun & Lin, 1997 ; Yáñez-Espinosa et al ., 2001 ; 邓传远等, 2004a , 2004b ; Verheyden et al ., 2005 ; Schmitz et al ., 2006 , 2007 ; Robert et al ., 2009 ), 因此, 木材结构与生境相适应的研究还有广阔的空间, 特别是红树植物生长在陆海交界的生态脆弱带, 其土壤理化因子呈高度动态变化(Lin, 1999 ), 应用更多土壤理化指标探讨其对木材结构的影响很有必要.本研究运用生态解剖学的方法和原理, 探讨了红海榄木材解剖特征与土壤理化因子的关系, 揭示了次生木质部解剖特征可塑性的生态适应意义. ...
... 植物生态解剖研究常因取样数量不足而丢失解剖特征变异幅度的全部信息, 或因生态因子的获取不全面而影响分析解剖特征与环境变量之间的关系(费松林等, 1999 ).红树植物许多物种常沿入海口呈带状分布, 取样数量可以很丰富.但生态因子除浸淹频度外, 常以海水或土壤的全盐量为指标表示盐度梯度的变化, 本研究表明, 随土壤全盐量的升高, 管孔密度增大, 而管孔径向直径与弦向直径等指标不受土壤全盐量的影响, 这与Schmitz等(2006) 以土壤水全盐量的变化对红茄苳(Rhizophora mucronata )次生木质部数量特征变动的影响的研究结果是一致的.但是, 与笼统的土壤全盐量相比, 土壤的离子含量更能影响红海榄次生木质部结构的数量特征, 如: 逐步回归分析揭示土壤Mn2+ 含量显著影响导管、射线和纤维组织的多项数量特征指标, 土壤Na+ 含量显著影响导管和射线组织的多项数量特征指标, 其他土壤理化指标, 如土壤全盐量、土壤Cl- 含量、土壤Ca2+ 含量、土壤有机质含量、土壤全磷含量和土壤pH值也显著影响次生木质部结构的数量特征(表3 ).因此, 以往用全盐量为指标探讨红树植物生境因子与木材结构的关系可能存在生态因子获取不全面的问题, 以后的研究应注意避免生态因子信息获取不足的倾向. ...
Sensing embolism in xylem vessels: The role of sucrose as a trigger for refilling.
2
2011
... 红海榄是热带或亚热带海岸潮间带生长的木本植物, 中国红树林生态系统的建群种之一, 高盐生境易产生渗透胁迫, 植物木质部必须在强负压下运输水分.植物对特殊生境的适应常表现为具有一些特化的特征, 本研究结果表明, 红海榄不同样地植物次生木质部具有一些共同的形态特征, 能促进水分输导的安全性, 对潮间带生境有高度的适应能力.具体表现在: (1)不同样地的研究植物都存在纤维状导管和环管管胞.环管管胞与导管相伴随, 当导管形成栓塞时, 环管管胞(抗负压, 不易产生空穴而形成栓塞)提供一个旁道使水分向上运输的功能不至于丧失(Carlquist, 2001 ).同时不同样地的红海榄还有纤维状导管, Carlquist (2001) 认为: 纤维状导管在促进水分输导安全性方面与管胞一样有效; (2) 许多导管壁的微观结构有利于水分输导的安全性.Carlquist (2001) 及Kohonen和Helland (2009)认为: 螺旋雕纹、附物等导管壁结构能增进水分的聚合力, 防止栓塞, 不同样地的研究植物都具有较显著的附物和螺旋雕纹, 可能是保证水分输导安全性的结构; (3)许多红树植物(杯萼海桑、银叶树、 角果木、木榄、木果楝和榄李)具有生长轮已有报道, 研究表明红树植物生长轮的形成是土壤水溶液盐度在一年内周期性变动的结果(Robert et al ., 2011 ).本研究观察表明, 红海榄具有生长轮, 且生长轮是以密度大、管孔小的导管为界, 使红海榄1个生长轮内具宽、窄导管并存的特点, Tyree和Zimmermann (2002) 认为: 宽窄导管并存可以同时兼顾水分输导的安全性和有效性.同时, 也可观察到红海榄的复管孔由宽导管和窄导管共同组成(图2A -2C).因此, 生长轮以窄导管为界, 主要是时间序列上对土壤水溶液盐度波动(如一年中的旱雨季)的适应, 而复管孔宽窄导管并存及比例变动则更可能是空间序列上(不同生长环境土壤水溶液盐度波动)的适应; (4)薄壁细胞(包括射线细胞和轴向薄壁细胞)内具大量淀粉粒.已有研究表明木质部薄壁细胞内淀粉粒水解有助于高负压下栓塞导管的修复(Salleo et al ., 2009 ; Zwieniecki & Holbrook, 2009 ; Nardini et al ., 2011 ; Secchi & Zwieniecki, 2011 ). ...
... 一定范围内随着土壤Ca2+ 含量的提高, 管孔密度有增大的趋势.功能解剖学的研究表明气体栓塞(embolism)引起导管输水率的丧失与导管密度的大小成线性负相关(Tyree et al ., 1994 ).红树林生境土壤盐度高, 且随着土壤盐度的增大, 植物遭受“生理干旱”越严重, 而随着盐度的提高, 导管密度提高有利于水分输导的安全性.外源Ca2+ 在促进植物体内水分运输, 防止栓塞的功能方面已有许多研究(Plavcová & Hacke, 2011 ), 对杨树个体发育研究表明, 土壤盐胁迫影响植物对Ca2+ 、Mg2+ 和K+ 等矿质营养的吸收, 进而影响植物体内生长素的浓度, 导致导管密度降低(Junghans et al ., 2006 ).综合本研究和前人的研究可以推测, 外源Ca2+ 在次生木质部建成中对形成导管分子的多少有影响, 这种对次生木质部结构数量特征的影响可能是在高盐生境中的适应途径之一.随着土壤Ca2+ 含量的提高, 射线宽度、射线密度有增大的趋势, 射线是次生木质部中的贮藏组织, 射线细胞内的淀粉粒水解有助于高负压下栓塞导管的修复(Salleo et al ., 2009 ; Zwieniecki & Holbrook, 2009 ; Nardini et al ., 2011 ; Secchi & Zwieniecki, 2011 ).因此, Ca2+ 含量的提高, 射线宽度、射线密度有增大的趋势是红海榄在高盐生境下促进水分上升的另一结构适应途径. ...
扁担木叶片和次生木质部解剖和水分生理特征的可塑性
1
2006
... 红树林是生长于热带亚热带海岸潮间带、受到海水周期性浸淹的木本植物群落, 兼具陆地生态和海洋生态特性, 是海岸重要生态关键区, 在维护生物多样性和沿海地区生态安全等方面发挥着重要作用.红海榄(Rhizophora stylosa )主要分布于亚洲和大洋洲, 我国的海南、广东、广西和台湾有自然分布, 作为红树林生态系统的建群种之一, 红海榄群落多存在于泥滩深厚的中内滩, 形成致密而纯一的群落.红海榄独特的胎生现象和形态结构对独特生境(潮汐淹水和高盐生境)的适应已经有很多研究(Lin, 1999 ).植物茎干和枝条易受到环境诸因子变化带来的影响(费松林等, 1999 ; 史刚荣等, 2006 ; 史刚荣和刘蕾, 2006 ), 这种表型可塑性是植物克服环境异质性、适应环境的重要途径(Strauss-Debenedetti & Bazzaz, 1991 ), 因此, 生态解剖学方法可以揭示植物结构与环境相适应的关系.独特的红树林海岸潮间带生境为研究植物生态解剖提供了很好的样地和材料, 关于红树植物木材结构的生态解剖研究已有许多报道, 主要集中在盐度与水淹频度对木材结构的影响.最近红树植物木材结构的生态解剖研究表明, 一些红树植物在干旱季节形成层产生的导管比雨季产生的导管管径略小, 能增进水分输导的安全性, 小的纹孔膜孔径和(或)小的管间纹孔表面积能增进水分输导系统的栓塞阻力.红树植物一些特殊的木材形态结构, 如附物纹孔的结构与功能也有新报道(Sun & Lin, 1997 ; Yáñez-Espinosa et al ., 2001 ; 邓传远等, 2004a , 2004b ; Verheyden et al ., 2005 ; Schmitz et al ., 2006 , 2007 ; Robert et al ., 2009 ), 因此, 木材结构与生境相适应的研究还有广阔的空间, 特别是红树植物生长在陆海交界的生态脆弱带, 其土壤理化因子呈高度动态变化(Lin, 1999 ), 应用更多土壤理化指标探讨其对木材结构的影响很有必要.本研究运用生态解剖学的方法和原理, 探讨了红海榄木材解剖特征与土壤理化因子的关系, 揭示了次生木质部解剖特征可塑性的生态适应意义. ...
扁担木叶片和次生木质部解剖和水分生理特征的可塑性
1
2006
... 红树林是生长于热带亚热带海岸潮间带、受到海水周期性浸淹的木本植物群落, 兼具陆地生态和海洋生态特性, 是海岸重要生态关键区, 在维护生物多样性和沿海地区生态安全等方面发挥着重要作用.红海榄(Rhizophora stylosa )主要分布于亚洲和大洋洲, 我国的海南、广东、广西和台湾有自然分布, 作为红树林生态系统的建群种之一, 红海榄群落多存在于泥滩深厚的中内滩, 形成致密而纯一的群落.红海榄独特的胎生现象和形态结构对独特生境(潮汐淹水和高盐生境)的适应已经有很多研究(Lin, 1999 ).植物茎干和枝条易受到环境诸因子变化带来的影响(费松林等, 1999 ; 史刚荣等, 2006 ; 史刚荣和刘蕾, 2006 ), 这种表型可塑性是植物克服环境异质性、适应环境的重要途径(Strauss-Debenedetti & Bazzaz, 1991 ), 因此, 生态解剖学方法可以揭示植物结构与环境相适应的关系.独特的红树林海岸潮间带生境为研究植物生态解剖提供了很好的样地和材料, 关于红树植物木材结构的生态解剖研究已有许多报道, 主要集中在盐度与水淹频度对木材结构的影响.最近红树植物木材结构的生态解剖研究表明, 一些红树植物在干旱季节形成层产生的导管比雨季产生的导管管径略小, 能增进水分输导的安全性, 小的纹孔膜孔径和(或)小的管间纹孔表面积能增进水分输导系统的栓塞阻力.红树植物一些特殊的木材形态结构, 如附物纹孔的结构与功能也有新报道(Sun & Lin, 1997 ; Yáñez-Espinosa et al ., 2001 ; 邓传远等, 2004a , 2004b ; Verheyden et al ., 2005 ; Schmitz et al ., 2006 , 2007 ; Robert et al ., 2009 ), 因此, 木材结构与生境相适应的研究还有广阔的空间, 特别是红树植物生长在陆海交界的生态脆弱带, 其土壤理化因子呈高度动态变化(Lin, 1999 ), 应用更多土壤理化指标探讨其对木材结构的影响很有必要.本研究运用生态解剖学的方法和原理, 探讨了红海榄木材解剖特征与土壤理化因子的关系, 揭示了次生木质部解剖特征可塑性的生态适应意义. ...
淮北相山三种群落中优势树种次生木质部的解剖学特征
1
2006
... 红树林是生长于热带亚热带海岸潮间带、受到海水周期性浸淹的木本植物群落, 兼具陆地生态和海洋生态特性, 是海岸重要生态关键区, 在维护生物多样性和沿海地区生态安全等方面发挥着重要作用.红海榄(Rhizophora stylosa )主要分布于亚洲和大洋洲, 我国的海南、广东、广西和台湾有自然分布, 作为红树林生态系统的建群种之一, 红海榄群落多存在于泥滩深厚的中内滩, 形成致密而纯一的群落.红海榄独特的胎生现象和形态结构对独特生境(潮汐淹水和高盐生境)的适应已经有很多研究(Lin, 1999 ).植物茎干和枝条易受到环境诸因子变化带来的影响(费松林等, 1999 ; 史刚荣等, 2006 ; 史刚荣和刘蕾, 2006 ), 这种表型可塑性是植物克服环境异质性、适应环境的重要途径(Strauss-Debenedetti & Bazzaz, 1991 ), 因此, 生态解剖学方法可以揭示植物结构与环境相适应的关系.独特的红树林海岸潮间带生境为研究植物生态解剖提供了很好的样地和材料, 关于红树植物木材结构的生态解剖研究已有许多报道, 主要集中在盐度与水淹频度对木材结构的影响.最近红树植物木材结构的生态解剖研究表明, 一些红树植物在干旱季节形成层产生的导管比雨季产生的导管管径略小, 能增进水分输导的安全性, 小的纹孔膜孔径和(或)小的管间纹孔表面积能增进水分输导系统的栓塞阻力.红树植物一些特殊的木材形态结构, 如附物纹孔的结构与功能也有新报道(Sun & Lin, 1997 ; Yáñez-Espinosa et al ., 2001 ; 邓传远等, 2004a , 2004b ; Verheyden et al ., 2005 ; Schmitz et al ., 2006 , 2007 ; Robert et al ., 2009 ), 因此, 木材结构与生境相适应的研究还有广阔的空间, 特别是红树植物生长在陆海交界的生态脆弱带, 其土壤理化因子呈高度动态变化(Lin, 1999 ), 应用更多土壤理化指标探讨其对木材结构的影响很有必要.本研究运用生态解剖学的方法和原理, 探讨了红海榄木材解剖特征与土壤理化因子的关系, 揭示了次生木质部解剖特征可塑性的生态适应意义. ...
淮北相山三种群落中优势树种次生木质部的解剖学特征
1
2006
... 红树林是生长于热带亚热带海岸潮间带、受到海水周期性浸淹的木本植物群落, 兼具陆地生态和海洋生态特性, 是海岸重要生态关键区, 在维护生物多样性和沿海地区生态安全等方面发挥着重要作用.红海榄(Rhizophora stylosa )主要分布于亚洲和大洋洲, 我国的海南、广东、广西和台湾有自然分布, 作为红树林生态系统的建群种之一, 红海榄群落多存在于泥滩深厚的中内滩, 形成致密而纯一的群落.红海榄独特的胎生现象和形态结构对独特生境(潮汐淹水和高盐生境)的适应已经有很多研究(Lin, 1999 ).植物茎干和枝条易受到环境诸因子变化带来的影响(费松林等, 1999 ; 史刚荣等, 2006 ; 史刚荣和刘蕾, 2006 ), 这种表型可塑性是植物克服环境异质性、适应环境的重要途径(Strauss-Debenedetti & Bazzaz, 1991 ), 因此, 生态解剖学方法可以揭示植物结构与环境相适应的关系.独特的红树林海岸潮间带生境为研究植物生态解剖提供了很好的样地和材料, 关于红树植物木材结构的生态解剖研究已有许多报道, 主要集中在盐度与水淹频度对木材结构的影响.最近红树植物木材结构的生态解剖研究表明, 一些红树植物在干旱季节形成层产生的导管比雨季产生的导管管径略小, 能增进水分输导的安全性, 小的纹孔膜孔径和(或)小的管间纹孔表面积能增进水分输导系统的栓塞阻力.红树植物一些特殊的木材形态结构, 如附物纹孔的结构与功能也有新报道(Sun & Lin, 1997 ; Yáñez-Espinosa et al ., 2001 ; 邓传远等, 2004a , 2004b ; Verheyden et al ., 2005 ; Schmitz et al ., 2006 , 2007 ; Robert et al ., 2009 ), 因此, 木材结构与生境相适应的研究还有广阔的空间, 特别是红树植物生长在陆海交界的生态脆弱带, 其土壤理化因子呈高度动态变化(Lin, 1999 ), 应用更多土壤理化指标探讨其对木材结构的影响很有必要.本研究运用生态解剖学的方法和原理, 探讨了红海榄木材解剖特征与土壤理化因子的关系, 揭示了次生木质部解剖特征可塑性的生态适应意义. ...
Plasticity and acclimation to light in tropical Moraceae of different sucessional positions.
1
1991
... 红树林是生长于热带亚热带海岸潮间带、受到海水周期性浸淹的木本植物群落, 兼具陆地生态和海洋生态特性, 是海岸重要生态关键区, 在维护生物多样性和沿海地区生态安全等方面发挥着重要作用.红海榄(Rhizophora stylosa )主要分布于亚洲和大洋洲, 我国的海南、广东、广西和台湾有自然分布, 作为红树林生态系统的建群种之一, 红海榄群落多存在于泥滩深厚的中内滩, 形成致密而纯一的群落.红海榄独特的胎生现象和形态结构对独特生境(潮汐淹水和高盐生境)的适应已经有很多研究(Lin, 1999 ).植物茎干和枝条易受到环境诸因子变化带来的影响(费松林等, 1999 ; 史刚荣等, 2006 ; 史刚荣和刘蕾, 2006 ), 这种表型可塑性是植物克服环境异质性、适应环境的重要途径(Strauss-Debenedetti & Bazzaz, 1991 ), 因此, 生态解剖学方法可以揭示植物结构与环境相适应的关系.独特的红树林海岸潮间带生境为研究植物生态解剖提供了很好的样地和材料, 关于红树植物木材结构的生态解剖研究已有许多报道, 主要集中在盐度与水淹频度对木材结构的影响.最近红树植物木材结构的生态解剖研究表明, 一些红树植物在干旱季节形成层产生的导管比雨季产生的导管管径略小, 能增进水分输导的安全性, 小的纹孔膜孔径和(或)小的管间纹孔表面积能增进水分输导系统的栓塞阻力.红树植物一些特殊的木材形态结构, 如附物纹孔的结构与功能也有新报道(Sun & Lin, 1997 ; Yáñez-Espinosa et al ., 2001 ; 邓传远等, 2004a , 2004b ; Verheyden et al ., 2005 ; Schmitz et al ., 2006 , 2007 ; Robert et al ., 2009 ), 因此, 木材结构与生境相适应的研究还有广阔的空间, 特别是红树植物生长在陆海交界的生态脆弱带, 其土壤理化因子呈高度动态变化(Lin, 1999 ), 应用更多土壤理化指标探讨其对木材结构的影响很有必要.本研究运用生态解剖学的方法和原理, 探讨了红海榄木材解剖特征与土壤理化因子的关系, 揭示了次生木质部解剖特征可塑性的生态适应意义. ...
Wood structure of Aegiceras corniculatum and its ecological adaptations to salinities.
2
1997
... 红树林是生长于热带亚热带海岸潮间带、受到海水周期性浸淹的木本植物群落, 兼具陆地生态和海洋生态特性, 是海岸重要生态关键区, 在维护生物多样性和沿海地区生态安全等方面发挥着重要作用.红海榄(Rhizophora stylosa )主要分布于亚洲和大洋洲, 我国的海南、广东、广西和台湾有自然分布, 作为红树林生态系统的建群种之一, 红海榄群落多存在于泥滩深厚的中内滩, 形成致密而纯一的群落.红海榄独特的胎生现象和形态结构对独特生境(潮汐淹水和高盐生境)的适应已经有很多研究(Lin, 1999 ).植物茎干和枝条易受到环境诸因子变化带来的影响(费松林等, 1999 ; 史刚荣等, 2006 ; 史刚荣和刘蕾, 2006 ), 这种表型可塑性是植物克服环境异质性、适应环境的重要途径(Strauss-Debenedetti & Bazzaz, 1991 ), 因此, 生态解剖学方法可以揭示植物结构与环境相适应的关系.独特的红树林海岸潮间带生境为研究植物生态解剖提供了很好的样地和材料, 关于红树植物木材结构的生态解剖研究已有许多报道, 主要集中在盐度与水淹频度对木材结构的影响.最近红树植物木材结构的生态解剖研究表明, 一些红树植物在干旱季节形成层产生的导管比雨季产生的导管管径略小, 能增进水分输导的安全性, 小的纹孔膜孔径和(或)小的管间纹孔表面积能增进水分输导系统的栓塞阻力.红树植物一些特殊的木材形态结构, 如附物纹孔的结构与功能也有新报道(Sun & Lin, 1997 ; Yáñez-Espinosa et al ., 2001 ; 邓传远等, 2004a , 2004b ; Verheyden et al ., 2005 ; Schmitz et al ., 2006 , 2007 ; Robert et al ., 2009 ), 因此, 木材结构与生境相适应的研究还有广阔的空间, 特别是红树植物生长在陆海交界的生态脆弱带, 其土壤理化因子呈高度动态变化(Lin, 1999 ), 应用更多土壤理化指标探讨其对木材结构的影响很有必要.本研究运用生态解剖学的方法和原理, 探讨了红海榄木材解剖特征与土壤理化因子的关系, 揭示了次生木质部解剖特征可塑性的生态适应意义. ...
... 实验材料2008年6月取自海南省东寨港红树林自然保护区6个不连续带状分布的种群, 其种群发育时间大致相同(根据保护区管理处的资料确定), 每一种群取3株标准木的成熟茎干, 用砂纸磨光茎干的横断面, 根据生长轮粗略判断树龄, 截取生长轮一致的茎段用FAA固定.实验材料均取自潮汐浸水时间较为一致的中潮滩(依保护区的观测资料确定).土壤样品采集方法是: 用内径为5 cm的PPT管钻取20-40 cm深的土样(20-40 cm层土样能较好代表红树林土壤理化因子的时空变动(Sun & Lin, 1997 ; Yáñez-Espinosa et al ., 2001 )), 在每一采样植物的四周采集4个土壤样品混合作为测试样, 采集后装入塑料袋, 带回实验室.2006-2008年间不同季节每采样植物附近如前述方法采集土壤测试样, 共8次采集土壤测试样, 取其平均值作为采样植物生长的土壤理化因子值. ...
1
1979
... 综上所述, 红树植物生长在高盐、水淹的独特的红树林生境, 红树植物在适应异质生境的过程中, 其形态结构必然会发生特化.本研究中红海榄木材结构受高盐等土壤理化因子影响严重, 遭受“生理干旱”胁迫明显, 依据Turner (1979) 对植物耐旱方式的分类, 红海榄既是高水势下耐旱型植物, 也是低水势下耐旱型植物, 上述的(1)、(2)和(3)表明红海榄具有与抗“生理干旱” (盐适应性相关)的形态结构, 在高负压下能持续吸收水分以延迟脱水的发生, 保持高的组织水势; 而上述的(4)倾向于支持红海榄为低水势下耐旱型植物, 具有效的溶质积累能力, 使红海榄忍耐脱水能力强. ...
Biophysical perspectives of xylem evolution: Is there a tradeoff of hydraulic efficiency for vulnerability to dysfunction?
1
1994
... 一定范围内随着土壤Ca2+ 含量的提高, 管孔密度有增大的趋势.功能解剖学的研究表明气体栓塞(embolism)引起导管输水率的丧失与导管密度的大小成线性负相关(Tyree et al ., 1994 ).红树林生境土壤盐度高, 且随着土壤盐度的增大, 植物遭受“生理干旱”越严重, 而随着盐度的提高, 导管密度提高有利于水分输导的安全性.外源Ca2+ 在促进植物体内水分运输, 防止栓塞的功能方面已有许多研究(Plavcová & Hacke, 2011 ), 对杨树个体发育研究表明, 土壤盐胁迫影响植物对Ca2+ 、Mg2+ 和K+ 等矿质营养的吸收, 进而影响植物体内生长素的浓度, 导致导管密度降低(Junghans et al ., 2006 ).综合本研究和前人的研究可以推测, 外源Ca2+ 在次生木质部建成中对形成导管分子的多少有影响, 这种对次生木质部结构数量特征的影响可能是在高盐生境中的适应途径之一.随着土壤Ca2+ 含量的提高, 射线宽度、射线密度有增大的趋势, 射线是次生木质部中的贮藏组织, 射线细胞内的淀粉粒水解有助于高负压下栓塞导管的修复(Salleo et al ., 2009 ; Zwieniecki & Holbrook, 2009 ; Nardini et al ., 2011 ; Secchi & Zwieniecki, 2011 ).因此, Ca2+ 含量的提高, 射线宽度、射线密度有增大的趋势是红海榄在高盐生境下促进水分上升的另一结构适应途径. ...
Xylem Structure and the Ascent of Sap. 2nd edn
2
2002
... 红海榄是热带或亚热带海岸潮间带生长的木本植物, 中国红树林生态系统的建群种之一, 高盐生境易产生渗透胁迫, 植物木质部必须在强负压下运输水分.植物对特殊生境的适应常表现为具有一些特化的特征, 本研究结果表明, 红海榄不同样地植物次生木质部具有一些共同的形态特征, 能促进水分输导的安全性, 对潮间带生境有高度的适应能力.具体表现在: (1)不同样地的研究植物都存在纤维状导管和环管管胞.环管管胞与导管相伴随, 当导管形成栓塞时, 环管管胞(抗负压, 不易产生空穴而形成栓塞)提供一个旁道使水分向上运输的功能不至于丧失(Carlquist, 2001 ).同时不同样地的红海榄还有纤维状导管, Carlquist (2001) 认为: 纤维状导管在促进水分输导安全性方面与管胞一样有效; (2) 许多导管壁的微观结构有利于水分输导的安全性.Carlquist (2001) 及Kohonen和Helland (2009)认为: 螺旋雕纹、附物等导管壁结构能增进水分的聚合力, 防止栓塞, 不同样地的研究植物都具有较显著的附物和螺旋雕纹, 可能是保证水分输导安全性的结构; (3)许多红树植物(杯萼海桑、银叶树、 角果木、木榄、木果楝和榄李)具有生长轮已有报道, 研究表明红树植物生长轮的形成是土壤水溶液盐度在一年内周期性变动的结果(Robert et al ., 2011 ).本研究观察表明, 红海榄具有生长轮, 且生长轮是以密度大、管孔小的导管为界, 使红海榄1个生长轮内具宽、窄导管并存的特点, Tyree和Zimmermann (2002) 认为: 宽窄导管并存可以同时兼顾水分输导的安全性和有效性.同时, 也可观察到红海榄的复管孔由宽导管和窄导管共同组成(图2A -2C).因此, 生长轮以窄导管为界, 主要是时间序列上对土壤水溶液盐度波动(如一年中的旱雨季)的适应, 而复管孔宽窄导管并存及比例变动则更可能是空间序列上(不同生长环境土壤水溶液盐度波动)的适应; (4)薄壁细胞(包括射线细胞和轴向薄壁细胞)内具大量淀粉粒.已有研究表明木质部薄壁细胞内淀粉粒水解有助于高负压下栓塞导管的修复(Salleo et al ., 2009 ; Zwieniecki & Holbrook, 2009 ; Nardini et al ., 2011 ; Secchi & Zwieniecki, 2011 ). ...
... 一定范围内随着土壤Na+ 和Mn2+ 含量的提高, 木质部导管分子有“小型化”的趋势(导管径向直径、50个最大导管径向直径、导管弦向直径、管孔平均面积、50个最大管孔平均面积呈减少的趋势).潮间带生境的最主要特点是含有大量的盐离子, 特别是Na+ 含量非常高(表1 ), 高浓度盐离子易造成低水势, 因此红海榄木质部导管内有较强的负压, 且随生境盐分含量的增加, 木质部导管内的负压增强, “小型化”导管抗负压、抗栓塞能力强, 有利于输导的安全性(Carlquist, 2001 ; Tyree & Zimmermann, 2002 ).Mn2+ 与木质部分子形成关系的个体发育研究主要集中在控制实验, 烟草属(Nicotiana )植物体外培养实验表明: Mn2+ 在愈伤组织形成和茎再生的过程中影响木质部分子的数量(导管密度)和木质化的程度(Santandrea et al ., 1998 ).本研究表明土壤Mn2+ 含量影响纤维壁厚, 但逐步回归分析没有揭示土壤Mn2+ 含量与导管密度具有显著相关性. ...
High-resolution time series of vessel density in Kenyan mangrove trees reveal a link with climate.
1
2005
... 红树林是生长于热带亚热带海岸潮间带、受到海水周期性浸淹的木本植物群落, 兼具陆地生态和海洋生态特性, 是海岸重要生态关键区, 在维护生物多样性和沿海地区生态安全等方面发挥着重要作用.红海榄(Rhizophora stylosa )主要分布于亚洲和大洋洲, 我国的海南、广东、广西和台湾有自然分布, 作为红树林生态系统的建群种之一, 红海榄群落多存在于泥滩深厚的中内滩, 形成致密而纯一的群落.红海榄独特的胎生现象和形态结构对独特生境(潮汐淹水和高盐生境)的适应已经有很多研究(Lin, 1999 ).植物茎干和枝条易受到环境诸因子变化带来的影响(费松林等, 1999 ; 史刚荣等, 2006 ; 史刚荣和刘蕾, 2006 ), 这种表型可塑性是植物克服环境异质性、适应环境的重要途径(Strauss-Debenedetti & Bazzaz, 1991 ), 因此, 生态解剖学方法可以揭示植物结构与环境相适应的关系.独特的红树林海岸潮间带生境为研究植物生态解剖提供了很好的样地和材料, 关于红树植物木材结构的生态解剖研究已有许多报道, 主要集中在盐度与水淹频度对木材结构的影响.最近红树植物木材结构的生态解剖研究表明, 一些红树植物在干旱季节形成层产生的导管比雨季产生的导管管径略小, 能增进水分输导的安全性, 小的纹孔膜孔径和(或)小的管间纹孔表面积能增进水分输导系统的栓塞阻力.红树植物一些特殊的木材形态结构, 如附物纹孔的结构与功能也有新报道(Sun & Lin, 1997 ; Yáñez-Espinosa et al ., 2001 ; 邓传远等, 2004a , 2004b ; Verheyden et al ., 2005 ; Schmitz et al ., 2006 , 2007 ; Robert et al ., 2009 ), 因此, 木材结构与生境相适应的研究还有广阔的空间, 特别是红树植物生长在陆海交界的生态脆弱带, 其土壤理化因子呈高度动态变化(Lin, 1999 ), 应用更多土壤理化指标探讨其对木材结构的影响很有必要.本研究运用生态解剖学的方法和原理, 探讨了红海榄木材解剖特征与土壤理化因子的关系, 揭示了次生木质部解剖特征可塑性的生态适应意义. ...
Effects of flooding on wood and bark anatomy of four species in a mangrove forest community.
2
2001
... 红树林是生长于热带亚热带海岸潮间带、受到海水周期性浸淹的木本植物群落, 兼具陆地生态和海洋生态特性, 是海岸重要生态关键区, 在维护生物多样性和沿海地区生态安全等方面发挥着重要作用.红海榄(Rhizophora stylosa )主要分布于亚洲和大洋洲, 我国的海南、广东、广西和台湾有自然分布, 作为红树林生态系统的建群种之一, 红海榄群落多存在于泥滩深厚的中内滩, 形成致密而纯一的群落.红海榄独特的胎生现象和形态结构对独特生境(潮汐淹水和高盐生境)的适应已经有很多研究(Lin, 1999 ).植物茎干和枝条易受到环境诸因子变化带来的影响(费松林等, 1999 ; 史刚荣等, 2006 ; 史刚荣和刘蕾, 2006 ), 这种表型可塑性是植物克服环境异质性、适应环境的重要途径(Strauss-Debenedetti & Bazzaz, 1991 ), 因此, 生态解剖学方法可以揭示植物结构与环境相适应的关系.独特的红树林海岸潮间带生境为研究植物生态解剖提供了很好的样地和材料, 关于红树植物木材结构的生态解剖研究已有许多报道, 主要集中在盐度与水淹频度对木材结构的影响.最近红树植物木材结构的生态解剖研究表明, 一些红树植物在干旱季节形成层产生的导管比雨季产生的导管管径略小, 能增进水分输导的安全性, 小的纹孔膜孔径和(或)小的管间纹孔表面积能增进水分输导系统的栓塞阻力.红树植物一些特殊的木材形态结构, 如附物纹孔的结构与功能也有新报道(Sun & Lin, 1997 ; Yáñez-Espinosa et al ., 2001 ; 邓传远等, 2004a , 2004b ; Verheyden et al ., 2005 ; Schmitz et al ., 2006 , 2007 ; Robert et al ., 2009 ), 因此, 木材结构与生境相适应的研究还有广阔的空间, 特别是红树植物生长在陆海交界的生态脆弱带, 其土壤理化因子呈高度动态变化(Lin, 1999 ), 应用更多土壤理化指标探讨其对木材结构的影响很有必要.本研究运用生态解剖学的方法和原理, 探讨了红海榄木材解剖特征与土壤理化因子的关系, 揭示了次生木质部解剖特征可塑性的生态适应意义. ...
... 实验材料2008年6月取自海南省东寨港红树林自然保护区6个不连续带状分布的种群, 其种群发育时间大致相同(根据保护区管理处的资料确定), 每一种群取3株标准木的成熟茎干, 用砂纸磨光茎干的横断面, 根据生长轮粗略判断树龄, 截取生长轮一致的茎段用FAA固定.实验材料均取自潮汐浸水时间较为一致的中潮滩(依保护区的观测资料确定).土壤样品采集方法是: 用内径为5 cm的PPT管钻取20-40 cm深的土样(20-40 cm层土样能较好代表红树林土壤理化因子的时空变动(Sun & Lin, 1997 ; Yáñez-Espinosa et al ., 2001 )), 在每一采样植物的四周采集4个土壤样品混合作为测试样, 采集后装入塑料袋, 带回实验室.2006-2008年间不同季节每采样植物附近如前述方法采集土壤测试样, 共8次采集土壤测试样, 取其平均值作为采样植物生长的土壤理化因子值. ...
Confronting Maxwell’s demon: Biophysics of xylem embolism repair.
2
2009
... 红海榄是热带或亚热带海岸潮间带生长的木本植物, 中国红树林生态系统的建群种之一, 高盐生境易产生渗透胁迫, 植物木质部必须在强负压下运输水分.植物对特殊生境的适应常表现为具有一些特化的特征, 本研究结果表明, 红海榄不同样地植物次生木质部具有一些共同的形态特征, 能促进水分输导的安全性, 对潮间带生境有高度的适应能力.具体表现在: (1)不同样地的研究植物都存在纤维状导管和环管管胞.环管管胞与导管相伴随, 当导管形成栓塞时, 环管管胞(抗负压, 不易产生空穴而形成栓塞)提供一个旁道使水分向上运输的功能不至于丧失(Carlquist, 2001 ).同时不同样地的红海榄还有纤维状导管, Carlquist (2001) 认为: 纤维状导管在促进水分输导安全性方面与管胞一样有效; (2) 许多导管壁的微观结构有利于水分输导的安全性.Carlquist (2001) 及Kohonen和Helland (2009)认为: 螺旋雕纹、附物等导管壁结构能增进水分的聚合力, 防止栓塞, 不同样地的研究植物都具有较显著的附物和螺旋雕纹, 可能是保证水分输导安全性的结构; (3)许多红树植物(杯萼海桑、银叶树、 角果木、木榄、木果楝和榄李)具有生长轮已有报道, 研究表明红树植物生长轮的形成是土壤水溶液盐度在一年内周期性变动的结果(Robert et al ., 2011 ).本研究观察表明, 红海榄具有生长轮, 且生长轮是以密度大、管孔小的导管为界, 使红海榄1个生长轮内具宽、窄导管并存的特点, Tyree和Zimmermann (2002) 认为: 宽窄导管并存可以同时兼顾水分输导的安全性和有效性.同时, 也可观察到红海榄的复管孔由宽导管和窄导管共同组成(图2A -2C).因此, 生长轮以窄导管为界, 主要是时间序列上对土壤水溶液盐度波动(如一年中的旱雨季)的适应, 而复管孔宽窄导管并存及比例变动则更可能是空间序列上(不同生长环境土壤水溶液盐度波动)的适应; (4)薄壁细胞(包括射线细胞和轴向薄壁细胞)内具大量淀粉粒.已有研究表明木质部薄壁细胞内淀粉粒水解有助于高负压下栓塞导管的修复(Salleo et al ., 2009 ; Zwieniecki & Holbrook, 2009 ; Nardini et al ., 2011 ; Secchi & Zwieniecki, 2011 ). ...
... 一定范围内随着土壤Ca2+ 含量的提高, 管孔密度有增大的趋势.功能解剖学的研究表明气体栓塞(embolism)引起导管输水率的丧失与导管密度的大小成线性负相关(Tyree et al ., 1994 ).红树林生境土壤盐度高, 且随着土壤盐度的增大, 植物遭受“生理干旱”越严重, 而随着盐度的提高, 导管密度提高有利于水分输导的安全性.外源Ca2+ 在促进植物体内水分运输, 防止栓塞的功能方面已有许多研究(Plavcová & Hacke, 2011 ), 对杨树个体发育研究表明, 土壤盐胁迫影响植物对Ca2+ 、Mg2+ 和K+ 等矿质营养的吸收, 进而影响植物体内生长素的浓度, 导致导管密度降低(Junghans et al ., 2006 ).综合本研究和前人的研究可以推测, 外源Ca2+ 在次生木质部建成中对形成导管分子的多少有影响, 这种对次生木质部结构数量特征的影响可能是在高盐生境中的适应途径之一.随着土壤Ca2+ 含量的提高, 射线宽度、射线密度有增大的趋势, 射线是次生木质部中的贮藏组织, 射线细胞内的淀粉粒水解有助于高负压下栓塞导管的修复(Salleo et al ., 2009 ; Zwieniecki & Holbrook, 2009 ; Nardini et al ., 2011 ; Secchi & Zwieniecki, 2011 ).因此, Ca2+ 含量的提高, 射线宽度、射线密度有增大的趋势是红海榄在高盐生境下促进水分上升的另一结构适应途径. ...
 , 郑俊鸣
, 郑俊鸣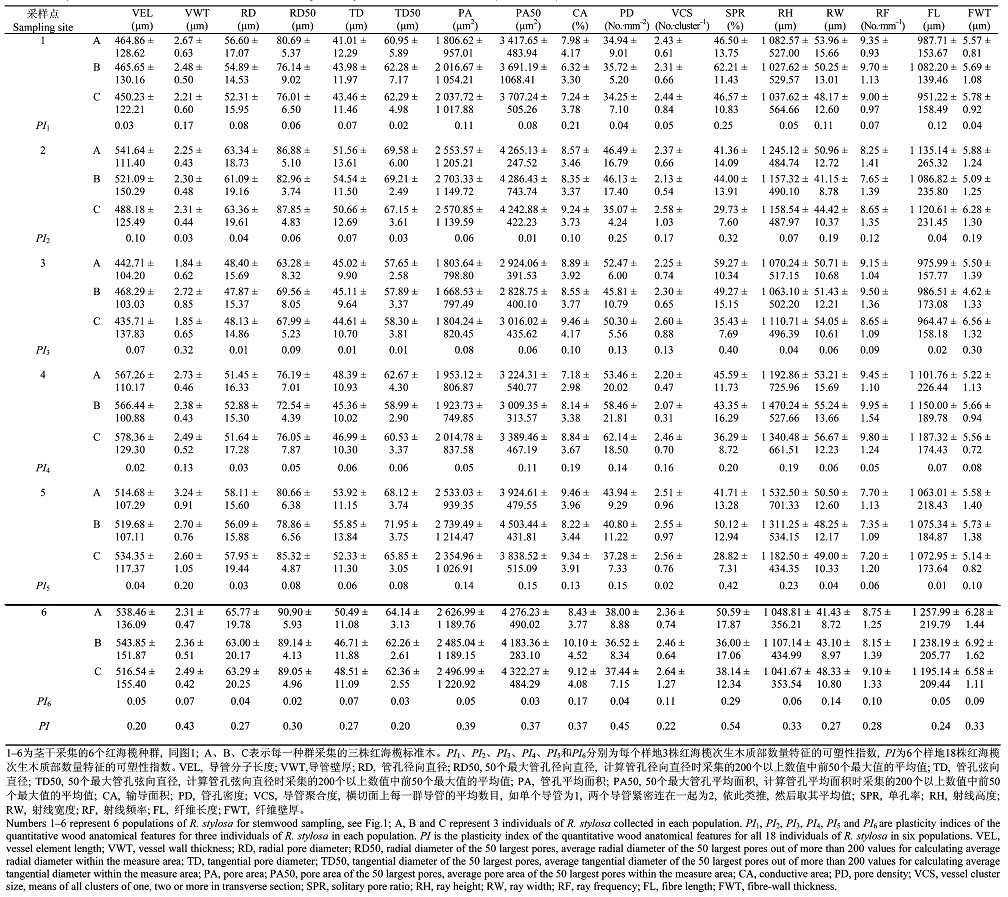



{kind=link}
{kind=link}
{kind=link}
{kind=link}