孙茜, 贺超, 贺学礼 , 赵丽莉
, 赵丽莉
河北大学生命科学学院, 河北保定 071002
SUN Qian, HE Chao, HE Xue-Li, ZHAO Li-Li
通讯作者:
收稿日期: 2015-03-4
接受日期: 2015-08-4
网络出版日期: 2015-09-03
版权声明: 2015 植物生态学报编辑部 本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。
基金资助:
展开
摘要
为了探明荒漠环境深色有隔内生真菌(dark septate endophytes, DSE)生态分布和定殖状况, 于2013年7月从宁夏银川、沙坡头和甘肃民勤地区采集沙冬青(Ammopiptanthus mongolicus)及其伴生植物根围0-10、10-20、20-30、30-40、40-50 cm共5个土层土壤样品和根样, 研究了DSE空间定殖规律及其与土壤因子的相关关系。结果表明: 沙冬青和伴生植物根系能被DSE侵染, DSE分布和定殖具有明显的空间异质性, 并与土壤因子密切相关。沙冬青DSE微菌核结构紧凑, 呈团块状, 而伴生植物DSE微菌核零散分布。同一样地, 沙冬青DSE菌丝定殖率、定殖强度和总定殖率较伴生植物高, 而沙冬青和伴生植物微菌核定殖率无显著差异。不同样地, 沙冬青和伴生植物微菌核定殖率无显著差异, 沙冬青DSE菌丝定殖率、定殖强度和总定殖率为沙坡头>银川>民勤; 伴生植物DSE菌丝定殖率、定殖强度和总定殖率为银川>民勤>沙坡头。通径分析和主成分分析表明, 土壤有机质、总球囊霉素、磷酸酶和速效钾是西北荒漠环境中DSE定殖的主要影响因子。
关键词:
Abstract
Dark septate endophytes (DSE) are commonly detected in plant roots in different ecosystems. However, so far our knowledge about the ecological significance of DSE in semiarid and arid lands is limited. The main objective of this study was to compare the colonization status and ecological distribution of DSE in Ammopiptanthus mongolicus and its associated plants in desert habitats.
In July 2013, soil and root samples of A. mongolicus and its associated plants were collected from three sites, including Yinchuan, Shapotou and Minqin located in northwest China. At each sampling site, samples were collected at five depth intervals, 0-10, 10-20, 20-30, 30-40 and 40-50 cm, respectively.
The roots of A. mongolicus and its associated plants were infected by DSE which could be characterized by typical septate hyphae and microsclerotia. Microsclerotia of A. mongolicus were compact lumps, while the associated plants had scattered microsclerotia. In the same site, microsclerotia colonization in A. mongolicus and its associated plants showed no significant difference, but hyphal colonization, colonization intensity and total colonization of A. mongolicus were much higher than that of its associated plants. In different sites, the colonization intensity and total colonization of DSE in A. mongolicus exhibited a pattern as Shapotou > Yinchuan > Minqin, and these indexes in associated plants were Yinchuan > Minqin > Shapotou. Plant species and soil layer had significant effects on the colonization and distribution of DSE as well as soil properties. The path coefficient and principal component analysis indicated that soil organic matter, total extractable glomalin, phosphatase and available K are the main influencing factors, which directly affected the colonization of DSE in desert environment, Northwest China.
Keywords:
深色有隔内生真菌(dark septate endophytes, DSE)泛指一群菌丝颜色深、有明显横隔、寄居于植物根内的小型真菌, 它们广泛分布于健康植物根表皮、皮层甚至维管组织细胞内或细胞间隙, 形成胞内或胞间深色有隔菌丝、微菌核或泡囊结构, 但不会产生典型的病理学特征。DSE可能具有与菌根真菌类似的生态学功能, 并在不同生态系统中扮演着重要角色(刘茂军等, 2009)。
Szymon等(2012)通过对24属25种药用植物研究发现, 有17种植物能被DSE定殖。Kovács和Szigetvári (2002)通过对半干旱沙化草原89种植物探查发现, 63种植物有DSE深色菌丝, 并在36种植物中检测到微菌核, 因此他们推测DSE在干旱沙漠环境中对植物生长有重要作用。Ruotsalainen等(2004)关于极端高寒植被的研究发现, 随着环境中非生物胁迫增加, DSE与植物相互作用方式的多样性和重要性显著增强。
Dhruba和Chaudhurl (2009)通过对印度水稻(Oryza sativa)根系DSE定殖结构研究发现, DSE横隔菌丝可以产生聚合厚垣孢子、深色厚壁长孢子和含有油滴状内含物的孢子球。姜桥等(2014)研究表明, DSE可与新疆沙冬青(Ammopiptanthus mongolicus)形成良好的根菌联系, 在宿主根皮层细胞间形成菌丝网络和胞内菌丝, 菌丝横隔明显。闫姣等(2014)对内蒙古荒漠北沙柳(Salix psammophila)根系DSE月份间定殖规律研究表明, DSE菌丝形态在月份间无明显区别, 但6月份显著多于8月份和10月份; 不同月份的微菌核形状无明显区别。
沙冬青(Ammopiptanthus mongolicus)是我国西北荒漠地区独有的旱生常绿豆科阔叶灌木, 是砾质及沙砾质荒漠化草原的建群种, 也是第三纪孑遗物种, 具有古老的生长史和发展史。沙冬青具有耐寒、抗旱、根系发达(杨期和等, 2004)等特点, 是防风治沙的理想树种。伴生植物与沙冬青相伴而生, 既有竞争抑制, 又有协同进化。近年来关于DSE真菌侵染沙冬青根组织及其定殖结构时有报道(张淑容等, 2013; 姜桥等, 2014), 但尚未对其伴生植物DSE的定殖规律进行系统研究。
本试验通过野外采样和室内分析, 对比分析荒漠环境沙冬青及其伴生植物DSE形态特征、定殖规律以及土壤因子相关性, 旨在阐明荒漠环境固有种与伴生种DSE定殖和生态分布规律, 为充分利用微生物资源和生物技术促进荒漠植被恢复和生态改良提供依据。
试验样地位于宁夏银川、沙坡头和甘肃民勤, 在每个样地分别随机选取生长良好的沙冬青及其伴生植物各4株, 除去土壤表层落叶, 在距植株主干0-30 cm范围内挖土壤剖面, 按0-10、10-20、20-30、30-40和40-50 cm 5个土层采集土样和根样约1 kg, 记录植物名称、采样地点、土壤类型等(表1)。样品装入隔热性能良好的塑料袋密封, 带回实验室。自然风干后, 过2 mm筛分离根样和土样。土样用于土壤理化性质测定, 将根样剪成1 cm长的根段, 用于DSE形态结构和定殖率观测。
表1 样地和沙冬青伴生植物种类
Table 1 Sampling sites and associated plant species of Ammopiptanthus mongolicus
| 样地 Sampling site | 伴生植物 Associated plants | 海拔 Elevation (m) | 经纬度 Latitude and longitude | 土壤类型 Type of soil |
|---|---|---|---|---|
| 宁夏银川 Yinchuan, Ningxia | 白刺 Nitraria tangutorum 沙蒿 Artemisia desertorum | 1 169 | 38.6° N, 106.57° E | 石质为主 Stony mainly |
| 宁夏沙坡头 Shapotou, Ningxia | 柠条锦鸡儿 Caragana korshinskii | 1 298 | 37.57° N, 104.97° E | 沙土为主 Sand mainly |
| 甘肃民勤 Minqin, Gansu | 无叶沙拐枣 Calligonum aphyllum 红花锦鸡儿 Caragana rosea | 1 350 | 38.58° N, 102.93° E | 荒漠沙质 Sandy desert |
用灼烧质量法测定土壤有机质, 碱解扩散法测定土壤碱解氮(N), 用0.5 mol·L-1 NaHCO3钼锑抗比色法测定土壤速效磷(P), 用火焰光度法测定土壤速效钾(K) (鲁如坤, 2000)。用改进的Tabatabai和Brimner法(宋勇春等, 2001)测定土壤磷酸酶。按Wright和Upadhyaya (1998)的方法测定球囊霉素。用pH 8.0的0.05 mol·L-1柠檬酸钠浸提液在121 ℃条件下所能提取的最大量球囊霉素即为总球囊霉素(TEG); 用pH 7.0的0.02 mol·L-1柠檬酸钠浸提液在121 ℃、30 min内获取的球囊霉素即为易提取球囊霉素(EEG)。
将根样放入装有10% KOH溶液的试管中, 于100 ℃恒温水浴中解离1 h。根样经0.5%酸性品红染色、乳酸甘油脱色(Zhao & He, 2007)后, 每一样本随机选取30个1 cm长的根段, 在显微镜下观察DSE形态结构和侵染状况, 统计根段中DSE菌丝、微菌核侵染根段数, 并计算每一根样DSE定殖率和定殖强度, 计算公式为:
定殖率(%) =定殖根段数/被镜检的根段总数×100%
定殖强度(%) = 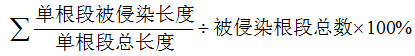
采用SPASS 19.0软件对DSE定殖情况和土壤因子进行单因素方差分析(one-way ANOVA)、土壤因子主成分分析, 并对DSE总定殖率与土壤因子间的关系进行通径分析。所有测定值均为平均值±标准偏差(mean ± SD)。
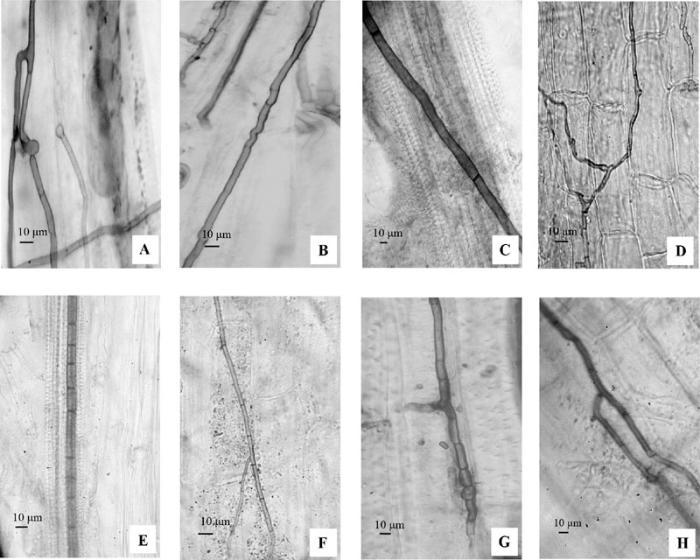
观测结果表明, DSE侵染沙冬青和伴生植物根系, 并在根组织形成菌丝网络(图1A、1D)、胞内菌丝(图1A、1G)和胞间菌丝(图1D、1F、1H), 并侵入维管组织(图1C、1E), 偶尔可见节间缩短的特化菌丝(图E), 菌丝颜色较深, 横隔明显缩短, 随后这类菌丝在皮层细胞或维管组织内(图2C、2E)堆积形成圆形膨大的厚垣细胞群, 称为微菌核。常见的有“脑”状(图2A、2G)、链状(图2B)、带状(图2E、2K)、叶状(图2D、2F、2J)。
图1 沙冬青及其伴生植物的深色有隔内生真菌(DSE)菌丝结构。 A, 银川沙冬青DSE菌丝。B, 沙坡头沙冬青DSE菌丝。C, 民勤沙冬青DSE菌丝侵入维管组织。D, 银川沙蒿DSE菌丝。E, 银川白刺DSE菌丝侵入维管组织。F, 沙坡头柠条锦鸡儿DSE菌丝。G, 民勤无叶沙拐枣DSE菌丝。H, 民勤红花锦鸡儿DSE菌丝。
Fig. 1 Hyphal structure of dark septate endophytes (DSE) in roots of Ammopiptanthus mongolicus and its associated plants. A, Hyphae of DSE in A. mongolicus in Yinchuan. B, Hyphae of DSE in A. mongolicus in Shapotou. C, Infection of DSE in vascular tissue of A. mongolicus in Minqin. D, Hyphae of DSE in Artemisia desertorum in Yinchuan. E, Infection of DSE in vascular tissue of Nitraria tangutorum in Yinchuan. F, Hyphae of DSE in Caragana korshinskii in Shapotou. G, Hyphae of DSE in Calligonum aphyllum in Minqin. H, Hyphae of DSE in C. rosea in Minqin.
图2 沙冬青及其伴生植物的深色有隔内生真菌(DSE)微菌核结构特征。A, 银川沙冬青DSE脑状微菌核。B, 银川沙冬青DSE链状微菌核。C, 沙坡头沙冬青DSE微菌核群。D, 沙坡头沙冬青DSE叶状微菌核。E, 民勤沙冬青DSE带状微菌核。F, 民勤沙冬青DSE叶状微菌核。G, 银川白刺DSE脑状微菌核。H, 银川沙蒿DSE零散状微菌核。I, 沙坡头柠条锦鸡儿DSE菌丝特化微菌核。J, 沙坡头柠条锦鸡儿DSE叶状微菌核。K, 民勤红花锦鸡儿DSE带状微菌核。L, 民勤无叶沙拐枣DSE叶状微菌核。
Fig. 2 Microsclerotia of dark septate endophytes (DSE) in roots of Ammopiptanthus mongolicus and its associated plants. A, Brain-like microsclerotia of DSE in A. mongolicus in Yinchuan. B, Chain-like microsclerotia of DSE in A. mongolicus in Yinchuan. C, Microsclerotia clusters of DSE in A. mongolicus in Shapotou. D, Leaf-like microsclerotia of DSE in A. mongolicus in Shapotou. E, Ribbon-like microsclerotia of DSE in A. mongolicus in Minqin. F, leaf-like microsclerotia of DSE in A. mongolicus in Minqin. G, Brain-like microsclerotia of DSE in Nitraria tangutorum in Yinchuan. H, Scattered microsclerotia of DSE in Artemisia desertorum in Yinchuan. I, Specialized microsclerotia of DSE in Caragana korshinskii in Shapotou. J, Leaf-like microsclerotia of DSE in C. korshinskii in Shapotou. K, Ribbon-like microsclerotia of DSE in C. rosea in Minqin. L, Leaf-like microsclerotia of DSE in Calligonum aphyllum in Minqin.
不同样地沙冬青与伴生植物DSE定殖结构有一定差异, 银川样地沙冬青菌丝粗, 横隔明显, 菌丝末端膨大(图1A); 伴生植物DSE菌丝细, 横隔不明显(图1D)。沙坡头样地沙冬青和伴生植物菌丝细(图1B)。民勤样地沙冬青菌丝粗(图1C), 伴生植物菌丝粗且横隔短(图1G)。沙冬青和伴生植物DSE微菌核都有离散型和聚合型, 其中, 银川样地沙冬青DSE微菌核多为微带条状(图2B), 聚合型为颜色较深的小团簇(图2A); 伴生植物DSE微菌核多为离散型(图2H)、颜色浅, 小团块状(图2G)。沙坡头样地沙冬青DSE微菌核呈团块状离散或微带条状(图2C、2D); 伴生植物DSE微菌核呈星点状离散型(图2J)。民勤样地沙冬青DSE微菌核呈聚合型, 多为大块状(图2E、2F); 伴生植物DSE微菌核排列紧密, 多为聚合型条带状和块状(图2K、2L)。
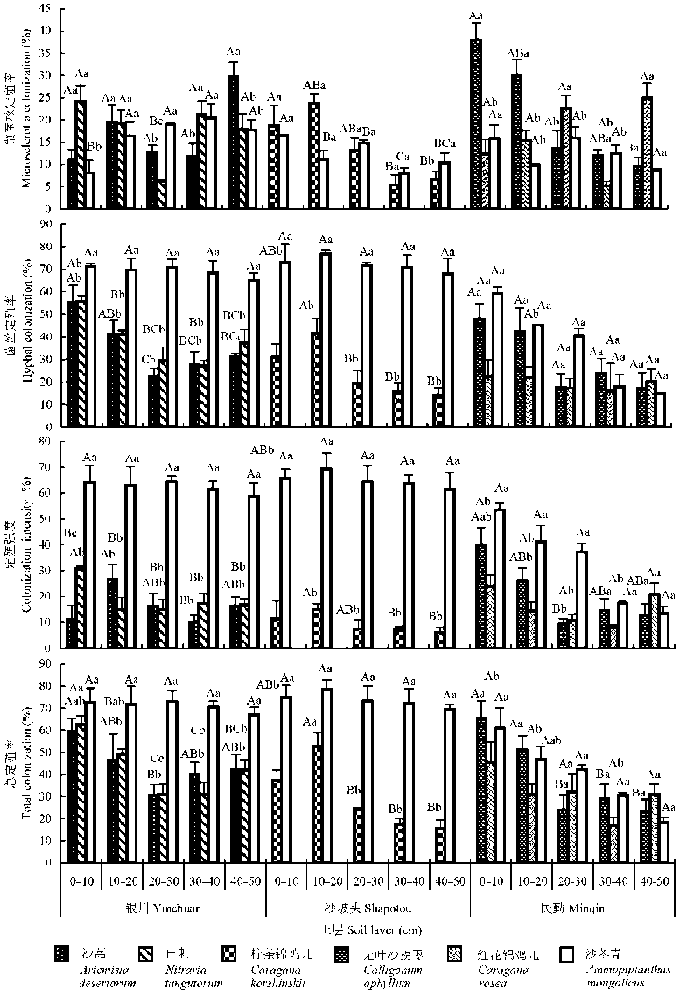
银川样地沙蒿(Artemisia desertorum) DSE菌丝定殖率和总定殖率最大值在0-10 cm土层, 10-20 cm土层定殖强度显著高于其他土层; 白刺(Nitraria tangutorum)菌丝定殖率、定殖强度和总定殖率最大值在0-10 cm土层, 与其他土层差异显著; 沙冬青DSE定殖率在土层间无显著差异。不同土层, 沙冬青DSE菌丝定殖率、定殖强度和总定殖率显著高于伴生植物(图3)。
图3 沙冬青及其伴生植物深色有隔内生真菌定殖率和定殖强度空间分布(平均值±标准偏差)。不同大写字母表示同一植物不同土层间差异显著(p < 0.05); 不同小写字母表示不同植物同一土层间差异显著(p < 0.05)。
Fig. 3 The spatial variation of the colonization of dark septate endophytes in roots of Ammopiptanthus mongolicus and its associated plants (mean ± SD). Different capital letters above the columns indicate significant difference between different soil layers of the same plant species (p < 0.05). Different lowercase letters above the columns indicate significant difference among different plants at the same soil layer (p < 0.05).
沙坡头样地柠条锦鸡儿(Caragana korshinskii) DSE定殖率最大值在0-20 cm土层, 并随土层加深而逐渐降低; 沙冬青DSE各项指标在土层间无明显差异。不同土层, 沙冬青DSE菌丝定殖率、定殖强度和总定殖率显著高于伴生植物(图3)。
民勤样地无叶沙拐枣(Calligonum aphyllum) DSE定殖率最大值在0-20 cm土层; 红花锦鸡儿(Caragana rosea) DSE定殖强度和总定殖率最大值在0-10 cm土层; 沙冬青DSE定殖率和定殖强度在土层间无显著差异。不同土层, 沙冬青仅DSE定殖强度显著高于伴生植物(图3)。
不同样地, 伴生植物DSE定殖率最大值主要集中在0-20 cm土层, 而沙冬青DSE定殖率在土层间差异不显著; 同一样地, 沙冬青DSE菌丝定殖率、定殖强度和总定殖率均高于伴生植物, 显著性为沙坡头>银川>民勤, 而微菌核定殖率差异不显著。不同样地, 沙冬青和伴生植物微菌核定殖率无显著差异; 伴生植物菌丝定殖率、定殖强度和总定殖率为 银川>民勤>沙坡头; 沙冬青菌丝定殖率、定殖强度和总定殖率为沙坡头>银川>民勤。
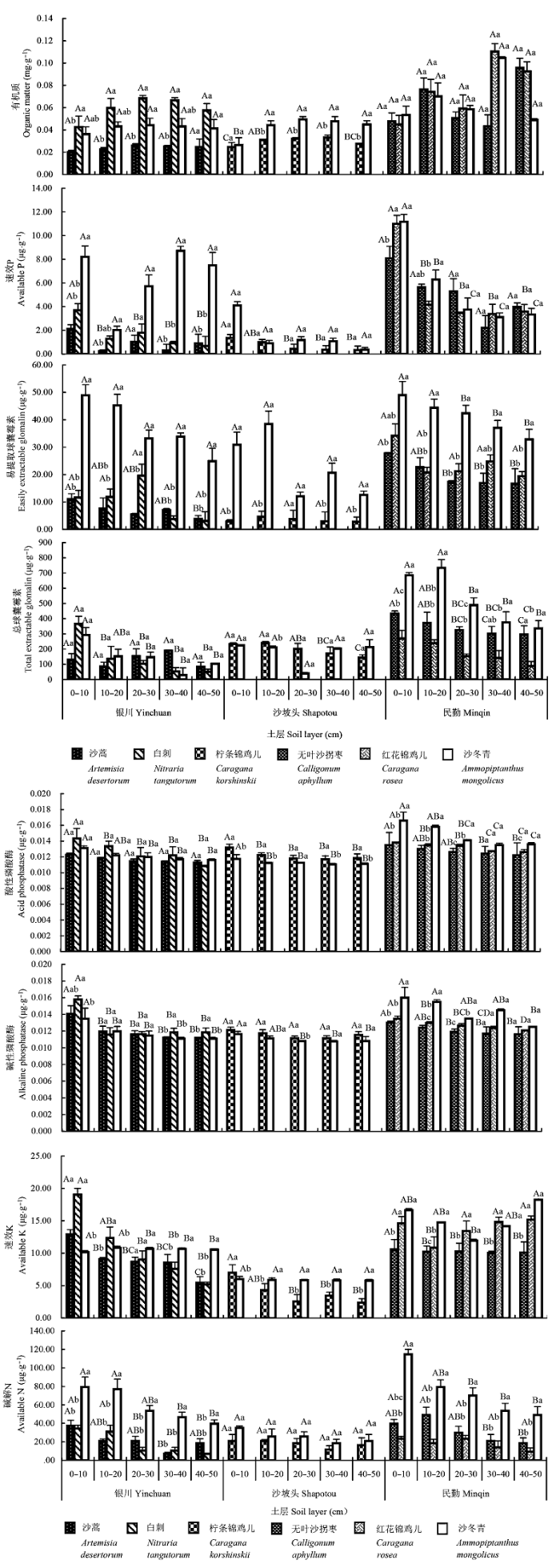
银川样地沙蒿各土层EEG、磷酸酶、速效K、碱解N最大值在0-10 cm土层; 白刺除有机质和EEG在土层间无显著差异, 其他指标最大值多在0-10 cm土层; 沙冬青0-10 cm土层TEG、磷酸酶和碱解N显著高于其他土层。沙冬青各土层速效P、EEG和碱解N显著高于伴生植物(图4)。
图4 沙冬青及其伴生植物根围土壤因子空间分布(平均值±标准偏差)。不同大写字母表示同一植物不同土层间差异显著(p < 0.05); 不同小写字母表示不同植物同一土层间差异显著(p < 0.05)。
Fig. 4 The spatial variation of soil properties in the rhizosphere of Ammopiptanthus mongolicus and its associated plants (mean ± SD). Different capital letters above the columns indicate significant difference between different soil layers of the same plant species (p < 0.05). Different lowercase letters above the columns indicate significant difference among different plants at the same soil layer (p < 0.05).
沙坡头样地柠条锦鸡儿有机质最大值在20-40 cm土层, 而0-20 cm土层速效P、TEG、酸性磷酸酶、速效K显著高于其他土层; 沙冬青各土层速效P、EEG、TEG、速效K、碱解N无显著差异, 而20-40 cm土层有机质高于其他土层, 磷酸酶最大值在0-10 cm土层。沙冬青各土层有机质含量、EEG、速效K均高于伴生植物(图4)。
民勤样地无叶沙拐枣0-20 cm土层EEG、TEG、磷酸酶、碱解N显著高于其他土层, 其中, EEG、TEG和磷酸酶随土层加深逐渐下降; 红花锦鸡儿速效P、TEG、磷酸酶最大值在0-20 cm土层, 其中磷酸酶、TEG随土层加深逐渐降低; 沙冬青除有机质外, 其他指标最大值多在0-10 cm土层, 其中, EEG、TEG、碱解N随土层加深逐渐降低。沙冬青各土层EEG、TEG、磷酸酶、速效K和碱解N显著高于伴生植物 (图4)。
不同样地, 沙冬青和伴生植物根围土壤速效P、EEG、TEG、磷酸酶、速效K和碱解N最大值多在0-10 cm土层, 而有机质最大值主要在20-40 cm土层; 沙冬青各土层EEG、TEG、速效K、碱解N显著高于其伴生植物。
银川样地土壤碱解N和EEG对DSE定殖贡献率最大, 其中碱解N是通过直接作用影响DSE定殖, EEG是通过碱解N起间接作用影响DSE定殖(表2)。沙坡头样地土壤TEG、酸性磷酸酶和碱解N对DSE定殖贡献率大, 其中TEG既通过直接作用对DSE定殖产生积极影响, 又通过酸性磷酸酶起间接作用影响DSE定殖; 同一样地, 酸性磷酸酶一方面直接影响DSE定殖, 一方面又间接通过TEG对DSE定殖产生正面影响; 碱解N主要是直接影响DSE定殖(表3)。民勤样地土壤有机质、EEG、TEG对DSE定殖贡献率大, 其中, 有机质和EEG通过直接作用影响DSE定殖; TEG一方面对DSE定殖有负作用, 一方面又可间接通过碱解N的负作用影响DSE定殖(表4)。
表2 银川样地土壤因子(xi)与深色有隔内生真菌总定殖率(y)的通径分析
Table 2 Path coefficient between soil factors (xi) and total colonization of dark septate endophytes (y) in Yinchuan
| 自变量 Independent variable | 与y的简单相关系数 Simple correlation with y | 通径系数的直接作用 Direct action of path coefficient | 间接通径系数(间接作用) Indirect path coefficient (indirect effect) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| x1 | x2 | x3 | x4 | x5 | x6 | x7 | x8 | |||
| x1 | -0.174 6 | -0.161 0 | - | 0.023 7 | -0.002 3 | -0.005 6 | 0.002 1 | -0.019 9 | -0.003 0 | -0.040 5 |
| x2 | 0.398 8 | 0.212 0 | -0.018 0 | - | -0.004 1 | -0.003 2 | 0.015 0 | 0.005 8 | -0.006 6 | 0.186 4 |
| x3 | 0.607 6 | -0.014 0 | -0.026 9 | 0.061 9 | - | 0.001 3 | 0.022 4 | 0.010 3 | -0.012 5 | 0.551 0 |
| x4 | 0.177 1 | 0.053 0 | 0.017 1 | -0.012 7 | -0.000 3 | - | 0.040 2 | 0.081 9 | 0.009 9 | 0.139 9 |
| x5 | 0.361 0 | 0.119 0 | -0.002 9 | 0.026 7 | -0.002 6 | 0.017 9 | - | 0.089 8 | 0.014 0 | 0.227 2 |
| x6 | 0.294 5 | 0.127 0 | 0.025 3 | 0.009 8 | -0.001 1 | 0.034 2 | 0.084 1 | - | 0.014 5 | 0.164 7 |
| x7 | -0.347 3 | 0.025 0 | 0.019 3 | -0.055 8 | 0.007 0 | 0.020 9 | 0.066 6 | 0.073 7 | - | -0.311 0 |
| x8 | 0.766 1 | 0.710 0 | 0.009 2 | 0.055 8 | -0.010 9 | 0.010 4 | 0.038 1 | 0.029 5 | -0.011 0 | - |
表3 沙坡头样地土壤因子(xi)与深色有隔内生真菌总定殖率(y)的通径分析
Table 3 Path coefficient between soil factors (xi) and total colonization of dark septate endophytes (y) in Shapotou
| 自变量Independent variable | 与y的简单相关系数 Simple correlation with y | 通径系数的直接作用 Direct action of path coefficient | 间接通径系数(间接作用) Indirect path coefficients (indirect effect) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| x1 | x2 | x3 | x4 | x5 | x6 | x7 | x8 | |||
| x1 | -0.073 2 | 0.039 0 | - | -0.006 1 | -0.006 8 | -0.072 5 | -0.062 1 | -0.012 3 | -0.001 7 | -0.048 5 |
| x2 | 0.222 6 | 0.032 0 | -0.007 5 | - | 0.002 4 | 0.329 6 | 0.259 6 | 0.035 6 | -0.078 0 | -0.053 3 |
| x3 | 0.249 3 | 0.110 0 | -0.002 4 | 0.000 7 | - | -0.194 1 | -0.066 0 | -0.012 8 | 0.115 0 | 0.267 9 |
| x4 | 0.359 9 | 0.529 0 | -0.005 3 | 0.019 9 | -0.040 4 | - | 0.334 8 | 0.051 5 | -0.115 5 | -0.077 0 |
| x5 | 0.406 6 | 0.388 0 | -0.006 2 | 0.021 4 | -0.018 7 | 0.456 5 | - | 0.052 7 | -0.094 1 | 0.018 5 |
| x6 | 0.357 0 | 0.060 0 | -0.008 0 | 0.019 0 | -0.023 4 | 0.454 4 | 0.341 1 | - | -0.084 7 | 0.002 2 |
| x7 | -0.075 5 | -0.174 0 | 0.000 4 | 0.014 3 | -0.072 7 | 0.351 3 | 0.209 9 | 0.029 2 | - | -0.246 1 |
| x8 | 0.389 0 | 0.370 0 | -0.005 1 | -0.004 6 | 0.079 6 | -0.110 0 | 0.019 4 | 0.036 0 | 0.115 7 | - |
表4 民勤样地土壤因子(xi)与深色有隔内生真菌总定殖率(y)的通径分析
Table 4 Path coefficient between soil factors (xi) and total colonization of dark septate endophytes (y) in Minqin
| 自变量Independent variable | 与y的简单相关系数 Simple correlation with y | 通径系数的直接作用 Direct action of path coefficient | 间接通径系数(间接作用) Indirect path coefficients (indirect effect) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| x1 | x2 | x3 | x4 | x5 | x6 | x7 | x8 | |||
| x1 | 0.502 1 | 0.358 0 | - | -0.026 6 | 0.037 5 | 0.106 9 | -0.051 3 | -0.018 9 | 0.018 3 | -0.002 3 |
| x2 | 0.179 2 | 0.190 0 | -0.050 1 | - | 0.011 0 | 0.042 3 | -0.001 4 | -0.002 1 | 0.000 5 | 0.016 2 |
| x3 | 0.438 0 | 0.305 0 | 0.044 0 | 0.006 8 | - | 0.088 6 | -0.014 8 | -0.002 3 | 0.008 7 | 0.009 7 |
| x4 | -0.511 6 | -0.226 0 | -0.007 1 | -0.030 2 | -0.101 6 | - | 0.034 8 | 0.008 7 | -0.013 3 | -0.143 9 |
| x5 | -0.371 3 | 0.078 0 | -0.235 6 | -0.003 6 | -0.058 0 | -0.118 6 | - | 0.021 6 | -0.021 6 | -0.007 7 |
| x6 | -0.264 9 | 0.034 0 | -0.199 0 | -0.011 6 | -0.021 0 | -0.067 8 | 0.049 5 | - | -0.011 2 | 0.004 8 |
| x7 | -0.429 0 | -0.031 0 | -0.210 5 | 0.002 9 | -0.086 0 | -0.113 8 | 0.054 4 | 0.012 3 | - | 0.000 7 |
| x8 | 0.136 0 | 0.041 0 | -0.019 7 | 0.075 2 | 0.072 0 | 0.046 3 | -0.014 7 | 0.004 0 | -0.000 5 | - |
表5 主成分载荷矩阵、特征值和贡献率
Table 5 Principle component (PC) loading matrix, eigenvalue and contribution rate
| 土壤因子 Soil factors | 第一主成分 PC1 | 第二主成分 PC2 |
|---|---|---|
| 有机质 Organic matter | 0.771 | 0.293 |
| 速效P Available P | 0.717 | 0.498 |
| 易提取球囊霉素 Easily extractable glomalin | 0.313 | 0.930 |
| 总球囊霉素 Total extractable glomalin | 0.856 | -0.356 |
| 酸性磷酸酶 Acid phosphatase | 0.969 | -0.072 |
| 碱性磷酸酶 Alkaline phosphatase | 0.947 | -0.173 |
| 速效K Available K | 0.759 | -0.576 |
| 碱解N Available N | 0.109 | 0.926 |
| 特征值 Eigenvalue | 4.364 | 2.550 |
| 贡献率 Contribution rate | 54.544 | 31.877 |
对比不同样地可知: 土壤碳氮含量可以影响DSE定殖; 球囊霉素既可直接影响DSE定殖, 又可通过影响其他土壤酶类或氮含量间接影响DSE定殖。
8种土壤因子不同程度地反映了DSE信息, 并且它们彼此之间有一定的相关性, 因而所得统计数据反映的信息在一定程度上有重叠。因此对各土壤因子进行主成分分析, 对其进行降维处理, 得到对DSE生长作用明显的指标。根据相关矩阵特征值大于1, 方差积累贡献率大于75%的原则, 选入了两个主成分, 其方差累计贡献率达到86%, 所以提取的主成分基本能反映全部信息。第一主成分中酸性磷酸酶载荷最大(比重占0.947), 有机质、TEG、碱性磷酸酶载荷较大(比重占0.759-0.947), 第二主成分中, EEG和碱解N载荷较大(比重占0.926-0.930)。由于第一主成分信息所占比重较大, 所以有机质、TEG、磷酸酶和速效K是主要土壤因子, 能综合反映西北荒漠环境土壤营养状况。
研究结果表明, 沙冬青及其伴生植物的根系能被DSE侵染, 并形成典型的微菌核和深色有隔菌丝, 但沙冬青DSE平均定殖率(60.58%)显著高于其伴生植物(37.57%)。这是因为沙冬青主根系粗壮, 侧根发达, 根幅常为冠幅的4倍以上, 而沙蒿和白刺等伴生植物根系入土浅, 不能为DSE提供更多的寄居环境。DSE较高定殖能够显著提高根系生物量, 有助于扩大根系生长范围(Newsham, 2011)。
不同样地, 沙冬青与伴生植物微菌核定殖率无明显差异, 但沙冬青菌丝定殖率显著高于伴生植物。微菌核是一种静止结构或繁殖器官, 菌丝是DSE与植物进行营养物质交换的通道(Peterson et al., 2008)。菌丝定殖率在一定程度上反映了植物根系物质交换活动强度。
沙质土壤表层营养物质流动性大, 根系较浅的植株只能在浅土层获取有限的营养物质, 而根系较深的植物能够利用更深土层养分, 这一研究结果可以解释沙坡头样地DSE的定殖规律。在高肥力土壤中DSE的矿化作用(Hoeksema et al., 2010)下降, 植物可以直接从土壤吸收所需营养物质, 使DSE与植物之间的互动联系降低, 这与民勤样地的研究结果相同。由此可见, DSE在不同植物间的定殖受到宿主根部结构和生境的双重影响。
球囊霉素是AM真菌产生的一种含金属离子的糖蛋白(Driver et al., 2005), 是土壤有机碳氮的重要来源, 能综合反映土壤中AM真菌群落动态, 促进植物根际土壤养分循环及其分解进程(陈颖等, 2009)。试验发现, 沙冬青及其伴生植物中均有AM真菌定殖, 在根内或根间与DSE形成致密的菌丝桥(张淑容等, 2013), 促进DSE矿化分解根际蛋白质、多糖和氨基酸等大分子, 使根系能够更好地利用无机氮, 促进植物生长(Newsham, 2011)。不同样地沙冬青及其伴生植物DSE定殖与磷酸酶呈正相关, 磷酸酶可将土壤有机磷酸酯水解为无机态磷酸, 为植物吸收利用(白春明等, 2009)。磷酸酶主要来自土壤微生物和植物根系的分泌(牛世全等, 2010)。DSE与其他土壤微生物有协同互助关系, 能扩大根系营养吸收范围, 有效地进行代谢物传递, 共同促进植物根围微生物活动, 改善植物根围土壤微环境(张淑容等, 2013)。
K虽然不是酶系统的组成成分, 但可参与酶活化, 影响植物光合速率及其光合产物合成诸多途径的运转效率(张淑容等, 2013)。土壤K主要通过质流和扩散方式穿过土壤颗粒移动到根部为植物吸收利用(占丽平等, 2012)。本试验中, 不同样地伴生植物根围速效K较沙冬青低, 这是由于沙冬青根系延伸范围广, 根毛数量多, 能够吸收到土壤中离根较远的K, 同时由于根系对K的吸收, 导致土壤K向根系表面的扩散作用增强(张富仓, 2001)。
植物体可以通过改变自身机能或生物间相互作用增强其环境适应性(van der Putten et al., 2010)。化石证据证实了早在4亿多年前就已经存在植物与真菌的联合体(Krings et al., 2007)。DSE通过在根部定殖对植物生长产生影响, 其对植物的负、中或正性效应取决于真菌种类、寄主植物和环境条件(Wu et al., 2010)。Gilbert等(2010)认为定殖于植物根系的DSE对于宿主营养和逆境适应是必需的, 因为特定植物的环境中有益DSE种群的遗传多样性能够扩展宿主植物的生态适应性, 使植物增强适应逆境的能力。在某些情况下, DSE能够改变植物之间的竞争结果: 当植物可以轻松获得养料时, DSE对植物生长无明显促进作用, 特别是土壤含N量高反而抑制DSE生长; 而贫瘠土壤中, DSE能够将有机物矿化成无机离子, 改变植物根围营养结构和碳循环(Hoeksema et al., 2010; Newsham, 2011; Reininger et al., 2012)。沙冬青发达的根系成为DSE良好的栖息地, 同时DSE定殖扩大了沙冬青根系延伸范围, 沙冬青根系与DSE形成的联合体(Barrow et al., 2008)有助于沙冬青适应极端荒漠环境。
致谢 感谢河北大学生命科学学院王晓乾、胡从从、陈严严、常辉同学协助野外样品采集工作。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
| [1] |
Study on relationships between arbuscular mycorrhizal fungi and soil enzyme activities of Astragalus adsurgens in the desert. Journal of Northwest A&F University ( 漠境沙打旺根围AM真菌与土壤酶活性的关系 . |
| [2] |
Do symbiotic microbes have a role in plant evolution, performance and response to stress? |
| [3] |
The spatio- temporal distribution of arbuscular mycorrhiza fungi and glomalin in the rhizosphere of Artemisia ordosica in desert. 荒漠油蒿根围AM真菌与球囊霉素的时空分布 . |
| [4] |
Association of dark septate endophytes with rice roots. |
| [5] |
Characterization of glomalin as a hyphal wall component of arbuscular mycorrhizal fungi. Soil Biology & |
| [6] |
Symbiosis as a source of selectable epigenetic variation: Taking the heat for the big guy.
|
| [7] |
A meta-analysis of context-dependency in plant response to inoculation with mycorrhizal fungi. |
| [8] |
Spatial distribution of AM and DSE fungi in the rhizosphere of Ammopiptanthus nanus. 新疆沙冬青AM和DSE真菌的空间分布 . |
| [9] |
Mycorrhizae and other root-associated fungal structures of the plants of a sandy grassland on the Great Hungarian Plain. |
| [10] |
Fungal endophytes in a 400-million-yr-old land plant: Infection pathways, spatial distribution, and host responses. |
| [11] |
Advances in the research of dark septate endophytes. 深色有隔内生真菌(DSE)研究进展 . |
| [12] |
|
| [13] |
A meta-analysis of plant responses to dark septate root endophytes. |
| [14] |
The relationships of soil microbial biomass, physicochemical factors and soil enzyme activities in Maqu swamp wetland of Gannan prefecture. 甘南玛曲沼泽湿地土壤微生物量、理化因子与土壤酶活的关系 . |
| [15] |
Associations between microfungal endophytes and roots: Do structural features indicate function? |
| [16] |
Host species and strain combination determine growth reduction of spruce and birch seedlings colonized by root-associated dark septate endophytes. |
| [17] |
Root fungus colonization along an altitudinal gradient in North Norway. |
| [18] |
Effect of phosphatase activity on soil organic phosphorus loss in the environment of clover growth. 菌根真菌磷酸酶活性对红三叶草生境中土壤有机磷亏缺的影响 . |
| [19] |
Fungal root endophyte associations of medicinal plants. |
| [20] |
Predicting species distribution and abundance responses to climate change: Why it is essential to include biotic interactions across trophic levels. |
| [21] |
A survey of soils for aggregate stability and glomalin, a glycoprotein produced by hyphae of arbuscular mycorrhizal fungi. |
| [22] |
The promoting role of an isolate of dark-septate fungus on its host plant Saussurea involucrata Kar. et Kir . |
| [23] |
Colonization of arbuscular mycorrhizal fungi and dark septate endophytes in roots of desert
荒漠北沙柳根系丛枝菌根真菌和黑隔内生真菌定殖状况 .
|
| [24] |
Characteristics of Ammopiptanthus nanus seed and factors affecting its germination. 矮沙冬青种子特性和萌发影响因素的研究 . |
| [25] |
Research advances on influence factors of soil potassium movement. 土壤钾素运移的影响因素研究进展 . |
| [26] |
Soil Root System and Its Numerical Simulation of Nutrient Transfer Mechanism .土壤-根系统养分迁移机制及其数值模拟 . |
| [27] |
Correlation study of AM and DSE fungi and soil factors in the rhizosphere of Ammopiptanthus mongolicus. Acta Botanica Boreali- 蒙古沙冬青根围AM和DSE真菌与土壤因子的相关性研究 . |
| [28] |
Arbuscular mycorrhizal fungi associated with the clonal plants in Mu Us sandland of China. |

Copyright © 2026 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19
![]()
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}