Dynamics and responses of sap flow of typical sand binding plants Haloxylon ammodendron to environmental variables
XU Shi-Qin1, 2, JI Xi-Bin1, *, , JIN Bo-Wen1
1Cold and Arid Region Environmental and Engineering Research Institute, Chinese Academy of Sciences, Lanzhou 730000, China2University of Chinese Academy of Sciences, Beijing 100049, China
Transpiration is one of important physiological activities for plants, which is regulated by many environmental variables. Our objectives were to understand the responses of sap flow density of Haloxylon ammodendron to environmental variables and simulate its diurnal change under different micrometeorological conditions.
Methods
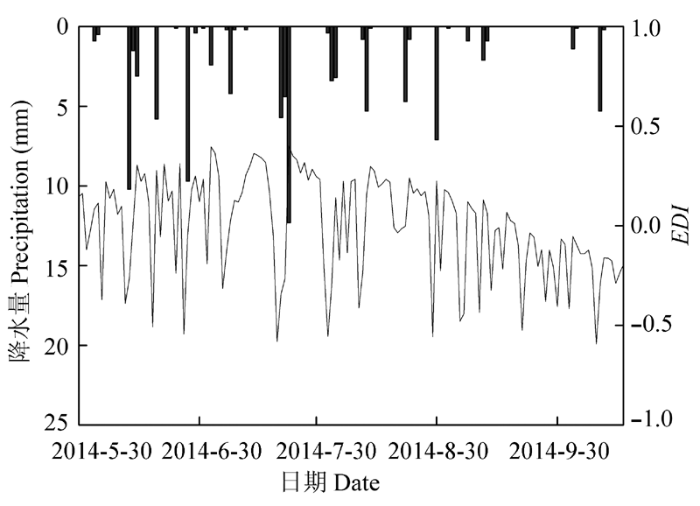
Sap flow in stems of H. ammodendron was measured with stem heat balance method using commercial sap-flow gauges from May to October, 2014, in the oasis-desert ecotone, located in the middle range of Hexi Corridor, Northwestern China.
Important findings
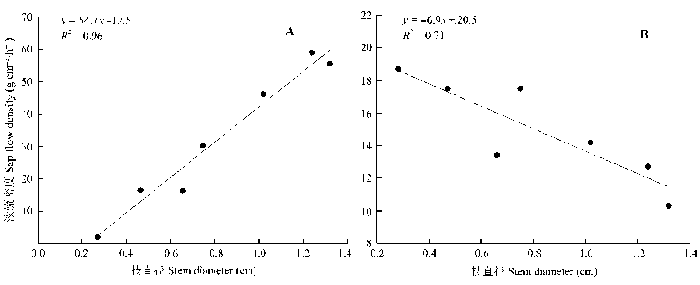
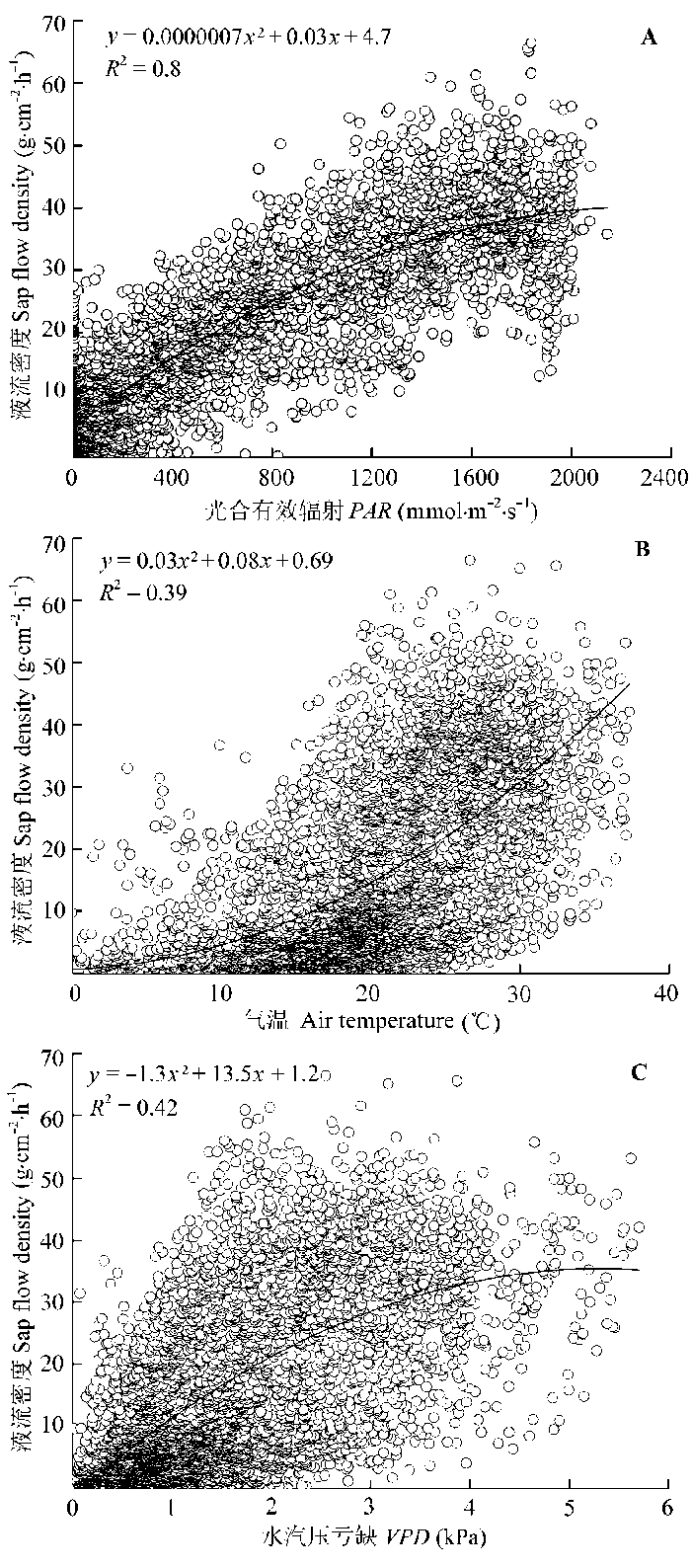
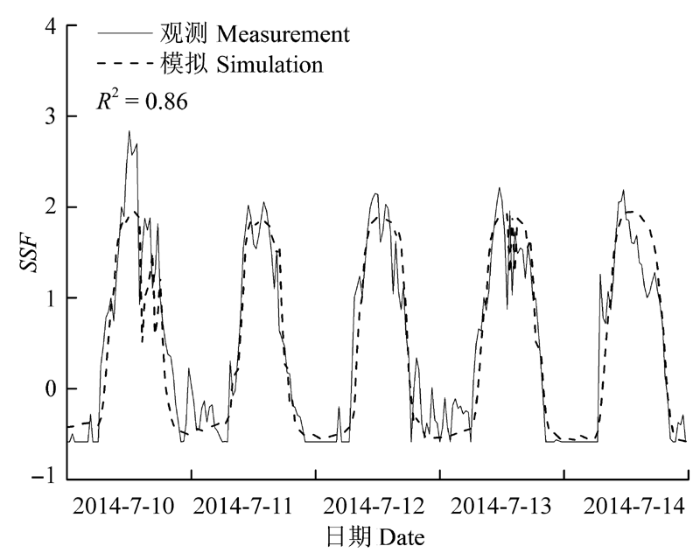
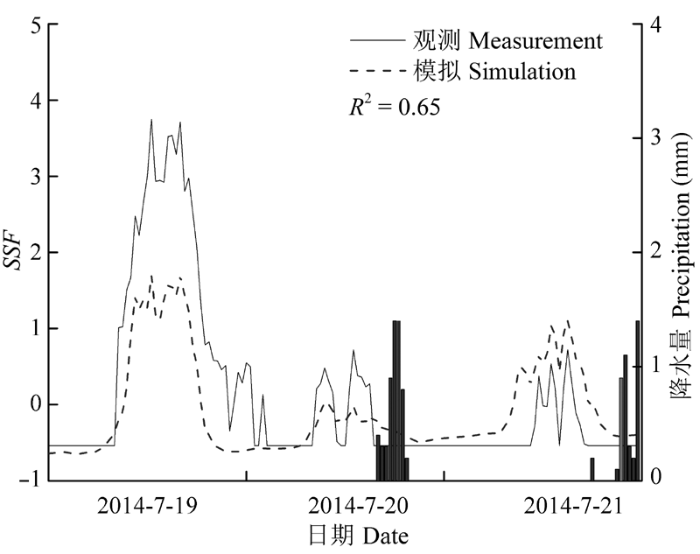
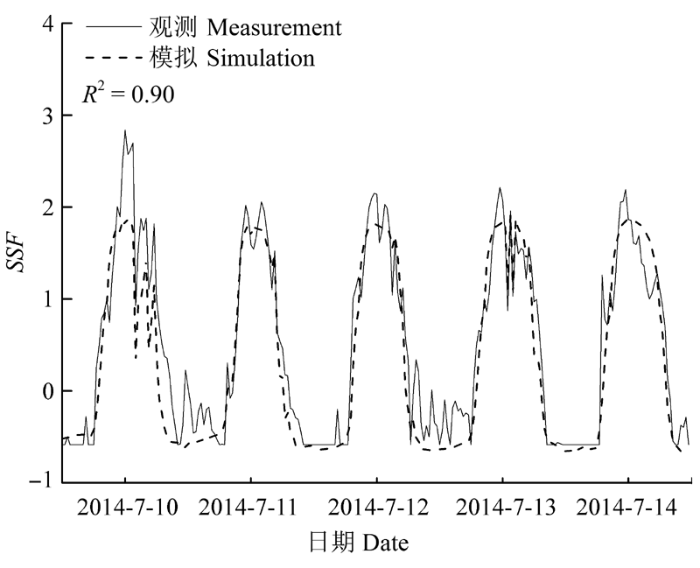
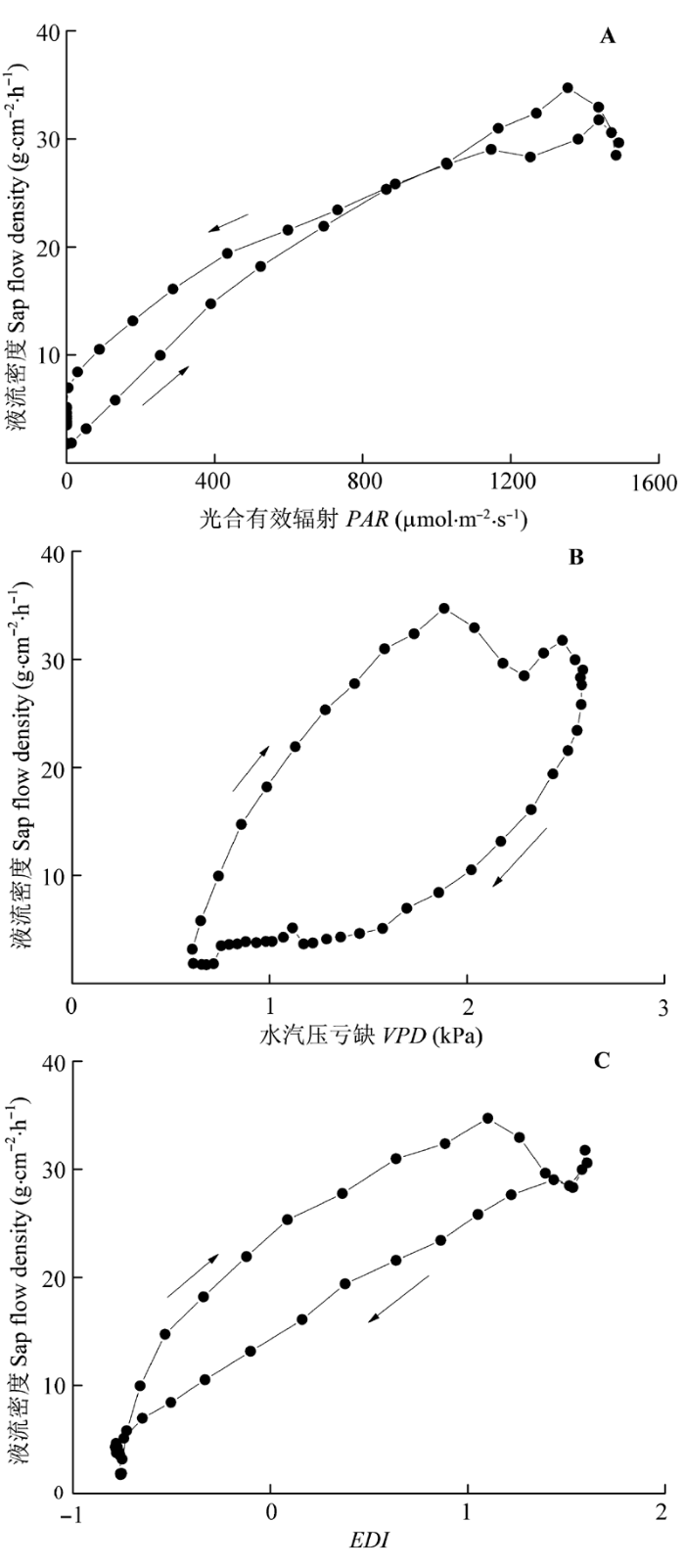
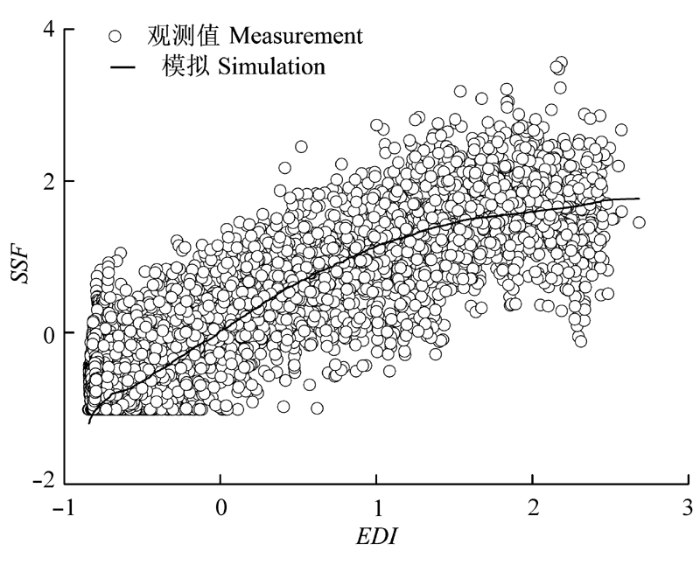
Sap flow velocity of H. ammodendron exhibited a positive relationship with stem diameter, but sap flow density (Js) decreased with stem diameter. The first three axes of principal component analysis (PCA) explained 49%, 15%, 12% of variances in the environmental datasets, respectively, and vapour pressure deficit (VPD), photosynthetically active radiation (PAR), temperature in the first axes indicated the atmospheric evaporative demand. A sigmoid function could explain 86% of the variation in Js in typical sunny days, while only 65% on rainy days. It was worth noting the simulated Js using the established sigmoid function agreed well with the measurements (R2 = 0.90) if the time lags of Js to principle environmental variables were taken into consideration. Plots of 30-min Js against PAR, VPD, and evaporative demand index (EDI) revealed a counter-clockwise hysteresis for PAR, but a clockwise hysteresis for VPD and EDI, it was possibly affected by water stress and time lags of sap flow density to principle environmental variables.
XUShi-Qin, JIXi-Bin, JINBo-Wen. Dynamics and responses of sap flow of typical sand binding plants Haloxylon ammodendron to environmental variables. Chinese Journal of Plant Ecology, 2015, 39(9): 890-900 https://doi.org/10.17521/cjpe.2015.0085
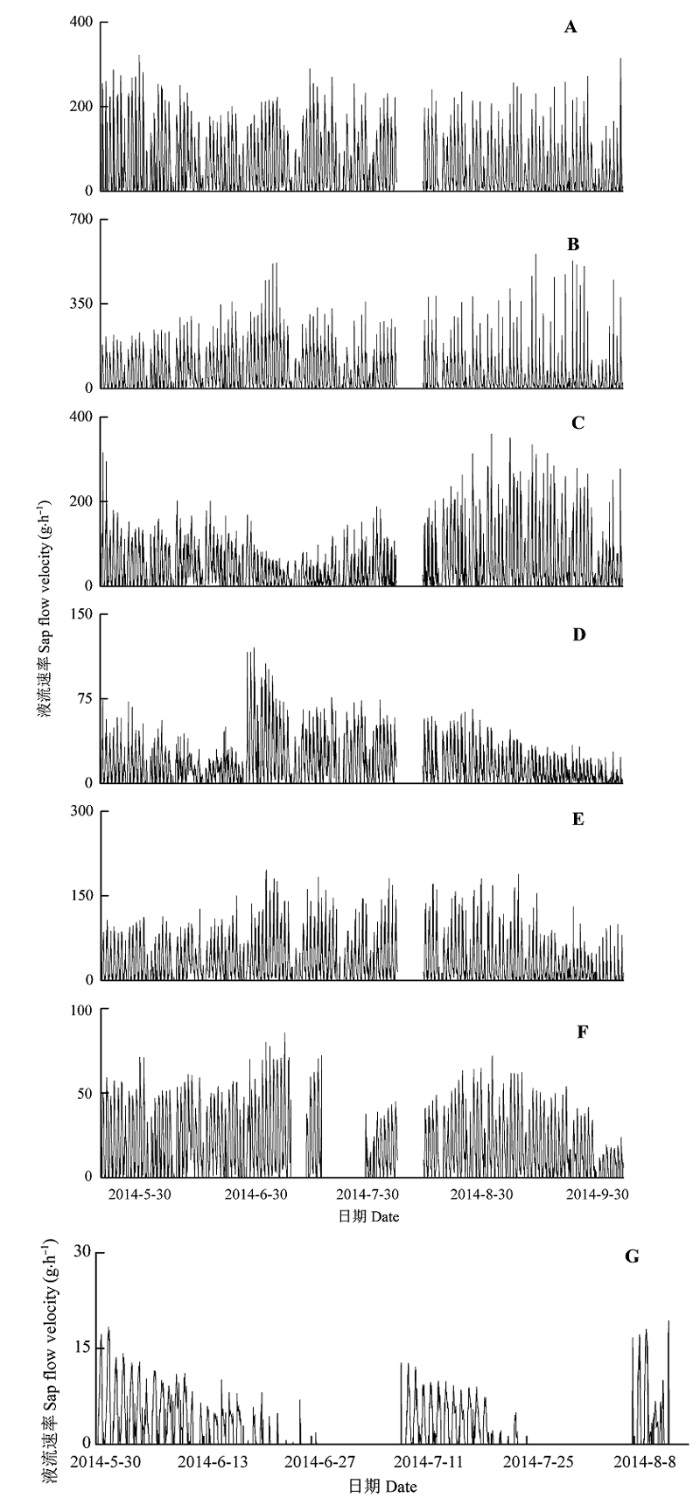
Fig. 1 Variation of sap flow velocity of Haloxylon ammodendron with different diameters. A-G, diameters of 1.32, 1.24, 1.02, 0.75, 0.66, 0.47, 0.27 cm.
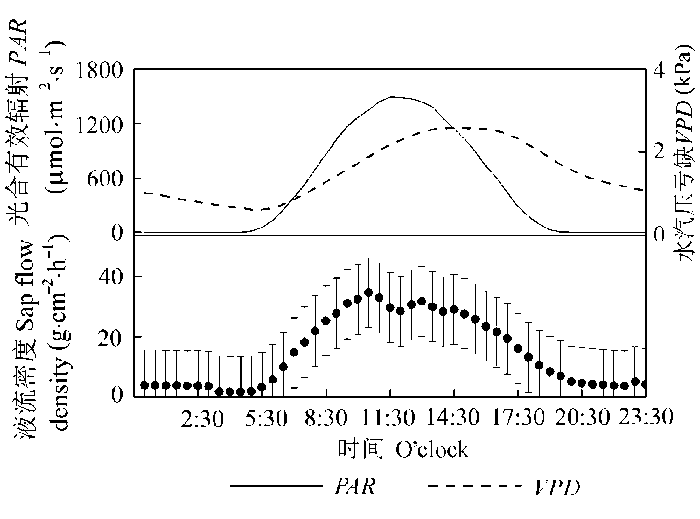
Fig. 3 The diurnal change of sap flow density (mean ± SD) and principle environmental variables during study period. PAR, photosynthetic active radiation; VPD, vapour pressure deficit.
Fig. 9 Hysteresis between sap flow density and photosynthetically active radiation (PAR)(A), vapour pressure deficit (VPD)(B), evaporative demand index (EDI)(C) .
Characteristics of dominant tree species stem sap flow and their relationships with environmental factors in a mixed conifer-broadleaf forest in Dinghushan, Guangdong Province of South China.
Chinese Journal of Applied Ecology, 23, 1159-1166.(in Chinese with English abstract)
Effects of planting density on the productivity and water use of tea (Camellia sinensis L.) clones: I. Measurement of water use in young tea using sap flow meters with a stem heat balance method.
Impact of stem water storage on diurnal estimates of whole-tree transpiration and canopy conductance from sap flow measurements in Japanese cedar and Japanese cypress trees.
Canopy transpiration and water fluxes in the xylem of the trunk of Larix and Picea trees — A comparison of xylem flow, porome- ter and cuvette measurements.
Regulation of water flux through trunks, branches, and leaves in trees of a lowland tropical forest.
1
1998
... 蒸腾是植物重要的生理活动之一, 该过程一方面促进植物体内水分和营养元素的运输, 另一方面通过气孔控制间接影响光合作用(Jones, 1983).植物茎干液流(stem sap flow)的90%以上通过叶片蒸腾散失到大气中(蒋高明, 2007), 因此能够将植物茎干液流量用于估算单株或群落蒸腾并获得植物重要的生理生态信息.茎干液流受到植物生理生化特征、环境要素及树形因子的综合影响(Andrade et al., 1998; Meinzer et al., 2001), 其中环境要素主要包括光合有效辐射(PAR)、水汽压亏缺(VPD)和气温等, 而长时间尺度液流变化则受区域气候的制约(Granier et al., 1992; Vertessy et al., 1995; McDowell et al., 2008).主成分分析(PCA)能将互有联系的环境要素分为不同的复合分量, 在保留原始数据主要信息的基础上实现数据降维, 该方法对于分析植物液流对环境因子的响应十分有效(O’Brien et al., 2004). ...
Environmental regulation of xylem sap flow and total conductance of Larix gmelinii trees in eastern Siberia.
1
1996
... 梭梭液流密度与VPD、EDI的日变化响应呈顺时针环状(图9A、9B), 相同VPD或EDI条件下上午液流密度更大, 这主要是因为虽然VPD和EDI在下午降低, 但是叶片气孔导度并没有恢复至原来水平(Arneth et al., 1996).液流密度与PAR响应的非对称性减弱且呈逆时针环状(图9C), 主要因为二者的日变化过程更为一致. ...
Measurement of mass flow of water in the stems of herbaceous plants.
1
1987
... 20世纪90年代以来, 茎干液流测定技术被广泛应用在植物单株及群落蒸腾研究上.目前常用的测量方法包括热脉冲(heat pulse method)(Granier, 1985, 1987), 热扩散(thermal dissipation method) (Smith & Allen, 1996)和热平衡(stem heat balance) (Baker & van Bavel, 1987), 它们均能获得植物茎干液流连续变化信息(Čermák et al., 1973; Edwards et al., 1996; Hall et al., 1998).但前两种方法需要计算树干边材面积, 耗时较多, 且不同方位、不同深度边材上植物液流速率存在的差异也会使测定结果的不确定性增大(Wullschleger & King, 2000; 徐飞等, 2012), 此外, 梭梭(Haloxylon ammodendron)茎干较细, 前两种测定技术在操作上并不可行.热平衡方法直接将加热器包裹在茎干外部对植物液流进行连续监测, 能有效地避免上述问题(Kigalu, 2007). ...
A new method of sap flow determination in trees.
1
1973
... 20世纪90年代以来, 茎干液流测定技术被广泛应用在植物单株及群落蒸腾研究上.目前常用的测量方法包括热脉冲(heat pulse method)(Granier, 1985, 1987), 热扩散(thermal dissipation method) (Smith & Allen, 1996)和热平衡(stem heat balance) (Baker & van Bavel, 1987), 它们均能获得植物茎干液流连续变化信息(Čermák et al., 1973; Edwards et al., 1996; Hall et al., 1998).但前两种方法需要计算树干边材面积, 耗时较多, 且不同方位、不同深度边材上植物液流速率存在的差异也会使测定结果的不确定性增大(Wullschleger & King, 2000; 徐飞等, 2012), 此外, 梭梭(Haloxylon ammodendron)茎干较细, 前两种测定技术在操作上并不可行.热平衡方法直接将加热器包裹在茎干外部对植物液流进行连续监测, 能有效地避免上述问题(Kigalu, 2007). ...
... 黄德卫等(2012)发现, 鼎湖山针阔混交林优势种液流峰值出现的时间晚于PAR峰值出现的时间, 与VPD的时滞效应在不同季节存在明显差异, 王华等(2008)发现马占相思(Acacia mangium)树干液流滞后于PAR, 而提前于VPD,王慧梅等(2011)发现兴安落叶松(Larix dahurica)树干液流峰值滞后于PAR约0.5 h, 提前于气温和空气相对湿度0-2 h.有研究认为植物茎干夜间水分的补充是导致时滞效应的重要原因(Phillips et al., 2003; Goldstein et al., 1998; Kumagai, 2009).此外植物茎干液流与环境要素时滞效应的差异还与植物木质部结构、树干储水性能及树形因子等因素有关(Granier et al., 2000; Ford et al., 2004; Chuang et al., 2006).有研究指出, 时滞现象通常在小时间尺度发生, 当上推至日或者更大时间尺度时, 这一现象不再明显(Oren et al., 1999; O’Brien et al., 2004). ...
A new method of sap flow measurement in tree stems.
1
1985
... 20世纪90年代以来, 茎干液流测定技术被广泛应用在植物单株及群落蒸腾研究上.目前常用的测量方法包括热脉冲(heat pulse method)(Granier, 1985, 1987), 热扩散(thermal dissipation method) (Smith & Allen, 1996)和热平衡(stem heat balance) (Baker & van Bavel, 1987), 它们均能获得植物茎干液流连续变化信息(Čermák et al., 1973; Edwards et al., 1996; Hall et al., 1998).但前两种方法需要计算树干边材面积, 耗时较多, 且不同方位、不同深度边材上植物液流速率存在的差异也会使测定结果的不确定性增大(Wullschleger & King, 2000; 徐飞等, 2012), 此外, 梭梭(Haloxylon ammodendron)茎干较细, 前两种测定技术在操作上并不可行.热平衡方法直接将加热器包裹在茎干外部对植物液流进行连续监测, 能有效地避免上述问题(Kigalu, 2007). ...
Evaluation of transpiration in a Douglas fir stand by means of sap flow measurements.
1
1987
... 20世纪90年代以来, 茎干液流测定技术被广泛应用在植物单株及群落蒸腾研究上.目前常用的测量方法包括热脉冲(heat pulse method)(Granier, 1985, 1987), 热扩散(thermal dissipation method) (Smith & Allen, 1996)和热平衡(stem heat balance) (Baker & van Bavel, 1987), 它们均能获得植物茎干液流连续变化信息(Čermák et al., 1973; Edwards et al., 1996; Hall et al., 1998).但前两种方法需要计算树干边材面积, 耗时较多, 且不同方位、不同深度边材上植物液流速率存在的差异也会使测定结果的不确定性增大(Wullschleger & King, 2000; 徐飞等, 2012), 此外, 梭梭(Haloxylon ammodendron)茎干较细, 前两种测定技术在操作上并不可行.热平衡方法直接将加热器包裹在茎干外部对植物液流进行连续监测, 能有效地避免上述问题(Kigalu, 2007). ...
Water balance, transpiration and canopy conductance in two beech stands.
1
2000
... 黄德卫等(2012)发现, 鼎湖山针阔混交林优势种液流峰值出现的时间晚于PAR峰值出现的时间, 与VPD的时滞效应在不同季节存在明显差异, 王华等(2008)发现马占相思(Acacia mangium)树干液流滞后于PAR, 而提前于VPD,王慧梅等(2011)发现兴安落叶松(Larix dahurica)树干液流峰值滞后于PAR约0.5 h, 提前于气温和空气相对湿度0-2 h.有研究认为植物茎干夜间水分的补充是导致时滞效应的重要原因(Phillips et al., 2003; Goldstein et al., 1998; Kumagai, 2009).此外植物茎干液流与环境要素时滞效应的差异还与植物木质部结构、树干储水性能及树形因子等因素有关(Granier et al., 2000; Ford et al., 2004; Chuang et al., 2006).有研究指出, 时滞现象通常在小时间尺度发生, 当上推至日或者更大时间尺度时, 这一现象不再明显(Oren et al., 1999; O’Brien et al., 2004). ...
Transpiration and stomatal conductance of two rain forest species growing in plantations (Simarouba amara and Goupia glabra) in French Guyana.
1
1992
... 蒸腾是植物重要的生理活动之一, 该过程一方面促进植物体内水分和营养元素的运输, 另一方面通过气孔控制间接影响光合作用(Jones, 1983).植物茎干液流(stem sap flow)的90%以上通过叶片蒸腾散失到大气中(蒋高明, 2007), 因此能够将植物茎干液流量用于估算单株或群落蒸腾并获得植物重要的生理生态信息.茎干液流受到植物生理生化特征、环境要素及树形因子的综合影响(Andrade et al., 1998; Meinzer et al., 2001), 其中环境要素主要包括光合有效辐射(PAR)、水汽压亏缺(VPD)和气温等, 而长时间尺度液流变化则受区域气候的制约(Granier et al., 1992; Vertessy et al., 1995; McDowell et al., 2008).主成分分析(PCA)能将互有联系的环境要素分为不同的复合分量, 在保留原始数据主要信息的基础上实现数据降维, 该方法对于分析植物液流对环境因子的响应十分有效(O’Brien et al., 2004). ...
... 蒸腾是植物重要的生理活动之一, 该过程一方面促进植物体内水分和营养元素的运输, 另一方面通过气孔控制间接影响光合作用(Jones, 1983).植物茎干液流(stem sap flow)的90%以上通过叶片蒸腾散失到大气中(蒋高明, 2007), 因此能够将植物茎干液流量用于估算单株或群落蒸腾并获得植物重要的生理生态信息.茎干液流受到植物生理生化特征、环境要素及树形因子的综合影响(Andrade et al., 1998; Meinzer et al., 2001), 其中环境要素主要包括光合有效辐射(PAR)、水汽压亏缺(VPD)和气温等, 而长时间尺度液流变化则受区域气候的制约(Granier et al., 1992; Vertessy et al., 1995; McDowell et al., 2008).主成分分析(PCA)能将互有联系的环境要素分为不同的复合分量, 在保留原始数据主要信息的基础上实现数据降维, 该方法对于分析植物液流对环境因子的响应十分有效(O’Brien et al., 2004). ...
1
2007
... 蒸腾是植物重要的生理活动之一, 该过程一方面促进植物体内水分和营养元素的运输, 另一方面通过气孔控制间接影响光合作用(Jones, 1983).植物茎干液流(stem sap flow)的90%以上通过叶片蒸腾散失到大气中(蒋高明, 2007), 因此能够将植物茎干液流量用于估算单株或群落蒸腾并获得植物重要的生理生态信息.茎干液流受到植物生理生化特征、环境要素及树形因子的综合影响(Andrade et al., 1998; Meinzer et al., 2001), 其中环境要素主要包括光合有效辐射(PAR)、水汽压亏缺(VPD)和气温等, 而长时间尺度液流变化则受区域气候的制约(Granier et al., 1992; Vertessy et al., 1995; McDowell et al., 2008).主成分分析(PCA)能将互有联系的环境要素分为不同的复合分量, 在保留原始数据主要信息的基础上实现数据降维, 该方法对于分析植物液流对环境因子的响应十分有效(O’Brien et al., 2004). ...
1
1983
... 蒸腾是植物重要的生理活动之一, 该过程一方面促进植物体内水分和营养元素的运输, 另一方面通过气孔控制间接影响光合作用(Jones, 1983).植物茎干液流(stem sap flow)的90%以上通过叶片蒸腾散失到大气中(蒋高明, 2007), 因此能够将植物茎干液流量用于估算单株或群落蒸腾并获得植物重要的生理生态信息.茎干液流受到植物生理生化特征、环境要素及树形因子的综合影响(Andrade et al., 1998; Meinzer et al., 2001), 其中环境要素主要包括光合有效辐射(PAR)、水汽压亏缺(VPD)和气温等, 而长时间尺度液流变化则受区域气候的制约(Granier et al., 1992; Vertessy et al., 1995; McDowell et al., 2008).主成分分析(PCA)能将互有联系的环境要素分为不同的复合分量, 在保留原始数据主要信息的基础上实现数据降维, 该方法对于分析植物液流对环境因子的响应十分有效(O’Brien et al., 2004). ...
Effects of planting density on the productivity and water use of tea (Camellia sinensis L.) clones: I. Measurement of water use in young tea using sap flow meters with a stem heat balance method.
1
2007
... 20世纪90年代以来, 茎干液流测定技术被广泛应用在植物单株及群落蒸腾研究上.目前常用的测量方法包括热脉冲(heat pulse method)(Granier, 1985, 1987), 热扩散(thermal dissipation method) (Smith & Allen, 1996)和热平衡(stem heat balance) (Baker & van Bavel, 1987), 它们均能获得植物茎干液流连续变化信息(Čermák et al., 1973; Edwards et al., 1996; Hall et al., 1998).但前两种方法需要计算树干边材面积, 耗时较多, 且不同方位、不同深度边材上植物液流速率存在的差异也会使测定结果的不确定性增大(Wullschleger & King, 2000; 徐飞等, 2012), 此外, 梭梭(Haloxylon ammodendron)茎干较细, 前两种测定技术在操作上并不可行.热平衡方法直接将加热器包裹在茎干外部对植物液流进行连续监测, 能有效地避免上述问题(Kigalu, 2007). ...
Sapflow measurements in forest stands: Methods and uncertainties.
Impact of stem water storage on diurnal estimates of whole-tree transpiration and canopy conductance from sap flow measurements in Japanese cedar and Japanese cypress trees.
1
2009
... 黄德卫等(2012)发现, 鼎湖山针阔混交林优势种液流峰值出现的时间晚于PAR峰值出现的时间, 与VPD的时滞效应在不同季节存在明显差异, 王华等(2008)发现马占相思(Acacia mangium)树干液流滞后于PAR, 而提前于VPD,王慧梅等(2011)发现兴安落叶松(Larix dahurica)树干液流峰值滞后于PAR约0.5 h, 提前于气温和空气相对湿度0-2 h.有研究认为植物茎干夜间水分的补充是导致时滞效应的重要原因(Phillips et al., 2003; Goldstein et al., 1998; Kumagai, 2009).此外植物茎干液流与环境要素时滞效应的差异还与植物木质部结构、树干储水性能及树形因子等因素有关(Granier et al., 2000; Ford et al., 2004; Chuang et al., 2006).有研究指出, 时滞现象通常在小时间尺度发生, 当上推至日或者更大时间尺度时, 这一现象不再明显(Oren et al., 1999; O’Brien et al., 2004). ...
Transpiration and stomatal conductance across a steep climate gradient in the southern Rocky Mountains.
1
2008
... 蒸腾是植物重要的生理活动之一, 该过程一方面促进植物体内水分和营养元素的运输, 另一方面通过气孔控制间接影响光合作用(Jones, 1983).植物茎干液流(stem sap flow)的90%以上通过叶片蒸腾散失到大气中(蒋高明, 2007), 因此能够将植物茎干液流量用于估算单株或群落蒸腾并获得植物重要的生理生态信息.茎干液流受到植物生理生化特征、环境要素及树形因子的综合影响(Andrade et al., 1998; Meinzer et al., 2001), 其中环境要素主要包括光合有效辐射(PAR)、水汽压亏缺(VPD)和气温等, 而长时间尺度液流变化则受区域气候的制约(Granier et al., 1992; Vertessy et al., 1995; McDowell et al., 2008).主成分分析(PCA)能将互有联系的环境要素分为不同的复合分量, 在保留原始数据主要信息的基础上实现数据降维, 该方法对于分析植物液流对环境因子的响应十分有效(O’Brien et al., 2004). ...
Regulation of water flux through tropical forest canopy trees: Do universal rules apply?
1
2001
... 蒸腾是植物重要的生理活动之一, 该过程一方面促进植物体内水分和营养元素的运输, 另一方面通过气孔控制间接影响光合作用(Jones, 1983).植物茎干液流(stem sap flow)的90%以上通过叶片蒸腾散失到大气中(蒋高明, 2007), 因此能够将植物茎干液流量用于估算单株或群落蒸腾并获得植物重要的生理生态信息.茎干液流受到植物生理生化特征、环境要素及树形因子的综合影响(Andrade et al., 1998; Meinzer et al., 2001), 其中环境要素主要包括光合有效辐射(PAR)、水汽压亏缺(VPD)和气温等, 而长时间尺度液流变化则受区域气候的制约(Granier et al., 1992; Vertessy et al., 1995; McDowell et al., 2008).主成分分析(PCA)能将互有联系的环境要素分为不同的复合分量, 在保留原始数据主要信息的基础上实现数据降维, 该方法对于分析植物液流对环境因子的响应十分有效(O’Brien et al., 2004). ...
Environmental and physiological regulation of transpiration in tropical forest gap species: The influence of boundary layer and hydraulic properties.
2
1995
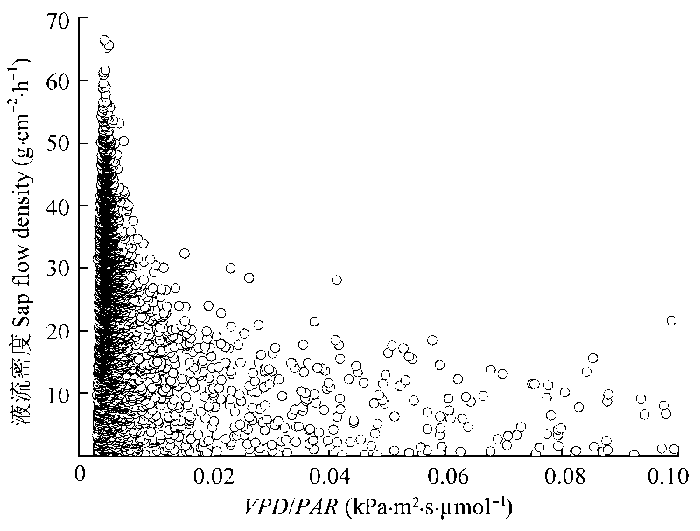
... 光是影响植物蒸腾的重要环境驱动因子, 但是对气孔导度变化具有消减作用(Meinzer et al., 1995), 从而影响茎干液流变化(Motzer et al., 2005).当VPD除以PAR以消除光的影响后发现, 液流密度对较低的VPD十分敏感, 且随着VPD的增加线性增加至其最大值后缓慢降低(图11), 而O’Brien等(2004)发现多种热带雨林植物茎干液流密度随VPD增加的速率同降低的速率基本一致, 这一差异表明在空气极端干旱时梭梭通过气孔调节尽可能减少水分损失(谭永芹等, 2011).Verbeeck等(2007)发现VPD是欧洲赤松(Pinus svlestris)树干液流的主要驱动因子, 当空气十分干燥时植物蒸腾受到抑制.大气环境蒸发力对植物液流的影响还受土壤水分状况以及植物导水率等因素的综合影响(Schulze et al., 1985; Meinzer et al., 1995; Motzer et al., 2005).Oguntunde (2005)发现当土壤严重干旱时, 空气蒸腾需求仅能解释木薯(Manihot esculenta)茎干液流24%的变化. ...
... ; Meinzer et al., 1995; Motzer et al., 2005).Oguntunde (2005)发现当土壤严重干旱时, 空气蒸腾需求仅能解释木薯(Manihot esculenta)茎干液流24%的变化. ...
Apparent responses of stomata to transpiration and humidity in a hybrid poplar canopy.
... 光是影响植物蒸腾的重要环境驱动因子, 但是对气孔导度变化具有消减作用(Meinzer et al., 1995), 从而影响茎干液流变化(Motzer et al., 2005).当VPD除以PAR以消除光的影响后发现, 液流密度对较低的VPD十分敏感, 且随着VPD的增加线性增加至其最大值后缓慢降低(图11), 而O’Brien等(2004)发现多种热带雨林植物茎干液流密度随VPD增加的速率同降低的速率基本一致, 这一差异表明在空气极端干旱时梭梭通过气孔调节尽可能减少水分损失(谭永芹等, 2011).Verbeeck等(2007)发现VPD是欧洲赤松(Pinus svlestris)树干液流的主要驱动因子, 当空气十分干燥时植物蒸腾受到抑制.大气环境蒸发力对植物液流的影响还受土壤水分状况以及植物导水率等因素的综合影响(Schulze et al., 1985; Meinzer et al., 1995; Motzer et al., 2005).Oguntunde (2005)发现当土壤严重干旱时, 空气蒸腾需求仅能解释木薯(Manihot esculenta)茎干液流24%的变化. ...
... ; Motzer et al., 2005).Oguntunde (2005)发现当土壤严重干旱时, 空气蒸腾需求仅能解释木薯(Manihot esculenta)茎干液流24%的变化. ...
Whole tree xylem sap flow responses to multiple environmental variables in a wet tropical forest.
4
2004
... 蒸腾是植物重要的生理活动之一, 该过程一方面促进植物体内水分和营养元素的运输, 另一方面通过气孔控制间接影响光合作用(Jones, 1983).植物茎干液流(stem sap flow)的90%以上通过叶片蒸腾散失到大气中(蒋高明, 2007), 因此能够将植物茎干液流量用于估算单株或群落蒸腾并获得植物重要的生理生态信息.茎干液流受到植物生理生化特征、环境要素及树形因子的综合影响(Andrade et al., 1998; Meinzer et al., 2001), 其中环境要素主要包括光合有效辐射(PAR)、水汽压亏缺(VPD)和气温等, 而长时间尺度液流变化则受区域气候的制约(Granier et al., 1992; Vertessy et al., 1995; McDowell et al., 2008).主成分分析(PCA)能将互有联系的环境要素分为不同的复合分量, 在保留原始数据主要信息的基础上实现数据降维, 该方法对于分析植物液流对环境因子的响应十分有效(O’Brien et al., 2004). ...
... 黄德卫等(2012)发现, 鼎湖山针阔混交林优势种液流峰值出现的时间晚于PAR峰值出现的时间, 与VPD的时滞效应在不同季节存在明显差异, 王华等(2008)发现马占相思(Acacia mangium)树干液流滞后于PAR, 而提前于VPD,王慧梅等(2011)发现兴安落叶松(Larix dahurica)树干液流峰值滞后于PAR约0.5 h, 提前于气温和空气相对湿度0-2 h.有研究认为植物茎干夜间水分的补充是导致时滞效应的重要原因(Phillips et al., 2003; Goldstein et al., 1998; Kumagai, 2009).此外植物茎干液流与环境要素时滞效应的差异还与植物木质部结构、树干储水性能及树形因子等因素有关(Granier et al., 2000; Ford et al., 2004; Chuang et al., 2006).有研究指出, 时滞现象通常在小时间尺度发生, 当上推至日或者更大时间尺度时, 这一现象不再明显(Oren et al., 1999; O’Brien et al., 2004). ...
Reliance on stored water increases with tree size in three species in the Pacific Northwest.
1
2003
... 黄德卫等(2012)发现, 鼎湖山针阔混交林优势种液流峰值出现的时间晚于PAR峰值出现的时间, 与VPD的时滞效应在不同季节存在明显差异, 王华等(2008)发现马占相思(Acacia mangium)树干液流滞后于PAR, 而提前于VPD,王慧梅等(2011)发现兴安落叶松(Larix dahurica)树干液流峰值滞后于PAR约0.5 h, 提前于气温和空气相对湿度0-2 h.有研究认为植物茎干夜间水分的补充是导致时滞效应的重要原因(Phillips et al., 2003; Goldstein et al., 1998; Kumagai, 2009).此外植物茎干液流与环境要素时滞效应的差异还与植物木质部结构、树干储水性能及树形因子等因素有关(Granier et al., 2000; Ford et al., 2004; Chuang et al., 2006).有研究指出, 时滞现象通常在小时间尺度发生, 当上推至日或者更大时间尺度时, 这一现象不再明显(Oren et al., 1999; O’Brien et al., 2004). ...
Canopy transpiration and water fluxes in the xylem of the trunk of Larix and Picea trees — A comparison of xylem flow, porome- ter and cuvette measurements.
... 光是影响植物蒸腾的重要环境驱动因子, 但是对气孔导度变化具有消减作用(Meinzer et al., 1995), 从而影响茎干液流变化(Motzer et al., 2005).当VPD除以PAR以消除光的影响后发现, 液流密度对较低的VPD十分敏感, 且随着VPD的增加线性增加至其最大值后缓慢降低(图11), 而O’Brien等(2004)发现多种热带雨林植物茎干液流密度随VPD增加的速率同降低的速率基本一致, 这一差异表明在空气极端干旱时梭梭通过气孔调节尽可能减少水分损失(谭永芹等, 2011).Verbeeck等(2007)发现VPD是欧洲赤松(Pinus svlestris)树干液流的主要驱动因子, 当空气十分干燥时植物蒸腾受到抑制.大气环境蒸发力对植物液流的影响还受土壤水分状况以及植物导水率等因素的综合影响(Schulze et al., 1985; Meinzer et al., 1995; Motzer et al., 2005).Oguntunde (2005)发现当土壤严重干旱时, 空气蒸腾需求仅能解释木薯(Manihot esculenta)茎干液流24%的变化. ...
干旱区五种木本植物枝叶水分状况与其抗旱性能
1
2011
... 光是影响植物蒸腾的重要环境驱动因子, 但是对气孔导度变化具有消减作用(Meinzer et al., 1995), 从而影响茎干液流变化(Motzer et al., 2005).当VPD除以PAR以消除光的影响后发现, 液流密度对较低的VPD十分敏感, 且随着VPD的增加线性增加至其最大值后缓慢降低(图11), 而O’Brien等(2004)发现多种热带雨林植物茎干液流密度随VPD增加的速率同降低的速率基本一致, 这一差异表明在空气极端干旱时梭梭通过气孔调节尽可能减少水分损失(谭永芹等, 2011).Verbeeck等(2007)发现VPD是欧洲赤松(Pinus svlestris)树干液流的主要驱动因子, 当空气十分干燥时植物蒸腾受到抑制.大气环境蒸发力对植物液流的影响还受土壤水分状况以及植物导水率等因素的综合影响(Schulze et al., 1985; Meinzer et al., 1995; Motzer et al., 2005).Oguntunde (2005)发现当土壤严重干旱时, 空气蒸腾需求仅能解释木薯(Manihot esculenta)茎干液流24%的变化. ...
Model analysis of the effects of atmospheric drivers on storage water use in Scots Pine.
1
2007
... 光是影响植物蒸腾的重要环境驱动因子, 但是对气孔导度变化具有消减作用(Meinzer et al., 1995), 从而影响茎干液流变化(Motzer et al., 2005).当VPD除以PAR以消除光的影响后发现, 液流密度对较低的VPD十分敏感, 且随着VPD的增加线性增加至其最大值后缓慢降低(图11), 而O’Brien等(2004)发现多种热带雨林植物茎干液流密度随VPD增加的速率同降低的速率基本一致, 这一差异表明在空气极端干旱时梭梭通过气孔调节尽可能减少水分损失(谭永芹等, 2011).Verbeeck等(2007)发现VPD是欧洲赤松(Pinus svlestris)树干液流的主要驱动因子, 当空气十分干燥时植物蒸腾受到抑制.大气环境蒸发力对植物液流的影响还受土壤水分状况以及植物导水率等因素的综合影响(Schulze et al., 1985; Meinzer et al., 1995; Motzer et al., 2005).Oguntunde (2005)发现当土壤严重干旱时, 空气蒸腾需求仅能解释木薯(Manihot esculenta)茎干液流24%的变化. ...
Relationships between stem diameter, sapwood area, leaf area and transpiration in a young mountain ash forest.
1
1995
... 蒸腾是植物重要的生理活动之一, 该过程一方面促进植物体内水分和营养元素的运输, 另一方面通过气孔控制间接影响光合作用(Jones, 1983).植物茎干液流(stem sap flow)的90%以上通过叶片蒸腾散失到大气中(蒋高明, 2007), 因此能够将植物茎干液流量用于估算单株或群落蒸腾并获得植物重要的生理生态信息.茎干液流受到植物生理生化特征、环境要素及树形因子的综合影响(Andrade et al., 1998; Meinzer et al., 2001), 其中环境要素主要包括光合有效辐射(PAR)、水汽压亏缺(VPD)和气温等, 而长时间尺度液流变化则受区域气候的制约(Granier et al., 1992; Vertessy et al., 1995; McDowell et al., 2008).主成分分析(PCA)能将互有联系的环境要素分为不同的复合分量, 在保留原始数据主要信息的基础上实现数据降维, 该方法对于分析植物液流对环境因子的响应十分有效(O’Brien et al., 2004). ...
马占相思树干液流与光合有效辐射和水汽压亏缺间的时滞效应
1
2008
... 黄德卫等(2012)发现, 鼎湖山针阔混交林优势种液流峰值出现的时间晚于PAR峰值出现的时间, 与VPD的时滞效应在不同季节存在明显差异, 王华等(2008)发现马占相思(Acacia mangium)树干液流滞后于PAR, 而提前于VPD,王慧梅等(2011)发现兴安落叶松(Larix dahurica)树干液流峰值滞后于PAR约0.5 h, 提前于气温和空气相对湿度0-2 h.有研究认为植物茎干夜间水分的补充是导致时滞效应的重要原因(Phillips et al., 2003; Goldstein et al., 1998; Kumagai, 2009).此外植物茎干液流与环境要素时滞效应的差异还与植物木质部结构、树干储水性能及树形因子等因素有关(Granier et al., 2000; Ford et al., 2004; Chuang et al., 2006).有研究指出, 时滞现象通常在小时间尺度发生, 当上推至日或者更大时间尺度时, 这一现象不再明显(Oren et al., 1999; O’Brien et al., 2004). ...
马占相思树干液流与光合有效辐射和水汽压亏缺间的时滞效应
1
2008
... 黄德卫等(2012)发现, 鼎湖山针阔混交林优势种液流峰值出现的时间晚于PAR峰值出现的时间, 与VPD的时滞效应在不同季节存在明显差异, 王华等(2008)发现马占相思(Acacia mangium)树干液流滞后于PAR, 而提前于VPD,王慧梅等(2011)发现兴安落叶松(Larix dahurica)树干液流峰值滞后于PAR约0.5 h, 提前于气温和空气相对湿度0-2 h.有研究认为植物茎干夜间水分的补充是导致时滞效应的重要原因(Phillips et al., 2003; Goldstein et al., 1998; Kumagai, 2009).此外植物茎干液流与环境要素时滞效应的差异还与植物木质部结构、树干储水性能及树形因子等因素有关(Granier et al., 2000; Ford et al., 2004; Chuang et al., 2006).有研究指出, 时滞现象通常在小时间尺度发生, 当上推至日或者更大时间尺度时, 这一现象不再明显(Oren et al., 1999; O’Brien et al., 2004). ...
 , 金博文
, 金博文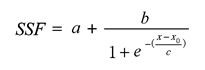
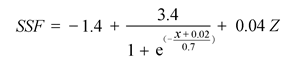


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}