版权声明: 2016 植物生态学报编辑部 本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。
展开
摘要
森林生态系统的产水量与固碳效益之间存在着一种可交易的平衡关系。基于WaSSI-C水碳耦合模型和趋势分析, 研究了1982-2006年川西杂古脑河上游22个子流域内不同植被类型空间分布对水碳平衡的影响并分析了其水碳耦合关系, 发现: 1)针叶林主导的流域在生长季增加土壤水分入渗的功能明显高于其他植被类型, 但不足以补偿其高蒸散带来的水分消耗, 因而其年平均土壤含水量明显低于高山草甸和混交林类型; 且森林土壤含水量随着森林覆盖率的升高而降低。2) 25年的土壤水分蓄变量的平均值, 高山草甸流域为-44 mm, 混交林为-18 mm, 针叶林为-5 mm, 说明川西亚高山植被的整体维持稳定产水量及其潜力在下降, 其中高山草甸流域下降趋势尤为显著。3)流域产流量和净生态系统生产力具有显著负相关性, 且不同植被组成对固碳和产水效益的转化具有重要影响: 高山草甸主导的子流域具有较高的产水量和较低的固碳能力, 常绿针叶林主导的子流域具有较高固碳能力和较低产水量, 且森林覆盖率越高, 产水量越低。三种植被类型的净生态系统生产力在研究期间均呈现上升趋势, 且高山草甸的上升趋势显著。
关键词:
Abstract
Aims There is increasing concern on the trade-off between carbon sequestration and water yield of forest ecosystems. Our objective was to explore the effects of vegetation composition on water and carbon trade-off in the sub-alpine watersheds of western Sichuan during 1982-2006.Methods The WaSSI-C, which is an eco-hydrological model with coupled water and carbon cycles, was employed to calculate the key components in water balance and carbon sequestration for the 22 sub-catchments in the upper reaches of Zagunao River. Spearman’s Rho trend analysis was used to examine the trends in runoff and net ecosystem productivity. Important findings Compared with either subalpine meadow or mixed forest dominated catchments, the conifer-dominated catchments had much higher water loss due to high evapotranspiration, and the loss was not offset by its higher soil water infiltration during the growing season. The change in soil water storage for subalpine meadow, mixed forest and coniferous forest are -44 mm, -18 mm and -5 mm, respectively, which indicated significant decline in soil water storage and thus water yield particularly in alpine meadow catchments. Significant negative relationship was found between runoff and net ecosystem productivity, the alpine meadow as the dominant vegetation type showed high water yield and low carbon sequestration, and the conifer-dominant and mixed forest vegetation showed low water yield and high carbon sequestration, moreover, the higher the forest coverage, the lower the water yield. Upward trends in net ecosystem productivity were observed in the three vegetation types during the study period and the alpine meadow type was significant.
Keywords:
为减缓气候变暖的影响, 进行大规模植被建设以增加陆地生态系统对CO2的吸收早已被联合国气候变化公约和政府间气候变化专门委员会等作为实施碳循环管理以减少碳排放的重要举措(IPCC, 2007; Canadell & Raupach, 2008)。为此, 我国在六大林业工程基础上, 承诺在2020年前, 比2005年净增森林面积4000万km2和林木蓄积13亿m3, 藉以实现通过增加森林碳汇来减缓气候变化。如此规模巨大的植被恢复与全球气候变化的影响相交互, 必将对区域水文过程、水量平衡和水土资源产生重要的影响。可见, 我国大规模植被建设与恢复工程背后隐藏着加剧我国区域水资源短缺和水资源供需不平衡的危机(Sun et al., 2006; McVicar et al., 2007)。森林生态系统的碳循环与水循环过程存在相互作用和相互耦合的关系(于贵瑞等, 2014)。一方面, 森林生态系统作为调节区域气候和水文过程的媒介和载体, 其覆盖度的增加能够起到蓄涵水源和保持水土的作用; 但另一方面, 森林生态系统也是水分的消耗者, 因此, 森林固碳与产水效益之间存在着权衡关系, 例如Jackson等(2005)认为大面积的人工林会导致土壤干化, 并首次提出碳水交易的问题, 但是在多种植被类型共存、气候变化协同影响的区域如何平衡水与碳之间的关系, 仍缺乏操作依据。
川西亚高山森林位于长江流域上游重要的水源地, 森林的采伐与恢复操作在1998年之前曾经非常频繁。随着天然林保护工程和退耕还林工程的实施, 川西亚高山区域的森林分布现状和水资源格局发生了一些重大变化。典型的岷江上游流域的植被组成在历史上主要由稀疏的天然森林植被、广阔的草原和高山草甸构成。1954年, 老龄针叶林是岷江上游的主导森林类型, 而森林大规模采伐以后, 1965年的森林植被主导类型就转变为次生林和人工针叶林, 岷江上游森林恢复时期21%的灌丛转化为郁闭森林、疏林地以及人工针叶林, 而17%的农田已转变为灌丛和草地(Zhou et al., 2008)。Zhang等(2012)基于长期水文数据(1953-1996), 结合时间序列分析和双累积曲线, 对岷江上游杂谷脑流域的气候变化和森林采伐对径流的影响做了定量分析, 结果表明平均年径流量随森林采伐而显著增加, 采伐对径流的贡献量为38 mm·a-1, 而气候变化对径流的贡献量为-27 mm·a-1。气候变化的作用主要可归因于增温提高了植被活动强度, 提高了植被固碳能力, 从而增加了流域水分消耗(Sun et al., 2008)。这种正负并存的相关性结果表明了森林采伐和植被恢复对径流的影响存在着抵消关系。同时, 还发现随森林植被的恢复, 森林采伐对径流的影响不断削弱, 直到采伐后20年水文功能上完全恢复, 从而在水碳循环中建立新的平衡关系。
因此, 我们需要分析这种新的水碳平衡关系与植被组成的关系及演变方向是什么。本文以生态水文模型WaSSI-C为工具, 分析经过较大程度人工和气候变化干扰后杂谷脑流域的不同植被组成的水碳耦合关系, 以揭示其区域特征和演变方向, 为该区天然林保护、植被恢复与流域经营管理提供科学 参考。
本研究选取杂古脑河上游流域为研究区(102.58-103.22° E, 31.18-31.93° N)。该流域位于中国四川省的西北部, 出口为杂古脑水文站, 流域总面积为2528 km2。杂古脑河隶属于长江流域重要水系岷江流域的主要支流之一。该流域坐落在川西高山峡谷地带, 流域海拔1823-5769 m, 平均海拔3859 m, 其中56.8%的区域海拔在3800 m以上, 有3.5%的区域被冰川和永久积雪覆盖。
杂古脑流域夏季凉爽, 冬季寒冷, 属于典型的高山气候。年平均气温为11.2 ℃, 最高气温在7月, 可达26.9 ℃, 最低气温在1月, 可达-10 ℃。另外, 由于杂古脑流域海拔较高, 高山林立, 区域之间的气候差异较大。受海拔和地形条件的影响, 流域多年平均气温空间变异很大(-1.7-12 ℃), 沿东南–西北方向递减。夏季主要受印度洋西南季风的影响, 形成5至9月的雨季。年降水量在627.5-1478.0 mm范围内波动, 平均为1067.6 mm, 流域雨季降水量为880 mm, 占年降水量的约80%。
流域主要的植被类型为高山草甸、亚高山常绿针叶林和亚高山灌丛, 主要针叶树种为岷江冷杉(Abies faxoniana)和云杉(Picea asperata)。研究区的植被类型和生境随海拔及坡向存在显著的分异: 阔叶林主要分布在海拔1800-2500 m的区域内, 针叶林主要分布在海拔2500-3500 m, 高山草甸主要分布在海拔4500 m以上区域。该研究区经历了1950到1978年大面积的采伐, 之后逐渐减少, 到1998年完全停止采伐。此后植被逐渐恢复, 采伐20年后植被完全恢复为次生阔叶林。人工恢复方式主要以营造人工林为主, 其中主要树种为云杉; 人为干扰过于频繁的地段部分退化为灌丛或者草甸。
本研究的主要方法建立在水碳耦合模型WaSSI-C的基础上。WaSSI模型是美国林务局东部森林环境威胁评估中心EFETC开发的月尺度生态系统水碳耦合模型(Sun et al., 2011a, 2011b)。WaSSI-C是由水分供需计算模型(WaSSI)和碳循环经验模型集成的月尺度生态系统水碳耦合模型。该模型采用分布式与集总式结合的方法, 可将栅格化遥感、植被、水文、土壤等数据输入模型进行运算, 并采用水文响应单元(HRU)进行子流域参数汇总。有关模型用到的数据集、子流域划分依据、参数率定和检验可参考我们前期发表的论文(刘宁等, 2013a; 刘宁等, 2013b)。
根据岷江上游区域范围内及周边51个雨量站收集的降雨数据, 并根据一个四变量的插值方式获得研究期间每月的降水空间数据, 相关技术方法见孙鹏森等(2004)。WaSSI-C内部的水文模型为萨克拉门托水文模型所需, 土壤参数基于中国科学院南京土壤研究所的区域土壤基础数据库(史学正等, 2007)。土壤水分重要参数的率定表见刘宁等(2013a)。模型参数化需要的遥感数据(例如归一化植被指数(NDVI)、叶面积指数(LAI)等)来源于MODIS全球共享数据。
本研究进行流域生态水文过程模拟的时间段是1982-2006年。统计分析是以HRU为基本单位进行的, 可对包括蒸散(ET)、径流(RUNOFF)、总生态系统生产力(GEP)、净生态系统碳交换(NEE)、生态系统呼吸消耗量(REC)等在内的水、碳循环关键过程进行模拟。根据地形条件和植被分布, 以及模型运行的效率系数, 杂谷脑流域最终被确定为22个子流域(刘宁等, 2013a), 作为基本的HRU进行统计分析。
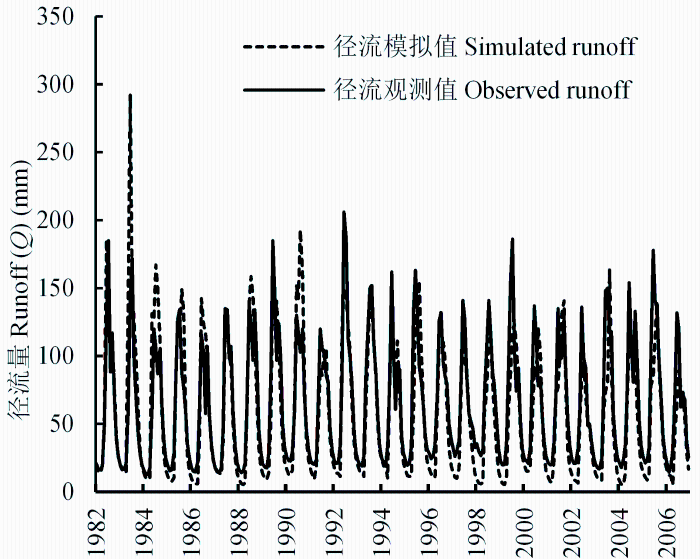
模拟时间段可分为两段: 参数率定期(1988- 1996年)和验证期(1997-2006年)。模型经过参数率定、校验后,最后模拟出基于月尺度的流域水碳循环过程的各个分量的时空变化过程。利用已发表数据、公共数据以及实测数据,我们对各个分量进行了验证。其中,流域径流量(Q), 利用流域总出口水文站(杂谷脑水文站)实测水文数据进行验证。结果表明, WaSSI-C率定期检验的模拟的决定系数R2 = 0.86 (p < 0.001), 总体检验Nash-Sutcliffe效率系数为0.82 (刘宁等, 2013b), 月尺度水文过程模拟效果达到统计要求(图1)。由于杂谷脑水文站以上22个子流域没有水文站数据验证, 推定各子流域汇水出口的径流输出量模拟值均达到此检验标准, 并在水量平衡的计算中采用模拟值。
其他关键参数的验证: WaSSI-C模拟的蒸散ET (下称ET)分别通过国际上公共数据和已发表的数据进行了检验。结果显示, 在HRU上, ET与MODIS/ET数据, 拟合线斜率为1.41, R2 = 0.9; ET与Zhang等(2010)的全球ET数据拟合斜率为1.13, R2 = 0.86。WaSSI-C模拟的GEP(下称GEP)模拟值与MODIS/ GEP值对比的R2为0.89, 拟合线的斜率为0.85; 在全流域水平的验证R2为0.91, 拟合线的斜率为0.88 (刘宁等, 2012)。
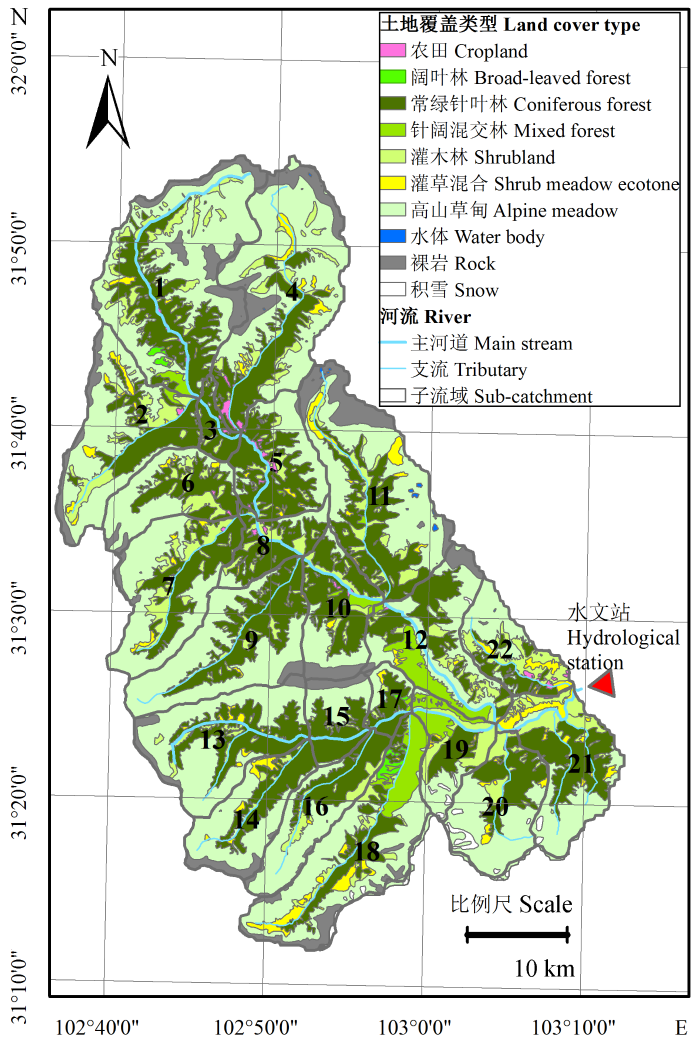
植被分类数据最初来源于TM (2000年)影像的分类结果, 并根据岷江流域内756个验证点数据达到87.6%的可靠性(Sun et al., 2008)。杂古脑流域的植被类型主要为高山草甸和亚高山常绿针叶林, 二者的比例之和占流域总面积的80%左右。流域植被的分布具有严格的垂直地带性, 从高海拔到低海拔依次为高山草甸、灌丛、常绿针叶林、针阔混交林和阔叶林(图2)。基于子流域划分方案可以得到每个子流域的植被类型比例, 同时基于其主要植被类型, 将这些子流域划分为3类: 高山草甸主导的流域(高山草甸覆盖率≥0.5)、常绿针叶林主导的流域(森林覆盖率≥50%且针叶林覆盖率>50%)和混交类型(森林覆盖率≥50%, 且针叶林<50%)。混交类型中没有主导植被类型, 其中包含针阔混交林, 例如针叶树种和桦木(Betula platyphylla, Betula utilis)次生林
以及针叶林和常绿灌丛。大面积分布的川滇高山栎(Quercus aquifolioides)中有部分区域达到郁闭状态, 因此, 在总体森林覆盖率的统计过程中, 将其视为森林进行统计。其中以高山草甸主导的子流域包括子流域编号1、2、4、6、7、9、13、14、16、20; 以常绿针叶林主导的子流域包括: 3、5、8、10、15、17, 其中子流域3的常绿针叶林比例占到91%; 没有针阔混交和灌丛植被类型的子流域包括: 11、12、18、19、21、22。每个子流域植被类型的比例如表1所示。
图1 杂谷脑水文站月尺度水文过程观测与模拟数据的比较。
Fig. 1 A comparison of monthly runoffs (simulated vs observed) guaged by Zagunao Hydrology Station.
图2 杂古脑河上游各子流域的土地覆盖类型分布图(图中数字表示子流域编号)。
Fig. 2 Land cover types for sub-catchments in upper reaches of Zagunao river (numbers in graph are serial numbers of sub-catchments).
表1 杂谷脑河流域上游各子流域的地形及植被组成
Table 1 Topography and vegetation composition for sub-catchments in upper reaches of Zagunao river
| 子流域编号 Serial number of sub-catchment | 地形特征 Topography characteristics | 植被组成 Vegetation composition | ||||||
|---|---|---|---|---|---|---|---|---|
| 面积 Area (km2) | 坡度 Slope (°) | 平均海拔 Mean altitude (m) | 针叶林覆盖率 Coniferous forest coverage (%) | 森林覆盖率 Forest coverage (%) | 高山草甸覆盖率 Alpine meadow coverage (%) | 主导植被类型 Dominant vegetation type | ||
| 3 | 20.0 | 51.5 | 4 077 | 91 | 99 | 0 | 针叶林 Coniferous forest | |
| 5 | 89.2 | 56.4 | 3 838 | 54 | 65 | 33 | ||
| 8 | 49.8 | 59.4 | 3 924 | 61 | 81 | 19 | ||
| 10 | 84.4 | 63.2 | 3 948 | 50 | 75 | 25 | ||
| 15 | 47.6 | 76.5 | 3 355 | 50 | 64 | 36 | ||
| 17 | 25.6 | 68.8 | 3 647 | 60 | 68 | 32 | ||
| 11 | 195.9 | 58.4 | 4 010 | 28 | 53 | 42 | 混交林 Mixed forest | |
| 12 | 127.9 | 69.3 | 3 322 | 28 | 6 | 38 | ||
| 18 | 171.2 | 65.9 | 3 937 | 18 | 5 | 44 | ||
| 19 | 59.6 | 71.6 | 3 866 | 24 | 76 | 23 | ||
| 21 | 112.5 | 66.7 | 4 017 | 39 | 58 | 34 | ||
| 22 | 66.0 | 59.1 | 3 543 | 22 | 48 | 43 | ||
| 1 | 251.5 | 49.4 | 4 004 | 27 | 48 | 51 | 高山草甸 Alpine meadow | |
| 2 | 146.2 | 54.9 | 3 995 | 25 | 47 | 50 | ||
| 4 | 221.7 | 58.9 | 3 371 | 21 | 41 | 55 | ||
| 6 | 82.5 | 59.3 | 3 672 | 33 | 52 | 49 | ||
| 7 | 132.7 | 61.3 | 3 446 | 30 | 47 | 51 | ||
| 9 | 149.1 | 60.9 | 3 628 | 37 | 43 | 57 | ||
| 13 | 139.8 | 73.6 | 3 348 | 32 | 43 | 55 | ||
| 14 | 87.9 | 69.6 | 3 998 | 33 | 45 | 51 | ||
| 16 | 81.2 | 70.6 | 3 348 | 36 | 51 | 50 | ||
| 20 | 96.9 | 66.6 | 4 016 | 29 | 43 | 54 | ||
三组主导植被类型基本上代表了该区域森林植被分布的现状。目前该区域的主要植被类型包括以岷江冷杉为优势树种的原始老龄针叶林、以云杉为主的人工针叶林、以云冷杉和桦木为主要树种的针阔混交林、以川滇高山栎为主要优势树种的常绿阔叶林。
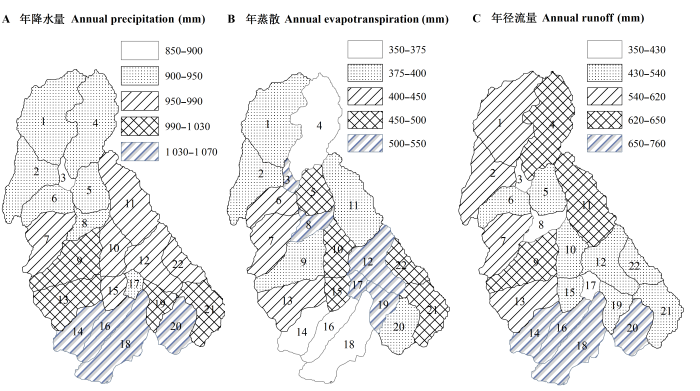
从流域的降水格局看, 流域年降水量在890- 1073 mm之间, 且具有明显的空间分布特征, 年降水量由南向北逐渐减少(图3A)。其中子流域1、3、6的年降水量均小于940 mm, 属于降水相对较少的区域; 流域14、16、18、20的年降水量均在1030 mm以上, 降水充沛。总体上流域年降水在900 mm以上, 属于降水充足地区。
流域年蒸散量也具有明显的垂直分布特征, 处于高海拔的子流域蒸散量较低; 相反, 处于低海拔的河道下游子流域具有较高的年蒸散量(图3B), 这与气温和植被类型梯度是一致的。流域的年蒸散量在367-542 mm之间, 年蒸散量大于500 mm的子流域分别为3、8、12、17、19, 这些子流域主要以常绿针叶林和混交林为主导。而流域年径流(Q)的情况正好相反, 位于高海拔的草甸主导的植被类型产流量较高, 而中低海拔的亚高山森林产流量较低(图3C)。在季节尺度上, 杂古脑流域的水文过程具有明显的季节变异性。据统计, 夏季(6-8月)由于受雨季和高温的影响, 该流域具有较高的蒸散量和流域产流量, 整个夏季的蒸散量可达216 mm, 占全年总蒸散量的50%; 径流量可达360 mm, 占全年径流量的51%。流域冬季主要受北方寒冷气候影响, 整个流域被积雪覆盖, 此时流域的蒸散量最低, 整个冬季的蒸散量仅有21 mm, 而冬季的产流量却有45 mm, 二者之和是流域冬季降水量的2倍多(降水量为27 mm), 可见该流域冬季仍然存在较大地下水补给径流量, 这与在相邻的黑水流域进行的同位素观测结果(Liu et al., 2008)是一致的。
图3 杂古脑河上游各子流域的年降水量(A)、年蒸散量(B)和年径流量(C)的空间分布(图中数字表示子流域编号)。
Fig. 3 Spatial patterns of mean annual precipitation (A), evapotranspiration (B) and runoff (C) of Zagunao upper reaches (numbers in graph are serial numbers of sub-catchments).
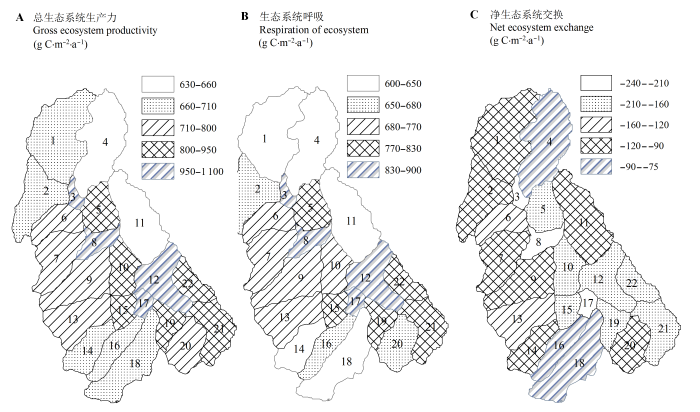
模型模拟的碳循环过程的参数包括总生态系统生产力(GEP)和净生态系统交换(NEE), 以常绿针叶林和混交林主导的流域具有较高的生产力, 这些流域的GEP在637-1070 g C·m-2·a-1之间, NEE在 -240- -70 g C·m-2·a-1之间, 其中以常绿针叶林主导的子流域固碳能力最大, 如子流域3的GEP为1066 g C·m-2·a-1, NEE为-240 g C·m-2·a-1。尽管子流域16和18具有充足的降水(年降水量>1000 mm), 但是两个子流域的海拔均在4000 m以上, 另外这两个子流域的主导植被类型均为高山草甸, 因此有>70%的水分转换为地表径流流出, 表现出低固碳能力。子流域16和18的GEP和NEE分别为704、670和-87、-76 g C·m-2·a-1 (图4)。
在季节尺度上, 杂古脑流域的碳循环过程参数也表现出明显的季节变异性。其中夏季(6-8月)流域植被进行旺盛生长, 整个夏季的GEP可达435 g C·m-2·a-1, 占全年总GEP的53%; NEE可达-102 g C·m-2·a-1, 占全年总NEE的86%。可见尽管其他季节植被仍在生长, 但是却有更多的有机物被呼吸消耗。流域冬季主要受北方寒冷气候的影响, 整个流域处于积雪覆盖的区域, 此时流域的植被基本停止生长, 但是仍有少量的呼吸消耗。空间分布上各个季节的碳循环通量分布具有相同的变化特征, 即海拔较低, 以常绿针叶林或者是混交林主导的流域具有较高的生产力, 相反, 以高山草甸主导的区域海拔较高温度较低, 因而具有较低的生产力。
图4 杂古脑流域平均年总生态系统生产力(A)、生态系统呼吸(B)和净生态系统交换(C)的空间分布(图中数字表示子流域 编号)。
Fig. 4 Spatial patterns of annual gross ecosystem productivity (A), net ecosystem exchange (B), respiration of ecosystem (C) of Zagunao upper reaches (numbers in graph are serial numbers of sub-catchments).
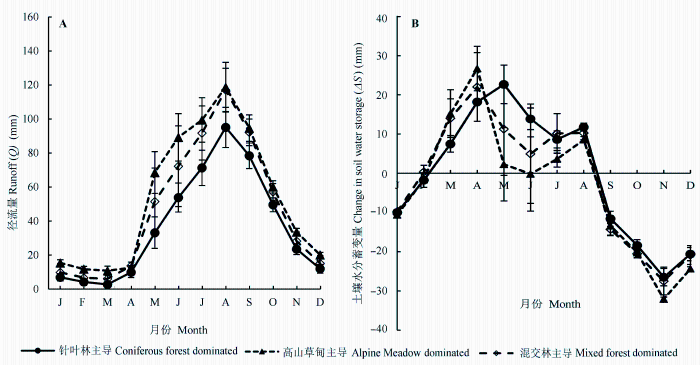
本研究对22个子流域的分类统计结果表明, 以针叶林主导和混交林类型的流域产流量明显低于草甸类型的流域, 这说明无论在哪个季节, 森林覆盖率高的植被类型产流量明显小于草甸主导的类型; 但同时在森林主导的流域内, 针叶林覆盖率高的流域产流量小于那些针阔混交的类型(图5A)。由于产流量仅是森林植被水源涵养功能的一个指标, 据此还不能完全判断森林对水源的涵养功能不及草甸。我们采用研究期间25年土壤水分蓄变量(ΔS = P - ET - Q)的平均值来分析, 不同植被类型将降水转化为土壤水分储存, 并实现消洪补枯的能力。结果表明, 在生长季, 针叶林和混交林ΔS明显高于高山草甸, 而非生长季没有明显差异(图5B)。
图5 不同主导植被类型下流域水量平衡要素的季节动态(平均值±标准误差)。
Fig. 5 Seasonal dynamics of water balance components under different dominating vegetation type (mean ± SE).
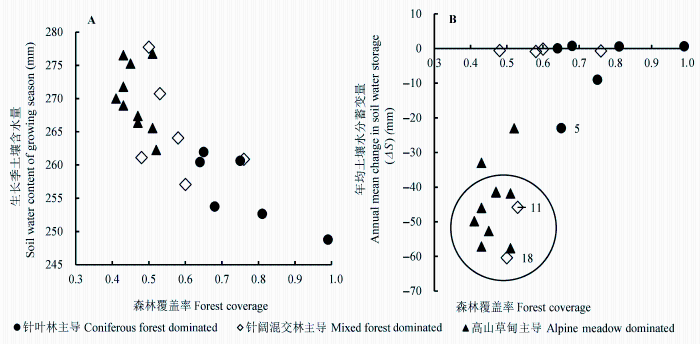
图6 土壤含水量(A)、土壤蓄水变量(B)与植被覆盖率之间的关系(图B中的数字为子流域编号, 圆圈内为水分流失严重的子流域)。
Fig. 6 The relationships between forest coverage and soil water content (A), and soil water storage change (B) (Numbers in Figure B denote sub-catchments and the severe water loss sub-catchments are inside circle).
进一步分析森林覆盖率与生长季(5-10月)土壤含水量之间的关系发现, 尽管森林覆盖率高的类型土壤水分蓄变量(ΔS)高于其他两个类型, 但其土壤含水量的绝对值(SWC)仍然低于其他两个类型, 且随着森林覆盖率的升高, 生长季土壤含水量呈现显著的线性下降趋势(图6A)。用25年的平均ΔS显示每个子流域的总体水量平衡状态, 发现针叶林主导的流域ΔS为-5 mm, 混交林主导的流域ΔS为-18 mm, 两者虽然略有亏缺, 但基本维持平衡状态, 而高山草甸主导的流域出现较为严重的水分失衡, ΔS为-44 mm (图6B)。
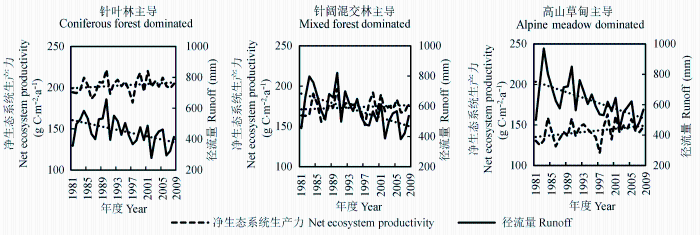
为了定量地探讨流域产水与固碳的权衡关系, 经常以生态系统的产水量(Q, 即径流量)和净生态系统生产力(NEP)作为其服务功能的重要标准, 本文采用这两个指标进行衡量。我们采用生长季(4-10月)时间序列进行统计, 这样可以排除因落叶树种或者草本在非生长季的变化因素, 以便不同主导类型之间比较研究。Spearman’s Rho时间序列检验发现, 在研究时间(1982-2008年)内, 各植被类型主导的流域的径流量(Q)呈现显著的且一致的下降趋势(统计量z均为负值且显著), 其中草甸主导的流域(z = -3.0, p < 0.01)和混交林主导的流域(z = -2.8, p < 0.01)显著度高于针叶林主导的流域(z = -2.5, p < 0.05), 三类主导植被类型的流域模拟结果基本一致, 且总体模拟结果基本符合杂谷脑水文站实测水文数据。NEP的趋势变化方向恰好与流域径流量的变化趋势相反, 总体呈现上升趋势, 其中针叶林(z = 1.2, p > 0.1)和混交林主导(z = 1.3, p > 0.1)的植被类型上升趋势不显著, 而草甸主导的类型上升显著(z = 2.09, p < 0.05)(图7)。
图7 不同主导植被类型流域的生长季平均径流量与净生态系统生产力的变化趋势检验(Spearman’s Rho)。
Fig. 7 Spearman’s Rho trend analysis of runoff and net ecosystem productivity for different vegetation dominated catchments.
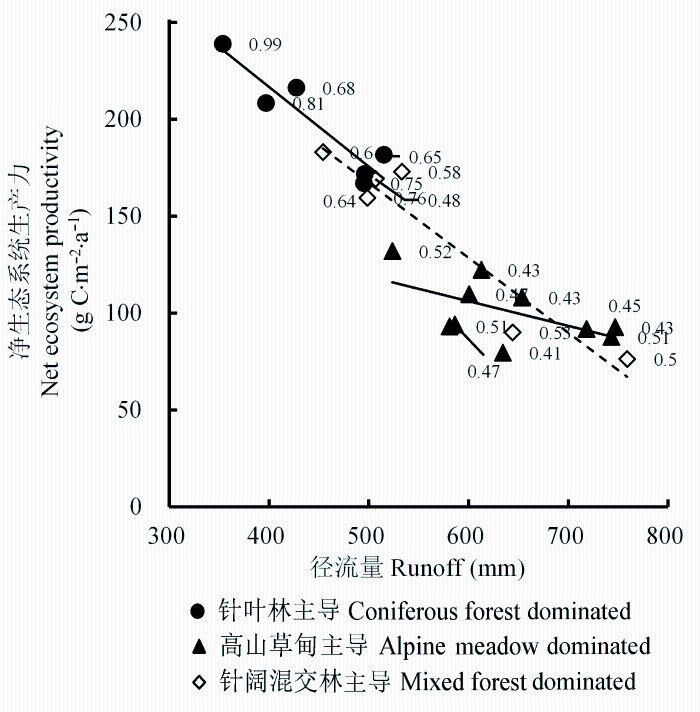
另外, 对22个不同植被组成类型子流域的多年平均Q与NEP进行回归分析, 发现二者存在显著的负相关关系(NEP = -0.4 × Q + 365, R2 = 0.84, p < 0.001)(图8), 即二者存在显著的权衡关系。通过分析各植被类型在总体拟合回归线上的分布, 我们发现以下三个规律: 1)不同主导植被类型的子流域具有明显的分界, 以高山草甸主导的子流域具有较高的产水量, 而固碳能力较低, 这些子流域一般分布在径流量600-800 mm、固碳能力在50-120 g C·m-2·a-1的值域范围内。与此相反, 常绿针叶林主导的流域具有较低的产水量, 而拥有较高的固碳能力, 分布在径流量350-550 mm之间, 固碳能力在120-250 g C·m-2·a-1之间。没有主导植被类型混交林的流域分布在二者之间具有中等的产水量和固碳能力。2)无主导植被类型的混交林子流域具有较高的水碳转换比例(拟合方程的斜率为0.56), 而高山草甸为主的子流域的水碳转换效率较低(拟合方程的斜率为0.12)(图8), 针叶林的水碳转换比率居中, 约为0.41。3)在针叶林和混交林类型中, 森林覆盖率决定不同植被组成的流域在回归拟合线上的位置。森林覆盖率高的流域位于水碳平衡线的左侧, 即高碳固定, 低产流量; 而森林覆盖率低的流域位于水碳平衡线的右侧, 即高产流量, 低碳固定(图8)。
图8 杂谷脑河流域上游产水量与净生态系统生产力的关系(图中数值标志为子流域森林覆盖率)。
Fig. 8 Relationship between water yield and net ecosystem productivity in the upper reaches of Zagunao river (numerical values in plot denote forest coverage for sub-catchment).
森林植被对水文的影响已经争论了一个多世纪。迄今为止, 比较一致的观点认为森林采伐或森林面积的减少将增加流域径流(Bosch & Hewlett, 1982; Andréassian, 2004)。但在我国长江流域早期的观点认为流域产水量随森林覆盖率的减少而减少(马雪华, 1987; 程根伟, 1993)。石培礼和李文华(2001)也认为, 除长江中上游外, 森林砍伐会降低植被层的蒸发散, 增加河川径流。
长江中上游区域与世界上大多数的区域真的会有区别吗?本研究表明, 在生长季, 森林调节土壤水分的功能高于高山草甸, 原因是森林具有良好的截留降雨的能力, 同时林下土壤发育较好, 含蓄水源的功能得以发挥。同时, 本研究还发现了重要的时间节点: 4月中旬和8月中旬, ΔS发生了明显的变化。4月中旬ΔS迅速降低与融雪径流有关, 而8月中旬ΔS的快速降低与雨季结束有关, 而此时森林植被生长仍茂盛, 因而土壤水分大量消耗。而11月到4月份, ΔS不断上升, 这主要与积雪效应有关, 这部分水分由于没有发生转移, 模型运算过程中仍被视作土壤储存的水分(图4B)。
总体上看, WaSSI-C模型对于森林年蒸散量的计算结果与Jiang等(2004)以及Zhang等(2008)在岷江上游区域的蒸散估算基本一致,均为480-520 mm; 高山草甸主导的子流域一般处于高海拔地区, 年蒸散量较低, 如子流域1、4、14、16、18等, 这些子流域的年蒸散量均在400 mm以下。根据Budyko理论曲线,杂古脑流域的多年平均干燥指数(DI)较小(<0.6), 因此可知该流域蒸散并非受水分限制,而是属于能量限制的区域, 流域蒸散量主要受太阳辐射、气温和风速等能量输入的条件限制(Budyko, 1974)。土壤平均含水量(SWC)是衡量年际间水量平衡的重要指标, 我们的研究结果是针叶林<混交林<高山草甸, 由此可以推断, 尽管森林, 尤其是针叶林在增加土壤水分入渗方面虽然具有一定的功能, 但其作用不足以补偿其水分消耗(高蒸散)而带来的土壤失水现象, 换言之, 森林的土壤含水量随着覆盖率的升高而降低, 总体上森林相比其他类型是高耗水的。
但是, 森林维持水文系统的稳定性高于草甸。综合平均25年的土壤水分蓄变量(ΔS)值, 我们发现了研究区域内不同植被类型土壤水分平衡的变化趋势, 该区域约1/2的流域处于水分减少的状态, 其中高山草甸的流域减少最显著。针叶林、混交林大部分类型水文系统基本维持平衡, 但略有水分亏缺。这一结果与其产水能力的分析并不矛盾, 这反而说明, 处于较高海拔区域的高山草甸植被类型受到增温的影响较为显著, 从而使其活动强度增加, 物候期延长并致使蒸散升高, 改变了原来的水文平衡状态(Sun et al., 2008), 另有研究表明, 川西地区季节性积雪和融雪也在减少(刘巧等, 2011)。这些原因最终会导致流域水资源供给减少, 从长远来看, 其下游的针叶林生长也会受到影响。另外部分森林类型 (子流域编号5, 11, 18)中也明显发生土壤水分亏缺, 虽然这几个流域总体森林覆盖率高于50%, 但是其针叶林比率较低, 植被类型多为次生阔叶林或者郁闭灌木, 草甸的比例均高达40%, 所以其土壤水分状况类似于草甸。
流域的管理, 需要综合考虑不同植被类型的综合效益, 制定合理的经营管理措施, 因而需要进行量化的分析。水碳效益之间的权衡可以辅助量化地确定森林固碳和产水目标位置。从不同主导植被类型NEP和Q之间的关系(图8)以及变化趋势(图7)可以看出, NEP和Q之间存在着此消彼长的权衡关系。水碳平衡本质上是生态系统自身特征导致的其对蓝水和绿水之间的长期分配关系, 即生态系统维持生产力与产水量平衡的状态。图8中总体拟合线的斜率应该体现不同流域的特点, 这包含植被组成特征、气温、降水等自然条件, 也可能与植被的水分利用效率和蒸散的限制机制有关, 例如水分限制或能量限制的改变(McVicar et al., 2012), 尚待进一步研究。在一个流域中, 不同植被类型的水分利用效率混合在一起, 形成流域整体的水分利用效率, 从而决定着生态系统的水分是用于形成生产力还是输出为产水量, 这是水碳效益权衡关系的生态学机理。通过调整各植被类型的组成和比例可以实现水碳平衡拟合线重心转移, 达到相应的目标效益。例如流域森林的面积比重增加或覆盖率增加, 其拟合线重心会向左侧移动, 区域整体会增大固碳效益, 而减少产流量; 反之则增加产流, 减少固碳(图8)。
植被结构组成的调整方向和强度应该根据流域的总体水分承载力决定, 具体可参考子流域的年水分蓄变量(ΔS)指标, 使其总体维持稳定, 不可盲目偏重单一效益。以杂谷脑流域为例, 通过NEP-Q的相关分析我们看出, 高山草甸主要分布于高海拔地区, 本身固碳能力较低, 并且水分利用效率低, 因此要产生相同产量的碳, 必然会消耗更多的水分。尽管常绿针叶林主导的子流域具有较高的固碳能力和水文调节功能, 但是其对径流的影响较大, 水碳之间的转换效率也不高。近期欧洲的研究结果发现, 混交林在养分利用、水分利用以及光能利用方面均高于纯林, 在应对气候变化方面具有优势(Forrester, 2014, 2015)。在本研究中,从拟合线的斜率来看, 混交林类型具有稍高于针叶林的水碳转换效率。但是由于目前该混交林类型在流域的总体森林覆盖中的比重不高, 因此不是主导整个杂谷脑流域的水碳耦合特征的类型。
从图8的拟合关系线上不同子流域的分布位置看, 当前不同植被组成的子流域基本上均匀分布, 没有发生重心偏移, 从而流域上整体水碳关系维持在大致平衡的状态。区域整体的拟合线斜率和针叶林主导的植被类型比较接近, 说明该区域植被特征仍然维持以亚高山暗针叶林为主体。但是, 随着区域性增温等自然因素的变化, 高山草甸主导的类型短期内覆盖度和植被指数以及生产力提高显著(Sun et al., 2008), 从而使得草甸类型的流域在水碳平衡线整体上向左侧移动, 势必加重水分需求。本研究也发现该类型流域产水量显著降低、而生产力显著上升的状况, 但同时, 其土壤水分蓄变量ΔS多年平均值为负值, 说明这种趋势是气温升高造成的短期效应, 具有不可持续性, 如果水分减少的趋势不改变, 未来将可能出现季节性土壤干化现象, 从而影响整个流域上下游植被的水分供应。现在已经发现部分造林地人工云杉林有枯梢现象, 因此不应大力发展高密度的人工纯林, 应依据自然条件, 适当增加混交林的比例。在低海拔区域进行植被恢复, 将已有的人工云杉林地进行疏伐时, 应充分权衡其水分需求和水量平衡状况, 以提高水分利用效率, 实现水碳双赢。
杂古脑流域水碳循环过程具有明显的时空变异性, 并且其空间分布与植被类型和海拔具有较强的相关性。主要表现在: 以高山草甸为主的高海拔区域, 具有高径流量、低蒸散量和低固碳能力; 相反, 以常绿针叶林和混交林植被类型为主的较低海拔区域, 具有高蒸散量、高固碳能力和低径流量。
不同植被类型相比, 针叶林在生长季增加土壤水分入渗方面具有一定的功能, 但其作用不足以补偿其水分消耗带来的土壤含水量降低, 因而针叶林下土壤水分低于高山草甸和混交林类型。土壤含水量随着森林覆盖率的升高逐渐降低, 森林的耗水是显著的。
通过25年的土壤水分蓄变量(ΔS)值, 我们发现了研究区域不同植被类型土壤水分平衡的变化趋势, 该区域约一半处于水分减少的状态, 其中高山草甸的流域减少最为显著。针叶林、混交林大部分类型基本维持水分平衡。这说明从趋势上看, 高山草甸流域的产水能力在下降, 一方面可能由于本身受增温的影响而生产力增加, 另一方面, 季节性积雪和融雪也在减少。这最终会对流域水资源供给以及其下游的针叶林产生长远影响。
三种植被类型的NEP在研究期间均呈现上升趋势, 且高山草甸的上升趋势显著。流域产水量(Q)和固碳(NEP)存在显著的负相关性, 水碳之间存在定量的交易和转换关系, 固碳则必然以耗水为代价, 区域性的水碳平衡拟合线是进行水碳关系调整的依据。从经营管理的角度讲, 调整森林覆盖率或者植被类型组成均可以实现水碳效益的转移。在西南地区低海拔区域进行植被恢复时, 应充分权衡其水分需求和水量平衡状况, 以提高水分利用效率, 实现水碳双赢。
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
| [1] |
Waters and forests: From historical controversy to scientific debate. |
| [2] |
A review of catchment experiments to determine the effect of vegetation changes on water yield and evapotranspiration. |
| [3] |
|
| [4] |
Managing forests for climate change mitigation. |
| [5] |
Evaluation on protective forest works at the upper reaches of the Changjiang River through the forest hydrologic effect. 从森林水文作用看长江上游防护林工程. 山地研究, |
| [6] |
The spatial and temporal dynamics of species interactions in mixed-487 species forests: From pattern to process. |
| [7] |
Transpiration and water-use efficiency in mixed-species forests versus monocultures: Effects of tree size, stand density and season. |
| [8] |
|
| [9] |
Trading water for carbon with biological carbon sequestration.
|
| [10] |
The influence of vegetation type on the hydrological process at the landscape scale.
|
| [11] |
a). Determination of the response unit scale of WASSI-C eco-hydrological model―A case study on the upper Zagunao River watershed. a). WASSI-C生态水文模型响应单元空间尺度的确定——以杂古脑流域为例. 植物生态学报, |
| [12] |
b). Coupling simulation of water-carbon processes for catchment―Calibration and validation of the WaSSI-C model, b). 流域水碳过程耦合模拟——WaSSI-C模型的率定与检验. 植物生态学报, |
| [13] |
Surface ablation features and recent variation of lower ablation area of the Hailuogou Glacier, Mt. Gongga. 嘎山海螺沟冰川消融区表面消融特征及近期变化. 冰川冻土, |
| [14] |
Characteristics of water isotopes and hydrograph separation during the wet season in the Heishui River, China. |
| [15] |
Preliminary study on hydrologic function of fir forest in Miyaluo region of Sichuan. 四川米亚罗地区高山冷杉林水文作用的研究. 林业科学, |
| [16] |
Developing a decision support tool for China’s re-vegetation program: Simulating regional impacts of afforestation on average annual streamflow in the Loess Plateau, |
| [17] |
Less bluster ahead? Overlooked ecohydrological implica- tions of global trends of terrestrial near-surface wind speeds.
|
| [18] |
Influence of forest cover change on hydrological process and watershed runoff. 森林植被变化对水文过程和径流的影响效应. 自然资源学报, |
| [19] |
Soil information system of China (SISChina) and its application. 中国土壤信息系统(SISChina)及其应用基础研究. 土壤,
|
| [20] |
a). A general predictive model for estimating monthly ecosystem evapotrans- piration. |
| [21] |
b). Upscaling key ecosystem functions across the conterminous United States by a water-centric ecosystem model. Journal of Geophysical Research, 116, |
| [22] |
Potential water yield reduction due to forestation across China. |
| [23] |
Hydrologic effects of NDVI time series in a context of climatic variability in an upstream catchment of the Minjiang River.
|
| [24] |
Estimation of precipitation using altitude and prevailing wind direction effect index in Mountainous region. 基于地形和主风向效应模拟山区降水空间分布, |
| [25] |
Fundamental scientific issues, theoretical framework and relative research methods of carbon-nitrogen-water coupling cycles in terrestrial ecosystems. 陆地生态系统碳-氮-水耦合循环的基本科学问题、理论框架与研究方法.
|
| [26] |
A continuous satellite-derived global record of land surface evapotranspiration from 1983 to 2006.
|
| [27] |
Evapotranspiration in the meso-scale forested watersheds in Minjiang Valley, West China.
|
| [28] |
The effect of forest harvesting and climatic variability on runoff in a large watershed: The case study in the upper Minjiang River of Yangtze River Basin. |
| [29] |
Vegeta- tion dynamics and forest landscape restoration in the upper Min River watershed, Sichuan, China.
|

/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}